Submitted:
30 September 2025
Posted:
01 October 2025
You are already at the latest version
Abstract
The evolving needs of society, shaped by lifestyle changes and growing health awareness, have intensified interest in sustainable foods with enhanced nutritional benefits. Berry crops are central to this trend, serving as rich dietary sources of bioactive compounds that support human health. Renowned for their high levels of diverse antioxidants, they offer benefits that extend well beyond basic nutrition. This review explores the diversity and global cultivation of major berry crops, including blueberries, raspberries, cranberries, blackberries, and grapes, with emphasis on their nutritional value and antioxidant profiles. It also examines their domestication history, wild relatives, and commercial cultivars, while offering insight into the genetic and phenotypic diversity underpinning their rich chemical composition. Furthermore, the review highlights the use of modern genomic and breeding tools to enhance bioactive compound content. Advances in cultivation and molecular breeding technologies, including transcriptomics, genome-wide association studies (GWAS), and genome editing, offer promising opportunities to maximize the antioxidant potential of these high-value crops, particularly in the face of a changing climate.
Keywords:
berry crops
; grapevine
; antioxidants
; nutritional value
; breeding
; genomic tools
1. Introduction
1.1. A multitude of Berry Crops
Berry crops, grapes among them, include a diverse group of fruits that differ botanically yet share similar cultivation practices, postharvest behavior, and health-promoting properties [1](Kumar et al., 2018). A true berry is a simple fruit that develops from a single ovary and contains multiple seeds within a fleshy pericarp, such as blueberries (Vaccinium spp.), cranberries (Vaccinium macrocarpon), and grapes (Vitis vinifera) [2]. In contrast, fruits such as raspberries (Rubus idaeus) and blackberries (Rubus fruticosus), though commonly referred to as berries, are in fact aggregate fruits, consisting of numerous small drupelets that originate from a single flower with multiple ovaries [3]. Despite this structural difference, all these fruits are commonly grouped under “berry crops” due to their overlapping agricultural, nutritional, and commercial significance [4].
Among major cultivated berries, raspberries (Rubus idaeus), native to Europe and northern Asia, yield about 940,000 tons globally, with Russia, Mexico, Serbia, Poland, and the United States as leading producers. Blueberries (Vaccinium spp.), native to North America, total about 1.1 million tons, dominated by the United States, Peru, Canada, Chile, and Spain. Cranberries (Vaccinium macrocarpon), also of North American origin, produce around 470,000 tons, largely from the United States, Canada, and Chile. In contrast, grapes (Vitis vinifera), native to the Near East, far exceed berry crops with over 72.5 million tons, produced mainly in China, Italy, Spain, the United States, and France [5] (Table 1).
Berry cultivation reflects a combination of historical domestication patterns, ecological adaptations, and modern agricultural practices. Raspberries, blackberries, blueberries, and cranberries are primarily associated with temperate or specialized agro-ecological zones, whereas grapes represent one of the most extensively cultivated fruit crops worldwide. A comparative overview of their production statistics, major producing countries, and key soil–climate requirements is presented in the following table [5].
1.2. Nutritional and Antioxidant Value of Berries as Pharmaceutical and Nutraceutical Sources
For centuries, fruits such as berries have been valued in traditional medicine and human diets for their health-promoting properties. Despite limited scientific understanding, their historical use was based on empirical knowledge; however, advances in phytochemistry, particularly through metabolomics, have revealed the complex mixture of bioactive compounds responsible for their beneficial effects on human health [6].
Berries are an abundant source of antioxidants, particularly flavonoids. Among these, flavonols such as quercetin and kaempferol present in raspberries, cranberries, and grapes have been reported to significantly contribute to their antioxidant capacity and associated health benefits [7]. Quercetin, present in raspberries and cranberries, demonstrates significant anti-inflammatory, antiviral, and antihistamine activities and has been studied as an supportive therapy for arthritis, allergies, and viral infections [8,9,10]. Anthocyanins, like cyanidin-3-glucoside (C3G), malvidin-3-glucoside (M3G), peonidin-3-glucoside (P3G), and delphinidin-3-glucoside (D3G) are highly concentrated in blueberries, blackberries, and red or purple grapes, where they are primarily responsible for the characteristic pigmentation [8]. Beyond their coloring function, anthocyanins have been extensively studied for neuroprotective and cardiovascular effects, including potential roles in mitigating neurodegenerative diseases such as Alzheimer’s and Parkinson’s, improving endothelial function, and reducing blood pressure [11,12]. In addition, grapes and cranberries are rich sources of flavan-3-ols, particularly catechin and epicatechin, which are concentrated in the seeds [13]. The tannin group is also well represented in berry crops, with hydrolysable tannins such as ellagic acid detected in abundance [14,15]. These compounds have been associated with cardiovascular protection, antimicrobial and antiviral activities, anti-inflammatory effects, and potential roles in metabolic health and cancer prevention [16]. In addition, condensed tannins, particularly procyanidins, are predominantly found in grape seeds, cranberries, and blackberries, further contributing to the bioactive profile of these fruits [17]. Stilbenoids such as resveratrol and pterostilbene are primarily concentrated in red and purple grapes. Resveratrol and related compounds are multifunctional polyphenols with potent antioxidant and anti-inflammatory properties, acting through the reduction of oxidative stress and modulation of inflammatory pathways, and they have been widely associated with cardioprotective effects [18]. Proanthocyanidins, particularly A-type forms, which are abundant in cranberries, are known for inhibiting bacterial adhesion in urinary tract infections and increasingly investigated for their roles in gut microbiome modulation and vascular health. [19]
Phenolic acids such as chlorogenic, caffeic, and gallic acids are widely distributed in blueberries, grapes, and cranberries [20]. These compounds have been reported to modulate glucose metabolism, enhance insulin sensitivity, and display antiviral activity, thereby playing an important role in the management of metabolic disorders such as diabetes and obesity [21]. Minor bioactive compounds in berries include vitamins C and E, carotenoids such as β-carotene and lutein, and terpenes including limonene, myrcene, linalool, and pinene. These compounds contribute to antioxidant capacity and characteristic aromas, and have been shown to enhance cellular antioxidant defenses, neutralize free radicals, and suppress inflammatory gene expression, thereby protecting against oxidative stress, inflammation, and cellular damage [22]. All the above-mentioned compounds form a robust biochemical platform that is increasingly leveraged in therapeutic and preventive healthcare applications.
Furthermore, micronutrients, such as calcium, magnesium, and iron present in varying amounts across berries play crucial roles in antioxidant defense mechanisms and metabolic homeostasis [23]. Calcium and magnesium contribute to proper enzymatic function and cellular signaling relevant to oxidative stress modulation [24,25] while iron, although required in small amounts, is vital for oxygen transport and redox reactions, and interacts with polyphenols and vitamin C absorption [26]. This synergy between these antioxidants, vitamins, and essential nutrients makes berries a highly nutritious choice for anyone looking to boost their health, reduce inflammation, and protect against chronic diseases [27,28].
Based on the recognized nutritional and pharmaceutical value of the antioxidants of berries, current research is directed toward tools that can increase their accumulation in the fruits. In this context, agronomical techniques, such as priming and grafting, can stimulate antioxidant pathways by improving plant resilience under stress conditions. At the same time, molecular breeding approaches, including marker-assisted selection, QTL mapping, and genome editing, offer targeted opportunities to enhance antioxidant biosynthesis. The integration of these strategies provides a comprehensive framework that connects crop improvement with the development of berry fruits enriched in antioxidants and enhanced health benefits (Figure 1).
2. Agronomical Practices for Increasing Antioxidant Concentration
2.1. Priming
Environmental changes have intensified abiotic stress in plants, reducing yield and threatening food security [29]. One adaptive mechanism that improves plant resilience is priming, a process that prepares plants to respond more efficiently to future stresses [30].
Priming, also referred to as sensitization or hardening, induces a “primed state” (PS) that enhances the plant’s ability to activate defense mechanisms when challenged again [31] This state is triggered by mild pre-exposure to stress or by the application of natural or synthetic agents. Biochemical mechanisms underlying PS include the accumulation of inactive signaling proteins or transcription factors, activation of mitogen-activated protein kinases (MAPKs), changes in redox status, epigenetic modifications, and breakdown of certain cellular compounds [32]. Once primed, plants exhibit enhanced redox balance and faster stress responses, improving resilience and growth [33].
Notably, priming strategies have been shown to enhance the antioxidant capacity of berry crops, thereby improving both stress resilience and nutritional quality. For instance, in blueberries, foliar application of polyamines such as putrescine and spermidine significantly increased antioxidant potential, with the fruit development stage identified as the most favorable period for treatment. Such findings provide valuable guidance for growers to optimize fertilization practices during critical phases of berry growth and maturation [34]. Likewise, in blackberries, priming with Bacillus amyloliquefaciens has been used to investigate the role of the flavonol–anthocyanin pathway in the adaptation of plants to biotic stress. This treatment modulated blackberry metabolism and identified key target genes, providing insights for improving plant adaptability and fruit quality [35]. Additionally, a biostimulant derived from Ascophyllum nodosum, applied as a foliar spray to raspberry, was found to enhance yield by over 30% while maintaining most nutritional traits. Notably, magnesium content increased in primed fruits, highlighting its potential as a sustainable tool to improve berry yield and quality [36]. Similarly, pre-harvest foliar treatments with ethephon (Eth) at the ‘veraison stage’ resulted in increased levels of total phenolics and anthocyanin and enhanced antioxidant capacity accompanied by an increased expression of the anthocyanin biosynthesis genes, such as chalcone synthase (CHS), flavanone 3-hydroxylase (F3H), and flavanol 3-o-glucosyl transferase (UFGT) in ‘Crimson Seedless’ grape berry skins [37]. Moreover, postharvest treatment with melatonin reduced grape berry abscission and decay, enhanced amino acid accumulation, and activated endogenous melatonin and phenolic biosynthesis pathways, highlighting the potential of improving postharvest quality through the modulation of secondary metabolism [38].
2.2. Grafting
Grafting, a technique that is widely used in perennial crops, has been adopted more in berry species such as raspberry and blueberry with the aim of improving plant’s stress tolerance and fruit quality [39]. By combining the root system of a robust rootstock with the scion of a high-yielding or high-quality cultivar, growers can modify physiological traits in a manner that is not possible through conventional breeding. Even though grafting is more commonly linked to disease resistance and soil adaptability, recent studies indicate it may also affect the antioxidant capacity of berry fruits. Changes in nutrient uptake, water-use efficiency, and hormonal signaling can result from physiological interactions between the rootstock and scion. These changes are closely related to the fruit’s generation of secondary metabolites [40].
The accumulation of phenolic compounds, anthocyanins, and flavonoids can be indirectly enhanced by specific rootstocks, according to evidence from grafted raspberry and blueberry systems [39]. Increased photosynthetic activity, a more stable water balance during drought, and improved root-to-shoot nutrient transport, particularly minerals like magnesium and zinc that are essential in antioxidant enzyme function, are frequently linked to these gains [41].
Specifically for grapes, grafting has notably improved antioxidant capacity by adjusting the scion’s physiological and biochemical reactions through rootstock selection. According to the study of Krishankumar et al. (2025), the use of drought-tolerant rootstocks, like Ramsey, R110 and 1103 Paulsen, positively resulted to a significant increase of important antioxidant enzyme activities, including glutathione reductase (GR), ascorbate peroxidase (APX), peroxidase (POD), catalase (CAT), and superoxide dismutase (SOD), under drought stress [42]. Furthermore, Klimek et al. (2024) showed that different rootstocks significantly affected the content of bioactive compounds in Regent grapes [43]. The DPPH radical scavenging measurement in grafted grapes revealed variations in L-ascorbic acid, phenolic acids, total flavonoids, anthocyanins, and tannins, all of which are directly related to antioxidant activity. Together, these findings prove that choosing the right rootstocks greatly increases the antioxidant profile and nutritional value of grapes in addition to increasing their resistance to abiotic stress.
3. Berry Genomics and Breeding for Enhancing Antioxidant Capacity
According to the literature, breeding programs targeting improved tolerance of berries to abiotic stresses such as extreme temperatures, drought, salinity, and iron deficiency have also contributed significantly to enhancing their antioxidant capacity [44]. Building on these health-promoting characteristics, improving the antioxidant content is becoming a central breeding target, guiding current efforts to combine nutritional quality with agronomic performance [45,46]. To achieve these goals, modern breeding strategies integrate advanced genomics and molecular tools [47]. Until now, conventional breeding has been carried out mainly through time-consuming, controlled crossings between selected parents, followed by long-term field evaluations of berry seedlings for traits such as yield, vigor, disease resistance, and fruit quality. Then promising genotypes are clonally propagated to establish new cultivars [47,48].
At the genomic level, high-quality reference genomes have been assembled for several berry crops, facilitating the discovery of gene families involved in secondary metabolite biosynthesis [49,50]. Multi-omic studies have elucidated metabolic pathways and the expression dynamics of regulating genes in response to both developmental and environmental stimuli. Integrated transcriptomic and metabolic profiling reveals that phenolic acid and flavonol biosynthesis in Rubus chingii Hu (Chinese raspberry) begins at the onset of fruit development. Afterwards, these compounds are systematically accumulated or metabolized into related derivatives [51]. The exploitation of pathways such as those for the phenylpropanoid or the flavonoid biosynthesis are essential to antioxidant production and are tightly coupled with responses to abiotic stresses in berry crops like blackberry [35].
Moreover, Genome Wide Association Studies (GWAS) have improved trait mapping in berries through investigations of the heritability of quality traits and their associations with known QTLs [52]. At the same time, berry breeding increasingly integrates conventional methods with modern molecular approaches, and CRISPR/Cas9-mediated gene editing has already been validated in blueberry, paving the way for precise trait improvement, including antioxidant enhancement without yield penalties [53]. However, GWAS and other genomic tools like CRISPR/Cas9, remain largely unexplored for the exploration and utilization of antioxidant capacity in raspberry breeding.
3.1. Blueberry (Vaccinium corymbosum)
3.1.1. Domestication and Wild Relatives
Blueberry is a widely cultivated native fruit in North America [54]. Domestication began in the early 20th century through extensive interspecific hybridization programs [55]. The genus Vaccinium evolved in eastern North America, with biogeographic origins centered in the Appalachian Mountains and Atlantic coastal plains, where natural populations exhibit maximum genetic diversity [56]. Evolutionary relationships within section Cyanococcus reveal three main lineages: (1) northern species (V. corymbosum, V. angustifolium), (2) southern species (V. darrowii, V. tenellum, V. virgatum), and (3) western species (V. ovalifolium, V. deliciosum) [57]. Polyploidy evolution is prominent, with diploid (2n = 2x = 24), tetraploid (2n = 4x = 48), and hexaploid (2n = 6x = 72) cytotypes, where allopolyploidy has driven speciation and adaptive radiation across diverse ecological niches [58,59]. Cultivated highbush blueberry (V. corymbosum) is tetraploid (2n = 4x = 48), with a monoploid genome size of ~2.53 pg [58,60]. Other cultivated groups include lowbush (V. angustifolium, diploid) and rabbiteye (V. virgatum, hexaploid), with extensive interspecific hybridization among them [61].
Wild blueberry species harbor broad genetic diversity with desirable traits that can be exploited in breeding programs. While most studies on their genetic variability do not directly assess antioxidant capacity, they often provide indirect insights, such as the presence and stress-induced accumulation of metabolites and flavonoids, that are increasingly targeted to enhance the antioxidant potential of cultivated blueberries [62]. More specifically, wild Vaccinium species have been recognized as tolerant genetic material with high nutritional value against various abiotic and biotic stressors, such as low chilling requirements in V. darrowii [63], and heat and drought tolerance in V. myrsinites and V. darrowii [64]. Traits like fruit firmness, late ripening, and disease resistance are well-documented in V. virgatum [55]. Moreover, diploid southern species such as V. tenellum, V. elliottii, and V. myrsinites contribute specialized metabolites and stress tolerance [64]. Furthermore, V. angustifolium (Lowbush, 2n=24), a wild-managed species native to northeastern North America, has been recognized as a source of cold hardiness and unique flavonoids [65]. High-quality genome assemblies are now available for V. darrowii (~1.06 Gb, >34,000 protein-coding genes) and V. corymbosum (‘W8520’ assembly, ~393 Mb), providing essential resources for elucidating the genetic basis of antioxidant capacity in berries and for uncovering the molecular mechanisms underlying its biosynthesis [63,66].
Blueberry commercial cultivation focuses on three main groups with distinct antioxidant profiles: a) Northern Highbush (V. corymbosum) cultivars dominate global production and include early-season genotypes such as ‘Duke’ and ‘Rubel’ known for high anthocyanin content and excellent antioxidant capacity [67], and mid-season standards like ‘Bluecrop’and ‘Frienship’, recognized for a balanced anthocyanin profile and high phenolic diversity [68]. Late-season cultivars such as ‘Elliott’, are distinguished by elevated proanthocyanidin content and extended shelf-life due to antioxidant stability, high total phenolic content and superior ORAC values [69,70]. According to Stevenson & Scalzo (2012), several Northern Highbush cultivars such as ‘Rubel’, ‘Duke’, and ‘Elliott’ consistently exhibit elevated anthocyanin and total phenolic content, reinforcing their value as benchmark genotypes for antioxidant potential. b) Southern Highbush (V. corymbosum × V. darrowii) hybrids include ‘Jewel’, a mid-season genotype with elevated flavonol content and enhanced UV protection compounds [55], ‘Legacy’ a mid- to late genotype with high total anthocyanins and phenolics and ‘Emerald’, which features a low-chill requirement and unique anthocyanin glycosylation patterns introgressed from V. darrowii [69]. c) Rabbiteye (V. virgatum) includes early and late-season harvests cultivars, like ‘Tifblue’ which contribute further diversity, serving as industry standards due to its high tannin content and antioxidant stability [55,70,71]. Late-ripening cultivars such as ‘Velluto Blue’ and ‘Centra Blue’ exhibit the highest fruit yields, anthocyanin contents, and estimated total anthocyanin harvestable from a given area [69]. Additionally, Rabbiteye cultivars, such as ‘Ono’, ‘Centurion’, and ‘Southland’, have been highlighted for their superior antioxidant richness, often surpassing both highbush groups in anthocyanin and phenolic content [72]. All the above-mentioned cultivars represent promising candidates for breeding programs focused on phytochemical enhancement regarding antioxidant properties.
