Submitted:
16 September 2025
Posted:
17 September 2025
You are already at the latest version
Abstract
The search for reliable biomarkers of blastocyst competence in IVF has historically emphasized morphological features of the trophectoderm and inner cell mass. However, parallel studies on mitochondrial biology have revealed a unique “oocyte–blastocyst mitochondrial dichotomy,” in which mitochondria shift functionally and structurally between fertilization and implantation. Contrary to the prevailing view that early embryonic mitochondria primarily drive ATP synthesis, evidence suggests that oocyte and preimplantation blastocyst mitochondria operate in a constrained metabolic state, with limited oxidative phosphorylation capacity due to anoxic conditions and immature cristae structure. Instead, these mitochondria appear to play a more critical role in biosynthetic precursor generation, redox regulation, and the metabolic programming of embryonic stem cells.We revisited this dichotomy using metabolomic profiling of spent blastocyst culture media, leading to the development of a non-invasive pre-implantation metabolic test for blastocyst competence (PMT-BC). Shotgun metabolomics revealed key biomarker patterns, not limited to classical substrates of glycolysis or oxidative metabolism, but including conjugated TCA intermediates, amino acid derivatives, and notably heme-associated metabolites. The detection of heme-a and heme-c derivatives aligns with their established roles in cytochrome function, oxidative signalling, and pluripotent stem cell activation. Importantly, inhibition of heme synthesis blocks the transition of naïve to primed pluripotent states, underscoring the developmental importance of these metabolites.Together, these findings highlight that oocyte-specific mitochondrial forms and their metabolic outputs are central to early embryogenesis, implantation potential, and stem cell priming. By integrating metabolomic biomarker discovery with mitochondrial biology, this work advances both embryo selection strategies in IVF and our understanding of the metabolic foundations of embryonic stem cell activation.
Keywords:
Oocyte-Blastocyst Mitochondria
; Embryomic Stem Cell activation
; Priming 2C ESCs differentiation
; mitochondrial TCA metabolites
; Heme synthesis
; pre-implantation embryo selection
; IVF
Introduction
Many advances in medical technology result when different research and innovation areas coincide. Here the search for biomarkers of blastocyst-embryo competence has developed parallel, but independent, to studies on mitochondrial biology and specifically that of the subcellular organelle anatomy of oocytes and blastocysts.
The search for morphological markers of development and hence competence of the IVF blastocyst -embryo to successfully implant, has relied on the gross tissue and, cell to cell structures. The form and function of trophectoderm and inner cell mass which are responsible for implantation and the triggering of pluripotent stem cells to form the true embryo.
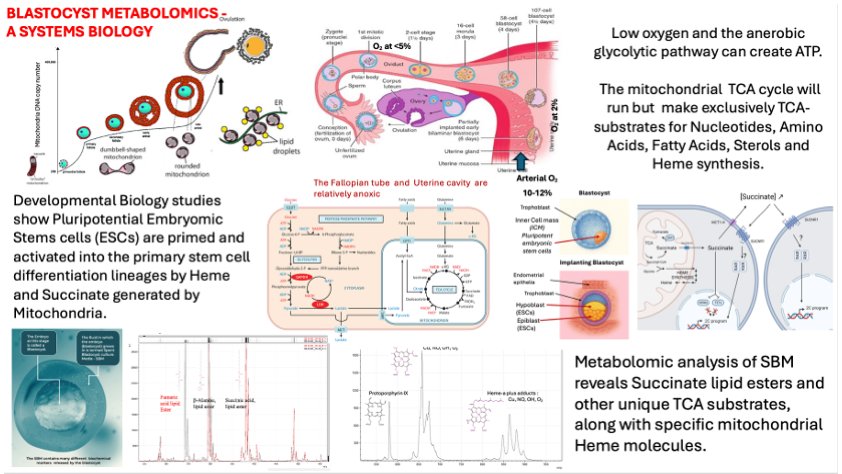
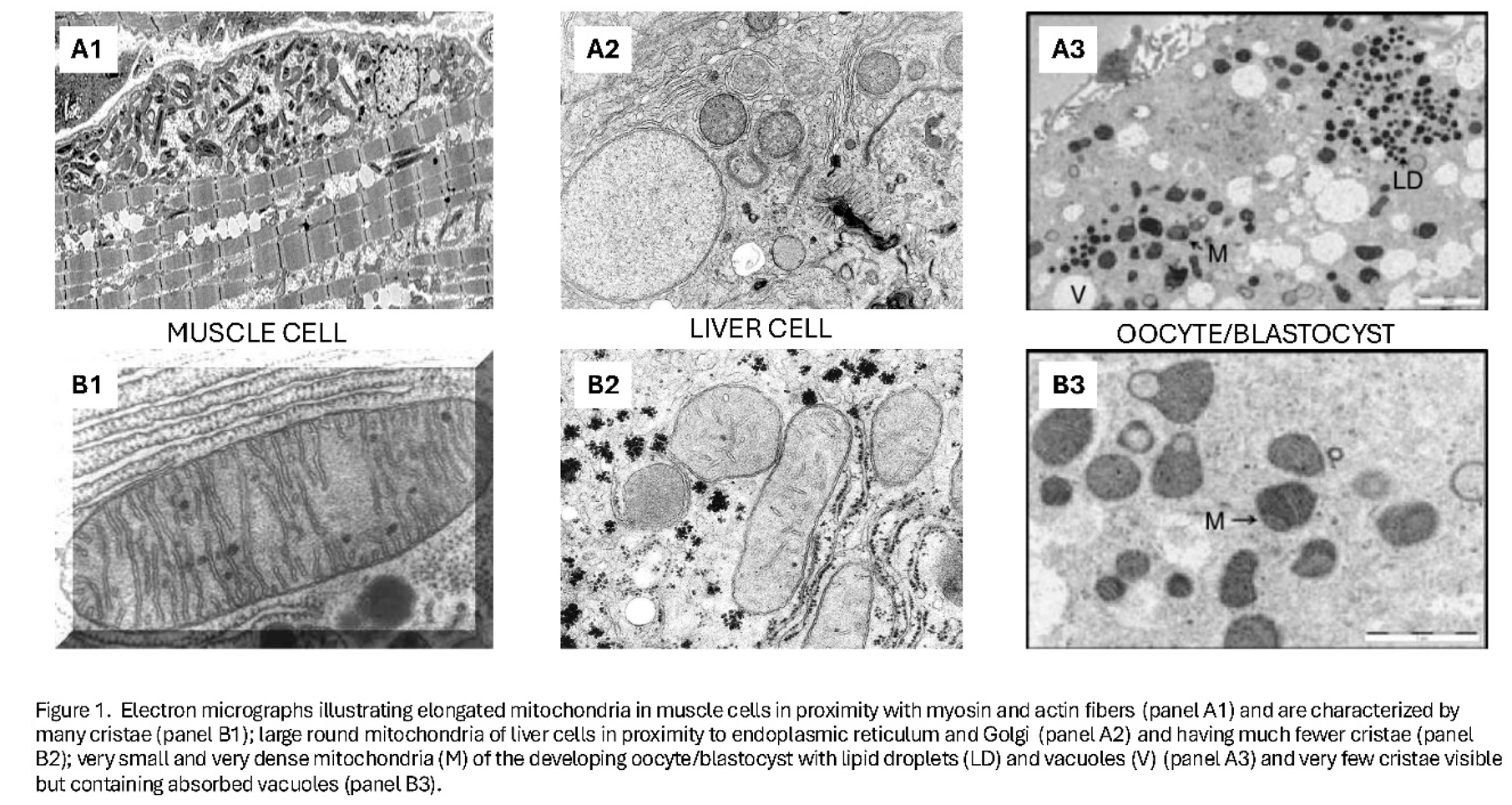
However, subcellularly and equally as remarkable as the oocyte haploid nucleus, which fuses with the sperm haploid nucleus, and the new diploid genome replicating; are the oocyte mitochondria. Many thousands of them, more than any other single cell, and their micro-anatomy is unique [1,2,3,4] (Figure 1 and Figure 2).
