
Submitted:
24 September 2025
Posted:
26 September 2025
You are already at the latest version
Abstract
Gene expression data from our publicly accessible transcriptomic dataset obtained by profiling the left ventricle myocardia of mouse models of Chagasic cardiomyopathy and post-ischemic heart failure were re-analyzed from the perspective of the Genomic Fabric Paradigm. In addition to the regulation of the gene expression levels, we determined the changes in the strength of the homeostatic control of transcript abundance and the remodeling of the gene networks responsible for the mitochondrial respiration. The analysis revealed that most of the mitochondrial genes assigned to the five complexes of the respiratory chain were significantly down-regulated by both Chagas disease and ischemia but exhibit outstanding recovery of the normal expression levels following direct injection of bone marrow-derived stem cells. However, instead of regaining the original expression control and gene networking, the treatment induced novel mitochondrial arrangements, suggesting that multiple transcriptomic topologies might be compatible with any given physiological or pathological state. The study confirmed several established mechanisms, and introduce new signals, especially Cox4i2, Cox6b1, Cox7b, Ndufb11, and Tmem186, that warrant further investigations. Their broad rescue with cell therapy underscores mitochondria as a convergent, tractable target for cardiac repair.
Keywords:
cellular respiration
; expression control
; expression correlation
; genomic fabric
; NADH dehydrogenase
; oxidative phosphorylation
1. Introduction
Mitochondria are critical organelles that regulate cell metabolism and survival, especially in the heart, where mitochondria comprise 30 - 40% of cardiomyocyte volume and play a primarily role for energy production, calcium homeostasis and regulation of cellular apoptosis. Mitochondrial dysfunction leads to impaired cardiomyocyte function, a hallmark of heart failure [1]. Since mitochondria are essential for the cellular energy-demanding cardiac contractile function, a decline in mitochondrial biogenesis and function [2] was associated with the development of ventricular dysfunction caused by myocardial infarction [3], type 2 diabetes [4], Chagas disease [5], or anticancer drug-related cardiotoxicity [6] among several cardiovascular afflictions.
Decades after initial infection with the parasitic euglenoid Trypanosoma cruzi [7], transmitted by the so-called “kissing bug” [8], ~30% of individuals can develop chronic Chagasic cardiomyopathy (CCC), a congestive heart failure and dilated cardiomyopathy [9,10,11]. It is estimated that CCC affects about 7 million of people worldwide, most of them in Latin America [12], and became endemic recently even in the United States [13]. Although the pathogenesis of CCC remains a matter of debate [14,15], the involvement of cardiac mitochondria was first demonstrated by Garg et al. in 2003 [16] and confirmed by Báez et al. in 2011 [17]. They found that a year post-infection parasite persistence and inflammation were associated with structural and functional alterations in mouse cardiac mitochondria in a parasite strain-dependent manner [18]. Common comorbidities include dyslipidemia, hypertension [19] and might lead to cryptogenic stroke [20]. As expected for any infectious disease, CCC triggers the immune response [21] that changes during the development of the disease [22].
Transcriptomic studies on cardiomyocytes and heart left ventricle of CCC mice revealed remodeling of immune response [23], extracellular matrix, cell adhesion [24], intercellular communication via gap junction channels [25,26], mitochondrial oxidative phosphorylation [27], JAK/STAT signaling and cell cycle [28] functional pathways.
The progenitor cells are well recognized for their therapeutic potential in cardiac regeneration [29]. We found that injecting bone marrow-derived stem cells restores most of the heart function and recovers most of the normal heart gene expression profile [30,31].
Myocardial infarction [32], described as the cardiomyocytes’ death caused by insufficient oxygen supply, whose definition and management are still under debate [33,34], is directly related to mitochondrial dysfunction [35] and affects several functional pathways [36,37].
Previous studies on mouse models have shown that myocardial infraction induced by interruption of blood supply [38] activates strong inflammatory response and leads to ventricular remodeling and ischemic heart failure (IHF) [39]. These effects were also reversed by injecting bone marrow-derived mononuclear stem cells into the cardiac scar tissue [40]. Whatever the cause, heart failure has severe consequences on all organs, including triggering various mental disorders [41].
This research investigated whether there are mitochondria-related genomic alterations in failing hearts due to CCC and IHF before and after stem cell treatment. We analyzed the KEGG-constructed oxidative phosphorylation functional pathway [42] as arranged in the mitochondrial module of the KEGG-constructed cardiovascular disease pathway of diabetic cardiomyopathy [43]. The study did not stop at the alteration of the gene expression profile but extended to encompass the entire mitochondrial transcriptomic topology, i.e. to include the control of transcripts’ abundances and the transcriptomic networks, as common in our Genomic Fabric Paradigm [31].
2. Materials and Methods
2.1. Experimental Data
We re-analyzed publicly accessible gene expression data generated in former Iacobas’ lab by profiling the transcriptomes of the left heart ventricle myocardium of mouse models of CCC [44] and IHF [45] before and after the treatment with bone-marrow derived mononuclear cells [46,47]. The treating stem cells were collected from femurs and tibiae of C57Bl/6 adult mice and purified by centrifugation in Ficoll gradient [30]. The Chagas disease was induced by infecting adult C57Bl/6j with trypomastigotes of Colombian Trypanosoma cruzi strain [48], while the permanent myocardial infarcts were obtained by ligating the descending branch of the left coronary artery [39]. The experimental protocols and raw and normalized expression data are fully described and publicly accessible in the Gene Expression Omnibus (GEO) of the (USA) National Center for Biotechnology Information (NCBI) [44,45,46,47]. For each disease, we examined two conditions: IN (infected/infarcted – not treated) and IT (infected/infarcted – treated) and compared with CN (healthy/control – not treated), all in four biological replicates.
2.2. Primary Independent Characteristics of Individual Genes
A transcript was considered as properly quantified if the probing spot had a foreground fluorescence signal more than twice the background and no corrupted or saturated pixels in any microarray. Background subtracted foreground fluorescence signals were averaged for spots probing redundantly the same gene. According to our standard procedure [31], each properly quantified gene is characterized in each condition by three independent characteristics: AVE, REV and COR.
AVE is the average expression level across biological replicas normalized to that of the median gene expression in each sample. This type of normalization minimizes the errors caused by the differential impact of technical noise in the preparation and hybridization of the fluorescent samples.
REV is defined as the mid-interval of the chi-square estimate of the coefficient of variation of the expression levels on all spots probing redundantly the same gene across biological replicas of one condition. REV is used to calculate the REC (relative expression control) and RCS (relative control strength) as:
where REC is the Relative Expression Control and 〈REV〉(c) is the median REV of all genes quantified in condition: c = CN, IN, IT. A positive/negative REC value indicates that the expression of that gene is more/less controlled (i.e. more/less stably expressed across biological replicas) than the median in the condition. The control level of a gene indicates how much critical is that gene for the cell survival and phenotypic expression, with the top positive value pointing to the most important mitochondrial gene for that condition.
COR is the Pearson pair-wise correlation coefficient of the (log2) expression levels of two genes. There are three (p < 0.05) statistically significant cases when each of the two genes “i” and “j” in condition “c” are probed by single microarray spots:
1) synergistically expressed, i.e. their expression levels oscillate in phase across biological replicates.
2) antagonistically expressed, i.e. their expression levels oscillate in antiphase across biological replicates.
3) independently expressed genes, i.e. expression of one gene has no influence on the expression of the other gene.
COR analysis was used to identify the (p < 0.05) significant transcriptomic networks based on the “Principle of transcriptomic stoichiometry” [49], requiring coordinated expressions of genes whose encoded products are linked in a functional pathway. The overall expression correlation of the mitochondrial genes responsible for adjacent complexes (Cx) of the respiratory chain was measured by their coordination (COORD) percentage:
where: SYN, ANT, IND are the percentages of the p < 0.05 significant synergistically, antagonistically and independently expressed gene pairs out of all pairs that can be formed between the two complexes.
REC and COR values were used to hierarchize the genes according to their Gene Commanding Height (CGH) score [50]:
2.3. Transcriptomic Changes of Individual Genes and Functional Pathways
Our standard protocol [51] considers the expression of a gene as statistically significantly regulated (here in the left heart ventricle of the diseased mouse w/o cell treatment (d = IN, IT) with respect to the control counterpart CN) if the absolute expression fold-change exceeds the appropriate cut-off and the p-value of the heteroscedastic t-test of the means equality is less than 0.05. The absolute fold-change cut off (CUT) is computed for each gene by considering both the expression variabilities in the compared conditions and the technical noises of the probing microarray spots in the two sets of biological replicas.
Contributions of individual genes to the transcriptomic alteration in condition “d” (CCC, IHF) with respect to their healthy counterparts were measured by their WIR (Weighted Individual (gene) Regulation) score:
The overall alteration of a functional pathway Φ in a particular condition (CCC or IHF) was measured by the median WIR of the pathway genes. A positive median WIR indicates that total number of (significantly and not significantly) upregulated genes in the pathway exceeded the number of the downregulated and vice-versa.
The alteration of expression control in CCC and IHF mice w/o cell treatment with respect to the condition CN was measured by the Fold-Change (FC) defined as:
The fold-change alteration (negative for downregulation) of GCH in a treated (IT) or untreated (IN) Chagas and post-ischemic heart failure is computed as:
3. Results
3.1. Both Chagasic Disease and Post-Ischemic Heart Failure Are Characterized by Substantial Downregulation of Mitochondrial Genes
Figure 1 presents the mitochondrial genes that were significantly regulated (according to the composite criterion of the absolute fold-change and p-value (5)) in Chagasic disease and post-ischemic heart failure. Thus, in the untreated CCC, four of the five complexes contain only downregulated genes, with Cx 4 making the exception having one upregulated (Cox6a1) although two (Cox5a, Cox6b1) were downregulated. The percentage of down-regulated genes is even larger in the post-ischemic heart despite the upregulation of Ndufa9.
The large numbers of the downregulated genes in the five complexes indicate a significant reduction of the oxidative phosphorylation, and by consequence less production of ATP. Remarkably, in both cardiomyopathies, Ca2+ storage in the mitochondrion was diminished by downregulation of one of the involved genes (Vdac1 in CCC and Slc25a5 in IHF mice). The downregulation of these transporters led consistently to Ca2+ accumulation in the cardiomyocyte that affected the contractility function of this heart muscle cell.
3.2. Cell Treatment Restores the Normal Expression of Most Mitochondrial Genes Altered in Both Chagasic Disease and Post-Ischemic Heart Failure
As illustrated in Figure 2, we found that the expressions of most mitochondrial genes that have been altered in both Chagasic and ischemic mice were markedly restored following the cell treatment.
Moreover, in the case of CCC mice, the initially not affected Ndufb11 was upregulated by treatment, while the originally downregulated Ndufs4 was even switched to upregulated. Of note is that normal Ca2+ storage in the mitochondrion was restored in the treated CCC mice but remained diminished in the treated IHF now because of the downregulation of Vdac2 despite recovering the normal expression of initially downregulated Slc25a5.
3.3. The Largest Mitochondrial Gene Contributors to the Transcriptomic Alterations in Both Chagasic and Post-Ischemic Mice
Figure 1 and Figure 2 considered implicitly significantly regulated genes as equal contributors to the transcriptomic alteration presenting the overall change as percentages of up- and downregulated genes that is an oversimplified description of the biological reality. A better representation of the contribution would be by the pair (expression ratio, p-value) of each affected gene. However, we think that a more comprehensive characterization is by the weighted individual (gene) regulation (WIR), computed with formula (6), that incorporates the total expression change AVE(CN)(|x|-1) and the statistical confidence (1 – p-val) of this change.
Table 1 presents the largest five contributors (as absolute value of WIR) in Chagasic (CCC) and ischemic (IHF) mice, together with their normal average expression levels (AVE), expression ratios (X) and p-values of their regulation with respect to the normal expression. As expected, all top contributors had negative expression ratios.
The effectiveness of the cell treatment on restoring the expression profiles of the mitochondrial genes is evident by the change of the median WIR from -18.74 in infected not treated to -4.50 after treatment in CCC mice and from -16.83 to -1.81 in IHF mice. These results indicate both substantial reduction of the mitochondrial function caused by the parasitic infection and partial recovery following the cell treatment. While in CCC mice, the non-mitochondrial gene Pln was the largest contributor (WIR = - 222), in the ischemic mice one mitochondrial gene, Cox5a, tops the list with the impressive WIR = - 2603(!).
Interestingly, the median WIR value for the entire transcriptome was positive in all conditions, meaning that overall, the contributions of the up-regulated genes (like those involved in the inflammatory response as previously reported [23,39]) exceeded those of the down-regulated. As expected, the median WIRs of the entire transcriptome were larger for both untreated CCC (0.67 vs. 0.13) and IHF (1.63 vs. 0.70) mice, pointing to an overall reduction of the alteration of the expression profile.
3.4. The Most and the Least Controlled Mitochondrial Genes in CCC and IHF Mice
The experimental design with four biological replicates allowed us to estimate the Relative Expression Control (REC) of individual genes in each condition and how much both treated and untreated cardiomyopathies affected them. Table 2 presents the REC values (computed according to equation 1) of the most (REC > 0) and the least (REC < 0) controlled mitochondrial genes in all profiled conditions (healthy, treated and untreated CCC and IHF mice). We listed also the most and the least controlled genes in the entire transcriptome and the fold-change (negative for downregulation) of diseased states with respect to the healthy mice.
3.5. Both CCC and IHF Alter the Transcriptomic Networks of the Mitochondrial Genes by Partially Decoupling the Oxidative Phosphorylation Complexes
Figure 3 presents the (p < 0.05) significant transcriptomic networks coupling the five complexes of the oxidative phosphorylation in the heart left ventricles of healthy and untreated CCC and IHF mice. One observes that the adjacent complexes are coupled in all conditions mostly by synergistic expression correlations and that the coordination degree (computed with formula (2) is significantly lower in the untreated cardiomyopathies, indicating a partial decoupling of the complexes. The coordinated percentages were computed for the 80 possible gene-pairs between Cx1 and Cx2, 27 between Cx2 and Cx3, 84 between Cx3 and Cx4, and 140 between Cx4 and Cx5.
The reduction was much more considerable for the untreated IHF mice, going to even negative values (COORD(Cx1-Cx2) = -2.50%, COORD(Cx3-Cx4) = -1.19%, which means that more significantly independently that synergistically + antagonistically expressed gene pairs were identified. For instance, the number of synergistically + antagonistically expressed gene pairs between complexes Cx1 and Cx2 decreased from 18 (= 18 + 0) in CN to 15 (= 15 + 0) in CCC and to 6 (4 + 2) in IHF, while the number of independently expressed pairs goes from 5 in CN to 4 in CCC and 8 in IHF. In general, there are fewer antagonistically than independently expressed gene pairs, explaining the synchrony of the respiratory chain.
