
Submitted:
20 July 2024
Posted:
22 July 2024
You are already at the latest version
Abstract
Seedling establishment is often a critical bottleneck to revegetation of mine tailings and similar substrates with a low capacity to retain water and plant nutrients. Biochar and deactivated yeast are potential sustainable, low-cost materials with high nutrient- and water-holding capacity that could be used in this context as seed coatings to aid in seedling establishment on challenging sub-strates. We conducted a greenhouse study to assess the effectiveness of biochar and deactivated yeast, applied alone in a factorial combination, as seed coatings, on the germination, establish-ment, and early growth of white clover (Trifolium repens) and purple prairie clover (Dalea pur-purea). Coated seeds were applied to a mine tailing, a coarse granitic sand, and potting soil mix substates; seedling establishment and growth were monitored over 75 days. Results indicate strong interactive effects of seed coatings with species and substrate. Biochar coatings enhanced seedling establishment of Trifolium, with biochar or biochar plus yeast coatings giving the best results. In some cases, these effects persisted throughout the experiment: biochar coatings resulted in a ~5-fold increase in Trifolium biomass at harvest for plants in the potting soil mix but had neutral effects on sand or tailings. Biochar seed coatings also enhanced Dalea germination in some cases, but benefits did not persist. Our results indicate that biochar-based seed coatings can have lasting effects on plant growth well beyond germination, but also emphasize highly spe-cies-specific responses that highlight the need for further study of broader patterns and mecha-nisms.
Keywords:
biochar
; charcoal
; clover
; ecological restoration
; inactive brewer’s yeast
; mine tailings
; revegetation
; seed coatings
; seed germination
; seedling establishment
1. Introduction
Mine tailings and other mine waste materials impact an estimated 50 million km2 of the earth’s surface [1,2], presenting an enormous challenge for sustainable land management. Restoration projects for mine tailing sites are challenged by poor soil quality and elevated concentrations of potentially toxic elements (PTEs). Tailing substrates are characterized by low organic matter, limited bioavailable nutrients, lack of soil macrostructure, and low water-holding capacity [3]. In addition, the weathering of sulfide minerals often causes acidic mine drainage and wider contamination with PTEs, which can have long-lasting impacts on local ecosystems. These factors often drastically limit the establishment and survival of plant species on tailings [4].
“Biochar” refers to pyrolyzed biomass material prepared for and used as a soil amendment [5]. Biochar has been found to increase plant growth and overall biomass production, particularly on coarse-textured, acidic substrates [6,7], and in the context of ecological restoration on mine tailings and degraded soils [8,9,10]. Biochar enhances substrate nutrient status and water holding capacity [11] and can reduce bioavailability of PTEs [8].
Biochar directly provides significant amounts of bioavailable P, K, Ca, and Mg, but most N present is either lost due to volatilization, or covalently bound in the biochar structure during pyrolysis [12,13]. Biochar has also been found to immobilize soil N [14], which can cause N deficiencies that negatively impact plant growth [15]. The tendency of biochar to induce N deficiency has motivated applied research on use of N-rich co-amendments with biochar, such as urea [16], composts [17], and manures [18]. Spent deactivated yeast is a major waste product of the brewing industry with a notably high N content and very low C:N ratio [19]. Recycling of deactivated brewer’s yeast is difficult due to the high volumes produced and short storage lifespan [20]. Its high N content and low cost make deactivated yeast of interest as a co-amendment with biochar, and one recent study has shown exceptional plant growth responses to mixtures of biochar and deactivated yeast [21].
The cost of biochar is a disincentive for its use in large-scale restoration projects. Concentrating the application of biochar and other amendments at the establishment site for seeds or seedlings could potentially greatly reduce costs. In particular, seed coatings are potentially a low-cost pathway to help seeds germinate and establish within harsh tailing environments. Biochar has recently been explored as a seed coating in ecological restoration [22,23], as well as in agricultural systems [24,25]. In the context of mine tailings restoration, biochar-based seed coatings might specifically be beneficial by improving soil water retention, increasing soil pH, adsorbing heavy metals, providing key nutrients, and enhancing soil microbial communities [26]. Seed coatings in general can be used to alter seed properties including size, weight, surface roughness or to add active ingredients to promote seedling establishment and early growth; seed coatings typically consist of binding agents, fillers and active ingredients [27]. In agriculture, seed coating treatments can including abscisic acid, salicylic acid, soil surfactants, and pest repellants have shown beneficial impacts on seedling germination, establishment, and growth [28]. Selection of appropriate binding agents is also important; for example, a recent study indicates that polyvinyl acetate, which has been commonly used in seed coating applications, has phytotoxic effects and can greatly reduce germination rates [29].
There has been less attention to deactivated yeast as a possible component in seed coatings, though live yeasts have been used in this context as antagonists to common pathogenic fungi [30]. Several recent studies have described growth benefits for plants after yeast biofertilizer supplementation [19,21,31,32]. Lonhienne et al. [32] used both inactive and active yeast as a fertilizer for tomato seedlings. Both yeast fertilizers increased root N and P, while inactive yeast increased overall root biomass, which was theorized to be a result of more rapid breakdown and dispersal than active yeast within the soil, allowing more rapid plant nutrient uptake. This presents a potential advantage of inactive yeast over live microorganisms. Inactive brewer’s yeast (Saccharomyces cerevisiae) is primarily made up of proteins (45-60%) and nucleic acids (6-15%) with a total N of ~9% producing very low C:N ratios [20]. Deactivated yeast also has shown PTE sorption in a number of studies, with sorption capacities of 1-20, 10-100, and 10-300 mg/g, for Cu, Cd, and Pb, respectively [33]. Uptake mechanisms for PTEs by yeast may be a combination of redox reactions, complexation, electrostatic attraction and ion exchange similar to biochar [34]. Yeast additions can also improve microbial communities [35]. The combination of biochar and deactivated yeast has not, to our knowledge, previously been examined in the context of seed coatings.
In the present study we assess the potential of biochar and yeast seed coatings, examining responses on three substrates including a mine tailing, a granitic sand, and a standard potting mix. Seed coating treatments include a factorial combination of biochar, deactivated yeast, and both ingredients, with starch used as a binding agent. We hypothesized that: (1) biochar and yeast coatings will increase the establishment and performance of seedlings with the greatest effects on the harsh tailings soil; (2) the combination of biochar plus yeast will have maximally beneficial effects; (3) although positive effects on seed establishment are expected, longer-term effects on growth will be limited due to the small volume of soil amendments associated with the seed coatings.
2. Materials and Methods
2.1. Species Selection
Two clover species, Trifolium repens L. (white clover, hereinafter Trifolium), a non-native species, and Dalea purpurea Vent. (purple prairie clover, hereinafter Dalea), a native species to eastern and central North America, were chosen for the study due to their rapid germination rates and identity as legumes. Clovers are desirable for phytoremediation because of their capacity to fix atmospheric N and rapid growth and survival in harsh environments. N-fixing legumes also have commonly been found to show high growth responses to biochar, likely due to fact that N fixation can offset biochar-related N deficits [36]. White clover showed increased in shoot and total biomass in cadmium-contaminated soil amended with biochar [37]. Increases in abundance of Trifolium species have also been noted in several studies examining biochar responses of plant communities in a restoration context [38,39,40,41]. Studies of soil remediation using clovers native to North America are limited; however, tallgrass prairie species including Dalea transplanted onto coal mine tailings grew with “exceptional vigor” [42].
