
Submitted:
23 September 2025
Posted:
24 September 2025
You are already at the latest version
Abstract
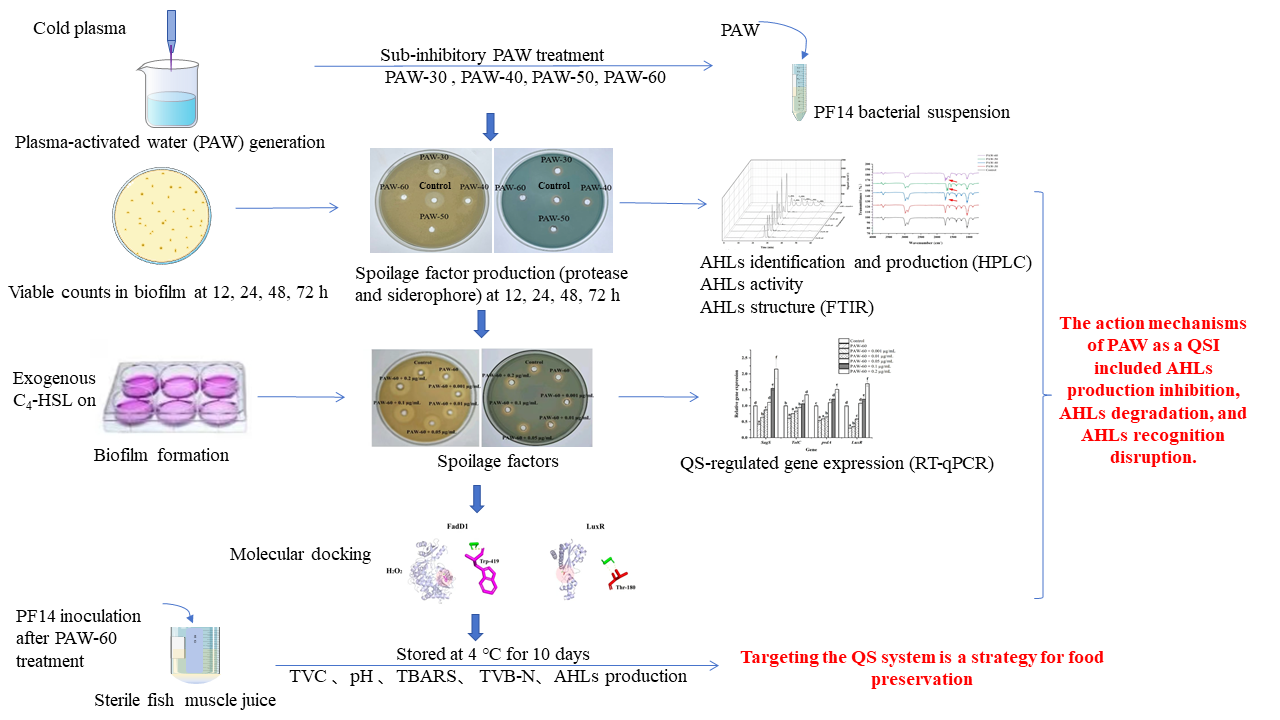
Plasma-activated water (PAW) is an emerging disinfectant, but few studies have explored its role as a quorum sensing inhibitor (QSI) for biofilm suppression, and the mechanisms of action remain unclear. Herein, we investigated the effects of PAW on biofilm formation and spoilage factors secretion in Pseudomonas fluorescens under sub-inhibitory conditions. The results showed that PAW-60 treatment achieved a maximum reduction of 1.29 log CFU/mL in P. fluorescens biofilm after 12 h incubation. Furthermore, PAW-60 completely inhibited protease production (100%) and decreased siderophore production by 31.87%. C4-HSL was identified as the dominant signaling molecule, and its production was reduced by 34.34% to 84.07% after selected PAW treatments. Meanwhile, C4-HSL activity was significantly decreased by 42.58% to 65.38% (P < 0.05). FTIR analysis detected the formation of a new C=O group after PAW treatment, indicating that PAW degraded AHLs by oxidative modification. Exogenous C4-HSL gradually restored biofilm biomass, spoilage factors production, and QS-related gene expression levels, implying that the inhibitory effects of PAW primarily resulted from the disruption of AHLs transduction. Molecular docking showed that long-lived reactive species in PAW could bind to AHLs synthetic protein (FadD1) and receptor protein (LuxR) via hydrogen bonding. PAW-60 treatment effectively reduced spoilage activity of P. fluorescens inoculated to fish muscle juice and extended its shelf life from 8 to 10 days during 4 °C storage. These findings revealed that PAW mitigated biofilm formation and the spoilage process of food by blocking signaling transduction, which involved AHLs production inhibition, degradation, and recognition disruption.
Keywords:
plasma-activated water
; quorum sensing inhibitor (QSI)
; AHLs degradation
; biofilm inhibition
; spoilage factors
; Pseudomonas fluorescens
1. Introduction
Quorum sensing (QS) is a cell-to-cell communication mechanism employed by bacteria to coordinate collective behaviors in response to population density (Bassler, 1999). The QS system operates through the synthesis, release, and reception of signaling molecules. When the signaling molecules reach a critical threshold, they will bind to the cognate receptor and trigger expression of target genes, including bioluminescence, virulence factors, spoilage factors, antibiotic resistance, biofilm formation, etc. (Miller et al., 2001). The signaling molecules of QS are classified into three major categories: acyl homoserine lactones (AHLs) in Gram-negative bacteria, autoinducing peptides (AIPs) in Gram-positive bacteria, and autoinducer-2 (AI-2) in interspecies communication. AHLs consist of a homoserine lactone ring with a fatty acyl side chain ranging from 4 to 18 carbons (C4-C18) in length (Fuqua et al., 2002). The homoserine lactone (HSL) ring is synthesized from S-adenosylmethionine (SAM), while the acyl side chain derives either from acyl carrier protein (ACP) or acyl-CoA precursors. The two components are conjugated by LuxI-type synthetase to form AHLs. LuxR-type protein specifically recognizes and binds AHLs, enabling target gene expression in a cell-density-dependent manner. Biofilms are structured microbial communities encased with self-produced extracellular polymeric matrix. Compared to planktonic bacteria, biofilms significantly enhance resistance to environmental stresses, including chemical disinfectants, antibiotics, thermal fluctuations, etc, rendering them remarkably difficult to eradicate (Lin et al., 2023). Excessive use of antibiotics or chemical sanitizers has resulted in the emergence of multidrug-resistant bacteria, which is a major public health concern (Mohanty et al., 2025). Recent studies have demonstrated that quorum-sensing inhibitors (QSIs) can suppress biofilm development through disrupting the QS system without directly inactivating or killing bacteria (Yin et al., 2022; Ge et al., 2024; Nayak et al., 2025). Thus, QSIs reduced the potential for inducing microbial resistance. This attribute confers them an outstanding advantage over conventional antibiotics or chemical sanitizers, which exert selective pressure by directly killing bacteria (Alonso et al., 2024). Different QSIs exhibit distinct mechanisms of action, which mainly include (1) inhibition of QS signal molecular synthesis by interfering with LuxI-type protein or signal precursors, such as some natural extracts (Gutierrez-Pacheco et al., 2019), (2) signal degradation via structural modification, such as quorum-quenching (QQ) enzymes (Wang et al., 2023), and based on the catalytic sites of AHLs, QQ enzymes are normally divided into three types: AHL lactonases (open the homoserine lactone ring), AHL acylase (break the amide linkage between homoserine lactone and acyl chain), and AHL oxidoreductase (oxidize the carbon of acyl chain or reduce the C3 carboxyl group), (3) blockage of QS signal reception by disrupting LuxR-type protein or recognition (Kalia, 2013). Over the past decade, research on QSIs has mainly focused on essential oils (EOs), plant extracts (Guo et al., 2023), or QQ enzymes (Wang et al., 2023). Although promising anti-biofilm effects have been demonstrated, the practical application of these quorum sensing inhibitors (QSIs) remains limited due to inherent drawbacks: plant extracts suffer from poor water solubility, high volatility, and sensitivity to light and oxygen, while QQ enzymes are often costly. Therefore, there is an urgent need to develop more accessible, stable, and cost-effective QSIs.
Pseudomonas fluorescens has been identified as a specific spoilage organism (SSO), especially in protein-rich foods, such as aquatic, meat, and dairy products (Ge et al., 2024). P. fluorescens exhibits potent spoilage activity through extracellular secretion of protease, lipase, and siderophore. These spoilage factors decompose food into amines, ammonia, aldehydes, sulfides, free fatty acids, etc, causing rapid quality degradation and offensive off-flavors (Liu et al., 2024). Due to its heat-resistant nature, protease is more challenging to inactivate than the bacterial cells. Consequently, spoilage factor production is also a particular concern. In addition, as a psychrotrophic bacterium, P. fluorescens maintains metabolic activity and cellular replication even at refrigerating temperatures, posing significant challenges for food quality and safety (Yang et al., 2024). Previous studies have shown that the spoilage ability of P. fluorescens is closely related to the QS system (Li et al., 2018; Tang et al., 2019; Wang et al., 2021). AHLs are the common QS signaling molecules in P. fluorescens. Therefore, targeting AHLs in P. fluorescens could be a promising strategy for food spoilage control.
Plasma is a partially or wholly ionized gas, containing a mixture of reactive species, charged particles, neutral atoms, etc (Zhao et al., 2019). Plasma is categorized into non-thermal (cold) and thermal types, with the former being suitable for laboratory applications. Plasma-activated water (PAW) is generated by exposing water to cold plasma discharge. The interactions between plasma and water initiate a series of chemical reactions, enabling PAW with unique properties due to the generation of reactive oxygen species (ROS) and reactive nitrogen species (RNS) (Zhao et al., 2020b). PAW has shown a significant antibacterial effect and is considered a potential green disinfectant. The ability of plasma or PAW to inhibit biofilm formation has been reported (Cui et al., 2021; Basiri et al., 2023), while most studies have focused on bactericidal effects under lethal conditions, its capacity to act as a QSI remains largely unexplored. Cai et al. (2025) found that AHLs production and biofilm formation of P. aeruginosa were reduced after PAW treatment. Li et al. (2019) showed that PAW inhibited biofilm formation and downregulated QS-related virulence gene expression of Enterococcus faecalis. However, the QS pathway involves multiple steps, including the production, release, and recognition of signaling molecules. To the best of our knowledge, the action mechanisms of PAW as a QSI against biofilm formation have not been reported, and the relationship between the spoilage process of food and the QS signaling molecules has not been illustrated.
In this study, the effects of PAW on biofilm formation, spoilage factors secretion, as well as the production, activity, and structure of AHLs in P. fluorescens were examined under sub-inhibitory conditions. The impacts of exogenous AHLs on biofilm formation, spoilage factors, and QS-related gene expression levels were measured. In addition, molecular docking interactions between long-lived reactive species in PAW and critical QS pathway proteins were investigated. Finally, to evaluate the spoilage potential of P. fluorescens after PAW treatment, the bacterium was inoculated into fish muscle juice and stored at 4 °C for 10 days, during which total viable counts (TVC), pH, thiobarbituric acid reactive substances (TBARS), total volatile basic nitrogen (TVB-N), and AHLs production were measured.
2. Materials and Methods
2.1. Strains and Culture Preparation
P. fluorescens employed in this investigation was originally isolated from a large yellow croaker, which was marked as PF14. The biosensor strain of Agrobacterium tumefaciens KYC55 (KYC55) and Chromobacterium violaceum 026 (CV026) were kindly provided by Prof. Mingsheng Dong (Nanjing Agricultural University, Nanjing, China) and Prof. Junli Zhu (Zhejiang Gongshang University, Hangzhou, China), respectively. Neither KYC55 nor CV026 produces AHLs, but they can sense exogenous AHLs to produce β-galactosidase and violacein, respectively. PF14 was overnight-cultured in tryptic soy broth (TSB, HangZhouBaiSi Biochemical Technology Co., Ltd., Hangzhou, China) at 28 ℃. Following incubation, bacterial cells were harvested via centrifugation (12,000× g, 5 min), the supernatant was removed, and the pellet was resuspended in sterile deionized water to the initial culture volume.
2.2. PAW Generation Under Sub-Inhibitory Conditions
For PAW generation, an atmospheric plasma jet (PG-1000ZD, Nanjing Suman Plasma Technology Co., Ltd., Nanjing, China) was employed. The plasma system was operated at 300 W and 20 kHz with a 30 L/min compressed air flow. The plasma nozzle was positioned 10 cm above the water surface during generation. PAW was obtained by exposing 200 mL of sterile deionized water to the plasma jet for varying durations (30, 40, 50, and 60 s), which were designated as PAW-30 through PAW-60. Our previous study showed that no significant inhibitory effect on bacterial counts or growth curves was observed after selected PAW treatments (Zhao et al., 2025), indicating that PAW treatments were under sub-inhibitory conditions of PF14. For PAW treatment, 1 mL of bacterial suspension was transferred to 9 mL of freshly prepared PAW, and the contact time of the mixture was 2 min. The bacterial suspension treated with sterile deionized water was used as the control.
