Submitted:
21 September 2025
Posted:
22 September 2025
You are already at the latest version
Abstract
Osteoarthritis (OA) is the most common joint disorder globally, affecting approxi-mately 595 million individuals and representing the first cause of chronic pain and disability. Recently, the infrapatellar fat pad (IFP), an intracapsular adipose tissue in the human knee joint, was recognized as an active and metabolically significant contributor to the pathophysiology of OA through the release of pro-inflammatory cytokines, adi-pokines, and growth factors that sustain inflammatory response, fibrotic remodeling, and neurogenic pain. The present review provides an overview of the pathophysiological significance of the IFP in OA and current and promising therapeutic strategies targeting this adipose structure. We summarize the available preclinical and translational evidence on conservative therapies, minimally invasive interventions, and surgical options as well as IFP-derived mesenchymal stromal cells as a potential cell source for cartilage repair. Overall, preclinical research indicates that the modulation of IFP inflammation and fi-brosis could alleviate pain and delay the progression of the disease. The superficial lo-cation and its central role in the pathogenesis of OA make the IFP a promising therapeutic target in knee OA (KOA).
Keywords:
osteoarthritis
; Infrapatellar Fat Pad
; adipose tissue
; adipokines
; pathophysiology
; treatment
1. Introduction
Osteoarthritis (OA) is the most common joint disorder worldwide, and is recognized as a primary cause of chronic pain and disability in adults. According to the data available from the Global Burden of Disease Study 2021, it has an impact on approximately 7.6% of the world's population in 2020 (95% UI: 6.8–8.4%), affecting 595 million people (with a 95% uncertainty interval of 535–656 million) [1]. It is a degenerative joint disease involving the progressive disruption of articular cartilage, subchondral bone remodeling, synovial joint inflammation, and changes in periarticular structures.
Previously considered as a purely mechanical condition resulting from joint "wear and tear," OA is currently recognized as a complex, multifactorial disease that affects all components of the joint, including articular cartilage, subchondral bone, ligaments, and surrounding muscles. As defined by the Osteoarthritis Research Society International (OARSI), OA is characterized by joint impairment driven by cellular stress responses and the breakdown of the extracellular matrix [2]. The onset of the disease is determined by acute injuries or repetitive microtraumas which break the homeostatic balance of joint tissues and lead to metabolic dysregulation and may interact with genetic susceptibility and other environmental factors [3]. But a key driver in the pathophysiology of OA is represented by low-grade chronic inflammation affecting not only the cartilage but also the joint structures and surrounding tissues. Elevated levels of pro-inflammatory cytokines such as Interleukin (IL) -1β, Tumor Necrosis Factor (TNF)-α, and IL-6 in joint tissues promote the expression of catabolic enzymes, including matrix metalloproteinases (MMP) and aggrecanases, as well as molecules like nitric oxide (NO) and Prostaglandin E2 (PGE2), which collectively degrade the extracellular matrix and inhibit cartilage repair [4,5]. Synovitis is a frequent finding in OA and results from the activation of synovial cells by damage-associated molecular patterns (DAMPs), stimulating cytokine production and immune cell infiltration [6]. Oxidative stress exacerbates these inflammatory processes enhancing pro-inflammatory signaling and inducing chondrocyte apoptosis and senescence [7]. Furthermore, emerging evidence suggests that alterations in gut microbiota composition may disrupt host-microbe homeostasis, provoking immune activation and stimulating the gut-joint axis, thereby playing a role in the development of OA [8].
Taken together these events promote progressive morphological alterations such as degradation of cartilage, remodeling of subchondral bone, formation of osteophytes, and inflammation of the synovial membrane, ultimately leading to the loss of normal joint function and structural integrity.
Although any synovial (diarthrodial) joint could be affected, hips, knees, and interphalangeal and metacarpophalangeal joints of hands are most commonly affected sites [9,10]. Multiple factors, such as a structural disorder in joints, the aging process, and genetic predisposition, contribute to the pathogenesis of the disease.
Obesity is a major contributing factor through complex biomechanical and metabolic mechanisms acting at both systemic and local tissue levels [11,12]. Fatty tissue functions as a metabolically active secretory organ rather than an inert lipid reservoir, promoting the secretion of pro-inflammatory mediators, regulatory hormones, and additional bioactive mediators, collectively referred to as adipokines [13,14]. These secreted substances modulate critical biological functions including metabolic homeostasis, immune responses and they are critically involved in the regulation of bone remodeling, exerting their effects through endocrine, paracrine, and autocrine pathways. They mediate the activity of osteoblasts and osteoclasts, influencing the dynamic equilibrium between bone formation and resorption and affecting cartilage metabolism [15]. Leptin, for instance, has been shown to exert both anabolic and catabolic effects on bone, depending on whether it acts centrally via the hypothalamus or peripherally on bone cells. It induces chondrocytes to produce pro-inflammatory cytokines, adhesion molecules and matrix-degrading enzymes, thereby accelerating extracellular matrix breakdown. Adiponectin shows a pro-osteogenic effect during bone remodelling [16,17] and inhibits osteoclastogenesis [18]. Although sometimes linked to bone-protective effects, it also induces the release of interleukins, MMPs and nitric oxide (NO) in chondrocytes, which can promote cartilage damage. Other adipokines, such as resistin and visfatin, have been implicated in promoting a pro-inflammatory environment that may enhance osteoclast activity and bone resorption [19,20]. Visfatin, highly expressed in the IFP near osteophyte formation, reinforces local inflammation and enhances chondrocyte production of cytokines and chemokines, contributing further to cartilage catabolism [21].
Infrapatellar Fat Pad (IFP), synovium and osteophyte cells are active producers of inflammatory cytokines and adipokines. These molecules, in particular leptin and adiponectin, can upregulate inflammatory mediators including PGE2, IL-6 and IL-8, TNF-α, and vascular cell adhesion molecule 1 (VCAM-1) within the synovial fluid of the knee [22,23] modulating the infiltration of inflammatory cells into cartilage, finally inducing a degenerative cascade that exacerbates the progression of OA. Correspondingly, studies have demonstrated that adipokine concentrations are markedly elevated in individuals with OA, and these increased levels remain significantly correlated with disease severity even after adjusting for confounding factors such as age, sex, and Body Mass Index (BMI) [24,25].
The dysregulation of adipokine signaling promotes chronic low-grade inflammation, which collectively accelerate cartilage degradation through upregulation of MMP and aggrecanases. Furthermore, obesity- associated metabolic syndrome factors, including insulin resistance and dyslipidemia, contribute to synovial inflammation and chondrocyte dysfunction through advanced glycation end-product formation and oxidative stress pathways [26,27].
This review provides a comprehensive analysis of emerging therapeutic options targeting the IFP in Knee Osteoarthritis (KOA), examining their effects on inflammatory factors, fibrotic pathways and nervous sensitization. We combine preclinical and translational data to provide evidence of the central role of the IFP in OA pathophysiology, discussing future directions for clinical application.
Pathophysiological Roles of the Infrapatellar Fat Pad in OA
The IFP, also known as Hoffa's fat pad, is an intra-articular but extra-synovial adipose tissue situated in the anterior region of the knee joint (Figure 1). This specialized fibrofatty tissue is strategically located between the patellar ligament and synovial membrane extending from the inferior patellar pole to the tibial tuberosity [28]. Functionally, it is a complex biomechanical structure that serves as a sophisticated cushion that optimizes joint movement by dynamically accommodating synovial fluid redistribution throughout the flexion-extension cycle. Being an intra-articular fat depot, it is in direct anatomical continuity with synovial and cartilaginous structures making it particularly relevant to joint homeostasis [29].
In addition to its mechanical function, the IFP acts as a metabolically active organ and a major player in obesity-related OA pathogenesis. The IFP is a significant source of cytokines (IL-6 , IL-1β, TNF-α), adipokines (leptin, resistin, adiponectin), and growth factors (like vascular endothelial growth factor, VEGF) [30] (Figure 2). Aging and obesity promote adipocyte hypertrophy and immune cell infiltration, especially pro-inflammatory M1 polarized macrophages and T cells, which contribute to local inflammation [31]. Simultaneously, the collection of senescent cells in the IFP drives the senescence-associated secretory phenotype (SASP) [32], releasing matrix-degrading enzymes and enhancing cytokine networks. These changes may also negatively impact adjacent synovium and cartilage via paracrine effects, eliciting synovitis and promoting cartilage degradation.
Fibrotic remodeling represents the main of the OA changes in the IFP, with an extraordinary collection of extracellular matrix proteins, mostly collagen types I and III [33].
Remodeling is the final result of interconnected mechanisms, with Transforming growth factor-β (TGF-β) signaling pathways being the primary secretary, and pro-fibrotic M2 macrophages and constant low-grade inflammation offering a continuous stimule for matrix production [34]. The fibrotic remodeling changes the essential biomechanical features of the IFP, alters its compliance, compromises its capacity to absorb joint stress effectively, and accelerates the deteriorative changes in the whole compartment [35]. Experimental results verify this mechanism demonstrating that post-inflammatory irreversible structural changes in the IFP plays a key role in sustaining chronic pain in rat OA models [36].
The neurogenic inflammation within the IFP also plays a central role in mediating OA pain. The IFP is densely innervated by sensory and autonomic nerve fibers, especially by branches of the tibial and peroneal nerves [37]. In OA, the IFP shows increased expression of substance P and calcitonin gene-related peptide (CGRP), which play an important role in pain transmission and neurogenic inflammation [38,39]. Histological analysis showed an increase in the number of CGRP-positive nerve fiber endings in fibrous IFP, especially in the areas of neovascularization [40], with further increase as the OA progresses [41]. Additionally, IFP contains substance-P sensory nerve fibers that promote macrophage activation, which subsequently release of pro-inflammatory cytokines, such as IL-1β and IL-6 [28]. The presence of synovitis is associated with diminished pressure pain thresholds (PPTs) of the patella on Magnetic Resonance Imaging (MRI), which indicates an increased pain sensitivity and diminished pain functionality. Synovitis has been shown to be significantly associated with reduced PPTs at the patella, showing an increase in local pain sensitivity [42]. This finding suggests that synovial inflammation contributes to peripheral sensitization, in which nociceptive neurons in the joint become hyper-responsive to mechanical stimuli. The lowering of PPTs indicates a lower tolerance to pressure-induced pain, which hesitates into impaired pain inhibitory mechanisms and reduced functional capacity in patients with joint pathology. These changes in nociceptive processing are consistent with findings from previous studies linking synovial inflammation to clinical pain severity and structural progression in OA [43]. Furthermore, the role of synovitis in amplifying nociceptive signaling aligns with evidence showing that inflammatory mediators released within the synovial membrane, such as cytokines and prostaglandins, can directly sensitize joint afferents, thereby enhancing the subjective experience of pain [44].
The coexistence of inflammation, fibrosis and neo-innervation create the condition for a maladaptive pain amplification circuit potentially leading to a critical transition from acute-to-chronic OA pain while increasing overall nociceptive burden typical of knee KOA [45].
All the above-mentioned mechanisms combined with its subcutaneous localization, making IFP an easily accessible site for minimal invasive interventions, a newer therapeutic target to treat KOA.
2. Therapeutic Strategies Targeting the Infrapatellar Fat Pad in Osteoarthritis
In the following sections we will delineate the currently available therapeutic options targeting the IFP in KOA, ranging from conservative, minimally invasive and surgical treatments.
2.1. Conservative Therapies (Figure 3)
2.1.1. Anti-Inflammatory Agents
When comparing the inflammatory profile of the IFP from KOA patients and subjects with chondral injuries, patterns closely related to inflammation are exhibited.
Studies demonstrated that the IFP from OA patients secretes significantly higher levels of prostanoids (PGE2, PGF2α, and PGD2) compared to IFP from subjects with isolated chondral injuries, suggesting a distinctive inflammatory profile associated with OA pathogenesis. The secretion pattern of prostanoids in OA IFP samples can be stratified into two distinct subgroups: one characterized by markedly elevated prostanoid levels indicative of heightened inflammatory activity, and the other showing comparatively moderate reductions in prostanoid production [46]. This heterogeneity in prostanoid secretion reflects varying degrees of inflammatory microenviroment within the joint and may influence disease severity and symptoms [46]. Notably, the selective COX-2 inhibitor celecoxib has been shown to exert anti-inflammatory effects in both subgroups, primarily through the suppression of prostanoid synthesis by the IFP. The efficacy of celecoxib appears particularly pronounced in individuals with higher baseline prostanoid secretion, underscoring its role in modulating inflammation mediated by lipid-derived mediators in OA. These findings not only highlight the IFP as a dynamic source of pro-inflammatory prostanoids but also support targeted therapeutic strategies aiming to modulate local inflammatory pathways to alleviate joint inflammation and pain [46].
