
Submitted:
25 September 2025
Posted:
26 September 2025
You are already at the latest version
Abstract
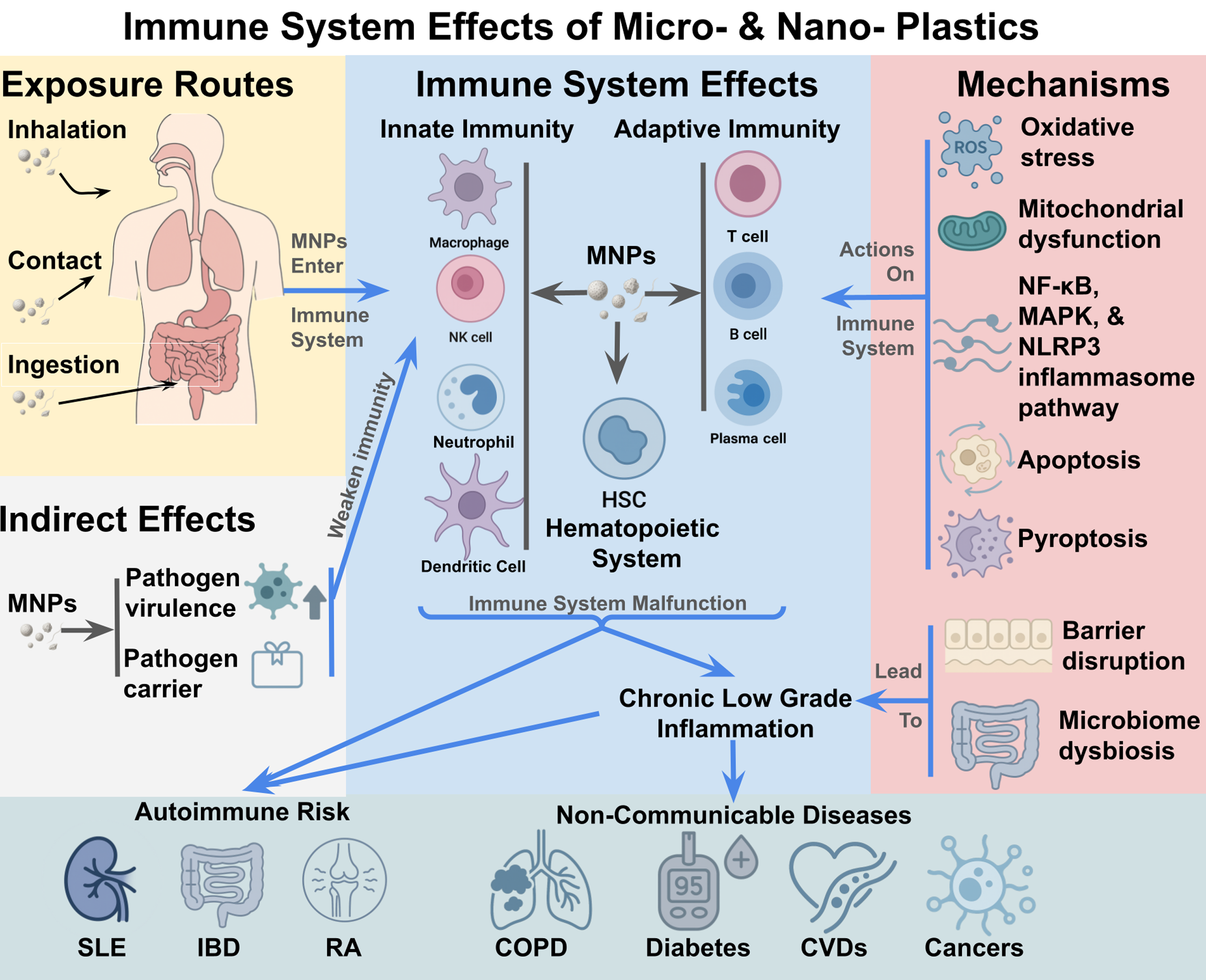
Micro- and nanoplastics (MNPs) are emerging environmental immunotoxins with widespread human exposure through ingestion, inhalation, and dermal contact. Detected in the placenta, lungs, blood, bone marrow, and brain, MNPs accumulate in immune organs where they disrupt innate and adaptive cell functions. Experimental evidence shows that MNPs impair macrophage phagocytosis, skew dendritic cell maturation, trigger neutrophil extracellular traps, and alter T and B cell responses. Mechanistically, these effects are driven by oxidative stress, mitochondrial dysfunction, and activation of NF-κB, MAPK, and NLRP3 inflammasome pathways, leading to apoptosis, pyroptosis, and chronic low-grade inflammation. Barrier disruption and microbiome dysbiosis further compromise immune tolerance, linking MNPs exposure to autoimmune risk. Preclinical models demonstrate exacerbation of systemic lupus erythematosus, inflammatory bowel disease, and rheumatoid arthritis, though human causal data remain limited. Standardized protocols, advanced imaging and omics, and longitudinal cohorts are urgently needed to establish causal links and inform public health strategies.
Keywords:
microplastics
; nanoplastics
; immune system
; inflammation
; signaling pathways
; oxidative stress
; NF-κB
; NLRP3
; cellular toxicity
; immunotoxicology
1. Introduction
Plastics, synthetic polymers valued for their durability, low production cost, and versatility, have become ubiquitous in modern life. Since the 1950s, cumulative global plastic production has surpassed 9 billion metric tons, with annual output exceeding 460 million tons by 2024 [1,2,3]. Despite extensive use, only about 9% of plastics are recycled [4], while most accumulate in landfills, terrestrial environments, and aquatic ecosystems [5,6]. Environmental stressors such as ultraviolet (UV) radiation, mechanical abrasion, thermal degradation, and biological activity drive the fragmentation of larger plastic debris into microplastics and nanoplastics (MNPs). These particles exhibit wide variation in size, shape, and polymer chemistry, properties that underlie their diverse environmental persistence and biological effects.
Growing evidence indicates that the health risks posed by MNPs are considerable. Beyond their intrinsic properties, these particles can adsorb and transport toxic chemicals, including bisphenol A (BPA), phthalates, and polyfluorinated compounds, which may amplify their biological impact. MNPs have been detected in several human tissues, including placenta, lungs, blood, and, most recently, the brain, pointing to systemic exposure through inhalation, ingestion, and dermal penetration [7]. Although the direct toxicological effects in humans remain incompletely understood, experimental models consistently demonstrate adverse impacts across multiple organ systems, notably the reproductive, respiratory, and nervous systems. These findings frame MNPs exposure as an emerging global health concern.
The immune system represents a first line of defense against foreign materials, with crucial roles in host protection, tissue repair, and surveillance against cellular damage. Barrier tissues such as the skin, respiratory tract, and gastrointestinal epithelium are the primary sites where MNPs are likely to interact with immune cells. As non-self particles, MNPs and their associated contaminants are subject to immune recognition and processing. Toxicological effects arising from these interactions are therefore expected to manifest initially within immune compartments, with broader implications for systemic immune competence [8]. Immune impairment caused by persistent MNPs exposure could contribute to a wide spectrum of disorders, making the study of MNPs–immune system interactions a priority.
This review summarizes current knowledge on the biodistribution of MNPs, their effects on immune cell function, roles in immune regulation, and underlying molecular mechanisms. These insights provide a foundation for developing preventive and therapeutic strategies aimed at mitigating the immunotoxic consequences of MNPs exposure.
2. Characteristics and Immunological Relevance of MNPs
2.1. Definition of Micro- and Nanoplastics
The concept of microplastics (MPs) was introduced by Thompson et al. (2004), who described the widespread occurrence of plastic fragments smaller than 5 mm in marine environments, marking a turning point in plastic pollution research [7,9]. The term nanoplastics (NPs) first appeared in the literature in 1995 [10]; however, at that time it referred to engineered, “intelligent” plastic materials [11] rather than nanoscale environmental contaminants [12]. The contemporary definition—plastic particles <1000 nm with potential ecological and health impacts—was established in the following decade. One of the first uses of this definition was in 2010, when Bhattacharya et al. demonstrated that nanoscale plastic particles could interact with ecological systems and bind environmental contaminants [13]. This work helped establish NPs as a distinct category of pollutants and set the stage for toxicological and ecological studies.
2.2. Routes and Mechanisms of MNPs Entry into the Human Body
2.2.1. Ingestion
Ingestion is now recognized as a major pathway for human exposure to MNPs. Food and water are frequently contaminated, and packaging materials provide additional sources [14]. Once ingested, MNPs can enter the bloodstream and distribute to multiple organs [15]. Experimental studies indicate that ingestion disrupts the gut microbiota, induces intestinal inflammation, and contributes to systemic immune effects [8,16,17].
Absorption across the gastrointestinal barrier occurs primarily through transcytosis in enterocytes and microfold (M) cells, while larger particles may pass via paracellular pathways [18,19]. Polystyrene (PS) NPs can traverse tight epithelial junctions, entering systemic circulation [20]. Nanoparticles are typically internalized through transcytosis, whereas microparticles may cross via damaged or loosened epithelial junctions [21]. In addition, MNPs can erode epithelial surfaces and promote oxidative stress–mediated apoptosis, thereby increasing gut permeability [22]. Particle size, epithelial integrity, and uptake mechanisms strongly influence gastrointestinal translocation.
2.2.2. Inhalation
Inhalation has emerged as another significant route of exposure. Indoor environments are major contributors, with estimates suggesting that adults inhale up to 68,000 PM10-sized plastic particles daily [23]. Particles in the 1–10 µm range are of particular concern, as they can reach the alveoli, cross epithelial barriers, and activate oxidative and inflammatory pathways [24,25].
Particle deposition in the respiratory tract is governed by diffusion, gravitational settling, and inertial impaction, enabling even nanosized plastics to reach alveolar sacs [25]. Once deposited, NPs can interact with pulmonary surfactant, sometimes impairing its function [26,27]. In vitro studies show that airborne NPs suppress immunoregulatory cytokines (e.g., IL-2, TGF-β), promote neutrophil recruitment, and trigger airway inflammation [28]. Chronic exposure may compromise epithelial integrity, facilitating deeper penetration and systemic immune effects [29]. Together, these findings establish inhalation—particularly in enclosed, plastic-rich settings—as a biologically significant pathway of MNPs exposure.
Figure 1.
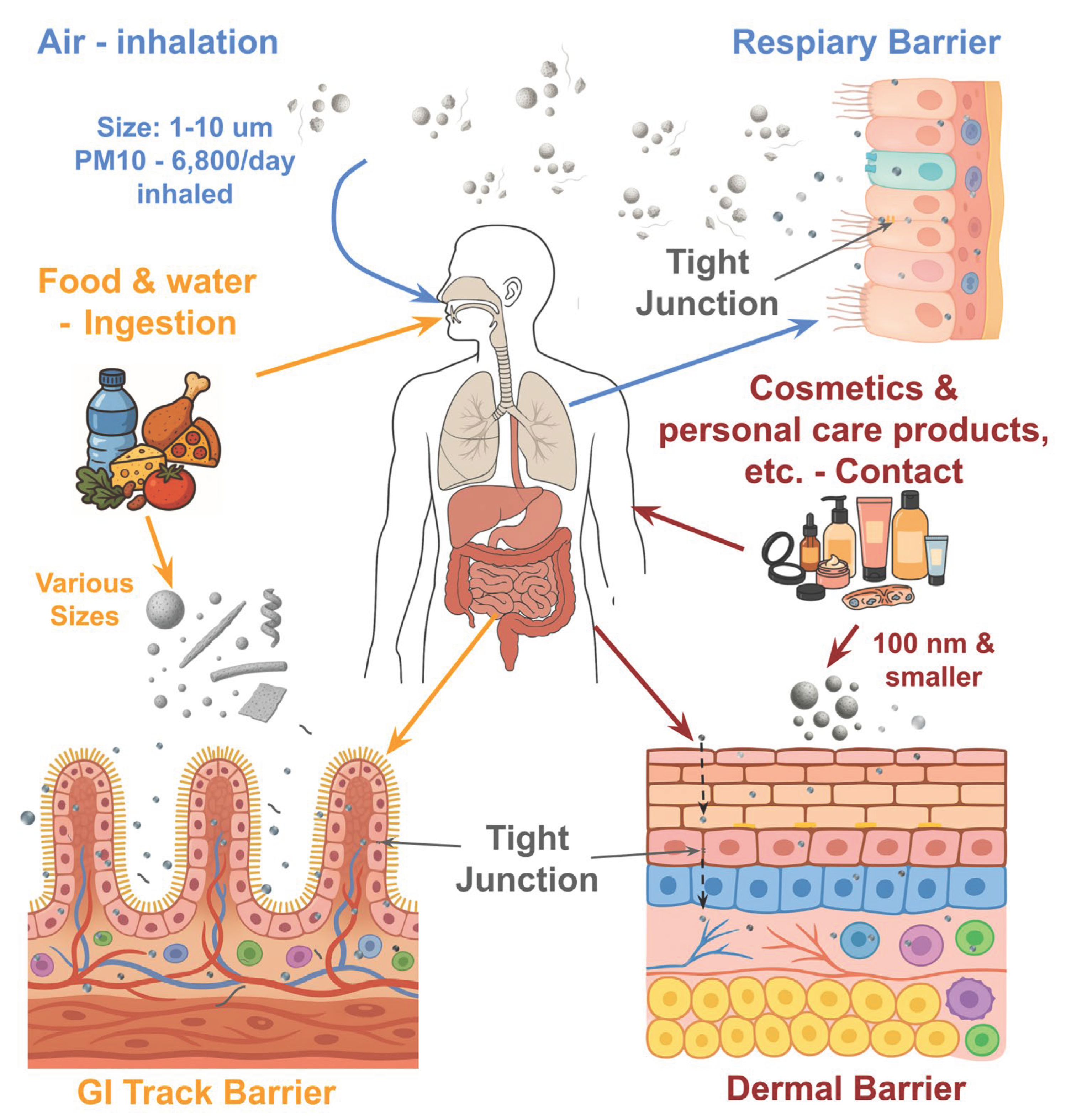
Routes of human exposure to MNPs and epithelial barrier interactions. MNPs enter the human body primarily through inhalation, ingestion, and dermal contact. Inhalation: Airborne MNPs, particularly in the 1–10 µm range, including PM10 particles (~6,800 particles inhaled daily), can deposit along the respiratory tract and cross the respiratory epithelial barrier via disrupted tight junctions; Ingestion: Contaminated food, drinking water, and packaging introduce MNPs of various sizes into the gastrointestinal tract, where they may translocate across the intestinal epithelial barrier through paracellular transport or uptake by enterocytes and M cells; Dermal contact: Cosmetics, personal care products, and medical materials represent additional sources. Nanoplastics smaller than 100 nm can penetrate the skin, particularly through hair follicles or compromised barrier regions, interacting with underlying immune cells. Across these exposure routes, epithelial barrier disruption and tight junction impairment facilitate systemic translocation of MNPs, potentially leading to immune activation and downstream health effects.