3.6. Stem Cell Treatment Benefits for the Transcriptomic Coupling of the Oxidative Phosphorylation Complexes
Figure 4 presents the (p < 0.05) significant gene expression correlations in treated CCC and IHF mice and the coordination degrees in all five conditions. As summarized in Figure 4c, both cardiomyopathies reduced the expression coordination degrees among all pairs of coupled complexes. However, the stem cell treatment restored partially the coordination degree in the infarcted mice while even substantially increasing it in the CCC mice.
One important observation is that although the cell treatment restored the normal expression levels for most mitochondrial genes, the transcriptomic coupling of the complexes exhibits different than normal patterns. Importantly, the patterns of treated CCC and IHF are also largely distinct to one another.
3.7. Both CCC and IHF Alter the Hierarchy of Mitochondrial Genes
Table 3 presents the most prominent five mitochondrial genes in treated and untreated Chagasic and ischemic mice according to their Gene Commanding Height (GCH) scores computed with formula (3). It contains also the fold-change (GCH-FC, negative for downregulation) of the GCHs in disease states with respect to the healthy condition, computed with formula (8). For comparison, the table presents also the GCH scores of the top five mitochondrial genes in healthy mice and their GCHs in the untreated CCC and IHF mice.
4. Discussion
Mitochondrial respiratory alterations and excessive generation of reactive oxygen species (ROS) are regarded as the main causes of heart failure [52,53], given that mitochondria provide about 90% of the energy used by heart in normal functioning [54,55]. Using the Genomic Fabric Paradigm [51], which provides the most theoretically possible comprehensive transcriptomic characterization, our present study has brought new insights into the current understanding of mitochondrial genes involvement underlying Chagas disease [56] and post-ischemic heart failure [57]. Although several previous studies specifically analyzed alteration of the expression of genes/proteins that regulate mitochondrial oxidative phosphorylation [16,17], none of them studied the alteration and recovery of the gene expression control and transcriptomic networking.
Our transcriptomic characterization of untreated CCC and IHF hearts indicated a unified failure of mitochondrial energetics that were substantially reversed by bone-marrow-derived cell therapy. At the level of Complex 1 [58], the downregulation of 11/32 Nduf genes in untreated CCC and 22/32 in untreated IHF mice likely predict a severe block in NADH oxidation, a surge in reactive-oxygen species, and consequent energy depletion. Complex 1 (NADH ubiquinone oxidoreductase [59]) is responsible in NADH oxidation and proton pumping along the electron transport chain [60]. For instance, the downregulation of Ndufa7 found by us (x = -1.92 in the untreated CCC and x = - 1.72 in untreated IHF) was reported also in cardiac hypertrophy [61]. Although Ndufa10, known for its role in spontaneous hypertension [62,63] was equally downregulated (x = -1.60) in both untreated cardiomyopathies, its downregulation was statistically significant only in IHF mice (p(IHF) = 0.009, p(CCC) = 0.142). Interestingly, while Ndufa9 was (even not statistically significant) downregulated in untreated CCC, it was upregulated (x = 4.18) in untreated IHF as happens in high-intensity training [62].
The collapse of 2/3 of the quantified Complex 2 genes, critical for the Krebs cycle [64,65,66,67], in both untreated heart diseases investigated by us, resulted from the significant down-regulation of Sdha (x(CCC) = -2.49, x(IHF) = -2.29) and Sdhb (x(CCC) = -2.07, x(IHF) = -1.59).
Electron flow stops at Complex 3: the hinge protein Uqcrch shows one of the strongest negative WIR scores (WIR = -44.38), consistent with knockout data in which its loss alone causes contractile failure [68], thereby supporting and broadening those functional observations to two distinct heart-failure models. Complex 3 (CoQ: cytochrome c-oxidoreductase [69]) deficiency is responsible for fewer mitochondrial disorders [70] although carbonylation of genes like Uqcrc2 (found as downregulated in both untreated CCC and IHF) is a direct response to the stress of Trypanosoma cruzi infection [71].
Complex 4 is compromised as suppression of the ATP/ADP sensor Cox4i1 in CCC, which matches marked reduction in state-3 respiration and cytochrome-c-oxidase deficits documented clinically [72]. The downregulation in untreated IHF mice of Cox7b (x = -2.3, WIR = -80) likely undermines Complex 4 assembly, highlighting a novel candidate for future functional probing. Cox5a, the largest contributor listed in Table 1 to the downregulation of Cx4 in IHF (WIR = -2603) is considered a biomarker for the blood stasis syndrome [67], cyanotic heart disease [73] and acute myocardial infarction [74].
Adaptive changes in energy preservation were evident at Complex 5, where the ATP synthase ε-subunit Atp5f1e was down-regulated (x = -2.22) in the ischemic hearts and then normalized after therapy. This not only agrees with, but now quantifies, the prior enzymology-based findings [75,76]. Our results about the downregulation of Cx 5 contradict the report of its upregulation found by another group [77] through meta-analysis of 13 patients with heart failure subjected to heart transplantation compared to 10 healthy counterparts. However, Cpt2 (Carnitine palmitoyltransferase 2), not affected by either CCC and IHF, is down-regulated in treated IHF, that, according to some research [78], might result in rapamycin-resistant, acetylation-independent hypertrophy.
By comparing Figure 1 and Figure 2, one observes that the cell treatment triggered marked recovery of the expression levels of almost all mitochondrial genes in both CCC and IHF mice. The recovery happens in all five oxidative phosphorylation complexes. While the commonly decreased expression of Ndufs1 in infarcted heart [79] is restored to the normal level by the cell treatment in our model, it remains (even less but still) downregulated in the treated CCC mice (from x(IN) = -3.02 to x(IT) = -2.20). However, the downregulation of Ndufs4 in T. cruzi infected mice was reversed to upregulation by the cell treatment, confirming the cardioprotection role of its upregulation [80].
A very important part of the study refers to the control of the transcripts’ abundances by the cellular homeostatic mechanisms and its modification during the disease development and following treatment. Among the most controlled genes (Table 2), there were 5 subunits of NADH:ubiquinone oxidoreductase (i.e. mitochondrial complex I), including Ndufa10, Ndufb10, Ndufb5, Ndufb11, and Ndufaf4. These genes can affect mitochondria function profoundly. Especially Ndufa10, the mitochondrial gene with the largest REC (relative expression control) in healthy mice whose RCS (relative control strength) was markedly reduced in both treated (by 32.80x) and untreated (by 22.70x) CCC, meaning a high expression flexibility to adapt to changeable environmental conditions that might eventually be used to alleviate diabetic cardiomyopathy [81]. Ndufa10 has been also implicated with diabetic cardiomyopathy [63] and ischemic injury [72]. Note also the substantial reduction of the expression control in both treated and untreated CCC mice for other stably expressed MITO genes in the healthy mice: Cox7b and Ndufb10. To our best knowledge, this is the first report revealing the significant alteration of the expression control of these mitochondrial gene in Chagasic heart. Ndufb10 observed destabilization agrees with its known role in holoenzyme assembly failure [82].
The high RCS of Tmem186 in the entire normal transcriptome was significantly reduced (RCS-FC = -39.13) in untreated IHF that persisted (RCS-FC = -22.50) after treatment, but the role of this gene is much less understood. Tmem186 is a component of the mitochondrial Complex 1 intermediate assembly [58]. In contrast, IHF increased by 36.56x the control strength of the antigen Cd164 but the cell treatment restores the normal transcription control.
There are several other interesting data in Table 2. For instance, while the RCS of Vdac2, whose downregulation is one of the main responsible factors for heart failure [83], increased by 2.89x in the not-treated CCC mice (IN) with respect to the healthy animals (CN), it decreased in treated mice (IT) by 3.23x. The control of Cox6b1, known to relieving hypoxia/reoxygenation injury [83] increased by 8.31x in the untreated IHF but decreased by 2.48 in treated IHF. The situation is opposite for Ndufaf4 whose expression control decreased in untreated CCC but increased in treated CCC. We found substantial changes in ischemic mice, with some genes exhibiting more expression control (e.g. Atp5f1e, Cox6b1). The control of Atp13a2 decreased in untreated IHF to considerably increase (by 14.82x) in treated IHF.
All these differences indicate that the homeostatic control mechanisms to keep the expression levels within certain intervals are neither uniform among genes nor similarly altered by each of the two cardiomyopathies or restored following the same therapeutic approach. A very important observation comparing data from Table 2 with Figure 1 and Figure 2 is that, even though the cell treatment restored the normal expression of several genes, their expression control was not. For instance, in the ischemic mice, the expression of the significantly down-regulated Cox6b1 (x(IN) = -2.87) was restored (x(IT) = 1.1) by the treatment but its RCS was decreased by 2.48x, while the recovered down-regulation of Atp5f1e (x(IN) = -2.22) was accompanied by an increase of the RCS by 2.77x). This observation indicates that whereas the treatment recovered the expression profile of most mitochondrial genes, it did not restore the initial control of the transcripts’ abundances.
Recent work in acute experimental models of Chagas disease showed that an imbalance between ROS and NO creates a strongly proarrhythmic substrate independent of major structural remodeling and directly links mitochondrial redox output to electrical instability [2,84]. The same study demonstrates that NO inhibition reverses early after-depolarizations and action potential alternans, underscoring redox–calcium crosstalk as a therapeutic lever. Our findings of Complexes 1-5 suppression and Vdac1 dysregulation align with this mechanism and suggest that transcriptomic loss of the ETC (electron transfer chain) control may be an upstream driver of the ROS/NO dis-equilibrium [85]. It was proved that excessive ROS generation is associated with low oxygen levels as in acute hypoxia [86] or in the ischemic heart.
Additional analysis highlighted that mitochondrial injury is shaped by both inflammatory environment and genotype [87]. This supports our observation that nuclear encoded ETC subunits (Ndufb11, Cox7b) are vulnerable nodes that could synergize with inflammatory signaling to depress ATP output. It also strengthens the rationale to stratify future cohorts by mitochondrial genotype and inflammatory profile when validating control metrics [81,88]. There was evidence for adipose tissue as a chronic reservoir for Trypanosoma cruzi and a source of adipokines-associated mitochondrial stress, implying that systemic metabolic inputs can burden cardiac bioenergetics [85]. This concept aligns with our Cpt2 signal finding that reduced fatty acid oxidation capacity in the heart may be compounded by parasite driven adipose dysfunction and argues for integrating peripheral metabolic tissues into future mechanistic and therapeutic designs [89].
Figure 3 presents for the first time to our knowledge the transcriptomic networks that couple the respiratory chain complexes. It shows also how infection with Trypanosoma cruzi and ischemia alter the transcriptomic coupling, indicating profound remodeling of the oxidative phosphorylation functional pathway. The most dramatic alteration occurred between Cx1 and Cx2 in the IHF mice, where the coordination degree decreased from +16.25% to -2.50%, meaning a significant decoupling of the paired genes between the two complexes. The marked coupling decrease between the adjacent complexes is an additional explanation of the mitochondrial failure in the two investigated cardiomyopathies.
Understanding the importance of some of these expression correlations for the normal heart function and what pathological consequences bring their alterations might open a very interesting and challenging field of research. For instance, Chagas disease turned the significant synergism of the downregulated Nfdufa6 and Sdha, whose simultaneous upregulation is an indication of acute myeloid leukemia [90], into statistically significant independence. We found that the pairs Uqcrb, Coxcb, and Atp5pf, Cox5b, reported as simultaneously upregulated in paradoxical low transvalvular flow and low gradient patients who develop advanced heart failure symptoms [91], are synergistically expressed in normal conditions. The synergism of the downregulated by ischemia Sdhb and Uqcrffs1 supports the efforts to improve the diagnosis and gene targeted therapies of Alzheimer’s disease [92].
As illustrated in Figure 4c, cell therapy recovered part of the decrease of the four coordination degrees in the IHF mice and even increased them over the original levels in the CCC mice. In the CCC mice the cell therapy not only restored the normal expressions of the down regulated Ndufv1 and Sdhb (downregulated also in diabetic patients caused by high fat diet [93]) but also made significant their synergistic expression. To prove this assertion, one can use an online available calculator [94] to determine the p-values of the statistical significance of the expression correlations in each the three conditions: COR(CN) = 0.832 (p-value = 0.168), COR(IN) = 0.397 (p-value = 0.603), COR(IT) = 0.996 (p-value = 0.004). Interestingly, Ndufa13 and Sdhd, that are independently expressed in healthy mice housed in normal atmospheric conditions, maintained their independence in untreated IHF but became synergistically expressed in both treated CCC and IHF mice. The treatment turned the practically neutral correlations: COR(CN) = 0.320 (p-val = 0.680), COR(IN) = 0.222 (0.778) between the downregulated Cox17 and Atp5mc1 in ischemic mice into a significant antagonistic expression: COR(IT) = -0.952 (p-val = 0.048). However, together with NDUFB1, COX17 and ATP5MC1, were reported as upregulated in Parkinson’s, Alzheimer’s and Huntington’s diseases [95].
Nevertheless, we have to recognize that the correlation analysis cannot determine what gene of the pair is the master, i.e. its expression level commands the expression of the other, nor about the energy coupling [96] and the electric charges transfer within or between the respiratory complexes [97,98,99].
The Prominence analysis ranked the mitochondrial genes according their importance for the functioning the cell powerhouse. From Table 3 one may learn that:
i) gene hierarchy is altered in cardiomyopathy as revealed by the differences between the top five genes in healthy mice compared to the sets of five in the other heart conditions.
ii) each type of cardiomyopathy induces distinct alteration of the gene hierarchy (see GCH differences between CCC and IHF for the most prominent genes in healthy mice.
iii) restoration of the normal expression level is not accompanied by the reinstate of the gene in the right hierarchy, indicated by the non-unit GCH-FC. For instance, Uqcrh, found by us as down-regulated in both untreated CCC and IHF mice, was reported as upregulated in patients with hypertrophic cardiomyopathy [100]. Cell treatment restored the normal expression in the CCC but not in the IHF mice. However, the restored expression level is not followed by the full restoration of its GCH after the treatment. Instead, its RCS became 6.67x stronger than in the normal condition (even stronger than in the untreated CCC, where it was 4.66x).
iv) cell treatment has different effects on the two types of heart afflictions. For instance, the somehow low GCH of Cox6b1 healthy mice that was considerably raised in both untreated cardiomyopathies (GCH-FC(IN-CCC) = 4.81, GCH-FC(IN-IHF) = 5.53), is raised even more in treated CCC (GCH-FC) = 7.34) but downgraded in treated IHF (GCH-FC = -2.10).