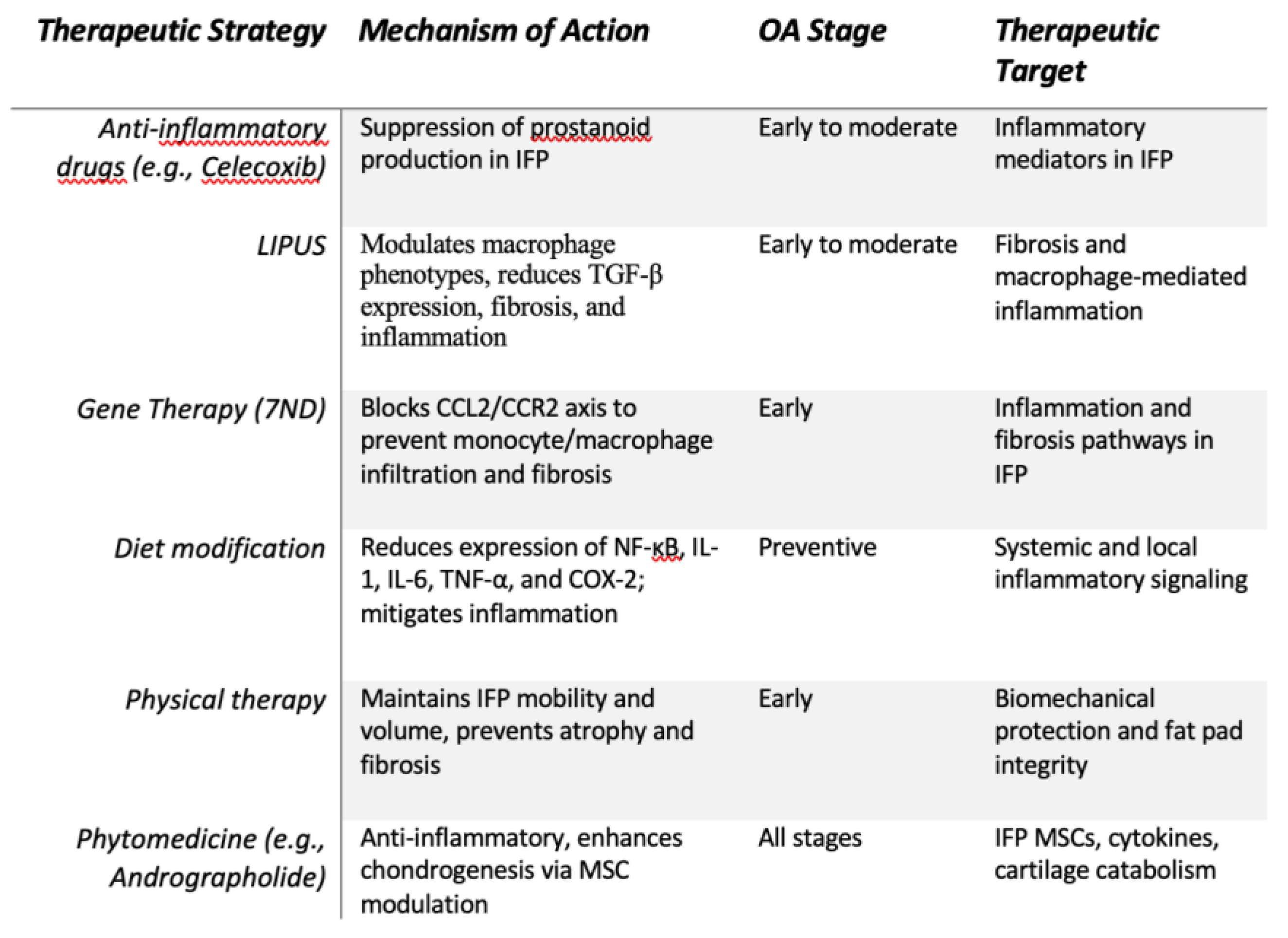
Figure 3.
Conservative Therapies. Summary of conservative therapeutic strategies for knee osteoarthritis, focusing on early and preventive interventions targeting inflammation, fibrosis, and biomechanical dysfunction. Treatments include pharmacological agents, lifestyle modifications, gene-based approaches, and phytomedicine, all aiming to modulate key pathological mechanisms in the IFP and surrounding joint environment.
Figure 3.
Conservative Therapies. Summary of conservative therapeutic strategies for knee osteoarthritis, focusing on early and preventive interventions targeting inflammation, fibrosis, and biomechanical dysfunction. Treatments include pharmacological agents, lifestyle modifications, gene-based approaches, and phytomedicine, all aiming to modulate key pathological mechanisms in the IFP and surrounding joint environment.
2.1.2. Low-Intensity Pulsed Ultrasound on the Fibrosis of the IFP
Low-Intensity Pulsed Ultrasound therapy (LIPUS) has demonstrated efficacy in alleviating KOA discomfort [47].
According to thermogenic mechanisms and high-frequency oscillatory stimulation, LIPUS exposure results in reduced edema and nociception and increased function [48,49].
Research conducted by Kitagawa and colleagues [50] demonstrated that LIPUS attenuated IFP fibrosis via multiple pathways. It inhibits Hypoxia Inducible Factor-1 (HIF-1) function, which correlates with macrophage phenotypic transformation [51], promoting M2 macrophage differentiation while reducing M1 phenotype (proinflammatory) prevalence. Furthermore, LIPUS has been documented to increase macrophage populations [52], decreases macrophage migration into the synovial membrane and reduce pro-inflammatory mediator release [53], showing a regulatory mechanism that promote modulation of macrophage activity through these combined effects [54].
Additionally, LIPUS leads to the downregulation of the genetic transcription of TGF-β [55], suppressing TGF-β-mediated fibrotic processes in the cultured synovial fibroblasts [56], inhibiting the osteoclastogenesis through the interference of the TGF-β1/Smad3 pathway [57], and subsequently the subchondral bone resorption.
2.1.3. Gene therapy
As previously mentioned fibrosclerosis of Hoffa's fat pad has a central role in the pathogenesis of degenerative knee arthropathy through histiocytic invasion, angiogenesis, and expansion of nociceptive innervation, resulting in persistent pain [40].
Monocyte chemoattractant protein-1 (CCL2/MCP-1), a strong chemotactic factor for histiocytes, shows increased expression in various fibrotic disorders like hepatic cirrhosis [58], nephrosclerosis [59], and progressive systemic sclerosis [60].
The CCL2/C-C Motif Chemokine Receptor 2 (CCR2) signaling pathway is the most frequently documented mechanism responsible for the recruitment of circulating monocytes in OA [61,62] as well as for pain development in murine OA models [63]. Elevated levels of CCL2 have been observed in the IFP, synovial tissue [64,65] and blood serum of individuals with OA [66]. N-terminally truncated or chemically altered CCL2 variants have been identified as functional receptor antagonists. Research suggests that targeting CCL2 could be a promising strategy to inhibit osteoclast formation, as the 7ND variant has been shown to prevent human osteoclast differentiation [67]. In addition, administration of 7ND markedly decreased osteolysis induced by wear particles and reduced both inflammatory cell infiltration and osteoclast numbers [68].
In a study conducted by Yoshimura et al. [69], employing a rat inflammatory arthropathy model, gene delivery of 7ND significantly suppressed the histological fibrosis in the IFP, decreased macrophage infiltration in synovial tissues, and led to a notable reduction in local CCL2 levels, apparently produced by infiltrating macrophages, ultimately inhibiting CCL2-mediated macrophage migration, and lowering the production of inflammatory cells and additional chemokines in the IFP.
Targeting CCL2 appears to play a central role in mitigating IFP fibrosis and limiting the inflammatory activation process42. Nevertheless, the precise in vivo mechanisms through which 7ND exerts its antagonistic effects still remain insufficiently understood. Additional investigations are required to fully clarify biological pathways influenced by 7ND.
2.1.4. Diet
Several studies, using high fat-diet-induced murine OA models, reported that IFP volume or area was increased after administration of lipid-enriched diet [70,71,72].
In humans, examining the association between anthropometric indices and size of the IFP, some studies found a positive relationship, while others reported no significant correlation. According to at least 2 studies no association between adiposity measures and IFP sizes was observed in normal-weight as well as obese subjects [73,74]. In contrast, other studies have shown significant correlations between IFP volume and body mass in females but not males [75].
Radakovich et al. indicated that the size of IFP was not dependent on body weight in the guinea pigs exposed to different dietary conditions. At the same time, the gene expression profiles within the IFP significantly varied. Specifically, this study showed higher transcription of pro-inflammatory genes (like Nuclear factor kappa light chain enhancer of activated B cells, NF-κB) in the IFP of hyperlipidic diet-fed guinea pigs than in the obese guinea and hypocaloric standard diet- fed pigs, enhancing expression of inflammatory signaling molecules including IL-1, IL-6, and TNF-a [76,77] that may directly act on pathologic processes related with inflammation. In that context, the COX-2 cascade and MCP-1 which is known to stimulate MMP activity, were overexpressed in hyperlipidic diet cohort inducing breakdown of type II and IV collagen within articular cartilage. Taken together, this evidence demonstrated that immune stimulation is greater in the knee joint of obese and hyperlipidic diet groups versus lean. In addition, these data support the concept that low-caloric diets with a low-fat content result in reduced knee joint inflammation.
2.1.5. Exercise
It has been established that the IFP exhibits dynamic movement during knee joint motion [78]. Several studies suggest that the IFP plays a functional role promoting efficient joint movement, maintaining joint space, and acting as a cushion to absorb mechanical loads, thereby protecting the joint structures [79,80].
Histological analyses knee immobilization in rat models [81,82] and in OA rat models induced by monoiodoacetate [83] demonstrated pathological changes in the IFP, including adipose tissue atrophy, fibrosis, and vascular congestion, particularly in the anterior region. These degenerative changes are associated with impaired joint function and are believed to contribute to joint contractures.
Therapeutic interventions involving joint mobilization and range-of-motion (ROM) exercises have been shown to reduce adipose tissue atrophy and preserve the structural integrity of the IFP [82]. For instance, Takeda et al. [84] demonstrated that joint movement not only reduced adipose cell degeneration in the IFP compared to non-treated controls, but may also exert a prophylactic effect. Nonetheless, the precise mechanisms underlying these therapeutic effects remain unclear and warrant further investigation to optimize treatment strategies.
2.1.6. Phytomedicine
Phytomedicine has been increasingly explored as a complementary approach for managing KOA, with several botanical compounds demonstrating anti-inflammatory and analgesic properties [85].
Among the phytotherapeutic compounds andrographolide, a bioactive labdane diterpenoid isolated from andrographis paniculata foliage, has demonstrated consistent therapeutic potential in OA management through multiple molecular mechanisms, including some that involve mesenchymal cells within the suprapatellar fat pad [86].
Experimental studies using both in vivo and in vitro models revealed that this compound exerts chondroprotective effects by suppressing multiple degradative enzymes, including MMP-1, MMP-3, and inducible nitric oxide synthase (iNOS), while simultaneously enhancing tissue inhibitor of metalloproteinase-1 (TIMP-1) expression in human chondrocytes exposed to IL-1β [87]. Notably, this natural compound facilitates the differentiation of mesenchymal stem cells derived from human suprapatellar adipose tissue toward osteogenic and chondrogenic lineages, evidenced by enhanced expression of specific markers including Runt-related transcription factor 2 (RUNX2), osteopontin, SOX9, and aggrecan, while concurrently suppressing adipogenic transcription factors Peroxisome Proliferator-Activated Receptor Gamma 2 (PPAR-γ2) and lipoprotein lipase [88]. These multifaceted mechanisms make andrographolide a promising therapeutic candidate for OA management.
Although the body of evidence supporting herbal medicine is increasing, study quality remains disparate and many trials have been hampered by small sample size, short treatment periods and lack of standardization of the herbal preparations. Furthermore, the bioavailability of several herbal compounds represents a challenge leading to development of improved formulations and combined therapies [89]. However, although limited, herbal medicines present a broadly safe possibility to the many who are seeking alternatives or complements for conventional therapies.
2.2. Minimally Invasive Treatments (Figure 4)
2.2.1. Intrarticular Injective Therapies
Hyaluronic acid (HA), a macromolecular glycosaminoglycan, is a key substance produced by the synovial membrane and present in hyaline cartilage [90]. Intra-articular administration of HA provides mechanical viscosupplementation to articular surfaces, diminishes cartilaginous degradation, promotes trophic support to chondrocytes, and enhances synthesis of native hyaluronan, consequently slowing the progression of OA [91,92].
Studies of Chen and Qu [93,94] collectively highlight that targeting the IFP with HA can effectively modulate local inflammation and promote both structural preservation and symptomatic relief in OA. In particular in the study of Chen [94] the researchers found, employing an in vitro model, that treatment with a combination of HA and platelet-rich plasma (PRP) significantly reduced the secretion of pro-inflammatory cytokines and adipokines from the IFP adipocytes. This anti-inflammatory effect helped to restore chondrocytes’ ability to produce a cartilage-like extracellular matrix, suggesting that HA can positively influence the local joint environment and protect cartilage by modifying the behavior of inflamed IFP cells. Study of Qu [93] extended this concept into an in vivo setting using a rat model of OA. Here, HA was applied in the form of a biodegradable sheet placed directly onto the IFP. The treatment led to a marked reduction in fibrotic tissue remodeling and nerve fiber ingrowth within the fat pad, both known contributors to pain in OA. Together, these studies suggest that HA does more than lubricate the joint; it actively modulates the inflammatory and fibrotic processes within the IFP that contribute to both cartilage degeneration and pain. Whether delivered as an injectable compound (with PRP) or as a physical implant (in sheet form), HA appears capable of altering the IFP's pathological signaling in OA, offering a potential disease-modifying and analgesic effect.