Figure 1.
Routes of human exposure to MNPs and epithelial barrier interactions. MNPs enter the human body primarily through inhalation, ingestion, and dermal contact. Inhalation: Airborne MNPs, particularly in the 1–10 µm range, including PM10 particles (~6,800 particles inhaled daily), can deposit along the respiratory tract and cross the respiratory epithelial barrier via disrupted tight junctions; Ingestion: Contaminated food, drinking water, and packaging introduce MNPs of various sizes into the gastrointestinal tract, where they may translocate across the intestinal epithelial barrier through paracellular transport or uptake by enterocytes and M cells; Dermal contact: Cosmetics, personal care products, and medical materials represent additional sources. Nanoplastics smaller than 100 nm can penetrate the skin, particularly through hair follicles or compromised barrier regions, interacting with underlying immune cells. Across these exposure routes, epithelial barrier disruption and tight junction impairment facilitate systemic translocation of MNPs, potentially leading to immune activation and downstream health effects.
2.2.3. Dermal Contact
Although less studied, dermal contact is a plausible route of MNPs exposure, especially through personal care products and medical materials. NPs in cosmetics, topical formulations, and prosthetic devices have been shown to provoke oxidative stress and inflammation in skin cells [15,30,31].
While the skin provides an effective barrier, particles <100 nm can penetrate under certain conditions [32,33]. Ex vivo models show that 100 nm fluorescent PS nanoparticles can infiltrate the epidermis and accumulate in keratinocytes and fibroblasts, especially when the stratum corneum is disrupted [34]. These effects have been linked to IL-17 pathway activation and fibroblast transformation. Particles as small as 40 nm may even reach Langerhans cells via hair follicles [35]. Penetration is therefore strongly size-dependent and enhanced when skin integrity is compromised, suggesting that dermal exposure may be relevant for NPs in consumer and medical products.
Together, ingestion, inhalation, and dermal contact demonstrate that MNPs have multiple entry points into the body. Once inside, these particles can spread to immune-related organs and tissues, shaping their potential immunological consequences.
2.3. Biodistribution and Immune System Impacts
Experimental studies demonstrate that MNPs distribute widely throughout the body, with particle size being a key determinant. Nanoparticles ~10 nm have been detected in blood, liver, spleen, thymus, lymph nodes, lungs, heart, and brain, whereas particles 100–250 nm remain more localized to blood, liver, and spleen [36]. Microparticles <5 µm can accumulate in macrophages, lymph nodes, spleen, and circulation, while those <10 µm may translocate from the gut into systemic sites, including liver, kidneys, and brain [37]. At the cellular level, intravenously administered 500 nm particles were rapidly taken up by granulocytes and bone marrow cells, accumulating in macrophage- and granulocyte-rich regions of the spleen, including the marginal zone and red pulp [38].
In humans, MPs have recently been detected in bone marrow, with average concentrations of ~51 µg/g and particle sizes <100 µm [39]. This suggests bone marrow may act as a reservoir for plastic accumulation, raising concerns about disruption of hematopoiesis and possible links to hematologic malignancies [40]. Animal studies further show that the thymus can accumulate PS particles, potentially affecting T-cell development [41,42]. Avian models confirm thymic structural disruption, cytokine upregulation, and apoptosis-related gene activation following exposure [43].
These findings demonstrate that MNPs distribute across central and peripheral immune compartments—including bone marrow, spleen, thymus, and circulation—posing risks for immune dysfunction and long-term disruption of hematopoietic and immunological balance.
2.4. Indirect Effects of MNPs on the Immune System
2.4.1. Pathogen Carriage and Increased Susceptibility
MNPs can act as carriers for microorganisms, altering microbial properties and intensifying immune responses [44,45]. They also provide surfaces for pathogen colonization, thereby amplifying infectious risks [46]. Laboratory findings show that positively charged NPs enhance Escherichia coli O157:H7 biofilm formation and toxin production, making the bacteria more virulent and resistant to host defenses [47,48].
MNPs may also interact with viruses. In bacteriophage models, up to 98% of viral particles adsorbed onto PS MPs, with efficiency influenced by particle size and surface chemistry [49]. Environmental studies suggest that MPs promote viral persistence and infectivity through charge effects, biofilms, and particle aging [50]. In amphibian models, exposure to MPs reduced antiviral resistance during ranavirus infection [51]. These observations support a dual role of MNPs as both vectors and amplifiers of pathogens, thereby elevating infection risk and complicating immune defense.
2.4.2. Gut Microbiota Disruption and Cytotoxicity
MNPs exposure can profoundly alter intestinal microbial composition and barrier integrity. In murine models, PS MPs disrupted microbiota balance, impaired gut permeability, and induced systemic inflammation [52]. Polyethylene MPs shifted bacterial communities, reduced bile acid metabolism, and promoted inflammation [53]. Reviews confirm that MNPs exposure destabilizes microbial homeostasis and facilitates immune activation [42].
Beyond microbiota disruption, MNPs exert direct cytotoxicity on intestinal epithelial and immune cells. PS MPs induce pro-inflammatory signaling, apoptosis, and cytotoxicity in intestinal cell lines [54]. Mechanistic studies link these effects to oxidative stress, mitochondrial dysfunction, and autophagy, often exacerbated by inhibition of cytoprotective enzymes such as heme oxygenase-1 [35]. Collectively, these findings suggest that MNPs compromise gut health both by reshaping host–microbe interactions and by directly damaging epithelial and immune cells, leading to chronic immune dysregulation.
3. Direct Effects of MNPs on the Immune System
3.1. Effects on Innate Immune Cells
3.1.1. Effects of MNPs on Macrophages
Macrophages are key sentinels that recognize and clear foreign particulates, including MNPs. Exposure to MNPs induces oxidative stress, mitochondrial dysfunction, and pro-inflammatory signaling. Polyethylene MPs, for example, activate NF-κB signaling (IKKα/β, p65), trigger Nod-like receptor protein 3 (NLRP3) inflammasome assembly, and elevate TNF-α, IL-1β, IL-6, and IL-8 while suppressing IL-4 and IL-10 [55]. The NLRP3 inflammasome appears to be a central sensor of plastic particles and a candidate biomarker for immunotoxicity [56].
Beyond signaling, MNPs impose metabolic and structural stress, promoting apoptosis and weakening cellular defenses [55,57]. PS MPs can also induce macrophage extracellular traps (METs) in the liver, responses that propagate inflammation and exacerbate tissue injury [58]. Together, these data indicate that MNPs both activate damaging inflammatory cascades and erode macrophage metabolic integrity, favoring chronic inflammation and impaired resolution.
3.1.1.1. Effects on Macrophages Phagocytosis
Macrophages internalize MNPs via phagocytosis and endocytosis. Murine macrophages efficiently take up 50- and 500-nm PS particles in time- and dose-dependent fashion, with uptake driving M1-biased polarization (↑iNOS, TNF-α; ↓IL-10, Arg-1) [59]. Uptake is size dependent, very small particles may poorly engage receptors, whereas large particles resist internalization [59,60]. Internalized plastics perturb redox balance, mitochondria, lysosomes, and surface receptors (e.g., CD86, PD-L1, CD204) in size- and dose-dependent patterns [61]. Environmentally weathered PVC, PP, and PA6.6 fragments are similarly engulfed by THP-1 derived macrophages and induce lysosomal stress, mitochondrial dysfunction, and reduced viability even without strong NF-κB mediated cytokine release [62]. Thus, plastic uptake reprograms phenotype and metabolism, disrupting macrophage homeostasis.
3.1.1.2. Effects on Macrophage Polarization
Polarization shifts shape macrophage function. In vitro, PS NPs (50–500 nm; ~50 µg/mL) bias RAW264.7 cells toward M1 (↑CD86, iNOS, TNF-α; ↓CD206, IL-10, Arg-1), via receptor-dependent endocytosis and downstream inflammatory cascades [59]. Findings vary by model: some THP-1 studies show minimal polarization [60], whereas murine pregnancy models report M2 skewing with immunosuppressive cytokines and developmental toxicity [60]. In vivo, chronic PS NPs exposure in ApoE⁻/⁻ mice augments aortic M1 phagocytosis, upregulates MARCO, perturbs lipid handling, and yields foamy macrophages within lesions [60]; testicular exposure promotes M1 polarization through reactive oxygen species (ROS)-driven MIF/NF-κB activation with metabolic disruption [63]. Overall, MNPs frequently favor M1 programs through ROS- and NF-κB/MIF-linked mechanisms, but responses are context- and cell-type dependent.
3.1.1.3. Effects on Macrophage Cytokine Secretion
MNPs exposure commonly elevates IL-6, TNF-α, and IL-1β, driven by oxidative stress and MAPK/NF-κB signaling. RAW264.7 cells exposed to surface-functionalized PS NPs (PS-COOH, PS-NH₂) show increased ROS, activation of MAPK/NF-κB, and heightened IL-6/TNF-α; antioxidants blunt these effects [64]. Systematic review report consistent cytokine imbalance and inflammasome activation across models [65]. Notably, environmentally relevant fragments (PVC, PP, PA6.6, PS) internalized by THP-1 macrophages did not always trigger strong NF-κB or cytokines yet still caused cytotoxicity, lysosomal stress, and mitochondrial dysfunction [62]. Thus, cytokine outcomes vary with particle properties and weathering, but oxidative injury is a unifying feature.
3.1.2. Effects on the Dendritic Cells (DCs)
DCs internalize MNPs and undergo phenotypic and functional changes. Human monocyte-derived DCs efficiently take up virgin and weathered PS particles (0.2–10 µm), especially with plasma-protein coronas; weathered 0.2 µm particles increase CD83/CD86 and enhance T-cell proliferation, consistent with adjuvant-like activity [66]. Irregularly shaped NPs (50–310 nm; PVC/PS fragments) elicit stronger cytokine release than spheres, underscoring shape and composition effects [67].
Taken together, these findings indicate that MNPs can directly activate dendritic cells through uptake-mediated modulation of surface markers and cytokine-driven inflammatory signaling. Such effects may disrupt antigen presentation and contribute to dysregulated adaptive immune responses, highlighting dendritic cells as critical targets in the immunotoxicity of plastic particles.
3.1.2.1. Antigen Presentation
3.1.2.2. Maturation
MNPs trigger pattern recognition receptor (PRR) engagement (e.g., Toll like receptors, TLRs), activating NF-κB/MAPK and upregulating CD80/CD86/CD83 with IL-6, IL-12, and TNF-α secretion [66]. Chronic low-dose exposure may sustain semi-mature, pro-inflammatory DC states that misprime T cells and erode tolerance [68].
Because DCs orchestrate T-cell priming, these alterations provide a mechanistic link between plastic exposure and downstream adaptive immune imbalance.
3.1.3. Effects on Neutrophils
MNPs strongly modulate neutrophil activation and fate. Mouse neutrophils bind/internalize PS MPs, mount vigorous inflammatory responses, and undergo cell death [69]. PS NPs induce robust neutrophil extracellular traps (NETs) formation through ROS- and PAD4-dependent histone citrullination [70]; in vivo, PS NPs drive hepatic neutrophil infiltration and NETs via a ROS–NLRP3 axis, and genetic/pharmacologic NLRP3 blockade or DNase I reduces cytokines and tissue damage [71].