5. Conclusions
Taken together, the findings of this study establish an exhaustive profile of mitochondrial collapse in Chagas disease and post ischemic heart failure. The investigation demonstrated that targeted cell therapy can partially restore the functional integrity of key ETC components. It further confirmed several established mechanisms, and introduce new signals. Their broad rescue with cell therapy underscores mitochondria as a convergent, tractable target for cardiac repair.
Interestingly, although the cell treatment recovers the normal expression of the most of the regulated genes, it does not restore the normal expression control or the inter-complexes transcriptomic coupling. Moreover, both treated cardiomyopathies present different expression control and correlation patterns to each-other. Since the treated animals displayed almost normal electrophysiological parameters and excitatory and contractility properties of the ventricular myocardium, our results suggest that a physiological state is compatible with several transcriptomic topologies. This conclusion points to the need of defining a kind of “transcriptomic entropy of a state”.
Author Contributions
Conceptualization, D.A.I., L.X. and D.D.; methodology, S.I. and D.A.I.; software, D.A.I.; validation, S.I. and D.A.I.; formal analysis, D.A.I.; investigation, S.I. and D.A.I.; resources, D.A.I.; data curation, L.X.; writing—original draft preparation, D.A.I; writing—review and editing, S.M. and D.D.; visualization, D.A.I; supervision, D.A.I.; project administration, D.A.I.; funding acquisition, D.A.I. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
microarray protocol and raw data can be found in the databases: https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE17363, https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE18703, https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE24088, https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE29769.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Hinton, A. Jr.; Claypool, S.M.; Neikirk, K.; Senoo, N.; Wanjalla, C.N.; Kirabo, A.; Williams, C.R. Mitochondrial Structure and Function in Human Heart Failure. Circ Res 2024, 135, 372–396. [Google Scholar] [CrossRef] [PubMed]
- Murphy E, Ardehali H, Balaban RS, DiLisa F, Dorn GW 2nd, Kitsis RN, Otsu K, Ping P, Rizzuto R, Sack MN et al. American Heart Association Council on Basic Cardiovascular Sciences, Council on Clinical Cardiology, and Council on Functional Genomics and Translational Biology. Mitochondrial Function, Biology, and Role in Disease: A Scientific Statement From the American Heart Association. Circ Res. 2016, 118, 1960–1991. [Google Scholar] [CrossRef]
- Jia, D.; Hou, L.; Lv, Y.; Xi, L.; Tian, Z. Postinfarction exercise training alleviates cardiac dysfunction and adverse remodeling via mitochondrial biogenesis and SIRT1/PGC-1alpha/PI3K/Akt signaling. J Cell Physiol 2019, 234, 23705–23718. [Google Scholar] [CrossRef] [PubMed]
- Koka, S.; Aluri, H.S.; Xi, L.; Lesnefsky, E.J.; Kukreja, R.C. Chronic inhibition of phosphodiesterase 5 with tadalafil attenuates mitochondrial dysfunction in type 2 diabetic hearts: potential role of NO/SIRT1/PGC-1alpha signaling. Am J Physiol Heart Circ Physiol 2014, 306, H1558–H1568. [Google Scholar] [CrossRef]
- Nunes, J.P.S.; Roda, V.M.P.; Andrieux, P.; Kalil, J.; Chevillard, C.; Cunha-Neto, E. Inflammation and mitochondria in the pathogenesis of chronic Chagas disease cardiomyopathy. Exp Biol Med (Maywood). 2023, 248, 2062–2071. [Google Scholar] [CrossRef]
- Zhu, S.G.; Kukreja, R.C.; Das, A.; Chen, Q.; Lesnefsky, E.J.; Xi, L. Dietary nitrate supplementation protects against Doxorubicin-induced cardiomyopathy by improving mitochondrial function. J Am Coll Cardiol 2011, 57, 2181–2189. [Google Scholar] [CrossRef]
- Schaub, G.A. Trypanosoma cruzi/Triatomine Interactions—A Review. Pathogens 2025, 14, 392. [Google Scholar] [CrossRef]
- Ouali, R.; Bousbata, S. Rhodnius prolixus (kissing bug). Trends Parasitol, 1471. [Google Scholar] [CrossRef]
- Mukherjee, S.; Belbin, T.J.; Spray, D.C.; Iacobas, D.A.; Weiss, L.M.; Kitsis, R.N.; Wittner, M.; Jelicks, L.A.; Scherer, P.E.; Ding, A.; et al. Microarray analysis of changes in gene expression in a murine model of chronic chagasic cardiomyopathy. Parasitol Res 2003, 91, 187–196. [Google Scholar] [CrossRef]
- Lopez, M.; Tanowitz, H.B.; Garg, N.J. Pathogenesis of Chronic Chagas Disease: Macrophages, Mitochondria, and Oxidative Stress. Curr Clin Microbiol Rep, 2018, 5, 45–54. [Google Scholar] [CrossRef]
- Hernandez, S.; Srikanth, K.K.; Bommireddi, A.; Leong, T.K.; Miller, D.A.; Ambrosy, A.P.; Zaroff, J. Chagas Disease in Northern California: Observed Prevalence, Clinical Characteristics, and Outcomes Within an Integrated Health Care Delivery System. Perm J. 2025, 1–9. [Google Scholar] [CrossRef]
- da Silva, J.C.; da Silva, M.T.S.; Bezerra Dos Santos, L.; Abdala, M.G.G.; Lopes, A.B.O.; de Oliveira, G.A.; Guedes-da-Silva, F.H.; Rigoni, T.D.S.; Damasceno, F.S.; Barros-Neto, J.A.; et al. Epidemiological assessment of the first year of mandatory notification of chronic Chagas disease in Alagoas. Northeast Brazil. Acta Trop. 2025, 270, 107805. [Google Scholar] [CrossRef] [PubMed]
- Beatty, N.L.; Hamer, G.L.; Moreno-Peniche, B.; Mayes, B.; Hamer, S.A. Chagas Disease, an Endemic Disease in the United States. Emerg Infect Dis. 2025, 31, 1691–1697. [Google Scholar] [CrossRef] [PubMed]
- Bunkofske, M.E.; Sanchez-Valdez, F.J.; Tarleton, R.L. The importance of persistence and dormancy in Trypanosoma cruzi infection and Chagas disease. Curr Opin Microbiol. 2025, 86, 102615. [Google Scholar] [CrossRef] [PubMed]
- Telleria, J.; Costales, J.A. An Overview of Trypanosoma cruzi Biology Through the Lens of Proteomics: A Review. Pathogens 2025, 14, 337. [Google Scholar] [CrossRef]
- Garg, N.; Popov, V.L.; Papaconstantinou, J. Profiling gene transcription reveals a deficiency of mitochondrial oxidative phosphorylation in Trypanosoma cruzi-infected murine hearts: implications in chagasic myocarditis development. Biochim Biophys Acta 2003, 1638, 106–120. [Google Scholar] [CrossRef]
- Baez, A.; Lo Presti, M.S.; Rivarola, H.W.; Mentesana, G.G.; Pons, P.; Fretes, R.; Paglini-Oliva, P. Mitochondrial involvement in chronic chagasic cardiomyopathy. Trans R Soc Trop Med Hyg 2011, 105, 239–246. [Google Scholar] [CrossRef]
- Echeverría, L.E.; Serrano-García, A.Y.; Rojas, L.Z.; Silva-Sieger, F.; Navarro, M.; Aguilera, L.; Gómez-Ochoa, S.A.; Morillo, C.A. Beyond cardiac embolism and cryptogenic stroke: unveiling the mechanisms of cerebrovascular events in Chagas disease. Lancet Reg Health Am. 2025, 50, 101203. [Google Scholar] [CrossRef]
- Costa, C.J.D.N.; da Silva, P.S.; Saraiva, R.M.; Sangenis, L.H.C.; de Holanda, M.T.; Sperandio da Silva, G.M.; Mendes, F.S.N.S.; Xavier, I.G.G.; Costa, H.S.; Gonçalves, T.R.; et al. Food insecurity is associated with decreased quality of life in patients with chronic Chagas disease. PLoS One. 2025, 20, e0328466. [Google Scholar] [CrossRef]
- Echeverría, L.E.; Serrano-García, A.Y.; Rojas, L.Z.; Silva-Sieger, F.; Navarro, M.; Aguilera, L.; Gómez-Ochoa, S.A.; Morillo, C.A. Beyond cardiac embolism and cryptogenic stroke: unveiling the mechanisms of cerebrovascular events in Chagas disease. Lancet Reg Health Am. 2025, 50, 101203. [Google Scholar] [CrossRef]
- Souza-Silva, T.G.; Figueiredo, A.; Morais, K.L.P.; Apostólico, J.; Pantaleao, A.; Mutarelli, A.; Araújo, S.S.; Nunes, M.D.C.P.; Gollob, K.J.; Dutra, W.O. Single-cell targeted transcriptomics reveals subset-specific immune signatures differentiating asymptomatic and cardiac patients with chronic Chagas disease. J Infect Dis. 2025, jiaf269. [Google Scholar] [CrossRef]
- Duque, C.; So, J.; Castro-Sesquen, Y.E.; DeToy, K.; Gutierrez Guarnizo, S.A.; Jahanbakhsh, F.; Malaga Machaca, E.; Miranda-Schaeubinger, M.; Chakravarti, I.; Cooper, V.; et al.; Chagas Working Group Immunologic changes in the peripheral blood transcriptome of individuals with early-stage chronic Chagas cardiomyopathy: a cross-sectional study. Lancet Reg Health Am. 2025, 45, 101090. [Google Scholar] [CrossRef] [PubMed]
- Soares MB, de Lima RS, Rocha LL, Vasconcelos JF, Rogatto SR, dos Santos RR, Iacobas S, Goldenberg RC, Iacobas DA, Tanowitz HB. et al. Gene expression changes associated with myocarditis and fibrosis in hearts of mice with chronic chagasic cardiomyopathy. J Infect Dis 2010, 202, 416–426. [Google Scholar] [CrossRef] [PubMed]
- Goldenberg, R.C.; Iacobas, D.A.; Iacobas, S.; Rocha, L.L.; da Silva de Azevedo, Fortes, F. ; Vairo, L.; Nagajyothi, F.; Campos de Carvalho. A.C.; Tanowitz, H.B. et al. Transcriptomic alterations in Trypanosoma cruzi-infected cardiac myocytes. Microbes Infect 2009, 11, 1140–1149. [Google Scholar] [CrossRef] [PubMed]
- Adesse, D.; Goldenberg, R.C.; Fortes, F.S.; Jasmin, Iacobas, D. A.; Iacobas, S.; Campos de Carvalho, A.C.; de Narareth Meirelles, M.; Huang, H.; Soares, M.B. et al. Gap junctions and chagas disease. Adv Parasitol 2011, 76, 63–81. [Google Scholar] [CrossRef]
- Xinxin, Z.; Pan, H.; Qiao, L. Research progress of connexin 43 in cardiovascular diseases. Front Cardiovasc Med. 2025, 12, 1650548. [Google Scholar] [CrossRef]
- Caetano-da-Silva, J.E.; Gonçalves-Santos, E.; Domingues, E.L.B.C.; Caldas, I.S.; Lima, G.D.A.; Diniz, L.F.; Gonçalves, R.V.; Novaes, R.D. The mitochondrial uncoupler 2,4-dinitrophenol modulates inflammatory and oxidative responses in Trypanosoma cruzi-induced acute myocarditis in mice. Cardiovasc Pathol. 2024, 72, 107653. [Google Scholar] [CrossRef]
- Nisimura, L.M.; Coelho, L.L.; de Melo, T.G.; Vieira, P.C.; Victorino, P.H.; Garzoni, L.R.; Spray, D.C.; Iacobas, D.A.; Iacobas, S.; Tanowitz, H.B.; et al. Trypanosoma cruzi Promotes Transcriptomic Remodeling of the JAK/STAT Signaling and Cell Cycle Pathways in Myoblasts. Front Cell Infect Microbiol 2020, 10, 255. [Google Scholar] [CrossRef]
- Narimani, S.; Rahbarghazi, R.; Salehipourmehr, H.; Taghavi Narmi, M.; Lotfimehr, H.; Mehdipour, R. Therapeutic Potential of Endothelial Progenitor Cells in Angiogenesis and Cardiac Regeneration: A Systematic Review and Meta-Analysis of Rodent Models. Adv Pharm Bull. 2025, 15, 268–283. [Google Scholar] [CrossRef]
- Soares MB, Lima RS, Souza BS, Vasconcelos JF, Rocha LL, Dos Santos RR, Iacobas S, Goldenberg RC, Lisanti MP, Iacobas DA. et al. Reversion of gene expression alterations in hearts of mice with chronic chagasic cardiomyopathy after transplantation of bone marrow cells. Cell Cycle 2011, 10, 1448–1455. [Google Scholar] [CrossRef]
- Iacobas, D.A.; Iacobas, S.; Tanowitz, H.B.; Campos de Carvalho, A.; Spray, D.C. Functional genomic fabrics are remodeled in a mouse model of Chagasic cardiomyopathy and restored following cell therapy. Microbes Infect 2018, 20, 185–195. [Google Scholar] [CrossRef]
- Saha, T.; Soliman-Aboumarie, H. Review of Current Management of Myocardial Infarction. J Clin Med. 2025, 14, 6241. [Google Scholar] [CrossRef]
- Mueller, C.; White, H.D.; Lopez-Ayala, P.; de Silva, R.; Kaski, J.C. Great debate: the universal definition of myocardial infarction is flawed and should be put to rest. Eur Heart J. 2025, ehaf641. [Google Scholar] [CrossRef] [PubMed]
- Taggart C, Ferry AV, Chapman AR, Schulberg SD, Bularga A, Wereski R, Boeddinghaus J, Kimenai DM, Lowry MTH, Chew DP. et al. The assessment and management of patients with type 2 myocardial infarction: an international Delphi study. Eur Heart J Qual Care Clin Outcomes. [CrossRef]
- Sun, S.; Zhang, Z.; Li, Y.; Zhang, H.; Guo, H.; Chen, G.; Wei, P.; Lin, F.; Zhao, G. Mitochondrial dysfunction as a central hub linking Na+/Ca2+ homeostasis and inflammation in ischemic arrhythmias: therapeutic implications. Front Cardiovasc Med. 2025, 12, 1506501. [Google Scholar] [CrossRef] [PubMed]
- Meco, M.; Giustiniano, E.; Nisi, F.; Zulli, P.; Agosteo, E. MAPK, PI3K/Akt Pathways, and GSK-3β Activity in Severe Acute Heart Failure in Intensive Care Patients: An Updated Review. J. Cardiovasc. Dev. Dis. 2025, 12, 266. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Ji, H.; Modarresi Chahardehi, A. JAK/STAT pathway in myocardial infarction: Crossroads of immune signaling and cardiac remodeling. Mol Immunol. 2025, 186, 206–217. [Google Scholar] [CrossRef]
- Deng, G.; Yang, Y.; Qing, O.; Linhui, J.; Haotao, S.; Liu, C.; Li, G.; Nasser, M.I. Chrysin Attenuates Myocardial Cell Apoptosis in Mice. Cardiovasc Toxicol. [CrossRef]
- Lachtermacher, S.; Esporcatte, B.L.; Montalvao, F.; Costa, P.C.; Rodrigues, D.C.; Belem, L.; Rabischoffisky, A.; Faria Neto, H.C.; Vasconcellos, R.; Iacobas, S. et. Cardiac gene expression and systemic cytokine profile are complementary in a murine model of post-ischemic heart failure. Braz J Med Biol Res 2010, 43, 377–389. [Google Scholar] [CrossRef]
- Lachtermacher, S.; Esporcatte, B.L.; Fortes, Fda, S. ; Rocha, N.N.; Montalvao, F.; Costa, P.C.; Belem, L.; Rabischoffisky, A.; Faria Neto, H.C.; Vasconcellos, R. et al. Functional and transcriptomic recovery of infarcted mouse myocardium treated with bone marrow mononuclear cells. Stem Cell Rev Rep 2012, 8, 251–261. [Google Scholar] [CrossRef]
- Patel, P.; Yang, F.; Iacobas, D.A.; Xi, L. Mental disorders after myocardial infarction: potential mediator role for chemokines in heart-brain interaction? J Geriatr Cardiol 2024, 21, 913–926. [Google Scholar] [CrossRef]
- Oxidative phosphorilation. Available online: https://www.kegg.jp/pathway/mmu00190 (accessed on 1 September 2025).