3.1.2. Modern Genomic and Breeding Tools for Enhancing the Antioxidant Profile of Blueberries
Several genes and gene families have been studied in relation to the antioxidant capacity of blueberries, and their roles have been linked to genotypes with desirable traits and high nutritional value. For instance, the MATE transporter gene family, which encodes transport proteins that facilitate the movement of anthocyanins across cell membranes during fruit ripening, has been characterized as V. corymbosum [73]. Additionally, genes involved in the flavonoid biosynthetic pathway include those encoding CHS, chalcone isomerase (CHI), F3H, flavonoid 3′-hydroxylase (F3′H), flavonoid 3′,5′-hydroxylase (F3′5′H), dihydroflavonol 4-reductase (DFR), anthocyanidin synthase (ANS), (UFGT), and leucoanthocyanidin reductase (LAR), are reported to be upregulated during fruit ripening and correlate with anthocyanin accumulation [74,75]. Notably, VcGSTF genes show high expression during ripening and are implicated in anthocyanin stabilization and vacuolar transport [74]. In research by Liu et al. (2021), a genome wide analysis of the Caffeic Acid O-Methyltransferase (COMT) gene family was conducted in V. corymbosum, identifying 92 COMT genes involved in phenylpropanoid metabolism [75]. Moreover, several gene families encoding enzymes involved in the phenylpropanoid pathway, such as phenylalanine ammonia lyase (PAL), anthocyanidin reductase (ANR), cinnamate-4-hydroxylase (C4H), 4-coumarate:CoA ligase (4CL), flavonol synthase (FLS), hydroxycinnamoyl-CoA:shikimate hydroxycinnamoyl transferase (HCT), have been identified in blueberry under stress conditions and are associated with antioxidant responses [76,77]. Terpene synthases (TPSs) have also been characterized in blueberries, contributing to volatile profiles and secondary metabolite diversity relevant to antioxidant potential [78]. Table 2 summarizes the genes and their corresponding enzyme classes investigated in blueberries with respect to antioxidant capacity. Despite the availability of high-quality genome assemblies, the precise roles, annotations, and functional characterizations of numerous genes in blueberry remain unresolved and continue to be active areas of research.
Advanced omic approaches such as transcriptome sequencing (RNA-seq) offer powerful insights into antioxidant-related genes, their expression dynamics, and their contribution to key metabolic traits in berry crops. RNA-seq analysis of 196 F1 genotypes, developed from the crosses of the cultivars ‘Draper-selection-44392” and ‘Jewel’, differing in anthocyanin glycosylation and acylation identified several differentially expressed genes (DEGs). For the major-effect QTL on chromosome 4 controlling glycosylation, three strong candidate genes were detected, including the UDP-glucosyltransferase (Vcev1_p0.Chr04.11988) and two endotransglycosylation (Vcev1_p0.Chr04.11996 and Vcev1_p0.Chr04.11998). Among them, Vcev1_p0.Chr04.11988 was significantly up-regulated in high-glycosylated genotypes and validated by qRT-PCR, supporting its role as a key regulator of anthocyanin glycosylation. For acylation, a major locus on chromosome 2 contained two acyltransferase genes (Vcev1_p0.Chr02.03371 and Vcev1_p0.Chr02.03383), both up-regulated in high-acylated genotypes [81].
The chromosome-scale, haplotype-resolved assembly of tetraploid highbush blueberry (Draper’) provided a framework for annotating candidate genes, including UGTs, acyltransferases, and HCT, many colocalizing with antioxidant QTLs [66]. Multi-year QTL mapping in a V. corymbosum var. caesariense × V. darrowii population identified a major chlorogenic acid locus on Vc02 overlapping HCT/UGCT, and 67 QTLs for anthocyanins across Vc01–Vc12 explaining 11–67% of variance. Major hotspots included 34 QTLs on Vc04 (~115 cM) linked to glucoside-enriched anthocyanins and 15 QTLs on Vc01 (18–35 cM) linked to reduced arabinose/galactose conjugates, with additional loci on Vc02 and Vc08–Vc12 [82]. GWAS with an 89K SNP array identified 47 SNP–trait associations for flavonoid metabolites, revealing novel biosynthetic genes and regulatory networks shaping antioxidant diversity [81]. These studies highlight the polygenic control of antioxidant metabolism in polyploid blueberry.
3.2. Red Raspberry (Rubus idaeus L.)
3.2.1. Domestication and Wild Relatives
Red raspberry, a member of the Rosoideae subfamily (Rosaceae), comprises over 740 described species worldwide, with two recognized diploid subspecies: the European R. idaeus subsp. idaeus and the North American R. idaeus subsp. strigosus [83,84]. Cultivated raspberries are diploid (2n = 2x = 14), with domestication involving both subspecies across more than 2,000 years of cultivation in Europe and approximately 150 years in North America [85,86]. Evolutionary origins trace to Eurasian forests, with natural distribution extending from Western Europe through Siberia to eastern Asia, and separate colonization of North America during Pleistocene glacial periods [87]. Phylogenetic analysis using chloroplast genome sequences places raspberry within subgenus Idaeobatus, with sister relationships to R. corchorifolius and close affinity to Korean raspberry (R. crataegifolius) [88]. Biogeographic reconstruction indicates Eurasia as the center of diversification.
Wild relatives and inter-subspecific hybrids (R. idaeus × R. strigosus) are maintained in germplasm banks and serve as crucial donors of antioxidant-related traits, with genomic analyses confirming synteny across the seven chromosomes [84,86,89]. Species diversity within raspberry germplasm includes multiple interfertile taxa. The R. idaeus complex comprises the European subspecies idaeus (2n=2x = 14), a source of high sugar content, and diverse flavor profiles [90], and the North American subspecies R. strigosus (2n =2x = 14), which contributes cold hardiness, disease resistance, and unique anthocyanin profiles [86,91]. Related species provide specialized traits: R. occidentalis (black raspberry, 2n=2x = 14) is notable for exceptional anthocyanin content and unique cyanidin-3-rutinoside accumulation [92]. Wild population diversity studies using SSR and SRAP markers reveal significant population structure across natural ranges, with diversity centers in the Caucasus region (European populations) and the Pacific Northwest (North American populations), both serving as priority collection sites [87,93]. Germplasm repositories maintain over 1,500 accessions globally, with core collections at USDA Corvallis (~450 accessions), Agriculture Canada (~320 accessions), and major European collections, representing maximum allelic diversity for breeding applications [84].
Commercial raspberry production focuses on antioxidant-rich cultivars such as ‘Haida’, ‘Chilli wack’ and ‘Meeker’, spanning different ripening seasons. European-derived cultivars are reported with several important commercial traits such as high anthocyanin content, excellent antioxidant stability during storage [94], elevated ellagitannin levels and superior ORAC values [95]. Several early-season cultivars also contain high total phenolic content and spine-free canes that facilitate harvest [96]. North American selections include genotypes with cold-hardy characteristics notable for unique anthocyanin glycosylation patterns and high vitamins, as well as others which combine disease resistance with elevated flavonol content and extended shelf-life due to antioxidant stability [94]. Interspecific hybrids (R. idaeus × R. occidentalis) contribute to genetic diversity, providing genotypes with exceptional anthocyanin profiles (including cyanidin and pelargonidin derivatives) and dual-purpose potential for both fresh consumption and processing [92]. Beyond established cultivars, advanced breeding lines specifically selected for enhanced antioxidant profiles exhibit 40–60% higher total phenolic content than standard cultivars, with specialized metabolite compositions tailored for functional food applications [95,97]. Notably, cultivars such as ‘Saanich’, ‘KiwiGold’, and ‘Summit’ have demonstrated superior antioxidant activity and mineral content, with ‘Saanich’ showing exceptional iron and manganese levels and ‘KiwiGold’ ranking among the highest in potassium and phenolics [45]. Additionally, non-commercial cultivars like ‘Adelita’, ‘Glen Dee’, ‘Himbo Top’, ‘San Rafael’, and ‘Tula Magic’ have shown promising antioxidant potential and polyphenol content, suggesting their suitability for future breeding programs targeting nutritional enhancement [98]. Recent evaluations of cultivars grown in northern Mexico further highlight ‘Red Autumn Bliss’ for its high anthocyanin concentration and antioxidant capacity, and ‘Yellow Autumn Bliss’ for its exceptional levels of total phenolics, ellagic acid, ascorbic acid, and polyphenolic summation, reinforcing their value in antioxidant-focused breeding strategies [99].
3.2.2. Modern Genomic and Breeding Tools for Enhancing the Antioxidant Character of Raspberry
Raspberry antioxidant quality is regulated by constitutive genes that sustain baseline metabolite production during fruit development. Three polyketide synthase (PKS) genes (PKS1, PKS2, and PKS3) have been characterized from raspberry cell cultures, with RiPKS1 encoding a typical chalcone synthase (CHS) active in naringenin production, RiPKS3 producing mainly p-coumaroyltriacetic acid lactone (CTAL), RiPKS2, an inactive form, and a newly identified, PKS4, with a crucial role in enhancing ketone levels in raspberry leaves and fruit [100,101]. Another study investigated antioxidant-related gene expression during fruit development in yellow, red, and black raspberries, revealing cultivar- and stage-specific patterns. Genes encoding antioxidant enzymes (SOD, POD, Peroxiredoxin-PRDX, CAT) and flavonoid/anthocyanin biosynthesis enzymes (PAL, FLS, ANR, LAR, UFGT) were found differentially expressed, with expression generally peaking at ripening stages. Transcription factor genes belonging to the basic Helix–Loop–Helix (bHLH), myeloblastosis (MYB), and WRKY families also displayed distinct expression profiles, demonstrating their regulatory role in antioxidant metabolism, [102]. Baseline phenolic acid biosynthesis relies on phenylpropanoid pathway genes such as PAL, C4H, and 4CL, which have been characterized in raspberry [51,103]. All the above-mentioned studies provide a valuable reference for understanding the molecular basis of antioxidant regulation in raspberry fruit. However, only a limited number of these genes have been functionally annotated and characterized (Table 3), and further research is required to exploit their roles in enhancing the antioxidant capacity of raspberries.
Genomic resources for raspberry are rapidly expanding and include a chromosome-scale diploid reference genome (~240 Mb, 2n = 2x = 14) assembled using PacBio long reads and Hi-C scaffolding, comprising 32,831 predicted genes with comprehensive functional annotation [89]. The genome architecture reveals seven linkage groups with high synteny to Fragaria and other Rosaceae species. Comparative genomics highlights gene family expansions in secondary metabolite biosynthesis, particularly phenylpropanoid-related genes—as well as disease resistance genes. Prominent tandem gene clusters encode flavonoid biosynthetic enzymes, terpene synthases, and glycosyltransferases. QTL mapping and SNP-based studies have identified loci associated with anthocyanin composition and antioxidant traits [104,105]. A review study by McCallum et al., (2018), in raspberry further reported QTLs directly associated with anthocyanin traits, antioxidant capacity, and phenolic content [106]. These include QTLs for anthocyanin composition, such as specific glycosides of cyanidin and pelargonidin, and the mapping of candidate genes underlying anthocyanin/phenolic biosynthesis (structural and regulatory genes/transcription factors such as bHLH, MYB, and basic-leucine zipper transcription factor-bZIP), which often colocalize with QTLs for pigmentation and anthocyanin-related traits. Investigation of gene relative expression as well as transcriptome resources provide datasets related to antioxidant characteristics at diverse developmental stages, tissues, and environmental conditions, supporting functional genomics and breeding applications [89,102].
3.3. Blackberry (Rubus occidentalis/Rubus fruticosus agg.)
3.3.1. Domestication and Wild Relatives
Blackberry (Rubus subgenus Rubus) represents a taxonomically and genetically complex crop group characterized by polyploidy, hybridization, and broad ecological adaptation. The cultivation history spans more than 150 years, though systematic breeding only intensified from the 1960s onward [107,108]. Evolutionary origins are geographically diverse: European species such as the R. fruticosus aggregate contributed to Old World germplasm, while R. allegheniensis and R. argutus formed the basis of North American breeding programs [84]. The species exhibit remarkable cytogenetic diversity, with chromosome numbers ranging from diploid (2x = 14) to dodecaploid (12x = 84), though most commercial cultivars are tetraploid (2n = 4x = 28) or hexaploid (2n = 6x = 42) [108]. Domestication has relied heavily on interspecific hybridization between taxonomically distinct lineages, generating highly heterozygous populations and enabling transgressive segregation for traits such as flavor, yield, and antioxidant content [107]. Genomic resources now support the study of this complexity, with R. occidentalis (black raspberry, ~290 Mb) sequenced using PacBio and Hi-C technologies [89], while the R. argutus ‘Hillquist’ genome (~270–298 Mb) offers a high-quality chromosome-scale reference for dissecting antioxidant-related traits in polyploid backgrounds [86].
Wild Rubus relatives remain an essential reservoir of antioxidant and adaptive diversity. The R. fruticosus aggregate, comprising more than 400 microspecies (agamospecies) maintained through apomixis, provides stable sources of anthocyanins, ellagitannins, and pathogen resistance [84]. European populations exhibit anthocyanin concentrations as high as 589 mg/100 g FW and unique ellagitannin profiles, though this value applies to select R. fruticosus accessions used in food colorant studies—not necessarily wild populations broadly [92]. North American species contribute complementary adaptive traits: R. allegheniensis combines cold hardiness and unique terpene profiles; R. argutus contributes heat tolerance, extended harvest windows, and high phenolic diversity; and R. ursinus, the Pacific dewberry, imparts exceptional flavor and rare anthocyanin glycosides [109]. Further genetic breadth derives from Mexican wild species with tropical adaptation and continuous fruiting potential, though direct citation is limited and may be supported by regional germplasm surveys or USDA reports. Asian taxa such as R. corchorifolius and R. phoenicolasius contribute disease resistance, stress tolerance, and novel bioactive compounds [110]. Comparative studies using AFLP markers reveal that North American blackberry cultivars have a narrower genetic base compared with European wild populations, underscoring the need to broaden germplasm pools in breeding programs [111]. Conservation initiatives now safeguard over 800 accessions globally, capturing diverse lineages to ensure sustainable exploitation of antioxidant-related traits [112].
Commercial blackberry cultivars display marked variation in antioxidant composition, reflecting their diverse pedigrees. Erect thorny selections have been reported with high total phenolic content and anthocyanin levels, showing excellent ORAC values and storage stability [113]. Thornless erect cultivars display high ellagitannin accumulation, malvidin-rich anthocyanin profiles, and shelf-life extension, while semi-erect genotypes achieve total antioxidant capacities up to 11,200 μmol TE/100 g, with consistent phenolic retention under stress [95]. Trailing cultivars from the Pacific Northwest dominate the processing and fresh markets due to their distinctive flavor, anthocyanin diversity, and superior postharvest vitamin C retention [94,108]. Recent metabolomic and pharmacological profiling identified several high-antioxidant cultivars such as ‘Kiowa’, which exhibits the highest antioxidant and anti-inflammatory activity, driven by high levels of quercitrin, luteolin, and cyanidin 3-O-glucoside. Cultivar ‘10-5n-2’ contains strong total anthocyanin and polyphenol content, while ‘Chester’ is noted for strong fruit firmness and high antioxidant retention; and ‘Shuofeng’ combines high yield with excellent antioxidant capacity. Additional cultivars such as ‘Thorn Free’, have been reported with notably high cyanidin-3-glucoside and total phenols concentrations as well as ‘Loch Ness’, which exhibits very high total phenols and strong antioxidant capacity, further exemplify the nutritional potential of thornless selections [114], while ‘TripleCrown’ and ‘Encore’ exhibit exceptionally high antioxidant activity, with DPPH values of 184.43 and 316.02 µg/ml, respectively, and strong FRAP values, highlighting their potential for breeding antioxidant-rich genotypes [45]. Complementing cultivated germplasm, wild relatives such as R. ulmifolius, R. perrobustus, R. wimmerianus, R. pedemontanus, and R. grabowskii also display exceptionally high phenolic content in fruits and leaves, offering unique opportunities for bioactive compound enrichment in breeding [115].
3.3.2. Modern Genomic and Breeding Tools for Enhancing the Antioxidant Character of Blackberry
Blackberry fruit quality depends on constitutive antioxidant production systems that operate throughout normal fruit development. In the study of Huang et al. (2023), the authors investigated the molecular basis of antioxidant accumulation during fruit ripening in blackberry by monitoring the expression of structural and defense-related genes [116]. They collected blackberry fruits at different developmental stages (green, red, and fully black) and quantified the gene expression through transcriptome profiling combined with quantitative RT-PCR. They reported that genes such as PAL, CHS, CHI, and F3H exhibited higher expression at early developmental stages, establishing the metabolic framework for downstream flavonoid biosynthesis. In contrast, the late anthocyanin biosynthetic genes, namely DFR, ANS, and UFGT, showed pronounced up-regulation during the red-to-black transition, coinciding with the rapid accumulation of anthocyanins. Additionally, antioxidant defense enzymes including SOD, CAT, APX, and POD, were significantly induced at the late ripening stage, reflecting their role in mitigating oxidative stress during fruit maturation [116]. Table 4 presents the genes that have been functionally annotated and characterized till today according to the international literature findings.