Dietary PS MPs amplify NETs in liver injury models through enhanced ROS [72]; PS NPs directly trigger NETosis via NADPH-oxidase-linked ROS and PAD4 activation, with neutrophil elastase contributing to chromatin decondensation [70]. TLR-like signaling during plastic phagocytosis further escalates NETosis and shortens neutrophil survival [69]. These pathways implicate neutrophils as key effectors of plastic-induced inflammatory injury
3.1.4. Effects on Natural Killer (NK) Cells
Direct data are limited, but oral PS MPs increase NK cells infiltration in the small intestine with villous changes and heightened inflammation in mice [73]. Given the cross-cellularity of MPs induced ROS and inflammatory signaling, similar oxidative constraints likely impair NK cell cytotoxicity and cytokine balance [74]. MPs also activate NF-κB/MAPK pathways central to NK cell receptor signaling (e.g., NKG2D, NKp46, DNAM-1); polypropylene NPs trigger lung inflammation via p38–NF-κB, suggesting plausible interference with NK cell function [75]. Persistent epithelial-barrier injury and cytokine dysregulation could raise NK cell activation thresholds, blunt surveillance, and favor tumor or viral escape.
3.2. Effects on Adaptive Immune Cells
3.2.1. Disruption of T Cell Compartments
MNPs foster oxidative and inflammatory milieus that impair T-cell activation, differentiation, and survival. Excess ROS activates NF-κB, p38 MAPK, and STAT3, skewing T-helper polarization and undermining Treg development, with potential exhaustion phenotypes and reduced surveillance [76]. Antigen-presenting cell (APC) dysfunction after plastic exposure further weakens priming [77]. Although direct TCR-signaling studies remain sparse, the cumulative evidence indicates meaningful downstream T-cell disruption.
3.2.1.1. Regulatory T (Treg) Cells
Polyethylene MPs reduce Treg frequencies and expand Th17 populations with increased IL-1α in mice, consistent with impaired tolerance and inflammatory skewing [8,78]. Mechanistically, activation of TLR4, AP-1, and IRF5 is implicated [8,78], while MAPK, NF-κB, and AKT signaling elevate IL-1β/IL-6, destabilizing Treg homeostasis and shifting Th2/Th17 ratios [77]. These changes favor chronic inflammation and may lower barriers to tumorigenesis.
3.2.1.2. T Helper (Th) Cells
Early-life exposures can program long-term Th imbalance. Lactational MPs alter immune status in male offspring, consistent with durable effects on Th differentiation [79]. Across models, increased IL-6 and IL-1β—drivers of Th17 polarization—are recurrent under plastic exposure [68]. Furthermore, mechanistic studies show that circulating MPs can suppress T helper cell cytokine production and interfere with T cell activation, thereby blunting Th cell–mediated immunity, and circulating MPs may blunt Th cytokine production and activation [59]. Net effects likely depend on exposure window, cytokine context, and APC competence.
3.2.1.3. Natural Killer T (NKT) Cells
Direct studies are lacking, but the observed cytokine pattern under MNPs exposure (↑IFN-γ, TNF-α, IL-1β; ↓IL-4, IL-10, TGF-β) [76] and sustained ROS/NF-κB/STAT3/p38 signaling [80] suggest potential impairment of NKT cells development and cytotoxic/regulatory functions. Such disruptions could contribute to systemic dysregulation and disease susceptibility.
3.2.1.4. Memory T Cells
Chronic exposure may erode memory T-cell durability by sustaining ROS-driven NF-κB/p38/STAT3 activation and pro-inflammatory cytokines, while APC dysfunction and altered IL-6/IL-1β signaling reduce recall efficiency [78,81]. MPs induced damage-associated molecular patterns (DAMP) release and TLR signaling may also impose trained-immunity–like reprogramming in innate cells, indirectly skewing memory T-cell reactivation.
3.2.2. Effects on B Cells
In rainbow trout cultures, 0.8–3.1 µm MPs are phagocytosed by B-lineage cells, reducing developing non-phagocytic B cells and downregulating RAG1 and Ig heavy chains (μ, τ), consistent with impaired lymphopoiesis and antibody potential [82]. More broadly, NPs induce ROS-mediated immune modulation and NF-κB activation across cell types, mechanisms that plausibly compromise B-cell viability and function [65]. Particle internalization can elicit oxidative stress, DAMP release, TLR perturbation, and cytokine dysregulation that hinder B-cell differentiation [78].
3.2.2.1. Memory B Cells (Emerging View)
Direct evidence is not yet available, but ROS, APC impairment, cytokine imbalance, and MP-induced dysbiosis [42] threaten germinal center dynamics and memory formation—raising concerns about humoral durability.
3.2.2.2. Regulatory B Cells (Bregs)
3.2.2.3. Long-Lived Plasma Cells
Survival depends on bone-marrow niches (stromal cues; IL-6, APRIL, BAFF) [84]. Systemic MNPs and their ROS-inflammatory signature may perturb these niches and depress plasma-cell longevity; NPs accumulate in immune populations and reduce viability [65], and broader hematopoietic interference could secondarily weaken antibody maintenance.
3.3. Disruption of Hematopoietic Stem Cell (HSC) Function
MPs reduce HSC self-renewal through microbiota–metabolite (e.g., hypoxanthine)–Wnt perturbations, impairing hematopoiesis and functional output [85]. Clinical correlations link higher MPs burdens in donor samples with poorer HSC transplantation outcomes, suggesting translational relevance. Mechanistically, MNPs disrupt hematopoiesis via gut dysbiosis, metabolic derangements, and cytokine shifts [86]; ~80 nm particles impair bone-marrow HSC renewal/differentiation in vivo [87]. Probiotics or melatonin partially rescue these defects, supporting microbiome-dependent oxidative and inflammatory pathways in HSC integrity. Overall, MNPs destabilize hematopoietic homeostasis by injuring stem-cell maintenance through microbiome signaling, oxidative damage, and metabolic dysfunction, with implications for blood production, immune competence, and HSC transplantation success.
Figure 2.
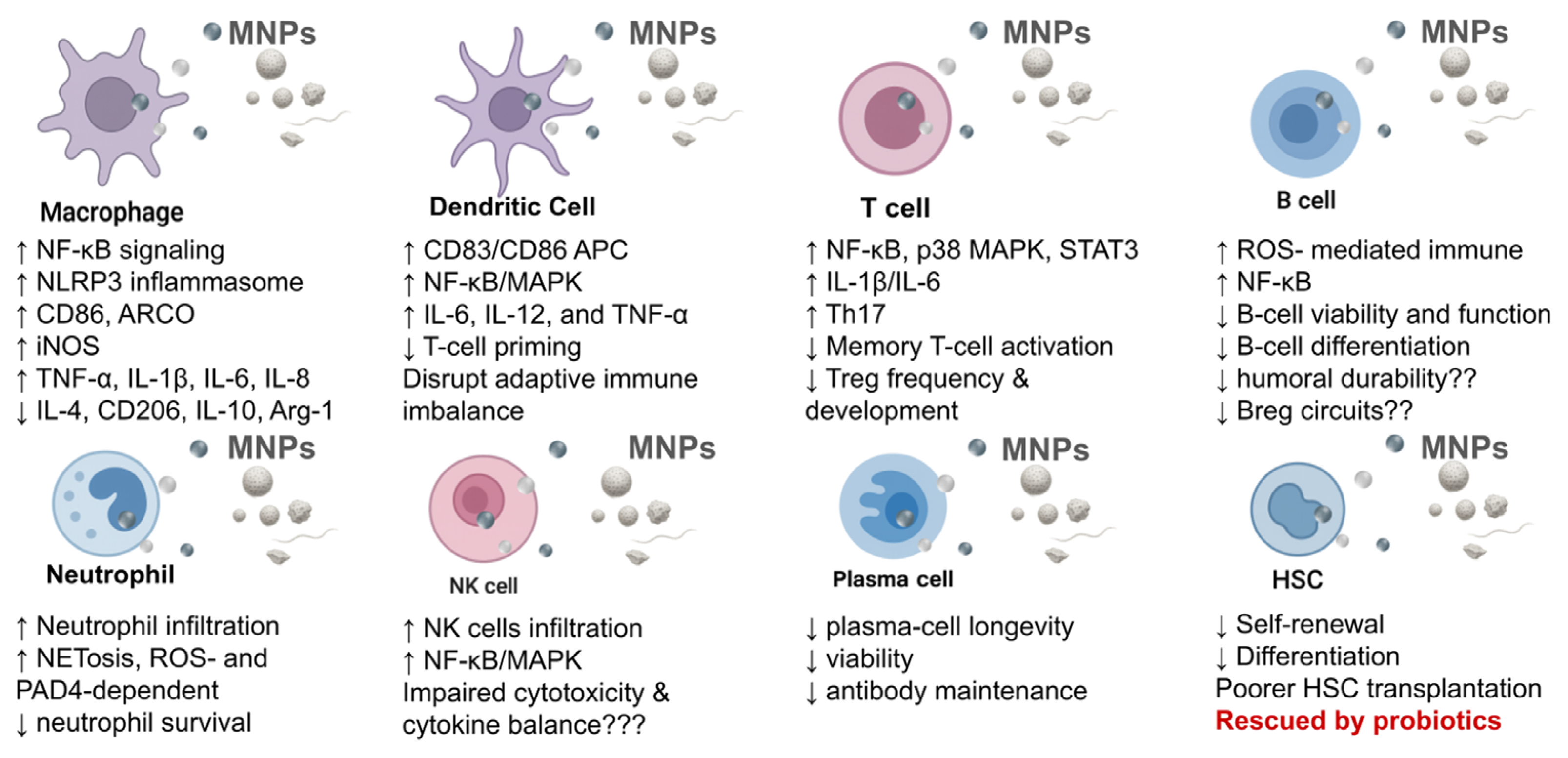
Immunotoxic effects of MNPs on immune cells. MNPs impair both innate and adaptive immune functions, including macrophage and dendritic cell activation, neutrophil NETosis, and NK cell dysfunction. They also disrupt T- and B-cell balance, reduce plasma-cell viability, and impair hematopoietic stem cell renewal, collectively promoting chronic inflammation and immune dysregulation.
Figure 2.
Immunotoxic effects of MNPs on immune cells. MNPs impair both innate and adaptive immune functions, including macrophage and dendritic cell activation, neutrophil NETosis, and NK cell dysfunction. They also disrupt T- and B-cell balance, reduce plasma-cell viability, and impair hematopoietic stem cell renewal, collectively promoting chronic inflammation and immune dysregulation.
4. Molecular Mechanisms and Signaling Pathways
4.1. Oxidative Stress and Mitochondrial Dysfunction
Exposure to MNPs consistently provokes oxidative stress, with excessive ROS driving DNA damage, protein oxidation, and lipid peroxidation. These injuries activate inflammatory programs and cellular senescence and are documented across cell lines, organoids, and animal models, where MNPs exposure disrupts redox balance and contributes to immune dysregulation and tissue injury [81]. Systematic review concur that NPs promote pro-inflammatory signaling, blunt antioxidant defenses, and impair tissue function, placing oxidative stress at the center of MNPs immunotoxicity [65].
Mitochondria are prominent targets. MNPs compromise organelle ultrastructure and function, lowering ATP output, collapsing the mitochondrial membrane potential, destabilizing mitochondrial DNA, and triggering selective autophagy [88]. They can also perturb mitochondrial biogenesis, dynamics, and the the mitochondrial unfolded protein response, culminating in energy failure, sustained ROS, chronic inflammation, and apoptosis [89]. Together, these effects link cellular oxidative injury to systemic immune dysfunction.
4.1.1. ROS Overproduction and Disruption of Antioxidant Defenses
Common polymers (e.g., PS, polyethylene) compromise membranes and the electron transport chain, increasing superoxide and hydrogen peroxide and driving lipid peroxidation; malondialdehyde is a frequent readout [90]. In parallel, antioxidant systems (SOD, CAT, GPx, glutathione reductase) are downregulated or overwhelmed, and Nrf2 activation is insufficient, reinforcing oxidative imbalance [90]. Dose–size relationships are evident: in AML12 cells, large MPs (~50 µm) raised intracellular ROS at higher concentrations (≈0.5 mg/mL) with limited compensatory antioxidant responses, leading to mitochondrial dysfunction and apoptosis [91]. In intestinal tissues, MPs elevate ROS, weaken barrier integrity, suppress antioxidant enzymes, and activate NF-κB–dependent inflammation [92]. Mechanistically, particle internalization or surface contact injures mitochondria and engages NADPH oxidases, converging on redox-sensitive MAPK, NF-κB, and p53 pathways that propagate cytokine expression and apoptosis/pyroptosis/ferroptosis. A self-reinforcing loop of ROS-induced antioxidant failure sustains organ dysfunction and chronic disease risk [90].
4.1.2. Loss of Mitochondrial Membrane Potential (Δψm)
Reduced Δψm is an early hallmark of MNPs injury and a prelude to apoptosis. In macrophages, NPs lower Δψm, activate cGAS–STING and NF-κB signaling, and drive apoptosis [60]. Epithelial models show Δψm loss with concomitant autophagy and inflammation even at non-cytotoxic doses, with higher exposures worsening depolarization and membrane compromise [81]. Across species, similar patterns emerge: in Eisenia fetida, 100 nm NPs increased ROS, suppressed antioxidant enzymes, and depolarized mitochondria, triggering early apoptotic features [93].
