Submitted:
17 September 2025
Posted:
22 September 2025
You are already at the latest version
Abstract
Alkaloids are among the most important secondary metabolites, and the discovery of their biosynthetic pathways is a key focus for pharmacological applications. Nowadays, these discoveries rely heavily on data from genomes, transcriptomes, and metabolomes. Since the 2000s, RNA sequencing has become a common approach for uncovering metabolic pathways by correlating plant transcript expression with metabolomic profiling. While the biosynthesis routes for isoquinoline alkaloids (IsAs) are well-documented in the literature, those for Amaryllidaceae alkaloids (AmAs) remain poorly described. In this review, we compile the available information on the genomic and transcriptomic approaches used in plants producing IsAs and AmAs and explore how the similarities between these two groups contribute to understanding the current challenges in elucidating AmAs pathways. Additionally, we compare historical approaches to discovering IsAs and AmAs pathways by analyzing key representatives—lycorine and galanthamine (AmAs) and morphine (IsAs)—while highlighting the role of various enzyme families involved in the synthesis of these alkaloids.
Keywords:
isoquinoline alkaloids
; Amaryllidaceae alkaloids
; transcriptomes
; RNA sequencing
; functional genes
; secondary metabolism
1. Introduction
Secondary plant metabolites (SPMs) are chemical phytocompounds that enable plants to compete in their environment. These molecules are of immense interest due to their high diversity [1] and industrial applications. They are produced by plant cells through metabolic pathways derived from primary metabolic processes, such as nitrogen metabolism, through secondary modifications, including deaminations, decarboxylations, and hydroxylations, among others [2,3]. On the basis of their chemical structure, SPMs can be classified as phenolics, alkaloids, saponins, terpenes, lipids, and glycosylated carbohydrates [3]. Among these, alkaloids are a diverse group of organic compounds often referred to as plant defense resources against potential predators [4], although many can be produced by different taxa such as fungi, bacteria, and animals. Alkaloids contain at least one nitrogen atom and can be classified according to the origin and position of this atom in the molecule as: 1) true alkaloids, which contain nitrogen atoms derived from amino acids, 2) protoalkaloids, which are alkaloid precursors that lack some structural features of true alkaloids, such as the presence of a quaternary nitrogen atom, and 3) pseudoalkaloids, which are compounds that resemble alkaloids in some ways but do not directly derive nitrogen atoms from amino acids but do so through mechanisms such as transamination reactions [5]. They can also be classified according to a defined chemical-related skeleton (Figure 1) [1] and are of great interest as plant natural products (PNPs) due to their biological activities, which include but are not limited to, antiviral, antibacterial, anti-inflammatory, and antineoplastic properties, as well as herbicidal, insecticidal and fungicidal [6,7,8].
Isoquinoline alkaloids are a structural class present in at least 28 plant families, distributed across 17 orders, with families belonging to Ranunculales order the most diverse producers [9]. The Ranunculales Isoquinoline alkaloids (IsAs) represent the largest group of well-documented alkaloids derived from benzopyridines synthesized from L-phenylalanine and/or L-tyrosine [10]. The synthesis involves the condensation of tyramine and L-DOPA to form dopamine, a substituted phenethylamine that is central to many biochemical pathways and produces a vast diversity of compounds [11]. In the IsAs pathway, dopamine is condensed to form another central intermediate known as norcoclaurine, which has a nitrogen heterocycle and two phenolic rings [12]. Several reactions involving methyltransferases and oxidoreductases then occur to produce reticuline, an important molecule that can undergo intramolecular phenol-couplings. Specifically, the para-ortho coupling leads to the most well-documented route from this family: the morphine pathway [13] (Figure 2).
On the other hand, the Amaryllidaceae alkaloids (AmAs) are a class of compounds restricted mainly to the Amaryllidaceae family, specifically Amaryllidoideae subfamily, but also present in Aspargaceae, the sister family within the same order Asparagales [14] (Figure 2). Despite IsAs being present in the Amarillydacea family, the metabolic pathway differs from what happens in other families. These alkaloids are derived from the amino acids L-phenylalanine and L-tyrosine. L-phenylalanine undergoes some reactions from the phenylpropanoid pathway leading to 3,4-dihydroxybenzaldehyde (3,4-DHBA), while tyramine is produced from L-tyrosine [15]. These two compounds undergo a condensation reaction to produce the imine norcraugsodine, followed by a reduction to produce norbelladine [16,17] and a methylation leading to 4'-O-methylnorbelladine (4OMET), a protoalkaloid of two phenolic rings connected by a nitrogen atom [17,18]. From this point, 4OMET can undergo three different types of intramolecular phenol-coupling reactions, para-para, para-ortho, or ortho-para couplings, producing compounds that can incorporate benzopyridine structures, such as the alkaloid haemanthamine. While similar phenol-coupling reactions, para-ortho, ortho-para, or ortho-ortho, occur in reticuline as part of the IsAs pathway, they are catalyzed by distinct enzymes. All couplings are mediated by a family of cytochrome P450-dependent oxidase enzymes known as CYPs. Galanthamine is one of the major examples of para-ortho phenolic coupling derivatives and is the only alkaloid approved by the FDA for the treatment of neurodegenerative diseases such as Alzheimer's disease due to its anticholinesterase inhibitory activity [19,20]. Lycorine, an alkaloid derived from ortho-para-phenol coupling, has shown cytotoxic and antitumor properties [21], while haemanthamine, crinine and montanine are some alkaloids resulting from the para-para phenol coupling of 4OMET (Figure 2), which have displayed anxiolytic, antidepressant and anticonvulsant activities, along with immunomodulatory properties. Over the past five years, researchers have described new alkaloids derived from norbelladine, as opposed to the previously mentioned 4OMET-derived alkaloids. These new alkaloids are known as Cherylline-type and Norbelladine-type, and their biosynthetic pathway remains fully unknown [22,23,24]. Figure 3 shows a comparation between IsAs and AmAs metabolic pathway.
Although AmAs have been extensively extracted and described from many plants, the molecular understanding of their synthesis remains incomplete due to the lack of genomic sequencing information for the plants in the Amaryllidaceae family that produce them, and complete transcriptomes are limited to only a few genera within them[24]. Furthermore, metabolomic studies have shown that certain alkaloids can be produced by specific species within a genus. For example, all Phaedranassa herbs have been reported to produce galanthamine and lycorine-type alkaloids, with the latter being the predominant one in most cases, except for P. cinerea which is the only species that does not appear to produce lycorine-type alkaloids [25]. Several omics approaches have been explored to understand how alkaloids are produced. Genomics, transcriptomics, and metabolomics are the main tools used to elucidate the molecular mechanisms and pathways involved in alkaloid production. However, genomic DNA sequencing has remained elusive because of its complexity and size. Instead, transcriptomics is currently the most widely researched and applied approach, as it allows the study of functional and differentially expressed genes (DEGs) and it is often used to complement metabolomic profiling of plants under specific conditions. Most of the research in this field focuses on gaining a differential understanding of plant tissues, elicitors, and abiotic factors [26].
While the biosynthetic pathways of isoquinoline alkaloids (IsAs) and Amaryllidaceae alkaloids (AmAs) share several common elements, such as the use of similar precursors and the involvement of key enzymes like methyltransferases and oxidoreductases, they also exhibit significant differences in taxonomic distribution, metabolite structure, and the current state of characterization. Table 1 synthesizes these similarities and differences, highlighting opportunities for biotechnological development in both groups. With this in mind, in this review, we seek to identify the challenges in elucidating AmAs biosynthetic pathways by contrasting them with the advances made in elucidating IsAs alkaloid metabolic pathways over the past 10 years. Specifically, we summarize the available studies on plants producing AmAs and IsAs, detailing the technologies employed and the enzymes identified in each case. Furthermore, we discuss how this accumulated information has been utilized to reconstruct metabolic pathways for specific alkaloids in both groups
2. Historical Landmarks in IsAs and AmAs Pathway Elucidation
The elucidation of alkaloid biosynthetic pathways has a long history that predates modern omics technologies. Morphine, for example, has been recognized for over two hundred years since its isolation in 1805 by Friedrich Wilhelm Adam Serturner, who referred to it as "Principium somniferum" [27]. The biosynthesis pathway of morphine was reviewed in 1967, proposing major steps from tyrosine to morphine via intermediates including norlaudanosoline, reticuline, salutaridine, salutaridinol-1, thebaine, and codeine, employing tracer experiments validated through analogous chemical transformations [28]. This suggests an understanding of at least five out of the twelve currently known intermediates leading directly from norlaudanosoline to morphine by 1967. In the case of pathway IsAs, the initial proposal in 1981 was for norlaudanosoline as the first natural intermediate, with 3,4-dihydroxyphenylacetaldehyde (3,4-DHPA) as the precursor, determined using 3H-radiolabeled dopamine as substrate and cell-free systems as enzyme sources [29]. However, subsequent tracer experiments with 13C-radiolabeled demonstrated that the enzyme does not produce norlaudanosoline in plants. Instead, it produces norcoclaurine from 4-hydroxyphenylacetaldehyde, as a result, the enzyme was renamed norcoclaurine/norlaudanosoline synthase (NCS) in 1987 [30]. Besides radiolabeling, studies related to gene expression were performed as tools for pathway elucidation, in situ RNA hybridization was used to establish differential and organ-dependent TYDC accumulation, suggesting a coordinated regulation of specific alkaloid biosynthetic genes [31].
In 2001, the biotechnological production of morphinan alkaloids started to be a goal by the hand of recombinant DNA for heterologous expression and enzyme purification, which allowed the characterization of enzymes such as 7-O-Acetyltransferase [32]. Later, immunofluorescence labeling, and shotgun proteomics were used to establish that morphine biosynthesis occurs in two specific cell types [13], by 2015 a genetic locus designated STORR ((S)- to (R)-reticuline) responsible for encoding cytochrome P450-oxydoreductase fusion protein, was sequenced and microbial-based morphinan production was practically a reality [33]. In 2019, a new enzyme responsible for catalyzing a reaction previously assumed as spontaneous was identified; the isomerization of neopinone to codeinone [34]. To date, several synthetic systems have been engineered to produce morphine [35,36] and even feedback inhibition in those systems has started to be studied to improve production [37]. These findings underline the ever-evolving nature of alkaloid pathway research and the need for ongoing investigation and reassessment of our understanding.
AmAs have been known since 1878 when the first alkaloid of its kind, lycorine, was isolated from Narcissus pseudonarcissus by A. W. Gerrard [38] during investigations into the poisonous properties of the Amaryllidaceae plants. Further studies on lycorine were carried out in Lycoris species, with initial reports in 1897, followed by the extraction and description of other alkaloids in the 1920s. Early studies aimed to understand the carbon origins of AmAs using radiolabeling experiments, which led to a partial understanding of the phenylpropanoid pathway that produces 3,4-DHBA and clarified that all AmAs are derived from norbelladine [39], which is derived from the amino acid precursors phenylalanine and tyrosine. Radiolabeling was also used to understand galanthamine biosynthesis by application of 13C-labelled 4′-O-methylnorbelladine to different organs of Leucojum aestivum [40]. Despite knowledge of the main steps on the biosynthetic pathway for AmAs [41], it remained largely unexplored until the last decade, with the discovery of norbelladine-4'-O-methyltransferase (N4OMT) as the first specific enzyme in the pathway responsible for the methylation of norbelladine, leading to the central intermediate 4OMET, which can undergo intramolecular phenol couplings [42]. By 2022, only three additional native enzymes have been described in Amaryllidaceae: norbelladine synthase (NBS) [43] and norcraugsodine/noroxomaritidine reductase (NR) [44], both of which are active prior to the phenol coupling reaction; and noroxomaritidine synthase (CYP96T1), the enzyme responsible for the para-para phenol coupling reaction [45]. In 2024, the remaining phenol-coupling enzymes: Demethylnarwedine synthase (CYP96T6) and Noroxopluviine synthase (CYP96T5), as well as a short-chain alcohol dehydrogenase/reductase (SDR1), a cinnamoyl-CoA reductase-type (SDR2), a CYP71-family protein (CYP71DW1), a 2-oxoglutarate dependent dioxygenase (ODD2), a 11-hydroxyvittatine O-methyltransferase (OMT1), a gamma-tocopherol methyltransferase (NMT1), and an aldo-keto reductase (AKR1) have been described, reporting the galanthamine and haemantamine biosynthetic pathways [46]. In the same year, the first monocot enzymes capable of catalyzing the three regioselective phenol couplings, CYP96T1 and CYP96T2 from Leucojum aestivum was also published [47]
Although IsAs have been studied for a longer period than AmAs, it is important to identify the essential similarities between the main components of both pathways. Both pathways share a core compound created through the condensation of two molecules: norcoclaurine (IsAs) and norbelladine (AmAs). Additionally, both compounds are capable of undergoing modifications that generate a molecule able to perform intramolecular phenol-coupling rearrangements through CYPs: reticuline (IsAs) and 4OMET (AmAs). Figure 4 illustrates the major discoveries in each pathway over time. Genomic and transcriptomic data have played a crucial role in discovering five out of seven genes responsible for morphine biosynthesis since 2010. However, the sequence of enzymes characterized thus far in the AmAs pathway has been elucidated solely with the aid of transcriptomes. Surprisingly, N4OMT was partially purified in 1963 [48], eighteen years before the first purification of (S)-norlaudanosoline synthase in the morphine pathway [29]. Since 1981, research into the AmAs pathway has not advanced as rapidly as that of the IsAs pathway. This is especially true when considering that the morphine pathway involves more steps than those involved in the synthesis of lycorine or galanthamine.
2.1. Mining Functional Genes on IsAs Biosynthesis
Isoquinoline alkaloids (IsAs) are renowned for their diversity and prevalence across multiple plant species, leading to extensive research on their biosynthesis. Transcriptomic studies have covered more than 70 species across 20 families (Table 2). To generate transcriptomes of IsAs-producing plants, technologies like third-generation sequencing have been used. In addition, genomic data from various families has been analyzed in tandem with transcriptomic data, specifically, just over 20 genomes have been reviewed within the Ranunculales order. However, despite significant efforts, certain biosynthetic pathways for specific IsAs remain only partially understood [49]. A systematic search of the Web of Science database yielded 237 articles on IsAs and transcriptomes data, 104 of which focused on plants (Figure 5). Transcriptomic analyses across various IsAs-producing genera have provided valuable insights into conditions, elicitors, and DEGs associated with the production of specific alkaloids. Notably, certain enzymes are predominantly studied within specific plant families.
