
Submitted:
25 September 2025
Posted:
26 September 2025
You are already at the latest version
Abstract
A pair of connected flowers preserved in a mid-Cretaceous (early Cenomanian) Myanmar amber is described and named Antiquigemina pilosa Wang and Li gen. et sp. nov.. Antiquigemina pilosa has bisexual flowers with a calyx of more than 3 sepals, a corolla of five (or six) petals, a whorl of more than 4 stamens and a tricarpellate pistil with a half inferior ovary. The differences in style divergence and calyx presence between paired flowers indicate different developmental stages. Antiquigemina pilosa is a core eudicot with a pentamerous or hexamerous perianth, in which sepals are obviously smaller than petals. Antiquigemina pilosa provides first-hand evidence for the early diversification of core eudicots.
Keywords:
angiosperm
; fossil flower
; core eudicots
; Myanmar amber
; paired flowers
1. Introduction
Core eudicots, characterized by pentamerous whorled flower parts, distinct sepals and petals, and tricolpate pollens [1], constitute a major part of the diversity of extant angiosperms. The earliest fossil records of core eudicots are found near the boundary of the Early Cretaceous and Late Cretaceous, indicating the radiation of core eudicots during that time [2,3,4,5,6,7,8,9]. These findings correspond with recent molecular estimates revealed that core eudicots originating during the Barremian to Aptian, with most subordinate orders originating from that time to the Late Cretaceous [10,11,12]. However, owing to the relatively limited fossil records of early core eudicots, the details of this radiation remain elusive [2,3,4,5,6,7,8,9]. Flowers preserved within Myanmar ambers from the latest Albian–earliest Cenomanian offer a great chance for understanding the radiation of core eudicots. More than 30 species of fossil flowers have been described within Myanmar ambers, with several of them identified as core eudicots [2,3,4,5,6,7].
In this work, we report a new pair of flowers directly linked through their pedicels and a short stalk, preserved within an amber from the Mid-Cretaceous. These flowers provided a direct evidence indicating the early diversification of core eudicots.
2. Materials and Methods
The samples were collected from the Noije Bum 2001 Summit Site, Hukawng Valley, Kachin, Myanmar (26°20′N, 96°36′E) (Figure 1). The age of the sample is the earliest Cenomanian, late Cretaceous (98.79 ± 0.62 Ma) according to the U–Pb dating of zircons [13]. The sample was immersed in oil and then photographed via a ZEISS Axio Zoom V16 stereomicroscope. The samples were scanned via a Zeiss Xradia 520 versa X-ray microscope for micro-CT. The 3D reconstruction and virtual sections were generated via VGStudio MAX 3.0. Floral diagrams were illustrated, and all the figures were organized for publication in Adobe Illustrator CC 2015.
3. Systematics
Angiospermae
Eudicots
Family Incertae sedis
Antiquigemina Wang and Li gen. nov.
Generic diagnosis: Flowers in pairs, each subtended by a bract. Flowers bisexual. The hypanthium formed by a receptacle, with at least 3 sepals, 5 or 6 petals and a whorl of at least 4 stamens. The stamens comprising a filament and a dorsifixed bilocular anther. Pistil solitary, tricarpellate, with bifurcated style. Ovary inferior or half inferior.
Antiquigemina pilosa Wang and Li gen. et sp. nov.
Holotype: PB206703, housed in the collection department of Nanjing Institute of Geology and Palaeontology.
Specific diagnosis: In addition to that of the genus, pedicels and bracts covered by trichomes. Sepals and petals triangular, involute, covered by unicellular, unbranched soft hairs. Sepals smaller than petals. Calyx caducous. Ovary half inferior.
Etymology: Antiqui- from the Latin word antiquus, emphasizing the relatively early age. Gemina from the zodiac sign Gemini, standing for the paired flowers. pilosa for trichomes or hairs covering bracts, pedicels and petals.
Description
A pair of flowers were preserved within a piece of brownish amber (Figure 2A–J). Each flower grew in the axil of the bract borne on a common fragmented stalk. The glabrous stalk measures 1.2 mm in length and 0.5 mm in diameter (Figure 2K). The bracts were covered by trichomes ranging from 1.4 mm long and 0.6 mm wide to 2.7 mm long and 0.8 mm wide (Figure 2K). These flowers were designated flower A and flower B according to the bract size: flower A has the larger bract and flower B the smaller bract (Figure 2A–C).
Flower A and flower B are similar in size, measuring 2.2–2.7 mm in diameter. Both flowers are bisexual, each consisting of a slender pedicel, a hypanthium formed by a receptacle, a calyx, a corolla, androecia, and a pistil (Videos S1, S2). The pedicels measure 4.9 mm in length and 220–300 μm in width and are covered by trichomes (Figure 2A, K). The petals and sepals are separated, triangular in shape, and reflexing outwards (Figure 2D–E, G–H). Unicellular, unbranched soft hairs are found on the petals under a stereomicroscope (Figure 3A–B). Flower B has both a calyx and a corolla (Figure 2H–I). The corolla comprises five petals measuring 1.0–1.2 mm in width at their widest point and 1.7–3.2 mm in length. The calyx comprises at least three sepals, which are smaller than the petals, measuring 0.7–1.0 mm in width at their widest point and 1.2–1.3 mm in length. Flower A has a corolla of only five petals (Figure 2E–F), each measuring 0.7–1.2 mm in width at the widest point and 1.2–2.0 mm in length. The calyx in flower A is absent. Both flower A and flower B have a petal split into two lobes (Figure 2E–F, H–J).
