Submitted:
21 September 2025
Posted:
23 September 2025
You are already at the latest version
Abstract
Forests are fragile ecosystems that are imperiled by various factors associated with human activity and climate change. They necessitate new and effective solutions for their natural and efficient regeneration. The germination of forest seeds is essential for ecosystem restoration and the safeguarding of biodiversity. This process is affected by various biotic and abiotic elements, such as the quality of the current soil microbiome and algae. These organisms are crucial in facilitating forest regeneration by enhancing seed physiology, augmenting stress resilience, improving nutrient acquisition, stimulating phytohormone production, and protecting against diseases. The soil microbial-algal consortium stimulates seed hydration and metabolic activation through external bioactive signals and availability of amino acids and polysaccharides, the specifics of which remain inadequately clarified. Recent studies suggest that the functional actions of the soil microbiome and algae may be a new way to increase forest seed germination in general, especially for preserving threatened and endangered species. In order to restore forest health, this review identifies the function mechanisms through which the microbiome and algae complex impacts seed germination, indicates the limitations of past studies to improve or conserve seed germination, and discusses more sustainable advanced biotechnological approaches that integrate the benefits of the microbiome and algae complex, such as bioformulations and seed coatings containing microbial and algal extracts.
Keywords:
forest seeds
; germination
; mechanisms
; algae
; microbiome
; ecosystem
1. Introduction
The 21st century has witnessed an alarming expansion of extreme and unusual environmental disturbances (fires, droughts, greenhouse effect, floods, storms, etc.) involved in the functional and structural alteration of forest ecosystems. In turn, the degradation of their resilience feeds the initial disruptions, in addition to exacerbating desertification, the depletion of mineral and edaphic biomass resources, and the emergence of forest islands, thus creating a destructive ecological cycle harmful to all forms of life and disruptive to trophic networks (plants–animals–microorganisms) [1,2]. It is important to point out that anthropogenic pressures, such as the overexploitation of forest resources (timber, medicinal plants, etc.), urban expansion, and soil and atmospheric pollution, are implicated in forest decline. The restoration of a balanced, stable, and sustainable ecosystem regime primarily depends on the regeneration of adaptive plant communities [3].
The forest population constitutes a complex, dynamic, and self-organized bio-entity. Its renewal forms a vital and fundamental framework for the biodiversity perpetuation of intra- and interspecific genetic and phenotypic heritage. In reality, it serves as the cradle for numerous biogeochemical connections that maintain the harmony, sustainability, integrity, and purity of natural ecosystems. It plays an active role in atmospheric carbon retention within the woody and herbaceous plant biomass; in aquifer recharge and hydrogeological filtration; in the planetary nitrogen cycle; in the enhancement of the soil’s chemical and microbiological footprint; and, above all, in the rehabilitation of the biotope for various living species [4].
The functional endurance of forests is essentially correlated with the generation of new seeds that are tolerant, robust, and continuously evolving genetically in terms of morphology, physiology, immunity, reproduction, and ecological adaptation, in order to be able to withstand and overcome the biotic and abiotic hazards of the environment that impede the development of new trees [5].
Germination embodies the vital spark that transforms a dormant seed into a dynamic young seedling, orchestrating its profound biological processes and interactions with the surrounding environment. It serves as a selective demographic filter for forest populations capable of readjusting their behaviors and physiologies in response to ecological stresses, where the self-healing of forest ecosystems often depends on the initial pool of dispersed seeds and their genetic quality, germination rate, survival, and growth speed of new seedlings, especially during the first weeks of development. In contrast, the alteration of the seeds’ germination cycle significantly disturbs the phytocoenotic heterogeneity of forests, promoting the invasive dominance of opportunistic and parasitic plants, alongside the degradation of the zoocenosis and microcenosis [5,6,7].
Seed germination is often triggered by the imbibition process, which hinders the genetic expression of abscisic acid and stimulates the release of ethylene gas to awaken the embryo and induce the hydrolytic enzymatic catabolism of carbon reserves, in order to obtain the energy necessary to initiate the differentiated development of the plumule and radicle. This bioactive physiological progression depends on the compatibility of pedoclimatic parameters (temperature, aeration, light, physicochemical soil profile, etc.), as well as competition and allelopathic phenomena; however, it is fundamentally regulated by a range of phytohormones (gibberellins, auxins, and cytokinins) present in delicate concentrations, whether endogenous (produced by the seed), exogenous (synthesized by the rhizospheric microbiome and microalgae), endophytic, and/or epiphytic (elaborated by the seed microbiome) [6,8].
The microbiome corresponds to a constellation of microorganisms and their genetic heritage, including bacteria, archaea, yeasts, molds, protozoa, and algae, which weave intimate and complementary metabolic, genetic, and immune interactions with those of the seed to support the germination process [9]. The microbiome is mainly established in the superficial and deep compartments of seeds and in their spermosphere, giving them a miraculous genetic and functional extension, whereby the resulting super-organism, called the “Holobiont” is endowed with a palpably enhanced adaptive capacity [10]. Among the fundamental components of the microbiome, microalgae such as cyanobacteria, chlorophytes, etc., can be distinguished. These are pigmented, hydrophilic microorganisms of various shapes. They are photosynthetic and oxygen-producing, and some can even capture and solubilize atmospheric nitrogen [11]. Spermospheric microalgae effectively contribute to the initial development of seeds by injecting the forest soil with nutrients and other auxiliary bioactive substances, such as phytohormones, enzymes, antioxidants, antibiotics, osmotic regulators, and transcriptomic and metabolic modulators [11,12].
Undoubtedly, germination is the starting point for the rejuvenation and expansion of the forest cover; however, it is currently highly threatened by anarchy, insufficiency, and failure due to the challenges associated with the annual loss of thousands of hectares of forests. This loss is responsible for the fragmentation of the green landscape, causing a drastic decline in seed stock and variability, as well as their genetic erosion and isolation due to the parallel disappearance of zoochorous dispersal vectors and the degradation of mutualistic and symbiotic microfauna, and this engenders disrupted reproductive phenology, forced altitudinal and latitudinal migration of plant species, and premature death of seedlings [5,7].
Researchers think that the success of the forest regeneration trajectory is closely correlated by the deciphering of the complex molecular interactions between the seeds and their microbiome and the algae of their biotope, in order to be able to select forest-specific bio-inoculants that sponsor the germinative potential of the seeds, and that amplify their adaptive physiological plasticity, particularly during prolonged periods of saline, oxidative and nutritive stress, and most importantly far from chemical inputs, within the framework of the improvement of sustainability and ecosystemic resilience [6,9].
To revitalize forest ecosystems, this review aims to scrutinize the functional dynamics through which the invisible microbiome–algae biounit orchestrates the germinative potential of seeds. In addition, it underlines the gaps of previous scientific research devoted to improving and boosting the first breath of plant life. This review also presents an analysis of pioneering and sustainable biotechnological approaches exploiting microbial and algal consortia in the development of new biofertilizers and enriched seed coatings, providing a biological lever that promotes forest robustness and regeneration.
2. Overview of Microbiomes and Algae in Forest Ecosystems
Recent scientific research considers the forest soil microbiome as a functionally holistic microbial meta-community, evolved and integrated by trophic networks, molecular transformation mechanisms, and continuous allelic migration between taxa within the mineral matrix of the forest soil [13]. This invisible ecosystem encompassed within the forest macrosystem, forms an indispensable biomotor of forest soil fertility, whose synergistic microbiotic architecture of bacteria, protists, fungi, archaea, algae, and viruses is tightly governed by tree root exudates, the availability of animal and plant necromass material, and the soil’s physicochemical gradient [14,15].
The taxonomic, genetic, and metabolic biodiversity of the soil microbiome constitutes a crucial biological exchange platform between the plant world and the litho-hydro-atmospheric sphere [13]. Almost one-third of its bacterial compartment is dominated by non-sporulating Gram-negative microbes of the phylum Acidobacteriota, such as Acidobacterium capsulatum, Acidipila rosea, Edaphobacter modestus, Terriglobus roseus, and others, which thrive in acidic soils of boreal and temperate forests due to their highly developed acid-tolerant enzymatic systems that pump excess protons elsewhere. These bacteria, with their flexible oligotrophic physiology, can finely adjust to unfavorable pedoclimatic conditions, whose their advanced fermentative and respiratory metabolic machineries are capable of dissecting massive organic polymers (cellulose, chitosan, hemicellulose, and lignin), and thereby sequestering carbon in the soil. Moreover, they are implicated in the biogeochemical cycles of nitrogen, iron, and sulfur, supporting the chemical profile of the soil [16].
Proteobacteria is another important phylogenetic clade of Gram-negative microbes, enriched with Plant Growth-Promoting Bacteria such as Rhizobium spp., Azospirillum brasilense, Pseudomonas fluorescens, Methylobacterium spp., Bradyrhizobium spp., Burkholderia spp., etc., involved in the ecological configuration of the edaphic microbiome by their prodigious murphy and metabolic heterogeneity, whose we can distinguish phototrophic, heterotrophic, chemotrophic, azototrophic, and methylotrophic bacteria that play a key role in optimizing the nutrient load of the forest soil in organic acids, hydrogen sulfide, nitrite, ammonia, ferrous iron, nitrate, and methylamines [17]. In order to preserve their position within the edaphic microbiome, these bacteria act as plant growth bio-modulators by releasing auxiliary phytohormonal substances such as cytokinins, auxins, gibberellins, and abscisic acid, or their precursors as in the case of ethylene (1-aminocyclopropane-1-carboxylate deaminase); by excreting antibiotics such as phenazine, pyoluteorin, pyrrolnitrin, and diacetylphloroglucinol to biocontrol fungal and bacterial progression in the forest soil; by biopolymerizing glycoproteic biofilms to safeguard their physiological balance; and by counteracting the bioaccumulation of pollutants and chemical inputs through a superlytic enzymatic arsenal [17,18]. Besides these bacterial phyla, Verrucomicrobia is another taxonomic group (Opitutus terrae, Candidatus Xiphinematobacter spp., Candidatus Udaeobacter copiosus, Luteolibacter pohnpeiensis, etc.), non-demanding, Gram-negative, and with a verrucose morphology. It is widespread in most forest soils (16–18% of the microbiome), especially in those rich in humus and plant litter, from which it derives energy mainly through aerobic respiration, in synchrony with the reduction of the structural complexity of the forest soil’s organic fractions [19,20].
