Submitted:
01 October 2025
Posted:
02 October 2025
You are already at the latest version
Abstract
Type 2 diabetes mellitus (T2DM) is a global health burden characterized by hyperglycemia, chronic inflammation, and oxidative stress, which together drive vascular dysfunction and micro- and macrovascular complications. Red blood cells (RBCs) are particularly vulnerable to oxidative and inflammatory injury, resulting in altered morphology, reduced deformability, increased aggregation, and impaired membrane fluidity. These abnormalities elevate blood viscosity, disrupt microcirculatory flow, and promote pathological thrombogenesis, thereby exacerbating cardiovascular risk in T2DM. While conventional therapies target glycemic control and thrombosis, there remains a need for adjunctive interventions that address the underlying hemorheological derangements. Earthing (grounding), defined as direct contact with the Earth’s surface, provides a continuous influx of free electrons that may neutralize reactive oxygen species and modulate inflammatory responses. Evidence indicates that grounding can improve redox balance, normalize diurnal cortisol rhythms, and enhance blood rheology by increasing RBC zeta potential, reducing aggregation, and lowering viscosity. Furthermore, mitochondrial studies indicate that grounding stabilizes electron transport, reduces reactive oxygen species generation, and improves ATP synthesis, providing a mechanistic basis for its systemic effects. Collectively, this suggests that earthing can be a safe, low-cost, and non-pharmacological adjunct with potential to ameliorate RBC dysfunction and vascular complications in T2DM. Rigorous clinical trials are warranted to establish its efficacy and therapeutic integration.
Keywords:
earthing
; inflammation
; Type 2 diabetes mellitus
; red blood cells
1. Introduction
Diabetes mellitus is among the most prevalent chronic conditions worldwide and represents the second leading cause of reduced global healthy life expectancy.[1,2] Type 2 diabetes mellitus (T2DM) accounts for over 90% of global diabetes cases and its incidence continues to rise at an alarming rate. T2DM is now recognized as the third leading risk factor for premature mortality worldwide, primarily due to complications driven by persistent hyperglycemia, chronic inflammation, and oxidative stress.[3] These interlinked pathophysiological processes contribute significantly to the development of diabetes-related complications, including microvascular complications such nephropathy, retinopathy, neuropathy, and cardiovascular disease.[3]
Obesity-associated insulin resistance, activation of the innate immune system, and chronic increased production of cytokines and adipokines are important links between obesity and T2DM.[4] Novel data further suggest that chronic adipose tissue inflammation and β-cell stress cause an activation of the adaptive immune system as well, which may also participate in the progression of the inflammatory response.[5]
While traditionally viewed through a metabolic lens, increasing evidence suggests that T2DM is also driven by strong inflammatory mechanisms.[6] Elevated levels of pro-inflammatory cytokines such as tumor necrosis factor-alpha (TNF-α), interleukin-6 (IL-6), and interleukin-1 (IL-1) have been implicated in the development and progression of T2DM-related comorbidities.[7] Excessive production of TNF-α contributes to increased vascular permeability, neural damage, and hypercoagulability factors that collectively promote complications such as diabetic polyneuropathy.[7] Similarly, TNF-α and IL-1 are central to the pathogenesis of diabetic retinopathy, with chronic low-grade inflammation being identified as a key driver.[7] Notably, the synergistic effects of oxidative stress and inflammatory cytokines accelerate the tissue damage that manifests in diabetic complications.[3,8,9] Oxidative stress when increased, induces hematological alteration, which affects the function, structure, and metabolism of red blood cells (RBCs), white blood cells (WBCs), and platelets.[10] In the presence of long-lasting hyperglycemia, the morphology, metabolism, and function of RBCs are inevitably subject to a series of changes that further affect hemorheology and microcirculation.[9] Under hyperglycemic conditions, autoxidation of glucose occurs, which is considered the major mechanism for free radical formation in RBCs.
Oxidative stress, marked by an imbalance between the production of reactive oxygen species (ROS) and the antioxidant defense system, plays a crucial role in T2DM pathogenesis.[3] Excessive ROS levels can directly damage cellular components. RBCs, which are in continuous contact with plasma ROS, absorb these reactive molecules over time, leading to oxidative damage of the RBC membrane.[3] This damage compromises RBC deformability, reduces microvascular perfusion, and promotes endothelial dysfunction through ROS transfer to the vascular lining.[11] Alterations in membrane protein expression, such as increased externalization of phosphatidylserine or overexpression of adhesion molecules, further impair RBC flow and contribute to pathological clot formation.[11] In both micro- and macrovascular domains, these haematological changes, mediated by chronic inflammation, are significant contributors to disease progression.[7]
In T2DM, chronic systemic inflammation is associated with structural and functional impairments of RBCs. Morphological studies have demonstrated disrupted membrane architecture, often attributed to inflammation and iron overload.[12] Additionally, they revealed that diabetic RBCs become elongated, display altered flexibility, and intertwine with spontaneously forming fibrin fibers, suggesting a compromised ultrastructure and interaction with clotting proteins.[13] At the molecular level, RBC membrane integrity is regulated by lipid bilayer asymmetry, which is maintained by specific enzymes, lippases, floppases, and scramblases, playing a key role in disrupting membrane asymmetry.[14] Pathological conditions, such as elevated ROS and calcium levels in T2DM, activate scramblase and inhibit flippase, resulting in the externalization of PS.
This exposure acts as a signal for eryptosis, ultimately leading to vesiculation, microparticle shedding, and altered rheology.[15,16,17,18,19]
Emerging evidence from alternative and integrative medicine highlights earthing (or grounding) as a novel intervention for managing inflammation and oxidative stress. Chronic inflammation and ROS-driven damage are implicated not only in diabetes but also in many degenerative diseases and aging processes.[20] Conventional treatment with anti-inflammatory agents, while effective, often carries undesirable side effects. In contrast, alternative approaches emphasize lifestyle changes, dietary modulation, and non-pharmacological interventions.[21] We use the terms free or mobile electrons to distinguish them from charged ions, which diffuse much more slowly through tissue fluids. Research conducted for more than a decade has demonstrated that earth’s charge and storehouse of electrons represent a major natural resource of health and healing.
The earth’s surface maintains a subtle negative electric charge, sustained by atmospheric and solar activity, forming a virtually unlimited reservoir of free electrons.[22] These electrons can enter the human body through direct skin contact with the earth, via walking barefoot on natural surfaces or using conductive grounding systems. This contact is hypothesized to facilitate the neutralization of positively charged free radicals (ROS), thereby mitigating oxidative stress and inflammatory responses.
Grounding has been shown to influence the electrical stability of biological systems, restore circadian rhythms, and potentially improve immune function[23]. As humans are fundamentally bioelectrical entities, with vital processes in the heart, brain, endocrine, and immune systems regulated by electric signaling—exogenous electron supplementation through grounding could have therapeutic potential. Preliminary findings suggest grounding may reduce markers of inflammation, promote improved blood flow, and enhance overall homeostasis.
2. Inflammation in Diabetes: A Driver of RBC Dysfunction
Inflammation is an essential biological process that supports tissue repair, defends against pathogens, and maintains homeostasis through tightly regulated immune responses[24]. Hence, the immune and metabolic systems are interconnected, sharing signalling pathways and molecular mediators.[24] When this interplay becomes dysregulated, the result is a sustained, maladaptive inflammatory response.[24] This persistent state, referred to as metaflammation, is a hallmark of many metabolic disorders, including T2DM.[24] Unlike acute inflammation, which is self-limiting and resolves once tissue integrity is restored[25], metaflammation is subtle but continuous, driven by metabolic stressors such as hyperglycemia[26], altered lipid metabolism[26], and excess adiposity.[26] In T2DM, this chronic inflammatory burden is responsible for most of the vascular damage, oxidative stress, and cellular dysfunction that contribute to disease progression and complications.
