Submitted:
30 September 2025
Posted:
02 October 2025
You are already at the latest version
Abstract
Among bioactive peptides, those with antimicrobial activity have attracted increasing attention due to their potential as alternatives to traditional antibiotics. Antimicrobial peptides (AMPs) are small molecules, typically composed of 5 to 40 amino acid residues, with low cytotoxicity and minimal side effects. They act through multiple mechanisms and interact with the immune system, offering broad-spectrum activity against bacteria, viruses, fungi, and parasites. This review presents the main aspects of AMPs, including their biochemical characteristics, sources, mechanisms of action, and computational tools used for their analysis and discovery. It also examines recent progress in clinical trials and the current limitations that restrict the development and commercialization of AMPs. The review discusses the application of AMPs beyond human medicine, including their use in food preservation to prevent microbial contamination and in veterinary medicine to control infections in livestock and reduce dependence on conventional antibiotics. AMPs are versatile antimicrobial agents, but they require multidisciplinary strategies to ensure their effective implementation in the health, food, and veterinary sectors.
Keywords:
application
; bioactive peptide
; characteristic
; peptide database
1. Introduction
Bioactive peptides (BAPs) are small molecules that usually contain 2-20 amino acid residues and have molecular masses less than 6000 Da [1]. However, BAPs with longer chains, ranging up to 40 amino acids have also been reported, likely due to improvements in peptide synthesis and manufacturing technologies [2]. The biological activity of peptides depends on their amino acid characteristics and sequences, which determine their physiological actions. BAPs have been gaining increasing attention in biomedical, food, and pharmaceutical sciences because of their specific functional properties, such as antimicrobial, antioxidant, immunomodulatory, and antitumor activities [3,4]. These amino acid oligomers are present in a wide variety of organisms and play key roles in cellular communication, immune defense, and metabolic regulation [4]. Although numerous studies have reported various potential biological activities of peptides, their antimicrobial potential remains highly promising [5].
Antimicrobial peptides (AMPs) are molecules capable of combating disease-causing microorganisms such as bacteria, viruses, fungi, and parasites through different mechanisms. The first report on the identification of AMPs was published in 1939 by René Dubos, who isolated a compound (gramicidin) from a Bacillus lineage that could protect mice against pneumococcal infections [6]. Subsequently, a series of AMPs have been identified in a wide range of organisms, which employ them in defense strategies against pathogens [7]. The Antimicrobial Peptide Database 3 (APD3), which was updated in September 2025, contains data for 5,680 peptides. This includes 3,351 natural AMPs, 1,733 synthetic AMPs, and 329 predicted AMPs. Among them, 4,865 have antibacterial activity, 270 antiviral, 1,664 antifungal and 348 antiparasitic [8].
AMPs typically have 5 to 40 amino acid residues, with a positive net charge and cationic features. Unlike conventional antibiotics, AMPs kill microbial cells by disruption of membrane integrity or the ionic gradient, through interactions with the negatively charged cellular membrane. They can also inhibit protein, DNA, and RNA synthesis or interact with specific intracellular targets [9,10].
2. Broad-Spectrum Activity of Peptides
Antimicrobial peptides can act against different microorganisms, and several databases compiling various aspects related to AMPs have been established. Figure 1 shows the types of activities associated with AMPs: antibacterial, antiviral, antifungal, and antiparasitic activities.
Antimicrobial peptides share common characteristics but exhibit specific properties. Moreover, they can act through multiple mechanisms, thereby affecting various biological processes of the infectious agent and/or enhancing the host immune response [7].
2.1. Antibacterial Activity
Most AMPs have been reported to show antibacterial effects; this may be related to the large number of studies seeking approaches to combat antibiotic resistance (AMR), a major challenge to global public health that has resulted in hard-to-treat infections and a shortage of effective therapies [18,19]. Moreover, the lack of effective actions to control these microorganisms can have devastating consequences, claiming millions of lives and increasing treatment costs to over USD 100 trillion annually [20]. The organization of pathogenic microorganisms into biofilms prevents antibiotics from interacting with the bacterial cells [21]. Peptides have also been explored as potential solutions to counteract the aggravating factors in AMR [22].
2.2. Antiviral Activity
Despite the predominance of AMPs combating bacterial infections, AMPs targeting viral infections have gained importance because of the high number of deaths caused by these infections. This has led the World Health Organization (WHO) to develop a list of priority diseases to accelerate research and development. In July 2024, the list included 28 viral families, and the priority diseases, which are associated with a higher risk of epidemics or pandemics, included Ebola virus disease, Marburg virus disease, Lassa fever, Crimean-Congo hemorrhagic fever, Middle East respiratory syndrome (MERS), severe acute respiratory syndrome (SARS), other known viral diseases, and the still-unknown "Disease X.” Concerns regarding the risks associated with these diseases are reported monthly by the WHO, with records showing that they affect various regions of the world [23].
In addition to mortality rates, the emergence of new viruses, viral resistance mechanisms, delays in vaccine development, and the severe adverse effects of antiviral drugs have contributed to the complexity of this problem. In this context, natural and synthetic antiviral peptides (AVPs) have emerged as a new class of antiviral agents offering several advantages [24].
2.3. Antifungal Activity
The growing incidence of fungal infections has become a topic of concern since millions of individuals are estimated to die annually as a result of such infections. Some of the main human fungal pathogens include Candida albicans, Cryptococcus neoformans, Aspergillus fumigatus, and the emerging multidrug-resistant Candida auris. The increase in fungal infections is associated with several factors. When considering the currently available treatment options, the limited number of antifungal drug classes combined with the development of resistance across different genera has driven the search for integrated and effective strategies [25].
Another factor that favors fungal infections is the growing number of vulnerable individuals resulting from various treatments involving immunosuppressive and chemotherapeutic drugs. In this context, antifungal peptides (AFPs) possess properties that could help overcome the limitations imposed by current therapies and can be applied to the control of fungal contamination in food products [26].
2.4. Antiparasitic Activity
Among the AMPs identified in various databases, those with antiparasitic activity were the least frequently represented. Parasitism is a biological interaction that can lead to disease and, in severe cases, result in host death. Among the most relevant human parasites are protozoans (e.g., Toxoplasma gondii, Plasmodium species, and Trypanosoma cruzi), which cause infections classified by the WHO as neglected diseases and, consequently, receive little investment for the development of new treatments. For some groups, the available drugs are toxic and only partially effective, and the emergence of resistant strains has driven the search for alternative strategies [27]. In this scenario, recent advances research on antiparasitic peptides has demonstrated that these molecules can act through specific mechanisms and targets and may, therefore, represent innovative solutions for combating these pathogens [28].
3. Approaches for Obtaining AMPs
With advances in biotechnology and protein engineering, the demand for efficient, scalable, and selective peptide production methods has substantially increased. Different routes are employed to produce peptides of interest, including chemical synthesis, enzymatic hydrolysis of proteins, extraction from natural sources, and expression through recombinant technologies. Each method shows specific advantages and limitations in terms of cost, purity, yield, scalability, and the possibility of structural modifications (Table 1) [29,30].
3.1. Peptide Production via Chemical Synthesis
Chemical peptide synthesis is one of the most widely used approaches for producing specific peptide chains, enabling the precise modification of amino acid sequences with high reproducibility and structural control. This method is particularly important in the industrial and pharmaceutical contexts, where the production of peptides with well-defined characteristics (such as stability, enzymatic resistance, or optimized biological activity) is essential [3]. One of the major advantages of this approach is the possibility of incorporating non-natural amino acids or artificial post-translational modifications such as cyclization, glycosylation, and even fluorescent or radioactive labeling, which significantly broadens the applications of the peptides in pharmacokinetic and therapeutic studies [29].
The classic solid-phase peptide synthesis (SPPS) method introduced by Merrifield remains the most widely adopted method, primarily because of its compatibility with automated and scalable processes. In this process, peptide assembly begins at the carboxyl end, which is anchored to a solid resin. Protected amino acids are added individually in alternating steps of coupling and deprotection until the full peptide chain is built [31].
The choice of coupling reagents, such as O-benzotriazole-N,N,N′,N′-tetramethyl-uronium-hexafluoro-phosphate (HBTU), O-(7-azabenzotriazol-1-yl)-N,N,N,N′-tetrame thyluronium hexafluorophosphate (HATU), or N,N′- diisopropylcarbodiimide (DIC), as well as protection systems (e.g., Fmoc–9-fluorenylmethyloxycarbonyl and Boc-tert-butyloxycarbonyl) directly influences the reaction efficiency, final purity, and failure rates during synthesis [4].
In certain situations, hybrid approaches combining solid- and liquid-phase syntheses are used to produce larger peptides or peptides with more complex cyclic structures. A well-known example is cyclotide kalata B1, a cysteine-rich peptide found in plants and characterized by a cyclic cystine knot (CCK) motif. Kalata B1 shows strong antimicrobial properties and is being studied as a structural framework for the development of therapeutic peptides, including receptor agonists and enzyme inhibitors [29].
Despite its versatility, chemical synthesis faces major challenges in scaling up the production of long or structurally intricate peptides in a cost-effective manner. Additionally, traces of leftover organic solvents and toxic reagents, such as dimethylformamide (DMF) and carbodiimide-based activators, require thorough purification, which can substantially increase production costs [3].
3.2. Peptide Production via Enzymatic Pathways
The production of peptides through enzymatic hydrolysis is a widely explored alternative, especially when seeking a more sustainable process with lower environmental impact and greater cleavage selectivity. This approach is based on the controlled action of proteases on natural proteins derived from milk, algae, eggs, fish, or plants, generating BAP fragments with various functions such as antioxidants, antihypertensive, antimicrobial, and other activities [3,31].
In many cases, these peptides remain inactive while embedded in the native structure of their parent proteins and are only released through specific enzymatic cleavage. These bioactive fragments are typically hidden within full-length proteins and become active under controlled hydrolytic conditions, allowing the generation of peptides with targeted biological functions [31,32]. This process supports the idea that proteins serve as reservoirs for latent peptides with distinct physiological effects. The choice of enzymes, such as trypsin, chymotrypsin, pepsin, alcalase, flavourzyme, or microbial proteases, depends on the desired cleavage pattern and the nature of the protein source [3].
Recent technological developments have facilitated the combination of physical techniques with enzymatic hydrolysis to boost both the yield and specificity of peptides. Pretreatments such as ultrasound-assisted extraction (UAE), pulsed electric fields (PEF), and high hydrostatic pressure (HHP) can help improve enzyme access to the inner regions of the protein, increase the efficiency of hydrolysis, and enable the production of peptides with specific bioactivities [31].
For instance, enzymatic hydrolysis of marine microalgae such as Tetraselmis suecica has been shown to produce peptides with strong antimicrobial properties. One variant, AQ-1766, demonstrated significant efficacy against a wide range of pathogenic bacteria, including Escherichia coli, Staphylococcus aureus, Salmonella typhimurium, Listeria monocytogenes, and Pseudomonas aeruginosa. These peptides typically disrupt microbial membranes, highlighting their potential in food preservation and therapeutic applications [32].
Enzymatic processes offer several advantages, including high specificity, environmental compatibility, and production of peptides that are generally considered safe, often eliminating the need for further chemical modifications. Nonetheless, this approach presents challenges such as variability in raw materials, high enzyme costs, and the need for additional purification steps, such as ultrafiltration, chromatography, and lyophilization [3,32].
In this context, the use of artificial intelligence (AI) and machine learning (ML) to predict the bioactivity of enzymatically generated peptides has gained importance. These computational tools can help optimize peptide sequences, predict therapeutic potential, and identify the most effective fermentation or hydrolysis conditions [3].
3.3. Peptide Extraction from Natural Sources
Direct extraction of peptides from natural sources is one of the oldest methods still in use today, especially for AMPs and immunomodulatory compounds. A wide range of organisms, including plants, marine animals, bacteria, and fungi, naturally produce BAPs with important biological roles in pathogen defense, cell signaling, and metabolic regulation [4,29].
In plants, many of these defense-related peptides are rich in cysteine and are stabilized by disulfide bonds, which help maintain their three-dimensional structures. Well-known peptide families exhibiting potent antifungal and antibacterial activities, such as defensins, cyclotides, and thionins, can be isolated from plant tissues such as leaves, seeds, and roots using conventional extraction methods such as maceration, solvent precipitation, chromatography, and ultrafiltration. The antimicrobial activity of these molecules is generally associated with their cationic nature and their interactions with anionically charged membranes of microorganisms [29].
Antimicrobial peptides have been extensively described in marine organisms such as sponges, mollusks, echinoderms, and fish. These molecules include arenicins, piscidins, and holothuroidins, which inhibit the growth of multidrug-resistant bacteria, fungi, and viruses even in extreme environments. Extraction of these molecules involves isolation from tissues or fluids (such as mucus, hemolymph, or pseudocoelom), followed by purification using reversed-phase HPLC, size-exclusion or ion-exchange chromatography and subsequent structural analysis by mass spectrometry and NMR imaging [4].
Despite the high therapeutic potential and natural diversity of these peptides, direct extraction from natural sources faces major limitations such as low yields, seasonality, and complexity of the extracts; these factors limit the ability to obtain such compounds on an industrial scale. Therefore, many natural peptides are cloned or synthesized using recombinant or chemical methods once their sequences have been identified [32].
3.4. Peptide Production via Recombinant Technology
Recombinant DNA technology is widely used for large-scale peptide production, especially for peptides that cannot be feasibly obtained using natural extraction or traditional chemical synthesis. In this approach, genes encoding the peptides of interest are inserted into plasmid vectors and expressed in hosts, such as bacteria, yeast, filamentous fungi, or even mammalian cells [33].
Recombinant production is particularly advantageous for producing peptides with specific post-translational modifications or when utilizing cellular systems for proper folding and disulfide bridge formation, as in the case of cyclotides and defensins. For small peptides, techniques such as fusion expression with carrier proteins (e.g., glutathione S-transferase [GST] or maltose-binding protein [MBP]) are used to increase solubility and facilitate purification [29]
A relevant example is the recombinant production of bacteriocins by genetically modified Lactobacillus strains, which allows the use of generally recognized as safe (GRAS) systems for producing bioactive compounds with applications in functional or therapeutic foods. Other strategies, such as the use of strong promoters, directed mutagenesis, and codon optimization, have also been explored to increase [33].
The main challenges of this approach include peptide toxicity to the host organism, the formation of inclusion bodies, and the need for additional purification steps, typically involving chromatographic techniques. Despite these challenges, recombinant methods are considered one of the most promising methods for producing BAPs with high purity and relatively low cost, particularly for pharmaceutical and industrial applications [29,33].
4. Biochemical Characteristics of AMPs
4.1. Structure
The most common secondary structures in AMPs are α-helices and β chains. Other types of structures include spirals, linear peptides, and cyclic peptides stabilized by disulfide bridges (Figure 2) [34]. Cyclotides, which show a three-dimensional structure containing loops and disulfide bridges, exhibit favorable biological activity and show resistance to proteolytic enzymes [35]. The combination of α-helix and β chains is a critical condition for antimicrobial activity. The α-helix structure favors membrane ruptures and β chains and cyclical structures undergo internalization into the cell [36]. Other tryptophan-rich AMPs can show transmembrane effects that favor the formation of ion channels and cell rupture [37].
Specific peptides from plants show specific conformations because they are rich in specific amino acids such as thionine and cysteine, favoring conformations with antifungal and antimicrobial activities. In addition, they can induce ribosome inactivation and trypsin inhibitor activity [38].
4.2. Charge, pH, saline concentration
For antimicrobial function, the best peptides are those with a positive charge under physiological conditions and amphipathic chains containing between 5 and 100 amino acids. The positive charge interacts better with the anionic membranes of pathogens, promoting better interactions and efficiency in the destruction and lysis capacity. Charges between +14 and +16 are the most common found among AMPs [40,41].
However, excess positive charges can affect active structures and decrease the antimicrobial capacity [42]. According to APD3, 87% of active peptides are cationic, 7% are neutral, and 6% are negative, and most contain chains rich in arginine and lysine [8]. Some anionic AMPs identified in invertebrates and plants play important roles in innate immune signaling [43].
Furthermore, the pH and saline concentration can also affect the antimicrobial potential of highly active structures. High salinity is a determinant of peptide activity. Most structures are stable in the pH range of 4 to 6 because an acidic pH favors the protonation of some amino acids and the action of positive charges against the bacterial cell wall [44].
4.3. Hidrofobicity
The hydrophobicity of AMPs modulates their efficiency and specificity. Most AMPs contain approximately 50% hydrophobic residues. This favors the entry of these peptides into the lipid bilayer of pathogens [41].
There is no perfect combination of positive charge and hydrophobic chain length that can maximize activity and reduce cytotoxicity, instead the balance between positive charges and hydrophobicity modulates the interaction of the peptide with microorganisms. Higher hydrophobicity at a given charge has been shown to increase the antimicrobial effect, whereas lower hydrophobicity decreases this effect [40]. However, this relationship is not always proportional, and reports have shown that increments in hydrophobicity can reduce antimicrobial activity and toxic effects in mammals [41]. Therefore, the ratio of charge to hydrophobicity is a key factor in the Pasupuleti development of AMPs. Brogden [45] proposed that ratios ranging from 1:1 to 2:1 between the hydrophobic and basic residues may be an efficient combination. Thus, amphipathic chain characteristics and positive charges can improve the efficiency of antimicrobial activity [46]
4.4. Size
The size of a peptide with antimicrobial activity can vary from six to 60 amino acid residues [45]. This size is primarily correlated with toxicity in mammals. However, to achieve amphipathic characteristics, at least seven to eight amino acids are required [40]. APD3 shows that 90% of AMPs have less than 50 amino acids, and functional peptides have chains of 22 to 30 amino acids [8].
5. Antimicrobial Peptide Action Mechanism
The amino acid sequence, hydrophobicity, charge, and three-dimensional structure are essential for understanding the mode of action of AMPs [47]. Variations in these aspects across AMP classes ensure specific interactions with their respective targets [48]. AMPs can interact directly with the target microorganism (cell surface or intracellular components), leading to its death, or act indirectly by modulating the host immune system to combat the infection [49]. In addition, many AMPs possess the ability to act on multiple targets and, consequently, exert more than one convergent mechanism of action contributing to their antimicrobial activity.
5.1. Extracellular Target AMPs
Several factors influence the ability of AMPs to permeate and destabilize microbial membranes, such as amino acid sequence, membrane lipids, and peptide concentration. Bacterial membranes are negatively charged due to the presence of phospholipid head groups such as phosphatidylglycerol (PG), cardiolipin (CL), or phosphatidylserine (PS). Additionally, the presence of lipopolysaccharides (LPS) in Gram-negative bacteria and teichoic acids in Gram-positive bacteria contributes to this negative charge [50,51].