ROS damages the electron transport chain and peroxidizes mitochondrial membranes, promoting mitochondrial permeability transition pore opening, collapse of Δψm, cytochrome-c release, and caspase-9/-3 activation. In parallel, NF-κB amplifies inflammatory outputs, while cGAS–STING, MAPKs, and p53 integrate mitochondrial distress with immune and death signaling, coordinating progression from stress adaptation to apoptosis.
4.2. Inflammatory Signaling Pathways
MNPs activate core inflammatory circuits, notably NF-κB, MAPKs (p38/JNK/ERK), and the NLRP3 inflammasome. Zebrafish and mammalian models show NF-κB activation in gut and lung following PS exposure, consistent with conserved mechanisms across tissues and species [94,95]. In lung epithelium, polypropylene NPs drive p38–NF-κB signaling, oxidative stress, mitochondrial damage, and IL-6/TNF-α secretion; blocking p38 or ROS dampens these effects [75]. TLR4–Nox2–dependent oxidative bursts further amplify inflammation and cellular senescence, linking barrier injury to chronic immune activation [92].
4.2.1. NF-κB Pathway
In macrophages, surface-functionalized NPs are internalized, increase ROS, and activate MAPK/NF-κB, elevating IL-6 and TNF-α; carboxylated particles can elicit stronger NF-κB activation [64]. Polypropylene NPs induce lung inflammation via p38-dependent NF-κB activation with concordant mitochondrial injury and cytokine upregulation, in vivo and in vitro [75]. Lactational MPs exposure upregulates TLR4–MyD88 and phosphorylated NF-κB in colon, consistent with barrier leakage and LPS co-stimulation [96].
4.2.2. Mitogen-Activated Protein Kinase (MAPK) Pathways
PS MPs activate p38 in reproductive models, elevating IL-6/IL-1β and impairing fertility; MPs also interface with endoplasmic reticulum (ER) stress, autophagy (JNK/ERK), and AKT/mTOR nodes, underscoring broad stress-pathway engagement [77]. In macrophages, smaller PS NPs evoke stronger MAPK activation than larger MPs, increase ROS, bias M1 polarization (↑TNF-α/IL-6, ↓M2 markers), and show responses contingent on particle size/shape [60]. Overall, ROS generated by MNPs exposure activates p38/JNK/ERK, reprogramming transcription toward inflammation, apoptosis, and tissue damage, with implications for organ function, fertility, and carcinogenesis [77].
4.2.3. NLRP3 Inflammasome
Across species, MNPs activate NLRP3 via ROS and ER stress. In poultry, chronic MPs exposure upregulated ER stress markers (ATF6, PERK, eIF2α) and NLRP3 components (NLRP3, ASC, caspase-1) along with IL-1β and GSDMD, yielding histological injury [97]. In THP-1–derived macrophages, pristine and modified plastics promoted caspase-1 activation and IL-1β release; effects were attenuated in NLRP3-deficient cells [98]. Aquatic models show analogous NLRP3 activation and IL-1β induction with oxidative stress and tissue inflammation [99].
4.3. Apoptosis and Cell Death Programs
Apoptosis is frequently initiated through the intrinsic (mitochondrial) pathway after MNPs exposure. In trophoblasts, NPs (50–200 µg/mL) increase ROS, decrease Δψm, suppress Bcl-2, and activate caspase-2/-3, culminating in apoptosis [100]. Similar cascades are reported in lung epithelia, where antioxidant failure, mitochondrial damage, and caspase-3 activation are prominent [100,101,102]. Ferroptosis, characterized by iron-dependent lipid peroxidation, GSH depletion, and GPX4 inhibition, has also been observed in intestinal epithelium exposed to 100 nm particles [103]. These modes can co-occur, indicating size-, dose-, and context-dependent toxicity.
4.3.1. Intrinsic and Extrinsic Apoptosis
4.3.2. Autophagy Dysregulation
NPs increase LC3-II and p62 with Δψm loss and ROS, indicating impaired flux rather than efficient clearance [108]. Inhaled or dust-associated MPs may suppress autophagy via PI3K/AKT/mTOR, accompanying a higher Bax/Bcl-2 ratio and caspase-3 activation [109]. ER stress–UPR signaling (e.g., GRP78, MAPK14, XBP1) can also engage autophagy; prolonged exposure shifts the response from adaptive to cytotoxic, culminating in autophagic cell death [110,111].
4.3.3. Pyroptosis
MNPs can initiate canonical pyroptosis characterized by caspase-1 activation, GSDMD cleavage, and IL-1β/IL-18 release [112]. NLRP3 serves as a principal upstream sensor in multiple models [56], and neuronal exposures to NPs show inflammasome assembly and caspase-1/GSDMD activation with increased IL-1β, implicating pyroptosis in neuroinflammation [113].
4.3.4. Convergence of Programmed Cell-Death Pathways
MNPs engage overlapping death programs, apoptosis, ferroptosis, pyroptosis, necroptosis, and dysregulated autophagy, largely via ROS, lipid peroxidation, mitochondrial and lysosomal injury, and redox failure [112]. In the gut, ROS-driven apoptosis compromises barrier integrity and may facilitate systemic particle spread [114], while 100 nm NPs preferentially induce ferroptosis [103]. This convergence underscores multi-modal cytotoxicity that threatens tissue homeostasis and immune competence.
5. Immunomodulatory and Autoimmune Implications
5.1. MNPs and Chronic Low-Grade Inflammation
MNPs sustains a state of chronic low-grade inflammation characterized by persistent immune signaling, tissue stress, and progressive dysfunction. Key pathways include NLRP3 inflammasome and NF-κB activation, which together maintain pro-inflammatory cytokine release [115]. Ongoing accumulation also perturbs the microbiome and drives systemic inflammatory responses through continuous exposure to inflammatory mediators [77]. In the lung, polypropylene NPs elicit mitochondrial injury and activate the p38 MAPK–NF-κB axis, a mechanism that can propagate persistent inflammation across tissues [75].
5.2. Disruption of Immune Tolerance Via Barrier Dysfunction and Innate Activation
Environmental plastic particles compromise epithelial barriers in the skin, gut, and airways, increasing permeability, promoting dysbiosis, and facilitating antigen translocation. The epithelial-barrier framework posits that such damage elevates epithelial cytokines (IL-25, IL-33, TSLP), weakens peripheral tolerance, and predisposes to allergy and autoimmunity [8]. NPs exposure further induces oxidative stress and cytokine imbalance, overriding regulatory circuits and sustaining inflammation [65]. MPs also engage TLRs and promote DAMP release, reinforcing innate activation and interfering with tolerance maintenance [78].
5.3. Immunomodulatory and Autoimmune Consequences of MNPs Exposure
Across models, MNPs disturb the gut–immune axis, hematopoiesis, and immune cell activation, shifting the balance between tolerance and autoimmunity [68]. Aerosolized NPs induce pro-inflammatory signaling in airway epithelium, increase neutrophil recruitment, and reduce modulators such as IL-21, IL-2, IL-15, CXCL10, and TGF-β, thereby weakening regulatory responses and tissue repair [28]. In a murine arthritis model, plastic exposure worsened synovial inflammation and joint pathology, linking particles to disease exacerbation [116]. Within the epithelial-barrier framework, MPs damage mucosal integrity, fuel dysbiosis, and promote IL-25/IL-33/TSLP-driven activation, providing a plausible route to chronic inflammatory priming and autoimmunity [117]. Review study further emphasizes ROS excess, skewed cytokine profiles, and pro-inflammatory signaling as canonical features of NPs immunotoxicity [65]; MPs also engage TLR pathways and DAMP signaling to tilt responses toward self-reactivity⁴⁴. Notably, mice exposed to MPs developed prolonged post-viral arthritis despite minimal microbiome shifts, indicating that MNPs can prime exaggerated secondary immune responses.
5.4. Potential Links to Specific Autoimmune Diseases
5.4.1. Systemic Lupus Erythematosus (SLE)
In mice, MPs exposure induced lupus-like features and exacerbated disease in MRL/lpr animals, including expansion of double-negative T cells, increased plasma cells, higher anti-dsDNA/ANA titers, elevated IL-6/TNF-α, altered CD4⁺/CD8⁺ ratios, and renal pathology consistent with lupus nephritis. Proteomics implicated aberrant MHC-II regulation and histone modifications in renal injury, suggesting inflammatory and epigenetic contributions to systemic autoimmunity [118].
5.4.2. Inflammatory Bowel Disease (IBD)
Epidemiology associates higher fecal MPs burdens with IBD severity [119]. Complementary models, in mice and intestinal organoids, show that plastic exposure exacerbates colitis via barrier disruption, dysbiosis, and innate activation; organoids secrete more inflammatory cytokines and in vivo tissues display greater ulceration and pathology [120,121]. These observations align with barrier-failure mechanisms thought to promote chronic intestinal inflammation and systemic immune dysregulation [8,117].
5.4.3. Rheumatoid Arthritis (RA) and TLR/NF-κB Signaling
In vivo, MPs worsen synovitis, cartilage damage, and matrix degradation, with histological confirmation [116,122]. Mechanistically, oxidative stress and barrier injury increase DAMP release and TLR engagement, sustaining NF-κB-dependent cytokine production and synovial inflammation. Similar TLR4–MyD88–NF-κB activation is observed in colonic tissue after PS exposure [96], and persistent NF-κB activation has been linked to chronic inflammation with potential oncogenic sequelae [123]. Cardiac models indicate that the same TLR4–NF-κB axis can mediate plastic induced myocardial inflammation, underscoring a conserved pathway across organs [124].
While direct epidemiologic links in humans remain limited, convergent mechanistic and preclinical evidence supports a multi-hit model wherein MNPs compromise barriers, induce oxidative stress, and activate TLR/NF-κB signaling to sustain inflammation and erode tolerance, elevating autoimmune risk.
6. Current Knowledge Gaps and Research Challenges
6.1. Lack of Human Cohort and Longitudinal Studies
MNPs relies on experimental models, environmental sampling, or short-term exposures, while population-level outcomes over time remain largely unexplored. Although exposure routes have been documented, epidemiological data directly linking MPs exposure to disease progression are lacking [14,125]. Systematic reviews emphasize that the field has focused disproportionately on environmental burdens and biomarker detection rather than longitudinal exposure disease relationships. Biomarkers of exposure are now being reported, but causal inference remains untested. Standardized exposure assessments and long-term studies are urgently needed to establish a credible evidence base for public health and policy development [126,127].
Evidence of human exposure is unequivocal: MPs have been detected in the digestive system, blood, lungs, and other tissues [128]. However, the health implications of these findings remain speculative. While analytical methods for detection have advanced rapidly, interpretation of long-term consequences lags behind. Commentaries from medical institutions describe this field as still in its infancy, with major challenges in evaluating population-level exposures and health outcomes.
In sum, key gaps include: (i) the absence of longitudinal cohort studies; (ii) lack of standardized protocols for exposure assessment; and (iii) inability to establish causality without prospective data. Addressing these limitations will require integrating exposure biomarkers into large-scale cohorts such as the UK Biobank or Nurses’ Health Study. Standardization of sampling, measurement, and reporting protocols is critical for comparability, while multi-omics and exposome approaches could integrate environmental, metabolic, and clinical data to clarify mechanistic pathways.
6.2. Poor Standardization of MNPs Parameters
6.2.1. Experimental Inconsistencies
Most laboratory studies employ pristine, spherical PS beads, whereas real-world exposures involve heterogeneous fibers, fragments, and weathered particles. This discrepancy raises concerns about ecological and toxicological relevance [129].
6.2.2. Variable Particle Sizes and Dosing Protocols
6.2.3. Sampling and Quality Control
Protocols for particle preparation, homogenization, and contamination control differ widely, with most studies lacking standardized reference materials. Surveys suggest nearly 80% of researchers view this lack of harmonized methods as a primary barrier [132]. New approaches such as cryomilling can generate more environmentally relevant fragments, but they are not yet standardized [133].
Collectively, the progress will require: (i) using particle types that mimic environmental conditions, including weathered fragments and fibers; (ii) aligning doses with realistic human exposures; (iii) adopting validated reference materials, harmonized sampling/extraction protocols, and strict contamination controls; and (iv) promoting interlaboratory collaborations and shared reference standards.
6.3. Interactions Between MNPs, Co-Pollutants, and the Gut Microbiota
6.3.1. Co-Exposures with Pollutants
Interactions between MNPs and other contaminants such as heavy metals remain poorly characterized, especially regarding their combined effects on gut microbiota. Despite the likelihood of real-world co-exposures, systematic studies in humans are virtually absent [134].
6.3.2. Gut Microbiota Disruption
MNPs consistently induce dysbiosis, often reducing Bacteroidetes and beneficial genera in animal and in vitro models. However, the mechanistic basis of these alterations remains unclear. Evidence suggests changes in microbial composition, metabolism, and microbial colonization of MPs, but the biochemical pathways remain speculative [135,136].
