
Submitted:
15 September 2025
Posted:
17 September 2025
You are already at the latest version
Abstract
Synthetic biology is undergoing a pivotal shift from single-species engineering to rational design of synthetic microbial consortia, enabling more complex, efficient, and resilient functionalities. This transition responds to the growing demand for microbial assemblies with synchronized behavior, division of labor, and ecological stability for applications in synthetic biology, biomanufacturing, bioremediation, and living therapeutics. The current single-species-centric paradigm cannot fully capture the emergent properties of microbial communities, such as temporal coordination, interspecies communication, ecological interactions, and niche specialization. Unlocking the full potential of multicellular synthetic biology therefore requires a fundamentally new design framework. In this perspective article, we introduce the symphony metaphor as a conceptual framework for engineering synthetic consortia, where microbes act as individual musicians, communication pathways as notes, genetic oscillators as rhythm, and artificial ecological dynamics as harmony, conducted through the integration of computational and experimental approaches. We synthesize advances in four interconnected areas: microbial communication as the language of microbial ensembles; oscillatory modules for temporal synchronization; synthetic ecology for stable, niche-specialized communities with controlled population size; and integrative modelling frameworks that unite metabolic and ecological design. We also outline future priorities, including modular and scalable engineering toolkits, real-time monitoring with feedback control, structural elucidation of signaling molecules, process-scale strategies for industrial deployment, and the creation of dedicated software platforms for designing multi-microbial systems. This framework aims to reshape synthetic biology to better address the complexities of multicellular systems, enhancing the rational design of synthetic microbial consortia for applications in sustainability, health, and industry.
Keywords:
synthetic biology
; synthetic microbial consortia
; synthetic ecology
; interspecies communication
; genetic oscillators
1. Introduction: From Unicellular to Multicellular Synthetic Biology
Synthetic biology has evolved from its early focus on the engineering of single microbial species into a broader, more ambitious discipline that aims to design and control complex, multicellular systems [1,2,3,4,5,6]. The formative years of the field were dominated by model chassis such as Escherichia coli, Corynebacterium glutamicum, Bacillus subtilis, Saccharomyces cerevisiae and Pichia pastoris, which provided tractable platforms for developing foundational genetic circuits, biosensors, and metabolic pathways [2,7,8,9,10]. These monoculture-based systems were instrumental for proof-of-concept demonstrations and for establishing the design–build–test–learn framework [7]. However, they could not reproduce the emergent behaviors and ecological interactions that define microbial life in natural environments [11,12]. Current biological synthetic processes, therefore, usually rely on fully sterile conditions and liquid feedstocks, making them costly and far removed from “real-life” microbial ecosystems such as those underpinning food industry fermentations (e.g., beer, cocoa, wine, kimchi, or citric acid production).
In nature, microbes rarely exist in isolation. They form self-organized communities whose functionality arises from competitive and cooperative interactions, including syntropy, cross-feeding, and mutualism [11,13]. Reproducing these properties in engineered systems requires moving beyond monoculture engineering toward the rational design of synthetic microbial consortia [3,5,11,14]. Such consortia can be tailored to perform complex, distributed tasks through mechanisms that are difficult or impossible to encode within a single organism. The demand for these systems is rising across diverse fields: in biomanufacturing and synthetic biology, division of labor reduces metabolic burden and increases yields, and can improve gene (cluster) integration and genetic stability; in bioremediation, cooperative degradation pathways enable the breakdown of complex pollutant mixtures; and in living therapeutics, engineered communities can deliver drugs, modulate immunity, or restore microbiome balance, outperforming single-strain probiotics [1,5,6,14,15,16,17,18,19,20].
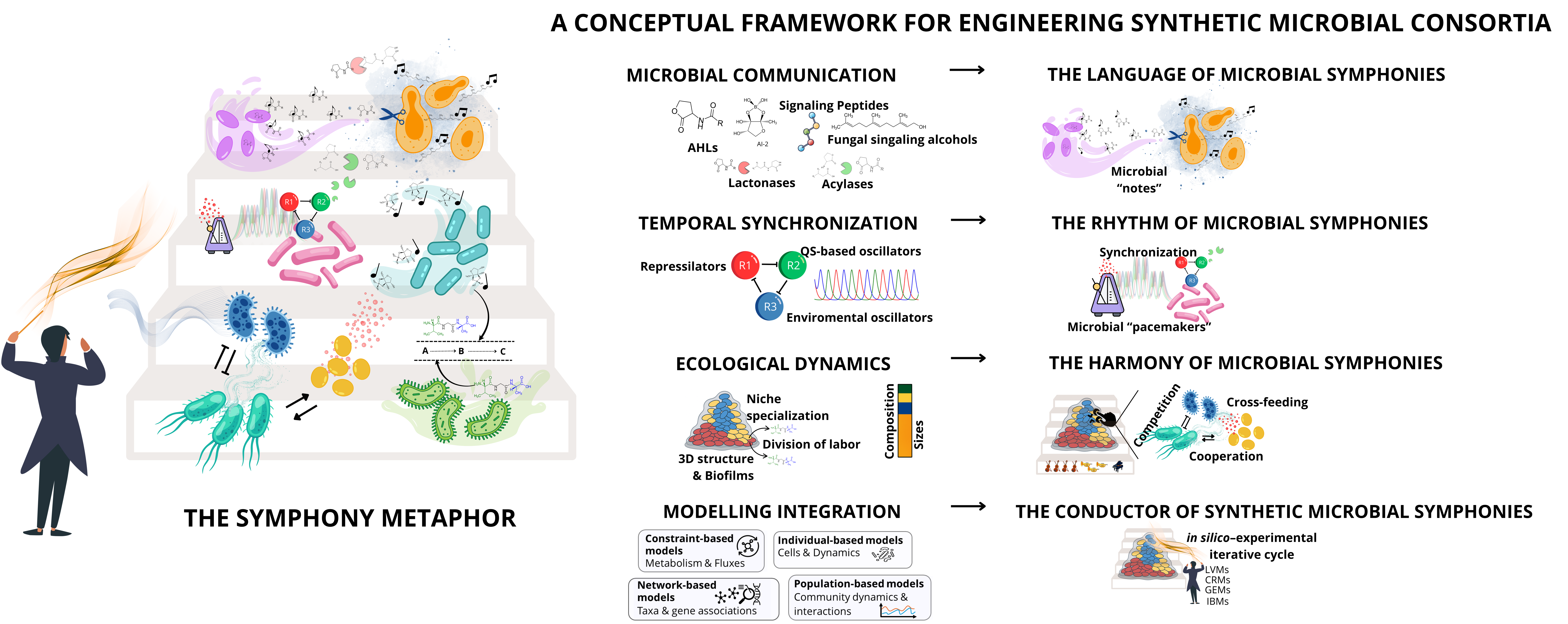
Designing synthetic consortia with predictable functionality remains a major challenge. These systems must integrate multiple layers of control (Figure 1), including precise microbial communication, robust temporal synchronization, and stable ecological structuring, while also being guided by predictive modelling that captures both metabolic and ecological dynamics, including metabolite transport and kinetics between the species or strains [3,5,11,14]. Figure 1 presents this vision and summarizes four fundamental objectives for the development of synthetic consortia synchronized with predictable functionalities: (i) optimizing microbial communication through high-precision quorum sensing systems, (ii) achieving precise temporal synchronization through genetic oscillators, (iii) simulating and stabilizing ecological interactions that promote coexistence and functional division of labor, and (iv) designing microbial communities based on integrative in silico models.
This combination of communication, timing, ecological harmony, and computational integration defines the frontier of multicellular synthetic biology. This perspective article aims to advance that frontier by introducing the symphony metaphor as a conceptual framework for designing synthetic consortia, examining recent progress in quorum sensing, temporal synchronization, synthetic ecology, and integrative modelling, and outlining future priorities to enhance the rational design of multi-microbial systems for applications in sustainability, health, and industry.
2. From Circuits to Symphonies: A Paradigm Shift
From its inception, synthetic biology has been guided by metaphors that both defined its conceptual boundaries and inspired its methodologies. In its early days, the field drew heavily on the analogy to electronic circuits, with genes envisioned as interchangeable parts, promoters as switches, and regulatory networks as logic gates [21,22]. This metaphor helped create a structured design language, simplifying complex cellular processes into standardized, modular units. It enabled predictability and modularity in single-species systems, but its linear and deterministic logic mirrored the behavior of isolated devices more than the complex and adaptive dynamics of living communities [21,23,24].
As the scope of synthetic biology expanded to encompass multi-species systems, the circuit metaphor began to show its limitations. While effective for describing controlled processes in isolated chassis, it fell short in representing the bidirectional feedback between species, the adaptive plasticity of microbial ecosystems, and the emergent properties that arise only in the context of community life [11,13,25]. Addressing these complexities requires a novel conceptual framework that captures diversity, coordination, and adaptive stability, qualities that are difficult to convey through the rigid imagery of electronic wiring diagrams. In this regard, we propose the symphony metaphor as such a framework for engineering synthetic microbial consortia. In this analogy, diverse microbial species act as specialized musicians, each contributing unique metabolic capabilities and ecological roles. Their interactions are coordinated through molecular "notes" and "rhythms", such as interspecies communication networks (including quorum sensing, quorum quenching, and metabolic cross-feeding) together with genetic oscillators. These oscillators provide a shared temporal framework, aligning the activity of different community members in recurring cycles and thereby preventing desynchronization that could disrupt collective performance.
Ecological stability emerges as harmony, achieved through niche partitioning, resource sharing, spatial partitioning and three-dimensional structuring (e.g., biofilms), as well as population size control and the mitigation of competitive exclusion. Overseeing these dynamics is the conductor, embodied by integrative modelling frameworks such as constraint-based modelling approaches and consumer–resource models (CRMs), which not only guide design choices and optimize interactions but can also act as composers, rationally engineering new ensemble arrangements to ensure the collective output remains balanced and functional over time. Figure 2 illustrates the conceptual shift from circuits (logic—linear, deterministic, and chassis-bound) to symphonies, where diverse microbial ‘musicians’ coordinate through communication, rhythm, and ecological harmony under the guidance of integrative modelling.
This metaphor aligns naturally with the emergent properties observed in both natural and synthetic communities. Just as an orchestra produces music that transcends the sum of its instruments, microbial consortia exhibit functionalities such as robustness, adaptability, and division of labor that cannot be reduced to the performance of an individual species [11,13,26,27,28]. Far from being purely aesthetic, the symphony metaphor maps directly onto the design principles explored in the following sections: Microbial communication (Section 3) defines the informational vocabulary for coordination, genetic oscillators (Section 4) set the tempo, ecological interactions (Section 5) sustain harmony, and integrated modelling (Section 6) balances metabolic potential with ecological constraints.