The androecium comprises at least four stamens in a whorl, mostly alternate to the petals but one opposite in flower B (Figure 2H, 3C–E; Videos S1, S2). A stamen consists of a filament and anther. The filament exserts above the inner margin of the hypanthium, tapering distally, and measuring 1.1–1.2 mm in length and 80–120 μm in diameter at the base (Figure 3C). The anther is dorsifixed and bilocular (Figure 3F–G). Each locule is ellipsoid in shape, measuring 1.3 mm in length, 430 μm in width and 310 μm in height, latrorse, and dehisces longitudinally (Figure 3F–G). The anther is absent in several stamens (Figure 3C). The pistil is located in the center of the flower and includes a stigma, style and ovary. The style of flower A bifurcated at an angle of 30 degrees (Figure 3C, H) with two branches measuring 1.0–1.3 mm, whereas the pistil tip of flower B barely bifurcated (Figure 3I–J). The stigma and style measure 1.9–2.1 mm in length and 310 μm in diameter at the style base. The hypanthium encloses more than 80% of the length of the ovary, suggesting a half inferior ovary. The ovaries are tricarpellate (Figure 3K). The placenta and ovules are obscure.
Remarks
There are minor differences between the results of the stereomicroscopic observations and the micro-CT reconstructions (Figure 2A–C). Micro-CT failed to reveal all the anthers and some stamens because of their weak density contrast against the amber matrix. The pistil of flower B can be observed via micro-CT reconstruction but cannot be observed directly under a stereomicroscope (Figures 2A–B, 3C), probably because of differential preservation. The characteristics based on observations using both methods are more credible.
Although the sepals are similar to the petals in shape, the sizes of the sepals (0.7–1.0 mm in width and 1.2–1.3 mm in length) are distinct from those of the petals (1.0–1.2 mm at their widest point and 1.7–3.2 mm in length), suggesting that the perianth parts can be distinguished as calyx and corolla in Antiquigemina pilosa.
In Antiquigemina pilosa, the spacing between sepals or stamens is uneven. It is impossible to determine whether it is original or artefacts. Therefore, we use “at least” to describe this situation.
Lobed petals in both flower A and flower B may lead to two different interpretations of flower merosity (Figure 2E–F, H–I). Each lobe is considered a single petal, and the number of petals is six rather than five. The sum of the widths of adjacent lobes equals that of a normal petal, suggesting that their forms are more likely a consequence of the splitting of the original petal, thus making alternative interpretations less likely. Lobed petals in Antiquigemina pilosa may reflect that the earliest corolla have high developmental plasticity, as the ancestral petals sometimes may possess the plasticity of lobation. On the other hand, lobed petals may result from accidental external damage. Basing on current evidence, none of these interpretations can be completely confirmed or excluded. Therefore, it is still necessary to include both interpretations of five and six petals in our treatment.
Several stamens comprise only a filament, without anther, suggesting that they might be staminodes. However, the alternative possibility that the anthers abscised naturally or broke off due to taphonomic reasons cannot be excluded. Thus in this paper, we collectively referred all filaments with or without an anther as stamens.
4. Discussion
4.1. Development of the Paired Flowers
The differences between flower A and flower B include the presence of the calyx and the bifurcation of the style. Considering that these two flowers are physically connected, there are three possible explanations for this phenomenon: 1) functional specialization between the two flowers; 2) preservation bias between the two flowers; and 3) different developmental stages of the flowers. However, the flower pair of Antiquigemina pilosa is unlikely to have functional specialization given that both are bisexual and of similar size. The absence of sepals in flower A should result from a secondary cause, as explained by 2) or 3). Thus, in the original state, the two flowers should have identical floral organization.
Bifurcation of the style may be a sign of gynoecium maturation. For example, the degree between style lobes of Litchi chinensis gradually increases during gynoecium development [15]. Similarly, the obvious bifurcated style in flower A may indicate that the flower is more mature than that in flower B, whose stigma only shows a slight bifurcation, suggesting that these flowers are in different (rather than the same) developmental stages. The developmental stage of flower B later than flower A can also account for the absence of sepals in flower A. Three sepals take up half a whorl in flower B, but they are absent in flower A, probably due to Antiquigemina pilosa’s caducous calyx. Sepals in a caducous calyx fall off earlier than other flower parts, and not withering when fallen. When fossilized, the sepals in flower A may have fallen completely, whereas flower B still has three unfolded sepals. The less severe calyx abscission in flower B than in flower A also aligns with the developmental order of flower B after flower A.
The paired flowers at different developmental stages provide clues for categorizing the inflorescence of Antiquigemina pilosa. The paired flowers of A. pilosa may be the distal part of a botryoid inflorescence or a cyme [16]. Consecutive development is common within the distal flower pairs of these inflorescences [16]. The alternative hypothesis is that paired flowers in A. pilosa are equal to paired flowers in Caprifoliaceae s.l. [17] or part of the paired-flower cyme as in Lamiales [18]. This hypothesis, which is consistent with asynchronously developed paired flowers, can further explain why there are two bracts associated with A. pilosa. However, considering that the formation of such inflorescences involves complex specializations [18,19,20,21,22], more evidence is needed to determine whether these specializations emerged during the early diversification of core eudicots that occurred 99 million years ago.