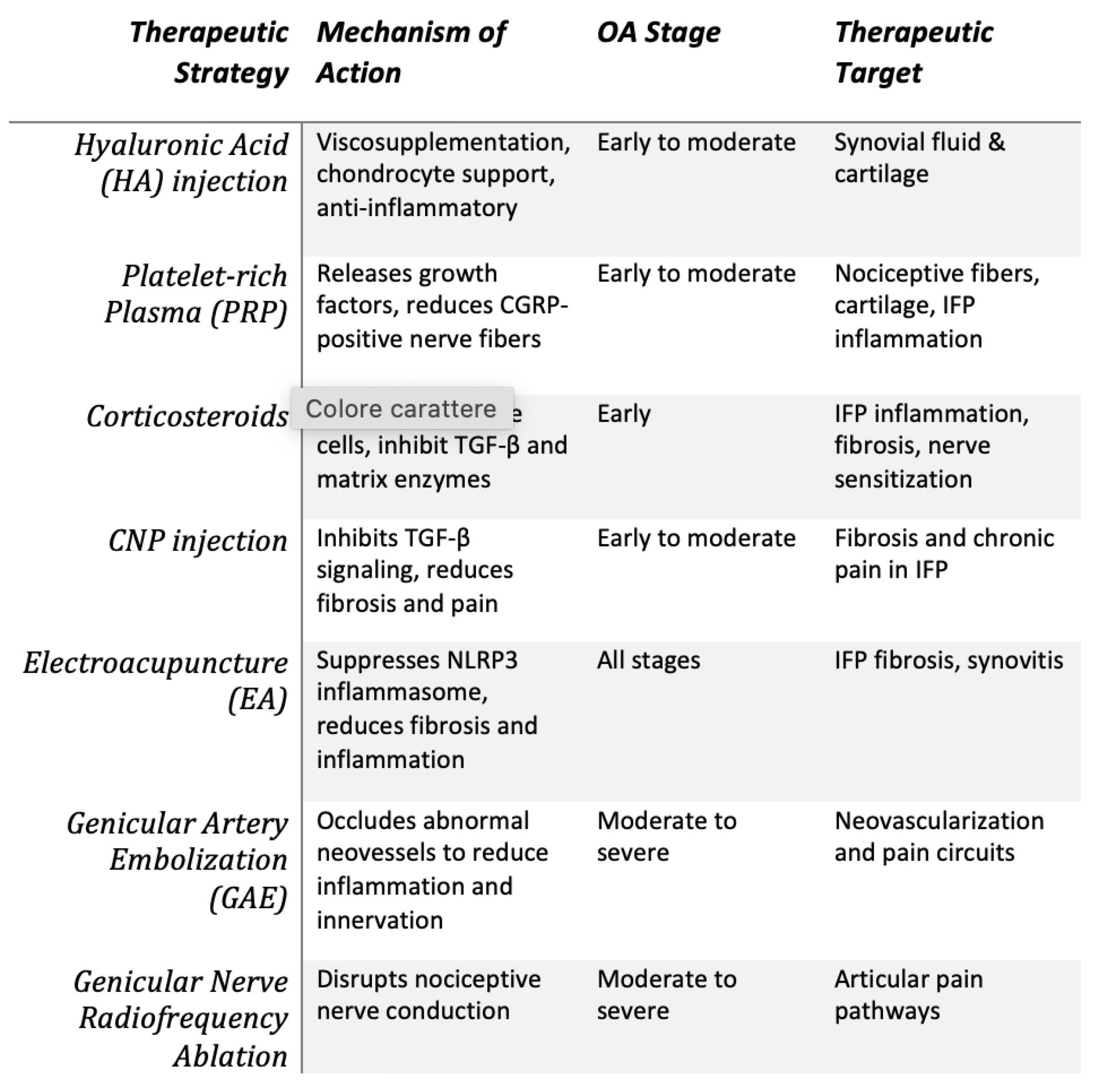
Figure 4.
Minimally Invasive Treatments. Overview of minimally invasive treatment strategies for KOA, highlighting therapeutic mechanisms, target tissues, and applicable disease stages. Approaches such as intra-articular injections, neuro-modulatory techniques, and vascular interventions address key pathological processes including inflammation, fibrosis, neo-innervation, and nociceptive signaling.
Figure 4.
Minimally Invasive Treatments. Overview of minimally invasive treatment strategies for KOA, highlighting therapeutic mechanisms, target tissues, and applicable disease stages. Approaches such as intra-articular injections, neuro-modulatory techniques, and vascular interventions address key pathological processes including inflammation, fibrosis, neo-innervation, and nociceptive signaling.
Plateled rich plasma (PRP). Recent advances in tissue biology elucidated the crucial role of growth factors (GFs) in maintaining tissue homeostasis and orchestrating reparative responses to pathological insults, with extensive in vitro and in vivo studies examining their impact on chondral regeneration [95]. PRP intrarticular injection causes an initial burst, then a sustained release of biologically active substances, including key growth factors such as platelet-derived growth factor (PDGF), TGF-β, insulin-like growth factor I (IGF-I), and VEGF [96]. These signaling proteins play essential roles in tissue repair processes, including the prevention of chondrocyte apoptosis, promotion of angiogenesis and osteogenesis, regulation of the inflammatory response, and stimulation of collagen production. Moreover, additional components released by platelets, such as fibrin, contribute to tissue regeneration by serving as both a structural matrix and a chemoattractant, facilitating the recruitment of stem cells and other reparative cell populations to the site of injury [97].
Research conducted by Araya and colleagues [98] revealed that intra-articular administration of pure PRP can reduce pain and suppress the advancement of synovial inflammation, infrapatellar adipose tissue architectural alterations, and articular cartilage deterioration in the short-term period. Furthermore, it was detected a reduction in the expression of CGRP nerve fibers in IFP of KOA patients treated with pure PRP. The diminished presence of CGRP-positive nerve fibers after PRP treatment suggests that PRP has the ability to modulate the local neural environment mitigating nociceptive sensitization. Notably, this effect appears to be independent of the degree of cartilage damage within the knee joint [99]. Even in cases where the chondral injury is severe, the beneficial influence of PRP on nerve fiber density and pain modulation within the fat pad remains evident. This observation is consistent with findings from prior studies, which have similarly indicated that PRP can exert analgesic effects through mechanisms beyond cartilage repair [100,101].
Corticosteroids (CCS). Steroids modulate immune cells in the IFP, as they regulate the polarization of macrophages [102], promoting immune cells switching from the pro-inflammatory M1 phenotype into an anti-inflammatory M2 phenotype and inhibiting T-cell activity and proliferation [103]. Moreover, CCS also have powerful anti-catabolic effects as they inhibit the production of MMPs and other proteolytic enzymes responsible for cartilage degradation.
The analgesic efficacy of CCS in IFP-related pathology acts through multiple mechanisms, including the suppression of inflammatory mediator production, attenuation of peripheral nerve sensitization, and modulation of nociceptive signal transduction [104,105]. Additionally, these therapeutic agents exhibit anti-fibrotic properties by interfering with TGF-β signaling pathways, suggesting their potential role in reducing excessive collagen deposition and promoting favorable tissue remodeling within the fat pad [106,107].
This multifaceted mechanism of action positions CCS as valuable therapeutic interventions for addressing both the inflammatory and structural alterations characteristic of IFP pathology in OA conditions. According to numerous studies, the difference in effect of the steroid can be particularly appreciated in the early stages of the disease, where proinflammatory mediators are predominantly secreted by tissue macrophages and fibroblast-like synoviocytes located within the synovial membrane [108].
Research conducted by Heard and colleagues [109] revealed that in a surgically induced IFP impingement, a single intra-articular injection of CCS administered during the operative procedure effectively attenuated the acute inflammatory response within the articular space, primarily by suppressing cellular proliferation. However, this intervention proved inadequate for sustaining joint protection through the 9-week postoperative period.
It was also demonstrated that MPA (Methylprednisolone acetate) attenuated the IL1β-mediated transcriptional upregulation of MMPs genes within the hyaline cartilage of various joint surfaces (including the kneecap and trochlear groove, distal femoral articular surfaces, and proximal tibial articular surface), synovial tissue, and IFP, demonstrating a dose-dependent suppressive effect [110].
The GLITTERS randomized controlled trial, the first that investigated corticosteroid administration directly into the IFP rather than the intra-articular space, failed to demonstrate favorable outcomes. The study revealed that targeted corticosteroid infiltration of the infrapatellar adipose tissue showed no significant efficacy in mitigating articular pain or diminishing the volume of effusion-synovitis in patients with inflammatory KOA [111]. Nevertheless, the emergence of encouraging trends warrants further investigation through large-scale, multicentric clinical trials to comprehensively evaluate the therapeutic potential and clinical relevance of this intervention.
C-type natriuretic peptides (CNP). C-type natriuretic peptide belongs to the natriuretic peptide superfamily and has been documented to suppress the transcriptional activity of type I collagen in both pulmonary alveolar epithelial cells and cardiac myocytes through downregulation of the TGF-β signaling cascade [112,113].
Research conducted by An and colleagues [114] demonstrated that the administration of CNP via intra-articular injection effectively inhibited fibroproliferative alterations within the IFP, resulting in significant mitigation of chronic knee pain. Furthermore, local articular administration of CNP was shown to attenuate the progressive deterioration of hyaline cartilage. These findings indicate that CNP represents a promising disease-modifying OA drug.
- Comparison between injective therapies
A meta-analysis of Gupta et collegues [115] demonstrated the superior therapeutic effectiveness of intra-synovial administration of PRP in association with HA, in providing sustained pain relief and enhanced joint function for KOA throughout a monitoring period of twelve months, when compared with isolated PRP administration, concurrent HA and CCS administration, or isolated administration of HA, CCS, or normal saline/control injections. In fact, studies have shown that the combination of PRP and HA exerts a synergistic effect, enhancing the activity of key molecular mediators involved in tissue homeostasis. These include pro-inflammatory mediators, catabolic enzymes, cytokines, and various growth factors [116].
Qiao et al. demonstrated that CCS provided superior short-term pain relief compared to HA, corroborating earlier findings by Naimi, who reported that CCS significantly reduced pain and improved functional outcomes in the early post-treatment phase (within 6 weeks) relative to placebo. Nonetheless, no significant clinical advantage was observed when CCS was directly compared to HA [117]. Over an extended follow-up period, however, HA was associated to more pronounced and sustained analgesic effect than CCS [118]. CCS and HA demonstrated comparable outcomes to placebo in some investigations [119].
Nevertheless, other studies reported divergent findings, indicating that both CCS and HA may exhibit superior efficacy relative to placebo [120,121].
The temporal persistence of therapeutic benefits following PRP administration continues to be a subject of uncertainty. In their investigation, Filardo and colleagues [95] examined the sustainability of positive outcomes from PRP treatment throughout a two-year observational period. Their findings demonstrated that while all assessed clinical parameters exhibited significant deterioration at the 24-month timepoint relative to the 12-month assessment, these measurements remained superior to pre-treatment values. The median timeframe for sustained clinical benefit was determined to be 9 months.
Autologous platelet concentrate therapy constitutes a readily available, cost-effective, and minimally invasive therapeutic approach [122]. While the management of KOA through concurrent administration of platelet-rich plasma and HA may incur can be more expensive and procedural complexity, it remains a potentially favorable alternative when considering the monetary implications and possible complications deriving from surgical intervention.
2.2.2. Electroacupuncture
Electroacupuncture (EA), extensively used in clinical settings, involves the application of pulsed electrical stimulation to acupuncture points following needle insertion. This non-invasive therapeutic modality is broadly employed in the management of KOA, with various acupuncture techniques having demonstrated clinical efficacy [123]. As a conventional intervention for pain management, its therapeutic properties including analgesia, sedative effects, enhancement of circulatory function, and modulation of muscular tone have been validated through extensive clinical evidence, leading to improvement of joint pain, edema and functional mobility associated with KOA.
Research conducted by Zhang and colleagues [124] revealed that EA not only mitigated cartilaginous deterioration but also attenuated synovial inflammatory processes and IFP fibrotic changes, suppressing NLR family pyrin domain containing 3 (NLRP3) inflammasome activation, thereby reducing the inflammatory microenvironment characteristic of KOA.
2.2.3. Genicular Artery Embolization (GAE)
The primary blood supply to the IFP originates from branches of the genicular arterial system, which penetrate the tissue and form an extensive network of anastomotic connections throughout the structure [125]. This rich periarticular vascular network surrounding the knee joint is clinically significant, as it influences the fat pad’s response to injury, its healing potential, and its involvement in various pathological conditions affecting the knee [126].
Within this perspective, GAE has been introduced during the last ten years as an innovative percutaneous intervention for managing pain associated with degenerative knee joint disease [127]. This procedure specifically targets the abnormal neovascularization in arthritic knee joint, a vascular network believed to contribute to inflammatory processes, promoting nociception and synovial hyperplasia [128]. Through the selective obstruction of pathological branches of the genicular vessels, GAE diminishes blood flow to the synovium and reduce inflammatory activity, consequently providing pain relief while preserving the mechanical integrity of the joint.
A meta-analysis conducted by Abussa and colleagues [129] revealed that GAE produces significant analgesia and improved joint functionality in a significant number of patients with KOA with approximately 70–80% achieving therapeutically significant improvement within six months post-procedure. The investigation additionally demonstrated decreased dependence on pain medications following the intervention.
