
Submitted:
24 September 2025
Posted:
24 September 2025
You are already at the latest version
Abstract
Neurodegenerative diseases such as Alzheimer’s disease (AD) and amyotrophic lateral sclerosis (ALS) have been associated with long-term effects of concussions, particularly in contact sport athletes. Lipid dysregulation has emerged as a potential contributor to neuronal damage and could serve as an early biomarker of brain pathology. This study aimed to investigate serum lipid biomarkers in retired rugby players with a history of concussion to explore their diagnostic relevance and potential link to neurodegenerative disease risk. A comparative analysis was conducted comparing serum levels of ApoE4, 24-hydroxycholesterol (24-HC), 25-HC, 27-HC, total triglycerides, and ceramide between concussed retired rugby players and non-contact healthy controls. ELISA-based quantification and statistical comparisons were used to identify significant group differences. Retired rugby players exhibited significantly lower serum 24-HC and significantly higher levels of ApoE4, 27-HC, triglycerides, and ceramide compared to controls. No significant difference was found in 25-HC, though some concussed individuals showed elevated levels. These findings align with previous reports in AD, PD, and ALS populations, supporting lipid dysregulation as a hallmark of neurodegenerative progression. Alterations in lipid profiles among concussed athletes suggest potential roles for ApoE4, 24-HC, 27-HC, ceramide, and triglycerides as early biomarkers for concussion-induced neurodegeneration. These findings may guide future diagnostics and post-care strategies for at-risk populations.
Keywords:
lipids
; biomarkers
; concussion
; brain injury
; sport
; neurodegenerative diseases
1. Introduction
Rugby, a well-known contact sport, carries a notably high risk of physical injury—particularly concussions—due to its inherently aggressive nature, which includes frequent and repeated tackles and collisions [1]. Concussions are a form of traumatic brain injury resulting from rapid head movements involving acceleration and deceleration. They often manifest as brief loss of consciousness, impaired reflexes, or amnesia [2]. Typical symptoms include headaches, dizziness, sleep irregularities, balance issues, disorientation, cognitive disturbances, and mood changes [3]. These injuries are particularly concerning because of their association with long-term deficits in cognition, motor abilities, and mental health [4,5,6].
Research has shown that repeated concussions can have cumulative effects, increasing the likelihood of future brain injuries and deteriorating neurological health. Tsushima et al. [7] reported that athletes with a prior concussion were three to five times more likely to sustain another compared to those without such a history. Moreover, experiencing concussions can significantly impair cognitive functions such as memory, executive functioning, and motor coordination. Retired athletes across various sports who have experienced concussions tend to report greater cognitive difficulties [8]. In a study by Kerr et al. [9] involving 204 former collegiate football players (84.3% with a history of concussion), the researchers explored the relationship between previous concussions and negative health outcomes. Their findings revealed associations between concussion history and several adverse effects, including poor general physical and mental health, depression, and increased risk of alcohol dependence.
While physical activity generally promotes overall health, there are instances where the potential risks may surpass the advantages of engaging in exercise. In this regard, there is growing concern about the health and well-being of former contact sport athletes, particularly in relation to the potential long-term effects of concussions and other brain injuries. For example, former professional and collegiate athletes in sports such as soccer and American football have been found to experience significantly higher rates of amyotrophic lateral sclerosis (ALS) [10]. In a large-scale mortality study involving 3,439 National Football League (NFL) players, Lehman et al. [11] reported that deaths due to neurodegenerative diseases—including Alzheimer’s disease and ALS—occurred at a rate nearly three times higher than in the general population. An earlier investigation by Guskiewicz et al. [12] involved administering a general health questionnaire to 2,552 retired professional football players, 61% of whom reported experiencing at least one concussion, and 24% reported three or more during their careers. The study found that players with a greater history of repeated concussions were at increased risk for developing dementia-related conditions. Additional research has consistently shown a strong association between traumatic brain injury (TBI) and elevated risk for cognitive decline and neurodegenerative disorders, including Alzheimer’s disease. Regrettably, most neurodegenerative diseases that lead to dementia remain undiagnosable and untreatable during their early, prodromal stages due to the lack of clinically validated biomarkers [13]. In addition, there is currently a gap in research, as no studies have specifically explored potential concussion-related biomarkers that may be associated with neurodegenerative conditions in retired athletes with a history of sports-related concussions (see the exceptional Alanazi et al., 2024) [14].
The current study aims to investigate lipid biomarkers, originally associated with cardiovascular diseases, as potential indicators of neurodegeneration in a well-defined cohort of retired rugby players with a history of multiple concussions (chronic state), compared to healthy controls. The lipid biomarkers examined include Apolipoprotein E4 (ApoE4), 24S-hydroxycholesterol (24-HC), 25-hydroxycholesterol (25-HC), 27-hydroxycholesterol (27-HC), ceramide, and total triglycerides.
ApoE is a central transporter of extracellular lipids and cholesterol within the brain, facilitating cholesterol exchange between neuronal and glial cells [15] (Posse de Chaves and Narayanaswami, 2008). Under normal physiological conditions, ApoE is primarily synthesized by astrocytes, though its expression can be induced in neurons during stress. It is most abundantly expressed in the midbrain and peripheral liver [16] (Konings et al., 2021). The role of ApoE—particularly the E4 isoform (ApoE4)—in concussion and traumatic brain injury (TBI) has been extensively explored, as recovery outcomes vary significantly among individuals. Notably, the ApoE4 genotype has been linked to poorer long-term outcomes post-TBI and neurodegenerative disease [17,18] (Huang and Mahley, 2014; McFadyen et al., 2021) and an increased risk of developing chronic traumatic encephalopathy (CTE) [19] (Atherton et al., 2022). ApoE4 is also a well-established genetic risk factor for early-onset Alzheimer’s disease (AD) [20] (Fernández-Calle et al., 2022), largely due to its impaired capacity to clear amyloid-beta (Aβ) compared to ApoE2 and ApoE3 isoforms [21] (Castellano et al., 2011). In neurons, ApoE4 is associated with the formation of neurofibrillary tangles (NFTs) and mitochondrial dysfunction, while astrocyte-derived ApoE4 may drive Aβ accumulation [22] (Zhang et al., 2023). These pathological changes promote neuronal injury and neuroinflammation, disrupting memory and learning [22] (Zhang et al., 2023). Astrocytic overexpression of ApoE4 has also been shown to enhance tau phosphorylation and aggregation [23] (Jablonski et al., 2021). Furthermore, ApoE4-related disruptions in cholesterol homeostasis may impair glucose metabolism, further contributing to AD pathology [24,25] (Wu et al., 2018; Sun et al., 2023). Animal studies reveal that ApoE4 delays blood–brain barrier (BBB) repair after injury [26] (Main et al., 2018) and compromises BBB integrity [27] (Bell et al., 2012), consistent with human findings showing increased BBB permeability in older ApoE4 carriers compared to ApoE3 homozygotes, irrespective of cognitive function [28] (Montagne et al., 2020). In military veterans with TBI history, ApoE4 was associated with poorer memory, slower processing speed, broader cognitive deficits [29] (Merritt et al., 2018), and damaging lipid droplets in Alzheimer’s disease microglia [30] (Haney et al., 2024). These findings suggest that post-TBI ApoE4 alterations may underlie molecular vulnerabilities that increase neurodegeneration risk [31] (Muza et al., 2019). Therefore, the present study will evaluate serum ApoE4 levels in retired rugby players with and without a history of concussions to assess its utility as a potential diagnostic biomarker.
Lipids are a diverse group of organic molecules that play essential roles in cellular structure, energy storage, and signaling pathways (Fahy et al., 2009) [32]. Remarkably, the brain contains one of the highest concentrations of lipids in the human body—second only to adipose tissue—with lipids accounting for over 50% of its dry weight (Bruce et al., 2017) [33]. These molecules are fundamental to key neurological functions such as axonal insulation, neurite outgrowth, and synapse formation. Consequently, disruptions in lipid metabolism have been increasingly linked to neurodegenerative processes and brain disorders (Cermenati et al., 2015) [43]. Additionally, brain lipid regulation is closely intertwined with oxidative stress, neuroinflammation, and the maintenance of energy balance. Dysregulated lipid metabolism in neuroglial cells can adversely affect normal neuronal function and signaling (Yang et al., 2022) [35]. Following traumatic brain injury (TBI), measurable alterations in lipid profiles have been detected in peripheral fluids such as blood, plasma, and serum, highlighting their potential as accessible biomarkers (Nessel et al., 2021) [36]. In this context, a deeper understanding of lipid dynamics may not only enhance the diagnosis of concussion-related injuries but also inform early intervention strategies for mitigating long-term neurodegenerative risks.