The Papaveraceae family has been the most extensively studied, revealing the presence of CYP719B1, a key P450-dependent oxidoreductase responsible for the phenol-phenol coupling of (S)-reticuline. This coupling reaction is the most studied in the production of IsAs [113], and is regulated by specific transcription factors and miRNAs in response to alkaloid elicitors. For example, the WRKY transcription factor family regulates morphinan biosynthesis [92]. The STORR protein, a gene fusion of a P450 enzyme and an oxidoreductase, is another important gene, it plays a key role in morphinan alkaloids pathway, transforming (S)-Reticuline into (R)-Reticuline directing metabolites into the morphinan branch of the pathway [98]. A large diversity of CYPs has been described in different species, many of which show organ specificity. Genomic data suggests that specific plant species produce exclusive families of alkaloids. Therefore, having sanguinarine, morphine, and papaverine in the same plant is unlikely. Regarding DEGs during bulb development in Corydalis yanhusuo, a study revealed several significant findings [86]. Using RNA-Seq, it identified dynamic changes in transcript abundance across three developmental stages: Day 0 (bulb initiation), Day 10 (early expansion), and Day 30 (maturation), 1387 unigenes were co-expressed on Day 0 and Day 10, while 917 were co-expressed on Day 10 and Day 30, and 662 unigenes were co-expressed between Day 0 and Day 30, indicating distinct gene expression patterns throughout development. Moreover, alkaloid biosynthesis was found to be developmentally regulated, as their accumulation increased as the bulbs matured. This suggests that alkaloid biosynthesis is transcriptionally regulated during bulb development.
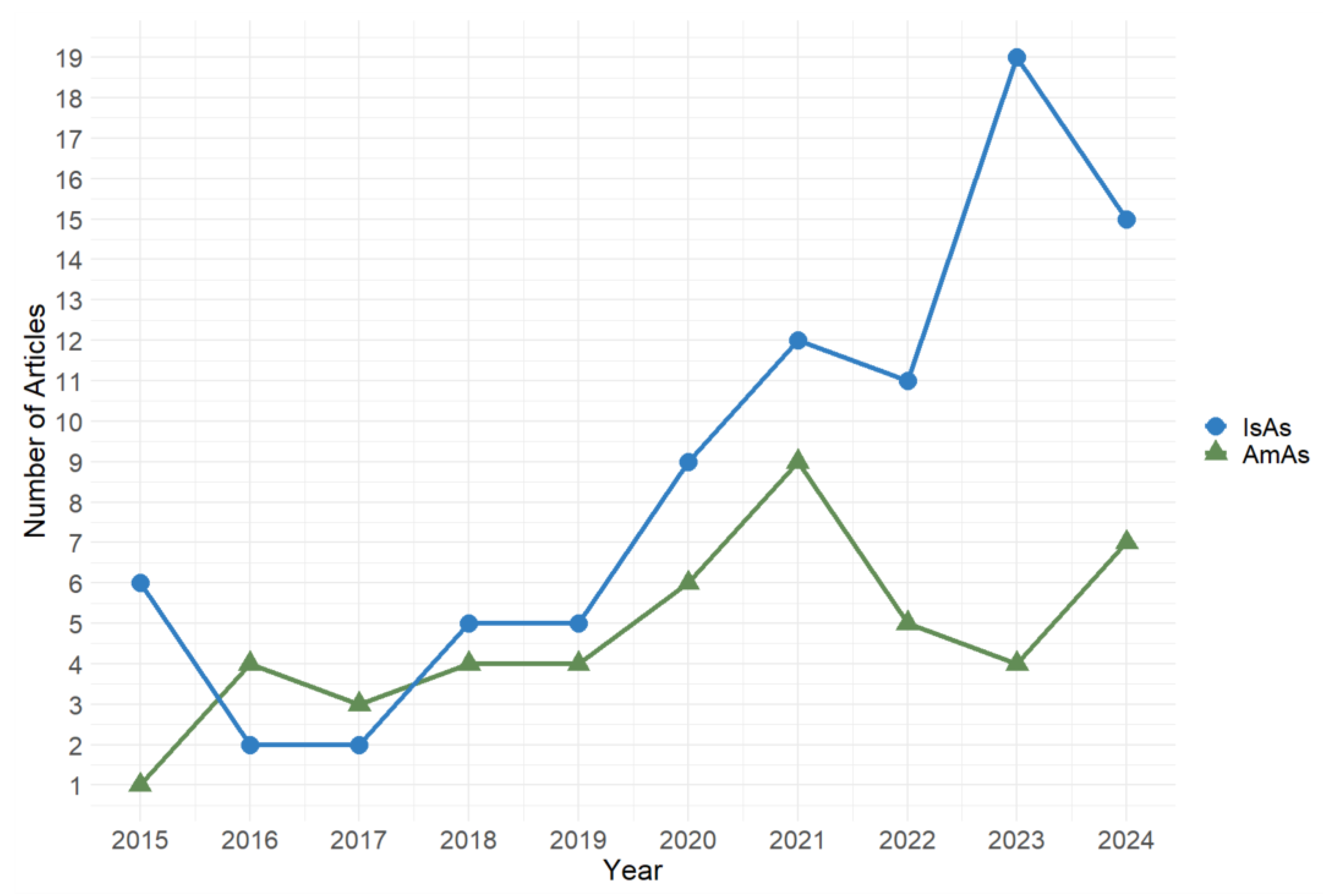
Figure 5.
Number of publications with transcriptomic related data on IsAs and AmAs over the years. Temporal evolution of the number of scientific publications mentioning IsAs (blue) and alkaloids from the AmAs (green). An increase in the number of publications for both categories has been observed in recent years, indicating a growing interest in the research of these bioactive compounds.
Figure 5.
Number of publications with transcriptomic related data on IsAs and AmAs over the years. Temporal evolution of the number of scientific publications mentioning IsAs (blue) and alkaloids from the AmAs (green). An increase in the number of publications for both categories has been observed in recent years, indicating a growing interest in the research of these bioactive compounds.
The combination of transcriptomic and metabolomic analyses related to plant developmental stages in members of the Ranunculacea family, such as Coptis chinensis, revealed that four-year-old rhizomes exhibited higher levels of IsAs alkaloids compared to two-year-old rhizomes [114]. Transcriptome analysis identified 464 DEGs, with 36 associated with the IsAs biosynthesis pathway. The study highlighted that the rapid growth stage is crucial for alkaloid accumulation. During this period, plants are more likely to produce higher amounts of bioactive compounds, whereas their accumulation may be less pronounced during the slower growth stages. Although no CYP719B enzymes have been reported in this family, CYP719A enzymes are present and are responsible for the formation of methylenedioxy bridges [115]. Additionally, CYP80G2 plays a key role in the C-C phenol-coupling reaction in Coptis japonica, contributing to aporphine biosynthesis [116]. Studies on the Berberidaceae family have revealed the presence of hydroxylases, N-methyltransferases, 4- and 6-O-methyltransferases, and synthases such as norcoclaurine synthase (NCS) and coclaurine synthase (CS) [57,58,59,65]. In addition, studies on Menispermaceae have identified DEGs between roots and leaves, including CYP80G, a phenol-phenol coupling enzyme involved in the production of corytuberine [57,59,65,75,76]. Studies in other families have focused on the effects of elicitors, such as those reported in the Araceae family, where factors such as light wavelength and hormones have been studied. Gene ontology analysis has suggested their influence on IsAs production [51,52].
Methyltransferases play a central role in the biosynthesis of IsAs due to their crucial function in generating alkaloid diversity [117,118]. Alkaloid skeletons are often modified through methylation events, which typically occur after condensation reactions catalyzed by lyases—more common in the IsAs pathway than in the AmAs pathway [73,119]. These methylation events, often followed by further methylations or coupled with oxidoreductase activity, lead to rearrangements of alkaloid skeletons, forming distinct alkaloid families. For example, dopamine-derived colchicine alkaloids undergo multiple methylation and oxidoreduction steps, producing complex multimethylated structures with seven-membered rings [120]. Downstream in the synthesis of (S)-reticuline, the molecule may interact with methyltransferases or oxidoreductases. These enzymatic activities contribute to ring closure, radical reduction and substrate stereospecificity [121,122]. Thus, methyltransferases and oxidoreductases play a crucial role in shaping the structural features and stereochemistry of IsAs during their biosynthesis.
2.2. Mining Functional Genes on AmAs Biosynthesis
RNA-Seq-based strategies, combined with isotopically labeled substrates, have become the primary tool for understanding the biosynthesis of AmAs. The enzymes have been identified using a similar methodology, which involves targeted or untargeted transcriptomics combined with metabolomic analyses, cloning of candidate genes, heterologous expression, in vitro enzymatic assays, metabolite characterization, and studies with isotope-labeled precursors. Recent studies highlight the use of transcriptomic analyses targeted at the developmental stage of different sections of basal tissue, as well as transient expression mediated by Agrobacterium tumefaciens in systems such as Nicotiana benthamiana [46]. The number of transcriptomic studies focused on this topic has been increasing steadily (Figure 4). Research in this area began in when Wang et al [123] published the first transcriptome assembly for an Amaryllidaceae species, Lycoris aurea, in response to different elicitors and tissues. In this study, 116,507 singletons were identified, related to responses to methyl jasmonate (MeJA) and salicylic acid (SA) in various tissues. Since then, additional transcriptomes for Amaryllidaceae species have been sequenced (Table 3), and in some cases, the proposed transcripts have been cloned to assess the functionality of the predicted enzymes [42,43,44,124]. Research has primarily focused on Leucojum and Narcissus species in Canada, while Lycoris species have been the most studied in China. To date, only one approach using third-generation sequencing has been conducted, while the majority of transcriptomic data from this family has been generated through second-generation sequencing technologies, and the only genome assembly available is for Narcissus pseudonarcissus (GCA_032191725.1), as part of a whole genome shotgun sequencing project. While genome sequences for other Amaryllidoideae species are registered in NCBI BioProjects, these data have not yet been made publicly available.
From the first transcriptome published, 25 hits for transcripts encoding phenylalanine ammonia-lyase (PAL), 6 for tyrosine decarboxylase (TYDC), 91 for O-methyltransferases (OMT), 214 for plant cytochromes P450 (P450s) and 191 for N-methyltransferases (NMT) were identified as potential candidates related to AmAs biosynthesis, but non validated [123]. From that point onwards, thirteen enzymes are related to the production of 4OMET have been exclusively characterized in the AmAs pathway including norbelladine O-methyltransferase (N4OMT) and norbelladine synthase (NBS). Enzymes related to and after the para-para coupling of 4OMET include noroxomaritidine synthase (CYP96T1), noroxomaritidine reductase (NR), normaritidine synthase (SDR2), vitattine synthase (CYP71DW1), vitattine hydroxylase (ODD2), and 11-hydroxyvitattine-O-methyltransferase (OMT1). Enzymes related to and after the para-ortho coupling of 4OMET include nornarwedine synthase (CYP96T6), nornarwedine methyltransferase (NMT1), and narwedine reductase (AKR1). And enzymes related to and after the ortho-para coupling of 4OMET include noroxopluviine synthase (CYP96T5) and noroxopluviine reductase (SDR1) to produce norpluviine.
N4OMT was the first enzyme characterised by Kilgore´s team in 2014 through illumina sequencing and transcriptome assembly. Candidate genes were identified using HAYSTACK, followed by PCR, cloning in E. coli BL21(DE3), protein purification, and kinetic characterization [42]. CYP96T1 and NR were described in 2016, with CYP96T1 being the enzyme responsible for the para-para coupling of 4OMET and a low-rate para-ortho coupling of the same compound as a by-product [45]. NR is a short-chain alcohol dehydrogenase/reductase responsible for the reaction following the para-para coupling of 4OMET.

Table 3.
Transcriptome sequencing studies on Amaryllidaceae species with data corelated to AmAs production.
Table 3.
Transcriptome sequencing studies on Amaryllidaceae species with data corelated to AmAs production.
| Amaryllidaceae species | Sequencing year | Sequencing country | Sequencing platforms |
Reference |
|---|---|---|---|---|
| Crinum x powellii | 2023 | Canada | Illumina NovaSeq 6000 | [125] |
|
Leucojum aestivum |
2022 | Canada | Illumina HiSeq 2000 | [126] |
|
Lycoris aurea |
2013 | China | 454 GS FLX | [123] |
| 2016 | China | Illumina HiSeq 2000 | [127] | |
| 2016 | China | Illumina HiSeq 2000 | [128] | |
| 2017 | China | Illumina HiSeq 2000 | [129] | |
| 2024 | China | Illumina HiSeq X Ten | [130] | |
|
Lycoris chinensis |
2022 | China | Illumina HiSeq 2500 | [131] |
|
Lycoris longituba |
2020 | China | Illumina HiSeq X Ten | [132] |
| 2021 | China | Illumina NovaSeq 6000 | [119] | |
|
Lycoris radiata |
2019 | China | Illumina NextSeq 500 | [133] |
| Narcissus cyclamineus | 2024 | USA | Illumina HiSeq 4000/PacBio Iso-Seq | [46] |
| Narcissus papyraceus | 2019 | Canada | Illumina HiSeq 2000 | [134] |
| Narcissus pseudonarcissus | 2017 | Canada | Illumina HiSeq 2000 | [135] |
| 2021 | U.K. | Illumina HiSeq 2500 | [136] |
* Tables may have a footer.
It is important to note that NR was found to be able to reduce norcraugsodine, an imine Schiff base that is an intermediate in the formation of norbelladine from 3,4-DHBA [137]. Condensation between tyramine and 3,4-DHBA is thought to occur spontaneously, followed by reduction by an enzyme such as NR. However, it is unclear whether the condensation takes place in the active site of the enzyme. It was not until 2018 that the main enzyme responsible for norbelladine formation was described, when Desgagné's group reported an NBS as the first relevant synthase specific to the AmAs pathway [43].
DEGs related to alkaloid biosynthesis vary depending on the tissue type and the developmental stage at which samples are collected [43,134]. Bulbs tend to accumulate higher concentrations of AmAs like galanthamine and lycorine, while proteins related to earlier steps in primary metabolism, such as TYDC, are found more broadly across tissues [136,138]. This suggests that gene expression of downstream biosynthetic pathway genes is tissue-specific, with potential metabolite translocation occurring within the plant. Mehta et al [46] studied DEGs related to alkaloid biosynthesis in Narcisus, finding differences in metabolite accumulation patterns across tissues. They proposed two possible explanations: either local synthesis of metabolites or transport from other tissues. Their results showed that younger foliar tissues accumulated fewer alkaloids than older tissues but still exhibited active biosynthesis, suggesting a strategy for resource allocation in toxin production during early development, and allows the identification and characterization of enzymes involved in galanthamine and hemanthamine biosynthesis such as NtAR1, NtCTP96T6 and NtNMT1.
The regulation of AmAs has also been studied in the family. In fact, one transcriptome research in Lycoris aurea showed that some transcripts can be related to AmAs by modulating the expression of some enzymes in the pathway in response to elicitors. Specifically, miR396 was found to be increased as well as galanthamine synthesis in response to MeJA (Xu et al. 2016). This study is the only one that has explored the role of miRNAs in the regulation of expression of genes related to alkaloid synthesis in Amaryllidaceae, and the authors reported that no miRNAs were found to target OMTs or NMTs. Furthermore, a direct relationship between a TYDC and the same miRNA was found later in a more specific study, where it was reported as responsible for tyramine accumulation in different tissues [139].