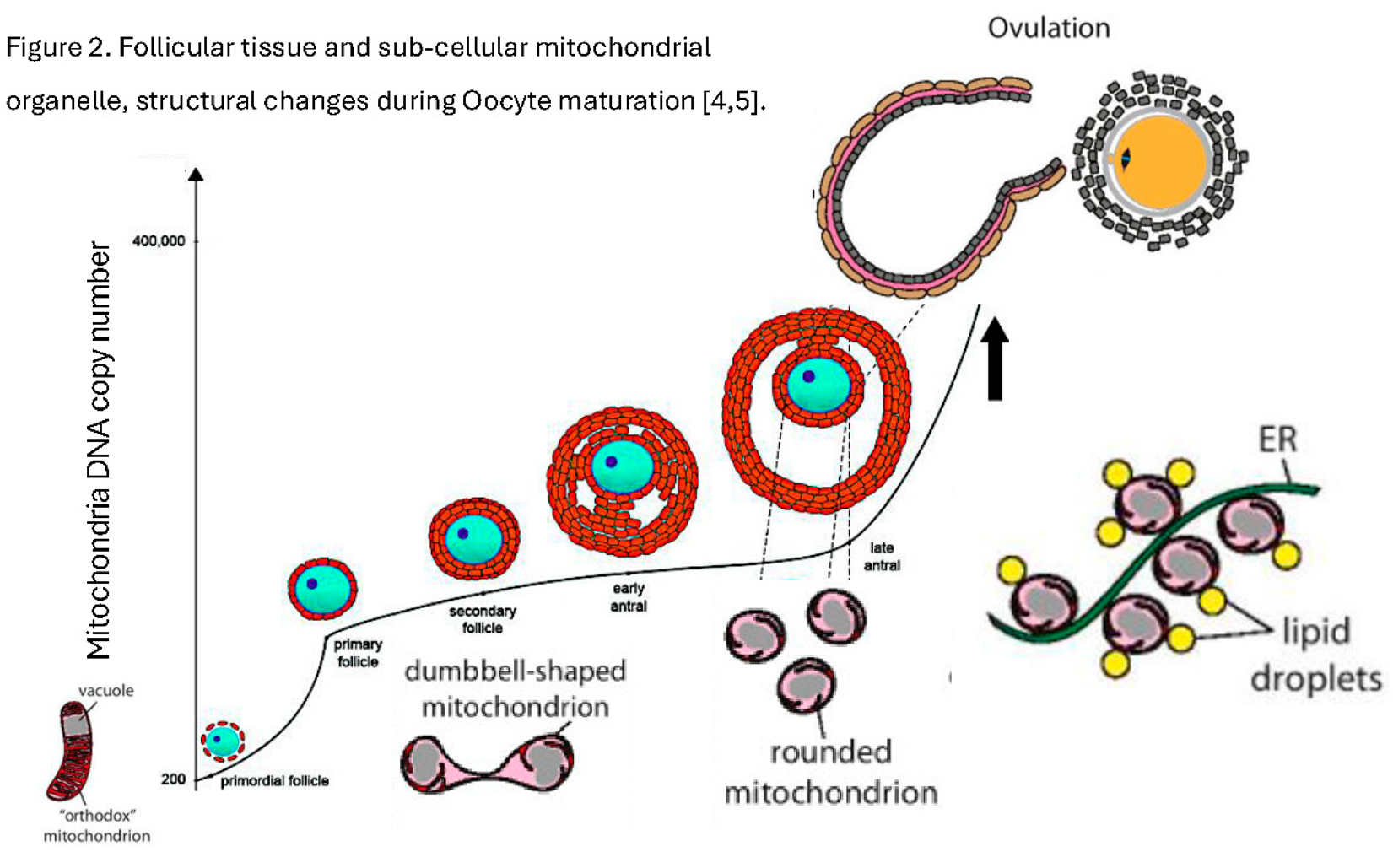
The mitochondrial dichotomy refers to the marked shift in mitochondrial function, morphology, and metabolic role between the fertilized oocyte and the implanting blastocyst. This difference reflects the unique energetic and developmental requirements of early human embryogenesis [4,5] (Table 1).

The importance of these unique oocyte blastocyst mitochondria were recognised and formed the basis of the fertility therapeutics company OVA Sciences and Autologous Germline Mitochondrial Energy Transfer (AUGMENT). The concept is based on the fact that oocytes from older women, or those with reduced ovarian reserve, often show mitochondrial dysfunction [6,7]. The presumed mechanism was impaired energy production and this may reduce fertilization, blastocyst formation, and implantation rates [8].
On this bases of these assumptions there was an immediate rush to therapeutic interventions such as AUGMENT. This Involved isolating putative oogonial stem cells from the patient’s ovarian tissue [9], extracting mitochondria, and injecting them into her oocytes at the time of ICSI [10] and thus increase healthy mitochondrial copy number using the patient’s own genetic material. Early reports claimed improved pregnancy rates [11] . However, this has not been validated [12,13] and the focus on mitochondrial ATP production as the rationale has proved a false distraction [8].
The common preconception is that mitochondria just, and uniquely, produce ATP. Furthermore, given all the cellular functions occurring after fertilization, the assumption is that all these mitochondria are only required to produce the ATP that drives DNA synthesis, mitosis and gene expression.
The first falsehood is that ATP is only synthesised by Oxidative Phosphorylation (OXPHOS) in the mitochondria and driven by ATP synthase coupled H+ ion channels. ATP is also generated by the glycolytic and purine salvage pathways [14] (Figure 3). One might question whether the ATP produced by these pathways is sufficient to maintain a cells integrity and function. However red blood cells, which survive and function for 90days, not only do not contain a nucleus they also do not contain any Mitochondria. Apart from stores and recycling ADP+Pi pathways they rely entirely on the Embden-Meyerhof pathway [15] (see Figure 3).
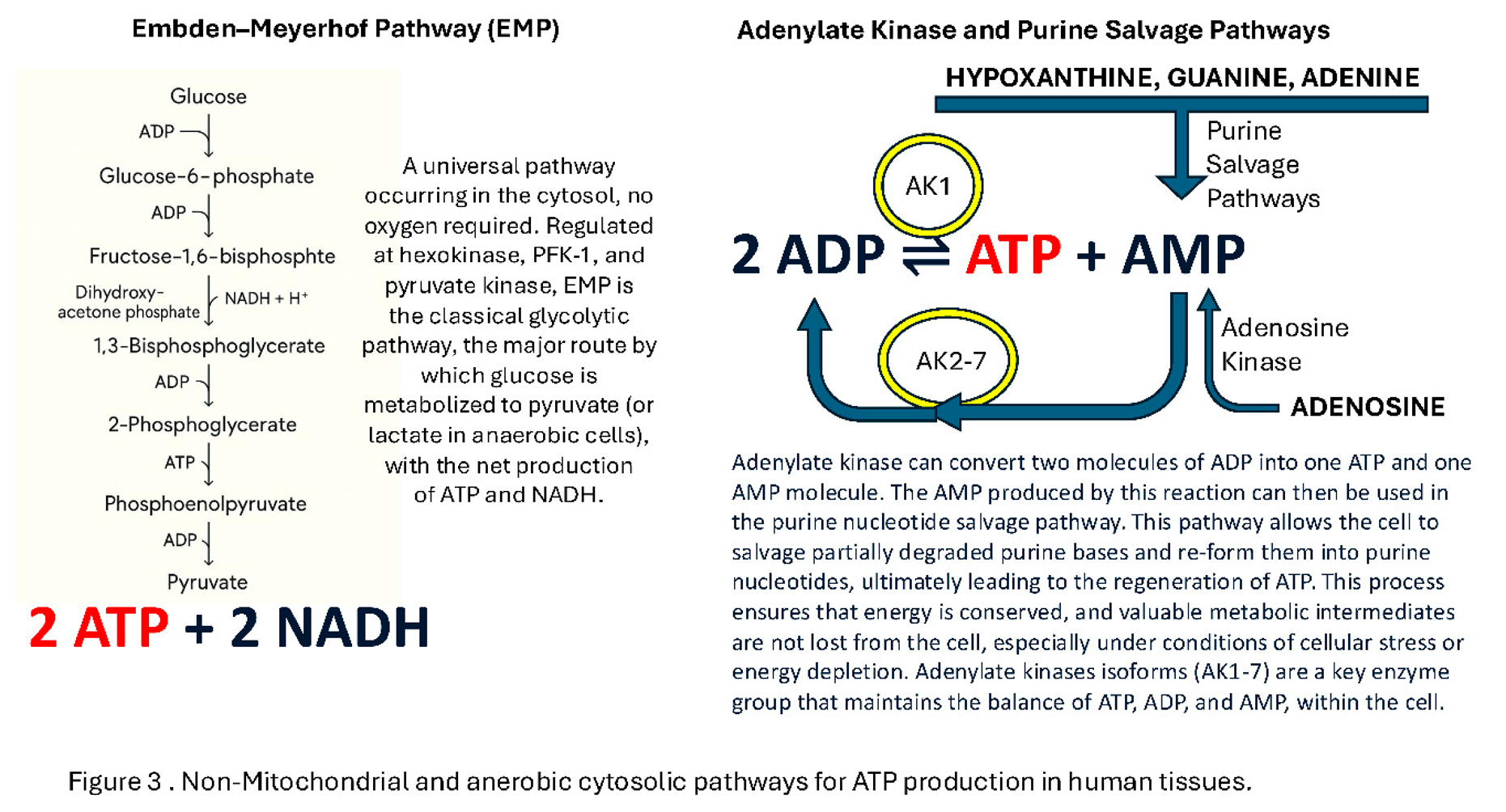
The second falsehood is that oocyte and preimplantation blastocyst mitochondria are actively generating ATP: The electron transport chain (ETC) relies entirely on electron transfer driving proton pump channel complexes straddling the mitochondrial inner membrane, hence infolded cristae, and consuming atomic oxygen, at the final stage to make H20 and converting ADP to ATP. However:
- A)
- B)
- C)
Indeed, the mitochondria of the oocyte and preimplantation blastocyst are structural distinct from forms found in any other cells (see Figure 2). Thus, all mitochondria are not the same and specific mitochondria phenotype match their desired metabolic function within a cell (see Figure 1).