Oxysterols—oxidized derivatives of cholesterol such as 24-HC, 25-HC, and 27-HC—play pivotal roles in both physiological regulation and pathological progression, particularly in the context of neurodegenerative disorders, including Alzheimer's disease (AD), Parkinson's disease (PD), and amyotrophic lateral sclerosis (ALS) (de Freitas et al., 2021) [37]. These metabolites arise via distinct enzymatic pathways: 24-HC is predominantly synthesized in the brain through the action of cholesterol 24-hydroxylase (CYP46A1) (Sun et al., 2016) [38], 25-HC is produced in macrophages and glial cells via cholesterol 25-hydroxylase (CH25H) in response to Toll-like receptor 4 (TLR-4) activation (Romero et al., 2024) [39], and 27-HC is generated by CYP27A1, which is widely expressed across various human tissues (Marwarha and Ghribi, 2015) [40]. These distinct hydroxylation sites on the cholesterol molecule contribute to their varying cytotoxic effects on neural cells and are believed to play critical roles in the pathogenesis of neurodegeneration (Nakazawa et al., 2017) [41]. Elevated levels of 24-HC in serum have been associated with TBIs and linked to neurodegenerative conditions such as multiple sclerosis and PD. Conversely, reduced 24-HC concentrations may reflect neuronal loss and brain atrophy, as observed in AD and multiple sclerosis (Leoni et al., 2002; [42] Björkhem et al., 2013) [43]. Due to its enhanced ability to cross biological membranes, including the blood-brain barrier, 24-HC holds promise as a serum biomarker for brain injury (Lu et al., 2021) [44]. Likewise, 25-HC has been shown to induce apoptosis in cultured motor neurons, with high levels negatively impacting neuronal viability (Odnoshivkina et al., 2022) [45]. Clinical findings by Kim et al. (2017) [46] revealed significantly elevated 25-HC levels in both serum and cerebrospinal fluid (CSF) of ALS patients compared to controls, implicating its role in disease pathology. Regarding 27-HC, elevated levels have been observed in patients with mild cognitive impairment, suggesting a link to early AD development (Zhang et al., 2019) [47]. However, contrasting evidence from Wuolikainen et al. (2014) [48] showed decreased 27-HC plasma levels in ALS patients relative to controls, indicating complex and disease-specific patterns in oxysterol metabolism.
Ceramide, a bioactive lipid composed of a sphingosine backbone linked to a fatty acid, is a central component of sphingolipid metabolism and plays a critical role in cellular signaling and homeostasis (Mencarelli and Martinez–Martinez, 2013) [49]. The regulation of intracellular ceramide levels is vital, as alterations in ceramide and sphingolipid profiles have been implicated in the pathophysiology of various neurological, neuroinflammatory, and age-associated disorders (Mencarelli & Martinez–Martinez, 2013) [49]. Elevated ceramide concentrations have been linked to mitochondrial dysfunction, impaired autophagic processes, and disrupted amyloid-β clearance, all of which are recognized contributors to the development and progression of Alzheimer’s disease (AD) (Chowdhury et al., 2022 [50]; Custodia et al., 2022 [51]; Tringali and Giussani, 2022) [52]. Notably, Han et al. (2002) [53] observed increased ceramide levels during the early stages of AD, with a subsequent decline as the disease advanced, suggesting a dynamic role in disease progression. Post-mortem analysis by He et al. (2010) [54] further confirmed elevated ceramide concentrations in brain tissues of AD patients compared to controls. Similarly, Fernández-Irigoyen et al. (2021) [55] reported significantly higher ceramide levels in the plasma of patients with Parkinson’s disease (PD) relative to healthy individuals. In preclinical research, Barbacci et al. (2017) [56] utilized mass spectrometry imaging to examine ceramide distribution in a rat model of controlled cortical impact (CCI), revealing elevated ceramide in damaged brain regions. In parallel, Ojo et al. (2019) [57] analyzed phospholipid alterations, including ceramide dysregulation, in mouse models of repetitive mild TBI and AD. Their findings identified both shared and unique lipidomic signatures, highlighting ceramide’s potential role in the transition from TBI to neurodegenerative disease. More recently, Liu et al. (2024) [58] demonstrated that mice with increased serum ceramide exhibited poorer cognitive performance, as evidenced by reduced discrimination indices, suggesting that circulating ceramide may serve as a viable biomarker for cognitive impairment.
Triglycerides are a class of lipids primarily involved in the storage and transport of energy. They originate from two main sources: dietary absorption through the gastrointestinal tract and endogenous synthesis by the liver. Structurally, triglycerides are composed of three fatty acids bound to a glycerol molecule, and their diversity is vast, with an estimated 6,000 distinct forms based on variations in fatty acid composition and configuration (Bernath et al., 2020) [59]. Triglycerides are fundamental constituents of triglyceride-rich lipoproteins, such as chylomicrons and very low-density lipoproteins (VLDL), and also exist in remnant particles that persist following lipoprotein catabolism (Do et al., 2013). The relationship between serum triglyceride levels and cognitive function has been explored in several human studies. Parthasarathy et al. (2017) [60], in a cohort of 251 older adults, reported an inverse correlation between serum triglyceride levels and executive function, although no association was found with memory performance. These findings suggest that elevated triglyceride levels may selectively impair certain cognitive domains. Similarly, Bernath et al. (2020) [59] conducted a cross-sectional analysis involving 689 participants, including both Alzheimer’s disease (AD) patients and healthy controls. Their results indicated that lower triglyceride levels were associated with better cognitive outcomes and reduced brain atrophy, implying a potential protective role. Complementing this, Zhao et al. (2019) [61] assessed serum lipid profiles in a large sample of 1,762 individuals aged 40 to 85. They found that higher triglyceride levels were significantly associated with cognitive decline in middle-aged men (≤55 years), further supporting a link between dyslipidemia and cognitive impairment. Despite these associations, human studies specifically examining serum triglycerides in individuals with a history of concussion or traumatic brain injury (TBI) remain lacking. However, animal models provide preliminary insights. For instance, Kuo et al. (2020) [62] demonstrated that rats subjected to TBI and fed a high-fat diet exhibited elevated triglyceride levels, which correlated with increased injury severity. Additionally, Hahnefeld et al. (2022) [63] reported that triglyceride accumulation contributed to behavioral changes post-TBI, highlighting a potential connection between altered lipid metabolism and neurobehavioral outcomes following brain injury.
Given their association with neuronal injury and degeneration, 24-HC, 25-HC, 27-HC, ceramide, and total triglycerides represent promising biomarker candidates for the early detection of neurodegenerative risk. This is particularly relevant for retired rugby players, who are frequently exposed to repeated head trauma. Early identification of oxysterol and other lipid imbalances in this population could inform timely therapeutic strategies aimed at reducing long-term cognitive and neurological burden.
2. Results
A total random selection of 45 males (with a history of concussions, n=26, and control n=19) participated in this study. Participants’ information is given in Table 1 below.
2.1. Serum Biomarker Levels in Concussed and Control Groups
A series of related serum biomarkers was explored (Figure 1). Concentrations of serum 24-HC were significantly lower in the retired rugby group compared to the healthy control group (* p < 0.01). There was a highly significant elevation of serum ApoE4, ceramide, and total triglycerides concentrations in the serum of retired rugby players who had suffered multiple concussions when compared to the non-contact control group (** p < 0.01). Moreover, there was a significant difference in 27-HC levels between the concussed and control groups (* p < 0.05), while there was no difference between the levels of 25-HC in serum between the two groups.
2.2. Correlations Among Biomarkers
A standard regression Pearson’s correlation analysis was performed between all the markers. Unexpectedly, there were insignificant correlations between 24-HC levels and total 25-HC (R2 = 0.03717, p = 0.41); 24-HC and 27-HC (R2 = 0.1508, p = 0.09); 25-HC and 27-HC (R2 = 0.03090, p = 0.45); 25-HC and ceramide (R2 = 0.06039, p = 0.2963); 27-HC and ceramide (R2 = 0.06558, p = 0.27); 25-HC and triglycerides (R2 = 0.02654, p = 0.49); 27-HC and triglycerides (R2 = 0.04482, p = 0.37); ceramide and triglycerides (R2 = 0.1939, p = 0.052); ApoE4 and 25-HC (R2 = 0.00003048, p = 0.98); ApoE4 and 27-HC (R2 = 0.01551, p = 0.6009);
Interestingly, there was a significant correlation between reduced 24-HC levels and higher total triglycerides (R2 = 0.38, p = 0.003) and between lower 24-HC and elevated ceramide levels (R2 =0.2354, p = 0.03). Moreover, there was a highly significant correlation between 24-HC and ApoE4 (R2 = 0.4902, p = 0.0006). There were also strong positive correlations between ApoE4 and ceramide (R2 = 0.2447, p = 0.0266) and between ApoE4 and total triglycerides (R2 = 0.3693, p = 0.0045) (Figure 2).