T2DM is increasingly becoming recognized not only as a metabolic disorder of glucose regulation, but also as a state of chronic low-grade inflammation.[27] The chronic inflammatory state in T2DM is reflected in elevated levels of systemic inflammatory markers, many of which are directly influenced by hyperglycemia, insulin resistance, and adipose tissue dysfunction (Table 1).
The imbalance between pro-oxidant processes and the body’s antioxidant defense systems results in excessive production of reactive oxygen species (ROS) such as superoxide anion (O2-), hydrogen peroxide (H2O2), and hydroxyl radical (OH). These highly reactive molecules can overwhelm normal cellular detoxification and repair mechanisms, leading to oxidative modification of proteins, lipids, and nucleic acids.[35] Such alterations impair structural integrity and disrupt critical cellular functions, including membrane fluidity, enzyme activity, and signalling pathways.[35] For example, H2O2 and O2- play an important role in the signal transduction in the pancreatic β-cells in the regulation of glucose stimulated insulin secretion.[35,36] In an oxidative stressed environment, mitochondrial and metabolic dysfunction occur due to excessive ROS from hyperglycemia or fatty acid oxidation.[35] As a result, less insulin is secreted from the β cells and prevents insulin signalling transduction needed in typical cell functioning.[35]
The body’s endogenous antioxidant defenses, including enzymatic systems such as superoxide dismutase (SOD), catalase, and glutathione peroxidase, as well as non-enzymatic antioxidants like glutathione, vitamin C, and vitamin E, work to neutralize ROS and limit their damage.[35,36] In T2DM, chronic hyperglycemia, mitochondrial dysfunction, and activation of nicotinamide adenine dinucleotide phosphate (NADPH) oxidase tip the balance toward sustained ROS generation.[35,36] This persistent oxidative burden promotes endothelial injury, impairs insulin signaling, and activates redox-sensitive transcription factors such as NF-κB[36], driving further production of pro-inflammatory cytokines.[35,36] In this way, oxidative stress not only sustains chronic low-grade inflammation but also accelerates the vascular and metabolic complications characteristic of T2DM.
Chronic hyperglycemia promotes the overproduction of ROS through mitochondrial dysfunction and the activation of NADPH oxidase.[36] Excess ROS not only damages lipids, proteins, and DNA, but also activates redox-sensitive transcription factors such as NF-κB, leading to increased expression of inflammatory mediators[36]. This creates a vicious cycle in which oxidative stress and inflammation reinforce each other, accelerating disease progression. Elevated systemic inflammatory markers, particularly high-sensitivity C-reactive protein (hs-CRP), IL-6, and TNF-α, are consistently observed in individuals with T2DM and correlate with poor glycemic control, insulin resistance, and increased cardiovascular risk.[36]
Under physiological conditions, RBCs maintain a highly specialized and flexible membrane architecture that enables them to deform and pass through narrow capillaries without damage.[37] This membrane consists of a lipid bilayer rich in phospholipids and cholesterol, supported by an underlying spectrin-actin cytoskeleton.[37,38] Embedded membrane proteins, such as band 3 and glycophorins, help preserve the cell’s biconcave shape and regulate ion transport.[37] (
). Importantly, the external surface carries a net negative charge, determined largely by sialic acid residues on glycoproteins, which contributes to electrostatic repulsion between cells and prevents aggregation.[37] Under normal metabolic conditions, RBCs are continuously exposed to low levels of ROS, primarily generated from hemoglobin autooxidation, mitochondrial remnants in reticulocytes, and enzymatic activity in surrounding tissues.[37] These ROS, including O2-, H2O2, and OH, are efficiently neutralized by the RBC’s antioxidant systems.[37] These key defenses include enzymatic pathways such as SOD, catalase, and glutathione peroxidase, as well as non-enzymatic antioxidants like glutathione (GSH).[37] Together, these mechanisms maintain redox balance, protecting membrane lipids and proteins from oxidative damage.[37]
In T2DM, chronic hyperglycemia and associated metabolic disturbances significantly affect the structural and functional properties of the RBC membrane.[39,40] Persistent high glucose levels promote non-enzymatic glycation of membrane proteins, particularly spectrin, ankyrin, and band 3, altering their conformation and impairing cytoskeletal stability.[39] Glycation also affects membrane lipids, increasing the cholesterol-to-phospholipid ratio and reducing membrane fluidity.[39] These changes compromise the RBC’s ability to deform, which is essential for efficient passage through microvessels.[39] Oxidative stress plays a central role in exacerbating these alterations. Excess ROS promotes lipid peroxidation[40] within the bilayer, generating reactive aldehydes such as malondialdehyde (MDA) and 4-hydroxynonenal (4-HNE).[39] These lipid breakdown products stiffen the membrane and disrupt the asymmetrical distribution of phospholipids, further impairing flexibility.[39,40] Concurrent oxidative modification of membrane proteins leads to crosslinking, aggregation, and altered interactions with the underlying cytoskeleton.[39]
Additionally, in T2DM a reduced membrane fluidity is consistently observed and is linked to both the degree of glycemic control and the severity of oxidative stress. [39] Decreased activity of endogenous antioxidant systems, which contributes to an imbalance favouring pro-oxidant processes.[39] The resulting damage reduces the negative surface charge (zeta potential) of RBCs due to loss of sialic acid residues, predisposing cells to aggregation and increasing whole blood viscosity.[39]
These RBC membrane alterations have significant pathophysiological consequences. Impaired deformability and increased aggregation slow microcirculatory flow, reduce oxygen delivery to tissues, and contribute to vascular complications such as retinopathy, nephropathy, and peripheral neuropathy.[39,40]
3. The Role of RBC Dysfunction in Microvascular Complications
Like platelets and fibrin networks in blood clotting, RBCs have also been found to contribute to coagulation. Emerging evidence indicates that they can enhance coagulation efficiency and promote platelet aggregation.[41] Among the spectrum of rheological properties, the deformability, aggregation and fluidity of RBCs have been associated with diabetes and diabetic complications.[42] The inherent deformability of RBCs influences blood viscosity, stemming from their unique dynamic cell membrane structure, as illustrated in Figure 1.[42] This characteristic of RBCs, namely deformability, improves blood flow in microvessels and large arteries at high shear rates. The major determinants of deformation response include cell geometry, cell shape and internal viscosity (i.e., mean cell haemoglobin concentration and components of the RBC membrane).[43] The RBC of the diabetic subject is more rigid and has decreased deformability due to the interaction of haemoglobin with the membrane. In the absence of shear stress, when RBCs are placed in suspension, they adhere face-to-face to aggregates called rouleaux.[44]
A mechanism that favours increased tendency of RBC aggregation is the decreased ionic charge of the membrane. Microscopic examination of the RBC aggregate reveals an increase in aggregate shape and size when compared to healthy controls. The para meter RBC deformability, unlike aggregation, is modified by the composition of the membrane, cytoplasmic contents age of the RBCs.[45] In diabetic patients, the content of protein of the RBC membrane, particularly the glycoproteins, is decreased, while sialidase activity is increased, thus decreasing sialic acids on RBC surfaces. This results in the superficial negative charge of the cells to decrease, and RBC aggregation is enhanced, impacting tissue perfusion and consequently blood flow and oxygen transport.[42]