2.2. Growth Media
Northern Ontario hosts one of the world’s largest gold camps in orogenic deposits of the Abitibi greenstone belt. The Delnite mine tailings used in this study have particularly elevated concentrations of As, and Cr, among other metals including Co and Cu [41]. High levels of As are commonly associated with gold in these deposits. The PTE typically found in orogenic gold deposits reflect a metamorphosed carbonaceous shale source. Organometallic complexes in these shales can host high levels of Mo, As, Zi, and Ni, while pyrite in the same sediments hosts gold and sulfur [43]. Orogenic gold deposits including the historic Delnite mine typically contain 3-5% sulfide minerals the majority of which are pyrite (FeS2), pyrrhotite (Fe7S8) and arsenopyrite (FeAsS) [44]. Historical mine tailings were sourced from the property located along the Porcupine-Destor deformation zone, which hosts many exploited Archean gold deposits. The pH was measured as 8.80 ± 0.06 and electrical conductivity (EC) 91.90 ± 8.72 μS cm-1 with bulk density of 1.08 g/cm3. Gold mine tailings including Delnite are generally characterized by low aggregation and cohesion as well as fine texture which makes sites highly susceptible to erosion by wind or water [35]. Further details on the tailings used are given by [41].
PRO-MIX® HP® high porosity mycorrhizae growing medium (Pro-mix, Rivière-du-Loup, Quebec, Canada) was used as a control substrate. The mixture is composed of sphagnum peat moss, perlite, limestone, a wetting agent and mycorrhizae (Rhizophagus irregularis). The pH measured 6.67 ± 0.56 with EC of 286 ± 225 μS cm-1. A granitic sand sourced from near Haliburton, ON, was used as a textural analog for the Delnite tailings. Its pH was measured as 6.16 ± 0.31 with EC of 79 ± 10 μS cm-1.
2.3. Biochar and Deactiated Yeast Properites
Biochar and yeast used were the same as in a prior study [21]. Biochar was produced at 625 °C from mixed conifer feedstock using slow pyrolysis with a residence time of twenty minutes. It has very fine grain size <0.042 mm and is hydrophobic, with a pH of 7.88 ± 0.03 and electrical conductivity (EC) of 86.07 ± 1.63 μS cm-1, and bulk density of 0.158 g cm-3 [21]. The species S. cerevisiae produced the inactive brewer’s yeast used. The deactivated yeast had pH 6.20 ± 0.01, EC 1027 ± 88 μS cm-1, and bulk density 1.08 g cm-3 [21]. Additional information on these materials is given by Sifton et al. [21].
2.5. Experimental Design and Implementation
The overall experimental design consisted of a 2 x 2 x 3 factorial combination of biochar seed coating, yeast seed coating, and substrate type, with 6 replicates per treatment, implemented for each of the two species (2 x 2 x 3 x 6 x 2 = 144 pots in total). Three coated seeds of a designated species and treatment were added to every pot. Seed coatings were produced using an adhesive mixture of 4% tapioca starch solution which was heated and stirred for 5-10 minutes. One mL of adhesive was used for every nine seeds. The mass of treatment applications was determined by average seed mass, which was 0.0060 g and 0.0019 g for Trifolium and Dalea, respectively. Yeast treatment per seed was 0.5x the seed mass and biochar treatment was 2x the seed mass; the yeast plus biochar treatment was additive (i.e., 0.5x seed mass of yeast plus 2x seed mass of biochar).
Dalea seeds were scarified and stored at 2-3 °C for 7 days to improve germination rates. Trifolium seeds did not require scarification and were stored at room temperature prior to coating. Seed coatings and seeds were applied to 1.5 x 1.5 cm squares of filter paper (Whatman #2), marked with their species and treatment. The coated seeds were left to air dry for two hours before sowing. Square plastic plant pots measuring 10 x 10 cm were lined with polysynthetic fabric mesh used to retain substrate media. Each pot received 240 mL of the designated substrate. Pots were arranged in a randomized block design with six replicates to reduce spatial effects.
Seeds were sown on Nov. 21, 2023 and plants harvested Feb. 3, 2024, giving a total experimental runtime of 75 days. The average temperature within the greenhouse was 19.8°C with extremes of 38.8°C and 3.3°C. Humidity varied between 2.7% and 61.8% with an average of 25.8%. Pots were watered to field capacity every two days depending on conditions within the greenhouse. Supplemental light was provided by sodium vapor lamps for 12 hours per day. Germination was recorded every four days during the first 3 weeks to track germination rates and seedling survivorship was recorded once weekly after the germination phase. A PAR sensor (LI-190R, Li-Cor, Lincoln, NE, USA) was used to record measurements of ambient light levels which were taken every three weeks at noon. Ambient light averaged 569 µmol m-2 s-1.
At harvest soil samples were added to test tubes with a 1:5 ratio of soil to deionized water then placed in an orbital shaker at 85 rpm for 24 hours. For all pots with live seedlings, the number of survivors and leaf numbers were recorded. Above and below ground biomass were separated for each pot, oven dried for 48 hours at 60°C, and weighed to the nearest 0.01 mg. Leaf area was recorded for each pot using an optical area meter scanner (LI-3100, Li-Cor, Lincoln, NE, USA). Leaf area ratio was calculated as leaf area per total plant biomass.
2.6. Statistical Analysis
Data were first analyzed using (generalized) linear mixed model analyses with biochar, yeast, and substrate as fixed factors, and block as a random factor. Block was not significant in any case, and so 3-way analysis of variance (ANOVA) was used. All analyses were performed separately by species. Counts of live seedlings after germination and at harvest were assessed using proportional odds ordered logistic function (“polr” function in R) to compare treatment effects. Values of leaf area, leaf number, soil pH, soil EC, aboveground biomass, belowground biomass and total biomass measured at harvest were assessed using the linear model (“lm”) function. Leaf area was transformed using a log function to normalize the data. Significant results were defined by p-values under 0.05. Post-hoc contrasts were preformed using the Tukey HSD procedure; in the case of seedling density data, the non-parametric Dunn post-hoc test was used (with a p-value <0.1 considered significant due to the lower test power). Analyses were performed in the R programming environment (R version 4.2.3) [45], using the mass and car packages.
3. Results
3.1. Germination and Early Seedling Establishment
The vast majority of germinations observed occurred during the first two weeks of the experiment (Figure 1). In general, Trifolium germinated faster than Dalea. After one week, biochar had a significant (p<0.05) positive effect on the germination and subsequent establishment of Trifolium (Figure 1A; Table 1). There was also a significant interactive effect of the substrate x biochar interaction with the strongest positive effect observed on the potting soil control. In addition, the biochar plus yeast treatment gave the best early germination response in the case of Trifolium on the sand substrate (Figure 1A). For Dalea at one week, there was a significant substrate x biochar (Table 1), corresponding to a stronger positive effect of biochar on the sand and tailings substrates (Figure 1D). There was also a significant biochar x yeast interaction term in Dalea consistent with non-additive effects, with the biochar plus yeast treatment showing reduced performance relative to either treatment alone (Figure 1D).