2.3. Enumeration of Bacterial Cells in Biofilm
Biofilm biomass of PF14 was quantified based on Li et al. (2022). A sterile glass slice (20 mm × 20 mm) was placed in a 6-well microplate, then 0.5 mL of PAW-treated bacterial suspension was 1:10 (v/v) inoculated to TSB in each well before incubation at 28 °C for 12, 24, 48, and 72 h. The glass slice was rinsed thrice using 0.01% PBS (Bkmam, China) after incubation. Each slice was transferred to a sterilized beaker containing 10 mL of 0.01% PBS and subjected to ultrasonication (Ymnl-1000Y, Nanjing Immanuel Instrument Equipment Co., Ltd., Nanjing, China) at 40 kHz/L (40 W) for 10 min. The detached bacteria were decimally diluted in 0.1% (w/v) peptone water (Guangdong Huankai Microbial SCI&TECH. Co., Ltd., Guangzhou, China) and plated on tryptic soy agar (TSA, HangZhouBaiSi Biochemical Technology Co., Ltd., Hangzhou, China). The colonies between 30–300 were counted after incubation at 28 ℃ for 48 h.
2.4. Spoilage Factors Assay
Protease and siderophore are the main spoilage factors of P. fluorescens, which were determined by the plate perforation method (Ding et al., 2017). PF14 was incubated at 28 °C for 12, 24, 48, and 72 h, respectively, after PAW treatment as described in Section 2.2. Cell-free supernatants were obtained by centrifugation (12,000 × g, 5 min) and sterile filtration (0.22 μm filter). The production of protease and siderophore was measured using plates containing 1.5% (v/v) skimmed milk and Chrome Azurol Sulphonate (CAS) plates (Hopebio, Qingdao, China), respectively. The wells in the plate were punched using a sterilized Oxford cup, and 200 µL of the supernatant was pipetted into the well. After incubation at 28 °C for 24 h, the diameter of the transparent hydrolysis circle and the orange circle was measured as protease and siderophore production, respectively.
2.5. AHLs Production Assay
The type of AHLs in P. fluorescens and the effect of PAW on the production of AHLs were determined using high-performance liquid chromatography (HPLC). As described in Section 2.2, 3 mL of the PAW-treated bacterial suspension was transferred to 27 mL of TSB and incubated at 28 ℃ for 24 h. The bacterial suspension treated with sterile deionized water was used as the control. AHLs were extracted according to the method of Li et al. (2018) with slight modifications. The culture was centrifuged (12,000 × g, 5 min) after incubation, and the supernatants were extracted three times with an equal volume of ethyl acetate with 0.1% (v/v) acetic acid. The extracts were evaporated to dryness in a rotary evaporator at 30 °C, and the residue was dissolved in 1 mL of methanol and passed through a 0.22 μm sterile filter. Standard solutions of C4-HSL, C6-HSL, C8-HSL, C10-HSL, C12-HSL, and C14-HSL (Sigma Aldrich, US) were prepared to determine the type and production of AHLs. The extracts and standard solutions were analyzed using an HPLC system (Thermo Fisher Scientific Co., Ltd., US), which was equipped with Aminex HPLC Columns (Bio-Rad, US) and a UV detector at 210 nm. The mobile phase consists of water (solvent A) and methanol (solvent B), both of which contain 0.1% formic acid. The gradient elution procedure was performed as follows: 0–20 min, 70% B; 20–40 min, 90% B; 40–60 min, 70% B (Jahid et al., 2015).
2.6. AHLs Activity Assay
Exogenous AHLs can induce KYC55 to express β-galactoside, which can hydrolyze o-nitro-β-d-galactoside (ONPG) to yellow ortho-nitrophenol (Gui et al., 2017), thus AHLs activity can be reflected by measuring β-galactoside activity. AHLs activity was determined according to the method of Xu et al. (2024) with slight modifications. KYC55 was cultured overnight (28 ℃, 16 h) in TSB supplemented with 1 µg/mL of tetracycline, 100 µg/mL of gentamicin, and 100 µg/mL of spectinomycin. The AHLs crude was prepared as described in Section 2.5. A 200 μL of the crude was pipetted to 2 mL of TSB containing 1% (v/v) overnight-cultured KYC55 and incubated at 28 °C for 16 h, followed by measurement of OD600. A 0.9 mL of Z-buffer, 45 μL of chloroform, and 15 μL of 0.1% SDS were added to a 5 mL sterile centrifuge tube containing 100 μL of KYC55 bacterial suspension, and mixed thoroughly. Then, 200 μL of 4 mg/mL ONPG was transferred to the centrifuge tube, and the mixture was incubated at 28 °C after mixing. A 600 μL of 1 M Na2CO3 was pipetted immediately into the mixture to terminate the reaction when the solution turned yellow, and the time (T) the solution took to change color was recorded. Finally, the absorbance of the supernatant at 420 nm was measured after centrifugation (12,000 × g, 5 min). β-galactosidase activity was calculated using the following formula:
β-galactosidase activity = (1000 × OD420)/(OD600 × T × 0.1)
2.7. AHLs Structure Assay
FTIR was used to investigate the impact of PAW treatment on AHLs structure, which was determined by referring to the method of Flynn et al. (2016) with slight modifications. A 0.1 mL of 1 mg/mL AHLs was pipetted to 0.9 mL of PAW, and the spectra were measured at 4000–400 cm- after 2 min exposure using FTIR (Nicolet iS50, Thermo Fisher Scientific Co., Ltd., US) with a resolution of 32 cm- and scanning times of 32. The 100 μg/mL of the standard AHLs solution was used as the control.
2.8. Determination of Biofilm Formation with Exogenous C4-HSL
The effect of exogenous C4-HSL on the biofilm formation of PF14 after PAW-60 treatment was determined by the crystal violet staining method (Cui et al., 2021). In brief, a sterilized glass slice (20 mm × 20 mm) served as the biofilm carrier and was placed in a 6-well plate. Each well was immersed with 4.5 mL of TSB containing different concentrations of C4-HSL (0, 0.01, 0.05, 0.1, and 0.2 µg/mL), then 0.5 mL of bacterial suspension was transferred to the well after PAW-60 treatment and incubated at 28 ℃ for 24 h. The bacterial suspension treated with sterile deionized water served as the control. After incubation, the biofilm on the glass slide was rinsed three times using 0.01% PBS to remove planktonic bacteria, and stained with 0.5% crystal violet for 10 min, then the unbound crystal violet was washed off with 0.01% PBS. Finally, 95% ethanol was transferred to dissolve the residual crystal violet, and the absorbance at 590 nm was measured.
2.9. Determination of Spoilage Factors with Exogenous C4-HSL
Protease and siderophore production were determined by the plate perforation method. As described in Section 2.8, PF14 was incubated in TSB containing different concentrations of C4-HSL (0, 0.01, 0.05, 0.1, and 0.2 µg/mL) for 24 h after PAW-60 treatment. The production of protease and siderophore was determined as described in Section 2.4.
2.10. Determination of Gene Expression with Exogenous C4-HSL
Four QS-regulated genes (biofilm formation, SgaS; spoilage factors: TolC, pvdA; AHLs receptor: LuxR) were selected based on the transcriptomic annotation (data not shown). The selected gene expression was determined by RT-qPCR. PF14 was incubated at 28 ℃ for 24 h after PAW-60 treatment according to Section 2.8. The supernatant was removed after centrifugation at 12,000 × g for 5 min at 4 ℃, and the pellets were collected. Bacterial RNA was extracted according to the instructions of the RNA Extraction Kit (Sangon Biotech Co., Ltd., Shanghai, China). The cDNA was synthesized using the HisyGo RT Red SuperMix for qPCR (+gDNA Wiper) kit (Vazyme, China). RT-qPCR analysis was performed according to the SYBR Green I kit (Vazyme, Nanjing, China). 16S rRNA of PF14 was used as the internal reference gene, and the primers used in this study are listed in Table 1.
2.11. Molecular Docking
The action mechanisms between long-lived reactive species (H2O2, NO2-, and NO3-) in PAW and key biomolecules (acyl-CoA synthetase of AHLs, FadD1; AHLs receptor, LuxR) were studied by molecular docking. The biomolecules (FadD1, LuxR) were selected based on our previous transcriptomic results, which demonstrated that the genes were down-regulated after PAW-60 treatment (data not shown). Swissmodl software was used to predict the molecular structure of proteins, and water molecules and metal ions of proteins were removed using Pymol software. Molecular docking was performed using AutoDock Vina, and the results were visualized using Pymol.
2.12. In Vivo Spoilage Potential of P. fluorescens in Fish Muscle Juice Assay
To evaluate the effect of PAW on the spoilage potential of P. fluorescens, PF14 was inoculated into sterile fish muscle juice after PAW-60 treatment and stored at 4 ℃ for 10 days, during which TVC, pH, TBARS, TVB-N, and AHLs production in fish muscle juice were detected every 2 days.
2.12.1. Fish Muscle Juice Contamination
The large yellow croakers were purchased from Metro supermarket (Zhenjiang, China) and transported to the laboratory on ice within 1 h. Sterile fish muscle juice was prepared according to the method of Dalgaard (1995). Briefly, the fish was cleaned, minced, and boiled in an appropriate amount of sterilized water for 5 min. The fish muscle juice was obtained by filtration, centrifugation, and sterilization at 121 ℃ for 15 min. Then trimethylamine oxide, L-cysteine, and L-methionine were added to the fish muscle juice to reach a final concentration of 1.6, 40, and 40 mg/L, respectively, to compensate for the nutrient loss caused by dilution and sterilization. After PAW-60 treatment as described in Section 2.2, the mixture was immediately tenfold-diluted to achieve a concentration of 3–4 log CFU/mL. Then, 5 mL of the diluted solution was transferred to 45 mL of sterile fish muscle juice to reach an inoculated level of 2–3 log CFU/mL and stored at 4 °C for 10 days. The bacteria treated with sterile deionized water were used as the control. During storage, 10% (v/v) fresh PAW-60 or sterile deionized water was added to fish muscle juice at 12-hour intervals.
2.12.2. TVC and pH Analysis
The TVC of the fish muscle juice was determined by the standard plate counting method according to the reference of Liu et al. (2025). In brief, 1 mL of the fish juice was transferred to 9 mL of sterile water and tenfold diluted using 0.1% (w/v) peptone water. The 100 μL of appropriate dilution was plated on the TSA, and the colonies were counted after incubation at 28 ℃ for 48 h. The pH of the fish muscle juice was determined using a pH meter (Mettler-Toledo LE438).
2.12.3. TBARS and TVB-N Analysis
TBARS of the fish muscle juice was detected according to Gan et al. (2024) with slight modifications. A 5 mL of the fish muscle juice was mixed thoroughly with 50 mL of trichloroacetic acid and filtered using Whatman filter paper. After that, 5 mL of the filtrate was mixed with 5 mL of 0.02 M thiobarbituric acid (TBA). The mixture was vortexed thoroughly before being subjected to a water bath (90 ℃, 30 min). The absorbance of the solution was measured at 590 nm after cooling down to room temperature. A standard curve of malondialdehyde (MDA) was plotted to transform the concentration into absorbance values. TBARS was expressed as mg MDA/100 mL. TVB-N was quantified according to the Chinese National Standard GB 5009.228–2016. In brief, 25 mL of the fish muscle juice was homogenized with 100 mL of deionized water and filtered after 30 min. The filtrate was acidified with 10 mL of perchloric acid, and basic nitrogen compounds were steam-distilled before titration with 0.01 M HCl. TVB-N of the fish muscle juice was expressed as mg N per 100 mL.
2.12.4. AHLs Production Analysis
The biosensor strain of CV026 does not produce AHLs, but it can sense exogenous AHLs to produce violacein, and violacein can dissolve in dimethylsulfoxide (DMSO), and thus the absorbance of violacein can semi-quantitatively determine AHLs production. The AHLs secreted by PF14 to the fish muscle juice were detected using CV026 according to the method of Li et al. (2020) with some modifications. A 20 mL of the fish muscle juice was centrifuged (12,000× g, 5 min), and AHLs in the supernatants were extracted as described in Section 2.5. The crude extract was dissolved in methanol and stored at -20 ℃ for later use. CV026 was inoculated into TSB supplemented with 20 µg/mL kanamycin and incubated at 28 ℃ for 16 h. The 200 μL of the AHLs extracts was transferred to 5 mL of fresh TSB inoculated with 1% (v/v) overnight-cultured CV026. After incubation at 28 ℃ for 24 h, 1 mL of the solution was centrifuged (12,000× g, 5 min), and the pellets were resuspended in 1 mL of DMSO. The suspension was vortexed thoroughly to allow a full dissolution of violacein before centrifugation (12,000 × g, 5 min), and the absorbance of the supernatant was detected at 590 nm.
