
Submitted:
22 September 2025
Posted:
23 September 2025
You are already at the latest version
Abstract
Plant-derived wastes are increasingly explored as organic matter sources for sustainable agriculture. Tea waste, a by-product of industrial tea processing, is often regarded as an environmental pollutant, yet its potential for agricultural use remains conditional and requires careful evaluation. This study evaluated the effects of factory-derived tea waste on kale (Brassica oleracea var. acephala) under drought stress. Plants were grown in soils amended with 5% or 10% tea waste and subjected to mild (75% FC) and moderate (50% FC) water deficits, compared with full irrigation (100% FC). Fifteen morphological and physiological parameters were assessed, and data were analyzed using principal component analysis (PCA) and correlation heatmaps to identify trait associations and stress markers. Drought stress significantly reduced all growth and yield traits, with stronger effects under severe water deficit. Tea waste generally exacerbated stress impacts, increasing damage indices, reducing plant height, and lowering chlorophyll values. However, 10% tea waste under non-stress conditions increased plant and root dry weights without negatively affecting other traits, suggesting a partial nutrient contribution. In contrast, 5% tea waste aggravated stress effects, likely due to phenolic and caffeine toxicity. Overall, raw tea waste was unsuitable for kale production under drought conditions. To harness its potential, bioactive compounds must be removed, and the material stabilized through composting or biochar conversion for safe integration into drought-resilient systems.
Keywords:
brassica spp.
; climate resilience
; drought stress
; leafy vegetables
; tea processing waste
; water deficit
1. Introduction
Kale (Brassica oleracea var. acephala) is a widely cultivated leafy vegetable that has gained global attention for its rich phytochemical profile and associated health benefits [1]. As one of the oldest domesticated morphotypes of Brassica oleracea with origins in the Eastern Mediterranean, it remains an important crop worldwide [2]. Traditionally embedded in Northern European cuisines, particularly in winter, kale has recently become popular as a superfood due to its health-promoting nutrient content [3]. Like other Brassicaceae, kale is rich in glucosinolates, including high glucobrassicin levels suitable for human consumption [4]. It provides fiber, potassium, and calcium with higher bioavailability than milk, as well as prebiotic carbohydrates, unsaturated fatty acids, and a range of vitamins, while anti-nutritional factors such as oxalates, tannins, and phytates can occur at relatively high concentrations [5]. Relative tolerance to adverse climates, low production costs, and broad genetic diversity across local populations and cultivars characterized by variation in leaf color, morphology, and flavor -highlight its economic and genetic importance [6,7,8]. Because kale is cultivated in temperate, subtropical, and tropical regions, understanding its resilience to water stress is a global priority. Recent bibliometric work in ornamental plants has also shown increasing research interest in drought stress globally, especially in physiological, molecular, and antioxidant response pathways [9].
Leafy vegetables such as kale are highly sensitive to environmental conditions. Optimal growth occurs at temperatures below 22°C, while temperatures above 25°C induce heat and moisture stress, often resulting in bitterness [10]. Water management is critical, as irrigation frequency and volume strongly influence biomass production, metabolism, and nutritional quality. Kale shows measurable responses to drought (changes in metabolome, leaf physiology), making it a good model leafy vegetable to understand amendment effects under water stress [11]. Stressful conditions such as drought or seasonal heat decrease chlorophyll and carotenoids in kale, while nutrient content also shifts under stress [12]. Drought stress reduces growth, physiological activity, and the accumulation of primary and secondary metabolites, with biomass losses that may exceed 10 percent within two weeks [10,13,14].
Drought and desertification represent major threats to agriculture. Rising evaporative demand coupled with insufficient soil moisture intensifies agricultural and ecological drought, while reduced river flow and surface storage contribute to hydrological drought [15]. Many production regions already face more frequent and severe droughts that depress yields across Africa, Asia, and Europe, with root crops and vegetables particularly vulnerable [16].
Improving soil water-holding capacity is a practical strategy to alleviate drought stress. Soil organic matter enhances aggregate stability, porosity, and infiltration, and organic amendments improve soil structure and microbial activity, thereby increasing resilience to water deficit [17,18]. Higher soil organic carbon reduces bulk density and can raise infiltration rates up to three times, restoring critical soil functions in degraded soils [19]. Plant-derived carbon inputs promote macroaggregate formation through microbial and plant mucilage that bind mineral particles [20]. The use of organic residues as soil conditioners supports circular agriculture by reducing waste, reusing resources, and producing high-value inputs, while also providing safe disposal pathways [21,22,23]. Studies on leafy vegetables illustrate that drought reduces yield and nutrient content, but that amendment (or fertility) can modulate these effects [24]. However, the behavior of organic amendments varies with composition and interactions with soil minerals, requiring crop- and site-specific evaluation [25].
Among organic residues with agronomic potential, tea waste is especially abundant and underutilized. As the world’s most popular drink, tea continues to experience growth in production. Tea waste, a byproduct of black tea processing, accumulates in large volumes at factories where landfill disposal or incineration creates environmental problems. In 2020, global tea consumption was about 6.3 million metric tons and is expected to reach 7.4 million metric tons by 2025, highlighting the scale of residues [26]. Compositionally similar to processed tea, tea waste contains structural proteins, lignin, cellulose, hemicellulose, secondary metabolites, and minerals. It also contains bioactive/allelochemical compounds (e.g. caffeine, polyphenols) that may have beneficial or inhibitory effects depending on concentration [27]. Some studies report phytotoxic effects of tea waste (or its derivatives) on seed germination, indicating potential limitations if used at high rates or without proper treatment [28]. Incorporating tea waste into soil has been shown to improve physical and biological properties, stimulate plant growth and yield, and enhance tolerance to certain abiotic stresses [29,30,31,32,33]. Nevertheless, very few studies have evaluated its effects on leafy vegetables, and no research has specifically addressed its role under drought conditions. This knowledge gap is particularly relevant given the dual challenges of crop productivity and waste management in modern agriculture. No prior study has evaluated the combined effects of tea waste amendment and drought stress in kale cultivation, representing a critical knowledge gap.
This study evaluates the effects of tea waste amendments on kale under controlled water-deficit conditions. By testing different incorporation levels and irrigation regimes, we aimed to assess whether tea waste can be used as a sustainable soil amendment to improve drought resilience in kale cultivation.
2. Materials and Methods
2.1. Plant Material
Two kale (Brassica oleracea var. acephala) cultivars were used: Karadeniz Yaprak (Naz Seed Production Agricultural Production Livestock Industry and Trade Ltd. Co., Türkiye; VK1) and КЕЙЛ (Semenabulgaria Company, Plovdiv, Bulgaria; VK2). Karadeniz Yaprak has flat, broad, waxy leaves, whereas КЕЙЛ is curly-leaved with smaller, serrated blades.
2.2. Tea Waste and Soil Characteristics
Tea waste was obtained from a private tea processing factory in Rize Province, Turkey. This material, consisting of leaf residues, fibers, and fine particles excluded during the processing of green tea into black tea, was left to decompose naturally under outdoor conditions for eight months at the Faculty of Agriculture, Recep Tayyip Erdoğan University. Representative image is presented in Figure 1, and its basic properties are summarized in Table 1. Soil characteristics used for the experiment are shown in Table 2.
2.3. Seedling Production
Seeds were sown in trays filled with a 1:1 (v/v) peat–perlite mixture to produce seedlings per cultivar. A balanced NPK fertilizer (20–20–20) was applied during early growth. At the 3–4 true leaf stage, seedlings were transplanted into pots containing the experimental soil–tea waste mixtures, with one seedling planted per pot.
2.4. Pot Experiment and Drought Stress Application
Soil and tea waste mixtures were prepared at 0% (TW0, control), 5% (TW1), and 10% (TW2) tea waste (w/w) and filled into 2 L polyethylene pots (17 × 13.3 cm). One seedling was planted per pot. Drought stress was applied two weeks after transplanting following the method of [34]. The field capacity of the pots was determined gravimetrically by saturating and draining two randomly selected pots per treatment. Irrigation regimes were imposed as follows:
DS0: 100% field capacity (FC) (well-watered control)
DS1: 75% of FC (mild stress)
DS2: 50% of FC (moderate stress)
A randomized block design was employed, with five replicates per treatment and five plants in each replicate. Drought treatments were maintained for 21 days, with irrigation based on the water requirements of the control group. The factorial treatment structure, including the two kale varieties, three tea waste levels, and three drought stress regimes, is summarized in Table 3, while a schematic representation of the experimental layout is provided in Figure 2.
2.5. Observations and Measurements
Following 21 days of stress exposure, trait evaluation was conducted. Damage index (0–5): Visual scoring of drought damage following by [35,36]. The point values and corresponding morphological appearances are as follows. 0: Plants are not affected by drought stress at all, 1: Slowdown in growth, 2: Beginning of wilting in lower leaves, 3: Curling and wilting in upper leaves, 4: Severe wilting and yellowing of leaves, beginning of drying at leaf edges, 5: Wilting of the plant and drying of lower leaves.
Measurements of plant height (cm) extended from the root collar up to the shoot apex. Leaf thickness (mm) was obtained from the fourth true leaf using a digital caliper (0.01 mm precision). Petiole length (cm) was taken from the leaf base to the blade. Leaf number was counted as the total number of true leaves per plant. Chlorophyll index was assessed with a SPAD-502 chlorophyll meter (Konica Minolta, Japan) on the third true leaf of three plants per replicate, and mean values were calculated.