The intelligent metabolic networks of these bacteria, indispensable to germination, confer them a high fertilizing potential by strengthening the soil status in carbon dioxide, ammonium, nitrate, inorganic phosphate, and short-chain fatty acids, in addition that they ensure them an exceptional resilience to inauspicious biotic and abiotic conditions through polysaccharide biofilms that are favorable to phytomicrobial defense and cooperation [19,21].
Otherwise, Actinobacteria represent a lineage of filamentous, Gram-positive microbes, mostly strict aerobes and non-sporulating (e.g., Streptomyces spp., Micromonospora spp., Nocardia spp., Kitasatospora spp., etc.), which profoundly shape the structure of the edaphic microbiocenosis. The genome of these bacteria is characterized by its richness in guanine–cytosine pairs, expressing a wide range of bioactive secondary metabolites and catalytic enzymes (cellulase, amylase, xylanase, hemicellulases, proteases, lipases, ligninases, peroxidases, laccases, phosphatases, chitinases, and nitrogenases) that contribute to the bioremediation of the edaphic niche by degrading hydrocarbons, cyclic paraffins, and other aromatic pollutants [22,23]. Certain species, such as Frankia casuarinae, Frankia discariae, Frankia alni, and Frankia elaeagni, are endowed with a remarkable power to establish symbiotic root nodules in actinorhizal plants so as to inject them with soluble nitrogen, whereas others such as Streptomyces spp. and Micromonospora spp., secrete broad-spectrum antimicrobial biomolecules (streptomycin, chloramphenicol, vancomycin, tetracycline, neomycin, clavulanate, and actinomycin) to modulate competitive interactions and ensure the survival of the holobiont [23].
In parallel, the fungal pool represents an intrinsic pillar of the soil microbial flora, strongly supporting the fertile sustainability of forest ecosystems, within which two highly bio-engaged categories “the saprophytes and the mycorrhizal fungi” [24]. Soil saprophytic fungi are mainly dominated by the presence of aerobic molds (Aspergillus niger, Mucor hiemalis, Chaetomium globosum, Rhizopus stolonifer, Trichoderma harzianum, Penicillium chrysogenum, Fusarium solani, Humicola grisea, etc.), which possess rapid sexual or asexual dissemination processes at the expense of organic wastes (carcasses, compost, excrements, dead wood and leaves, and other plant litter) to prevent their toxic accumulation for natural fauna and flora. These filamentous fungi are ecologically superflexible and exhibit an extraordinary bioenergetic potential owing to their strong secretory capacity of extracellular degrading enzymes (cellulases, amylases, lipases, mannanases, xylanases, chitinase, laccases, peroxidase, proteases etc) which depolymerize a vast spectrum of giant molecules such as cellulose, chitin, lignin, starch, proteins, and hemicellulose, thereby elaborating humus and releasing other carbon, nitrogen, phosphorus, and potassium substrates accessible to trees and other microbial partners [25,26].
Mycorrhizal fungi form a distinct class of saprophytes that are indispensable for the cohesion and fertilization of soil aggregates. They are obligatory heterotrophic filamentous organisms, whose energy metabolism and growth are fueled by sugars (glucose, maltose, sucrose, fructose, and/or phosphorylated hexoses) assimilated from tree roots. They constitute a subterranean, reticular, and complex biological infrastructure the “wood-wide web” that links plant roots, and facilitates the exchange of water, nutrients (carbon dioxide, nitrate, ammonium, inorganic phosphate, organic acids, etc.), and even bioactive metabolites (phytohormones, polyphenols, enzymes, antibiotics etc.) between neighboring trees and/or between trees and the soil microbial population [27]. Although the majority of mycorrhizal fungi are strict symbionts with a constrained metabolic horizon, certain species are able to express specific extracellular enzymes (cellulases, glucanases, pectinases, phosphatases, xylanases, proteases, phytases, catalases, and oxidoreductases) in order to metabolize surrounding biological residues, access essential nutrient oligoelements (nitrogen, sulfur, phosphate, etc.), detoxify free radicals, and supply the soil with organic acids such as oxalic acid and citric acid. Other mycorrhizal fungal taxa are genetically more advanced by biosynthesizing molecules of antibacterial and antifungal interest (terpenoids, peptaibols, glomalin, and other volatile derivatives) to defend themselves against the dissemination of invasive microorganisms and to protect the plant host from certain root diseases. In parallel, some species are able to elaborate cryoprotective molecules that increase the physiological and metabolic plasticity of the holobiont against thermal fluctuations [27,28]. It should be noted that the family of mycorrhizal fungi is subdivided into two groups according to their root position, whose the endomycorrhizal fungi (Glomus intraradices, Glomus etunicatum, Acaulospora scrobiculata, Scutellospora calospora, Acaulospora laevis, Gigaspora margarita, etc.) are installed within the root cortidal cells by forming visicles and arbuscules, and the ectomycorrhizal fungi (Amanita phalloides, Boletus pinophilus, Laccaria bicolor, Russula cyanoxantha, Suillus luteus, Tricholoma matsutake, etc.) are adhered to the roots without invading the cells by emerging giving a network called the Hartig net [29,30].
The development of molecular biology has revealed that certain species from the archaeal kingdom (Nitrososphaera viennensis, Candidatus, Nitrosotenuis uzonensis, Methanobacterium formicicum, Methanocella arvoryzae, Methanosaeta concilii, Nitrososphaera gargensis, etc.) are strongly present in the microbiome of forest soils, particularly those that are poorly aerated, peat-rich, dry, saline, acidic, and/or low in nutrient resources. These microorganisms are endowed with emergent properties of resilience and functional redundancy, whose they express thousands of genes to maintain the geochemical flow of nitrogen, methane, sulfur and carbon by playing a crucial role in the underground economy of forest soils [31].
On the other hand, [32] has revealed that the forest soil microbiome is crowded with protists such as Amoebozoa, Apicomplexa, Cercozoa, Ciliophora, etc., and with viruses such as bacteriophages, phytoviruses and mycoviruses, which they weave super-important symbiotic, parasitic, predatory or disease-carrying interactions with trees and other microorganisms, to balance the bacterial and fungal load of the microbiome, to supervise the rate of afforestation and to support horizontal gene mobilization and genomic adaptation of the holobiont.
For a long time, the ecobiological signature of algae on the biogeochemical balance of forest ecosystems was underestimated. However, the rise of soil microbiology has re-evaluated their importance, pointing out that the multifunctional and protective properties of these cryptic autotrophic organisms make them a pioneering contributor to the architecture and dynamics of the microbiological and inert crust of the soil, in addition to supporting the mineral nutrition of trees. Macroalgae are excluded from this concept since they are multicellular and occupy marine aquatic environments. In contrast, microalgae are concerned since they are unicellular, photosynthetic species, mostly strict aerobes and sometimes facultative aero-anaerobes, occupying especially the superficial horizons of hydromorphic, peaty, and acidic forest soils in order to access light, oxygen, and fresh organic debris easily. The taxonomy of microalgae reveals that certain species belong to the phylum of cyanobacteria (blue-green algae such as Nostoc commune, Scytonema sp., Microcoleus vaginatus, Tolypothrix sp., Anabaena sp., Calothrix sp., etc.), which are heterocystous prokaryotes, characterized by photosynthetic infrastructures rich in chlorophyll A, phycoerythrin, and phycocyanin [33,34,35]. On the other hand, the great part of these microorganisms belong to the domain of eukaryotes, among which are distinguished: Chlorophyta (green algae such as Chlorella vulgaris, Klebsormidium flaccidum, Trebouxia spp., Scenedesmus obliquus, Chlamydomonas reinhardtii, etc.) endowed with intelligent light receptors of the chlorophyll A and B type; Xanthophyta (yellow-green algae such as Botrydium granulatum, Characiopsis spp., Heterococcus spp., Tribonema spp., etc.), whose their photosynthetic apparatus is composed of carotenoids, chlorophylls A and C, and rarely fucoxanthin; Eustigmatophyta, whose their plastids contain chlorophyll A and violaxanthin, as in Eustigmatos magnus, Eustigmatos vischeri, Eustigmatos polyphem, Monodopsis subterranea, Pseudocharaciopsis minuta; Chrysophyta (golden algae such as Mallomonas spp., Dinobryon spp., Ochromonas spp., Synura spp., etc.), characterized by the photosynthetic pigments fucoxanthin and chlorophylls A and C; Dinophyta, whose their photosynthetic system is composed of peridinin and chlorophylls A and C, as in Ceratium hirundinella, Symbiodinium spp., and Alexandrium spp.; Diatoms (Bacillariophyceae such as Navicula spp., Eunotia spp., Cymbella spp., Pinnularia spp., Surirella spp., etc.), whose their photosynthetic bioenergy depends on the presence of fucoxanthin and chlorophylls A and C; and Cryptophyta such as Cryptomonas spp., Komma caudata, Rhodomonas spp., Chroomonas spp., etc., whose their photosynthetic cycle is sustained by phycocyanin, chlorophylls A and C, and phycoerythrin [33,36]. Besides the fact that the edaphic algal microflora is capable of synthesizing its own energy sources (reducing and complex sugars) while releasing oxygen into nature and supplying other heterotrophic microbial contingencies of the microbiome, certain species are diazotrophic due to their genetic repertoire of nitrogenases, that convert gaseous nitrogen into edaphic ammonium that is bioavailable for the nitrogen metabolism of trees and seeds, whereas other microalgal strains shape soil microhabitats by generating exopolysaccharides and mucilages and by establishing symbiotic interactions with fungi, particularly ascomycetes forming forest lichens that increase hydration, infiltration, and stabilization of the soil molecular aggregates. Microalgae possess a highly developed ecophysiological plasticity to overcome periods of unfavorable thermal, water, light, and nutrient fluctuations, through the strengthening of their protective intracellular osmolyte pool (glycerol and trehalose) and the formation of thick zygotes and akinetes, whose their revitalization significantly shapes the physical, biological, and functional engineering of the soil [37,38].
This heterogeneity of taxonomic lineages renders the edaphic microbiome a genuine living ecological memory, which responds to pedoclimatic fluctuations through the adjustment of the holobiont’s metabolic functions in order to prolong its lifetime [15]. Table 1 summarizes the different types of microbiomes and algae that play a crucial role in the proper functioning of forest ecosystems and plant development.
3. Functional Mechanisms Enhancing Seed Germination
Microbes and algae (Figure 1), are biological resources that can naturally promote seed germination in forests by preparing and helping seeds for germination to support the early life stages of forest plants [39].