2.13. Statistical Analysis
In this study, each group of experiments was performed in triplicate, and the values were expressed as mean ± standard deviation. SPSS 16.0 software was used for statistical analysis. Analysis of variance (ANOVA) was used to evaluate the differences between groups, and a level of p < 0.05 was considered statistically significant.
3. Results and Discussion
3.1. The Effect of PAW on Biofilm Biomass
Biofilm formation of PF14 during 72 h incubation was quantitatively analyzed by calculating viable counts in the biofilm. As shown in Figure 1, all the selected PAW treatments decreased biofilm formation compared to the control, with a significant difference observed after incubation for 12 and 24 h (p < 0.05), and prolonging PAW generation time progressively increased repression rates. The biofilm at 12 h incubation was significantly decreased by 0.10, 0.35, 0.86, and 1.29 log CFU/mL, respectively, after PAW-30, PAW-40, PAW-50, and PAW-60 treatment (p < 0.05). Moreover, Figure 1 shows that biofilm formation reaches a maximal level at 24 h before a subsequent decline until 72 h. Thus, the peak biofilm was obtained at 24 h with a value of 7.08, 6.95, 6.73, 6.41, and 6.05 Log CFU/mL for control, PAW-30, PAW-40, PAW-50, and PAW-60, respectively. The trend in biofilm biomass can be attributed to the maturation phase within 24 h, followed by the dispersal phase during 24 to 72 h. The strengthened inhibitory effect resulted from the accumulated reactive species in PAW when PAW generation time was extended (Zhao et al., 2020a), which enhanced disruption of the QS pathway and led to suppression of biofilm formation. A previous study has also shown that PAW treatments mitigated biofilm biomass of Enterococcus faecalis in the ultra-low dose ROS group during 72 h incubation, and no inhibitory effect against planktonic bacteria was observed at 0 h (Li et al., 2019).
3.2. The Effect of PAW on Spoilage Factors
Proteases can degrade proteins into sulfur-containing compounds and volatile nitrogen metabolites, leading to food spoilage. Siderophore is a small molecule with specific chelation of iron ion (III), which enables bacteria to acquire iron under nutrient-limited conditions and improve survival competitiveness (Wang et al., 2021). As presented in Figure 2a,b, the production of protease and siderophore was decreased after PAW treatments, and the suppression rates were enhanced with prolonged PAW generation time. Figure 2a shows that the diameter of the hydrolysis circle at 12 h incubation was significantly decreased from 27.70 to 26.25, 12.50 mm after PAW-30 and PAW-40 treatment, respectively (p < 0.05), and no hydrolysis circle was observed after PAW-50 and PAW-60 treatment, indicating a complete inhibition of protease production. However, there was no significant difference in siderophore production at 12 h incubation after PAW-30 and PAW-40 treatment, and a significant reduction of 26.37% and 31.87% was obtained after PAW-50 and PAW-60 treatment, respectively (p < 0.05). In addition, similar to biofilm formation, both protease and siderophore production peaked at 24 h and then decreased until 72 h. The trend was consistent with previous studies (Chen et al., 2023; Cui et al., 2023), which demonstrated that spoilage factor production was positively correlated with biofilm formation. As the spoilage factor production is regulated by the QS system via a series of cascading reactions (Wang et al., 2021), the decrease in spoilage factor production can be attributed to the disruption of the QS pathway after PAW treatment. Though the study of PAW on spoilage factor production has not been covered, the effects of many other antibacterial substances have been reported (Li et al., 2018; Machado et al., 2020; Lang et al., 2025). Li et al. (2016) found that the protease production of P. fluorescens was decreased by 58.50% under the sub-MIC concentration of 0.1 µg/mL cinnamaldehyde. Wang et al. (2021) presented that the siderophore production of P. fluorescens was decreased after Cytidine-5′-monophosphate (5′-CMP) and 5′-adenylic acid treatment.
3.3. The Effect of PAW on AHLs Production of PF14
The type and production of AHLs in PF14 were measured by comparing with standard AHLs. As shown in Figure 3a, C4-HSL was identified as the most dominant AHLs after incubation for 24 h. The type of AHLs was consistent with the previous study of Tang et al. (2019), who also detected C4-HSL as the main AHLs in P. fluorescens after 24 h incubation. Furthermore, the production of C4-HSL was remarkably decreased from 312.80 to 205.39, 184.47, 94.78, and 49.82 µg/mL, respectively, after PAW-30, PAW-40, PAW-50, and PAW-60 treatment, with a reduction of 34.34%, 41.03%, 69.70%, and 84.07%, respectively (Figure 3b). The decrease in AHLs production indicated that QS signaling transduction was disrupted after PAW treatment. Since biofilm formation and spoilage factor production are QS-regulated phenotypes, the disruption of the QS pathway caused a reduction in biofilm biomass and spoilage factor production, which was demonstrated in Section 3.1 and 3.2, respectively. The study of Cai et al. (2025) demonstrated 42.00% and 100% (undetectable level) reduction of C4-HSL production in P. aeruginosa after PAW and plasma-activated lactic acid (PALA) treatment, respectively. Flynn et al. (2016) described that no violacein production in the bio-reporter CV026 was observed after AHLs were exposed to plasma for 60 s, indicating a complete decomposition. Yin et al. (2022) also reported a significant decrease in C4-HSL in P. aeruginosa after exposure to sub-MICs of soy isoflavones, and a dose-dependent manner was observed. Fidaleo et al. (2013) revealed that triclosan inhibited overall AHLs production by preventing the activity of acyl carrier protein reductase FabL protein.
3.4. The Effect of PAW on AHLs Activity of PF14
As illustrated in Figure 4, PAW treatments attenuated AHLs activity, and PAW-60 showed the maximal repression. There was no significant reduction in the activity of AHLs after PAW-30 treatment compared to the control, while PAW-40, PAW-50, and PAW-60 significantly reduced the activity of AHLs by 42.58%, 55.99%, and 65.38%, respectively (p < 0.05). The weakened activity of AHLs resulted from oxidative degradation by reactive species generated in PAW. Though the study of PAW on AHLs activity is quite rare, the effects of other antibacterial substances on AHLs activity have been reported. Zhang et al. (2018) showed that the activity of AHLs in P. fluorescens and Erwinia carotovora was decreased during the monitoring of 24 h under different sub-MICs of hexanal. Zhao et al. (2021) found that the AHLs activity of Aeromonas vickerii was reduced by 39.60% when the concentration of garlic extract was 1.20 mg/mL. Frey et al. (2010) also reported a decreased AHLs activity after hydroxyl radical treatment. Shen et al. (2021) discovered PF-1240 (a new QQ enzyme) reduced AHLs activity via quenching AHLs with different carbon chain lengths.
3.5. The Effect of PAW on C4-HSL Structure
According to Section 3.3, the dominant signaling molecule of PF14 was C4-HSL, and thus C4-HSL was selected as the subject of this study. As shown in Figure 5, PAW-30 did not change the structure of C4-HSL compared with the standard. A vibration at approximately 1710 cm- was observed after PAW-40, PAW-50, and PAW-60 treatments, indicating the generation of a new C=O group in the C4-HSL (Flynn et al., 2016). The vibration of the new functional group was gradually strengthened with prolonged PAW generation time, suggesting that C4-HSL oxidation was enhanced. The acyl side chain of C4-HSL underwent oxidative modification via ROS in PAW, leading to structural alterations and compromised signaling potency. PAW-30 did not cause structural changes in C4-HSL, which explained the maintenance of AHLs activity in Section 3.4. PAW-60 exhibited the most pronounced oxidative degradation to C4-HSL, thereby the greatest reduction in C4-HSL activity (Section 3.4). Flynn et al. (2016) reported an OH functional group and a new C=O group generation after four types of AHLs exposure to plasma for 60 s using UHPLC-MS, and a complete decomposition after 240 s exposure. They also found that shorter-chain AHLs were more resistant to plasma degradation than longer-chain AHLs. Chowdhary et al. (2007) reported that CYP102A1, a well-known cytochrome from Bacillus megaterium, efficiently oxidized AHLs, and the oxidation mainly happened at the ω-1, ω-2, and ω-3 carbons of the acyl side chain.
3.6. The Effect of Exogenous C4-HSL on the Biofilm Formation of PF14
As shown in Figure 6, PAW-60 significantly reduced the biofilm biomass of PF14 by 33.03% after 24 h of incubation compared with the control (p < 0.05). The biofilm biomass of PF14 was increased with the supplementation of exogenous C4-HSL in a dose-dependent manner, demonstrating the vital role of the QS signaling molecule in biofilm formation. There was no significant difference compared with the control when exogenous C4-HSL was 0.05 µg/mL, and biofilm biomass was significantly increased by 26.60% and 56.88%, respectively, when the exogenous supplementation was 0.1 and 0.2 µg/mL (p < 0.05). Li et al. (2019) reported that the biofilm formation of P. fluorescens was significantly increased by 127.49% when the exogenous C4-HSL was 2 µg/mL (p < 0.05). The findings further indicated that PAW treatment suppressed biofilm formation by disrupting QS signaling transduction, and the supplementation of exogenous C4-HSL can restore biofilm formation.
3.7. The Effect of Exogenous C4-HSL on the Spoilage Factors of PF14
As shown in Figure 7, both protease (a,b) and siderophore (c,d) production were gradually restored with the supplementation of exogenous C4-HSL. Similar to biofilm formation, there was no significant difference in protease and siderophore production compared with the control when the exogenous C4-HSL was 0.05 µg/mL. Protease and siderophore production increased significantly by 30.86% and 24.31%, respectively, at the maximum supplementation of 0.2 µg/mL (p < 0.05). The results indicated that AHLs modulated spoilage factor production in P. fluorescens, and PAW acted as a disruptor of AHLs transduction. Li et al. (2019) showed that 2 µg/mL of exogenous C4-HSL stimulated the protease production of P. fluorescens. Zhang et al. (2023) reported that 100 ng/mL of exogenous C8-HSL resulted in a 59.68% recovery of protease activity in P. fluorescens after being treated with sub-MIC hexanal.
3.8. The Effect of Exogenous C4-HSL on the Gene Expression of PF14
Gene transcription was determined using RT-qPCR. As shown in Figure 8, genes involved in biofilm formation (SgaS), spoilage factors (TolC, pvdA), and AHLs receptor (LuxR) were all down-regulated after PAW-60 treatment, and the supplementation of exogenous C4-HSL stimulated the gene expression in a dose-dependent manner. There was no significant difference in gene expression of biofilm formation and spoilage factor production compared with the control when the exogenous C4-HSL was 0.05 µg/mL, which was consistent with the results of biofilm formation (Section 3.6) and spoilage factor production (Section 3.7). Moreover, the expression of gene SgaS, TolC, pvdA, and LuxR was significantly increased by 2.15-fold, 1.35-fold, 1.51-fold, and 1.69-fold, respectively, at the maximal supplementation of 0.2 µg/mL (p < 0.05). Gene up-regulation also explained the stimulatory effect of exogenous C4-HSL on biofilm formation, spoilage factor production, and AHLs transduction.
3.9. Molecular Docking Analysis
The binding interactions between long-lived reactive species of H2O2, NO2-, and NO3- in PAW and AHLs synthetic (FadD1) and AHLs receptor (LuxR) proteins were investigated. Figure 9a shows that H2O2 formed 1 hydrogen bond with Trp-419; NO2- formed 6 hydrogen bonds with Leu-275, Tyr-23, Ser-33, Val-29, Gln-32, and Phe-37; and NO3- formed 7 hydrogen bonds with Leu-282, Ser-284, Asn-45, and Leu-46. Figure 9b presents that H2O2, NO2-, and NO3- form 2, 5, and 4 hydrogen bonds with the amino acid residue of LuxR, respectively. H2O2 interacted with Thr-180. NO2- interacted with Ile-145, His-120, Gly-121, and Trp-98. NO3- interacted with Ser-137, Thr-58, and Asp-83. These findings demonstrated that the three long-lived reactive species in PAW could bind to various amino acid residues of AHLs synthetic proteins and receptor proteins through hydrogen bonding, thereby disrupting AHLs transduction of the QS pathway, which further inhibited biofilm formation and spoilage factor production. Ge et al. (2024) reported that benzyl isothiocyanate formed pi-pi, pi-alkyl bonds, and van der Waals forces with different amino acid residues of the LuxR-type protein, thus suppressing biofilm formation in P. fluorescens. Yang et al. (2025) described that peptide Ser-Phe formed hydrogen bonds with several different amino acids in the AI-2 synthetic protein of LuxS and AI-2 receptor protein of LuxP, thereby destroying the AI-2 QS system in Vibrio parahaemolyticus.