Average leaf weight (g) was derived from three leaves per replicate weighed on a precision scale. Leaf area (cm²) was quantified from scanned images (HP Scanjet G2410) using WinDIAS 3.2 software. Leaf water content (RWC, %) was calculated according to Smart and Bingham [37] using fresh, turgid, and dry weights (samples dried at 85 °C for 24 h).
For biomass parameters, shoot fresh weight (g plant⁻¹) was determined by harvesting and weighing aboveground biomass, while shoot dry weight (g plant⁻¹) was obtained after drying the same samples at 65 °C to constant weight. Root length (cm) was taken after washing roots, and root diameter (cm) was read with a digital caliper. Root fresh weight (g plant⁻¹) was recorded immediately after washing, and root dry weight (g plant⁻¹) was determined after drying samples at 65 °C to constant weight.
2.6. Statistical Analyses
Data were analyzed using JMP Pro 13.0 software. Analysis of variance (ANOVA) was performed, and mean separations were conducted using the least significant difference (LSD) test. Principal component analysis (PCA) was performed in RStudio to identify major sources of variation, and correlation patterns were visualized using polar heatmaps and hierarchical clustering.
3. Results
The statistical evaluation of the greenhouse pot experiment, which investigated plant growth and yield under drought stress, is summarized in Table 4. Variance analysis revealed significant effects for damage index, plant height, leaf thickness, leaf length, leaf number, chlorophyll content, leaf weight, leaf area, leaf water content, root length, and root dry weight. These differences were associated with variety, tea waste, drought stress, and their interactions. Fresh shoot weight showed no significant response to the variety × tea waste × drought stress interaction, nor to the variety × drought stress interaction; however, both the variety × tea waste and tea waste × drought Stress interactions were significant. For dry weight, significant effects were observed for variety, tea waste, and drought stress, with additional significance in the variety × tea waste, tea waste × drought stress, and three-way interactions, while the variety × drought stress effect remained nonsignificant. Root diameter was significantly affected by most factors, except for the three-way interaction. In contrast, root dry weight was unaffected by the variety × drought stress and three-way interactions, but significant differences emerged for the variety × tea waste and tea waste × drought stress combinations. Detailed results for all traits and interactions are provided in Table 4.
Damage index. The three-way interaction showed that VK1 responded similarly under DS2 in both TW1 and TW2, whereas VK2 responded consistently to DS1 across all media (Table 5). In variety × tea waste, damage was higher in TW2 for VK1 and in TW1 for VK2 (Table 6). In variety × drought stress, controls of both cultivars grouped together, and DS2 produced the strongest effect in VK2 (Table 7). In tea waste × drought stress, DS2 increased damage in every medium, and higher tea waste aggravated symptoms (Table 8).
Plant height. VK1 was most reduced by DS1–DS2 in TW1, while VK2 was more sensitive to DS1–DS2 in TW0 (Table 5). With variety × tea waste, both cultivars were taller in TW2, peaking in VK2 (Table 6). In variety × drought stress, control plants again grouped together and DS2 had the largest impact on VK2 (Table 7). Tea waste × drought stress showed DS2 depressed height in all media and greater tea waste further reduced growth (Table 8).
Leaf thickness. VK1 declined under DS1–DS2 in TW1 and TW2, whereas VK2 was less affected overall; both were most impacted in TW2 with DS2 (Table 5). In variety × tea waste, VK1 was thicker in TW0, whereas VK2 was thicker in TW0 as well (Table 6). In variety × drought stress, DS1–DS2 grouped for VK1 and DS0–DS1 for VK2 (Table 7). No differences appeared in TW1 across irrigations in tea waste × drought stress (Table 8).
Petiole length. Longest values occurred in VK1 under TW0–DS0 and in VK2 under TW2–DS0; shortest in VK1 under TW2–DS2 and in VK2 under TW1–DS1 (Table 5). Tea waste shortened petioles in VK1 but lengthened them in VK2 (Table 6). VK1 showed similar DS1–DS2 values, while VK2 decreased with increasing stress (Table 7). Under Tea Waste × Drought Stress, petioles shortened with stress, yet 10% tea waste increased length in DS0, giving the maximum in TW2–DS0 (Table 8).
Leaf number. VK1 was unchanged in TW0 and TW2 relative to control, and VK2 remained stable in TW1 under DS1 (Table 5). With variety × tea waste, VK1 held steady in TW2, while VK2 was higher in TW1–TW2 (Table 6). Both cultivars declined under stress, with VK1 grouping at DS1–DS2 and VK2 showing the steepest drop at DS2 (Table 7). DS2 reduced leaf number in all media and higher tea waste did not intensify this decline (Table 8).
Chlorophyll (SPAD). Values rose under DS2 in TW1–TW2 for both cultivars (Table 5). Without tea waste the highest SPAD was recorded, indicating reductions with tea waste addition (Table 6). Controls had higher SPAD in both varieties; VK2 grouped at DS1–DS2, and VK1 peaked at DS2 (Table 7). The highest SPAD appeared in TW0, yet stress combined with tea waste tended to raise SPAD relative to stressed soils without tea waste (Table 8).
Leaf weight. VK1 peaked at TW2–DS0 and VK2 at TW1–DS0 (Table 5). TW2 produced the greatest weights in both cultivars (Table 6). Controls were highest and weights declined with stress (Table 7). Stress reduced leaf weight in all media, while 10% tea waste improved weight under DS0 (Table 8).
Leaf area. VK1 was maximal at TW0–DS0 and TW2–DS0, VK2 at TW1–DS0 (Table 5). TW2 enlarged leaf area in both cultivars; TW1 decreased it in VK1 but increased it in VK2 (Table 6). Area decreased progressively with stress (Table 7). Ten percent tea waste increased area only under DS0 and did not offset drought reductions (Table 8).
Leaf water content. Highest values occurred in DS0 for both cultivars, with VK2 maintaining relatively higher values under DS1–DS2 in TW1 (Table 5). VK1 was lowest in TW0, whereas VK2 reached a maximum in TW2 (Table 6). Water content declined as stress intensified, with VK2 highest at DS0 (Table 7). DS2 lowered water content in all media and tea waste amplified the stress effect (Table 8).
Shoot fresh weight. The three-way interaction was not significant (Table 5). VK1 showed similar values in TW0 and TW2, while VK2 peaked in TW2 (Table 6). Fresh weight decreased with stress in both cultivars, with a nonsignificant variety × drought stress interaction (Table 7). Under tea waste × drought stress, fresh weight declined with stress; 10% tea waste did not reduce DS0 values, which grouped with control (Table 8).
Shoot dry weight. Drought reduced biomass in both cultivars. VK1 was highest at DS0 in TW0–TW2; VK2 peaked at TW2–DS0 (Table 5). Dry weight increased with tea waste level, maximizing at TW2 (Table 6). Values fell with stress, although the variety × drought stress interaction was not significant (Table 7). Drought reduced dry weight in all media, and TW2 did not depress biomass under DS0 (Table 8).
Root length. Drought reduced length in both cultivars (Table 5). Tea waste shortened roots in VK1 but 5% tea waste lengthened them in VK2 (Table 6). Both cultivars were longer at DS0, with VK2 more stress-sensitive (Table 7). Ten percent tea waste increased root length only without stress (Table 8).
Root diameter. No significant three-way interaction (Table 5). VK1 was greatest in TW0, VK2 in TW1 (Table 6). Diameter decreased with stress, especially in VK2 (Table 7). DS2 reduced diameter across treatments with similar effects in TW0 and TW2 (Table 8).
Root fresh weight. VK1 performed best at TW0–DS0; VK2 showed no clear pattern (Table 5). VK1 was highest in TW0 and lowest in TW1; VK2 was similar in TW0–TW1 (Table 6). Fresh weight declined with stress and VK2 was more affected (Table 7). DS2 reduced root fresh weight in all media and tea waste negatively influenced VK1 (Table 8).
Root dry weight. The three-way interaction was not significant (Table 5). Drought reduced root dry weight in both cultivars; VK1 maintained higher values at TW0–DS0 and VK2 showed no distinct response (Table 6). Values decreased with stress without a significant variety × drought stress effect (Table 7). DS2 reduced root dry weight in all media, while 10% tea waste increased it under DS0 (Table 8).