The effects of treatments on germination diminished over time (Figure 1B,C,E,F). After two weeks, there was only a marginally significant (p<0.1) substrate x yeast interaction for Dalea and biochar x yeast interaction for Trifolium at day 75 (Table 1). Although not statistically significant, the early positive effects of biochar and biochar plus yeast apparent at 7 days persisted through the experiment in the case of Trifolium (Figure 1B,C).
3.2. Seedling Biomass
Plant biomass at harvest was much greater on the potting soil than either sand or tailings substrates for both species (Figure 2; Table 2). The biochar, yeast and combination coatings had a strong positive effect on aboveground, belowground and total biomass for the Trifolium in potting soil (Figure 2A-C; Table 2); however, effects on the other substrates were not significant. This pattern corresponds to significant biochar x substrate interactions in each case (Table 2). In contrast, Dalea only showed a significant effect of substrate on biomass (Figure 2D-F; Table 2). In addition, there were significant 2- and 3-way interactive effects of the yeast seed coating treatments for Trifolium, mainly attributable to positive effects of the yeast and yeast plus biochar coatings on the control (potting soil) substrate, but not on sand or tailings substrates (Figure 2A-C). Yeast-alone coatings showed the highest biomass for Dalea on tailings (Figure 2D), but this apparent effect was not statistically significant. Although both species are nodulating N-fixers, no recognizable root nodules were observed during root processing.
3.3. Seedling Leaf Development and Morphometrics
Leaf area at harvest showed a strong biochar main effect in Trifolium with interaction terms similar to those for biomass, corresponding to a strong positive effect on the control substrate only (Figure 3A; Table 2). In the case of Dalea there was a marginally significant negative effect of biochar on leaf area (Figure 3D; Table 2). The two species also showed markedly different responses of morphometrics. Trifolium showed a large reduction in leaf area ratio on sand and tailings substrates compared to the potting soil control, while Dalea showed the opposite pattern (Figure 3 B,E; Table 2). In addition, Dalea showed a strong reduction in root mass fraction on tailings compared to the other substrates (Figure 3F; Table 2). Seed coating effects on leaf area ratio and root mass fraction were not significant (Figure 3; Table 2).
Leaf area per container at the time of harvest showed a significant effect for the biochar treatment for Trifolium in the potting soil (p<0.001) (Figure 2; Table 2). Of note is the large difference in leaf area between the potting soil substrate and other substrates for Trifolium and the large difference in leaf area for Trifolium vs. Dalea in the potting soil. The highest leaf area was observed for Trifolium grown in potting soil with the biochar only treatment.
4. Discussion
Overall, the seed coatings tested had positive effects on seedling establishment and growth performance. Both species generally responded best to the biochar-only treatment in terms of the early germination and seedling establishment. There were also significant interactive effects such that in some treatments the biochar plus yeast combination produced favorable responses; however, contrary to our initial hypothesis, this was not generally the case. In addition, we found that seed coatings could have long-lasting effects on plant performance; in particular, the biochar-only seed coating greatly increased final biomass production in Trifolium on the potting soil substrate. In contrast, Dalea showed no such long-term seed coating benefit.
Species-specific responses to biochar treatments have been recorded in a number of studies, though the mechanisms involved are generally not yet well understood. A glasshouse study by Gale et al. [15] showed highly species-specific responses to the same biochar among 13 herbaceous plant species, with both negative and positive responses observed; Trifolium repens was among the majority species showing a strong positive response. Similarly, in a field study examining biochar effects on tree seed germination and radicle extension growth, there was a strong positive effect on seedling performance overall, but great variation among species with both positive and negative responses observed [46]. Biochar can immobilize soil N [14], which may negatively affect species with high nitrogen demands [15]. The poorer response of Dalea could possibly be explained by this effect, implying higher seedling N demands than Trifolium.
Results for yeast in the seed coatings are somewhat ambiguous. The ANOVAs for Trifolium showed significant interaction 2- and 3-way interaction terms for the yeast addition treatments (Table 2), but the main pattern accounting for this was that biochar-alone coatings provided stronger performance than coatings with yeast on the potting soil substrate (Figure 2A-C). However, for Trifolium on the sand substrate early seedling establishment was superior for the biochar plus yeast treatment (Figure 1A). Dalea quantitatively showed the highest biomass and leaf area values for the yeast-alone coating treatment on tailings (Figures 2,3D-F); however, this pattern was not statistically significant and Dalea plants showed low growth in all treatments. Deactivated yeast thus plausibly has benefits as a seed coating, either alone or in combination with biochar, particularly on adverse substrates such as sand or mine tailings; however, further studies are required on this point.
The benefits to plant performance provided by seed coatings were very short-lived on the sand and tailings substrates. This likely reflects the very low nutrient status and low water and nutrient holding capacity of these substrates, plus metals toxicity of the tailings substrate. The marked reduction in root allocation in Dalea grown on tailings likely reflects the toxicity of the substrate and the roots’ role as metal uptake sites. Metals including cadmium, copper and zinc have been found to cause reductions of root length up to 60% attributed to various disruptions of cellular function [47]. Additional soil amendments at the time of sowing of coated seeds may be necessary to improve root biomass and phytostabilization.
Trifolium seeds responded more positively to biochar coatings than Dalea, but only in the potting soil. The minimal effect of biochar on the sand and tailings substrates contradicts findings that biochar has stronger impacts on low-fertility soils [48]. Several factors may have contributed to this. The quantities of biochar added were almost certainly insufficient to increase bulk soil pH. The liming effect is one of the main factors thought to improve soil characteristics by decreasing heavy metal bioavailability and buffering high acidities [49]. The tailings used in the present study were alkaline, such that any liming effect of biochar is unlikely to further limit metal bioavailability. The elemental composition of the tailings used is also important when considering the neutral to negative response of seeds with biochar coatings. The majority of PTEs are cationic with one notable exception: anionic arsenic, which is increasingly bioavailable at higher pH, presenting a potential limitation for biochar in remediation [49]. Arsenic is commonly found at tailings sites in Northeast Ontario due to its spatial association with orogenic gold deposits which are the most productive in the region, including the Delnite tailings used in the present study (with As content of 320 ± 9 ppm [41]).
The results of the biochar seed coatings on the potting soil, although not the initial focus of this study, were the most surprising. Despite the very small volumes of biochar used, Trifolium seedlings with biochar-only coatings showed a ~5-fold increase in biomass at harvest, ~11 weeks after sowing. The mechanism for this effect is uncertain; however, one possibility is that the biochar treatment provided key micronutrients during the earliest stages of growth. Biochars commonly contain appreciable quantities of important plant micronutrients including B, Fe, Mn, Mo, and Zn [50]. A number of studies have recorded increased plant micronutrient uptake in response to biochar additions [21,41,51], though reductions are also possible due to liming effects. Long-persistent effects of biochar-based seed coatings might also be mediated by reductions in seedling stress or microbial interactions.
The limited persistence of positive effects from either biochar or deactivated yeast coatings on the sand and tailings substrates suggests that although the coatings can improve germination, further soil amendments are needed to promote long-term establishment of species. High-carbon wood ash biochar from bioenergy facilities is a likely cost-effective option [40,41], but a range of possibilities exist, including reclaimed topsoil or peat, wood mulches, and lignin residues [52,53,54].