Among features that are implicated in increased blood viscosity[46], in diabetics, especially those with complications[47], is not only ascribed to a decrease in RBC deformability[48], but to changes in RBC membrane fluidity as well.[49,50] The diffusion of protein and lipid molecules within the membrane is known as membrane fluidity. Thus, changes in membrane fluidity of type 2 diabetics may influence the dynamic properties of the RBC membrane, as membrane fluidity is a critical prerequisite for the functioning of biological membranes.[51] Uncontrolled blood glucose levels coupled with oxidative stress not only damage RBC but also affects membrane fluidity and cause diabetes complications since cellular metabolism and signal transduction are related to membrane fluidity.[42]
Accelerated atherosclerosis is the main underlying factor contributing to the high risk of atherothrombotic events in patients with diabetes mellitus, and athero thrombotic complications are the main cause of mortality.[52,53] Evidence from recent studies shows an association between RBC defects and increased thrombotic risk.[54] The adhesion of RBCs to vascular endothelial cells may work in concert with heightened leukocyte–endothelial cell interactions, contributing to vascular injury and increasing the risk of thrombosis in diabetes mellitus. Beyond the role of cell surface CXCL16, endothelial-RBC–RBC interactions have also been linked to endothelial CD36 and thrombospondin. In diabetes, additional factors such as the glycation of Band 3 protein and the activation of the receptor for advanced glycation end products (AGEs) are thought to further promote RBC endothelial cell adhesion, potentially exacerbating vascular inflammation and increasing endothelial permeability.[55] Hypercoagulability in diabetes mellitus is driven by an increased proportion of phosphatidylserine (PS) exposing RBCs and elevated levels of circulating RBC-derived microparticles that express PS. The externalization of PS on the RBC membrane provides a surface for the assembly of prothrombinase and tenase complexes, which subsequently enhance thrombin generation and promote clot formation. Additionally, increased interactions between RBCs and platelets may further contribute to RBC-mediated thrombogenesis, highlighting their role in the procoagulant state observed in diabetes.[55] In diabetic patients, the RBC membrane becomes rigid and non-deformable. A decrease in the cholesterol to phospholipid ratio is responsible for this abnormality. Not only is the cholesterol component of the membrane increased, but there is a four-fold increase in the phospholipid concentration, which results in a significant decrease ratio.[56]
A mechanism that favours increased tendency of RBC aggregation is the decreased ionic charge of the membrane. Microscopic examination of the RBC aggregate reveals an increase in aggregate shape and size when compared to healthy controls. The para meter RBC deformability, unlike aggregation, is modified by the composition of the membrane, cytoplasmic contents and age of the RBC.[45]
The property of aggregation is guided by the composition of the RBC membrane and plasma proteins, fibrinogen and globulin. When fibrinogen levels are increased and albumin is decreased, aggregation is enhanced. The RBCs in diabetic patients are exposed to many risk factors, such as hyperglycemia, hyperosmolarity, oxidative stress, inflammation, and lipid metabolism disorder, which consequently result in increased aggregation, reduced cell deformability, and reduced membrane fluidity. The result is catastrophic changes on the RBCs, eventually giving rise to microcirculation disorder and diabetic complications.[42]
Red blood cells play a crucial role in maintaining redox balance and actively contribute to the regulation of vascular tone, particularly under hypoxic and ischemic conditions, through the release of nitric oxide (NO) bioactivity and adenosine triphosphate (ATP). Interest in the function of RBCs has grown following recent findings that highlight significant alterations in their behaviour under various pathological conditions. These changes include enhanced adhesion properties, increased production of ROS, as well as modifications in protein composition and enzymatic activities. In addition to serving as indicators of heightened oxidative stress, dysfunctional RBCs exhibit a diminished capacity to export NO bioactivity, largely due to increased arginase activity. Notably, these functional changes in RBCs have critical implications for cardiovascular diseases. Research has demonstrated that altered RBC function contributes to endothelial dysfunction and exacerbates cardiac injury during ischemia-reperfusion in individuals with diabetes.[57]
4. Earthing: A Novel Anti-Inflammatory Approach
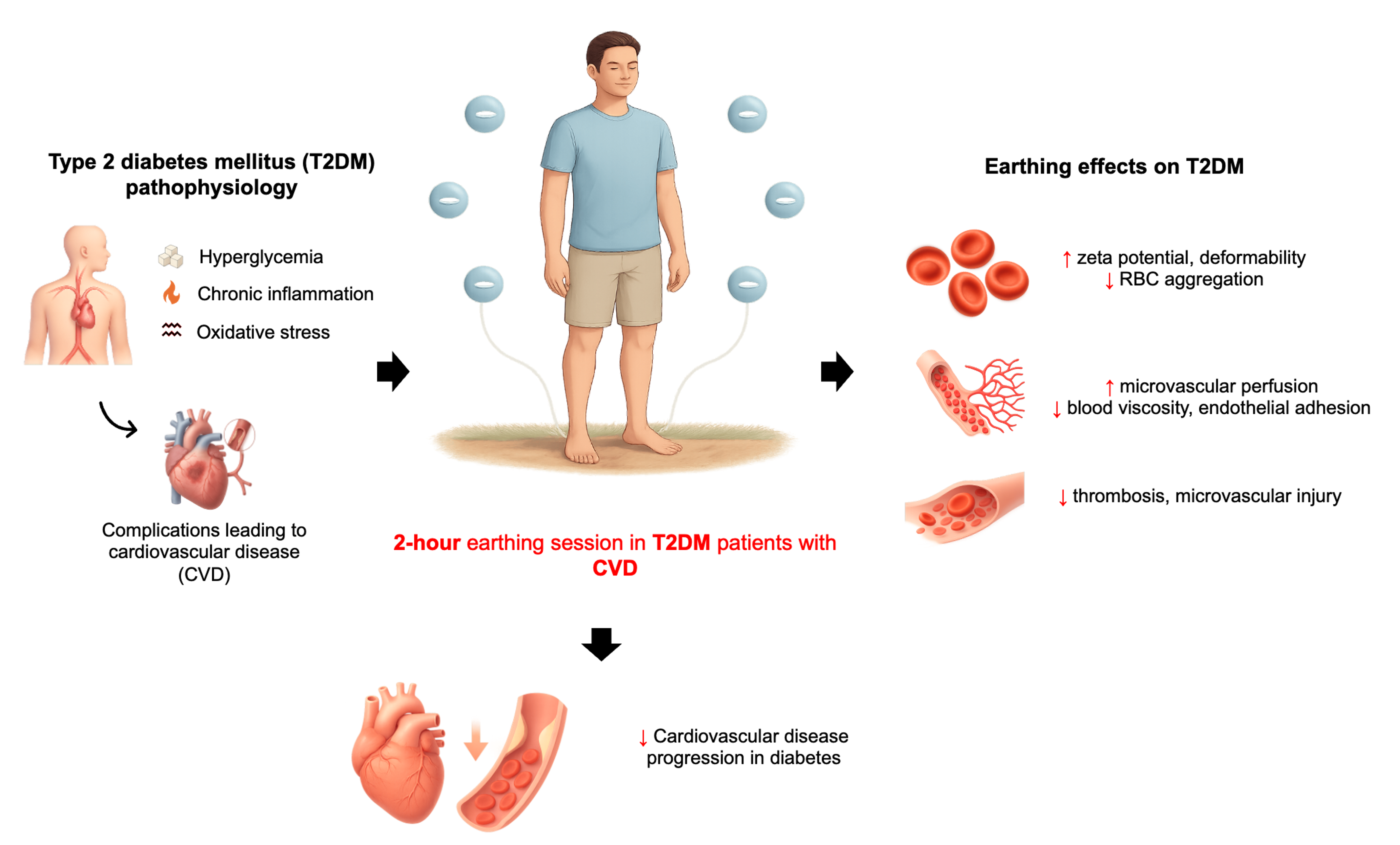
Earthing, also known as grounding refers to the direct contact of the human body to the earth’s surface.[58,59] This can include walking barefoot on grass, soil, or sand.[58]. Alternatively, this can also be simulated using a conductive system indoors that connects to the ground.[58] Earthing is based on the premise that the earth’s negative surface maintains a continuous supply of free electrons, which can enter the body and neutralize positively charged free radicals (Figure 2).[58] It is hypothesized that the neutralization of these excess free radicals or ROS can reduce the oxidative stress and inflammation via scavenging of these radicals, thereby preventing oxidative damage in surrounding tissues.[58]
Schumann resonances are natural electromagnetic waves that exist between the earth’s surface and the ionosphere.[60] The earth has a resonance frequency of approximately 7.83 Hz, cycling roughly eight times per second.[60] This frequency is maintained by a global electrical circuit powered by three main sources: the solar wind interacting with the magnetosphere, ionospheric winds, and lightning storms, mostly near the equator but occurring worldwide.[60] Lightning transfers large amounts of positive charges to the upper atmosphere and negative charges to the earth’s surface, flooding it with free electrons.[60] When these electrons enter the human body, they can produce beneficial physiological effects.[60] Published studies on earthing indicate not only anti-inflammatory effects but also a range of potential health benefits, including improved sleep[61], reduced pain[62], lower perceived stress[61], normalization of diurnal cortisol rhythms[61], enhanced glucose regulation[63], better blood fluidity[64], and modulation of immune responses.[65] (Table 2). Currently, patients who present with chronic inflammation are prescribed an anti-inflammatory medication, but these often have serious side effects for long-term usage.[60] Thus, better alternatives are needed. As such, integrative medical practitioners are rather supporting the use of non-inflammatory diets and grounding techniques instead.[60]
Grounding has shown a significant effect on blood rheology improvement, specifically the zeta potential and RBC aggregation.[66] Zeta potential indicates the negative charge across the surface of a RBC. The more negative the charge, the better the RBCs can repel each other, making the blood more coagulable.[66] In a study by Chevalier et al., 10 healthy participants were grounded for 2 hours using electrode patches.[64] Blood samples were collected before and after grounding to assess the effects on blood rheology.[64] The results showed a reduction in blood viscosity and RBC aggregation, along with an increase in zeta potential, suggesting improved blood flow characteristics.[64] This suggests that the net negative charge of the grounded electrons is able to increase the negative charge across RBCs and potentially neutralize any excess free radicals that are generated in normal cellular functioning to maintain hemodynamic stability.