3. Discussion
Given their established links to neurodegenerative conditions such as Alzheimer’s disease (AD) and amyotrophic lateral sclerosis (ALS), this paper assesses the concentrations of key lipid biomarkers: ApoE4, 24-HC, 25-HC, 27-HC, triglycerides, and ceramide between retired rugby players with a history of concussion and matched healthy controls. These markers were examined to determine whether measurable differences exist that may serve a diagnostic purpose. This paper highlights that several lipid species may contribute meaningfully to the early detection of concussion-related pathology and the long-term risk of neurodegeneration, with potential implications for revising concussion protocols in high-impact sports such as rugby.
We found a significant increase in serum ApoE4 concentrations in concussed individuals compared to the control group (Figure 1). This finding aligns with previous research, indicating that the presence of the ApoE4 allele is associated with a heightened risk of both neurodegenerative diseases and concussion-related injuries such as traumatic brain injuries [17,64]. (Zhou et al., 2008; Huang & Mahley, 2014). Supporting this, Hellstrøm et al. (2022) [65] reported that individuals carrying the ApoE4 allele exhibited significantly greater verbal memory impairments following TBI compared to non-carriers. Additionally, ApoE4 is believed to contribute to AD pathology through mechanisms involving oxidative stress, mitochondrial dysfunction, and disrupted sleep patterns (Sun et al., 2023) [25]. Yang et al. (2015) [66] also found a notably higher frequency of the ApoE4 allele among patients with mild TBI, and comorbid dementia compared to healthy controls. Similarly, Merritt et al. (2018) [29] observed that ApoE4 was associated with more pronounced cognitive deficits in military veterans with a history of mild TBI than in non-carriers. While these studies demonstrate a clear association between ApoE4 and adverse outcomes in both TBI and AD, limited research has directly assessed ApoE4 serum concentrations in individuals with a history of concussion compared to non-concussed controls. This study addresses that gap, introducing a novel approach that enhances the understanding of ApoE4’s diagnostic potential in the context of sports-related brain injury and neurodegenerative disease.
The findings revealed significantly reduced serum 24-HC concentrations in concussed individuals compared to the control group (Figure 1). This observation corresponds with previous studies in neurological disorders. For instance, lower 24-HC levels have been documented in older patients with multiple sclerosis (MS), Parkinson’s disease (PD), and ALS, likely reflecting neuronal loss or impaired metabolic activity in cholesterol catabolism (Lee et al., 2009; [67] La Marca et al., 2016) [68]. Conversely, studies such as Weiner et al. (2008) [69] did not report a difference in 24-HC following TBI, although their sample involved younger individuals, which may account for this discrepancy. Overall, our findings support the hypothesis that decreased 24-HC may indicate reduced neuronal viability and suggest its potential utility as a biomarker for concussion-induced neural damage.
Regarding 25-HC, no significant group differences were observed; however, several concussed individuals exhibited elevated levels relative to controls (Figure 1). Prior research on ALS patients showed mixed outcomes—Kim et al. (2017) [46] reported increased serum and CSF 25-HC, whereas Wuolikainen et al. (2014) [48] found no significant alterations. The lack of statistical significance in our study may be attributed to the small sample size, the variability of ELISA assays, or undiagnosed head injuries in the control cohort.
Serum levels of 27-HC were found to be elevated in both the concussed and control groups (Figure 1). These findings partially mirror reports of increased 27-HC in CSF of ALS patients (Kim et al., 2017) [46], possibly due to impaired clearance or upregulated synthesis. In contrast, reduced plasma 27-HC was observed by Wuolikainen et al. (2014) [48] in ALS, suggesting divergent peripheral and central regulation mechanisms. This divergence emphasizes the need to distinguish between systemic and CNS-specific oxysterol pathways in neurodegenerative risk.
Total serum triglyceride levels were also significantly elevated in the retired rugby players compared to controls (Figure 1). This novel finding aligns with studies linking triglyceride dysregulation to brain atrophy and reduced cognitive function (Bernath et al., 2020 [59]; Zhao et al., 2019) [61]. Furthermore, elevated triglyceride concentrations have been associated with a heightened risk of dementia and AD (Tian et al., 2023) [70]. Animal studies further substantiate these associations—Kuo et al. (2020) [62] and Hahnefeld et al. (2022) [46] both reported that increased triglycerides exacerbated injury and behavioral deficits post-TBI.
Ceramide levels were notably higher in the concussed group relative to controls (Figure 1), representing another novel contribution. While ceramide has been extensively studied in AD and PD, its association with sports-related concussion has not previously been investigated. Prior work demonstrated elevated ceramide in early AD and PD cases (Han et al., 2002 [53]; He et al., 2010 [54]; Fernández-Irigoyen et al., 2021) [55]. Animal models similarly reveal ceramide accumulation in brain regions impacted by injury (Barbacci et al., 2017 [56]; Ojo et al., 2019) [57]. Most recently, Liu et al. (2024) [58] linked elevated ceramide with cognitive impairment in mice, suggesting serum ceramide as a promising marker of neural dysfunction.
With reference to the correlation between biomarkers, we found a negative correlation between 24-HC and triglycerides. Previous studies discussed the association between 24-HC and triglycerides. For instance, Dunk et al. [71] (2024) found that there is an association between 24-HC and triglycerides, suggesting that plasma oxysterols may be a modifiable risk factor for dementia. Lin et al (2018) [72] observed that the plasma 24-HC was correlated with total plasma cholesterol (R = 0.659, p= 0.001), affecting the risk of age-related macular degeneration (AMD) development and progression. Though not a direct measure of triglycerides, total cholesterol correlation often implies a broader relationship across lipid fractions, including TG. Furthermore, key circulating oxysterols, including 24-HC, were found to be positively correlated with triglycerides, indicating systemic lipid interactions (Passarelli et al., 2022) [73]. These studies suggest that 24-HC levels are significantly associated with systemic lipid markers, including triglycerides. They reinforce the biological link between brain cholesterol metabolism (via 24-HC) and peripheral lipid status. We also observed a negative correlation between 24-HC and ceramide. Previous research suggested that 24-HC is substantially reduced in the presence of neurodegenerative disease. Because the final effect of a neurodegenerative process is a loss of active neural cells, a reduction in 24-hydroxylase activity, with subsequent decline in the formation of 24-HC and its lower efflux from the brain into circulation, is likely to be the outcome in these disorders (Leoni et al., 2013) [74]. Prior studies also denoted that higher levels of ceramide are associated with increased AD pathology and inflammation (Teitsdottir et al., 2021) [75]. Currently, no studies have reported direct correlations between 24-HC and ceramide levels in neurological patients or within the context of concussion/TBI. This highlights the novelty of the present study, which explores lipid biomarkers and their interrelationships as potential tools for diagnosing concussions and predicting neurodegenerative disease. We also observed a negative correlation between ApoE4 and 24-HC. This finding is inconsistent with a study on AD patients that reported no effect of ApoE4 positivity on serum 24-HC levels in AD subjects (Roy et al., 2019 [76]. Furthermore, there were also strong positive correlations between ApoE4 and ceramide and between ApoE4 and total triglycerides. In brain tissues from AD patients, especially those carrying the ApoE4 allele (Bandaru et al. 2009) [77] observed significantly elevated ceramide levels were observed compared to ApoE3 carriers, along with disrupted sphingomyelin metabolism. Similarly,
Taken together, these results suggest that lipids—particularly 24-HC, 27-HC, triglycerides, and ceramide—warrant further investigation as potential biomarkers of concussion-related neurodegeneration. This study represents one of the first attempts to explore lipidomics in the context of repeated head trauma in contact sport athletes and contributes valuable evidence for refining diagnostic strategies and long-term monitoring.
4. Materials and Methods
4.1. Study Design and Setting
The current research analyzed a range of selected biomarkers in retired rugby players with a history of concussion and non-contact sports controls from the UK Rugby Health Project [6]. The UK Rugby Health Project was initiated in 2016 as an extension to the inaugural New Zealand Rugby Health Project [78], and the methods and results so far have been published elsewhere [1,6,78,79].