5. Conclusions
Increasing vegetation cover and improving plant diversity on tailings sites can help restore natural soil profiles, increase soil stability and improve the overall hydrologic cycle [55]. Restoration of mine tailings, including sand-capped tailings structures, is challenged by low nutrient availability, low water holding capacity and extreme pH values which inhibit germination and growth of plant species. The treatments used in the present study may improve success of direct seeding, which has been challenged by low seedling establishment rates typically under 10% [23,28]. Successful direct seeding would greatly reduce restoration costs especially when compared to other common restoration methods including planting of seedlings [56]. Our results indicate that biochar and deactivated yeast can promote seedling establishment and growth; however, responses are strongly species- and substrate-specific. A focus on tailoring seed coatings to target substrates, including more acidic tailings or tailings with varying PTE composition, is a priority for future work.
Author Contributions
Conceptualization, J.C. and S.C.T.; methodology, E.T. and S.C.T.; formal analysis and visualization, J.C., E.T. and S.C.T.; investigation, J.C. and E.T.; resources, S.C.T.; data curation, J.C.; writing—original draft preparation, J.C.; writing—review and editing, E.T. and S.C.T.; supervision, S.C.T.; project administration, S.C.T.; funding acquisition, S.C.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by grants from the Canadian Natural Sciences and Engineering Research Council, with partner contributions from Seed the North, Inc., and BC Biocarbon, Inc.
Data Availability Statement
Data for this article can be found in the University of Toronto Dataverse research data repository: (link to be provided on acceptance).
Acknowledgments
We thank Melanie Sifton and Tony Ung for assistance with materials and greenhouse management. We thank Fajar Handayani and Samuel Sherman for assistance with the experimental setup.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Werner, T.T.; Mudd, G.M.; Schipper, A.M.; Huijbregts, M.A.; Taneja, L.; Northey, S.A. Global-scale remote sensing of mine areas and analysis of factors explaining their extent. Glob. Environ. Change 2020, 60, 1–10. [Google Scholar] [CrossRef]
- Liang, T.; Werner, T.T.; Heping, X.; Jingsong, Y.; Zeming, S. A global-scale spatial assessment and geodatabase of mine areas. Global Planet. Change 2021, 204, 1–11. [Google Scholar] [CrossRef]
- Mendez, M.O.; Maier, R.M. Phytostabilization of Mine Tailings in Arid and Semiarid Environments—An Emerging Remediation Technology. Environ. Health Perspect. 2007, 116, 278–383. [Google Scholar] [CrossRef]
- Karaca, O.; Cameselle, C.; Reddy, K.R. Mine tailing disposal sites: Contamination problems, remedial options and phytocaps for sustainable remediation. Rev Environ Sci Biotechnol 2018, 17, 205–228. [Google Scholar] [CrossRef]
- Lehmann, J.; Joseph, S. Biochar for Environmental Management: An Introduction. In Biochar for Environmental Management, 2nd ed.; Lehmann, J., Joseph, S., Eds.; Routledge: Abingdon, England, 2015; pp. 1–14. [Google Scholar]
- Biederman, L.A.; Harpole, W.S. Biochar and its effects on plant productivity and nutrient cycling: A meta-analysis. GCB Bioenergy 2013, 5, 202–214. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, A.; Ji, C.; Joseph, S.; Bian, R.; Li, L.; Pan, G.; Paz-Ferreiro, J. Biochar’s effect on crop productivity and the dependence on experimental conditions—A meta-analysis of literature data. Plant Soil 2013, 373, 583–594. [Google Scholar] [CrossRef]
- Beesley, L.; Moreno-Jimenez, E.; Gomez-Eyles, J.L.; Harris, E.; Robinson, B.; Sizmur, T. A review of biochars' potential role in the remediation, revegetation and restoration of contaminated soils. Environ Pollut. 2011, 159, 3269–3282. [Google Scholar] [CrossRef]
- Thomas, S.C.; Gale, N. Biochar and forest restoration: A review and meta-analysis of tree growth responses. New Forest 2015, 46, 931–946. [Google Scholar] [CrossRef]
- Rodriguez-Franco, C.; Page-Dumroese, D.S. Woody biochar potential for abandoned min land restoration in the US: A review. Biochar 2021, 3, 7–22. [Google Scholar] [CrossRef]
- Fellet, G.; Marchiol, L.; Delle Vedove, G.; Peressotti, A. Application of biochar on mine tailings: Effects and perspectives for land reclamation. Chemosphere 2011, 83, 1262–1267. [Google Scholar] [CrossRef]
- Ippolito, J.A.; Cui, L.; Kammann, C.; Wrage-Monnig, N.; Estavillo, J.M.; Fuertes-Mendizabal, T.; Cayuela, M.L.; Sigua, G.; Novak, J.; Spokas, K.; Borchard, N. Feedstock choice, pyrolysis temperature and type influence biochar characteristics: A comprehensive meta-data analysis review. Biochar 2020, 2, 421–438. [Google Scholar] [CrossRef]
- Tomczyk, A.; Sokolowska, Z.; Boguta, P. Biochar physicochemical properties: Pyrolysis temperature and feedstock kind effects. Rev Environ Sci Biotechnol 2020, 19, 191–215. [Google Scholar] [CrossRef]
- Clough, T.J.; Condron, L.M.; Kamman, C.; Muller, C. A Review of Biochar and Soil Nitrogen Dynamics. Agronomy 2013, 3, 275–293. [Google Scholar] [CrossRef]
- Gale, N.V.; Halim, M.A.; Horsburgh, M.; Thomas, S.C. Comparative responses of early-successional plants to charcoal soil amendments. Ecosphere 2017, 8, 1–18. [Google Scholar] [CrossRef]
- Shi, W.; Ju, Y.; Bian, R.; Li, L.; Joseph, S.; Mitchell, D.R. . & Pan, G. Biochar bound urea boosts plant growth and reduces nitrogen leaching. Sci. Total Environ. 2020, 701, 1–9. [Google Scholar]
- Agegnehu, G.; Srivastava, A.K.; Bird, M.I. The role of biochar and biochar-compost in improving soil quality and crop performance: A review. Appl. Soil Ecol. 2017, 119, 156–170. [Google Scholar] [CrossRef]
- Inal, A.; Gunes, A.Y. D. I. N.; Sahin, O.Z. G. E.; Taskin, M.B.; Kaya, E.C. Impacts of biochar and processed poultry manure, applied to a calcareous soil, on the growth of bean and maize. Soil Use Manage. 2015, 31, 106–113. [Google Scholar] [CrossRef]