Membrane-active AMPs are typically amphipathic and positively charged (rich in lysine and arginine residues) at physiological pH 7.4. Due to their net positive charge and hydrophobicity, AMPs interact with negatively charged bacterial membranes, where the negative charge arises from lipid head groups and other anionic membrane components. This interaction is mediated by electrostatic forces on the bacterial surface, altering the cell’s electrochemical potential, disrupting lipid organization, and leading to leakage of intracellular contents [52,53].
In general, AMPs follow two major types of membrane action: pore formation, which includes two models: barrel-stave and toroidal pore and non-pore mechanisms, including: the carpet model and detergent-like action [49]. Barrel-stave model: AMPs insert perpendicularly into the lipid bilayer, forming oligomers that generate a transmembrane pore with a hydrophilic interior. Hydrophobic residues interact with the fatty acid chains within the membrane, stabilizing pore formation [51]. The peptide Mo-CBP3-Pep-CPAIQRCC and magainin2-GIGKFLHAS KKFGKAFVGEIMNS are examples of peptides that act via this model due to their hydrophobicity and α-helical structure [54,55]. Toroidal pore model: Peptides interact with the polar head groups of membrane lipids to form pores, causing the lipid bilayer to bend around the peptide and form a continuous curvature, resulting in stable channels that allow the passage of ions and other molecules [50,56,57].
Peptides such as YK5-NKVKEWIKYLKSLFK and MSI-78-GIGKFLKKAKKFGKA FVKILKK exemplify this mechanism by forming toroidal pores in membranes [58,59]. Carpet model: Peptides accumulate on the membrane surface at high concentrations, aligning parallel to the membrane to form a dense layer resembling a carpet. When a threshold concentration is reached, this layer disrupts and lyses the membrane without forming pores [51,60]. Cecropin P1-SWLSKTAKKLENSAKKRISEGIAIAIQGGPR acts via this model, coating the lipid bilayer and inducing membrane rupture [61]. Detergent-like model: Similar to the carpet model, but with lipid solubilization leading to micelle formation and complete bilayer disintegration. This results in dissipation of the electrochemical gradient. Although structurally predisposed to pore formation, the peptide PepD2M-Myr-WKKLKKLLKKLKKL-NH2 acts via a detergent-like mechanism, collapsing the membrane into lipid aggregates [62].
Some AMPs act via alternative or combined mechanisms. The S-thanatin-GSKKPVPIIYCNRRSGKCQRM peptide exerts multiple actions, combining membrane depolarization and interference with the bacterial respiratory chain, while maintaining low toxicity to mammalian cells due to its selectivity for lipid composition [63]. Melimine-TLISWIKNKRKQRPRVSRRRRRRGGRRRR and Mel4-KNKRKRRRRRRG GRRRR peptides, acting against Pseudomonas aeruginosa, follow a sequence of events including LPS neutralization, membrane depolarization, intracellular material release, and ultimately cell lysis [64]. The hRNase 7 peptide directly interacts with the OprI lipoprotein of P. aeruginosa, promoting increased membrane permeability and cellular content leakage [65]. Unlike the previously mentioned peptides, α-defensin HD6-CLL QGRCNCNLKLGCKSGFCGGCVNLP acts by forming nanonets on the bacterial surface without directly inducing cell death. This physical net traps bacteria and prevents adhesion and invasion, protecting the intestinal epithelium [66]. The F6-KGGDLGLFEPTL peptide fraction derived from egg yolk also demonstrates that cationic and anionic peptides can act synergistically, while cationic peptides destabilize the membrane via direct interaction, anionic peptides use metal ions to facilitate penetration [52].
Antifungal peptides have emerged as a promising alternative in combating fungal infections, especially in light of the global rise in resistance to conventional antifungals such as azoles, polyenes, and echinocandins [67]. The mechanism of action of AFPs involves the destabilization of the microbial cell membrane, a process mediated by electrostatic interactions between the cationic residues of the peptides and the anionic phospholipids of the membrane, such as phosphatidylglycerol. These peptides can create pores or disrupting membrane permeability, which compromises cellular structural integrity, therefore the result is cell death. Some examples include Mo-CBP3-Pep-CPAIQRCC and RcAlb-Pep-AKLIPTIAL, which facilitate pore formation in fungal membranes and induce oxidative stress, thereby contributing to hyphal damage [68]. The AFP, derived from Aspergillus giganteus, interacts with the fungal membrane without forming pores but alters local membrane fluidity and compromises structural integrity, ultimately resulting in fungal cell destruction. Some antifungal peptides, such as CAADIVGQCPAKLK—derived from thaumatin—act through receptor-dependent mechanisms. This peptide interacts with the PHO36 receptor, found in Saccharomyces cerevisiae and Candida albicans, triggering apoptotic pathways mediated by oxidative stress [69].
The AVPs are strongly related to their physicochemical structure, which enables specific interactions with viral components or essential cellular elements involved in the viral replication cycle. Among these interactions, those with viral membranes, cellular receptors, or viral enzymes are particularly noteworthy [70]. Inhibition of viral entry is one of the most common mechanisms, especially for enveloped viruses. Membrane active peptides display broad-spectrum antiviral properties by disrupting the stability of the viral envelope. According to Hoffmann et al. [71], these molecules interact directly with the viral lipid bilayer, hindering endosomal fusion and inducing virion aggregation, effects reported for pathogens such as influenza, dengue, and herpes simplex viruses. Their specificity is linked to the pronounced curvature of viral membranes and to the lack of repair or homeostatic mechanisms in these structures. In vitro studies, the peptide p5(coil)-GGGYSKGGKG GGKGGKGGGKGGKGGGKGGKG prevents viral attachment to host cells by binding to a heparan sulfate proteoglycan (HSPG) located on the cell surface, thereby blocking the entry of human and murine cytomegaloviruses (HCMV and MCMV) [72]. Another example is Enfuvirtide (T20)-Ac-YTSLIHSLIEESQNQQEKNEQELLELDKWASLWNWF-NH2, which interferes with the formation of the six-helix bundle (6-HB) required for membrane fusion, while peptide C60 reinforces the pre-formed 6-HB, preventing further fusion events, particularly in endocytic pathways, with demonstrated efficacy against HIV [73]. In influenza A, inhibitory activity has also been observed for peptides such as C18-ARLPR and C12-Hp-KKWK, which target hemagglutinin subunits HA1 and HA2, respectively, blocking membrane fusion. Another peptide with a similar mode of action is C20-Jp-Hp-ARLPRKKWK, which blocks the conformational rearrangement of HA2 and interferes with endosomal acidification crucial step for viral activation [74]. Peptides such as cecropin B-(NH2)-KWKVFKKIEKMGRNIRNGIVKAGPAIAVLGEAKAL-(CONH2), and CAP37-NQGRHFCGGALIHARFVMTAASCFQ have shown efficacy against non-enveloped viruses like adenovirus and rotavirus by promoting capsid disintegration or preventing viral entry through particle aggregation [57].
5.2. Intracelular target AMPs
By crossing the cell membrane without the need to induce lysis, AMPs interact with intracellular components such as DNA, RNA, enzymes, and metabolic systems, ultimately leading to cell death. Peptides such as KT2-NGVQPKYKWWK WWKKWW-NH2-, RT2-NGVQPKYRWWRWWRRWW-NH2, and PR-39-RRRPRPPYLPRPRPPPFF PPRLPPRIPPGFPPRFPPRFP-NH₂ exemplify cell entry without pore formation or membrane lysis, a process facilitated by interactions with lipopolysaccharides and exploitation of the bacterial electrochemical gradient. Once inside the cell, they bind to DNA, thereby inhibiting transcription and translation [75,76]. Buforin II-TRSSRAGLQFPVGRVHRLLRK, NDEELNKLM, and RSSKRRQ are also capable of translocating across the membrane without causing lysis and accumulate in the cytoplasm, where they strongly bind to bacterial DNA and RNA, resulting in inhibition of their functions [77,78]. Similarly, the AMP Oct-P2-MFLVVKVLKYVV interacts directly with plasmid DNA, promoting its aggregation and thereby impairing its functionality [79] .
Some AMPs directly interfere with bacterial enzymatic activity. The peptide APKHKEMPFPKYP acts against Staphylococcus aureus by inhibiting critical enzymes such as DNA gyrase and dihydrofolate reductase (DHFR) through hydrogen bonding, hydrophobic interactions, and π–π stacking. This interaction blocks DNA replication and cellular metabolism, ultimately leading to bacterial death [80]. The induction of reactive oxygen species (ROS) is a mechanism observed in several AMPs. The peptide LTX-315-KKWWKKW–Dip–K-NH₂ (Dip = β-difenílalanina) induces oxidative stress by increasing the production of nitric oxide and hypochlorite, disrupting NAD+ metabolism and ATP production, and thereby compromising bacterial viability [81]. Peptides such as CM15-KWKLFKKIGAVLKVL and MM63:CHx37 also induce oxidative stress by generating superoxide, hydroxyl radicals, and hydrogen peroxide, especially under aerobic conditions, while LL-37-LLGDFFRKSFKKLGKRLLRFFLSKSA causes oxidative stress by interfering with the cytochrome oxidase-bd complex in the periplasm [82]. The peptide [K4K15]-CZS-1-GFLKIVKGVGKVALKAVSKLF not only permeabilizes membranes but also disrupts bacterial metabolism by inhibiting essential pathways such as fatty acid biosynthesis, phospholipid metabolism, and the tricarboxylic acid (TCA) cycle, thereby hindering cellular recovery and enhancing bacterial death [83].
In addition to cell membrane action of AFPs, many antifungal peptides target essential intracellular components related to fungal metabolism. AMPs such as histatin5 (Hst5)-DSHAKRHHGYKRKFHEKHHSHRGY can interfere with genomic DNA without forming pores in the fungal membrane. As a result, these peptides cause morphogenetic defects, such as irregular chitin distribution, abnormal hyphal branching, and eventual cell lysis [84]. Metchnikowin (Mtk)-MQLNLGAIFLALLGVMATATSVLAEPHRHQG PIFDTRPSPFNPNQPRPGPIY, for example, derived from Drosophila melanogaster, acts specifically against Fusarium graminearum by inhibiting succinate-coenzyme Q reductase (SQR), an enzyme essential for mitochondrial energy production. Furthermore, Mtk interferes with fungal cell wall synthesis by inhibiting β (1,3)-glucanosyltransferase Gel1, a crucial enzyme for cell wall formation in Fusarium [85].
The peptide IJ4-Ac-KW∆FWK∆FAK∆FAKNH2 exhibits antifungal activity against various Candida species and other filamentous fungi by oxidative stress and stimulating the production of reactive oxygen species (ROS), which compromise fungal cellular components [86]. Similarly, the peptide AMT-RRWWRF acts interacting with fungal membranes and causing structural disruption, while also targeting intracellular processes. This peptide can inhibit lanosterol demethylase, an important enzyme for ergosterol biosynthesis, a major constituent of the fungal cell membrane [87].
The AVPs may also function through diverse pathways beyond extracellular binding. For example, WL-1-GWKRIKQRIKDKLRNL combats herpes simplex virus type 1 (HSV-1) by blocking viral attachment, suppressing replication, and downregulating pro-inflammatory cytokine production [57]. Direct inhibition of viral enzymes, such as influenza polymerase or essential replication proteases, has been observed for P9 -NGAICWGPCPTAFRQIGNCGHFKVRCCKIR and LVLQTM [57].
5.3. Imunomodulatory AMPs
The AMPs that do not exert their effects primarily through microbial killing can modulate the host immune system, thereby assisting in the clearance of pathogens. The human peptide LL-37-LLGDFFRKSFKKLGKRLLRFFLSKSA is a well-studied example of an immune regulator. Its activity involves binding to bacterial components such as LPS, which prevents excessive macrophage stimulation and decreases TNF-α production. Furthermore, LL-37 disrupts the interaction between LPS and LBP (binding protein), reducing TLR4 activation and limiting the development of sepsis. The LL-37 peptide promotes the recruitment of immune cells by inducing chemokines such as IL-8 and MCP-1 and increasing the expression of chemokine receptors such as IL-8RB, CXCR-4, and CCR2 [88]. The peptide’s activity is dose dependent, so at low concentrations it functions as an “immune sentinel,” while at higher levels it promotes cell migration. According to Fjell et al. [89], LL-37’s antimicrobial action is strengthened by its ability to directly penetrate bacterial membranes without the need for active endocytosis, whereas its immunomodulatory effects rely on its translocation into eukaryotic cells.
The peptides HV2-RRVHVHVDPGVHVHVRR-NH2 and PA-13-KIAKRIWKI LRRR also exhibit immunomodulatory activity. Their mechanism involves neutralizing LPS from Gram-negative bacteria, thereby inhibiting TNF-α production in RAW264.7 macrophage cells. This action is essential for protecting the host against endotoxic shock, which can result from severe Gram-negative [90]. Another immunomodulatory peptide is equine β-defensin 1 – MRTSYLLLFTLCLLLSEMASGGNFLTGLGHRSDHYNCVSSG GQCLYSACPIFTKIQGTCYRGKAKCCK- (eBD-1), which acts on macrophages infected by Staphylococcus aureus by activating the PI3K-AKT-NF-κB signaling pathway. Activation of this cascade leads to increased expression of proinflammatory cytokines such as TNF-α, IL-6, CXCL10, CD40, and RANTES. Inhibition of this pathway significantly reduces cytokine expression, underscoring its importance in the immune response. Moreover, eBD-1 stimulates the expression of phagocytosis-related proteins such as CD16 and paxillin, thereby promoting efficient pathogen clearance [91].
6. Clinical trial of AMPs
Over 5,000 antimicrobial peptides have been described, yet only 31 have reached preclinical trials (Supplementary 1), 38 are in phases I-III (Supplementary 2), and 17 have been FDA-approved and are currently on the market (Table 2). This scenario is primarily attributable to the pharmacokinetic and regulatory challenges associated with the use of AMPs in vivo, such as selective toxicity, low bioavailability, proteolytic degradation, and a lack of standardization in establishing specific legislative guidelines [92,93,94]. Nevertheless, AMPs remain promising alternatives for treating antibiotic resistance because they offer rapid antimicrobial activity, fewer adverse reactions, and broad therapeutic applications. This has led to a growing search for peptides with desirable characteristics as well as for peptide-engineering techniques and delivery systems that optimize their pharmacological profile [92,93].
Peptides exhibit low bioavailability owing to their instability at acidic pH, enzymatic action, and poor intestinal mucosal absorption, which is why most AMPs are administered via injection [94]. However, this characteristic can be advantageous depending on the type of treatment, as observed for surotomycin, an antibacterial peptide in phase III clinical trials (NCT01598311). This peptide is minimally absorbed, making it ideal for treating Clostridium difficile-associated diarrhea and achieving the required high drug concentrations in the colon. Additionally, Surotomycin is a cyclic lipopeptide, ensuring greater stability and proteolytic resistance [114,115].
An important consideration is the potential cytotoxic effect of AMPs, which is associated with their low target specificity and biochemical properties such as hydrophobicity, amphipathicity, and charge. Excessive hydrophobicity increases lytic activity, whereas cationicity enhances the selectivity toward bacterial membranes. [89,92,116]. Furthermore, studies have highlighted the importance of selecting appropriate in vivo and in vitro models, particularly for erythrocyte-based assays, since interspecies differences can affect the dose dependence of peptides [117]. Murepavadin is an example of an unsuccessful molecule that reached a phase III trial (NCT03582007) for the oral treatment of resistant Pseudomonas aeruginosa infections but exhibited dose-dependent nephrotoxicity [118,119].
To improve the success of clinical trials, certain strategies are employed, such as topical or inhalable administration, which minimize systemic exposure. Examples such as XOMA-629, Omiganan, and PL-5 are applied topically as sprays or creams and are in phase III clinical trials. Meanwhile, nanotechnology-based systems using metallic nanomaterials (e.g., gold and silver nanoparticles) or polymeric materials (e.g., chitosan and lipid-based formulations), particularly with techniques like oil-in-water nanoemulsions, liposomes, and micelles, can optimize AMP delivery, absorption, and half-life [120,121]. Nisin A, an oral AMP used in preclinical testing, demonstrated greater stability and antimicrobial activity against Staphylococcus aureus strains when formulated in chitosan/sodium alginate (CS/SA) microspheres [122].
Additionally, chemical modifications, such as substituting l-amino acids with their d-enantiomers, N-methylation, PEGylation, glycosylation, cyclization, and lipidation, can enhance AMP performance. These structural changes can improve solubility and stability in aqueous solutions, reduce renal clearance, extend antimicrobial function, and influence peptide activity through multiple mechanisms [123,124,125]. For example, oritavancin, a semisynthetic lipoglycopeptide antibiotic derived from vancomycin, is used to treat Gram-positive bacteria, including resistant strains. Its spectrum of activity and half-life are extended by its cyclic structure, lipophilic aromatic side chain, and amino acid d-enantiomers [125,126].
7. Computational Approaches for AMPs Discovery and Design
Recent studies have shown notable advancements in computational methods for the discovery and optimization of AMPs. The existing research typically follows two complementary strategies: (1) bioinformatics-based mining of natural sequences to identify novel AMPs and (2) rational design of synthetic AMPs, either by modifying known templates through structure-based approaches or by employing de novo computational methods [127]. These strategies are summarized in Figure 3.
Quantitative structure-activity relationship (QSAR) modeling is one of the longest-standing computational approaches in AMP research. This method predicts biological activity on the basis of the molecular structure, operating under the principle that structurally similar compounds exhibit similar functions [128]. Traditional QSAR employs linear regression models, in which biological activity is calculated as the weighted sum of molecular descriptors. More advanced ML-based QSAR utilizes nonlinear algorithms, including random forest, artificial neural networks, K-nearest neighbor, and support vector machines, to capture complex structure-activity relationships through hierarchical feature interactions rather than simple weighted sums [129,130]. Overall, the previously cited ML strategies and other approaches (e.g., hidden Markov models, logistic regression, fuzzy K-nearest neighbor, discriminant analysis), have substantially enhanced AMP classification accuracy within a hierarchical multilevel description [131,132].
Discriminative models such as convolutional neural networks (CNNs), deep neural networks (DNNs), and recurrent neural networks (RNNs) are often integrated into pipelines for tasks such as AMP classification, property prediction, and bioactivity categorization [132,133]. Building on these advances, Brizuela et al. [134] identified five hierarchical levels of tool functionality: (1) distinguishing AMPs from non-AMPs, (2) categorizing inputs by bioactivity, (3) target classification and prediction, (4) predicting AMP properties, and (5) estimating minimum inhibitory concentrations (MICs).
With the emergence of new methods, researchers are employing approaches combining evolutionary algorithms, codon-based optimization, and AI to design AMPs with enhanced antimicrobial activity and reduced host toxicity [127]. For de novo AMP design, deep learning-based generative models, such as variational autoencoders (VAEs), diffusion models, and generative adversarial networks (GANs), have shown notable success when trained on large datasets of known AMPs. These models generate novel peptide sequences while optimizing desired properties [135].