- Diabetic cardiomyopathy. Available online: https://www.genome.jp/pathway/mmu05415 (accessed on 1 September 2025).
- Gene expression changes associated with myocarditis and fibrosis in hearts of mice with chronic chagasic cardiomyopathy. Available online: https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE17363 (accessed on 1 September 2025).
- Cardiac gene expression and systemic cytokine profile are complementary in a murine model of post ischemic heart failure. Available online: https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE18703 (accessed on 1 September 2025).
- Therapy with bone marrow cells recovers gene expression alterations in hearts of mice with chronic chagasic cardiomyopathy. Available online: https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE24088 (accessed on 1 September 2025).
- Functional and Transcriptomic Recovery of Infarcted Mouse Myocardium Treated with Bone Marrow Mononuclear Cells. Available online at: https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE29769. Accessed on Sept 1st, 2025.
- Adesse, D.; Iacobas, D.A.; Iacobas, S.; Garzoni, L.R.; Meirelles Mde, N.; Tanowitz, H.B.; Spray. D.C. Transcriptomic signatures of alterations in a myoblast cell line infected with four distinct strains of Trypanosoma cruzi. Am J Trop Med Hyg 2010, 82, 846–854. [Google Scholar] [CrossRef]
- Iacobas, D.A.; Iacobas, S.; Lee, P.R.; Cohen, J.E.; Fields, R.D. Coordinated Activity of Transcriptional Networks Responding to the Pattern of Action Potential Firing in Neurons. Genes. 2019, 10, 754. [Google Scholar] [CrossRef]
- Iacobas, D.A. Biomarkers, Master Regulators and Genomic Fabric Remodeling in a Case of Papillary Thyroid Carcinoma. Genes. 2020, 11, 1030. [Google Scholar] [CrossRef]
- Iacobas, D.A. ; Xi, L Theory and Applications of the (Cardio) Genomic Fabric Approach to Post-Ischemic and Hypoxia-Induced Heart Failure. J Pers Med 2022, 12. [Google Scholar] [CrossRef]
- Parker, A.M.; Lees, J.G.; Murray, A.J.; Velagic, A.; Lim, S.Y.; De Blasio, M.J.; Ritchie, R.H. Precision Medicine: Therapeutically Targeting Mitochondrial Alterations in Heart Failure. JACC Basic Transl Sci. 2025, 10, 101345. [Google Scholar] [CrossRef]
- Mongelli, A.; Mengozzi, A.; Geiger, M.; Gorica, E.; Mohammed, S.A.; Paneni, F.; Ruschitzka, F.; Costantino, S. ; Mitochondrial epigenetics in aging and cardiovascular diseases. Front Cardiovasc Med. 2023, 10, 1204483. [Google Scholar] [CrossRef]
- Pietrangelo, D.; Lopa, C.; Litterio, M.; Cotugno, M.; Rubattu, S.; Lombardi, A. Metabolic Disturbances Involved in Cardiovascular Diseases: The Role of Mitochondrial Dysfunction, Altered Bioenergetics and Oxidative Stress. Int J Mol Sci. 2025, 26, 6791. [Google Scholar] [CrossRef]
- Dhalla, N.S.; Ostadal, P.; Tappia, P.S. Involvement of Oxidative Stress in Mitochondrial Abnormalities During the Development of Heart Disease. Biomedicines. 2025, 13, 1338. [Google Scholar] [CrossRef]
- Mueller, M.; Blandino, A.; Scherer, D.; Zulantay, I.; Apt, W.; Varela, N.M.; Llancaqueo, M.; Garcia, L.; Ortiz, L.; Nicastri, E.; et al. Small-RNA sequencing identifies serum microRNAs associated with abnormal electrocardiography findings in patients with Chagas disease. J Infect. 2025, 106613. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Xu, H. Diagnosis and treatment of post-acute myocardial infarction ventricular aneurysm: A review. Medicine 2025, 104, e43696. [Google Scholar] [CrossRef] [PubMed]
- Formosa, L.E.; Muellner-Wong, L.; Reljic, B.; Sharpe, A.J.; Jackson, T.D.; Beilharz, T.H.; Stojanovski, D.; Lazarou, M.; Stroud, D.A.; Ryan, M.T. Dissecting the Roles of Mitochondrial Complex I Intermediate Assembly Complex Factors in the Biogenesis of Complex I. Cell Rep. 2020, 31, 107541. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Zhao, H.; Luo, L.; Wu, W. Nicotinamide Adenine Dinucleotide Supplementation to Alleviate Heart Failure: A Mitochondrial Dysfunction Perspective. Nutrients. 2025, 17, 1855. [Google Scholar] [CrossRef]
- Bozdemir, N.; Cakir, C.; Topcu, U.; Uysal, F. A Comprehensive Review of Mitochondrial Complex I During Mammalian Oocyte Maturation. Genesis. 2025, 63, e70017. [Google Scholar] [CrossRef]
- Shi, X.; Zhang, Y.; Chen, R.; Gong, Y.; Zhang, M.; Guan, R.; Rotstein, O.D.; Liu, X.; Wen, X.Y. ndufa7 plays a critical role in cardiac hypertrophy. J Cell Mol Med. 2020, 24, 13151–13162. [Google Scholar] [CrossRef]
- Wyckelsma, V.L.; Levinger, I.; McKenna, M.J.; Formosa, L.E.; Ryan, M.T.; Petersen, A.C.; Anderson, M.J.; Murphy, R.M. Preservation of skeletal muscle mitochondrial content in older adults: relationship between mitochondria, fibre type and high-intensity exercise training. J Physiol. 2017, 595, 3345–3359. [Google Scholar] [CrossRef]
- Guo, P.; Hu, S.; Liu, X.; He, M.; Li, J.; Ma, T.; Huang, M.; Fang, Q.; Wang, Y. CAV3 alleviates diabetic cardiomyopathy via inhibiting NDUFA10-mediated mitochondrial dysfunction. J Transl Med 2024, 22, 390. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Jin, X.; Xia, L.; Wang, X.; Yu, Y.; Liu, C.; Shao, D.; Fang, N.; Meng, C. Characterization of mitochondrial NADH dehydrogenase 1alpha subcomplex 10 variants in cardiac muscles from normal Wistar rats and spontaneously hypertensive rats: Implications in the pathogenesis of hypertension. Mol Med Rep 2016, 13, 961–966. [Google Scholar] [CrossRef] [PubMed]
- Iverson, T.M.; Singh, P.K.; Cecchini, G. An evolving view of complex II-noncanonical complexes, megacomplexes, respiration, signaling, and beyond. J Biol Chem. 2023, 299, 104761. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Fasham, J.; Al-Hijawi, F.; Qutob, N.; Gunning, A.; Leslie, J.S.; McGavin, L.; Ubeyratna, N.; Baker, W.; Zeid, R.; et al. Consolidating biallelic SDHD variants as a cause of mitochondrial complex II deficiency. Eur J Hum Genet. 2021, 29, 1570–1576. [Google Scholar] [CrossRef]
- Zhang, Y.; Wei, J.; Qiao, L.; Yu, R.; Ren, H.; Zhao, A.; Sun, Y.; Wang, A.; Li, B.; Wang, X. et. Exploring the biological basis for the identification of different syndromes in ischemic heart failure based on joint multi-omics analysis. Front Pharmacol. 2025, 16, 1641422. [Google Scholar] [CrossRef]
- Spielmann, N.; Schenkl, C.; Komlodi, T.; da Silva-Buttkus, P.; Heyne, E.; Rohde, J.; Amarie, O.V.; Rathkolb, B.; Gnaiger, E.; Doenst, T.; et al. Knockout of the Complex III subunit Uqcrh causes bioenergetic impairment and cardiac contractile dysfunction. Mamm Genome 2023, 34, 229–243. [Google Scholar] [CrossRef]
- Čunátová, K.; Fernández-Vizarra, E. Pathological variants in nuclear genes causing mitochondrial complex III deficiency: An update. J Inherit Metab Dis. 2024, 47, 1278–1291. [Google Scholar] [CrossRef]
- Sinkler, C.A.; Kalpage, H.; Shay, J.; Lee, I.; Malek, M.H. Grossman, L.I.; Huttemann, M. Tissue- and Condition-Specific Isoforms of Mammalian Cytochrome c Oxidase Subunits: From Function to Human Disease. Oxid Med Cell Longev 2017, 1534056. [Google Scholar] [CrossRef]
- Wen, J.J.; Garg, N. Oxidative modification of mitochondrial respiratory complexes in response to the stress of Trypanosoma cruzi infection. Free Radic Biol Med. 2004, 37, 2072–2081. [Google Scholar] [CrossRef]
- Pandey, R.; Velasquez, S.; Durrani, S.; Jiang, M.; Neiman, M.; Crocker, J.S.; Benoit, J.B.; Rubinstein, J.; Paul, A.; Ahmed, R.P. MicroRNA-1825 induces proliferation of adult cardiomyocytes and promotes cardiac regeneration post-ischemic injury. Am J Transl Res 2017, 9, 3120–3137. [Google Scholar]
- Elbatarny, M.; Lu, Y.T.; Hu, M.; Coles, J.; Mital, S.; Ross-White, A.; Honjo, O.; Barron, D.J.; Gramolini, A.O. Systems biology approaches investigating mitochondrial dysfunction in cyanotic heart disease: a systematic review. EBioMedicine 2025, 118, 105839. [Google Scholar] [CrossRef]
- Qiu, J.; Gu, Y. Analysis of the prognostic value of mitochondria-related genes in patients with acute myocardial infarction. BMC Cardiovasc Disord. 2024, 24, 408. [Google Scholar] [CrossRef] [PubMed]
- Hejzlarova, K.; Mracek, T.; Vrbacky, M.; Kaplanova, V.; Karbanova, V.; Nuskova, H.; Pecina, P.; Houstek, J. Nuclear genetic defects of mitochondrial ATP synthase. Physiol Res 2014, 63, S57–S71. [Google Scholar] [CrossRef] [PubMed]
- Mayr, J.A.; Havlickova, V.; Zimmermann, F.; Magler, I.; Kaplanova, V.; Jesina, P.; Pecinova, A.; Nuskova, H.; Koch, J.; Sperl, W.; et al. Mitochondrial ATP synthase deficiency due to a mutation in the ATP5E gene for the F1 epsilon subunit. Hum Mol Genet 2010, 19, 3430–3439. [Google Scholar] [CrossRef] [PubMed]
- Pereyra, A.S.; Hasek, L.Y.; Harris, K.L.; Berman, A.G.; Damen, F.W.; Goergen, C.J.; Elli,s J. M. Loss of cardiac carnitine palmitoyltransferase 2 results in rapamycin-resistant, acetylation-independent hypertrophy. J Biol Chem 2017, 292, 18443–18456. [Google Scholar] [CrossRef]
- Giménez-Escamilla, I.; Benedicto, C.; Pérez-Carrillo, L.; Delgado-Arija, M.; González-Torrent, I.; Vilchez, R.; Martínez-Dolz, L.; Portolés, M.; Tarazón, E.; Roselló-Lletí, E. Alterations in Mitochondrial Oxidative Phosphorylation System: Relationship of Complex V and Cardiac Dysfunction in Human Heart Failure. Antioxidants 2024, 13, 285. [Google Scholar] [CrossRef]