Forest soil microbes comprise diverse communities of interacting microorganisms that inhabit the rhizosphere, plant roots, and surrounding soil. These communities include bacteria, fungi, and archaea, as well as their symbiotic counterparts, protists, and viruses [40]. Table 2 highlights their vital ecosystem functions through biophysical, biochemical, and ecological mechanisms, such as nutrient availability for new tissue growth in germinating seeds, soil structure improvement, and pathogen suppression.
Microbes maintain nutrient cycling by degrading organic matter, detoxifying toxins, and maintaining mineral balance [46]. These microbes may also regulate the redox state of nutrients required for the formation of organic compounds. They also stabilize nutrients in available forms, mineralize mineral nutrients, and regulate nitrogen fixation and phosphorus acquisition [47]. In nutrient-deficient soils, the direct transfer of nutrients from microbiome cells to developing seedlings improves their availability [48]. Some microbes promote seed germination through the biosynthesis of plant hormones, such as auxins, gibberellins, and cytokinins. These hormones directly stimulate seed metabolism, promoting cell division and elongation, which accelerates germination and root development [49].
Furthermore, forest soil microbes can alter soil structural properties by influencing the soil matrix and their ability to form multi-species biofilms, which promotes seed germination, improving soil properties, water retention and gas exchange that are factors directly related to seed germination [50]. Moreover, the presence of a healthy microbiome on and around the seed can protect it from environmental stressors and pathogens, by inducing systemic resistance in the plant to stressors such as drought and salinity by producing osmoprotectants, which help the plant retain water [51]. In addition, it acts as a biological control agent, producing compounds that inhibit the growth of harmful pathogens present in the soil, which helps create a protective barrier around the seed, thus reducing the risk of disease and rot [52].
Algae form a diverse community ranging from unicellular microalgae to aquatic macroalgae. In forests, algae are deposited on surfaces by rain or wind and act as biological promoters of seed germination [53]. Therefore, the role of algae in forest regeneration is crucial. These photosynthetic organisms, along with their microbiomes, promote germination by sharing similar biological functions. They convert inorganic carbon into biomass, which contributes significantly to nutrient and oxygen levels during the early stages of seed germination and throughout subsequent stages. They also accumulate phosphate and distribute nutrients to the surrounding environment. According to [54] soils in evergreen broadleaf forests typically contain Pseudanabaena, which fixes atmospheric nitrogen (N2) and accumulates inorganic nutrients such as phosphorus and potassium. While chlorophyceous, xanthophyceous, and cyanophyceous algae fix carbon through photosynthesis and then secrete carbohydrates, organic acids, and amino acids into the soil [55]. In addition, a portion of this fixed carbon forms extracellular polymeric materials that bind sand particles, altering soil texture to improve water and mineral nutrient retention [38]. [56] highlighted that Cyanobacteria also utilizes both atmospheric carbon dioxide and nitrogen to provide organic carbon and nitrogen compounds, enhancing soil fertility. Meanwhile, algae with filamentous structures facilitate the aggregation of soil particles, and some species produce exopolysaccharides capable of adhering to soil particles. Furthermore, some algae, whose secondary metabolites inhibit the growth of fungi such as Fusarium [57].
4. Microbiome–Algae Interactions in the Soil–Seed Interface
4.1. Synergistic Effects on Nutrient Cycling and Bioactive Compound Production
Microbiome’s soil and algae often interact synergistically at the seed-soil interface, enhancing nutrient cycling and producing compounds that stimulate germination. In biological communities of the soil crust, co-inoculation of bacteria and algae has been shown to significantly increase soil nutrient availability beyond that achieved by single inoculants. For instance, [58] reported that co-inoculation increased ammonium and nitrate levels by 10–17% compared to controls. Other studies have also shown that treatments with microbial agents improved soil fertility, nitrogen content and microbial quantity [59]. Particularly, [60] revealed that algae contribute through photosynthesis-driven carbon fixation and by releasing organic exudates rich in sugars, amino acids, and other nutrients. These algal exudates feed heterotrophic microbes and drive nutrient cycling processes such as nitrogen fixation and phosphorus solubilization by bacteria. In turn, microbial metabolism provides inorganic nutrients that algae can assimilate. This positive feedback loop accelerates nutrient turnover in the seed microsite, creating a more fertile zone around the germinating seed [61]. Microbiome-algae consortia also generate bioactive compounds that can directly influence seed germination. Most terrestrial algae produce phytohormones and metabolites that stimulate growth. Furthermore, seed dressing with microalgae has been shown to increase seedling vigor and growth, an effect attributed to the algal production of growth-promoting substances and vitamins [62].
4.2. Competitive and Facilitative Relationships
Within the seed microsite, algae and other microorganisms engage in competitive and facilitative interactions that shape the balance of the community. Competition can occur for resources such as minerals, water, or space in the seed layer. For instance, algae and bacteria can both use available nitrogen or carbon sources; an uncontrolled algal bloom can temporarily immobilize nutrients or overshadow some microbes. However, in many cases, a facilitative and mutualistic relationship dominates [63]. Algae often produce oxygen and organic matter through photosynthesis, which benefits nearby aerobic bacteria and fungi. Algal photosynthesis increases local O₂ levels in moist soil films, mitigating hypoxia around the seeds and thus favoring aerobic microbes, beneficial to plants, over anaerobic competitors or pathogens. As illustrated in Figure 2, soil bacteria support algae by releasing CO₂ and transforming soil nutrients into forms that algae can absorb [64]. For instance, bacterial ammonification and nitrification provide inorganic nitrogen that algae can assimilate, while bacterial release of B vitamins and other growth factors can stimulate algal proliferation, a well-documented interdependence in algal-bacterial consortia. These exchanges create a syntrophic partnership: algae feed the microbes’ photosynthate and exopolysaccharides, and the microbes reciprocate with nutrients and CO₂, reducing direct competition [65].
4.3. Impact on Seed Microenvironment and Germination Rates
By modifying the soil environment, microbiome-algae interactions can have profound effects on seed microclimate and germination success. An important impact is improving soil structure and moisture retention around the seed. According to [66], algal filaments and exopolysaccharides act as biopolymers and bind soil particles into stable aggregates. In several experiments, inoculation with algal-bacterial consortia increased the percentage of protective macroaggregates in the soil, halved the proportion of destabilized microaggregates, and significantly improved aggregate stability indices compared to uninoculated soil. This aggregation, coupled with the hygroscopic nature of algal polysaccharides, allows the soil near the seed to retain more water [67]. In fact, co-inoculated soils maintain a higher volumetric moisture content, creating a constantly moist microenvironment that prevents seed desiccation during the critical stages of imbibition and root emergence. Furthermore, as emphasized by [68], the improved structure also increases aeration and reduces soil compaction around the roots of germinating seedlings.
5. Forest Seed Germination: Processes and Ecological Significance
5.1. Physiological Stages of Seed Germination in Forest Species
Seed germination is a physiological process that goes through several stages, and forest seeds follow the same stages as in other plants; although, often with adaptations of dormancy appropriate to their ecosystem. The process begins with imbibition, where the dry seed rapidly absorbs water upon contact with moist soil [69]. Imbibition causes the seed to swell and reactivate metabolic pathways. In forest soils, this initial water absorption can be influenced by the soil’s water-holding capacity and temperature. Once a critical level of hydration is reached, the seed enters the lag phase, during which cellular metabolism accelerates. A recent study [70] highlights that enzymes are synthesized or activated to mobilize stored reserves in the endosperm or cotyledons. For instance, in many seeds, enzymes such as α-amylase begin to convert starch into sugars to feed the growing embryo, a process under hormonal control by gibberellins and other phytohormones. If the forest seed was dormant, dormancy should be broken during this phase: in some species, this occurs after a required trigger. Once any barriers to dormancy are overcome such as the release of inhibitors, the softening of the hard layer, or the fulfillment of stratification requirements, the embryo resumes growth [71]. The final stage is root elongation, where the root breaks through the seed coat and enters the soil. This marks the visible event of germination. In forest species, root emergence can be relatively slow. The time from imbibition to root emergence varies greatly: seeds of fast-germinating pioneer trees can complete it within a few days, while some dormant forest seeds take weeks or months. For instance, Quercus species germinate within 2–3 weeks, while Horsfieldia hainanensis showed a prolonged germination period of several months due to a state of physiological dormancy. After root emergence, the hypocotyl and shoot begin to grow, completing germination and entering seedling establishment. During these stages, [72] documented that the seed relies on internal reserves until it can photosynthesize; thus, seed size and reserve composition might significantly influence the duration of a forest seedling in low-light forest understory conditions before it needs external resources.
5.2. Differences Between Pioneer and Climax Forest Seeds
Forest seeds exhibit diverse germination traits and strategies, but there is a major ecological dichotomy between pioneer and climax species. These two groups have evolved contrasting seed traits to adapt to their regeneration sites [73]. Table 3 summarizes the key differences in seed traits and germination behavior between pioneer and climax forest species.
These changes reflect adaptive strategies. Pioneer species, faced with unpredictable opportunities driven by disturbances, struggle with their large numbers and ability to remain dormant until a gap opens up. Their small, dormant seeds can be widely dispersed and lie in wait as a continuous seed bank [76]. For instance, [75] highlighted that tropical pioneers in the Amazon produce seeds that require high irradiance or diurnal temperature fluctuations to induce germination, ensuring that they germinate only in the gaps of fallen trees with direct sunlight. In contrast, climax species invest each offspring with a large, well-stocked seed that can germinate under the parent canopy or in small gaps. Since such seeds cannot germinate for long in the ground, they usually germinate in the next favorable season. The lack of a long-term seed bank in climax species means that these trees rely more on consistent forest conditions and reliable seasonal signals; it also makes them more sensitive to disturbances that remove mature trees without immediate regeneration [74]. Ecologically, pioneer seeds promote rapid colonization and forest succession, while climax seeds support the maintenance and slow turnover of mature forest stands.