Metaphors have long-shaped scientific paradigms [29,30]. The electronics analogy (see above) provided a scaffold for early synthetic biology, just as the ancient concept of musica universalis (“music of the spheres”) portrayed the cosmos as a harmonious orchestration of celestial motion, an idea later refined by Johannes Kepler in Harmonices mundi to express the mathematical proportionalities of planetary orbits [31]. Likewise, modern neuroscience often uses the “neural orchestra” metaphor, where different brain regions coordinate like sections of an orchestra to produce coherent cognitive function [32]. By adopting the symphony metaphor, we aim to shift the emphasis from isolated circuit logic to coordinated ensemble performance, prioritizing coordination over isolation, diversity over uniformity, and adaptive balance over rigid control. This reframing enables multicellular synthetic biology to confront the interconnected challenges of sustainability, health, and industry by designing systems that function as living, evolving performances rather than static devices.
3. Microbial Communication: The Language of Microbial Symphonies
Interspecies communication has emerged as a central design principle in synthetic biology, with quorum sensing as one of the most versatile systems. Quorum sensing is a signalling process that coordinates collective microbial behavior within and between species, through a multilayered regulatory architecture that couples small-molecule autoinducers with transcriptional regulators and post-transcriptional checkpoints [33,34,35,36]. Recent advances have refined classic LuxI/LuxR-type modules, expanded multi-signal architectures with tunable activation thresholds, and developed strategies to directly couple quorum sensing nodes to metabolic outputs, enabling precise control over community-wide phenotypes [11,37,38,39,40,41,42,43,44,45,46,47]. The repertoire of channels has broadened beyond canonical acyl-homoserine lactone (AHL) systems to include peptide-based signaling and hybrid hierarchies, offering orthogonal modes of coordination for complex consortia [33,37,41,42,43,45,48,49] (Figure 3).
Counterbalancing these activating circuits are quorum quenching mechanisms, mediated by enzymes such as acylases, lactonases, and oxidoreductases, which degrade or modify signaling molecules to disrupt communication channels [50] (Figure 3). Quorum quenching has been applied in synthetic contexts to reduce crosstalk, prevent overactivation, and stabilize population dynamics by dampening runaway feedback. Synthetic circuits employing quorum quenching analogues illustrate how inhibitory channels can be integrated alongside activating ones, creating bidirectional “dialogues” within consortia [51].
Together, quorum sensing and quorum quenching form a dynamic molecular language of activation and inhibition. Yet engineering them for predictable functionality faces persistent challenges. Crosstalk between homologous regulators can blur signal specificity, host metabolism introduces background noise, and environmental variables such as pH, ionic strength, light (e.g., UVA radiation[52]), or enzymatic turnover alter activation, diffusion and degradation rates, shifting activation thresholds [40,41,45,53]. In structured environments, uneven spatial distribution and growth heterogeneity can desynchronize population-level responses. Overcoming these constraints requires designs that incorporate insulation and feedback, as well as spatially aware placement of signal senders and receivers. Recent strategies to improve precision include the use of insulated promoters and engineered receiver specificity to suppress crosstalk, feedback controllers to sharpen activation thresholds, and spatiotemporal patterning such as growth-phase-gated oscillators to maintain coherence [11,33,39,40,41,42,43,44,46,47,54,55]. Multi-channel systems with differential sensitivities can establish hierarchical programs that unfold in stages, enabling reliable division of labor and phase-separated bioprocessing [33,39,43,44,49,56,57].
Beyond bacterial systems, new opportunities lie in the quorum sensing and quorum quenching chemistries of fermentative yeasts. These channels could provide orthogonal signaling with distinct stability and diffusion profiles in acidic, nutrient-rich fermentations, contexts where bacterial signals often underperform [43,58,59,60]. These studies suggest the feasibility of porting yeast-derived signaling into synthetic circuits, yet systematic discovery, characterization, and standardization remain largely untapped. Another emerging frontier is the design of chemically modified autoinducers to expand quorum sensing functionality. Tailored modifications can alter sensitivity, dynamic range, and degradation kinetics, enabling finer tuning, multiplexing, and interference-resistant coordination [46,56,57,61]. Where natural chemical space is limited, customized synthetic quorum sensing analogs could create entirely new orthogonal channels [62,63,64], ideally even controllable by an external stimulus. A valuable resource in this context is the Quorumpeps® database (https://quorumpeps.ugent.be/search/), which catalogues the chemical space, microbial origin, and functional diversity of quorum-sensing peptides [65]. Beyond serving as a reference library, such datasets provide a foundation for rational design of custom-modified peptides and can be mined with machine learning tools to explore novel functional variants and expand orthogonal quorum sensing channels.
Nevertheless, quorum sensing and quorum quenching are only part of the ensemble. Microbial consortia also communicate through metabolite cross-feeding, contact-dependent signaling, and environmental sensing. Exchange of amino acids, vitamins, or other intermediates creates mutualistic couplings that stabilize composition and enhance population evenness [66,67,68]. Engineered amino acid cross-feeding in bacterial consortia, for example, has been shown to buffer competitive exclusion and reinforce robustness even in complex environments such as the mammalian gut [66]. These exchanges act as a complementary layer of molecular “notes” expanding the score beyond canonical signaling molecules.
Within the symphony metaphor, interspecies communication supplies the “notes” that score community-wide programs. Distinct channels orchestrate specific roles such as switching between biofilm and planktonic states, partitioning metabolic labor, or synchronizing production phases across species [11,33,37,39,42,46,47]. In biofilms, graded-to-binary quorum sensing and quorum quenching responses regulate matrix formation, stress tolerance, and dispersal timing, aligning local microenvironments with the needs of the larger community [33,40,41,54,69,70]. When linked to pathway branches, different quorum sensing modules can allocate upstream precursor synthesis to one population and downstream conversion to another, reducing metabolic burden and improving yield stability under fluctuating conditions [33,35,37,39,41,42,46,48,56,57,71,72]. Through feedback loops and spatial organization, microbial ensembles can also “improvise” in response to environmental fluctuations, much like an orchestra adapts to subtle changes in tempo or acoustics.
Looking forward, the challenge is to enrich this repertoire. Moving beyond classic bacterial models, future work must explore the diverse communication chemistries of fungi, yeasts, and archaea, while also developing synthetic analogues that resist interference and expand the functional space of signals. By integrating natural diversity with designed molecules, synthetic biology can compose richer and more interference-resistant communication networks. Mastering this expanded molecular language will transform microbial consortia into true symphonies, capable of precision, adaptability, and resilience in real-world applications.
4. Temporal Synchronization: Setting the Rhythm of Microbial Symphonies
Temporal synchronization in synthetic consortia is orchestrated by oscillatory genetic modules that periodically modulate gene expression, metabolic fluxes, or intercellular signals, generating predictable cycles of activity [14,33,40,73,74,75]. These modules, ranging from repressilator-based circuits to coupled quorum sensing oscillators, act as rhythmic scaffolds that align the activities of distinct community members. In multi-species settings, oscillators can be programmed to control production phases, synchronize stress responses, or trigger sequential activation of biosynthetic pathways, ensuring coordinated function even across spatially separated populations [14,54,74,76,77,78,79].
Maintaining stable rhythmicity under real-world conditions remains a central challenge. Oscillatory genetic modules typically operate on relatively fast timescales (ranging from minutes to a few hours) whereas environmental fluctuations in temperature, nutrient availability, pH and other factors often unfold over much longer periods (hours to days). These mismatched frequencies can desynchronize population-level rhythms, as slow environmental drifts shift oscillation periods or distort amplitudes [74,76,80,81,82,83]. Designing oscillators that remain robust across both short-term stochastic noise and longer-term environmental perturbations is therefore essential for predictable community-level synchronization. In structured environments such as biofilms or packed-bed reactors, uneven signal diffusion can introduce phase lags between subpopulations. Genetic instability through mutation or evolutionary drift may gradually dampen oscillatory behavior. Strategies to address these issues include incorporating negative feedback loops to sharpen phase transitions, coupling oscillators to environmental cues via multi-signal synchronization schemes, and adopting noise-resistant architectures that minimize stochastic fluctuations [74,76,80,81].
Several case studies highlight the versatility of engineered rhythmic modules. The original bacterial repressilator and synchronized quorum sensing loops have been adapted to control spatial pattern formation, timed metabolite release, and alternating biofilm morphologies [36,79,84,85,86]. In bioprocessing, oscillators have enabled “just-in-time” division of labor, where one population produces precursors during the high phase and another converts them during the low phase, reducing metabolic interference [27,84,87,88,89]. Other advances include temperature-gated oscillators to coordinate fermentation steps and light-responsive modules that synchronize photosynthetic and heterotrophic partners in mixed consortia [48,90,91]. These examples demonstrate how oscillators serve not only as abstract timing devices but also as practical controllers of spatial organization, metabolite flow, and cooperative behavior. Figure 4 illustrates how oscillatory genetic modules act as a metronome, aligning diverse microbial populations to shared rhythmic cycles that sustain coherence and coordinated outputs. The figure highlights the key oscillator types (i.e., repressilators, quorum sensing–based circuits, and environmentally gated modules), and links them to practical applications such as division of labor, biosynthetic sequencing, and stress-response synchronization.
Within the symphony metaphor, temporal synchronization functions as the metronome that keeps every microbial “musician” in time. In this symphony, interspecies communication not only provides the “notes” but also interacts with genetic oscillators and feedback loops to set the “rhythm”, coordinating the timing of gene expression and metabolic activity across the community. Rhythmic alignment prevents discordant activity that can waste resources or destabilize the community. A consortium whose oscillators drift out of phase resembles an orchestra losing its tempo, with coherence and performance rapidly degrading. Robust temporal control therefore becomes essential for predictable multicellular behavior. Achieving it will require integrating oscillatory modules with feedback regulation, multi-signal coupling, and ecological context, enabling synthetic communities to sustain a stable rhythm that withstands environmental variability while maintaining coordinated function over time.
5. Synthetic Ecology: Ecological Dynamics as the Harmony of the Symphony
Ecological stability denotes the capacity of a community to maintain its structure and function through time, encompassing persistence, resistance to perturbations, and resilience in recovering from disturbances [92]. In practice, stability emerges from the dynamic balance of competitive and cooperative interactions, often reinforced by niche partitioning, where populations specialize in distinct metabolic roles or environmental microhabitats. In engineered systems, stability also depends on controlling not only which species are present but also their relative abundances. Just as orchestras balance entire sections of violins against a single piano, synthetic consortia require fine-tuned population ratios to avoid dominance or collapse. Spatial partitioning and three-dimensional structuring (e.g., biofilms, encapsulation, microcompartments) further define where each microbial “instrument” is placed within the ensemble, shaping diffusion, nutrient access, and contact-dependent interactions. Such strategies reduce direct competition, enhance functional diversity, and buffer communities against destabilizing perturbations [93]. Importantly, stability is not solely a product of pairwise relationships but often arises from higher-order interactions that generate emergent properties beyond the sum of individual components [94,95,96].