Computational tools such as AlphaFold [136], RoseTTAFold [137,138] and PEP-FOLD [39] can be employed to predict AMP secondary and tertiary structures. Once structural models are available, molecular docking approaches, using platforms such as AutoDock [139] HADDOCK [140] and CABS-dock [141] allow the evaluation of potential interactions with target membranes or proteins. Subsequently, molecular dynamics simulations with packages such as GROMACS [142], CHARMM36 [143] or the coarse-grained MARTINI force field [144] provide insights into AMP stability, binding mechanisms, and dynamic behavior in biological environments. Table 3 shows the common tools for AMP analysis, supporting researchers in the identification of AMPs and the generation of novel peptide sequences.
Regardless of how the models are constructed, tool performance improves proportionally with database accuracy and proper structural organization [129,170]. Supplementary Table 3 presents the widely used databases containing AMP data. These databases typically provide comprehensive annotations on AMPs, including details such as amino acid sequence (composition and length), source organisms (bacteria, fungi, plants, or animals), biological activity (e.g., antibacterial, antiviral, or antifungal effects), physicochemical properties (hydrophobicity and charge), secondary structures (α-helices, β-sheets, mixed α/β, or non-α/β folds), mechanisms of action (molecular targets), and additional data [127].
8. AMPs and their Use in the Food Industry
Antimicrobial peptides can be potentially used for food preservation, mainly because they are considered ecological, safe, and economically viable in comparison with chemical preservatives [171]. Moreover, plant- and animal-based foods can be enzymatically hydrolyzed to generate peptides with antimicrobial activity [172]. Research has highlighted the challenges in applying AMPs in food systems, such as the use of nanotechnology through nanoencapsulation, which can enhance the stability, antimicrobial spectrum, and efficacy.
Microbial fermentation can generate BAPs from proteins of plant or animal origin. These peptides exhibit antimicrobial, antioxidant, antihypertensive, and anti-inflammatory properties, and show the potential for disease prevention and health promotion. Various microorganisms, including lactic acid bacteria, Bacillus spp., and fungi (Aspergillus, Rhizopus, Saccharomyces), can contribute to the release of these compounds during the fermentation of foods such as milk, meat, cereals, legumes, and seafood [173]. Bacteriocins are AMPs derived from probiotics and produced by lactic acid bacteria in traditional fermented foods. They have emerged as sustainable alternatives to antimicrobials and a means of food biopreservation [171,174]. These bacteriocins act by forming pores and interfering with vital cellular processes and are selective, which preserve the body's beneficial microbiota [175].
The concept of "Clean Label" foods (foods produced with natural compounds) has gained prominence due to consumers' demand for healthier diets. This growing demand has pressured the food industry to replace synthetic preservatives with sustainable alternatives [176]. In this context, the use of bacteriocins and protective cultures has emerged as a promising approach for food preservation. These active cultures release antimicrobial substances directly into food, thereby increasing microbiological safety through a comprehensive effect against harmful bacteria, and are compatible with food systems [177,178]. Despite these benefits, issues related to transparency, regulation, and social acceptance persist [179].
The combined use of probiotics and antimicrobials has been investigated as a novel approach to improve food safety and extend the shelf life of products. Probiotics can modulate microbiota, block pathogens, and simultaneously enhance the efficacy of AMPs. Synergistic approaches include the local production of antimicrobials through probiotic cultures and the strengthening of host immune defenses [180]. Analyses of toxicological risks, potential negative effects on beneficial microbiota, and mechanisms of bacterial resistance are important before these compounds can be used commercially [181]. Although these compounds have great potential as natural alternatives to artificial preservatives, information on their safety on a large scale is currently lacking. Obstacles include standardizing toxicological testing, investigating the effects on gut microbiota, and monitoring bacterial resistance [182]. From this perspective, integrating ethical principles into the development and application of food biotechnology is essential. Innovation should be guided not only by technological efficiency but also by environmental sustainability, social justice, and public health protection [172].
9. AMPs and Their Use in Veterinary Medicine
The abuse and inappropriate use of antibiotics has caused AMR to become a major public health concern. One aspect of AMR has been discussed in Swann's report published in 1969, which highlights the influence of AMR in animals and the latent concern that this may negatively affect humans [183].
In livestock animals, antibiotics are prophylactically used to promote growth. This approach employs sub-therapeutic doses, which are known to select for resistant organisms [184,185]. Moreover, contrary to popular belief, domestic animals can influence this phenomenon. The indiscriminate use of antibiotics to prevent and treat diseases generates selective pressure that could lead to the transfer of AMR microorganisms to humans as a result of the close relationship between the two [186]. Wild animals may also aggravate this problem, since they could function as an ecological pool of resistant organisms however, the transmission routes involving wild animals are difficult to untangle [187]. In this scenario, the application of AMPs in the veterinary field may substantially contribute to resolving the issue at hand, facilitating the treatment of pathologies and the modulation of the immune system [188].
Antimicrobial peptides can be directly applied as growth-promoting agents. A recombinantly expressed piscidin from Epinephelus lanceolatus was used to supplement chicken feed. Animals fed 1.5%, 3%, and 6% recombinant E. lanceolatus peptide showed better growth than those in the control groups, with an increase of approximately 15%, 20%, and 16% in weight gain, respectively [189]. Another study on piglets performed with recombinant bovine lactoferrampin–lactoferricin as a dietary supplement suggested that it may improve the digestive microbiome in weaned piglets [190]. Similarly, dietary supplementation of weaning pig feed with 60 mg/kg of the antimicrobial peptide P5 (KWKKLLKKPLLKKLLKKL-NH2) improved nutrient digestibility and growth performance [191].
Antimicrobial peptides also have potential therapeutic applications. In a previous study, MPX (H-INWKGIAAMAKKLL-NH2), a peptide found in wasp venom, was found to counteract biofilm formation in vivo in a mouse model infected with Actinobacillus pleuropneumoniae, a highly infectious and relevant pathogen in the pig industry. This peptide destroyed the bacterial cell membrane and significantly reduced biofilm formation. MPX also prevented the death of mice infected with a lethal dose of A. pleuropneumoniae [192]. In a study conducted on chickens, the peptide HJH-3 (VNFKLLSHSLLVTLRSHL) showed good antibacterial activity against Salmonella pullorum, with low hemolysis and no cytotoxicity, and prevented death in chickens infected with a lethal dose of S. Pullorum [193].
Furthermore, animals can be rich sources of AMPs, since these amino acid sequences are ubiquitous in the host defense system. Horses, for example, may be an interesting source of these substances, which encompass a wide range of peptides subdivided into several classes, including lysozymes, cathelicidins, defensins, and NK lysins [194]. Recently, a study integrating deep learning and bioinformatics attempted to describe new AMPs from the ruminant gastrointestinal microbiome, and the authors identified a peptide that can inhibit methicillin-resistant Staphylococcus aureus (MRSA). Fish are also a promising source of antimicrobial substances, since they are constantly exposed to pathogens [195]. Fish gills, spleen, skin, intestines, and other organs are rich with AMPs [196,197].
Two recent reviews have focused on marine organisms and their application in aquaculture. García-Beltrán et al. [198]called attention to algae and cyanobacteria as sources of AMPs. Their study identified a wide range of activities, including antibacterial, antiparasitic, antifungal, antioxidant, and anti-inflammatory activities. Rodrigues et al. [199], on the other hand, focused on the AMPs derived from marine invertebrates. Their study identified approximately 350 peptides from 10 different invertebrate phyla, some of which showed broad-spectrum in vitro activities against pathogens, including fungi, Gram-positive bacteria, Gram-negative bacteria, and viruses. The only caveat described in their paper was the lack of in vivo studies reported in literature, highlighting a research gap that should be addressed [199].
Based on the aforementioned scenarios, the application of AMPs in veterinary medicine can be safely assumed to yield positive results. Nevertheless, more research is needed to draw definite conclusions about the efficacy and viability of the application of these compounds. However, these peptides offer concrete alternatives to antibiotics, since they are abundant in a wide range of sources, and therefore constitute a strong contender for antibiotic use.
10. Conclusions
Considering the potential advantages of using BAPs as therapeutic agents, along with their market appeal, studies that refine the mode of action of these biomolecules have gained relevance and require advanced and clinical practice. The diversity of sources and methods to obtain BAPs make them a rich and inexhaustible source of possibilities for the promotion of human and veterinary health.
The application of omics techniques, which combine the results of bioinformatics analyses, has led to the identification of several peptide sequences with biological activities, allowing the generation of these peptides through chemical synthesis or gene expression. Several servers and databases are available for researching biological activities and designing peptide amino acid sequences. These bioinformatics tools, combined with AI, can predict the biological activity of a given amino acid sequence and the possible changes and modifications that can optimize the biological activity of AMPs.
Supplementary Materials
The following supporting information can be downloaded at: X [199,200,201,202,203,204,205,206,207,208,209,210,211,212,213,214,215,216,217,218,219,220,221,222,223,224,225,226,227,228,229,230,231,232,233,234,235,236,237,238,239,240,241,242,243,244,245,246,247,248,249,250,251,252,253,254,255,256,257,258,259,260,261,262,263,264,265,266]. Table S1: AMPs in pre-clinical. All information was collected and reviewed in the DRAMP database (http://dramp.cpu-bioinfor.org/) [15] (accessed August 15, 2025). Table S2: AMPs in clinical trial. Data was collected and reviewed in the DRAMP database (http://dramp.cpu-bioinfor.org/), Chinese Clinical Trial Registry [ChiCTR] (https://www.chictr.org.cn/indexEN.html), European Union Clinical Trials Register (https://www.clinicaltrialsregister.eu/ctr-search/search), and ClinicalTrials.gov (https://www.clini caltrials.gov/) (accessed August 28, 2025). Table S3: AMPs databases. Data collected on September 5.
Author Contributions
Conceptualization, H.C. validation, H.C.; formal analysis, H.V, C.L.M.; writing—original draft preparation, C.L.M, J.V.F.L., M.S.C.R., R.A.N., A.O.F., I.C.S., T.N.C.M., N.G.R.G., H.C.; figure preparation: N.G.R.G., C.L.M., M.S.C.R., J.V.F.L. writing—review and editing, H. C., C.L.M.; supervision, H.C. All authors have read and agreed to the published version of the manuscript.”
Funding
FAPESP (2020/14426-3 and 2024/18725-6), CNPq (306023/2022-9) and CAPES (finance code 001).
Acknowledgments
We thank the Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) grants numbers 2020/14426-3 and 2024/18725-6 and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) grant number 306023/2022-9 and Fundação Coordenação de Aperfeiçoamento do Pessoal do Ensino Superior, CAPES), from Brazil Finance code 001.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| BAP | Bioactive Peptide |
| AMP | Antimicrobial Peptide |
| APD3 | Antimicrobial Peptide Database 3 |
| AMR | Antibiotic Resistance |
| WHO | World Health Organization |
| MERS | Middle East respiratory syndrome |
| SARS | Severe acute respiratory syndrome |
| AVP | Antiviral peptides |
| AFP | Antifungal peptides |
| SPPS | Solid-phase Peptide Synthesis |
| HBTU | O-benzotriazole-N,N,N′,N′-tetramethyl-uronium-hexafluoro-phosphate |
| HATU | O-(7-azabenzotriazol-1-yl)-N,N,N′,N′-tetramethyluronium hexafluorophosphate |
| DIC | N,N′-diisopropylcarbodiimide |
| CCK | Cyclic Cystine Knot |
| DMF | Dimethylformamide |
| UAE | Ultrasound-assisted extraction |
| PEF | Pulsed electric fields |
| HHP | High hydrostatic pressure |
| AI | Artificial Intelligence |
| ML | Machine Learning |
| GST | Gglutathione S-transferase |
| MBP | Maltose-binding protein |
| GRAS | Generally recognized as safe |
| PG | Phosphatidylglycerol |
| CL | Cardiolipin |
| PS | Phosphatidylserine |
| LPS | Lipopolysaccharides |
| HSPG | Heparan sulfate proteoglycan |
| HCMV | Human cytomegaloviruses |
| MCMV | Murine cytomegaloviruses |
| DHFR | Dihydrofolate reductase |
| TCA | Tricarboxylic acid |
| SQR | Succinate-coenzyme Q reductase |
| ROS | Reactive oxygen species |
| HSV-1 | Simplex virus type 1 |
| LBP | Binding protein |
| FDA | Food and Drug Administration |
| IV | Intravenous injections |
| IM | Intramuscular injections |
| SC | Subcutaneous injections |
| CS/SA | Chitosan/sodium alginate |
| QSAR | Quantitative Structure-Activity Relationship |
| CNNs | Convolutional Neural Networks |
| DNNs | Deep Neural Networks |
| RNNs | Recurrent Neural Networks |
| VAEs | Variational Autoencoders |
| GANs | Generative Adversarial Networks |
| MRSA | Methicillin-resistant Staphylococcus aureus |
| DL | Deep learning |
| SVM | Support vector machines |
| AIR | Ambiguous interaction restraints (AIRs). |
References
- Sarmadi, B.H.; Ismail, A. Antioxidative peptides from food proteins: A review. Peptides 2010, 31, 1949–1956. [Google Scholar] [CrossRef] [PubMed]
- Lau, J.L.; Dunn, M.K. Therapeutic peptides: Historical perspectives, current development trends, and future directions. Bioorganic & Medicinal Chemistry 2018, 26, 2700–2707. [Google Scholar] [CrossRef]
- Habib, H.M.; Ismail, R.; Agami, M.; El-Yazbi, A.F. Exploring the impact of bioactive peptides from fermented Milk proteins: A review with emphasis on health implications and artificial intelligence integration. Food Chemistry 2025, 481, 144047. [Google Scholar] [CrossRef]
- Kanaujia, K.A.; Wagh, S.; Pandey, G.; Phatale, V.; Khairnar, P.; Kolipaka, T.; … Kumar, S. Harnessing marine antimicrobial peptides for novel therapeutics: A deep dive into ocean-derived bioactives. International Journal of Biological Macromolecules 2025, 307, 142158. [Google Scholar] [CrossRef]
- Sánchez, A.; Vázquez, A. Bioactive peptides: A review. Food Quality and Safety 2017, 1, 29–46. [Google Scholar] [CrossRef]
- Van Epps, H.L. René Dubos: unearthing antibiotics. The Journal of Experimental Medicine 2006, 203, 259–259. [Google Scholar] [CrossRef]
- Boparai, J.K.; Sharma, P.K. Mini Review on Antimicrobial Peptides, Sources, Mechanism and Recent Applications. Protein & Peptide Letters 2019, 27, 4–16. [Google Scholar] [CrossRef]
- Wang, G.; Li, X.; Wang, Z. APD3: the antimicrobial peptide database as a tool for research and education. Nucleic Acids Research 2016, 44(D1), D1087–D1093. [Google Scholar] [CrossRef]
- Roudi, R.; Syn NL,Roudbary, M. Antimicrobial Peptides As Biologic and Immunotherapeutic Agents against Cancer: A Comprehensive Overview. Frontiers in Immunology 2017, 8. [CrossRef]
- Lei, J.; Sun, L.; Huang, S.; Zhu, C.; Li, P.; He, J.; … He, Q. The antimicrobial peptides and their potential clinical applications. American journal of translational research 2019, 11, 3919–3931. [Google Scholar] [PubMed]
- Pirtskhalava, M.; Amstrong AA, Grigolava M, Chubinidze M, Alimbarashvili E, Vishnepolsky B, … Tartakovsky, M. DBAASP v3: database of antimicrobial/cytotoxic activity and structure of peptides as a resource for development of new therapeutics. Nucleic Acids Research 2021, 49(D1), D288–D297. [CrossRef]
- Gawde, U.; Chakraborty, S.; Waghu FH, Barai RS, Khanderkar A, Indraguru R, … Idicula-Thomas, S. CAMPR4: a database of natural and synthetic antimicrobial peptides. Nucleic Acids Research 2023, 51(D1), D377–D383. [CrossRef]
- Singh, S.; Chaudhary, K.; Dhanda SK, Bhalla S, Usmani SS, Gautam A, … Raghava, G.P.S. SATPdb: a database of structurally annotated therapeutic peptides. Nucleic Acids Research 2016, 44(D1), D1119–D1126. [CrossRef]
- Quiroz, C.; Saavedra YB, Armijo-Galdames B, Amado-Hinojosa J, Olivera-Nappa Á, Sanchez-Daza A,Medina-Ortiz, D. Peptipedia: a user-friendly web application and a comprehensive database for peptide research supported by Machine Learning approach. Database 2021, 2021. [CrossRef]
- Ma, T.; Liu, Y.; Yu, B.; Sun, X.; Yao, H.; Hao, C.; … Zheng, H. DRAMP 4.0: an open-access data repository dedicated to the clinical translation of antimicrobial peptides. Nucleic acids research 2025, 53(D1), D403–D410. [Google Scholar] [CrossRef] [PubMed]
- Mondal, R.K.; Sen, D.; Arya, A.; Samanta, S.K. Developing anti-microbial peptide database version 1 to provide comprehensive and exhaustive resource of manually curated AMPs. Scientific Reports 2023, 13, 17843. [Google Scholar] [CrossRef]
- Das, D.; Jaiswal, M.; Khan FN, Ahamad S,Kumar, S. PlantPepDB: A manually curated plant peptide database. Scientific Reports 2020, 10, 2194. [CrossRef]
- O’Neill. (2016). Tackling Drug-Resistant Infections Globally: Final Report and Recommendations. Review on Antimicrobial Resistance. Retrieved from https://amr-review.org/sites/default/files/160525_Final%20paper_with%20cover.pdf.
- World Health Organization. (2024). Pathogens prioritization: a scientific framework for epidemic and pandemic research. Available at: https://www.who.int/publications/m/item/pathogens-prioritization-a-scientific-framework-for-epidemic-and-pandemic-research-preparedness. Accessed at 12/07/2025.
- Poudel, A.N.; Zhu, S.; Cooper, N.; Little, P.; Tarrant, C.; Hickman, M.; Yao, G. The economic burden of antibiotic resistance: A systematic review and meta-analysis. PLOS ONE 2023, 18, e0285170. [Google Scholar] [CrossRef]
- Al-Madboly, L.A.; Aboulmagd, A.; El-Salam, M.A.; Kushkevych, I.; El-Morsi, R.M. Microbial enzymes as powerful natural anti-biofilm candidates. Microbial Cell Factories 2024, 23, 343. [Google Scholar] [CrossRef]
- Yang, A.; Bai, Y.; Zhang, Y.; Xiao, R.; Zhang, H.; Chen, F.; Zeng, W. Detection and Treatment with Peptide Power: A New Weapon Against Bacterial Biofilms. ACS Biomaterials Science & Engineering 2025, 11, 806–819. [Google Scholar] [CrossRef]
- World Health Organization. (2025). Disease Outbreak News; Ebola virus disease in the Democratic Republic of the Congo. Available at: https://www.who.int/emergencies/disease-outbreak-news/item/2025-DON580. Accessed at 07/08/2025.