The mitochondria are the primary site of aerobic energy metabolism, producing adenosine triphosphate (ATP) through oxidative phosphorylation.[67] Electrons derived from the metabolic substrates are transferred along the electron transport chain (ETC) in the inner mitochondrial membrane.[67] This flow of electrons through complexes I – IV is coupled to proton pumping, which generates the mitochondrial membrane potential (MMP).[67] This, in turn, drives ATP synthase to phosphorylate ADP to ATP, which is used in most biochemical pathways in the body.[67] A byproduct of this process is the partial reduction of oxygen to form ROS (as O2-), which is then converted to H2O2.[67] In this process, ROS is primarily produced in complex I and III, where electron carriers are overproduced or the MMP is hyperpolarized.[67] While low levels of ROS are needed as signalling molecules in some biologically pathways, excess ROS contributes to oxidative stress, which damages proteins, DNA and leads to metabolic disorders.[67] Thus, the mitochondria are a tightly regulated process that operates a specific balance of maximizing ATP production and preventing electron leakage and ROS generation.[67]
Grounding, as previously mentioned, refers to a direct electrical connection with the earth’s surface, which serves as a reservoir of free electrons.[67] From a biophysical standpoint, mitochondria operate as nano-electrical circuits, where electron flow along the ETC can be influenced by the electrical environment.[67] A study conducted by Giulivi et al., was the first to test grounding directly in isolated mitochondria using optical probes to avoid artifacts from metal electrodes.[67] They demonstrated that when mitochondria were grounded, ATP production increased by 5-11%, ROS generation decreased by 22-23% and MMP decreased by 5-6%.[67] This suggested that grounding was subtly able to modify the electrical conditions of the inner membrane thereby stabilizing electron flow and reducing electron oxidation to free radical states.[67]
In metabolic disorders such as T2DM, hyperpolarization of the MMP occurs and causes an excess of electrons in the ETC[68]. This results in reverse electron transport at complex I, causing an increase in ROS generation.[67] Under the grounding conditions of Giulivi et al., the MMP decreased by 5-6% which yielded a 30% reduction in ROS.[67] This suggests that the mechanism of grounding is heavily dependent on ROS generation on MMP.[67] This means that grounding pushes the mitochondria into a state where electron transport is efficient for ATP synthesis and less prone to the uncontrolled reduction of oxygen.[67] From a biophysical perspective, the grounding connection may allow redistribution of charge across the mitochondrial system, reducing local electron “pressure” in the ETC.[67] This would prevent over-reduction of the ubiquinone pool and limit superoxide formation at Complexes I and III.[67] The consequence is not specifically reduced oxidative stress but improved coupling efficiency. Therefore, more electrons are used productively for ATP generation rather than leaking as ROS.[67]
Because mitochondrial ROS are also signals that trigger inflammation via damage-associated molecular patterns, grounding may have systemic effects beyond bioenergetics, including modulation of redox-sensitive transcription, immune activation, and cellular stress responses.[67] The Giulivi et al., findings therefore provide a mechanistic basis for grounding as a natural regulator of oxidative stress, acting not as a direct antioxidant but as a biophysical modifier of mitochondrial electron transport.
Furthermore, emerging research has suggested that earthing can also play an important role in enhancing wound healing by exerting systemic anti-inflammatory and circulatory effects. Clinical case studies have documented significant improvements in difficult-to-heal wounds, particularly among elderly patients with compromised circulation. For example, an 84-year-old woman with an 8-month non-healing ankle wound achieved rapid pain relief and complete wound closure within two weeks of daily 30-minute earthing sessions.[59] Additionally, thermal imaging studies further demonstrated enhanced peripheral circulation and lymphatic flow following grounding, providing objective evidence of improved nutrient and oxygen delivery to injured tissue.[65] Moreover, Oschman et al.,[58] and Sokal et al.,[63] suggested that earthing acts as a “universal regulating factor” by modulating bioelectrical signalling, promoting tissue repair, and counteracting chronic inflammation. Observational reports from clinicians integrating earthing into practice describe accelerated recovery from trauma, improved skin colour and tissue vitality, reduced oedema, and rapid resolution of pain around wounds. Given that impaired wound healing is a major complication in diabetes due to chronic inflammation, vascular dysfunction, and oxidative stress, these findings suggest that earthing may hold particular promise as a supportive therapy for diabetic wound care. Collectively, these findings point to earthing as a low-cost, non-invasive, and easily accessible adjunct for wound care, with potential clinical significance in populations burdened by chronic, non-healing wounds, an area where conventional therapies often fail. While many reports remain preliminary, the consistent demonstration of faster tissue repair, reduced inflammation, and improved circulation highlights the need for larger, controlled clinical trials to fully establish Earthing as a supportive strategy in integrative wound management.
5. Clinical Implications and Future Research Directions
An array of abnormalities in diabetic RBCs has been identified and includes, amongst others: (a) altered membrane phospholipid asymmetry[69], (b) increased aggregation[70], (c) reduced deformability[71], (d) decreased membrane fluidity[72]. These changes exacerbate endothelial dysfunction and play a role in the development of diabetic complications, including retinopathy, nephropathy, and cardiovascular disease.[73,74]
The array of RBC abnormalities in diabetes mellitus is well described, comprising reduced membrane fluidity, diminished negative surface charge, and increased aggregation, all contributing to elevated blood viscosity and impaired microvascular perfusion.[75,76] These abnormalities exacerbate endothelial dysfunction and play a role in the development of diabetic complications, including retinopathy, nephropathy, and cardiovascular disease.[73,74]
Research performed using non-diabetic adults suggests that earthing (grounding) may impede some of these hemorheological derangements. Grounding has been shown to increase RBC zeta potential, reduce aggregation, and lower blood viscosity, which collectively lead to improved microcirculatory flow.[64,77] In addition, small clinical trials in patients with diabetes reported improvements in blood glucose control during night-time earthing, though no RBC parameters were evaluated.[63,78] The latter findings suggest that earthing could represent a simple, low-cost, and non-pharmacological adjunct to standard diabetes care, with potential to improve vascular health and decrease risk for complications.