Despite significant advances, one of the key unresolved questions in AmAs biosynthesis is the specific pathway leading to the production of 3,4-dihydroxybenzaldehyde (3,4-DHBA) (Figure 3). Although caffeic acid has been detected in Amaryllidaceae and is believed to act as an intermediate in 3,4-DHBA synthesis, no enzymes from this family have been fully characterized to bridge the gaps in the pathway, despite the proposal of candidates such as CSE and HBS [24,124]. Recently, the metabolite dihydrocaffeic acid methyl ester was found in Pancratium maritimum [140] and it was also described in Hippeastrum vittatum 17 years prior [141]. It is also possible that 3,4-DHBA formation may be accomplished by CoA coupling through enzymes like FerB; however, this pathway has only been proposed through retrobiosynthesis data [142].
3. Integrative Approaches on Biosynthetic Understanding
Authors should discuss the results and how they can be interpreted from the perspective of previous studies and of the working hypotheses. The findings and their implications should be discussed in the broadest context possible. Future research directions may also be highlighted. The enzymes involved in the proposed biosynthetic pathway of AmAs consist of oxidoreductases, hydrolases, methyltransferases, and lyases, with oxidoreductases being the most frequently found. This enzyme distribution resembles that of IsAs biosynthesis, although a quick glance at the KEGG pathway map (https://www.genome.jp/pathway/map00950) reveals a more complex route for IsAs, with a predominance of methyltransferases and synthases. Each enzyme group exhibits distinct characteristics, such as specificity for co-substrates [143,144]. Enzymes involved in IsAs biosynthesis have been extensively characterized through genomic and transcriptomic approaches, in contrast to Amaryllidaceae alkaloids AmAs, where there is still a significant lack of genomic information. Until recently, two major gaps remained in the AmAs biosynthetic pathway: (1) the specific enzymes responsible for producing 3,4-DHBA and (2) the C–C coupling reactions catalyzed by cytochrome P450s (CYPs), oxidoreductases, and methyltransferases downstream of 4OMET. However, recent advances have begun to shed light on the second gap, thanks to the research teams of Sattely’s [46] and Desgagne-Penix´s [47] who have described a variety of CYPs involved in these key C–C coupling reactions.
It is interesting to note that information about enzymes capable of ortho-para coupling is still needed for both IsAs and AmAs, despite the importance of compounds produced through this type of rearrangement in both groups of alkaloids—lycorine-type alkaloids in AmAs and isoboldine-type alkaloids in IsAs. While several lycorine CYPs have been proposed [46,47], the pathway remains to be fully understood, as deep as galanthamine and hemanthamine. In contrast, for isoboldine, no CYP has yet been proposed, leaving a significant gap in understanding this pathway [145]. Nature has played a significant role in the evolution of enzymes that exhibit precise control over both the site and stereoselectivity of bond formation. Among these enzymes, CYPs, laccases, and peroxidases are particularly noteworthy, with CYPs being the most extensively studied in alkaloid biosynthetic pathways. These enzymes initiate radical mechanisms in phenolic rings by abstracting a hydrogen atom. Interestingly, only CYPs have been characterized as being responsible for intermolecular couplings, whereas all three enzyme types—CYPs, laccases, and peroxidases—can facilitate intramolecular couplings [146]. This suggests that research focusing on enzymes beyond CYPs, such as laccases and peroxidases, could provide valuable insights into alkaloid biosynthesis. Expanding the scope of these enzymes might uncover new mechanisms and enzyme functions involved in the formation of complex alkaloid structures.
It is important to not only explore the proposed reactions found in other plants but also to investigate chemical similarities with other SPMs to identify enzymatic activities responsible for such biotransformations. Regarding the first gap, insights suggest that the native pathways could be elucidated by integrating data from methyltransferases and oxidoreductases [73,147] (Table 4). For both IsAs and AmAs, many enzymes involved have shown the ability to efficiently catalyze a wide variety of substrates. However, it is essential to note that assays testing enzyme specificity have only been conducted within compounds of the same biosynthetic pathways. This limits our understanding of their broader catalytic potential across different SPMs, suggesting a need for more diverse experimental setups to explore substrate flexibility.
Analysis based on chemical structures of the metabolites can be accomplished to gain insights into what is needed to be looked for. For example, noticing that the structure of AmAs after the phenol-phenol coupling has tetrahydrofurans (galanthamine) or even dioxolanes (lycorine, crinine and haemanthamine) which are products of cyclization that can be catalyzed by CYPs enzymes, similar compounds can be addressed in other routes like from IsAs and analyzed to discover potential enzymes capable of making such cyclizations. Nornarwedine, so far known as the p-o precursor of galanthamine, has an immediate tetrahydrofuran ring produced from 4OMET after the phenol-phenol coupling that seems to be spontaneously produced 148; however, this rearrangement does not occur in IsAs biosynthesis of (S)-salutaridine, the p-o morphine precursor where the formation of a tetrahydrofuran ring happens through the incorporation of acetate which provides the radicals needed to produce thebaine and later this molecule undergoes several demethylation and isomerization reactions leading to morphine [34] which could be seen as the structural analogue to galanthamine among IsAs.
Despite the various technologies used for RNA-Seq, the geographic distribution of research on IsAs-producing plants is notably more diverse compared to that on Amaryllidaceae. China leads investigations related to omics approaches for both IsAs and AmAs (Figure 6), highlighting its dominance in plant natural product (PNP) research. This underscores the need for a more globally diversified interest and research effort in this area, as advancements in understanding these pathways and their enzymes hold great potential for biotechnological applications across different regions. Expanding research beyond current geographic boundaries could enhance knowledge and exploration of these valuable compounds. Different technologies for RNA-Seq have been employed to study the transcriptomes of IsAs and AmAs-producing plants. However, it is crucial to note that sequencing technologies are continuously evolving, making it easier to obtain relevant information. For instance, several Illumina platforms such as HiSeq 2500, HiSeq 3000, HiSeq 4000, HiSeq X Ten, and HiSeq X Five have been declared discontinued [148]. This exemplifies the rapid advancement in sequencing technologies, underscoring the importance of keeping pace with new developments to facilitate more efficient and comprehensive transcriptomic analyses.
4. Challenges and Opportunities
DNA sequencing has complemented RNA-Seq data, providing deeper insights into these processes. By 2022, sequencing a complete human genome cost approximately 1,000 USD, with results obtainable within a few days [149], by 2024, the company 3Billion estimates the cost of whole genome sequencing at approximately 600 USD, while RNA sequencing is projected to range between 36.9 and 173 USD per sample, showing a clear difference in price and accessibility. Beyond common sequencing challenges, plant genomes tend to be larger in size due to their high content of transposable elements and polyploidy, characterized by the presence of more than one set of chromosomes, including identical homologues, which are difficult to distinguish during genome assembly [150]. Moreover, the lack of bioinformatic tools specifically tailored for polyploids complicates the generation of reference genomes [151], as these identical sequences frequently result in chimeric contigs with switch errors or false alingments [152]. This issue is particularly relevant for the Ranunculaceae family, where species such as Ranunculus kuepferi exhibit diploid, triploid, and autotetraploid cytotypes in the Alps [153], as well as for the second major clade of the Amaryllidaceae in the Americas, the Andean clade, which consists of tetraploid species [154]. Exploring polyploid genomes not only enhances our understanding of enzyme variability and metabolic diversity but also provides a foundation for investigating the biosynthesis of alkaloids.
Establishing connections between different groups of alkaloids is essential for elucidating the molecular mechanisms underlying the production of these diverse compounds. Research into IsAs has made significant strides, particularly in identifying the enzymes responsible for key biotransformations. Among the most extensively studied enzyme groups are methyltransferases and oxidoreductases (including CYPs). These enzymes have even been used in studies of evolutionary ancestry [73] and linked to polyploidy events that drive the specificity of IsAs production in particular taxa [97,98]. Over the last two decades, transcriptomic analyses of IsA-producing plants have led to the near-complete elucidation of the biosynthetic pathway for key alkaloids such as morphine, However, research on the molecular mechanisms underlying the biosynthesis of AmAs has progressed more slowly. Since 2018, transcriptomes in the genera Leucojum sp., Narcissus sp. and Lycoris sp. have been continuously studied (Table 3), from these studies, elicitors effects have been more understood and other important data has been obtained like NBS being localized in the cytosol [126] and S-adenosyl methionine (SAM) synthetases and decarboxylases have been found co-expressing among previously known transcripts [136]. Despite enzymes from AmAs pathway have been described since 2014: NBS, NR, N4OMT and CYP96T1, being NBS the last one described 5 years ago and its specific mechanism coupled to NR is the most recently studied [44], the non-enzymatic components of the pathway and the regulatory mechanisms behind biosynthesis remain poorly understood, leaving a gap in knowledge that could be addressed through genomics and transcriptomics, as well as through a deeper understanding of the impact of elicitors and plant developmental stages on the biosynthesis of both IsAs and AmAs.
While challenges in genome and transcriptome sequencing of AmAs-producing plants persist, these limitations also present opportunities to advance integrative and collaborative approaches. In this context, discussing emerging opportunities in biotechnology, such as engineering biosynthetic pathways in microbial systems, offers a promising perspective toward industrial and pharmacological applications. The discovery of complete biosynthetic gene clusters that can be used to produce AmAs or IsAs is key for protein engineering and bioprospecting of enzymes and metabolites. Such discoveries hold immense potential for biotechnological developments, especially in the realm of SPMs. Although Amaryllidaceae plants have long been valued for their pharmacological properties, most research has focused on extracting raw components from these plants. Investigations into in vitro cultures, using callus or parenchyma cells under controlled conditions, aim to produce AmAs with better reproducibility. However, there is limited discussion on the cost-effectiveness of these experiments. Microbial cell factories have successfully been used to produce SPMs in various microorganisms [155], and IsAs are no exception [156,157,158,159]. Continuing to develop similar approaches for AmAs, based on fully elucidated biosynthetic pathways through multi-omics integration, would be of tremendous value.
These advancements pave the way for multiple practical applications in biotechnology, pharmacology, and industrial production. For instance, microbial engineering has enabled the use of engineered microorganisms to produce high-value compounds like morphine, reducing dependency on plant extraction, which is often costly and environmentally unsustainable. Beyond healthcare, IsAs have shown bioactivity as templates for herbicides and pesticides, which could lead to environmentally friendly agricultural solutions. Similarly, leveraging these techniques for AmAs could provide controlled and scalable production platforms for compounds like galanthamine and lycorine with pharmacological applications for Alzheimer’s treatment, and cytotoxic and antitumor activities respectively, presenting potential for novel chemotherapeutic agents.
5. Conclusions
Historically, the elucidation of IsAs pathways began earlier than that of AmAs pathways. Both have undergone similar approaches, which have been shaped by technological advancements over the years, allowing for different investigative strategies. A clear difference lies in the availability of genomic data, which is significantly greater for IsAs-producing plants compared to those in the Amaryllidaceae family, along with a predominant reliance on transcriptomic approaches for studying AmAs. Thus, expanding multi-omics data in the elucidation of metabolic pathways is essential, as it facilitates a comprehensive and integrative understanding of the processes involved. Interdisciplinary collaborations and investments in genomic and transcriptomic resources are essential to fully unlock alkaloid potential and realize their economic and societal benefits. Bridging basic science with applied research, the elucidation of these pathways offers innovative solutions across industries and creates opportunities for the development of biotechnological applications, facilitating the scalable production of compounds with pharmaceutical and industrial relevance.
Acknowledgments
We gratefully acknowledge Minciencias, the Colombian Ministry of Science, Technology, and Innovation, for its funding support through the grant No. 86978, and the Assistant Professorship Funds from Universidad de los Andes cod. INV-2023-158-2701.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Hoffmann, D. Medical Herbalism: The Science Principles and Practices Of Herbal Medicine; 1st ed.; Healing Arts Press: Rochester, Vt, 2003. [Google Scholar]
- Seigler, D.S. Plant Secondary Metabolism; Springer US, 1998;
- Hussein, R.A.; El-Anssary, A.A. Plants Secondary Metabolites: The Key Drivers of the Pharmacological Actions of Medicinal Plants. In Herbal Medicine; Builders, P.F., Ed.; IntechOpen, 2019.
- Taylor, S.L.; Hefle, S.L. Naturally Occurring Toxicants in Foods. In Foodborne Diseases (Third Edition); et al., C.E.R.D., Ed.; Academic Press, 2017; pp. 327–344.
- Pallardy, S.G. Chapter 9 - Nitrogen Metabolism. In Physiology of Woody Plants (Third Edition); Academic Press, 2008; pp. 233–254.
- Rajput, A.; Sharma, R.; Bharti, R. Pharmacological Activities and Toxicities of Alkaloids on Human Health. Mater Today Proc 2022, 48, 1407–1415. [Google Scholar] [CrossRef]
- Thawabteh, A.; Juma, S.; Bader, M.; Karaman, D.; Scrano, L.; Bufo, S.A.; Karaman, R. The Biological Activity of Natural Alkaloids against Herbivores, Cancerous Cells and Pathogens. Toxins 2019, Vol. 11, Page 656 2019, 11, 656. [Google Scholar] [CrossRef]
- Adamski, Z.; Blythe, L.L.; Milella, L.; Bufo, S.A. Biological Activities of Alkaloids: From Toxicology to Pharmacology. Toxins (Basel) 2020, 12, 210. [Google Scholar] [CrossRef]
- dos Santos, A.R.; Vaz, N.P. Isoquinoline Alkaloids and Chemotaxonomy. 2019, 167–193. [CrossRef]
- Dey, P.; et al. Chapter 15 - Analysis of Alkaloids (Indole Alkaloids, Isoquinoline Alkaloids, Tropane Alkaloids). In Recent Advances in Natural Products Analysis; et al., A.S.S., Ed.; Elsevier, 2020; pp. 505–567.
- Bozarth, M. Dopamine. In; 2017; pp. 1173–1177.
- Samanani, N.; Facchini, P.J. Purification and Characterization of Norcoclaurine Synthase: THE FIRST COMMITTED ENZYME IN BENZYLISOQUINOLINE ALKALOID BIOSYNTHESIS IN PLANTS. Journal of Biological Chemistry 2002, 277, 33878–33883. [Google Scholar] [CrossRef]
- Onoyovwe, A.; Hagel, J.M.; Chen, X.; Khan, M.F.; Schriemer, D.C.; Facchini, P.J. Morphine Biosynthesis in Opium Poppy Involves Two Cell Types: Sieve Elements and Laticifers. Plant Cell 2013, 25, 4110. [Google Scholar] [CrossRef]
- Schenck, C.A.; Maeda, H.A. Tyrosine Biosynthesis, Metabolism, and Catabolism in Plants. Phytochemistry 2018, 149, 82–102. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Xu, X.-H. Amaryllidaceae Alkaloids. In Natural Products: Phytochemistry, Botany and Metabolism of Alkaloids, Phenolics and Terpenes; Ramawat, K.G., Mérillon, J.-M., Eds.; Springer, 2013; pp. 479–522.