4.2. Comparison with Extant Angiosperms
The solitary tricarpellate pistil of Antiquigemina pilosa is distinct from extant basal angiosperms, including the ANA clade [23] and magnoliids [24]. Five distinct petals in a whorl of A. pilosa distinguish itself from monocots, which are characterized by trimerous flowers [1]. An alternative interpretation of petal number, treating each lobe of the lobed petal in pentamerous interpretation as a distinct petal (Figure 2F, I), is still at odds with monocots. Two whorls of similar trimerous perianths whose bases fit tightly, forming a perianth plane in some monocots (e.g., Xerophyta humilis), may be similar to the corolla of A. pilosa under micro-CT. However, the three sepals in flower B of A. pilosa defy this interpretation, as the gap in this whorl suggests that their calyx is composed of more than three members.
Among eudicots, a nearly inferior tricarpellate ovary as observed in Antiquigemina pilosa is relatively rare and might be informative of the affinities of A. pilosa. Flowers with pentamerous perianths and a tricarpellate ovary in some basal eudicots have been explained as a transitional form toward the typical pentamerous flower in core eudicots according to flower development [1,25]. The pentamerous perianths, tricarpellate ovary and position of stamens in A. pilosa indicate that A. pilosa probably have not yet evolved into a typical pentamerous flower. However, the perianths in A. pilosa are obviously differentiated into sepals and petals, suggesting that A. pilosa is closer to core eudicots than to basal eudicots, whose calyx and corolla are usually poorly differentiated [1].
The tricarpellate ovary has also evolved independently within several different orders of core eudicots [26]. The relatively limited space for gynoecium development compared with the outer whorls probably leads to a reduction in carpel numbers [27]. Derivation through the specialization of pentamerous flowers may serve as an alternative hypothesis for the tricarpellate ovary of A. pilosa.
A bisexual, pentamerous or hexamerous flowers with inferior or half inferior tricarpellate ovaries can be found only in the following extant angiosperm families: Saxifragaceae (Saxifragales) [28]; Rhamnaceae (Rosales) [pp. 320–338] [29]; Myrtaceae (Myrtales) [30]; Ancistrocladaceae (Caryophyllales) [31]; Hydrangeaceae [pp. 202–215] [29]; Loasaceae (Cornales) [pp. 239–254] [29]; Alseuosmiaceae, Argophyllaceae, Campanulaceae (Asterales) [32]; Araliaceae, Torricelliaceae (Apiales) [33]; Viburnaceae, Caprifoliaceae s.l. (Dipsacales) [34]. A detailed comparison between Antiquigemina pilosa and these families can be found in Table 1. Among these families, families in Cornales, Asterales and Dipsacales can be distinguished from A. pilosa by their connate petals. Sepals in A. pilosa are distinct, lamellate, different from the tube-formed calyx with minute lobes in Araliaceae and Torricelliaceae. Disc that exists in Myrtaceae, Rhamnaceae and Loasaceae is absent in A. pilosa. Ancistrocladaceae usually has more whorls of stamens than A. pilosa, and their anther attachment is also different. The lobed ovary in Saxifragaceae and the papillate stigma in Hydrangeaceae do not exist in A. pilosa.
Overall, the characteristic assemblage of Antiquigemina pilosa is not completely comparable to that of any extant angiosperm family. This reduces the possibility of Antiquigemina pilosa being a specialized form evolving to an extant core eudicot families. Considering the relatively early age of A. pilosa, A. pilosa is more likely being a unique extinct core eudicot that gives rise to no extant angiosperms. Thus, we put A. pilosa into incertae sedis at the family level.
4. Conclusions
The description of Antiquigemina pilosa enhances our knowledge of the diversity of the mid-Cretaceous Myanmar Amber Biota. This plant displays morphological characteristics of core eudicots, but the systematic relationships of A. pilosa to other angiosperms remain elusive. The atypical character combination, on one hand, reveals a unique transitional, unstable, uncanalized status of eudicot flowers in their early evolution, on the other hand, underscores the existence of non-basal eudicots in the mid-Cretaceous. Nonetheless, it is necessary to emphasize that the diversification of core eudicots was already underway about 100 Ma ago. It is expected that, as more digging is carried out, more aspects of the early evolution of eudicots will be unveiled.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Video S1. 3D reconstruction of flower A.Video S2. 3D reconstruction of flower B.
Author Contributions
Resources, Weijia Huang; conceptualization, Weijia Huang and Xin Wang; methodology, Fengyan Li and Xin Wang; investigation, Fengyan Li; writing—original draft preparation, Fengyan Li; writing—review and editing, Xin Wang.
Funding
This research was funded by the National Key Research and Development Program of China, grant number 2024YFF0807601.
Acknowledgments
We appreciate Suping Wu for her help in micro-CT reconstruction and Yehao Wang for his help in amber grinding and polishing.
Conflicts of interest
The authors declare that they have no conflicts of interest.