Building on ecological theory, recent advances in synthetic ecology have applied tools from metabolic engineering, model-guided design, and directed evolution to construct microbial consortia with predefined and stable interaction architectures [38,94,96,97,98,99,100,101]. These efforts translate abstract ecological concepts into tangible design strategies by reconstructing and modeling distinct interaction modes (e.g., mutualism, competition, commensalism, interference, exploitation) in simplified systems, providing a quantitative basis to predict their dynamics [38,94]. Non-transitive motifs such as “rock-paper-scissors” dynamics can also be intentionally engineered to stabilize coexistence and prevent dominance by a single strain [94].
Nevertheless, several challenges limit the stability of engineered communities. Evolutionary escape mutants can erode engineered dependencies, cheaters can exploit cross-feeding or quorum sensing networks without contributing, and environmental heterogeneity can amplify stochastic fluctuations that drive compositional drift. Unintended interactions, such as cross-inhibition by secondary metabolites, may further destabilize networks [99,101]. Moreover, interaction outcomes are highly context-dependent: relationships cooperative in controlled conditions may invert into competition or interference under fluctuating resources or spatial constraints [97]. Even cooperative architectures may collapse due to emergent dynamics, underscoring the need to evaluate function at the systems level rather than only at pairwise interactions [94].
Addressing these issues requires adaptive ecological scaffolds that integrate genetic feedback circuits, spatial partitioning, and environmental modulation. Keystone species may serve as ecological anchors, buffering variability and maintaining coherence even under perturbations [97]. However, overreliance on keystones is risky: their centrality can create bottlenecks, render them evolutionary liabilities, or shift their ecological roles across environments, ultimately risking community collapse [102]. Thus, the design of stable consortia must incorporate redundancy and distributed resilience to avoid overdependence on single nodes. Methods such as auxotrophy-based dependencies, feedback-controlled kill-switches, or tunable metabolic cross-feeding can actively regulate consortium composition, ensuring that no strain overwhelms the ensemble [103,104,105,106]. These strategies allow population ratios to be dynamically optimized for specific tasks. Table 1 summarizes key design principles and challenges in synthetic ecology, illustrating how interaction modes (from predator–prey dynamics to engineered mutualisms and non-transitive motifs) can be orchestrated into stable consortia. Just as musical scores specify both harmony and counterpoint, these ecological scaffolds, combined with spatial partitioning and division of labor, provide the structural framework for robust microbial symphonies.
Parallels to natural fermentation ecosystems provide concrete illustrations of these principles. During wine fermentation, one of the oldest biotechnological processes known to humankind, yeasts and other microorganisms grow and interact simultaneously [107]. These interactions are often physiological and metabolic in nature, occurring either directly through cell-to-cell mechanisms [108], or indirectly via physicochemical changes in the medium caused by one strain affecting another [109]. While the wine industry has historically used pure cultures for fermentation, recent years have seen a shift towards cocultivation strategies. This approach addresses modern challenges such as reducing alcohol content and enhancing aroma production[110,111]. However, designing a microbial consortium for wine fermentation is challenging because Saccharomyces cerevisiae is highly effective in alcoholic fermentation, outcompeting and displacing other yeast and bacterial species that are dominant in the grape must and at the beginning of the process. Therefore, understanding the interaction mechanisms developed during multispecies fermentation is important for properly designing the most interesting combination of yeast strains and species [112]. Nevertheless, a clear example of division of labour can also be observed during winemaking, as most wines undergo malolactic fermentation after alcoholic fermentation. During this process, yeast species, primarily S. cerevisiae, are replaced by lactic acid bacteria, predominantly Oenococcus oeni [113,114]. These bacteria decarboxylate L-malic acid into L-lactic acid and CO₂, a transformation that reduces wine acidity, softens mouthfeel, and contributes to microbial stability. Beyond deacidification, malolactic fermentation also generates aroma-active compounds that shape the sensory profile, further illustrating how sequential microbial succession underpins complementary roles in the overall fermentation process. In cheese rinds, spatial partitioning and cross-feeding between yeasts and filamentous fungi stabilize multispecies assemblies, while bacteriocin-producing bacteria demonstrate competitive inhibition strategies [115]. Cocoa fermentations further exemplify mutualistic and commensal interactions, where yeasts liberate metabolites that fuel bacterial succession, echoing synthetic cross-feeding designs [116,117].
Other traditional beverages illustrate similar ecological motifs. Kombucha, for example, is driven by a naturally assembled, back-slopped consortium of yeasts and acetic acid bacteria (AAB) embedded within a floating cellulose-based SCOBY (Symbiotic Culture of Bacteria and Yeast). Yeast invertase hydrolyzes sucrose into glucose and fructose, which are fermented to ethanol, CO₂, and glycerol. AAB then oxidize ethanol to acetic acid and glucose to gluconic and glucuronic acids, establishing reciprocal cross-feeding and acidification that suppresses competitors. As a cellulose-producing Komagataeibacter pellicle develops at the air–liquid interface, oxygen transfer becomes spatially structured, reinforcing niche partitioning between aerobes and facultative fermenters. Feedback loops further stabilize the process: low ethanol stimulates bacterial cellulose formation, while AAB-derived acetic acid can enhance yeast ethanol production, steering succession toward a sour, low-pH endpoint [118,119,120]
Kefir offers another example of a structured, cooperative consortium. Kefir grains are gelatinous starter cultures containing ~10⁸ CFU/g lactic acid bacteria, 10⁶–10⁷ CFU/g yeasts, and ~10⁵ CFU/g acetic acid bacteria embedded in a polysaccharide matrix. Their spatial organization is stratified: rod-shaped LAB dominate the outer layer, yeasts predominate toward the core, and intermediate zones contain a balanced mix, creating gradients along the core-to-surface axis. Within the grains, yeasts and lactobacilli co-grow in balanced proportions, while broader symbioses involving yeasts, lactobacilli, and streptococci shape both fermentation dynamics and flavor. Yeasts create conditions favorable for bacterial growth and release metabolites that contribute to the beverage’s organoleptic profile. Together, bacteria and yeasts form a cooperative metabolic network that sustains lactic and alcoholic fermentations in parallel [121,122,123].
More broadly, natural fermented food systems (e.g., wine, cheese, cocoa, kombucha, and kefir) highlight how sequential inoculation, spatial organization, and metabolite cross-feeding stabilize consortia and shape functionality [124,125,126]. A recurring theme is niche complementarity: yeasts, lactic acid bacteria, acetic acid bacteria, and other microbes occupy distinct metabolic roles, reducing competition and enhancing resilience [126]. Division of labor, ecological succession, and complementarity together enable robustness and reproducibility [94,100]. Translating these lessons into synthetic systems requires identifying key ecological interactions, engineering them into defined strains, and maintaining them under controlled yet realistic conditions [127]. While heterologous pathway transfer into microorganisms is now well established, efficient transport of metabolites and cofactors in and out of cells or organelles remains a major challenge. For example, S-adenosylmethionine (SAM), an essential cofactor in many pathways that also shows metabolic toxicity at elevated concentrations, is sequestered in vacuoles by some producer yeasts, making it inaccessible to partner organisms within a consortium[128,129,130,131,132].
Within the symphony metaphor, ecological design defines not only the harmonic progression but also the ensemble composition. Some instruments are represented by entire sections, like violins, while others, such as timpani or piano, play solo roles. Similarly, microbial consortia must balance population sizes across functions, while spatial partitioning determines where each section “sits” in the orchestral arrangement. When numbers or placement are mistuned, harmony collapses; when balanced, the ensemble achieves coherence and stability.
Synthetic ecology can thus be envisioned as the art of composing microbial harmony, a dynamic equilibrium of diverse, specialized players whose coordinated interactions sustain robust and predictable functionality. Building and maintaining this harmony will depend on uniting ecological theory, advanced modelling, and insights from both engineered and traditional systems to create ecological networks that are resilient, adaptable, and functionally rich. Future progress will require explicitly incorporating higher-order ecological motifs into design principles. Although pairwise interactions have long formed the default modeling substrate, emergent higher-order interactions increasingly appear to dictate collective behavior in complex microbial consortia. Nevertheless, the debate remains open: in certain contexts, higher-order interactions may promote stability, while in others (especially when structured) they could undermine or fundamentally reshape community dynamics [94,133,134]. Equally critical is the development of adaptive monitoring frameworks capable of tracking dynamics in real time and reshaping scaffolds as communities evolve, ensuring that higher-order motifs remain functional under changing conditions [97]. Such adaptive monitoring must ultimately converge with predictive and integrative modelling, as discussed in the following section.
6. Data Integration and Modelling: The Conductor of Synthetic Microbial Orchestras
Model microbial communities or consortia, which provide simplified systems resembling natural microbial diversity, offer opportunities to investigate population dynamics and understand the role of community diversity in ecosystem performance. The complexity of synthetic microbial consortia calls for computational frameworks that unite ecological and metabolic processes into coherent design tools. Computational models can facilitate the strategic design of synthetic communities and help forecast behaviours well beyond what can be feasibly tested experimentally. A diverse set of approaches has emerged, capturing complementary dimensions of community dynamics [13,15,45,135,136,137,138,139,140,141,142,143]. These models can be broadly grouped into four categories: network-based, population-based, individual-based, and constraint-based, each contributing its own “voice” to the orchestral score of synthetic ecology (Figure 5).
Network-based models analyse abundances of microbial taxa or functions across communities. Grounded in the assumption that non-random co-occurrence patterns reflect ecological processes, they can identify potential interactions and core features [144]. Species that co-occur more often than expected by chance are more likely to share positive interactions, while organisms with high centrality in network analyses may act as keystone taxa, offering promising candidates for synthetic consortia.
Population-based models include Lotka-Volterra models (LVMs) and consumer-resource models (CRMs). LVMs represent population-level behaviour through interaction coefficients that capture growth, inhibition, and stability patterns, forming the bassline of populations and interactions [135,145,146,147]. They have been refined with non-linear feedbacks and environmental dependencies to improve stability predictions [145,146,147,148,149]. CRMs describe species interactions through the uptake and transformation of shared resources, predicting coexistence, competitive exclusion, and niche differentiation, the rhythmic foundation of resources and consumption [136,150,151,152]. Modern CRMs incorporate adaptive uptake strategies and variable stoichiometry, reflecting metabolic plasticity in dynamic environments [136,152,153,154].
Individual-based models (IBMs) simulate the behaviours of single cells in spatial environments, incorporating stochasticity, diffusion, and microcolony formation, much like solo voices that capture the dynamics of individual cells [138,141,142,155]. Increasingly, IBMs integrate mechanistic gene regulatory networks, coupling intracellular oscillators with spatial diffusion to capture emergent synchronization [138,141,156].