- Bastida, J.; Berkov, S.; Torras Claveria, L.; Pigni, N.B.; Andrade, J.P.; Martínez, V.; Codina Mahrer, C.; Viladomat Meya, F. Chemical and Biological Aspects of Amaryllidaceae Alkaloids. In Recent Advances in Pharmaceutical Sciences; Elsevier, 2011; pp. 65–100.
- Berkov, S.; Osorio, E.; Viladomat, F.; Bastida, J. Chemodiversity, Chemotaxonomy and Chemoecology of Amaryllidaceae Alkaloids. In The Alkaloids: Chemistry and Biology; Knölker, H.-J., Ed.; Academic Press, 2020; Vol. 83, pp. 113–185.
- Dey, A.; Mukherjee, A. Chapter 6 - Plant-Derived Alkaloids: A Promising Window for Neuroprotective Drug Discovery. In Discovery and Development of Neuroprotective Agents from Natural Products; Brahmachari, G., Ed.; Elsevier (Natural Product Drug Discovery), 2018; pp. 237–320.
- Marco, L.; do Carmo Carreiras, M. Galanthamine, a Natural Product for the Treatment of Alzheimer’s Disease. Recent Pat CNS Drug Discov 2006, 1, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Kausar, S.; Mustafa, H.G.; Altaf, A.A.; Mustafa, G.; Badshah, A. ; others Galantamine. In Reference Module in Biomedical Sciences; Elsevier, 2019.
- Lamoral-Theys, D.; Andolfi, A.; Van Goietsenoven, G.; Cimmino, A.; Le Calvé, B.; Wauthoz, N.; Mégalizzi, V.; Gras, T.; Bruyère, C.; Dubois, J.; et al. Lycorine, the Main Phenanthridine Amaryllidaceae Alkaloid, Exhibits Significant Antitumor Activity in Cancer Cells That Display Resistance to Proapoptotic Stimuli: An Investigation of Structure−Activity Relationship and Mechanistic Insight. ACS Publications 2009, 52, 6244–6256. [Google Scholar] [CrossRef] [PubMed]
- Ka, S.; Masi, M.; Merindol, N.; Di Lecce, R.; Plourde, M.B.; Seck, M.; Górecki, M.; Pescitelli, G.; Desgagne-Penix, I.; Evidente, A. Gigantelline, Gigantellinine and Gigancrinine, Cherylline- and Crinine-Type Alkaloids Isolated from Crinum Jagus with Anti-Acetylcholinesterase Activity. Phytochemistry 2020, 175, 112390. [Google Scholar] [CrossRef]
- Mamun, A.; Pidaný, F.; Hulcová, D.; Maříková, J.; Kučera, T.; Schmidt, M.; Catapano, M.C.; Hrabinová, M.; Jun, D.; Múčková, L.; et al. Amaryllidaceae Alkaloids of Norbelladine-Type as Inspiration for Development of Highly Selective Butyrylcholinesterase Inhibitors: Synthesis, Biological Activity Evaluation, and Docking Studies. Int J Mol Sci 2021, 22, 8308. [Google Scholar] [CrossRef]
- Desgagné-Penix, I. Biosynthesis of Alkaloids in Amaryllidaceae Plants: A Review. Phytochemistry Reviews 2021, 20, 409–431. [Google Scholar] [CrossRef]
- Moreno, R.; Tallini, L.R.; Salazar, C.; Osorio, E.H.; Montero, E.; Bastida, J.; Oleas, N.H.; Acosta León, K. Chemical Profiling and Cholinesterase Inhibitory Activity of Five Phaedranassa Herb. (Amaryllidaceae) Species from Ecuador. Molecules 2020, 25, 2092. [Google Scholar] [CrossRef] [PubMed]
- Georgiev, V.; Ivanov, I.; Pavlov, A. Recent Progress in Amaryllidaceae Biotechnology. Molecules 2020, 25, 4670. [Google Scholar] [CrossRef]
- Krishnamurti, C.; Rao, S.C. The Isolation of Morphine by Serturner. Indian J Anaesth 2016, 60, 861–862. [Google Scholar] [CrossRef] [PubMed]
- Kirby, G.W. Biosynthesis of the Morphine Alkaloids. Science (1979) 1967, 155, 170–173. [Google Scholar] [CrossRef]
- Rueffer, M.; El-Shagi, H.; Nagakura, N.; H Zenk, M. (S)-Norlaudanosoline Synthase: The First Enzyme in the Benzylisoquinoline Biosynthetic Pathway. FEBS Lett 1981, 129, 5–9. [Google Scholar] [CrossRef]
- Loeffler, S.; Stadler, R.; Nagakura, N.; Zenk, M.H. Norcoclaurine as Biosynthetic Precursor of Thebaine and Morphine. J Chem Soc Chem Commun 1987, 1160–1162. [Google Scholar] [CrossRef]
- Facchini’, P.J.; De Luca, V. Phloem-Specific Expression of Tyrosine/Dopa Decarboxylase Genes and the Biosynthesis of Isoquinoline Alkaloids in Opium Poppy. Plant Cell 1995, 7, 1811–1821. [Google Scholar] [CrossRef]
- Grothe, T.; Lenz, R.; Kutchan, T.M. Molecular Characterization of the Salutaridinol 7-O-Acetyltransferase Involved in Morphine Biosynthesis in Opium Poppy Papaver Somniferum. Journal of Biological Chemistry 2001, 276, 30717–30723. [Google Scholar] [CrossRef] [PubMed]
- Winzer, T.; Kern, M.; King, A.J.; Larson, T.R.; Teodor, R.I.; Donninger, S.L.; Li, Y.; Dowle, A.A.; Cartwright, J.; Bates, R.; et al. Plant Science. Morphinan Biosynthesis in Opium Poppy Requires a P450-Oxidoreductase Fusion Protein. Science 2015, 349, 309–312. [Google Scholar] [CrossRef]
- Dastmalchi, M.; Chen, X.; Hagel, J.M.; Chang, L.; Chen, R.; Ramasamy, S.; Yeaman, S.; Facchini, P.J. Neopinone Isomerase Is Involved in Codeine and Morphine Biosynthesis in Opium Poppy. Nature Chemical Biology 2019 15:4 2019, 15, 384–390. [Google Scholar] [CrossRef]
- Hao, L. Evaluation of Biosynthetic Pathway and Engineered Biosynthesis of Morphine with CRISPR. BIO Web Conf 2023, 59, 01022. [Google Scholar] [CrossRef]
- Narcross, L.; Pyne, M.E.; Kevvai, K.; Siu, K.-H.; Dueber, J.E.; Martin, V.J.J. Benzylisoquinoline Alkaloid Production in Yeast via Norlaudanosoline Improves Selectivity and Yield. bioRxiv 2023, 2023.05.19.541502. [Google Scholar] [CrossRef]
- Ozber, N.; Yu, L.; Hagel, J.M.; Facchini, P.J. Strong Feedback Inhibition of Key Enzymes in the Morphine Biosynthetic Pathway from Opium Poppy Detectable in Engineered Yeast. ACS Chem Biol 2023, 18, 419–430. [Google Scholar] [CrossRef]
- Gerrard, A.W. The Proximate Principles of the Narcissus Pseudonarcissus. The pharmaceutical journal and transactions. Soc. 1878, 214–215. [Google Scholar]
- Barton, D.; Cohen, T. Some Biogenetic Aspects of Phenol Oxidation. In Festschrift Prof Dr Arthur Stoll; Birkhäuser Verlag: Basel, 1957; p. 117. [Google Scholar]
- Eichhorn, J.; Takada, T.; Kita, Y.; Zenk, M.H. Biosynthesis of the Amaryllidaceae Alkaloid Galanthamine. Phytochemistry 1998, 49, 1037–1047. [Google Scholar] [CrossRef]
- El Tahchy, A.; Boisbrun, M.; Ptak, A.; Dupire, F.; Chrétien, F.; Henry, M.; Chapleur, Y.; Laurain-Mattar, D. New Method for the Study of Amaryllidaceae Alkaloid Biosynthesis Using Biotransformation of Deuterium-Labeled Precursor in Tissue Cultures. Acta Biochim Pol. 2010, 57, 75–82. [Google Scholar] [CrossRef]
- Kilgore, M.B.; Augustin, M.M.; Starks, C.M.; O’Neil-Johnson, M.; May, G.D., C. J.A.; Kutchan, T.M. Cloning and Characterization of a Norbelladine 4′-O-Methyltransferase Involved in the Biosynthesis of the Alzheimer’s Drug Galanthamine in Narcissus Sp. Aff. Pseudonarcissus. PLoS One 2014, 9, e103223. [Google Scholar] [CrossRef]
- Singh, A.; Massicotte, M.A.; Garand, A.; Tousignant, L.; Ouellette, V.; Bérubé, G.; Desgagné-Penix, I. Cloning and Characterization of Norbelladine Synthase Catalyzing the First Committed Reaction in Amaryllidaceae Alkaloid Biosynthesis. BMC Plant Biol 2018, 18, 1–12. [Google Scholar] [CrossRef]
- Majhi, B.B.; Gélinas, S.E.; Mérindol, N.; Ricard, S.; Desgagné-Penix, I. Characterization of Norbelladine Synthase and Noroxomaritidine/Norcraugsodine Reductase Reveals a Novel Catalytic Route for the Biosynthesis of Amaryllidaceae Alkaloids Including the Alzheimer’s Drug Galanthamine. Front Plant Sci 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Kilgore, M.B.; Augustin, M.M.; May, G.D.; Crow, J.A.; Kutchan, T.M. CYP96T1 of Narcissus Sp. Aff. Pseudonarcissus Catalyzes Formation of the Para-Para’ C-C Phenol Couple in the Amaryllidaceae Alkaloids. Front Plant Sci 2016, 7, 225. [Google Scholar] [CrossRef]
- Mehta, N.; Meng, Y.; Zare, R.; Kamenetsky-Goldstein, R.; Sattely, E. A Developmental Gradient Reveals Biosynthetic Pathways to Eukaryotic Toxins in Monocot Geophytes. Cell 2024, 187, 5620–5637.e10. [Google Scholar] [CrossRef]
- Desgagne-Penix, I. Elucidating the Enzyme Network Driving Amaryllidaceae Alkaloids Biosynthesis. 2024. [CrossRef]
- Mann, J.D.; Fales, H.M.; Mudd, S.H. Alkaloids and Plant Metabolism: VI. O-METHYLATION IN VITRO OF NORBELLADINE, A PRECURSOR OF AMARYLLIDACEAE ALKALOIDS. Journal of Biological Chemistry 1963, 238, 3820–3823. [Google Scholar] [CrossRef]
- Pyne, M.E.; Martin, V.J.J. Microbial Synthesis of Natural, Semisynthetic, and New-to-Nature Tetrahydroisoquinoline Alkaloids. Curr Opin Green Sustain Chem 2022, 33, 100561. [Google Scholar] [CrossRef]
- Shi, P.; Gu, M. Transcriptome Analysis and Differential Gene Expression Profiling of Two Contrasting Quinoa Genotypes in Response to Salt Stress. BMC Plant Biol 2020, 20, 568. [Google Scholar] [CrossRef]
- Guo, C.; Chen Y; Wu D; Du Y; Wang M; Liu C; Chu J; Yao X Transcriptome Analysis Reveals an Essential Role of Exogenous Brassinolide on the Alkaloid Biosynthesis Pathway in Pinellia Ternata. Int J Mol Sci 2022, 23, 10898. [Google Scholar] [CrossRef]
- Zhong Y; Wang L; Ma Z; Du X Physiological Responses and Transcriptome Analysis of Spirodela Polyrhiza under Red, Blue, and White Light. Planta 2021, 255, 11. [CrossRef]
- Li, W.; Xu, P.; Qian, C.; Zhao, X.; Xu, H.; Li, K. The Combined Analysis of the Transcriptome and Metabolome Revealed the Possible Mechanism of Flower Bud Formation in Amorphophallus Bulbifer. Agronomy 2024, 14, 519. [Google Scholar] [CrossRef]
- Chen C; Shi X; Zhou T.; Li W; Li S; Bai G Full-Length Transcriptome Analysis and Identification of Genes Involved in Asarinin and Aristolochic Acid Biosynthesis in Medicinal Plant Asarum Sieboldii. Genome 2021, 64, 639–653. [CrossRef] [PubMed]
- Cui X; Meng F; Pan X; Qiu X; Zhang S; Li C; Lu S Chromosome-Level Genome Assembly of Aristolochia Contorta Provides Insights into the Biosynthesis of Benzylisoquinoline Alkaloids and Aristolochic Acids. Hortic Res 2022, 9, uhac005. [CrossRef]
- Zhao M; Ren Y; Li, Z. Transcriptome Profiling of Jerusalem Artichoke Seedlings (Helianthus Tuberosus L.) under Polyethylene Glycol-Simulated Drought Stress. Ind Crops Prod 2021, 170, 113696. [CrossRef]
- Farrow, S.C.; Hagel, J.M.; Facchini, P.J. Transcript and Metabolite Profiling in Cell Cultures of 18 Plant Species That Produce Benzylisoquinoline Alkaloids. Phytochemistry 2012, 77, 79–88. [Google Scholar] [CrossRef]
- Marques, J. V; Dalisay, D.S.; Yang, H.; Lee, C.; Davin, L.B.; Lewis, N.G. A Multi-Omics Strategy Resolves the Elusive Nature of Alkaloids in Podophyllum Species. Mol Biosyst 2014, 10, 2838–2849. [Google Scholar] [CrossRef]
- Hagel, J.M.; Morris, J.S.; Lee, E.J.; Desgagné-Penix, I.; Bross, C.D.; Chang, L.; Chen, X.; Farrow, S.C.; Zhang, Y.; Soh, J.; et al. Transcriptome Analysis of 20 Taxonomically Related Benzylisoquinoline Alkaloid-Producing Plants. BMC Plant Biol 2015, 15, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Lee, E.-J.; Chang, L.; Facchini, P. Genes Encoding Norcoclaurine Synthase Occur as Tandem Fusions in the Papaveraceae. Sci Rep 2016, 6, 39256. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.-L.; Li, X.; Liu, P.; Ma, L.; Wu, W.; Zhang, X.; Li, Z.; Huang, B. Transcriptomic Analysis of Eruca Vesicaria Subs. Sativa Lines with Contrasting Tolerance to Polyethylene Glycol-Simulated Drought Stress. BMC Plant Biol 2019, 19, 419. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Liu, X.; Li, W.; Wang, X.; Zhong, X.; Gao, Y.; Xu, H.; Hu, H.; Zhang, L.; Cheng, X.; et al. The Regulatory Metabolic Networks of the Brassica Campestris L. Hairy Roots in Response to Cadmium Stress Revealed from Proteome Studies Combined with a Transcriptome Analysis. Ecotoxicol Environ Saf 2023, 263, 115214. [Google Scholar] [CrossRef]
- Dai, H. -f.; Jiang, B.; Zhao, J. -s.; Li, J. -c.; Sun, Q. -m. Metabolomics and Transcriptomics Analysis of Pollen Germination Response to Low-Temperature in Pitaya (Hylocereus Polyrhizus). Front Plant Sci 2022, 13, 1–15. [Google Scholar] [CrossRef]
- Watkins, J.L.; Li, Q.; Yeaman, S.; Facchini, P.J. Elucidation of the Mescaline Biosynthetic Pathway in Peyote (Lophophora Williamsii). The Plant Journal 2023, 116, 635–649. [Google Scholar] [CrossRef]
- Li, J.; Zhen, W.; Long, D.; Ding, L.; Gong, A.; Xiao, C.; Jiang, W.; Liu, X.; Zhou, T.; Huang, L. De Novo Sequencing and Assembly Analysis of the Pseudostellaria Heterophylla Transcriptome. PLoS One 2016, 11, e0164235. [Google Scholar] [CrossRef]
- Liu, E.; Xu, L.; Luo, Z.; Li, Z.; Zhou, G.; Gao, H.; Fang, F.; Tang, J.; Zhao, Y.; Zhou, Z.; et al. Transcriptomic Analysis Reveals Mechanisms for the Different Drought Tolerance of Sweet Potatoes. Front Plant Sci 2023, 14, 1136709. [Google Scholar] [CrossRef]