- Qi, B.; Song, L.; Hu, L.; Guo, D.; Ren, G.; Peng, T.; Liu, M.; Fang, Y.; Li, C.; Zhang, M.; et al. Cardiac-specific overexpression of Ndufs1 ameliorates cardiac dysfunction after myocardial infarction by alleviating mitochondrial dysfunction and apoptosis. Exp Mol Med. 2022, 54, 946–960. [Google Scholar] [CrossRef]
- Shao, M.; Li, L.; Ma, L.; Song, C.; Li, W.; Zhang, Y.; Cheng, W.; Chen, Y.; Yang, Y.; Wang, Q.; et al. Activation of RXRα exerts cardioprotection through transcriptional upregulation of Ndufs4 in heart failure. Sci Bull 2024, 69, 1202–1207. [Google Scholar] [CrossRef]
- Guo, P.; Hu, S.; Liu, X.; He, M.; Li, J.; Ma, T.; Huang, M.; Fang, Q.; Wang, Y. CAV3 alleviates diabetic cardiomyopathy via inhibiting NDUFA10-mediated mitochondrial dysfunction. J Transl Med. 2024, 22, 390. [Google Scholar] [CrossRef]
- Friederich MW, Erdogan AJ, Coughlin CR, 2nd, Elos MT, Jiang H, O'Rourke CP, Lovell MA, Wartchow E, Gowan K, Chatfield KC, Chick WS, Spector EB, Van Hove JLK and Riemer J. Mutations in the accessory subunit NDUFB10 result in isolated complex I deficiency and illustrate the critical role of intermembrane space import for complex I holoenzyme assembly. Hum Mol Genet 2017, 26, 702–716. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, Y.; Wan, J.; Zhang, P.; Pei, F. COX6B1 relieves hypoxia/reoxygenation injury of neonatal rat cardiomyocytes by regulating mitochondrial function. Biotechnol Lett. 2019, 41, 59–68. [Google Scholar] [CrossRef]
- Nogueira, S.S.; Souza, M.A.; Santos, E.C.; Caldas, I.S.; Gonçalves, R.V.; Novaes, R.D. Oxidative stress, cardiomyocytes premature senescence and contractile dysfunction in in vitro and in vivo experimental models of Chagas disease. Acta Trop. 2023, 244, 106950. [Google Scholar] [CrossRef] [PubMed]
- Santos-Miranda, A.; Joviano-Santos, J.V.; Ribeiro, G.A.; Botelho, A.F.M.; Rocha, P.; Vieira, L.Q.; Cruz, J.S.; Roman-Campos, D. Reactive oxygen species and nitric oxide imbalances lead to in vivo and in vitro arrhythmogenic phenotype in acute phase of experimental Chagas disease. PLoS Pathog 2020, 16, e1008379. [Google Scholar] [CrossRef] [PubMed]
- Hernansanz-Agustín, P.; Enríquez, J.A. Generation of Reactive Oxygen Species by Mitochondria. Antioxidants 2021, 10, 415. [Google Scholar] [CrossRef] [PubMed]
- Frade, A.F.; Guerin, H.; Nunes, J.P.S.; Silva, L.; Roda, V.M.P.; Madeira, R.P.; Brochet, P.; Andrieux, P.; Kalil, J.; Chevillard, C.; et al. Cardiac and Digestive Forms of Chagas Disease: An Update on Pathogenesis, Genetics, and Therapeutic Targets. Mediators Inflamm 2025, 8862004. [Google Scholar] [CrossRef]
- Ma, Y.H.; Yang, Y.; Li, J.H.; Yao, B.C.; Chen, Q.L.; Wang, L.Q.; Guo, Z.G.; Guo, S.Z. NDUFB11 and NDUFS3 regulate arterial atherosclerosis and venous thrombosis: Potential markers of atherosclerosis and venous thrombosis. Medicine 2023, 102, e36133. [Google Scholar] [CrossRef]
- Nagajyothi, J.F.; Weiss, L.M. Advances in understanding the role of adipose tissue and mitochondrial oxidative stress in Trypanosoma cruzi infection. F1000Res. 2019, 8, F1000. [Google Scholar] [CrossRef]
- Abdel-Aziz, A.K. OXPHOS mediators in acute myeloid leukemia patients: Prognostic biomarkers and therapeutic targets for personalized medicine. World J Surg Oncol. 2024, 22, 298. [Google Scholar] [CrossRef]
- Elkenani, M.; Barallobre-Barreiro, J.; Schnelle, M.; Mohamed, B.A.; Beuthner, B.E.; Jacob, C.F.; Paul, N.B.; Yin, X.; Theofilatos, K.; Fischer, A. et. Cellular and extracellular proteomic profiling of paradoxical low-flow low-gradient aortic stenosis myocardium. Front Cardiovasc Med. 2024, 11, 1398114. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Bai, J.; Zhong, S.; Zhang, R.; Zhang, X.; Xu, Y.; Zhao, M.; Zhao, C.; Zhou, Z. Molecular Signatures of Mitochondrial Complexes Involved in Alzheimer's Disease via Oxidative Phosphorylation and Retrograde Endocannabinoid Signaling Pathways. Oxid Med Cell Longev. 2022, 2022, 9565545. [Google Scholar] [CrossRef] [PubMed]
- Sparks, L.M.; Xie, H.; Koza, R.A.; Mynatt, R.; Hulver, M.W.; Bray, G.A.; Smith, S.R. A high-fat diet coordinately downregulates genes required for mitochondrial oxidative phosphorylation in skeletal muscle. Diabetes. 2005, 54, 1926–1933. [Google Scholar] [CrossRef] [PubMed]
- Social Science Statistics. Available online: https://www.socscistatistics.com/pvalues/pearsondistribution.aspx (accessed on 1 September 2025).
- Ahmad, B.; Dumbuya, J.S.; Li, W.; Tang, J.X.; Chen, X.; Lu, J. Evaluation of GFM1 mutations pathogenicity through in silico tools, RNA sequencing and mitophagy pahtway in GFM1 knockout cells. Int J Biol Macromol. 2025, 304 Pt 2 Pt 2, 140970. [Google Scholar] [CrossRef]
- Lemeshko, V.V. Mechanism of Na+ ions contribution to the generation and maintenance of a high inner membrane potential in mitochondria. Biochim Biophys Acta Bioenerg. 2025, 149571. [Google Scholar] [CrossRef]
- Fedor, J.G.; Jones, A.J.Y.; Di Luca, A.; Kaila, V.R.I.; Hirst, J. Correlating kinetic and structural data on ubiquinone binding and reduction by respiratory complex I. Proc Natl Acad Sci U S A. 2017, 114, 12737–12742. [Google Scholar] [CrossRef]
- Galemou Yoga, E.; Haapanen, O.; Wittig, I.; Siegmund, K.; Sharma, V.; Zickermann, V. Mutations in a conserved loop in the PSST subunit of respiratory complex I affect ubiquinone binding and dynamics. Biochim Biophys Acta Bioenerg. 2019, 1860, 573–581. [Google Scholar] [CrossRef]
- Röpke, M.; Saura, P.; Riepl, D.; Pöverlein, M.C.; Kaila, V.R.I. Functional Water Wires Catalyze Long-Range Proton Pumping in the Mammalian Respiratory Complex I. J Am Chem Soc. 2020, 142, 21758–21766. [Google Scholar] [CrossRef]
- Chen, P.; Yawar, W.; Farooqui, A.R.; Ali, S.; Lathiya, N.; Ghous, Z.; Sultan, R.; Alhomrani, M.; Alghamdi, S.A.; Almalki, A.A.; et al. Transcriptomics data integration and analysis to uncover hallmark genes in hypertrophic cardiomyopathy. Am J Transl Res. 2024, 16, 637–653. [Google Scholar] [CrossRef]
- Iacobas, S.; Ede, N.; Iacobas, D.A. The Gene Master Regulators (GMR) Approach Provides Legitimate Targets for Personalized, Time-Sensitive Cancer Gene Therapy. Genes. 2019, 10, 560. [Google Scholar] [CrossRef]
Figure 1.
Significantly regulated mitochondrial genes in: (a) Chagasic disease and in (b) post-ischemic heart failure. Regulated genes: Atp5f1d/e (ATP synthase, H+ transporting, mitochondrial F1 complex, delta/epsilon subunit), Atp5mc2 (ATP synthase membrane subunit c locus 2), Atp5pb/o (ATP synthase peripheral Uqcrh stalk-membrane subunit b/OSCP), Cox4i1 (Cytochrome c oxidase subunit IV isoform 1), Cox17 (Cytochrome c oxidase, subunit XVII assembly protein homolog (yeast)), Cox5a (Cytochrome c oxidase, subunit Va), Cox6a1/6b1 (Cytochrome c oxidase, subunit VI a/b, polypeptide 1), Cox7a2/b (Cytochrome c oxidase, subunit VIIa 2/VIIb), Cpt2 (Carnitine palmitoyltransferase 2), Cyc1 (Cytochrome c-1), Ndufa6/7/12 (NADH:ubiquinone oxidoreductase subunit A6/7/12), Ndufb8/9/10 (NADH: ubiquinone oxidoreductase subunit beta8/9/10), Ndufc1/2 (NADH:ubiquinone oxidoreductase subunit C1/C2), Ndufs1/4/6/7/8 (NADH dehydrogenase (ubiquinone) Fe-S protein 1/4/6/7/8), Ndufv3 (NADH:ubiquinone oxidoreductase core subunit V3), Pdha1 (Pyruvate dehydrogenase E1 alpha 1), Pdk2 (Pyruvate dehydrogenase kinase, isoenzyme 2), Sdha/b (Succinate dehydrogenase Complex, subunit A/B), Slc25a5 (Solute carrier family 25 (mitochondrial carrier, adenine nucleotide translocator), member 5), Uqcrc2 (Ubiquinol cytochrome c reductase core protein 2), Uqcrfs1 (Ubiquinol-cytochrome c reductase, Rieske iron-sulfur polypeptide 1), Uqcrh (Ubiquinol-cytochrome c reductase hinge protein), Uqcrb/h (Ubiquinol-cytochrome c reductase binding/hinge protein), Vdac1/2 (Voltage-dependent anion channel 1/2).
Figure 1.
Significantly regulated mitochondrial genes in: (a) Chagasic disease and in (b) post-ischemic heart failure. Regulated genes: Atp5f1d/e (ATP synthase, H+ transporting, mitochondrial F1 complex, delta/epsilon subunit), Atp5mc2 (ATP synthase membrane subunit c locus 2), Atp5pb/o (ATP synthase peripheral Uqcrh stalk-membrane subunit b/OSCP), Cox4i1 (Cytochrome c oxidase subunit IV isoform 1), Cox17 (Cytochrome c oxidase, subunit XVII assembly protein homolog (yeast)), Cox5a (Cytochrome c oxidase, subunit Va), Cox6a1/6b1 (Cytochrome c oxidase, subunit VI a/b, polypeptide 1), Cox7a2/b (Cytochrome c oxidase, subunit VIIa 2/VIIb), Cpt2 (Carnitine palmitoyltransferase 2), Cyc1 (Cytochrome c-1), Ndufa6/7/12 (NADH:ubiquinone oxidoreductase subunit A6/7/12), Ndufb8/9/10 (NADH: ubiquinone oxidoreductase subunit beta8/9/10), Ndufc1/2 (NADH:ubiquinone oxidoreductase subunit C1/C2), Ndufs1/4/6/7/8 (NADH dehydrogenase (ubiquinone) Fe-S protein 1/4/6/7/8), Ndufv3 (NADH:ubiquinone oxidoreductase core subunit V3), Pdha1 (Pyruvate dehydrogenase E1 alpha 1), Pdk2 (Pyruvate dehydrogenase kinase, isoenzyme 2), Sdha/b (Succinate dehydrogenase Complex, subunit A/B), Slc25a5 (Solute carrier family 25 (mitochondrial carrier, adenine nucleotide translocator), member 5), Uqcrc2 (Ubiquinol cytochrome c reductase core protein 2), Uqcrfs1 (Ubiquinol-cytochrome c reductase, Rieske iron-sulfur polypeptide 1), Uqcrh (Ubiquinol-cytochrome c reductase hinge protein), Uqcrb/h (Ubiquinol-cytochrome c reductase binding/hinge protein), Vdac1/2 (Voltage-dependent anion channel 1/2).
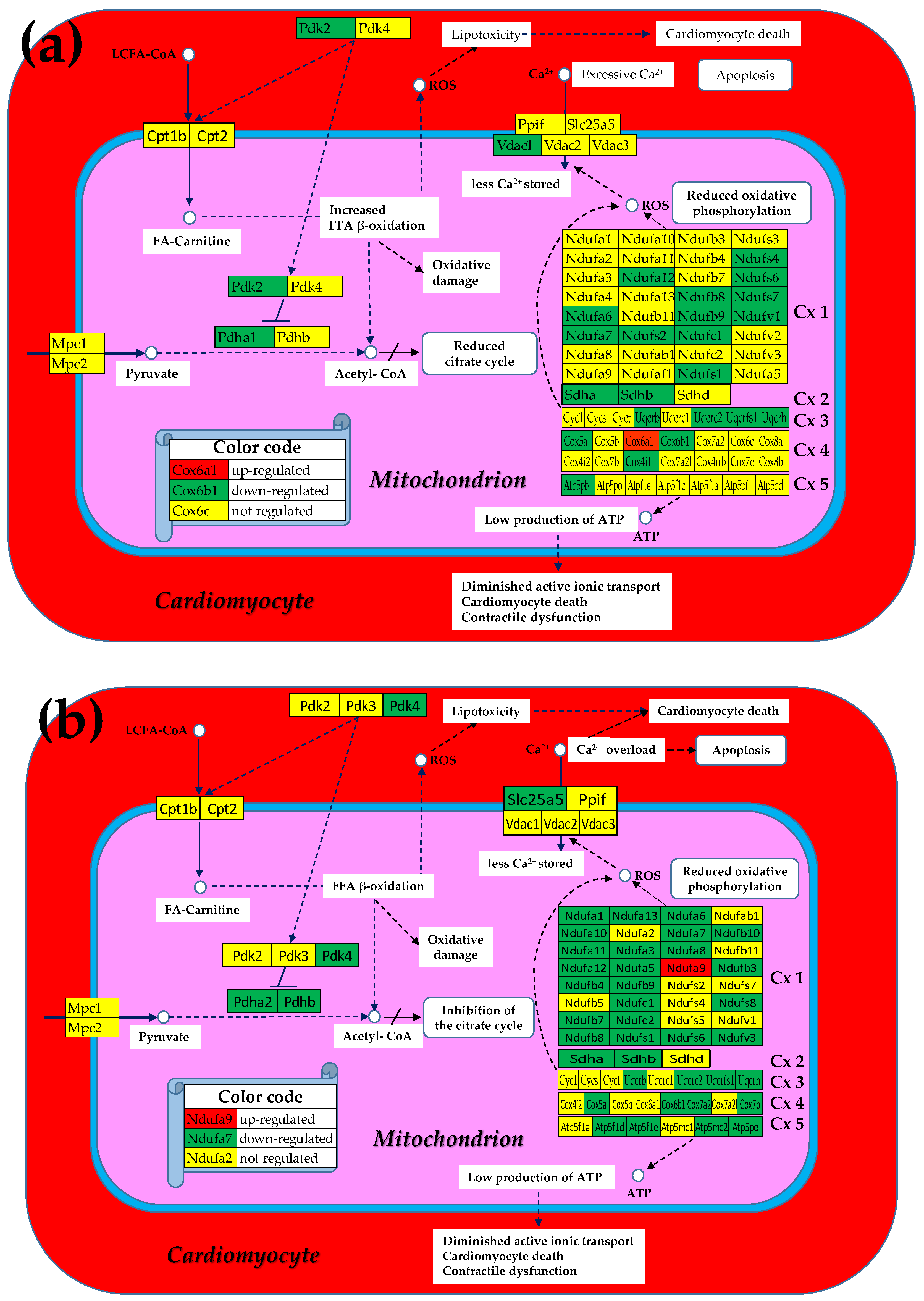
Figure 2.