4.2. Study Participants
Former male rugby players and non-contact sport athletes took part in the study and were recruited from September 2016 to December 2018 using past player/athlete associations, printed and televised media reports, word of mouth, and social media. Participants in the current study were those who had attended an in-person clinical appointment and provided a fasted blood sample. The group of concussed in this study were retired rugby players who reported 5 or more diagnosed concussions during their sporting career (n = 26), and the control group was both retired rugby players and retired non-contact sports participants with no reported concussions (n = 19). The number of replicate analyses reflected the limited availability of sample volumes for randomly selected age-matched groups of participants, which reflects a limitation of this present study. No significant differences were seen in participant ages or mean years at and since retirement from sport (p > 0.05). The severity of the individual concussions was not monitored, and only the group with over 5 concussions was explored in this initial study to explore the extremes in the first instance. Further studies will follow to meet these limitations.
4.3. Biomarker Assays
4.4. ELISA Assays
Assays performed using serum followed manufacturer instructions for 24-HC (abcam, catalog no: ab204530), 25-HC (BT lab, catalog no: EA0143Hu), 27-HC (abbexa, catalog no: abx257403), ceramide (AFG Scientific, catalog no: EK710698), and total triglycerides (abbexa, catalog no: abx257659).
4.5. Statistical Analyses
Data were processed in Microsoft Excel 2023 and GraphPad Prism software version 10 was used for all statistical analyses, including the calculation of serum and exosome concentrations, means, standard deviations (SD), coefficient of determination (R2), and p values (where * significant p < 0.05; ** highly significant p < 0.01; n.s. denotes non-significant). The test carried out was a Mann–Whitney U test. All quantitative data are expressed as mean values ± SD of the mean for individual cases.
5. Conclusions
This study highlights the potential of lipid biomarkers, particularly 24-HC, 27-HC, ceramide, and triglycerides, as indicators of concussion-related neurodegenerative risk in retired rugby players. The observed alterations in lipid levels mirror patterns reported in neurodegenerative diseases such as AD, PD, and ALS, reinforcing the biological relevance of these markers. While 25-HC showed no significant difference, its elevation in some concussed individuals warrants further exploration. These findings support the use of lipid profiling as a non-invasive approach to detect early neurodegenerative changes, offering valuable insight for clinical monitoring, early intervention, and refining concussion management protocols in high-impact sports.
6. Patents
There are no patents resulting from the work reported in this manuscript.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, P.C., K.H.; methodology, N.A,.; validation, P.C. formal analysis, N.A,P.C.; investigation, N.A., T.R., I.E..; resources, P.C., K.H..; data curation, N.A., P.C.; writing—original draft preparation, N.A..; writing—review and editing, P.C., K.H., I.E..; supervision, P.C.,K.H.; project administration, P.C., K.H., I.E..; funding acquisition, P.C.,K.H. All authors have read and agreed to the published version of the manuscript.” Please turn to the CRediT taxonomy for the term explanation. Authorship must be limited to those who have contributed substantially to the work reported.
Funding
Funding was provided by the Auckland University of Technology, Durham University (UK), and the Saudi Arabian Scholarship fund for PhD studentship (NA).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Data supporting reported results generated during the study can be found in the paper.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Hind, K.; Konerth, N.; Entwistle, I.; Theadom, A.; Lewis, G.; King, D.; Chazot, P.; Hume, P. Cumulative sport-related injuries and longer term impact in retired male Elite-and Amateur-Level rugby code athletes and non-contact athletes: A retrospective study. Sports Med. 2020, 50, 2051–2061. [Google Scholar] [CrossRef] [PubMed]
- Shaw, N.A. The neurophysiology of concussion. Prog. Neurobiol. 2002, 67, 281–344. [Google Scholar] [CrossRef] [PubMed]
- Stillman, A.; Alexander, M.; Mannix, R.; Madigan, N.; Pascual-Leone, A.; Meehan, W.P. Concussion: Evaluation and management. Cleve Clin. J. Med. 2017, 84, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Moretti, L.; Cristofori, I.; Weaver, S.M.; Chau, A.; Portelli, J.N.; Grafman, J. Cognitive decline in older adults with a history of traumatic brain injury. Lancet Neurol. 2012, 11, 1103–1112. [Google Scholar] [CrossRef]
- Martini, D.N.; Broglio, S.P. Long-term effects of sport concussion on cognitive and motor performance: A review. Int. J. Psychophysiol. 2018, 132, 25–30. [Google Scholar] [CrossRef]
- Hind, K.; Konerth, N.; Entwistle, I.; Hume, P.; Theadom, A.; Lewis, G.; King, D.; Goodbourn, T.; Bottiglieri, M.; Ferraces-Riegas, P.; et al. Mental health and wellbeing of retired elite and amateur rugby players and non-contact athletes and associations with sports-related concussion: The UK Rugby Health Project. Sports Med. 2022, 52, 1419–1431. [Google Scholar] [CrossRef]
- Tsushima, W.T.; Siu, A.M.; Ahn, H.J.; Chang, B.L.; Murata, N.M. Incidence and risk of concussions in youth athletes: Comparisons of age, sex, concussion history, sport, and football position. Arch. Clin. Neuropsychol. 2019, 34, 60–69. [Google Scholar] [CrossRef]
- Cunningham, J.; Broglio, S.P.; O′Grady, M.; Wilson, F. History of sport-related concussion and long-term clinical cognitive health outcomes in retired athletes: A systematic review. J. Athl. Train. 2020, 55, 132–158. [Google Scholar] [CrossRef]
- Kerr, Z.Y.; Thomas, L.C.; Simon, J.E.; McCrea, M.; Guskiewicz, K.M. Association between history of multiple concussions and health outcomes among former college football players: 15-year follow-up from the NCAA concussion study (1999–2001). Am. J. Sports Med. 2018, 46, 1733–1741. [Google Scholar] [CrossRef]
- Lacorte, E.; Ferrigno, L.; Leoncini, E.; Corbo, M.; Boccia, S.; Vanacore, N. Physical activity, and physical activity related to sports, leisure and occupational activity as risk factors for ALS: A systematic review. Neurosci. Biobehav. Rev. 2016, 66, 61–79. [Google Scholar] [CrossRef]
- Lehman, E.J.; Hein, M.J.; Baron, S.L.; Gersic, C.M. Neurodegenerative causes of death among retired National Football League players. Neurology 2012, 79, 1970–1974. [Google Scholar] [CrossRef] [PubMed]