5.3. Case Studies of Threatened and Endangered Forest Seeds
Understanding seed germination traits is particularly critical for the conservation of threatened forest species. Many endangered trees present germination challenges, such as strict dormancy, low viability, or unstable seeds, which hinder natural regeneration and ex situ propagation [77]. For instance, a recent analysis of 953 tropical tree species on the Osa Peninsula of Costa Rica found that over a third are likely to be unstable, and this percentage increases to nearly half among species classified as endangered or endemic. Unstable seeds cannot withstand drying or freezing, and thus cannot be stored in seed vaults for long-term storage. They also remain viable for only short periods in the forest, creating a narrow window for germination. This trait poses a serious obstacle to conservation: if habitat conditions are unfavorable during the short period of seed viability, an entire reproductive cycle may be lost. Conservationists are exploring alternatives such as embryo cryopreservation or clonal propagation for such “exceptional” species [78]. Even forest species with orthodox seeds may be threatened if they have complex dormancy states or habitat-specific germination traits. The Hainan ironwood, an endangered tree in Asia, illustrates this, its seeds exhibiting a deep physiological state that requires a long post-maturation period, which historically ensured germination only under stable old-growth conditions [79]. In disturbed or changing climates, the necessary conditions may not occur, leading to poor regeneration. Similarly, many rare softwood trees have dormant seeds that require very specific stratification regimes. The critically endangered Yangbi maple, known from only a handful of individuals in China, has seeds that take an average of ~44 days to germinate under laboratory conditions, with only ~50% germination within 7–10 weeks. This slow and partial germination indicates a combination of dormancy and possibly low embryo vigor. Intensive studies on A. yangbiense showed that careful temperature controls and possibly hormonal treatments were necessary to improve germination, and ex situ seedling cultivation has been key to its rescue [80]. Another case is the tropical legume Karomia gigas, a critically endangered tree in East Africa. It produces very few seeds in the wild, and those seeds have low innate viability and possibly inhibitory compounds [81]. Germination trials have had limited success, hampering recovery efforts. Conservationists have had to resort to vegetative propagation and the establishment of living conservation orchards, as seed-based regeneration has been highly unreliable. This pattern is seen in many endangered forest plants: seed dormancy and low germination are often cited as barriers to population restoration [82]. In response, researchers are developing species specific germination protocols, for example, using steam or heat shock to induce germination in rare Fynbos shrubs, or applying GA₃ to stimulate germination in dormant conifer seeds of endangered spruce species [83]. Table 3 already highlighted that many climax species do not form permanent seed banks; accordingly, if mature trees are removed, the lack of a seed bank means that the species cannot easily be reestablished. This has been observed with the endangered Wollemi pine in Australia although its seeds are orthodox, in the wild it reproduces mostly by cuttings, and seedling recruitment is rare [84]. Conservation efforts have been to collect seeds immediately after the first few years and germinate them in nursery conditions to create reserve populations. Threatened forest seeds often incorporate the most extreme germination traits: long dormancy cycles, resistance, or very narrow germination requirements [85]. These traits, while once adaptive in ancient, stable forests, now complicate conservation under rapidly changing conditions. The ecological importance is twofold: such seeds highlight the importance of preserving intact habitats, and they spur innovation in seed technology and ex situ propagation. By studying the germination ecology of threatened trees, scientists might design interventions ranging from simulating environmental cues to biotechnological aids to prevent species extinction [86,87]. Effective conservation of forest biodiversity thus depends on combining ecological understanding of seed germination processes with practical strategies to overcome the germination barriers presented by many climax forest species. Every threatened seed that is persuaded to germinate in a nursery is not just a new plant, but a lifeline for the future of its species in a stressed ecosystem [88].
6. Biotechnological Applications for Enhancing Forest Seed Germination
Biotechnological applications using microbiome–algae consortia present sustainable options to stimulate forest seed germination and seedling establishment. The use of biotechnological applications involving microbiome–algae consortia presents a viable method for increasing forest seed germination and promoting large-scale reforestation programs. Recent findings of [89] suggested that seed bio-priming with microbial inoculants and algal extracts can enhance seed vigor, quicken germination time, and enhance tolerance to abiotic stresses such as salinity and drought, due to promoting phytohormone production, nutrient mobilization and activating resistance pathways. Seed coatings containing beneficial bacteria (e.g., Pseudomonas, Rhizobium, Bacillus) and mycorrhizal fungi, along with cyanobacteria and green algae, enhance nutrient uptake, soil stability, and biological protection against pathogens [90]. Additionally, biopolymers and exopolysaccharides from microalgae can be added to seed coating matrices to support seed soaking, and improve soil aggregation and mineral retention, thereby it provides a hospitable microhabitat for seedling establishment [91]. The application of engineered microbial consortia also allows for biofertilizers to be designed to help with dormancy breaking, seed coat softening, and early root elongation, which are especially relevant to forest regeneration in degraded soil systems [92]. All of these environmentally sustainable approaches could effectively replace chemical seed treatments, aiding in forest ecosystem resilience and biodiversity conservation. Taken together, these advances illustrate an important transformative potential of biotechnological approaches based on microbiome–algae, which are ecological alternatives to chemical management. These approaches provide scalable methods to facilitate forest recovery, support ecosystem resilience, and promote long-term biodiversity conservation.
7. Challenges and Limitations
Despite the potential benefit of using microbiome–algae consortia in promoting the germination of forest seeds, a number of associated difficulties remain. Most notably, variability and complexity in seed–microbiome interactions can be a significant limitation as they depend on the species, environment, and developmental stage [93]. Microbial community stability on seeds may be disrupted by abiotic stressors (e.g., drought, salinity, and temperature), thus making it difficult to predict an outcome from inoculation. In addition, there are strain-specific responses to microbial inoculants, which do not lend to universal bioformulations; some consortia promote germination, while others may compete with seeds or the local microbiome and therefore, be less effective [94]. A further major obstacle to success is the limited ability of inoculated microbes to survive and establish in natural soils, where native communities will outcompete introduced strains. Additionally, while microalgae–bacteria consortia are useful in theory, they require very specific nutrient and humidity conditions that may not always be available in nutrient-poor or degraded forest soils to thrive truly [95]. Other limitations associated with the technical and logistical challenges of larger-scale production, shelf-life of bioinoculants, and cost-prevention will limit bioinoculant applications. Finally, a standardized protocol for determining germination-promoting quality for forest species has not yet been established; this reduces the capacity to compare results and apply laboratory findings to field settings. Consequently, although microbiome-algae consortia provide exciting opportunities for improving forest seed germination, it will be critical to overcome biological, ecological, and technical barriers to ensure that results found in the laboratory can be reliably scaled and applied for field use.
8. Future Research Directions
Future studies should emphasize multi-omics approaches such as metagenomics, metabolomics, and transcriptomics to explore functional interactions between seed-associated microbiomes and their algal partners in multiple ecological contexts. According to [16], the application of machine learning and predictive modeling represents an exciting opportunity for identifying the best fitting microbial consortia for specific forest species and restoration conditions. In addition, advancements in synthetic biology could enable the design of microbial strains with abiotic stress tolerance and optimized phytohormone production, thereby is essential improving plant establishment and resilience [96,97].
From an applied perspective, microbial–algal bio-priming seed coatings with biopolymers provide a sustainable alternative to chemical treatments, which have shown potential to increase germination performance under degraded soil conditions [98]. Incorporating biotechnological developments alongside traditional ecological restoration methods—such as direct seeding and assisted natural regeneration—could greatly facilitate forest restoration in biodiversity hot spots [99,100]. Furthermore, as indicated by [101,102], future studies are needed to assess policy, regulatory, and socio-economic aspects to support safely, scalable, and broadly acceptable microbial–algal bioformulations for deployment in large-scale reforestation programs. In general, the integration of multi-omics science, synthetic biology, and microbial–algal bio-priming into existing ecological restoration techniques in association with supportive policy and socio-economic systems offers great potential for expanding viable and resilient forest regeneration in response to a changing environment.
9. Conclusions
Forests represent one of most complex and critical ecosystems on the planet; however, their stability and ability to recover from disturbance are increasingly compromised by human-caused disturbances, climate change, and other changes to ecosystems. At the center of any renewal process is the successful germination of forest seeds, a phenomenon involving complex physiological, molecular, and ecological interactions. This review emphasized that seed germination is not simply the outcome of genetic and hormonal regulation. However, it will also be shaped and regulated by the microbiome–algae consortia that surround seeds in the soil. It highlights that the seed–microbiome–algae might act as an essential functional unit for supporting biodiversity in forests, improving soil fertility, and mitigating against environmental stress. It also explains that microbiome–algae interactions contribute to plant seed vigor and germination by their roles in nutrient cycling, soil stabilization, and biocontrol.
Nevertheless, the review determined that there are still significant challenges. This study showed that seed-associated microbiome variability varies by species and environmental conditions, and the complexity of ecological relationships is still an area of research. Furthermore, the current study demonstrates that advances in synthetic biology and precision biotechnology, especially for engineered microbial strains could potentially assist in enhancing forest regeneration in disturbed areas.
In conclusion, this research demonstrates that safeguarding and rehabilitating forest ecosystems in the 21st century will mostly depend upon our capability to leverage the ecological and bio-technological potential of microbiome–algae–seed interactions. Integration of ecological theory with applied technology could provide viable and practical solutions to strengthen forest resilience, conserve biodiversity, and enhance ecological sustainability on a global scale.
Author Contributions
Conceptualization, B.Xh. and R.S.; methodology, B.Xh., R. S. and H. S.; writing—original draft preparation, B.Xh., R.S., H.S., H.K., and G.Gj; writing—review and editing, B.Xh. and R.S.; visualization, R.S., and H.K.; supervision, B.Xh. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Aguirre-Gutiérrez, J.; Díaz, S.; Rifai, S.W.; Corral-Rivas, J.J.; Nava-Miranda, M.G.; González-M, R.; Hurtado-M, A.B.; Revilla, N.S.; Vilanova, E.; Almeida, E.; de Oliveira, E.A. Tropical forests in the Americas are changing too slowly to track climate change. Science 2025, 387, eadl5414. [Google Scholar] [CrossRef] [PubMed]
- Manojkumar, S.; Maurya, L.L.; Kumar, S.; George, E. Impact of Forest Degradation on Forest Regeneration. In Forest Degradation and Management. 2025 (pp. 195–206). Springer, Cham.