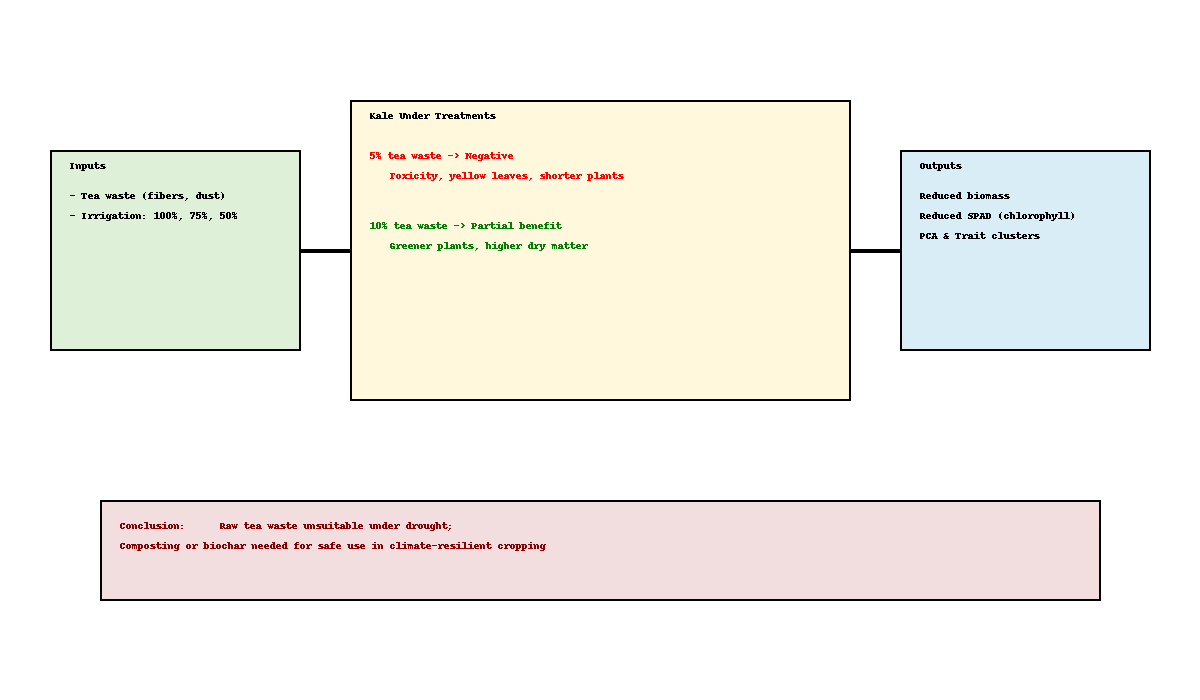
To assess the reliability of the parameters used to evaluate the effects of mild (DS1) and moderate (DS2) drought stress in soils amended with tea waste, a principal component analysis (PCA) was conducted. Multivariate analyses highlighted distinct patterns among morphological and physiological traits of kale under tea waste and drought stress. The polar heatmap with dendrogram (Figure 3) revealed two major trait clusters. The analysis showed that the 15 traits clustered into a two-factor structure, explaining 78.3% of the total variance. The first factor accounted for 65.4% and the second for 12.9%, confirming the validity of the factorial design in explaining treatment effects. Growth-related parameters such as plant height (PH), leaf area (LA), leaf weight (LW), plant fresh weight (PFW), plant dry weight (PDW), root dry weight (RDW), and root diameter (RD) grouped together, indicating their strong interdependence in determining biomass production. In contrast, relative water content (RWC), leaf length (LL), leaf number (LN), leaf thickness (LT), and root fresh weight (RFW) formed a separate cluster, reflecting physiological adaptation under stress. The damage index (DI) and chlorophyll content (SPAD) were associated as a distinct pair, underscoring their role as stress indicators.
The correlation heatmap (Figure 4) supported these findings, showing strong positive associations (>0.90) among PH, LA, LW, PFW, and PDW, and similarly strong correlations between root traits and aboveground biomass. RWC was moderately correlated with LL and LN, confirming their functional linkage observed in the dendrogram. DI exhibited strong negative correlations with nearly all growth and biomass traits, validating its role as a direct measure of stress impact. SPAD, however, displayed only weak associations with growth traits, clustering instead with LT and LN as part of a physiological response group.
4. Discussion
4.1. Drought Stress Effects on Kale Growth and Biomass
Drought stress causes significant biomass loss in kale [10,24], and our findings are consistent with these reports. In our study, both mild (75% FC) and moderate (50% FC) deficits reduced fresh and dry biomass, with Karadeniz Yaprak less affected than КЕЙЛ, though both suffered yield losses. Even mild drought therefore poses economic risks in kale cultivation. In contrast, Barickman et al. [14] reported a 22.5% increase in fresh weight when comparing kale grown under higher irrigation thresholds (0.35 vs. 0.15 m³m³VWC). The discrepancy likely reflects differences in experimental design and, importantly, the genotypes tested. Indeed, previous studies have reported that some kale cultivars may be more drought tolerant while others are more sensitive [11,38], which may explain the different biomass responses observed across studies.
4.2. Multivariate Assessment of Drought, Tea Waste, and Varietal Responses (PCA)
In our study, PCA revealed two main trait clusters that explained 78.3% of the total variance. Biomass-related traits grouped together, while water relations and leaf traits formed a second cluster, and SPAD with damage index acted as stress indicators. Similar clustering has been reported in Brassica oleracea, where PCA distinguished tolerant from sensitive accessions [38], and in kale, where root traits and osmolytes contributed to resilience under combined stress [11]. Comparable results in wheat also confirm PCA as a robust tool for detecting interdependent traits and identifying reliable markers of drought tolerance [39].
4.3. Physiological and Biochemical Responses Under Drought and Tea Waste Amendment
Drought stress alters pigments and secondary metabolism in Brassica crops. Barickman et al. [14] found reduced neoxanthin and anteraxanthin but increased glucobenzene, progoitrin, and total phenolics under water deficit. In our study, tea waste under drought exacerbated these negative effects, consistent with reports that unprocessed tea residues can be phytotoxic due to phenolics and caffeine [40,41]. By contrast, under full irrigation, 10% tea waste increased dry weight, likely due to nutrient supply [31], whereas 5% was insufficient to offset toxicity, reducing chlorophyll stability. This aligns with Ekbiç et al. [42], who recommend composting to reduce phytotoxicity. Reviews further confirm that tea waste requires processing for safe agronomic use [26,27].
4.4. Trait Interrelationships Revealed by Correlation Heatmap
Correlation analysis showed strong positive associations among growth and biomass traits, reflecting their coordinated role in yield formation. RWC correlated moderately with leaf number and length, while the damage index was negatively associated with nearly all growth traits, confirming its reliability as a stress marker. SPAD was weakly related to biomass, clustering instead with leaf thickness and number, suggesting a separate physiological response. Similar trait interdependencies have been reported in Brassica oleracea [11,38] and other Brassica species, where pigments often decouple from biomass under drought [43,44].
4.5. Role of Organic Amendments in Mitigating Drought Stress
Organic matter contributes to improved soil aggregation, enhances nutrient cycling, and increases water-holding capacity, thereby supporting plant tolerance to drought. Biochar, vermicompost, and rice husks have shown beneficial effects [45]. In kale, biochar substitution for organic fertilizers improved yield and soil fertility [46], and farm-waste compost enhanced biomass and chlorophyll content [47]. In our study, tea waste acted as a dose-dependent amendment: 10% partly mitigated drought stress by buffering toxicity and stabilizing pH [48,49], whereas 5% aggravated stress, likely due to exposure to allelochemicals.
4.6. Composition and Variability of Tea Waste
Tea waste composition varies with cultivar, processing, harvest, and season [50]. It contains catechins, gallic acid, caffeine, and other polyphenols, which contribute to strong antioxidant activity [40,41] but may act as phytotoxins in soils. This variability helps explain why low application aggravated stress, while 10% partly buffered toxicity through added organic matter and nutrient release. Thus, the agronomic performance of tea waste is highly context-dependent, reflecting both composition and application rate.
4.7. Valorization Potential of Tea Waste
As the world’s most consumed beverage, tea generates substantial processing by-products that pose environmental concerns while offering opportunities for resource utilization [27]. Processing is essential to reduce toxicity and unlock agronomic value. Yıldırım et al. [30] showed that composted tea waste enriched with organic fertilizers enhanced seedling growth and chlorophyll in maize. Similarly, Kang et al. [46] and Thepsilvisut et al. [47] reported positive effects of biochar and compost in kale. These findings support the view that raw factory-derived tea waste is unsuitable under drought stress, but composting or biochar conversion can transform it into a valuable soil conditioner within sustainable agriculture.
4.8. Comparative Stress Physiology with Other Crops
Species respond differently to drought. For example, Cebeci [51] compared eggplant genotypes with their wild relatives under gradually increased drought stress and found that wild relatives often retained higher biomass, leaf water content, and superior physiological traits under 50–75% water deficit, highlighting both intra- and inter-species variation in drought tolerance. Moreover, Kıran and Baysal Furtana [34] reported that eggplant exposed to combined drought and salinity exhibited increased chlorophyll and antioxidant activity, whereas in our study kale subjected to drought with tea waste showed reduced pigment stability. Similarly, Azotobacter inoculation in eggplant improved antioxidant enzymes and proline accumulation, thereby buffering oxidative damage [52]. Collectively, these comparisons suggest that while unprocessed tea waste may intensify stress in kale, biostimulants can mitigate drought injury in other crops, underscoring the importance of species- and amendment-specific responses.
4.9. Agronomic Potential and Limitations of Tea Waste Under Drought Stress
This study highlights both the promise and the challenges of using factory-derived tea waste to enhance kale performance under drought stress. While higher application rates partly buffered drought effects, raw residues generally aggravated stress, underscoring the risks of direct use. Varietal differences (Karadeniz Yaprak vs. КЕЙЛ) highlight the role of genetic diversity, consistent with findings that kale accessions differ in stress physiology [11] and that B. oleracea genotypes can be screened using indices such as STI [38]. Collectively, these results indicate that future breeding and management strategies must combine genotype selection with safe processing of agro-industrial residues to enhance climate resilience in kale production.