References
- Endress, P.K. Flower structure and trends of evolution in eudicots and their major subclades. Ann. Missouri Bot. Gard. 2010, 97, 541–583. [Google Scholar] [CrossRef]
- Poinar, G.Jr.; Chambers, K.L.; Buckley, R. Eoepigynia burmensis gen. and sp. nov., an early Cretaceous eudicot flower (Angiospermae) in Burmese Amber. J. Bot. Res. Inst. Texas 2007, 1, 91–96. [Google Scholar]
- Poinar, G.Jr.; Chambers, K.L.; Buckley, R. An early Cretaceous angiosperm fossil of possible significance in rosid floral diversification. J. Bot. Res. Inst. Texas 2008, 2, 1183–1192. [Google Scholar]
- Chambers, K.; Poinar, G.Jr.; Buckley, R. Tropidogyne, a new genus of Early Cretaceous Eudicots (Angiospermae) from Burmese Amber. Novon 2010, 20, 23–29. [Google Scholar] [CrossRef]
- Poinar, G.O.Jr.; Chambers, K.L. Tropidogyne pentaptera, sp. nov., a new mid-Cretaceous fossil angiosperm flower in Burmese amber. Palaeodiversity 2017, 10, 135–140. [Google Scholar] [CrossRef]
- Poinar, G.O.Jr.; Chambers, K.L. Tropidogyne lobodisca sp. nov., a third species of the genus from mid-Cretaceous Myanmar amber. J. Bot. Res. Inst. Texas 2019, 13, 461–466. [Google Scholar] [CrossRef]
- Liu, Z.J.; Huang, D.Y.; Cai, C.Y.; Wang, X. The core eudicot boom registered in Myanmar amber. Sci. Rep. 2018, 8, 16765. [Google Scholar] [CrossRef]
- Friis, E.M.; Pedersen, K.R.; Crane, P.R. The emergence of core eudicots: new floral evidence from the earliest Late Cretaceous. Proc. R. Soc. B 2016, 283, 20161325. [Google Scholar] [CrossRef]
- Manchester, S.R.; Dilcher, D.L.; Judd, W.S.; Corder, B.; Basinger, J.F. Early Eudicot flower and fruit: Dakotanthus gen. nov. from the Cretaceous Dakota Formation of Kansas and Nebraska, USA. Acta Palaeobot. 2018, 58, 27–40. [Google Scholar] [CrossRef]
- Li, H.T.; Yi, T.S.; Gao, L.M.; Ma, P.F.; Zhang, T.; Yang, J.B.; Gitzendanner, M.A.; Fritsch, P.W.; Cai, J.; Luo, Y.; et al. Origin of angiosperms and the puzzle of the Jurassic gap. Nat. Plants 2019, 5, 461–470. [Google Scholar] [CrossRef]
- Zhang, G.J.; Ma, H. Nuclear phylogenomics of angiosperms and insights into their relationships and evolution. J. Integr. Plant Biol. 2024, 66, 546–578. [Google Scholar] [CrossRef]
- Ma, X.Y.; Zhang, C.; Yang, L.X.; Hedges, S.B.; Zhong, B.J. New insights on angiosperm crown age based on Bayesian node dating and skyline fossilized birth-death approaches. Nat. Commun. 2025, 16, 2265. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.H.; Grimaldi, D.A.; Harlow, G.E.; Wang, J.; Wang, J.; Yang, M.C.; Lei, W.Y.; Li, Q.L.; Li, X.H. Age constraint on Burmese amber based on U–Pb dating of zircons. Cretac. Res. 2012, 37, 155–163. [Google Scholar] [CrossRef]
- Cruickshank, R.D.; Ko, K. Geology of an amber locality in the Hukawng Valley, northern Myanmar. J. Asian Earth Sci. 2003, 21, 441–455. [Google Scholar] [CrossRef]
- Stern, R.A.; Eisenstein, D.; Voet, H.; Gazit, S. Female ‘Mauritius’ litchi flowers are not fully mature at anthesis. J. Hortic. Sci. 1997, 72, 19–25. [Google Scholar] [CrossRef]
- Endress, P.K. Disentangling confusions in inflorescence morphology: Patterns and diversity of reproductive shoot ramification in angiosperms. J. Syst. Evol. 2010, 48, 225–239. [Google Scholar] [CrossRef]