The adverse event profile of GAE appears highly favorable, with absence of major complications, no evidence of chronic articular damage, and no detrimental impact on subsequent operative procedures, making it a promising therapeutic option for patients where surgery is contraindicated [130].
However, therapeutic efficacy is not uniformly achieved [129]. Therefore, careful candidate identification and appropriate prognostic counseling remains crucial.
2.2.4. Genicular Nerve-targeted Cooled and Pulsed Radiofrequency Ablation
The IFP, is richly innervated, creating a complex neural network that contributes significantly to knee pain experience in KOA patients. The fat pad is primarily innervated by branches of the femoral, common peroneal, and saphenous nerves, with particular contribution from the infrapatellar branch of the saphenous nerve and articular branches that form part of the genicular nerve complex [131,132].
The dense neural supply of the IFP includes both sensory and sympathetic nerve fibers, containing numerous nociceptors and mechanoreceptors that make it particularly sensitive to mechanical stress and inflammatory mediators [39]. In KOA, the fat pad often becomes fibrotic, inflamed, and hyperinnervated, leading to enhanced pain transmission and contributing to the overall pain experience. Radiofrequency ablation has emerged as promising interventional approach for managing chronic knee pain by targeting the neural structures innervating the joint, including those supplying the IFP.
The technique involves using thermal or pulsed energy to disrupt nerve conduction, thereby interrupting pain signal transmission [133,134].
Conventional radiofrequency ablation creates thermal lesions at temperatures exceeding 80 °C [135], while cooled radiofrequency maintains tissue temperatures around 60-70 °C through internal probe cooling, allowing for larger lesion volumes [136].
A systematic review and meta-analysis by Soetjahjo et al. [137] found that both cooled and pulsed Radiofrequency Ablation (RFA) techniques targeting genicular nerves provided significant pain reduction in KOA patients at all follow-up intervals (1, 3, 6, and 12 months- post-treatment), with no significant difference in analgesic effectiveness between the two methods. Pain scores, measured using Visual Analogue Scale (VAS) or Numeric Rating Scale (NRS), showed substantial improvement particularly at the 6-month mark. However, the techniques differed in their impact on physical function as measured by Western Ontario and McMaster Universities Osteoarthritis Index (WOMAC) scores. While pulsed RFA demonstrated significant functional improvement for up to 3 months, cooled RFA showed no significant functional improvement at any follow-up period. The meta-analysis revealed also that both techniques were generally safe with minimal adverse events reported, including only minor complications such as injection site pain, numbness, and stiffness that resolved quickly. Both techniques offer effective pain relief for KOA patients for at least 6 months, though the long-term benefits beyond 12 months remain uncertain, possibly due to nerve regeneration.
This suggests that these non-conventional RFA techniques provide a valuable minimally invasive treatment option for managing chronic KOA pain when conservative therapies have failed.
2.3. Surgery (Figure 5)
Evidence demonstrates that specific anatomical regions within the IFP, specifically the upper and lower sections, exhibit heightened vulnerability to biomechanical stress and loading forces during articular motion [138]. Such focal loading patterns induce hypoxic tissue damage, inflammatory responses, and promote releasing of inflammatory mediators and adipose-derived factors, thereby intensifying the manifestations of KOA [139].
The significant nociceptive innervation throughout the IFP makes also these compression zones primary generators of articular pain, perpetuating the pathological cascade [140].
Figure 5.
Surgical therapy. Surgical resection of the IFP as a therapeutic strategy for moderate to severe knee osteoarthritis (OA). This intervention targets fibrotic and inflamed IFP tissue, aiming to reduce inflammation and alleviate pain by modifying IFP structure and its pathological contribution to OA progression.
Figure 5.
Surgical therapy. Surgical resection of the IFP as a therapeutic strategy for moderate to severe knee osteoarthritis (OA). This intervention targets fibrotic and inflamed IFP tissue, aiming to reduce inflammation and alleviate pain by modifying IFP structure and its pathological contribution to OA progression.

Studies demonstrated that selective removal of the IFP, whether through partial or total excision, successfully diminished articular inflammation while preserving chondral structure, thereby slowing the progression of KOA [141,142].
Notably, targeted excision of adipose tissue enhanced biomechanical stability of the joint, maintained the structural integrity of the subchondral osseous tissue, and diminished degradation of the articular surface. Furthermore, immunohistochemistry evaluation revealed significant suppression of pro-inflammatory mediators, including IL-6, TNF-a, and MMP-3, alongside optimized retention of type II collagen [143].
The best approach to the IFP during total knee arthroplasty in patients with KOA remains controversial. A systematic review by Yao et al. [144] found inconsistent evidence regarding whether IFP resection leads to significantly inferior outcomes compared to preservation. Subsequently, Rajbhandari et al. [145] reported that IFP resection yielded marginally superior patient-reported functional outcomes as measured by Oxford Knee Scores; however, no significant difference in patient satisfaction was observed between the resection and preservation groups based on SF-12 scores.
3. The Use of Infrapatellar Mesenchymal Stromal Cells in Joint Cartilage Repair
Multipotent stromal cells (MSCs) exhibit immunomodulatory and nutritive properties involving anti-inflammatory, vasculogenic, and fibrosis-inhibiting mechanisms [146].
Research demonstrated that progenitor cells exhibiting stem cell properties are found within IFP, sharing characteristics with, though not being identical to, mesenchymal stromal cells of bone marrow origin [147]. These Infrapatellar Fat Pad-derived Stem Cells (IFPSCs) demonstrate reparative capacity and immunomodulatory properties, modulating macrophage phenotypic polarization, while exhibiting superior cartilage-forming potential compared to other mesenchymal stromal cells [148,149,150] due to their proximity to the knee joint and similarity to subcutaneous adipose tissue cells.
The mesenchymal-derived extracellular vesicles from the infrapatellar adipose tissue (MSCIPFP-Exos) mitigated OA progression in vivo through suppression of apoptosis, induction of extracellular matrix production, and downregulation of degradative mediators in vitro. Furthermore, MSCIPFP-Exos substantially elevated autophagic activity within cartilage cells, mediated in part through microRNA 100-5p (miR100-5p)-dependent suppression of mTOR autophagy signaling cascade [151].
Research demonstrates that bone morphogenetic protein-14 (BMP-14) and bone morphogenetic protein-7 (BMP-7) exhibit significantly elevated expression levels within the synovial membrane compared to hyaline cartilage [152], which may substantially influence the proliferation, lineage commitment, and biological activity of mesenchymal stromal cells derived from the IFP [153,154].
For these reasons, the stromal cells of IFP tissue have been focused on as potential therapeutic cell source for localized chondral defect in knee joint due to their anatomical location and surgical accessibility [155,156].
In addition, IFPSCs exhibit intrinsic migratory abilities toward proliferating chondrocytes, suggesting their potential involvement in the endogenous repair of articular cartilage. This chemotactic behavior shows that IFPSCs are not only passive progenitor cells but may actively home to sites of tissue damage, where their regenerative functions can be most effectively exerted. Moreover, when cultured within a HA-enriched environment, IFPSCs exhibit significantly enhanced chondrogenic differentiation capacity, characterized by increased expression of cartilage-specific genes and improved extracellular matrix synthesis [157].
These findings highlight the dual role of HA as both a structural component and a bioactive modulator within the joint microenvironment. By influencing stem cell behavior, particularly migration and lineage commitment, HA may serve as a key factor in leading more effective cartilage repair. Consequently, combined use of an HA-enriched microenvironment into IFPSCs-based therapeutic strategies could substantially improve the quality and long-term efficacy of joint cartilage repair, especially in degenerative or post-traumatic conditions [150,158].
Although the matrix microenvironment can stimulate stem cell proliferation and differentiation potential, acellular matrix derived from aged cells fails to support the revitalization of mature stromal stem cells in vitro, hampering individualized cellular therapy.
Research conducted by Wang and colleagues [159] demonstrated that viral vector-mediated gene transfer altered the cartilage-forming, and fat-forming capabilities of IPFSCs, affecting mitotically exhausted IPFSCs proliferation rates and lineage commitment preferences. These observations suggest that both acellular matrix cultivation and life-extension methodologies may be employed for proliferation-exhausted adult stem cells' proliferation and cell-type specialization, representing an opportunity for future cellular-mediated tissue repair therapies.
Preconditioning represents a promising strategy for enhancing the therapeutic potential of extracellular vesicles derived from IFPSCs in KOA management.
Specifically, research conducted by Wu and colleagues [160] demonstrated that TNF-a priming enhanced extracellular vesicle production from IFPSCs compared to non-primed cells (via activation of the Phosphoinositide 3-kinase/ Protein Kinase B, PI3K/AKT, signaling cascade), improving therapeutic efficacy in relieving disease-associated joint alterations in murine OA models.
4. Discussion
This review presents an in-depth evaluation of novel therapeutic approaches targeting the IFP in KOA, with particular focus on their impact on inflammatory mediators, fibrotic remodeling processes, and nociceptive sensitization. By integrating findings from both preclinical models and translational research, we highlight the pivotal role of the IFP in the pathophysiological mechanisms underlying OA and examine translational relevance for patient care.
It has been studied how, in patients with OA, the IFP not merely represents a fat storage and a biomechanical shock absorber, but also an active inflammatory organ, displaying hypertrophic adipocytes, infiltration of proinflammatory M1 macrophages, and a senescence-associated secretory phenotype (SASP) [31,32].
This structure also stimulates the secretion of prostaglandins such as PGE2, PGF2a, and PGD2, which contribute, through paracrine stimulation, to the development of a chronic inflammatory trigger in the synovium and cartilage of adjacent tissues [161].
Among the pathological changes of the IFP in KOA, fibrotic remodeling is the most important, predominantly driven by the TGF-β signaling pathway and profibrotic M1 macrophages [33]. These structural changes alter the biomechanical properties of the tissue, promoting a change in loading distribution and thus supporting degeneration of joint structures.
The IFP is also a densely innervated structure, influencing pain stimulation. In KOA, increased expression of CGRP and substance P within IFP gives rise to a neurogenic inflammatory loop that contributes to the transition from acute to chronic OA pain [40,41].
Several treatments have been developed specifically to target the IFP, ranging from pharmacological therapies to interventional procedures. The use of nonsteroidal anti-inflammatory drugs, particularly celecoxib, has demonstrated efficacy in inhibiting prostanoid production, particularly in patients with significant inflammatory profiles [46]. However, inflammatory patterns are highly heterogeneous, so phenotyping of the IFP may be useful to establish personalized therapy and achieve improved therapeutic outcomes.
LIPUS, a minimally invasive technique, shows therapeutic potential in reducing IFP fibrosis through several mechanisms, such as HIF-1 inhibition and macrophage phenotypic modulation [47,51]. Specifically, this minimally invasive approach inhibits TGF-β signaling and stimulates macrophage differentiation into M2, reducing the population of proinflammatory M1 macrophages [52].
Gene therapy has also shown promising results, demonstrating how CCL2 receptor antagonists can suppress OA-related histopathological changes in the IFP, reducing macrophage infiltration into synovial tissues and suppressing additional intrinsic chemokine production [69]. Intra-articular injective therapies have also demonstrated efficacy, particularly when used in combination (e.g., PRP with HA), thanks to a synergistic mechanism that simultaneously modulates inflammation and biomechanical function [116]. However, the limited duration of the therapeutic effect, approximately 9 months, should prompt clinicians to consider treatment cycles rather than single administrations.
Genicular artery embolization has shown significant functional improvement in 70–80% of patients, underscoring the important role of treating the vascular component, which promote chronic pain, by ablating neovascularized areas [129].
Regarding the surgical approach, selective excision of the IFP has been proven to reduce the progression of KOA, although there is no conclusive evidence for removing this structure during knee replacement surgery in patients with KOA, and high-quality studies highlighting the best surgical strategy are lacking [141,142].
The most innovative therapeutic approach involves the use of IFP-derived mesenchymal stromal cells, which have a natural tissue regenerative ability. These cells exhibit enhanced chondrogenic potential than other mesenchymal populations, suppress apoptosis, and stimulate matrix production, contributing to reduce the rate of OA progression [155,156].
Despite these advances, significant gaps persist in our knowledge and capacity to therapeutically target the IFP.
The pathological involvement of the IFP varies significantly among patients, and it would be important to identify biomarkers to stratify patients, working toward patient-specific treatment. Novel imaging tool able to detect more accurately the level of tissue inflammation, not just structural characteristics, are also needed. Moreover, the time course of IFP pathology during disease development has not been fully defined, limiting optimization of intervention timing.
Additionally, the predominance of rodent models in translation of preclinical findings may not fully capture the complexity of human IFP pathology, particularly regarding chronic OA development and the impact of comorbidities.
Future research must give priority to the development of non-invasive IFP phenotyping methods, advanced imaging techniques, and artificial intelligence-enhanced analysis to promote a personalized therapeutic approach.
5. Conclusion
In conclusion, the IFP emerges as a promising therapeutic target in OA management, offering multiple intervention opportunities across the spectrum of conservative, minimally invasive, and surgical approaches. The convergence of mechanistic understanding, preclinical efficacy data, and early clinical observations supports the continued investigation of IFP-targeted therapies. However, the translation of these insights into standardized clinical practice requires more standardized studies like randomized controlled trials specifically designed to evaluate IFP-targeted interventions, the development of reliable biomarkers for patient selection and the creation of algorithms that consider IFP assessment into routine OA management protocols.