Constraint-based metabolic models focus on intracellular metabolism at the reaction level. Based on genome-scale metabolic models (GEMs) and flux balance analysis, they provide the harmonic detail of networks and fluxes by mapping nutrient requirements, reaction fluxes, and potential metabolic exchanges [135,137,157,158]. Originally restricted to individual species, they are now applied at the scale of communities and microbiomes, enabling analysis of metabolic division of labor and cross-feeding [159,160,161,162]. Importantly, their flexibility allows integration of multi-omics data and experimental observations as “constraints,” refining predictions and improving accuracy [157].
A critical frontier now lies in combining these approaches into hybrid frameworks that integrate metabolic and ecological variables. Constraint-based metabolic models, based on GEMs, usually assume steady-state and are therefore static in time, but they can be coupled with dynamic frameworks to orchestrate synthetic biology applications. Embedding flux predictions from GEMs into network-based models [163,164], Lotka–Volterra models [165], or individual-based models [166] enables simulations that account for both intracellular metabolism and interspecies interactions. For example, Ravikrishnan and colleagues proposed a method that combines GEMs with network-based analysis to infer all possible pathways between a source and a target metabolite, helping identify potential nutrient exchanges between organisms [164]. Brunner and Chia integrated constraint-based models with LVMs to predict probiotic engraftment in gut communities. Bauer, Zimmerman, and colleagues developed a tool that combines metabolic reaction information from GEMs with IBMs to simulate growth and interactions in space and time. While powerful, these hybrid models remain computationally demanding and require estimation of large numbers of parameters, limiting their routine use. Nevertheless, their critical value lies in supporting the rational design of consortia with balanced resource allocation, stable coexistence, and optimized metabolic output [135,167,168]. They also help identify synergistic strain combinations, optimal feedstocks, and environmental conditions that maximize intended functions [15].
Complementary multi-omics provide the quantitative “score” for this orchestration. Metagenomics, metatranscriptomics, metaproteomics, and (meta-)metabolomics together trace the flow of information from genetic potential to realized function, linking community membership to activity and phenotype. These datasets can constrain and validate models by supplying taxonomic and functional potential (metagenomics), real-time expression states (metatranscriptomics), realized enzymatic functions (metaproteomics), and metabolic outputs (metabolomics). Integrated omics therefore enable data-anchored parameterization and hypothesis testing, reducing uncertainty in model predictions [169,170,171]. Translating this integration into practice requires careful bioinformatics design: selecting omics layers matched to hypotheses, planning sampling depth and platforms to minimize bias, provisioning high-performance computing or cloud resources, and applying statistical frameworks that account for heterogeneity, compositionality, sparsity, and missingness, all within reproducible pipelines.
The value of modelling integration increases when coupled to iterative experimental validation. This process can be likened to a conductor refining an orchestra’s performance through repeated rehearsals. In practice, the conductor is also a composer: iterative model-guided design, experimental testing, and feedback refinement form the design-build-test-learn cycle. Like a musical score revised between rehearsals, models evolve with each iteration to compose new arrangements of microbial interactions. Instruments available to the conductor extend beyond in silico predictions. Real-time monitoring technologies such as online flow cytometry [172], optical biosensors [173], and Raman spectroscopy [174] provide continuous measurements of growth rates, metabolite levels, and population structures. Feeding these data into models allows dynamic recalibration, turning the conductor’s role from passive prediction to active adjustment by setting tempo, balance, and harmony in real time to maintain coherence across the ensemble.
Modelling integration therefore acts as the conductor of the microbial symphony, bringing together metabolic detail and ecological realism to orchestrate complex, stable, and productive communities. Figure 5 illustrates these complementary frameworks, their integration, and their iteration within model-experiment feedback cycles, supported by a comparative table of their focus, scale, strengths, and limitations. The convergence of static and dynamic models, reinforced by continuous data-driven refinement, offers a path to designing synthetic ecosystems that are not only predictable but also resilient and scalable for industrial, environmental, and therapeutic applications.
7. Future Perspectives and Conclusion: The Symphony of Multimicrobial Synthetic Biology
The shift from single-species engineering to multicellular synthetic biology represents a profound transformation in the way microbial systems are designed, implemented, and scaled. In this symphony, engineered microbes act as musicians with distinct capabilities. Their coordination depends on precise communication “notes”, temporal “rhythms” and ecological “harmony”, orchestrated through the integration of genetic, metabolic and ecological design principles. This perspective moves beyond the notion of synthetic consortia as simple aggregates of engineered strains, instead framing them as coordinated ensembles whose performance depends on interactions across multiple scales.
Future advances in synthetic consortia design have the potential to address global challenges in sustainability, health, and industrial biotechnology. For sustainability, distributed metabolic tasks can optimize biomass conversion, waste valorization, and carbon capture in bioenergy systems [175,176]. In health, engineered microbiomes may deliver targeted therapeutics or modulate immune responses with greater stability and specificity than single-strain probiotics [177,178]. Industrially, cooperative microbial production systems can increase yields, reduce metabolic burden, and enhance robustness in large-scale bioprocesses [179]. Realizing these possibilities will require modular and scalable design frameworks that can be rapidly adapted to new hosts, metabolic pathways, and environments [180], as well as real-time monitoring and feedback control systems capable of tracking interspecies dynamics and maintaining desired community structures [90,181]. Integration of structural elucidation tools, ideally such as NMR spectroscopy (yet to become more sensitive vs. mass spectrometry), will be essential for validating the identity and stability of signaling molecules in situ, ensuring fidelity in quorum sensing and metabolic coordination [182,183]. Process-scale engineering must bridge the gap between laboratory-scale demonstrations and industrial deployment, tackling oxygen transfer, intermediate build-up and metabolite transport, mixing efficiency, spatial heterogeneity, relative abundances of strains and species, and evolutionary stability in large-scale fermenters.
A critical barrier to progress lies in the lack of dedicated software platforms for consortia design [184]. Existing tools have streamlined the engineering of single-organism constructs, but they generally lack support for interspecies interactions, spatial organization, and dynamic feedback loops. While some initiatives begin to address aspects of ecological modelling, there is still no widely adopted environment that integrates genetic circuit design with ecological dynamics and process-scale simulation. Developing such a resource remains an urgent priority, as it would enable the predictive design of consortia from the molecular to the reactor scale.
Table 2 outlines the outstanding challenges and priorities that must be addressed for multimicrobial synthetic biology to mature, mapping the path from current limitations to future opportunities across communication, synchronization, stability, modelling, and process-scale translation. The future of multicellular synthetic biology will depend on the ability to integrate predictive modelling, high-resolution experimental data, and iterative design into a continuous feedback loop, in which in silico predictions inform strain engineering and cultivation strategies, while experimental results refine the underlying models. This process mirrors the work of a conductor, adjusting tempo, balance, and harmony to maintain coherence across the ensemble. By fully embracing the symphony metaphor and committing to the integration of communication, temporal synchronization, and ecological design, the field can produce microbial systems that are functional, robust, and adaptable to the demands of sustainability, health, and industry. Realizing this vision will require closing methodological gaps across molecular characterization, process engineering, and computational modelling, and orchestrating these dimensions into a coherent discipline of multicellular synthetic biology.
Acknowledgments
IATA-CSIC received funding from the Spanish government, ref. MCIN/AEI/10.13039/501100011033, as a ‘Severo Ochoa’ Center of Excellence (CEX2021-001189-S). VRM was supported by the Spanish Ministry of Science and Innovation (RYC2023-042907-I).
References
- Contreras-Salgado, E.A.; Sánchez-Morán, A.G.; Rodríguez-Preciado, S.Y.; Sifuentes-Franco, S.; Rodríguez-Rodríguez, R.; Macías-Barragán, J.; Díaz-Zaragoza, M. Multifaceted Applications of Synthetic Microbial Communities: Advances in Biomedicine, Bioremediation, and Industry. Microbiol. Res. 2024, 15, 1709–1727. [CrossRef]
- Bittihn, P.; Din, M.O.; Tsimring, L.S.; Hasty, J. Rational engineering of synthetic microbial systems: from single cells to consortia. Curr. Opin. Microbiol. 2018, 45, 92–99. [CrossRef]
- Rapp, K.M.; Jenkins, J.P.; Betenbaugh, M.J. Partners for life: building microbial consortia for the future. Curr. Opin. Biotechnol. 2020, 66, 292–300. [CrossRef]
- Ben Said, S.; Tecon, R.; Borer, B.; Or, D. The engineering of spatially linked microbial consortia – potential and perspectives. Curr. Opin. Biotechnol. 2020, 62, 137–145. [CrossRef]
- Mccarty, N.S.; Ledesma-Amaro, R. Synthetic biology tools to engineer microbial communities for biotechnology. Trends Biotechnol. 2019, 37, 181–197. [CrossRef]
- Brenner, K.; You, L.; Arnold, F.H. Engineering microbial consortia: a new frontier in synthetic biology. Trends Biotechnol. 2008, 26, 483–489. [CrossRef]
- Jeong, K.J. Advancing microbial engineering through synthetic biology. J. Microbiol. 2025, 63, e2503100. [CrossRef]
- León-Buitimea, A.; Balderas-Cisneros, F.d.J.; Garza-Cárdenas, C.R.; Garza-Cervantes, J.A.; Morones-Ramírez, J.R. Synthetic Biology Tools for Engineering Microbial Cells to Fight Superbugs. Front. Bioeng. Biotechnol. 2022, 10, 869206. [CrossRef]
- Liu, Y.; Su, A.; Li, J.; Ledesma-Amaro, R.; Xu, P.; Du, G.; Liu, L. Towards next-generation model microorganism chassis for biomanufacturing. Appl. Microbiol. Biotechnol. 2020, 104, 9095–9108. [CrossRef]
- Kim, J.; Salvador, M.; Saunders, E.; González, J.; Avignone-Rossa, C.; Jiménez, J.I.; Pinheiro, V.B. Properties of alternative microbial hosts used in synthetic biology: towards the design of a modular chassis. Essays Biochem. 2016, 60, 303–313. [CrossRef]
- Wintermute, E.H.; Silver, P.A. Dynamics in the mixed microbial concourse. Genes Dev. 2010, 24, 2603–2614. [CrossRef]
- Konopka, A. What is microbial community ecology? ISME J 3, 1223–1230 (2009).