- Landrein, S.; Prenner, G. Unequal twins? Inflorescence evolution in the twinflower tribe Linnaeeae (Caprifoliaceae s.l.). Int. J. Plant Sci. 2013, 174, 200–233. [Google Scholar] [CrossRef]
- Weber, A. Pair-flowered cymes in the Lamiales: structure, distribution and origin. Ann. Bot. 2013, 112, 1577–1595. [Google Scholar] [CrossRef]
- Haber, J. Comparative anatomy and morphology of flowers and inflorescences of the Proteaceae. I. Some Australian taxa. Phytomorphology 1959, 9, 325–358. [Google Scholar]
- Haber, J. Comparative anatomy and morphology of flowers and inflorescences of the Proteaceae. II. Some American taxa. Phytomorphology 1961, 11, 1–16. [Google Scholar]
- Johnson, L.A.S.; Briggs, B.G. Evolution in the Proteaceae. Aust. J. Bot. 1963, 11, 21–61. [Google Scholar] [CrossRef]
- Douglas, A.W.; Tucker, S.C. Inflorescence ontogeny and floral organogenesis in Grevilleoideae (Proteaceae), with emphasis on the nature of the flower pairs. Int. J. Plant Sci. 1996, 157. [Google Scholar] [CrossRef]
- Endress, P.K.; Igersheim, A. Gynoecium structure and evolution in basal angiosperms. Int. J. Plant Sci. 2000, 161, S211–S223. [Google Scholar] [CrossRef]
- Igersheim, A.; Endress, P.K. Gynoecium diversity and systematics of the Magnoliales and winteroids. Bot. J. Linn. Soc. 1997, 124, 213–271. [Google Scholar] [CrossRef]
- Ronse De Craene, L.P.; Quandt, D.; Wanntorp, L. Floral development of Sabia (Sabiaceae): Evidence for the derivation of pentamery from a trimerous ancestry. Am. J. Bot. 2015, 102, 336–349. [Google Scholar] [CrossRef]
- Soltis, D.E.; Senters, A.E.; Zanis, M.J.; Kim, S.; Thompson, J.D.; Soltis, P.S.; Ronse De Craene, L.P. .; Endress, P.K.; Farris, J.S. Gunnerales are sister to other core eudicots: Implications for the evolution of pentamery. Am. J. Bot. 2003, 90, 461–470. [Google Scholar] [CrossRef]
- Ronse De Craene, L.P. Meristic changes in flowering plants: How flowers play with numbers. Flora. 2016, 221, 22–37. [Google Scholar] [CrossRef]
- Kubitzki, K. The families and genera of vascular plants. Volume IX flowering plants. eudicots: Berberidopsidales, Buxales, Crossosomatales, Fabales p.p., Geraniales, Gunnerales, Myrtales p.p., Proteales, Saxifragales, Vitales, Zygophyllales, Clusiaceae Alliance, Passifloraceae Alliance, Dilleniaceae, Huaceae, Picramniaceae, Sabiaceae; Springer-Verlag Berlin Heidelberg GmbH: Germany, 2007; pp. 418–435. [Google Scholar]
- Kubitzki, K. The families and genera of vascular plants. Volume VI flowering plants. eudicots. dicotyledons: Celastrales, Oxalidales, Rosales, Cornales, Ericales; Springer-Verlag Berlin Heidelberg GmbH: Germany, 2004; pp. 202–215. [Google Scholar]
- Kubitzki, K. The families and genera of vascular plants. Volume X flowering plants. eudicots: Sapindales, Cucurbitales, Myrtaceae; Springer-Verlag Berlin Heidelberg GmbH: Germany, 2011; pp. 212–271. [Google Scholar]
- Kubitzki, K.; Bayer, C. The families and genera of vascular plants. Volume V flowering plants. eudicots. dicotyledons: Malvales, Capparales and Nonbetalain Caryophyllales; Springer-Verlag Berlin Heidelberg GmbH: Germany, 2003; pp. 25–27. [Google Scholar]