5. Conclusions
Climate projections indicate that drought stress will increasingly threaten agricultural production, with kale cultivation particularly vulnerable to irregular rainfall and water scarcity. Our findings show that even mild drought stress caused severe reductions in growth and yield. Tea waste as a soil amendment had a dose-dependent effect: a low dose (5%) aggravated stress due to phenolic compounds, whereas a higher dose (10%) improved dry matter accumulation and partially alleviated drought effects through its mineral contribution. Thus, raw factory-derived tea waste is unsuitable for direct use under stress conditions, but when applied at higher rates or after processing, it can provide agronomic benefits.
Tea waste has significant value as an agro-industrial by-product, yet its safe recycling requires processing (e.g., composting or biochar conversion). In practice, future research should: (i) evaluate diverse kale genotypes under drought × amendment interactions, (ii) employ integrative indices such as the Stress Tolerance Index (STI), (iii) select for agronomic and physiological traits including chlorophyll stability and antioxidant activity, and (iv) combine breeding efforts with sustainable management practices to enhance resilience in real-world production systems.
Author Contributions
Conceptualization, H.F.B.; formal analysis, H.F.B.; funding acquisition, H.F.B.; investigation, A.O. H.F.B.; project administration, H.F.B.; resources, A.O.; supervision, H.F.B.; writing—original draft, H.F.B.; writing—review and editing, H.F.B. All authors have read and agreed to the published version of the manuscript.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Šamec, D.; Urlić, B.; Salopek-Sondi, B. Kale (Brassica oleracea var. acephala) as a superfood: Review of the scientific evidence behind the statement. Crit. Rev. Food Sci. 2019, 59, 2411–2422. [Google Scholar]
- Balkaya, A.; Yanmaz, R. Promising kale (Brassica oleracea var. acephala) populations from Black Sea region, Turkey. N. Z. J. Crop Hortic. Sci. 2005, 33, 1–7. [Google Scholar]
- Hahn, C.; Howard, N. P.; Albach, D. C. Different shades of kale—approaches to analyze kale variety interrelations. Genes 2022, 13. [Google Scholar] [CrossRef]
- Ferioli, F.; Manco, M.; Giambanelli, E.; D’Antuono, L.F.; Albuquerque, T.G.; Sanches-Silva, A.; Koçaoğlu, B.; Hayran, O. Variability of glucosinolates and phenolics in local kale populations from Turkey, Italy and Portugal. In Proceedings of the 9th International Food Data Conference, Lisbon, Portugal, Instituto Nacional de Saúde Doutor Ricardo Jorge, IP. 2011, 14–17 September.
- Satheesh, N.; Workneh Fanta, S. Kale: Review on nutritional composition, bioactive compounds, antinutritional factors, health-beneficial properties and value-added products. Cogent Food Agric. 2020, 6, 1811048. [Google Scholar] [CrossRef]
- Cartea, M. E.; Picoaga, A.; Soengas, P.; Ordás, A. Morphological characterization of kale populations from northwestern Spain. Euphytica 2003, 129, 25–32. [Google Scholar] [CrossRef]
- Pipan, B.; Neji, M.; Meglič, V.; Sinkovič, L. Genetic diversity of kale (Brassica oleracea L. var. acephala) using agro-morphological and simple sequence repeat (SSR) markers. Genet Resour Crop Evol. 2024, 71, 1221–1239. [Google Scholar]
- Ortega-Hernández, E.; Antunes-Ricardo, M.; Jacobo-Velázquez, D. A. Improving the health-benefits of kales (Brassica oleracea L. var. acephala DC) through the application of controlled abiotic stresses: A Review. Plants 2021, 10. [Google Scholar] [CrossRef]
- Karagüzel, Ü.Ö. Blossoms amid drought: A bibliometric mapping of research on drought stress in ornamental plants (1995–2025). Front. Plant Sci. 2025, 16, 1644092. [Google Scholar]
- Pathirana, I.; Thavarajah, P.; Siva, N.; Wickramasinghe, A.N.; Smith, P. Moisture deficit effects on kale (Brassica oleracea L. var. acephala) biomass, mineral, and low molecular weight carbohydrate concentrations. Sci. Hortic. 2017, 226, 216–222. [Google Scholar] [CrossRef]
- Bauer, N.; Tkalec, M.; Major, N.; Vasari, A.T.; Tokić, M.; Vitko, S.; Ban, D.; Ban, S.G.; Salopek-Sondi, B. Mechanisms of kale (Brassica oleracea var. acephala) tolerance to individual and combined stresses of drought and elevated temperature. Int. J. Mol. Sci. 2022, 23, 11494. [Google Scholar] [CrossRef]
- Wang, Y.; Miao, H.; Zhang, F.; Sun, B.; Wang, Q. Seasonal variation in nutritional substances in varieties of leafy Chinese kale (Brassica oleracea var. alboglabra): A pilot trial. Agronomy 2025, 15, 671. [Google Scholar] [CrossRef]
- Issarakraisila, M.; Ma, Q.; Turner, D.W. Photosynthetic and growth responses of juvenile Chinese kale (Brassica oleracea var. alboglabra) and caisin (Brassica rapa subsp. parachinensis) to waterlogging and water deficit. Sci. Hortic. 2007, 111, 107–113. [Google Scholar] [CrossRef]
- Barickman, T.C.; Ku, K.M.; Sams, C.E. Differing precision irrigation thresholds for kale (Brassica oleracea L. var. acephala) induce changes in physiological performance, metabolites, and yield. Environ. Exp. Bot. 2020, 180, 104253. [Google Scholar] [CrossRef]
- Seneviratne, S.I.; Zhang, X.; Adnan, M.; Badi, W.; Dereczynski, C.; Di Luca, A.; Ghosh, S.; Iskandar, I.; Kossin, J.; Lewis, S.; Otto, F.; Pinto, I.; Satoh, M.; Vicente-Serrano, S.M.; Wehner, M.; Zhou, B. Weather and climate extreme events in a changing climate. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., Huang, M., Leitzell, K., Lonnoy, E., Matthews, J.B.R., Maycock, T.K., Waterfield, T., Yelekçi, O., Yu, R., Zhou, B., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA. 2021; pp. 1513–1766. [Google Scholar]
- Caretta, M.A.; Mukherji, A.; Arfanuzzaman, M.; Betts, R.A.; Gelfan, A.; Hirabayashi, Y.; Lissner, T.K.; Liu, J.; Lopez Gunn, E.; Morgan, R.; Mwanga, S.; Supratid, S. Water. In Climate Change 2022: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Pörtner, H.-O., Roberts, D.C., Tignor, M., Poloczanska, E.S., Mintenbeck, K., Alegría, A., Craig, M., Langsdorf, S., Löschke, S., Möller, V., Okem, A., Rama, B., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA. 2022; pp. 551–712. [Google Scholar] [CrossRef]
- Bot, A.; Benites, J. The Importance of Soil Organic Matter: Key to Drought-Resistant Soil and Sustained Food Production; FAO Soils Bulletin 80; Food and Agriculture Organization of the United Nations: Rome, Italy. 2005. [Google Scholar]
- Hueso, S.; Hernández, T.; García, C. Resistance and resilience of the soil microbial biomass to severe drought in semiarid soils: The importance of organic amendments. Appl. Soil Ecol. 2011, 50, 27–36. [Google Scholar] [CrossRef]
- Franzluebbers, A.J. Water infiltration and soil structure related to organic matter and its stratification with depth. Soil Tillage Res. 2002, 66, 197–205. [Google Scholar] [CrossRef]
- Even, R.J.; Cotrufo, M.F. The ability of soils to aggregate, more than the state of aggregation, promotes protected soil organic matter formation. Geoderma 2024, 442, 116760. [Google Scholar] [CrossRef]
- Pérez-Lucas, G.; Martínez-Zapata, A.; Navarro, S. Valorization of agro-industrial wastes as organic amendments to reduce herbicide leaching into soil. J. Xenobiotics. 2025, 15, 100. [Google Scholar] [CrossRef]
- Ahmad, R.; Jilani, G.; Arshad, M.; Zahir, Z.A.; Khalid, A. Bio-conversion of organic wastes for their recycling in agriculture: An overview of perspectives and prospects. Ann. Microbiol. 2007, 57, 471–479. [Google Scholar] [CrossRef]
- Hiranmai, R.Y.; Neeraj, A.; Vats, P. Improvement of soil health and crop production through utilization of organic wastes: A sustainable approach. Int. J. Recycl. Org. Waste Agric. 2024, 13, 1. [Google Scholar]