- Berg, N.I.v.D.; Machado, D.; Santos, S.; Rocha, I.; Chacón, J.; Harcombe, W.; Mitri, S.; Patil, K.R. Ecological modelling approaches for predicting emergent properties in microbial communities. Nat. Ecol. Evol. 2022, 6, 855–865. [CrossRef]
- Kim, H.; Jin, X.; Glass, D.S.; Riedel-Kruse, I.H. Engineering and modeling of multicellular morphologies and patterns. Curr. Opin. Genet. Dev. 2020, 63, 95–102. [CrossRef]
- Ibrahim, M.; Raajaraam, L.; Raman, K. Modelling microbial communities: Harnessing consortia for biotechnological applications. Comput. Struct. Biotechnol. J. 2021, 19, 3892–3907. [CrossRef]
- Sgobba, E.; Wendisch, V.F. Synthetic microbial consortia for small molecule production. Curr. Opin. Biotechnol. 2020, 62, 72–79. [CrossRef]
- Che, S.; Men, Y. Synthetic microbial consortia for biosynthesis and biodegradation: promises and challenges. J. Ind. Microbiol. Biotechnol. 2019, 46, 1343–1358. [CrossRef]
- Jia, X.; Liu, C.; Song, H.; Ding, M.; Du, J.; Ma, Q.; Yuan, Y. Design, analysis and application of synthetic microbial consortia. Synth. Syst. Biotechnol. 2016, 1, 109–117. [CrossRef]
- Ortiz-Marquez, J.C.F.; Nascimento, M.D.; Zehr, J.P.; Curatti, L. Genetic engineering of multispecies microbial cell factories as an alternative for bioenergy production. Trends Biotechnol. 2013, 31, 521–529. [CrossRef]
- Shong, J.; Diaz, M.R.J.; Collins, C.H. Towards synthetic microbial consortia for bioprocessing. Curr. Opin. Biotechnol. 2012, 23, 798–802. [CrossRef]
- Szymanski, E.; Scher, E. Models for DNA Design Tools: The Trouble with Metaphors Is That They Don’t Go Away. ACS Synth. Biol. 2019, 8, 2635–2641. [CrossRef]
- McLeod, C.; Nerlich, B. Synthetic biology, metaphors and responsibility. Life Sci. Soc. Policy 2017, 13, 1–13. [CrossRef]
- Teo, J.J.Y.; Woo, S.S.; Sarpeshkar, R. Synthetic Biology: A Unifying View and Review Using Analog Circuits. IEEE Trans. Biomed. Circuits Syst. 2015, 9, 453–474. [CrossRef]
- Boudry, M.; Pigliucci, M. The mismeasure of machine: Synthetic biology and the trouble with engineering metaphors. Stud. Hist. Philos. Sci. Part C: Stud. Hist. Philos. Biol. Biomed. Sci. 2013, 44, 660–668. [CrossRef]
- Alfinito, E.; Beccaria, M. Resonance for Life: Metabolism and Social Interactions in Bacterial Communities. Biophysica 2025, 5, 12. [CrossRef]
- Rafieenia, R.; Atkinson, E.; Ledesma-Amaro, R. Division of labor for substrate utilization in natural and synthetic microbial communities. Curr. Opin. Biotechnol. 2022, 75, 102706. [CrossRef]
- Tsoi, R.; Wu, F.; Zhang, C.; Bewick, S.; Karig, D.; You, L. Metabolic division of labor in microbial systems. Proc. Natl. Acad. Sci. 2018, 115, 2526–2531. [CrossRef]
- Bassler, B.L.; Losick, R. Bacterially Speaking. Cell 2006, 125, 237–246. [CrossRef]
- Montuschi, E. Metaphor in Science. in A Companion to the Philosophy of Science 277–282 (Wiley, 2017). [CrossRef]
- Grinter, A. Metaphors in Science: Lessons for Developing an Ecological Paradigm. Telos 2020, 2020, 77–91. [CrossRef]
- Kepler, J. Ioannis Keppleri Harmonices Mundi Libri V. (sumptibus Godofredi Tampachii ..., excudebat Ioannes Plancus, 1619). [CrossRef]
- Brittan, F. The Neural Orchestra: Cognitive Instrumentalities. J. Am. Music. Soc. 2024, 77, 1–63. [CrossRef]
- Boo, A.; Amaro, R.L.; Stan, G.-B. Quorum sensing in synthetic biology: A review. Curr. Opin. Syst. Biol. 2021, 28. [CrossRef]
- Abisado, R.G.; Benomar, S.; Klaus, J.R.; Dandekar, A.A.; Chandler, J.R.; Garsin, D.A. Bacterial Quorum Sensing and Microbial Community Interactions. mBio 2018, 9, e02331-17. [CrossRef]
- An, J.H.; Goo, E.; Kim, H.; Seo, Y.-S.; Hwang, I. Bacterial quorum sensing and metabolic slowing in a cooperative population. Proc. Natl. Acad. Sci. 2014, 111, 14912–14917. [CrossRef]
- Hooshangi, S.; E Bentley, W. From unicellular properties to multicellular behavior: bacteria quorum sensing circuitry and applications. Curr. Opin. Biotechnol. 2008, 19, 550–555. [CrossRef]
- Jiang, Y.; Wu, R.; Zhang, W.; Xin, F.; Jiang, M. Construction of stable microbial consortia for effective biochemical synthesis. Trends Biotechnol. 2023, 41, 1430–1441. [CrossRef]
- Li, S.; Xiao, J.; Sun, T.; Yu, F.; Zhang, K.; Feng, Y.; Xu, C.; Wang, B.; Cheng, L. Synthetic microbial consortia with programmable ecological interactions. Methods Ecol. Evol. 2022, 13, 1608–1621. [CrossRef]
- Wu, S.; Xu, C.; Liu, J.; Liu, C.; Qiao, J. Vertical and horizontal quorum-sensing-based multicellular communications. Trends Microbiol. 2021, 29, 1130–1142. [CrossRef]
- Winkle, J.J.; Karamched, B.R.; Bennett, M.R.; Ott, W.; Josić, K.; You, L. Emergent spatiotemporal population dynamics with cell-length control of synthetic microbial consortia. PLOS Comput. Biol. 2021, 17, e1009381. [CrossRef]
- Stephens, K.; Bentley, W.E. Synthetic Biology for Manipulating Quorum Sensing in Microbial Consortia. Trends Microbiol. 2020, 28, 633–643. [CrossRef]
- Kim, J.K.; Chen, Y.; Hirning, A.J.; Alnahhas, R.N.; Josić, K.; Bennett, M.R. Long-range temporal coordination of gene expression in synthetic microbial consortia. Nat. Chem. Biol. 2019, 15, 1102–1109. [CrossRef]
- Kylilis, N.; Tuza, Z.A.; Stan, G.-B.; Polizzi, K.M. Tools for engineering coordinated system behaviour in synthetic microbial consortia. Nat. Commun. 2018, 9, 1–9. [CrossRef]
- Parkin, J. M., Hsiao, V. & Murray, R. M. Engineering pulsatile communication in bacterial consortia. bioRxiv 111906, (2017).
- Scott, S.R.; Hasty, J. Quorum Sensing Communication Modules for Microbial Consortia. ACS Synth. Biol. 2016, 5, 969–977. [CrossRef]
- Ge, C.; Yu, Z.; Sheng, H.; Shen, X.; Sun, X.; Zhang, Y.; Yan, Y.; Wang, J.; Yuan, Q. Redesigning regulatory components of quorum-sensing system for diverse metabolic control. Nat. Commun. 2022, 13, 1–12. [CrossRef]
- Miano, A.; Liao, M.J.; Hasty, J. Inducible cell-to-cell signaling for tunable dynamics in microbial communities. Nat. Commun. 2020, 11, 1–8. [CrossRef]
- Bacchus, W.; Fussenegger, M. Engineering of synthetic intercellular communication systems. Metab. Eng. 2013, 16, 33–41. [CrossRef]
- Marchand, N. & Collins, C. H. Peptide-based communication system enables Escherichia coli to Bacillus megaterium interspecies signaling. Biotechnol Bioeng 110, 3003–3012 (2013).
- Fetzner, S. Quorum quenching enzymes. J. Biotechnol. 2015, 201, 2–14. [CrossRef]
- Kamath, A.; Shukla, A.; Patel, D. Quorum Sensing and Quorum Quenching: Two sides of the same coin. Physiol. Mol. Plant Pathol. 2022, 123. [CrossRef]
- Pezzoni, M.; Pizarro, R.A.; Costa, C.S. Role of quorum sensing in UVA-induced biofilm formation in Pseudomonas aeruginosa. Microbiology 2020, 166, 735–750. [CrossRef]
- Tanouchi, Y.; Tu, D.; Kim, J.; You, L.; Asthagiri, A. Noise Reduction by Diffusional Dissipation in a Minimal Quorum Sensing Motif. PLOS Comput. Biol. 2008, 4, e1000167–e1000167. [CrossRef]
- Chen, Y.; Kim, J.K.; Hirning, A.J.; Josić, K.; Bennett, M.R. Emergent genetic oscillations in a synthetic microbial consortium. Science 2015, 349, 986–989. [CrossRef]
- McMillen, D.; Kopell, N.; Hasty, J.; Collins, J.J. Synchronizing genetic relaxation oscillators by intercell signaling. Proc. Natl. Acad. Sci. 2002, 99, 679–684. [CrossRef]
- Wu, J.; Bao, M.; Duan, X.; Zhou, P.; Chen, C.; Gao, J.; Cheng, S.; Zhuang, Q.; Zhao, Z. Developing a pathway-independent and full-autonomous global resource allocation strategy to dynamically switching phenotypic states. Nat. Commun. 2020, 11, 1–14. [CrossRef]
- Gu, F.; Jiang, W.; Mu, Y.; Huang, H.; Su, T.; Luo, Y.; Liang, Q.; Qi, Q. Quorum Sensing-Based Dual-Function Switch and Its Application in Solving Two Key Metabolic Engineering Problems. ACS Synth. Biol. 2020, 9, 209–217. [CrossRef]
- Leguina, A.C.d.V.; Nieto, C.; Pajot, H.F.; Bertini, E.V.; Mac Cormack, W.; de Figueroa, L.I.C.; Nieto-Peñalver, C.G. Inactivation of bacterial quorum sensing signals N-acyl homoserine lactones is widespread in yeasts. Fungal Biol. 2018, 122, 52–62. [CrossRef]
- Jagtap, S. S., Bedekar, A. A. & Rao, C. V. Quorum Sensing in Yeast. in 235–250 (2020). [CrossRef]
- Li, L.; Pan, Y.; Zhang, S.; Yang, T.; Li, Z.; Wang, B.; Sun, H.; Zhang, M.; Li, X. Quorum sensing: cell-to-cell communication in Saccharomyces cerevisiae. Front. Microbiol. 2023, 14, 1250151. [CrossRef]
- Zargar, A.; Quan, D.N.; Bentley, W.E. Enhancing Intercellular Coordination: Rewiring Quorum Sensing Networks for Increased Protein Expression through Autonomous Induction. ACS Synth. Biol. 2016, 5, 923–928. [CrossRef]
- Geske, G.D.; O’nEill, J.C.; Blackwell, H.E. Expanding dialogues: from natural autoinducers to non-natural analogues that modulate quorum sensing in Gram-negative bacteria. Chem. Soc. Rev. 2008, 37, 1432–1447. [CrossRef]
- Igarashi, J. & Suga, H. Custom Synthesis of Autoinducers and Their Analogues. in 265–274 (2011). [CrossRef]
- Roy, V.; Smith, J.A.I.; Wang, J.; Stewart, J.E.; Bentley, W.E.; Sintim, H.O. Synthetic Analogs Tailor Native AI-2 Signaling Across Bacterial Species. J. Am. Chem. Soc. 2010, 132, 11141–11150. [CrossRef]
- Wynendaele, E.; Bronselaer, A.; Nielandt, J.; D’hOndt, M.; Stalmans, S.; Bracke, N.; Verbeke, F.; Van De Wiele, C.; De Tré, G.; De Spiegeleer, B. Quorumpeps database: chemical space, microbial origin and functionality of quorum sensing peptides. Nucleic Acids Res. 2012, 41, D655–D659. [CrossRef]
- Ziesack, M.; Gibson, T.; Oliver, J.K.W.; Shumaker, A.M.; Hsu, B.B.; Riglar, D.T.; Giessen, T.W.; DiBenedetto, N.V.; Bry, L.; Way, J.C.; et al. Engineered Interspecies Amino Acid Cross-Feeding Increases Population Evenness in a Synthetic Bacterial Consortium. mSystems 2019, 4. [CrossRef]
- Li, X.; Wu, S.; Dong, Y.; Fan, H.; Bai, Z.; Zhuang, X. Engineering Microbial Consortia towards Bioremediation. Water 2021, 13, 2928. [CrossRef]
- Kerner, A.; Park, J.; Williams, A.; Lin, X.N.; Sandler, S.J. A Programmable Escherichia coli Consortium via Tunable Symbiosis. PLOS ONE 2012, 7, e34032. [CrossRef]
- Wang, Y., Bian, Z. & Wang, Y. Biofilm formation and inhibition mediated by bacterial quorum sensing. Appl Microbiol Biotechnol 106, 6365–6381 (2022).