Remaining significantly regulated mitochondrial genes after stem cell treatment in: (a) Chagase disease cardiomyopathy and in (b) post-ischemic heart failure. Regulated genes: Atp5po (ATP synthase peripheral Uqcrh stalk-membrane subunit OSCP), Cpt2 (Carnitine palmitoyltransferase 2), Ndufb1/10/11 (NADH: ubiquinone oxidoreductase subunit beta1/10/11), Ndufc2 (NADH:ubiquinone oxidoreductase subunit C2), Ndufs1/4/8 (NADH dehydrogenase (ubiquinone) Fe-S protein 1/4/8), Uqcrh (Ubiquinol-cytochrome c reductase hinge protein), Vdac2 (Voltage-dependent anion channel 2).
Figure 2.
Remaining significantly regulated mitochondrial genes after stem cell treatment in: (a) Chagase disease cardiomyopathy and in (b) post-ischemic heart failure. Regulated genes: Atp5po (ATP synthase peripheral Uqcrh stalk-membrane subunit OSCP), Cpt2 (Carnitine palmitoyltransferase 2), Ndufb1/10/11 (NADH: ubiquinone oxidoreductase subunit beta1/10/11), Ndufc2 (NADH:ubiquinone oxidoreductase subunit C2), Ndufs1/4/8 (NADH dehydrogenase (ubiquinone) Fe-S protein 1/4/8), Uqcrh (Ubiquinol-cytochrome c reductase hinge protein), Vdac2 (Voltage-dependent anion channel 2).
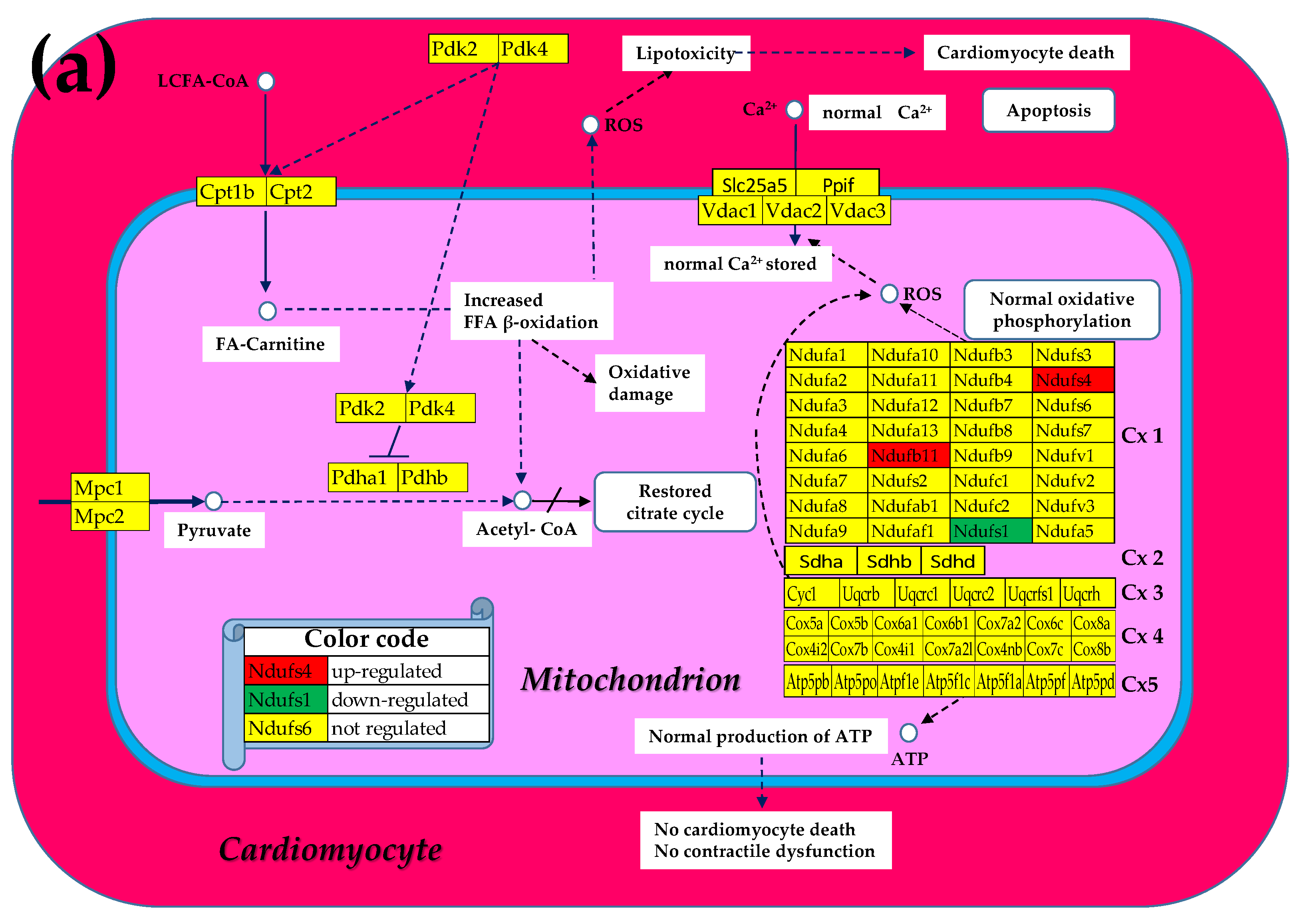
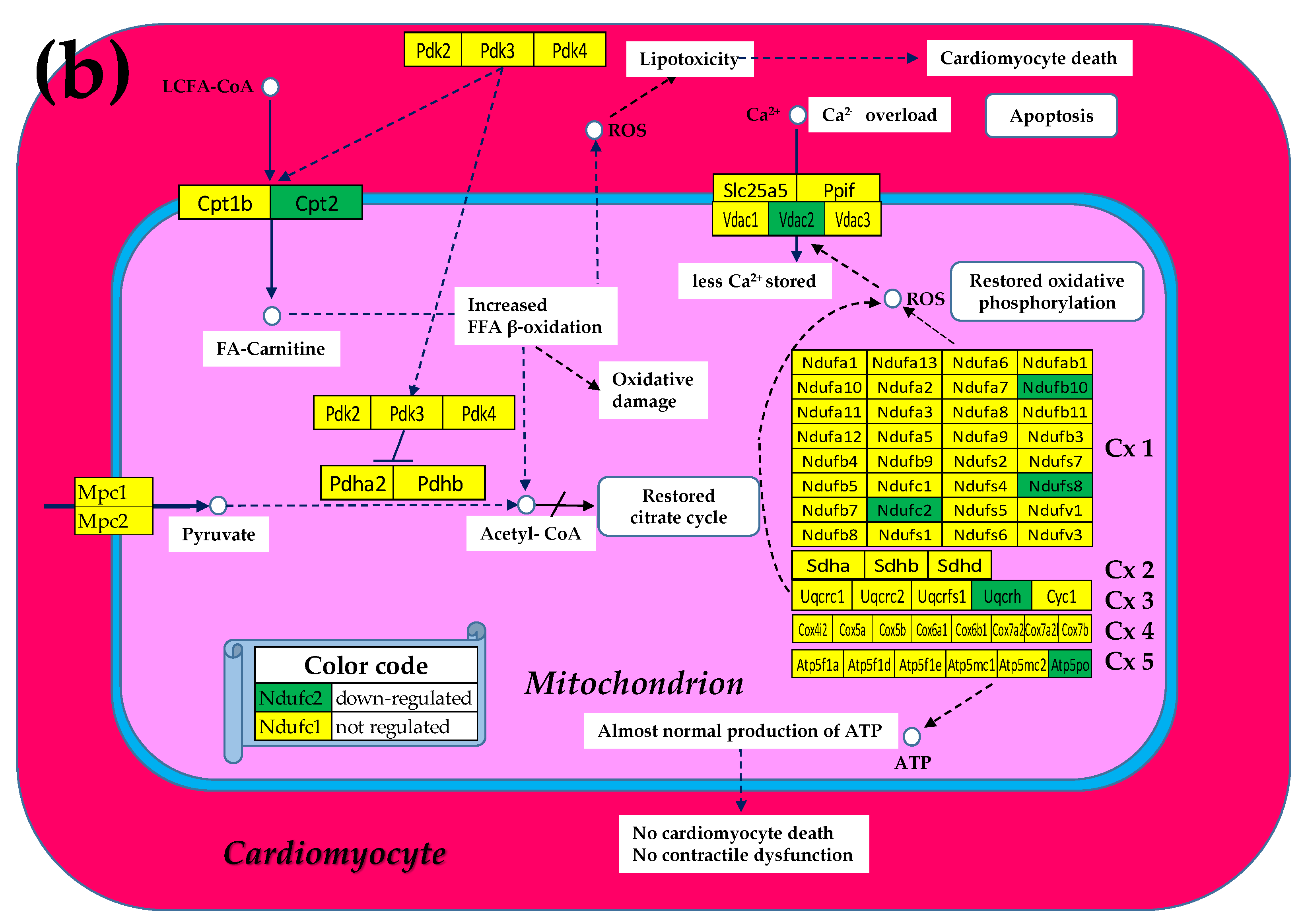
Figure 3.
The (p < 0.05) significant transcriptomic networks coupling the five complexes of the oxidative phosphorylation in the heart left ventricles of healthy and untreated CCC and IHF mice. (a) Healthy mice, (b) untreated CCC mice, (c) untreated IHF mice. Continuous red/blue lines indicate (p < 0.05) significant synergistic/antagonistic expression correlations of the paired genes, while dashed black lines point to significantly independently expressed gene pairs. Missing lines means that the expression correlation was not 0.05 statistically significant. Red/green background of the gene symbols indicates significant up-/down-regulation, while yellow background means not significant change of the expression level with respect to the healthy mice.
Figure 3.
The (p < 0.05) significant transcriptomic networks coupling the five complexes of the oxidative phosphorylation in the heart left ventricles of healthy and untreated CCC and IHF mice. (a) Healthy mice, (b) untreated CCC mice, (c) untreated IHF mice. Continuous red/blue lines indicate (p < 0.05) significant synergistic/antagonistic expression correlations of the paired genes, while dashed black lines point to significantly independently expressed gene pairs. Missing lines means that the expression correlation was not 0.05 statistically significant. Red/green background of the gene symbols indicates significant up-/down-regulation, while yellow background means not significant change of the expression level with respect to the healthy mice.
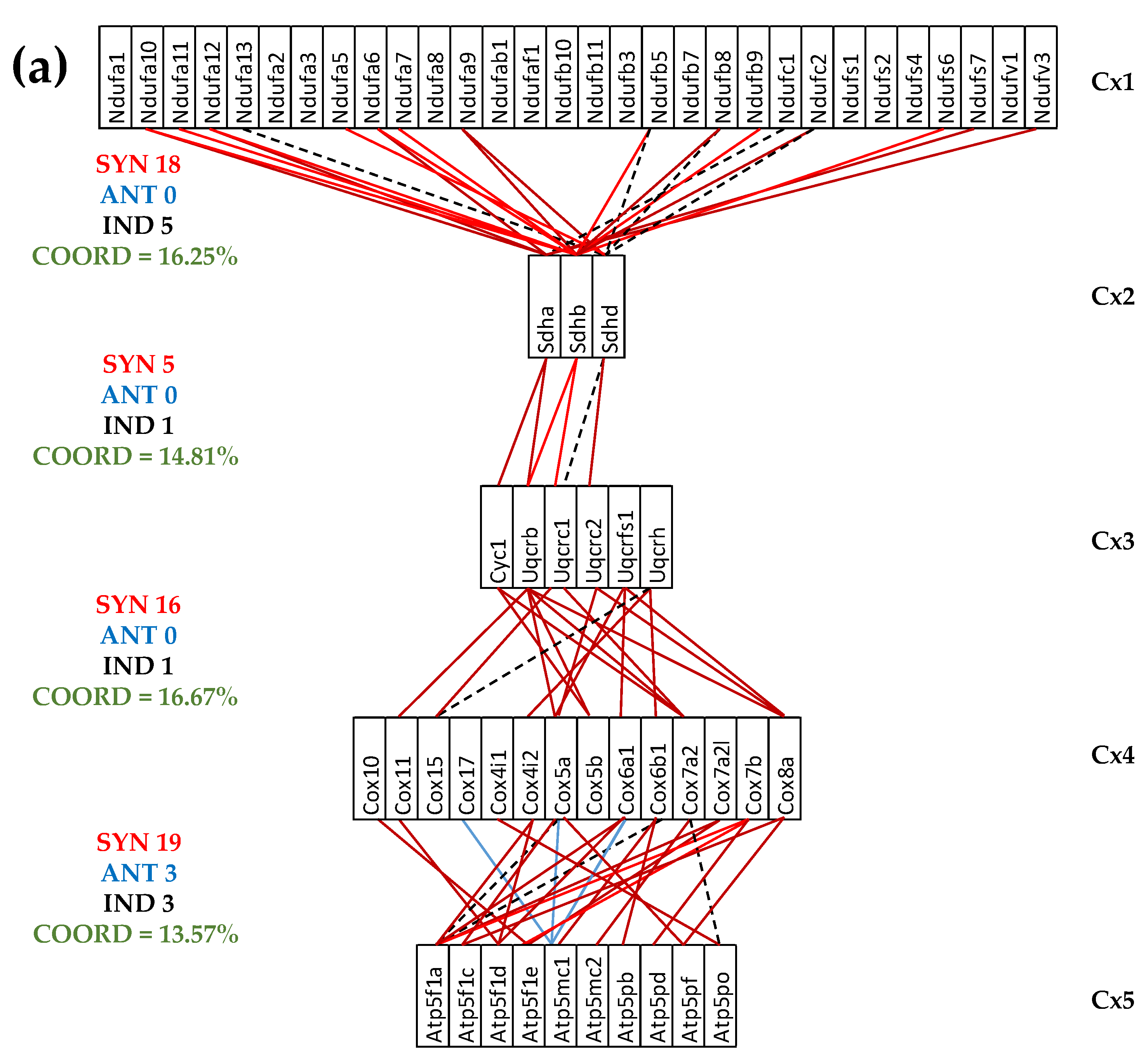
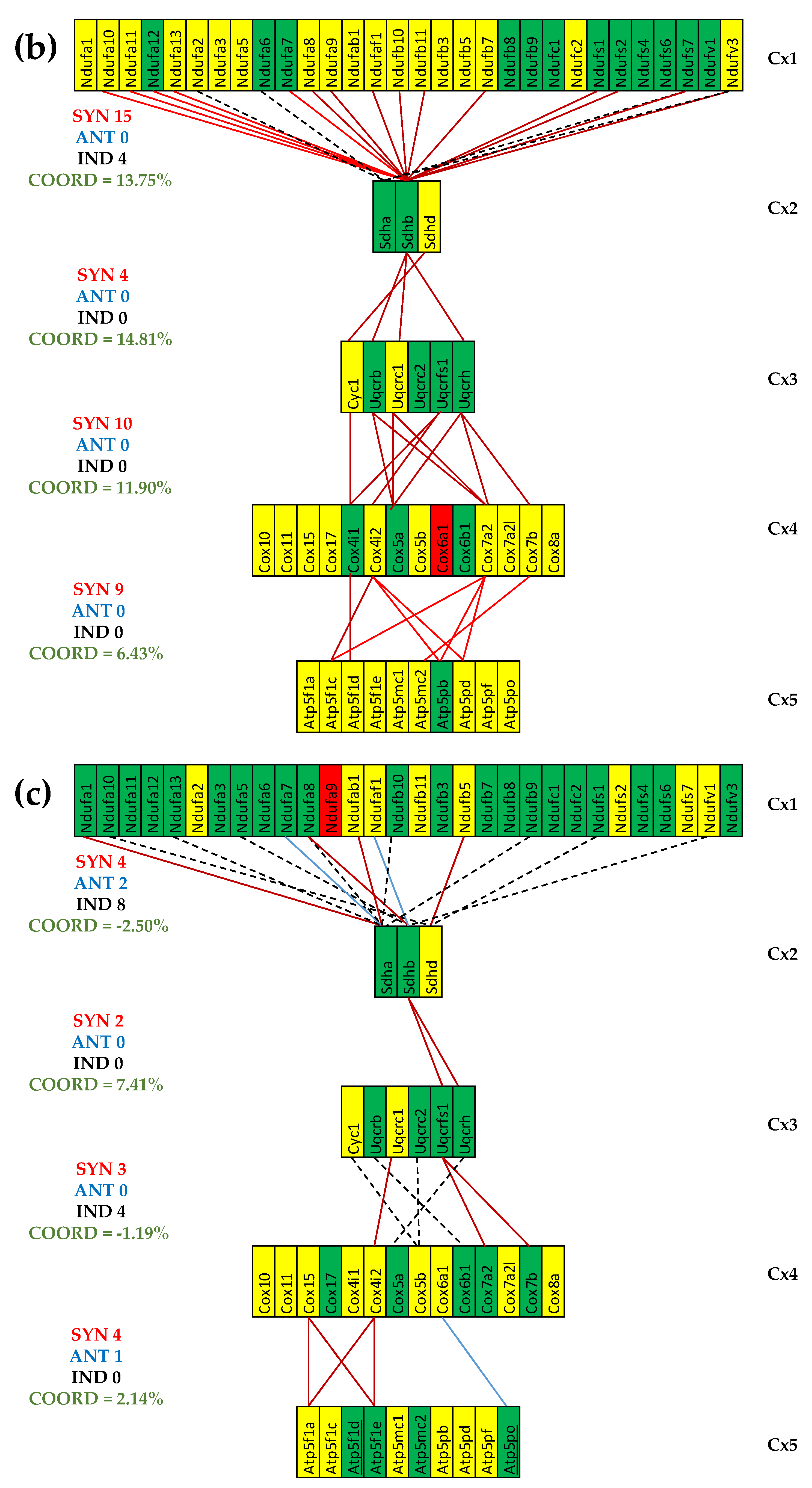
Figure 4.