- Park, T.; Fischer, S.; Lambert, C.; Hilger, T.; Jordan, I.; Cadisch, G. Combined effects of drought and soil fertility on the synthesis of vitamins in green leafy vegetables. Agriculture 2023, 13, 984. [Google Scholar] [CrossRef]
- Lehmann, J.; Kleber, M. The contentious nature of soil organic matter. Nature. 2015, 528, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Debnath, B.; Haldar, D.; Purkait, M.K. Potential and sustainable utilization of tea waste: A review on present status and future trends. J. Environ. Chem. Eng. 2021, 9, 106179. [Google Scholar] [CrossRef]
- Çakmak, T.G.; Saricaoglu, B.; Ozkan, G.; Tomas, M.; Capanoglu, E. Valorization of tea waste: Composition, bioactivity, extraction methods, and utilization. Food Sci. Nutr. 2024, 12, 3112–3124. [Google Scholar] [CrossRef]
- Tunklová, B.; Jeníček, L.; Malaťák, J.; Neškudla, M.; Velebil, J.; Hnilička, F. Properties of biochar derived from tea waste as an alternative fuel and its effect on phytotoxicity of seed germination for soil applications. Materials 2022, 15, 8709. [Google Scholar] [CrossRef]
- Karataş, A. Is tea waste a promising co-substrate for optimizing the cultivation, growth, and yield of Charleston pepper (Capsicum annuum L.)? Res. Agric. Sci. 2024, 55, 183–192. [Google Scholar] [CrossRef]
- Yıldırım, G.H.; Ay, E.B.; Şahin, M.D. The effects of tea wastes prepared using different composting methods on the seedling growth and selected biochemical properties of maize (Zea mays var. indurata). Food Sci. Nutr. 2025, 13, e70670. [Google Scholar] [CrossRef]
- Pekşen, A.; Yakupoğlu, G. Tea waste as a supplement for the cultivation of Ganoderma lucidum. World J. Microbiol. Biotechnol. 2009, 25, 611–618. [Google Scholar] [CrossRef]
- Ekbiç, E.; Keskin, A. Effects of tea waste compost applications on onion grown under salt stress conditions. Akademik Ziraat Dergisi 2018, 7, 1–8. (In Turkish) [Google Scholar] [CrossRef]
- Özenç, D.B.; Hut, D. Effects of tea waste compost and salt applications on the growth of pepper plants. Topr. Bil. Bitki Besl. Derg. 2018, 6, 86–94. (In Turkish) [Google Scholar]
- Kıran, S.; Baysal Furtana, G. Responses of eggplant seedlings to combined effects of drought and salinity stress: Effects on photosynthetic pigments and enzymatic and non-enzymatic antioxidants. Gesunde Pflanz. 2023, 75, 2579–2590. [Google Scholar] [CrossRef]
- Kıran, S.; Kuşvuran, Ş.; Özkay, F.; Ellialtıoğlu, Ş. Changes in some morphological traits of salt-tolerant and sensitive eggplant genotypes under drought stress conditions. MKUJAS. 2016, 21, (2). (In Turkish) [Google Scholar]
- Kuşvuran, Ş.; Daşgan, H.Y.; Abak, K. Responses of different melon genotypes to drought stress. YYU J AGR SCI. 2011, 21, 209–219. (In Turkish) [Google Scholar]
- Smart, R.E.; Bingham, G.E. Rapid estimates of relative water content. Plant Physiol. 1974, 53, 258–260. [Google Scholar] [CrossRef]
- Ben Ammar, H.; Picchi, V.; Arena, D.; Treccarichi, S.; Bianchi, G.; Lo Scalzo, R.; Branca, F. Variation of bio-morphometric traits and antioxidant compounds of Brassica oleracea L. accessions in relation to drought stress. Agronomy 2022, 12, 2016. [Google Scholar] [CrossRef]
- Mohi-Ud-Din, M.; Hossain, M.A.; Rohman, M.M.; Uddin, M.N.; Haque, M.S.; Ahmed, J.U.; Hossain, A.; Hassan, M.M.; Mostofa, M.G. Multivariate analysis of morpho-physiological traits reveals differential drought tolerance potential of bread wheat genotypes at the seedling stage. Plants 2021, 10, 879. [Google Scholar] [CrossRef]
- Abdeltaif, S.A.; SirElkhatim, K.A.; Hassan, A.B. Estimation of phenolic and flavonoid compounds and antioxidant activity of spent coffee and black tea (processing) waste for potential recovery and reuse in Sudan. Recycling 2018, 3, 27. [Google Scholar] [CrossRef]
- Nadiah, N.I.; Uthumporn, U. Determination of phenolic and antioxidant properties in tea and spent tea under various extraction methods and determination of catechins, caffeine, and gallic acid by HPLC. IJASEIT 2015, 11, 10–18517. [Google Scholar]
- Ekbiç, H.B.; Akbulut, Ş.; Özenç, D.B. Effects of mixtures of hazelnut husk and tea waste compost on the development of 41B American grapevine rootstock cuttings grown under saline conditions. Akademik Ziraat Dergisi 2022, 11, 1–8. (In Turkish) [Google Scholar]
- Chevilly, S.; Dolz-Edo, L.; López-Nicolás, J.M.; Morcillo, L.; Vilagrosa, A.; Yenush, L.; Mulet, J.M. Physiological and molecular characterization of the differential response of broccoli (Brassica oleracea var. italica) cultivars reveals limiting factors for broccoli tolerance to drought stress. J. Agric. Food Chem. 2021, 69, 10394–10404. [Google Scholar] [CrossRef]
- Yoo, M.J.; Hwang, Y.; Koh, Y.M.; Zhu, F.; Deshpande, A.S.; Bechard, T.; Andreescu, S. Physiological and molecular modulations to drought stress in the Brassica species. Int. J. Mol. Sci. 2024, 25, 3306. [Google Scholar] [CrossRef] [PubMed]
- Yildirim, E.; Ekinci, M.; Turan, M. Impact of biochar in mitigating the negative effect of drought stress on cabbage seedlings. J. Soil Sci. Plant Nutr. 2021, 21, 2297–2309. [Google Scholar] [CrossRef]
- Kang, Y.G.; Choi, J.; Lee, J.Y.; Kim, J.H.; Kim, J.H.; Park, D.G.; Yun, Y.U. Response of biochar amendment for substituting organic fertilizer ingredients in kale-grown soils. Korean J. Agric. Sci. 2024, 51, 819–827. [Google Scholar] [CrossRef]
- Thepsilvisut, O.; Srikan, N.; Chutimanukul, P.; Athinuwat, D.; Chuaboon, W.; Marubodee, R.; Ehara, H. Practical guidelines for farm waste utilization in sustainable kale production. Horticulturae 2024, 10, 525. [Google Scholar] [CrossRef]
- Clemente, R.; Almela, C.; Bernal, M.P. A remediation strategy based on active phytoremediation followed by natural attenuation in a soil contaminated by pyrite waste. Environ. Pollut. 2006, 143, 397–406. [Google Scholar] [CrossRef]
- Marschner, H. Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: London, UK, 2012; ISBN 9780123849052. [Google Scholar] [CrossRef]
- Güçlü Üstündağ, Ö.; Erşan, S.; Özcan, E.; Özan, G.; Kayra, N.; Ekinci, F.Y. Black tea processing waste as a source of antioxidant and antimicrobial phenolic compounds. Eur. Food Res. Technol. 2016, 242, 1523–1532. [Google Scholar] [CrossRef]
- Cebeci, E. Comparative evaluation of eggplant genotypes with their wild relatives under gradually increased drought stress. Bragantia 2024, 83, e20230246. [Google Scholar] [CrossRef]
- Kıran, S.; Baysal Furtana, G.; Ellialtıoğlu, Ş.Ş. Physiological and biochemical assay of drought stress responses in eggplant (Solanum melongena L.) inoculated with commercial inoculant of Azotobacter chroococcum and Azotobacter vinelandii. Sci. Hortic. 2022, 305, 111394. [Google Scholar] [CrossRef]
Figure 1.
Appearance of factory-derived tea waste, consisting of leaf residues, fibers, and fine particles excluded during the processing of green tea into black tea.
Figure 1.
Appearance of factory-derived tea waste, consisting of leaf residues, fibers, and fine particles excluded during the processing of green tea into black tea.

Figure 2.
Schematic representation of the experimental design. The study was arranged as a factorial combination of two kale varieties (VK1: Karadeniz Yaprak, VK2: КЕЙЛ), three levels of tea waste amendment (TW0: 0%, TW1: 5%, TW2: 10%), and three drought stress regimes (DS0: 100% field capacity (FC), DS1: 75% of (FC): mild stress, DS2: 50% of FC: moderate stress).
Figure 2.
Schematic representation of the experimental design. The study was arranged as a factorial combination of two kale varieties (VK1: Karadeniz Yaprak, VK2: КЕЙЛ), three levels of tea waste amendment (TW0: 0%, TW1: 5%, TW2: 10%), and three drought stress regimes (DS0: 100% field capacity (FC), DS1: 75% of (FC): mild stress, DS2: 50% of FC: moderate stress).