- Tan, C.H.; Oh, H.-S.; Sheraton, V.M.; Mancini, E.; Loo, S.C.J.; Kjelleberg, S.; Sloot, P.M.A.; Rice, S.A. Convection and the Extracellular Matrix Dictate Inter- and Intra-Biofilm Quorum Sensing Communication in Environmental Systems. Environ. Sci. Technol. 2020, 54, 6730–6740. [CrossRef]
- Goo, E.; An, J.H.; Kang, Y.; Hwang, I. Control of bacterial metabolism by quorum sensing. Trends Microbiol. 2015, 23, 567–576. [CrossRef]
- Gupta, A.; Reizman, I.M.B.; Reisch, C.R.; Prather, K.L.J. Dynamic regulation of metabolic flux in engineered bacteria using a pathway-independent quorum-sensing circuit. Nat. Biotechnol. 2017, 35, 273–279. [CrossRef]
- Grandel, N.E.; Gamas, K.R.; Bennett, M.R. Control of synthetic microbial consortia in time, space, and composition. Trends Microbiol. 2021, 29, 1095–1105. [CrossRef]
- Fussenegger, M. Synchronized bacterial clocks. Nature 2010, 463, 301–302. [CrossRef]
- Ronda, C.; Wang, H.H. Engineering temporal dynamics in microbial communities. Curr. Opin. Microbiol. 2022, 65, 47–55. [CrossRef]
- Gurevich, L.; Cohen-Luria, R.; Wagner, N.; Ashkenasy, G. Robustness of synthetic circadian clocks to multiple environmental changes. Chem. Commun. 2015, 51, 5672–5675. [CrossRef]
- Tayar, A.M.; Karzbrun, E.; Noireaux, V.; Bar-Ziv, R.H. Synchrony and pattern formation of coupled genetic oscillators on a chip of artificial cells. Proc. Natl. Acad. Sci. 2017, 114, 11609–11614. [CrossRef]
- Song, H.; Payne, S.; Tan, C.; You, L. Programming microbial population dynamics by engineered cell–cell communication. Biotechnol. J. 2011, 6, 837–849. [CrossRef]
- Curatolo, A.I.; Zhou, N.; Zhao, Y.; Liu, C.; Daerr, A.; Tailleur, J.; Huang, J. Cooperative pattern formation in multi-component bacterial systems through reciprocal motility regulation. Nat. Phys. 2020, 16, 1152–1157. [CrossRef]
- Li, Z.; Yang, Q. Systems and synthetic biology approaches in understanding biological oscillators. Quant. Biol. 2017, 6, 1–14. [CrossRef]
- Potvin-Trottier, L., Lord, N. D., Vinnicombe, G. & Paulsson, J. Synchronous long-term oscillations in a synthetic gene circuit. Nature 538, 514–517 (2016).
- Nguyen, J.; Lara-Gutiérrez, J.; Stocker, R. Environmental fluctuations and their effects on microbial communities, populations and individuals. FEMS Microbiol. Rev. 2020, 45. [CrossRef]
- Cookson, N.A.; Tsimring, L.S.; Hasty, J. The pedestrian watchmaker: Genetic clocks from engineered oscillators. FEBS Lett. 2009, 583, 3931–3937. [CrossRef]
- Dinh, C.V.; Prather, K.L.J. Development of an autonomous and bifunctional quorum-sensing circuit for metabolic flux control in engineered Escherichia coli. Proc. Natl. Acad. Sci. 2019, 116, 25562–25568. [CrossRef]
- Yi, Q.; Zhou, T. Communication-induced multistability and multirhythmicity in a synthetic multicellular system. Phys. Rev. E 2011, 83, 051907. [CrossRef]
- Kong, W.; Qian, Y.; Stewart, P.S.; Lu, T. De novo engineering of a bacterial lifestyle program. Nat. Chem. Biol. 2022, 19, 488–497. [CrossRef]
- Hamrick, G.S.; Maddamsetti, R.; Son, H.-I.; Wilson, M.L.; Davis, H.M.; You, L. Programming Dynamic Division of Labor Using Horizontal Gene Transfer. ACS Synth. Biol. 2024, 13, 1142–1151. [CrossRef]
- Takhaveev, V.; Özsezen, S.; Smith, E.N.; Zylstra, A.; Chaillet, M.L.; Chen, H.; Papagiannakis, A.; Milias-Argeitis, A.; Heinemann, M. Temporal segregation of biosynthetic processes is responsible for metabolic oscillations during the budding yeast cell cycle. Nat. Metab. 2023, 5, 294–313. [CrossRef]
- Roell, G.W.; Zha, J.; Carr, R.R.; Koffas, M.A.; Fong, S.S.; Tang, Y.J. Engineering microbial consortia by division of labor. Microb. Cell Factories 2019, 18, 1–11. [CrossRef]
- Deter, H.S.; Lu, T. Engineering microbial consortia with rationally designed cellular interactions. Curr. Opin. Biotechnol. 2022, 76, 102730–102730. [CrossRef]
- Zhao, E.M.; Zhang, Y.; Mehl, J.; Park, H.; Lalwani, M.A.; Toettcher, J.E.; Avalos, J.L. Optogenetic regulation of engineered cellular metabolism for microbial chemical production. Nature 2018, 555, 683–687. [CrossRef]
- Grimm, V.; Wissel, C. Babel, or the ecological stability discussions: an inventory and analysis of terminology and a guide for avoiding confusion. Oecologia 1997, 109, 323–334. [CrossRef]
- Mougi, A.; Kondoh, M. Diversity of Interaction Types and Ecological Community Stability. Science 2012, 337, 349–351. [CrossRef]
- Gallardo-Navarro, O.; Aguilar-Salinas, B.; Rocha, J.; Olmedo-Álvarez, G. Higher-order interactions and emergent properties of microbial communities: The power of synthetic ecology. Heliyon 2024, 10, e33896. [CrossRef]
- Madsen, J.S.; Sørensen, S.J.; Burmølle, M. Bacterial social interactions and the emergence of community-intrinsic properties. Curr. Opin. Microbiol. 2018, 42, 104–109. [CrossRef]
- Stenuit, B.; Agathos, S.N. Deciphering microbial community robustness through synthetic ecology and molecular systems synecology. Curr. Opin. Biotechnol. 2015, 33, 305–317. [CrossRef]
- Román, M.S.; Arrabal, A.; Benitez-Dominguez, B.; Quirós-Rodríguez, I.; Diaz-Colunga, J. Towards synthetic ecology: strategies for the optimization of microbial community functions. Front. Synth. Biol. 2025, 3, 1532846. [CrossRef]
- Brüls, T.; Baumdicker, F.; Smidt, H. Editorial: Synthetic Microbial Ecology. Front. Microbiol. 2021, 12. [CrossRef]
- Escalante, A.E.; Rebolleda-Gã³Mez, M.; Benã tEz, M.; Travisano, M. Ecological perspectives on synthetic biology: insights from microbial population biology. Front. Microbiol. 2015, 6, 143–143. [CrossRef]
- De Roy, K.; Marzorati, M.; Abbeele, P.V.D.; Van de Wiele, T.; Boon, N. Synthetic microbial ecosystems: an exciting tool to understand and apply microbial communities. Environ. Microbiol. 2013, 16, 1472–1481. [CrossRef]
- Dunham, M.J. Synthetic ecology: A model system for cooperation. Proc. Natl. Acad. Sci. 2007, 104, 1741–1742. [CrossRef]
- Timóteo, S.; Albrecht, J.; Rumeu, B.; Norte, A.C.; Traveset, A.; Frost, C.M.; Marchante, E.; López-Núñez, F.A.; Peralta, G.; Memmott, J.; et al. Tripartite networks show that keystone species can multitask. Funct. Ecol. 2022, 37, 274–286. [CrossRef]
- Stephens, K.; Pozo, M.; Tsao, C.-Y.; Hauk, P.; Bentley, W.E. Bacterial co-culture with cell signaling translator and growth controller modules for autonomously regulated culture composition. Nat. Commun. 2019, 10, 1–11. [CrossRef]
- Fedorec, A.J.H.; Karkaria, B.D.; Sulu, M.; Barnes, C.P. Single strain control of microbial consortia. Nat. Commun. 2021, 12, 1–12. [CrossRef]
- Grandel, N. E. et al. Long-term homeostasis in microbial consortia via auxotrophic cross-feeding. Preprint at https://doi.org/10.1101/2025.01.08.631749 (2025).
- Li, X.; Zhou, Z.; Li, W.; Yan, Y.; Shen, X.; Wang, J.; Sun, X.; Yuan, Q. Design of stable and self-regulated microbial consortia for chemical synthesis. Nat. Commun. 2022, 13, 1–9. [CrossRef]
- Liu, Y.; Rousseaux, S.; Tourdot-Maréchal, R.; Sadoudi, M.; Gougeon, R.; Schmitt-Kopplin, P.; Alexandre, H. Wine microbiome: A dynamic world of microbial interactions. Crit. Rev. Food Sci. Nutr. 2015, 57, 856–873. [CrossRef]
- Petitgonnet, C.; Klein, G.L.; Roullier-Gall, C.; Schmitt-Kopplin, P.; Quintanilla-Casas, B.; Vichi, S.; Julien-David, D.; Alexandre, H. Influence of cell-cell contact between L. thermotolerans and S. cerevisiae on yeast interactions and the exo-metabolome. Food Microbiol. 2019, 83, 122–133. [CrossRef]
- Comitini, F.; Agarbati, A.; Canonico, L.; Ciani, M. Yeast Interactions and Molecular Mechanisms in Wine Fermentation: A Comprehensive Review. Int. J. Mol. Sci. 2021, 22, 7754. [CrossRef]
- Novello, V. & de Palma, L. Viticultural strategy to reduce alcohol levels in wine. in Alcohol level reduction in wine (ed. Teissedre, P.-L.) 41–47 (ŒNOVITI INTERNATIONAL Network, 2013).