- Qureshi, A. A review on current status of antiviral peptides. Discover Viruses 2025, 2, 3. [Google Scholar] [CrossRef]
- Puumala, E.; Fallah, S.; Robbins, N.; Cowen, L.E. Advancements and challenges in antifungal therapeutic development. Clinical Microbiology Reviews 2024, 37. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Singh, G.; Bhatti JS, Gill SK,Arya, S.K. Antifungal peptides: Therapeutic potential and challenges before their commercial success. International Journal of Biological Macromolecules 2025, 284, 137957. [CrossRef]
- Rivera-Fernández, N.; Anacleto-Santos, J.; Casarrubias-Tabarez, B.; López-Pérez, T. de J., Rojas-Lemus, M., López-Valdez, N., & Fortoul, T.I. Bioactive Peptides against Human Apicomplexan Parasites. Antibiotics 2022, 11, 1658. [CrossRef]
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. Antimicrobial Peptides: Classification, Design, Application and Research Progress in Multiple Fields. Frontiers in Microbiology 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Shirsat, H.; Datt, M.; Kale, A.; Mishra, M. Plant Defense Peptides: Exploring the Structure–Function Correlation for Potential Applications in Drug Design and Therapeutics. ACS Omega 2025, 10, 7583–7596. [Google Scholar] [CrossRef] [PubMed]
- Sutcliffe, R.; Doherty, C.P. A., Morgan, H.P., Dunne, N.J., & McCarthy, H.O. Strategies for the design of biomimetic cell-penetrating peptides using AI-driven in silico tools for drug delivery. Biomaterials Advances 2025, 169, 214153. [CrossRef]
- Meng, T.; Wen, J.; Liu, H.; Guo, Y.; Tong, A.; Chu, Y.; … Zhao, C. Algal proteins and bioactive peptides: Sustainable nutrition for human health. International Journal of Biological Macromolecules 2025, 303, 140760. [Google Scholar] [CrossRef]
- García-Encinas, J.P.; Ruiz-Cruz, S.; Juárez, J.; Ornelas-Paz, J. de J., Del Toro-Sánchez, C.L., & Márquez-Ríos, E. Proteins from Microalgae: Nutritional, Functional and Bioactive Properties. Foods 2025, 14, 921. [CrossRef]
- Sharma, A.; Lee, H.-J. Antimicrobial Activity of Probiotic Bacteria Isolated from Plants: A Review. Foods 2025, 14, 495. [Google Scholar] [CrossRef]
- Lewies, A.; Wentzel, J.; Jacobs, G.; Du Plessis, L. The Potential Use of Natural and Structural Analogues of Antimicrobial Peptides in the Fight against Neglected Tropical Diseases. Molecules 2015, 20, 15392–15433. [Google Scholar] [CrossRef]
- Grover, A.; Singh, S.; Sindhu, S.; Lath, A.; Kumar, S. Advances in cyclotide research: bioactivity to cyclotide-based therapeutics. Molecular Diversity 2025. [Google Scholar] [CrossRef]
- Masuda, R.; Dazai, Y.; Mima, T.; Koide, T. Structure-activity relationships and action mechanisms of collagen-like antimicrobial peptides. Biopolymers 2017, 108. [Google Scholar] [CrossRef]
- Lundbæk, J.A.; Collingwood, S.A.; Ingólfsson, H.I.; Kapoor, R.; Andersen, O.S. Lipid bilayer regulation of membrane protein function: gramicidin channels as molecular force probes. Journal of The Royal Society Interface 2010, 7, 373–395. [Google Scholar] [CrossRef] [PubMed]
- Slezina, M.P.; Odintsova, T.I. Plant Antimicrobial Peptides: Insights into Structure-Function Relationships for Practical Applications. Current Issues in Molecular Biology 2023, 45, 3674–3704. [Google Scholar] [CrossRef]
- Shen, Y.; Maupetit, J.; Derreumaux, P.; Tufféry, P. Improved PEP-FOLD Approach for Peptide and Miniprotein Structure Prediction. Journal of Chemical Theory and Computation 2014, 10, 4745–4758. [Google Scholar] [CrossRef]
- Bahar, A.; Ren, D. Antimicrobial Peptides. Pharmaceuticals 2013, 6, 1543–1575. [Google Scholar] [CrossRef] [PubMed]
- Pasupuleti, M.; Schmidtchen, A.; Malmsten, M. Antimicrobial peptides: key components of the innate immune system. Critical Reviews in Biotechnology 2012, 32, 143–171. [Google Scholar] [CrossRef] [PubMed]
- Tossi, A.; Sandri, L.; Giangaspero, A. Amphipathic, α-helical antimicrobial peptides. Biopolymers 2000, 55, 4–30. [Google Scholar] [CrossRef]
- Malik, E.; Dennison, S.; Harris, F.; Phoenix, D. pH Dependent Antimicrobial Peptides and Proteins, Their Mechanisms of Action and Potential as Therapeutic Agents. Pharmaceuticals 2016, 9, 67. [Google Scholar] [CrossRef]
- Walkenhorst, W.F.; Klein, J.W.; Vo, P.; Wimley, W.C. pH Dependence of Microbe Sterilization by Cationic Antimicrobial Peptides. Antimicrobial Agents and Chemotherapy 2013, 57, 3312–3320. [Google Scholar] [CrossRef]
- Brogden, K.A. Antimicrobial peptides: pore formers or metabolic inhibitors in bacteria? Nature Reviews Microbiology 2005, 3, 238–250. [Google Scholar] [CrossRef]
- Mahlapuu, M.; Björn, C.; Ekblom, J. Antimicrobial peptides as therapeutic agents: opportunities and challenges. Critical Reviews in Biotechnology 2020, 40, 978–992. [Google Scholar] [CrossRef]
- Sun, C.; Gu, L.; Hussain MA, Chen L, Lin L, Wang H, … Hou, J. Characterization of the Bioactivity and Mechanism of Bactenecin Derivatives Against Food-Pathogens. Frontiers in Microbiology 2019, 10. [CrossRef]
- Zhang, Q.-Y.; Yan, Z.-B.; Meng, Y.-M.; Hong, X.-Y.; Shao, G.; Ma, J.-J.; … Fu, C.-Y. Antimicrobial peptides: mechanism of action, activity and clinical potential. Military Medical Research 2021, 8, 48. [Google Scholar] [CrossRef]
- Chen, N.; Jiang, C. Antimicrobial peptides: Structure, mechanism, and modification. European Journal of Medicinal Chemistry 2023, 255, 115377. [Google Scholar] [CrossRef]
- Giuliani, A.; Pirri, G.; Nicoletto, S. Antimicrobial peptides: an overview of a promising class of therapeutics 2007, 2, 1–33. [CrossRef]
- Shai, Y. Mode of action of membrane active antimicrobial peptides. Peptide Science 2002, 66, 236–248. [Google Scholar] [CrossRef]
- Pimchan, T.; Tian, F.; Thumanu, K.; Rodtong, S.; Yongsawatdigul, J. Isolation, identification, and mode of action of antibacterial peptides derived from egg yolk hydrolysate. Poultry Science 2023, 102, 102695. [Google Scholar] [CrossRef]
- Espeche, J.C.; Martínez, M.; Maturana, P.; Cutró; A; Semorile, L.; Maffia, P.C.; Hollmann, A. Unravelling the mechanism of action of “de novo” designed peptide P1 with model membranes and gram-positive and gram-negative bacteria. Archives of Biochemistry and Biophysics 2020, 693, 108549. [CrossRef]
- Branco, L.A. C. , Souza, P.F.N., Neto, N.A.S., Aguiar, T.K.B., Silva, A.F.B., Carneiro, R.F., … Freitas, C.D.T. New Insights into the Mechanism of Antibacterial Action of Synthetic Peptide Mo-CBP3-PepI against Klebsiella pneumoniae. Antibiotics 2022, 11, 1753. [Google Scholar] [CrossRef]
- Ludtke, S.J.; He, K.; Heller, W.T.; Harroun, T.A.; Yang, L.; Huang, H.W. Membrane Pores Induced by Magainin. Biochemistry 1996, 35, 13723–13728. [Google Scholar] [CrossRef]
- Yang, L.; Harroun TA, Weiss TM, Ding L,Huang, H.W. Barrel-Stave Model or Toroidal Model? A Case Study on Melittin Pores. Biophysical Journal 2001, 81, 1475–1485. [CrossRef]
- Neghabi Hajigha, M.; Hajikhani, B.; Vaezjalali, M.; Samadi Kafil, H.; Kazemzadeh Anari, R.; Goudarzi, M. Antiviral and antibacterial peptides: Mechanisms of action. Heliyon 2024, 10, e40121. [Google Scholar] [CrossRef]
- Kim, M.; Son, J.; Kim, Y. Structural and Mechanismic Studies of Lactophoricin Analog, Novel Antibacterial Peptide. International Journal of Molecular Sciences 2021, 22, 3734. [Google Scholar] [CrossRef]
- Hallock, K.J.; Lee, D.-K.; Ramamoorthy, A. MSI-78, an Analogue of the Magainin Antimicrobial Peptides, Disrupts Lipid Bilayer Structure via Positive Curvature Strain. Biophysical Journal 2003, 84, 3052–3060. [Google Scholar] [CrossRef]
- Bechinger, B.; Lohner, K. Detergent-like actions of linear amphipathic cationic antimicrobial peptides. Biochimica et Biophysica Acta (BBA) - Biomembranes 2006, 1758, 1529–1539. [Google Scholar] [CrossRef]
- Gazit, E.; Miller IR, Biggin PC, Sansom MSP,Shai, Y. Structure and Orientation of the Mammalian Antibacterial Peptide Cecropin P1 within Phospholipid Membranes. Journal of Molecular Biology 1996, 258, 860–870. [CrossRef]
- Chen, N.; Jiang, C. Antimicrobial peptides: Structure, mechanism, and modification. European Journal of Medicinal Chemistry 2023, 255, 115377. [Google Scholar] [CrossRef]
- Wu, G.; Wu, H.; Fan, X.; Zhao, R.; Li, X.; Wang, S.; … Xi, T. Selective toxicity of antimicrobial peptide S-thanatin on bacteria. Peptides 2010, 31, 1669–1673. [Google Scholar] [CrossRef]
- Yasir, M.; Dutta, D.; Willcox, M.D.P. Comparative mode of action of the antimicrobial peptide melimine and its derivative Mel4 against Pseudomonas aeruginosa. Scientific Reports 2019, 9, 7063. [Google Scholar] [CrossRef]
- Lin, Y.-M.; Wu, S.-J.; Chang, T.-W.; Wang, C.-F.; Suen, C.-S.; Hwang, M.-J.; … Liao, Y.-D. Outer Membrane Protein I of Pseudomonas aeruginosa Is a Target of Cationic Antimicrobial Peptide/Protein. Journal of Biological Chemistry 2010, 285, 8985–8994. [Google Scholar] [CrossRef]
- Chu, H.; Pazgier, M.; Jung, G.; Nuccio, S.-P.; Castillo PA, de Jong MF, … Bevins, C. L. Human α-Defensin 6 Promotes Mucosal Innate Immunity Through Self-Assembled Peptide Nanonets. Science 2012, 337, 477–481. [Google Scholar] [CrossRef]
- Riciluca, K.C. T. , Oliveira, U.C., Mendonça, R.Z., Bozelli Junior, J.C., Schreier, S., & da Silva Junior, P.I. Rondonin: antimicrobial properties and mechanism of action. FEBS Open Bio 2021, 11, 2541–2559. [Google Scholar] [CrossRef]
- Lima, P.G.; Souza, P.F.N.; Freitas, C.D.T.; Bezerra, L.P.; Neto, N.A.S.; Silva, A.F.B.; … Sousa, D.O.B. Synthetic peptides against Trichophyton mentagrophytes and T. rubrum: Mechanisms of action and efficiency compared to griseofulvin and itraconazole. Life Sciences 2021, 265, 118803. [Google Scholar] [CrossRef]
- Lopes, F.E. S. , da Costa, H.P.S., Souza, P.F.N., Oliveira, J.P.B., Ramos, M.V., Freire, J.E.C., … Freitas, C.D.T. Peptide from thaumatin plant protein exhibits selective anticandidal activity by inducing apoptosis via membrane receptor. Phytochemistry 2019, 159, 46–55. [Google Scholar] [CrossRef]
- Skalickova, S.; Heger, Z.; Krejcova, L.; Pekarik, V.; Bastl, K.; Janda, J.; … Kizek, R. Perspective of Use of Antiviral Peptides against Influenza Virus. Viruses 2015, 7, 5428–5442. [Google Scholar] [CrossRef]
- Hoffmann, A.R.; Guha, S.; Wu, E.; Ghimire, J.; Wang, Y.; He, J.; … Wimley, W.C. Broad-Spectrum Antiviral Entry Inhibition by Interfacially Active Peptides. Journal of Virology 2020, 94. [Google Scholar] [CrossRef]
- Jackson, J.W.; Hancock, T.J.; Dogra, P.; Patel, R.; Arav-Boger, R.; Williams, A.D.; … Sparer, T.E. Anticytomegalovirus Peptides Point to New Insights for CMV Entry Mechanisms and the Limitations of In Vitro Screenings. mSphere 2019, 4. [Google Scholar] [CrossRef]
- Chao, L.; Lu, L.; Yang, H.; Zhu, Y.; Li, Y.; Wang, Q.; … Chen, Y.-H. Identification of a Human Protein-Derived HIV-1 Fusion Inhibitor Targeting the gp41 Fusion Core Structure. PLoS ONE 2013, 8, e66156. [Google Scholar] [CrossRef]
- Lin, D.; Li, F.; Wu, Q.; Xie, X.; Wu, W.; Wu, J.; … He, J. A “building block” approach to the new influenza A virus entry inhibitors with reduced cellular toxicities. Scientific Reports 2016, 6, 22790. [Google Scholar] [CrossRef]
- Anunthawan, T.; de la Fuente-Núñez, C.; Hancock, R.E. W. , & Klaynongsruang, S. Cationic amphipathic peptides KT2 and RT2 are taken up into bacterial cells and kill planktonic and biofilm bacteria. Biochimica et Biophysica Acta (BBA) - Biomembranes 2015, 1848, 1352–1358. [Google Scholar] [CrossRef]
- Sneideris, T.; Erkamp NA, Ausserwöger H, Saar KL, Welsh TJ, Qian D, … Knowles, T. P.J. Targeting nucleic acid phase transitions as a mechanism of action for antimicrobial peptides. Nature Communications 2023, 14, 7170. [Google Scholar] [CrossRef]
- Park, C.B.; Kim, H.S.; Kim, S.C. Mechanism of Action of the Antimicrobial Peptide Buforin II: Buforin II Kills Microorganisms by Penetrating the Cell Membrane and Inhibiting Cellular Functions. Biochemical and Biophysical Research Communications 1998, 244, 253–257. [Google Scholar] [CrossRef]
- Li, L.; Li, J.; Yu, X.; Cao, R.; Hong, M.; Xu, Z.; … Zhu, H. Antimicrobial peptides fight against Pseudomonas aeruginosa at a sub-inhibitory concentration via anti-QS pathway. Bioorganic Chemistry 2023, 141, 106922. [Google Scholar] [CrossRef]
- Chen, P.; Zhang, T.; Li, C.; Praveen, P.; Parisi, K.; Beh, C.; … Shang, C. Aggregation-prone antimicrobial peptides target gram-negative bacterial nucleic acids and protein synthesis. Acta Biomaterialia 2025, 192, 446–460. [Google Scholar] [CrossRef]
- Zhang, X.; Yang, J.; Suo, H.; Tan, J.; Zhang, Y.; Song, J. Identification and molecular mechanism of action of antibacterial peptides from Flavourzyme-hydrolyzed yak casein against Staphylococcus aureus. Journal of Dairy Science 2023, 106, 3779–3790. [Google Scholar] [CrossRef]
- Tang, Y.; Yang, C.; Zhao, J.; Heng, H.; Peng, M.; Sun, L.; … Chen, S. LTX-315 is a novel broad-spectrum antimicrobial peptide against clinical multidrug-resistant bacteria. Journal of Advanced Research 2025. [Google Scholar] [CrossRef]
- Choi, H.; Yang, Z.; Weisshaar, J.C. Oxidative stress induced in E. coli by the human antimicrobial peptide LL-37. PLOS Pathogens 2017, 13, e1006481. [Google Scholar] [CrossRef]
- Bermúdez-Puga, S.; Dias, M.; Freire de Oliveira, T.; Mendonça, C.M. N., Yokomizo de Almeida, S.R., Rozas, E.E., … Oliveira, R.P. de S. Dual antibacterial mechanism of [K4K15]CZS-1 against Salmonella Typhimurium: a membrane active and intracellular-targeting antimicrobial peptide. Frontiers in Microbiology 2023, 14. [CrossRef]
- Chadha, S. Combating fungal phytopathogens with human salivary antimicrobial peptide histatin 5 through a multi-target mechanism. World Journal of Microbiology and Biotechnology 2023, 39, 215. [Google Scholar] [CrossRef]
- Moghaddam, M.-R. B. , Gross, T., Becker, A., Vilcinskas, A., & Rahnamaeian, M. The selective antifungal activity of Drosophila melanogaster metchnikowin reflects the species-dependent inhibition of succinate–coenzyme Q reductase. Scientific Reports 2017, 7, 8192. [Google Scholar] [CrossRef]
- Maurya, I.K.; Thota, C.K.; Sharma, J.; Tupe, S.G.; Chaudhary, P.; Singh, M.K.; … Chauhan, V.S. Mechanism of action of novel synthetic dodecapeptides against Candida albicans. Biochimica et Biophysica Acta (BBA) - General Subjects 2013, 1830, 5193–5203. [Google Scholar] [CrossRef]
- Di Marino, S.; Scrima, M.; Grimaldi, M.; D’Errico, G.; Vitiello, G.; Sanguinetti, M.; … D’Ursi, A.M. Antifungal peptides at membrane interaction. European Journal of Medicinal Chemistry 2012, 51, 154–162. [Google Scholar] [CrossRef]
- Scott, M.G.; Davidson, D.J.; Gold, M.R.; Bowdish, D.; Hancock, R.E.W. The Human Antimicrobial Peptide LL-37 Is a Multifunctional Modulator of Innate Immune Responses. The Journal of Immunology 2002, 169, 3883–3891. [Google Scholar] [CrossRef]
- Fjell, C.D.; Hiss, J.A.; Hancock, R.E.W.; Schneider, G. Designing antimicrobial peptides: form follows function. Nature Reviews Drug Discovery 2012, 11, 37–51. [Google Scholar] [CrossRef]
- Dong, N.; Wang, C.; Zhang, T.; Zhang, L.; Xue, C.; Feng, X.; … Shan, A. Bioactivity and Bactericidal Mechanism of Histidine-Rich β-Hairpin Peptide Against Gram-Negative Bacteria. International Journal of Molecular Sciences 2019, 20, 3954. [Google Scholar] [CrossRef]
- Pei, L.; Hou, Y.; Feng, Y.; Li, F.; Su, H.; Zhang, Y.; … Cao, G. Equine β-defensin 1 regulates cytokine expression and phagocytosis in S. aureus-infected mouse monocyte macrophages via the Paxillin-FAK-PI3K pathway. International Immunopharmacology 2023, 123, 110793. [Google Scholar] [CrossRef]
- de Oliveira, K.B.S.; Leite, M.L.; Melo, N.T.M.; Lima, L.F.; Barbosa, T.C.Q.; Carmo, N.L.; … Franco, O.L. Antimicrobial Peptide Delivery Systems as Promising Tools Against Resistant Bacterial Infections. Antibiotics 2024, 13, 1042. [Google Scholar] [CrossRef]