6.3.3. Ecosystem Perspective
Research is fragmented and often restricted to specific organisms, limiting generalizability. Reviews stress the need for One Health approaches integrating human, animal, soil, and insect microbiomes [137].
Critical gaps include the synergistic effects of MNPs with heavy metals or organic pollutants, mechanisms underlying dysbiosis and microbial colonization, and the potential for gut microbes to biodegrade plastics. Priorities include: (i) developing human-relevant gut models such as organoid–microbiota co-cultures; (ii) applying metagenomics and metabolomics to resolve microbial and metabolic disruptions; (iii) standardizing co-exposure protocols; and (iv) launching epidemiological studies correlating MNPs exposure with microbiome shifts and health outcomes.
6.4. Challenges in Tracing MNPs In Vivo
6.4.1. Lack of Reliable Tracking Methods
6.4.2. Limitations in Imaging and quantification
Techniques such as FTIR, Raman, pyrolysis-GC-MS, and fluorescence microscopy are well-suited for environmental sampling but lack the sensitivity and resolution to detect NPs in complex tissues [140]. Fluorescent labeling introduces artifacts such as leakage, photobleaching, or aggregation, while tissue autofluorescence and the inability to detect particles <100 nm hinder accurate biodistribution mapping [141].
6.4.3. Lack of Standardization in Particle Characterization
Most studies use pristine PS spheres that differ substantially from environmental particles [142]. Moreover, many experiments apply extremely high doses of MNPs for short exposure periods, which fail to reflect realistic chronic, low-dose human exposure scenario. Experimental designs often apply high, acute doses rather than chronic, low-level exposures, limiting translational value [143].
Future research requires multi-modal tracers, such as, radiolabeled, isotopically tagged, or magnetically responsive plastics, suitable for long-term animal studies. Advances in PET, MRI, and hyperspectral imaging offer opportunities for real-time in vivo tracking, while mass spectrometry–based quantification could improve sensitivity and accuracy [144].
7. Future Perspectives and Research Directions
7.1. Advancing Human Epidemiological Evidence
The most urgent priority is the establishment of large-scale, longitudinal human studies that can directly link MNPs exposure to health outcomes. Cohorts should integrate exposure biomarkers, such as plastic particle quantification in blood, urine, and stool, into ongoing population studies (e.g., UK Biobank, Nurses’ Health Study). Harmonization of sampling, quantification, and reporting protocols will be critical to ensure comparability across populations and over time. Such studies will help shift the field from descriptive detection toward causal inference and disease risk prediction.
7.2. Improving Environmental and Experimental Realism
To enhance translational relevance, future work must adopt particle types and exposure conditions that mirror environmental reality. This includes using weathered fragments, fibers, and biofilm-coated plastics rather than pristine PS beads. Dose ranges should align with real-world estimates of chronic, low-dose human exposure. Development and adoption of standardized reference materials, as well as interlaboratory comparisons, will be essential for reproducibility.
7.3. Innovations in Detection, Imaging, and Tracing
Progress in vivo requires the development of advanced tracers and multi-modal imaging technologies. Radiolabeled or isotopically tagged plastics, combined with PET or MRI, may enable dynamic tracking of biodistribution and clearance. Hyperspectral imaging and single-particle mass spectrometry hold promise for tissue-level detection, while multi-omics readouts, such as proteomics, metabolomics, and epigenomics, could link exposure signatures to immune and metabolic outcomes. Integration of these technologies will be key to defining dose–response relationships and organ-specific effects.
7.4. Mechanistic Insights into Host–Microbe–Pollutant Interactions
Future studies should prioritize mechanistic dissection of how MNPs interact with the gut microbiota, immune cells, and co-pollutants. Human-relevant organoid–microbiota co-cultures, coupled with metagenomic and metabolomic profiling, can clarify microbial contributions to immune dysregulation and pollutant metabolism. Investigating how plastics alter microbial ecology and antimicrobial resistance will also be important for both health and ecological risk assessments.
7.5. Clinical and Immunological Endpoints
Research should move beyond descriptive inflammatory markers toward clinically relevant endpoints. For example, evaluating how MNPs influence vaccine responses, immune aging, autoimmune disease severity, and outcomes after hematopoietic stem cell transplantation will provide direct translational insights. Animal models should be optimized for chronic exposure windows, developmental periods, and comorbid conditions to better approximate human risk.
7.6. Policy, Mitigation, and Intervention Strategies
Finally, science must inform mitigation efforts. Standardized monitoring of human exposure, such as dietary intake, air sampling in enclosed environments, will support regulatory policies on plastic production, packaging, and waste management. Clinical trials exploring antioxidants, probiotics, or dietary interventions could help mitigate MNPs-associated oxidative stress and microbiome dysbiosis. At the public health level, integrating MNPs risk into environmental health frameworks will be critical for global prevention strategies.
8. Conclusions
MNPs have emerged as pervasive environmental contaminants with growing evidence for biological relevance in humans. Experimental and early epidemiological studies demonstrate that MNPs can enter the body through ingestion, inhalation, and dermal contact, accumulate in immune organs, and disrupt both innate and adaptive immune responses. At the molecular level, MNPs provoke oxidative stress, mitochondrial dysfunction, and activation of NF-κB, MAPK, and NLRP3 inflammasome pathways, leading to chronic inflammation, apoptosis, pyroptosis, and altered cell signaling. These mechanisms collectively compromise immune homeostasis, heighten susceptibility to infections, and may accelerate the development of autoimmune and chronic inflammatory diseases.
Despite these advances, major gaps remain in understanding human health risks. Current research is limited by reliance on short-term animal models, pristine particles, and supra-physiological exposures, with little data from longitudinal human cohorts. Standardization of exposure assessment, particle characterization, and experimental protocols is urgently needed. Future research must also expand to include human-relevant models, co-pollutant interactions, and microbiome dynamics. Bridging these gaps will require interdisciplinary efforts that integrate advanced imaging, multi-omics profiling, and large-scale epidemiology. Only through such approaches can we establish causal links between plastic exposure and disease, ultimately guiding effective mitigation strategies and informing public health policy.
Author Contributions
J.F. conceived the idea, initiated, and composed the manuscript. J.F. conducted the literature search and summary, Y.H. oversaw the manuscript preparation process. All authors critically reviewed and endorsed the final manuscript.
Funding
Not applicable.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| APC | Antigen presenting cell |
| BPA | Bisphenol A |
| Bregs | Regulatory B cells |
| DAMP | Damage-associated molecular patterns |
| DCs | Dendritic cells |
| ER | Endoplasmic reticulum |
| FTIR | Fourier Transform Infrared spectroscopy |
| GC-MS | Gas Chromatography-Mass Spectrometry |
| GSDMD | Gasdermin D |
| GSH | Glutathione |
| HSC | Hematopoietic stem cell |
| IBD | Inflammatory Bowel Disease |
| IFN-γ | Interferon-γ |
| IL | Interleukin |
| iNOS | Inducible Nitric Oxide Synthase |
| LPS | Lipopolysaccharide |
| MAPK | Mitogen-Activated Protein Kinase |
| MARCO | Macrophage receptor with collagenous structure |
| METs | Macrophage extracellular traps |
| MNPs | Micro- and Nano-Plastics |
| MPs | Microplastics |
| MRI | Magnetic resonance imaging |
| NETs | Neutrophil extracellular traps |
| NF-κB | Nuclear Factor kappa-light-chain-enhancer of activated B cells |
| NK | Natural Killer |
| NKT | Natural Killer T |
| NLRP3 | Nod-like receptor protein 3 |
| NPs | Nanoplastics |
| PET | Positron emission tomography |
| PPR | Pattern Recognition Receptor |
| PS | Polystyrene |
| RA | Rheumatoid Arthritis |
| ROS | Reactive oxygen species |
| SLE | Systemic Lupus Erythematosus |
| TCR | T cell receptor |
| TGF-β | Transforming growth factor beta |
| Th | Helper T cells |
| TLR | Toll like receptor |
| Treg | Regulatory T cells |
| UPR | Unfolded Protein Response |
| UV | Ultraviolet |
References
- Mohajan, H.K. Plastic Pollution: A Potential Threat on Health and Environment. Stud. Soc. Sci. Humanit. 2025, 4, 25–30. [Google Scholar] [CrossRef]
- Plastic Pollution. Available online: https://iucn.org/resources/issues-brief/plastic-pollution (accessed on 26 August 2025).
- Geyer, R.; Jambeck, J.R.; Law, K.L. Production, Use, and Fate of All Plastics Ever Made. Sci. Adv. 2017, 3, e1700782. [Google Scholar] [CrossRef]
- Abimbola, A.N.; Adejumobi, V.O.; Aribisala, O.C.; Oyeniyi, E.O. Influence of Plastic Waste Management on the Environment: A Review. Environ. Technol. Sci. J. 2023, 14, 56–64. [Google Scholar] [CrossRef]
- Jambeck, J.R.; Geyer, R.; Wilcox, C.; Siegler, T.R.; Perryman, M.; Andrady, A.; Narayan, R.; Law, K.L. Plastic Waste Inputs from Land into the Ocean. Science 2015, 347, 768–771. [Google Scholar] [CrossRef] [PubMed]
- Law, K.L.; Thompson, R.C. Microplastics in the Seas. Science 2014, 345, 144–145. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.C.; Courtene-Jones, W.; Boucher, J.; Pahl, S.; Raubenheimer, K.; Koelmans, A.A. Twenty Years of Microplastic Pollution Research—What Have We Learned? Science 2024, 386, eadl2746. [Google Scholar] [CrossRef]
- Li, B.; Ding, Y.; Cheng, X.; Sheng, D.; Xu, Z.; Rong, Q.; Wu, Y.; Zhao, H.; Ji, X.; Zhang, Y. Polyethylene Microplastics Affect the Distribution of Gut Microbiota and Inflammation Development in Mice. Chemosphere 2020, 244, 125492. [Google Scholar] [CrossRef]
- Thompson, R.C.; Olsen, Y.; Mitchell, R.P.; Davis, A.; Rowland, S.J.; John, A.W.G.; McGonigle, D.; Russell, A.E. Lost at Sea: Where Is All the Plastic? Science 2004, 304, 838–838. [Google Scholar] [CrossRef]
- Mcguinness, K. Nanoplastics . Futurist 1995, 29, 50–53. [Google Scholar]