- Kadereit, J.W.; Jeffrey, C. The families and genera of vascular plants. Volume VIII flowering plants. eudicots: Asterales; Springer-Verlag Berlin Heidelberg GmbH: Germany, 2007; pp. 7–56. [Google Scholar]
- Kadereit, J.W.; Bittrich, V. The families and genera of vascular plants. Volume XV flowering plants. eudicots: Apiales, Gentianales (except Rubiaceae); Springer International Publishing AG: Switzerland, 2018. [Google Scholar]
- Kadereit, J.W.; Bittrich, V. The families and genera of vascular plants. Volume XIV flowering plants. eudicots: Aquifoliales, Boraginales, Bruniales, Dipsacales, Escalloniales, Garryales, Paracryphiales, Solanales (except Convolvulaceae), Icacinaceae, Metteniusaceae, Vahliaceae; Springer International Publishing AG: Switzerland, 2016; pp. 17–401. [Google Scholar]
Figure 1.
A. Map of Myanmar, red delta indicates the position of Hukawng Valley; B. Geographical map of the amber locality in the Hukawng Valley (modified from Cruickshank and Ko, 2003 [14]).
Figure 1.
A. Map of Myanmar, red delta indicates the position of Hukawng Valley; B. Geographical map of the amber locality in the Hukawng Valley (modified from Cruickshank and Ko, 2003 [14]).
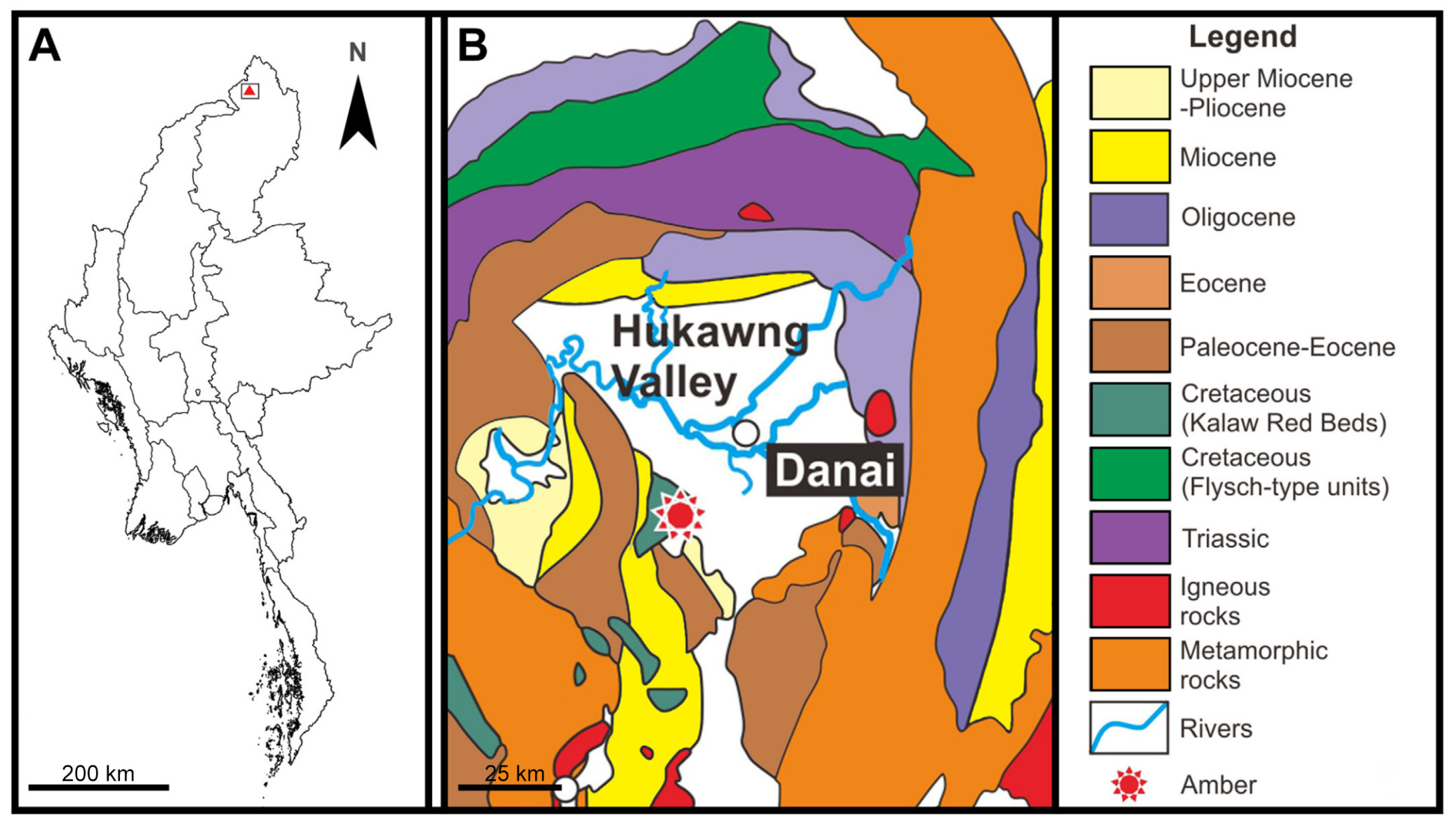
Figure 2.
Paired flowers of Antiquigemina pilosa (PB206703): (A–B) General morphology of A. pilosa from both sides of the amber. (C) Micro-CT reconstruction of A. pilosa. (D) Longitudinal section of flower A. (E) Bottom view of flower A. (F) Transverse section of flower A. (G) Longitudinal section of flower B. (H) Bottom view of flower B. (I) Transverse section of flower B. For (E–F) and (H–I), we numbered sepals and petals. Yellow arrows indicate the petals. Blue arrows indicate the sepals. Blue dotted lines indicate the outline of sepals. White arrows indicate the lobed petal. The pink arrow indicates an antipetalous stamen. (J) Flower diagram of flowers B (left) and A (right). Stamens with solid lines are observed via at least one method (stereomicroscopy or micro-CT). Stamens with broken lines are preserved only as filament residues. The petals with dotted line intervals within are the lobed petals. (K) Enlargement of A, showing the paired bracts on the fragmented stalk.
Figure 2.
Paired flowers of Antiquigemina pilosa (PB206703): (A–B) General morphology of A. pilosa from both sides of the amber. (C) Micro-CT reconstruction of A. pilosa. (D) Longitudinal section of flower A. (E) Bottom view of flower A. (F) Transverse section of flower A. (G) Longitudinal section of flower B. (H) Bottom view of flower B. (I) Transverse section of flower B. For (E–F) and (H–I), we numbered sepals and petals. Yellow arrows indicate the petals. Blue arrows indicate the sepals. Blue dotted lines indicate the outline of sepals. White arrows indicate the lobed petal. The pink arrow indicates an antipetalous stamen. (J) Flower diagram of flowers B (left) and A (right). Stamens with solid lines are observed via at least one method (stereomicroscopy or micro-CT). Stamens with broken lines are preserved only as filament residues. The petals with dotted line intervals within are the lobed petals. (K) Enlargement of A, showing the paired bracts on the fragmented stalk.