- Mahlapuu, M.; Björn, C.; Ekblom, J. Antimicrobial peptides as therapeutic agents: opportunities and challenges. Critical Reviews in Biotechnology 2020, 40, 978–992. [Google Scholar] [CrossRef]
- Bruno, B.J.; Miller, G.D.; Lim, C.S. Basics and Recent Advances in Peptide and Protein Drug Delivery. Therapeutic Delivery 2013, 4, 1443–1467. [Google Scholar] [CrossRef]
- Katsila, T.; Siskos AP,Tamvakopoulos, C. Peptide and protein drugs: The study of their metabolism and catabolism by mass spectrometry. Mass Spectrometry Reviews 2012, 31, 110–133. [Google Scholar] [CrossRef]
- Leuthner, K.D.; Yuen, A.; Mao, Y.; Rahbar, A. Dalbavancin (BI-387) for the treatment of complicated skin and skin structure infection. Expert Review of Anti-infective Therapy 2015, 13, 149–159. [Google Scholar] [CrossRef]
- Cortes-Penfield, N.; Oliver NT, Hunter A,Rodriguez-Barradas, M. Daptomycin and combination daptomycin-ceftaroline as salvage therapy for persistent methicillin-resistant Staphylococcus aureus bacteremia. Infectious Diseases 2018, 50, 643–647. [Google Scholar] [CrossRef]
- Kapić, E.; Becić, F.; Zvizdić, S. Enfuvirtide, mechanism of action and pharmacological properties. Medicinski arhiv 2005, 59, 313–6. [Google Scholar]
- Pavithrra, G.; Rajasekaran, R. Identification of Effective Dimeric Gramicidin-D Peptide as Antimicrobial Therapeutics over Drug Resistance: In-Silico Approach. Interdisciplinary Sciences: Computational Life Sciences 2019, 11, 575–583. [Google Scholar] [CrossRef]
- Vater, J.; Stein, T.H. (1999). Structure, Function, and Biosynthesis of Gramicidin S Synthetase. In Comprehensive Natural Products Chemistry (pp. 319–352). Elsevier. [CrossRef]
- Greig, S.L. Obiltoxaximab: First Global Approval. Drugs 2016, 76, 823–830. [Google Scholar] [CrossRef]
- Kaasch, A.J.; Seifert, H. Oritavancin: A Long-Acting Antibacterial Lipoglycopeptide. Future Microbiology 2016, 11, 843–855. [Google Scholar] [CrossRef]
- Mazur, N.I.; Löwensteyn, Y.N.; Terstappen, J.; Leusen, J.; Schobben, F.; Cianci, D.; … Schuurman, R. Daily intranasal palivizumab to prevent respiratory syncytial virus infection in healthy preterm infants: a phase 1/2b randomized placebo-controlled trial. eClinicalMedicine 2023, 66, 102324. [Google Scholar] [CrossRef]
- Avedissian, S.N.; Liu, J.; Rhodes, N.J.; Lee, A.; Pais, G.M.; Hauser, A.R.; Scheetz, M.H. A Review of the Clinical Pharmacokinetics of Polymyxin B. Antibiotics 2019, 8, 31. [Google Scholar] [CrossRef]
- Soon, R.L.; Nation, R.L.; Cockram, S.; Moffatt, J.H.; Harper, M.; Adler, B.; … Li, J. Different surface charge of colistin-susceptible and -resistant Acinetobacter baumannii cells measured with zeta potential as a function of growth phase and colistin treatment. Journal of Antimicrobial Chemotherapy 2011, 66, 126–133. [Google Scholar] [CrossRef]
- Mazumdar, S. Raxibacumab. mAbs 2009, 1, 531–538. [Google Scholar] [CrossRef]
- Zhanel, G.G.; Calic, D.; Schweizer, F.; Zelenitsky, S.; Adam, H.; Lagacé-Wiens, P.R.S.; … Karlowsky, J.A. New Lipoglycopeptides. Drugs 2010, 70, 859–886. [Google Scholar] [CrossRef]
- Romani, L.; Bistoni, F.; Gaziano, R.; Bozza, S.; Montagnoli, C.; Perruccio, K.; … Garaci, E. Thymosin α 1 activates dendritic cells for antifungal Th1 resistance through Toll-like receptor signaling. Blood 2004, 103, 4232–4239. [Google Scholar] [CrossRef]
- Vosloo, J.A.; Rautenbach, M. Following tyrothricin peptide production by Brevibacillus parabrevis with electrospray mass spectrometry. Biochimie 2020, 179, 101–112. [Google Scholar] [CrossRef]
- Gramenzi, A.; Cursaro, C.; Andreone, P.; Bernardi, M. Thymalfasin. BioDrugs 1998, 9, 477–486. [Google Scholar] [CrossRef]
- Rybak, M.J. The Pharmacokinetic and Pharmacodynamic Properties of Vancomycin. Clinical Infectious Diseases 2006, 42(Supplement_1), S35–S39. [CrossRef]
- Knox, C.; Wilson, M.; Klinger CM, Franklin M, Oler E, Wilson A, … Wishart, D.S. DrugBank 6.0: the DrugBank Knowledgebase for 2024. Nucleic Acids Research 2024, 52(D1), D1265–D1275. [CrossRef]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; … Bolton, E.E. PubChem 2025 update. Nucleic Acids Research 2025, 53(D1), D1516–D1525. [Google Scholar] [CrossRef]
- Ooijevaar, R.E.; van Beurden, Y.H.; Terveer, E.M.; Goorhuis, A.; Bauer, M.P.; Keller, J.J.; … Kuijper, E.J. Update of treatment algorithms for Clostridium difficile infection. Clinical Microbiology and Infection 2018, 24, 452–462. [Google Scholar] [CrossRef]
- Chandorkar, G.; Zhan, Q.; Donovan, J.; Rege, S.; Patino, H. Pharmacokinetics of surotomycin from phase 1 single and multiple ascending dose studies in healthy volunteers. BMC Pharmacology and Toxicology 2017, 18, 24. [Google Scholar] [CrossRef]
- Lei, J.; Sun, L.; Huang, S.; Zhu, C.; Li, P.; He, J.; … He, Q. The antimicrobial peptides and their potential clinical applications. American journal of translational research 2019, 11, 3919–3931. [Google Scholar]
- Greco, I.; Molchanova, N.; Holmedal, E.; Jenssen, H.; Hummel BD, Watts JL, … Svenson, J. Correlation between hemolytic activity, cytotoxicity and systemic in vivo toxicity of synthetic antimicrobial peptides. Scientific Reports 2020, 10, 13206. [Google Scholar] [CrossRef]
- Prasad, N.K.; Seiple, I.B.; Cirz, R.T.; Rosenberg, O.S. Leaks in the Pipeline: a Failure Analysis of Gram-Negative Antibiotic Development from 2010 to 2020. Antimicrobial Agents and Chemotherapy 2022, 66. [Google Scholar] [CrossRef]
- Dale, G.E.; Halabi, A.; Petersen-Sylla, M.; Wach, A.; Zwingelstein, C. Pharmacokinetics, Tolerability, and Safety of Murepavadin, a Novel Antipseudomonal Antibiotic, in Subjects with Mild, Moderate, or Severe Renal Function Impairment. Antimicrobial Agents and Chemotherapy 2018, 62. [Google Scholar] [CrossRef]
- Fadaka, A.O.; Sibuyi, N.R.S.; Madiehe, A.M.; Meyer, M. Nanotechnology-Based Delivery Systems for Antimicrobial Peptides. Pharmaceutics 2021, 13, 1795. [Google Scholar] [CrossRef]
- Haddadzadegan, S.; Dorkoosh, F.; Bernkop-Schnürch, A. Oral delivery of therapeutic peptides and proteins: Technology landscape of lipid-based nanocarriers. Advanced Drug Delivery Reviews 2022, 182, 114097. [Google Scholar] [CrossRef]
- Tang, T.; Chen, Y.; Zhao, Z.; Bai, Q.; Leisner JJ,Liu, T. Nisin-loaded chitosan/sodium alginate microspheres enhance the antimicrobial efficacy of nisin against Staphylococcus aureus. Journal of Applied Microbiology 2024, 135. [CrossRef]
- Hayes, H.C.; Luk, L.Y.P.; Tsai, Y.-H. Approaches for peptide and protein cyclisation. Organic & Biomolecular Chemistry 2021, 19, 3983–4001. [Google Scholar] [CrossRef]
- Bellavita, R.; Braccia, S.; Galdiero, S.; Falanga, A. Glycosylation and Lipidation Strategies: Approaches for Improving Antimicrobial Peptide Efficacy. Pharmaceuticals 2023, 16, 439. [Google Scholar] [CrossRef]
- Erak, M.; Bellmann-Sickert, K.; Els-Heindl, S.; Beck-Sickinger, A.G. Peptide chemistry toolbox – Transforming natural peptides into peptide therapeutics. Bioorganic & Medicinal Chemistry 2018, 26, 2759–2765. [Google Scholar] [CrossRef]
- Biondi, S.; Chugunova, E.; Panunzio, M. Chapter 8 - From Natural Products to Drugs: Glyco- and Lipoglycopeptides, a New Generation of Potent Cell Wall Biosynthesis Inhibitors. In Atta-ur-Rahman (Ed.) 2016, (Vol. 50, pp. 249–297). Elsevier. [CrossRef]
- Yan, J.; Cai, J.; Zhang, B.; Wang, Y.; Wong DF,Siu, S. W.I. Recent Progress in the Discovery and Design of Antimicrobial Peptides Using Traditional Machine Learning and Deep Learning. Antibiotics 2022, 11, 1451. [Google Scholar] [CrossRef]
- Wang, Y.; Ding, Y.; Wen, H.; Lin, Y.; Hu, Y.; Zhang, Y.; … Lin, Z. QSAR Modeling and Design of Cationic Antimicrobial Peptides Based on Structural Properties of Amino Acids. Combinatorial Chemistry & High Throughput Screening 2012, 15, 347–353. [Google Scholar] [CrossRef]
- Nedyalkova, M.; Paluch AS, Vecini DP,Lattuada, M. Progress and future of the computational design of antimicrobial peptides (AMPs): bio-inspired functional molecules. Digital Discovery 2024, 3, 9–22. [Google Scholar] [CrossRef]
- Cardoso, M.H.; Orozco, R.Q.; Rezende, S.B.; Rodrigues, G.; Oshiro, K.G.N.; Cândido, E.S.; Franco, O.L. (2020). Computer-Aided Design of Antimicrobial Peptides: Are We Generating Effective Drug Candidates? Frontiers in Microbiology, 10. [CrossRef]
- Ramazi, S.; Mohammadi, N.; Allahverdi, A.; Khalili, E.; Abdolmaleki, P. A review on antimicrobial peptides databases and the computational tools. Database 2022, 2022. [Google Scholar] [CrossRef] [PubMed]
- Agüero-Chapin, G.; Galpert-Cañizares, D.; Domínguez-Pérez, D.; Marrero-Ponce, Y.; Pérez-Machado, G.; Teijeira, M.; Antunes, A. Emerging Computational Approaches for Antimicrobial Peptide Discovery. Antibiotics 2022, 11, 936. [Google Scholar] [CrossRef] [PubMed]
- Mwangi, J.; Kamau, P.; Thuku, R.; Lai, R. Design Methods for Antimicrobial Peptides with Improved Performance. Zoological Research 2023, 0, 0–0. [Google Scholar] [CrossRef] [PubMed]
- Brizuela, C.A.; Liu, G.; Stokes, J.M.; de la Fuente-Nunez, C. Methods for Antimicrobial Peptides: Progress and Challenges. Microbial Biotechnology 2025, 18. [Google Scholar] [CrossRef]
- Zhou, X.; Liu, G.; Cao, S.; Lv, J. Deep Learning for Antimicrobial Peptides: Computational Models and Databases. Journal of Chemical Information and Modeling 2025, 65, 1708–1717. [Google Scholar] [CrossRef]
- Mirdita, M.; Schütze, K.; Moriwaki, Y.; Heo, L.; Ovchinnikov, S.; Steinegger, M. ColabFold: making protein folding accessible to all. Nature Methods 2022, 19, 679–682. [Google Scholar] [CrossRef]
- Baek, M.; DiMaio, F.; Anishchenko, I.; Dauparas, J.; Ovchinnikov, S.; Lee GR, … Baker, D. Accurate prediction of protein structures and interactions using a three-track neural network. Science 2021, 373, 871–876. [Google Scholar] [CrossRef]
- Humphreys, I.R.; Pei, J.; Baek, M.; Krishnakumar, A.; Anishchenko, I.; Ovchinnikov, S.; … Baker, D. Computed structures of core eukaryotic protein complexes. Science 2021, 374. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. Journal of Computational Chemistry 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Giulini, M.; Reys, V.; Teixeira, J.M. C., Jiménez-García, B., Honorato, R.V., Kravchenko, A., … Bonvin, A.M.J.J. (2025, May 7). HADDOCK3: A modular and versatile platform for integrative modelling of biomolecular complexes. [CrossRef]
- Kurcinski, M.; Jamroz, M.; Blaszczyk, M.; Kolinski, A.; Kmiecik, S. CABS-dock web server for the flexible docking of peptides to proteins without prior knowledge of the binding site. Nucleic Acids Research 2015, 43(W1), W419–W424. [Google Scholar] [CrossRef]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX, 2015; 1–2, 19–25. [Google Scholar] [CrossRef]
- Huang, J.; MacKerell, A.D. CHARMM36 all-atom additive protein force field: Validation based on comparison to NMR data. Journal of Computational Chemistry 2013, 34, 2135–2145. [Google Scholar] [CrossRef]
- Marrink, S.J.; Risselada, H.J.; Yefimov, S.; Tieleman, D.P.; de Vries, A.H. The MARTINI Force Field: Coarse Grained Model for Biomolecular Simulations. The Journal of Physical Chemistry B 2007, 111, 7812–7824. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.; Cao, Z.; Li, M.; Wang, S. ACEP: improving antimicrobial peptides recognition through automatic feature fusion and amino acid embedding. BMC Genomics 2020, 21, 597. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Martín, F.; Annaval, T.; Buchoux, S.; Sarazin, C.; D’Amelio, N. ADAPTABLE: a comprehensive web platform of antimicrobial peptides tailored to the user’s research. Life Science Alliance 2019, 2, e201900512. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Xu, F.; Wei, L.; Jiang, Y.; Chen, J.; Wei, L.; Wei, D.-Q. AFP-MFL: accurate identification of antifungal peptides using multi-view feature learning. Briefings in Bioinformatics 2023, 24. [Google Scholar] [CrossRef]
- Varadi, M.; Bertoni, D.; Magana, P.; Paramval, U.; Pidruchna, I.; Radhakrishnan, M.; … Velankar, S. AlphaFold Protein Structure Database in 2024: providing structure coverage for over 214 million protein sequences. Nucleic Acids Research 2024, 52(D1), D368–D375. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; … Hassabis, D. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Bajiya, N.; Choudhury, S.; Dhall, A.; Raghava, G.P.S. AntiBP3: A Method for Predicting Antibacterial Peptides against Gram-Positive/Negative/Variable Bacteria. Antibiotics 2024, 13, 168. [Google Scholar] [CrossRef]
- Veltri, D.; Kamath, U.; Shehu, A. Improving Recognition of Antimicrobial Peptides and Target Selectivity through Machine Learning and Genetic Programming. IEEE/ACM Transactions on Computational Biology and Bioinformatics 2017, 14, 300–313. [Google Scholar] [CrossRef]
- Torrent, M.; Nogués VM,Boix, E. A theoretical approach to spot active regions in antimicrobial proteins. BMC Bioinformatics 2009, 10, 373. [Google Scholar] [CrossRef]
- Torrent, M.; Di Tommaso, P.; Pulido, D.; Nogués MV, Notredame C, Boix E,Andreu, D. AMPA: an automated web server for prediction of protein antimicrobial regions. Bioinformatics 2012, 28, 130–131. [Google Scholar] [CrossRef] [PubMed]
- Salem, M.; Keshavarzi Arshadi, A.; Yuan, J.S. AMPDeep: hemolytic activity prediction of antimicrobial peptides using transfer learning. BMC Bioinformatics 2022, 23, 389. [Google Scholar] [CrossRef]
- Lawrence, T.J.; Carper, D.L.; Spangler, M.K.; Carrell, A.A.; Rush, T.A.; Minter, S.J.; …, L.a.b.b.é.; JL amPEPpy 1. 0: a portable and accurate antimicrobial peptide prediction tool. Bioinformatics 2021, 37, 2058–2060. [Google Scholar] [CrossRef]
- Burdukiewicz, M.; Sidorczuk, K.; Rafacz, D.; Pietluch, F.; Chilimoniuk, J.; Rödiger, S.; Gagat, P. Proteomic Screening for Prediction and Design of Antimicrobial Peptides with AmpGram. International Journal of Molecular Sciences 2020, 21, 4310. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Yan, J.; Un, C.; Wang, Y.; Campbell-Valois, F.-X.; Siu, S.W.I. BERT-AmPEP60: A BERT-Based Transfer Learning Approach to Predict the Minimum Inhibitory Concentrations of Antimicrobial Peptides for Escherichia coli and Staphylococcus aureus. Journal of Chemical Information and Modeling 2025, 65, 3186–3202. [Google Scholar] [CrossRef]
- Porto, W.F.; Pires, Á.S.; Franco, O.L. CS-AMPPred: An Updated SVM Model for Antimicrobial Activity Prediction in Cysteine-Stabilized Peptides. PLoS ONE 2012, 7, e51444. [Google Scholar] [CrossRef] [PubMed]
- Slavokhotova, A.A.; Shelenkov, A.A.; Rogozhin, E.A. Computational Prediction and Structural Analysis of α-Hairpinins, a Ubiquitous Family of Antimicrobial Peptides, Using the Cysmotif Searcher Pipeline. Antibiotics 2024, 13, 1019. [Google Scholar] [CrossRef]
- Azim, S.M.; Sharma, A.; Shatabda, S.; Dehzangi, A. (2021, October 29). DeepAmp: A Convolutional Neural Network Based Tool for Predicting Protein AMPylation Sites From Binary Profile Representation. [CrossRef]
- Xu, J.; Li, F.; Li, C.; Guo, X.; Landersdorfer, C.; Shen, H.-H.; … Song, J. iAMPCN: a deep-learning approach for identifying antimicrobial peptides and their functional activities. Briefings in Bioinformatics 2023, 24. [Google Scholar] [CrossRef]
- Zheng, W.; Wuyun, Q.; Li, Y.; Liu, Q.; Zhou, X.; Peng, C.; … Zhang, Y. (2025). Deep-learning-based single-domain and multidomain protein structure prediction with D-I-TASSER. Nature Biotechnology. [CrossRef]
- Porto, W.F.; Fensterseifer, I.C.M.; Ribeiro, S.M.; Franco, O.L. Joker: An algorithm to insert patterns into sequences for designing antimicrobial peptides. Biochimica et Biophysica Acta (BBA) - General Subjects 2018, 1862, 2043–2052. [Google Scholar] [CrossRef]