Consistent with their structure would be the direction of the TCA cycle to form the raw materials required for DNA replication, lipid membrane formation, amino acid formation and sterol and heme molecules; and not simply generating FADH and NADH substrate for the electron transport chain proton pumps found on cristae (see Figure 4).
Therefore only the mitochondria seen in mature ova would need to be selected and not those of other cells, even putative ovarian stem cells, in order to restore metabolic competence in maternal aged oocytes [7].
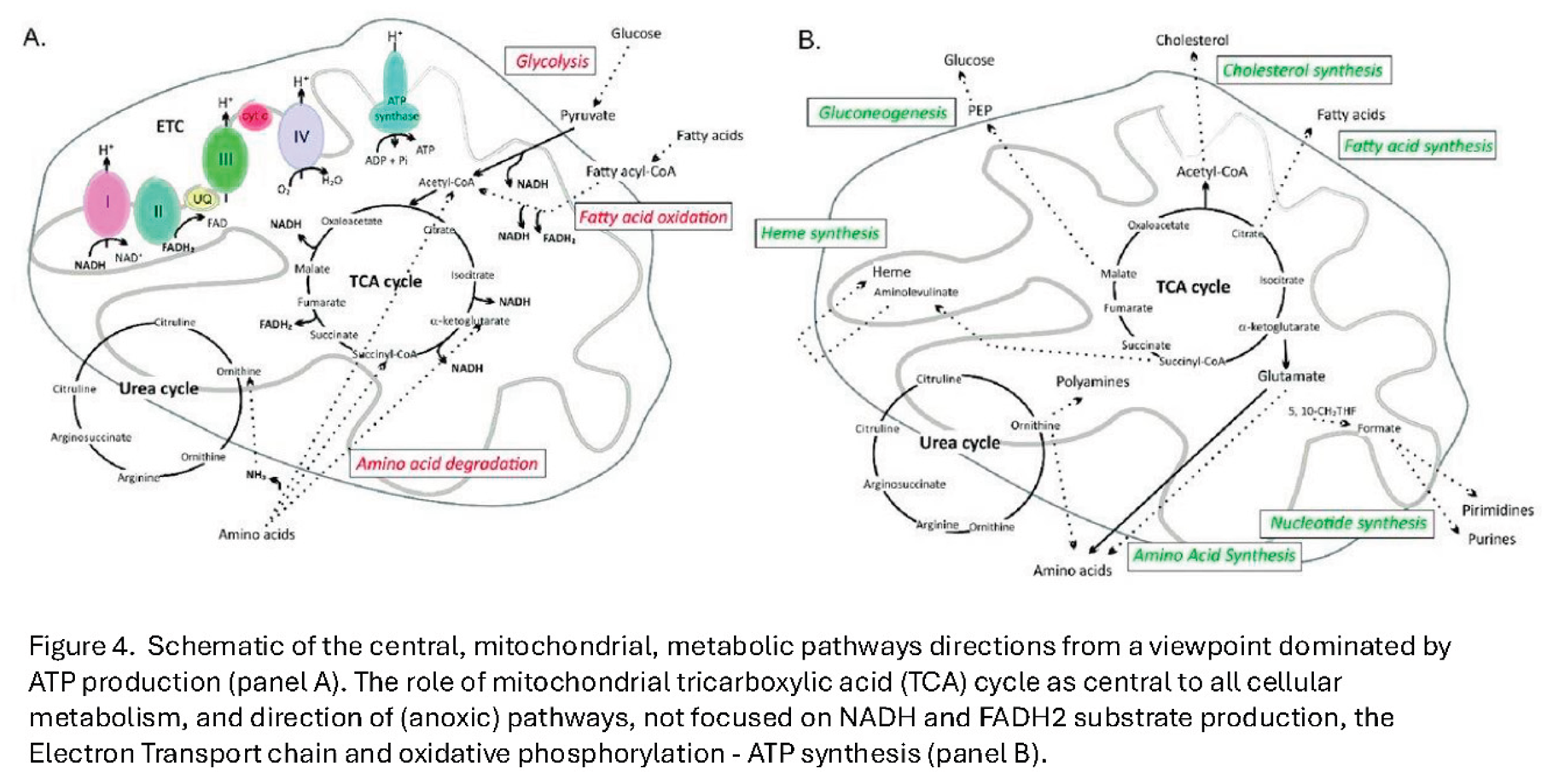
Why Revisit the Oocyte–Blastocyst Mitochondrial Dichotomy?
From these and our own metabolomic studies of spent blastocyst media we developed the concept of the Blastocyst metabolomic battery (Figure 5).
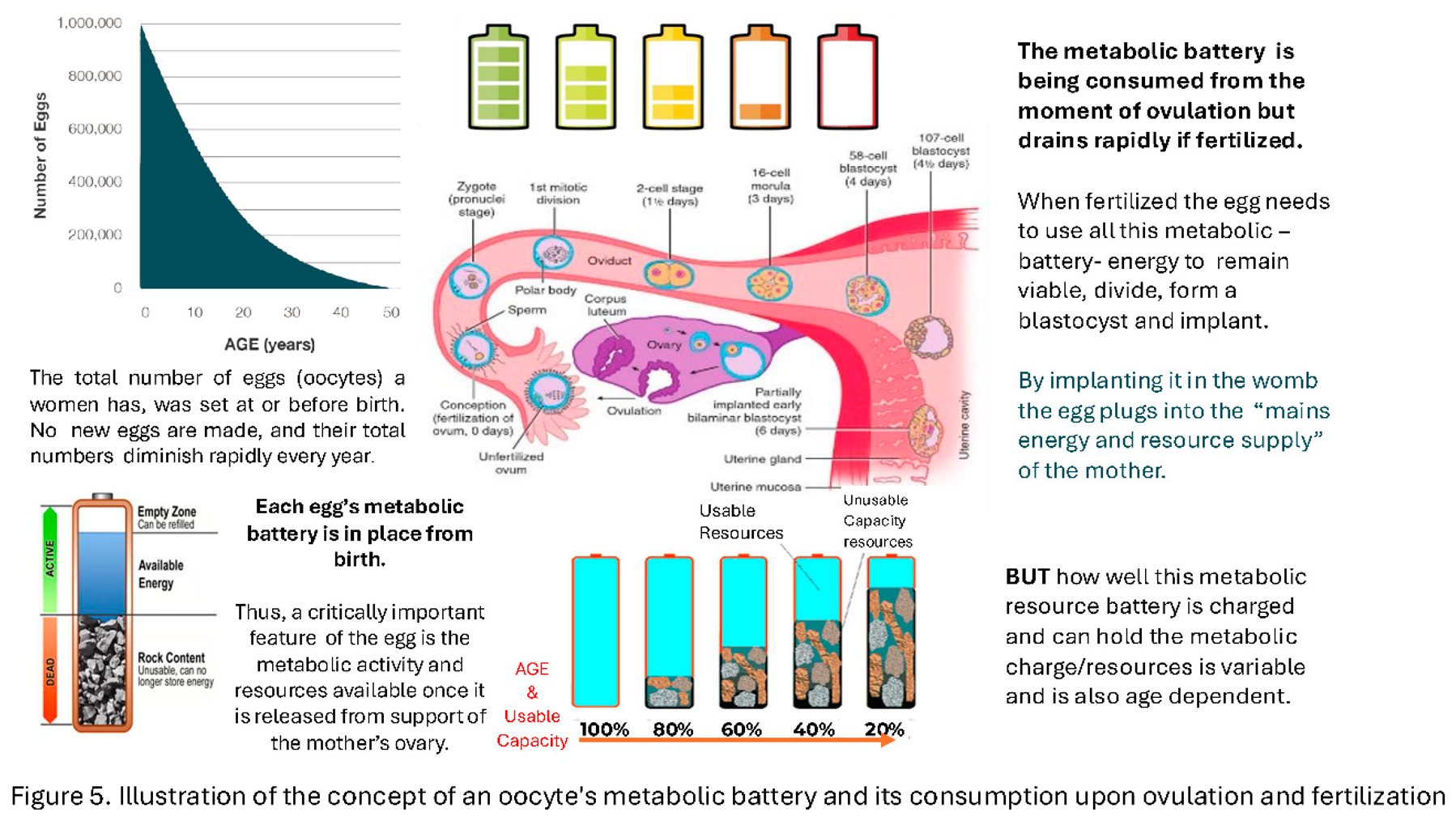
Once free of the ovarian follicle and maternal granulosa and blood supply, the oocyte has to rely on its internal supplies of resources. Upon fertilisation these resources are mobilised and, with anabolic and catabolic pathways active, consumed rapidly with some metabolites being excreted, shed or over spilling from the blastocyst. In the case of IVF, into the Spent Culture Media (SBM) [22,23].