3.10. The Effect of PAW on the Spoilage Potential of PF14 in Fish Muscle Juice
As seen in Figure 10a, TVC of the fish muscle juice increased significantly during the whole storage, and the value of PAW-60 was significantly lower than that of the control (p < 0.05). According to the International Committee on Microbiological Practices for Food (Wang et al., 2021), the maximum acceptable limit for the TVC in aquatic products is 7.00 log CFU/mL. The control and PAW-60 exceeded the acceptable limit on day 8 and day 10, respectively, with a value of 7.23 and 7.55 log CFU/mL. Moreover, PAW-60 decreased TVC by 0.79 log CFU/mL compared with the control on day 10. The results indicated that PAW-60 treatment slowed down the bacterial proliferation during the cold storage of 10 days.
Figure 10b shows that the pH of the fish juice was lower than the control after PAW-60 treatment, and there was a significant difference between the control and PAW-60 on day 8 and day 10 (p < 0.05). In addition, pH values presented a downward trend first (day 2), followed by an upward trend. This phenomenon was attributed to the glycolysis and ATP degradation at the early stage, producing lactic acid and decreasing pH. Subsequently, the nitrogen-containing compounds in the fish were broken down into alkaline substances with the extension of storage, increasing pH. The results were consistent with Cui et al. (2023), who also reported a similar pH trend on tuna fish chunks during 4 °C storage.
TBARS value represents the degree of fat oxidation, which is also an important indicator to evaluate food quality. Similar to TVC, PAW-60 reduced TBARS, and the values kept increasing during the storage, which reached 1.12 and 0.82 mg/kg for the control and PAW-60, respectively, on day 10 (Figure 10c). TVB-N is an indicator of protein oxidation, and seafood products will be considered completely spoiled when exceeds 30 mg/100 mL (Wang et al., 2021). After PAW-60 treatment, TVB-N was lower than that of the control during the whole storage, and the control exceeded the threshold on day 8 with a value of 31.85 mg/100 mL, while the value of PAW-60 was 24.92 mg/100 mL on the same day, and it increased to 33.83 mg/100 mL on day 10 (Figure 10d). TVB-N results were consistent with the TVC analysis, both of which reached the complete spoilage threshold on day 8 and day 10 for the control and PAW-60 treatment, respectively.
Figure 10e shows that AHLs production kept increasing during the storage, and the value of PAW-60 was lower than that of the control with a significant reduction of 26.85% and 17.14% on day 6 and day 8, respectively (p < 0.05). The results also manifested that AHLs production was positively correlated with the spoilage process of food, and lower AHLs production indicated less spoilage. Lu et al. (2005) reported that the QS signaling molecule of AI-2 was involved in the spoilage process of tomatoes.
The results illustrated that PAW-60 treatment attenuated the spoilage activity of PF14, resulting in reduced TVC, fat oxidation, and protein oxidation of fish muscle juice. In addition, the QS signaling molecule of AHLs played an imperative role in the spoilage process of food. Though the report of PAW on food preservation under sub-inhibitory conditions is rare, the bactericidal effects of PAW on various food preservation have been reported (Liao et al., 2018; Zhao et al., 2020b; Chanioti et al., 2023).
4. Conclusions
Our study demonstrated that biofilm formation and spoilage factors in P. fluorescens were mitigated by PAW treatment under sub-inhibitory conditions, and the inhibitory effect was positively correlated with PAW generation time. The structure of C4-HSL was oxidatively modified after selected PAW treatments except for PAW-30, which weakened AHLs activity. Meanwhile, AHLs production was also significantly decreased after PAW treatments except for PAW-30 (p < 0.05). Biofilm biomass, spoilage factors, and gene expression levels of biofilm formation (SgaS), spoilage factors (TolC, pvdA), and AHLs receptor (LuxR) were gradually restored with exogenous C4-HSL supplementation after PAW-60 treatment. The recovery further confirmed that the inhibitory effects of PAW on biofilm formation and spoilage factors secretion were principally attributed to the disruption of AHLs pathway. Molecular docking revealed that H2O2, NO2-, and NO3- in PAW interacted with various amino acid residues of AHLs synthetic (FadD1) and AHLs receptor protein (LuxR) through hydrogen bonding. Furthermore, the spoilage activity of P. fluorescens inoculated into fish muscle juice was attenuated after PAW treatment. It can be summarized that reactive species generated in PAW disrupted QS signaling transduction by oxidatively modifying AHLs structure and interacting with both AHLs synthetic and receptor proteins. The interference reduced AHLs production, activity, and recognition, which ultimately suppressed biofilm formation, spoilage factors, and the spoilage process of food. Therefore, targeting the QS system can be a strategy for food preservation.
Author Contributions
Yi-Ming Zhao: Writing–original draft, Formal analysis, Data curation, Conceptualization, Supervision, Funding acquisition, Review & Editing. Qingyun Zhang: Writing–original draft, Formal analysis, Data curation. Lin Zhang: Writing–original draft, Formal analysis, Data curation. Yulong Bao: Visualization, Review & Editing. Yiting Guo: Conceptualization, Review & Editing. Liurong Huang: Conceptualization, review & editing. Ronghai He: Conceptualization, review & editing. Haile Ma: Visualization, Review & Editing. Da-Wen Sun: Conceptualization, Review & Editing.
Acknowledgments
The authors would like to acknowledge the financial support from the National Natural Science Foundation of China (32202227), the China Postdoctoral Science Foundation (2023M731380), and the Senior Talent Program of Jiangsu University (21JDG044).
Conflicts of Interest
The authors declare that they have no conflict of interest.
References
- Alonso, V. P. P., M. M. Furtado, C. H. T. Iwase, J. Z. Brondi-Mendes, and M. d. S. Nascimento. 2024. Microbial resistance to sanitizers in the food industry. Critical reviews in food science and nutrition 64, 3: 654–669. [Google Scholar] [CrossRef]
- Basiri, N., M. Zarei, M. Kargar, and F. Kafilzadeh. 2023. Effect of plasma-activated water on the biofilm-forming ability of Salmonella enterica serovar Enteritidis and expression of the related genes. International journal of food microbiology 406, 110419. [Google Scholar] [CrossRef] [PubMed]
- Bassler, B. L. 1999. How bacteria talk to each other: regulation of gene expression by quorum sensing. Current opinion in microbiology 2, 6: 582–587. [Google Scholar] [CrossRef]
- Cai, Z., W. Zhang, G. Liao, C. Huang, J. Wang, and J. Zhang. 2025. Inhibiting mechanism of Pseudomonas aeruginosa biofilm formation-An innovational reagent of plasma-activated lactic acid. Journal of Water Process Engineering 69, 106613. [Google Scholar] [CrossRef]
- Chanioti, S., M. Giannoglou, P. Stergiou, and et al. 2023. Plasma-activated water for disinfection and quality retention of sea bream fillets: Kinetic evaluation and process optimization. Innovative Food Science & Emerging Technologies 85, 103334. [Google Scholar]
- Chen, J., Z. Sun, and J. Jin. 2023. Role of siderophore in Pseudomonas fluorescens biofilm formation and spoilage potential function. Food microbiology 109, 104151. [Google Scholar] [CrossRef]
- Chowdhary, P. K., N. Keshavan, H. Q. Nguyen, J. A. Peterson, J. E. González, and D. C. Haines. 2007. Bacillus megaterium CYP102A1 oxidation of acyl homoserine lactones and acyl homoserines. Biochemistry 46, 50: 14429–14437. [Google Scholar] [CrossRef]
- Cui, F., Q. Wang, J. Liu, D. Wang, J. Li, and T. Li. 2023. Effects of deletion of siderophore biosynthesis gene in Pseudomonas fragi on quorum sensing and spoilage ability. International journal of food microbiology 396, 110196. [Google Scholar] [CrossRef]
- Cui, H., H. Li, M. A. Abdel-Samie, D. Surendhiran, and L. Lin. 2021. Anti-Listeria monocytogenes biofilm mechanism of cold nitrogen plasma. Innovative Food Science & Emerging Technologies 67, 102571. [Google Scholar]
- Dalgaard, P. 1995. Qualitative and quantitative characterization of spoilage bacteria from packed fish. International journal of food microbiology 26, 3: 319–333. [Google Scholar] [CrossRef] [PubMed]
- Ding, T., T. Li, Z. Wang, and J. Li. 2017. Curcumin liposomes interfere with quorum sensing system of Aeromonas sobria and in silico analysis. Scientific reports 7, 1: 8612. [Google Scholar] [CrossRef]
- Fidaleo, M., A. Zuorro, and R. Lavecchia. 2013. Enhanced antibacterial and anti-quorum sensing activities of triclosan by complexation with modified β-cyclodextrins. World Journal of Microbiology and Biotechnology 29, 9: 1731–1736. [Google Scholar] [CrossRef]
- Flynn, P. B., A. Busetti, E. Wielogorska, and et al. 2016. Non-thermal Plasma Exposure Rapidly Attenuates Bacterial AHL-Dependent Quorum Sensing and Virulence. Sci Rep 6, 1: 26320. [Google Scholar] [CrossRef]
- Frey, R. L., L. He, Y. Cui, and et al. 2010. Reaction of N-acylhomoserine lactones with hydroxyl radicals: rates, products, and effects on signaling activity. Environmental science & technology 44, 19: 7465–7469. [Google Scholar]
- Fuqua, C., and E. P. Greenberg. 2002. Listening in on bacteria: acyl-homoserine lactone signalling. Nature reviews Molecular cell biology 3, 9: 685–695. [Google Scholar] [CrossRef]
- Gan, J., M. Mukaddas, Y. Tao, H. Liu, K. Ye, and G. Zhou. 2024. High-voltage electrostatic field with 35 kV-15 min could reduce Pseudomonas spp. to maintain the quality of pork during− 1° C storage. Innovative Food Science & Emerging Technologies 94, 103700. [Google Scholar]