- Atakpama, W.; Egbelou, H.; Bali, B.E.; Ahouandjinou, E.B.; Dibegdina, M.; Batawila, K. Challenges for the conservation and restoration of the forest ecosystems of the Oti-Kéran-Mandouri Biosphere Reserve in Togo. Ann. Rech. For. Alge rie. 2025, 15, 17–30. [Google Scholar]
- Liu, X.; Feng, Y.; Hu, T.; Luo, Y.; Zhao, X.; Wu, J.; Maeda, E.E.; Ju, W.; Liu, L.; Guo, Q.; Su, Y. Enhancing ecosystem productivity and stability with increasing canopy structural complexity in global forests. Science Advances. 2024, 10, eadl1947. [Google Scholar] [CrossRef]
- Lussier, N.M.; Crafford, R.E.; Reid, J.L.; Kwit, C. Seeding success: Integrating seed dispersal networks in tropical forest restoration. Biotropica. 2025, 57, e13347. [Google Scholar] [CrossRef]
- Bargah, A.S.; Sharma, D.; Kumar, R.; Nag, R.; Pradhan, R. Enhancing germination of forest tree seeds in Chhattisgarh through PGR-based treatments: A review. Journal of Advances in Biology & Biotechnology. 2025, 28, 851–863. [Google Scholar]
- Mamani, G.Q.; Duarte, M.L.; Almeida, L.S.; Martins Filho, S. Non-parametric survival analysis in seed germination of forest species. Journal of Seed Science. 2024, 46, e202446036. [Google Scholar] [CrossRef]
- Zuhri, M.; Setiawan, N.N.; Dewi, S.P.; Sulistyawati, E. Seed germination variability and its association with functional traits in submontane tropical forest species of Indonesia: recommendations for direct seeding. Forest Science and Technology. 2025, 21, 64–73. [Google Scholar] [CrossRef]
- Yang, P.; Lu, L.; Condrich, A.; Muni, G.A.; Scranton, S.; Xu, S.; Xia, Y.; Huang, S. Innovative Approaches for Engineering the Seed Microbiome to Enhance Crop Performance. Seeds. 2025, 4, 24. [Google Scholar] [CrossRef]
- Ahmed, S.; Singh, P.; Middleton, J.; Merritt, D.; Jenkins, S.; Nichols, P. Impact of seed maturation on the morphology, nutrition, microbiome composition and germinability of subterranean clover (Trifolium subterraneum) seeds. Grass and Forage Science. 2025, 80, e12725. [Google Scholar] [CrossRef]
- Murugesan, S.; Priya, P.; Sivamurugan, V. Microbial Interactions in Soil Algae. In Soil Algae: Morphology, Ecology and Biotechnological Applications. 2025 (pp. 1–35). Singapore: Springer Nature Singapore.
- Shukla, S.; Upadhyay, D.; Mishra, A.; Jindal, T.; Shukla, K. The Impact on Soil Ecology of the Algal Community. In Soil Algae: Morphology, Ecology and Biotechnological Applications. 2025 (pp. 193–219). Singapore: Springer Nature Singapore.
- Onet, A.; Grenni, P.; Onet, C.; Stoian, V.; Crisan, V. Forest soil microbiomes: a review of key research from 2003 to 2023. Forests. 2025, 16, 148. [Google Scholar] [CrossRef]
- Gomes, S.I.; Gundersen, P.; Bezemer, T.M.; Barsotti, D.; D’Imperio, L.; Georgopoulos, K.; Justesen, M.J.; Rheault, K.; Rosas, Y.M.; Schmidt, I.K.; Tedersoo, L. Soil Microbiome Inoculation for Resilient and Multifunctional New Forests in Post-Agricultural Landscapes. Global Change Biology. 2025, 31, e70031. [Google Scholar] [CrossRef]
- Jansson, J.K.; McClure, R.; Egbert, R.G. Soil microbiome engineering for sustainability in a changing environment. Nature Biotechnology. 2023, 41, 1716–1728. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, E.A.; Suleymanov, A.R.; Nikitin, D.A.; Semenov, M.V.; Abakumov, E.V. Machine learning-based mapping of Acidobacteriota and Planctomycetota using 16 S rRNA gene metabarcoding data across soils in Russia. Scientific Reports. 2025, 15, 23763. [Google Scholar] [CrossRef]
- Bereczki, K.; Tóth, E.G.; Szili-Kovács, T.; Megyes, M.; Korponai, K.; Lados, B.B.; Illés, G.; Benke, A.; Márialigeti, K. Soil Parameters and Forest Structure Commonly Form the Microbiome Composition and Activity of Topsoil Layers in Planted Forests. Microorganisms. 2024, 12, 1162. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Wu, S.; Pan, H.; Lu, X.; Liu, Y.; Yang, L. Effect of forest fires on the alpha and beta diversity of soil bacteria in taiga forests: proliferation of rare species as successional pioneers. Forests. 2024, 15, 606. [Google Scholar] [CrossRef]
- Rakitin, A.L.; Kulichevskaya, I.S.; Beletsky, A.V.; Mardanov, A.V.; Dedysh, S.N.; Ravin, N.V. Verrucomicrobia of the family Chthoniobacteraceae participate in xylan degradation in boreal peat soils. Microorganisms. 2024, 12, 2271. [Google Scholar] [CrossRef]
- Wang, Y.; Lv, Y.; Kavana, D.J.; Zhang, G.; He, S.; Yu, B. The Characteristics of Soil Microbial Community Structure, Soil Microbial Respiration and their Influencing Factors of Three Vegetation Types in Alpine Wetland Ecosystem. Wetlands. 2024, 44, 104. [Google Scholar] [CrossRef]
- Di, X.; Li, P.; Wang, J.; Nowak, V.; Zhi, S.; Jin, M.; Liu, L.; He, S. Genome Mining Analysis Uncovers the Previously Unknown Biosynthetic Capacity for Secondary Metabolites in Verrucomicrobia. Marine Biotechnology. 2024, 26, 1324–1335. [Google Scholar] [CrossRef]
- Rebai, H.; Sholkamy, E.N.; Abdelhamid, M.A.; Prakasam Thanka, P.; Aly Hassan, A.; Pack, S.P.; Ki, M.R.; Boudemagh, A. Soil actinobacteria exhibit metabolic capabilities for degrading the toxic and persistent herbicide Metribuzin. Toxics. 2024, 12, 709. [Google Scholar] [CrossRef]
- Kapoor, P.; Joshi, P.; Pal, M.; Parkash, V. Actinobacteria as Proficient Biocontrol Agents for Combating Fungal Diseases in Forest Plant Species. Journal of Basic Microbiology. 2025, 2, e70030. [Google Scholar] [CrossRef]
- Akter, S.; Mahmud, U.; Shoumik, B.A.; Khan, M.Z. Although invisible, fungi are recognized as the engines of a microbial powerhouse that drives soil ecosystem services. Archives of Microbiology. 2025, 207, 79. [Google Scholar] [CrossRef]
- Li, X.; Qu, Z.; Zhang, Y.; Ge, Y.; Sun, H. Soil fungal community and potential function in different forest ecosystems. Diversity. 2022, 14, 520. [Google Scholar] [CrossRef]
- Ning, Q.; Chen, L.; Zhang, C.; Ma, D.; Li, D.; Han, X.; Cai, Z.; Huang, S.; Zhang, J. Saprotrophic fungal communities in arable soils are strongly associated with soil fertility and stoichiometry. Applied Soil Ecology. 2021, 159, 103843. [Google Scholar] [CrossRef]
- Fall, A.F.; Nakabonge, G.; Ssekandi, J.; Founoune-Mboup, H.; Apori, S.O.; Ndiaye, A.; Badji, A.; Ngom, K. Roles of arbuscular mycorrhizal fungi on soil fertility: contribution in the improvement of physical, chemical, and biological properties of the soil. Frontiers in fungal biology. 2022, 3, 723892. [Google Scholar] [CrossRef]
- Chaudhary, A.; Poudyal, S.; Kaundal, A. Role of arbuscular mycorrhizal fungi in maintaining sustainable agroecosystems. Applied Microbiology. 2025, 5, 6. [Google Scholar] [CrossRef]
- Ediriweera, A.N.; Lu, W.; Perez Moreno, J.; Kalamulla, R.; Mayadunna, N.; Pelewatta, A.; Dissanayake, G.; Maduwanthi, I.; Wijesooriya, M.; Dai, D.Q.; Yapa, N. Ectomycorrhizal fungal symbiosis on plant nutrient acquisition in tropical ecosystems. New Zealand Journal of Botany. 2025, 2, 1–24. [Google Scholar] [CrossRef]
- Ballaoui, M.; El Gabardi, S.; Chliyeh, M.; Selmaoui, K.; Saidi, N.; Alaoui, M.A.; Mouden, N.; Benkirane, A.R.; Ouazzani Touhami, A.; Douira, A. Diversity of Endomycorrhizal Fungi in the Rhizosphere of Fig Trees in the Region of Ifrane (Middle Atlas Region of Northern Morocco). In Advanced Systems for Environmental Monitoring, IoT and the application of Artificial Intelligence. 2024 (pp. 109–122). Cham: Springer Nature Switzerland.
- Song, B.; Fang, J.; Yu, Z.; Liu, Z.; Li, N.; Pena, R.; Hu, Z.; Xu, Z.; Adams, J.M.; Razavi, B.S. The development of biological soil crust along the time series is mediated by archaeal communities. Geoderma. 2024, 449, 117022. [Google Scholar] [CrossRef]
- Ma, L.; Zhang, L.; Zhang, S.; Zhou, M.; Huang, W.; Zou, X.; He, Z.; Shu, L. Soil protists are more resilient to the combined effect of microplastics and heavy metals than bacterial communities. Science of the total environment. 2024, 906, 167645. [Google Scholar] [CrossRef] [PubMed]
- Lorentz, J.F.; Calijuri, M.L.; Rad, C.; Cecon, P.R.; Assemany, P.P.; Martinez, J.M.; Kholssi, R. Microalgae biomass as a conditioner and regulator of soil quality and fertility. Environmental Monitoring and Assessment. 2024, 196, 198. [Google Scholar] [CrossRef] [PubMed]
- Yeheyo, H.A.; Ealias, A.M.; George, G.; Jagannathan, U. Bioremediation potential of microalgae for sustainable soil treatment in India: a comprehensive review on heavy metal and pesticide contaminant removal. Journal of Environmental Management. 2024, 363, 121409. [Google Scholar] [CrossRef]
- Zhang, C.; Yu, X.; Laipan, M.; Wei, T.; Guo, J. Soil health improvement by inoculation of indigenous microalgae in saline soil. Environmental Geochemistry and Health. 2024, 46, 23. [Google Scholar] [CrossRef]
- Singh, J.; Saxena, R.C. An introduction to microalgae: diversity and significance. In Handbook of marine microalgae. 2015 (pp. 11–24). Academic Press.