- Vargas, M.F.; Mestre, M.V.; Vergara, C.; Maturano, P.; Petrignani, D.; Pesce, V.; Vasquez, F. Residual brewer’s Saccharomyces cerevisiae yeasts as biofertilizers in horticultural seedlings: Towards a sustainable industry and agriculture. Front. Ind. Microbiol. 2024, 2, 1–11. [Google Scholar] [CrossRef]
- Puligundla, P.; Mok, C.; Park, S. Advances in the valorization of spent brewer's yeast. IFSET 2020, 62, 1–18. [Google Scholar] [CrossRef]
- Sifton, M.A.; Smith, S.M.; Thomas, S.C. Biochar-biofertilizer combinations enhance growth and nutrient uptake in silver maple grown in an urban soil. PLoS ONE 2023, 1–31. [Google Scholar] [CrossRef]
- Williams, M.I.; Dumroese, R.K.; Page-Dumroese, D.S.; Hardegree, S.P. Can biochar be used as a seed coating to improve native plant germination and growth in arid conditions? J. Arid Environ. 2016, 125, 8–15. [Google Scholar] [CrossRef]
- Law, Y.K.; Lee, C.K.; Pang, C.C.; Hau, B.C. H.; Wu, J. Vegetation regeneration on natural terrain landslides in Hong Kong: Direct seeding of native species as a restoration tool. Land Degrad. Dev. 2023, 34, 751–762. [Google Scholar] [CrossRef]
- Zhang, K.; Khan, Z.; Yu, Q.; Qu, Z.; Liu, J.; Luo, T.; Zhu, K.; Bi, J.; Hu, L.; Luo, L. Biochar Coating Is a Sustainable and Economical Approach to Promote Seed Coating Technology, Seed Germination, Plant Performance, and Soil Health. Plants 2022, 11, 1–23. [Google Scholar] [CrossRef]
- Zhang, K.; Han, X.; Fu, Y.; Zhou, Y.; Khan, Z.; Bi, J.; Hu, L.; Luo, L. Biochar Coating as a Cost-Effective Delivery Approach to Promoting Seed Quality, Rice Germination, and Seedling Establishment. Plants 2023, 12, 1–16. [Google Scholar] [CrossRef]
- Kelly, C.N.; Peltz, C.D.; Stanton, M.; Rutherford, D.W.; Rostad, C.E. Biochar application to hardrock mine tailings: Soil quality, microbial activity, and toxic element sorption. Appl. Geochem. 2014, 43, 35–48. [Google Scholar] [CrossRef]
- Pedrini, S.; Merritt, D.J.; Stevens, J.; Dixon, K. Seed coating: Science or marketing spin? Trends Plant Sci. 2017, 22, 106–116. [Google Scholar] [CrossRef]
- Pedrini, S.; Balestrazzi, A.; Madsen, M.D.; Bhalsing, K.; Hardegree, S.P.; Dixon, K.W.; Kildisheva, O.A. Seed enhancement: Getting seeds restoration-ready. Restor. Ecol. 2020, 28, 266–275. [Google Scholar] [CrossRef]
- Thomas, S.C.; Liu, Y.; Tang, E. Polyvinyl acetate binders undermine the effectiveness of biochar-based seed coatings. Land 2024, in review.
- Abdel-Razik, S.A.; Sallam, N.M.; Eraky, A.M.; Hassan, M.H. A. Integrated control of root rot and wilt disease of faba bean by soil amendment with suppressive compost in combination with seed coating with an antagonistic yeast. Arch. Phytopathol. Pflanzenchutz 2012, 45, 1692–1704. [Google Scholar] [CrossRef]
- Muhammed, A.A.; Thomas, K.; Bin-Hamed, U. Feasibility of using brewers spent grain as a fertilizer in agriculture. Int. J. Technol. 2015, 10, 23–31. [Google Scholar] [CrossRef]
- Lonhienne, T.; Mason, M.G.; Ragan, M.A.; Hugenholtz, P.; Schmidt, S.; Paungfoo-Lonhienne, C. Yeast as a Biofertilizer Alters Plant Growth and Morphology. Crop Sci. 2014, 54, 785–790. [Google Scholar] [CrossRef]
- Wang, J.; Chen, C. Biosorption of heavy metals by Saccharomyces cerevisiae: A review. Biotechnol. Adv. 2006, 24, 427–451. [Google Scholar] [CrossRef]
- Pescatore, A.; Grassi, C.; Rizzo, A.M.; Orlandini, S.; Napoli, M. Effects of biochar on berseem clover (Trifolium alexandrinum, L.) growth and heavy metal (Cd, Cr, Cu, Ni, Pb, and Zn) accumulation. Chemosphere 2022, 287, 1–10. [Google Scholar] [CrossRef]
- Fashola, M.O.; Ngole-Jeme, V.M.; Babalola, O.O. Heavy Metal Pollution from Gold Mines: Environmental Effects and Bacterial Strategies for Resistance. IJERPH 2016, 13, 1–20. [Google Scholar] [CrossRef]
- Farhangi-Abriz, S.; Ghassemi-Golezani, K.; Torabian, S.; Qin, R. A meta-analysis to estimate the potential of biochar in improving nitrogen fixation and plant biomass of legumes. Biomass Convers. and Biorefin. 2024, 14, 3293–3303. [Google Scholar] [CrossRef]
- Zheng, W.; Wang, Y.; Mo, J.; Zeng, P.; Chen, J.; Sun, C. Effects of biochar application and flucation on the growth, and cadmium and nutrient uptake of Trifolium repens with different planting densities in Cd-contaminated soils. Front. Plant Sci. 2023, 14, 1269082. [Google Scholar] [CrossRef]
- van de Voorde, T.F.; Bezemer, T.M.; Van Groenigen, J.W.; Jeffery, S.; Mommer, L. Soil biochar amendment in a nature restoration area: Effects on plant productivity and community composition. Ecol. Appl. 2014, 24, 1167–1177. [Google Scholar] [CrossRef]
- Bieser, J.M.; Thomas, S.C. Biochar and high-carbon wood ash effects on soil and vegetation in a boreal clearcut. Canadian J. For. Res. 2019, 49, 1124–1134. [Google Scholar] [CrossRef]
- Williams, J.M.; Thomas, S.C. Effects of high-carbon wood ash biochar on volunteer vegetation establishment and community composition on metal mine tailings. Restor. Ecol. 2023, 31, 1–15. [Google Scholar] [CrossRef]
- Williams, J.M.; Thomas, S.C. High carbon wood ash biochar for mine tailings restoration: A field assessment of planted tree performance and metals uptake. Sci. Total Environ. 2023, 901, 1–16. [Google Scholar] [CrossRef]
- Bjugstad, A.J.; Warren, C. Perennial forbs for wildlife habitat restoration on mined lands in the northern Great Plains. In Proceedings of the 62nd annual conference of the Western Association of Fish and Wildlife Agencies, Las Vegas, Nevada, United States of America, 19-22 July 1982. [Google Scholar]
- Large, R.R.; Bula, S.W.; Maslennikov, V.V. A Carbonaceous Sedimentary Source-Rock Model for Carlin-Type and Orogenic Gold Deposits. Econ. Geol. 2011, 106, 331–358. [Google Scholar] [CrossRef]
- Groves, D.I.; Goldfarb, R.J.; Gebre-Miriam, M.; Hagemann, S.G.; Robert, F. Orogenic gold deposits: A proposed classification in the context of their crustal distribution and relationship to other gold deposit types. Ore Geol. Rev. 1998, 13, 7–27. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria. 2023.