- Ge, Z., X. Du, and J. Liu. 2024. Benzyl isothiocyanate suppresses biofilms and virulence factors as a quorum sensing inhibitor in Pseudomonas fluorescens. LWT 204, 116387. [Google Scholar] [CrossRef]
- Gui, M., R. Wu, L. Liu, S. Wang, L. Zhang, and P. Li. 2017. Effects of quorum quenching by AHL lactonase on AHLs, protease, motility and proteome patterns in Aeromonas veronii LP-11. International journal of food microbiology 252, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Guo, N., X. Bai, Y. Shen, and T. Zhang. 2023. Target-based screening for natural products against Staphylococcus aureus biofilms. Critical reviews in food science and nutrition 63, 14: 2216–2230. [Google Scholar] [CrossRef]
- Gutierrez-Pacheco, M. M., A. T. Bernal-Mercado, F. J. Vazquez-Armenta, and et al. 2019. Quorum sensing interruption as a tool to control virulence of plant pathogenic bacteria. Physiological and Molecular Plant Pathology 106, 281–291. [Google Scholar] [CrossRef]
- Jahid, I. K., M. F. R. Mizan, A. J. Ha, and S.-D. Ha. 2015. Effect of salinity and incubation time of planktonic cells on biofilm formation, motility, exoprotease production, and quorum sensing of Aeromonas hydrophila. Food microbiology 49, 142–151. [Google Scholar] [CrossRef]
- Kalia, V. C. 2013. Quorum sensing inhibitors: an overview. Biotechnology advances 31, 2: 224–245. [Google Scholar] [CrossRef]
- Lang, X., W. Lan, S. Liu, and X. Sun. 2025. Quorum sensing inhibitory of plant extracts on specific spoilage organisms and the potential utilization on the preservation of aquatic products. Chemical Engineering Journal 506, 160259. [Google Scholar] [CrossRef]
- Li, P., J. Mei, and J. Xie. 2022. Carbon dioxide can inhibit biofilms formation and cellular properties of Shewanella putrefaciens at both 30° C and 4° C. Food Research International 161, 111781. [Google Scholar] [CrossRef]
- Li, T., X. Sun, and H. Chen. 2020. Methyl anthranilate: a novel quorum sensing inhibitor and anti-biofilm agent against Aeromonas sobria. Food microbiology 86, 103356. [Google Scholar] [CrossRef] [PubMed]
- Li, T., D. Wang, and N. Liu. 2018. Inhibition of quorum sensing-controlled virulence factors and biofilm formation in Pseudomonas fluorescens by cinnamaldehyde. International journal of food microbiology 269, 98–106. [Google Scholar] [CrossRef]
- Li, T. T., F. C. Cui, F. L. Bai, G. H. Zhao, and J. R. Li. 2016. Involvement of Acylated Homoserine Lactones (AHLs) of Aeromonas sobria in Spoilage of Refrigerated Turbot Scophthalmus maximus. SENSORS 16, 7. [Google Scholar] [CrossRef]
- Li, Y., J. Pan, D. Wu, Y. Tian, J. Zhang, and J. Fang. 2019. Regulation of Enterococcus faecalis biofilm formation and quorum sensing related virulence factors with ultra-low dose reactive species produced by plasma activated water. Plasma Chemistry and Plasma Processing 39, 35–49. [Google Scholar] [CrossRef]
- Liao, X., Y. Su, and D. Liu. 2018. Application of atmospheric cold plasma-activated water (PAW) ice for preservation of shrimps (Metapenaeus ensis). Food Control 94, 307–314. [Google Scholar] [CrossRef]
- Lin, L., P. Zhang, and X. Chen. 2023. Inhibition of Staphylococcus aureus biofilms by poly-L-aspartic acid nanoparticles loaded with Litsea cubeba essential oil. International Journal of Biological Macromolecules 242, 124904. [Google Scholar] [CrossRef]
- Liu, J., Y. Wang, P. Yang, and et al. 2025. Quality decline of prepared dishes stored at 4° C: Microbial regulation of nitrite and biogenic amine formation. Food microbiology 128, 104730. [Google Scholar] [CrossRef]
- Liu, L., J. Li, M. Tu, and et al. 2024. Complete genome sequence provides information on quorum sensing related spoilage and virulence of Aeromonas salmonicida GMT3 isolated from spoiled sturgeon. Food Research International 196, 115039. [Google Scholar] [CrossRef]
- Lu, L., M. E. Hume, and S. D. Pillai. 2005. Autoinducer-2–like activity on vegetable produce and its potential involvement in bacterial biofilm formation on tomatoes. Foodbourne Pathogens & Disease 2, 3: 242–249. [Google Scholar]
- Machado, I., L. R. Silva, E. D. Giaouris, L. F. Melo, and M. Simões. 2020. Quorum sensing in food spoilage and natural-based strategies for its inhibition. Food Research International 127, 108754. [Google Scholar] [CrossRef]
- Miller, M. B., and B. L. Bassler. 2001. Quorum sensing in bacteria. Annual Reviews in Microbiology 55, 1: 165–199. [Google Scholar] [CrossRef]
- Mohanty, D., M. Suar, and S. K. Panda. 2025. Nanotechnological interventions in bacteriocin formulations–advances, and scope for challenging food spoilage bacteria and drug-resistant foodborne pathogens. Critical reviews in food science and nutrition 65, 6: 1126–1143. [Google Scholar] [CrossRef]
- Nayak, S. R. R., P. Pohokar, and A. Das. 2025. Chalcone derivative enhance poultry meat preservation through quorum sensing inhibition against Salmonella (Salmonella enterica serovar Typhi) contamination. Food Control 171, 111155. [Google Scholar] [CrossRef]
- Shen, Y., F. Cui, D. Wang, T. Li, and J. Li. 2021. Quorum quenching enzyme (pf-1240) capable to degrade ahls as a candidate for inhibiting quorum sensing in food spoilage bacterium Hafnia alvei. Foods 10, 11: 2700. [Google Scholar] [CrossRef] [PubMed]
- Tang, R., J. Zhu, L. Feng, J. Li, and X. Liu. 2019. Characterization of LuxI/LuxR and their regulation involved in biofilm formation and stress resistance in fish spoilers Pseudomonas fluorescens. International journal of food microbiology 297, 60–71. [Google Scholar] [CrossRef]
- Wang, D., F. Cui, L. Ren, J. Li, and T. Li. 2023. Quorum-quenching enzymes: Promising bioresources and their opportunities and challenges as alternative bacteriostatic agents in food industry. Comprehensive Reviews in Food Science and Food Safety 22, 2: 1104–1127. [Google Scholar] [CrossRef]
- Wang, Y., Y. Wang, J. Chen, and et al. 2021. Screening and preservation application of quorum sensing inhibitors of Pseudomonas fluorescens and Shewanella baltica in seafood products. LWT 149, 111749. [Google Scholar] [CrossRef]
- Xu, C., L. Ni, and C. Du. 2024. Decoding Microcystis aeruginosa quorum sensing through AHL-mediated transcriptomic molecular regulation mechanisms. Science of the Total Environment 926, 172101. [Google Scholar] [CrossRef]
- Yang, W., S. Liu, and X. Liu. 2025. Novel quorum-sensing inhibitor peptide SF derived from Penaeus vannamei myosin inhibits biofilm formation and virulence factors in Vibrio parahaemolyticus. LWT, 117542. [Google Scholar] [CrossRef]
- Yang, X., W. Lan, and X. Sun. 2024. Effects of chlorogenic acid-grafted-chitosan on biofilms, oxidative stress, quorum sensing and c-di-GMP in Pseudomonas fluorescens. International Journal of Biological Macromolecules 273, 133029. [Google Scholar] [CrossRef]
- Yin, L., Y. Zhang, F. Azi, and et al. 2022. Inhibition of biofilm formation and quorum sensing by soy isoflavones in Pseudomonas aeruginosa. Food Control 133, 108629. [Google Scholar] [CrossRef]
- Zhang, Y., J. Kong, F. Huang, and et al. 2018. Hexanal as a QS inhibitor of extracellular enzyme activity of Erwinia carotovora and Pseudomonas fluorescens and its application in vegetables. Food Chem 255, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y., H. Yu, Y. Xie, Y. Guo, Y. Cheng, and W. Yao. 2023. Inhibitory effects of hexanal on acylated homoserine lactones (AHLs) production to disrupt biofilm formation and enzymes activity in Erwinia carotovora and Pseudomonas fluorescens. J Food Sci Technol 60, 1: 372–381. [Google Scholar] [CrossRef]
- Zhao, D., Y. Ding, W. Chen, J. Hu, and X. Zhou. 2021. AHLs Based Quorum Sensing in Aeromonas veronii bv.veronii and Its Interruption from Garlic Extract. Journal of Chinese Institute Of Food Science and Technology 21, 2: 28–36. [Google Scholar]
- Zhao, Y.-M., M. de Alba, D.-W. Sun, and B. Tiwari. 2019. Principles and recent applications of novel non-thermal processing technologies for the fish industry—A review. Critical reviews in food science and nutrition 59, 5: 728–742. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.-M., L. Zhang, Y. Bao, and et al. 2025. The inhibitory mechanisms of plasma-activated water on biofilm formation of Pseudomonas fluorescens by disrupting quorum sensing. In Food Research International. p. 117436. [Google Scholar]
- Zhao, Y. M., S. Ojha, C. Burgess, D. W. Sun, and B. Tiwari. 2020a. Inactivation efficacy and mechanisms of plasma activated water on bacteria in planktonic state. Journal of applied microbiology 129, 5: 1248–1260. [Google Scholar] [CrossRef]
- Zhao, Y. M., A. Patange, D. W. Sun, and B. Tiwari. 2020b. Plasma-activated water: Physicochemical properties, microbial inactivation mechanisms, factors influencing antimicrobial effectiveness, and applications in the food industry. Comprehensive Reviews in Food Science and Food Safety 19, 6: 3951–3979. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Enumeration of biofilm biomass in PF14 after PAW treatments during 72 h incubation. The results are presented as mean ± SD, (n=3). Different letters indicate significant differences (p ˂ 0.05).
Figure 1.
Enumeration of biofilm biomass in PF14 after PAW treatments during 72 h incubation. The results are presented as mean ± SD, (n=3). Different letters indicate significant differences (p ˂ 0.05).
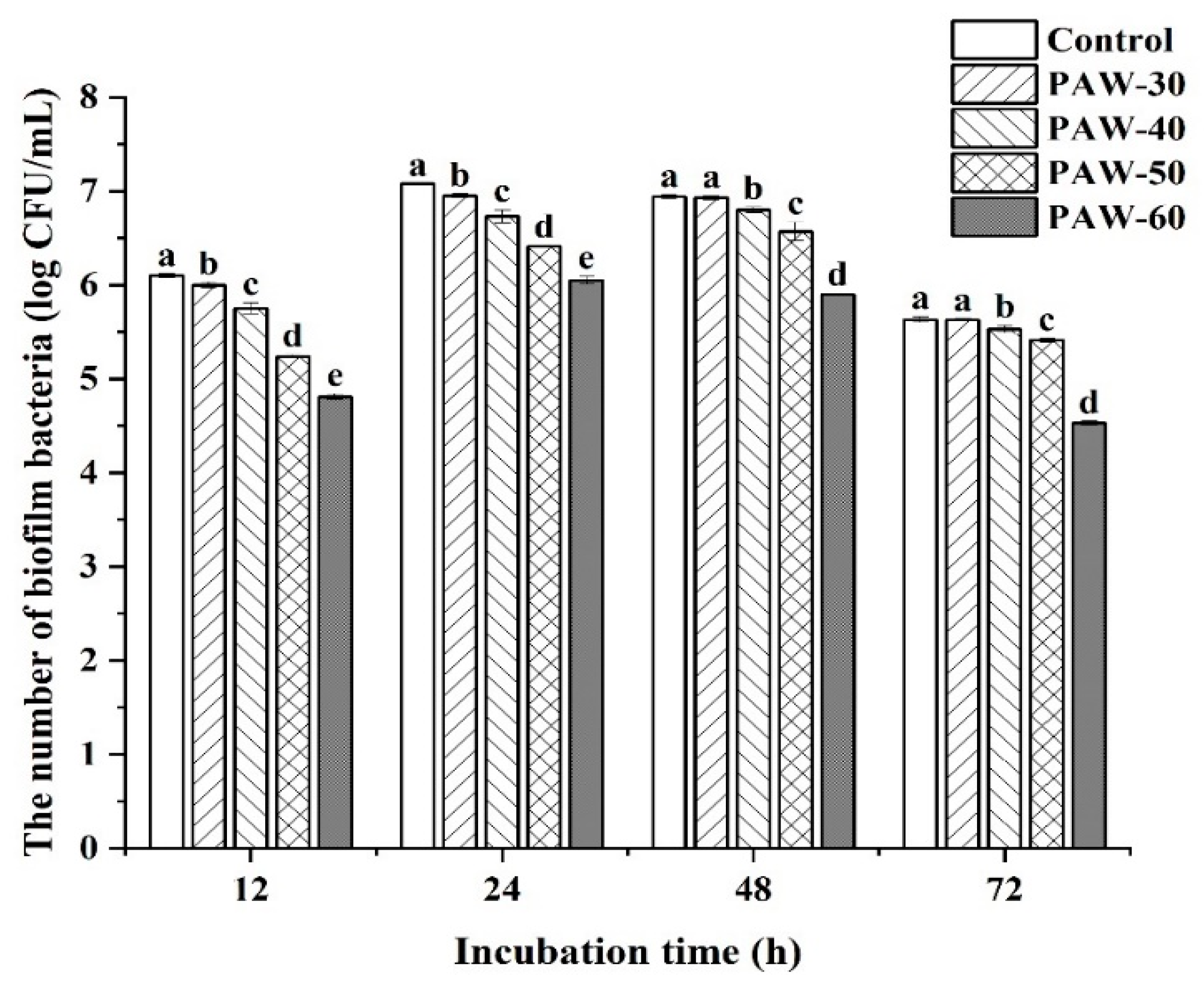
Figure 2.
The effect of PAW on spoilage factors of (a,b) protease and (c,d) siderophore in PF14 during 72 h incubation. The results are presented as mean ± SD, (n=3). Different letters indicate significant differences (p ˂ 0.05).
Figure 2.
The effect of PAW on spoilage factors of (a,b) protease and (c,d) siderophore in PF14 during 72 h incubation. The results are presented as mean ± SD, (n=3). Different letters indicate significant differences (p ˂ 0.05).