Supplementary Materials
The following supporting information can be downloaded at Preprints.Org.
Author Contributions
All authors have read and agreed to the published version of the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| BMI | Body Mass Index |
| BMP-7 | Bone Morphogenetic Protein-7 |
| BMP-14 | Bone Morphogenetic Protein-14 |
| CCL2 | Monocyte chemoattractant protein-1 |
| CCR2 | C-C Motif Chemokine Receptor 2 |
| CCS | Corticosteroids |
| CNP | C-type natriuretic peptide |
| DAMPS | Damage Associated Molecular Patterns |
| EA | Electroacupuncture |
| GAE | Genicular Artery Embolization |
| GFs | Growth Factors |
| HA | Hyaluronic Acid |
| HIF-1 | Activation of hypoxia-inducible factor 1 |
| IFP | Infrapatellar Fat Pad |
| IFPSCs | Infrapatellar Fat Pad-derived Stem Cells |
| IL-6 | Interleukin-6 |
| IL-1 | Interleukin-1 |
| iNOS | Inducible Nitric Oxide Synthase |
| KOA | Knee Osteoarthritis |
| LIPUS | Low-intensity pulsed ultrasound |
| MCP-1 | Monocyte chemoattractant protein-1 |
| miR100-5p | MicroRNA 100-5-p |
| MSCIPFP-Exos | Mesenchymal-derived extracellular vesicles from the infrapatellar adipose tissue |
| MMP-1 | Matrix Metalloproteinase 1 |
| MMP-3 | Matrix Metalloproteinase 3 |
| MPA | Methylprednisolone acetate |
| MRI | Magnetic Resonance |
| m-TOR | Mechanistic Target of Rapamycin |
| NF-κB | Nuclear factor kappa light chain enhancer of activated B cells |
| NLRP3 | NLR family pyrin domain containing 3 |
| NO | Nitric Oxide |
| NRS | Numeric Rating Scale |
| OA | Osteoarthritis |
| PGE2 | Prostaglandin E2 |
| PGF2α | Prostaglandin F2α |
| PGD2 | Prostaglandin D2 |
| PI3K/AKT | Phosphoinositide 3-kinase/ Protein Kinase B. |
| PPAR-γ2 | Peroxisome Proliferator-Activated Receptor Gamma 2 |
| PPTs | Pressure Pain Thresholds |
| PRP | Plateled Rich Plasma |
| RFA | Radiofrequency Ablation |
| RUNX2 | Runt-related transcription factor 2 |
| SASP | Secretory phenotype |
| SOX-9 | SRY-related HMG-box gene 9 |
| SMAD3 | Mothers against decapentaplegic homolog 3 |
| TGF-β | Transforming growth factor—β |
| TIMP | Tissue Inhibitor of Metalloproteinases |
| TNF-α | Tumor necrosis factor |
| VAS | Visual Analogue Scale |
| VCAM1 | Vascular Cell Adhesion Molecule 1 |
| VEGF | Vascular endotelial growth factor |
| WOMAC | Western Ontario and McMaster Universities Osteoarthritis Index |
References
- Steinmetz JD, Culbreth GT, Haile LM, et al. Global, regional, and national burden of osteoarthritis, 1990–2020 and projections to 2050: a systematic analysis for the Global Burden of Disease Study 2021. Lancet Rheumatol. 2023;5(9):e508-e522. [CrossRef]
- OARSI (Osteoathritis Research Society International). https://oarsi.org/research/standardization-osteoarthritis-definitions.
- Tang S, Zhang C, Oo WM, et al. Osteoarthritis. Nat Rev Dis Primers. 2025;11(1):10. [CrossRef]
- Knights AJ, Redding SJ, Maerz T. Inflammation in osteoarthritis: the latest progress and ongoing challenges. Curr Opin Rheumatol. 2023;35(2):128-134. [CrossRef]
- Goldring MB, Otero M. Inflammation in osteoarthritis. Curr Opin Rheumatol. 2011;23(5):471-478. [CrossRef]
- Chen D, Shen J, Zhao W, et al. Osteoarthritis: toward a comprehensive understanding of pathological mechanism. Bone Res. 2017;5(1):16044. [CrossRef]
- Mathiessen A, Conaghan PG. Synovitis in osteoarthritis: current understanding with therapeutic implications. Arthritis Res Ther. 2017;19(1):18. [CrossRef]
- Kloppenburg M, Namane M, Cicuttini F. Osteoarthritis. The Lancet. 2025;405(10472):71-85. [CrossRef]
- Wei Y, Bai L. Recent advances in the understanding of molecular mechanisms of cartilage degeneration, synovitis and subchondral bone changes in osteoarthritis. Connect Tissue Res. 2016;57(4):245-261. [CrossRef]
- Coaccioli S, Sarzi-Puttini P, Zis P, Rinonapoli G, Varrassi G. Osteoarthritis: New Insight on Its Pathophysiology. J Clin Med. 2022;11(20):6013. [CrossRef]
- Binvignat M, Sellam J, Berenbaum F, Felson DT. The role of obesity and adipose tissue dysfunction in osteoarthritis pain. Nat Rev Rheumatol. 2024;20(9):565-584. [CrossRef]
- Zapata-Linares N, Eymard F, Berenbaum F, Houard X. Role of adipose tissues in osteoarthritis. Curr Opin Rheumatol. 2021;33(1):84-93. [CrossRef]
- Coelho M, Oliveira T, Fernandes R. State of the art paper Biochemistry of adipose tissue: an endocrine organ. Archives of Medical Science. 2013;2:191-200. [CrossRef]
- Trayhurn P, Drevon CA, Eckel J. Secreted proteins from adipose tissue and skeletal muscle – adipokines, myokines and adipose/muscle cross-talk. Arch Physiol Biochem. 2011;117(2):47-56. [CrossRef]
- Neumann E, Junker S, Schett G, Frommer K, Müller-Ladner U. Adipokines in bone disease. Nat Rev Rheumatol. 2016;12(5):296-302. [CrossRef]
- Kanazawa I, Yamaguchi T, Yano S, Yamauchi M, Yamamoto M, Sugimoto T. Adiponectin and AMP kinase activator stimulate proliferation, differentiation, and mineralization of osteoblastic MC3T3-E1 cells. BMC Cell Biol. 2007;8(1):51. [CrossRef]
- Lin YY, Chen CY, Chuang TY, et al. Adiponectin receptor 1 regulates bone formation and osteoblast differentiation by GSK-3β/β-Catenin signaling in mice. Bone. 2014;64:147-154. [CrossRef]
- Mohammadi SM, Saniee N, Borzoo T, Radmanesh E. Osteoporosis and Leptin: A Systematic Review. Iran J Public Health. Published online January 15, 2024. [CrossRef]
- Cheleschi S, Gallo I, Barbarino M, et al. MicroRNA Mediate Visfatin and Resistin Induction of Oxidative Stress in Human Osteoarthritic Synovial Fibroblasts Via NF-κB Pathway. Int J Mol Sci. 2019;20(20):5200. [CrossRef]
- Economou A, Mallia I, Fioravanti A, et al. The Role of Adipokines between Genders in the Pathogenesis of Osteoarthritis. Int J Mol Sci. 2024;25(19):10865. [CrossRef]
- Cheleschi S, Tenti S, Barbarino M, et al. Exploring the Crosstalk between Hydrostatic Pressure and Adipokines: An In Vitro Study on Human Osteoarthritic Chondrocytes. Int J Mol Sci. 2021;22(5):2745. [CrossRef]
- Klein-Wieringa IR, Kloppenburg M, Bastiaansen-Jenniskens YM, et al. The infrapatellar fat pad of patients with osteoarthritis has an inflammatory phenotype. Ann Rheum Dis. 2011;70(5):851-857. [CrossRef]
- Zhang C, Lin Y, Yan CH, Zhang W. Adipokine Signaling Pathways in Osteoarthritis. Front Bioeng Biotechnol. 2022;10. [CrossRef]
- de Boer TN, van Spil WE, Huisman AM, et al. Serum adipokines in osteoarthritis; comparison with controls and relationship with local parameters of synovial inflammation and cartilage damage. Osteoarthritis Cartilage. 2012;20(8):846-853. [CrossRef]
- Bas S, Finckh A, Puskas GJ, et al. Adipokines correlate with pain in lower limb osteoarthritis: different associations in hip and knee. Int Orthop. 2014;38(12):2577-2583. [CrossRef]
- Tu C, He J, Wu B, Wang W, Li Z. An extensive review regarding the adipokines in the pathogenesis and progression of osteoarthritis. Cytokine. 2019;113:1-12. [CrossRef]
- Wang T, He C. Pro-inflammatory cytokines: The link between obesity and osteoarthritis. Cytokine Growth Factor Rev. 2018;44:38-50. [CrossRef]
- Clockaerts S, Bastiaansen-Jenniskens YM, Runhaar J, et al. The infrapatellar fat pad should be considered as an active osteoarthritic joint tissue: a narrative review. Osteoarthritis Cartilage. 2010;18(7):876-882. [CrossRef]
- Wang MG, Seale P, Furman D. The infrapatellar fat pad in inflammaging, knee joint health, and osteoarthritis. npj Aging. 2024;10(1):34. [CrossRef]
- Yue S, Zhai G, Zhao S, et al. The biphasic role of the infrapatellar fat pad in osteoarthritis. Biomedicine & Pharmacotherapy. 2024;179:117364. [CrossRef]
- Frasca D, Blomberg BB. Adipose tissue, immune aging, and cellular senescence. Semin Immunopathol. 2020;42(5):573-587. [CrossRef]
- Jeon OH, David N, Campisi J, Elisseeff JH. Senescent cells and osteoarthritis: a painful connection. Journal of Clinical Investigation. 2018;128(4):1229-1237. [CrossRef]
- Bolia IK, Mertz K, Faye E, et al. Cross-Communication Between Knee Osteoarthritis and Fibrosis: Molecular Pathways and Key Molecules. Open Access J Sports Med. 2022;Volume 13:1-15. [CrossRef]
- Hengtrakool P, Leearamwat N, Sengprasert P, et al. Infrapatellar fat pad adipose tissue-derived macrophages display a predominant CD11c+CD206+ phenotype and express genotypes attributable to key features of OA pathogenesis. Front Immunol. 2024;15. [CrossRef]
- Fontanella CG, Belluzzi E, Pozzuoli A, et al. Mechanical behavior of infrapatellar fat pad of patients affected by osteoarthritis. J Biomech. 2022;131:110931. [CrossRef]
- Inomata K, Tsuji K, Onuma H, et al. Time course analyses of structural changes in the infrapatellar fat pad and synovial membrane during inflammation-induced persistent pain development in rat knee joint. BMC Musculoskelet Disord. 2019;20(1):8. [CrossRef]
- Leese J, Davies DC. An investigation of the anatomy of the infrapatellar fat pad and its possible involvement in anterior pain syndrome: a cadaveric study. J Anat. 2020;237(1):20-28. [CrossRef]
- Walsh DA, Mapp PI, Kelly S. Calcitonin gene-related peptide in the joint: contributions to pain and inflammation. Br J Clin Pharmacol. 2015;80(5):965-978. [CrossRef]
- Bohnsack M, Meier F, Walter GF, et al. Distribution of substance-P nerves inside the infrapatellar fat pad and the adjacent synovial tissue: a neurohistological approach to anterior knee pain syndrome. Arch Orthop Trauma Surg. 2005;125(9):592-597. [CrossRef]
- Onuma H, Tsuji K, Hoshino T, et al. Fibrotic changes in the infrapatellar fat pad induce new vessel formation and sensory nerve fiber endings that associate prolonged pain. Journal of Orthopaedic Research. 2020;38(6):1296-1306. [CrossRef]
- Aikawa J, Uchida K, Takano S, et al. Expression of calcitonin gene-related peptide in the infrapatellar fat pad in knee osteoarthritis patients. J Orthop Surg Res. 2017;12(1):65. [CrossRef]
- Neogi T, Guermazi A, Roemer F, et al. Association of Joint Inflammation With Pain Sensitization in Knee Osteoarthritis: The Multicenter Osteoarthritis Study. Arthritis & Rheumatology. 2016;68(3):654-661. [CrossRef]