Genomic resources for blackberry are rapidly advancing, with recent efforts focused on developing chromosome-scale reference genomes for diploid R. argutus (‘Hillquist’, ~298 Mb) and tetraploid BL1 (~919 Mb), assembled using PacBio, Oxford Nanopore, and Hi-C technologies [123,124]. These assemblies reveal complex subgenome relationships shaped by ancient polyploidization and diploidization, with gene family expansions in secondary metabolite biosynthesis—particularly phenylpropanoid and flavonoid pathways—and stress-response genes. Transcriptomic datasets across fruit developmental stages and tissue types support functional annotation and breeding applications [125]. Resequencing of seven tetraploid cultivars uncovered candidate genes for antioxidant traits and fruiting habits [124]. Early diversity studies using AFLP markers laid the foundation for pangenome initiatives, suggesting an accessory genome comprising 15–20% of total genes enriched for secondary metabolism and environmental response [126]. Likewise, Chizk et al. (2023), performed a large-scale GWAS on 300 tetraploid blackberry genotypes to identify genetic factors controlling fruit firmness and red drupelet reversion (RDR). Their findings demonstrated that postharvest quality traits are moderately heritable and influenced by multiple small-effect QTLs, including loci linked to polygalacturonase, pectin methylesterase, and glucanase related genes. Overall, the research highlighted that postharvest quality is genetically complex but provided candidate loci and datasets for future genomic selection [127].
3.4. Cranberry (Vaccinium macrocarpon Ait.)
3.4.1. Domestication and Wild Relatives
American cranberry is a diploid species (2n = 2x = 24) native to acidic bog ecosystems of North America, cultivated primarily in the United States, Canada, and Chile for its high polyphenol content and associated health benefits. It represents one of only three widely cultivated fruit crops native to North America, with domestication dating back approximately 200 years ago [54,128]. Natural populations span acidic wetlands from Newfoundland to North Carolina and west to Minnesota, with genetic diversity centers in Wisconsin, Massachusetts, and New Jersey bog regions [128]. Phylogenetic analyses using chloroplast markers indicate that V. macrocarpon diverged from other cranberry species 5–10 million years ago, followed by population expansion after Pleistocene glacial retreats [129,130]. Early domestication involved selection from wild bog populations, showing minimal genetic bottlenecks compared to other crops. Modern breeding began in the 1950s, initially targeting fruit quality and yield, and later expanded to include enhanced antioxidant content and anthocyanin profiles, as confirmed by genotyping and metabolomic studies of breeding cycles [131,132].
Cranberry genetic resources include both wild populations and related species. Wisconsin bog populations harbor the largest genetic diversity, with over 150 distinct genotypes serving as sources of disease resistance and unique antioxidant profiles [128,133]. Massachusetts populations provide cold hardiness traits and elevated proanthocyanidin content [134], while New Jersey Pine Barrens populations contribute heat tolerance, unique A-type proanthocyanidin variants, and extended harvest characteristics [54]. Canadian populations exhibit extreme cold tolerance, specialized flavonoid profiles, and late season ripening traits [135]. Related species, including V. oxycoccos (small cranberry) and V. microcarpum, are genetically distinct wild relatives of V. macrocarpon. They contribute specialized traits such as cold hardiness, disease resistance, high anthocyanin content, unique glycosylation patterns, and stress tolerance [54]. Microsatellite marker analyses of wild cranberry (V. macrocarpon) clones from Newfoundland and Labrador revealed significant population structure correlated with geography, along with high allelic richness and diverse antioxidant properties, highlighting these Canadian populations as priority resources for cranberry conservation and breeding [135]. The USDA’s National Clonal Germplasm Repository (NCGR) maintains over 300 cranberries and cranberry-wild relative accessions (including ~146 V. macrocarpon) in its Vaccinium collection. Genetic studies of cultivars and accessions have identified dozens of unique genotypes among these accessions for example, 64 unique genotypes out of 271 plants in 77 accessions confirming substantial genotypic diversity in the repository [136] . Recent chromosome-level genome assemblies for V. macrocarpon and V. microcarpum further provide foundational resources for trait and antioxidant biosynthesis analyses [133].
Commercial cranberry cultivars are selected for enhanced antioxidant profiles across early, mid, and late harvest seasons. Early-season cultivars are noted for high anthocyanin and total phenolic content, excellent processing quality, elevated proanthocyanidin levels, and superior antioxidant stability [134]. Heritage cultivars such as ‘Early Black’ and ‘Howes’ also fall into this category, showing strong antioxidant capacity and minimal genetic bottlenecks, with ‘Howes’ exhibiting exceptionally high anthocyanin concentrations and ‘Early Black’ known for stable phenolic content across ripening stages [137]. Mid-season cultivars such as ‘Stevens’ serve as industry standards with balanced antioxidant profiles and high A-type proanthocyanidin content, while others like ‘Ben Lear’ exhibit elevated total phenolics, unique anthocyanin glycosylation patterns, and strong cold hardiness. ‘McFarlin’, another heritage cultivar, contributes moderate antioxidant levels and maintains high genetic diversity, making it valuable for breeding and conservation. Late-season cultivars are recorded with exceptional anthocyanin concentrations and the highest observed ORAC values, with selections like ‘Bergman’ noted for elevated flavonol content and superior antioxidant retention during processing [138]. Additional high-antioxidant cultivars include ‘Crimson Queen’, known for its high anthocyanin and proanthocyanidin content with excellent color stability; ‘Mullica Queen’, which demonstrates elevated total phenolics and strong antioxidant retention during processing; ‘Scarlet Knight’, rich in flavonols and anthocyanins and bred for functional food applications; ‘HyRed’, valued for its deep red pigmentation and high ORAC values; and ‘Demoranville’, which combines high proanthocyanidin levels with improved disease resistance [132]. Specialized selections provide deep red pigmentation, diverse anthocyanin profiles, enhanced proanthocyanidin content, and improved disease resistance, making them ideal for premium fresh markets and functional food applications [137]. Finaly ‘Kalnciema Agrā’ and ‘Kalnciema Tumšā’ exhibit high proanthocyanidin and triterpenoid content, contributing to strong antioxidant activity. ‘Searles’ is notable for elevated proanthocyanidins, while ‘Black Veil’ stands out with high flavonol levels. ‘Red Star’ combines high chlorogenic acid with triterpenoids, and ‘Pilgrim’, despite lower anthocyanins, shows significant antioxidant capacity due to its triterpenoid content [137,139].
3.4.2. Modern Genomic and Breeding Tools for Enhancing the Antioxidant Character of Cranberry
Cranberries are rich in phenolic compounds, particularly flavonoids and proanthocyanidins, which have been widely studied for their diverse biological activities [140]. Breeding strategies have primarily focused on traits prioritized by the industry, such as fruit quality characteristics—including firmness, size, and anthocyanin content—as well as resistance to fruit rot, which are considered the most desirable attributes in new cranberry cultivar releases [141]. Several studies suggest that advanced DNA-based selection strategies could significantly benefit the global cranberry industry [141], while others emphasize that integrating genetic diversity analysis with antioxidant profiling using DNA markers (ISSR, EST-SSR, EST-PCR) is crucial for conserving wild cranberry germplasm and identifying diverse, antioxidant-rich genotypes for future breeding and crop improvement [135]. Vorsa et al. in 2019 reviewed the biology, domestication history, key traits, and breeding challenges of American cranberry, highlighting recent molecular advances and proposing MAS as a crucial tool for future genetic improvement [142]. They noted QTLs identified for important traits like productivity, berry size and anthocyanins content pointing the value of molecular tools and MAS to overcome traditional breeding limitations and accelerate genetic improvement. Debnath et al. (2012) used DNA-based markers, particularly ISSRs, to analyze genetic diversity in wild and cultivated berry crops, showing that these markers effectively differentiated genotypes and are useful for cultivar identification and breeding, while also reporting wide variation in antioxidant activity that did not align with molecular diversity patterns, highlighting the need to combine both approaches for germplasm conservation and crop improvement [143].
Few studies have applied GWAS in cranberry breeding, identifying marker–trait associations for key agronomic traits such as average fruit weight and fruit rot resistance [144]. Other research has uncovered extensive gene presence–absence variation related to disease resistance and fruit quality, with these resources now publicly available to accelerate molecular breeding in Vaccinium crops [131]. However, breeding efforts specifically targeting fruit antioxidant improvement remain largely unexplored in cranberries. Moreover, while genetic tools such as CRISPR/Cas9 have begun to be applied in other berry species like blueberries [145,146], these efforts have primarily focused on traits unrelated to antioxidant capacity. To date, no reports exist on the application of such genome-editing tools in cranberry.
3.5. Grapevine (Vitis vinifera L.)
3.5.1. Domestication and Wild Relatives
V. vinifera is one of the most economically important crops cultivated worldwide. Global vineyard coverage spans about 7.2 million hectares, producing 77.8 million tons of grapes annually of which 47.4% is used for winemaking, 44.5% for fresh consumption, and 8% for dried grapes (OIV). Grape species are classified into two subgenera: Muscadinia, (2n = 40) and the larger subgenus Euvitis (including V. vinifera), with a basic chromosome number of 2n = 38. Grapevine domestication dates back approximately 8,000–11,000 years and archaeological and genetic evidence points to Western Asia (a region between the Black Sea and Iran) as the primary center of origin, though multiple domestication centers have also been proposed [145,146,147]. The Vitis genus comprises 60–80 species characterized by exceptionally high genetic diversity. Ampelographic and genetic analyses suggest that between 6,000 and 10,000 V. vinifera cultivars are currently cultivated worldwide [148]. Global germplasm inventory repositories, such as the Vitis International Variety Catalogue (VIVC), have catalogued thousands of cultivars and wild accessions, providing valuable resources for elucidating phylogenetic relationships and trait variation.
Wild Vitis relatives exhibit important traits not found in V. vinifera that have been associated mainly with resistance/tolerance to biotic/abiotic stresses [149]. Historically, European V. vinifera grapevines were grafted onto American varieties derived from wild Vitis species to combat the catastrophic infestation of the soil born aphid phylloxera (Daktulosphaira vitifoliae) [150]. Moreover, over the years, breeding efforts utilizing crosses of V. vinifera with wild species such as V. arizonica and Muscadinia rotundifolia managed to develop lines with resistance to devastating bacterial and fungal diseases like ‘Pierce’s disease-PD’, downy mildew (Plasmopara viticola) and (powdery mildew (Erysiphe necator), respectively [151].
In the context of antioxidant capacity, the wild relative Muscadinia rotundifolia (Michx.) Small, a grape species native to the southeastern United States, has drawn much interest recently from the food, pharmaceutical, and nutraceutical industries owing to their unique chemical composition and nutritional value [152]. Muscadine grapes are rich in distinctive primary and secondary metabolites, including notable phenolics like gallic acid, ellagic acid, proanthocyanidins, catechins, quercetin, resveratrol, and myricetin. Muscadine-derived bioactive compounds exhibit strong antioxidant, anticancer, antimutagenic, antimicrobial, and anti-inflammatory properties, and thus hold tremendous health promoting potential [153]. They are also essential contributors to grape quality traits and their wide flavor diversity.
3.5.2. Modern Genomic and Breeding Tools for Enhancing the Antioxidant Character of Grapevine
The grapevine was among the first fruit crops to be sequenced, with the reference genome of the near-homozygous Pinot Noir PN40024 line published by Jaillon et al. (2007) [154] . The diploid genome is made up of 19 chromosomes and covers about 487 Mb. The assembly has been improved to chromosome-scale resolution by further updates that included BAC-by-BAC sequencing, Illumina short reads, and long-read data [155]. Gene families engaged in secondary metabolism were revealed by annotation, particularly those associated with the biosynthesis of terpene and polyphenol compounds, which include antioxidants like anthocyanins and stilbenes (e.g., resveratrol) [156]. These genomic resources have allowed identification of key genes such as VvDXS1, VvbZIP61 (terpene synthesis) VvCHS, VvMYBA1 and VvSTS (polyphenol synthesis) facilitating marker-assisted breeding and functional genomics in grapevines. Table 5 presents the genes that have been functionally annotated and characterized regarding their association with antioxidant levels.
Advances in DNA sequencing technologies have yielded enhanced grapevine reference genome assemblies [157] and a non-reference pangenome derived from wild grapevine accessions [158]. These genomic resources provide powerful tools for elucidating the molecular basis of key morphological, physiological, and biochemical traits that influence yield, fruit quality, and stress tolerance. Notably, the recent development of a comprehensive grapevine pangenome reference map built from resequencing data of 466 cultivars and capturing an extensive array of both short variants and structural variants (SVs)—is expected to shed light on complex polygenic trait associations and accelerate genomic-assisted breeding efforts [159].
Transcriptomics and metabolomics are increasingly employed to investigate molecular processes in grapevines and have been focusing either on plant and fruit development or on interactions with abiotic and biotic factors. Integrating these approaches has become critically important for understanding key physiological and metabolic pathways [160]. Insights gained from such analyses can be directly applied in breeding programs, using genes linked to improved fruit quality—such as those enhancing the accumulation of health-promoting compounds like antioxidants—as molecular markers for crop improvement.
Over the past two decades, a wealth of transcriptome and metabolome data has been produced which greatly aid the investigation of grapevine berry physiology, secondary metabolism, and quality traits [160,161,162,163,164]. Integrated “omics” datasets can be compiled into user-friendly resources for the research community, enabling exploration of potential gene–gene and gene–metabolite interactions that can later be validated through hypothesis-driven experiments. One such resource is the Grapevine Transcriptomic and Metabolomic Integrated Database (TransMetaDb), available through the Vitis Visualization (VitViz) platform (https://tomsbiolab.com/vitviz which provides a valuable tool for linking antioxidant potential with berry development and metabolism [164].

Table 5.
Genes analyzed for their functional involvement in the antioxidant capacity of grapevine.
| Enzyme Class | Gene name | Functional Role/ Association with antioxidant levels | References |
| 1-deoxy-d-xylulose-5-phosphate synthase (DXS) | VvDXS1 | Rate-limiting enzyme of the MEP pathway; Major determinant of monoterpenoid content | Yang et al., 2017 [165] Zhang et al., 2025 [167] |
| Basic-leucine zipper transcription factor | VvbZIP61 | Monoterpene metabolism Associated with increased levels of monoterpenes |
Zhang et al., 2023 [166] |
| Isopentenyl pyrophosphate synthases (IPPS) (geranylgeranyl pyrophosphate synthase) |
VvGGPPS-LSU | Enzyme of the MEP terpene biosynthesis pathway; Associated with increased accumulation of monoterpenoid and norisoprenoid levels. |
Zhang et al., 2025 [167] |
| WRKY transcription factors |
VvWRKY24 | A key regulator of isoprenoid metabolism. Associated with enhanced levels of β-damascenone, an isoprenoid important for berry and wine aroma |
Wei et al., 2025 [168] |
| MYB transcription factors |
VvMYBA1 | A key positive regulator of anthocyanin biosynthesis | Liu et al., 2023 [169] |
| Chalcone synthase (CHS) | VvCH2 | A key enzyme in the phenylpropanoid metabolism committed to the synthesis of flavonoids | Lai et al., 2025 [170] |
| Stilbene synthases (STS) | VvSTS | A key enzyme of the phenylpropanoid pathway committed to the synthesis of resveratrol | Lai et al., 2025 [170] |
In grapevine, antioxidants comprise large classes of compounds, including secondary metabolites involved in shaping the aroma and flavor of grapes and wine (e.g., terpenoids), compounds related to health benefits (e.g., polyphenols, resveratrol, anthocyanins), and compounds determining the responses to biotic and abiotic stresses (e.g., stilbenes, anthocyanins, terpenes) [171]. Despite the importance of antioxidant content as a key quality trait in grapevine breeding, the genetic basis underlying the biosynthesis and accumulation of aroma/flavor-related terpenoid compounds or phenolic, flavonoid and anthocyanin molecules remains obscure due to the complexity of the respective metabolic pathways that involve a multitude of enzymes and gene networks [172]. Moreover, reliable molecular markers linked to these traits are largely lacking, posing a challenge for the implementation of marker-assisted selection (MAS) in antioxidant-enhancement breeding and pointing to the necessity of adopting modern genomic tools and molecular breeding strategies [173].
- Antioxidants related to the aromatic profile of grapes and wine
The complex bouquet of grape aroma is attributed to a diverse array of volatile organic compounds, such as benzenoids, esters, norisoprenoids, and terpenoids [174,175]. Distinct classes of terpenoids like mono- and sesquiterpenes contribute significantly to the characteristic aromatic profiles of individual grapevine cultivars. For example, monoterpenoid alcohols such as linalool, nerol, and geraniol are key contributors to the floral and citrus-like aromas typical of highly aromatic varieties such as Muscat and Gewürztraminer. On the other hand, sesquiterpenes, such as rotundone, are associated with spicy, peppery notes, particularly characteristic of the Syrah cultivar [176]. Despite numerous investigations, thus far, the genetic and regulatory aspects of terpenoid production have not been fully uncovered due to the complexity of the particular biosynthetic pathway. Monoterpenoids and sesquiterpenes are synthesized via the mevalonate pathway (MVA) and the methylerythritol phosphate (MEP) pathway which are compartmentalized into the cytoplasm and plastids, respectively (Figure 2). The 1-deoxy-d-xylulose-5-phosphate synthase (DXS) and HMG-CoA reductase (HMGR) are the major contributors to metabolic flux control of the pathways, respectively whereas terpene synthases (TS) functioning in the final steps are associated with the synthesis of distinct terpenoid molecules from prenyldiphosphate precursors [177]. A large family of terpene synthases (TS) encoded by 69 genes identified in the grapevine genome is considered to play critical roles in terpenoid biosynthesis [178]. Structurally diverse terpene synthases acting on prenyldiphosphate precursors are considered to drive the wide range of terpenes produced in grapes. These terpenes can then undergo secondary modifications, such as glycosylation and oxidation, further expanding the diversity of grape and wine aromas and flavors [177]. In addition, transcription factors such as members of the NAC, MYB, and AP2/ERF families, have been suggested to play roles in the transcriptional regulation of terpene synthesis genes in grapevine [178]. However, the functional characterization of most TS genes/enzymes or transcription factor genes has been limited. Similarly, the oxidation, reduction, and glycosylation processes that lead to the production of monoterpenoid derivatives -also key aroma compounds- are poorly understood [167].