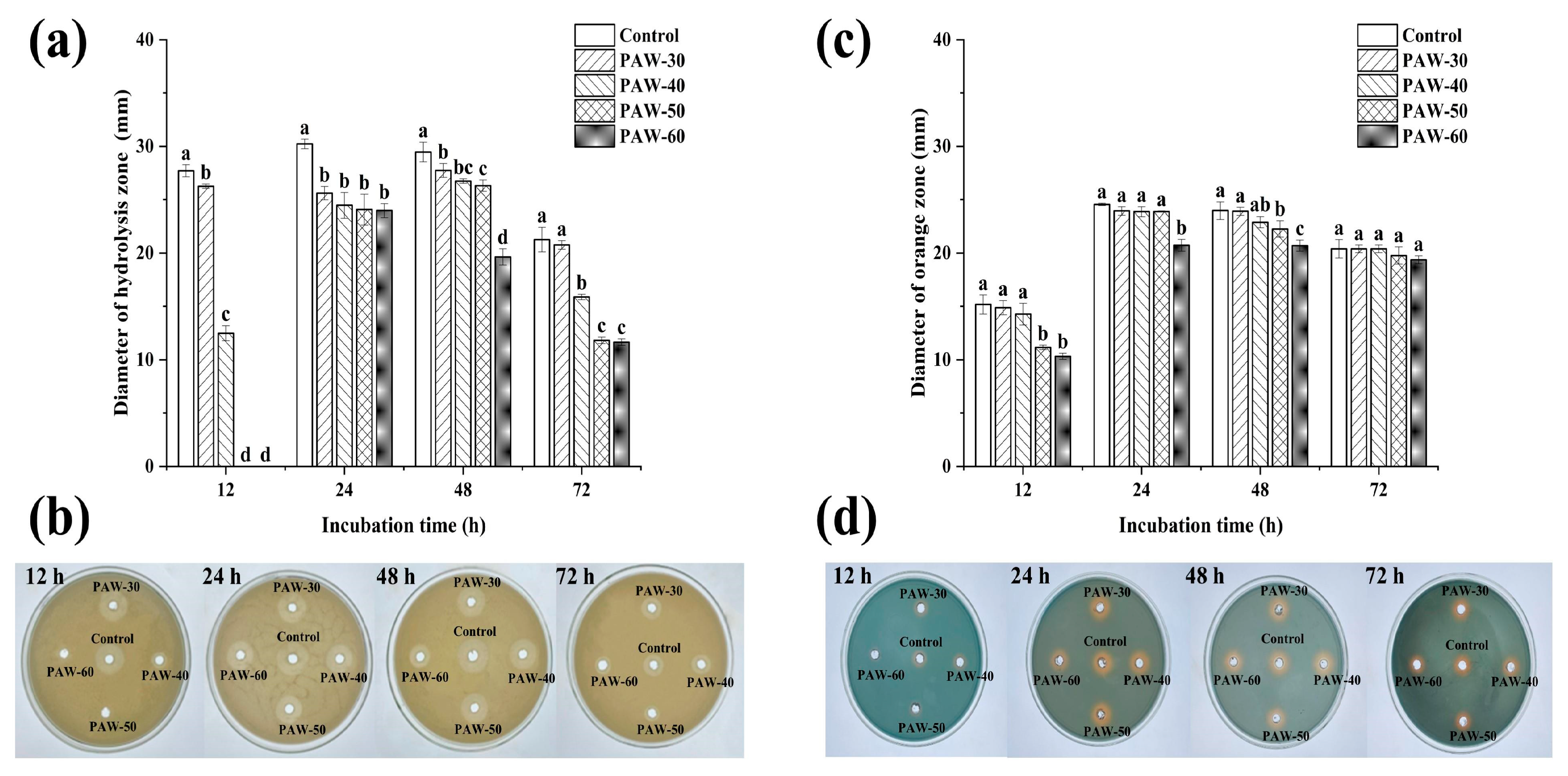
Figure 3.
(a) HPLC chromatogram of standard AHLs and AHLs type in PF14. (b) The effect of PAW on AHLs production of PF14. The results are presented as mean ± SD, (n=3). Different letters indicate significant differences (p ˂ 0.05).
Figure 3.
(a) HPLC chromatogram of standard AHLs and AHLs type in PF14. (b) The effect of PAW on AHLs production of PF14. The results are presented as mean ± SD, (n=3). Different letters indicate significant differences (p ˂ 0.05).
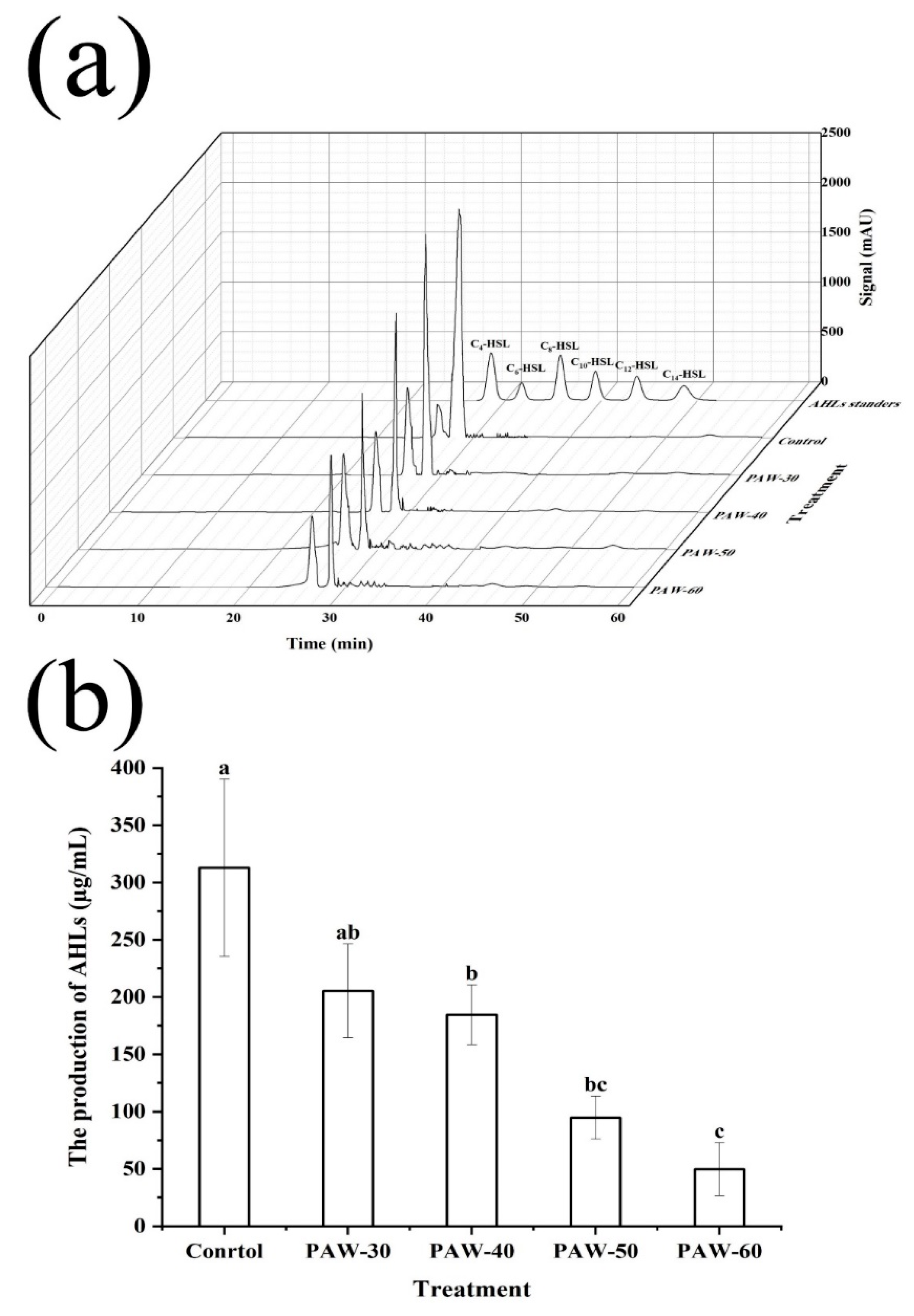
Figure 4.
The effect of PAW on AHLs activity of PF14. The results are presented as mean ± SD, (n=3). Different letters indicate significant differences (p ˂ 0.05).
Figure 4.
The effect of PAW on AHLs activity of PF14. The results are presented as mean ± SD, (n=3). Different letters indicate significant differences (p ˂ 0.05).
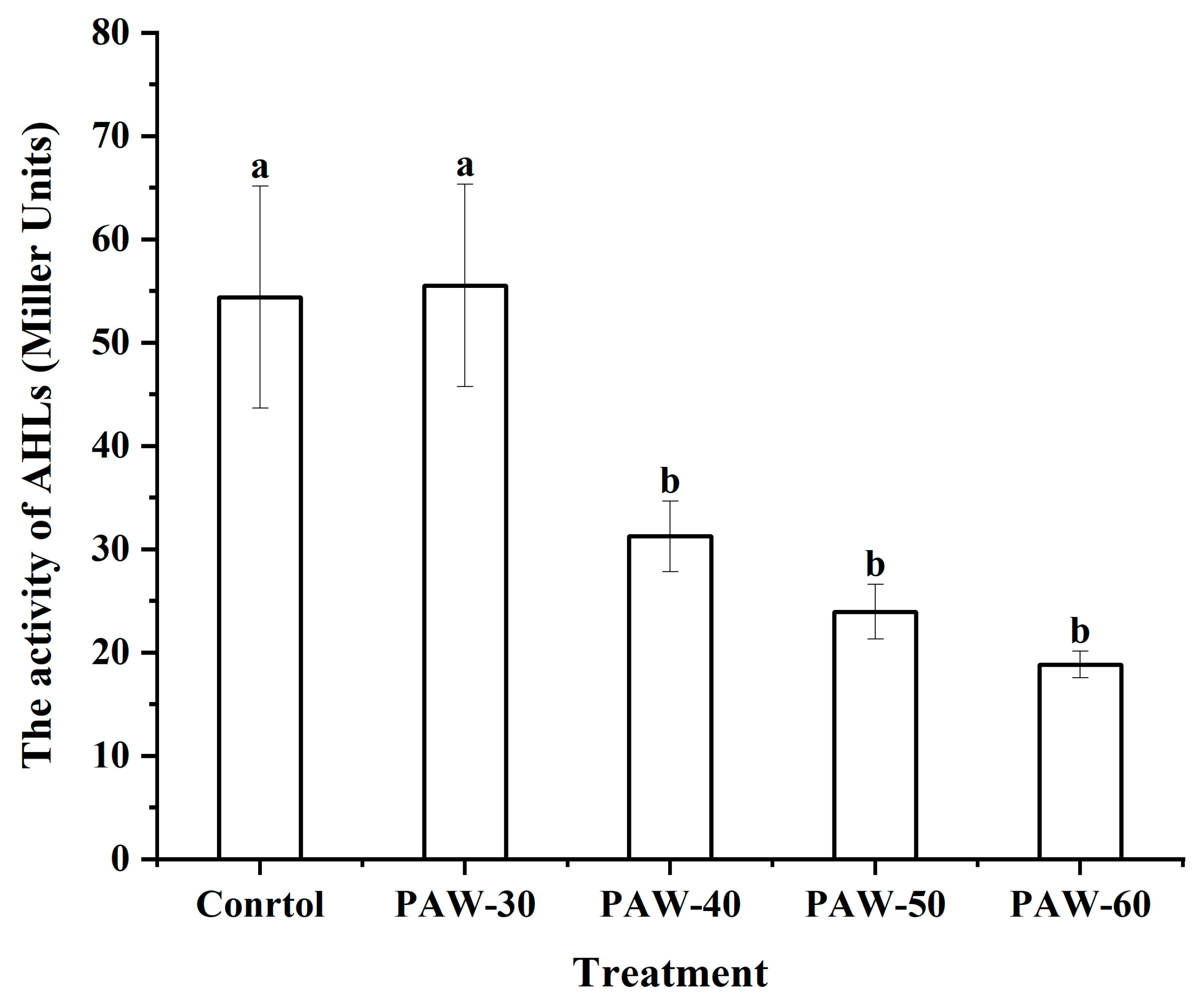
Figure 5.
The FTIR spectra of C4-HSL after PAW treatments.