- Guskiewicz, K.M.; Marshall, S.W.; Bailes, J.; McCrea, M.; Cantu, R.C.; Randolph, C.; Jordan, B.D. Association between recurrent concussion and late-life cognitive impairment in retired professional football players. Neurosurgery 2005, 57, 719–726. [Google Scholar] [CrossRef] [PubMed]
- Halicki, M.J.; Hind, K.; Chazot, P.L. Blood-based biomarkers in the diagnosis of Chronic Traumatic Encephalopathy: Research to date and future directions. Int. J. Mol. Sci. 2023, 24, 12556. [Google Scholar] [CrossRef] [PubMed]
- Alanazi, N.; Fitzgerald, M.; Hume, P.; Hellewell, S.; Horncastle, A.; Anyaegbu, C.; Papini, M.G.; Hargreaves, N.; Halicki, M.; Entwistle, I.; Hind, K.; Chazot, P. Concussion-related biomarker variations in retired rugby players and implications for neurodegenerative disease risk: the UK Rugby Health Study. International journal of molecular sciences 2024, 25, 7811. [Google Scholar] [CrossRef] [PubMed]
- Posse de Chaves, E.; Narayanaswami, V. Apolipoprotein E and cholesterol in aging and disease in the brain. Future Lipidol 2008, 3, 505–530. [Google Scholar] [CrossRef]
- Konings, S.C.; Torres-Garcia, L.; Martinsson, I.; Gouras, G.K. Astrocytic and neuronal apolipoprotein E isoforms differentially affect neuronal excitability. Frontiers in Neuroscience 2021, 15, 734001. [Google Scholar] [CrossRef]
- Huang, Y.; Mahley, R.W. Apolipoprotein E: structure and function in lipid metabolism, neurobiology, and Alzheimer's diseases. Neurobiology of disease 2014, 72, 3–12. [Google Scholar] [CrossRef]
- McFadyen, C.A.; Zeiler, F.A.; Newcombe, V.; Synnot, A.; Steyerberg, E.; Gruen, R.L.; Rosand, J.; Palotie, A.; Maas, A.I.; Menon, D.K. Apolipoprotein E4 polymorphism and outcomes from traumatic brain injury: a living systematic review and meta-analysis. Journal of neurotrauma 2021, 38, 1124–1136. [Google Scholar] [CrossRef]
- Atherton, K.; Han, X.; Chung, J.; Cherry, J.D.; Baucom, Z.; Saltiel, N.; Nair, E.; Abdolmohammadi, B.; Uretsky, M.; Khan, M.M.; Shea, C. Association of APOE genotypes and chronic traumatic encephalopathy. JAMA neurology 2022, 79, 787–796. [Google Scholar] [CrossRef]
- Fernández-Calle, R.; Konings, S.C.; Frontiñán-Rubio, J.; García-Revilla, J.; Camprubí-Ferrer, L.; Svensson, M.; Martinson, I.; Boza-Serrano, A.; Venero, J.L.; Nielsen, H.M.; Gouras, G.K. APOE in the bullseye of neurodegenerative diseases: impact of the APOE genotype in Alzheimer’s disease pathology and brain diseases. Molecular neurodegeneration 2022, 17, 62. [Google Scholar] [CrossRef]
- Castellano, J.M.; Kim, J.; Stewart, F.R.; Jiang, H.; DeMattos, R.B.; Patterson, B.W.; Fagan, A.M.; Morris, J.C.; Mawuenyega, K.G.; Cruchaga, C.; Goate, A.M. Human apoE isoforms differentially regulate brain amyloid-β peptide clearance. Science translational medicine 2011, 3, 89ra57. [Google Scholar] [CrossRef]
- Zhang, L.; Xia, Y.; Gui, Y. Neuronal ApoE4 in Alzheimer’s disease and potential therapeutic targets. Frontiers in aging neuroscience 2023, 15, 1199434. [Google Scholar] [CrossRef] [PubMed]
- Jablonski, A.M.; Warren, L.; Usenovic, M.; Zhou, H.; Sugam, J.; Parmentier-Batteur, S.; Voleti, B. Astrocytic expression of the Alzheimer’s disease risk allele, ApoEε4, potentiates neuronal tau pathology in multiple preclinical models. Scientific Reports 2021, 11, 3438. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Zhang, X.; Zhao, L. Human ApoE isoforms differentially modulate brain glucose and ketone body metabolism: implications for Alzheimer's disease risk reduction and early intervention. Journal of Neuroscience 2018, 38, 6665–6681. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.Y.; Wang, Z.; Huang, H.C. Roles of apoE4 on the pathogenesis in Alzheimer’s disease and the potential therapeutic approaches. Cellular and molecular neurobiology 2023, 43, 3115–3136. [Google Scholar] [CrossRef]
- Main, B.S.; Villapol, S.; Sloley, S.S.; Barton, D.J.; Parsadanian, M.; Agbaegbu, C.; Stefos, K.; McCann, M.S.; Washington, P.M.; Rodriguez, O.C.; Burns, M.P. Apolipoprotein E4 impairs spontaneous blood brain barrier repair following traumatic brain injury. Molecular neurodegeneration 2018, 13, 1–18. [Google Scholar] [CrossRef]
- Bell, R.D.; Winkler, E.A.; Singh, I.; Sagare, A.P.; Deane, R.; Wu, Z.; Holtzman, D.M.; Betsholtz, C.; Armulik, A.; Sallstrom, J.; Berk, B.C. Apolipoprotein E controls cerebrovascular integrity via cyclophilin A. Nature 2012, 485, 512–516. [Google Scholar] [CrossRef]
- Montagne, A.; Nation, D.A.; Sagare, A.P.; Barisano, G.; Sweeney, M.D.; Chakhoyan, A.; Pachicano, M.; Joe, E.; Nelson, A.R.; D’Orazio, L.M.; Buennagel, D.P. APOE4 leads to blood–brain barrier dysfunction predicting cognitive decline. Nature 2020, 581, 71–76. [Google Scholar] [CrossRef]
- Merritt, V.C.; Clark, A.L.; Sorg, S.F.; Evangelista, N.D.; Werhane, M.L.; Bondi, M.W.; Schiehser, D.M.; Delano-Wood, L. Apolipoprotein E (APOE) ε4 genotype is associated with reduced neuropsychological performance in military veterans with a history of mild traumatic brain injury. Journal of clinical and experimental neuropsychology 2018, 40, 1050–1061. [Google Scholar] [CrossRef]
- Haney, M.S.; Pálovics, R.; Munson, C.N.; Long, C.; Johansson, P.K.; Yip, O.; Dong, W.; Rawat, E.; West, E.; Schlachetzki, J.C.; Tsai, A. APOE4/4 is linked to damaging lipid droplets in Alzheimer’s disease microglia. Nature 2024, 628, 154–161. [Google Scholar] [CrossRef]
- Muza, P.; Bachmeier, C.; Mouzon, B.; Algamal, M.; Rafi, N.G.; Lungmus, C.; Abdullah, L.; Evans, J.E.; Ferguson, S.; Mullan, M.; Crawford, F. APOE genotype specific effects on the early neurodegenerative sequelae following chronic repeated mild traumatic brain injury. Neuroscience 2019, 404, 297–313. [Google Scholar] [CrossRef]
- Fahy, E.; Subramaniam, S.; Murphy, R.C.; Nishijima, M.; Raetz, C.R.; Shimizu, T.; Spener, F.; Van Meer, G.; Wakelam, M.J.; Dennis, E.A. Update of the LIPID MAPS comprehensive classification system for lipids1. Journal of lipid research 2009, 50, S9–S14. [Google Scholar] [CrossRef]
- Bruce, K.D.; Zsombok, A.; Eckel, R.H. Lipid processing in the brain: a key regulator of systemic metabolism. Frontiers in endocrinology 2017, 8, 60. [Google Scholar] [CrossRef] [PubMed]
- Cermenati, G.; Mitro, N.; Audano, M.; Melcangi, R.C.; Crestani, M.; De Fabiani, E.; Caruso, D. Lipids in the nervous system: from biochemistry and molecular biology to patho-physiology. Biochimica et Biophysica Acta (BBA)-Molecular and Cell Biology of Lipids 2015, 1851, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Wang, X.; Zhang, L.; Fang, Y.; Zheng, Q.; Liu, X.; Yu, W.; Chen, S.; Ying, J.; Hua, F. Lipid metabolism and storage in neuroglia: role in brain development and neurodegenerative diseases. Cell & bioscience 2022, 12, 106. [Google Scholar]