- Tang, W.; Dai, R.; Yan, W.; Zhang, W.; Bin, Y.; Xia, E.; Xia, J. Identifying multi-functional bioactive peptide functions using multi-label deep learning. Briefings in Bioinformatics 2022, 23. [Google Scholar] [CrossRef]
- Müller, A.T.; Gabernet, G.; Hiss, J.A.; Schneider, G. modlAMP: Python for antimicrobial peptides. Bioinformatics 2017, 33, 2753–2755. [Google Scholar] [CrossRef]
- Phillips, J.C.; Braun, R.; Wang, W.; Gumbart, J.; Tajkhorshid, E.; Villa, E.; … Schulten, K. Scalable molecular dynamics with NAMD. Journal of Computational Chemistry 2005, 26, 1781–1802. [Google Scholar] [CrossRef]
- Singh, H.; Singh, S.; Singh Raghava, G.P. (2019, February 25). Peptide Secondary Structure Prediction using Evolutionary Information. [CrossRef]
- Xu, J.; Li, F.; Leier, A.; Xiang, D.; Shen, H.-H. ; Marquez Lago TT, … Song, J. Comprehensive assessment of machine learning-based methods for predicting antimicrobial peptides. Briefings in Bioinformatics, 2021; 22. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. Journal of Molecular Graphics 1996, 14, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Ma, Y.; Yu, K.; Dong, J.; Zeng, W. Integrated computational approaches for advancing antimicrobial peptide development. Trends in Pharmacological Sciences 2024, 45, 1046–1060. [Google Scholar] [CrossRef]
- Bahrami, S.; Andishmand, H.; Pilevar, Z.; Hashempour-baltork, F.; Torbati, M.; Dadgarnejad, M.; … Azadmard-Damirchi, S. Innovative perspectives on bacteriocins: advances in classification, synthesis, mode of action, and food industry applications. Journal of Applied Microbiology 2024, 135. [Google Scholar] [CrossRef] [PubMed]
- Parada Fabián, J.C.; Álvarez Contreras, A.K.; Natividad Bonifacio, I.; Hernández Robles, M.F.; Vázquez Quiñones, C.R.; Quiñones Ramírez, E.I.; Vázquez Salinas, C. Toward safer and sustainable food preservation: a comprehensive review of bacteriocins in the food industry. Bioscience Reports 2025, 45, 277–302. [Google Scholar] [CrossRef]
- Peres Fabbri, L.; Cavallero, A.; Vidotto, F.; Gabriele, M. Bioactive Peptides from Fermented Foods: Production Approaches, Sources, and Potential Health Benefits. Foods 2024, 13, 3369. [Google Scholar] [CrossRef] [PubMed]
- Bisht, V.; Das, B.; Hussain, A.; Kumar, V.; Navani, N.K. Understanding of probiotic origin antimicrobial peptides: a sustainable approach ensuring food safety. npj Science of Food 2024, 8, 67. [Google Scholar] [CrossRef]
- Pérez de la Lastra, J.M.; González-Acosta, S.; Otazo-Pérez, A.; Asensio-Calavia, P.; Rodríguez-Borges, V.M. Antimicrobial Peptides for Food Protection: Leveraging Edible Mushrooms and Nano-Innovation. Dietetics 2025, 4, 9. [Google Scholar] [CrossRef]
- Chauhan, K.; Rao, A. Clean-label alternatives for food preservation: An emerging trend. Heliyon 2024, 10, e35815. [Google Scholar] [CrossRef]
- Kumar, L.; Tyagi, P.; Lucia, L.; Pal, L. Innovations in Edible Packaging Films, Coatings, and Antimicrobial Agents for Applications in Food Industry. Comprehensive Reviews in Food Science and Food Safety 2025, 24. [Google Scholar] [CrossRef]
- Choi, D.; Bedale, W.; Chetty, S.; Yu, J. Comprehensive review of clean-label antimicrobials used in dairy products. Comprehensive Reviews in Food Science and Food Safety 2024, 23. [Google Scholar] [CrossRef] [PubMed]
- Min, K.H.; Kim, K.H.; Ki, M.-R.; Pack, S.P. Antimicrobial Peptides and Their Biomedical Applications: A Review. Antibiotics 2024, 13, 794. [Google Scholar] [CrossRef] [PubMed]
- Sengkhui, S.; Klubthawee, N.; Aunpad, R. A novel designed membrane-active peptide for the control of foodborne Salmonella enterica serovar Typhimurium. Scientific Reports 2023, 13, 3507. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Dekker, M.; Heising, J.; Zhao, L.; Fogliano, V. Food matrix design can influence the antimicrobial activity in the food systems: A narrative review. Critical Reviews in Food Science and Nutrition 2024, 64, 8963–8989. [Google Scholar] [CrossRef]
- Chu, Z.; Wang, H.; Dong, B. Research on Food Preservation Based on Antibacterial Technology: Progress and Future Prospects. Molecules 2024, 29, 3318. [Google Scholar] [CrossRef]
- Swann, M. Swann. (1969). Report. H.M.S.O.
- Rodrigues, G.; Maximiano MR,Franco, O. L. Antimicrobial peptides used as growth promoters in livestock production. Applied Microbiology and Biotechnology 2021, 105, 7115–7121. [Google Scholar] [CrossRef]
- Vercelli, C.; Gambino, G.; Amadori, M.; Re, G. Implications of Veterinary Medicine in the comprehension and stewardship of antimicrobial resistance phenomenon. From the origin till nowadays. Veterinary and Animal Science 2022, 16, 100249. [Google Scholar] [CrossRef]
- Pomba, C.; Rantala, M.; Greko, C.; Baptiste KE, Catry B, van Duijkeren E, … Törneke, K. Public health risk of antimicrobial resistance transfer from companion animals. Journal of Antimicrobial Chemotherapy 2016, dkw481. [Google Scholar] [CrossRef]
- Vittecoq, M.; Godreuil, S.; Prugnolle, F.; Durand, P.; Brazier, L.; Renaud, N.; … Renaud, F. Antimicrobial resistance in wildlife. Journal of Applied Ecology 2016, 53, 519–529. [Google Scholar] [CrossRef]
- Valdez-Miramontes, C.E.; De Haro-Acosta, J.; Aréchiga-Flores, C.F.; Verdiguel-Fernández, L.; Rivas-Santiago, B. Antimicrobial peptides in domestic animals and their applications in veterinary medicine. Peptides 2021, 142, 170576. [Google Scholar] [CrossRef]
- Tai, H.-M.; Huang, H.-N.; Tsai, T.-Y.; You, M.-F.; Wu, H.-Y.; Rajanbabu, V.; … Chen, J.-Y. Dietary supplementation of recombinant antimicrobial peptide Epinephelus lanceolatus piscidin improves growth performance and immune response in Gallus gallus domesticus. PLOS ONE 2020, 15, e0230021. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.-S.; Shao, H.; Li, T.-J.; Tang, Z.-R.; Huang, R.-L.; Wang, S.-P.; … Yin, Y.-L. Dietary Supplementation with Bovine Lactoferrampin–Lactoferricin Produced by Pichia pastoris Fed-batch Fermentation Affects Intestinal Microflora in Weaned Piglets. Applied Biochemistry and Biotechnology 2012, 168, 887–898. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.H.; Ingale, S.L.; Kim, J.S.; Kim, K.H.; Lohakare, J.; Park, Y.K.; … Chae, B.J. Effects of dietary supplementation with antimicrobial peptide-P5 on growth performance, apparent total tract digestibility, faecal and intestinal microflora and intestinal morphology of weanling pigs. Journal of the Science of Food and Agriculture 2013, 93, 587–592. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhao, X.; Zhu, C.; Zhao, Y.; Liu, S.; Xia, X.; … Hu, J. The antimicrobial peptide MPX kills Actinobacillus pleuropneumoniae and reduces its pathogenicity in mice. Veterinary Microbiology 2020, 243, 108634. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, Q.; Dong, M.; Song, H.; Hang, B.; Sun, Y.; … Hu, J. Evaluation of the efficacy of the antimicrobial peptide HJH-3 in chickens infected with Salmonella Pullorum. Frontiers in Microbiology 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Bruhn, O.; Grötzinger, J.; Cascorbi, I.; Jung, S. Antimicrobial peptides and proteins of the horse - insights into a well-armed organism. Veterinary Research 2011, 42, 98. [Google Scholar] [CrossRef]
- Shen, H.; Li, Y.; Pi, Q.; Tian, J.; Xu, X.; Huang, Z.; … Mao, S. (2025). Unveiling novel antimicrobial peptides from the ruminant gastrointestinal microbiomes: A deep learning-driven approach yields an anti-MRSA candidate. Journal of Advanced Research. [CrossRef]
- Rakers, S.; Niklasson, L.; Steinhagen, D.; Kruse, C.; Schauber, J.; Sundell, K.; Paus, R. Antimicrobial Peptides (AMPs) from Fish Epidermis: Perspectives for Investigative Dermatology. Journal of Investigative Dermatology 2013, 133, 1140–1149. [Google Scholar] [CrossRef] [PubMed]
- Shabir, U.; Ali, S.; Magray AR, Ganai BA, Firdous P, Hassan T,Nazir, R. Fish antimicrobial peptides (AMP’s) as essential and promising molecular therapeutic agents: A review. Microbial Pathogenesis 2018, 114, 50–56. [Google Scholar] [CrossRef]
- García-Beltrán, J.M.; Arizcun, M.; Chaves-Pozo, E. Antimicrobial Peptides from Photosynthetic Marine Organisms with Potential Application in Aquaculture. Marine Drugs 2023, 21, 290. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, T.; Guardiola FA, Almeida D,Antunes, A. Aquatic Invertebrate Antimicrobial Peptides in the Fight Against Aquaculture Pathogens. Microorganisms 2025, 13, 156. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Falla, T.J. Antimicrobial peptides: therapeutic potential. Expert Opinion on Pharmacotherapy 2006, 7, 653–663. [Google Scholar] [CrossRef]
- Spindler, E.C.; Hale, J.D.F.; Giddings, T.H.; Hancock, R.E.W.; Gill, R.T. Deciphering the Mode of Action of the Synthetic Antimicrobial Peptide Bac8c. Antimicrobial Agents and Chemotherapy 2011, 55, 1706–1716. [Google Scholar] [CrossRef]
- Stern, N.J.; Svetoch, E.A.; Eruslanov, B.V.; Perelygin, V.V.; Mitsevich, E.V.; Mitsevich, I.P.; … Seal, B.S. Isolation of a Lactobacillus salivarius Strain and Purification of Its Bacteriocin, Which Is Inhibitory to Campylobacter jejuni in the Chicken Gastrointestinal System. Antimicrobial Agents and Chemotherapy 2006, 50, 3111–3116. [Google Scholar] [CrossRef]
- Park, C.B.; Kim, H.S.; Kim, S.C. Mechanism of Action of the Antimicrobial Peptide Buforin II: Buforin II Kills Microorganisms by Penetrating the Cell Membrane and Inhibiting Cellular Functions. Biochemical and Biophysical Research Communications 1998, 244, 253–257. [Google Scholar] [CrossRef]
- Cursino, L.; Smajs, D.; Smarda, J.; Nardi, R.M. D. , Nicoli, J.R., Chartone-Souza, E., & Nascimento, A.M.A. Exoproducts of the Escherichia coli strain H22 inhibiting some enteric pathogens both in vitro and in vivo. Journal of Applied Microbiology 2006, 100, 821–829. [Google Scholar] [CrossRef]
- Nagant, C.; Tré-Hardy, M.; El-Ouaaliti, M.; Savage, P.; Devleeschouwer, M.; Dehaye, J.-P. Interaction between tobramycin and CSA-13 on clinical isolates of Pseudomonas aeruginosa in a model of young and mature biofilms. Applied Microbiology and Biotechnology 2010, 88, 251–263. [Google Scholar] [CrossRef]
- Kang, B.S.; Seo, J.-G.; Lee, G.-S.; Kim, J.-H.; Kim, S.Y.; Han, Y.W.; … Park, Y.M. Antimicrobial activity of enterocins from Enterococcus faecalis SL-5 against Propionibacterium acnes, the causative agent in acne vulgaris, and its therapeutic effect. The Journal of Microbiology 2009, 47, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Shriwastav, S.; Kaur, N.; Hassan, M.; Ahmed Mohammed, S.; Chauhan, S.; Mittal, D.; … Bibi, A. Antimicrobial peptides: a promising frontier to combat antibiotic resistant pathogens. Annals of Medicine and Surgery 2025, 87. Retrieved from https://journals.lww.com/annals-of-medicine-and-surgery/fulltext/2025/04000/antimicrobial_peptides__a_promising_frontier_to.45.aspx.
- Koo, H.B.; Seo, J. Antimicrobial peptides under clinical investigation. Peptide Science 2019, 111, e24122. [Google Scholar] [CrossRef]
- Harris, S.M.; Zhang, L.; Parente, J.; Rodeheaver, G.T.; Falla, T.J. 111 HB-50: A Pre-Clinical Study of a Prophylactic for Wound Infection. Wound Repair and Regeneration 2004, 12. [Google Scholar] [CrossRef]
- Kharrat, O. ; Yamaryo-Botté; Y; Nasreddine, R.; Voisin, S.; Aumer, T.; Cammue, B.P. A., … Landon, C. The antimicrobial activity of ETD151 defensin is dictated by the presence of glycosphingolipids in the targeted organisms. Proceedings of the National Academy of Sciences, 2025; 122. [Google Scholar] [CrossRef]
- Sebők, C.; Tráj, P.; Mackei, M.; Márton RA, Vörösházi J, Kemény Á, … Mátis, G. Modulation of the immune response by the host defense peptide IDR-1002 in chicken hepatic cell culture. Scientific Reports 2023, 13, 14530. [Google Scholar] [CrossRef]
- Turovskiy, Y.; Ludescher RD, Aroutcheva AA, Faro S,Chikindas, M. L. Lactocin 160, a Bacteriocin Produced by Vaginal Lactobacillus rhamnosus, Targets Cytoplasmic Membranes of the Vaginal Pathogen, Gardnerella vaginalis. Probiotics and Antimicrobial Proteins 2009, 1, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Reddy, K.V. R. , Aranha, C., Gupta, S.M., & Yedery, R.D. Evaluation of antimicrobial peptide nisin as a safe vaginal contraceptive agent in rabbits: in vitro and in vivo studies. Reproduction 2004, 128, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Pantarat, N.; Knappe, D. ; Reynolds EC, Hoffmann R,OBrien-Simpson, N.M. (2025). Novel action of proline-rich antimicrobial peptides Api88, Api137, Onc72 and Onc112 against Pseudomonas aeruginosa and Escherichia coli in ion-rich environments. bioRxiv. [CrossRef]
- Dabour, N.; Zihler, A.; Kheadr, E.; Lacroix, C.; Fliss, I. In vivo study on the effectiveness of pediocin PA-1 and Pediococcus acidilactici UL5 at inhibiting Listeria monocytogenes. International Journal of Food Microbiology 2009, 133, 225–233. [Google Scholar] [CrossRef]
- Molchanova, N.; Hansen, P.; Franzyk, H. Advances in Development of Antimicrobial Peptidomimetics as Potential Drugs. Molecules 2017, 22, 1430. [Google Scholar] [CrossRef]
- Castiglione, F.; Cavaletti, L.; Losi, D.; Lazzarini, A.; Carrano, L.; Feroggio, M.; … Selva, E. A Novel Lantibiotic Acting on Bacterial Cell Wall Synthesis Produced by the Uncommon Actinomycete Planomonospora sp. Biochemistry 2007, 46, 5884–5895. [Google Scholar] [CrossRef]
- Crost, E.H.; Ajandouz, E.H.; Villard, C.; Geraert, P.A.; Puigserver, A.; Fons, M. Ruminococcin C, a new anti-Clostridium perfringens bacteriocin produced in the gut by the commensal bacterium Ruminococcus gnavus E1. Biochimie 2011, 93, 1487–1494. [Google Scholar] [CrossRef]
- Rossetto, G.; Bergese, P.; Colombi, P.; Depero LE, Giuliani A, Nicoletto SF,Pirri, G. Atomic force microscopy evaluation of the effects of a novel antimicrobial multimeric peptide on Pseudomonas aeruginosa. Nanomedicine: Nanotechnology 2007, Biology and Medicine, 3, 198–207. [CrossRef]
- Dourado, F.S.; Leite, J.R.S.A.; Silva, L.P.; Melo, J.A.T.; Bloch, C.; Schwartz, E.F. Antimicrobial peptide from the skin secretion of the frog Leptodactylus syphax. Toxicon 2007, 50, 572–580. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.B.; Conlon, J.M.; Iwamuro, S.; Knoop, F.C. Antimicrobial peptides from the skin of the Japanese mountain brown frog, Rana ornativentris. The Journal of Peptide Research 2001, 58, 349–356. [Google Scholar] [CrossRef]
- Bulger, E.M.; Maier, R.V.; Sperry, J.; Joshi, M.; Henry, S.; Moore, F.A.; … Shirvan, A. A Novel Drug for Treatment of Necrotizing Soft-Tissue Infections. JAMA Surgery 2014, 149, 528. [Google Scholar] [CrossRef]
- Sugimoto, A.; Maeda, A.; Itto, K.; Arimoto, H. Deciphering the mode of action of cell wall-inhibiting antibiotics using metabolic labeling of growing peptidoglycan in Streptococcus pyogenes. Scientific Reports 2017, 7, 1129. [Google Scholar] [CrossRef]
- Mba, I.E.; Nweze, E.I. Antimicrobial Peptides Therapy: An Emerging Alternative for Treating Drug-Resistant Bacteria. The Yale journal of biology and medicine 2022, 95, 445–463. [Google Scholar]
- Jadi, P.K.; Sharma, P.; Bhogapurapu, B.; Roy, S. Alternative Therapeutic Interventions: Antimicrobial Peptides and Small Molecules to Treat Microbial Keratitis. Frontiers in Chemistry 2021, 9. [Google Scholar] [CrossRef]
- Shriwastav, S.; Kaur, N.; Hassan, M.; Ahmed Mohammed, S.; Chauhan, S.; Mittal, D.; … Bibi, A. Antimicrobial peptides: a promising frontier to combat antibiotic resistant pathogens. Annals of Medicine and Surgery 2025, 87. Retrieved from https://journals.lww.com/annals-of-medicine-and-surgery/fulltext/2025/04000/antimicrobial_peptides__a_promising_frontier_to.45.aspx.