Approaching IVF success improvement from a different, non-invasive, angle we and others have been pursuing proteomic and metabolomic biomarker assessment of IVF Blastocyst-embryo SBM [24,25,26,27]. Not knowing what the metabolites of such pathways maybe, and with no pre conceived hypothesis to follow; this is based on a more shotgun approach to biomarker discovery [25]. This approach is only now possible due to developments in mass, nuclear magnetic resonance and Raman spectroscopies [28,29]. These “Omics” technologies allow the rapid and sensitive resolution of thousands of metabolite molecules without knowing precisely what every resolved (e.g. mass) peak are. Using advanced computing the challenge of associating the relative intensities of these many molecules with outcomes is made significantly faster by advanced computing Ai and Machine Learning [30]. This allowed us to identify mass spectrometry characterised mass peak biomarkers of significance to implantation and viability [25].
On hindsight the signature of these blastocyst metabolic activity was not going to be the, predominantly, classically understood substrates of anaerobic respiration such as glucose, pyruvate, lactate, NADH, FADH and ATP. Indeed this transpired during our studies of metabolites identified in the development of our pre-implantation metabolic test for blastocyst competence, PMT-BC [25]. However, what has been found advances our understanding of oocyte-blastocyst mitochondrial metabolism, its importance in form and function, and signally during this very early stage of embryonic stem cell developmental biology to form the true embryo.
Analysis of the mass spectral peaks found to be important markers in the PMT-BC algorithm includes key TCA substrates (e.g. succinate and fumarate) and amino acids (e.g. b-alanine) but conjugated as esters to long chain aliphatic (lipid) groups. Functionally these would be insert into, and held as, inert stores within cellular membranes and lipid rich granules (see Figure 6).
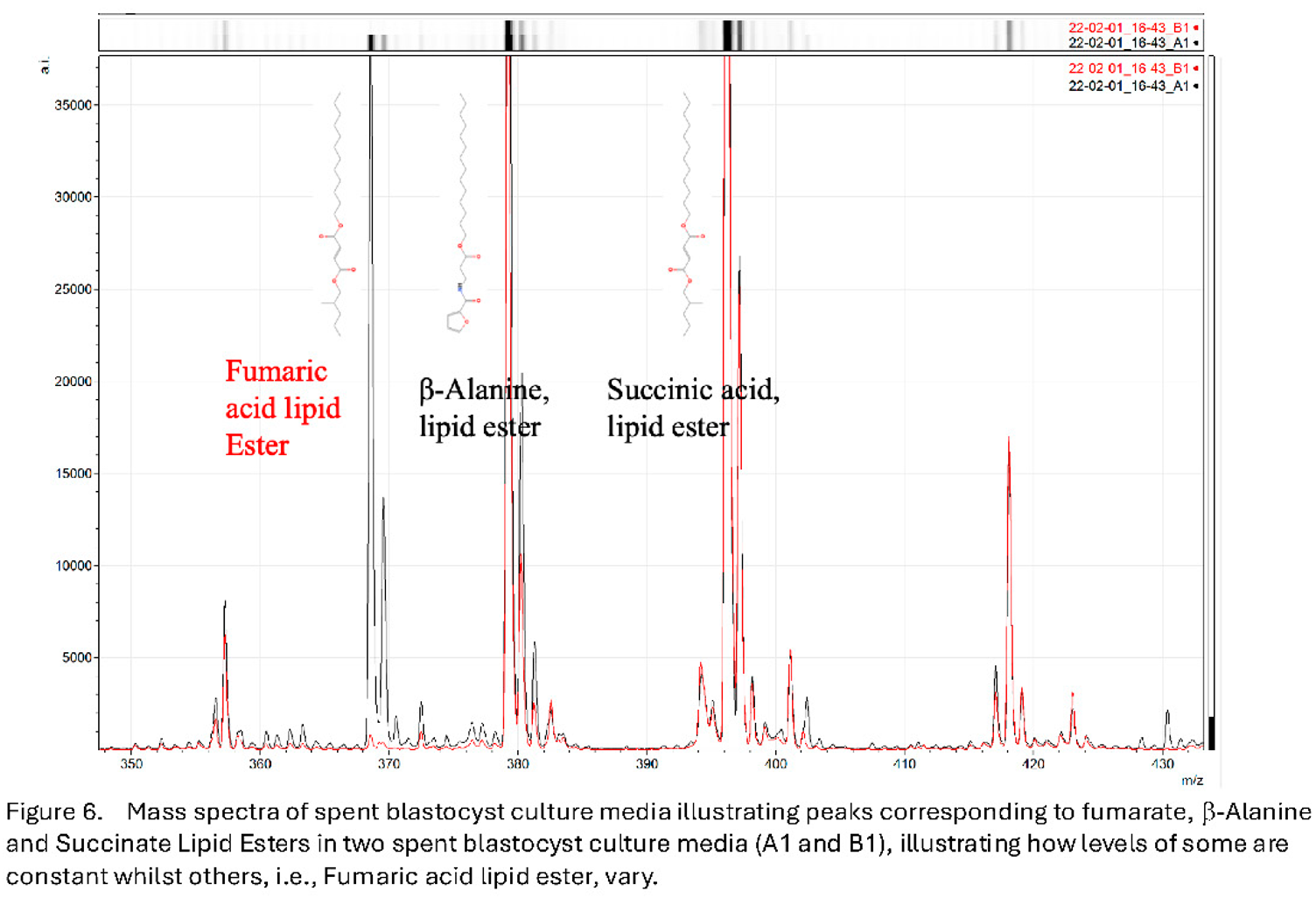
Furthermore, particularly associated with viability are the appearance of peaks corresponding to mass spectra of protoporphyrin IX and heme-c & heme-a variously conjugated with oxygen (O2), nitric oxide (NO) and Copper (Cu) (see Figure 7).
Significantly heme-b, heme-o, and heme-d were not detected in the SBM mass spectra (Figure 8).
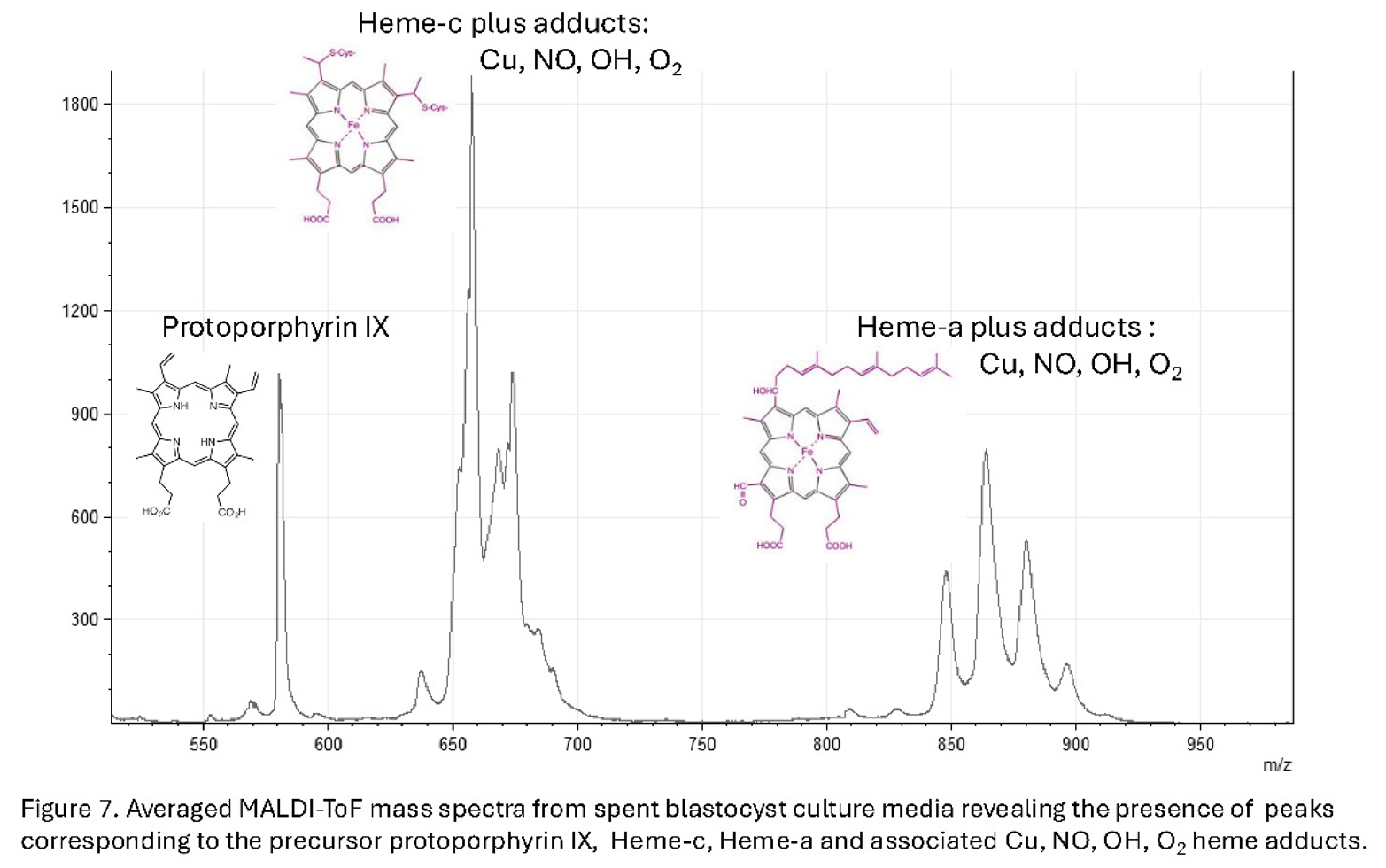
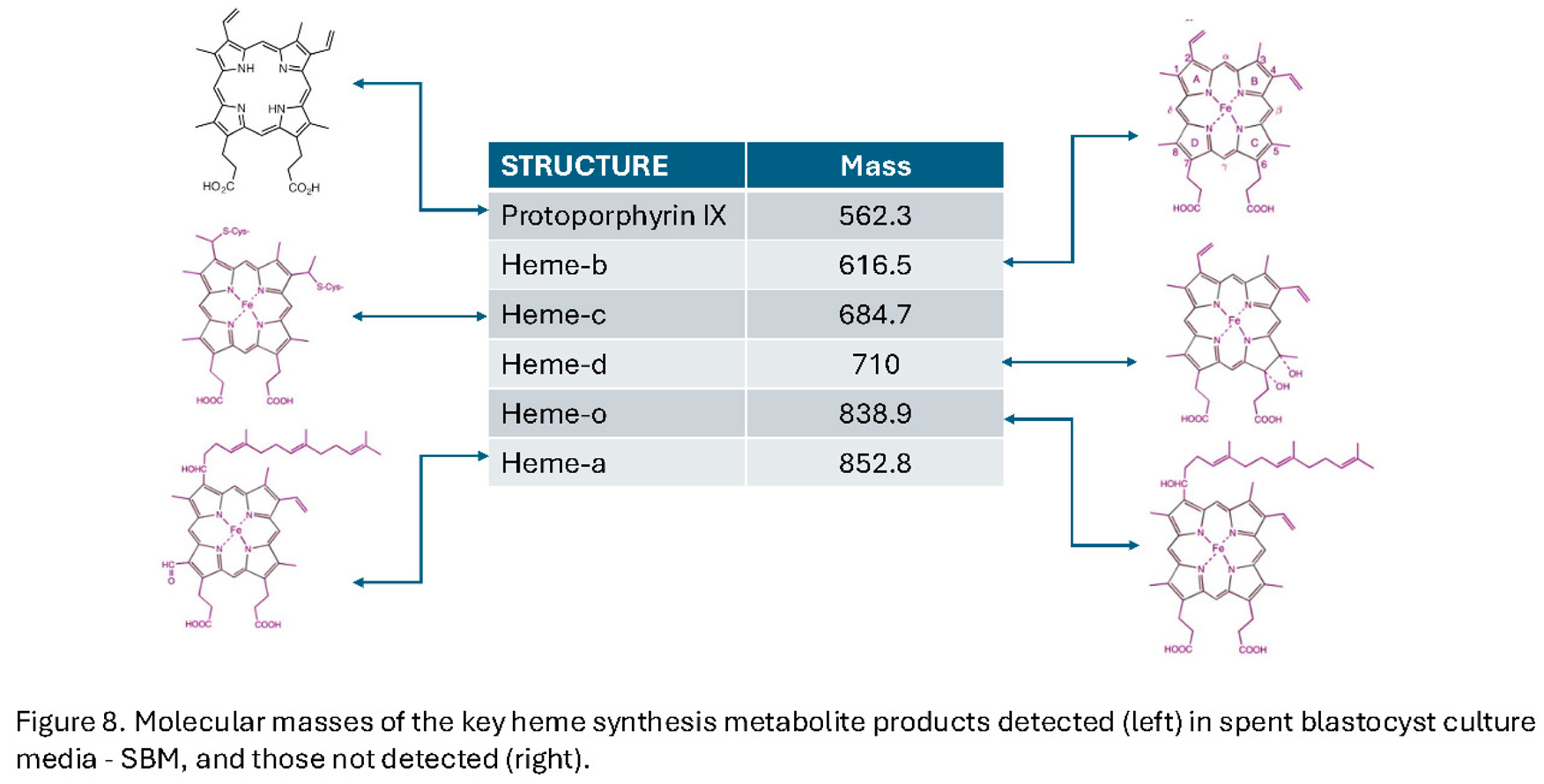