- Yu, Q.; Chuang, C.-Y.A.; Jiang, Y.; Zhong, H.; Cundy, A.; Kwong, R.W.M.; Min, C.; Zhu, X.; Ji, R. Exploring Environmental Nanoplastics Research: Networks and Evolutionary Trends. Rev. Environ. Contam. Toxicol. 2023, 261, 12. [Google Scholar] [CrossRef]
- Lai, H.; Liu, X.; Qu, M. Nanoplastics and Human Health: Hazard Identification and Biointerface. Nanomaterials 2022, 12, 1298. [Google Scholar] [CrossRef]
- Bhattacharya, P.; Lin, S.; Turner, J.P.; Ke, P.C. Physical Adsorption of Charged Plastic Nanoparticles Affects Algal Photosynthesis. J. Phys. Chem. C 2010, 114, 16556–16561. [Google Scholar] [CrossRef]
- Nawab, A.; Ahmad, M.; Khan, M.T.; Nafees, M.; Khan, I.; Ihsanullah, I. Human Exposure to Microplastics: A Review on Exposure Routes and Public Health Impacts. J. Hazard. Mater. Adv. 2024, 16, 100487. [Google Scholar] [CrossRef]
- Jahedi, F.; Jaafarzadeh Haghighi Fard, N. Micro- and Nanoplastic Toxicity in Humans: Exposure Pathways, Cellular Effects, and Mitigation Strategies. Toxicol. Rep. 2025, 14, 102043. [Google Scholar] [CrossRef] [PubMed]
- Niccolai, E.; Colzi, I.; Amedei, A. Adverse Effects of Micro- and Nanoplastics on Humans and the Environment. Int. J. Mol. Sci. 2023, 24, 15822. [Google Scholar] [CrossRef]
- Sajedi, S.; An, C.; Chen, Z. Unveiling the Hidden Chronic Health Risks of Nano- and Microplastics in Single-Use Plastic Water Bottles: A Review. J. Hazard. Mater. 2025, 495, 138948. [Google Scholar] [CrossRef] [PubMed]
- Covello, C.; Di Vincenzo, F.; Cammarota, G.; Pizzoferrato, M. Micro(Nano)Plastics and Their Potential Impact on Human Gut Health: A Narrative Review. Curr. Issues Mol. Biol. 2024, 46, 2658–2677. [Google Scholar] [CrossRef]
- Zhou, L.; Ran, L.; He, Y.; Huang, Y. Mechanisms of Microplastics on Gastrointestinal Injury and Liver Metabolism Disorder (Review). Mol. Med. Rep. 2025, 31, 98. [Google Scholar] [CrossRef] [PubMed]
- Vagner, M.; Boudry, G.; Courcot, L.; Vincent, D.; Dehaut, A.; Duflos, G.; Huvet, A.; Tallec, K.; Zambonino-Infante, J.-L. Experimental Evidence That Polystyrene Nanoplastics Cross the Intestinal Barrier of European Seabass. Environ. Int. 2022, 166, 107340. [Google Scholar] [CrossRef] [PubMed]
- Prata, J.C. Microplastics and Human Health: Integrating Pharmacokinetics. Crit. Rev. Environ. Sci. Technol. 2023, 53, 1489–1511. [Google Scholar] [CrossRef]
- Wang, Y.-F.; Wang, X.-Y.; Chen, B.-J.; Yang, Y.-P.; Li, H.; Wang, F. Impact of Microplastics on the Human Digestive System: From Basic to Clinical. World J. Gastroenterol. 2025, 31. [Google Scholar] [CrossRef]
- Yakovenko, N.; Pérez-Serrano, L.; Segur, T.; Hagelskjaer, O.; Margenat, H.; Roux, G.L.; Sonke, J.E. Human Exposure to PM10 Microplastics in Indoor Air. PLOS ONE 2025, 20, e0328011. [Google Scholar] [CrossRef]
- Ningrum, P.T.; Keman, S.; Sulistyorini, L.; Sudiana, I.K.; Hidayat, A.; Negoro, A.H.S.; Junaidi, H.; Kustin, K. A Systematic Review of the Effects of Airborne Microplastic Contamination on Human Lungs. Afr. J. Reprod. Health 2024, 28. [Google Scholar] [CrossRef]
- Gou, Z.; Wu, H.; Li, S.; Liu, Z.; Zhang, Y. Airborne Micro- and Nanoplastics: Emerging Causes of Respiratory Diseases. Part. Fibre Toxicol. 2024, 21, 50. [Google Scholar] [CrossRef]
- Mousseau, F.; Berret, J.-F. The Role of Surface Charge in the Interaction of Nanoparticles with Model Pulmonary Surfactants. Soft Matter 2018, 14, 5764–5774. [Google Scholar] [CrossRef]
- Radiom, M.; Sarkis, M.; Brookes, O.; Oikonomou, E.K.; Baeza-Squiban, A.; Berret, J.-F. Pulmonary Surfactant Inhibition of Nanoparticle Uptake by Alveolar Epithelial Cells. Sci. Rep. 2020, 10, 19436. [Google Scholar] [CrossRef]
- Breidenbach, J.D.; French, B.W.; Shrestha, U.; Adya, Z.K.; Wooten, R.M.; Fribley, A.M.; Malhotra, D.; Haller, S.T.; Kennedy, D.J. Acute Exposure to Aerosolized Nanoplastics Modulates Redox-Linked Immune Responses in Human Airway Epithelium. Antioxidants 2025, 14, 424. [Google Scholar] [CrossRef]
- Pat, Y.; Yazici, D.; D’Avino, P.; Li, M.; Ardicli, S.; Ardicli, O.; Mitamura, Y.; Akdis, M.; Dhir, R.; Nadeau, K.; et al. Recent Advances in the Epithelial Barrier Theory. Int. Immunol. 2024, 36, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Pileta-Labañino, M.; Crespo-Barrera, P.; Navarro-Frómeta, A. Human Skin and Micro- and Nanoplastics: A Mini-Review. MOJ Ecol. Environ. Sci. 2024, 9. [Google Scholar] [CrossRef]
- Aristizabal, M.; Jiménez--Orrego, K.V.; Caicedo--León, M.D.; Páez--Cárdenas, L.S.; Castellanos--García, I.; Villalba--Moreno, D.L.; Ramírez--Zuluaga, L.V.; Hsu, J.T.S.; Jaller, J.; Gold, M. Microplastics in Dermatology: Potential Effects on Skin Homeostasis. J. Cosmet. Dermatol. 2024, 23, 766–772. [Google Scholar] [CrossRef] [PubMed]
- Hoang, H.G.; Nguyen, N.S.H.; Zhang, T.; Tran, H.-T.; Mukherjee, S.; Naidu, R. A Review of Microplastic Pollution and Human Health Risk Assessment: Current Knowledge and Future Outlook. Front. Environ. Sci. 2025, 13. [Google Scholar] [CrossRef]
- Menichetti, A.; Mordini, D.; Montalti, M. Penetration of Microplastics and Nanoparticles Through Skin: Effects of Size, Shape, and Surface Chemistry. J. Xenobiotics 2025, 15, 6. [Google Scholar] [CrossRef]
- Martin, L.; Simpson, K.; Brzezinski, M.; Watt, J.; Xu, W. Cellular Response of Keratinocytes to the Entry and Accumulation of Nanoplastic Particles. Part. Fibre Toxicol. 2024, 21, 22. [Google Scholar] [CrossRef]
- Thapliyal, C.; Negi, S.; Nagarkoti, S.; Daverey, A. Mechanistic Insight into Potential Toxic Effects of Microplastics and Nanoplastics on Human Health. Discov. Appl. Sci. 2025, 7, 645. [Google Scholar] [CrossRef]
- López de las Hazas, M.-C.; Boughanem, H.; Dávalos, A. Untoward Effects of Micro- and Nanoplastics: An Expert Review of Their Biological Impact and Epigenetic Effects. Adv. Nutr. 2022, 13, 1310–1323. [Google Scholar] [CrossRef] [PubMed]
- Jayavel, S.; Govindaraju, B.; Michael, J.R.; Viswanathan, B. Impacts of Micro and Nanoplastics on Human Health. Bull. Natl. Res. Cent. 2024, 48, 110. [Google Scholar] [CrossRef]
- Yang, Y.-W.; Luo, W.-H. Cellular Biodistribution of Polymeric Nanoparticles in the Immune System. J. Controlled Release 2016, 227, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Wang, L.; Wang, X.; Li, D.; Wang, H.; Xu, H.; Liu, Y.; Kang, R.; Chen, Q.; Zheng, L.; et al. Discovery and Analysis of Microplastics in Human Bone Marrow. J. Hazard. Mater. 2024, 477, 135266. [Google Scholar] [CrossRef]
- Deng, X.; Gui, Y.; Zhao, L. The Micro(Nano)Plastics Perspective: Exploring Cancer Development and Therapy. Mol. Cancer 2025, 24, 30. [Google Scholar] [CrossRef] [PubMed]
- Park, S.B.; Jung, W.H.; Choi, K.J.; Koh, B.; Kim, K.Y. A Comparative Systematic Analysis of The Influence of Microplastics on Colon Cells, Mouse and Colon Organoids. Tissue Eng. Regen. Med. 2022, 20, 49–58. [Google Scholar] [CrossRef]
- Hirt, N.; Body-Malapel, M. Immunotoxicity and Intestinal Effects of Nano- and Microplastics: A Review of the Literature. Part. Fibre Toxicol. 2020, 17, 57. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yin, K.; Hou, L.; Zhang, Y.; Lu, H.; Ma, C.; Xing, M. Polystyrene Microplastics Mediate Inflammatory Responses in the Chicken Thymus by Nrf2/NF-κB Pathway and Trigger Autophagy and Apoptosis. Environ. Toxicol. Pharmacol. 2023, 100, 104136. [Google Scholar] [CrossRef]
- Yang, W.; Li, Y.; Boraschi, D. Association between Microorganisms and Microplastics: How Does It Change the Host–Pathogen Interaction and Subsequent Immune Response? Int. J. Mol. Sci. 2023, 24, 4065. [Google Scholar] [CrossRef]
- Stunnenberg, M.; de Roda Husman, A.M. It’s a Matter of Microbes: A Perspective on the Microbiological Aspects of Micro- and Nanoplastics in Human Health. Front. Nanotechnol. 2024, 6. [Google Scholar] [CrossRef]
- Zhi, L.; Li, Z.; Su, Z.; Wang, J. Immunotoxicity of Microplastics: Carrying Pathogens and Destroying the Immune System. TrAC Trends Anal. Chem. 2024, 177, 117817. [Google Scholar] [CrossRef]
- Gross, N.; Muhvich, J.; Ching, C.; Gomez, B.; Horvath, E.; Nahum, Y.; Zaman, M.H. Effects of Microplastic Concentration, Composition, and Size on Escherichia Coli Biofilm-Associated Antimicrobial Resistance. Appl. Environ. Microbiol. 2025, 91, e02282–24. [Google Scholar] [CrossRef]
- Nath, J.; Banerjee, G.; De, J.; Dsouza, N.; Sur, S.; Scott, J.W.; Banerjee, P. Nanoplastics-Mediated Physiologic and Genomic Responses in Pathogenic Escherichia Coli O157:H7. J. Nanobiotechnology 2025, 23, 304. [Google Scholar] [CrossRef]
- Lu, J.; Yu, Z.; Ngiam, L.; Guo, J. Microplastics as Potential Carriers of Viruses Could Prolong Virus Survival and Infectivity. Water Res. 2022, 225, 119115. [Google Scholar] [CrossRef]
- Wang, X.; Zheng, K.; Wang, Y.; Hou, X.; He, Y.; Wang, Z.; Zhang, J.; Chen, X.; Liu, X. Microplastics and Viruses in the Aquatic Environment: A Mini Review. Front. Microbiol. 2024, 15. [Google Scholar] [CrossRef] [PubMed]
- Cai, B.; De Jesus Andino, F.; McGrath, J.L.; Romanick, S.S.; Robert, J. Ingestion of Polyethylene Terephthalate Microplastic Water Contaminants by Xenopus Laevis Tadpoles Negatively Affects Their Resistance to Ranavirus Infection and Antiviral Immunity. Environ. Pollut. 2024, 356, 124340. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Lu, L.; Tu, W.; Luo, T.; Fu, Z. Impacts of Polystyrene Microplastic on the Gut Barrier, Microbiota and Metabolism of Mice. Sci. Total Environ. 2019, 649, 308–317. [Google Scholar] [CrossRef]
- Li, L.; Yang, J.; Zhou, Q.; Peijnenburg, W.J.G.M.; Luo, Y. Uptake of Microplastics and Their Effects on Plants. In Microplastics in Terrestrial Environments: Emerging Contaminants and Major Challenges; He, D., Luo, Y., Eds.; Springer International Publishing: Cham, 2020; ISBN 978-3-030-56271-7. [Google Scholar]
- Mattioda, V.; Benedetti, V.; Tessarolo, C.; Oberto, F.; Favole, A.; Gallo, M.; Martelli, W.; Crescio, M.I.; Berio, E.; Masoero, L.; et al. Pro-Inflammatory and Cytotoxic Effects of Polystyrene Microplastics on Human and Murine Intestinal Cell Lines. Biomolecules 2023, 13, 140. [Google Scholar] [CrossRef]
- Wang, L.; Pei, W.; Li, J.; Feng, Y.; Gao, X.; Jiang, P.; Wu, Q.; Li, L. Microplastics Induced Apoptosis in Macrophages by Promoting ROS Generation and Altering Metabolic Profiles. Ecotoxicol. Environ. Saf. 2024, 271, 115970. [Google Scholar] [CrossRef]
- Alijagic, A.; Hedbrant, A.; Persson, A.; Larsson, M.; Engwall, M.; Särndahl, E. NLRP3 Inflammasome as a Sensor of Micro- and Nanoplastics Immunotoxicity. Front. Immunol. 2023, 14. [Google Scholar] [CrossRef]
- Cui, Y.; Wu, Y.; Shi, P.; Ni, Y.; Zeng, H.; Zhang, Z.; Zhao, C.; Sun, W.; Yi, Q. Mitigating Microplastic-Induced Organ Damage: Mechanistic Insights from the Microplastic-Macrophage Axes. Redox Biol. 2025, 84, 103688. [Google Scholar] [CrossRef]