Early QTL mapping and GWAS studies demonstrated that a major QTL linked to aroma profiling and monoterpenoid content was associated with the VvDXS1gene. A series of causal SNPs were identified mainly in the coding region of the gene affecting the catalytic activity of the DXS1 enzyme, as well as in the regulatory region affecting gene expression levels [165,179,180,181,182]. Thus, DXS1 was proposed as an effective molecular marker for an aroma-related quality trait; nevertheless, various aromatic varieties and their aroma/flavor variabilities were found unrelated to DXS1, highlighting the need for further gene mining and development of additional genetic markers.
An integrative approach combining QTL mapping, transcriptomic analysis, and calli transformation revealed additional genes associated with monoterpenoid composition and abundance [166]. The F1 progeny of a cross between an aromatic (Beifeng) and a neutral (3–34) grape variety identified stable QTLs associated with 11 monoterpenes. In parallel, RNA seq analysis from 34 diverse cultivars with distinct aromatic profiles, indicated a significant correlation between the expression of genes mapping to these QTLs and monoterpenoid content of grapevine berries. One of those was identified as the VvbZIP61 gene encoding a basic-leucine zipper transcription factor known to regulate various plant processes. VvbZIP61 implication in monoterpenoid metabolism was further supported by its overexpression in V. amurensis calli, which led to a significant increase in the accumulation of several monoterpene compounds [166].
To identify additional candidate genes associated with monoterpenoid levels, Zhang et al. (2025) employed GWAS on an F1 population derived from crosses between a typical muscat variety (Muscat of Alexandria) and a nonaromatic variety (Christmas Rose). A range of significant SNPs were identified, all located on chromosome 5, forming a monoterpenoid-related marker cluster. 25 lead SNPs appearing stable over a two-year period were validated in a germplasm population of 97 diverse Vitis species and were found highly associated with monoterpenoid levels. The underlying genes encode the DXS enzyme, a glycoside hydrolase, an ABC transporter, and a geranylgeranyl pyrophosphate synthase-large subunit (VvGGPPS-LSU) involved in the terpene synthesis pathway. Notably, the functionality of the latter in increasing monoterpenoid levels was verified in vivo through transformation and overexpression in transgenic calli, reinforcing the reliability of the marker cluster. Importantly, the 25 lead SNP markers were converted to a KASP marker set, enabling direct molecular breeding applications [167].
Besides terpenoid compounds, norisoprenoids, which are derived from the oxidative cleavage of carotenoids, constitute another class of volatile aroma compounds with major contribution to the floral and fruity notes of grape berries and wine. Among them, β-damascenone is the most abundant norisoprenoid found in grapevine [183]. A recent study reported that a WRKY transcription factor, ViWRKY24, plays a role as a key regulator of β-damascenone accumulation in grape berries. Functional characterization through overexpression and gene silencing in grape leaves, berries, and calli revealed that VviWRKY24 modulates norisoprenoid metabolic flux and alters norisoprenoid composition, specifically enhancing β-damascenone levels [168].
The studies described above will illuminate further the regulation of terpenoid and isoprenoid metabolic pathways and facilitate grapevine improvement as it pertains to antioxidant related aroma/flavor traits.
-Antioxidants related to phenolic compounds and health benefits
In the context of the rising interest in nutraceuticals and phenolic antioxidants found in the skin, flesh, and seeds of grapevine berries, Muscadine grapes (Muscadinia rotundifolia Michx.), a native grape of the southeastern United States, have been gaining much attention in recent years. Muscadine grapes have distinctive flavor and aroma properties and are a major source of essential oils, vitamins, minerals, and antioxidants. Notably, they accumulate significantly higher levels of bioactive polyphenolic compounds compared to other grape species [184,185], enhancing their value both nutritionally and commercially.
Antioxidant activity is higher in skin than pulp and increasing the antioxidant capacity of muscadine berry skin has been an important breeding target. Although many studies have reported on the phenolic content and antioxidant activities in muscadine berries [186], the genetic basis of varied antioxidant profiles has not been studied until recently. Transcriptomic analysis of two contrasting muscadine genotypes differing in phenolic content and antioxidant capacity revealed distinct transcript profiles across berry development that was accompanied by differential total phenolic/total flavonoid contents and antioxidant activities [187]. Differential gene expression was witnessed predominately at the veraison stage and involved genes coding for key enzymes in the polyphenol biosynthetic pathway which were found significantly upregulated in the C5 genotype pointing to genotype-specific regulation. On the other hand, genes of the non-flavonoid pathway—stilbene synthases (STS1, STS2, STS3)— exhibited higher expression levels at the veraison and ripe berry stages in the C6 genotype, indicating, enhanced, genotype-specific production of stilbenoid compounds in the late berry developmental stages [187]. Similarly, RNA-seq was employed to perform comparative transcriptomic analysis of phenolic pathway associated genes in two muscadine grape varieties, Paulk and Supreme, during berry development. Differential gene expression was observed for VvPAL and VvCHS which manifested in a genotype- and tissue-specific manner. Conversely, VvMYBA1 and VvMYBCS1 were significantly upregulated in veraison and mature grape berries in both varieties, suggesting a positive contribution to anthocyanin accumulation [188]. These findings underscore the importance of specific metabolic genes and transcriptional regulators in shaping antioxidant biosynthesis, providing valuable targets for breeding programs aimed at enhancing antioxidant capacity and fruit quality.
To elucidate the genetic architecture of antioxidant quantitative traits in Muscadinia grape, Park et al. (2022) conducted a multi-locus GWAS analysis to identify genomic loci associated with total phenolic and flavonoid content and the quantitative property of their antioxidant activity [173]. 350 genotypes were analyzed through a combination of phenotypic (phenolic content determination and antioxidant activity), berry skin transcriptomic, and GWAS analyses. 12 Quantitative Trait Nucleotides (QTNs) were significantly correlated with four antioxidant-related traits, and candidate genes were found to be positively and negatively associated with antioxidant activities. Among the positively correlated candidate genes, a UDP-glycosyltransferase was identified as a strong candidate significantly correlating with high antioxidant values. UDP-glycosyltransferases are known to be involved in flavonoid and anthocyanin biosynthesis, indicating that the muscadine gene identified in this study may play an important role in enhanced antioxidant capacity. Notably, the gene coding for 4-hydroxy-4-methyl-2-oxoglutarate aldolase was negatively associated with antioxidant values. This gene is involved in the degradation pathway of gallic acid—a key phenolic compound—in bacteria [188], suggesting that it may act as a negative regulator of phenolic antioxidant activity in muscadine grapes. In another study, the role of the VvMYBA1 transcription factor in anthocyanin biosynthesis was further elucidated using a combination of CRISPR/Cas9-mediated knockout of VvBBX44, a gene encoding a repressor of VvMYBA1, and transient transformation of ‘Jingxiu’ grapevine berries. Loss of VvBBX44 function led to increased anthocyanin accumulation, whereas overexpression of VvMYBA1 increased VvBBX44 expression [189]. These results demonstrate that VvMYBA1 and VvBBX44 act together in a regulatory feedback loop to fine-tune anthocyanin biosynthesis, maintaining anthocyanin levels in the desired range.
Along these lines, a notable recent study has reported on the use of targeted editing through CRISPR/Cas9 technology to induce a metabolic shift from the flavonoid to the stilbenoid pathway, thereby promoting resveratrol biosynthesis and accumulation in V. davidii transgenic cells [170]. This approach leveraged the fact that both resveratrol (a stilbenoid compound) and flavonoids are derived from the phenylpropanoid biosynthesis pathway, where stilbene synthase (STS) and chalcone synthase (CHS) compete for the same substrate to produce stilbenoids and flavonoids, respectively (Figure 3). CRISPR/Cas9-based targeted editing of the VdCH2 gene resulted in efficient CHS2 knock-out leading to suppression of the flavonoid pathway and reduction in flavonoid levels, but to a significant increase in stilbenoid compounds such as resveratrol and its derivatives. These findings offer novel strategies for enhancing targeted antioxidant content in grapevine through redirecting metabolic flux from the flavonoid to the stilbenoid pathway and present an effective alternative to the challenging and costly extraction of resveratrol, a highly valuable antioxidant with beneficial antioxidant properties [170,190].
The findings above set the basis for a deeper understanding of polyphenol accumulation and antioxidant activity and pave the way for enhancing the nutraceutical value of berries through modern breeding and metabolic engineering approaches.
4. Conclusions
Berry crops are highly valued for their rich antioxidant capacity, largely attributed to anthocyanins, flavonols, proanthocyanidins, vitamins, and carotenoids, which play important roles in reducing oxidative stress and lowering the risk of chronic diseases in humans. Until recently, breeding programs rarely focused directly on antioxidant traits. Instead, breeding for antioxidants was achieved indirectly, mostly through the selection of genotypes with increased tolerance to environmental stresses. Despite the availability of high-quality genome assemblies for several berry species, the specific roles, annotation, and functional characterization of many genes underlying antioxidant biosynthesis remain insufficiently explored. However, due to the rising demand for functional foods, the targeted breeding of berries for enhanced antioxidant potential has become an emerging priority. This shift is supported by the exploitation of the wide genetic diversity found in wild relatives, which serve as reservoirs of valuable traits, and by the integration of modern molecular and genetic breeding tools, including QTL mapping, marker-assisted selection, genomic selection, and CRISPR/Cas9-based gene editing [191]. The combination of advanced molecular and genetic breeding tools provides powerful opportunities to accelerate the development of cultivars that combine high nutritional quality with resilience and productivity, contributing both to human health and to sustainable berry production systems.
Author Contributions
Conceptualization, V.P., E.T., and A.K.; writing—original draft preparation, G.M., A.V., A.K and M.G.; writing—review and editing, V.P., E.T., A.K., G.M., A.V., M.G. and E.M.A.; visualization, M.G. and A.K.; supervision, V.P., E.T., and E.M.A.; project administration, V.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kumar, S.; Baghel, M.; Yadav, A.; Dhakar, M.K. Postharvest Biology and Technology of Berries. In Postharvest Biology and Technology of Temperate Fruits; Mir, S.A., Shah, M.A., Mir, M.M., Eds.; Springer International Publishing: Cham, 2018; pp. 349–370 ISBN 978-3-319-76842-7.
- Dickenson, V. Berries; Reaktion Books, 2020.
- Kaume, L.; Howard, L.R.; Devareddy, L. The Blackberry Fruit: A Review on Its Composition and Chemistry, Metabolism and Bioavailability, and Health Benefits. J. Agric. Food Chem. 2012, 60, 5716–5727. [CrossRef]
- Skrovankova, S.; Sumczynski, D.; Mlcek, J.; Jurikova, T.; Sochor, J. Bioactive Compounds and Antioxidant Activity in Different Types of Berries. International journal of molecular sciences 2015, 16, 24673–24706. [CrossRef]
- FAOSTAT Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 24 September 2025).
- Pedisić, S.; Zorić, Z.; Repajić, M.; Levaj, B.; Dobrinčić, A.; Balbino, S.; Čošić, Z.; Dragović-Uzelac, V.; Elez Garofulić, I. Valorization of Berry Fruit By-Products: Bioactive Compounds, Extraction, Health Benefits, Encapsulation and Food Applications. Foods 2025, 14, 1354. [CrossRef]
- Cosme, F.; Pinto, T.; Aires, A.; Morais, M.C.; Bacelar, E.; Anjos, R.; Ferreira-Cardoso, J.; Oliveira, I.; Vilela, A.; Gonçalves, B. Red Fruits Composition and Their Health Benefits—A Review. Foods 2022, 11, 644. [CrossRef]
- Li, X.; Sun, J.; Chen, Z.; Jiang, J.; Jackson, A. Characterization of Carotenoids and Phenolics during Fruit Ripening of Chinese Raspberry (Rubus Chingii Hu). RSC advances 2021, 11, 10804–10813. [CrossRef]
- La Torre, C.; Loizzo, M.R.; Frattaruolo, L.; Plastina, P.; Grisolia, A.; Armentano, B.; Cappello, M.S.; Cappello, A.R.; Tundis, R. Chemical Profile and Bioactivity of Rubus Idaeus L. Fruits Grown in Conventional and Aeroponic Systems. Plants 2024, 13, 1115. [CrossRef] [PubMed]
- Bhat, I.U.H.; Bhat, R. Quercetin: A Bioactive Compound Imparting Cardiovascular and Neuroprotective Benefits: Scope for Exploring Fresh Produce, Their Wastes, and by-Products. Biology 2021, 10, 586. [CrossRef]
- Zaa, C.A.; Marcelo, Á.J.; An, Z.; Medina-Franco, J.L.; Velasco-Velázquez, M.A. Anthocyanins: Molecular Aspects on Their Neuroprotective Activity. Biomolecules 2023, 13, 1598. [CrossRef] [PubMed]
- Norouzkhani, N.; Afshari, S.; Sadatmadani, S.-F.; Mollaqasem, M.M.; Mosadeghi, S.; Ghadri, H.; Fazlizade, S.; Alizadeh, K.; Akbari Javar, P.; Amiri, H. Therapeutic Potential of Berries in Age-Related Neurological Disorders. Frontiers in Pharmacology 2024, 15, 1348127. [CrossRef]
- Padilla-González, G.F.; Grosskopf, E.; Sadgrove, N.J.; Simmonds, M.S. Chemical Diversity of Flavan-3-Ols in Grape Seeds: Modulating Factors and Quality Requirements. Plants 2022, 11, 809. [CrossRef] [PubMed]
- Noce, A.; Di Daniele, F.; Campo, M.; Di Lauro, M.; Pietroboni Zaitseva, A.; Di Daniele, N.; Marrone, G.; Romani, A. Effect of Hydrolysable Tannins and Anthocyanins on Recurrent Urinary Tract Infections in Nephropathic Patients: Preliminary Data. Nutrients 2021, 13, 591. [CrossRef]
- Sójka, M.; Hejduk, A.; Piekarska-Radzik, L.; Ścieszka, S.; Grzelak-Błaszczyk, K.; Klewicka, E. Antilisterial Activity of Tannin Rich Preparations Isolated from Raspberry (Rubus Idaeus L.) and Strawberry (Fragaria X Ananassa Duch.) Fruit. Scientific Reports 2025, 15, 10196. [CrossRef]
- Cosme, F.; Aires, A.; Pinto, T.; Oliveira, I.; Vilela, A.; Gonçalves, B. A Comprehensive Review of Bioactive Tannins in Foods and Beverages: Functional Properties, Health Benefits, and Sensory Qualities. Molecules 2025, 30, 800. [CrossRef]
- Sławińska, N.; Prochoń, K.; Olas, B. A Review on Berry Seeds—A Special Emphasis on Their Chemical Content and Health-Promoting Properties. Nutrients 2023, 15, 1422. [CrossRef]
- Duta-Bratu, C.-G.; Nitulescu, G.M.; Mihai, D.P.; Olaru, O.T. Resveratrol and Other Natural Oligomeric Stilbenoid Compounds and Their Therapeutic Applications. Plants 2023, 12, 2935. [CrossRef]
- González de Llano, D.; Moreno-Arribas, M.; Bartolomé, B. Cranberry Polyphenols and Prevention against Urinary Tract Infections: A Brief Review. 2021.
- Golovinskaia, O.; Wang, C.-K. Review of Functional and Pharmacological Activities of Berries. Molecules 2021, 26, 3904. [CrossRef]
- Singh, A.K.; Singla, R.K.; Pandey, A.K. Chlorogenic Acid: A Dietary Phenolic Acid with Promising PharmacotherapeuticPotential. CMC 2023, 30, 3905–3926. [CrossRef]
- Bezerra, M.; Ribeiro, M.; Cosme, F.; Nunes, F.M. Overview of the Distinctive Characteristics of Strawberry, Raspberry, and Blueberry in Berries, Berry Wines, and Berry Spirits. Comp Rev Food Sci Food Safe 2024, 23, e13354. [CrossRef] [PubMed]
- Bai, Q.; Shen, Y.; Huang, Y. Advances in Mineral Nutrition Transport and Signal Transduction in Rosaceae Fruit Quality and Postharvest Storage. Frontiers in Plant Science 2021, 12, 620018. [CrossRef] [PubMed]
- Martins, V.; Soares, C.; Spormann, S.; Fidalgo, F.; Gerós, H. Vineyard Calcium Sprays Reduce the Damage of Postharvest Grape Berries by Stimulating Enzymatic Antioxidant Activity and Pathogen Defense Genes, despite Inhibiting Phenolic Synthesis. Plant Physiology and Biochemistry 2021, 162, 48–55. [CrossRef] [PubMed]
- Ahmed, N.; Zhang, B.; Bozdar, B.; Chachar, S.; Rai, M.; Li, J.; Li, Y.; Hayat, F.; Chachar, Z.; Tu, P. The Power of Magnesium: Unlocking the Potential for Increased Yield, Quality, and Stress Tolerance of Horticultural Crops. Frontiers in Plant Science 2023, 14, 1285512. [CrossRef]
- Karimi, R.; Koulivand, M.; Ollat, N. Soluble Sugars, Phenolic Acids and Antioxidant Capacity of Grape Berries as Affected by Iron and Nitrogen. Acta Physiol Plant 2019, 41, 117. [CrossRef]
- Yadav, R.B. Potential Benefits of Berries and Their Bioactive Compounds as Functional Food Component and Immune Boosting Food. In Immunity Boosting Functional Foods to Combat COVID-19; CRC Press, 2021; pp. 75–90.