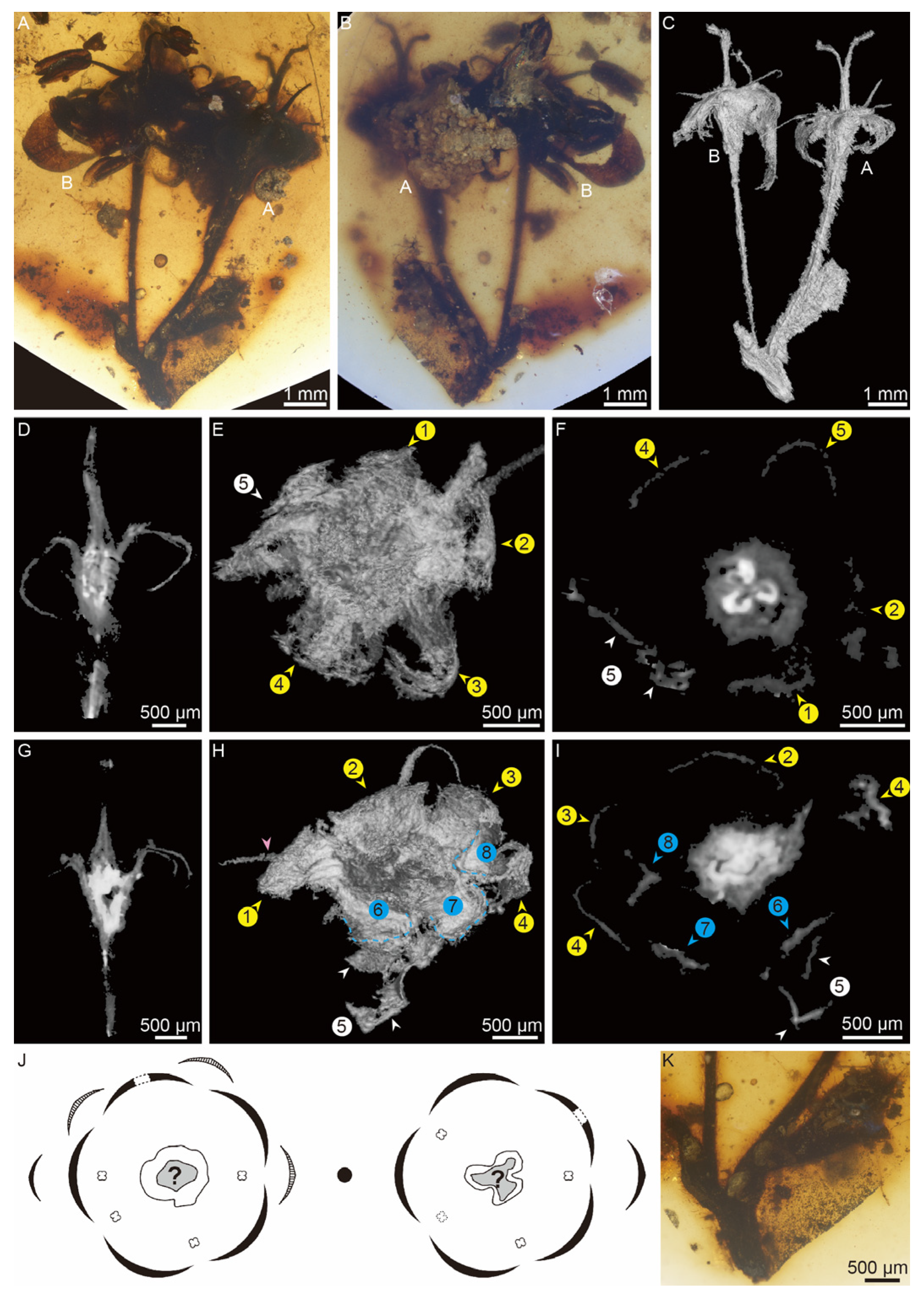
Figure 3.
Hairs, stamens and pistils of Antiquigemina pilosa: (A–B) Unicellular unbranched hairs on petals. (C) Enlargement of Figure 1A showing stamens. The white arrows indicate the stamens in flower B, and the blue arrows indicate the stamens in flower A. (D–E) Side view of flowers A and B showing the positions of the stamens. The white arrows indicate stamen alternate petals. The pink arrow indicates an antipetalous stamen. The green arrows indicate two prominent stamen bases. (F–G) Dorsifixed anthers. The white arrows indicate the position of the attachment of the anther. (H) Bifurcated style in flower A. (I) Slightly bifurcated style of flower B. (J) Transverse section from the yellow broken line in I. (K) Transverse section of a tricarpellate ovary in flower A.
Figure 3.
Hairs, stamens and pistils of Antiquigemina pilosa: (A–B) Unicellular unbranched hairs on petals. (C) Enlargement of Figure 1A showing stamens. The white arrows indicate the stamens in flower B, and the blue arrows indicate the stamens in flower A. (D–E) Side view of flowers A and B showing the positions of the stamens. The white arrows indicate stamen alternate petals. The pink arrow indicates an antipetalous stamen. The green arrows indicate two prominent stamen bases. (F–G) Dorsifixed anthers. The white arrows indicate the position of the attachment of the anther. (H) Bifurcated style in flower A. (I) Slightly bifurcated style of flower B. (J) Transverse section from the yellow broken line in I. (K) Transverse section of a tricarpellate ovary in flower A.