The (p < 0.05) significant transcriptomic networks coupling the five complexes of the oxidative phosphorylation in the heart left ventricles of treated CCC and IHF mice. (a) CCC treated mice, (b) IHF treated mice, (c) coordination degrees between adjacent complexes of the oxidative phosphorylation functional pathway in the left heart ventricle of healthy (denoted by CN) untreated CCC (CCI) and IHF (IHI) and treated CCC (CCT) and IHF (IHT) mice. Continuous red/blue lines indicate (p < 0.05) significant synergistic/antagonistic expression correlations of the paired genes, while dashed black lines point to independently expressed gene pairs. Red/green background of the gene symbols indicates significant up-/down-regulation, while yellow background means not significant change of the expression level with respect to the healthy mice. Note the coordination reduction in both untreated cardiomyopathies, reaching even negative values for two pairs of complexes (Cx1-Cx2, Cx3-Cx4) in IHF mice. (e) IHF treated mice.
Figure 4.
The (p < 0.05) significant transcriptomic networks coupling the five complexes of the oxidative phosphorylation in the heart left ventricles of treated CCC and IHF mice. (a) CCC treated mice, (b) IHF treated mice, (c) coordination degrees between adjacent complexes of the oxidative phosphorylation functional pathway in the left heart ventricle of healthy (denoted by CN) untreated CCC (CCI) and IHF (IHI) and treated CCC (CCT) and IHF (IHT) mice. Continuous red/blue lines indicate (p < 0.05) significant synergistic/antagonistic expression correlations of the paired genes, while dashed black lines point to independently expressed gene pairs. Red/green background of the gene symbols indicates significant up-/down-regulation, while yellow background means not significant change of the expression level with respect to the healthy mice. Note the coordination reduction in both untreated cardiomyopathies, reaching even negative values for two pairs of complexes (Cx1-Cx2, Cx3-Cx4) in IHF mice. (e) IHF treated mice.
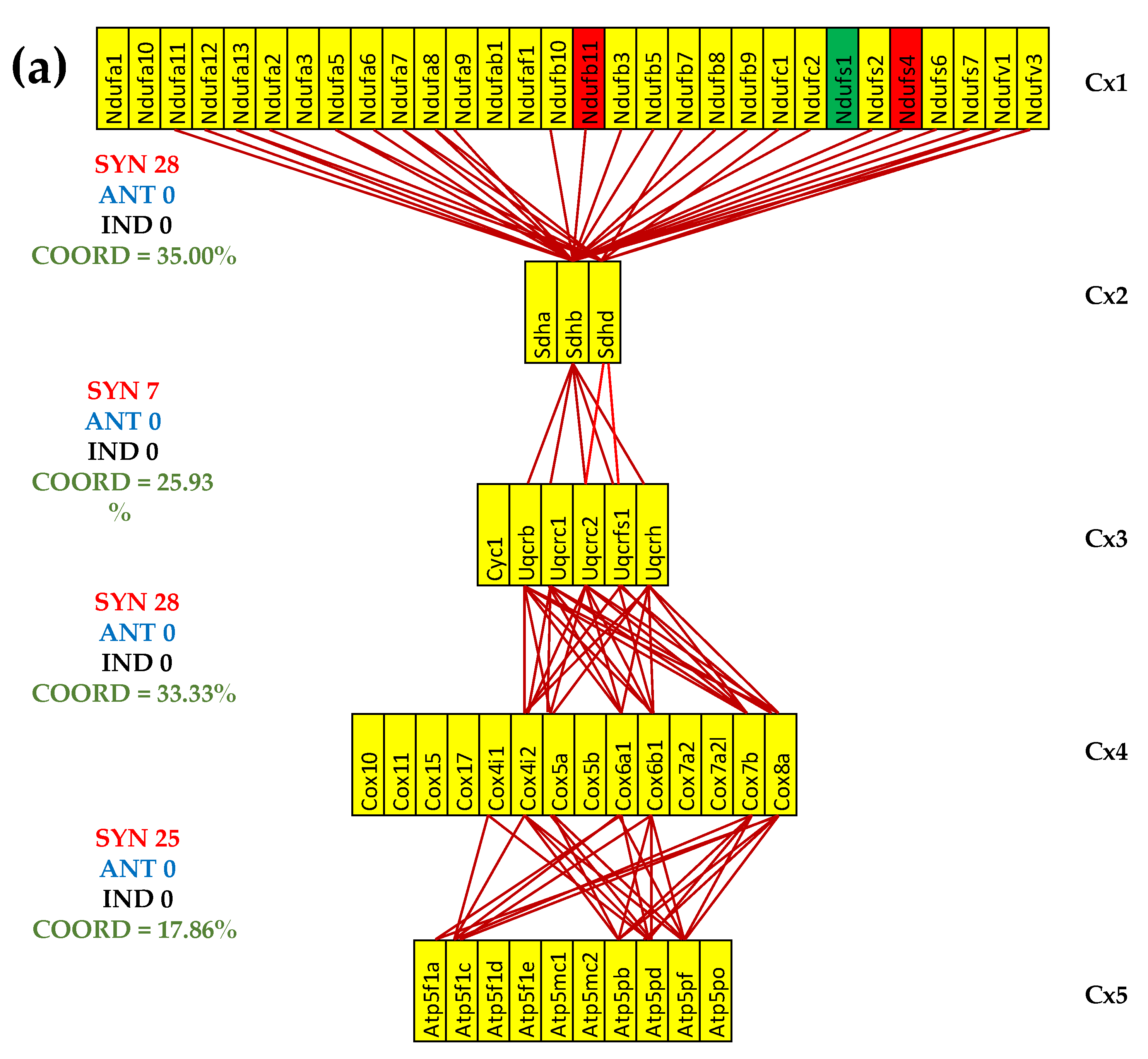
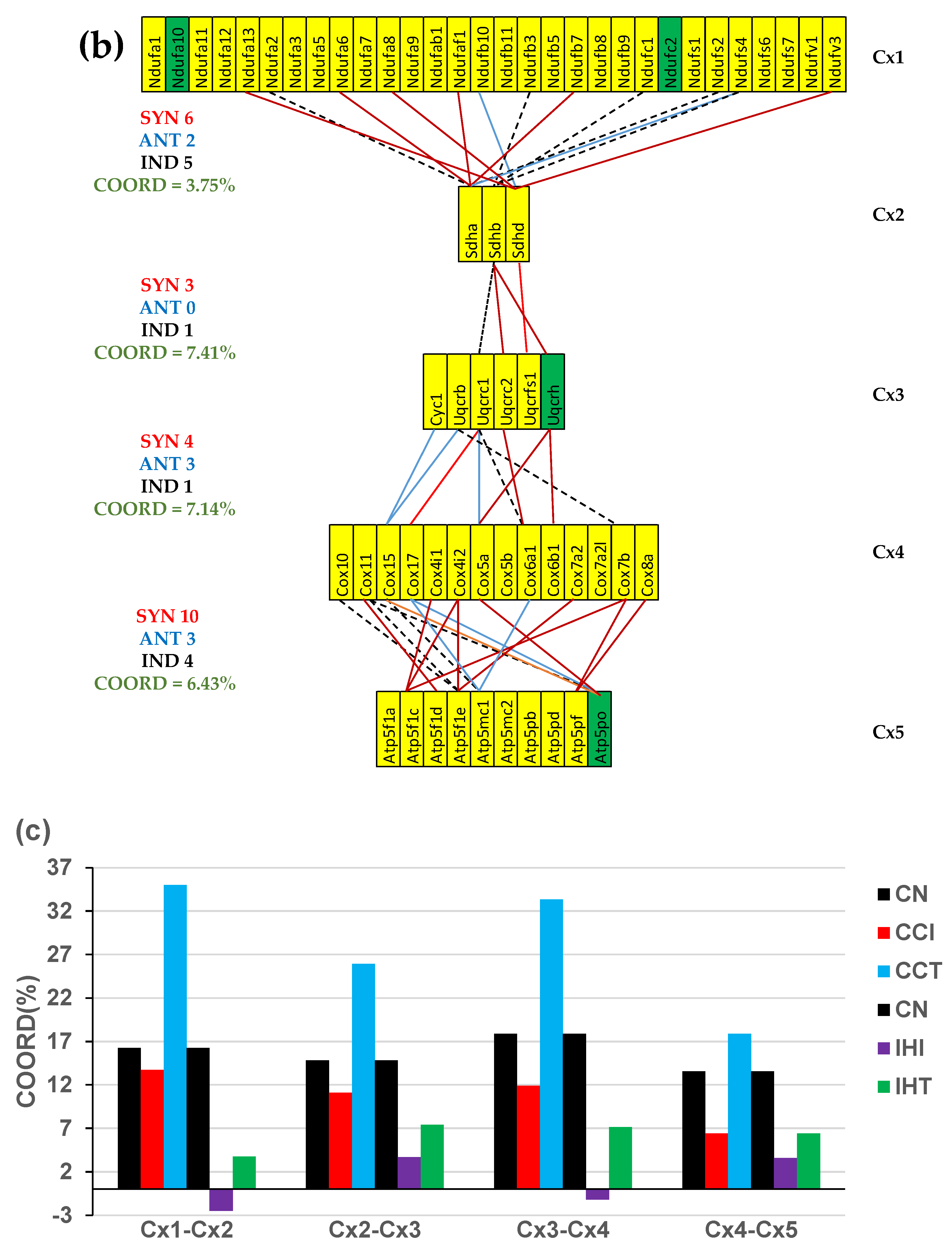

Table 1.
The largest mitochondrial (MITO) gene contributors to the heart transcriptome alteration in Chagasic (CCC) and ischemic (IHF) mice. AVE = average expression in the control mice, X-IN = expression ratio in untreated infected/infarcted with respect to control (negative for down-regulation), X-IT = expression ratio in treated infected/infarcted with respect to control (negative for down-regulation), |WIR| = Absolute value of the Weighted Individual (gene) Regulation.
Table 1.