Protoporphyrin IX is the precursor caging structure for iron held in either the ferrous Fe-2 or ferric Fe-3 , ionic forms – the heme’: Heme-a is a specialized form of heme found within the respiratory enzyme cytochrome oxidase of mitochondria. Heme c is found in a large group of heme prosthetic containing proteins termed generically cytochromes c (cyt c). These are a diverse family of proteins essential for electron transport in photosynthesis and respiration, The functional prosthetic Heme-c itself can be fine-tuned over a wider range of oxidation-reduction potentials than heme-b. Heme-b being the form found in haemoglobin and myoglobin. Heme C also plays an important role in apoptosis where just a few molecules of cytoplasmic cytochrome c, which must still contain heme-c, leads to programmed cell death [31,32]. Hem-o, is an intermediate in heme-a synthesis and a cofactor for terminal oxidases in the electron transport chain, primarily found in bacteria like Escherichia coli and some archaea and eukaryotes. Heme-d is only found in bacteria and in specific enzyme pathways, such as nitrate reduction [33].
Synthesised by and interplay between the mitochondrial matrix pathways and the cellular cytosol, heme’ a & c are critical as the core functional moieties in large carrier heme-protein complexes of the electron transport chain, i.e. cytochrome oxidase, reductase and cytochrome c. Unlike in blood haemoglobins and muscle myoglobin (heme-b containing), these particular heme proteins trap oxygen, oxygen species and oxygen radicals between the porphyrin (heme) caged Fe+3 and Cu+1 metal ions. Passing electrons to and from the ETC proton pump complexes, straddling the mitochondrial inner membrane, generates the reservoir of H+ ions between the mitochondrial inner and outer lipid bilayers. Passing back down the H+ ion concentration gradient drives mitochondrial ATP synthetase and effects Oxidative Phosphorylation (OXPHOS); cytochrome c splitting O2 to monomeric O-2 radicals which combine with these H+ ions to form H20 [34] (see Figure 4a).
Thus, synthesis of the precursor non-protein key organic components, that are the functional moieties of the ETC proteins of mitochondrial oxidative OXPHOS, coupled with micronutrients Cu, Fe and Mn [35], only activates at the time (and spatial alignment) of implantation and ICM stem cell growth & differentiation; when huge excess ATP synthesis is needed. This is both logical and developmentally, a key spatial-temporal event window mechanism.
Moreover, heme is involved in a considerable number of other metabolic systems [36,37]. Recent studies on activation of pluripotent embryonic stem cells has shown that development of the naïve stem cells is completely blocked by Heme synthesis inhibition [38]. Indeed, free heme(s) have been shown to signal activation, or priming of what would be the blastocyst ICM naïve embryonic pluripotent stem cells; and free inter cellular succinate drives primed naïve embryonic stem cells into the next stage of stem cell development [39]. Thus, early mammalian embryonic development is tied directly to mitochondrial biochemistry and ordered metabolomics of these specialist oocyte morphological mitochondria (see Figure 9).