- Yin, K.; Wang, D.; Zhang, Y.; Lu, H.; Hou, L.; Guo, T.; Zhao, H.; Xing, M. Polystyrene Microplastics Promote Liver Inflammation by Inducing the Formation of Macrophages Extracellular Traps. J. Hazard. Mater. 2023, 452, 131236. [Google Scholar] [CrossRef]
- Jiang, W.; Liu, Y.; Wu, Y.; Zhang, L.; Zhang, B.; Zhou, S.; Zhang, P.; Xu, T.; Wu, M.; Lv, S. Polystyrene Nanoplastics of Different Particle Sizes Regulate the Polarization of Pro-Inflammatory Macrophages. Sci. Rep. 2024, 14, 16329. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, M.G.; Casati, L.; Sauro, G.; Taurino, G.; Griffini, E.; Milani, C.; Ventura, M.; Bussolati, O.; Chiu, M. Biological Effects of Micro-/Nano-Plastics in Macrophages. Nanomaterials 2025, 15, 394. [Google Scholar] [CrossRef] [PubMed]
- Collin-Faure, V.; Vitipon, M.; Torres, A.; Tanyeres, O.; Dalzon, B.; Rabilloud, T. The Internal Dose Makes the Poison: Higher Internalization of Polystyrene Particles Induce Increased Perturbation of Macrophages. Front. Immunol. 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- van den Berg, A.E.T.; Adriaans, K.J.; Parker, L.A.; Höppener, E.M.; Dusza, H.M.; Legler, J.; Pieters, R.H.H. Top-down Generated Micro- and Nanoplastics Reduce Macrophage Viability without Eliciting a pro-Inflammatory Response. Microplastics Nanoplastics 2025, 5, 32. [Google Scholar] [CrossRef]
- Li, S.; Liu, L.; Luo, G.; Yuan, Y.; Hu, D.; Xiao, F. The Crosstalk between M1 Macrophage Polarization and Energy Metabolism Disorder Contributes to Polystyrene Nanoplastics-Triggered Testicular Inflammation. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2023, 180, 114002. [Google Scholar] [CrossRef]
- Chen, J.; Chen, X.; Xuan, Y.; Shen, H.; Tang, Y.; Zhang, T.; Xu, J. Surface Functionalization-Dependent Inflammatory Potential of Polystyrene Nanoplastics through the Activation of MAPK/ NF-κB Signaling Pathways in Macrophage Raw 264.7. Ecotoxicol. Environ. Saf. 2023, 251, 114520. [Google Scholar] [CrossRef]
- Skaba, D.; Fiegler-Rudol, J.; Dembicka-Mączka, D.; Wiench, R. Nanoplastics and Immune Disruption: A Systematic Review of Exposure Routes, Mechanisms, and Health Implications. Int. J. Mol. Sci. 2025, 26, 5228. [Google Scholar] [CrossRef]
- van den Berg, A.E.T.; Plantinga, M.; Vethaak, D.; Adriaans, K.J.; Bol-Schoenmakers, M.; Legler, J.; Smit, J.J.; Pieters, R.H.H. Environmentally Weathered Polystyrene Particles Induce Phenotypical and Functional Maturation of Human Monocyte-Derived Dendritic Cells. J. Immunotoxicol. 2022, 19, 125–133. [Google Scholar] [CrossRef]
- Weber, A.; Schwiebs, A.; Solhaug, H.; Stenvik, J.; Nilsen, A.M.; Wagner, M.; Relja, B.; Radeke, H.H. Nanoplastics Affect the Inflammatory Cytokine Release by Primary Human Monocytes and Dendritic Cells. Environ. Int. 2022, 163, 107173. [Google Scholar] [CrossRef] [PubMed]
- Lopez, G.L.; Lamarre, A. The Impact of Micro- and Nanoplastics on Immune System Development and Functions: Current Knowledge and Future Directions. Reprod. Toxicol. 2025, 135, 108951. [Google Scholar] [CrossRef] [PubMed]
- Park, K.-M.; Kim, B.; Woo, W.; Kim, L.K.; Hyun, Y.-M. Polystyrene Microplastics Induce Activation and Cell Death of Neutrophils through Strong Adherence and Engulfment. J. Hazard. Mater. 2024, 480, 136100. [Google Scholar] [CrossRef]
- Zhu, X.; Peng, L.; Song, E.; Song, Y. Polystyrene Nanoplastics Induce Neutrophil Extracellular Traps in Mice Neutrophils. Chem. Res. Toxicol. 2022, 35, 378–382. [Google Scholar] [CrossRef] [PubMed]
- Chi, Q.; Xu, T.; He, Y.; Li, Z.; Tang, X.; Fan, X.; Li, S. Polystyrene Nanoparticle Exposure Supports ROS-NLRP3 Axis-Dependent DNA-NET to Promote Liver Inflammation. J. Hazard. Mater. 2022, 439, 129502. [Google Scholar] [CrossRef]
- Ma, S.; Xiao, Y.; Zhang, X.; Xu, Y.; Zhu, K.; Zhang, K.; Li, X.; Zhou, H.; Chen, G.; Guo, X. Dietary Exposure to Polystyrene Microplastics Exacerbates Liver Damage in Fulminant Hepatic Failure via ROS Production and Neutrophil Extracellular Trap Formation. Sci. Total Environ. 2024, 907, 167403. [Google Scholar] [CrossRef]
- Saijo, Y.; Hasegawa, Y.; Okamura, T.; Ono, Y.; Ichikawa, T.; Nakanishi, N.; Tsuchimura, Y.; Morioka, T.; Tanaka, S.; Takano, H.; et al. Negative Impact of Oral Exposure to Polystyrene Microplastics on Glucose Tolerance and Intestinal Environment in Mice Is Independent of Particle Size. Environ. Sci. Eur. 2025, 37, 125. [Google Scholar] [CrossRef]
- Ali, N.; Katsouli, J.; Marczylo, E.L.; Gant, T.W.; Wright, S.; Serna, J.B. de la The Potential Impacts of Micro-and-Nano Plastics on Various Organ Systems in Humans. eBioMedicine 2024, 99. [Google Scholar] [CrossRef]
- Woo, J.-H.; Seo, H.J.; Lee, J.-Y.; Lee, I.; Jeon, K.; Kim, B.; Lee, K. Polypropylene Nanoplastic Exposure Leads to Lung Inflammation through P38-Mediated NF-κB Pathway Due to Mitochondrial Damage. Part. Fibre Toxicol. 2023, 20, 2. [Google Scholar] [CrossRef]
- Trushina, E.N.; Riger, N.A.; Mustafina, O.K.; Timonin, A.N. [Effect of microand nanoplastics as food contaminants on the immune system]. Vopr. Pitan. 2023, 92, 6–15. [Google Scholar] [CrossRef]
- Cheng, Y.; Yang, Y.; Bai, L.; Cui, J. Microplastics: An Often-Overlooked Issue in the Transition from Chronic Inflammation to Cancer. J. Transl. Med. 2024, 22, 959. [Google Scholar] [CrossRef]
- Yang, W.; Jannatun, N.; Zeng, Y.; Liu, T.; Zhang, G.; Chen, C.; Li, Y. Impacts of Microplastics on Immunity. Front. Toxicol. 2022, 4, 956885. [Google Scholar] [CrossRef]
- Shang, Q.; Wu, H.; Wang, K.; Zhang, M.; Dou, Y.; Jiang, X.; Zhao, Y.; Zhao, H.; Chen, Z.-J.; Wang, J.; et al. Exposure to Polystyrene Microplastics during Lactational Period Alters Immune Status in Both Male Mice and Their Offspring. Sci. Total Environ. 2024, 951, 175371. [Google Scholar] [CrossRef]
- Dan, K.-B.; Yoo, J.Y.; Min, H. The Emerging Threat of Micro- and Nanoplastics on the Maturation and Activity of Immune Cells. Biomol. Ther. 2025, 33, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Mahmud, F.; Sarker, D.B.; Jocelyn, J.A.; Sang, Q.-X.A. Molecular and Cellular Effects of Microplastics and Nanoplastics: Focus on Inflammation and Senescence. Cells 2024, 13, 1788. [Google Scholar] [CrossRef] [PubMed]
- Zwollo, P.; Quddos, F.; Bagdassarian, C.; Seeley, M.E.; Hale, R.C.; Abderhalden, L. Polystyrene Microplastics Reduce Abundance of Developing B Cells in Rainbow Trout (Oncorhynchus Mykiss) Primary Cultures. Fish Shellfish Immunol. 2021, 114, 102–111. [Google Scholar] [CrossRef]
- Catalán, D.; Mansilla, M.A.; Ferrier, A.; Soto, L.; Oleinika, K.; Aguillón, J.C.; Aravena, O. Immunosuppressive Mechanisms of Regulatory B Cells. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef]
- Slamanig, S.A.; Nolte, M.A. The Bone Marrow as Sanctuary for Plasma Cells and Memory T-Cells: Implications for Adaptive Immunity and Vaccinology. Cells 2021, 10, 1508. [Google Scholar] [CrossRef]
- Jiang, L.; Ye, Y.; Han, Y.; Wang, Q.; Lu, H.; Li, J.; Qian, W.; Zeng, X.; Zhang, Z.; Zhao, Y.; et al. Microplastics Dampen the Self-Renewal of Hematopoietic Stem Cells by Disrupting the Gut Microbiota-Hypoxanthine-Wnt Axis. Cell Discov. 2024, 10, 35. [Google Scholar] [CrossRef] [PubMed]
- Jing, J.; Zhang, L.; Han, L.; Wang, J.; Zhang, W.; Liu, Z.; Gao, A. Polystyrene Micro-/Nanoplastics Induced Hematopoietic Damages via the Crosstalk of Gut Microbiota, Metabolites, and Cytokines. Environ. Int. 2022, 161, 107131. [Google Scholar] [CrossRef]
- Zhang, L.; Jing, J.; Han, L.; Liu, Z.; Wang, J.; Zhang, W.; Gao, A. Melatonin and Probiotics Ameliorate Nanoplastics-Induced Hematopoietic Injury by Modulating the Gut Microbiota-Metabolism. Nano Res. 2023, 16, 2885–2894. [Google Scholar] [CrossRef]
- Lee, S.E.; Yi, Y.; Moon, S.; Yoon, H.; Park, Y.S. Impact of Micro- and Nanoplastics on Mitochondria. Metabolites 2022, 12, 897. [Google Scholar] [CrossRef]
- Yöntem, F.D.; Ahbab, M.A. Mitochondria as a Target of Micro- and Nanoplastic Toxicity. Camb. Prisms Plast. 2024, 2, e6. [Google Scholar] [CrossRef]
- Kadac-Czapska, K.; Ośko, J.; Knez, E.; Grembecka, M. Microplastics and Oxidative Stress—Current Problems and Prospects. Antioxidants 2024, 13, 579. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Liu, R.; Guo, S.; Li, S.; Huang, Z.; Wang, Y.; Yu, C.; Hou, Z.; Zhang, Y.; Zhang, Y.; et al. Large-Sized Polystyrene Microplastics Induce Oxidative Stress in AML12 Cells. Sci. Rep. 2025, 15, 26616. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, Y.; Zhang, J.; Feng, G.; Miao, S.; Lu, R.; Tian, X.; Ye, Y. Antioxidant Intervention Against Microplastic Hazards. Antioxidants 2025, 14, 797. [Google Scholar] [CrossRef]
- Shi, H.; Wang, Y.; Li, X.; Wang, X.; Qi, Y.; Hu, S.; Liu, R. Polystyrene Nanoplastics Elicit Multiple Responses in Immune Cells of the Eisenia Fetida (Savigny, 1826). Toxics 2025, 13, 18. [Google Scholar] [CrossRef]
- Chakraborty, S.; Banerjee, M.; Jayaraman, G.; Rajeswari V, D. Evaluation of the Health Impacts and Deregulation of Signaling Pathways in Humans Induced by Microplastics. Chemosphere 2024, 369, 143881. [Google Scholar] [CrossRef]
- Krause, S.; Ouellet, V.; Allen, D.; Allen, S.; Moss, K.; Nel, H.A.; Manaseki-Holland, S.; Lynch, I. The Potential of Micro- and Nanoplastics to Exacerbate the Health Impacts and Global Burden of Non-Communicable Diseases. Cell Rep. Med. 2024, 5, 101581. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, S.; Liu, S.; Wang, Z.; Li, F.; Bu, Q.; An, X. Polystyrene Microplastics Induce Potential Toxicity through the Gut-Mammary Axis. Npj Sci. Food 2025, 9, 139. [Google Scholar] [CrossRef]
- Lu, H.; Guo, T.; Zhang, Y.; Liu, D.; Hou, L.; Ma, C.; Xing, M. Endoplasmic Reticulum Stress-Induced NLRP3 Inflammasome Activation as a Novel Mechanism of Polystyrene Microplastics (PS-MPs)-Induced Pulmonary Inflammation in Chickens. J. Zhejiang Univ. Sci. B 2024, 25, 233–243. [Google Scholar] [CrossRef]
- Busch, M.; Bredeck, G.; Waag, F.; Rahimi, K.; Ramachandran, H.; Bessel, T.; Barcikowski, S.; Herrmann, A.; Rossi, A.; Schins, R.P.F. Assessing the NLRP3 Inflammasome Activating Potential of a Large Panel of Micro- and Nanoplastics in THP-1 Cells. Biomolecules 2022, 12, 1095. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Xu, R.; Wang, F.; Geng, Y.; Xu, T.; Zhu, M.; Lv, H.; Xu, S.; Guo, M. Polyethylene Microplastics Trigger Cell Apoptosis and Inflammation via Inducing Oxidative Stress and Activation of the NLRP3 Inflammasome in Carp Gills. Fish Shellfish Immunol. 2023, 132, 108470. [Google Scholar] [CrossRef]
- Wan, S.; Wang, X.; Chen, W.; Wang, M.; Zhao, J.; Xu, Z.; Wang, R.; Mi, C.; Zheng, Z.; Zhang, H. Exposure to High Dose of Polystyrene Nanoplastics Causes Trophoblast Cell Apoptosis and Induces Miscarriage. Part. Fibre Toxicol. 2024, 21, 13. [Google Scholar] [CrossRef] [PubMed]