- Alvarez, A.L.; Weyers, S.L.; Goemann, H.M.; Peyton, B.M.; Gardner, R.D. Microalgae, soil and plants: A critical review of microalgae as renewable resources for agriculture. Algal Research. 2021, 54, 102200. [Google Scholar] [CrossRef]
- Abinandan, S.; Subashchandrabose, S.R.; Venkateswarlu, K.; Megharaj, M. Soil microalgae and cyanobacteria: the biotechnological potential in the maintenance of soil fertility and health. Critical reviews in biotechnology. 2019, 39, 981–998. [Google Scholar] [CrossRef]
- Gerna, D.; Clara, D.; Antonielli, L.; Mitter, B.; Roach, T. Seed imbibition and metabolism contribute differentially to initial assembly of the soybean holobiont. Phytobiomes journal. 2023, 8, 21–33. [Google Scholar] [CrossRef]
- Li, F.; Zi, H.; Sonne, C.; Li, X. Microbiome sustains forest ecosystem functions across hierarchical scales. Eco-Environment & Health. 2023, 2, 24–31. [Google Scholar]
- Li, D.; Chen, W.; Luo, W.; Zhang, H.; Liu, Y.; Shu, D.; Wei, G. Seed microbiomes promote Astragalus mongholicus seed germination through pathogen suppression and cellulose degradation. Microbiome. 2025, 13, 23. [Google Scholar]
- Rafiq, I.; Reshi, Z.A.; Bashir, I. Diversity and Plant Growth-Promoting Activities of Culturable Seed Endophytes in Abies pindrow (Royle ex D. Don) Royle: Their Role in Seed Germination and Seedling Growth. Current Microbiology. 2025, 82, 82. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, D.E.; Sevanto, S.; Peterson, S.; Ryan, M.; Dunbar, J. Effects of soil microbes on functional traits of loblolly pine (Pinus taeda) seedling families from contrasting climates. Frontiers in Plant Science. 2020, 10, 1643. [Google Scholar] [CrossRef]
- Ugarte, R.M.; Martínez, M.H.; Díaz-Santiago, E.; Pugnaire, F.I. Microbial controls on seed germination. Soil Biology and Biochemistry. 2024, 199, 109576. [Google Scholar]
- Eldridge, D.J.; Travers, S.K.; Val, J.; Ding, J.; Wang, J.T.; Singh, B.K.; Delgado-Baquerizo, M. Experimental evidence of strong relationships between soil microbial communities and plant germination. Journal of Ecology. 2021, 109, 2488–2498. [Google Scholar] [CrossRef]
- Gupta, A.; Gupta, R.; Singh, R.L. Microbes and environment. In Principles and applications of environmental biotechnology for a sustainable future. 2016 (pp. 43–84). Singapore: Springer Singapore.
- Mahala, D.M.; Maheshwari, H.S.; Yadav, R.K.; Prabina, B.J.; Bharti, A.; Reddy, K.K.; Kumawat, C.; Ramesh, A. Microbial transformation of nutrients in soil: an overview. Rhizosphere microbes: Soil and plant functions. 2021; 175-211.
- Singh, B.K.; Hu, H.W.; Macdonald, C.A.; Xiong, C. Microbiome-facilitated plant nutrient acquisition. Cell Host & Microbe. 2025, 33, 869–881. [Google Scholar]
- Vincze, É.B.; Becze, A.; Laslo, É.; Mara, G. Beneficial soil microbiomes and their potential role in plant growth and soil fertility. Agriculture. 2024, 14, 152. [Google Scholar] [CrossRef]
- Lladó, S.; López-Mondéjar, R.; Baldrian, P. Drivers of microbial community structure in forest soils. Applied microbiology and biotechnology. 2018, 102, 4331–4338. [Google Scholar] [CrossRef] [PubMed]
- Al-Turki, A.; Murali, M.; Omar, A.F.; Rehan, M.; Sayyed, R.Z. Recent advances in PGPR-mediated resilience toward interactive effects of drought and salt stress in plants. Frontiers in microbiology. 2023, 14, 1214845. [Google Scholar] [CrossRef]
- Kholostiakov, V.; Burns, B.; Ridgway, H.; Padamsee, M. Variation in seed-borne microbial communities of Metrosideros excelsa Sol. ex Gaertn. with consequences for germination success. New Zealand Journal of Botany. 2025, 63, 1981–2004. [Google Scholar] [CrossRef]
- Liu, B.W.; Li, S.Y.; Zhu, H.; Liu, G.X. Phyllosphere eukaryotic microalgal communities in rainforests: Drivers and diversity. Plant Diversity. 2023, 45, 45–53. [Google Scholar] [CrossRef]
- Durán, P.; Flores-Uribe, J.; Wippel, K.; Zhang, P.; Guan, R.; Melkonian, B.; Melkonian, M.; Garrido-Oter, R. Shared features and reciprocal complementation of the Chlamydomonas and Arabidopsis microbiota. Nature Communications. 2022, 13, 406. [Google Scholar] [CrossRef]
- Alobwede, E.; Leake, J.R.; Pandhal, J. Circular economy fertilization: Testing micro and macro algal species as soil improvers and nutrient sources for crop production in greenhouse and field conditions. Geoderma. 2019, 334, 113–123. [Google Scholar] [CrossRef]
- Lee, S.M.; Ryu, C.M. Algae as new kids in the beneficial plant microbiome. Frontiers in Plant Science. 2021, 12, 599742. [Google Scholar] [CrossRef]
- Torres-Cortés, G.; Bonneau, S.; Bouchez, O.; Genthon, C.; Briand, M.; Jacques, M.A.; Barret, M. Functional microbial features driving community assembly during seed germination and emergence. Frontiers in plant science. 2018, 9, 902. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Xie, X.; Chen, M.; Qiao, F.; Liu, X.; Zhao, Y.; Peng, X.; Liu, F. Co-Inoculation Between Bacteria and Algae from Biological Soil Crusts and Their Effects on the Growth of Poa annua and Sandy Soils Quality. Microorganisms. 2025, 13, 1778. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, J.; Zhu, X.; Miao, T.; Hou, H.; Chen, G. The effects of microbial composite fertilizer CAMP on soil improvement and growth of Chinese cabbage. North Hortic. 2018, 12, 112–118. [Google Scholar]
- Treves, H.; Küken, A.; Arrivault, S.; Ishihara, H.; Hoppe, I.; Erban, A.; Höhne, M.; Moraes, T.A.; Kopka, J.; Szymanski, J.; Nikoloski, Z. Carbon flux through photosynthesis and central carbon metabolism show distinct patterns between algae, C3 and C4 plants. Nature plants. 2022, 8, 78–91. [Google Scholar] [CrossRef]
- Li, D.; Dong, H.; Cao, X.; Wang, W.; Li, C. Enhancing photosynthetic CO2 fixation by assembling metal-organic frameworks on Chlorella pyrenoidosa. Nature Communications. 2023, 14, 5337. [Google Scholar] [CrossRef]
- Mutum, L.; Janda, T.; Darkó, É.; Szalai, G.; Hamow, K.Á.; Molnár, Z. Outcome of microalgae biomass application on seed germination and hormonal activity in winter wheat leaves. Agronomy. 2023, 13, 1088. [Google Scholar] [CrossRef]
- Hibbing, M.E.; Fuqua, C.; Parsek, M.R.; Peterson, S.B. Bacterial competition: surviving and thriving in the microbial jungle. Nature reviews microbiology. 2010, 8, 15–25. [Google Scholar] [CrossRef]
- Ren, C.G.; Kong, C.C.; Li, S.M.; Wang, X.J.; Yu, X.; Wang, Y.C.; Qin, S.; Cui, H.L. Symbiotic microalgae and microbes: a new frontier in saline agriculture. Frontiers in Microbiology. 2025, 16, 1540274. [Google Scholar] [CrossRef] [PubMed]
- Cirri, E.; Pohnert, G. Algae− bacteria interactions that balance the planktonic microbiome. New Phytologist. 2019, 223, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Olofintila, O.E.; Noel, Z.A. Soybean and cotton spermosphere soil microbiome shows dominance of soilborne copiotrophs. Microbiology Spectrum. 2023, 11, e00377–23. [Google Scholar] [CrossRef]
- Gonzalez-Gonzalez, L.M.; De-Bashan, L.E. The potential of microalgae–bacteria consortia to restore degraded soils. Biology. 2023, 12, 693. [Google Scholar] [CrossRef] [PubMed]
- de Goede, S.P.; Hannula, S.E.; Jansen, B.; Morriën, E. Fungal-mediated soil aggregation as a mechanism for carbon stabilization. The ISME Journal. 2025, 19, wraf074. [Google Scholar] [CrossRef]
- Özden E, Ermiş S, Yildirim E, Demir İ. Perspective Chapter: Insights into Seed Germination–Physiological and Environmental Mechanisms. 2025.
- Fernández-Pascual, E.; Carta, A.; Mondoni, A.; Cavieres, L.A.; Rosbakh, S.; Venn, S.; Satyanti, A.; Guja, L.; Briceño, V.F.; Vandelook, F.; Mattana, E. The seed germination spectrum of alpine plants: a global meta-analysis. New phytologist. 2021, 229, 3573–3586. [Google Scholar] [CrossRef]
- Lamont, B.B.; Pausas, J.G. Seed dormancy revisited: Dormancy-release pathways and environmental interactions. Functional Ecology. 2023, 37, 1106–1125. [Google Scholar] [CrossRef]
- Carrera-Castaño, G.; Calleja-Cabrera, J.; Pernas, M.; Gómez, L.; Oñate-Sánchez, L. An updated overview on the regulation of seed germination. Plants. 2020, 9, 703. [Google Scholar] [CrossRef] [PubMed]
- Manohan, B.; Shannon, D.P.; Tiansawat, P.; Chairuangsri, S.; Jainuan, J.; Elliott, S. Use of functional traits to distinguish successional guilds of tree species for restoring forest ecosystems. Forests. 2023, 14, 1075. [Google Scholar] [CrossRef]
- Tiebel, K.; Huth, F.; Wagner, S. Soil seed banks of pioneer tree species in European temperate forests: a review. iForest-Biogeosciences and Forestry. 2018, 11, 48. [Google Scholar] [CrossRef]
- Carvalho, A.S.; De Andrade, L.G.; De Andrade, A.C. Germination of small tropical seeds has distinct light quality and temperature requirements, depending on microhabitat. Plant Biology. 2021, 23, 981–991. [Google Scholar] [CrossRef] [PubMed]
- Qiu, T.; Andrus, R.; Aravena, M.C.; Ascoli, D.; Bergeron, Y.; Berretti, R.; Berveiller, D.; Bogdziewicz, M.; Boivin, T.; Bonal, R.; Bragg, D.C. Limits to reproduction and seed size-number trade-offs that shape forest dominance and future recovery. Nature communications. 2022, 13, 2381. [Google Scholar] [CrossRef]
- Iralu, V.; Barbhuyan, H.S.; Upadhaya, K. Ecology of seed germination in threatened trees: a review. Energy, Ecology and Environment. 2019, 4, 189–210. [Google Scholar] [CrossRef]
- Ley-Lopez, J.M.; Wawrzyniak, M.K.; Chacon-Madrigal, E.; Chmielarz, P. Seed traits and tropical arboreal species conservation: a case study of a highly diverse tropical humid forest region in Southern Costa Rica. Biodiversity and Conservation. 2023, 32, 1573–1590. [Google Scholar] [CrossRef]
- Liu, X.; Xiao, Y.; Wang, Y.; Wang, R.; Huang, R.; Liang, H.; Jiang, Y.; Jiang, Y. Seed germination ecology of endangered plant Horsfieldia hainanensis Merr. In China. BMC Plant Biology. 2024, 24, 486. [Google Scholar] [CrossRef]
- Kim, S.; Ko, C.H.; Kwon, H.C.; Rhie, Y.H.; Lee, S.Y. Effect of Temperature and Covering Structures in Seed Dormancy and Germination Traits of Manchurian Striped Maple (Acer tegmentosum Maxim.) Native to Northeast Asia. Plants. 2025, 14, 767. [Google Scholar] [CrossRef]
- Linan, A.G.; Gereau, R.E.; Sucher, R.; Mashimba, F.H.; Bassuner, B.; Wyatt, A.; Edwards, C.E. Capturing and managing genetic diversity in ex situ collections of threatened tropical trees: A case study in Karomia gigas. Applications in Plant Sciences. 2024, 12, e11589. [Google Scholar] [CrossRef] [PubMed]
- Beckman, E.; Meyer, A.; Pivorunas, D.; Hoban, S.; Westwood, M. Conservation Gap Analysis of Native. 2021.