- Zeng, W.; Sun, Z.; Lai, Z.; Yang, S.; Chen, H.; Yang, X.; Tao, J.; Tang, X. Determination of the MiRNAs Related to Bean Pyralid Larvae Resistance in Soybean Using Small RNA and Transcriptome Sequencing. Int J Mol Sci 2019, 20, 2966. [Google Scholar] [CrossRef] [PubMed]
- Chacon, D.S.; Torres, T.; da Silva, I.; de Araújo TF; Roque, A.; Pinheiro, F.; Selegato, D.; Pilon, A.; Reginaldo, F.; da Costa, C.; et al. Erythrina Velutina Willd. Alkaloids: Piecing Biosynthesis Together from Transcriptome Analysis and Metabolite Profiling of Seeds and Leaves. J Adv Res 2021, 34, 123–136. [CrossRef]
- Zhao, X.; Li, X.; Bao, A.; Zhang, X.; Xu, Y.; Li, Y. Metabolome and Transcriptome Integration Explored the Mechanism of Browning in Glycyrrhiza Uralensis Fisch Cells. Front Plant Sci 2024, 15. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Luo, Y.; Wang, J.; Ji, T.; Yuan, L.; Kai, G. Integrated Analysis of the Transcriptome, Metabolome and Analgesic Effect Provide Insight into Potential Applications of Different Parts of Lindera Aggregata. Food Research International 2020, 138, 109799. [Google Scholar] [CrossRef]
- Han, H.; Zhang, L.; Li, S.; Zhao, R.; Wang, F.; Dong, R.; Wang, X. Transcriptome and Metabolome Integrated Analysis Reveals the Mechanism of Cinnamomum Bodinieri Root Response to Alkali Stress. Plant Mol Biol Report 2023, 41, 470–488. [Google Scholar] [CrossRef]
- Cheng, X.; Li, D.; Jiang, Z.; Qu, C.; Yan, H.; Wu, Q. Metabolite Profiling and Transcriptomic Analyses Demonstrate the Effects of Biocontrol Agents on Alkaloid Accumulation in Fritillaria Thunbergii. BMC Plant Biol 2023, 23, 435. [Google Scholar] [CrossRef]
- Cheng, W.; Yao, Y.; Wang, Q.; Chang, X.; Shi, Z.; Fang, X.; Chen, F.; Chen, S.; Zhang, Y.; Zhang, F.; et al. Characterization of Benzylisoquinoline Alkaloid Methyltransferases in Liriodendron Chinense Provides Insights into the Phylogenic Basis of Angiosperm Alkaloid Diversity. The Plant Journal 2022, 112, 535–548. [Google Scholar] [CrossRef]
- Liu, J.; Han, L.; Li, G.; Zhang, A.; Liu, X.; Zhao, M. Transcriptome and Metabolome Profiling of the Medicinal Plant Veratrum Mengtzeanum Reveal Key Components of the Alkaloid Biosynthesis. Front Genet 2023, 14, 1023433. [Google Scholar] [CrossRef]
- Zhang, Y.; Kang, Y.; Xie, H.; Wang, Y.; Li, Y.; Huang, J. Comparative Transcriptome Analysis Reveals Candidate Genes Involved in Isoquinoline Alkaloid Biosynthesis in Stephania Tetrandra. Planta Med 2020, 86, 1258–1268. [Google Scholar] [CrossRef]
- Li, K.; Chen, X.; Zhang, J.; Wang, C.; Xu, Q.; Hu, J.; Kai, G.; Feng, Y. Transcriptome Analysis of Stephania Tetrandra and Characterization of Norcoclaurine-6-O-Methyltransferase Involved in Benzylisoquinoline Alkaloid Biosynthesis. Front Plant Sci 2022, 13, 874583. [Google Scholar] [CrossRef]
- Huang, H.; Wei, Y.; Huang, S.; Lu, S.; Su, H.; Ma, L.; Huang, W. Integrated Metabolomic and Transcriptomic Analyses Provide Insights into Regulation Mechanisms during Bulbous Stem Development in the Chinese Medicinal Herb Plant, Stephania Kwangsiensis. BMC Plant Biol 2024, 24, 276. [Google Scholar] [CrossRef]
- Yang, M.; Zhu, L.; Li, L.; Li, J.; Xu, L.; Feng, J.; Liu, Y. Digital Gene Expression Analysis Provides Insight into the Transcript Profile of the Genes Involved in Aporphine Alkaloid Biosynthesis in Lotus (Nelumbo Nucifera). Front Plant Sci 2017, 8. [Google Scholar] [CrossRef]
- Sun, H.; Song, H.; Deng, X.; Liu, J.; Yang, D.; Zhang, M.; Wang, Y.; Xin, J.; Chen, L.; Liu, Y.; et al. Transcriptome-Wide Characterization of Alkaloids and Chlorophyll Biosynthesis in Lotus Plumule. Front Plant Sci 2022, 13. [Google Scholar] [CrossRef]
- Li, Y.; Shi, L.-C.; Cushman, S.A. Transcriptomic Responses and Physiological Changes to Cold Stress among Natural Populations Provide Insights into Local Adaptation of Weeping Forsythia. Plant Physiology and Biochemistry 2021, 165, 94–103. [Google Scholar] [CrossRef]
- Jiao, C.; Wei, M.; Fan, H.; Song, C.; Wang, Z.; Cai, Y.; Jin, Q. Transcriptomic Analysis of Genes Related to Alkaloid Biosynthesis and the Regulation Mechanism under Precursor and Methyl Jasmonate Treatment in Dendrobium Officinale. Front Plant Sci 2022, 13. [Google Scholar] [CrossRef]
- Desgagné-Penix, I.; Khan, M.F.; Schriemer, D.C.; Cram, D.; Nowak, J.; Facchini, P.J. Integration of Deep Transcriptome and Proteome Analyses Reveals the Components of Alkaloid Metabolism in Opium Poppy Cell Cultures. BMC Plant Biol 2010, 10, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Priyal, A.; Tripathi, H.; Yadav, D.K.; Khan, F.; Gupta, V.; Shukla, R.K.; Darokar, M.P. Functional Annotation of Expressed Sequence Tags of Papaver Somniferum. Plant Omics 2012, 5. [Google Scholar]
- Zeng, J.; Liu, Y.; Liu, W.; Liu, X.; Liu, F.; Huang, P.; Zhu, P.; Chen, J.; Shi, M.; Guo, F.; et al. Integration of Transcriptome, Proteome and Metabolism Data Reveals the Alkaloids Biosynthesis in Macleaya Cordata and Macleaya Microcarpa. PLoS One 2013, 8, e53409. [Google Scholar] [CrossRef]
- Pathak, S.; Lakhwani, D.; Gupta, P.; Mishra, B.K.; Shukla, S.; Asif, M.H.; Trivedi, P.K. Comparative Transcriptome Analysis Using High Papaverine Mutant of Papaver Somniferum Reveals Pathway and Uncharacterized Steps of Papaverine Biosynthesis. PLoS One 2013, 8, e65622. [Google Scholar] [CrossRef] [PubMed]
- Liao, D.; Wang, P.; Jia, C.; Sun, P.; Qi, J.; Zhou, L.; Li, X. Identification and Developmental Expression Profiling of Putative Alkaloid Biosynthetic Genes in Corydalis Yanhusuo Bulbs. Sci Rep 2016, 6, 19460. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, P.; Pathak, S.; Lakhwani, D.; Gupta, P.; Asif, M.H.; Trivedi, P.K. Comparative Analysis of Transcription Factor Gene Families from Papaver Somniferum: Identification of Regulatory Factors Involved in Benzylisoquinoline Alkaloid Biosynthesis. Protoplasma 2016, 253, 857–871. [Google Scholar] [CrossRef] [PubMed]
- He, S.M.; Song, W.L.; Cong, K.; Wang, X.; Dong, Y.; Cai, J.; Zhang, J.J.; Zhang, G.H.; Yang, J.L.; Yang, S.C.; et al. Identification of Candidate Genes Involved in Isoquinoline Alkaloids Biosynthesis in Dactylicapnos Scandens by Transcriptome Analysis. Sci Rep 2017, 7, 9119. [Google Scholar] [CrossRef]
- Liu, X.; Liu, Y.; Huang, P.; Ma, Y.; Qing, Z.; Tang, Q.; Cao, H.; Cheng, P.; Zheng, Y.; Yuan, Z.; et al. The Genome of Medicinal Plant Macleaya Cordata Provides New Insights into Benzylisoquinoline Alkaloids Metabolism. Mol Plant 2017, 10, 975–989. [Google Scholar] [CrossRef]
- Kim, D.; Jung, M.; Ha, I.J.; Lee, M.Y.; Lee, S.-G.; Shin, Y.; Subramaniyam, S.; Oh, J. Transcriptional Profiles of Secondary Metabolite Biosynthesis Genes and Cytochromes in the Leaves of Four Papaver Species. Data (Basel) 2018, 3, 55. [Google Scholar] [CrossRef]
- Oh, J.; Shin, Y.; Ha, I.J.; Lee, M.Y.; Lee, S.-G.; Kang, B.-C.; Kyeong, D.; Kim, D. Transcriptome Profiling of Two Ornamental and Medicinal Papaver Herbs. Int J Mol Sci 2018, 19, 3192. [Google Scholar] [CrossRef]
- Guo, L.; Winzer, T.; Yang, X.; Li, Y.; Ning, Z.; He, Z.; Teodor, R.; Lu, Y.; Bowser, T.A.; Graham, I.A.; et al. The Opium Poppy Genome and Morphinan Production. Science 2018, 362, 343–347. [Google Scholar] [CrossRef]
- Pourmazaheri, H.; Soorni, A.; Baghban Kohnerouz, B.; Khosravi Dehaghi, N.; Kalantar, E.; Omidi, M.; Naghavi, M.R. Comparative Analysis of the Root and Leaf Transcriptomes in Chelidonium Majus L. PLoS One 2019, 14, e0215165. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, Z.; Li, M.; Luo, J.; Chen, F.; Gong, Y.; Li, Y.; Wei, Y.; Su, Y.; Kong, L. Transcriptomic Profiles of 33 Opium Poppy Samples in Different Tissues, Growth Phases, and Cultivars. Sci Data 2019, 6, 66. [Google Scholar] [CrossRef]
- Yamada, Y.; Nishida, S.; Shitan, N.; Sato, F. Genome-Wide Identification of AP2/ERF Transcription Factor-Encoding Genes in California Poppy (Eschscholzia Californica) and Their Expression Profiles in Response to Methyl Jasmonate. Sci Rep 2020, 10, 18066. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Lin, H.; Tang, Y.; Huang, L.; Xu, J.; Nian, S.; Zhao, Y. Integration of Full-Length Transcriptomics and Targeted Metabolomics to Identify Benzylisoquinoline Alkaloid Biosynthetic Genes in Corydalis Yanhusuo. Hortic Res 2021, 8, 16. [Google Scholar] [CrossRef]
- Pei, L.; Wang, B.; Ye, J.; Hu, X.; Fu, L.; Li, K.; Ni, Z.; Wang, Z.; Wei, Y.; Shi, L.; et al. Genome and Transcriptome of Papaver Somniferum Chinese Landrace CHM Indicates That Massive Genome Expansion Contributes to High Benzylisoquinoline Alkaloid Biosynthesis. Hortic Res 2021, 8, 5. [Google Scholar] [CrossRef]
- Catania, T.; Li, Y., W. T.; Harvey, D.; Meade, F.; Caridi, A.; Leech, A.; Larson, T.R.; Ning, Z.; Chang, J.; Van de Peer, Y.; et al. A Functionally Conserved STORR Gene Fusion in Papaver Species That Diverged 16.8 Million Years Ago. Nat Commun 2022, 13, 3150. [Google Scholar] [CrossRef]
- George, S.; Manoharan, D.; Li, J.; Britton, M.; Parida, A. Drought and Salt Stress in Chrysopogon Zizanioides Leads to Common and Specific Transcriptomic Responses and May Affect Essential Oil Composition and Benzylisoquinoline Alkaloids Metabolism. Curr Plant Biol 2017, 11–12, 12–22. [Google Scholar] [CrossRef]
- Li, B.; Zhang, P.; Wang, F.; Li, R.; Liu, J.; Wang, Q.; Liu, W.; Wang, B.; Hu, G. Integrated Analysis of the Transcriptome and Metabolome Revealed Candidate Genes Involved in GA3-Induced Dormancy Release in Leymus Chinensis Seeds. Int J Mol Sci 2021, 22, 4161. [Google Scholar] [CrossRef]
- Pal, T.; Malhotra, N.; Chanumolu, S.K.; Chauhan, R.S. Next-Generation Sequencing (NGS) Transcriptomes Reveal Association of Multiple Genes and Pathways Contributing to Secondary Metabolites Accumulation in Tuberous Roots of Aconitum Heterophyllum Wall. Planta 2015, 242, 239–258. [Google Scholar] [CrossRef]
- He, S.M.; Liang, Y.L.; Cong, K.; Chen, G.; Zhao, X.; Zhao, Q.M.; Zhang, J.J.; Wang, X.; Dong, Y.; Yang, J.L.; et al. Identification and Characterization of Genes Involved in Benzylisoquinoline Alkaloid Biosynthesis in Coptis Species. Front Plant Sci 2018, 9, 328684. [Google Scholar] [CrossRef]
- Shen, Y.; Xu, S.; Mengyan, W.; Yongbin, T.; Xu, C.; Hailey, H.; Kai, T.; Lan, M.; Ji, C.; Weiping, Z.; et al. Systematic Estimation of Potential Risk Caused by the Replacement of Aconite’s Cultivar. Pharmacogn Mag 2019, 15, 722. [Google Scholar] [CrossRef]
- Zhong, F.; Huang, L.; Qi, L.; Ma, Y.; Yan, Z. Full-Length Transcriptome Analysis of Coptis Deltoidea and Identification of Putative Genes Involved in Benzylisoquinoline Alkaloids Biosynthesis Based on Combined Sequencing Platforms. Plant Mol Biol 2020, 102, 477–499. [Google Scholar] [CrossRef] [PubMed]
- Hao, D.C.; Li, P.; Xiao, P.G.; He, C.N. Dissection of Full-Length Transcriptome and Metabolome of Dichocarpum (Ranunculaceae): Implications in Evolution of Specialized Metabolism of Ranunculales Medicinal Plants. PeerJ 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.M.; Tan, J.P.; Cheng, S.Y.; Chen, Z.X.; Ye, J.B.; Zheng, J.R.; Xu, F.; Zhang, W.W.; Liao, Y.L.; Yang, X.Y. Comparative Transcriptome Analysis Provides Novel Insights into the Molecular Mechanism of Berberine Biosynthesis in Coptis Chinensis. Sci Hortic 2022, 291, 110585. [Google Scholar] [CrossRef]
- Duan, Y.; You, J.; Wang, J.; Tang, T.; Guo, X.; Wang, F.; Wang, X.; Mu, S.; Wang, Q.; Niu, X.; et al. Transcriptome Analysis Reveals the Potential Mechanism of Altering Viability, Yield, and Isoquinoline Alkaloids in Coptis Chinensis through Cunninghamia Lanceolata Understory Cultivation. Chemical and Biological Technologies in Agriculture 2024, 11, 1–15. [Google Scholar] [CrossRef]
- Li, X.; Xu, B.; Sahito, Z.A.; Chen, S.; Liang, Z. Transcriptome Analysis Reveals Cadmium Exposure Enhanced the Isoquinoline Alkaloid Biosynthesis and Disease Resistance in Coptis Chinensis. Ecotoxicol Environ Saf 2024, 271. [Google Scholar] [CrossRef]
- Zhong, F.; Ke, W.; Li, Y.; Chen, X.; Zhou, T.; Xu, B.; Qi, L.; Yan, Z.; Ma, Y. Comprehensive Analysis of the Complete Mitochondrial Genomes of Three Coptis Species (C. Chinensis, C. Deltoidea and C. Omeiensis): The Important Medicinal Plants in China. Front Plant Sci 2023, 14, 1166420. [Google Scholar] [CrossRef]
- Huang, P.; Li, Z.; Wang, H.; Huang, J.; Tan, G.; Fu, Y.; Liu, X.; Zheng, S.; Xu, P.; Sun, M.; et al. A Genome Assembly of Decaploid Houttuynia Cordata Provides Insights into the Evolution of Houttuynia and the Biosynthesis of Alkaloids. Hortic Res 2024, 11, uhae203. [Google Scholar] [CrossRef]