- Ciani, M.; Morales, P.; Comitini, F.; Tronchoni, J.; Canonico, L.; Curiel, J.A.; Oro, L.; Rodrigues, A.J.; Gonzalez, R. Non-conventional Yeast Species for Lowering Ethanol Content of Wines. Front. Microbiol. 2016, 7, 642–642. [CrossRef]
- Planells-Cárcel, A.; Quintas, G.; Pardo, J.; Garcia-Rios, E.; Guillamón, J.M. Exploring Proteomic and Metabolomic Interactions in a Yeast Consortium Designed to Enhance Bioactive Compounds in Wine Fermentations. Food Front. 2025, 6, 1544–1557. [CrossRef]
- Alexandre, H., Costello, P. J., Remize, F., Guzzo, J. & Guilloux-Benatier, M. Saccharomyces cerevisiae–Oenococcus oeni interactions in wine: current knowledge and perspectives. Int J Food Microbiol 93, 141–154 (2004).
- Balmaseda, A.; Rozès, N.; Bordons, A.; Reguant, C. Molecular adaptation response of Oenococcus oeni in non-Saccharomyces fermented wines: A comparative multi-omics approach. Int. J. Food Microbiol. 2022, 362, 109490. [CrossRef]
- Kovács, Á.T. A fungal scent from the cheese. Environ. Microbiol. 2020, 22, 4524–4526. [CrossRef]
- Fernández-Niño, M.; Rodríguez-Cubillos, M.J.; Herrera-Rocha, F.; Anzola, J.M.; Cepeda-Hernández, M.L.; Mejía, J.L.A.; Chica, M.J.; Olarte, H.H.; Rodríguez-López, C.; Calderón, D.; et al. Dissecting industrial fermentations of fine flavour cocoa through metagenomic analysis. Sci. Rep. 2021, 11, 8638. [CrossRef]
- Herrera-Rocha, F.; Cala, M.P.; Mejía, J.L.A.; Rodríguez-López, C.M.; Chica, M.J.; Olarte, H.H.; Fernández-Niño, M.; Barrios, A.F.G. Dissecting fine-flavor cocoa bean fermentation through metabolomics analysis to break down the current metabolic paradigm. Sci. Rep. 2021, 11, 1–18. [CrossRef]
- Bishop, P.; Pitts, E.R.; Budner, D.; Thompson-Witrick, K.A. Kombucha: Biochemical and microbiological impacts on the chemical and flavor profile. Food Chem. Adv. 2022, 1. [CrossRef]
- Aung, T.; Lee, W.-H.; Eun, J.-B. Metabolite profiling and pathway prediction of laver (Porphyra dentata) kombucha during fermentation at different temperatures. Food Chem. 2022, 397, 133636. [CrossRef]
- Laureys, D.; Britton, S.J.; De Clippeleer, J. Kombucha Tea Fermentation: A Review. J. Am. Soc. Brew. Chem. 2020, 78, 165–174. [CrossRef]
- Arslan, S. A review: chemical, microbiological and nutritional characteristics of kefir. CyTA - J. Food 2014, 13, 340–345. [CrossRef]
- Ströher, J.A.; Oliveira, W.d.C.; de Freitas, A.S.; Salazar, M.M.; Silva, L.d.F.F.d.; Bresciani, L.; Flôres, S.H.; Malheiros, P.d.S. A Global Review of Geographical Diversity of Kefir Microbiome. Fermentation 2025, 11, 150. [CrossRef]
- Prado, M.R.; Blandón, L.M.; Vandenberghe, L.P.S.; Rodrigues, C.; Castro, G.R.; Thomaz-Soccol, V.; Soccol, C.R. Milk kefir: composition, microbial cultures, biological activities, and related products. Front. Microbiol. 2015, 6, 1177. [CrossRef]
- Louw, N.L.; Lele, K.; Ye, R.; Edwards, C.B.; Wolfe, B.E. Microbiome Assembly in Fermented Foods. Annu. Rev. Microbiol. 2023, 77, 381–402. [CrossRef]
- Sooresh, M.M.; Willing, B.P.; Bourrie, B.C.T. Opportunities and Challenges of Understanding Community Assembly in Spontaneous Food Fermentation. Foods 2023, 12, 673. [CrossRef]
- Xin, Y.; Qiao, M. Towards microbial consortia in fermented foods for metabolic engineering and synthetic biology. Food Res. Int. 2025, 201, 115677. [CrossRef]
- Mittermeier, F.; Bäumler, M.; Arulrajah, P.; Lima, J.d.J.G.; Hauke, S.; Stock, A.; Weuster-Botz, D. Artificial microbial consortia for bioproduction processes. Eng. Life Sci. 2022, 23, e2100152. [CrossRef]
- Dippe, M.; Brandt, W.; Rost, H.; Porzel, A.; Schmidt, J.; Wessjohann, L.A. Rationally engineered variants of S-adenosylmethionine (SAM) synthase: reduced product inhibition and synthesis of artificial cofactor homologues. Chem. Commun. 2015, 51, 3637–3640. [CrossRef]
- Wessjohann, L., Bauer, A., Dippe, M., Ley, J. & Geißler, T. Biocatalytic Synthesis of Natural Products by <scp> O </scp> -Methyltransferases. in Applied Biocatalysis: From Fundamental Science to Industrial Applications 121–146 (Wiley, 2016). [CrossRef]
- Remines, M.; Schoonover, M.G.; Knox, Z.; Kenwright, K.; Hoffert, K.M.; Coric, A.; Mead, J.; Ampfer, J.; Seye, S.; Strome, E.D.; et al. Profiling the compendium of changes in Saccharomyces cerevisiae due to mutations that alter availability of the main methyl donor S-Adenosylmethionine. G3 Genes|Genomes|Genetics 2024, 14. [CrossRef]
- Hammer, S.K.; Avalos, J.L. Harnessing yeast organelles for metabolic engineering. Nat. Chem. Biol. 2017, 13, 823–832. [CrossRef]
- Tan, L.; Zhang, Y.; Liu, P.; Wu, Y.; Huang, Z.; Hu, Z.; Liu, Z.; Wang, Y.; Zheng, Y. System metabolic engineering modification of Saccharomyces cerevisiae to increase SAM production. Bioresour. Bioprocess. 2025, 12, 19. [CrossRef]
- Venturelli, O.S.; Carr, A.V.; Fisher, G.; Hsu, R.H.; Lau, R.; Bowen, B.P.; Hromada, S.; Northen, T.; Arkin, A.P. Deciphering microbial interactions in synthetic human gut microbiome communities. Mol. Syst. Biol. 2018, 14, e8157. [CrossRef]
- Morin, M.A.; Morrison, A.J.; Harms, M.J.; Dutton, R.J. Higher-order interactions shape microbial interactions as microbial community complexity increases. Sci. Rep. 2022, 12, 1–16. [CrossRef]
- Wu, S.; Qu, Z.; Chen, D.; Wu, H.; Caiyin, Q.; Qiao, J. Deciphering and designing microbial communities by genome-scale metabolic modelling. Comput. Struct. Biotechnol. J. 2024, 23, 1990–2000. [CrossRef]
- Plata, G.; Srinivasan, K.; Krishnamurthy, M.; Herron, L.; Dixit, P.; Gibbons, J.G. Designing host-associated microbiomes using the consumer/resource model. mSystems 2025, 10, e0106824. [CrossRef]
- Cerk, K.; Ugalde-Salas, P.; Nedjad, C.G.; Lecomte, M.; Muller, C.; Sherman, D.J.; Hildebrand, F.; Labarthe, S.; Frioux, C. Community-scale models of microbiomes: Articulating metabolic modelling and metagenome sequencing. Microb. Biotechnol. 2024, 17, e14396. [CrossRef]
- Ni, C.; Lu, T. Individual-Based Modeling of Spatial Dynamics of Chemotactic Microbial Populations. ACS Synth. Biol. 2022, 11, 3714–3723. [CrossRef]
- León, D.S.; Nogales, J. Toward merging bottom–up and top–down model-based designing of synthetic microbial communities. Curr. Opin. Microbiol. 2022, 69, 102169. [CrossRef]
- Leggieri, P.A.; Liu, Y.; Hayes, M.; Connors, B.; Seppälä, S.; O'MAlley, M.A.; Venturelli, O.S. Integrating Systems and Synthetic Biology to Understand and Engineer Microbiomes. Annu. Rev. Biomed. Eng. 2021, 23, 169–201. [CrossRef]
- Jayathilake, P.G.; Gupta, P.; Li, B.; Madsen, C.; Oyebamiji, O.; González-Cabaleiro, R.; Rushton, S.; Bridgens, B.; Swailes, D.; Allen, B.; et al. A mechanistic Individual-based Model of microbial communities. PLOS ONE 2017, 12, e0181965–e0181965. [CrossRef]
- Hellweger, F.L.; Clegg, R.J.; Clark, J.R.; Plugge, C.M.; Kreft, J.-U. Advancing microbial sciences by individual-based modelling. Nat. Rev. Microbiol. 2016, 14, 461–471. [CrossRef]
- Bernstein, H.C.; Carlson, R.P. Microbial Consortia Engineering for Cellular Factories: In Vitro to In Silico Systems. Comput. Struct. Biotechnol. J. 2012, 3, e201210017. [CrossRef]
- Toju, H.; Abe, M.S.; Ishii, C.; Hori, Y.; Fujita, H.; Fukuda, S. Scoring Species for Synthetic Community Design: Network Analyses of Functional Core Microbiomes. Front. Microbiol. 2020, 11, 1361. [CrossRef]
- Gonze, D., Coyte, K. Z., Lahti, L. & Faust, K. Microbial communities as dynamical systems. Curr Opin Microbiol 44, 41–49 (2018).