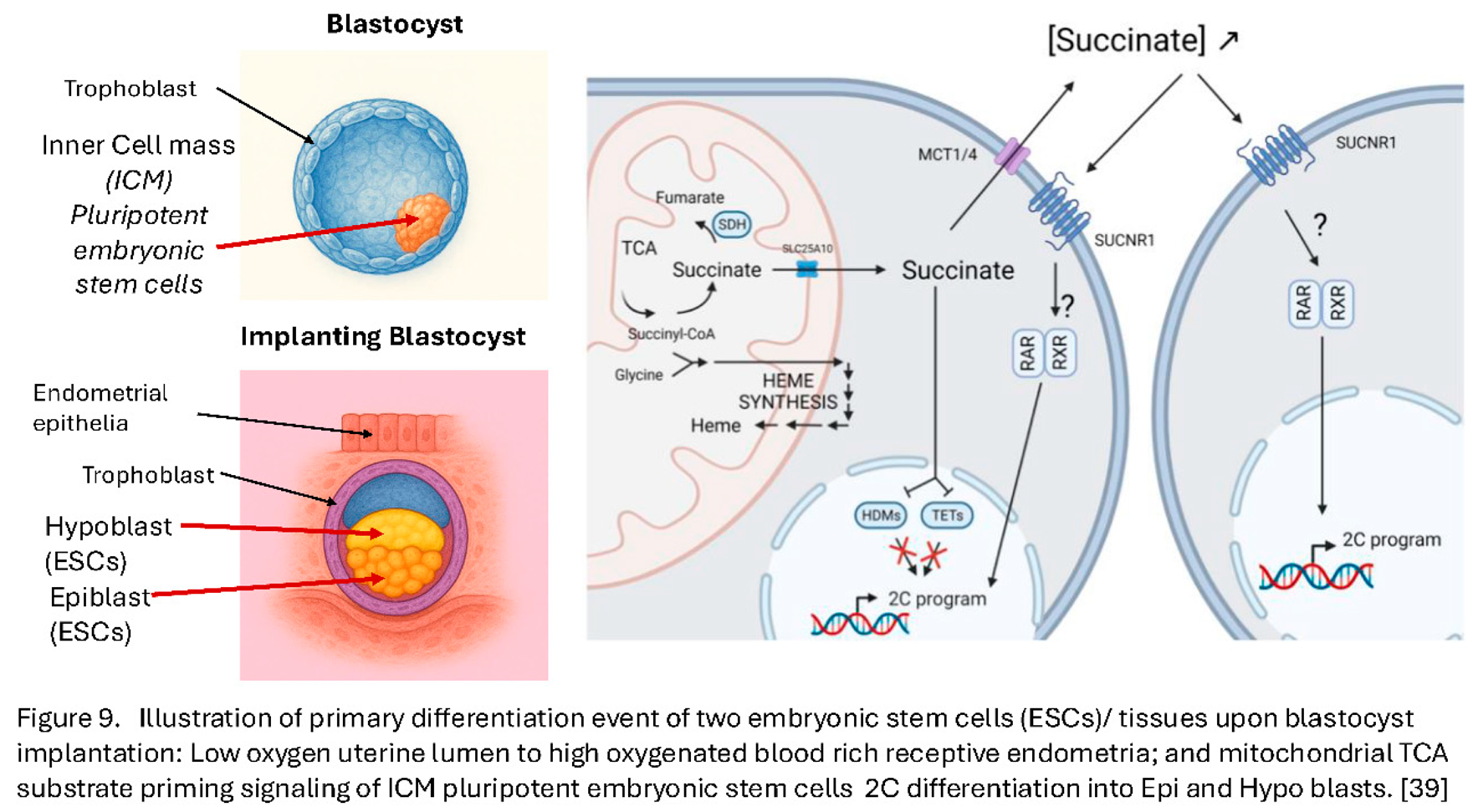
Two main states of pluripotent stem cells have been described: the naïve embryonic stem cells (ESCs), resembling the inner cell mass (ICM) of the pre-implantation blastocyst-embryo, and the primed ESCs, mirroring the epiblast and hypoblast of the post- implantation stage (see Figure 9). While these cells represent closely timed stages in embryo development, they display dramatic differences, such as developmental potential, epigenetic landscape, X- chromosome inactivation patterns [40,41,42,43], but critically also metabolic activity [39,44,45]. Embryomic stem cell culture is known to be very heterogenous in terms of pluripotent/epigenetic marker expression [46] and a small population of mouse ESCs, grown in naïve in vitro conditions, displays features of the two- cell-type stage embryo (2C- like population or 2CLCs) exhibiting extended potential [39,47]. This subset also displays the expression of 2C- specific genes such as the Zscan4 cluster, the retro- transposable element MuERVL and/or the master regulator DUX [39,47,48,49], concomitant with a disappearance of chromocenters and loss of the core pluripotency protein OCT4 [39,47].
Thus, shotgun analysis for biomarker metabolites that indicate IVF Embryo- blastocyst viable implantation potential has coincided with advancing studies on pluripotent naïve embryonic stem cell activation. Amongst our key biomarker ratios are heme and succinate ester lipid precursor metabolites (see Figure 6 and Figure 7) that have been implicated in ICM ESCs activation [39].
Conclusions
Mitochondrial augmentation in IVF has continued and now refers to a group of experimental assisted reproduction techniques aimed at improving oocyte and embryo competence by increasing the quantity or functional quality of mitochondria in the oocyte, [11,50,51], (see Table 2).

However, the mitochondria harvested need to be the specialist oocyte differentiated forms, and in most cases will be of donor oocyte origin. Pioneered in the late 1990s [50] cytoplasm from young women’ donor oocytes (containing healthy, oocyte specific differentiated, mitochondria) is injected into the patient’s oocyte. This led to increased pregnancy rates, but raised ethical and safety concerns because it results in three-parent genetic mitochondrial heteroplasmy. The FDA halted its use in the US due to germline modification implications. However, specifically to prevent inheritance of mitochondrial genetic disorders, now the entire doner oocyte cell has its nucleus removed and replaced by the parental nucleus so that the mitochondrial entire genetic makeup is that of the donor alone [51,52,53,54,55,56].
Specific case by case permission is required currently but has been successful and garnered international news headlines [57,58,59].
Like the common misconception that a “genetic” test is all we need, the simple assumption that all Mitochondria are the same, i.e. a power house only producing ATP, is a gross simplification that it is not just wrong, but damagingly wrong:
The interplay of any one genetic mutation effecting the complex multigene activation and built in redundancy/backup processes of living organisms is generally small. We all carry genetic mutations-variants but the phenomic’ magnitude of their individual effects are usually small, and only clearly manifest in a minority, although a significant minority, of disorders. So for complex diseases and disorders multiple genetic features must co-exist and single genetic tests can only give an association, or range in probability of the disorder manifesting. Thus, PGT-A is a single genetic test (but of gross chromosomal replication errors) and its value is limited to a defined group of advanced maternal age IVF patients and/or repeat miscarriage [60].
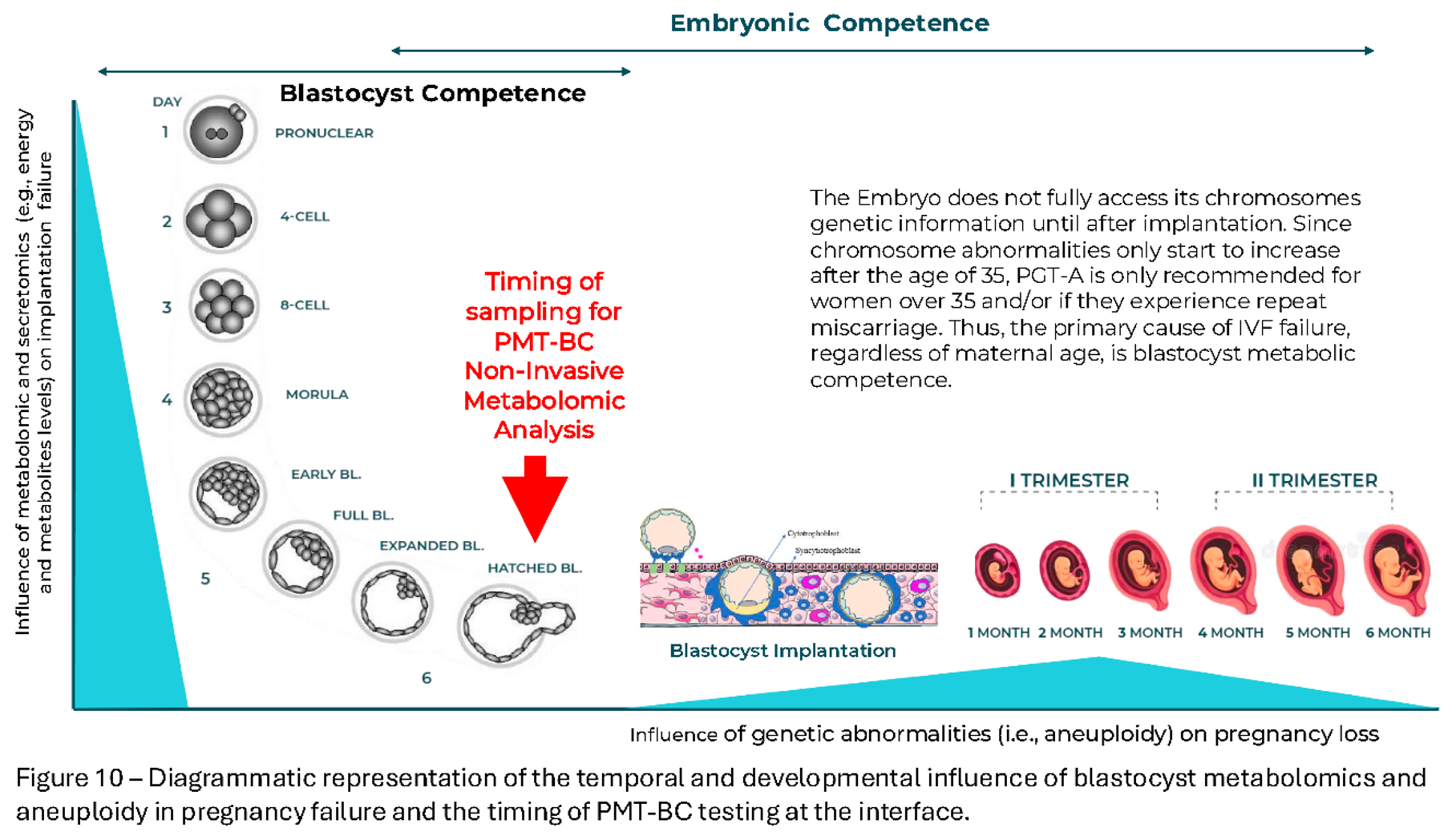
In the case of metabolomics and mitochondrial transfer it would strongly suggest the opposite. Instead of just applying the technique to a very limited minority, with known mitochondrial disease inheritance; donor oocyte specific mitochondrial transfer or enucleated Ova from young donors being utilised in IVF of older women, may well solve a significant number of failed IVF cases. However, this is by definition heteroplasmy, three genetic contributors. Only metabolomic competence selection of blastocysts (or if possible ova) can even attempt to solve the IVF success rate problem for all, by selection of the metabolomic (mitochondrial) fittest for IVF-transfer (Figure 10).