- Yang, C.; Wu, P.; Yao, X.; Sheng, Y.; Zhang, C.; Lin, P.; Wang, K. Integrated Transcriptome and Metabolome Analysis Reveals Key Metabolites Involved in Camellia Oleifera Defense against Anthracnose. Int J Mol Sci 2022, 23, 536. [Google Scholar] [CrossRef]
- Miao, Y.; Li, H.; Pan, J.; Zhou, B.; He, T.; Wu, Y.; Zhou, D.; He, W.; Chen, L. Salicylic Acid Modulates Secondary Metabolism and Enhanced Colchicine Accumulation in Long Yellow Daylily (Hemerocallis Citrina). AoB Plants 2024, 16, plae029. [Google Scholar] [CrossRef]
- Guo, J.; Huang, Z.; Sun, J.; Cui, X.; Liu, Y. Research Progress and Future Development Trends in Medicinal Plant Transcriptomics. Front Plant Sci 2021, 12, 691838. [Google Scholar] [CrossRef]
- Min, X.; Zhu, T.; Hu, X.; Hou, C.; He, J.; Liu, X. Transcriptome and Metabolome Analysis of Isoquinoline Alkaloid Biosynthesis of Coptis Chinensis in Different Years. Genes (Basel) 2023, 14, 2232. [Google Scholar] [CrossRef]
- Ikezawa, N.; Tanaka, M.; Nagayoshi, M.; Shinkyo, R.; Sakaki, T.; Inouye, K.; Sato, F. Molecular Cloning and Characterization of CYP719, a Methylenedioxy Bridge-Forming Enzyme That Belongs to a Novel P450 Family, from Cultured Coptis Japonica Cells. Journal of Biological Chemistry 2003, 278, 38557–38565. [Google Scholar] [CrossRef] [PubMed]
- Ikezawa, N.; Iwasa, K.; Sato, F. Molecular Cloning and Characterization of CYP80G2, a Cytochrome P450 That Catalyzes an Intramolecular C-C Phenol Coupling of (S)-Reticuline in Magnoflorine Biosynthesis, from Cultured Coptis Japonica Cells. Journal of Biological Chemistry 2008, 283, 8810–8821. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Park, N. Il; Park, Y.; Park, K.C.; Kim, E.S.; Son, Y.K.; Choi, B.S.; Kim, N.S.; Choi, I.Y. O- and N-Methyltransferases in Benzylisoquinoline Alkaloid Producing Plants. Genes Genomics 2024, 46, 367–378. [Google Scholar] [CrossRef]
- Facchini, P.J.; Morris, J.S. Molecular Origins of Functional Diversity in Benzylisoquinoline Alkaloid Methyltransferases. Front Plant Sci 2019, 10, 1058. [Google Scholar] [CrossRef]
- Li, Q.; Xu, J.; Zheng, Y.; Zhang, Y.; Cai, Y. Transcriptomic and Metabolomic Analyses Reveals That Exogenous Methyl Jasmonate Regulates Galanthamine Biosynthesis in Lycoris Longituba Seedlings. Front Plant Sci 2021, 12, 713795. [Google Scholar] [CrossRef] [PubMed]
- Nett, R.S.; Lau, W.; Sattely, E.S. Discovery and Engineering of Colchicine Alkaloid Biosynthesis. Nature 2020, 584, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Ounaroon, A.; Decker, G.; Schmidt, J.; Lottspeich, F.; Kutchan, T.M. (R,S)-Reticuline 7-O-Methyltransferase and (R,S)-Norcoclaurine 6-O-Methyltransferase of Papaver Somniferum – CDNA Cloning and Characterization of Methyl Transfer Enzymes of Alkaloid Biosynthesis in Opium Poppy. The Plant Journal 2003, 36, 808–819. [Google Scholar] [CrossRef]
- Guo, D.; Kong, S.; Sun, Y.; Li, X.; Pan, H. Development of an Artificial Biosynthetic Pathway for Biosynthesis of (S)-Reticuline Based on HpaBC in Engineered Escherichia Coli. Biotechnol Bioeng 2021, 118, 4635–4642. [Google Scholar] [CrossRef]
- Wang, R.; Xu, S.; Jiang, Y.; Jiang, J.; Li, X.; Liang, L.; He, J.; Peng, F.; Xia, B. De Novo Sequence Assembly and Characterization of Lycoris Aurea Transcriptome Using GS FLX Titanium Platform of 454 Pyrosequencing. PLoS One 2013, 8, e60449. [Google Scholar] [CrossRef]
- Karimzadegan, V.; Koirala, M.; Sobhanverdi, S.; Merindol, N.; Majhi, B.B.; Gélinas, S.E.; Timokhin, V.I.; Ralph, J.; Dastmalchi, M.; Desgagné-Penix, I. Characterization of Cinnamate 4-Hydroxylase (CYP73A) and p-Coumaroyl 3′-Hydroxylase (CYP98A) from Leucojum Aestivum, a Source of Amaryllidaceae Alkaloids. Plant Physiology and Biochemistry 2024, 210, 108612. [Google Scholar] [CrossRef]
- Koirala, M.; Cristine Goncalves dos Santos, K.; Gélinas, S.E.; Ricard, S.; Karimzadegan, V.; Lamichhane, B.; Sameera Liyanage, N.; Merindol, N.; Desgagné-Penix, I. Auxin and Light-Mediated Regulation of Growth, Morphogenesis, and Alkaloid Biosynthesis in Crinum x Powellii “Album” Callus. Phytochemistry 2023, 216. [Google Scholar] [CrossRef]
- Tousignant, L.; Diaz-Garza, A.M.; Majhi, B.B.; Gélinas, S.-E.; Singh, A.; Desgagne-Penix, I. Transcriptome Analysis of Leucojum Aestivum and Identification of Genes Involved in Norbelladine Biosynthesis. Planta 2022, 255, 30. [Google Scholar] [CrossRef]
- Xu, S.; Jiang, Y.; Wang, N.; Xia, B.; Jiang, Y.; Li, X.; Zhang, Z.; Li, Y.; Wang, R. Identification and Differential Regulation of MicroRNAs in Response to Methyl Jasmonate Treatment in Lycoris Aurea by Deep Sequencing. BMC Genomics 2016, 17, 789. [Google Scholar] [CrossRef]
- Ma, R.; Xu, S.; Zhao, Y.; Xia, B.; Wang, R. Selection and Validation of Appropriate Reference Genes for Quantitative Real-Time PCR Analysis of Gene Expression in Lycoris Aurea. Front Plant Sci 2016, 7, 536. [Google Scholar] [CrossRef]
- Wang, R.; Xu, S.; Wang, N.; Xia, B.; Jiang, Y.; Wang, R. Transcriptome Analysis of Secondary Metabolism Pathway, Transcription Factors, and Transporters in Response to Methyl Jasmonate in Lycoris Aurea. Front Plant Sci 2017, 7. [Google Scholar] [CrossRef]
- Zhou, Z.; Wu, M.; Sun, B.; Li, J.; Li, J.; Liu, Z.; Gao, M.; Xue, L.; Xu, S.; Wang, R. Identification of Transcription Factor Genes Responsive to MeJA and Characterization of a LaMYC2 Transcription Factor Positively Regulates Lycorine Biosynthesis in Lycoris Aurea. J Plant Physiol 2024, 296. [Google Scholar] [CrossRef]
- Zhang, F.; Cheng, G.; Shu, X.; Wang, N.; Wang, Z. Transcriptome Analysis of Lycoris Chinensis Bulbs Reveals Flowering in the Age-Mediated Pathway. Biomolecules 2022, 12, 899. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Xu, J.; Yang, L.; Zhou, X.; Cai, Y.; Zhang, Y. Transcriptome Analysis of Different Tissues Reveals Key Genes Associated With Galanthamine Biosynthesis in Lycoris Longituba. Front Plant Sci 2020, 11, 519752. [Google Scholar] [CrossRef] [PubMed]
- Park, C.H.; Yeo, H.J.; Park, Y.E.; Baek, S.A.; Kim, J.K.; Park, S.U. Transcriptome Analysis and Metabolic Profiling of Lycoris Radiata. Biology (Basel) 2019, 8, 63. [Google Scholar] [CrossRef]
- Hotchandani, T.; de Villers, J.; Desgagné-Penix, I. Developmental Regulation of the Expression of Amaryllidaceae Alkaloid Biosynthetic Genes in Narcissus Papyraceus. Genes (Basel) 2019, 10, 594. [Google Scholar] [CrossRef]
- Singh, A.; Desgagné-Penix, I. Transcriptome and Metabolome Profiling of Narcissus Pseudonarcissus “King Alfred” Reveal Components of Amaryllidaceae Alkaloid Metabolism. Sci Rep 2017, 7, 1–14. [Google Scholar] [CrossRef]
- Ferdausi, A.; Chang, X.; Jones, M. Transcriptomic Analysis for Differential Expression of Genes Involved in Secondary Metabolite Production in Narcissus Pseudonarcissus Field Derived Bulb and in Vitro Callus. Ind Crops Prod 2021, 168, 113615. [Google Scholar] [CrossRef]
- Kilgore, M.B.; Holland, C.K.; Jez, J.M.; Kutchan, T.M. Identification of a Noroxomaritidine Reductase with Amaryllidaceae Alkaloid Biosynthesis Related Activities. Journal of Biological Chemistry 2016, 291, 16740–16752. [Google Scholar] [CrossRef] [PubMed]
- Mu, H.M.; Wang, R.; Li, X.D.; Jiang, Y.M.,F.P. ; Xia, B. Alkaloid Accumulation in Different Parts and Ages of Lycoris Chinensis. Zeitschrift für Naturforschung C 2010, 65, 458–462. [Google Scholar] [CrossRef]
- Wang, R.; Han, X.; Xu, S.; Xia, B.; Jiang, Y.; Xue, Y.; Wang, R. Cloning and Characterization of a Tyrosine Decarboxylase Involved in the Biosynthesis of Galanthamine in Lycoris Aurea. PeerJ 2019, 7, e6729. [Google Scholar] [CrossRef] [PubMed]
- Youssef D. T., A.; Shaala, L.A.; Altyar, A.E. Cytotoxic Phenylpropanoid Derivatives and Alkaloids from the Flowers of Pancratium Maritimum L. Plants 2022, 11, 476. [Google Scholar] [CrossRef]
- Youssef, D.T.A. Cytotoxic Phenolics from the Flowers of Hippeastrum Vittatum. Bulletin of Pharmaceutical Sciences. Assiut 2005, 28, 143–148. [Google Scholar] [CrossRef]
- Takos, A.M.; Rook, F. Towards a Molecular Understanding of the Biosynthesis of Amaryllidaceae Alkaloids in Support of Their Expanding Medical Use. Int J Mol Sci 2013, 14, 11713–11741. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Luo, T.; Li, J.; Li, G.; Zhou, D.; Liu, T.; Zou, X.; Pandey, A.; Luo, Z. Transcriptomics-Based Identification and Characterization of 11 CYP450 Genes of Panax Ginseng Responsive to MeJA. Acta Biochim Biophys Sin (Shanghai) 2018, 50, 1094–1103. [Google Scholar] [CrossRef]
- Wang, WF.; Xiao, H.; Zhong, JJ. Biosynthesis of a Novel Ganoderic Acid by Expressing CYP Genes from Ganoderma Lucidum in Saccharomyces Cerevisiae. Appl Microbiol Biotechnol 2022, 106, 523–534. [Google Scholar] [CrossRef]
- Sun, J.; Zhan, X.; Wang, W.; Yang, X.; Liu, Y.; Yang, H.; Deng, J.; Yang, H. Natural Aporphine Alkaloids: A Comprehensive Review of Phytochemistry, Pharmacokinetics, Anticancer Activities, and Clinical Application. J Adv Res 2024, 63, 231–253. [Google Scholar] [CrossRef]
- Zetzsche, L.E.; Narayan, A.R.H. Broadening the Scope of Biocatalytic C–C Bond Formation. Nat Rev Chem 2020, 4, 334–346. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Qiao, C.; Pang, J.; Zhang, G.; Luo, Y. The Versatile O-Methyltransferase LrOMT Catalyzes Multiple O-Methylation Reactions in Amaryllidaceae Alkaloids Biosynthesis. Int J Biol Macromol 2019, 141, 680–692. [Google Scholar] [CrossRef]
- Pervez, M.T.; Ul Hasnain, M.J.; Abbas, S.H.; Moustafa, M.F.; Aslam, N.; Shah, S.S.M. A Comprehensive Review of Performance of Next-Generation Sequencing Platforms. Biomed Res Int 2022, e3457806. [Google Scholar] [CrossRef]
- Purushothaman, S.; Meola, M.; Egli, A. Combination of Whole Genome Sequencing and Metagenomics for Microbiological Diagnostics. International Journal of Molecular Sciences 2022, Vol. 23, Page 9834 2022, 23, 9834. [Google Scholar] [CrossRef]
- Song, B.; Buckler, E.S.; Stitzer, M.C. New Whole-Genome Alignment Tools Are Needed for Tapping into Plant Diversity. Trends Plant Sci 2024, 29, 355–369. [Google Scholar] [CrossRef]
- Sun, Y.; Shang, L.; Zhu, Q.H.; Fan, L.; Guo, L. Twenty Years of Plant Genome Sequencing: Achievements and Challenges. Trends Plant Sci 2022, 27, 391–401. [Google Scholar] [CrossRef]
- Kong, W.; Wang, Y.; Zhang, S.; Yu, J.; Zhang, X. Recent Advances in Assembly of Complex Plant Genomes. Genomics Proteomics Bioinformatics 2023, 21, 427–439. [Google Scholar] [CrossRef] [PubMed]
- Schinkel, C.C.F.; Kirchheimer, B.; Dullinger, S.; Geelen, D.; De Storme, N.; Hörandl, E. Pathways to Polyploidy: Indications of a Female Triploid Bridge in the Alpine Species Ranunculus Kuepferi (Ranunculaceae). Plant Systematics and Evolution 2017, 303, 1093–1108. [Google Scholar] [CrossRef] [PubMed]
- Meerow, A.W.; Gardner, E.M.; Nakamura, K. Phylogenomics of the Andean Tetraploid Clade of the American Amaryllidaceae (Subfamily Amaryllidoideae): Unlocking a Polyploid Generic Radiation Abetted by Continental Geodynamics. Front Plant Sci 2020, 11, 582422. [Google Scholar] [CrossRef]
- Fossati, E.; Narcross, L.; Ekins, A.; Falgueyret, J.-P.; Martin, VJJ. Synthesis of Morphinan Alkaloids in Saccharomyces Cerevisiae. PLoS One 2015, 10, e0124459. [CrossRef]
- Thodey K., G.S. ; Smolke, C.D. A Microbial Biomanufacturing Platform for Natural and Semisynthetic Opioids. Nat Chem Biol 2014, 10, 837–844. [Google Scholar] [CrossRef] [PubMed]
- Narcross, L.; Fossati, E.; Bourgeois, L.; Dueber, J.E.; Martin, V.J. Microbial Factories for the Production of Benzylisoquinoline Alkaloids. Trends Biotechnol 2016, 34, 228–241. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; et al. Bioinspired Scalable Total Synthesis of Opioids. CCL Chemistry 2021, 13, 371–380. [Google Scholar] [CrossRef]
- Li, X. ; others High-Efficiency Biocatalytic Conversion of Thebaine to Codeine. ACS Omega 2020, 5, 9339–9347. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Some representative skeletons of alkaloid diversity: (1) Indole, (2) Quinoline, (3) Isoquinoline, (4) Pyrrolizidine, (5) Tropane, (6) Imidazole, (7) Pyrroline, (8) Pyrrole, (9) Piperidine, and (10) Pyridine. Adapted from Singh, Poonam, and Geeta (2019). Narcissus pseudonarcissus (Amaryllidaceae). Created in BioRender.com.