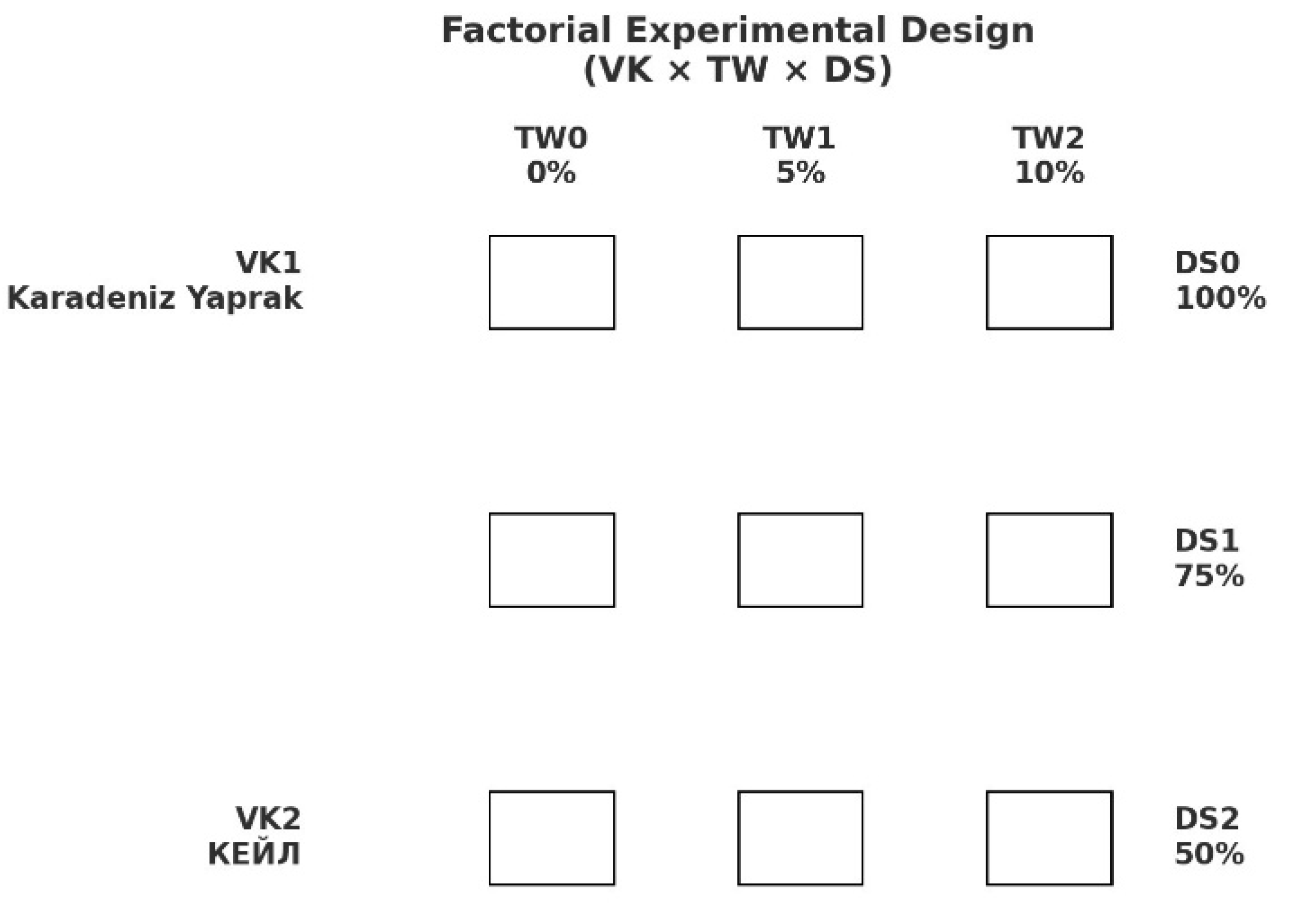
Figure 3.
Trait clustering revealed by a polar heatmap with dendrogram based on principal component analysis of kale varieties grown in tea waste–amended soils under different drought stress regimes.
Figure 3.
Trait clustering revealed by a polar heatmap with dendrogram based on principal component analysis of kale varieties grown in tea waste–amended soils under different drought stress regimes.

Figure 4.
Correlation heatmap showing relationships among morphological and physiological traits of kale cultivated in tea waste–amended soil under drought stress.
Figure 4.
Correlation heatmap showing relationships among morphological and physiological traits of kale cultivated in tea waste–amended soil under drought stress.
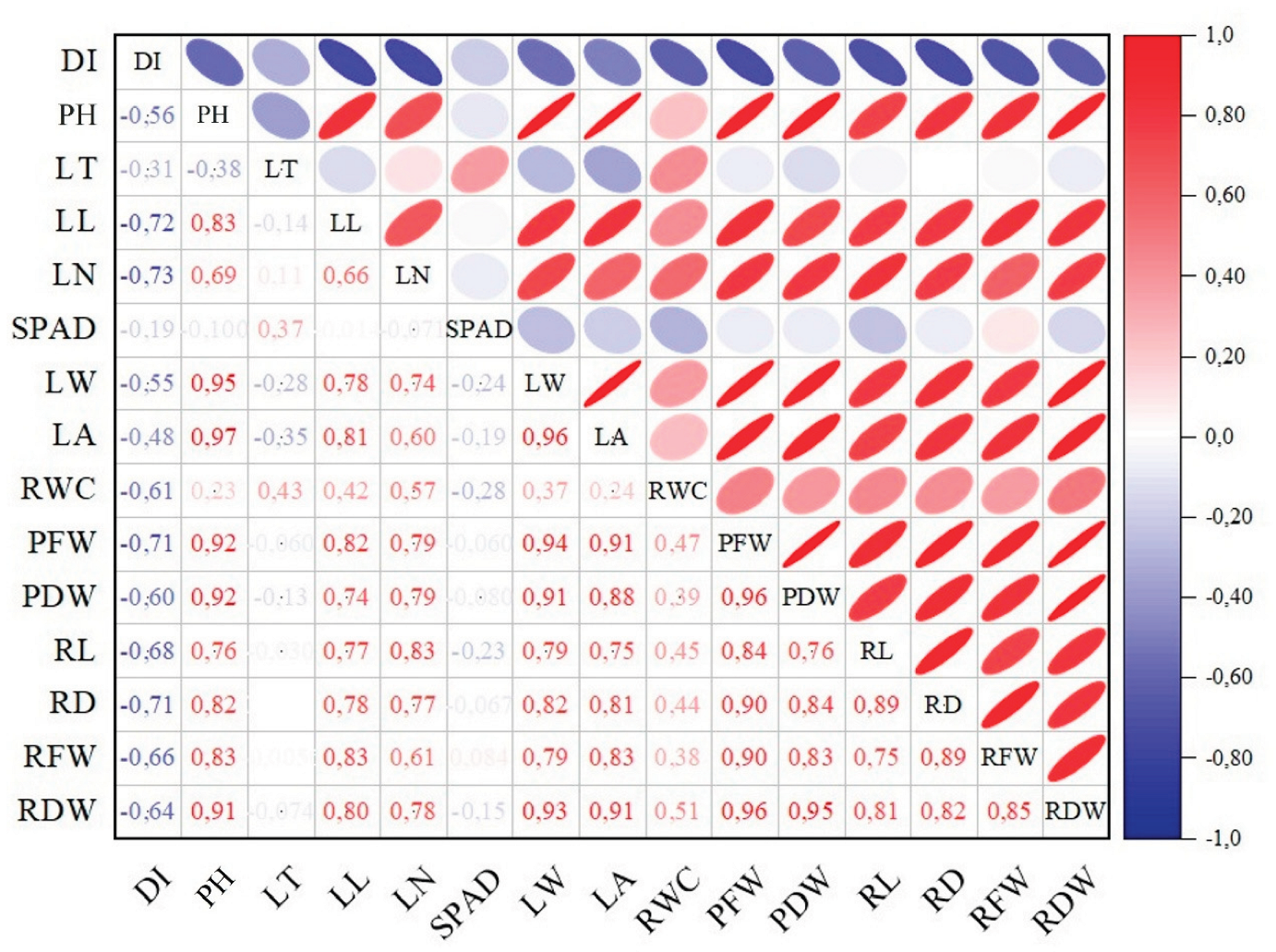

Table 1.
Physical and chemical properties of factory-derived tea waste applied as an organic amendment in kale cultivation.
Table 1.
Physical and chemical properties of factory-derived tea waste applied as an organic amendment in kale cultivation.
| Property | Value |
|---|---|
| pH | 5.57 |
| EC (dS m⁻¹) | 0.102 |
| Total N (%) | 2.19 |
| Organic matter (%) | 47 |
| Water holding capacity (%) | 96.02 |
Table 2.
Physical and chemical properties of soil used in the experiment.
| Property | Value |
|---|---|
| Texture class | Clayey |
| EC (dS m⁻¹, saturated paste) | 0.62 |
| pH (saturated paste) | 6.93 |
| CaCO₃ (%) | 1.80 |
| Organic matter (%) | 2.58 |
Table 3.
Factorial structure of kale variety (VK), tea waste level (TW), and drought stress regime (DS).
Table 3.
Factorial structure of kale variety (VK), tea waste level (TW), and drought stress regime (DS).
| Variety (VK) | Tea Waste (TW, %) | Irrigation Regime (RI) |
|---|---|---|
| Karadeniz Yaprak (VK1) | 0 (TW0) | DS0: Control (100% field capacity) DS1: Light stress (75% field capacity) DS2: Moderate stress (50% field capacity) |
| 5 (TW1) | DS0: Control (100% field capacity) DS1: Light stress (75% field capacity) DS2: Moderate stress (50% field capacity) |
|
| 10 (TW2) | DS0: Control (100% field capacity) DS1: Light stress (75% field capacity) DS2: Moderate stress (50% field capacity) |
|
| КЕЙЛ (VK2) | 0 (TW0) | DS0: Control (100% field capacity) DS1: Light stress (75% field capacity) DS2: Moderate stress (50% field capacity) |
| 5 (TW1) | DS0: Control (100% field capacity) DS1: Light stress (75% field capacity) DS2: Moderate stress (50% field capacity) |
|
| 10 (TW2) | DS0: Control (100% field capacity) DS1: Light stress (75% field capacity) DS2: Moderate stress (50% field capacity) |
Varieties: Karadeniz Yaprak (VK1) and КЕЙЛ (VK2). Tea waste was incorporated into soil at 0% (TW0), 5% (TW1), and 10% (TW2) (w/w). Drought stress treatments were imposed at 100% field capacity (DS0, control), 75% field capacity (FC) (DS1, mild stress), and 50% field capacity (FC) (DS2, moderate stress).