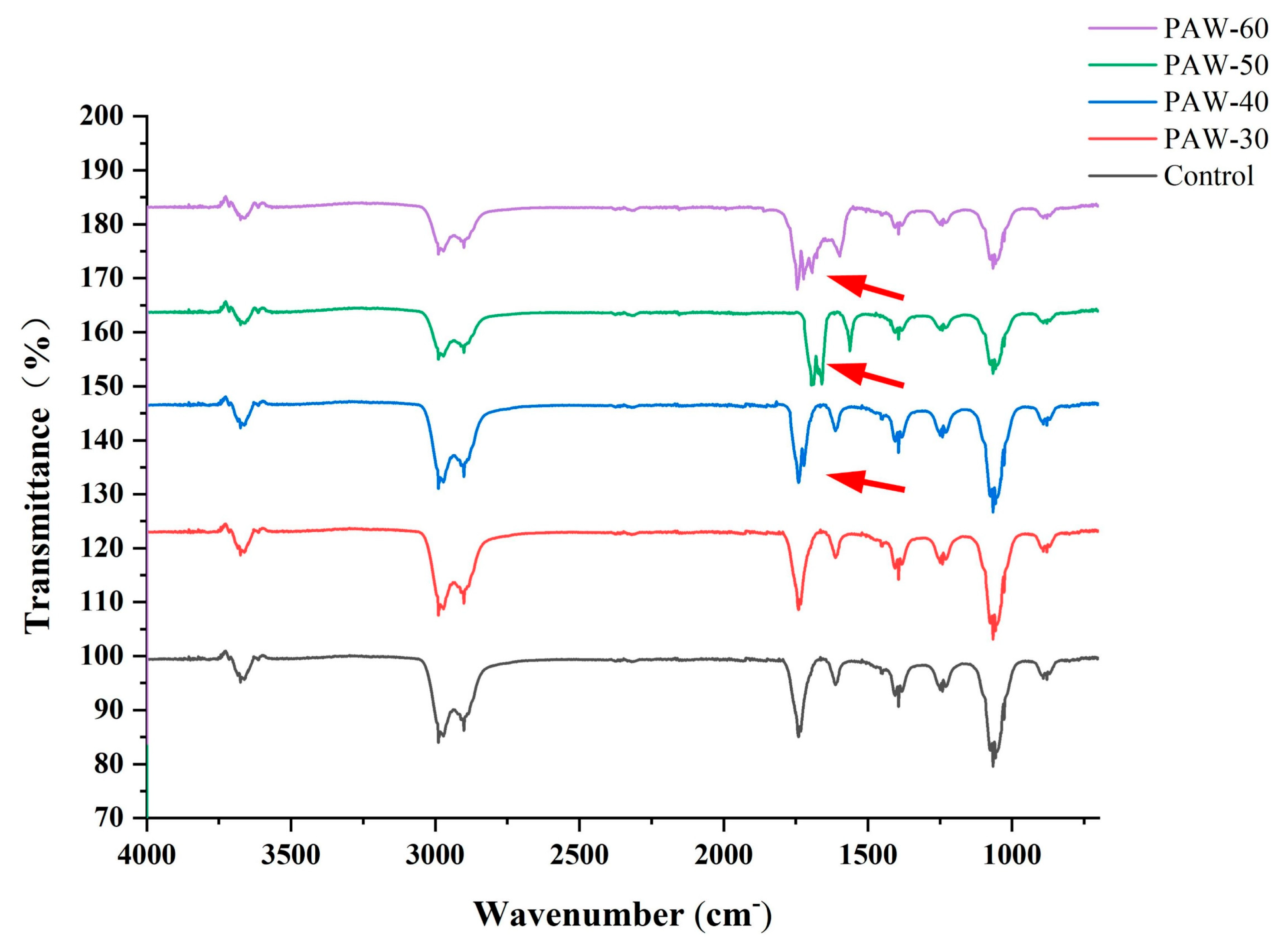
Figure 6.
The effect of exogenous C4-HSL on the biofilm formation of PF14 after PAW-60 treatment. The results are presented as mean ± SD, (n=3). Different letters indicate significant differences (p ˂ 0.05).
Figure 6.
The effect of exogenous C4-HSL on the biofilm formation of PF14 after PAW-60 treatment. The results are presented as mean ± SD, (n=3). Different letters indicate significant differences (p ˂ 0.05).
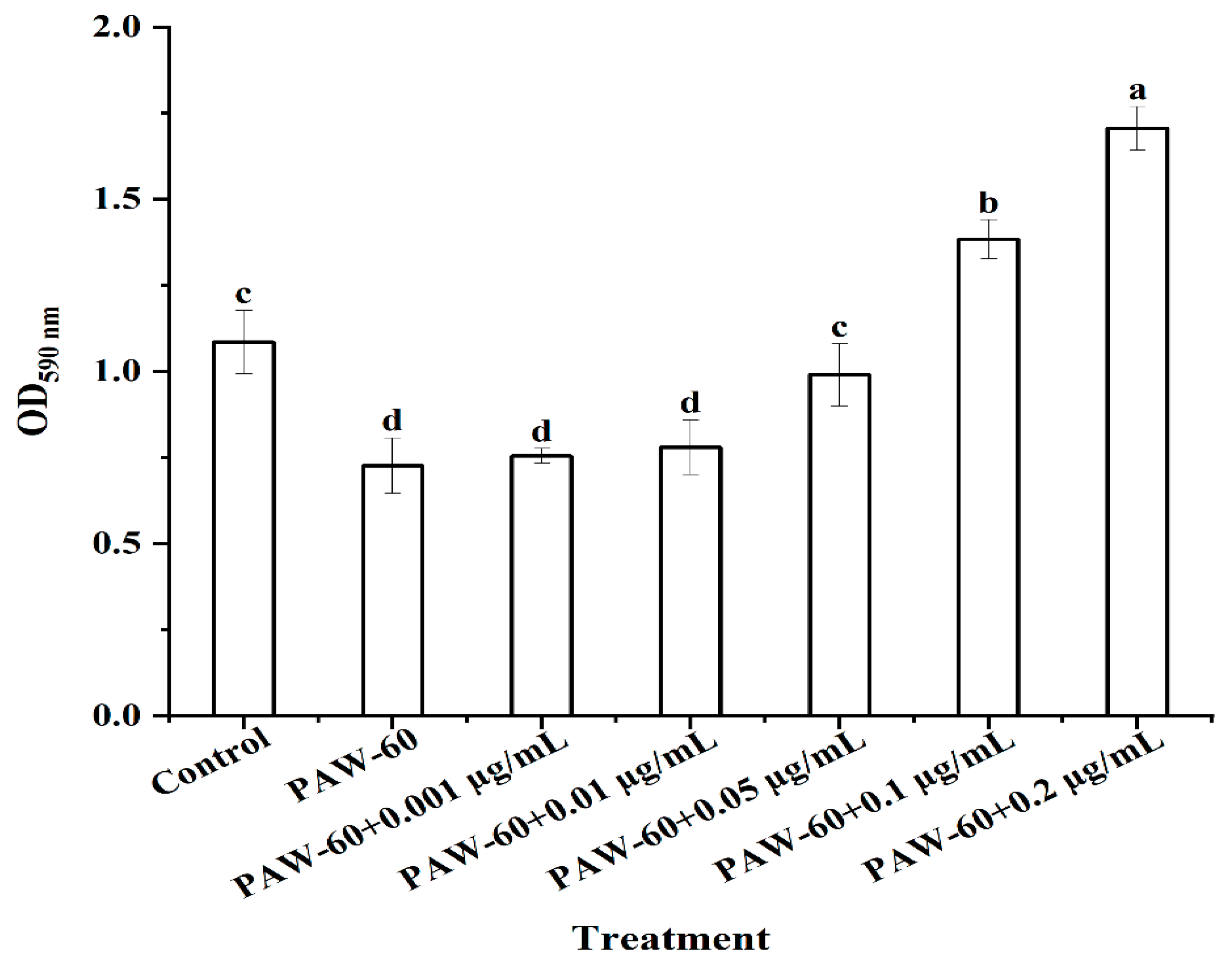
Figure 7.
The effect of exogenous C4-HSL on (a,b) protease and (c,d) siderophile production of PF14 after PAW-60 treatment. The results are presented as mean ± SD, (n=3). Different letters indicate significant differences (p ˂ 0.05).
Figure 7.
The effect of exogenous C4-HSL on (a,b) protease and (c,d) siderophile production of PF14 after PAW-60 treatment. The results are presented as mean ± SD, (n=3). Different letters indicate significant differences (p ˂ 0.05).
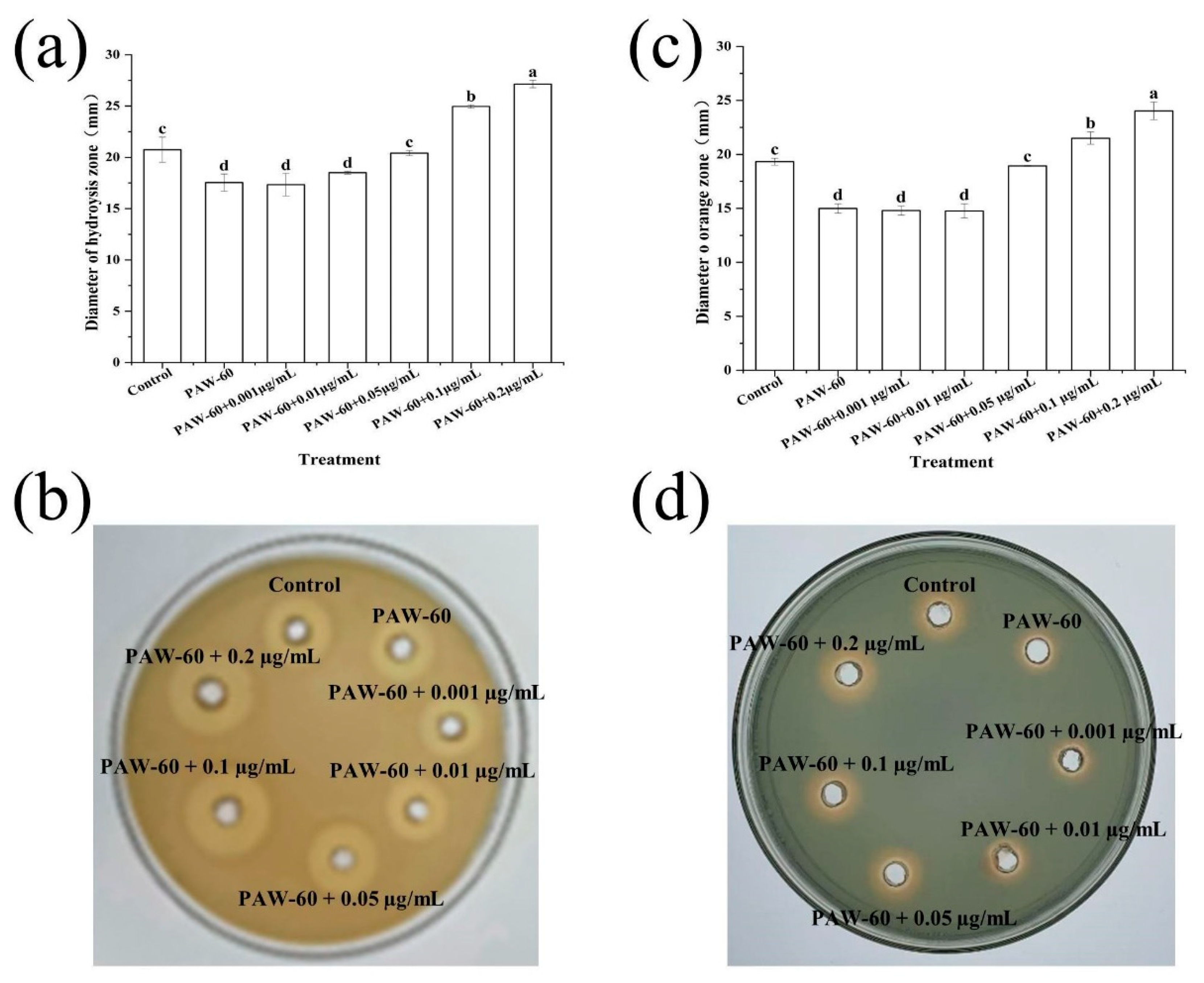
Figure 8.
The effect of exogenous C4-HSL on gene expression levels of PF14 after PAW-60 treatment. The results are presented as mean ± SD, (n=3). Different letters indicate significant differences (p ˂ 0.05).
Figure 8.
The effect of exogenous C4-HSL on gene expression levels of PF14 after PAW-60 treatment. The results are presented as mean ± SD, (n=3). Different letters indicate significant differences (p ˂ 0.05).