- Thomas, S.C. Biochar effects on germination and radicle extension in temperate tree seedlings under field conditions. Canadian J. For. Res. 2020, 51, 10–17. [Google Scholar] [CrossRef]
- Dijk, J.R.; Kranchev, M.; Blust, R.; Cuypers, A.; Vissenberg, K. Arabidopsis root growth and development under metal exposure presented in an adverse outcome pathway framework. Plant Cell Environ. 2022, 45, 737–750. [Google Scholar] [CrossRef]
- Crane-Droesch, A.; Abiven, S.; Jeffrey, S.; Torn, M.S. Heterogeneous global crop yield response to biochar: A meta-regression analysis. Environ. Res. Lett. 2013, 8, 1–9. [Google Scholar] [CrossRef]
- Joseph, S.; Cowie, L.A.; Zwieten, L.V.; Bolan, N.; Budai, A.; Buss, W.; Cayuela, M.L.; Graber, E.R.; Ippolito, J.A.; Kuzyakov, Y.; Luo, Y.; Ok, Y.S.; Palansooriya, K.N.; Shepherd, J.; Stephens, S.; Weng, Z.; Lehmann, J. How biochar works, and when it doesn't: A review of mechanisms controlling soil and plant responses to biochar. GCB Bioenergy 2021, 13, 1731–1764. [Google Scholar] [CrossRef]
- Ippolito, J.A.; Cui, L.; Kammann, C.; Wrage-Mönnig, N.; Estavillo, J.M.; Fuertes-Mendizabal, T.; Cayuela, M.L.; Sigua, G.; Novak, J.; Spokas, K.; Borchard, N. Feedstock choice, pyrolysis temperature and type influence biochar characteristics: A comprehensive meta-data analysis review. Biochar 2020, 2, 421–438. [Google Scholar] [CrossRef]
- Xu, Q.; Xu, Q.; Zhu, H.; Li, H.; Yin, W.; Feng, K.; Wang, S.; Wang, X. Does biochar application in heavy metal-contaminated soils affect soil micronutrient dynamics? Chemosphere, 2022, 290, 133349. [Google Scholar] [CrossRef]
- Ribeiro, R.A.; Giannini, T.C.; Gastauer, M.; Awade, M.; Siquiera, J.O. Topsoil application during the rehabilitation of a manganese tailing dam increases plant taxonomic, phylogenetic and functional diversity. J. Environ Manage 2018, 227, 1–9. [Google Scholar] [CrossRef]
- Navarro-Pedreno, J.; Almendro-Candel, M.B.; Gomez, I.; Jordan, M.M.; Pardo, F. Organic Mulching to Improve Mining Soil Restoration. In Assessment, Restoration and Reclamation of Mining Influenced Soils, 1st ed.; Bech, J., Bini, C., Pashkevich, M.A.; Academic Press: London, UK; pp. 375–386.
- Gul, S.; Yanni, S.F.; Whalen, J.K. Lignin controls on soil ecosystem services: Implications for biotechnological advances in biofuel crops. In Lignin, 1st ed.; Lu, F., Ed.; Nova Science Publishers Inc: New York, USA, 2014; pp. 375–416. [Google Scholar]
- Mango, L.; Kugedera, A.T. Effects of Re-vegetating Mine Tailings on Genetic Diversity and Role of Micro-fauna on Improving Mine Tailings (A Review). Int. J. Agric. & Agribus. 2019, 4, 108–123. [Google Scholar]
- Perez, D.R.; Gonzalez, F.; Ceballos, C.; Oneto, M.E.; Aronson, J. Direct seeding and outplantings in drylands of Argentinean Patagonia: Estimated costs, and prospects for large-scale restoration and rehabilitation. Restor. Ecol. 2019, 27, 1–12. [Google Scholar] [CrossRef]
Figure 1.
Seedling establishment by species, substrate and treatment, at 7, 14, and 75 days after seeds were sown, for (A-C) Trifolium and (D-F) Dalea. Significant effects of substrate (S), (B), yeast (Y) and interactions are indicated: * p< 0.05; full GLM results in Table 1. Letters indicate significant differences (p < 0.1) by post-hoc Dunn tests within each substrate. Means are plotted ±1 SE.
Figure 1.
Seedling establishment by species, substrate and treatment, at 7, 14, and 75 days after seeds were sown, for (A-C) Trifolium and (D-F) Dalea. Significant effects of substrate (S), (B), yeast (Y) and interactions are indicated: * p< 0.05; full GLM results in Table 1. Letters indicate significant differences (p < 0.1) by post-hoc Dunn tests within each substrate. Means are plotted ±1 SE.
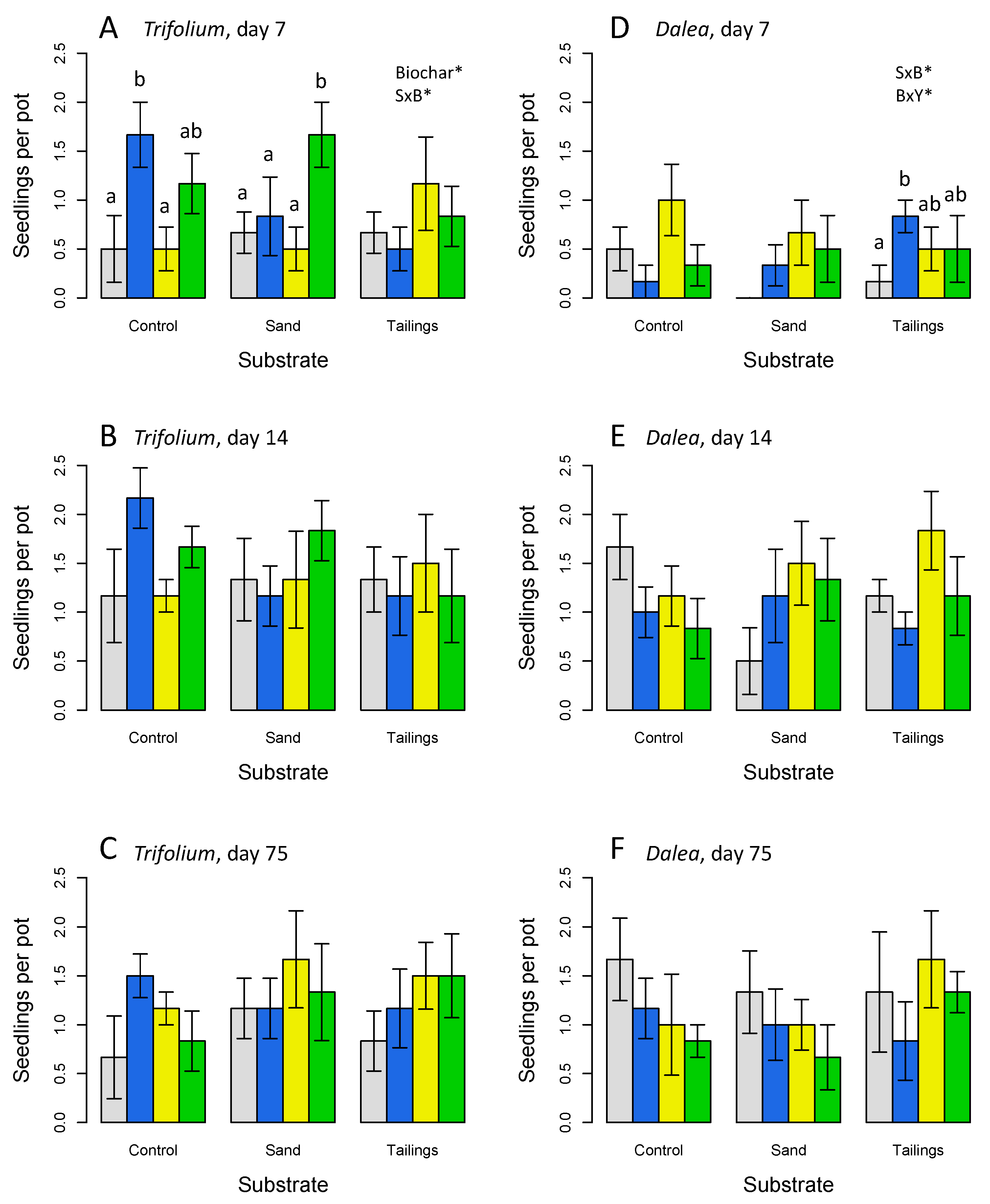
Figure 2.