- Scanzello CR, Goldring SR. The role of synovitis in osteoarthritis pathogenesis. Bone. 2012;51(2):249-257. [CrossRef]
- Schaible HG. Nociceptive neurons detect cytokines in arthritis. Arthritis Res Ther. 2014;16(5):470. [CrossRef]
- Zou Y, Liu C, Wang Z, Li G, Xiao J. Neural and immune roles in osteoarthritis pain: Mechanisms and intervention strategies. J Orthop Translat. 2024;48:123-132. [CrossRef]
- Timur UT, Caron MMJ, Bastiaansen-Jenniskens YM, et al. Celecoxib-mediated reduction of prostanoid release in Hoffa’s fat pad from donors with cartilage pathology results in an attenuated inflammatory phenotype. Osteoarthritis Cartilage. 2018;26(5):697-706. [CrossRef]
- ter Haar G. Therapeutic ultrasound. European Journal of Ultrasound. 1999;9(1):3-9. [CrossRef]
- Kitano M, Kawahata H, Okawa Y, et al. Effects of low-intensity pulsed ultrasound on the infrapatellar fat pad in knee osteoarthritis: a randomized, double blind, placebo-controlled trial. J Phys Ther Sci. 2023;35(3):2022-2085. [CrossRef]
- Feltham T, Paudel S, Lobao M, Schon L, Zhang Z. Low-Intensity Pulsed Ultrasound Suppresses Synovial Macrophage Infiltration and Inflammation in Injured Knees in Rats. Ultrasound Med Biol. 2021;47(4):1045-1053. [CrossRef]
- Kitagawa T, Kawahata H, Kudo S. Effect of Low-Intensity Pulsed Ultrasound on Macrophage Properties and Fibrosis in the Infrapatellar Fat Pad in a Carrageenan-Induced Knee Osteoarthritis Rat Model. Cureus. Published online April 29, 2024. [CrossRef]
- He S, Fan C, Ji Y, et al. SENP3 facilitates M1 macrophage polarization via the HIF-1α/PKM2 axis in lipopolysaccharide-induced acute lung injury. Innate Immun. 2023;29(1-2):25-34. [CrossRef]
- Gouda SAA, Aboulhoda BE, Abdelwahed OM, et al. Low-intensity pulsed ultrasound (LIPUS) switched macrophage into M2 phenotype and mitigated necroptosis and increased HSP 70 in gentamicin-induced nephrotoxicity. Life Sci. 2023;314:121338. [CrossRef]
- Nakamura T, Fujihara S, Yamamoto-Nagata K, Katsura T, Inubushi T, Tanaka E. Low-Intensity Pulsed Ultrasound Reduces the Inflammatory Activity of Synovitis. Ann Biomed Eng. 2011;39(12):2964-2971. [CrossRef]
- Qin H, Luo Z, Sun Y, et al. Low-intensity pulsed ultrasound promotes skeletal muscle regeneration via modulating the inflammatory immune microenvironment. Int J Biol Sci. 2023;19(4):1123-1145. [CrossRef]
- Gurkan I, Ranganathan A, Yang X, et al. Modification of osteoarthritis in the guinea pig with pulsed low-intensity ultrasound treatment. Osteoarthritis Cartilage. 2010;18(5):724-733. [CrossRef]
- Liao B, Guan M, Tan Q, et al. Low-intensity pulsed ultrasound inhibits fibroblast-like synoviocyte proliferation and reduces synovial fibrosis by regulating Wnt/β-catenin signaling. J Orthop Translat. 2021;30:41-50. [CrossRef]
- Yi X, Wu L, Liu J, Qin Y, Li B, Zhou Q. Low-intensity pulsed ultrasound protects subchondral bone in rabbit temporomandibular joint osteoarthritis by suppressing TGF-β1/Smad3 pathway. Journal of Orthopaedic Research. 2020;38(11):2505-2512. [CrossRef]
- Tacke F. Targeting hepatic macrophages to treat liver diseases. J Hepatol. 2017;66(6):1300-1312. [CrossRef]
- Mansour SG, Puthumana J, Coca SG, Gentry M, Parikh CR. Biomarkers for the detection of renal fibrosis and prediction of renal outcomes: a systematic review. BMC Nephrol. 2017;18(1):72. [CrossRef]
- Distler JHW, Akhmetshina A, Schett G, Distler O. Monocyte chemoattractant proteins in the pathogenesis of systemic sclerosis. Rheumatology. 2009;48(2):98-103. [CrossRef]
- Yuan GH, Masuko-Hongo K, Sakata M, et al. The role of C-C chemokines and their receptors in osteoarthritis. Arthritis Rheum. 2001;44(5):1056-1070. [CrossRef]
- Miller RE, Malfait AM. Can we target CCR2 to treat osteoarthritis? The trick is in the timing! Osteoarthritis Cartilage. 2017;25(6):799-801. [CrossRef]
- Miotla Zarebska J, Chanalaris A, Driscoll C, et al. CCL2 and CCR2 regulate pain-related behaviour and early gene expression in post-traumatic murine osteoarthritis but contribute little to chondropathy. Osteoarthritis Cartilage. 2017;25(3):406-412. [CrossRef]
- Favero M, El-Hadi H, Belluzzi E, et al. Infrapatellar fat pad features in osteoarthritis: a histopathological and molecular study. Rheumatology. 2017;56(10):1784-1793. [CrossRef]
- Bastiaansen-Jenniskens YM, Wei W, Feijt C, et al. Stimulation of Fibrotic Processes by the Infrapatellar Fat Pad in Cultured Synoviocytes From Patients With Osteoarthritis: A Possible Role for Prostaglandin F 2α. Arthritis Rheum. 2013;65(8):2070-2080. [CrossRef]
- Longobardi L, Jordan JM, Shi XA, et al. Associations between the chemokine biomarker CCL2 and knee osteoarthritis outcomes: the Johnston County Osteoarthritis Project. Osteoarthritis Cartilage. 2018;26(9):1257-1261. [CrossRef]
- Morrison NA, Day CJ, Nicholson GC. Dominant Negative <scp>MCP</scp> -1 Blocks Human Osteoclast Differentiation. J Cell Biochem. 2014;115(2):303-312. [CrossRef]
- Yao Z, Keeney M, Lin T, et al. Mutant monocyte chemoattractant protein 1 protein attenuates migration of and inflammatory cytokine release by macrophages exposed to orthopedic implant wear particles. J Biomed Mater Res A. 2014;102(9):3291-3297. [CrossRef]
- Yoshimura H, Nakagawa Y, Muneta T, Koga H. A CCL2/MCP-1 antagonist attenuates fibrosis of the infrapatellar fat pad in a rat model of arthritis. BMC Musculoskelet Disord. 2024;25(1):674. [CrossRef]
- Barboza E, Hudson J, Chang W, et al. Profibrotic Infrapatellar Fat Pad Remodeling Without M1 Macrophage Polarization Precedes Knee Osteoarthritis in Mice With Diet-Induced Obesity. Arthritis & Rheumatology. 2017;69(6):1221-1232. [CrossRef]
- Iwata M, Ochi H, Hara Y, et al. Initial Responses of Articular Tissues in a Murine High-Fat Diet-Induced Osteoarthritis Model: Pivotal Role of the IPFP as a Cytokine Fountain. PLoS One. 2013;8(4):e60706. [CrossRef]
- Chang W, DeMoe J, Kent C, et al. 130 INFRAPATELLAR FAT PAD HYPERTROPHY WITHOUT INFLAMMATION IN A DIET-INDUCED MOUSE MODEL OF OBESHY AND OSTEOARTHRITIS. Osteoarthritis Cartilage. 2011;19:S66. [CrossRef]
- Han W, Cai S, Liu Z, et al. Infrapatellar fat pad in the knee: is local fat good or bad for knee osteoarthritis? Arthritis Res Ther. 2014;16(4):R145. [CrossRef]
- Chuckpaiwong B, Charles HC, Kraus VB, Guilak F, Nunley JA. Age-associated increases in the size of the infrapatellar fat pad in knee osteoarthritis as measured by 3T MRI. Journal of Orthopaedic Research. 2010;28(9):1149-1154. [CrossRef]
- Diepold J, Ruhdorfer A, Dannhauer T, Wirth W, Steidle E, Eckstein F. Sex-differences of the healthy infra-patellar (Hoffa) fat pad in relation to intermuscular and subcutaneous fat content – Data from the Osteoarthritis Initiative. Annals of Anatomy - Anatomischer Anzeiger. 2015;200:30-36. [CrossRef]
- Cortez M, Carmo LS, Rogero MM, Borelli P, Fock RA. A High-Fat Diet Increases IL-1, IL-6, and TNF-α Production by Increasing NF-κB and Attenuating PPAR-γ Expression in Bone Marrow Mesenchymal Stem Cells. Inflammation. 2013;36(2):379-386. [CrossRef]
- Radakovich LB, Marolf AJ, Culver LA, Santangelo KS. Calorie restriction with regular chow, but not a high-fat diet, delays onset of spontaneous osteoarthritis in the Hartley guinea pig model. Arthritis Res Ther. 2019;21(1):145. [CrossRef]
- Hannon J, Bardenett S, Singleton S, Garrison JC. Evaluation, Treatment, and Rehabilitation Implications of the Infrapatellar Fat Pad. Sports Health: A Multidisciplinary Approach. 2016;8(2):167-171. [CrossRef]
- Guilak F, Fermor B, Keefe FJ, et al. The Role of Biomechanics and Inflammation in Cartilage Injury and Repair. Clin Orthop Relat Res. 2004;423:17-26. [CrossRef]
- Teichtahl AJ, Wulidasari E, Brady SRE, et al. A large infrapatellar fat pad protects against knee pain and lateral tibial cartilage volume loss. Arthritis Res Ther. 2015;17(1):318. [CrossRef]
- Takahashi I, Matsuzaki T, Kuroki H, Hoso M. Disuse histological changes of an unloading environment on joint components in rat knee joints. Osteoarthr Cartil Open. 2019;1(1-2):100008. [CrossRef]
- Watanabe M, Hoso M, Hibino I, Matsuzaki T, Kojima S. Histopathological Changes of Joint Capsule after Joint Immobility Compared with Aging in Rats. J Phys Ther Sci. 2010;22(4):369-374. [CrossRef]
- Trudel G, Uhthoff HK, Goudreau L, Laneuville O. Quantitative analysis of the reversibility of knee flexion contractures with time: an experimental study using the rat model. BMC Musculoskelet Disord. 2014;15(1):338. [CrossRef]
- Takeda K, Takeshima E, Kojima S, Watanabe M, Matsuzaki T, Hoso M. Daily and short-term application of joint movement for the prevention of infrapatellar fat pad atrophy due to immobilization. J Phys Ther Sci. 2019;31(11):873-877. [CrossRef]
- Park J, Lee SW. Medical treatment of osteoarthritis: botanical pharmacologic aspect. Journal of Rheumatic Diseases. 2024;31(2):68-78. [CrossRef]
- Wang Z, Efferth T, Hua X, Zhang X an. Medicinal plants and their secondary metabolites in alleviating knee osteoarthritis: A systematic review. Phytomedicine. 2022;105:154347. [CrossRef]
- Ding Q hai, Ji X wei, Cheng Y, Yu Y quan, Qi Y ying, Wang X hua. Inhibition of matrix metalloproteinases and inducible nitric oxide synthase by andrographolide in human osteoarthritic chondrocytes. Mod Rheumatol. 2013;23(6):1124-1132. [CrossRef]
- Kulsirirat T, Honsawek S, Takeda-Morishita M, et al. The Effects of Andrographolide on the Enhancement of Chondrogenesis and Osteogenesis in Human Suprapatellar Fat Pad Derived Mesenchymal Stem Cells. Molecules. 2021;26(7):1831. [CrossRef]
- Atul Bhattaram V, Graefe U, Kohlert C, Veit M, Derendorf H. Pharmacokinetics and Bioavailability of Herbal Medicinal Products. Phytomedicine. 2002;9:1-33. [CrossRef]
- Anil U, Markus DH, Hurley ET, et al. The efficacy of intra-articular injections in the treatment of knee osteoarthritis: A network meta-analysis of randomized controlled trials. Knee. 2021;32:173-182. [CrossRef]
- Komatsu S, Iwata H, Nabeshima T. Studies on the Kinetics, Metabolism and Re-utilisation after Intra-articular Administration of Hyaluronan to Rabbits. Arzneimittelforschung. 2011;49(05):427-433. [CrossRef]