Figure 1.
Some representative skeletons of alkaloid diversity: (1) Indole, (2) Quinoline, (3) Isoquinoline, (4) Pyrrolizidine, (5) Tropane, (6) Imidazole, (7) Pyrroline, (8) Pyrrole, (9) Piperidine, and (10) Pyridine. Adapted from Singh, Poonam, and Geeta (2019). Narcissus pseudonarcissus (Amaryllidaceae). Created in BioRender.com.
Figure 2.
Phylogenetic comparison between the main families producing IsAs and AmAs. The cladogram shows the relationship between key orders and families, highlighting Ranunculales and Asparagales, At the top, representative alkaloid structures from Ranunculales and Asparagales, At the top, representative alkaloid structures from Ranunculales are represented: (1) Morphine, (2) Isoboldine, (3) Corytuberine. At the bottom, examples of representative AmAs are shown, such as (4) Hemanthamine, (5) Galanthamine and (6) Lycorine. The flowers are Hepatica nobilis (Ranunculaceae) and Zephyranthes carinata (Amaryllidaceae). Created in BioRender.com.
Figure 2.
Phylogenetic comparison between the main families producing IsAs and AmAs. The cladogram shows the relationship between key orders and families, highlighting Ranunculales and Asparagales, At the top, representative alkaloid structures from Ranunculales and Asparagales, At the top, representative alkaloid structures from Ranunculales are represented: (1) Morphine, (2) Isoboldine, (3) Corytuberine. At the bottom, examples of representative AmAs are shown, such as (4) Hemanthamine, (5) Galanthamine and (6) Lycorine. The flowers are Hepatica nobilis (Ranunculaceae) and Zephyranthes carinata (Amaryllidaceae). Created in BioRender.com.
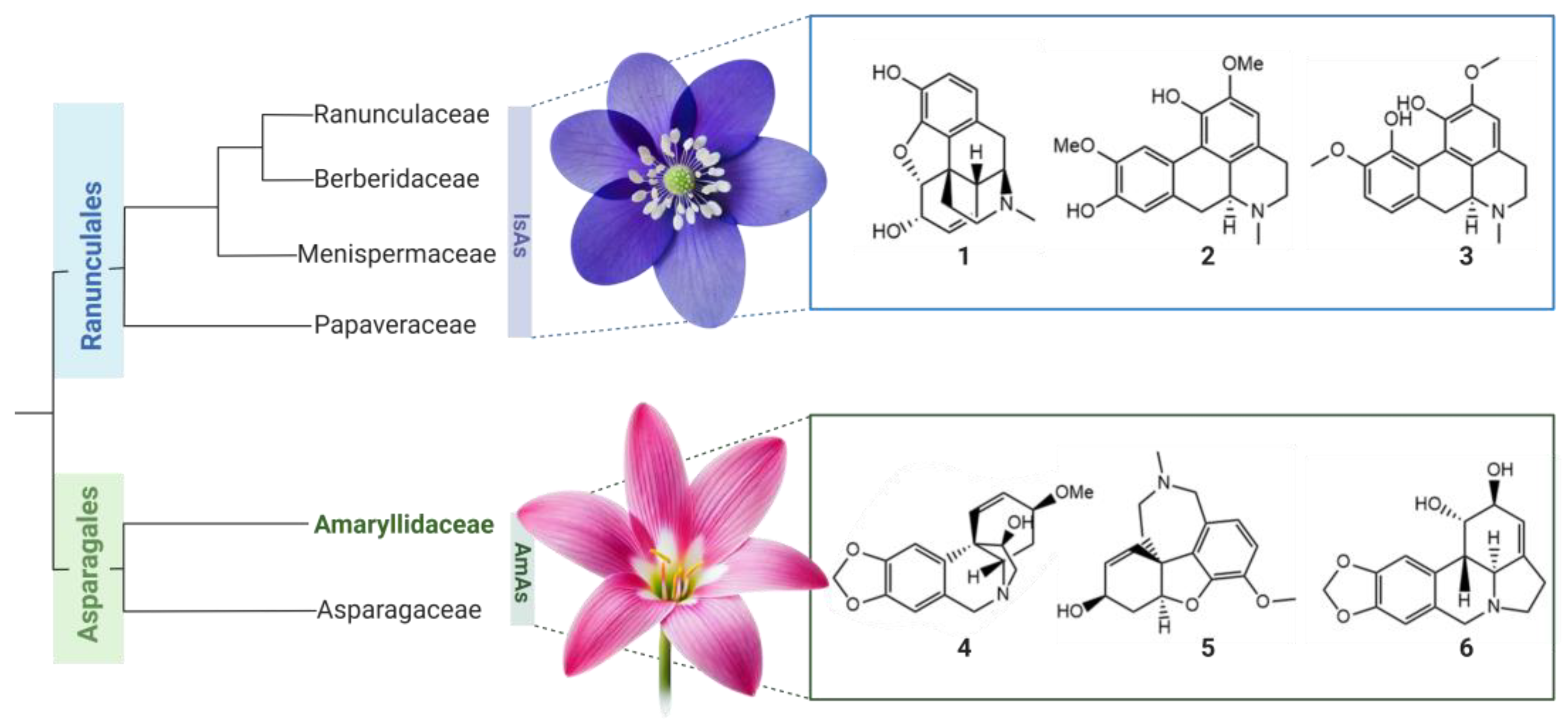
Figure 3.
IsAs and AmAs biosynthetic pathways comparation. The IsAs pathway is highlighted in blue, while the AmAs pathway is depicted in green. Solid lines indicate enzymatic reactions catalyzed by a single enzyme, whereas dashed lines represent multi-step reactions catalyzed by several enzymes. The structures of key intermediates are displayed, highlighting the specific carbon atoms involved in phenol-coupling reactions with distinct colors. 4-HPP; 4-hydroxyphenylpyruvate, 4-HPAA; 4-Hydroxyphenylacetaldehyde, L-DOPA; 3,4-Dihydroxy-L-phenylalanine, 3,4-DHPA; 3,4-Dihydroxyphenylacetaldehyde, 3,4-DHBA; 3,4-dihydroxybenzaldehyde, 4’OMET; 4’-O-methylnorbelladine. TAT; Tyrosine aminotransferase, TH; Tyrosine hydroxylase, AADC; Aromatic L-amino acid decarboxylase, TYDC; Tyrosine decarboxylase, PAL; Phenylalanine ammonia lyase, C4H; Trans-cinnamate 4-monooxygenase, 4CL: Coumarate-4-lyase, C3H: 4-coumarate 3-hydroxylase (ascorbate peroxidase), HCT; hydroxycinnamoyl transferase, C3´H; p-coumaroyl shikimate 3´-hydroxylase, CSE; Caffeoyl shikimate esterase, HBS; Hydroxy Benzaldehyde synthase, NBS: Norbelladine synthase (coupled with norcraugsodine/noroxomaritidine reductase – NR), NCS; Norcoclaurine synthase, CYP96: Family of enzymes capable of phenolic couplings. Created in BioRender.com.
Figure 3.
IsAs and AmAs biosynthetic pathways comparation. The IsAs pathway is highlighted in blue, while the AmAs pathway is depicted in green. Solid lines indicate enzymatic reactions catalyzed by a single enzyme, whereas dashed lines represent multi-step reactions catalyzed by several enzymes. The structures of key intermediates are displayed, highlighting the specific carbon atoms involved in phenol-coupling reactions with distinct colors. 4-HPP; 4-hydroxyphenylpyruvate, 4-HPAA; 4-Hydroxyphenylacetaldehyde, L-DOPA; 3,4-Dihydroxy-L-phenylalanine, 3,4-DHPA; 3,4-Dihydroxyphenylacetaldehyde, 3,4-DHBA; 3,4-dihydroxybenzaldehyde, 4’OMET; 4’-O-methylnorbelladine. TAT; Tyrosine aminotransferase, TH; Tyrosine hydroxylase, AADC; Aromatic L-amino acid decarboxylase, TYDC; Tyrosine decarboxylase, PAL; Phenylalanine ammonia lyase, C4H; Trans-cinnamate 4-monooxygenase, 4CL: Coumarate-4-lyase, C3H: 4-coumarate 3-hydroxylase (ascorbate peroxidase), HCT; hydroxycinnamoyl transferase, C3´H; p-coumaroyl shikimate 3´-hydroxylase, CSE; Caffeoyl shikimate esterase, HBS; Hydroxy Benzaldehyde synthase, NBS: Norbelladine synthase (coupled with norcraugsodine/noroxomaritidine reductase – NR), NCS; Norcoclaurine synthase, CYP96: Family of enzymes capable of phenolic couplings. Created in BioRender.com.
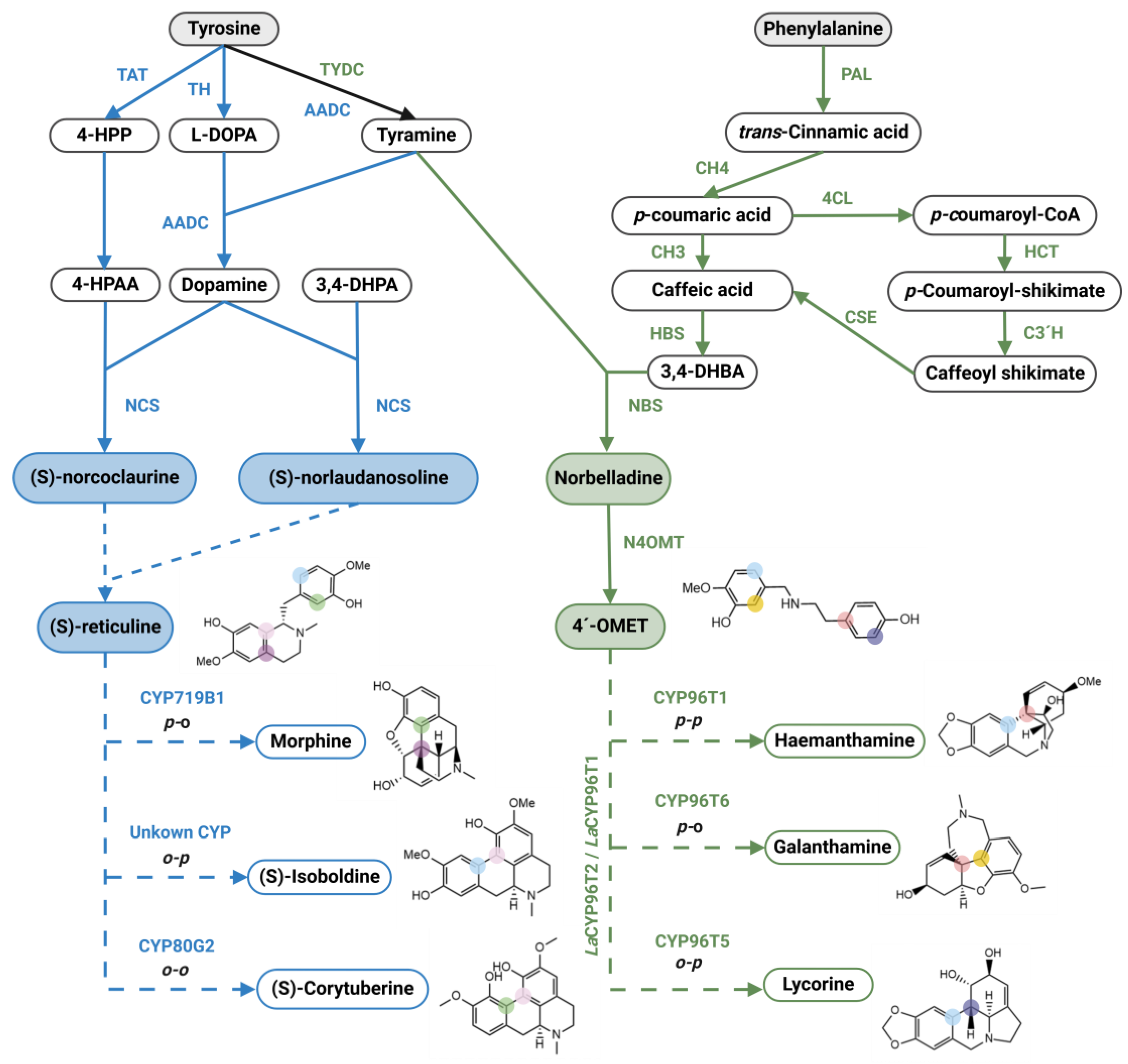
Figure 4.