Final biomass at harvest for (A,B) aboveground, (C,D) belowground, and (E,F) root biomass for Trifolium (A-C) and Dalea (D-F) grown for 75 days. Significant effects of substrate (S), (B), yeast (Y) and interactions are indicated: * p< 0.05; ** p< 0.01, *** p< 0.001; full ANOVA results in Table 2. Letters indicate significant differences (p < 0.05) by post-hoc HSD tests within each substrate. Means are plotted ±1 SE.
Figure 2.
Final biomass at harvest for (A,B) aboveground, (C,D) belowground, and (E,F) root biomass for Trifolium (A-C) and Dalea (D-F) grown for 75 days. Significant effects of substrate (S), (B), yeast (Y) and interactions are indicated: * p< 0.05; ** p< 0.01, *** p< 0.001; full ANOVA results in Table 2. Letters indicate significant differences (p < 0.05) by post-hoc HSD tests within each substrate. Means are plotted ±1 SE.
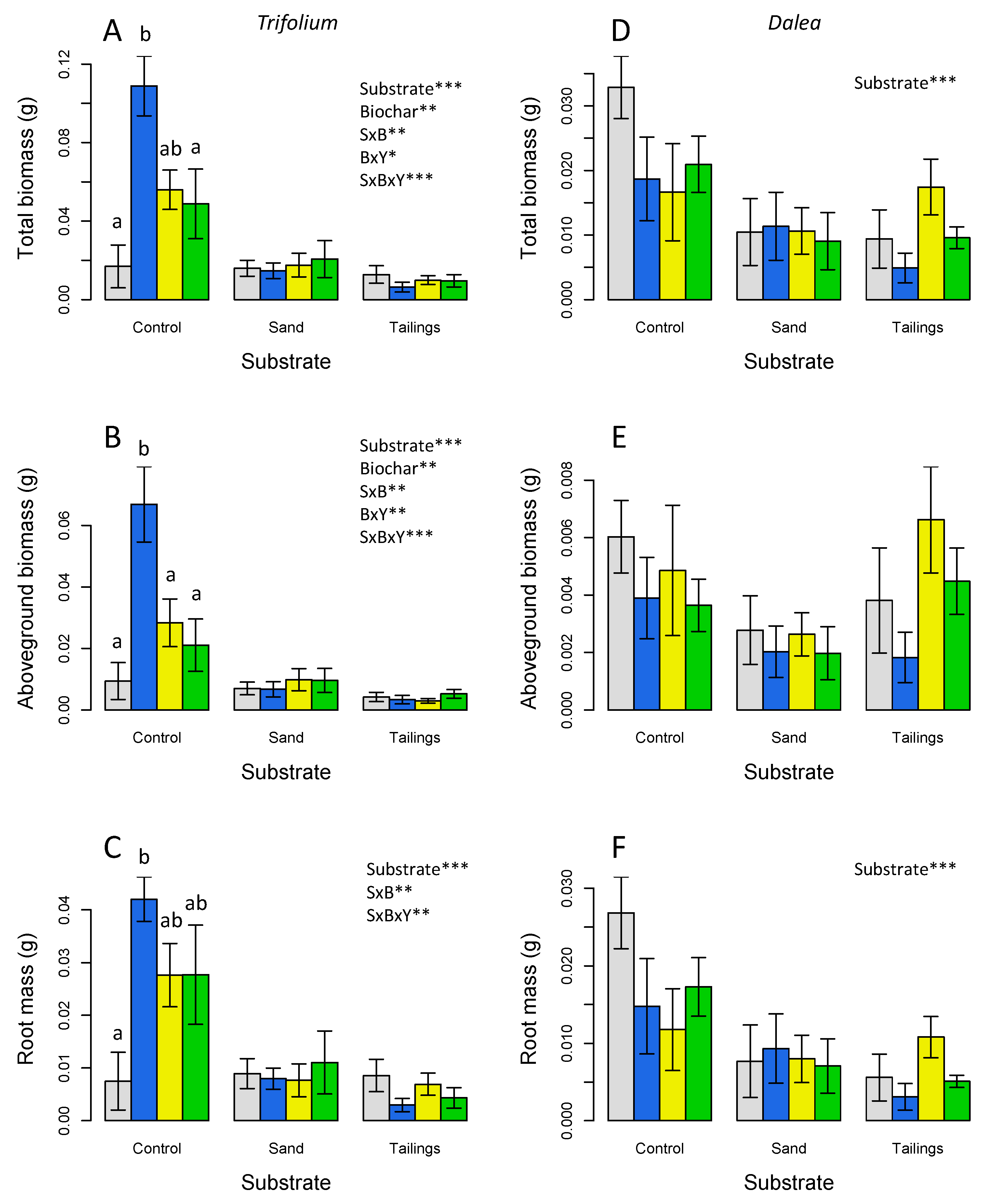
Figure 3.
Leaf area (A,D) leaf area ratio, (B,E), and root mass fraction (C,F) at harvest for Trifolium (A-C) and Dalea (D-F) grown for 75 days. Significant effects of substrate (S), (B), yeast (Y) and interactions are indicated: * p< 0.05; ** p< 0.01, *** p< 0.001; full ANOVA results in Table 2. Letters indicate significant differences (p < 0.05) by post-hoc HSD tests within each substrate. Means are plotted ±1 SE.
Figure 3.
Leaf area (A,D) leaf area ratio, (B,E), and root mass fraction (C,F) at harvest for Trifolium (A-C) and Dalea (D-F) grown for 75 days. Significant effects of substrate (S), (B), yeast (Y) and interactions are indicated: * p< 0.05; ** p< 0.01, *** p< 0.001; full ANOVA results in Table 2. Letters indicate significant differences (p < 0.05) by post-hoc HSD tests within each substrate. Means are plotted ±1 SE.

Table 1.
Analysis of deviance results (using proportional odds ordered logistic model) for germinant counts at days 7, 14, and 75 after seed additions. P-values ≤ 0.05 are in bold text.
Table 1.