- Nessel, I.; Michael-Titus, A.T. Lipid profiling of brain tissue and blood after traumatic brain injury: A review of human and experimental studies. In Seminars in Cell & Developmental Biology; Academic Press, April 2021; Volume 112, pp. 145–156. [Google Scholar]
- de Freitas, F.A.; Levy, D.; Zarrouk, A.; Lizard, G.; Bydlowski, S.P. Impact of oxysterols on cell death, proliferation, and differentiation induction: current status. Cells 2021, 10, 2301. [Google Scholar] [CrossRef]
- Sun, M.Y.; Linsenbardt, A.J.; Emnett, C.M.; Eisenman, L.N.; Izumi, Y.; Zorumski, C.F.; Mennerick, S. 24 (S)-Hydroxycholesterol as a modulator of neuronal signaling and survival. The Neuroscientist 2016, 22, 132–144. [Google Scholar] [CrossRef]
- Romero, J.; Toral-Rios, D.; Yu, J.; Paul, S.M.; Cashikar, A.G. 25-hydroxycholesterol promotes brain cytokine production and leukocyte infiltration in a mouse model of lipopolysaccharide-induced neuroinflammation. Journal of Neuroinflammation 2024, 21, 251. [Google Scholar] [CrossRef]
- Marwarha, G.; Ghribi, O. Does the oxysterol 27-hydroxycholesterol underlie Alzheimer's disease–Parkinson's disease overlap? Experimental gerontology 2015, 68, 13–18. [Google Scholar] [CrossRef]
- Nakazawa, T.; Miyanoki, Y.; Urano, Y.; Uehara, M.; Saito, Y.; Noguchi, N. Effect of vitamin E on 24 (S)-hydroxycholesterol-induced necroptosis-like cell death and apoptosis. The Journal of steroid biochemistry and molecular biology 2017, 169, 69–76. [Google Scholar] [CrossRef]
- Leoni, V.; Masterman, T.; Diczfalusy, U.; De Luca, G.; Hillert, J.; Björkhem, I. Changes in human plasma levels of the brain specific oxysterol 24S-hydroxycholesterol during progression of multiple sclerosis. Neuroscience letters 2002, 331, 163–166. [Google Scholar] [CrossRef]
- Björkhem, I.; Lövgren-Sandblom, A.; Leoni, V.; Meaney, S.; Brodin, L.; Salveson, L.; Winge, K.; Pålhagen, S.; Svenningsson, P. Oxysterols and Parkinson's disease: evidence that levels of 24S-hydroxycholesterol in cerebrospinal fluid correlates with the duration of the disease. Neuroscience letters 2013, 555, 102–105. [Google Scholar] [CrossRef]
- Lu, F.; Fan, S.; Romo, A.R.; Xu, D.; Ferriero, D.M.; Jiang, X. Serum 24S-hydroxycholesterol predicts long-term brain structural and functional outcomes after hypoxia-ischemia in neonatal mice. Journal of Cerebral Blood Flow & Metabolism 2021, 41, 312–323. [Google Scholar]
- Odnoshivkina, U.G.; Kuznetsova, E.A.; Petrov, A.M. 25-hydroxycholesterol as a signaling molecule of the nervous system. Biochemistry (Moscow) 2022, 87, 524–537. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Noh, M.Y.; Kim, H.; Cheon, S.Y.; Lee, K.M.; Lee, J.; Cha, E.; Park, K.S.; Lee, K.W.; Sung, J.J.; Kim, S.H. 25-Hydroxycholesterol is involved in the pathogenesis of amyotrophic lateral sclerosis. Oncotarget 2017, 8, 11855. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Xi, Y.; Yu, H.; An, Y.; Wang, Y.; Tao, L.; Wang, Y.; Liu, W.; Wang, T.; Xiao, R. 27-hydroxycholesterol promotes Aβ accumulation via altering Aβ metabolism in mild cognitive impairment patients and APP/PS1 mice. Brain Pathology 2019, 29, 558–573. [Google Scholar] [CrossRef]
- Wuolikainen, A.; Acimovic, J.; Lövgren-Sandblom, A.; Parini, P.; Andersen, P.M.; Björkhem, I. Cholesterol, oxysterol, triglyceride, and coenzyme Q homeostasis in ALS. Evidence against the hypothesis that elevated 27-hydroxycholesterol is a pathogenic factor. PloS one 2014, 9, p. [Google Scholar] [CrossRef]
- Mencarelli, C.; Martinez–Martinez, P. Ceramide function in the brain: when a slight tilt is enough. Cellular and Molecular Life Sciences 2013, 70, 181–203. [Google Scholar] [CrossRef]
- Chowdhury, M.R.; Jin, H.K.; Bae, J.S. Diverse roles of ceramide in the progression and pathogenesis of Alzheimer’s disease. Biomedicines 2022, 10, 1956. [Google Scholar] [CrossRef]
- Custodia, A.; Romaus-Sanjurjo, D.; Aramburu-Núñez, M.; Álvarez-Rafael, D.; Vázquez-Vázquez, L.; Camino-Castiñeiras, J.; Leira, Y.; Pías-Peleteiro, J.M.; Aldrey, J.M.; Sobrino, T.; Ouro, A. Ceramide/sphingosine 1-phosphate axis as a key target for diagnosis and treatment in Alzheimer’s disease and other neurodegenerative diseases. International Journal of Molecular Sciences 2022, 23, 8082. [Google Scholar] [CrossRef]
- Tringali, C.; Giussani, P. Ceramide and sphingosine-1-phosphate in neurodegenerative disorders and their potential involvement in therapy. International Journal of Molecular Sciences 2022, 23, 7806. [Google Scholar] [CrossRef]
- Han, X.; Holtzman, D.M.; McKeel, W.D., Jr.; Kelley, J.; Morris, J.C. Substantial sulfatide deficiency and ceramide elevation in very early Alzheimer's disease: potential role in disease pathogenesis. Journal of neurochemistry 2002, 82, 809–818. [Google Scholar] [CrossRef]
- He, X.; Huang, Y.; Li, B.; Gong, C.X.; Schuchman, E.H. Deregulation of sphingolipid metabolism in Alzheimer's disease. Neurobiology of aging 2010, 31, 398–408. [Google Scholar] [CrossRef]
- Fernández-Irigoyen, J.; Cartas-Cejudo, P.; Iruarrizaga-Lejarreta, M.; Santamaría, E. Alteration in the cerebrospinal fluid lipidome in Parkinson’s disease: a post-mortem pilot study. Biomedicines 2021, 9, 491. [Google Scholar] [CrossRef]
- Barbacci, D.C.; Roux, A.; Muller, L.; Jackson, S.N.; Post, J.; Baldwin, K.; Hoffer, B.; Balaban, C.D.; Schultz, J.A.; Gouty, S.; Cox, B.M. Mass spectrometric imaging of ceramide biomarkers tracks therapeutic response in traumatic brain injury. ACS chemical neuroscience 2017, 8, 2266–2274. [Google Scholar] [CrossRef] [PubMed]
- Ojo, J.O.; Algamal, M.; Leary, P.; Abdullah, L.; Mouzon, B.; Evans, J.E.; Mullan, M.; Crawford, F. Converging and differential brain phospholipid dysregulation in the pathogenesis of repetitive mild traumatic brain injury and Alzheimer’s disease. Frontiers in Neuroscience 2019, 13, 103. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Jin, Y.; Cheng, X.; Song, Q.; Wang, Y.; He, L.; Chen, T. The relevance between abnormally elevated serum ceramide and cognitive impairment in Alzheimer’s disease model mice and its mechanism. Psychopharmacology 2024, 241, 525–542. [Google Scholar] [CrossRef] [PubMed]
- Bernath, M.M.; Bhattacharyya, S.; Nho, K.; Barupal, D.K.; Fiehn, O.; Baillie, R.; Risacher, S.L.; Arnold, M.; Jacobson, T.; Trojanowski, J.Q.; Shaw, L.M. Serum triglycerides in Alzheimer disease: Relation to neuroimaging and CSF biomarkers. Neurology 2020, 94, e2088–e2098. [Google Scholar] [CrossRef]
- Parthasarathy, V.; Frazier, D.T.; Bettcher, B.M.; Jastrzab, L.; Chao, L.; Reed, B.; Mungas, D.; Weiner, M.; DeCarli, C.; Chui, H.; Kramer, J.H. Triglycerides are negatively correlated with cognitive function in nondemented aging adults. Neuropsychology 2017, 31, 682. [Google Scholar] [CrossRef]