- Lee CL, Wang YC, Huang HT, Chen CH, Chang KL, Tien YC. Efficacy of Intra-Articular Injection of Biofermentation-Derived High-Molecular Hyaluronic Acid in Knee Osteoarthritis: An Ultrasonographic Study. Cartilage. 2022;13(1). [CrossRef]
- Qu Z, Koga H, Tsuji K, et al. Hyaluronic acid sheet transplantation attenuates infrapatellar fat pad fibrosis and pain in a rat arthritis model. Journal of Orthopaedic Research. 2023;41(11):2442-2454. [CrossRef]
- Chen WH, Lin CM, Huang CF, et al. Functional Recovery in Osteoarthritic Chondrocytes Through Hyaluronic Acid and Platelet-Rich Plasma–Inhibited Infrapatellar Fat Pad Adipocytes. Am J Sports Med. 2016;44(10):2696-2705. [CrossRef]
- Filardo G, Kon E, Buda R, et al. Platelet-rich plasma intra-articular knee injections for the treatment of degenerative cartilage lesions and osteoarthritis. Knee Surgery, Sports Traumatology, Arthroscopy. 2011;19(4):528-535. [CrossRef]
- Sundman EA, Cole BJ, Karas V, et al. The Anti-inflammatory and Matrix Restorative Mechanisms of Platelet-Rich Plasma in Osteoarthritis. Am J Sports Med. 2014;42(1):35-41. [CrossRef]
- Sundman EA, Cole BJ, Karas V, et al. The Anti-inflammatory and Matrix Restorative Mechanisms of Platelet-Rich Plasma in Osteoarthritis. Am J Sports Med. 2014;42(1):35-41. [CrossRef]
- Araya N, Miyatake K, Tsuji K, et al. Intra-articular Injection of Pure Platelet-Rich Plasma Is the Most Effective Treatment for Joint Pain by Modulating Synovial Inflammation and Calcitonin Gene-Related Peptide Expression in a Rat Arthritis Model. Am J Sports Med. 2020;48(8):2004-2012. [CrossRef]
- Burchard R, Huflage H, Soost C, Richter O, Bouillon B, Graw JA. Efficiency of platelet-rich plasma therapy in knee osteoarthritis does not depend on level of cartilage damage. J Orthop Surg Res. 2019;14(1):153. [CrossRef]
- Xu J, Chen X, Zhang H, et al. Platelet-rich plasma relieves inflammation and pain by regulating M1/M2 macrophage polarization in knee osteoarthritis rats. Sci Rep. 2025;15(1):12805. [CrossRef]
- Yan X, Ye Y, Wang L, Xue J, Shen N, Li T. Platelet-rich plasma alleviates neuropathic pain in osteoarthritis by downregulating microglial activation. BMC Musculoskelet Disord. 2024;25(1):331. [CrossRef]
- Xie Y, Tolmeijer S, Oskam JM, Tonkens T, Meijer AH, Schaaf MJM. Glucocorticoids inhibit macrophage differentiation towards a pro-inflammatory phenotype upon wounding without affecting their migration. Dis Model Mech. 2019;12(5). [CrossRef]
- Lee MJ, Pramyothin P, Karastergiou K, Fried SK. Deconstructing the roles of glucocorticoids in adipose tissue biology and the development of central obesity. Biochimica et Biophysica Acta (BBA) - Molecular Basis of Disease. 2014;1842(3):473-481. [CrossRef]
- Jørgensen TS, Graven-Nielsen T, Ellegaard K, Danneskiold-Samsøe B, Bliddal H, Henriksen M. Intra-Articular Analgesia and Steroid Reduce Pain Sensitivity in Knee OA Patients: An Interventional Cohort Study. Pain Res Treat. 2014;2014:1-6. [CrossRef]
- Bensa A, Salerno M, Boffa A, et al. Corticosteroid injections for the treatment of osteoarthritis present a wide spectrum of effects ranging from detrimental to disease-modifying: A systematic review of preclinical evidence by the ESSKA Orthobiologic Initiative. Knee Surgery, Sports Traumatology, Arthroscopy. 2024;32(11):2725-2745. [CrossRef]
- Wen FQ, Kohyama T, Sköld CM, et al. Glucocorticoids Modulate TGF-β Production by Human Fetal Lung Fibroblasts. Inflammation. 2003;27(1):9-19. [CrossRef]
- Wen FQ, Kohyama T, Sköld CM, et al. Glucocorticoids Modulate TGF-β Production. Inflammation. 2002;26(6):279-290. [CrossRef]
- Lopes EBP, Filiberti A, Husain SA, Humphrey MB. Immune Contributions to Osteoarthritis. Curr Osteoporos Rep. 2017;15(6):593-600. [CrossRef]
- Heard BJ, Solbak NM, Chung M, et al. The infrapatellar fat pad is affected by injury induced inflammation in the rabbit knee: use of dexamethasone to mitigate damage. Inflammation Research. 2016;65(6):459-470. [CrossRef]
- Barton KI, Chung M, Frank CB, Shrive NG, Hart DA. Methylprednisolone acetate mitigates IL1β induced changes in matrix metalloproteinase gene expression in skeletally immature ovine explant knee tissues. Inflammation Research. 2021;70(1):99-107. [CrossRef]
- Zhang Y, Ruan G, Zheng P, et al. Efficacy and safety of GLucocorticoid injections into InfrapaTellar faT pad in patients with knee ostEoarthRitiS: a randomized clinical trial. Osteoarthritis Cartilage. 2025;33:S223. [CrossRef]
- Soeki T, Kishimoto I, Okumura H, et al. C-type natriuretic peptide, a novel antifibrotic and antihypertrophic agent, prevents cardiac remodeling after myocardial infarction. J Am Coll Cardiol. 2005;45(4):608-616. [CrossRef]
- Kimura T, Nojiri T, Hino J, et al. C-type natriuretic peptide ameliorates pulmonary fibrosis by acting on lung fibroblasts in mice. Respir Res. 2016;17(1):19. [CrossRef]
- An JS, Tsuji K, Onuma H, et al. Inhibition of fibrotic changes in infrapatellar fat pad alleviates persistent pain and articular cartilage degeneration in monoiodoacetic acid-induced rat arthritis model. Osteoarthritis Cartilage. 2021;29(3):380-388. [CrossRef]
- Gupta N, Khatri K, Lakhani A, et al. Long-term effectiveness of intra-articular injectables in patients with knee osteoarthritis: a systematic review and Bayesian network meta-analysis. J Orthop Surg Res. 2025;20(1):227. [CrossRef]
- José F.S.D. Lana AWSESEFVSCHCVSMAAHVAUPCMKOJMABMHASWDB. Randomized controlled trial comparing hyaluronic acid, platelet-rich plasma and the combination of both in the treatment of mild and moderate osteoarthritis of the knee. J Stem Cells Regen Med. 2016;12(2):69-78. [CrossRef]
- Anil U, Markus DH, Hurley ET, et al. The efficacy of intra-articular injections in the treatment of knee osteoarthritis: A network meta-analysis of randomized controlled trials. Knee. 2021;32:173-182. [CrossRef]
- He W wei, Kuang M jie, Zhao J, et al. Efficacy and safety of intraarticular hyaluronic acid and corticosteroid for knee osteoarthritis: A meta-analysis. International Journal of Surgery. 2017;39:95-103. [CrossRef]
- Qiao X, Yan L, Feng Y, et al. Efficacy and safety of corticosteroids, hyaluronic acid, and PRP and combination therapy for knee osteoarthritis: a systematic review and network meta-analysis. BMC Musculoskelet Disord. 2023;24(1):926. [CrossRef]
- Jones A, Doherty M. Intra-articular corticosteroids are effective in osteoarthritis but there are no clinical predictors of response. Ann Rheum Dis. 1996;55(11):829-832. [CrossRef]
- Lo GH, LaValley M, McAlindon T, Felson DT. Intra-articular Hyaluronic Acid in Treatment of Knee Osteoarthritis. JAMA. 2003;290(23):3115. [CrossRef]
- Jameson C. Autologous Platelet Concentrate for the Production of Platelet Gel. Lab Med. 2007;38(1):39-42. [CrossRef]
- Liu CY, Tu JF, Lee MS, et al. Is acupuncture effective for knee osteoarthritis? A protocol for a systematic review and meta-analysis. BMJ Open. 2022;12(1):e052270. [CrossRef]
- Zhang W, Zhang L, Yang S, Wen B, Chen J, Chang J. Electroacupuncture ameliorates knee osteoarthritis in rats via inhibiting NLRP3 inflammasome and reducing pyroptosis. Mol Pain. 2023;19. [CrossRef]
- Nemschak G, Pretterklieber ML. The Patellar Arterial Supply via the Infrapatellar Fat Pad (of Hoffa): A Combined Anatomical and Angiographical Analysis. Anat Res Int. 2012;2012:1-10. [CrossRef]
- Kohn D, Deiler S, Rudert M. Arterial blood supply of the infrapatellar fat pad. Arch Orthop Trauma Surg. 1995;114(2):72-75. [CrossRef]
- Taslakian B, Miller LE, Mabud TS, et al. Genicular artery embolization for treatment of knee osteoarthritis pain: Systematic review and meta-analysis. Osteoarthr Cartil Open. 2023;5(2):100342. [CrossRef]
- O’Brien S, Blakeney WG, Soares J. Selective Genicular Artery Embolization in the Management of Osteoarthritic Knee Pain—A Narrative Review. J Clin Med. 2024;13(11):3256. [CrossRef]
- Abussa R, Jeremic A. Bridging the Gap between Injections and Surgery: Meta-Analysis of Genicular Artery Embolization in Knee Osteoarthritis. Acad Radiol. Published online May 2025. [CrossRef]
- Ashraf N, Tahir MJ, Arabi TZ, et al. Safety and Efficacy of Genicular Artery Embolization for Knee Joint Osteoarthritis Associated Pain: A Systematic Review. Journal of Endovascular Therapy. Published online April 11, 2025. [CrossRef]
- Choi WJ, Hwang SJ, Song JG, et al. Radiofrequency treatment relieves chronic knee osteoarthritis pain: A double-blind randomized controlled trial. Pain. 2011;152(3):481-487. [CrossRef]
- Fonkoué L, Behets C, Kouassi JÉK, et al. Distribution of sensory nerves supplying the knee joint capsule and implications for genicular blockade and radiofrequency ablation: an anatomical study. Surgical and Radiologic Anatomy. 2019;41(12):1461-1471. [CrossRef]
- Chou SH, Shen PC, Lu CC, et al. Comparison of Efficacy among Three Radiofrequency Ablation Techniques for Treating Knee Osteoarthritis: A Systematic Review and Meta-Analysis. Int J Environ Res Public Health. 2021;18(14):7424. [CrossRef]
- Hong T, Wang H, Li G, Yao P, Ding Y. Systematic Review and Meta-Analysis of 12 Randomized Controlled Trials Evaluating the Efficacy of Invasive Radiofrequency Treatment for Knee Pain and Function. Biomed Res Int. 2019;2019:1-14. [CrossRef]
- Karaman H, Tüfek A, Kavak GÖ, et al. Intra-articularly applied pulsed radiofrequency can reduce chronic knee pain in patients with osteoarthritis. Journal of the Chinese Medical Association. 2011;74(8):336-340. [CrossRef]
- Menzies RD, Hawkins JK. Analgesia and Improved Performance in a Patient Treated by Cooled Radiofrequency for Pain and Dysfunction Postbilateral Total Knee Replacement. Pain Practice. 2015;15(6). [CrossRef]
- Soetjahjo B, Adriansyah D, Yudistira MB, Rahman AN, Herman H, Diwan S. Systematic Review The Analgesic Effectiveness of Genicular Nerve-targeted Cooled and Pulsed Radiofrequency Ablation for Osteoarthritis Knee Pain: A Systematic Review and Meta-analysis. www.painphysicianjournal.com.