- Kirkpatrick, D.L.; Powis, G. Clinically Evaluated Cancer Drugs Inhibiting Redox Signaling. Antioxidants & Redox Signaling 2017, 26, 262–273. [Google Scholar] [CrossRef]
- You, Y.; Liu, H.; Zhu, Y.; Zheng, H. Rational design of stapled antimicrobial peptides. Amino Acids 2023, 55, 421–442. [Google Scholar] [CrossRef] [PubMed]
- North, J.R.; Takenaka, S.; Rozek, A.; Kielczewska, A.; Opal, S.; Morici, L.A.; … Donini, O. A novel approach for emerging and antibiotic resistant infections: Innate defense regulators as an agnostic therapy. Journal of Biotechnology 2016, 226, 24–34. [Google Scholar] [CrossRef]
- Surur, A.S.; Sun, D. Macrocycle-Antibiotic Hybrids: A Path to Clinical Candidates. Frontiers in Chemistry 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Wu, J.; Chen, Y.; Fan, K.; Yu, X.; Li, X.; … Chen, M. Efficacy and Safety of PL-5 (Peceleganan) Spray for Wound Infections. Annals of Surgery 2023, 277, 43–49. [Google Scholar] [CrossRef]
- Khurshid, Z.; Najeeb, S.; Mali, M.; Moin SF, Raza SQ, Zohaib S, … Zafar, M. S. Histatin peptides: Pharmacological functions and their applications in dentistry. Saudi Pharmaceutical Journal 2017, 25, 25–31. [Google Scholar] [CrossRef]
- Wiig, M.E.; Dahlin, L.B.; Fridén, J.; Hagberg, L.; Larsen, S.E.; Wiklund, K.; Mahlapuu, M. PXL01 in Sodium Hyaluronate for Improvement of Hand Recovery after Flexor Tendon Repair Surgery: Randomized Controlled Trial. PLoS ONE 2014, 9, e110735. [Google Scholar] [CrossRef]
- McCullough, P.A.; Bennett-Guerrero, E.; Chawla, L.S.; Beaver, T.; Mehta, R.L.; Molitoris, B.A.; … Khan, S. ABT-719 for the Prevention of Acute Kidney Injury in Patients Undergoing High-Risk Cardiac Surgery: A Randomized Phase 2b Clinical Trial. Journal of the American Heart Association 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.L.; He, X.; Shi, W. Precision Reengineering of the Oral Microbiome for Caries Management. Advances in Dental Research 2019, 30, 34–39. [Google Scholar] [CrossRef]
- EGreber, K.; Dawgul, M. Antimicrobial Peptides Under Clinical Trials. Current Topics in Medicinal Chemistry 2016, 17, 620–628. [Google Scholar] [CrossRef]
- Håkansson, J.; Ringstad, L.; Umerska, A.; Johansson, J.; Andersson, T.; Boge, L.; … Mahlapuu, M. Characterization of the in vitro, ex vivo, and in vivo Efficacy of the Antimicrobial Peptide DPK-060 Used for Topical Treatment. Frontiers in Cellular and Infection Microbiology 2019, 9. [Google Scholar] [CrossRef]
- Khatib, M.N.; Shankar, A.H.; Kirubakaran, R.; Gaidhane, A.; Gaidhane, S.; Simkhada, P.; Quazi Syed, Z. Ghrelin for the management of cachexia associated with cancer. Cochrane Database of Systematic Reviews 2018, 2020. [Google Scholar] [CrossRef]
- Parisi, K.; McKenna JA, Lowe R, Harris KS, Shafee T, Guarino R, … Anderson, M. A. Hyperpolarisation of Mitochondrial Membranes Is a Critical Component of the Antifungal Mechanism of the Plant Defensin, Ppdef1. Journal of Fungi 2024, 10, 54. [Google Scholar] [CrossRef] [PubMed]
- Donini, O.; Watkins BA, Palardy J, Opal S, Sonis S, Abrams MJ,North, J. R. Reduced Infection and Mucositis In Chemotherapy-Treated Animals Following Innate Defense Modulation Using a Novel Drug Candidate. Blood 2010, 116, 3781–3781. [Google Scholar] [CrossRef]
- Mercer, D.K.; Robertson, J.C.; Miller, L.; Stewart, C.S.; O’Neil, D.A. NP213 (Novexatin®): A unique therapy candidate for onychomycosis with a differentiated safety and efficacy profile. Medical Mycology 2020, 58, 1064–1072. [Google Scholar] [CrossRef] [PubMed]
- Ming, L.; Huang, J.-A. The Antibacterial Effects of Antimicrobial Peptides OP-145 against Clinically Isolated Multi-Resistant Strains. Japanese Journal of Infectious Diseases 2017, 70, 601–603. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.-F.; Han, B.-C.; Lin, W.-Y.; Wang, S.-Y.; Linn TY, Hsu H-W, … Chang, W. -J. Efficacy of antimicrobial peptide P113 oral health care products on the reduction of oral bacteria number and dental plaque formation in a randomized clinical assessment. Journal of Dental Sciences 2024, 19, 2367–2376. [Google Scholar] [CrossRef]
- Cheng, K.-T.; Wu, C.-L.; Yip, B.-S.; Chih, Y.-H.; Peng, K.-L.; Hsu, S.-Y.; … Cheng, J.-W. The Interactions between the Antimicrobial Peptide P-113 and Living Candida albicans Cells Shed Light on Mechanisms of Antifungal Activity and Resistance. International Journal of Molecular Sciences 2020, 21, 2654. [Google Scholar] [CrossRef]
- Kowalski, R.P.; Romanowski, E.G.; Yates, K.A.; Mah, F.S. An Independent Evaluation of a Novel Peptide Mimetic, Brilacidin (PMX30063), for Ocular Anti-Infective. Journal of Ocular Pharmacology and Therapeutics 2016, 32, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.-R.; Yang, L.-M.; Wang, Y.-H.; Pang, W.; Tam, S.-C.; Tien, P.; Zheng, Y.-T. Sifuvirtide, a potent HIV fusion inhibitor peptide. Biochemical and Biophysical Research Communications 2009, 382, 540–544. [Google Scholar] [CrossRef]
- van derDoes, A.M.; Bogaards, S.J.P.; Ravensbergen, B.; Beekhuizen, H.; van Dissel, J.T.; Nibbering, P.H. Antimicrobial Peptide hLF1-11 Directs Granulocyte-Macrophage Colony-Stimulating Factor-Driven Monocyte Differentiation toward Macrophages with Enhanced Recognition and Clearance of Pathogens. Antimicrobial Agents and Chemotherapy 2010, 54, 811–816. [Google Scholar] [CrossRef]
- Ridyard, K.E.; Overhage, J. The Potential of Human Peptide LL-37 as an Antimicrobial and Anti-Biofilm Agent. Antibiotics (Basel Switzerland) 2021, 10. [CrossRef]
- Isaksson, J.; Brandsdal BO, Engqvist M, Flaten GE, Svendsen JSM,Stensen, W. A Synthetic Antimicrobial Peptidomimetic (LTX 109): Stereochemical Impact on Membrane Disruption. Journal of Medicinal Chemistry 2011, 54, 5786–5795. [Google Scholar] [CrossRef]
- Chen, X.; Li, S.; Yu, L.; Miller, A.; Du, L. Systematic optimization for production of the anti- MRSA antibiotics WAP-8294A in an engineered strain of Lysobacter enzymogenes. Microbial Biotechnology 2019, 12, 1430–1440. [Google Scholar] [CrossRef]
- Janec, K.J.; Yuan, H.; Norton Jr, J.E.; Kelner, R.H.; Hirt, C.K.; Betensky, R.A.; Guinan, E.C. rBPI 21 (opebacan) promotes rapid trilineage hematopoietic recovery in a murine model of high-dose total body irradiation. American Journal of Hematology 2018, 93, 1002–1013. [Google Scholar] [CrossRef]
- Schneider, T.; Gries, K.; Josten, M.; Wiedemann, I.; Pelzer, S.; Labischinski, H.; Sahl, H.-G. The Lipopeptide Antibiotic Friulimicin B Inhibits Cell Wall Biosynthesis through Complex Formation with Bactoprenol Phosphate. Antimicrobial Agents and Chemotherapy 2009, 53, 1610–1618. [Google Scholar] [CrossRef]
- Scott, M.G.; Dullaghan, E.; Mookherjee, N.; Glavas, N.; Waldbrook, M.; Thompson, A.; … Hancock, R.E.W. An anti-infective peptide that selectively modulates the innate immune response. Nature Biotechnology 2007, 25, 465–472. [Google Scholar] [CrossRef]
- Boakes, S.; Dawson, M.J. Discovery and Development of NVB302, a Semisynthetic Antibiotic for Treatment of Clostridium difficile Infection. In Natural Products (pp. 455–468). Wiley 2014. [CrossRef]
- Xiong, Y.Q.; Hady, W.A.; Deslandes, A.; Rey, A.; Fraisse, L.; Kristensen, H.-H.; … Bayer, A.S. Efficacy of NZ2114, a Novel Plectasin-Derived Cationic Antimicrobial Peptide Antibiotic, in Experimental Endocarditis Due to Methicillin-Resistant Staphylococcus aureus. Antimicrobial Agents and Chemotherapy 2011, 55, 5325–5330. [Google Scholar] [CrossRef]
- Iwasaki, M.; Akiba, Y.; Kaunitz, J.D. Recent advances in vasoactive intestinal peptide physiology and pathophysiology: focus on the gastrointestinal system. F1000Research 2019, 8, 1629. [Google Scholar] [CrossRef]
- Lee, H.-T.; Lee, C.-C.; Yang, J.-R.; Lai, J.Z.C.; Chang, K.Y. A Large-Scale Structural Classification of Antimicrobial Peptides. BioMed Research International 2015, 2015, 1–6. [Google Scholar] [CrossRef]
- Alcock, B.P.; Huynh, W.; Chalil, R.; Smith, K.W.; Raphenya, A.R.; Wlodarski, M.A.; … McArthur, A.G. CARD 2023: expanded curation, support for machine learning, and resistome prediction at the Comprehensive Antibiotic Resistance Database. Nucleic Acids Research 2023, 51, D690–D699. [Google Scholar] [CrossRef]
- Novković, M.; Simunić, J.; Bojović, V.; Tossi, A.; Juretić, D. DADP: the database of anuran defense peptides. Bioinformatics (Oxford, England) 2012, 28, 1406–1407. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.; Guan, J.; Xie, P.; Chung, C.-R.; Zhao, Z.; Dong, D.; … Lee, T.-Y. dbAMP 3.0: updated resource of antimicrobial activity and structural annotation of peptides in the post-pandemic era. Nucleic Acids Research 2025, 53, D364–D376. [Google Scholar] [CrossRef] [PubMed]
- Gautam, A.; Chaudhary, K.; Singh, S.; Joshi, A.; Anand, P.; Tuknait, A.; … Raghava, G.P.S. Hemolytik: a database of experimentally determined hemolytic and non-hemolytic peptides. Nucleic Acids Research 2014, 42, D444–D449. [Google Scholar] [CrossRef] [PubMed]
- Gómez, E.A.; Giraldo, P.; Orduz, S. InverPep: A database of invertebrate antimicrobial peptides. Journal of Global Antimicrobial Resistance 2017, 8, 13–17. [Google Scholar] [CrossRef]
- Bonin, N.; Doster, E.; Worley, H.; Pinnell LJ, Bravo JE, Ferm P, … Boucher, C. MEGARes and AMR++, v3.0: an updated comprehensive database of antimicrobial resistance determinants and an improved software pipeline for classification using high-throughput sequencing. Nucleic acids research 2023, 51, D744–D752. [Google Scholar] [CrossRef]
- D’Aloisio, V.; Dognini, P.; Hutcheon, G.A.; Coxon, C.R. PepTherDia: database and structural composition analysis of approved peptide therapeutics and diagnostics. Drug Discovery Today 2021, 26, 1409–1419. [Google Scholar] [CrossRef] [PubMed]
- Usmani, S.S.; Bedi, G.; Samuel, J.S.; Singh, S.; Kalra, S.; Kumar, P.; … Raghava, G.P.S. THPdb: Database of FDA-approved peptide and protein therapeutics. PLOS ONE 2017, 12, e0181748. [Google Scholar] [CrossRef] [PubMed]
- Piotto, S.P.; Sessa, L.; Concilio, S.; Iannelli, P. YADAMP: yet another database of antimicrobial peptides. International Journal of Antimicrobial Agents 2012, 39, 346–351. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Distribution of AMPs according to their specific activities in different databases. Information obtained from databases available at: DBAASP (https://dbaasp.org/home) [11]; CAMP (https://camp.bicnirrh.res.in/) [12]; SATPdb (http://crdd.osdd.net/raghava/satpdb/) [13]; APD3 (https://aps.unmc.edu/AP/) [8]; Peptipedia (https://app.peptipedia.cl/) [14]; DRAMP (http://dramp.cpu-bioinfor.org) [15]; AMPDB (https://bblserver.org.in/ampdb/) [16]; PlantPepDB (http://www.nipgr.ac.in/PlantPepDB/) [17](accessed on August 24).
Figure 1.
Distribution of AMPs according to their specific activities in different databases. Information obtained from databases available at: DBAASP (https://dbaasp.org/home) [11]; CAMP (https://camp.bicnirrh.res.in/) [12]; SATPdb (http://crdd.osdd.net/raghava/satpdb/) [13]; APD3 (https://aps.unmc.edu/AP/) [8]; Peptipedia (https://app.peptipedia.cl/) [14]; DRAMP (http://dramp.cpu-bioinfor.org) [15]; AMPDB (https://bblserver.org.in/ampdb/) [16]; PlantPepDB (http://www.nipgr.ac.in/PlantPepDB/) [17](accessed on August 24).
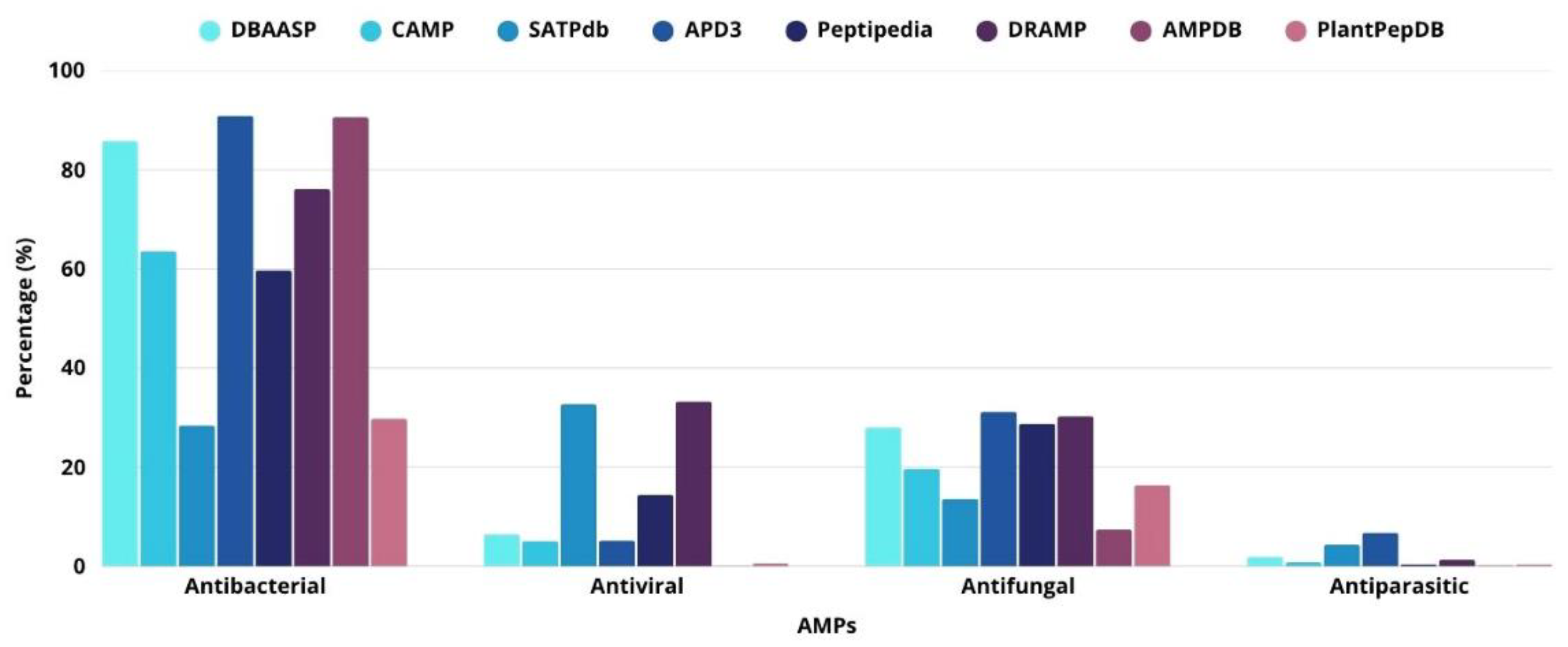
Figure 2.
The scheme represents the different sources of production, biological activities and biochemical influences of AMPs. The central part shows examples of structures described for AMPs. Information obtained from DRAMP (http://dramp.cpu-bioinfor.org/) [15]and predicted by PEP-FOLD3 (https://bioserv.rpbs.univ-paris-diderot.fr/index.html) [39] (accessed August 25, 2025) and figures constructed with clean png (https://www.cleanpng.com/).
Figure 2.
The scheme represents the different sources of production, biological activities and biochemical influences of AMPs. The central part shows examples of structures described for AMPs. Information obtained from DRAMP (http://dramp.cpu-bioinfor.org/) [15]and predicted by PEP-FOLD3 (https://bioserv.rpbs.univ-paris-diderot.fr/index.html) [39] (accessed August 25, 2025) and figures constructed with clean png (https://www.cleanpng.com/).