Future studies are therefore needed to demonstrate the relevance of these findings in diabetic populations. Randomized controlled trials should assess the effects of earthing on RBC zeta potential, aggregation indices, deformability, and whole blood viscosity in patients with type 2 diabetes. In addition, evidence on mechanisms activated by earthing that improve glycaemic control (HbA1c), and markers of endothelial dysfunction is needed. Longitudinal prospective studies are required to determine whether regular earthing confers measurable benefits, particularly in reducing the incidence of microvascular complications and cardiovascular events. Studies that investigate how grounding impacts oxidative stress, ion movement across cell membranes, or the expression of surface proteins could help to explain why these effects occur.
6. Conclusion
Accelerated atherosclerosis is the main underlying factor contributing to the high risk of atherothrombotic events in patients with diabetes mellitus and athero thrombotic complications are the main cause of mortality [52,53]. Despite being on currently recommended dual antiplatelet therapy, diabetes still poses an increased atherothrombotic risk.[79] It is thus evident that more effective therapies are warranted for this group of high-risk patients.
Given its safety, accessibility, and low cost, earthing holds promise as a complementary intervention, particularly in resource-limited settings where the burden of diabetes and its vascular complications is substantial. However, rigorous clinical evidence within diabetic populations remains limited. Carefully designed trials integrating hemorheological, metabolic, and vascular outcomes are required to determine whether earthing can significantly remodel the pathophysiological path of diabetic vascular disease.
References
- DeFronzo, R.A.; Ferrannini, E.; Groop, L.; Henry, R.R.; Herman, W.H.; Holst, J.J.; Hu, F.B.; Kahn, C.R.; Raz, I.; Shulman, G.I. Type 2 diabetes mellitus. Nature reviews Disease primers 2015, 1, 1–22. [Google Scholar]
- Dong, C.; Wu, G.; Li, H.; Qiao, Y.; Gao, S. Type 1 and type 2 diabetes mortality burden: predictions for 2030 based on Bayesian age-period-cohort analysis of China and global mortality burden from 1990 to 2019. Journal of diabetes investigation 2024, 15, 623–633. [Google Scholar] [PubMed]
- Oguntibeju, O.O. Type 2 diabetes mellitus, oxidative stress and inflammation: examining the links. Int. J. Physiol. Pathophysiol. Pharmacol. 2019, 11, 45. [Google Scholar] [PubMed]
- Grossmann, V.; Schmitt, V.H.; Zeller, T.; Panova-Noeva, M.; Schulz, A.; Laubert-Reh, D.; Juenger, C.; Schnabel, R.B.; Abt, T.G.; Laskowski, R. Profile of the immune and inflammatory response in individuals with prediabetes and type 2 diabetes. Diabetes care 2015, 38, 1356–1364. [Google Scholar]
- Itariu, B.K.; Stulnig, T.M. Autoimmune aspects of type 2 diabetes mellitus-a mini-review. Gerontology 2014, 60, 189–196. [Google Scholar]
- Adriaanse, M.C.; Drewes, H.W.; Van Der Heide, I.; Struijs, J.N.; Baan, C.A. The impact of comorbid chronic conditions on quality of life in type 2 diabetes patients. Quality of Life Research 2016, 25, 175–182. [Google Scholar] [CrossRef]
- Navarro, J.F.; Mora, C. Role of inflammation in diabetic complications. Nephrology dialysis transplantation 2005, 20, 2601–2604. [Google Scholar]
- Martín-Timón, I.; Sevillano-Collantes, C.; Segura-Galindo, A.; del Cañizo-Gómez, F.J. Type 2 diabetes and cardiovascular disease: have all risk factors the same strength? World J. Diabetes 2014, 5, 444. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, P.; Yan, Z.; Liu, Z.; Ma, Q.; Zhang, Z.; Wang, Y.; Su, Y. The relationship between erythrocytes and diabetes mellitus. Journal of Diabetes Research 2021, 2021, 6656062. [Google Scholar] [CrossRef]
- Essawi, K.; Dobie, G.; Shaabi, M.F.; Hakami, W.; Saboor, M.; Madkhali, A.M.; Hamami, A.A.; Allallah, W.H.; Akhter, M.S.; Mobarki, A.A. Comparative analysis of red blood cells, white blood cells, platelet count, and indices in type 2 diabetes mellitus patients and normal controls: association and clinical implications. Diabetes, Metabolic Syndrome and Obesity, 3132. [Google Scholar]
- Mohanty, J.G.; Nagababu, E.; Rifkind, J.M. Red blood cell oxidative stress impairs oxygen delivery and induces red blood cell aging. Frontiers in Physiology 2014, 5. [Google Scholar] [CrossRef]
- Lipinski, B.; Pretorius, E. Novel pathway of iron-induced blood coagulation: implications for diabetes mellitus and its complications. 2012.
- Jin, H.; Xing, X.; Zhao, H.; Chen, Y.; Huang, X.; Ma, S.; Ye, H.; Cai, J. Detection of erythrocytes influenced by aging and type 2 diabetes using atomic force microscope. Biochemical and biophysical research communications 2010, 391, 1698–1702. [Google Scholar] [CrossRef] [PubMed]
- Pretorius, E.; Oberholzer, H.M.; van der Spuy, W.J.; Swanepoel, A.C.; Soma, P. Qualitative scanning electron microscopy analysis of fibrin networks and platelet abnormalities in diabetes. Blood coagulation & fibrinolysis: an international journal in haemostasis and thrombosis 2011, 22, 463–467. [Google Scholar] [CrossRef]
- Pretorius, E.; Bester, J.; Vermeulen, N.; Lipinski, B.; Gericke, G.S.; Kell, D.B. Profound morphological changes in the erythrocytes and fibrin networks of patients with hemochromatosis or with hyperferritinemia, and their normalization by iron chelators and other agents. PloS one 2014, 9, e85271. [Google Scholar] [CrossRef] [PubMed]
- Lang, E.; Qadri, S.M.; Lang, F. Killing me softly - suicidal erythrocyte death. Int J Biochem Cell Biol 2012, 44, 1236–1243. [Google Scholar] [CrossRef]
- Lang, F.; Abed, M.; Lang, E.; Föller, M. Oxidative stress and suicidal erythrocyte death. Antioxid. Redox Signal. 2013. [Google Scholar] [CrossRef]
- Qadri, S.M.; Bauer, J.; Zelenak, C.; Mahmud, H.; Kucherenko, Y.; Lee, S.H.; Ferlinz, K.; Lang, F. Sphingosine but not sphingosine-1-phosphate stimulates suicidal erythrocyte death. Cellular physiology and biochemistry: international journal of experimental cellular physiology, biochemistry, and pharmacology 2011, 28, 339–346. [Google Scholar] [CrossRef]
- Bettiol, A.; Galora, S.; Argento, F.R.; Fini, E.; Emmi, G.; Mattioli, I.; Bagni, G.; Fiorillo, C.; Becatti, M. Erythrocyte oxidative stress and thrombosis. Expert reviews in molecular medicine 2022, 24, e31. [Google Scholar] [CrossRef]
- Sinatra, S.T.; Oschman, J.L.; Chevalier, G.; Sinatra, D. Electric nutrition: The surprising health and healing benefits of biological grounding (Earthing). Altern Ther Health Med 2017, 23, 8–16. [Google Scholar]
- Williams, E.; Heckman, S. The local diurnal variation of cloud electrification and the global diurnal variation of negative charge on the Earth. Journal of Geophysical Research: Atmospheres 1993, 98, 5221–5234. [Google Scholar] [CrossRef]
- Sinatra, S.T.; Sinatra, D.S.; Sinatra, S.W.; Chevalier, G. Grounding–The universal anti-inflammatory remedy. Biomedical journal 2023, 46, 11–16. [Google Scholar] [CrossRef]
- Oschman, J.L. Charge transfer in the living matrix. J. Bodywork Movement Ther. 2009, 13, 215–228. [Google Scholar]
- van de Vyver, M. Immunology of chronic low-grade inflammation: relationship with metabolic function. Journal of Endocrinology 2023, 257. [Google Scholar] [CrossRef] [PubMed]
- Sherwood, E.R.; Toliver-Kinsky, T. Mechanisms of the inflammatory response. Best practice & research Clinical anaesthesiology 2004, 18, 385–405. [Google Scholar]
- Mukherjee, S.; Kundu, R.; Vidaković, M. Metaflammation in obesity and diabetes. 2025, 15, 1540999.