- Stephen JM, Sopher R, Tullie S, Amis AA, Ball S, Williams A. The infrapatellar fat pad is a dynamic and mobile structure, which deforms during knee motion, and has proximal extensions which wrap around the patella. Knee Surgery, Sports Traumatology, Arthroscopy. 2018;26(11):3515-3524. [CrossRef]
- Ali MM, Phillips SA, Mahmoud AM. HIF1α/TET1 Pathway Mediates Hypoxia-Induced Adipocytokine Promoter Hypomethylation in Human Adipocytes. Cells. 2020;9(1):134. [CrossRef]
- Macchi V, Porzionato A, Sarasin G, et al. The Infrapatellar Adipose Body: A Histotopographic Study. Cells Tissues Organs. 2016;201(3):220-231. [CrossRef]
- Liu Y, Gao Q. Partial excision of infrapatellar fat pad for the treatment of knee osteoarthritis. J Orthop Surg Res. 2024;19(1):631. [CrossRef]
- Afzali MF, Radakovich LB, Sykes MM, et al. Early removal of the infrapatellar fat pad/synovium complex beneficially alters the pathogenesis of moderate stage idiopathic knee osteoarthritis in male Dunkin Hartley guinea pigs. Arthritis Res Ther. 2022;24(1):282. [CrossRef]
- Li Y, Lu P, Yao H, et al. Observation of the Effects of Infrapatellar Fat Pad Excision on the Inflammatory Progression of Knee Osteoarthritis in Mice. J Inflamm Res. 2025;Volume 18:6653-6672. [CrossRef]
- Yao B, Samuel LT, Acuña AJ, et al. Infrapatellar Fat Pad Resection or Preservation during Total Knee Arthroplasty: A Systematic Review. J Knee Surg. 2021;34(04):415-421. [CrossRef]
- Rajbhandari A, Banskota B, Bhusal R, Banskota AK. Effect of Infrapatellar Fat Pad Preservation vs Resection on Clinical Outcomes After Total Knee Arthroplasty in Patient with End-Stage Osteoarthritis. Indian J Orthop. 2023;57(6):863-867. [CrossRef]
- Caplan AI, Correa D. The MSC: An Injury Drugstore. Cell Stem Cell. 2011;9(1):11-15. [CrossRef]
- Wickham MQ, Erickson GR, Gimble JM, Vail TP, Guilak F. Multipotent Stromal Cells Derived From the Infrapatellar Fat Pad of the Knee. Clin Orthop Relat Res. 2003;412:196-212. [CrossRef]
- Sriwatananukulkit O, Tawonsawatruk T, Rattanapinyopituk K, Luangwattanawilai T, Srikaew N, Hemstapat R. Scaffold-Free Cartilage Construct from Infrapatellar Fat Pad Stem Cells for Cartilage Restoration. Tissue Eng Part A. 2022;28(5-6):199-211. [CrossRef]
- Sun Y, Chen S, Pei M. Comparative advantages of infrapatellar fat pad: an emerging stem cell source for regenerative medicine. Rheumatology. 2018;57(12):2072-2086. [CrossRef]
- Ding DC, Wu KC, Chou HL, Hung WT, Liu HW, Chu TY. Human Infrapatellar Fat Pad-Derived Stromal Cells have more Potent Differentiation Capacity than other Mesenchymal Cells and can be Enhanced by Hyaluronan. Cell Transplant. 2015;24(7):1221-1232. [CrossRef]
- Wu J, Kuang L, Chen C, et al. miR-100-5p-abundant exosomes derived from infrapatellar fat pad MSCs protect articular cartilage and ameliorate gait abnormalities via inhibition of mTOR in osteoarthritis. Biomaterials. 2019;206:87-100. [CrossRef]
- Jeon Y, Kim J, Cho JH, Chung H, Chae J. Comparative Analysis of Human Mesenchymal Stem Cells Derived From Bone Marrow, Placenta, and Adipose Tissue as Sources of Cell Therapy. J Cell Biochem. 2016;117(5):1112-1125. [CrossRef]
- Kouidhi M, Villageois P, Mounier CM, et al. Characterization of Human Knee and Chin Adipose-Derived Stromal Cells. Stem Cells Int. 2015;2015:1-11. [CrossRef]
- Koh YG, Choi YJ. Infrapatellar fat pad-derived mesenchymal stem cell therapy for knee osteoarthritis. Knee. 2012;19(6):902-907. [CrossRef]
- Pei M. Environmental preconditioning rejuvenates adult stem cells’ proliferation and chondrogenic potential. Biomaterials. 2017;117:10-23. [CrossRef]
- Meng H, Lu V, Khan W. Adipose Tissue-Derived Mesenchymal Stem Cells as a Potential Restorative Treatment for Cartilage Defects: A PRISMA Review and Meta-Analysis. Pharmaceuticals. 2021;14(12):1280. [CrossRef]
- Huang S, Song X, Li T, et al. Pellet coculture of osteoarthritic chondrocytes and infrapatellar fat pad-derived mesenchymal stem cells with chitosan/hyaluronic acid nanoparticles promotes chondrogenic differentiation. Stem Cell Res Ther. 2017;8(1):264. [CrossRef]
- Ahearne M, Liu Y, Kelly DJ. Combining Freshly Isolated Chondroprogenitor Cells from the Infrapatellar Fat Pad with a Growth Factor Delivery Hydrogel as a Putative Single Stage Therapy for Articular Cartilage Repair. Tissue Eng Part A. 2014;20(5-6):930-939. [CrossRef]
- Wang Y, Pei YA, Sun Y, Zhou S, Zhang XB, Pei M. Stem cells immortalized by hTERT perform differently from those immortalized by SV40LT in proliferation, differentiation, and reconstruction of matrix microenvironment. Acta Biomater. 2021;136:184-198. [CrossRef]
- Wu J, Wu J, Xiang W, et al. Engineering exosomes derived from TNF-α preconditioned IPFP-MSCs enhance both yield and therapeutic efficacy for osteoarthritis. J Nanobiotechnology. 2024;22(1):555. [CrossRef]
- Aikawa J, Uchida K, Takano S, et al. Regulation of calcitonin gene-related peptide expression through the COX-2/mPGES-1/PGE2 pathway in the infrapatellar fat pad in knee osteoarthritis. Lipids Health Dis. 2018;17(1):215. [CrossRef]
Figure 1.
A midsagittal section of the knee joint illustrates the anatomical position of the IFP, along with the other smaller adipose structures: the posterior fat pad, the anterior suprapatellar fat pad, and the posterior suprapatellar fat pad. The IFP is located inferior to the patella and posterior to the patellar tendon. Anteriorly, it is enclosed by the joint capsule, while its articular-facing surface is lined by synovial membrane. As such, the IFP is included within the joint capsule (intracapsular) but outside the synovial cavity (extrasynovial). Additionally, it lies in close proximity to the articular cartilage surfaces, highlighting its potential functional significance in joint biomechanics. Figure created with the assistance of artificial intelligence and scientifically validated by the authors.
Figure 1.
A midsagittal section of the knee joint illustrates the anatomical position of the IFP, along with the other smaller adipose structures: the posterior fat pad, the anterior suprapatellar fat pad, and the posterior suprapatellar fat pad. The IFP is located inferior to the patella and posterior to the patellar tendon. Anteriorly, it is enclosed by the joint capsule, while its articular-facing surface is lined by synovial membrane. As such, the IFP is included within the joint capsule (intracapsular) but outside the synovial cavity (extrasynovial). Additionally, it lies in close proximity to the articular cartilage surfaces, highlighting its potential functional significance in joint biomechanics. Figure created with the assistance of artificial intelligence and scientifically validated by the authors.
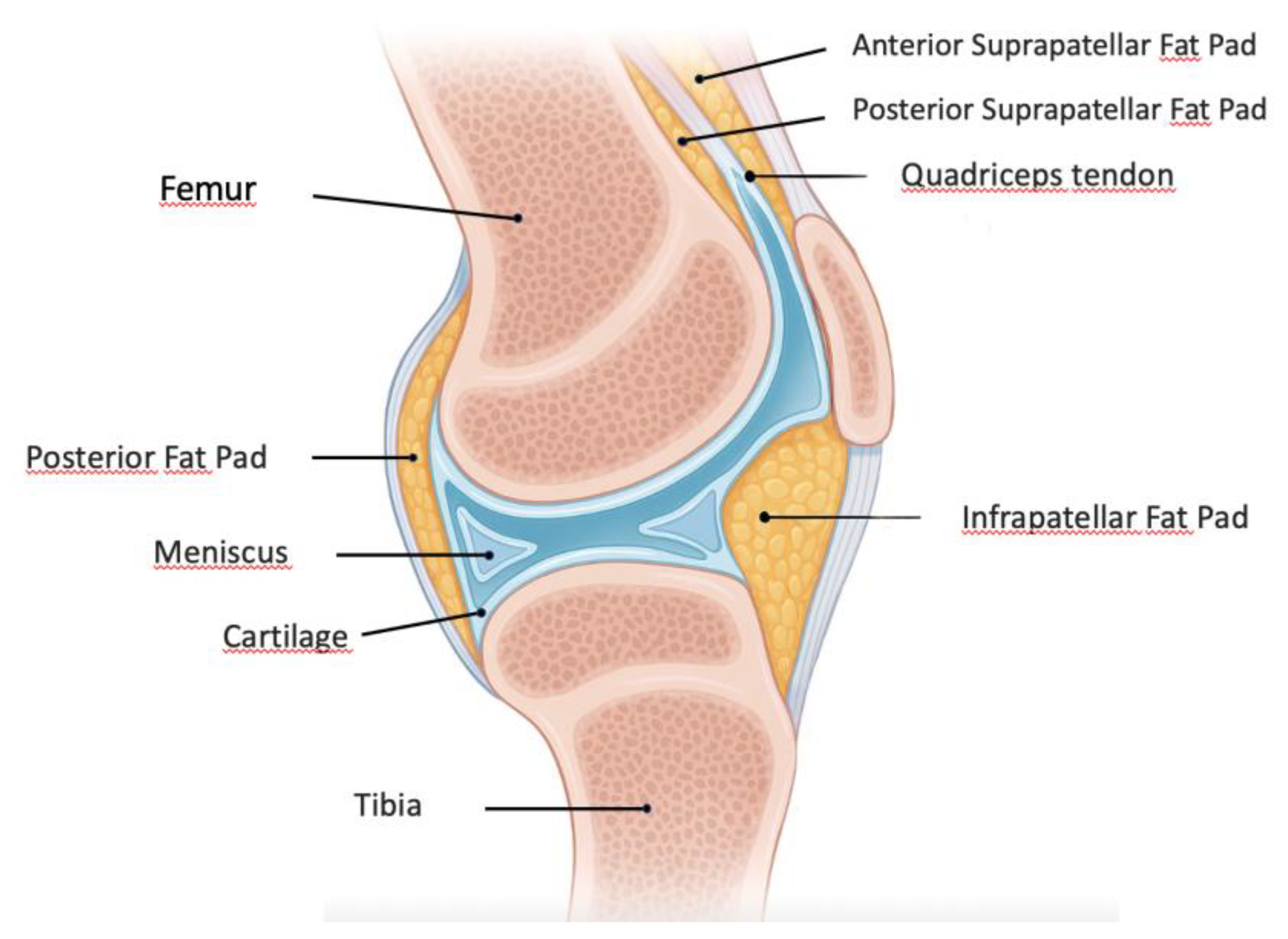
Figure 2.
Inflammatory signaling pathway between the IFP and articular cartilage. The IFP contains adipocytes, macrophages, and T cells within a highly vascularized environment. Upon activation, these cellular components release pro-inflammatory cytokines including TNF-α, IL-6, VEGF, and PGE2. These inflammatory mediators target the articular cartilage, potentially affecting chondrocyte metabolism and contributing to cartilage degradation. The proximity of the IFP to the cartilage-subchondral bone interface facilitates this paracrine signaling mechanism in knee joint pathophysiology. Figure created with the assistance of artificial intelligence and scientifically validated by the authors.
Figure 2.
Inflammatory signaling pathway between the IFP and articular cartilage. The IFP contains adipocytes, macrophages, and T cells within a highly vascularized environment. Upon activation, these cellular components release pro-inflammatory cytokines including TNF-α, IL-6, VEGF, and PGE2. These inflammatory mediators target the articular cartilage, potentially affecting chondrocyte metabolism and contributing to cartilage degradation. The proximity of the IFP to the cartilage-subchondral bone interface facilitates this paracrine signaling mechanism in knee joint pathophysiology. Figure created with the assistance of artificial intelligence and scientifically validated by the authors.
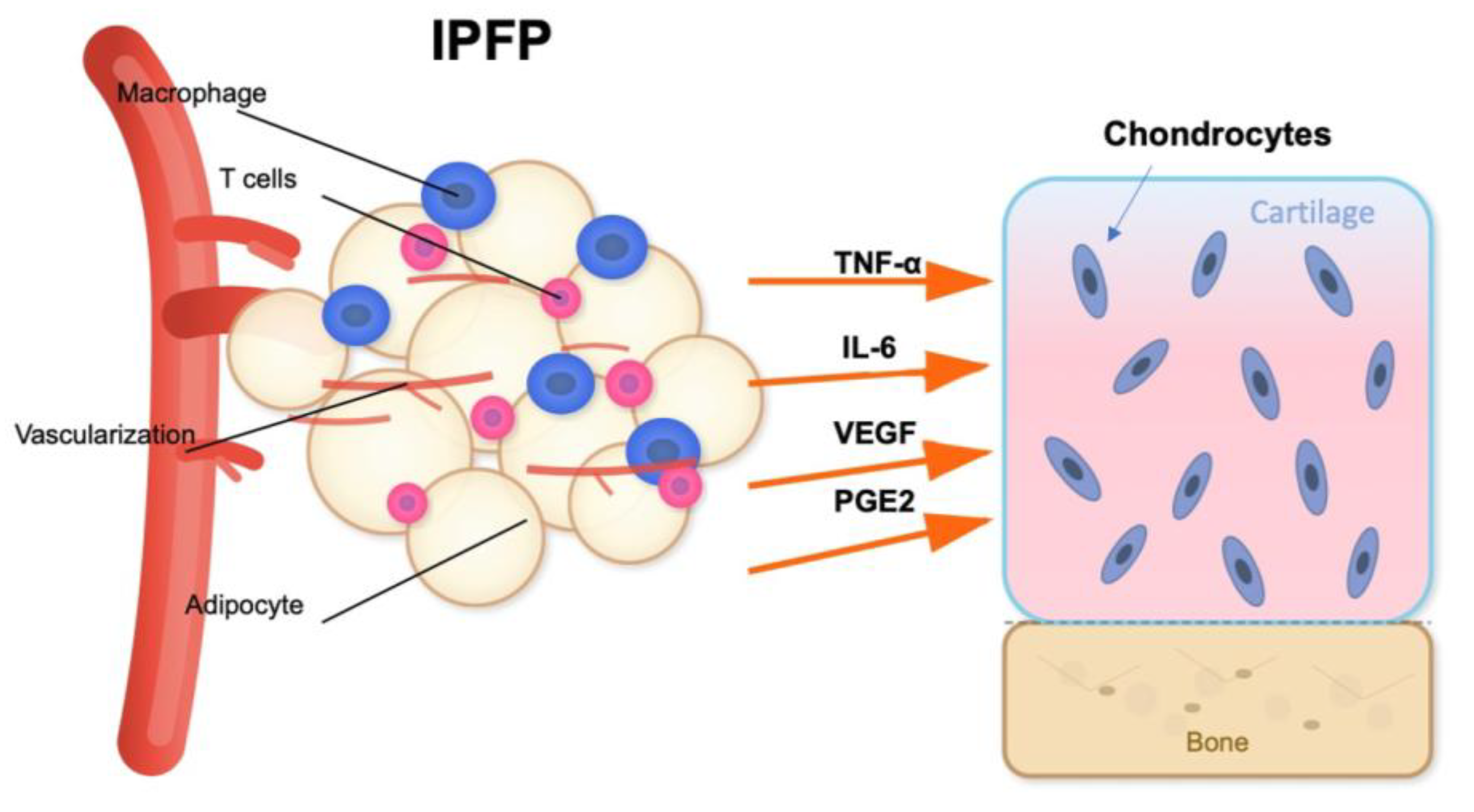
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