- Pei, J.; Chen, S.; Li, L.; Wang, K.; Pang, A.; Niu, M.; Peng, X.; Li, N.; Wu, H.; Nie, P. Impact of Polystyrene Nanoplastics on Apoptosis and Inflammation in Zebrafish Larvae: Insights from Reactive Oxygen Species Perspective. Sci. Total Environ. 2024, 948, 174737. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.E.; Kim, D.Y.; Jeong, T.S.; Park, Y.S. Micro- and Nano-Plastic-Induced Adverse Health Effects on Lungs and Kidneys Linked to Oxidative Stress and Inflammation. Life 2025, 15, 392. [Google Scholar] [CrossRef]
- Cheng, Y.; Chen, J.; Fu, R.; Zhang, P.; Chen, H.; Cao, H.; Jiang, Z.; Hong, Y.; Li, Y.; He, C.; et al. Molecular Mechanism Differences between Nanoplastics and Microplastics in Colon Toxicity: Nanoplastics Induce Ferroptosis-Mediated Immunogenic Cell Death, While Microplastics Cause Cell Metabolic Reprogramming. J. Nanobiotechnology 2025, 23, 505. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.; Liu, M.; Xiong, F.; Xu, K.; Huang, J.; Liu, J.; Wang, D.; Pu, Y. Polystyrene Micro- and Nanoplastics Induce Gastric Toxicity through ROS Mediated Oxidative Stress and P62/Keap1/Nrf2 Pathway. Sci. Total Environ. 2024, 912, 169228. [Google Scholar] [CrossRef]
- Liu, Y.; Li, X.; Xiong, Y. Chronic Polystyrene Microplastic Exposure Reduces Testosterone Levels in Mice through Mitochondrial Oxidative Stress and BAX/BCL2-Mediated Apoptosis. Toxics 2024, 12, 561. [Google Scholar] [CrossRef]
- Santos, D.; Luzio, A.; Félix, L.; Cabecinha, E.; Bellas, J.; Monteiro, S.M. Microplastics and Copper Induce Apoptosis, Alter Neurocircuits, and Cause Behavioral Changes in Zebrafish (Danio Rerio) Brain. Ecotoxicol. Environ. Saf. 2022, 242, 113926. [Google Scholar] [CrossRef]
- Wang, S.; Wu, H.; Shi, X.; Wang, Y.; Xu, S. Polystyrene Microplastics with Different Sizes Induce the Apoptosis and Necroptosis in Liver through the PTEN/PI3K/AKT/Autophagy Axis. Sci. Total Environ. 2023, 899, 165461. [Google Scholar] [CrossRef]
- Annangi, B.; Villacorta, A.; López-Mesas, M.; Fuentes-Cebrian, V.; Marcos, R.; Hernández, A. Hazard Assessment of Polystyrene Nanoplastics in Primary Human Nasal Epithelial Cells, Focusing on the Autophagic Effects. Biomolecules 2023, 13, 220. [Google Scholar] [CrossRef]
- Ma, Y.; Yu, J.; Sun, J.; Zhu, Y.; Li, X.; Liu, X.; Zhang, X.; Liu, L.; Li, L.; Yang, J.; et al. Dust Fall Microplastics from a Megacity of China Inhibit Autophagy via the PI3K/Akt/mTOR Pathway. Environ. Health 2025, 3, 469–481. [Google Scholar] [CrossRef]
- Alqahtani, S.; Alqahtani, S.; Saquib, Q.; Mohiddin, F. Toxicological Impact of Microplastics and Nanoplastics on Humans: Understanding the Mechanistic Aspect of the Interaction. Front. Toxicol. 2023, 5. [Google Scholar] [CrossRef] [PubMed]
- Florance, I.; Cordani, M.; Pashootan, P.; Moosavi, M.A.; Zarrabi, A.; Chandrasekaran, N. The Impact of Nanomaterials on Autophagy across Health and Disease Conditions. Cell. Mol. Life Sci. 2024, 81, 184. [Google Scholar] [CrossRef]
- Liu, H.; Li, H.; Chen, T.; Yu, F.; Lin, Q.; Zhao, H.; Jin, L.; Peng, R. Research Progress on Micro(Nano)Plastic-Induced Programmed Cell Death Associated with Disease Risks. Toxics 2024, 12, 493. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Zeng, Y.; Zhang, P.; Zhu, B.; Feng, J.; Deng, T.; Fu, Z.; Liu, C.; Chen, C.; Zhang, Y. Polystyrene Nanoplastics Trigger Pyroptosis in Dopaminergic Neurons through TSC2/TFEB-Mediated Disruption of Autophagosome-Lysosome Fusion in Parkinson’s Disease. J. Transl. Med. 2025, 23, 631. [Google Scholar] [CrossRef]
- Liang, B.; Zhong, Y.; Huang, Y.; Lin, X.; Liu, J.; Lin, L.; Hu, M.; Jiang, J.; Dai, M.; Wang, B.; et al. Underestimated Health Risks: Polystyrene Micro- and Nanoplastics Jointly Induce Intestinal Barrier Dysfunction by ROS-Mediated Epithelial Cell Apoptosis. Part. Fibre Toxicol. 2021, 18, 20. [Google Scholar] [CrossRef]
- Vanetti, C.; Broggiato, M.; Pezzana, S.; Clerici, M.; Fenizia, C. Effects of Microplastics on the Immune System: How Much Should We Worry? Immunol. Lett. 2025, 272, 106976. [Google Scholar] [CrossRef]
- Lihua, C.; Zhiyin, T. Microplastics Aggravates Rheumatoid Arthritis by Affecting the Proliferation/Migration/Inflammation of Fibroblast-like Synovial Cells by Regulating Mitochondrial Homeostasis. Int. Immunopharmacol. 2023, 120, 110268. [Google Scholar] [CrossRef]
- Zeyneloglu, C.; Babayev, H.; Ogulur, I.; Ardicli, S.; Pat, Y.; Yazici, D.; Zhao, B.; Chang, L.; Liu, X.; D’Avino, P.; et al. The Epithelial Barrier Theory Proposes a Comprehensive Explanation for the Origins of Allergic and Other Chronic Noncommunicable Diseases. FEBS Lett. 2025. [Google Scholar] [CrossRef]
- Chen, H.; Wan, L.; Qiu, Y.; Qiu, F.; Wen, C.; Mao, Y.; He, Z. Microplastics Exposure Induced and Exacerbated the Development of Systemic Lupus Erythematosus in Mice. Sci. Total Environ. 2024, 909, 168586. [Google Scholar] [CrossRef]
- Yan, Z.; Liu, Y.; Zhang, T.; Zhang, F.; Ren, H.; Zhang, Y. Analysis of Microplastics in Human Feces Reveals a Correlation between Fecal Microplastics and Inflammatory Bowel Disease Status. Environ. Sci. Technol. 2022, 56, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Zolotova, N.; Dzhalilova, D.; Tsvetkov, I.; Makarova, O. Influence of Microplastics on Morphological Manifestations of Experimental Acute Colitis. Toxics 2023, 11, 730. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Williams, A.M.; Gordon, E.B.; Rudolph, S.E.; Longo, B.N.; Li, G.; Kaplan, D.L. Biological Effects of Polystyrene Micro- and Nano-Plastics on Human Intestinal Organoid-Derived Epithelial Tissue Models without and with M Cells. Nanomedicine Nanotechnol. Biol. Med. 2023, 50, 102680. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; He, S.; Zhao, Y.; Jiang, M.; Wang, W.; Yang, L.; Du, W.; Zhuang, W. Microplastics Exposure Aggravates Synovitis and Pyroptosis in SLE by Activating NF-κB and NRF2/KEAP1 Signaling. Toxics 2024, 12, 840. [Google Scholar] [CrossRef]
- Kumar, N.; Pachar, A.K.; Singh, J.K.; Mandal, M.; Lamba, M.; Yadav, S.; Sharda, N.; Acharya, A. Investigating the Hypothesis Role of the Hidden Poison Microplastics in Lymphoma Development. Discov. Environ. 2025, 3, 73. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, F.; Xu, S.; Cui, J.; Li, K.; Shiwen, X.; Guo, M.-Y. Polystyrene Microplastics Induce Myocardial Inflammation and Cell Death via the TLR4/NF-κB Pathway in Carp. Fish Shellfish Immunol. 2023, 135, 108690. [Google Scholar] [CrossRef] [PubMed]
- Winiarska, E.; Jutel, M.; Zemelka-Wiacek, M. The Potential Impact of Nano- and Microplastics on Human Health: Understanding Human Health Risks. Environ. Res. 2024, 251, 118535. [Google Scholar] [CrossRef]
- Seewoo, B.J.; Goodes, L.M.; Mofflin, L.; Mulders, Y.R.; Wong, E.V.; Toshniwal, P.; Brunner, M.; Alex, J.; Johnston, B.; Elagali, A.; et al. The Plastic Health Map: A Systematic Evidence Map of Human Health Studies on Plastic-Associated Chemicals. Environ. Int. 2023, 181, 108225. [Google Scholar] [CrossRef]
- Valamontes, A. Impact of Microplastics on Global Public Health: A Systematic Review and Meta-Analysis 2025.
- Roslan, N.S.; Lee, Y.Y.; Ibrahim, Y.S.; Tuan Anuar, S.; Yusof, K.M.K.K.; Lai, L.A.; Brentnall, T. Detection of Microplastics in Human Tissues and Organs: A Scoping Review. J. Glob. Health 14, 04179. [CrossRef]
- Weis, J.S.; Palmquist, K.H. Reality Check: Experimental Studies on Microplastics Lack Realism. Appl. Sci. 2021, 11, 8529. [Google Scholar] [CrossRef]
- Cho, Y.M.; Choi, K.-H. The Current Status of Studies of Human Exposure Assessment of Microplastics and Their Health Effects: A Rapid Systematic Review. Environ. Anal. Health Toxicol. 2021, 36, e2021004. [Google Scholar] [CrossRef]
- US EPA, O. Microplastics Research. Available online: https://www.epa.gov/water-research/microplastics-research (accessed on 14 September 2025).
- Prata, J.C.; Padrão, J.; Khan, M.T.; Walker, T.R. Do’s and Don’ts of Microplastic Research: A Comprehensive Guide. Water Emerg. Contam. Nanoplastics 2024, 3, N. [Google Scholar] [CrossRef]
- Boettcher, H.; Kukulka, T.; Cohen, J.H. Methods for Controlled Preparation and Dosing of Microplastic Fragments in Bioassays. Sci. Rep. 2023, 13, 5195. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, Z.; Hsueh, Y.; Zhang, C.; Yu, J.; Zhu, J.; Niu, J.; Yin, N.; Zhang, J.; Cui, X.; et al. Interactions between Environmental Pollutants and Gut Microbiota: A Review Connecting the Conventional Heavy Metals and the Emerging Microplastics. Environ. Res. 2025, 269, 120928. [Google Scholar] [CrossRef] [PubMed]
- Demarquoy, J. Microplastics and Microbiota: Unraveling the Hidden Environmental Challenge. World J. Gastroenterol. 2024, 30, 2191–2194. [Google Scholar] [CrossRef]
- Tamargo, A.; Molinero, N.; Reinosa, J.J.; Alcolea-Rodriguez, V.; Portela, R.; Bañares, M.A.; Fernández, J.F.; Moreno-Arribas, M.V. PET Microplastics Affect Human Gut Microbiota Communities during Simulated Gastrointestinal Digestion, First Evidence of Plausible Polymer Biodegradation during Human Digestion. Sci. Rep. 2022, 12, 528. [Google Scholar] [CrossRef]
- Procopio, A.C.; Soggiu, A.; Urbani, A.; Roncada, P. Interactions between Microplastics and Microbiota in a One Health Perspective. One Health 2025, 20, 101002. [Google Scholar] [CrossRef] [PubMed]
- Prata, J.C. Microplastics in Wastewater: State of the Knowledge on Sources, Fate and Solutions. Mar. Pollut. Bull. 2018, 129, 262–265. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Zhang, Y.; Lemos, B.; Ren, H. Tissue Accumulation of Microplastics in Mice and Biomarker Responses Suggest Widespread Health Risks of Exposure. Sci. Rep. 2017, 7, 46687. [Google Scholar] [CrossRef] [PubMed]
- Löder, M.G.J.; Gerdts, G. Methodology Used for the Detection and Identification of Microplastics—A Critical Appraisal. In Marine Anthropogenic Litter; Bergmann, M., Gutow, L., Klages, M., Eds.; Springer International Publishing: Cham, 2015; pp. 201–227. ISBN 978-3-319-16510-3. [Google Scholar]
- Wu, P.; Huang, J.; Zheng, Y.; Yang, Y.; Zhang, Y.; He, F.; Chen, H.; Quan, G.; Yan, J.; Li, T.; et al. Environmental Occurrences, Fate, and Impacts of Microplastics. Ecotoxicol. Environ. Saf. 2019, 184, 109612. [Google Scholar] [CrossRef]
- Bucci, K.; Tulio, M.; Rochman, C.M. What Is Known and Unknown about the Effects of Plastic Pollution: A Meta-Analysis and Systematic Review. Ecol. Appl. 2020, 30, e02044. [Google Scholar] [CrossRef]
- Prata, J.C.; da Costa, J.P.; Lopes, I.; Duarte, A.C.; Rocha-Santos, T. Environmental Exposure to Microplastics: An Overview on Possible Human Health Effects. Sci. Total Environ. 2020, 702, 134455. [Google Scholar] [CrossRef]
- Revel, M.; Châtel, A.; Mouneyrac, C. Micro(Nano)Plastics: A Threat to Human Health? Curr. Opin. Environ. Sci. Health 2018, 1, 17–23. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