References
- S.A. Nottola, G. Macchiarelli, G. Coticchio, S. Bianchi, S. Cecconi, L. De Santis, G. Scaravelli, C. Flamigni, A. Borini, Ultrastructure of human mature oocytes after slow cooling cryopreservation using different sucrose concentrations, Human Reproduction, 2007, 22:(4):1123–1133. [CrossRef]
- R. Dumollard, J. Carroll, M.R. Duchen, K. Campbell, K. Swann, Mitochondrial function and redox state in mammalian embryos, Seminars in Cell & Developmental Biology, 2009 20(3):346-353. [CrossRef]
- Van Hoeck, V., Leroy, J. L. M. R., Arias Alvarez, M., Rizos, D., Gutierrez-Adan, A., Schnorbusch, K., Bols, P. E. J., Leese, H. J., & Sturmey, R. G. (2013). Oocyte developmental failure in response to elevated nonesterified fatty acid concentrations: mechanistic insights. REPRODUCTION, 2013, 145(1): 33-44. [CrossRef]
- Chiaratti MR. Uncovering the important role of mitochondrial dynamics in oogenesis: impact on fertility and metabolic disorder transmission. Biophysical Reviews. 2021;13(6):967-981. [CrossRef]
- Kirillova, A.; Smitz, J.E.J.; Sukhikh, G.T.; Mazunin, I. The Role of Mitochondria in Oocyte Maturation. Cells 2021, 10, 2484. [Google Scholar] [CrossRef] [PubMed]
- Woods, D.C.; Khrapko, K.; Tilly, J.L. Influence of Maternal Aging on Mitochondrial Heterogeneity, Inheritance, and Function in Oocytes and Preimplantation Embryos. Genes 2018, 9, 265. [Google Scholar] [CrossRef] [PubMed]
- Zhang D, Keilty D, Zhang ZF, Chian RC. Mitochondria in oocyte aging: current understanding. Facts Views Vis Obgyn. 2017, 9(1):29-38.
- Couzin-Frankel, J. Eggs' power plants energize new IVF debate. Firm adding energy-generating mitochondria to egg cells has already produced human pregnancies. SCIENCE, 2015, 348(6230): 14-15. [CrossRef]
- White YA, Woods DC, Takai Y, Ishihara O, Seki H, Tilly JL. Oocyte formation by mitotically active germ cells purified from ovaries of reproductive-age women. Nat Med. 2012,18(3):413-21. [CrossRef]
- Dale B, Wilding M, Botta G, et al. Pregnancy after cytoplasmic transfer in a couple suffering from idiopathic infertility: case report. Hum Reprod. 2001;16(7):1469–1472. [CrossRef]
- Fakih, M. H., El Shmoury, M., Szeptycki, J., Cruz, D., Lux, C.R., Verjee, S., Burgess, C.M., Cohn., G.M., Casper R.F. The AUGMENT treatment: Physician reported outcomes of the initial global patient experience. JFIV Reprod Med Genet 2015, 3:3. [CrossRef]
- Lee K. An Ethical and Legal Analysis of Ovascience – A Publicly Traded Fertility Company and its Lead Product AUGMENT. American Journal of Law & Medicine. 2018, 44(4):508-528. [CrossRef]
- https://www.technologyreview.com/2016/12/29/106805/turmoil-at-troubled-fertility-company-ovascience/.
- Dzeja, P.; Terzic, A. Adenylate Kinase and AMP Signalling Networks: Metabolic Monitoring, Signal Communication and Body Energy Sensing. 2009, 10, 1729–1772. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.L., Cox, M.M. Lehninger Principles of Biochemistry. 7th Edition, W.H. Freeman, New York, 2017. ISBN 10: 1319108245.
- Ng KYB, Mingels R, Morgan H, Macklon N, Cheong Y. In vivo oxygen, temperature and pH dynamics in the female reproductive tract and their importance in human conception: a systematic review. Hum Reprod Update. 2018, 24(1):15-34. [CrossRef]
- Brouillet, S., Baron, C., Barry, F. et al. Biphasic (5–2%) oxygen concentration strategy significantly improves the usable blastocyst and cumulative live birth rates in in vitro fertilization. Sci Rep 2021, 11:22461. [CrossRef]
- Au, H.-K., Yeh, T.-S., Kao, S.-H., Tzeng, C.-R., Hsieh, R.-H. Abnormal Mitochondrial Structure in Human Unfertilized Oocytes and Arrested Embryos. Annals of the New York Academy of Sciences 2005, 1042: 177-185. [CrossRef]
- Motta, P.M., Nottola, S.A., Makabe, S., Heyn, R. Mitochondrial morphology in human fetal and adult female germ cells, Human Reproduction, 2000, 15:129–147. [CrossRef]
- Rodríguez-Nuevo, A., Torres-Sanchez, A., Duran, J.M. et al. Oocytes maintain ROS-free mitochondrial metabolism by suppressing complex I. Nature 2022, 607:756–761. [CrossRef]
- Surmacki, J.M., Abramczyk, H., Sobkiewicz, B., Walczak-Jędrzejowska, R., Słowikowska-Hilczer, J., Marchlewska, K. The role of cytochrome c in mitochondrial metabolism of human oocytes. bioRxiv 2024.10.01.616010. [CrossRef]
- Baştu, E.; Parlatan, U.; Başar, G.; Yumru, H.; Bavili, N.; Sağ, F.; Bulgurcuoğlu, S.; Buyru, F. Spectroscopic analysis of embryo culture media for predicting reproductive potential in patients undergoing in vitro fertilization. Turk J Obstet Gynecol. 2017, 14(3):145-150. [CrossRef]
- Iles, R.K.; Sharara, F.I.; Zmuidinaite, R.; Abdo, G.; Keshavarz, S.; Butler, S.A. Secretome profile selection of optimal IVF embryos by matrix-assisted laser desorption ionization time-of-flight mass spectrometry. J Assist Reprod Genet. 2019, 36(6):1153-1160.
- Nagy, Z.P.; Sakkas, D.; Behr, B. Symposium: innovative techniques in human embryo viability assessment. Non-invasive assessment of embryo viability by metabolomic profiling of culture media ('metabolomics'). Reprod Biomed Online. 2008, 17(4):502-507. [CrossRef]
- Iles, RK, Zmuidinaite R, Iles JK, Nasser S. The influence of Hatching on blastocyst metabolomic analysis: Mass Spectral analysis of Spent blastocyst media in Ai/ML prediction of IVF Embryo implantation potential– To Hatch or not to Hatch? PREprints.org 2025. [CrossRef]
- Iles, R.K., Zmuidinaite, R., Iles, J.K., Nasser, S. The influence of defining desired outcomes on prediction algorithms: Mass Spectral analysis of Spent blastocyst media in Ai/ML prediction of IVF Embryo implantation potential – Implantation, or Viability, or both? PREprints, 2025. [CrossRef]
- Iles, RK, Zmuidinaite R, Iles JK, Nasser S. The development of a robust and user friendly preimplantation metabolic test of blastocyst competence. PMT-BC: Mass Spectral analysis of Spent blastocyst media in Ai/ML-Bayesian prediction of IVF Embryo implantation potential. 2025. [CrossRef]
- Tesaik , J. Noninvasive Biomarkers of Human Embryo Developmental Potential PREprints.org 2025. https://www.preprints.org/manuscript/202504.1568/v1.
- Zmuidinaite, R.; Sharara, F. I.; Iles, R.K. “Current advancements in noninvasive profiling of the embryo culture media secretome,” Int. J. Mol. Sci., 2021, 22(5):2513.