- Lontchi-Yimagou, E.; Sobngwi, E.; Matsha, T.E.; Kengne, A.P. Diabetes mellitus and inflammation. Current diabetes reports 2013, 13, 435–444. [Google Scholar]
- Pal, R.; Bhadada, S.K. AGEs accumulation with vascular complications, glycemic control and metabolic syndrome: A narrative review. Bone 2023, 176, 116884. [Google Scholar] [CrossRef]
- Luc, K.; Schramm-Luc, A.; Guzik, T.J.; Mikolajczyk, T.P. Oxidative stress and inflammatory markers in prediabetes and diabetes. J Physiol Pharmacol 2019, 70. [Google Scholar] [CrossRef]
- Elimam, H.; Abdulla, A.M.; Taha, I.M. Inflammatory markers and control of type 2 diabetes mellitus. Diabetes & Metabolic Syndrome: Clinical Research & Reviews 2019, 13, 800–804. [Google Scholar] [CrossRef] [PubMed]
- Dandona, P.; Aljada, A.; Bandyopadhyay, A. Inflammation: the link between insulin resistance, obesity and diabetes. Trends Immunol 2004, 25, 4–7. [Google Scholar] [CrossRef]
- Zand, H.; Morshedzadeh, N.; Naghashian, F. Signaling pathways linking inflammation to insulin resistance. Diabetes & Metabolic Syndrome: Clinical Research & Reviews 2017, 11, S307–S309. [Google Scholar] [CrossRef]
- Jalali, M.; Bahadoran, Z.; Mirmiran, P.; Azizi, F.; Hosseinpanah, F. Severity of adipose tissue dysfunction is associated with progression of pre-diabetes to type 2 diabetes: the Tehran Lipid and Glucose Study. BMC Public Health 2024, 24, 121. [Google Scholar] [CrossRef]
- Smith, U.; Kahn, B.B. Adipose tissue regulates insulin sensitivity: role of adipogenesis, de novo lipogenesis and novel lipids. J Intern Med 2016, 280, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Newsholme, P.; Cruzat, V.F.; Keane, K.N.; Carlessi, R.; de Bittencourt Jr, P.I.H. Molecular mechanisms of ROS production and oxidative stress in diabetes. Biochemical Journal 2016, 473, 4527–4550. [Google Scholar] [CrossRef]
- González, P.; Lozano, P.; Ros, G.; Solano, F. Hyperglycemia and Oxidative Stress: An Integral, Updated and Critical Overview of Their Metabolic Interconnections. International Journal of Molecular Sciences 2023, 24, 9352. [Google Scholar] [CrossRef]
- Orrico, F.; Laurance, S.; Lopez, A.C.; Lefevre, S.D.; Thomson, L.; Möller, M.N.; Ostuni, M.A. Oxidative Stress in Healthy and Pathological Red Blood Cells. Biomolecules 2023, 13, 1262. [Google Scholar] [CrossRef]
- Himbert, S.; Rheinstädter, M.C. Structural and mechanical properties of the red blood cell's cytoplasmic membrane seen through the lens of biophysics. Front Physiol 2022, 13, 953257. [Google Scholar] [CrossRef]
- Pitocco, D.; Hatem, D.; Riente, A.; Giulio, M.M.D.; Rizzi, A.; Abeltino, A.; Serantoni, C.; Tartaglione, L.; Rizzo, E.; Paoli, L.L. Evaluating red blood cells' membrane fluidity in diabetes: insights, mechanisms, and future aspects. Diabetes/Metabolism Research and Reviews 2025, 41, e70011. [Google Scholar] [CrossRef]
- Gabreanu, G.R.; Angelescu, S. Erythrocyte membrane in type 2 diabetes mellitus. Discoveries 2016, 4, e60. [Google Scholar] [CrossRef]
- Brown, G.E.; Ritter, L.S.; McDonagh, P.F.; Cohen, Z. Functional enhancement of platelet activation and aggregation by erythrocytes: role of red cells in thrombosis. PeerJ Preprints 2014, 2, e351v351. [Google Scholar]
- Wang, X.; Sahu, K.K.; Cerny, J. Coagulopathy, endothelial dysfunction, thrombotic microangiopathy and complement activation: potential role of complement system inhibition in COVID-19. Journal of thrombosis and thrombolysis 2021, 51, 657–662. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.; Ku, Y.; Babu, N.; Singh, M. Erythrocyte deformability and its variation in diabetes mellitus. Indian journal of experimental biology 2007, 45, 121. [Google Scholar]
- Desouky, O. Rheological and electrical behavior of erythrocytes in patients with diabetes mellitus. Rom J Biophys 2009, 19, 239–250. [Google Scholar]
- Singh, M.; Shin, S. Changes in erythrocyte aggregation and deformability in diabetes mellitus: a brief review. Indian journal of experimental biology 2009, 47, 7. [Google Scholar]
- McMillan, D.E. Plasma protein changes, blood viscosity, and diabetic microangiopathy. Diabetes 1976, 25, 858–864. [Google Scholar]
- Schut, N.; Van Arkel, E.; Hardeman, M.; Bilo, H.; Michels, R.; Vreeken, J. Blood and plasma viscosity in diabetes: possible contribution to late organ complications? Diabetes Research (Edinburgh, Scotland) 1992, 19, 31–35. [Google Scholar] [PubMed]
- McMillan, D.E.; Utterback, N.G.; Puma, J.L. Reduced erythrocyte deformability in diabetes. Diabetes 1978, 27, 895–901. [Google Scholar] [CrossRef] [PubMed]
- Bryszewska, M.; Leyko, W. Effect of insulin on human erythrocyte membrane fluidity in diabetes mellitus. Diabetologia 1983, 24, 311–313. [Google Scholar] [CrossRef] [PubMed]
- Juhan-Vague, I.; Rahmani-Jourdheuil, D.; Mishal, Z.; Roul, C.; Mourayre, Y.; Aillaud, M.; Vague, P. Correction by insulin added in vitro of abnormal membrane fluidity of the erythrocytes from type 1 (insulin-dependent) diabetic patients. Diabetologia 1986, 29, 417–420. [Google Scholar] [CrossRef]
- Emam, S.; Nikzamir, A.; Nakhjavani, M.; ESTEGHAMATI, A. Erythrocyte membrane fluidity in ageing, type 2 diabetes and stroke patients. 2008.