The largest mitochondrial (MITO) gene contributors to the heart transcriptome alteration in Chagasic (CCC) and ischemic (IHF) mice. AVE = average expression in the control mice, X-IN = expression ratio in untreated infected/infarcted with respect to control (negative for down-regulation), X-IT = expression ratio in treated infected/infarcted with respect to control (negative for down-regulation), |WIR| = Absolute value of the Weighted Individual (gene) Regulation.
| THE LARGEST CONTRIBUTORS TO THE MITOCHONDRIAL TRANSCRIPTOME ALTERATION IN CCC MICE | ||||||||
|---|---|---|---|---|---|---|---|---|
| Gene | Description | AVE | X-IN | P | |WIR| | X-IT | P | |WIR| |
| Cox4i1 | Cytochrome c oxidase subunit IV isoform 1 | 165 | -1.69 | 0.02 | 111 | -2.05 | 0.45 | 95 |
| Sdha | Succinate dehydrogenase complex, subunit A, flavoprotein | 61 | -2.49 | 0.00 | 91 | -1.71 | 0.16 | 36 |
| Atp5pb | ATP synthase peripheral stalk-membrane subunit b | 62 | -2.35 | 0.00 | 83 | -1.47 | 0.27 | 21 |
| Ndufa4 | Mlrq-like protein | 105 | -1.78 | 0.09 | 76 | -1.46 | 0.27 | 35 |
| Ndufs1 | NADH dehydrogenase (ubiquinone) Fe-S protein 1 | 34 | -3.02 | 0.01 | 68 | -2.20 | 0.05 | 39 |
| Average mitochondrial |WIR| | 54 | 49 | ||||||
| The largest overall contributors to the entire transcriptome alteration in CCC mice | ||||||||
| Pln | Phospholamban | 148 | -2.64 | 0.09 | 222 | -2.65 | 0.31 | 167 |
| Overall average |WIR| | 2.24 | 1.15 | ||||||
| THE LARGEST CONTRIBUTORS TO THE MITOCHONDRIAL TRANSCRIPTOME ALTERATION IN IHF MICE | ||||||||
| Gene | Description | AVE | X-IN | P | |WIR| | X-IT | P | |WIR| |
| Cox5a | Cytochrome c oxidase, subunit Va | 49 | -59.69 | 0.09 | 2603 | -2.29 | 0.28 | 45 |
| Uqcrh | Ubiquinol-cytochrome c reductase hinge protein | 59 | -2.64 | 0.02 | 94 | -1.94 | 0.03 | 54 |
| Cox7a2 | Cytochrome c oxidase, subunit VIIa 2 | 49 | -2.67 | 0.00 | 81 | -1.20 | 0.16 | 8 |
| Cox6b1 | Cytochrome c oxidase, subunit VIb polypeptide 1 | 49 | -2.66 | 0.00 | 80 | 1.09 | 0.48 | 2 |
| Cox7b | Cytochrome c oxidase subunit VIIb | 64 | -2.28 | 0.03 | 80 | -1.07 | 0.69 | 1 |
| Average mitochondrial |WIR| | 69 | 12 | ||||||
| The largest overall contributors to the entire transcriptome alteration in IHF mice | ||||||||
| Cox5a | Cytochrome c oxidase, subunit Va | 49 | -59.69 | 0.09 | 2603 | -2.29 | 0.28 | 45 |
| Overall average |WIR| | 1.63 | 0.70 | ||||||
Table 2.
The most and the least controlled mitochondrial genes in Chagasic and ischemic mice. Abbreviations: CN = Control (reference); IN = Infected/ischemic not treated; IT = Infected/ischemic treated; MITO = Mitochondrial; RCS-FC, Fold-change of the relative control strength in IN/IT with respect to CN. For reference, the table includes the RECs and the RCS-FCs of the most and least controlled genes in the entire profiled transcriptome in control and untreated and treated CCC mice. The gray background indicates the most representative genes in each condition.
Table 2.
The most and the least controlled mitochondrial genes in Chagasic and ischemic mice. Abbreviations: CN = Control (reference); IN = Infected/ischemic not treated; IT = Infected/ischemic treated; MITO = Mitochondrial; RCS-FC, Fold-change of the relative control strength in IN/IT with respect to CN. For reference, the table includes the RECs and the RCS-FCs of the most and least controlled genes in the entire profiled transcriptome in control and untreated and treated CCC mice. The gray background indicates the most representative genes in each condition.
| THE MOST AND THE LEAST CONTROLLED MITO GENES IN CCC MICE | RCS-FC | ||||||
|---|---|---|---|---|---|---|---|
| Gene | Description | CN | IN | IT | IN | IT | |
| Ndufa10 | NADH:ubiquinone oxidoreductase subunit A10 | 3.86 | -1.17 | -0.64 | -32.80 | -22.70 | |
| Cox7b | Cytochrome c oxidase subunit VIIb | 2.59 | -0.42 | -1.59 | -8.05 | -18.12 | |
| Ndufb10 | NADH:ubiquinone oxidoreductase subunit B10 | 2.15 | -0.95 | -0.68 | -8.56 | -7.13 | |
| Vdac2 | Voltage-dependent anion channel 2 | 1.06 | 2.59 | -0.63 | 2.89 | -3.23 | |
| Mpc2 | mitochondrial pyruvate carrier 2 | 0.32 | 1.54 | 0.68 | 2.34 | 1.28 | |
| Cox4nb | COX4 neighbor | 0.81 | 1.28 | 0.98 | 1.38 | 1.12 | |
| Ndufb5 | NADH:ubiquinone oxidoreductase subunit B5 | 0.43 | -0.53 | 1.66 | -1.95 | 2.33 | |
| Ndufb11 | NADH:ubiquinone oxidoreductase subunit B11 | 0.99 | 0.54 | 1.62 | -1.36 | 1.55 | |
| Ndufaf4 | NADH:ubiquinone oxidoreductase subunit A4 | 0.29 | -0.92 | 1.37 | -2.32 | 2.11 | |
| Sdhd | Succinate dehydrogenase complex, subunit D, integral membrane protein | -1.87 | -0.32 | -0.30 | 2.92 | 2.96 | |
| Cox5a | Cytochrome c oxidase, subunit Va | -1.58 | 1.17 | -2.53 | 6.69 | -1.94 | |
| Ndufa9 | NADH:ubiquinone oxidoreductase subunit A9 | -1.26 | -2.64 | -0.32 | -2.60 | 1.93 | |
| Cpt2 | Carnitine palmitoyltransferase 2 | 0.53 | -1.67 | 0.21 | -4.60 | -1.25 | |
| Pdk2 | Pyruvate dehydrogenase kinase, isoenzyme 2 | 0.60 | -1.43 | -0.67 | -4.09 | -2.41 | |
| Ppif | Peptidylprolyl isomerase F (cyclophilin F) | 0.57 | -1.41 | -0.56 | -3.94 | -2.18 | |
| Cox7b | Cytochrome c oxidase subunit VIIb | 2.59 | -0.42 | -1.59 | -8.05 | -18.12 | |
| Atp5pf | ATP synthase peripheral stalk subunit F6 | 1.52 | -0.81 | -1.58 | -5.01 | -8.53 | |
| Cox4i1 | Cytochrome c oxidase subunit IV isoform 1 | 0.66 | -0.20 | -1.56 | -1.81 | -4.65 | |
| THE MOST AND THE LEAST CONTROLLED MITO GENES IN IHF MICE | RCS-FC | ||||||
| Cox6b1 | Cytochrome c oxidase, subunit VIb polypeptide 1 | 1.36 | 4.38 | 0.05 | 8.13 | -2.48 | |
| Atp5f1e | ATP synthase F1 subunit epsilon | -0.79 | 2.68 | 0.68 | 11.07 | 2.77 | |
| Atp5mc2 | ATP synthase membrane subunit c locus 2 | 0.52 | 1.75 | -0.37 | 2.35 | -1.86 | |
| Ndufb11 | NADH dehydrogenase (ubiquinone) 1 beta subcomplex, 11 | 0.74 | 0.87 | 2.38 | 1.10 | 3.11 | |
| Ndufv1 | NADH dehydrogenase (ubiquinone) flavoprotein 1 | -1.14 | -0.97 | 1.79 | 1.12 | 7.58 | |
| Cpt1b | Carnitine palmitoyltransferase 1b, muscle | 0.16 | -1.42 | 1.64 | -2.99 | 2.79 | |
| Ndufa9 | NADH:ubiquinone oxidoreductase subunit A9 | -1.26 | -2.64 | -0.32 | -2.60 | 1.93 | |
| Cyc1 | Cytochrome c-1 | -1.05 | -1.67 | 0.93 | -1.54 | 3.95 | |
| Ndufa11 | NADH:ubiquinone oxidoreductase subunit A11 | -0.36 | -1.58 | -0.29 | -2.34 | 1.04 | |
| Cox5a | Cytochrome c oxidase, subunit Va | -1.58 | 1.17 | -2.53 | 6.69 | -1.94 | |
| Mpc1 | mitochondrial pyruvate carrier 1 | -1.05 | -0.92 | -1.71 | 1.10 | -1.58 | |
| Ndufc2 | NADH:ubiquinone oxidoreductase subunit C2 | 0.38 | -0.30 | -1.52 | -1.61 | -3.75 | |
| The most controlled genes in the entire transcriptome in CN and IHF mice | RCS-FC | ||||||
| Tmem186 | Transmembrane protein 186 | 5.19 | -0.10 | 0.70 | -39.13 | -22.50 | |
| Cd164 | CD164 antigen | 0.09 | 5.28 | 0.57 | 36.56 | 1.40 | |
| Atp13a2 | ATPase type 13A2 | 0.66 | -0.03 | 4.55 | -1.61 | 14.82 | |
| The least controlled genes in the entire transcriptome in CN and IHF mice | RCS-FC | ||||||
| Gmcl1 | Germ cell-less homolog 1 (Drosophila) | -2.40 | -0.33 | -2.50 | 4.19 | -1.07 | |
| Idh3g | Isocitrate dehydrogenase 3 (NAD+), gamma | -1.10 | -2.83 | 0.80 | -3.32 | 3.72 | |
| Tsc22d4 | TSC22 domain family, member 4 | 0.98 | 0.25 | -2.91 | -1.67 | -14.86 | |
Table 3.
The Gene Commanding Heights (GCH) of the five most prominent mitochondrial genes in control, treated and untreated Chagasic and ischemic mice. Abbreviations: IN = Infected/ischemic not treated; IT = Infected/ischemic treated; MITO = Mitochondrial; GCH-FC = Fold-change of the Gene Commanding Height, CCC = Chagasic mice, IHF = ischemic mice. For reference, the table includes the GCHs of the most prominent MITO genes in control mice and their scores in untreated CCC and IHF mice. .
Table 3.
The Gene Commanding Heights (GCH) of the five most prominent mitochondrial genes in control, treated and untreated Chagasic and ischemic mice. Abbreviations: IN = Infected/ischemic not treated; IT = Infected/ischemic treated; MITO = Mitochondrial; GCH-FC = Fold-change of the Gene Commanding Height, CCC = Chagasic mice, IHF = ischemic mice. For reference, the table includes the GCHs of the most prominent MITO genes in control mice and their scores in untreated CCC and IHF mice. .
| MOST PROMINENT MITO GENES IN CCC MICE | GCH | GCH-FC | ||||||
|---|---|---|---|---|---|---|---|---|
| GENE | Description | IN | IT | IN | IT | |||
| Cox4i2 | cytochrome c oxidase subunit 4I2 | 13.20 | 17.02 | 8.74 | 11.27 | |||
| Ndufb7 | NADH dehydrogenase (ubiquinone) 1 beta subcomplex, 7 | 11.73 | 17.82 | 14.09 | 21.42 | |||
| Cox6b1 | Cytochrome c oxidase, subunit VIb polypeptide 1 | 11.48 | 17.52 | 4.81 | 7.34 | |||
| Uqcrh | Ubiquinol-cytochrome c reductase hinge protein | 11.21 | 16.07 | 4.66 | 6.67 | |||
| Ndufs4 | NADH dehydrogenase (ubiquinone) Fe-S protein 4 | 11.00 | 12.23 | 4.66 | 5.18 | |||
| Ndufa7 | NADH dehydrogenase (ubiquinone) 1 alpha subcomplex, 7 | 7.48 | 19.80 | 1.42 | 3.77 | |||
| Ndufc1 | NADH:ubiquinone oxidoreductase subunit C1 | 4.60 | 19.71 | -1.18 | 3.63 | |||
| Ndufa2 | NADH dehydrogenase (ubiquinone) 1 alpha subcomplex, 2 | 6.54 | 19.70 | 7.64 | 23.01 | |||
| Ndufa10 | NADH dehydrogenase (ubiquinone) 1 alpha subcomplex 10 | 9.54 | 19.57 | 1.71 | 3.52 | |||
| Uqcrb | Ubiquinol-cytochrome c reductase binding protein | 9.95 | 19.55 | 2.44 | 4.80 | |||
| MOST PROMINENT MITO GENES IN IHF MICE | GCH | GCH-FC | ||||||
| Cox6b1 | Cytochrome c oxidase, subunit VIb polypeptide 1 | 24.16 | 2.08 | 5.53 | -2.10 | |||
| Atp5f1e | ATP synthase F1 subunit epsilon | 9.01 | 2.00 | 3.70 | -1.22 | |||
| Atp5mc2 | ATP synthase membrane subunit c locus 2 | 5.13 | 1.12 | 1.88 | -2.44 | |||
| Ndufs5 | NADH dehydrogenase (ubiquinone) Fe-S protein 5 | 4.22 | 1.40 | 1.13 | -2.65 | |||
| Ndufa1 | NADH dehydrogenase (ubiquinone) 1 alpha subcomplex, 1 | 4.21 | 3.60 | 1.38 | 1.18 | |||
| Ndufb11 | NADH dehydrogenase (ubiquinone) 1 beta subcomplex, 11 | 3.01 | 6.39 | -1.90 | 1.12 | |||
| Ndufs4 | NADH dehydrogenase (ubiquinone) Fe-S protein 4 | 0.80 | 5.92 | -9.72 | -1.31 | |||
| Ndufv1 | NADH dehydrogenase (ubiquinone) flavoprotein 1 | 0.72 | 4.48 | -3.68 | 1.68 | |||
| Cyc1 | Cytochrome c-1 | 0.52 | 4.26 | -4.69 | 1.73 | |||
| Ndufa1 | NADH dehydrogenase (ubiquinone) 1 alpha subcomplex, 1 | 4.21 | 3.60 | 1.38 | 1.18 | |||
| MOST PROMINENT MITO GENES IN HEALTHY MICE | GCH | CCC | IHF | |||||
| Ndufb10 | NADH dehydrogenase (ubiquinone) 1 beta subcomplex, 10 | 13.07 | 7.21 | 2.26 | ||||
| Ndufa10 | NADH dehydrogenase (ubiquinone) 1 alpha subcomplex 10 | 9.79 | 9.54 | 1.01 | ||||
| Uqcrb | Ubiquinol-cytochrome c reductase binding protein | 9.69 | 9.95 | 2.91 | ||||
| Ndufaf1 | NADH:ubiquinone oxidoreductase complex assembly factor 1 | 8.14 | 8.31 | 1.67 | ||||
| Ndufs4 | NADH dehydrogenase (ubiquinone) Fe-S protein 4 | 7.76 | 11.00 | 0.80 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