- Nile, S.H.; Park, S.W. Edible Berries: Bioactive Components and Their Effect on Human Health. Nutrition 2014, 30, 134–144. [CrossRef]
- Oshunsanya, S.O.; Nwosu, N.J.; Li, Y. Abiotic Stress in Agricultural Crops Under Climatic Conditions. In Sustainable Agriculture, Forest and Environmental Management; Jhariya, M.K., Banerjee, A., Meena, R.S., Yadav, D.K., Eds.; Springer Singapore: Singapore, 2019; pp. 71–100 ISBN 978-981-13-6829-5.
- Liu, H.; Able, A.J.; Able, J.A. Priming Crops for the Future: Rewiring Stress Memory. Trends in plant science 2022, 27, 699–716. [CrossRef]
- Pagano, A.; Macovei, A.; Balestrazzi, A. Molecular Dynamics of Seed Priming at the Crossroads between Basic and Applied Research. Plant Cell Rep 2023, 42, 657–688. [CrossRef] [PubMed]
- Turgut-Kara, N.; Arikan, B.; Celik, H. Epigenetic Memory and Priming in Plants. Genetica 2020, 148, 47–54. [CrossRef]
- Hönig, M.; Roeber, V.M.; Schmülling, T.; Cortleven, A. Chemical Priming of Plant Defense Responses to Pathogen Attacks. Frontiers in Plant Science 2023, 14, 1146577. [CrossRef]
- Karami, A.A.; Abdossi, V.; Ghanbari Jahromi, M.; Aboutalebi Jahromi, A. Optimizing Blueberry (Vaccinium Corymbosum L.) Yield with Strategic Foliar Application of Putrescin and Spermidine at Key Growth Stages through Biochemical and Anatomical Changes. Frontiers in Plant Science 2025, 16, 1564026. [CrossRef]
- Gutiérrez-Albanchez, E.; Gradillas, A.; García, A.; García-Villaraco, A.; Gutierrez-Mañero, F.J.; Ramos-Solano, B. Elicitation with Bacillus QV15 Reveals a Pivotal Role of F3H on Flavonoid Metabolism Improving Adaptation to Biotic Stress in Blackberry. PloS one 2020, 15, e0232626. [CrossRef] [PubMed]
- Kazakov, P.; Alseekh, S.; Ivanova, V.; Gechev, T. Biostimulant-Based Molecular Priming Improves Crop Quality and Enhances Yield of Raspberry and Strawberry Fruits. Metabolites 2024, 14, 594. [CrossRef] [PubMed]
- Salama, A.-M.; Abdelsalam, M.A.; Rehan, M.; Elansary, M.; El-Shereif, A. Anthocyanin Accumulation and Its Corresponding Gene Expression, Total Phenol, Antioxidant Capacity, and Fruit Quality of ‘Crimson Seedless’ Grapevine (Vitis Vinifera L.) in Response to Grafting and Pre-Harvest Applications. Horticulturae 2023, 9, 1001. [CrossRef]
- Wang, K.; Wu, D.; Bo, Z.; Chen, S.I.; Wang, Z.; Zheng, Y.; Fang, Y. Regulation of Redox Status Contributes to Priming Defense against Botrytis Cinerea in Grape Berries Treated with β-Aminobutyric Acid. Scientia Horticulturae 2019, 244, 352–364. [CrossRef]
- Zhu, B.; Guo, P.; Wu, S.; Yang, Q.; He, F.; Gao, X.; Zhang, Y.; Xiao, J. A Better Fruit Quality of Grafted Blueberry than Own-Rooted Blueberry Is Linked to Its Anatomy. Plants 2024, 13, 625. [CrossRef] [PubMed]
- Habibi, F.; Liu, T.; Folta, K.; Sarkhosh, A. Physiological, Biochemical, and Molecular Aspects of Grafting in Fruit Trees. Horticulture Research 2022, 9, uhac032. [CrossRef] [PubMed]
- Ulas, F.; Kılıç, F.N.; Ulas, A. Alleviate the Influence of Drought Stress by Using Grafting Technology in Vegetable Crops: A Review. Journal of Crop Health 2025, 77, 51. [CrossRef]
- Krishankumar, S.; Hunter, J.J.; Alyafei, M.; Souka, U.; Subramaniam, S.; Ayyagari, R.; Kurup, S.S.; Amiri, K. Influence of Different Scion-Rootstock Combinations on Sugars, Polyamines, Antioxidants and Malondialdehyde in Grafted Grapevines under Arid Conditions. Frontiers in Plant Science 2025, 16, 1559095. [CrossRef]
- Klimek, K.E.; Kapłan, M.; Maj, G.; Borkowska, A.; Słowik, T. Effect of ‘Regent’Grapevine Rootstock Type on Energy Potential Parameters. Journal of Water and Land Development 2024. [CrossRef]
- Rugienius, R.; Vinskienė, J.; Andriūnaitė, E.; Morkūnaitė-Haimi, Š.; Juhani-Haimi, P.; Graham, J. Genomic Design of Abiotic Stress-Resistant Berries. In Genomic Designing for Abiotic Stress Resistant Fruit Crops; Kole, C., Ed.; Springer International Publishing: Cham, 2022; pp. 197–249 ISBN 978-3-031-09874-1.
- Farajpour, M.; Ahmadi, S.R.Z.; Aouei, M.T.; Ramezanpour, M.R.; Sadat-Hosseini, M.; Hajivand, S. Nutritional and Antioxidant Profiles of Blackberry and Raspberry Genotypes. BMC Plant Biol 2025, 25, 380. [CrossRef]
- Lebedev, V.G.; Lebedeva, T.N.; Vidyagina, E.O.; Sorokopudov, V.N.; Popova, A.A.; Shestibratov, K.A. Relationship between Phenolic Compounds and Antioxidant Activity in Berries and Leaves of Raspberry Genotypes and Their Genotyping by SSR Markers. Antioxidants 2022, 11, 1961. [CrossRef]
- Krishna, P.; Pandey, G.; Thomas, R.; Parks, S. Improving Blueberry Fruit Nutritional Quality through Physiological and Genetic Interventions: A Review of Current Research and Future Directions. Antioxidants 2023, 12, 810. [CrossRef]
- Vorsa, N.; Zalapa, J. Domestication, Genetics, and Genomics of the American Cranberry. In Plant Breeding Reviews; Goldman, I., Ed.; Wiley, 2019; pp. 279–315 ISBN 978-1-119-61673-3.
- Thole, V.; Bassard, J.-E.; Ramírez-González, R.; Trick, M.; Ghasemi Afshar, B.; Breitel, D.; Hill, L.; Foito, A.; Shepherd, L.; Freitag, S.; et al. RNA-Seq, de Novo Transcriptome Assembly and Flavonoid Gene Analysis in 13 Wild and Cultivated Berry Fruit Species with High Content of Phenolics. BMC Genomics 2019, 20, 995. [CrossRef]
- Zhu, S.; Zhang, X.; Ren, C.; Xu, X.; Comes, H.P.; Jiang, W.; Fu, C.; Feng, H.; Cai, L.; Hong, D.; et al. Chromosome--level Reference Genome of Tetrastigma Hemsleyanum (Vitaceae) Provides Insights into Genomic Evolution and the Biosynthesis of Phenylpropanoids and Flavonoids. The Plant Journal 2023, 114, 805–823. [CrossRef] [PubMed]
- Chen, Z.; Jiang, J.; Shu, L.; Li, X.; Huang, J.; Qian, B.; Wang, X.; Li, X.; Chen, J.; Xu, H. Combined Transcriptomic and Metabolic Analyses Reveal Potential Mechanism for Fruit Development and Quality Control of Chinese Raspberry (Rubus Chingii Hu). Plant Cell Rep 2021, 40, 1923–1946. [CrossRef]
- Chizk, T.M.; Clark, J.R.; Johns, C.; Nelson, L.; Ashrafi, H.; Aryal, R.; Worthington, M.L. Genome-Wide Association Identifies Key Loci Controlling Blackberry Postharvest Quality. Frontiers in Plant Science 2023, 14, 1182790. [CrossRef]
- Omori, M.; Yamane, H.; Osakabe, K.; Osakabe, Y.; Tao, R. Targeted Mutagenesis of CENTRORADIALIS Using CRISPR/Cas9 System through the Improvement of Genetic Transformation Efficiency of Tetraploid Highbush Blueberry. The Journal of Horticultural Science and Biotechnology 2021, 96, 153–161. [CrossRef]
- Polashock, J.; Zelzion, E.; Fajardo, D.; Zalapa, J.; Georgi, L.; Bhattacharya, D.; Vorsa, N. The American Cranberry: First Insights into the Whole Genome of a Species Adapted to Bog Habitat. BMC Plant Biol 2014, 14, 165. [CrossRef]
- Retamales, J.B.; Hancock, J.F. Blueberries; Cabi, 2018; Vol. 27;
- Lyrene, P.M. Value of Various Taxa in Breeding Tetraploid Blueberries in Florida. Euphytica 1997, 94, 15–22, doi:10.1023/A:1002903609446. [CrossRef]
- Crowl, A.A.; Fritsch, P.W.; Tiley, G.P.; Lynch, N.P.; Ranney, T.G.; Ashrafi, H.; Manos, P.S. A First Complete Phylogenomic Hypothesis for Diploid Blueberries ( Vaccinium Section Cyanococcus ). American J of Botany 2022, 109, 1596–1606. [CrossRef]
- Rowland, L.J.; Ogden, E.L.; Ballington, J.R. Relationships among Blueberry Species within the Section Cyanococcus of the Vaccinium Genus Based on EST-PCR Markers. Can. J. Plant Sci. 2022, 102, 744–748. [CrossRef]
- Colle, M.; Leisner, C.P.; Wai, C.M.; Ou, S.; Bird, K.A.; Wang, J.; Wisecaver, J.H.; Yocca, A.E.; Alger, E.I.; Tang, H. Haplotype-Phased Genome and Evolution of Phytonutrient Pathways of Tetraploid Blueberry. GigaScience 2019, 8, giz012. [CrossRef]
- Mohammadpour, S.; Karimzadeh, G.; Ghaffari, S.M. Karyomorphology, Genome Size, and Variation of Antioxidant in Twelve Berry Species from Iran. Caryologia 2022, 75, 133–148. [CrossRef]
- Ballington, J.R. The Role of Interspecific Hybridization in Blueberry Improvement. In Proceedings of the IX International Vaccinium Symposium 810; 2008; pp. 49–60.
- Araniti, F.; Baron, G.; Ferrario, G.; Pesenti, M.; Della Vedova, L.; Prinsi, B.; Sacchi, G.A.; Aldini, G.; Espen, L. Chemical Profiling and Antioxidant Potential of Berries from Six Blueberry Genotypes Harvested in the Italian Alps in 2020: A Comparative Biochemical Pilot Study. Agronomy 2025, 15, 262. [CrossRef]
- Yu, J.; Hulse-Kemp, A.M.; Babiker, E.; Staton, M. High-Quality Reference Genome and Annotation Aids Understanding of Berry Development for Evergreen Blueberry (Vaccinium Darrowii). Horticulture Research 2021, 8. [CrossRef] [PubMed]
- Manzanero, B.R.; Kulkarni, K.P.; Vorsa, N.; Reddy, U.K.; Natarajan, P.; Elavarthi, S.; Iorizzo, M.; Melmaiee, K. Genomic and Evolutionary Relationships among Wild and Cultivated Blueberry Species. BMC Plant Biol 2023, 23, 126. [CrossRef]
- Debnath, S.C.; Bhatt, D.; Goyali, J.C. DNA-Based Molecular Markers and Antioxidant Properties to Study Genetic Diversity and Relationship Assessment in Blueberries. Agronomy 2023, 13, 1518. [CrossRef]
- Colle, M.; Leisner, C.P.; Wai, C.M.; Ou, S.; Bird, K.A.; Wang, J.; Wisecaver, J.H.; Yocca, A.E.; Alger, E.I.; Tang, H. Haplotype-Phased Genome and Evolution of Phytonutrient Pathways of Tetraploid Blueberry. GigaScience 2019, 8, giz012. [CrossRef]
- Rossi, G.; Woods, F.M.; Leisner, C.P. Quantification of Total Phenolic, Anthocyanin, and Flavonoid Content in a Diverse Panel of Blueberry Cultivars and Ecotypes. HortScience 2022, 57, 901–909. [CrossRef]
- Wang, S.Y.; Chen, C.-T.; Sciarappa, W.; Wang, C.Y.; Camp, M.J. Fruit Quality, Antioxidant Capacity, and Flavonoid Content of Organically and Conventionally Grown Blueberries. J. Agric. Food Chem. 2008, 56, 5788–5794. [CrossRef]
- Scalzo, J.; Stevenson, D.; Hedderley, D. Blueberry Estimated Harvest from Seven New Cultivars: Fruit and Anthocyanins. Food chemistry 2013, 139, 44–50. [CrossRef]
- Connor, A.M.; Luby, J.J.; Tong, C.B.; Finn, C.E.; Hancock, J.F. Genotypic and Environmental Variation in Antioxidant Activity, Total Phenolic Content, and Anthocyanin Content among Blueberry Cultivars. Journal of the American Society for Horticultural Science 2002, 127, 89–97. [CrossRef]
- Kalt, W.; McDonald, J.E.; Ricker, R.D.; Lu, X. Anthocyanin Content and Profile within and among Blueberry Species. Can. J. Plant Sci. 1999, 79, 617–623. [CrossRef]
- Stevenson, D.; Scalzo, J. Anthocyanin Composition and Content of Blueberries from around the World. Journal of Berry Research 2012, 2, 179–189. [CrossRef]
- Chen, L.; Liu, Y.; Liu, H.; Kang, L.; Geng, J.; Gai, Y.; Ding, Y.; Sun, H.; Li, Y. Identification and Expression Analysis of MATE Genes Involved in Flavonoid Transport in Blueberry Plants. PloS one 2015, 10, e0118578. [CrossRef]
- Zhang, Y.; Zhang, Z.; Guo, S.; Qu, P.; Liu, J.; Cheng, C. Characterization of Blueberry Glutathione S-Transferase (GST) Genes and Functional Analysis of VcGSTF8 Reveal the Role of ‘MYB/bHLH-GSTF’Module in Anthocyanin Accumulation. Industrial Crops and Products 2024, 218, 119006. [CrossRef]
- Liu, Y.; Wang, Y.; Pei, J.; Li, Y.; Sun, H. Genome-Wide Identification and Characterization of COMT Gene Family during the Development of Blueberry Fruit. BMC Plant Biol 2021, 21, 5. [CrossRef] [PubMed]
- Yang, B.; Song, Y.; Li, Y.; Wang, X.; Guo, Q.; Zhou, L.; Zhang, Y.; Zhang, C. Key Genes for Phenylpropanoid Metabolite Biosynthesis during Half-Highbush Blueberry (Vaccinium angustifolium×Vaccinium Corymbosum) Fruit Development. JBR 2022, 12, 297–311. [CrossRef]
- Günther, C.S.; Dare, A.P.; McGhie, T.K.; Deng, C.; Lafferty, D.J.; Plunkett, B.J.; Grierson, E.R.; Turner, J.L.; Jaakola, L.; Albert, N.W. Spatiotemporal Modulation of Flavonoid Metabolism in Blueberries. Frontiers in plant science 2020, 11, 545. [CrossRef]
- Ferrão, L.F.V.; Sater, H.; Lyrene, P.; Amadeu, R.R.; Sims, C.A.; Tieman, D.M.; Munoz, P.R. Terpene Volatiles Mediates the Chemical Basis of Blueberry Aroma and Consumer Acceptability. Food research international 2022, 158, 111468. [CrossRef]
- Chu, L.; Du, Q.; Li, A.; Liu, G.; Wang, H.; Cui, Q.; Liu, Z.; Liu, H.; Lu, Y.; Deng, Y. Integrative Transcriptomic and Metabolomic Analyses of the Mechanism of Anthocyanin Accumulation and Fruit Coloring in Three Blueberry Varieties of Different Colors. Horticulturae 2024, 10, 105. [CrossRef]
- Zhang, L.; Ma, J. Promoter Cloning of VcCHS Gene from Blueberries and Selection of Its Transcription Factors. Hortic. Environ. Biotechnol. 2025, 66, 983–992. [CrossRef]
- Mengist, M.F.; Grace, M.H.; Mackey, T.; Munoz, B.; Pucker, B.; Bassil, N.; Luby, C.; Ferruzzi, M.; Lila, M.A.; Iorizzo, M. Dissecting the Genetic Basis of Bioactive Metabolites and Fruit Quality Traits in Blueberries (Vaccinium Corymbosum L.). Frontiers in Plant Science 2022, 13, 964656. [CrossRef]
- Montanari, S.; Thomson, S.; Cordiner, S.; Günther, C.S.; Miller, P.; Deng, C.H.; McGhie, T.; Knäbel, M.; Foster, T.; Turner, J. High-Density Linkage Map Construction in an Autotetraploid Blueberry Population and Detection of Quantitative Trait Loci for Anthocyanin Content. Frontiers in Plant Science 2022, 13, 965397. [CrossRef] [PubMed]
- Meng, Q.; Manghwar, H.; Hu, W. Study on Supergenus Rubus L.: Edible, Medicinal, and Phylogenetic Characterization. Plants 2022, 11, 1211. [CrossRef] [PubMed]
- Foster, T.M.; Bassil, N.V.; Dossett, M.; Leigh Worthington, M.; Graham, J. Genetic and Genomic Resources for Rubus Breeding: A Roadmap for the Future. Horticulture research 2019, 6. [CrossRef]
- Graham, J.; Brennan, R. Introduction to the Rubus Genus. In Raspberry; Graham, J., Brennan, R., Eds.; Springer International Publishing: Cham, 2018; pp. 1–16 ISBN 978-3-319-99030-9.