Analysis of deviance results (using proportional odds ordered logistic model) for germinant counts at days 7, 14, and 75 after seed additions. P-values ≤ 0.05 are in bold text.
| Effect | 2 |
Trifolium df |
p |
Dalea df |
p | |
|---|---|---|---|---|---|---|
| Day 7 | ||||||
| Substrate | 0.659 | 2 | 0.7192 | 1.221 | 2 | 0.5431 |
| Biochar | 5.612 | 1 | 0.0178 | 0.018 | 1 | 0.8934 |
| Yeast | 0.756 | 1 | 0.3846 | 2.478 | 1 | 0.1155 |
| Substrate x biochar | 7.193 | 2 | 0.0274 | 6.037 | 2 | 0.0489 |
| Substrate x yeast | 1.600 | 2 | 0.4493 | 1.804 | 2 | 0.4058 |
| Biochar x yeast | 0.181 | 1 | 0.6702 | 4.078 | 1 | 0.0435 |
| Substrate x biochar x yeast | 3.388 | 2 | 0.1838 | 1.864 | 2 | 0.3937 |
| Day 14 | ||||||
| Substrate | 0.838 | 2 | 0.6577 | 0.777 | 2 | 0.6781 |
| Biochar | 1.338 | 1 | 0.2475 | 1.927 | 1 | 0.1651 |
| Yeast | 0.020 | 1 | 0.8864 | 1.959 | 1 | 0.1616 |
| Substrate x biochar | 3.596 | 2 | 0.1656 | 3.055 | 2 | 0.2171 |
| Substrate x yeast | 1.364 | 2 | 0.5057 | 5.152 | 2 | 0.0761 |
| Biochar x yeast | 0.029 | 1 | 0.8642 | 0.760 | 1 | 0.3834 |
| Substrate x biochar x yeast | 1.340 | 2 | 0.5118 | 1.608 | 2 | 0.4474 |
| Day 75 | ||||||
| Substrate | 1.656 | 2 | 0.4370 | 1.154 | 2 | 0.5616 |
| Biochar | 0.317 | 1 | 0.5735 | 1.676 | 1 | 0.1955 |
| Yeast | 1.310 | 1 | 0.2524 | 0.332 | 1 | 0.5642 |
| Substrate x biochar | 1.111 | 2 | 0.5739 | 0.181 | 2 | 0.9136 |
| Substrate x yeast | 0.845 | 2 | 0.6556 | 3.709 | 2 | 0.1566 |
| Biochar x yeast | 2.715 | 1 | 0.0994 | 0.103 | 1 | 0.7481 |
| Substrate x biochar x yeast | 1.442 | 2 | 0.4862 | 0.319 | 2 | 0.8525 |
Table 2.
Analysis variance for growth measurements at final harvest (75 days). P-values ≤ 0.05 are in bold text.
Table 2.
Analysis variance for growth measurements at final harvest (75 days). P-values ≤ 0.05 are in bold text.
| Effect | F |
Trifolium df |
p | F |
Dalea df |
p |
|---|---|---|---|---|---|---|
| Total biomass | ||||||
| Substrate | 33.15 | 2 | <0.0001 | 8.28 | 2 | 0.0007 |
| Biochar | 6.62 | 1 | 0.0126 | 1.91 | 1 | 0.1721 |
| Yeast | 0.18 | 1 | 0.6732 | 0.04 | 1 | 0.8360 |
| Substrate x biochar | 7.95 | 2 | 0.0009 | 0.41 | 2 | 0.6639 |
| Substrate x yeast | 0.68 | 2 | 0.5085 | 1.95 | 2 | 0.1511 |
| Biochar x yeast | 8.16 | 1 | 0.0059 | 0.58 | 1 | 0.4490 |
| Substrate x biochar x yeast | 11.31 | 2 | <0.0001 | 1.67 | 2 | 0.1970 |
| Aboveground biomass | ||||||
| Substrate | 28.71 | 2 | <0.0001 | 3.12 | 2 | 0.0514 |
| Biochar | 7.14 | 1 | 0.0097 | 3.58 | 1 | 0.0634 |
| Yeast | 1.15 | 1 | 0.2875 | 0.67 | 1 | 0.4178 |
| Substrate x biochar | 6.80 | 2 | 0.0022 | 0.27 | 2 | 0.7681 |
| Substrate x yeast | 2.52 | 2 | 0.0890 | 1.84 | 2 | 0.1684 |
| Biochar x yeast | 10.39 | 1 | 0.0020 | 0.03 | 1 | 0.8566 |
| Substrate x biochar x yeast | 12.07 | 2 | <0.0001 | 0.04 | 2 | 0.9579 |
| Root biomass | ||||||
| Substrate | 23.47 | 2 | <0.0001 | 9.93 | 2 | 0.0002 |
| Biochar | 3.34 | 1 | 0.0727 | 1.06 | 1 | 0.3068 |
| Yeast | 0.22 | 1 | 0.6432 | 0.29 | 1 | 0.5943 |
| Substrate x biochar | 5.98 | 2 | 0.0043 | 0.37 | 2 | 0.6946 |
| Substrate x yeast | 0.12 | 2 | 0.8900 | 1.59 | 2 | 0.2125 |
| Biochar x yeast | 3.00 | 1 | 0.0907 | 0.75 | 1 | 0.3898 |
| Substrate x biochar x yeast | 5.84 | 2 | 0.0048 | 2.26 | 2 | 0.1128 |
| Total leaf area | ||||||
| Substrate | 61.66 | 2 | <0.0001 | 1.35 | 2 | 0.2667 |
| Biochar | 14.12 | 1 | 0.0004 | 2.86 | 1 | 0.0963 |
| Yeast | 1.09 | 1 | 0.3007 | 0.25 | 1 | 0.6203 |
| Substrate x biochar | 13.83 | 2 | <0.0001 | 0.08 | 2 | 0.9241 |
| Substrate x yeast | 0.77 | 2 | 0.4661 | 2.04 | 2 | 0.1400 |
| Biochar x yeast | 13.44 | 1 | 0.0005 | 0.25 | 1 | 0.6168 |
| Substrate x biochar x yeast | 12.58 | 2 | <0.0001 | 0.01 | 2 | 0.9858 |
| Leaf area ratio | ||||||
| Substrate | 78.26 | 2 | <0.0001 | 3.50 | 2 | 0.0404 |
| Biochar | 2.20 | 1 | 0.1454 | 0.01 | 1 | 0.9127 |
| Yeast | 0.04 | 1 | 0.8333 | 0.26 | 1 | 0.6135 |
| Substrate x biochar | 2.12 | 2 | 0.1325 | 0.73 | 2 | 0.4894 |
| Substrate x yeast | 0.49 | 2 | 0.6175 | 1.86 | 2 | 0.1700 |
| Biochar x yeast | 0.06 | 1 | 0.8049 | 0.00 | 1 | 0.9793 |
| Substrate x biochar x yeast | 0.12 | 2 | 0.8830 | 1.09 | 2 | 0.3462 |
| Root mass fraction | ||||||
| Substrate | 1.20 | 2 | 0.3109 | 11.16 | 2 | 0.0001 |
| Biochar | 2.18 | 1 | 0.1470 | 0.11 | 1 | 0.7465 |
| Yeast | 0.20 | 1 | 0.6603 | 0.04 | 1 | 0.8523 |
| Substrate x biochar | 2.98 | 2 | 0.0610 | 1.17 | 2 | 0.3206 |
| Substrate x yeast | 3.10 | 2 | 0.0551 | 0.04 | 2 | 0.9569 |
| Biochar x yeast | 0.02 | 1 | 0.8816 | 0.04 | 1 | 0.8395 |
| Substrate x biochar x yeast | 0.31 | 2 | 0.7381 | 1.26 | 2 | 0.2947 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