- Pais, R.J.; Sharara, F.; Zmuidinaite, R.; Butler, S.; Keshavarz, S.; Iles, R. Bioinformatic identification of euploid and aneuploid embryo secretome signatures in IVF culture media based on MALDI-ToF mass spectrometry. J Assist Reprod Genet. 2020, 37(9):2189-2198. [CrossRef]
- Iqbal Hamza, I., Dailey. One ring to rule them all: Trafficking of heme and heme synthesis intermediates in the metazoans. Biochimica et Biophysica Acta (BBA) - Molecular Cell Research, 2012, 1823(9): 1617-1632. [CrossRef]
- Tsiftsoglou, A.S., Athina I. Tsamadou. Heme as key regulator of major mammalian cellular functions: Molecular, cellular, and pharmacological aspects. Pharmacology & Therapeutics, 2006, 111(2): 327-345. [CrossRef]
- Rinaldo, S., Brunori, M., & Cutruzzola, F. Ancient hemes for ancient catalysts. Plant Signaling & Behavior, 2008, 3(2):135–136. [CrossRef]
- Yoshikawa, S., Muramoto, K., Shinzawa-Itoh, K. The O2 reduction and proton pumping gate mechanism of bovine heart cytochrome c oxidase, Biochimica et Biophysica Acta (BBA) - Bioenergetics, 2011,1807(10):1279-1286. [CrossRef]
- Margarita G. Skalnaya, Alexey A. Tinkov, Yulia N. Lobanova, Jung-Su Chang, Anatoly V. Skalny . Serum levels of copper, iron, and manganese in women with pregnancy, miscarriage, and primary infertility, Journal of Trace Elements in Medicine and Biology, 2019, 56:124-130. [CrossRef]
- Yin, L., Wu, N., Curtain J.C., Qatanani, M., Szxwegold N.R., Reid, R.A., Waitt, G.M., Parks, D.J., Pearce, K.H., Wisely, G.B., Lazer, M.A. Rev-erbα, a Heme Sensor That Coordinates Metabolic and Circadian Pathways. Science 2007, 318:1786-1789. [CrossRef]
- Dunaway LS, Loeb SA, Petrillo S, Tolosano E, Isakson BE. Heme metabolism in nonerythroid cells J. Biol. Chem. 2024, 300(4):107132. [CrossRef]
- Chiabrando, D., Fiorito, V., Petrillo, S., Bertino, F., and Tolosano, E. HEME: a neglected player in nociception? Neurosci. Biobehav Rev. 2021, 124: 124–136.
- Detraux, D., Caruso, M., Feller, L., Fransolet, M., Meurant, S., Mathieu, J., Arnould, T., Renard, P. A critical role for heme synthesis and succinate in the regulation of pluripotent states transitions. eLife 2023, 12:e78546. [CrossRef]
- Gafni O, Weinberger L, Mansour AA, Manor YS,Chomsky E,Ben-Yosef D, Kalma Y, Viukov S, Maza I, Zviran A, Rais Y, Shipony Z, Mukamel Z, Krupalnik V, Zerbib M, Geula S, Caspi I, Schneir D, Shwartz T, Gilad S, Amann-Zalcenstein D, Benjamin S, Amit I, Tanay A Massarwa R, Novershtern N, Hanna JH. Derivation of novel human ground state naïve pluripotent stem cells, Nature 2013 504:282–286. [CrossRef]
- Ware CB, Nelson AM, Mecham B, Hesson J, Zhou W, Jonlin EC, Jimenez-Caliani AJ, Deng X, Cavanaugh C, Cook S, Tesar PJ, Okada J, Margaretha L, Sperber H, Choi M, Blau CA, Treuting PM, Hawkins RD, Cirulli V, Ruohola-Baker H. (2014) Derivation of naive human embryonic stem cells, PNAS 2014 111:4484–4489. [CrossRef]
- Theunissen TW Friedli M He Y Planet E O’Neil RC Markoulaki S Pontis J Wang H Iouranova A Imbeault M Duc J Cohen MA Wert KJ Castanon R Zhang Z Huang Y Nery JR Drotar J Lungjangwa T Trono D Ecker JR Jaenisch R. Molecular Criteria for Defining the Naive Human Pluripotent State. Cell Stem Cell 2016, 19:502–515. [CrossRef]
- Sahakyan A Kim R Chronis C Sabri S Bonora G Theunissen TW Kuoy E Langerman J Clark AT Jaenisch R Plath K. Human Naive Pluripotent Stem Cells Model X Chromosome Dampening and X Inactivation. Cell Stem Cell 2017, 20:87–101. [CrossRef]
- Sperber H Mathieu J Wang Y Ferreccio A Hesson J Xu Z Fischer KA Devi A Detraux D Gu H Battle SL Showalter M Valensisi C Bielas JH Ericson NG Margaretha L Robitaille AM Margineantu D Fiehn O Hockenbery D Blau CA Raftery D Margolin AA Hawkins RD Moon RT Ware CB Ruohola-Baker H. The metabolome regulates the epigenetic landscape during naive-to-primed human embryonic stem cell transition Nature Cell Biology 2015 17:1523–1535. [CrossRef]
- Zhou W Choi M Margineantu D Margaretha L Hesson J Cavanaugh C Blau CA Horwitz MS Hockenbery D Ware C Ruohola-Baker H. HIF1α induced switch from bivalent to exclusively glycolytic metabolism during ESC-to-EpiSC/hESC transition The EMBO Journal 2012, 31:2103–2116. [CrossRef]
- Sustáčková G Legartová S Kozubek S Stixová L Pacherník J Bártová E. Differentiation-independent fluctuation of pluripotency-related transcription factors and other epigenetic markers in embryonic stem cell colonies Stem Cells and Development 2012, 21:710–720. [CrossRef]
- Macfarlan TS Gifford WD Driscoll S Lettieri K Rowe HM Bonanomi D Firth A Singer O Trono D Pfaff SL. Embryonic stem cell potency fluctuates with endogenous retrovirus activity. Nature 2012, 487:57–63. [CrossRef]
- Ishiuchi T Enriquez-Gasca R Mizutani E Bošković A Ziegler-Birling C Rodriguez-Terrones D Wakayama T Vaquerizas JM Torres-Padilla ME. Early embryonic-like cells are induced by downregulating replication-dependent chromatin assembly Nature Structural & Molecular Biology 2015, 22:662–671. [CrossRef]
- Percharde M Lin CJ Yin Y Guan J Peixoto GA Bulut-Karslioglu A Biechele S Huang B Shen X Ramalho-Santos M. A LINE1-Nucleolin Partnership Regulates Early Development and ESC Identity Cell 2018, 174:391–405. [CrossRef]
- Cohen J, Scott R, Alikani M, Schimmel T, Munné S, Levron J, Wu L, Brenner C, Warner C, Willadsen S. Ooplasmic transfer in mature human oocytes. Mol Hum Reprod. 1998 4(3):269-80. [CrossRef]
- Hyslop, L., Blakeley, P., Craven, L. et al. Towards clinical application of pronuclear transfer to prevent mitochondrial DNA disease. Nature 2016 534, 383–386 (2016). [CrossRef]
- Newson AJ, Wilkinson S, Wrigley A. Ethical and legal issues in mitochondrial transfer. EMBO Mol Med. 2016 Jun 1;8(6):589-91. [CrossRef]
- Costa-Borges N, Nikitos E, Späth K, Miguel-Escalada I, Ma H, Rink K, Coudereau C, Darby H, Koski A, Van Dyken C, Mestres E, Papakyriakou E, De Ziegler D, Kontopoulos G, Mantzavinos T, Vasilopoulos I, Grigorakis S, Prokopakis T, Dimitropoulos K, Polyzos P, Vlachos N, Kostaras K, Mitalipov S, Calderón G, Psathas P, Wells D. First pilot study of maternal spindle transfer for the treatment of repeated in vitro fertilization failures in couples with idiopathic infertility. Fertil Steril. 2023 Jun;119(6):964-973. [CrossRef]
- Kang MH, Kim YJ, Lee JH. Mitochondria in reproduction. Clin Exp Reprod Med. 2023 50(1):1-11. [CrossRef]
- Raziye Melike Yildirim, Emre Seli, Mitochondria as therapeutic targets in assisted reproduction, Human Reproduction, 2024, 39(10): 2147-2159. [CrossRef]
- Subirá J, Soriano MJ, Del Castillo LM, de Los Santos MJ. Mitochondrial replacement techniques to resolve mitochondrial dysfunction and ooplasmic deficiencies: where are we now? Hum Reprod. 2025 Apr 1;40(4):585-600. [CrossRef]
- Callaway, E. ‘Landmark’ study: three-person IVF leads to eight healthy children. Long-awaited results suggest that mitochondrial donation can prevent babies from inheriting diseases caused by mutant mitochondria. Nature 2025, 643:891. [CrossRef]
- Hyslop, L.A., Blakely, E.L., Aushev, M., Marley, J., Takeda, Y., Pyle, A., Moody, E., Feeney, C., Dutton, J., Shaw, C. and Smith, S.J., 2025. Mitochondrial donation and preimplantation genetic testing for mtDNA disease. New England Journal of Medicine, 2025 393(5):438-449. [CrossRef]
- McFarland, R., Hyslop, L.A., Feeney, C., Pillai, R.N., Blakely, E.L., Moody, E., Prior, M., Devlin, A., Taylor, R.W., Herbert, M. and Choudhary, M. Mitochondrial Donation in a Reproductive Care Pathway for mtDNA Disease. N Engl J Med. 2025, 393(5):461-468.
- Ethics Committee of the American Society for Reproductive Medicine. Use of preimplantation genetic testing for monogenic adult-onset conditions: an Ethics Committee opinion. Fertil. Steril. 2024, 122 (4): 607–611. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