- Cuena Lombraña, A.; Dessì, L.; Podda, L.; Fois, M.; Luna, B.; Porceddu, M.; Bacchetta, G. The effect of heat shock on seed dormancy release and germination in two rare and Endangered Astragalus L. species (Fabaceae). Plants. 2024, 13, 484. [Google Scholar] [CrossRef]
- Mackenzie, B.D.; Clarke, S.W.; Zimmer, H.C.; Liew, E.C.; Phelan, M.T.; Offord, C.A.; Menke, L.K.; Crust, D.W.; Bragg, J.G.; McPherson, H. Ecology and conservation of a living fossil: Australias Wollemi Pine (Wollemia nobilis). 2021.
- Liu, X.; Xiao, Y.; Ling, Y.; Liao, N.; Wang, R.; Wang, Y.; Liang, H.; Li, J.; Chen, F. Effects of Seed Biological Characteristics and Environmental Factors on Seed Germination of the Critically Endangered Species Hopea chinensis (Merr.) Hand.-Mazz. in China. Forests. 2023, 14, 1975. [Google Scholar] [CrossRef]
- Klupczyńska, E.A.; Pawłowski, T.A. Regulation of seed dormancy and germination mechanisms in a changing environment. International journal of molecular sciences. 2021, 22, 1357. [Google Scholar] [CrossRef] [PubMed]
- Putri, K.P.; Yuniarti, N.; Aminah, A.; Suita, E.; Sudrajat, D.J.; Syamsuwida, D. Seed handling of specific forest tree species: Recalcitrant and intermediate seed. InIOP Conference Series: Earth and Environmental Science. 2020; (Vol. 522, No. 1, p. 012015). IOP Publishing.
- Smith, P.; Pence, V. The role of botanic gardens in ex situ conservation. Plant conservation science and practice: the role of botanic gardens. 2017; 102-33.
- Yang, P.; Lu, L.; Condrich, A.; Muni, G.A.; Scranton, S.; Xu, S.; Xia, Y.; Huang, S. Innovative Approaches for Engineering the Seed Microbiome to Enhance Crop Performance. Seeds. 2025, 4, 24. [Google Scholar] [CrossRef]
- Li, D.; Chen, W.; Luo, W.; Zhang, H.; Liu, Y.; Shu, D.; Wei, G. Seed microbiomes promote Astragalus mongholicus seed germination through pathogen suppression and cellulose degradation. Microbiome. 2025, 13, 23. [Google Scholar]
- Lorentz, J.F.; Calijuri, M.L.; Rad, C.; Cecon, P.R.; Assemany, P.P.; Martinez, J.M.; Kholssi, R. Microalgae biomass as a conditioner and regulator of soil quality and fertility. Environmental Monitoring and Assessment. 2024, 196, 198. [Google Scholar] [CrossRef]
- Abate, R.; Oon, Y.L.; Oon, Y.S.; Bi, Y.; Mi, W.; Song, G.; Gao, Y. Diverse interactions between bacteria and microalgae: A review for enhancing harmful algal bloom mitigation and biomass processing efficiency. Heliyon. 2024, 10. [Google Scholar] [CrossRef]
- Eldridge, D.J.; Travers, S.K.; Val, J.; Ding, J.; Wang, J.T.; Singh, B.K.; Delgado-Baquerizo, M. Experimental evidence of strong relationships between soil microbial communities and plant germination. Journal of Ecology. 2021, 109, 2488–2498. [Google Scholar] [CrossRef]
- Semple, A.; Pandhal, J. Engineering the phycosphere: fundamental concepts and tools for the bottom-up design of microalgal-bacterial consortia. Applied Phycology. 2025, 6, 21–51. [Google Scholar] [CrossRef]
- Koneru, H.; Bamba, S.; Bell, A.; Estrada-Graf, A.A.; Johnson, Z.I. Integrating microbial communities into algal biotechnology: a pathway to enhanced commercialization. Frontiers in Microbiology. 2025, 16, 1555579. [Google Scholar] [CrossRef]
- Zhang, D.; Xu, F.; Wang, F.; Le, L.; Pu, L. Synthetic biology and artificial intelligence in crop improvement. Plant Communications. 2025, 6. [Google Scholar] [CrossRef]
- Kozaeva, E.; Eida, A.A.; Gunady, E.F.; Dangl, J.L.; Conway, J.M.; Brophy, J.A. Roots of synthetic ecology: microbes that foster plant resilience in the changing climate. Current Opinion in Biotechnology. 2024, 88, 103172. [Google Scholar] [CrossRef]
- Fiodor, A.; Ajijah, N.; Dziewit, L.; Pranaw, K. Biopriming of seed with plant growth-promoting bacteria for improved germination and seedling growth. Frontiers in Microbiology. 2023, 14, 1142966. [Google Scholar] [CrossRef]
- Peddle, S.D.; Hodgson, R.J.; Borrett, R.J.; Brachmann, S.; Davies, T.C.; Erickson, T.E.; Liddicoat, C.; Muñoz-Rojas, M.; Robinson, J.M.; Watson, C.D.; Krauss, S.L.; Breed, M.F. Practical applications of soil microbiota to improve ecosystem restoration: current knowledge and future directions. Biol Rev Camb Philos Soc. 2025, 100, 1–18. [Google Scholar] [CrossRef]
- Williams, B.A.; Beyer, H.L.; Fagan, M.E.; Chazdon, R.; Schmoeller, M.; Sprenkle-Hyppolite, S.; Griscom, B.W.; Watson, J.E.M.; Tedesco, A.M.; Gonzalez-Roglich, M.; et al. Global potential for natural regeneration in deforested tropical regions. Nature. 2024, 636, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Chemla, Y.; Sweeney, C.J.; Wozniak, C.A.; Voigt, C.A. Design and regulation of engineered bacteria for environmental release. Nature Microbiology. 2025, 10, 281–300. [Google Scholar] [CrossRef] [PubMed]
- Keiper, F.; Atanassova, A. International synthetic biology policy developments and implications for global biodiversity goals. Frontiers in Synthetic Biology. 2025, 3, 1585337. [Google Scholar] [CrossRef]
Figure 1.
Functional mechanisms of microbes and algae on forest seed germination.

Figure 2.
Interactions showing nutrient exchanges and structural effects of algae microbiome consortia on the seed microenvironment.
Figure 2.
Interactions showing nutrient exchanges and structural effects of algae microbiome consortia on the seed microenvironment.
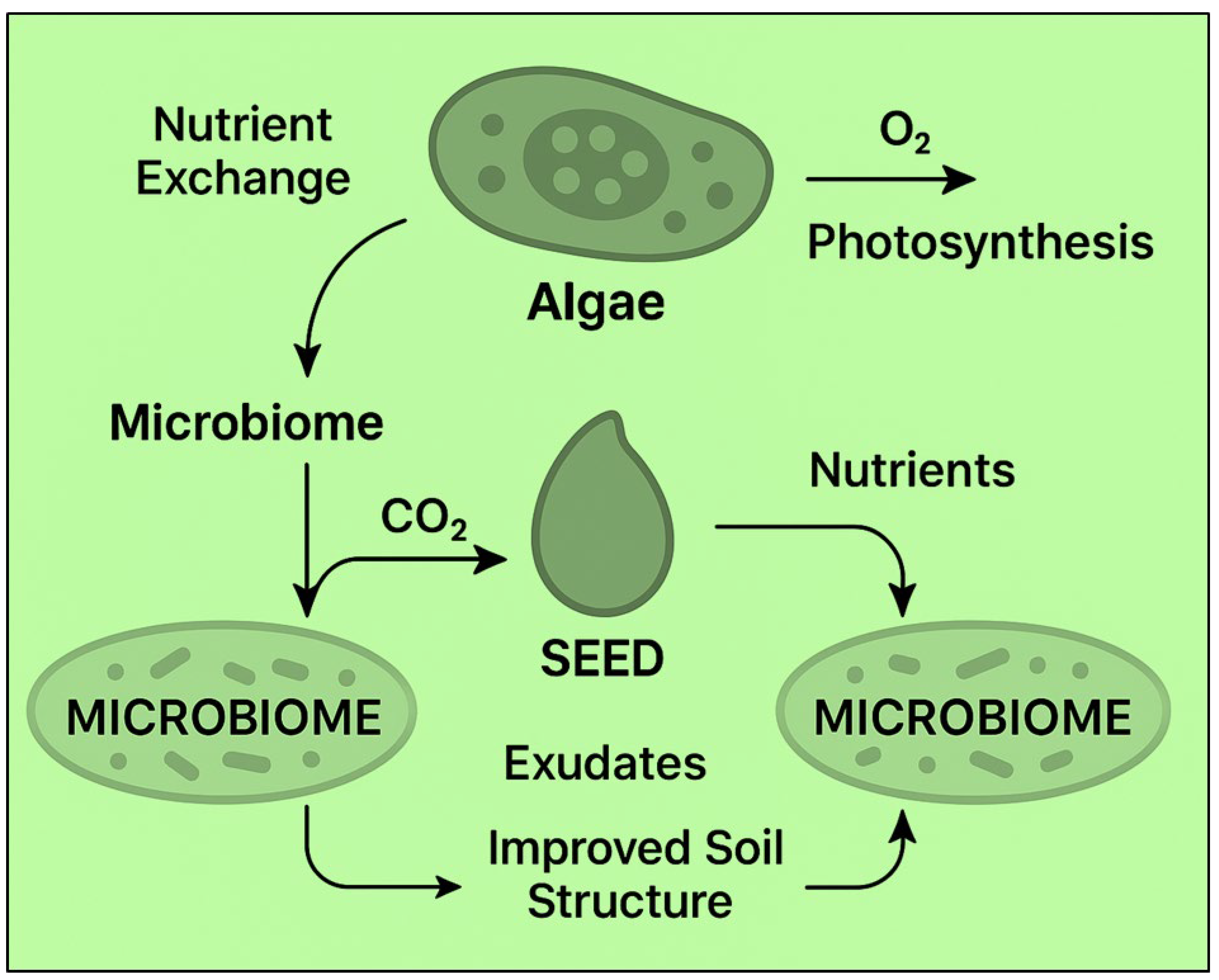

Table 2.