Landmarks on IsAs and AmAs pathways elucidation. The elucidation of metabolic pathways for IsAs and AmAs began with the isolation of compounds now characteristic of each group, such as morphine and lycorine. Initially, the proposed steps in these pathways were based on experiments involving radioactive tracers. Later, omics technologies enabled a more in-depth characterisation of the enzymes involved in each pathway, to the point where, today, biofactories can be assembled to produce these molecules. Created in BioRender.com.
Figure 4.
Landmarks on IsAs and AmAs pathways elucidation. The elucidation of metabolic pathways for IsAs and AmAs began with the isolation of compounds now characteristic of each group, such as morphine and lycorine. Initially, the proposed steps in these pathways were based on experiments involving radioactive tracers. Later, omics technologies enabled a more in-depth characterisation of the enzymes involved in each pathway, to the point where, today, biofactories can be assembled to produce these molecules. Created in BioRender.com.
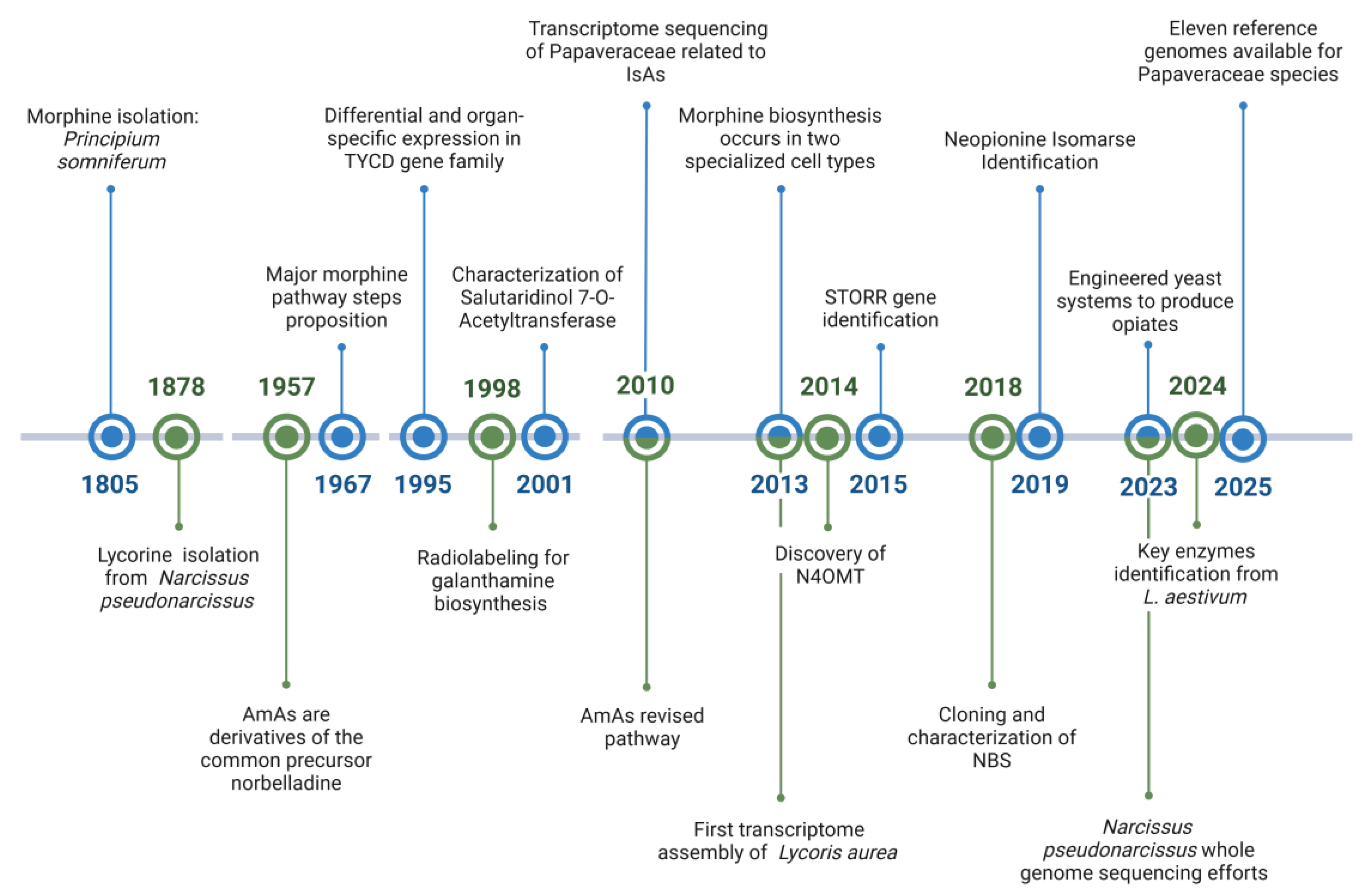
Figure 6.
Sequencing countries of alkaloid producing plants. here is a clear segregation in terms of the countries involved in studying IsAs and AmAs. While the sequencing of plants producing IsAs appears to be a global effort, only three countries are currently sequencing AmAs-producing plants.
Figure 6.
Sequencing countries of alkaloid producing plants. here is a clear segregation in terms of the countries involved in studying IsAs and AmAs. While the sequencing of plants producing IsAs appears to be a global effort, only three countries are currently sequencing AmAs-producing plants.
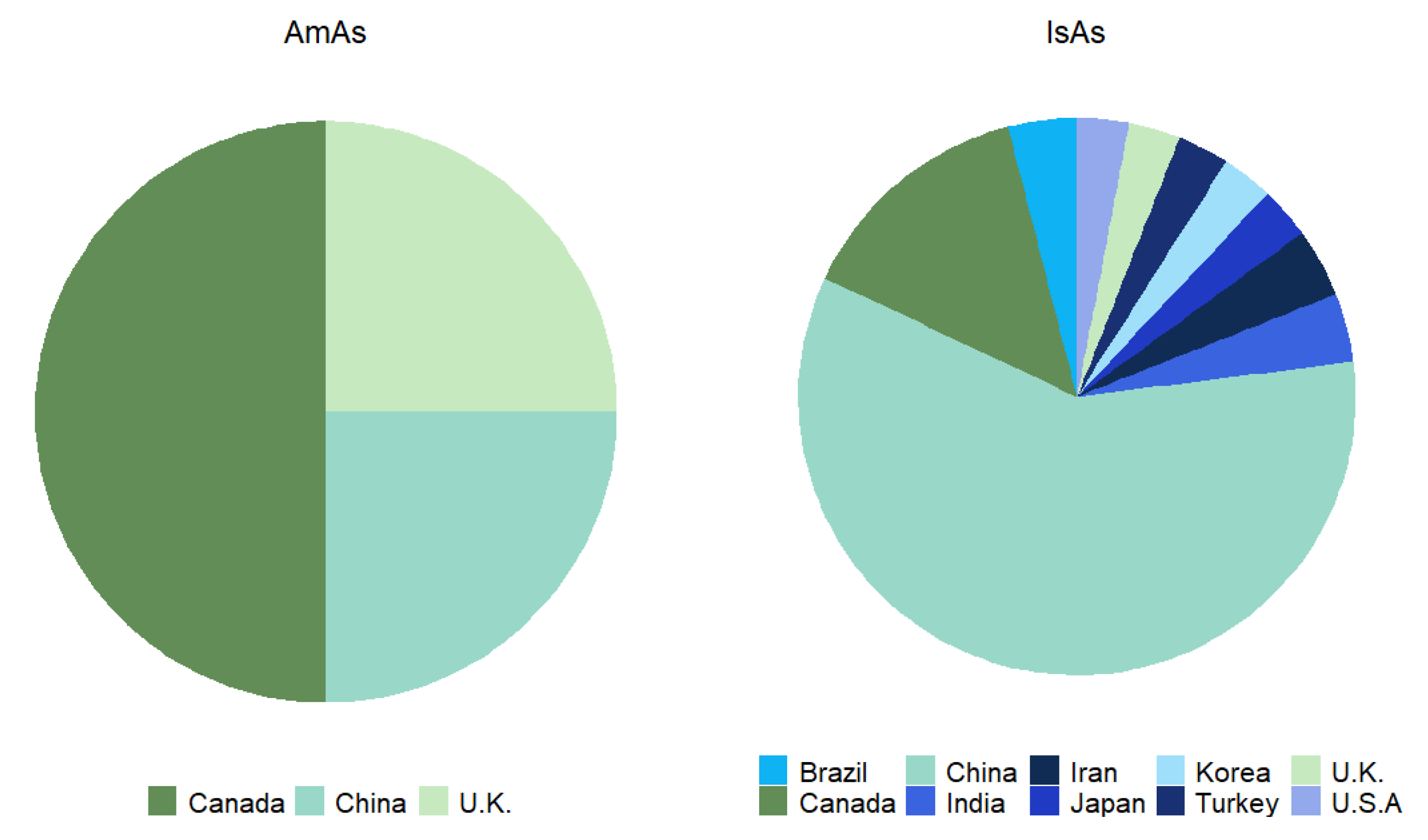
Table 1.
Comparative table between AmAs and IsAs.
| Aspect | Isoquinoline Alkaloids (IsAs) | Amaryllidaceae Alkaloids (AmAs) |
|---|---|---|
| Central Metabolic Pathways | Norcoclaurine → Reticuline → Morphine | Norbelladine → 4OMET → Galanthamine/Lycorine |
| Main Precursors | L-tyrosine, L-DOPA | L-tyrosine, L-phenylalanine |
| Key Enzymes | Methyltransferases, oxidoreductases (CYP719B, STORR) | Norbelladine synthase (NBS), CYP96T1, N4OMT |
| Phenolic Couplings | Intramolecular: Ortho-Ortho, Para-Ortho | Intramolecular: Para-Para, Para-Ortho, Ortho-Para |
| Taxonomic Distribution | 28 families in 17 orders (mainly Ranunculales) | Mainly in Amaryllidaceae, with exceptions in Asparagaceae |
| Progress in Elucidation | Pathway almost fully characterized | Pathway partially elucidated with major gaps (e.g., 3,4-DHBA) |
| Biotechnological Applications | Morphine production in microorganisms | Galanthamine production for Alzheimer’s treatment |
| Current Limitations | Structural complexity of final metabolites | Lack of integrated genomic and transcriptomic data |
Table 2.
Transcriptome sequencing studies on non-Amaryllidaceae families with data correlated to IsAs production.
Table 2.
Transcriptome sequencing studies on non-Amaryllidaceae families with data correlated to IsAs production.
| Family | Sequencing year | Sequencing country | Sequencing platforms |
Reference |
|---|---|---|---|---|
| Amaranthaceae | 2020 | China | Illumina HiSeq 2500 | [50] |
| Araceae | 2021, 2022 and 2024 | China | Illumina Nova seq 6000 and Illumina HiSeq 2000 | [51,52,53] |
| Aristolochiaceae | 2021 and 2022 | China | PacBio Iso-Seq, Illumina HiSeq2500 & Oxford Nanopore. | [54,55] |
| Asteraceae | 2021 | China | Illumina NovaSeq 6000 | [56] |
| Berberidaceae | 2012, 2014, 2015 and 2016 | Canada and U.S.A. | Illumina and 454 GS FLX pyrosequencing | [57,58,59,60] |
| Brassicaceae | 2019 and 2023 | China | Illumina HiSeq 2000 | [61,62] |
| Cactaceae | 2022 and 2023 | China and Canada | Illumina HiSeq X Ten and Illumina NovaSeq 6000 | [63,64] |
| Caryophyllaceae | 2016 | China | Illumina HiSeq 4000 | [65] |
| Convolvulaceae | 2023 | China | Illumina NovaSeq 6000 | [66] |
| Fabaceae | 2019, 2021 and 2024 | China and Brazil | Illumina HiSeq 2000 and Illumina NextSeq 500 | [67,68,69] |
| Lauraceae | 2020 and 2023 | China | bgiSeq-500 and Illumina | [70,71] |
| Liliaceae | 2023 | China | Illumina Novaseq6000 | [72] |
| Magnoliaceae | 2022 | China | Illumina HiSeq 2500 | [73] |
| Melanthiaceae | 2023 | China | Illumina HiSeq 2500 | [74] |
| Menispermaceae | 2012, 2015, 2016, 2020, 2022 and 2024 | Canada and China | Illumina, 454 GS FLX pyrosequencing, Illumina HiSeq X Ten, Illumina HiSeq-2500 | [57,59,60,75,76,77] |
| Nelumbonaceae | 2017 and 2022 | China | Illumina HiSeq 2000 |
[78,79] |
| Oleaceae | 2021 | China | Illumina HiSeq X Ten | [80] |
| Orchidaceae | 2022 | China | Illumina HiSeq 2500 | [81] |
| Papaveraceae | 2010, 2012 (3), 2013 (2), 2015 (2), 2016 (3), 2017 (2), 2018 (3), 2019 (2), 2020, 2021 (2) and 2022. | Canada, China, Iran, Japan, Korea, Turkey and U.K. | 454 GS FLX pyrosequencing, Illumina HiSeq 2000, Illumina HiSeq 2500, Illumina HiSeq 3000, Illumina HiSeq 4000, Illumina HiSeq X Ten and PacBio Iso-Seq | [82,57,83,84,85,59,60,86,87,88,89,90,91,92,93,94,95,96,97,98] |
| Poaceae | 2017 and 2021 | India and China | Illumina HiSeq 4000 | [99,100] |
| Ranunculaceae | 2012, 2015 (2), 2016, 2018, 2019, 2020, 2021, 2022, 2023 and 2024 | Canada, China | Illumina, 454 GS FLX pyrosequencing, Illumina HiSeq 2000, Illumina HiSeq 2500, Illumina HiSeq X Ten, PacBio Iso-Seq, Illumina NovaSeq 6000 | [57,59,60,101,10][] |
| Saururaceae | 2024 | China | Illumina/MGI-SEQ 2000 | [110] |
| Theaceae | 2022 | China | Illumina HiSeq 2000 | [111] |
| Xanthorrhoeaceae | 2024 | China | Illumina HiSeq xten/NovaSeq6000 | [112] |
* Tables may have a footer.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