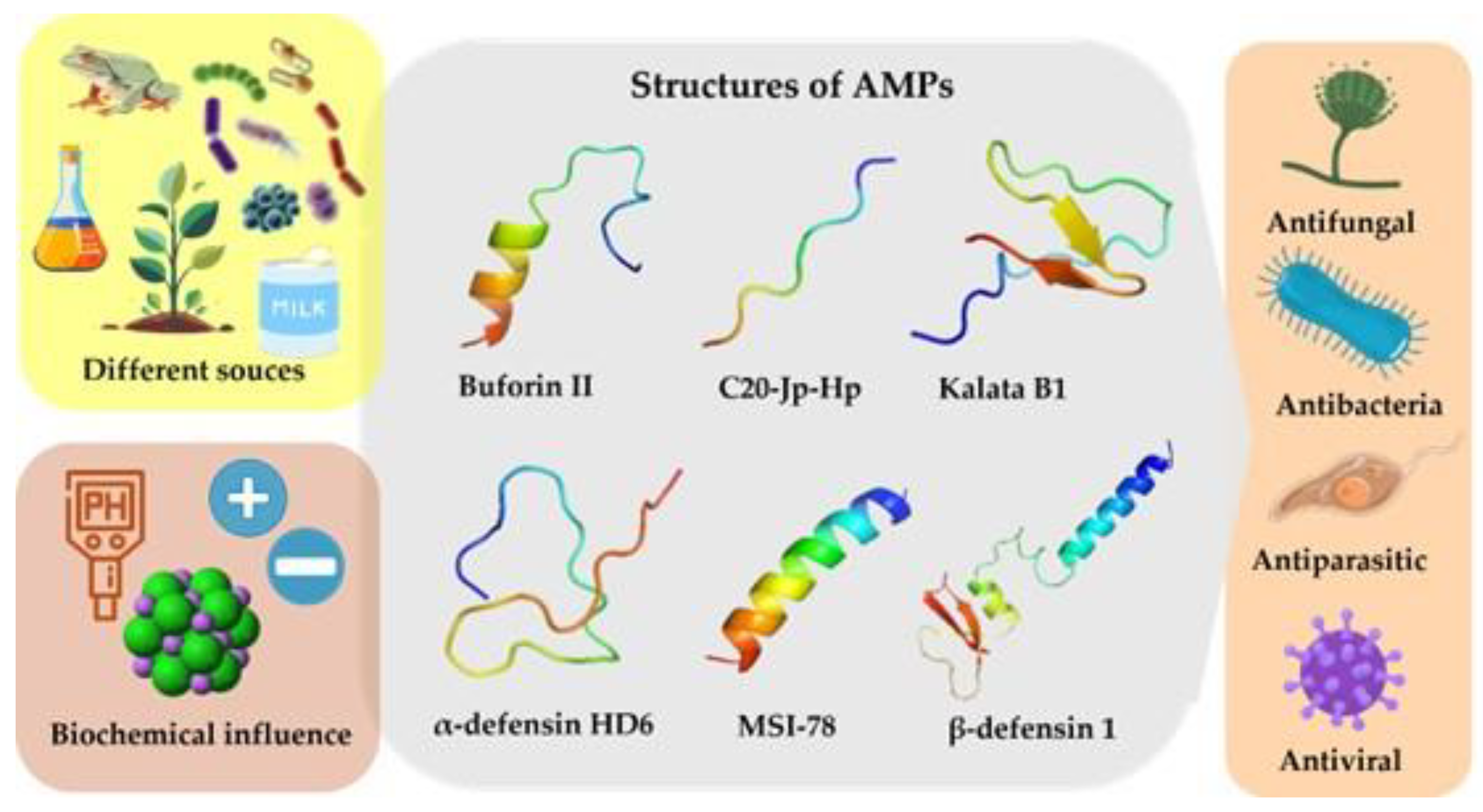
Figure 3.
Two complementary strategies guide the discovery of antimicrobial peptides (AMPs): the identification of novel candidates from natural sources and the computationally guided design of synthetic AMPs. Protein images used in this figure were obtained from the freely accessible BioArt resource (https://bioart.niaid.nih.gov/).
Figure 3.
Two complementary strategies guide the discovery of antimicrobial peptides (AMPs): the identification of novel candidates from natural sources and the computationally guided design of synthetic AMPs. Protein images used in this figure were obtained from the freely accessible BioArt resource (https://bioart.niaid.nih.gov/).
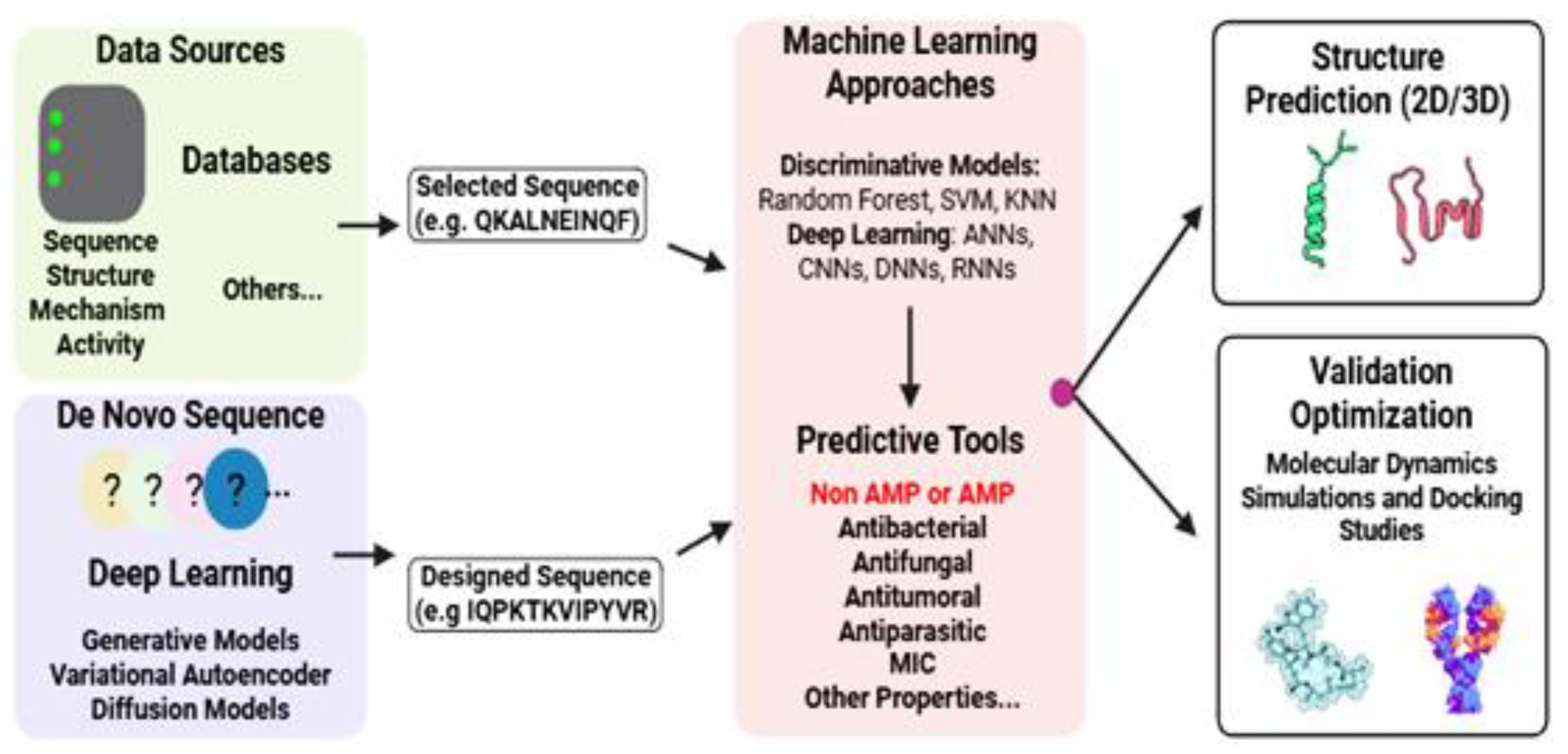

Table 1.
Comparative summary of the main characteristics of peptide production methods.
| Criterion | Chemical Synthesis | Enzymatic | Natural | Recombinant |
|---|---|---|---|---|
| Cost | High | Moderate | Variable | Low |
| Yield | Medium | Low | Very Low | High |
| Scalability | High | Moderate | Low | Very High |
| Technical Complexity | High | Low | Medium | High |
| Application Versatility | Very High | Moderate | Low | High |
Table 2.
AMPs approved for FDA and in market.
| Peptide Name | Sequence | Structure | Source | Biological Activity | Medical use | Target site | Delivery path | Company | Reference |
|---|---|---|---|---|---|---|---|---|---|
| Bacitracin | Leu-D-Glu Ile-Lys-D-Orn-Ile-D-Phe-His-D-Asp-Asn |  |
Bacillus licheniformis | Antibacterial | Prevent wound infections, pneumonia and empyema in infants, skin and eye infections. | Cell wall | Topical | Various companies | [95] |
| Dalbavancin Xydalba | Not available |  |
Semi synthetic |
Antibacterial | Acute bacterial skin infections, Osteomyelitis and septic arthritis | Cell wall | IV | DALVANCE® | [96] |
| Daptomycin Cubicin |
decanoyl-Trp-Asn-Asp-Thr-Gly- Orn-Asp-D-Ala-Asp-Gly-D-Ser-Glu(3R-Me)-Asp (Ph(2-NH2)) |  |
Streptomyces roseosporus | Antibacterial | Complicated skin infections (cSSSI) and bloodstream infections (bacteremia) | Cell membrane | IV | CUBICIN® | [97] |
| Enfuvirtide Fuzeon T20 |
YTSLIHSLIEESQNQQEKNEQELLELDKWASLWNWF |  |
Synthetic | Anti-HIV | Human Immunodeficiency Virus (HIV) Infections; AIDS | Fusion protein gp41 | SC | FUZEON® |
[98] |
| Gramicidin D | VGALAVVVWLW LWLW-ethanolamine |  |
Bacillus brevis | Antibacterial | Skin lesions, surface wounds and eye infections | Cell membrane | Topical | Various companies |
[99] |
| Gramicidin S | cyclo[Leu-D-Phe-Pro-Val-Orn-Leu-D-Phe-Pro-Val-Orn] | 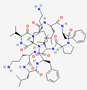 |
Bacillus brevis | Antibacterial | Against bacteria and fungi restricted use as spermicide and to treat genital ulcers | Cell membrane | Topical | Various companies | [100] |
| Obiltoxaximab | Not available | Not available | monoclonal antibody | Antibacterial | Treatment and prevention of inhalational anthrax | Antitoxin | IV | ANTHIM® | [101] |
| Oritavancin | Not available | Not available | Semi synthetic |
Antibacterial | Acute bacterial skin | Cell wall | IV | KIMYRSA™ ORBACTIV® | [102] |
| Palivizumab | Not available |  |
monoclonal antibody |
Antiviral | Prevent serious lung infections (such as pneumonia) that are caused by respiratory syncytial virus-RSV | Blocking viral replication | IM | SYNAGIS® | [103] |
| Polymyxin B | Not available |  |
Bacillus polymyxa | Antibacterial | Infections of the urinary tract, meninges, and blood stream | Cell membrane | IV | Various companies | [104] |
| Polymyxin E Colistin |
6-mh-DabTDab [γDablLDabDabT] |
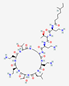 |
Bacillus polymyxa | Antibacterial | Acute or chronic infections due to gram-negative bacilli | Cell membrane | Oral | Various companies | [105] |
| Raxibacumab | Not available | Not available | monoclonal antibody | Antibacterial | Treatment and prevention of inhalational anthrax | Antitoxin | IV | RAXIBACUMAB ® | [106] |
| Telavancin TD-6424 |
Not available | 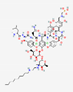 |
Semisynthetic | Antibacterial | Osteomyelitis and bacterial infections | Cell membrane | IV | VIBATIV® | [107] |
| Tyrothricin | Not available | 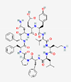 |
Brevibacillus parabrevis | Antibacterial Antifungal | Infected skin and infected oropharyngeal mucous membranes | Cell membrane | Topical/ Oral |
Various companies | [108,109] |
| Thymalfasin | Ac-Ser-Asp-Ala-Ala-Val-Asp-Thr-Ser-Ser-Glu-Ile-Thr-Thr-Lys-Asp-Leu-Lys-Glu-Lys-Lys-Glu-Val-Val-Glu-Glu-Ala-Glu-Asn-OH | Not available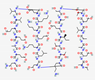
|
Synthetic | Antiviral | Hepatitis B and C. Boost the immune response in the treatment of other diseases. |
Immuno modulator |
IV | ZADAXIN® | [110] |
| Vancomycin | Not available | 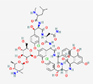 |
Amycolatopsis orientalis | Antibacterial | Septicemia, infective endocarditis, skin, bone and lower respiratory tract infections. | Cell wall | IV/Oral | Various companies | [111] |
* Intravenous injections (IV), Intramuscular injections (SC) Intramuscular injections (IM). All information was collected and reviewed in the DRAMP database (http://dramp.cpu-bioinfor.org/) [15], Drugbank (https://go.drugbank.com/) [112] and Pubchem (https://pubchem.ncbi.nlm.nih.gov/) [113] (accessed August 3, 2025).
Table 3.
AMPs tools for analysis.
| Database Name | Functionality | Additional Information | URL | Reference |
|---|---|---|---|---|
| ACEP | Identification of AMPs. | The classification method is based on deep learning (DL). | https://github.com/Fuhaoyi/ACEP | [145] |
| ADAPTABLE | Designing novel peptides, predicting their activities, and identifying functional motifs. | Webserver and data-miner of antimicrobial peptides. | http://gec.upicardie.fr/adaptable/ | [146] |
| AFP-MFL | A novel deep learning model that can predict antifungal peptides. | It only needs the peptide sequence to run. | https://inner.weigroup.net/AFPMFL/#/ | [147] |
| AlphaFold | To predict a protein/peptide’s 3D structure based on amino acid sequence. | AI system developed by Google DeepMind. | https://colab.research.google.com/github/sokrypton/ColabFold/blob/main/AlphaFold2.ipynb | [148,149] |
| AntiBP 3.0 | A web-tool for predicting, scanning and designing AMPs. | Based on machine learning techniques. | https://webs.iiitd.edu.in/raghava/antibp3/ | [150] |
| AMP- Scanner | A web server tool for predicting if it is an AMP based on amino acid sequence. | Only includes bacteria targets. | https://www.dveltri.com/ascan/ | [151] |
| AMPA | A web server tool for identifying active regions in antimicrobial proteins. | The algorithm uses an antimicrobial propensity scale to generate an antimicrobial profile. | https://tcoffee.crg.eu/apps/ampa/do | [152,153] |
| AMPDeep | Deep learning approach to predict hemolytic activity of AMPs. | It was built on Python. | https://github.com/milad73s/AMPDeep | [154] |
| amPEPpy | A python application for predicting antimicrobial peptide sequences. | The classification method is based on random forest. | https://github.com/tlawrence3/amPEPpy | [155] |
| AmpGram | A web server tool for identification of AMPs. | The classification method is based on random forest and n-gram analysis. | http://biongram.biotech.uni.wroc.pl /AmpGram/ | [156] |
| AutoDock | A tool to predict how small molecules bind to a receptor of know 3D structure. | It has two options: AutoDock-GPU and AutoDock Vina. | https://autodock.scripps.edu/ | [139] |
| AxPEP | It is a collection of sequence-based machine learning methods for AMPs prediction. | The classification method is based on random forest. | https://app.cbbio.online/ampep/home | [157] |
| CABS-dock | A web server tool for flexible protein-peptide docking. | It requires the sequence of a protein receptor and a peptide sequence. | https://biocomp.chem.uw.edu.pl/CABSdock | [141] |
| CHARMM36 | Broad-scope molecular simulation software for complex environments. | There is no cost to academic students. | https://academiccharmm.org/ | [143] |
| CS-AMPPred | An SVM-based (support vector machines) tool to predict antimicrobial activity in cysteine-knotted proteins. | Were based on 310 AMPs and 310 non-antimicrobial peptide sequences. | https://sourceforge.net/projects/csamppred/ | [158] |
| Cysmotif Searcher | A Perl package for revealing peptide sequences possessing cysteine motifs. | Cysteine motifs are common to various families of AMPs and others cysteine-rich peptides. | https://github.com/fallandar/cysmotifsearcher | [159] |
| deepAMP | A tool for predicting protein AMPylation sites from binary profile representation. | The classification method is based on a convolutional neural network. | https://github.com/MehediAzim/DeepAmp | [160] |
| GROMACS | Powerful open-source suite for molecular dynamics simulation and analysis. | Broad spectrum of calculation types, preparation and analysis tools. | https://manual.gromacs.org/archive/4.6.7/online/speptide.html | [142] |
| HADDOCK | A web platform for biomolecular docking simulations. | A data-driven docking approach guided by ambiguous interaction restraints (AIRs). | https://rascar.science.uu.nl/haddock2.4/ | [140] |
| iAMPCN | DL approach to identifying antimicrobial peptides and their functional activities. | It was built on Python. | https://github.com/joy50706/iAMPCN | [161] |
| iTASSER | DL approach to identifying antimicrobial peptides and their functional activities. | It was built on Python. | https://zhanggroup.org/I-TASSER/ | [162] |
| Joker | A web tool to predict structure of a protein. | It also predicts the function. | https://github.com/williamfp7/Joker | [163] |
| MARTINI | Coarse-grained force field for biomolecular simulations, parameterized using experimental data and atomistic simulations. | Based on a four-to-one mapping scheme. | https://cgmartini.nl/ | [144] |
| MLBP | Multi-label DL approach to identifying multi-functional bioactive peptide functions. | It was built on Python. | https://github.com/tangwending/MLBP | [164] |
| ModlAMP | A Python package for working with any sequence of natural amino acids. | It includes the following modules: sequence generation, sequence library analysis and description calculation. | https://modlamp.org/ | [165] |
| NAMD | A highly scalable software for parallel molecular dynamics simulations of large biomolecules. | It uses the program VHD for simulation setup and trajectory analysis. | https://www.ks.uiuc.edu/Research/namd/ | [166] |
| PEP2D | A web tool for predicting secondary structure of peptides. | The model was trained and tested based on 3100 peptide structures. | https://webs.iiitd.edu.in/raghava/pep2d/ | [167] |
| PEP-FOLD 4 | A de novo approach to predict peptide structure from amino acid sequences. | The peptides should have 5 to 50 amino acids. | https://bioserv.rpbs.univparisdiderot.fr/services/PEP-FOLD4/ #overview | [39] |
| RoseTTAFold | A Python-based AI tool for protein/peptide structure prediction. | The structure prediction method is based on DL. | https://github.com/RosettaCommons/RoseTTAFold?tab=readme-ov-file | [137] |
| sAMPpred-GAT | A web tool for identification of AMPs. | The program uses graphs constructed based on predicted peptide structures. | http://bliulab.net/sAMPpred-GAT/server | [168] |
| SWISS-MODEL | A web tool for protein structure prediction and modeling. | It supports interactive modeling for both simple and complex needs. | wissmodel.expasy.org | [169] |
| VMD | A program to display, animate and analyze large biomolecules systems. | It has 3-D graphics | https://www.ks.uiuc.edu/Research/vmd/ | [169] |
* All information was collected and reviewed on August 17.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