- Ferreiro, J.L.; Gómez-Hospital, J.A.; Angiolillo, D.J. Platelet abnormalities in diabetes mellitus. Diabetes and vascular disease research 2010, 7, 251–259. [Google Scholar] [CrossRef]
- Alzahrani, S.; Ajjan, R. Coagulation and fibrinolysis in diabetes. Diabetes and Vascular Disease Research 2010, 7, 260–273. [Google Scholar] [CrossRef]
- Byrnes, J.R.; Wolberg, A.S. Red blood cells in thrombosis. Blood, The Journal of the American Society of Hematology 2017, 130, 1795–1799. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.; Bissinger, R.; Shamaa, H.; Patel, S.; Bourne, L.; Artunc, F.; Qadri, S.M. Pathophysiology of Red Blood Cell Dysfunction in Diabetes and Its Complications. Pathophysiology 2023, 30, 327–345. [Google Scholar] [CrossRef]
- Jw, B. Role of oxidative stress in development of complication in diabetes. Diabetes 1991, 40, 405–412. [Google Scholar] [PubMed]
- Pernow, J.; Mahdi, A.; Yang, J.; Zhou, Z. Red blood cell dysfunction: a new player in cardiovascular disease. Cardiovascular research 2019, 115, 1596–1605. [Google Scholar] [CrossRef]
- Oschman, J.L.; Chevalier, G.; Brown, R. The effects of grounding (earthing) on inflammation, the immune response, wound healing, and prevention and treatment of chronic inflammatory and autoimmune diseases. Journal of inflammation research 2015, 83–96. [Google Scholar] [CrossRef]
- Menigoz, W.; Latz, T.T.; Ely, R.A.; Kamei, C.; Melvin, G.; Sinatra, D. Integrative and lifestyle medicine strategies should include Earthing (grounding): Review of research evidence and clinical observations. Explore 2020, 16, 152–160. [Google Scholar] [CrossRef]
- Sinatra, S.T.; Sinatra, D.S.; Sinatra, S.W.; Chevalier, G. Grounding – The universal anti-inflammatory remedy. Biomedical Journal 2023, 46, 11–16. [Google Scholar] [CrossRef]
- Chevalier, G.; Sinatra, S.T. Emotional stress, heart rate variability, grounding, and improved autonomic tone: clinical applications. Integrative Medicine 2011, 10, 16–21. [Google Scholar]
- Chevalier, G.; Patel, S.; Weiss, L.; Chopra, D.; Mills, P.J. The effects of grounding (earthing) on bodyworkers’ pain and overall quality of life: a randomized controlled trial. Explore 2019, 15, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Sokal, K.; Sokal, P. Earthing the human body influences physiologic processes. The Journal of Alternative and Complementary Medicine 2011, 17, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Chevalier, G.; Sinatra, S.T.; Oschman, J.L.; Delany, R.M. Earthing (grounding) the human body reduces blood viscosity—a major factor in cardiovascular disease. The journal of alternative and complementary medicine 2013, 19, 102–110. [Google Scholar] [CrossRef]
- Chevalier, G.; Melvin, G.; Barsotti, T. One-hour contact with the earth’s surface (Grounding) improves inflammation and blood flow—A randomized, double-blind, pilot study. Health 2015, 7, 1022–1059. [Google Scholar] [CrossRef]
- Chevalier, G.; Sinatra, S.T.; Oschman, J.L.; Sokal, K.; Sokal, P. Earthing: health implications of reconnecting the human body to the Earth′ s surface electrons. Journal of environmental and public health 2012, 2012, 291541. [Google Scholar] [CrossRef] [PubMed]
- Giulivi, C.; Kotz, R. Earthing effects on mitochondrial function: ATP production and ROS generation. FEBS Open Bio 2025. [Google Scholar] [CrossRef] [PubMed]
- Widlansky, M.E.; Wang, J.; Shenouda, S.M.; Hagen, T.M.; Smith, A.R.; Kizhakekuttu, T.J.; Kluge, M.A.; Weihrauch, D.; Gutterman, D.D.; Vita, J.A. Altered mitochondrial membrane potential, mass, and morphology in the mononuclear cells of humans with type 2 diabetes. Translational Research 2010, 156, 15–25. [Google Scholar] [CrossRef]
- Wali, R.K.; Jaffe, S.; Kumar, D.; Kalra, V.K. Alterations in organization of phospholipids in erythrocytes as factor in adherence to endothelial cells in diabetes mellitus. Diabetes 1988, 37, 104–111. [Google Scholar] [CrossRef]
- Babu, N. Alterations in aggregation parameters of erythrocytes due to hyper cholesterol in type-2 diabetes mellitus. The Open Circulation and Vascular Journal 2009, 2, 10–14. [Google Scholar] [CrossRef]
- Popel, A.S.; Johnson, P.C. Microcirculation and hemorheology. Annu. Rev. Fluid Mech. 2005, 37, 43–69. [Google Scholar] [CrossRef]
- Deuticke, B. Red cell membrane transport in health and disease. 2003.
- McMillan, D.E. Increased levels of acute-phase serum proteins in diabetes. Metabolism 1989, 38, 1042–1046. [Google Scholar] [CrossRef]
- Brownlee, M. The pathobiology of diabetic complications: a unifying mechanism. diabetes 2005, 54, 1615–1625. [Google Scholar] [CrossRef]
- Adak, S.; Chowdhury, S.; Bhattacharyya, M. Dynamic and electrokinetic behavior of erythrocyte membrane in diabetes mellitus and diabetic cardiovascular disease. Biochimica et Biophysica Acta (BBA)-General Subjects 2008, 1780, 108–115. [Google Scholar] [CrossRef]
- Kitagawa, S.; Nozaki, O.; Tsuda, T. Study of the relationship between electrophoretic mobility of the diabetic red blood cell and hemoglobin A1c by using a mini-cell electrophoresis apparatus. ELECTROPHORESIS: An International Journal 1999, 20, 2560–2565. [Google Scholar]
- Brown, D.; Chevalier, G.; Hill, M. Pilot study on the effect of grounding on delayed-onset muscle soreness. The Journal of Alternative and complementary Medicine 2010, 16, 265–273. [Google Scholar] [PubMed]
- Sokal, K.; Sokal, P. Earthing the human organism influences bioelectrical processes. The Journal of Alternative and Complementary Medicine 2012, 18, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Storey, R.F. What drives thrombogenesis despite antiplatelet therapy in diabetes mellitus? 2010, 7, 249–250.
Figure 1.
Proposed illustration of red blood cell membrane components in a normal (A) and glycated state (B), consisting of the phospholipid bilayer, spectrin-actin cytoskeleton responsible for RBC deformability, and embedded membrane proteins band 3 and glycophorins. Drawn by CC Nadasen using Illustrae.
Figure 1.
Proposed illustration of red blood cell membrane components in a normal (A) and glycated state (B), consisting of the phospholipid bilayer, spectrin-actin cytoskeleton responsible for RBC deformability, and embedded membrane proteins band 3 and glycophorins. Drawn by CC Nadasen using Illustrae.