- Zhao, B.; Shang, S.; Li, P.; Chen, C.; Dang, L.; Jiang, Y.; Wang, J.; Huo, K.; Deng, M.; Wang, J.; Qu, Q. The gender-and age-dependent relationships between serum lipids and cognitive impairment: a cross-sectional study in a rural area of Xi’an, China. Lipids in Health and Disease 2019, 18, 1–11. [Google Scholar] [CrossRef]
- Kuo, J.R.; Lim, S.W.; Zheng, H.X.; Ho, C.H.; Chang, C.H.; Chio, C.C.; Wee, H.Y. Triglyceride is a good biomarker of increased injury severity on a high fat diet rat after traumatic brain injury. Neurochemical Research 2020, 45, 1536–1550. [Google Scholar] [CrossRef]
- Hahnefeld, L.; Vogel, A.; Gurke, R.; Geisslinger, G.; Schäfer, M.K.; Tegeder, I. Phosphatidylethanolamine deficiency and triglyceride overload in perilesional cortex contribute to non-goal-directed hyperactivity after traumatic brain injury in mice. Biomedicines 2022, 10, 914. [Google Scholar] [CrossRef]
- Zhou, W.; Xu, D.; Peng, X.; Zhang, Q.; Jia, J.; Crutcher, K.A. Meta-analysis of APOE 4 allele and outcome after traumatic brain injury. Journal of neurotrauma 2008, 25, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Hellstrøm, T.; Andelic, N.; Holthe, Ø.Ø.; Helseth, E.; Server, A.; Eiklid, K.; Sigurdardottir, S. APOE-ε4 is associated with reduced verbal memory performance and higher emotional, cognitive, and everyday executive function symptoms two months after mild traumatic brain injury. Frontiers in neurology 2022, 13, 735206. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.T.; Hsiao, T.; Hsieh, C.J.; Chiang, Y.H.; Yen, T.C.; Chiu, W.T.; Lin, K.J.; Hu, C.J. Accumulation of amyloid in cognitive impairment after mild traumatic brain injury. Journal of the neurological sciences 2015, 349, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.Y.J.; Seet, R.C.; Huang, S.H.; Long, L.H.; Halliwell, B. Different patterns of oxidized lipid products in plasma and urine of dengue fever, stroke, and Parkinson's disease patients: cautions in the use of biomarkers of oxidative stress. Antioxidants & redox signaling 2009, 11, 407–420. [Google Scholar]
- La Marca, V.; Maresca, B.; Spagnuolo, M.S.; Cigliano, L.; Dal Piaz, F.; Di Iorio, G.; Abrescia, P. Lecithin-cholesterol acyltransferase in brain: Does oxidative stress influence the 24-hydroxycholesterol esterification? Neuroscience Research 2016, 105, 19–27. [Google Scholar] [CrossRef]
- Weiner, M.F.; Vega, G.L.; Diaz-Arrastia, R.; Moore, C.; Madden, C.; Hudak, A.; Lütjohann, D. Plasma 24S-hydroxycholesterol and other oxysterols in acute closed head injury. Brain injury 2008, 22, 611–615. [Google Scholar] [CrossRef]
- Tian, N.; Fa, W.; Dong, Y.; Liu, R.; Liu, C.; Liu, K.; Mao, M.; Zhu, M.; Liang, X.; Wang, N.; Ma, Y. Triglyceride–glucose index, Alzheimer's disease plasma biomarkers, and dementia in older adults: The MIND-China study. Alzheimer's & Dementia: Diagnosis, Assessment & Disease Monitoring 2023, 15, p. [Google Scholar]
- Dunk, M.M.; Rapp, S.R.; Hayden, K.M.; Espeland, M.A.; Casanova, R.; Manson, J.E.; Shadyab, A.H.; Wild, R.; Driscoll, I. Plasma oxysterols are associated with serum lipids and dementia risk in older women. Alzheimer's & Dementia 2024, 20, 3696–3704. [Google Scholar] [CrossRef]
- Lin, J.B.; Sene, A.; Santeford, A.; Fujiwara, H.; Sidhu, R.; Ligon, M.M.; Shankar, V.A.; Ban, N.; Mysorekar, I.U.; Ory, D.S.; Apte, R.S. Oxysterol signatures distinguish age-related macular degeneration from physiologic aging. EBioMedicine 2018, 32, 9–20. [Google Scholar] [CrossRef]
- Passarelli, M.N.; McDonald, J.G.; Thompson, B.M.; Arega, E.A.; Palys, T.J.; Rees, J.R.; Barry, E.L.; Baron, J.A. Association of demographic and health characteristics with circulating oxysterol concentrations. Journal of clinical lipidology 2022, 16, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Leoni, V.; Long, J.D.; Mills, J.A.; Di Donato, S.; Paulsen, J.S.; members of the PREDICT-HD study group. Plasma 24S-hydroxycholesterol correlation with markers of Huntington disease progression. Neurobiology of disease 2013, 55, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Teitsdottir, U.D.; Halldorsson, S.; Rolfsson, O.; Lund, S.H.; Jonsdottir, M.K.; Snaedal, J.; Petersen, P.H. Cerebrospinal fluid C18 ceramide associates with markers of Alzheimer’s disease and inflammation at the pre-and early stages of dementia. Journal of Alzheimer’s Disease 2021, 81, 231–244. [Google Scholar] [CrossRef] [PubMed]
- Roy, D.; Chakrabarti, S.S.; Banerjee, A.; Sharma, P.; Biswas, A.; Chakrabarti, S. Serum 24-hydroxycholesterol in probable Alzheimer's dementia: Reexploring the significance of a tentative Alzheimer's disease biomarker. Aging Medicine 2019, 2, 74–81. [Google Scholar] [CrossRef]
- Bandaru, V.V.; Troncoso, J.; Wheeler, D.; Pletnikova, O.; Wang, J.; Conant, K.; Haughey, N.J. ApoE4 disrupts sterol and sphingolipid metabolism in Alzheimer's but not normal brain. Neurobiology of aging 2009, 30, 591–599. [Google Scholar] [CrossRef]
- Hume, P.A.; Theadom, A.; Lewis, G.N.; Quarrie, K.L.; Brown, S.R.; Hill, R.; Marshall, S.W. A comparison of cognitive function in former rugby union players compared with former non-contact-sport players and the impact of concussion history. Sports Med. 2017, 47, 1209–1220. [Google Scholar] [CrossRef]
- Sharma, A.; Hind, K.; Hume, P.; Singh, J.; Neary, J.P. Neurovascular coupling by functional near infra-red spectroscopy and sport-related concussion in retired rugby players: The UK rugby health project. Front. Hum. Neurosci. 2020, 14, 42. [Google Scholar] [CrossRef]
- Entwistle, I.; Francis, P.; Lees, M.; Hume, P.; Hind, K. Lean Mass, Muscle Strength, and Muscle Quality in Retired Rugby Players: The UK Rugby Health Project. Int. J. Sports Med. 2022, 43, 958–963. [Google Scholar] [CrossRef]
- Finnoff, J.T.; Jelsing, E.J.; Smith, J. Biomarkers, genetics, and risk factors for concussion. PM&R 2011, 3, 452–459. [Google Scholar] [CrossRef]
- Pham, N.; Akonasu, H.; Shishkin, R.; Taghibiglou, C. Plasma soluble prion protein, a potential biomarker for sport-related concussions: A pilot study. PLoS ONE 2015, 10, e0117286. [Google Scholar] [CrossRef]
Figure 1.
Comparison of the serum concentration of 24-HC, 25-HC, 27-HC, ceramide, total triglycerides, and ApoE4 in a concussion vs. control group. Each dot represents an individual data point, with the bars representing the median and the range. Blue dots = the control group, and black dots = the concussion group. * p < 0.05; ** p < 0.01; ns not significant.
Figure 1.
Comparison of the serum concentration of 24-HC, 25-HC, 27-HC, ceramide, total triglycerides, and ApoE4 in a concussion vs. control group. Each dot represents an individual data point, with the bars representing the median and the range. Blue dots = the control group, and black dots = the concussion group. * p < 0.05; ** p < 0.01; ns not significant.
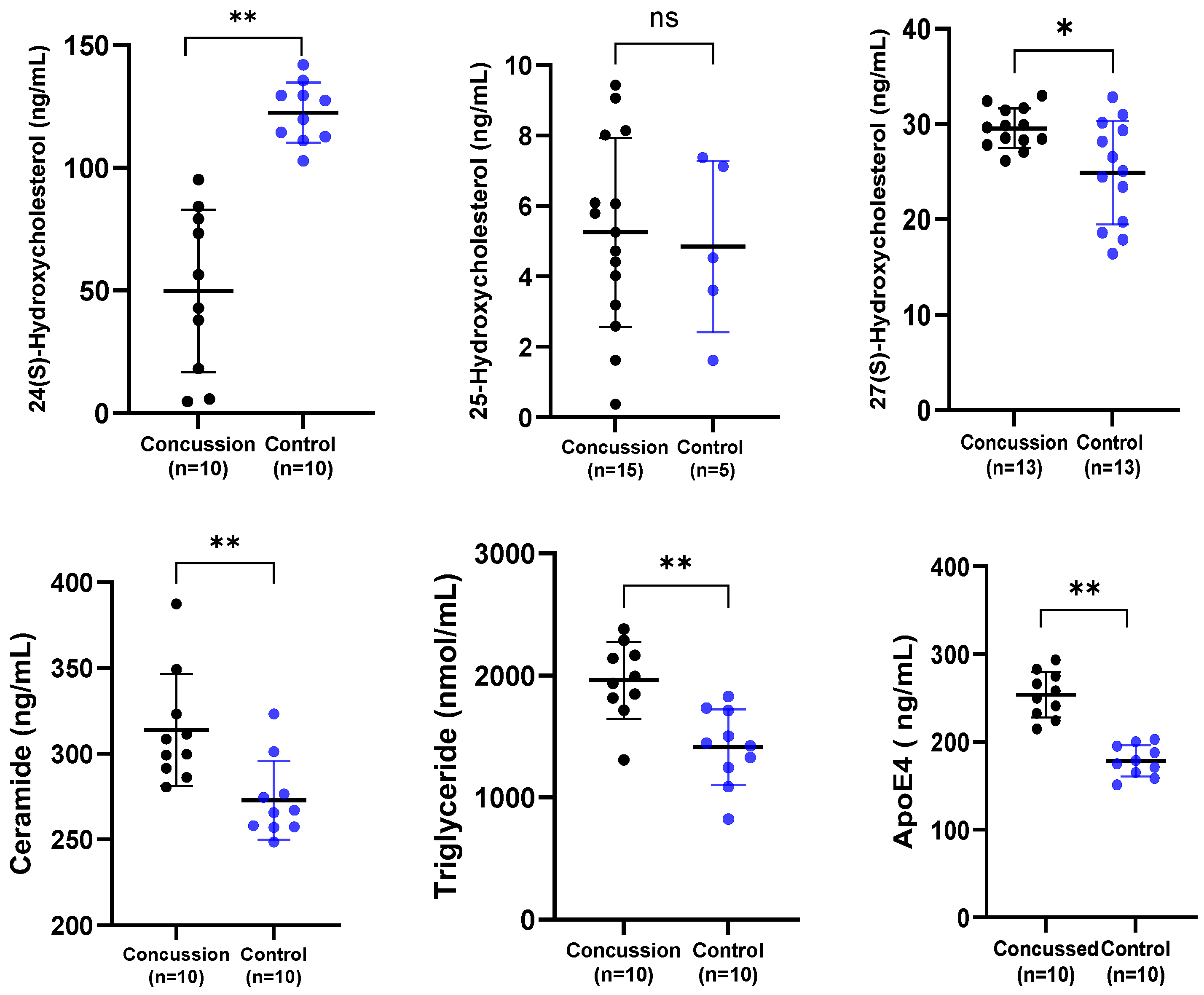
Figure 2.