Type of microbiomes and algae relevant in forest soils and its impact on initial stages of plant growth.
Table 2.
Type of microbiomes and algae relevant in forest soils and its impact on initial stages of plant growth.
| Microbial/Algal Group | Example Species | Main Functions | Impact on Germination/Seedlings |
|---|---|---|---|
| Acidobacteriota Bacteria [13,16] |
Acidobacterium capsulatum Acidipila rosea Edaphobacter modestus Terriglobus roseus |
Degradation of polymers (cellulose, lignin, chitin), N–Fe–S cycles, tolerance to acidic soils | Improve soil fertility → indirect stimulation of germination |
| Proteobacteria Bacteria (PGPB) [17,18] |
Rhizobium Azospirillum brasilense Pseudomonas fluorescens Methylobacterium Bradyrhizobium Burkholderia |
Nitrogen fixation, phytohormone production (IAA, cytokinins, gibberellins), antibiosis, biofilms | Accelerated germination, enhanced early growth, biocontrol |
| Verrucomicrobia Bacteria [19,20,21] |
Opitutus terrae Candidatus Udaeobacter copiosus Luteolibacter pohnpeiensis |
Aerobic metabolism, organic matter transformation, production of organic acids & nutrients | Increased fertility, improved mineral nutrition of seeds |
| Actinobacteria Bacteria [22,23] |
Streptomyces Micromonospora Nocardia Frankia spp. |
Production of enzymes (cellulases, ligninases, phosphatases), natural antibiotics, nitrogen-fixing symbioses | Favor germination by improving nutrition and pathogen protection |
| Saprophytic Fungi [25,26] |
Aspergillus niger Mucor hiemalis Chaetomium globosum Trichoderma harzianum Penicillium chrysogenum Fusarium solani Rhizopus stolonifer Humicola grisea |
Litter degradation, release of C–N–P–K, extracellular enzymes | Prepare the soil, but some (Fusarium) inhibit germination |
| Arbuscular Mycorrhizal Fungi (AMF) [29,30] |
Glomus intraradices Glomus etunicatum Acaulospora scrobiculata Gigaspora margarita |
Root symbiosis (arbuscules), P and N uptake, stress tolerance | Natural biopriming → improves germination and seedling vigor |
| Ectomycorrhizal Fungi (ECM) [29,30] |
Laccaria bicolor Russula cyanoxantha Suillus luteus Boletus pinophilus Amanita phalloides Tricholoma matsutake |
Mycelial network (Hartig net), mineral nutrition, inter-tree communication | Enhance germination and survival of forest species |
| Archaea [31] |
Nitrososphaera viennensis Candidatus Nitrosotenuis uzonensis Methanobacterium formicicum Methanocella arvoryzae Methanosaeta concilii Nitrososphaera gargensis |
Biogeochemical cycles (N, C, S, CH4), survival in extreme soils | Stabilize the ecosystem → indirect support of germination |
| Protists [32] |
Amoebozoa Apicomplexa Cercozoa Ciliophora Viruses Bacteriophages Phytoviruses Mycoviruses |
Regulation of microbial populations, symbiotic/parasitic interactions | Influence germination by balancing the microbiome |
| Cyanobacterial Algae (prokaryotes) [33,34,35] |
Nostoc commune Scytonema sp. Microcoleus vaginatus Tolypothrix spp. Anabaena spp. Calothrix spp. |
Photosynthesis, N2 fixation, polysaccharide production, lichenic symbioses | Improve nitrogen nutrition, soil humidity and stability → stimulated germination |
| Green Algae (Chlorophyta) [33] |
Chlorella vulgaris Klebsormidium flaccidum Scenedesmus obliquus |
Photosynthesis (chlorophyll A/B), biomass, exopolysaccharides | Biostimulants: accelerate germination and early growth |
| Yellow/Golden Algae (Xanthophyta, Chrysophyta, Diatoms) [33,36] |
Botrydium granulatum Ochromonas spp. Navicula spp. Eunotia spp. |
Carotenoid/fucoxanthin pigments, photosynthesis, exopolymers | Improve rhizospheric microhabitats, organic C input |
| Dinophyta & Cryptophyta Algae [33] |
Ceratium hirundinella Symbiodinium spp. Cryptomonas spp. |
Photosynthesis, antioxidant molecules, symbioses | Increase oxidative stress tolerance, germination under difficult conditions |
Table 2.
Examples of microbiomes associated with forest species and their functional relationships influencing germination.
Table 2.
Examples of microbiomes associated with forest species and their functional relationships influencing germination.
| Forest Species | Beneficial microbiomes | Mechanisms promoting growth | References |
|---|---|---|---|
| Astragalus membranaceus var. mongholicus |
Paenibacillus spp. Pseudomonas spp. |
Pathogen suppression by enrichment of antagonistic bacteria and reduction of pathogens (such as Fusarium)/Decomposition of seed coats would facilitate radicle emergence and be associated with better germination. | [41] |
| Abies pindrow | Bacterial (30 species)/consortium; fungal (18 species) (729 isolates) | indole-3-acetic acid (IAA) production-phytohormone, ammonia prodection, phosphate solubilization, Fusarium biocontrol | [42] |
| Pinus taeda | Arid community rich in Actinobacteria (Rubrobacterales) and Pleosporales fungi (dark septate fungi). | Stimulate germination via hormones and water retention, increase in root biomass (wet family), facilitation of nitrogen acquisition, but variable effects on photosynthesis and nutrition. | [43] |
|
Betula pendula Carpinus betulus Fagus sylvatica Fraxinus excelsior Larix decidua Picea abies Pinus sylvestris Quercus robur |
Russula spp. Laccaria spp. Tuber spp. Cenococcum geophilum Glomus spp. Claroideoglomus spp. Paraglomus spp. Pseudomonas spp. Bacillus spp. Burkholderia spp. Rhizobium spp. Actinobacteria divers |
Mycorrhization, nutrition (N, P), protection vs. pathogens (Pythium spp., Fusarium spp.), or on the contrary competition and infections. | [44] |
|
Eucalyptus largiflorens Eucalyptus camaldulensis Callitris glaucophylla |
Actinobacteria Proteobacteria Ascomycota Arbuscular mycorrhizae Aectomycorrhizal fungi |
Strong positive correlation between Arbuscular Mycorrhizal Fungi (AMF) and nitrogen-fixing plants/Secretion of phytohormones (auxins and cytokinins) stimulating germination/Promoting dormancy breaking by breaking the seed coat/Decomposition of organic matter to release nutrients available to the seed/Pathogen suppression | [45] |
Table 3.
The main differences in seed traits and germination behavior between pioneer and climax forest species retrieved and adopted from [74,75].
| Trait | Pioneer Forest Species Seeds | Climax Forest Species Seeds |
|---|---|---|
| Typical seed size & number | Small seeds, often produced in large quantities such as Cecropia or produces tiny wind seeds in the millions. | Larger seeds with higher individual mass, produced in fewer numbers. |
| Nutrient reserves | Minimal reserves per seed, seedlings depend quickly on external resources. Low reserve content limits survival in shade. | Substantial nutrient reserves in the seed endosperm/cotyledons; can sustain seedling growth for extended periods. Large reserves support emergence in deep shade or through thick leaf litter. |
| Dormancy and seed bank | Often physiologically or physically dormant at shedding; form persistent soil seed banks. Dormancy is broken by disturbance cues signalling an open habitat. Many pioneers require light to germinate and can remain viable in soil for years until a gap occurs. | Frequently non-dormant or only briefly dormant; many are recalcitrant. Those with dormancy often require specific cues like prolonged cold or chemical scarification by gut passage. Generally, climax species seeds germinate in the next suitable season or perish, rather than forming long soil banks. |
| Germination conditions | Require high-light, fluctuating temperature, or other gap conditions for germination. Germinate rapidly once conditions are met, exploiting ephemeral resource availability. Seedlings are shade-intolerant, needing direct sunlight soon after germination. For instance, pioneer tree seeds often germinate only in canopy gaps or open clearings. | Can germinate under low light or stable understory conditions. Some exhibit slow or extended germination, and may germinate opportunistically over time. Seedlings are often shade-tolerant, capable of establishing under an intact canopy using seed reserves. Large-seeded climax species may germinate in deep shade and wait for later light availability. |
| Seed/seedling strategy | Emphasize quick establishment and fast early growth. Seedlings typically allocate more to shoot elongation to outgrow competitors. Survival is low if suitable gaps are absent shortly after germination. | Emphasize survival in competitive, resource-limited settings. Seedlings often exhibit slow growth but high survivorship, allocating to root development and defense. They can endure shaded conditions until a light gap appears years later, thanks to the energy reserves from large seeds. |
| Examples | Pioneer: Betula pendula, tiny, wind-dispersed seeds; Cecropia peltata light demanding tropical pioneer with dust-like seeds; Pinus sylvestris in early post-fire succession. | Climax: Fagus sylvatica, large nut-like seeds that germinate under forest canopy; Shorea spp. winged large seeds with no dormancy, germinating soon after dropping in the rainy season; Pouteria spp. recalcitrant seeds that lose viability if dried. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