Table 4.
Variance analysis of kale traits under factorial combinations of variety, tea waste amendment, and drought stress.
Table 4.
Variance analysis of kale traits under factorial combinations of variety, tea waste amendment, and drought stress.
| Sources of Variation | df | DI | PH | LT | LL | LN | SPAD |
LW |
LA |
RWC |
PFW |
PDW |
RL |
RD |
RFW |
RDW |
| VK | 1 | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** |
| TW | 2 | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** |
| DS | 2 | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** |
| VK x TW | 2 | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** |
| VK x DS | 2 | ** | * | ** | * | ** | ** | ** | ** | ** | ns | ns | ** | * | ** | ns |
| TW x DS | 4 | ** | ** | ** | ** | ** | ** | * | ** | * | ** | ** | ** | ** | ** | * |
| VK x TW x DS | 4 | ** | ** | ** | ** | * | ** | ** | ** | ** | ns | * | ** | ns | ** | ns |
| Error | 68 | 0.0026 | 0.236 | 0.0005 | 0.0243 | 0.1369 | 0.7464 | 0.0101 | 0.58 | 0.181 | 1.008 | 0.035 | 0.194 | 0.037 | 0.027 | 0.005 |
| CV (%) | 4.39 | 3.32 | 4.14 | 2.07 | 4.82 | 1.86 | 4.29 | 1.40 | 0.55 | 8.89 | 11.30 | 1.610 | 4.23 | 5.16 | 12.71 |
Note: ns: Non significant *: p < 0.05, **: p < 0.01. Variety: VK, Tea Waste: TW, Drought Stress: DS, Damage Index: DI, Plant Height: PH, Leaf Thickness: LT, Leaf Length: LL, Leaf Number: LN, Chlorophyll Content: SPAD, Leaf weight: LW, Leaf area: LA, Leaf water content: LWC, Plant fresh weight: PFW, Plant dry weight: PDW, Root length: RL, Root diameter: RD, Root fresh weight: RFW, Root dry weight: RDW.
Table 5.
Interaction effects of variety, tea waste level, and drought stress on morphological and physiological traits of kale.
Table 5.
Interaction effects of variety, tea waste level, and drought stress on morphological and physiological traits of kale.
| Variety | TW % | DS | Dİ |
PH (cm) |
LT (mm) |
LL (cm) |
LN (number plant-1) |
SPAD |
LW (g) |
LA (cm2) |
RWC (%) |
PFW (g plant-1) |
RL (cm) |
RFW (g plant-1) |
| VK1 | TW0 | DS0 | 0.00 l | 21.67 a | 0.200 b | 12.44 a | 8.60 bc | 50.30 bc | 3.62 b | 98.6 a | 92.36 a | 2.84 a | 35.36 a | 7.64 a |
| DS1 | 0.34 k | 18.98 c | 0.188 cd | 10.20 b | 8.04 def | 50.36 bc | 3.18 c | 81.80 b | 71.54 j | 2.06 cd | 31.28 b | 3.90 cd | ||
| DS2 | 1.10 h | 16.48 e | 0.176 e | 8.08 d | 8.38 bcd | 52.60 a | 2.60 e | 58.88 g | 60.22 o | 1.80 ef | 29.72 e | 3.76 d | ||
| TW1 | DS0 | 0.72 i | 17.90 d | 0.140 j | 9.10 c | 7.78 efgh | 40.04 h | 3.04 d | 70.56 d | 84.41 e | 1.84 def | 30.50 cd | 3.46 e | |
| DS1 | 1.46 e | 15.86 f | 0.142 ij | 7.52 e | 7.60 fgh | 40.78 h | 2.68 e | 66.74 e | 77.57 h | 1.54 gh | 28.70 f | 2.86 g | ||
| DS2 | 1.74 c | 15.22 g | 0.156 gh | 7.26 f | 7.02 i | 43.0 fg | 2.20 g | 61.28 f | 70.46 k | 1.18 ij | 26.90 g | 2.50 h | ||
| TW2 | DS0 | 1.04 h | 19.90 b | 0.198 b | 7.92 d | 8.80 b | 42.32 g | 4.18 a | 97. 66 a | 86.52 d | 2.98 a | 35.78 a | 4.88 b | |
| DS1 | 1.36 fg | 18.54 c | 0.160 fg | 7.52 e | 8.04 def | 42.96 fg | 3.10 cd | 73.64 c | 78.56 g | 2.58 b | 25.28 h | 4.00 c | ||
| DS2 | 1.74 c | 17.62 d | 0.150 hi | 7.08 fg | 7.54 gh | 49.30 cd | 2.98 d | 70.10 d | 65.33 l | 2.00 cde | 24.58 i | 3.38 e | ||
| VK2 | TW0 | DS0 | 0.00 l | 13.64 h | 0.224 a | 6.94 g | 8.58 bc | 50.42 b | 2.38 f | 42.40 j | 91.78 b | 1.66 fg | 27.44 g | 3.14 f |
| DS1 | 1.42 ef | 8.80 l | 0.220 a | 5.28 j | 6.24 kj | 49.06 d | 1.02 k | 23.96 n | 82.74 f | 0.70 lm | 21.50 k | 2.36 h | ||
| DS2 | 1.96 a | 8.62 l | 0.200 b | 4.56 k | 5.82 k | 50.54 b | 0.80 l | 22.08 o | 63.30 m | 0.60 m | 17.16 m | 1.98 i | ||
| TW1 | DS0 | 1.34 g | 12.12 i | 0.204 b | 8.04 d | 8.24 cde | 39.90 h | 2.64 e | 48.92 h | 90.11 c | 1.28 i | 30.30 d | 3.14 f | |
| DS1 | 1.44 e | 11.62 i | 0.196 bc | 6.34 i | 8.00 defg | 49.04 d | 1.28 j | 27.78 m | 78.01 h | 1.32 hi | 28.48 f | 2.40 h | ||
| DS2 | 1.64 d | 9.52 k | 0.186 d | 5.16 j | 6.42 j | 43.6 f | 1.020 k | 25.52 n | 61.96 n | 0.92 kl | 25.64 h | 1.98 i | ||
| TW2 | DS0 | 0.48 j | 15.24 g | 0.204 b | 8.10 d | 9.42 a | 46.20 e | 2.42 f | 46.72 i | 90.56 c | 2.16 c | 30.86 bc | 3.08 f | |
| DS1 | 1.44 e | 11.52 i | 0.220 a | 6.94 g | 7.36 hi | 45.5 e | 1.66 h | 35.98 k | 84.83 e | 1.30 i | 22.68 j | 1.66 j | ||
| DS2 | 1.86 b | 10.46 j | 0.168 ef | 6.62 h | 6.42 j | 48.7 d | 1.42 i | 29.26 l | 72.85 i | 1.00 jk | 19.58 l | 1.36 k | ||
| LSD %5 | 0.065 | 0.614 | 0.023 | 0.197 | 0.467 | 1.090 | 0.127 | 0.959 | 0.537 | 0.236 | 0.555 | 0.208 |
Statistically significant means have been grouped according to the LSD (5%) test. The differences between means denoted by different letters are significant (p<0.05). VK1: Karadeniz Yaprak, CK2: КЕЙЛ, TW: Tea Waste, TW0: Soil, TW1: Soil with 5% Tea Waste added, TW2: Soil with 10% Tea Waste added, DS: Drought Stress, DS0: 100% full irrigation, DS1: 75% of full irrigation, DS2: 50% of full irrigation, DI: Damage Index, PH: Plant height, LT: Leaf thickness, LL: Leaf length, LN: Leaf number, SPAD: Chlorophyll content, LW: Leaf weight, LA: Leaf area, RWC: Leaf water content, PFW: Plant fresh weight, PDW: Plant dry weight, RL: Root length, RFW: Root fresh weight, RDW: Root dry weight.
Table 6.
Interaction effects of variety and tea waste level on morphological and physiological traits of kale under drought conditions.
Table 6.