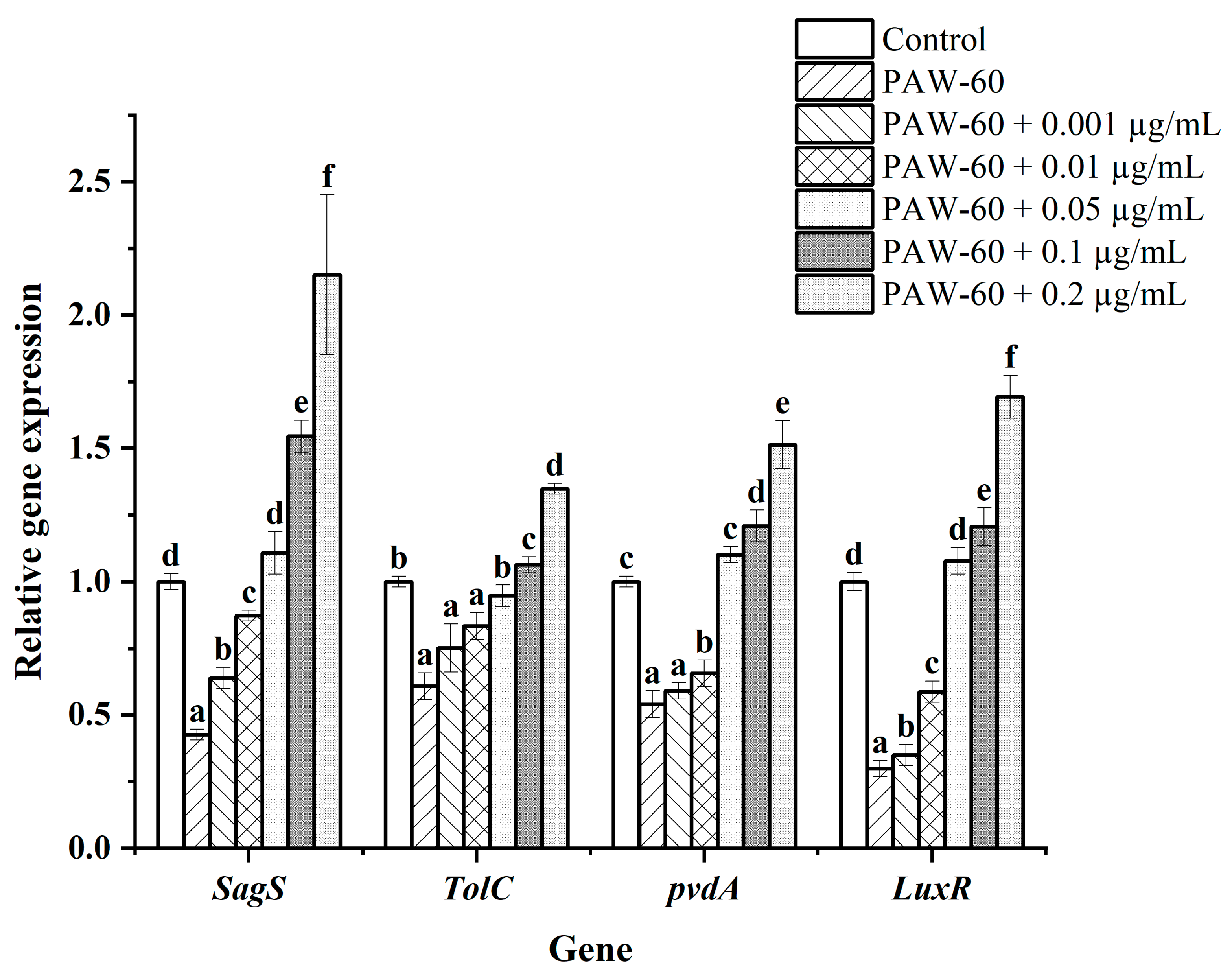
Figure 9.
Molecular docking plot of H2O2, NO2-, and NO3- in PAW with the (a) FadD1 and (b) LuxR.
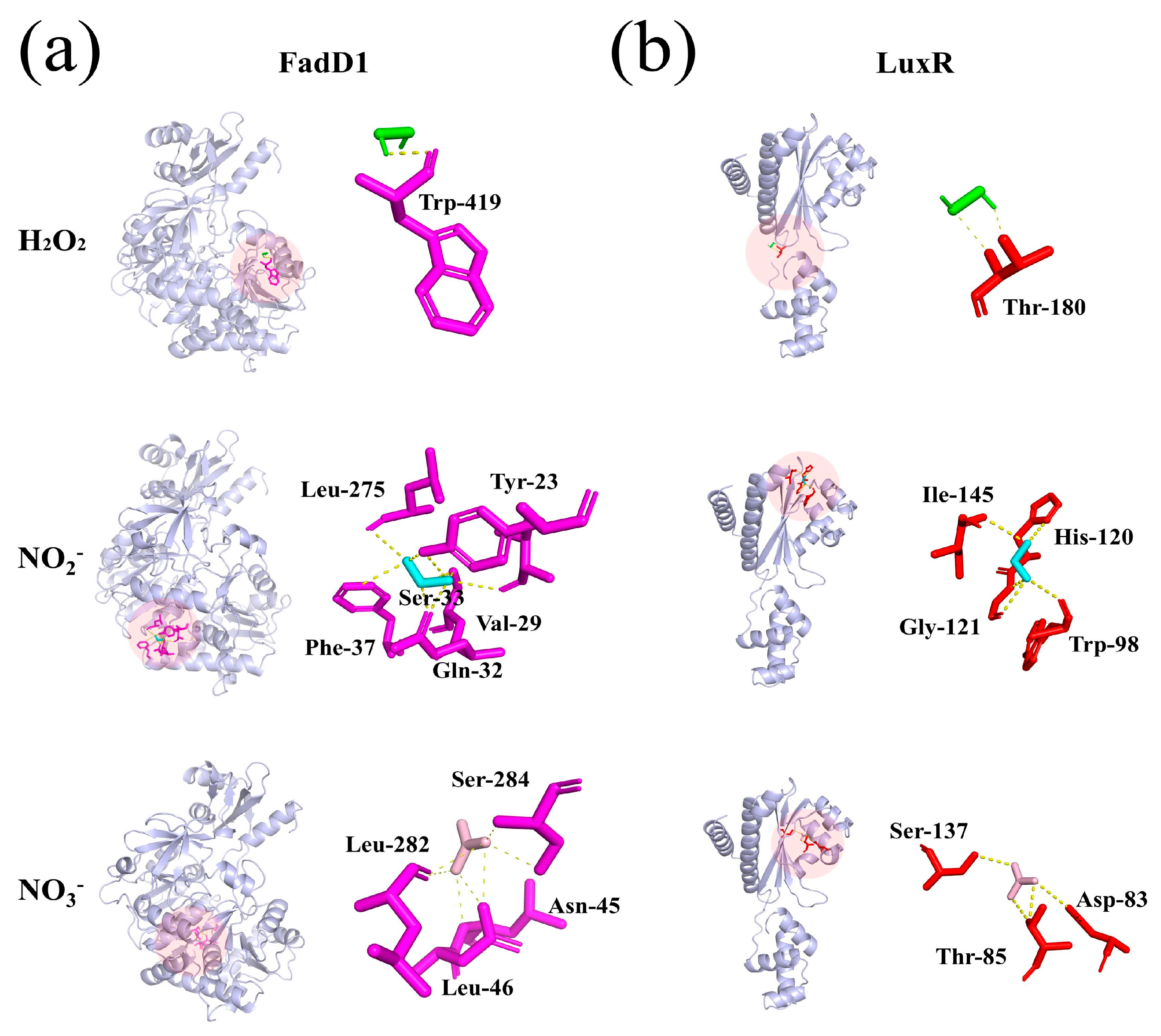
Figure 10.
The effect of PAW on (a) TVC, (b) pH, (c) TBARS, (d) TVB-N, and (e) AHLs production of fish muscle juice during 4 °C storage. The results are presented as mean ± SD, (n=3). Different uppercase letters indicate significant differences after the same treatment at different storage times (p ˂ 0.05), and different lowercase letters indicate significant differences at the same storage time after different treatments (p ˂ 0.05).
Figure 10.
The effect of PAW on (a) TVC, (b) pH, (c) TBARS, (d) TVB-N, and (e) AHLs production of fish muscle juice during 4 °C storage. The results are presented as mean ± SD, (n=3). Different uppercase letters indicate significant differences after the same treatment at different storage times (p ˂ 0.05), and different lowercase letters indicate significant differences at the same storage time after different treatments (p ˂ 0.05).
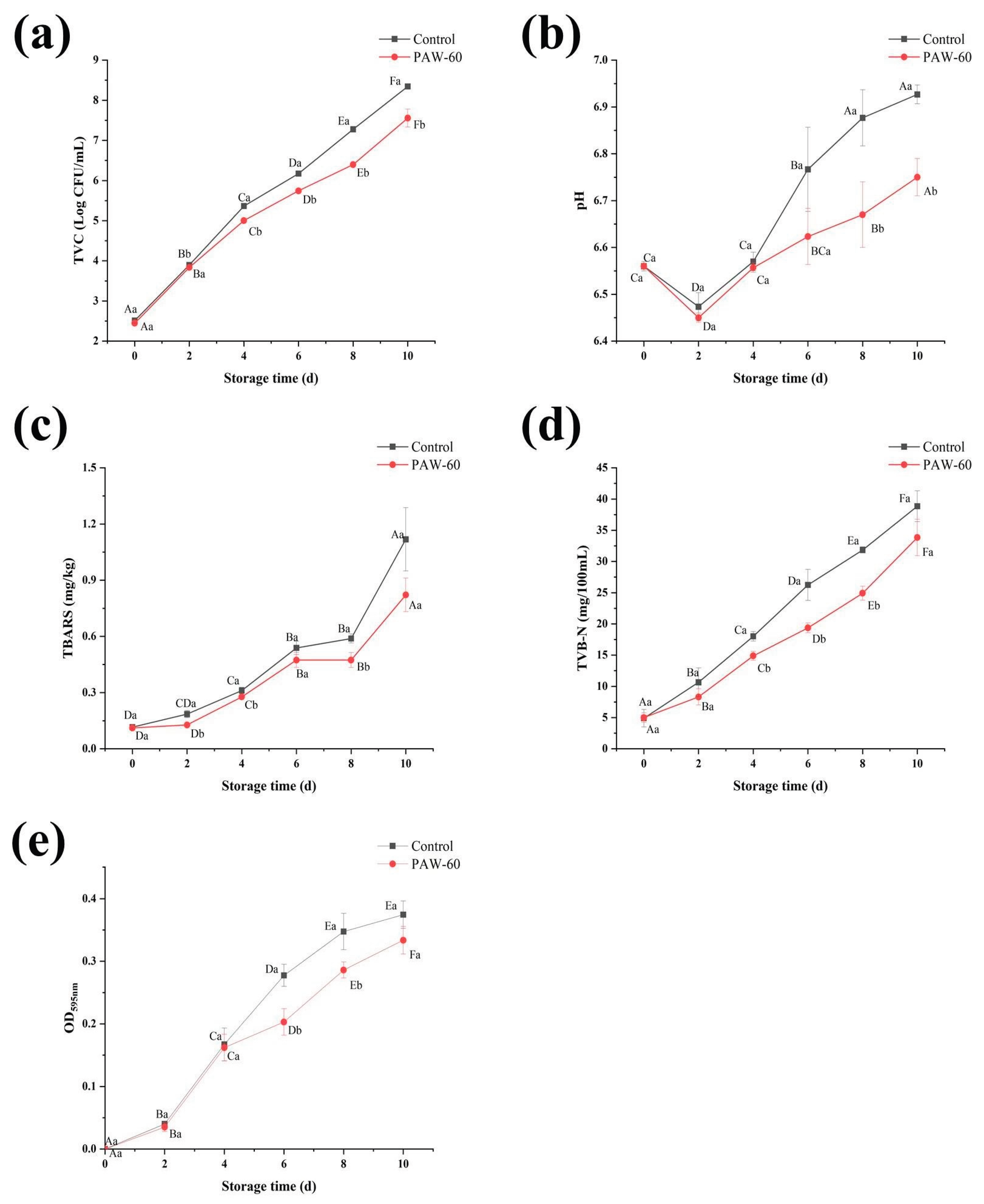

Table 1.
Primer sequences for RT-qPCR.
| Gene | Primer | Sequence (5′-3′) |
|---|---|---|
| 16S rRNA | 16S rRNA-F | GGAATCTGCCTGGTAGTGGG |
| 16S rRNA-R | CAGTTACGGATCGTCGCCTT | |
| SagS | sagS-F | GCTGAACTCGCTCAGGAACT |
| sagS-R | TGGCGCCAAACAGAAAATCG | |
| TolC | TolC-F | AACCGATTTGGTCAGCGTCT |
| TolC-R | CTTGTTCGTTGACGGCTTCG | |
| pvdA | pvdA-F | CCTGGTGACCCAGAGTGAAC |
| pvdA-R | GAGATCACACGCAACGCTTC | |
| LuxR | LuxR-F | GTGCCAACGCTATGCTGAAC |
| LuxR-R | TGCGATCCAAACAATGGCAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