- Davik, J.; Røen, D.; Lysøe, E.; Buti, M.; Rossman, S.; Alsheikh, M.; Aiden, E.L.; Dudchenko, O.; Sargent, D.J. A Chromosome-Level Genome Sequence Assembly of the Red Raspberry (Rubus Idaeus L.). Plos one 2022, 17, e0265096. [CrossRef]
- Kikuchi, S.; Mimura, M.; Naruhashi, N.; Setsuko, S.; Suzuki, W. Phylogenetic Inferences Using Nuclear Ribosomal ITS and Chloroplast Sequences Provide Insights into the Biogeographic Origins, Diversification Timescales and Trait Evolution of Rubus in the Japanese Archipelago. Plant Syst Evol 2022, 308, 20. [CrossRef]
- Zhang, H.; Yan, M.; Li, L.; Jiang, Z.; Xiong, Y.; Wang, Y.; Akogwu, C.O.; Tolulope, O.M.; Zhou, H.; Sun, Y.; et al. Assembly and Comparative Analysis of the Complete Mitochondrial Genome of Red Raspberry (Rubus Idaeus L.) Revealing Repeat-Mediated Recombination and Gene Transfer. BMC Plant Biol 2025, 25, 85. [CrossRef] [PubMed]
- VanBuren, R.; Wai, C.M.; Colle, M.; Wang, J.; Sullivan, S.; Bushakra, J.M.; Liachko, I.; Vining, K.J.; Dossett, M.; Finn, C.E. A near Complete, Chromosome-Scale Assembly of the Black Raspberry (Rubus Occidentalis) Genome. Gigascience 2018, 7, giy094. [CrossRef]
- Graham, J.; Brennan, R. Introduction to the Rubus Genus. In Raspberry; Graham, J., Brennan, R., Eds.; Springer International Publishing: Cham, 2018; pp. 1–16 ISBN 978-3-319-99030-9.
- Shoukat, S.; Mahmudiono, T.; Al-Shawi, S.G.; Abdelbasset, W.K.; Yasin, G.; Shichiyakh, R.A.; Iswanto, A.H.; Kadhim, A.J.; Kadhim, M.M.; Al–rekaby, H.Q. Determination of the Antioxidant and Mineral Contents of Raspberry Varieties. Food Science and Technology 2022, 42, e118521. [CrossRef]
- Toshima, S.; Fujii, M.; Hidaka, M.; Nakagawa, S.; Hirano, T.; Kunitak, H. Fruit Qualities of Interspecific Hybrid and First Backcross Generations between Red Raspberry and Rubus Parvifolius. Journal of the American Society for Horticultural Science 2021, 146, 445–451. [CrossRef]
- Guo, Z.; Fan, Z.; Li, X.; Du, H.; Wu, Z.; Li, T.; Yang, G. Combined Analysis of SRAP and SSR Markers Reveals Genetic Diversity and Phylogenetic Relationships in Raspberry (Rubus Idaeus L.). Agronomy 2025, 15, 1492. [CrossRef]
- Connor, A.M.; Stephens, M.J.; Hall, H.K.; Alspach, P.A. Variation and Heritabilities of Antioxidant Activity and Total Phenolic Content Estimated from a Red Raspberry Factorial Experiment. Journal of the American Society for Horticultural Science 2005, 130, 403–411. [CrossRef]
- Kostecka-Gugała, A.; Ledwożyw-Smoleń, I.; Augustynowicz, J.; Wyżgolik, G.; Kruczek, M.; Kaszycki, P. Antioxidant Properties of Fruits of Raspberryand Blackberry Grown in Central Europe. Open Chemistry 2015, 13, 000010151520150143. [CrossRef]
- Graham, J.; Woodhead, M. Raspberries and Blackberries: The Genomics of Rubus. In Genetics and Genomics of Rosaceae; Folta, K.M., Gardiner, S.E., Eds.; Springer New York: New York, NY, 2009; pp. 507–524 ISBN 978-0-387-77490-9.
- Lebedev, V.G.; Lebedeva, T.N.; Vidyagina, E.O.; Sorokopudov, V.N.; Popova, A.A.; Shestibratov, K.A. Relationship between Phenolic Compounds and Antioxidant Activity in Berries and Leaves of Raspberry Genotypes and Their Genotyping by SSR Markers. Antioxidants 2022, 11, 1961. [CrossRef]
- Ljujić, J.; Sofrenić, I.; \DJor\djević, I.; Macura, P.; Tešević, V.; Vujisić, L.; An\djelković, B. Antioxidant Potential and Polyphenol Content of Five New Cultivars of Raspberries. Macedonian pharmaceutical bulletin 2022, 68, 47–48. [CrossRef]
- Frías-Moreno, M.N.; Parra-Quezada, R.Á.; Ruíz-Carrizales, J.; González-Aguilar, G.A.; Sepulveda, D.; Molina-Corral, F.J.; Jacobo-Cuellar, J.L.; Olivas, G.I. Quality, Bioactive Compounds and Antioxidant Capacity of Raspberries Cultivated in Northern Mexico. International Journal of Food Properties 2021, 24, 603–614. [CrossRef]
- Zheng, D.; Schröder, G.; Schröder, J.; Hrazdina, G. Molecular and Biochemical Characterization of Three Aromatic Polyketide Synthase Genes from Rubus Idaeus. Plant Mol Biol 2001, 46, 1–15. [CrossRef]
- Kassim, A.; Poette, J.; Paterson, A.; Zait, D.; McCallum, S.; Woodhead, M.; Smith, K.; Hackett, C.; Graham, J. Environmental and Seasonal Influences on Red Raspberry Anthocyanin Antioxidant Contents and Identification of Quantitative Traits Loci (QTL). Molecular Nutrition Food Res 2009, 53, 625–634. [CrossRef] [PubMed]
- Huang, X.; Wu, Y.; Zhang, S.; Yang, H.; Wu, W.; Lyu, L.; Li, W. Changes in Antioxidant Substances and Antioxidant Enzyme Activities in Raspberry Fruits at Different Developmental Stages. Scientia Horticulturae 2023, 321, 112314. [CrossRef]
- Woodhead, M.; Weir, A.; Smith, K.; McCallum, S.; MacKenzie, K.; Graham, J. Functional Markers for Red Raspberry. Journal of the American Society for Horticultural Science 2010, 135, 418–427. [CrossRef]
- Bushakra, J.M.; Krieger, C.; Deng, D.; Stephens, M.J.; Allan, A.C.; Storey, R.; Symonds, V.V.; Stevenson, D.; McGhie, T.; Chagné, D.; et al. QTL Involved in the Modification of Cyanidin Compounds in Black and Red Raspberry Fruit. Theor Appl Genet 2013, 126, 847–865. [CrossRef] [PubMed]
- Bushakra, J.M.; Dossett, M.; Carter, K.A.; Vining, K.J.; Lee, J.C.; Bryant, D.W.; VanBuren, R.; Lee, J.; Mockler, T.C.; Finn, C.E.; et al. Characterization of Aphid Resistance Loci in Black Raspberry (Rubus Occidentalis L.). Mol Breeding 2018, 38, 83. [CrossRef]
- McCallum, S.; Simpson, C.; Graham, J. QTL Mapping and Marker Assisted Breeding in Rubus Spp. In Raspberry; Graham, J., Brennan, R., Eds.; Springer International Publishing: Cham, 2018; pp. 121–144 ISBN 978-3-319-99030-9.
- Salgado, A.; Clark, J.R. Evaluation of a New Type of Firm and Reduced Reversion Blackberry: Crispy Genotypes. In Proceedings of the XI International Rubus and Ribes Symposium 1133; 2015; pp. 405–410.
- Finn, C.E.; Clark, J.R. Blackberry. In Fruit Breeding; Badenes, M.L., Byrne, D.H., Eds.; Springer US: Boston, MA, 2012; pp. 151–190 ISBN 978-1-4419-0762-2.
- Van de Beek, A.; Widrlechner, M.P. North American Species of Rubus L.(Rosaceae) Described from European Botanical Gardens (1789-1823). Adansonia 2021, 43, 67–98. [CrossRef]
- Moreno-Medina, B.L.; Casierra-Posada, F.; Cutler, J. Phytochemical Composition and Potential Use of Rubus Species. Gesunde Pflanzen 2018, 70, 65–74. [CrossRef]
- Agar, G.; Halasz, J.; Ercisli, S. Genetic Relationships among Wild and Cultivated Blackberries ( Rubus Caucasicus L.) Based on Amplified Fragment Length Polymorphism Markers. Plant Biosystems - An International Journal Dealing with all Aspects of Plant Biology 2011, 145, 347–352. [CrossRef]
- Llauradó Maury, G.; Méndez Rodríguez, D.; Hendrix, S.; Escalona Arranz, J.C.; Fung Boix, Y.; Pacheco, A.O.; García Díaz, J.; Morris-Quevedo, H.J.; Ferrer Dubois, A.; Aleman, E.I. Antioxidants in Plants: A Valorization Potential Emphasizing the Need for the Conservation of Plant Biodiversity in Cuba. Antioxidants 2020, 9, 1048. [CrossRef]
- Croge, C.P.; Cuquel, F.L.; Pintro, P.T.; Biasi, L.A.; De Bona, C.M. Antioxidant Capacity and Polyphenolic Compounds of Blackberries Produced in Different Climates. HortScience 2019, 54, 2209–2213. [CrossRef]
- Memete, A.R.; Sărac, I.; Teusdea, A.C.; Budău, R.; Bei, M.; Vicas, S.I. Bioactive Compounds and Antioxidant Capacity of Several Blackberry (Rubus Spp.) Fruits Cultivars Grown in Romania. Horticulturae 2023, 9, 556. [CrossRef]
- Martins, A.; Barros, L.; Carvalho, A.M.; Santos-Buelga, C.; Fernandes, I.P.; Barreiro, F.; Ferreira, I.C. Phenolic Extracts of Rubus Ulmifolius Schott Flowers: Characterization, Microencapsulation and Incorporation into Yogurts as Nutraceutical Sources. Food & function 2014, 5, 1091–1100.
- Huang, X.; Wu, Y.; Zhang, S.; Yang, H.; Wu, W.; Lyu, L.; Li, W. Variation in Antioxidant Enzyme Activity and Key Gene Expression during Fruit Development of Blackberry and Blackberry–Raspberry Hybrids. Food Bioscience 2023, 54, 102892. [CrossRef]
- Vogt, T. Phenylpropanoid Biosynthesis. Molecular plant 2010, 3, 2–20. [CrossRef]
- Dao, T.T.H.; Linthorst, H.J.M.; Verpoorte, R. Chalcone Synthase and Its Functions in Plant Resistance. Phytochem Rev 2011, 10, 397–412. [CrossRef]
- Jez, J.M.; Bowman, M.E.; Dixon, R.A.; Noel, J.P. Structure and Mechanism of the Evolutionarily Unique Plant Enzyme Chalcone Isomerase. Nature structural biology 2000, 7, 786–791. [PubMed]
- Petit, P.; Granier, T.; d’Estaintot, B.L.; Manigand, C.; Bathany, K.; Schmitter, J.-M.; Lauvergeat, V.; Hamdi, S.; Gallois, B. Crystal Structure of Grape Dihydroflavonol 4-Reductase, a Key Enzyme in Flavonoid Biosynthesis. Journal of molecular biology 2007, 368, 1345–1357. [CrossRef] [PubMed]
- Saito, K.; Yonekura-Sakakibara, K.; Nakabayashi, R.; Higashi, Y.; Yamazaki, M.; Tohge, T.; Fernie, A.R. The Flavonoid Biosynthetic Pathway in Arabidopsis: Structural and Genetic Diversity. Plant Physiology and Biochemistry 2013, 72, 21–34. [CrossRef]
- Kobayashi, S.; Ishimaru, M.; Ding, C.K.; Yakushiji, H.; Goto, N. Comparison of UDP-Glucose: Flavonoid 3-O-Glucosyltransferase (UFGT) Gene Sequences between White Grapes (Vitis Vinifera) and Their Sports with Red Skin. Plant Science 2001, 160, 543–550. [CrossRef] [PubMed]
- Br\uuna, T.; Aryal, R.; Dudchenko, O.; Sargent, D.J.; Mead, D.; Buti, M.; Cavallini, A.; Hytönen, T.; Andres, J.; Pham, M. A Chromosome-Length Genome Assembly and Annotation of Blackberry (Rubus Argutus, Cv.”Hillquist”). G3 2023, 13, jkac289. [CrossRef]
- Paudel, D.; Parrish, S.B.; Peng, Z.; Parajuli, S.; Deng, Z. A Chromosome-Scale and Haplotype-Resolved Genome Assembly of Tetraploid Blackberry (Rubus L. Subgenus Rubus Watson). Horticulture Research 2025, 12, uhaf052. [CrossRef]
- Worthington, M.; Chizk, T.M.; Johns, C.A.; Nelson, L.D.; Silva, A.; Godwin, C.; Clark, J.R. Advances in Molecular Breeding of Blackberries in the Arkansas Fruit Breeding Program. In Proceedings of the XIII International Rubus and Ribes Symposium 1388; 2023; pp. 85–92.
- Naithani, S.; Deng, C.H.; Sahu, S.K.; Jaiswal, P. Exploring Pan-Genomes: An Overview of Resources and Tools for Unraveling Structure, Function, and Evolution of Crop Genes and Genomes. Biomolecules 2023, 13, 1403. [CrossRef]
- Chizk, T.M.; Clark, J.R.; Johns, C.; Nelson, L.; Ashrafi, H.; Aryal, R.; Worthington, M.L. Genome-Wide Association Identifies Key Loci Controlling Blackberry Postharvest Quality. Frontiers in Plant Science 2023, 14, 1182790. [CrossRef]
- Rodriguez-Bonilla, L.; Williams, K.A.; Rodriguez Bonilla, F.; Matusinec, D.; Maule, A.; Coe, K.; Wiesman, E.; Diaz-Garcia, L.; Zalapa, J. The Genetic Diversity of Cranberry Crop Wild Relatives, Vaccinium Macrocarpon Aiton and V. Oxycoccos L., in the US, with Special Emphasis on National Forests. Plants 2020, 9, 1446. [CrossRef] [PubMed]
- Schlautman, B.; Covarrubias-Pazaran, G.; Fajardo, D.; Steffan, S.; Zalapa, J. Discriminating Power of Microsatellites in Cranberry Organelles for Taxonomic Studies in Vaccinium and Ericaceae. Genet Resour Crop Evol 2017, 64, 451–466. [CrossRef]
- Zhidkin, R.R.; Matveeva, T.V. Phylogeny Problems of the Genus Vaccinium L. and Ways to Solve Them. Ecological genetics 2022, 20, 151–164. [CrossRef]
- Diaz-Garcia, L.; Covarrubias-Pazaran, G.; Johnson-Cicalese, J.; Vorsa, N.; Zalapa, J. Genotyping-by-Sequencing Identifies Historical Breeding Stages of the Recently Domesticated American Cranberry. Frontiers in Plant Science 2020, 11, 607770. [CrossRef] [PubMed]
- Urbstaite, R.; Raudone, L.; Janulis, V. Phytogenotypic Anthocyanin Profiles and Antioxidant Activity Variation in Fruit Samples of the American Cranberry (Vaccinium Macrocarpon Aiton). Antioxidants 2022, 11, 250. [CrossRef]
- Diaz-Garcia, L.; Garcia-Ortega, L.F.; González-Rodríguez, M.; Delaye, L.; Iorizzo, M.; Zalapa, J. Chromosome-Level Genome Assembly of the American Cranberry (Vaccinium Macrocarpon Ait.) and Its Wild Relative Vaccinium Microcarpum. Frontiers in Plant Science 2021, 12, 633310. [CrossRef]
- Vorsa, N.; Johnson-Cicalese, J. American Cranberry. In Fruit Breeding; Badenes, M.L., Byrne, D.H., Eds.; Springer US: Boston, MA, 2012; pp. 191–223 ISBN 978-1-4419-0762-2.
- Debnath, S.C.; An, D. Antioxidant Properties and Structured Biodiversity in a Diverse Set of Wild Cranberry Clones. Heliyon 2019, 5. [CrossRef]
- Schlautman, B.; Covarrubias-Pazaran, G.; Rodriguez-Bonilla, L.; Hummer, K.; Bassil, N.; Smith, T.; Zalapa, J. Genetic Diversity and Cultivar Variants in the NCGR Cranberry (Vaccinium Macrocarpon Aiton) Collection. J Genet 2018, 97, 1339–1351. [CrossRef]
- Šedbarė, R.; Sprainaitytė, S.; Baublys, G.; Viskelis, J.; Janulis, V. Phytochemical Composition of Cranberry (Vaccinium Oxycoccos L.) Fruits Growing in Protected Areas of Lithuania. Plants 2023, 12, 1974. [CrossRef]
- Vvedenskaya, I.O.; Vorsa, N. Flavonoid Composition over Fruit Development and Maturation in American Cranberry, Vaccinium Macrocarpon Ait. Plant Science 2004, 167, 1043–1054. [CrossRef]
- Šedbarė, R.; Pašakinskienė, I.; Janulis, V. Changes in the Composition of Biologically Active Compounds during the Ripening Period in Fruit of Different Large Cranberry (Vaccinium Macrocarpon Aiton) Cultivars Grown in the Lithuanian Collection. Plants 2023, 12, 202. [CrossRef] [PubMed]
- Sun, J.; Liu, W.; Ma, H.; Marais, J.P.J.; Khoo, C.; Dain, J.A.; Rowley, D.C.; Seeram, N.P. Effect of Cranberry ( Vaccinium Macrocarpon ) Oligosaccharides on the Formation of Advanced Glycation End-Products. Journal of Berry Research 2016, 6, 149–158. [CrossRef] [PubMed]
- Gallardo, R.K.; Klingthong, P.; Zhang, Q.; Polashock, J.; Atucha, A.; Zalapa, J.; Rodriguez-Saona, C.; Vorsa, N.; Iorizzo, M. Breeding Trait Priorities of the Cranberry Industry in the United States and Canada. HortScience 2018, 53, 1467–1474. [CrossRef]
- Vorsa, N.; Zalapa, J. Domestication, Genetics, and Genomics of the American Cranberry. In Plant Breeding Reviews; Goldman, I., Ed.; Wiley, 2019; pp. 279–315 ISBN 978-1-119-61673-3.