Interaction effects of variety and tea waste level on morphological and physiological traits of kale under drought conditions.
| Variety | TW | DI |
PH (cm) |
LT (mm) |
LL (cm) |
LN (number plant-1) |
SPAD |
LW (g) |
LA (cm2) |
RWC (%) |
PFW (g plant-1) |
PDW (g plant-1) |
RL (cm) |
RD (mm) |
RFW (g plant-1) |
RDW (g plant-1) |
| VK1 | TW0 | 0.48 f | 19.04 a | 0.188 c | 10.24 a | 8.34 a | 51.09 a | 3.13 b | 79.76 b | 74.71 e | 16.53 a | 2.23 b | 32.12 a | 5.49 a | 5.10 a | 0.77 a |
| TW1 | 1.31 c | 16.33 b | 0.146 e | 8.01 b | 7.47 b | 41.27 f | 2.64 c | 66.19 c | 77.47 c | 10.80 b | 1.52 c | 28.70 b | 4.47 d | 2.94 c | 0.61 b | |
| TW2 | 1.38 b | 18.68 a | 0.169 d | 7.51 c | 8.12 a | 44.86 d | 3.42 a | 80.47 a | 76.80 d | 16.13 a | 2.52 a | 28.55 b | 5.13 b | 4.09 b | 0.79 a | |
| VK2 | TW0 | 1.13 e | 10.35 e | 0.215 a | 5.59 f | 6.89 c | 50.01 b | 1.40 f | 29.48 f | 79.27 b | 7.27 d | 0.99 e | 22.03 e | 3.93 e | 2.49 d | 0.41 d |
| TW1 | 1.47 a | 11.08 d | 0.195 b | 6.51 e | 7.55 b | 44.18 e | 1.64 e | 33.41 e | 76.70 d | 7.93 d | 1.17 d | 28.14 c | 4.61 c | 2.51 d | 0.41 d | |
| TW2 | 1.26 e | 12.41 c | 0.197 b | 7.22 d | 7.73 b | 46.79 c | 1.83 d | 37.32 d | 82.74 a | 9.06 c | 1.49 c | 24.37 d | 3.83 e | 2.03 e | 0.55 c | |
| LSD %5 | 0.037 | 0.355 | 0.006 | 0.114 | 0.628 | 0.630 | 0.073 | 0.554 | 0.310 | 0.732 | 0.136 | 0.321 | 0.141 | 0.120 | 0.055 | |
Statistically significant means have been grouped according to the LSD (5%) test. The differences between means denoted by different letters are significant (p<0.05). VK1: Karadeniz Yaprak, CK2: КЕЙЛ, TW: Tea Waste, TW0: Soil, TW1: Soil with 5% Tea Waste added, TW2: Soil with 10% Tea Waste added, DS: Drought Stress, DS0: 100% full irrigation, DS1: 75% of full irrigation, DS2: 50% of full irrigation, DI: Damage Index, PH: Plant height, LT: Leaf thickness, LL: Leaf length, LN: Leaf number, SPAD: Chlorophyll content, LW: Leaf weight, LA: Leaf area, RWC: Leaf water content, PFW: Plant fresh weight, PDW: Plant dry weight, RL: Root length, RD: Root diameter, RFW: Root fresh weight, RDW: Root dry weight.
Table 7.
Interaction effects of variety and drought stress on morphological and physiological traits of kale cultivated in tea waste–amended soil.
Table 7.
Interaction effects of variety and drought stress on morphological and physiological traits of kale cultivated in tea waste–amended soil.
| Variety | DS | DI |
PH (cm) |
LT (mm) |
LL (cm) |
LN (number plant-1) |
SPAD |
LW (g) |
LA (cm2) |
RWC (%) |
PFW (g plant-1) |
PDW (g plant-1) |
RL (cm) |
RD (mm) |
RFW (g plant-1) |
RDW (g plant-1) |
| VK1 | DS0 | 0.58 e | 19.82 a | 0.179 b | 9.82 a | 8.39 b | 44.22 d | 3.61 a | 88.94 a | 87.76 b | 19.00 | 2.55 | 33.89 a | 5.57 a | 5.33 a | 0.90 |
| DS1 | 1.05 d | 17.79 b | 0.163 c | 8.47 b | 7.89 c | 44.70 d | 2.98 b | 74.06 b | 75. 89 d | 13.60 | 2.06 | 28.42 c | 5.02 b | 3.58 b | 0.68 | |
| DS2 | 1.52 b | 16.44 c | 0.161 c | 7.47 d | 7.65 c | 48.3 a | 2.59 c | 63.42 c | 65. 34 f | 10.87 | 1.66 | 27.00 d | 4.51 d | 3.21 c | 0.59 | |
| VK2 | DS0 | 0.61 e | 13.67 d | 0.210 a | 7.69 c | 8.75 a | 45.51 c | 2.48 d | 46.01 d | 90.81 a | 12.33 | 1.70 | 29.53 b | 4.81 c | 3.12 c | 0.61 |
| DS1 | 1.43 c | 10.65 e | 0.212 a | 6.19 e | 7.20 d | 47.87 ab | 1.32 e | 29.24 e | 81.86 c | 6.73 | 1.11 | 24.22 e | 4.06 e | 2.14 d | 0.47 | |
| DS2 | 1.82 a | 9.53 f | 0.184 b | 5.45 f | 6.22 e | 47.6 b | 1.08 f | 24.95 f | 66.03 e | 5.20 | 0.84 | 20.80 f | 3.51 f | 1.77 e | 0.30 | |
| LSD %5 | 0.037 | 0.355 | 0.006 | 0.114 | 0.628 | 0.630 | 0.073 | 0.554 | 0.310 | 0.321 | 0.141 | 0.120 | ||||
Statistically significant means have been grouped according to the LSD (5%) test. The differences between means denoted by different letters are significant (p<0.05). VK1: Karadeniz Yaprak, CK2: КЕЙЛ, TW: Tea Waste, TW0: Soil, TW1: Soil with 5% Tea Waste added, TW2: Soil with 10% Tea Waste added, DS: Drought Stress, DS0: 100% full irrigation, DS1: 75% of full irrigation, DS2: 50% of full irrigation, DI: Damage Index, PH: Plant height, LT: Leaf thickness, LL: Leaf length, LN: Leaf number, SPAD: Chlorophyll content, LW: Leaf weight, LA: Leaf area, RWC: Leaf water content, PFW: Plant fresh weight, PDW: Plant dry weight, RL: Root length, RD: Root diameter, RFW: Root fresh weight, RDW: Root dry weight.
Table 8.
Interaction effects of tea waste level and drought stress on morphological and physiological traits of kale.
Table 8.
Interaction effects of tea waste level and drought stress on morphological and physiological traits of kale.
|
TW |
DS | DI |
PH (cm) |
LT (mm) |
LL (cm) |
LN (number plant-1) |
SPAD |
LW (g) |
LA (cm2) |
RWC (%) |
PFW (g plant-1) |
PDW (g plant-1) |
RL (cm) |
RD (mm) |
RFW (g plant-1) |
RDW (g plant-1) |
| TW0 | DS0 | 0.00 i | 17.65 a | 0.212 a | 9.69 a | 8.59 b | 50.36 b | 3.00 b | 70.50 b | 92.07 a | 17.60 a | 2.25 a | 31.40 b | 5.77 a | 5.39 a | 0.76 b |
| DS1 | 0.88 g | 13.89 c | 0.204 b | 7.74 d | 7.14 d | 49.71 bc | 2.10 f | 52.88 e | 77.14 f | 10.20 cd | 1.38 e | 26.39 e | 4.53 de | 3.13 d | 0.56 d | |
| DS2 | 1.53 c | 12.55 d | 0.188 c | 6.32 h | 7.10 d | 51.57 a | 1.70 h | 40.48 i | 61.76 i | 7.90 e | 1.20 f | 23.44 g | 3.83 g | 2.87 e | 0.46 e | |
| TW 1 | DS0 | 1.03 f | 16.01 b | 0.172 d | 8.57 b | 8.01 c | 39.97 f | 2.84 c | 59.74 c | 87.26 c | 11.90 b | 1.56 d | 30.40 c | 4.71 c | 3.30 c | 0.63 c |
| DS1 | 1.45 d | 13.74 c | 0.169 d | 7.01 f | 7.80 c | 44.91 d | 1.98 g | 47.26 g | 77.79 e | 9.40 d | 1.43 de | 28.59 d | 4.67 cd | 2.63 f | 0.51 de | |
| DS2 | 1.69 b | 12.37 d | 0.171 d | 6.21 h | 6.72 e | 43.30 e | 1.61 i | 42.40 h | 66.21 h | 6.80 f | 1.05 f | 26.27 e | 4.24 f | 2.37 g | 0.39 f | |
| TW 2 | DS0 | 0.76 h | 17.57 a | 0.201 b | 8.01 c | 9.11 a | 44.26 d | 3.30 a | 72.19 a | 88.54 b | 17.50 a | 2.57 a | 33.32 a | 5.08 b | 3.98 b | 0.87 a |
| DS1 | 1.40 e | 15.03 b | 0.190 c | 7.23 e | 7.70 c | 44.24 d | 2.38 d | 54.81 d | 81.69 d | 10.90 c | 1.94 c | 23.98 f | 4.42 e | 2.83 e | 0.65 c | |
| DS2 | 1.80 a | 14.04 c | 0.159 d | 6.85 g | 6.98 de | 48.98 c | 2.20 e | 49.68 f | 69.09 g | 9.40 d | 1.50 de | 22.08 h | 3.95 g | 2.37 g | 0.48 e | |
| LSD %5 | 0.045 | 0.434 | 0.006 | 0.139 | 0.330 | 0.771 | 0.90 | 0.678 | 0.380 | 0.896 | 0.168 | 0.393 | 0.173 | 0.147 | 0.067 | |
Statistically significant means have been grouped according to the LSD (5%) test. The differences between means denoted by different letters are significant (p<0.05). VK1: Karadeniz Yaprak, CK2: КЕЙЛ, TW: Tea Waste, TW0: Soil, TW1: Soil with 5% Tea Waste added, TW2: Soil with 10% Tea Waste added, DS: Drought Stress, DS0: 100% full irrigation, DS1: 75% of full irrigation, DS2: 50% of full irrigation, DI: Damage Index, PH: Plant height, LT: Leaf thickness, LL: Leaf length, LN: Leaf number, SPAD: Chlorophyll content, LW: Leaf weight, LA: Leaf area, RWC: Leaf water content, PFW: Plant fresh weight, PDW: Plant dry weight, RL: Root length, RD: Root diameter, RFW: Root fresh weight, RDW: Root dry weight.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