Scatter plots to highlight the correlation between lipid biomarkers. Dots are individual data points for the concentration of the substances specified on graphs of specific serum samples (n = 20 or more).
Figure 2.
Scatter plots to highlight the correlation between lipid biomarkers. Dots are individual data points for the concentration of the substances specified on graphs of specific serum samples (n = 20 or more).
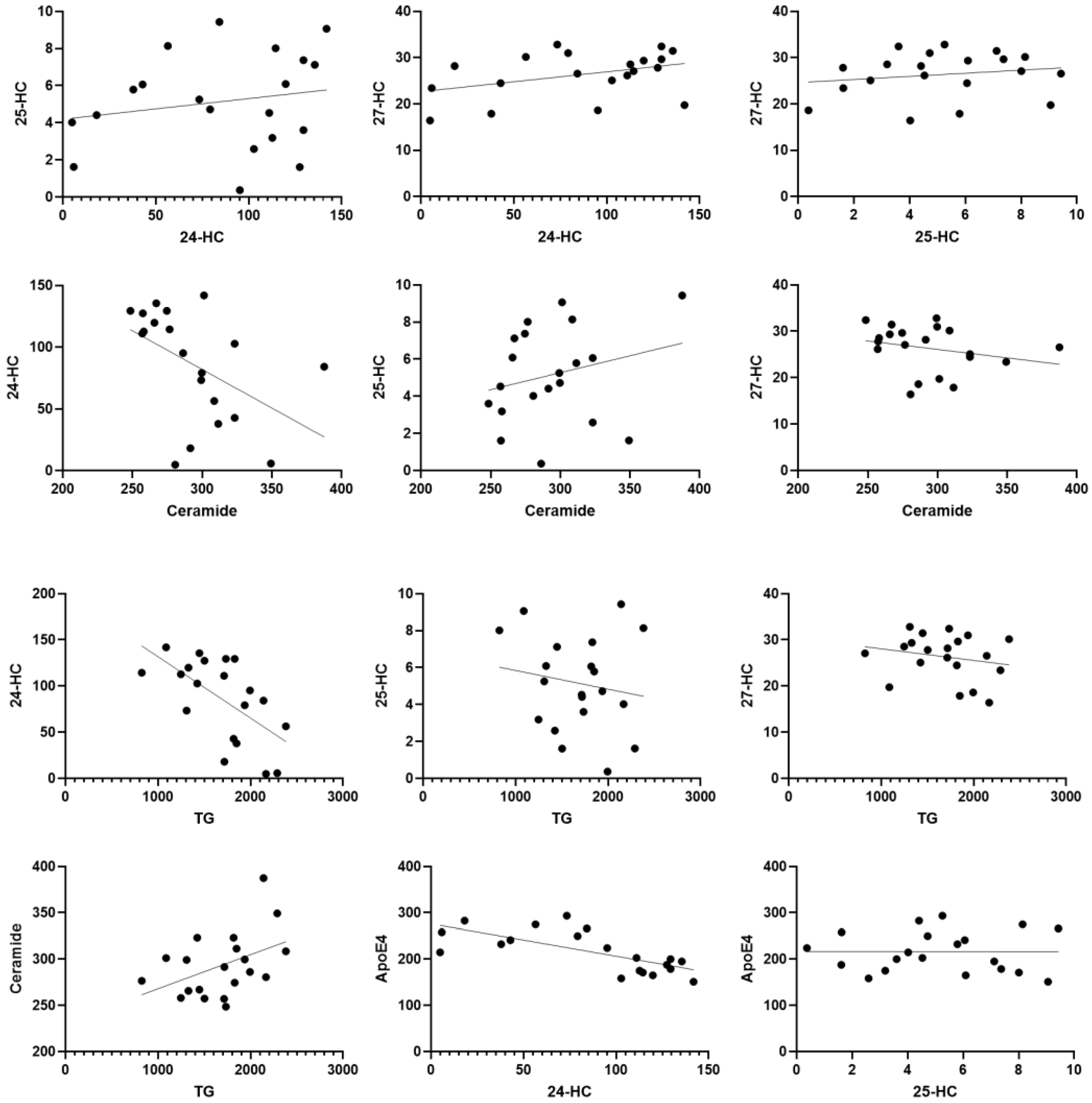
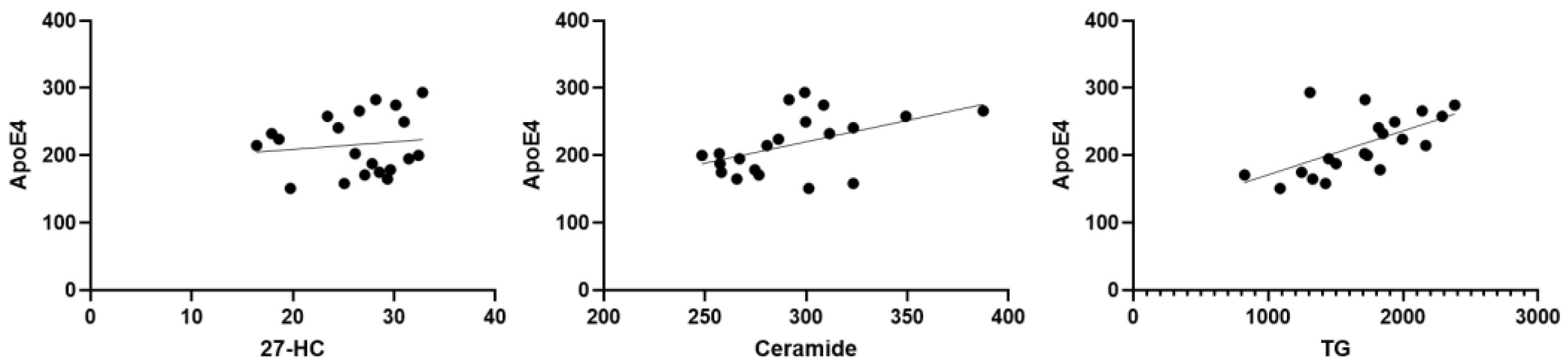

Table 1.
Descriptive characteristics.
| Concussed Group N = 26 |
Control Group N = 19 |
|
|---|---|---|
| Mean ages | 39.32±6.44 | 47.19±12.11 |
| Mean ages at retirement | 26.56±4.89 | 33.77±9.24 |
| Mean years since retirement from the sport | 7.04±5.23 | 8.33±4.29 |
| Playing position | 4 prop, 3 hooker, 1 forward, 2 s row, 1 fly half, 4 center, 4 wing, 3 backward, 1 openside flanker, 1 blindside flanker, 2 lock. |
1 blindside flanker, 2 backwards, 1 wing, 1 number 8, 1 prop, 1 standoff, 12 non-athletes. |
| Mean weight | 100.2±11.14 | 86.53±15.47 |
| Mean height | 183.81±7.09 | 178.5±6.33 |
| Rugby league (RL) or union (RU) | 12(RL), 14 (RU) | 5(RU), 2(RL), 12 (N/A) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