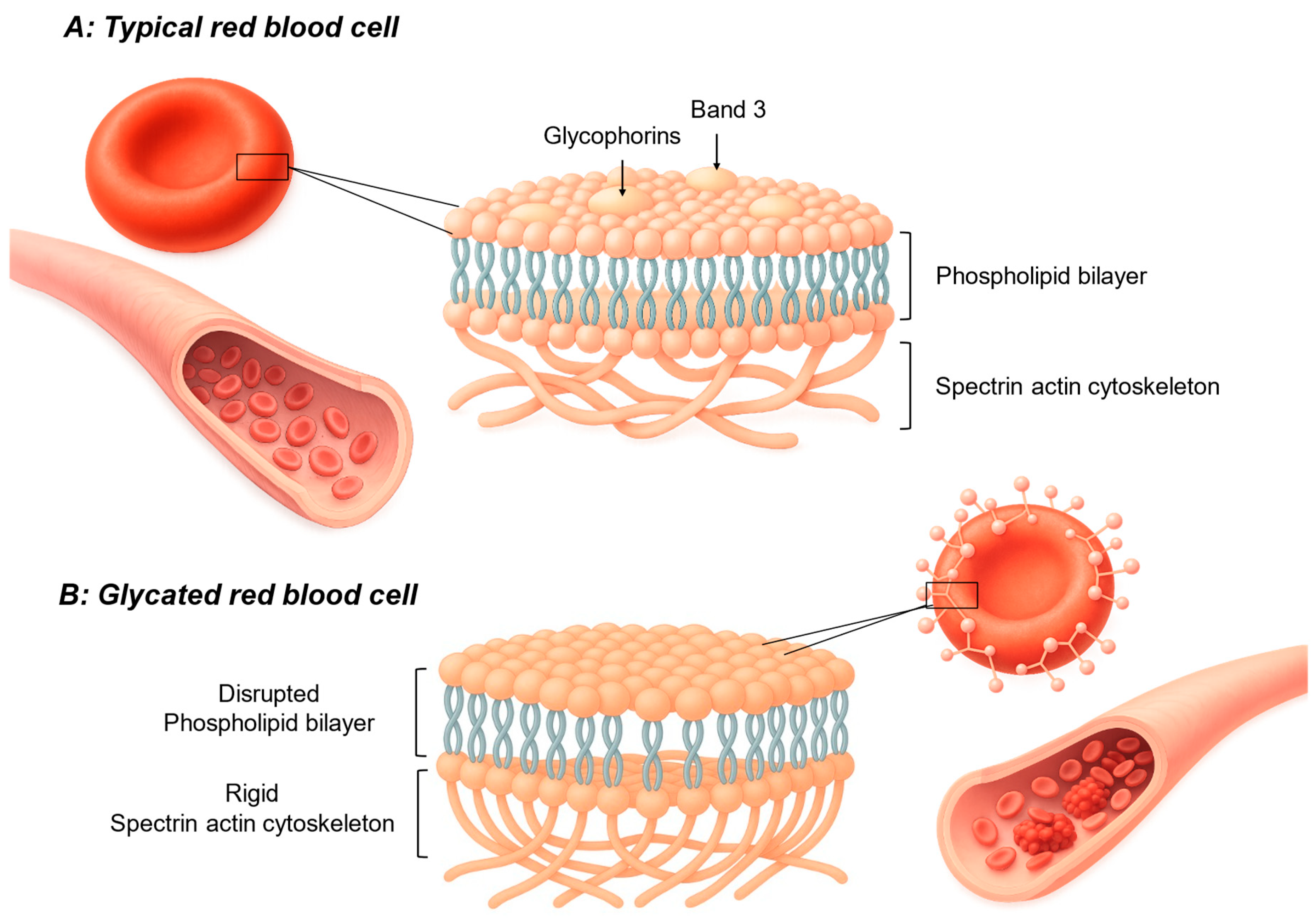
Figure 2.
Proposed illustration of grounding showing how the ground electrons can stabilize free radicals. Drawn by CC Nadasen using Illustrae.
Figure 2.
Proposed illustration of grounding showing how the ground electrons can stabilize free radicals. Drawn by CC Nadasen using Illustrae.
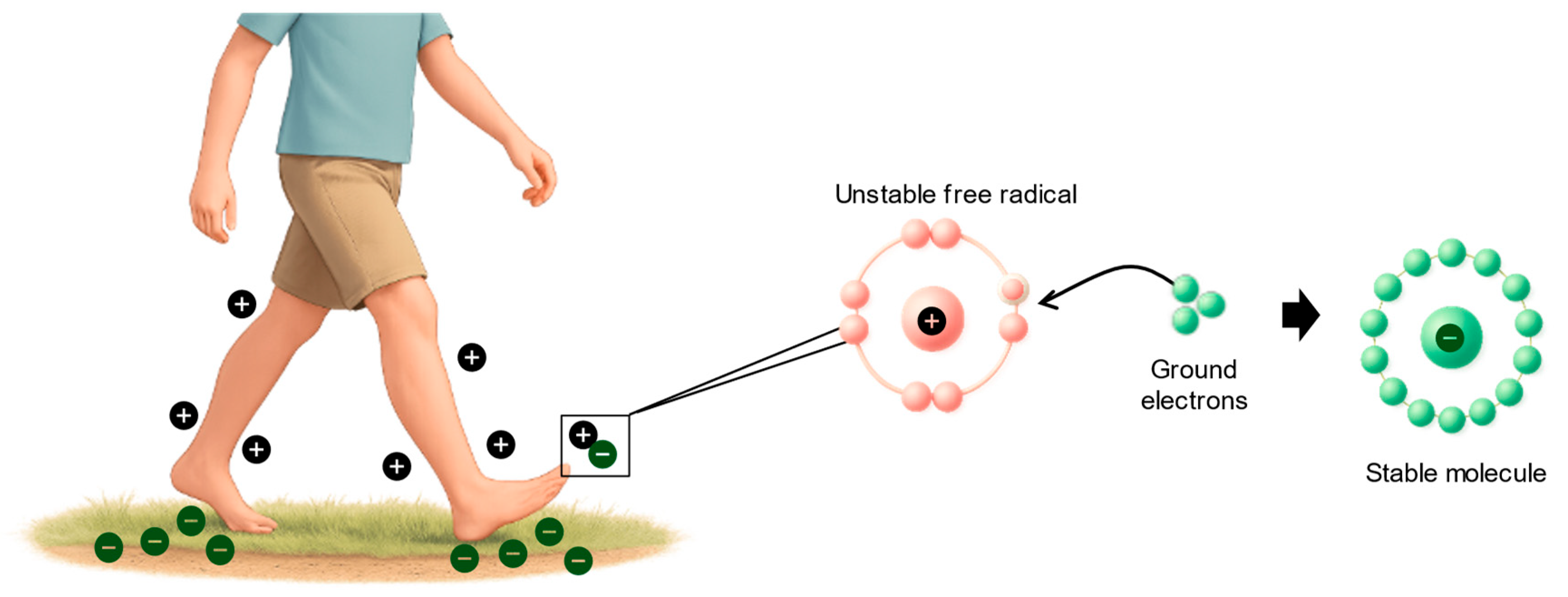

Table 1.
Inflammatory maker for pathophysiological factors of T2DM.
| Pathophysiological factor in T2DM | Mechanism driving inflammation | Typical systemic inflammatory markers |
| Hyperglycemia[28,29,30] | Formation of advanced glycation end products (AGEs) activates NF-κB signaling and stimulates cytokine release from endothelial and immune cells. | ↑ hs-CRP, ↑ IL-6, ↑ TNF-α |
| Insulin Resistance[31],[32] | Increased circulating free fatty acids activate Toll-like receptor pathways in macrophages/adipocytes, elevating cytokine production | ↑ TNF-α, ↑ IL-6, ↑ hs-CRP |
| Adipose Tissue Dysfunction[27],[33],[34] | Visceral fat immune cell infiltration shifts toward pro-inflammatory cytokine secretion and reduced adiponectin levels. | ↑ IL-6, ↑ TNF-α, ↑ hs-CRP, ↑ fibrinogen, ↑ serum amyloid A (SAA) |
Table 2.
Proposed anti-inflammatory mechanisms of grounding.
| Mechanism | Proposed biological pathway | Evidence/notes |
| Electron uptake & ROS buffering[59] | Ground contact establishes earth potential in body, allowing electron influx to neutralize ROS (O2-, H2O2, OH) at inflammatory sites, reducing oxidative tissue damage. | Hypothesis supported by thermal imaging and symptom changes; direct molecular proof in humans limited. |
| Autonomic recalibration (vagal tone)[59] | Increased parasympathetic activity activates the cholinergic anti-inflammatory pathway, suppressing cytokine production. | Observed HRV increases within minutes of grounding; small-scale human studies. |
| Normalization of cortisol rhythm[59] | Grounded sleep aligns diurnal cortisol profile, improving neuroendocrine-immune balance. | Small uncontrolled studies show corrected cortisol patterns. |
| Improved blood rheology[59] | Electron effects maintain RBC zeta potential, reduce aggregation, lower blood viscosity, and improve capillary flow. | Pilot perfusion studies and reduced viscosity observed in grounded participants. |
| Pain & thermal signature reduction[59] | Faster recovery from DOMS and cooler thermographic patterns suggest reduced inflammatory mediator activity. | Small RCTs and case series. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