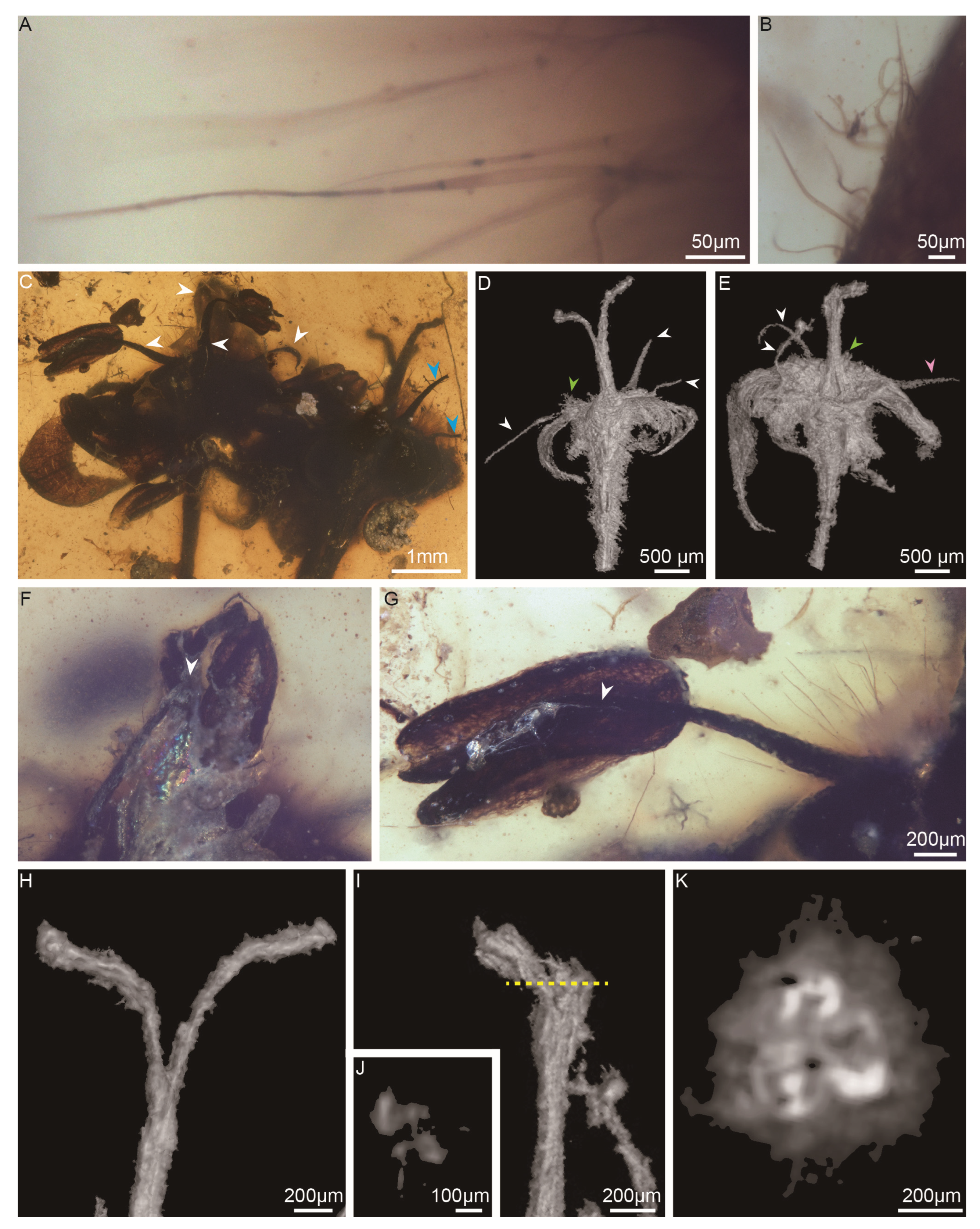

Table 1.
Comparison among Antiquigemina pilosa and the extant core eudicot families with bisexual pentamerous or hexamerous tricarpellate flowers.
Table 1.
Comparison among Antiquigemina pilosa and the extant core eudicot families with bisexual pentamerous or hexamerous tricarpellate flowers.
| Sepal number | Petal number | Stamen number | Stamen opposite to petals? | Anther attachment | Existence of disk | Style | Ovary position | Carpel number | Placentation | |
|---|---|---|---|---|---|---|---|---|---|---|
| A. pilosa | at least 3 | 5 or 6 | at least 4 | yes or no | dorsifixed | no | 2-lobed | half inferior | 3 | unknown |
| Saxifragaceae | (3–)5(–10) | (4)5(6) | 5/5+5 | no | basifixed | no | 2–3 divided | superior to inferior | 2–3 | axile or parietal |
| Rhamnaceae | (3)4–5(6) | (3)4–5(6) | 4–5 | yes | anther minute | yes | 2–4-lobed | superior to inferior | 2–4 | basal |
| Myrtaceae | 4–5 | 4–5 | numerous | / | dorsifixed | yes | solitary | half inferior to inferior | 1–5 | parietal, axile or basal |
| Ancistrocladaceae | 5 | 5 | 10(5, 15) | / | basifixed | no | 3(4)-lobed | half-inferior | 3(4) | pendulous |
| Hydrangeaceae | 4–12 | 4–12, connate | 4–numerous | / | basifixed | no | 1–12 divided | half inferior to inferior | 2–12 | axile at the base and parietal above |
| Loasaceae | (4–)5(–8), connate |
(4–)5(–8), connate |
numerous | / | basifixed | no | solitary | half inferior to inferior | 3–5 | parietal |
| Alseuosmiaceae | 4–5(6), connate | 4–5(6), connate | 4–5(6), connate | no | dorsifixed | yes | solitary | half inferior to inferior | 2–3 | axile |
| Argophyllaceae | 5, connate |
5, connate |
5 | no | basifixed | yes or no | solitary | half inferior to inferior | 1–3 | axile |
| Campanulaceae | (3–)5(–10), connate |
(3–)5(–10), connate | (3–)5(–10) | yes | basifixed | yes | 2–3-lobed | inferior, rarely half inferior or superior | 2–5(–10) | axile |
| Araliaceae | (3–)5(–12), connate |
(3–)5(–12), | 3 –numerous, | / | dorsifixed | yes | 1–numerous | inferior, rarely half inferior or superior | 2–5(–numerous) | apical |
| Torricelliaceae | (3–)5, connate | 5 | 5 | no | basi or dorsifixed | yes or no | 3 divided | inferior | 3 | apical or axile |
| Caprifoliaceae s.l. | 4–5(6), connate | 4–5(6), connate | 4–5 | no | dorsifixed | yes or no | solitary or 2–3-lobed | inferior | 3(2–5) | axile or pendulous |
| Viburnaceae | 2–5, connate |
3–5(6), conate |
3–5(6) | no | basi or dorsifixed | yes or no | solitary or 2–5-lobed | half inferior to inferior | 2-5 | axile or parietal |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