- Debnath, S.C.; Siow, Y.L.; Petkau, J.; An, D.; Bykova, N.V. Molecular Markers and Antioxidant Activity in Berry Crops: Genetic Diversity Analysis. Can. J. Plant Sci. 2012, 92, 1121–1133. [CrossRef]
- Diaz-Garcia, L.; Covarrubias-Pazaran, G.; Johnson-Cicalese, J.; Vorsa, N.; Zalapa, J. Genotyping-by-Sequencing Identifies Historical Breeding Stages of the Recently Domesticated American Cranberry. Frontiers in Plant Science 2020, 11, 607770. [CrossRef]
- Dong, Y.; Duan, S.; Xia, Q.; Liang, Z.; Dong, X.; Margaryan, K.; Musayev, M.; Goryslavets, S.; Zdunić, G.; Bert, P.-F.; et al. Dual Domestications and Origin of Traits in Grapevine Evolution. Science 2023, 379, 892–901. [CrossRef]
- Grassi, F.; De Lorenzis, G. Back to the Origins: Background and Perspectives of Grapevine Domestication. International Journal of Molecular Sciences 2021, 22, 4518. [CrossRef]
- Guzmán-Ardiles, R.E.; Pegoraro, C.; Da Maia, L.C.; Costa de Oliveira, A. Genetic Changes in the Genus Vitis and the Domestication of Vine. Frontiers in Plant Science 2023, 13, 1019311. [CrossRef]
- Wolkovich, E.M.; García de Cortázar-Atauri, I.; Morales-Castilla, I.; Nicholas, K.A.; Lacombe, T. From Pinot to Xinomavro in the World’s Future Wine-Growing Regions. Nature Climate Change 2018, 8, 29–37. [CrossRef]
- Kapazoglou, A.; Gerakari, M.; Lazaridi, E.; Kleftogianni, K.; Sarri, E.; Tani, E.; Bebeli, P.J. Crop Wild Relatives: A Valuable Source of Tolerance to Various Abiotic Stresses. Plants 2023, 12, 328. [CrossRef] [PubMed]
- Mudge, K.; Janick, J.; Scofield, S.; Goldschmidt, E.E. A History of Grafting. Horticultural reviews 2009, 35, 437–493.
- Migicovsky, Z.; Sawler, J.; Money, D.; Eibach, R.; Miller, A.J.; Luby, J.J.; Jamieson, A.R.; Velasco, D.; Von Kintzel, S.; Warner, J.; et al. Genomic Ancestry Estimation Quantifies Use of Wild Species in Grape Breeding. BMC Genomics 2016, 17, 478. [CrossRef] [PubMed]
- Darwish, A.G.; Das, P.R.; Ismail, A.; Gajjar, P.; Balasubramani, S.P.; Sheikh, M.B.; Tsolova, V.; Sherif, S.M.; El-Sharkawy, I. Untargeted Metabolomics and Antioxidant Capacities of Muscadine Grape Genotypes during Berry Development. Antioxidants 2021, 10, 914. [CrossRef] [PubMed]
- Mendonca, P.; Darwish, A.G.; Tsolova, V.; El-Sharkawy, I.; Soliman, K.F. The Anticancer and Antioxidant Effects of Muscadine Grape Extracts on Racially Different Triple-Negative Breast Cancer Cells. Anticancer research 2019, 39, 4043–4053. [CrossRef] [PubMed]
- French The Grapevine Genome Sequence Suggests Ancestral Hexaploidization in Major Angiosperm Phyla. nature 2007, 449, 463–467. [CrossRef] [PubMed]
- Canaguier, A.; Grimplet, J.; Di Gaspero, G.; Scalabrin, S.; Duchêne, E.; Choisne, N.; Mohellibi, N.; Guichard, C.; Rombauts, S.; Le Clainche, I. A New Version of the Grapevine Reference Genome Assembly (12X. v2) and of Its Annotation (VCost. V3). Genomics data 2017, 14, 56. [CrossRef]
- Vitulo, N.; Forcato, C.; Carpinelli, E.C.; Telatin, A.; Campagna, D.; D’Angelo, M.; Zimbello, R.; Corso, M.; Vannozzi, A.; Bonghi, C.; et al. A Deep Survey of Alternative Splicing in Grape Reveals Changes in the Splicing Machinery Related to Tissue, Stress Condition and Genotype. BMC Plant Biol 2014, 14, 99. [CrossRef] [PubMed]
- Shi, X.; Cao, S.; Wang, X.; Huang, S.; Wang, Y.; Liu, Z.; Liu, W.; Leng, X.; Peng, Y.; Wang, N. The Complete Reference Genome for Grapevine (Vitis Vinifera L.) Genetics and Breeding. Horticulture Research 2023, 10, uhad061. [CrossRef]
- Chougule, K.; Tello-Ruiz, M.K.; Wei, S.; Olson, A.; Lu, Z.; Kumari, S.; Kumar, V.; Contreras-Moreira, B.; Naamati, G.; Dyer, S. Pan Genome Resources for Grapevine. In Proceedings of the XI International Symposium on Grapevine Physiology and Biotechnology 1390; 2021; pp. 257–266.
- Liu, Z.; Wang, N.; Su, Y.; Long, Q.; Peng, Y.; Shangguan, L.; Zhang, F.; Cao, S.; Wang, X.; Ge, M. Grapevine Pangenome Facilitates Trait Genetics and Genomic Breeding. Nature Genetics 2024, 56, 2804–2814. [CrossRef]
- Theine, J.; Holtgräwe, D.; Herzog, K.; Schwander, F.; Kicherer, A.; Hausmann, L.; Viehöver, P.; Töpfer, R.; Weisshaar, B. Transcriptomic Analysis of Temporal Shifts in Berry Development between Two Grapevine Cultivars of the Pinot Family Reveals Potential Genes Controlling Ripening Time. BMC Plant Biol 2021, 21, 327. [CrossRef]
- Zenoni, S.; Dal Santo, S.; Tornielli, G.B.; D’Incà, E.; Filippetti, I.; Pastore, C.; Allegro, G.; Silvestroni, O.; Lanari, V.; Pisciotta, A. Transcriptional Responses to Pre-Flowering Leaf Defoliation in Grapevine Berry from Different Growing Sites, Years, and Genotypes. Frontiers in plant science 2017, 8, 630. [CrossRef]
- Zenoni, S.; Amato, A.; D’Incà, E.; Guzzo, F.; Tornielli, G.B. Rapid Dehydration of Grape Berries Dampens the Post-Ripening Transcriptomic Program and the Metabolite Profile Evolution. Horticulture Research 2020, 7. [CrossRef] [PubMed]
- Savoi, S.; Wong, D.C.J.; Arapitsas, P.; Miculan, M.; Bucchetti, B.; Peterlunger, E.; Fait, A.; Mattivi, F.; Castellarin, S.D. Transcriptome and Metabolite Profiling Reveals That Prolonged Drought Modulates the Phenylpropanoid and Terpenoid Pathway in White Grapes (Vitis Vinifera L.). BMC Plant Biol 2016, 16, 67. [CrossRef]
- Savoi, S.; Santiago, A.; Orduña, L.; Matus, J.T. Transcriptomic and Metabolomic Integration as a Resource in Grapevine to Study Fruit Metabolite Quality Traits. Frontiers in Plant Science 2022, 13, 937927. [CrossRef] [PubMed]
- Yang, X.; Guo, Y.; Zhu, J.; Shi, G.; Niu, Z.; Liu, Z.; Li, K.; Guo, X. Associations between the 1-Deoxy-d-Xylulose-5-Phosphate Synthase Gene and Aroma in Different Grapevine Varieties. Genes Genom 2017, 39, 1059–1067. [CrossRef]
- Zhang, Y.; Liu, C.; Liu, X.; Wang, Z.; Wang, Y.; Zhong, G.; Li, S.; Dai, Z.; Liang, Z.; Fan, P. Basic Leucine Zipper Gene VvbZIP61 Is Expressed at a Quantitative Trait Locus for High Monoterpene Content in Grape Berries. Horticulture Research 2023, 10, uhad151. [CrossRef]
- Zhang, H.-M.; Lyu, X.-J.; Sun, Z.-Y.; Sun, Q.; Wang, Y.-C.; Sun, L.; Xu, H.-Y.; He, L.; Duan, C.-Q.; Pan, Q.-H. GWAS Identifies a Molecular Marker Cluster Associated with Monoterpenoids in Grapes. Horticulture Research 2025, uhaf144. [CrossRef]
- Wei, Y.; Wang, Y.; Meng, X.; Yao, X.; Xia, N.; Zhang, H.; Meng, N.; Duan, C.; Pan, Q. VviWRKY24 Promotes β-Damascenone Biosynthesis by Targeting VviNCED1 to Increase Abscisic Acid in Grape Berries. Horticulture Research 2025, 12, uhaf017. [CrossRef]
- Liu, W.; Mu, H.; Yuan, L.; Li, Y.; Li, Y.; Li, S.; Ren, C.; Duan, W.; Fan, P.; Dai, Z. VvBBX44 and VvMYBA1 Form a Regulatory Feedback Loop to Balance Anthocyanin Biosynthesis in Grape. Horticulture Research 2023, 10, uhad176. [CrossRef]
- Lai, G.; Fu, P.; He, L.; Che, J.; Wang, Q.; Lai, P.; Lu, J.; Lai, C. CRISPR/Cas9-Mediated CHS2 Mutation Provides a New Insight into Resveratrol Biosynthesis by Causing a Metabolic Pathway Shift from Flavonoids to Stilbenoids in Vitis Davidii Cells. Horticulture Research 2025, 12, uhae268. [CrossRef]
- Hou, H.; Li, Y.; Zhou, S.; Zhang, R.; Wang, Y.; Lei, L.; Yang, C.; Huang, S.; Xu, H.; Liu, X. Compositional Analysis of Grape Berries: Mapping the Global Metabolism of Grapes. Foods 2024, 13, 3716. [CrossRef]
- Tello, J.; Moffa, L.; Ferradás, Y.; Gasparro, M.; Chitarra, W.; Milella, R.A.; Nerva, L.; Savoi, S. Grapes: A Crop with High Nutraceuticals Genetic Diversity. In Compendium of Crop Genome Designing for Nutraceuticals; Springer, 2023; pp. 945–984.
- Park, M.; Darwish, A.G.; Elhag, R.I.; Tsolova, V.; Soliman, K.F.; El-Sharkawy, I. A Multi-Locus Genome-Wide Association Study Reveals the Genetics Underlying Muscadine Antioxidant in Berry Skin. Frontiers in Plant Science 2022, 13, 969301. [CrossRef]
- Swallah, M.S.; Sun, H.; Affoh, R.; Fu, H.; Yu, H. Antioxidant Potential Overviews of Secondary Metabolites (Polyphenols) in Fruits. International Journal of Food Science 2020, 2020, 1–8. [CrossRef] [PubMed]
- Zhan, Z.; Gao, Y.; Chen, Y.; Xu, M.; Wang, H.; Cheng, J.; Zhang, B. Comprehensive Metabolomic Profiling of Aroma-Related Volatiles in Table Grape Cultivars across Developmental Stages. Food Quality and Safety 2025, 9, fyaf013. [CrossRef]
- Šikuten, I.; Štambuk, P.; Marković, Z.; Tomaz, I.; Preiner, D. Grape Volatile Organic Compounds (VOCs)-Analysis, Biosynthesis and Profiling. Journal of experimental botany 2025, eraf082.
- Bosman, R.N.; Lashbrooke, J.G. Grapevine Mono-and Sesquiterpenes: Genetics, Metabolism, and Ecophysiology. Frontiers in Plant Science 2023, 14, 1111392. [CrossRef] [PubMed]
- Wen, Y.-Q.; Zhong, G.-Y.; Gao, Y.; Lan, Y.-B.; Duan, C.-Q.; Pan, Q.-H. Using the Combined Analysis of Transcripts and Metabolites to Propose Key Genes for Differential Terpene Accumulation across Two Regions. BMC Plant Biol 2015, 15, 240. [CrossRef]
- Emanuelli, F.; Battilana, J.; Costantini, L.; Le Cunff, L.; Boursiquot, J.-M.; This, P.; Grando, M.S. A Candidate Gene Association Study on Muscat Flavor in Grapevine (Vitis Vinifera L.). BMC Plant Biol 2010, 10, 241. [CrossRef]
- Duchêne, E.; Butterlin, G.; Claudel, P.; Dumas, V.; Jaegli, N.; Merdinoglu, D. A Grapevine (Vitis Vinifera L.) Deoxy-d-Xylulose Synthase Gene Colocates with a Major Quantitative Trait Loci for Terpenol Content. Theor Appl Genet 2009, 118, 541–552. [CrossRef]
- Battilana, J.; Costantini, L.; Emanuelli, F.; Sevini, F.; Segala, C.; Moser, S.; Velasco, R.; Versini, G.; Grando, M.S. The 1-Deoxy-d-Xylulose 5-Phosphate Synthase Gene Co-Localizes with a Major QTL Affecting Monoterpene Content in Grapevine. Theor Appl Genet 2009, 118, 653–669. [CrossRef] [PubMed]
- Battilana, J.; Emanuelli, F.; Gambino, G.; Gribaudo, I.; Gasperi, F.; Boss, P.K.; Grando, M.S. Functional Effect of Grapevine 1-Deoxy-D-Xylulose 5-Phosphate Synthase Substitution K284N on Muscat Flavour Formation. Journal of Experimental Botany 2011, 62, 5497–5508. [CrossRef] [PubMed]
- Mendes-Pinto, M.M. Carotenoid Breakdown Products the—Norisoprenoids—in Wine Aroma. Archives of biochemistry and biophysics 2009, 483, 236–245. [CrossRef]
- Stringer, S.J.; Marshall, D.A.; Perkins-Veazie, P. Nutraceutical Compound Concentrations of Muscadine (Vitis Rotundifolia Michx.) Grape Cultivars and Breeding Lines. In Proceedings of the II International Symposium on Human Health Effects of Fruits and Vegetables: FAVHEALTH 2007 841; 2007; pp. 553–556.
- Sandhu, A.K.; Gu, L. Antioxidant Capacity, Phenolic Content, and Profiling of Phenolic Compounds in the Seeds, Skin, and Pulp of Vitis Rotundifolia (Muscadine Grapes) As Determined by HPLC-DAD-ESI-MSn. J. Agric. Food Chem. 2010, 58, 4681–4692. [CrossRef] [PubMed]
- Darwish, A.G.; Das, P.R.; Ismail, A.; Gajjar, P.; Balasubramani, S.P.; Sheikh, M.B.; Tsolova, V.; Sherif, S.M.; El-Sharkawy, I. Untargeted Metabolomics and Antioxidant Capacities of Muscadine Grape Genotypes during Berry Development. Antioxidants 2021, 10, 914. [CrossRef]
- Ismail, A.; Darwish, A.G.; Park, M.; Gajjar, P.; Tsolova, V.; Soliman, K.F.; El-Sharkawy, I. Transcriptome Profiling during Muscadine Berry Development Reveals the Dynamic of Polyphenols Metabolism. Frontiers in Plant Science 2022, 12, 818071. [CrossRef]
- Nogales, J.; Canales, Á.; Jiménez--Barbero, J.; Serra, B.; Pingarrón, J.M.; García, J.L.; Díaz, E. Unravelling the Gallic Acid Degradation Pathway in Bacteria: The Gal Cluster from Pseudomonas Putida. Molecular Microbiology 2011, 79, 359–374. [CrossRef] [PubMed]
- Liu, W.; Mu, H.; Yuan, L.; Li, Y.; Li, Y.; Li, S.; Ren, C.; Duan, W.; Fan, P.; Dai, Z. VvBBX44 and VvMYBA1 Form a Regulatory Feedback Loop to Balance Anthocyanin Biosynthesis in Grape. Horticulture Research 2023, 10, uhad176. [CrossRef]
- Căpruciu, R. Resveratrol in Grapevine Components, Products and by-Products—A Review. Horticulturae 2025, 11, 111. [CrossRef]
- Guo, X.; Hu, J.; Yang, S.; Wang, D.; Wang, J. Genome-Wide Analysis of the F3’5’H Gene Family in Blueberry (Vaccinium Corymbosum L.) Provides Insights into the Regulation of Anthocyanin Biosynthesis. Phyton (0031-9457) 2023, 92. [CrossRef]
Figure 1.
Exploitation of berry crops antioxidant capacity through agronomical practices and molecular breeding methods utilization.
Figure 1.
Exploitation of berry crops antioxidant capacity through agronomical practices and molecular breeding methods utilization.
Figure 2.
A general model of the mevalonate (MVA) and methylerythritol phosphate (MEP) pathways, operating in the cytosol and plastid, respectively, generate the prenyldiphosphate precursors required for terpene biosynthesis; downstream metabolic products and respective enzymes are indicated. AACT, acetyl-CoA acetyltransferase; CDP-ME, 4-diphosphocytidyl-2-C-methyl-D-erythritol; CDP-MEP, CDPME 2-phosphate; CMK, 4-(cytidine 5′-diphospho)-2-C-methyl-D-erythritol kinase; DMAPP, dimethyl allyl diphosphate; DXP, 1-deoxy-D-xylulose 5-phosphate; DXS, DXP synthase; DXR, 1-deoxy-D-xylulose 5-phosphate reductoisomerase; FPP, farnesyl diphosphate; FPPS, FPP synthase; G3P, glyceraldehyde 3-phosphate; GGPP, geranylgeranyl diphosphate; GGPPS, GGPP synthase; GPP, geranyl diphosphate; GPPS, GPP synthase; HDR, hydroxymethylbutenyl diphosphate reductase; HDS, 4-hydroxy-3-methylbut-2-en-1-yl diphosphate synthase; HMBPP, (E)-4-hydroxy-3-methylbut-2-en-1-yl diphosphate; HMG-CoA, hydroxymethylglutaryl-CoA; HMGR, HMG-CoA reductase; HMGS, HMG-CoA synthase; IDI, isopentenyl pyrophosphate isomerase; IPP, isopentenyl diphosphate; MCT, 2-C-methyl-D-erythritol 4-phosphate cytidylyltransferase; MECPD, 2-C-methyl-D-erythritol 2,4-cyclodiphosphate; MECPS, MECPD synthase; MVK, mevalonate kinase; MPDC, mevalonate diphosphate decarboxylase; MVP, mevalonate 5-phosphate; MVPP, mevalonate 5- pyrophosphate; PMK, phosphomevalonate kinase; TPS, terpene synthase (modified from Bosman et al., [177]).
Figure 2.
A general model of the mevalonate (MVA) and methylerythritol phosphate (MEP) pathways, operating in the cytosol and plastid, respectively, generate the prenyldiphosphate precursors required for terpene biosynthesis; downstream metabolic products and respective enzymes are indicated. AACT, acetyl-CoA acetyltransferase; CDP-ME, 4-diphosphocytidyl-2-C-methyl-D-erythritol; CDP-MEP, CDPME 2-phosphate; CMK, 4-(cytidine 5′-diphospho)-2-C-methyl-D-erythritol kinase; DMAPP, dimethyl allyl diphosphate; DXP, 1-deoxy-D-xylulose 5-phosphate; DXS, DXP synthase; DXR, 1-deoxy-D-xylulose 5-phosphate reductoisomerase; FPP, farnesyl diphosphate; FPPS, FPP synthase; G3P, glyceraldehyde 3-phosphate; GGPP, geranylgeranyl diphosphate; GGPPS, GGPP synthase; GPP, geranyl diphosphate; GPPS, GPP synthase; HDR, hydroxymethylbutenyl diphosphate reductase; HDS, 4-hydroxy-3-methylbut-2-en-1-yl diphosphate synthase; HMBPP, (E)-4-hydroxy-3-methylbut-2-en-1-yl diphosphate; HMG-CoA, hydroxymethylglutaryl-CoA; HMGR, HMG-CoA reductase; HMGS, HMG-CoA synthase; IDI, isopentenyl pyrophosphate isomerase; IPP, isopentenyl diphosphate; MCT, 2-C-methyl-D-erythritol 4-phosphate cytidylyltransferase; MECPD, 2-C-methyl-D-erythritol 2,4-cyclodiphosphate; MECPS, MECPD synthase; MVK, mevalonate kinase; MPDC, mevalonate diphosphate decarboxylase; MVP, mevalonate 5-phosphate; MVPP, mevalonate 5- pyrophosphate; PMK, phosphomevalonate kinase; TPS, terpene synthase (modified from Bosman et al., [177]).
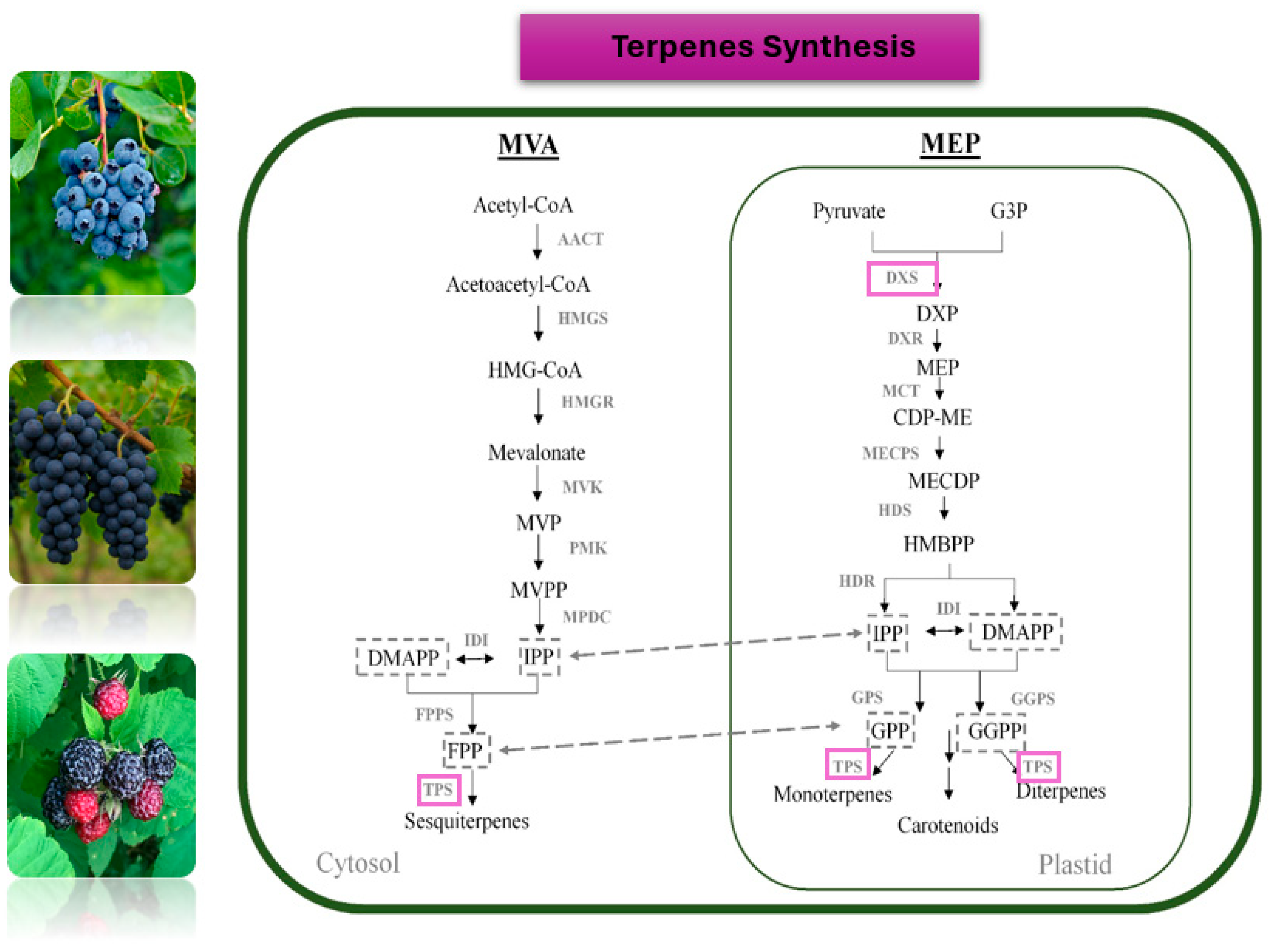
Figure 3.
A general model of the phenylpropanoid pathway; Targeted genome editing of chalcone synthase (CHS) using CRISPR/Cas9 reduces flux into the flavonoid branch of the phenylpropanoid pathway. This shift enhances substrate availability for stilbene synthase (STS), thereby promoting resveratrol biosynthesis and accumulation in transgenic cells. Both flavonoids and stilbenoids originate from the same precursor, p-coumaroyl-CoA, highlighting the competitive relationship between the two pathways.
Figure 3.
A general model of the phenylpropanoid pathway; Targeted genome editing of chalcone synthase (CHS) using CRISPR/Cas9 reduces flux into the flavonoid branch of the phenylpropanoid pathway. This shift enhances substrate availability for stilbene synthase (STS), thereby promoting resveratrol biosynthesis and accumulation in transgenic cells. Both flavonoids and stilbenoids originate from the same precursor, p-coumaroyl-CoA, highlighting the competitive relationship between the two pathways.
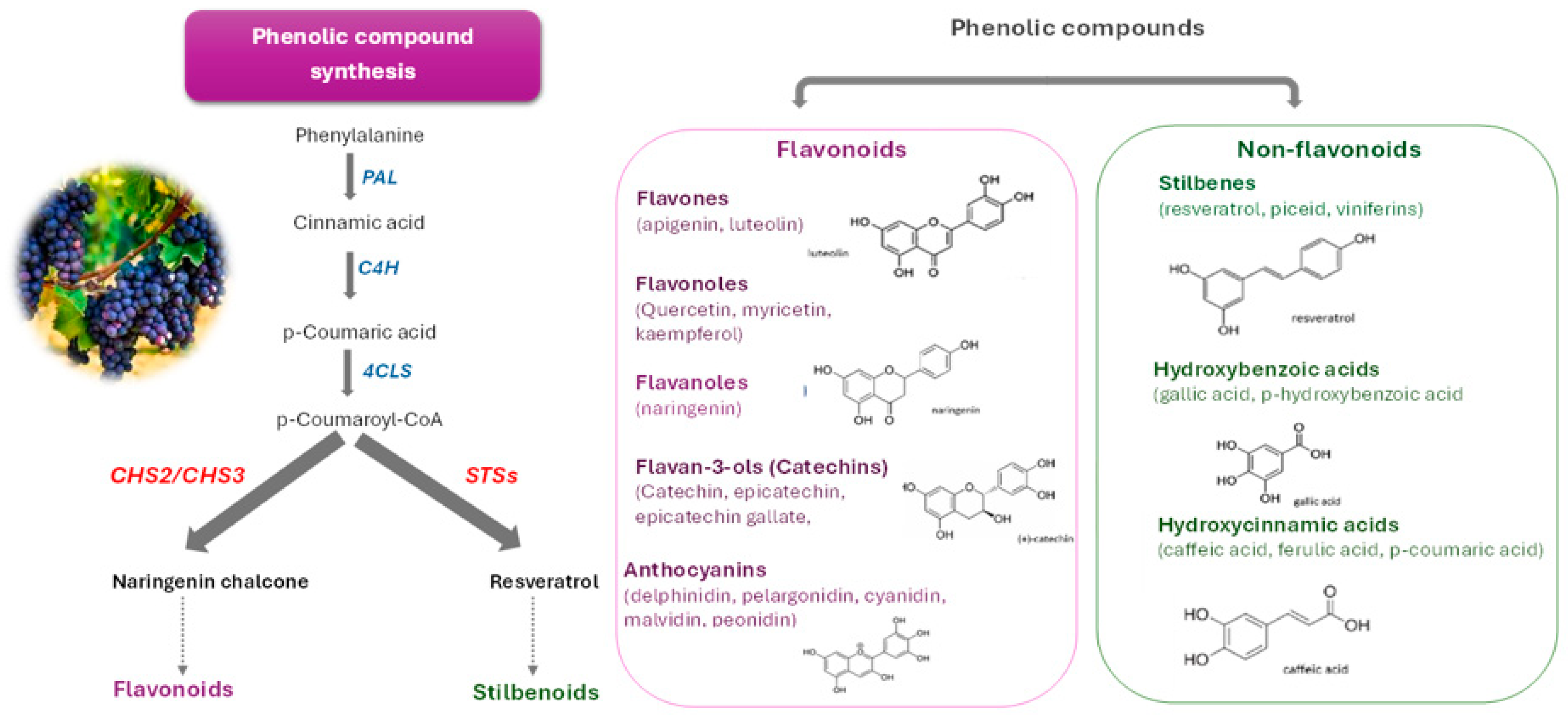
Table 1.
Origins, major producer countries, and growing requirements of key berry crops and grapes (FAOSTAT 2023).
Table 1.
Origins, major producer countries, and growing requirements of key berry crops and grapes (FAOSTAT 2023).
| Crop | Scientific name | Native origin | Global production | Main producers (tn) | Soil & climate requirements |
| Raspberry | Rubus idaeus | Europe, Northern Asia | ~940,000 tn | Russia (219k), Mexico (165k), Serbia (122k), Poland (118k), USA (100k) | Fertile, well-drained soils; cool winters for dormancy and flowering |
| Blueberry | Vaccinium spp. | North America | ~1.1 million tn | USA (300k), Peru (230k), Canada (165k), Chile (125k), Spain (70k) | Acidic soils; consistent moisture; frost protection |
| Cranberry | Vaccinium macrocarpon | Northeastern North America | ~470,000 tn | USA (300k), Canada (151k), Turkey (12k) | Acidic, water-saturated soils; benefit from regular field renewal |
| Grape | Vitis vinifera | Near East | >72.5 million tn | China (12.5m), Italy (8.1m), Spain 5.9m), USA (5.4m), France (6.2m) | Wide soil tolerance; good drainage; warm, dry summers |
Table 2.
Genes analyzed for their functional involvement in the antioxidant capacity of blueberries.
Table 2.
Genes analyzed for their functional involvement in the antioxidant capacity of blueberries.
| Enzyme Class | Gene | Functional Role | Reference |
| Multidrug and Toxic Compound Extrusion Transporters (MATE) | VcMATE2 | Facilitate anthocyanin movement across cellular membranes during ripening | Chen et al., 2015 [73] |
| VcMATE3 | |||
| VcMATE5 | |||
| VcMATE7 | |||
| VcMATE8 | |||
| VcMATE9 | |||
| Glutathione S-transferases (GSTs) | VcGSTF8 | Highly expressed during fruit ripening; strong correlation with anthocyanin accumulation |
Ζhang et al., 2024 [74] |
| VcGSTF20 | |||
| VcGSTF22 | |||
| O-methyltransferases (COMTs) | VcCOMT40 | Highly expressed during fruit development; involved in lignin biosynthesis and anthocyanin modification | Liu et al., 2021 [75] |
| VcCOMT92 | |||
| Flavonoid Biosynthesis Enzymes | VcCHS | Initiates flavonoid biosynthesis; upregulated during ripening | Chu et al., 2024; Zhang et al., 2025 [79,80] |
| VcCHI | Converts chalcone to naringenin; active in ripening fruit | ||
| VcF3H | Hydroxylates flavonoids contribute to anthocyanin diversity | ||
| VcF3′H | Adds hydroxyl group; modifies anthocyanin structure | ||
| VcF3′5′H | Adds hydroxyl groups for delphinidin-type anthocyanins | ||
| VcDFR | Converts dihydroflavonols to leucoanthocyanidins | ||
| VcANS | Synthesizes anthocyanidins; active during ripening | ||
| VcUFGT | Glycosylates anthocyanins for stability and vacuolar transport | ||
| Flavonol Synthase (FLS) | VcFLS homologs | Produces flavonols like quercetin and kaempferol in fruit skin | Günther et al., 2020 [77] |
| Leucoanthocyanidin Reductase (LAR) | VcLAR | Synthesizes catechin; contributes to proanthocyanidin biosynthesis | |
| Anthocyanidin Reductase (ANR) | VcANR1 | Produces epicatechin; active in fruit tissues |
Table 3.
Genes analyzed for their functional involvement in the antioxidant capacity of raspberries.
Table 3.
Genes analyzed for their functional involvement in the antioxidant capacity of raspberries.
| Enzyme Class | Gene | Functional Role | Reference |
| Polyketide synthase (PKS) | RiPKS1 | Encodes a typical chalcone synthase (CHS) active in naringenin production | Zheng et al., 2001 & Kassim et al., 2009 [100,101] |
| RiPKS2 | Characterized from raspberry cell cultures but found inactive due to specific amino acid exchanges | ||
| RiPKS3 | Produced mainly p-coumaroyltriacetic acid lactone (CTAL) | ||
| RiPKS5 | Catalyzes first step in flavonoid biosynthesis | Woodhead et.al, (2010) [103] | |
| 4-coumarate:CoA ligase (4CL) | Ri4CL1 | Catalyzes activation of 4-coumarate to CoA esters used in phenylpropanoid pathway | |
| Ri4CL2 | |||
| Ri4CL3 | |||
| Anthocyanidin synthase (ANS) | RiANS | Conversion of leucocyanidins to anthocyanidins | |
| Lipoxygenase (LOX) | RiLOX | Involved in lipoxygenase pathway | |
| Terpene synthase (TPS) | RiTerpSynth | Involved in monoterpene biosynthesis | |
| 3-hydroxy-3-methylglutaryl-coenzyme A (HMG CoA) reductase | ERubLR_SQ8.1_H09 | Involved in mevalonic acid pathway | |
| Isopentenyl-diphosphate delta-isomerase (IDI/IPI) | ERubLR_SQ13.1_F09 | ||
| Aconitase (ACO) | ERubLR_SQ13.2_C12 |
Table 4.
Genes analyzed for their functional involvement in the antioxidant capacity of blackberries.
Table 4.
Genes analyzed for their functional involvement in the antioxidant capacity of blackberries.
| Enzyme Class | Gene | Functional Role | References |
| Phenylalanine ammonia-lyase (PAL) | RuPAL | Converting L-phenylalanine to trans-cinnamic acid, providing precursors for flavonoids, lignin, and phenolic acids | Vogt et al., 2010 [117] |
| Chalcone synthase (CHS) | RuCHS | Catalyzes the formation of naringenin chalcone | Dao et al., 2011 [118] |
| Chalcone isomerase (CHI) | RuCHI | Catalyzes the stereospecific isomerization of chalcones into flavanones | Jez et al., 2000 [119] |
| Dihydroflavonol 4-reductase (DFR) | RuDFR | Reduces dihydroflavonols to leucoanthocyanidins | Petit et al., 2007 [120] |
| Anthocyanidin synthase (ANS) | RuANS | Oxidizes leucoanthocyanidins to anthocyanidins | Saito et al., 2013 [121] |
| UDP-glucose:flavonoid 3-O-glucosyltransferase (UFGT) | RuUFGT | Glycosylates unstable anthocyanidins at the 3-hydroxyl position | Kobayashi et al., 2001 [122] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