- Joseph, T.A.; Shenhav, L.; Xavier, J.B.; Halperin, E.; Pe’eR, I.; Dakos, V. Compositional Lotka-Volterra describes microbial dynamics in the simplex. PLOS Comput. Biol. 2020, 16, e1007917. [CrossRef]
- Remien, C.H.; Eckwright, M.J.; Ridenhour, B.J. Structural identifiability of the generalized Lotka–Volterra model for microbiome studies. R. Soc. Open Sci. 2021, 8. [CrossRef]
- Jin, Q. Building microbial kinetic models for environmental application: A theoretical perspective. Appl. Geochem. 2023, 158. [CrossRef]
- Marschmann, G.L.; Tang, J.; Zhalnina, K.; Karaoz, U.; Cho, H.; Le, B.; Pett-Ridge, J.; Brodie, E.L. Predictions of rhizosphere microbiome dynamics with a genome-informed and trait-based energy budget model. Nat. Microbiol. 2024, 9, 421–433. [CrossRef]
- Blumenthal, E.; Mehta, P. Geometry of ecological coexistence and niche differentiation. Phys. Rev. E 2023, 108, 044409–044409. [CrossRef]
- Muscarella, M.E.; O’dWyer, J.P. Species dynamics and interactions via metabolically informed consumer-resource models. Theor. Ecol. 2020, 13, 503–518. [CrossRef]
- Ho, P.-Y.; Nguyen, T.H.; Sanchez, J.M.; DeFelice, B.C.; Huang, K.C. Resource competition predicts assembly of gut bacterial communities in vitro. Nat. Microbiol. 2024, 9, 1036–1048. [CrossRef]
- Pacciani-Mori, L.; Giometto, A.; Suweis, S.; Maritan, A.; Maslov, S. Dynamic metabolic adaptation can promote species coexistence in competitive microbial communities. PLOS Comput. Biol. 2020, 16, e1007896. [CrossRef]
- Pacciani-Mori, L.; Giometto, A.; Suweis, S.; Maritan, A.; Maslov, S. Dynamic metabolic adaptation can promote species coexistence in competitive microbial communities. PLOS Comput. Biol. 2020, 16, e1007896. [CrossRef]
- Ferrer, J.; Prats, C.; López, D. Individual-based Modelling: An Essential Tool for Microbiology. J. Biol. Phys. 2008, 34, 19–37. [CrossRef]
- Saula, A. Y., Rowlatt, C. & Bowness, R. Use of Individual-Based Mathematical Modelling to Understand More About Antibiotic Resistance Within-Host. in 93–108 (2024). [CrossRef]
- Mahadevan, R.; Henson, M.A. Genome-Based Modeling and Design of Metabolic Interactions in Microbial Communities. Comput. Struct. Biotechnol. J. 2012, 3, e201210008. [CrossRef]
- Orth, J. D., Thiele, I. & Palsson, B. Ø. What is flux balance analysis? Nat Biotechnol 28, 245–248 (2010).
- Quinn-Bohmann, N.; Carr, A.V.; Diener, C.; Gibbons, S.M. Moving from genome-scale to community-scale metabolic models for the human gut microbiome. Nat. Microbiol. 2025, 10, 1055–1066. [CrossRef]
- Marcelino, V.R.; Welsh, C.; Diener, C.; Gulliver, E.L.; Rutten, E.L.; Young, R.B.; Giles, E.M.; Gibbons, S.M.; Greening, C.; Forster, S.C. Disease-specific loss of microbial cross-feeding interactions in the human gut. Nat. Commun. 2023, 14, 1–11. [CrossRef]
- Zorrilla, F.; Buric, F.; Patil, K.R.; Zelezniak, A. metaGEM: reconstruction of genome scale metabolic models directly from metagenomes. Nucleic Acids Res. 2021, 49, e126–e126. [CrossRef]
- Diener, C.; Gibbons, S.M.; Resendis-Antonio, O.; Chia, N. MICOM: Metagenome-Scale Modeling To Infer Metabolic Interactions in the Gut Microbiota. mSystems 2020, 5. [CrossRef]
- Levy, R.; Borenstein, E. Metabolic modeling of species interaction in the human microbiome elucidates community-level assembly rules. Proc. Natl. Acad. Sci. 2013, 110, 12804–12809. [CrossRef]
- Ravikrishnan, A.; Nasre, M.; Raman, K. Enumerating all possible biosynthetic pathways in metabolic networks. Sci. Rep. 2018, 8, 1–11. [CrossRef]
- Brunner, J.D.; Chia, N. Metabolic model-based ecological modeling for probiotic design. eLife 2024, 13. [CrossRef]
- Bauer, E.; Zimmermann, J.; Baldini, F.; Thiele, I.; Kaleta, C.; Maranas, C.D. BacArena: Individual-based metabolic modeling of heterogeneous microbes in complex communities. PLOS Comput. Biol. 2017, 13, e1005544. [CrossRef]
- Khandelwal, R.A.; Olivier, B.G.; Röling, W.F.M.; Teusink, B.; Bruggeman, F.J.; Vera, J. Community Flux Balance Analysis for Microbial Consortia at Balanced Growth. PLOS ONE 2013, 8, e64567. [CrossRef]
- Harcombe, W.R.; Riehl, W.J.; Dukovski, I.; Granger, B.R.; Betts, A.; Lang, A.H.; Bonilla, G.; Kar, A.; Leiby, N.; Mehta, P.; et al. Metabolic Resource Allocation in Individual Microbes Determines Ecosystem Interactions and Spatial Dynamics. Cell Rep. 2014, 7, 1104–1115. [CrossRef]
- McDaniel, E.A.; Wahl, S.A.; Ishii, S.; Pinto, A.; Ziels, R.; Nielsen, P.H.; McMahon, K.D.; Williams, R.B. Prospects for multi-omics in the microbial ecology of water engineering. Water Res. 2021, 205, 117608. [CrossRef]
- Walsh, A.M.; Leech, J.; Huttenhower, C.; Delhomme-Nguyen, H.; Crispie, F.; Chervaux, C.; Cotter, P.D. Integrated molecular approaches for fermented food microbiome research. FEMS Microbiol. Rev. 2023, 47. [CrossRef]
- Arıkan, M.; Muth, T. Integrated multi-omics analyses of microbial communities: a review of the current state and future directions. Mol. Omics 2023, 19, 607–623. [CrossRef]
- Mermans, F.; Chatzigiannidou, I.; Teughels, W.; Boon, N.; Newton, R.J. Quantifying synthetic bacterial community composition with flow cytometry: efficacy in mock communities and challenges in co-cultures. mSystems 2025, 10, e0100924. [CrossRef]
- Sharma, K.; Sharma, M. Optical biosensors for environmental monitoring: Recent advances and future perspectives in bacterial detection. Environ. Res. 2023, 236, 116826. [CrossRef]
- Li, J.; Luo, C.; Cai, X.; Zhang, D.; Guan, G.; Li, B.; Zhang, G. Microbial consortium assembly and functional analysis via isotope labelling and single-cell manipulation of polycyclic aromatic hydrocarbon degraders. ISME J. 2024, 18. [CrossRef]
- Zhou, S.-P.; Ke, X.; Jin, L.-Q.; Xue, Y.-P.; Zheng, Y.-G. Sustainable management and valorization of biomass wastes using synthetic microbial consortia. Bioresour. Technol. 2024, 395, 130391. [CrossRef]
- Troiano, D.T.; Studer, M.H.-P. Microbial consortia for the conversion of biomass into fuels and chemicals. Nat. Commun. 2025, 16, 1–12. [CrossRef]
- Patra, D. Synthetic Biology-Enabled Engineering of Probiotics for Precision and Targeted Therapeutic Delivery Applications. Exon 2024, 1, 54–66. [CrossRef]
- Murali, S.K.; Mansell, T.J. Next generation probiotics: Engineering live biotherapeutics. Biotechnol. Adv. 2024, 72, 108336. [CrossRef]
- Mao, J.; Zhang, H.; Chen, Y.; Wei, L.; Liu, J.; Nielsen, J.; Chen, Y.; Xu, N. Relieving metabolic burden to improve robustness and bioproduction by industrial microorganisms. Biotechnol. Adv. 2024, 74, 108401. [CrossRef]
- Abdi, G. et al. Scaling Up Nature’s Chemistry: A Guide to Industrial Production of Valuable Metabolites. in Advances in Metabolomics 331–375 (Springer Nature Singapore, Singapore, 2024). [CrossRef]
- Kong, W.; Meldgin, D.R.; Collins, J.J.; Lu, T. Designing microbial consortia with defined social interactions. Nat. Chem. Biol. 2018, 14, 821–829. [CrossRef]
- Chigozie, V.U.; Saki, M.; O Esimone, C. Molecular structural arrangement in quorum sensing and bacterial metabolic production. World J. Microbiol. Biotechnol. 2025, 41, 1–20. [CrossRef]
- Dias, D.A.; Jones, O.A.; Beale, D.J.; Boughton, B.A.; Benheim, D.; Kouremenos, K.A.; Wolfender, J.-L.; Wishart, D.S. Current and Future Perspectives on the Structural Identification of Small Molecules in Biological Systems. Metabolites 2016, 6, 46. [CrossRef]
- Oliveira, S.M.; Densmore, D. Hardware, Software, and Wetware Codesign Environment for Synthetic Biology. BioDesign Res. 2022, 2022. [CrossRef]
- Singh, A.; Lankapalli, A.K.; Mendem, S.K.; Semmler, T.; Ahmed, N.; Rappuoli, R. Unraveling the evolutionary dynamics of toxin-antitoxin systems in diverse genetic lineages of Escherichia coli including the high-risk clonal complexes. mBio 2024, 15, e0302323. [CrossRef]
- Balagaddé, F. K. et al. A synthetic Escherichia coli predator–prey ecosystem. Mol Syst Biol 4, (2008).
- Hong, Y.-J.; Cai, Y.; Antoniewicz, M.R. Cross-feeding of amino acid pathway intermediates is common in co-cultures of auxotrophic Escherichia coli. Metab. Eng. 2025, 88, 172–179. [CrossRef]
- Hanly, T.J.; Urello, M.; Henson, M.A. Dynamic flux balance modeling of S. cerevisiae and E. coli co-cultures for efficient consumption of glucose/xylose mixtures. Appl. Microbiol. Biotechnol. 2011, 93, 2529–2541. [CrossRef]
- Zhou, K.; Qiao, K.; Edgar, S.M.; Stephanopoulos, G. Distributing a metabolic pathway among a microbial consortium enhances production of natural products. Nat. Biotechnol. 2015, 33, 377–383. [CrossRef]
- Singh, G.; Yadav, M.; Ghosh, C.; Rathore, J.S. Bacterial toxin-antitoxin modules: classification, functions, and association with persistence. Curr. Res. Microb. Sci. 2021, 2, 100047. [CrossRef]
- Lin, J.; Guo, Y.; Yao, J.; Tang, K.; Wang, X. Applications of toxin-antitoxin systems in synthetic biology. Eng. Microbiol. 2023, 3, 100069. [CrossRef]
- Gupta, S.; Ross, T.D.; Gomez, M.M.; Grant, J.L.; Romero, P.A.; Venturelli, O.S. Investigating the dynamics of microbial consortia in spatially structured environments. Nat. Commun. 2020, 11, 1–15. [CrossRef]
- Spaccasassi, A.; Utz, F.; Dunkel, A.; Börner, R.A.; Ye, L.; De Franceschi, F.; Bogicevic, B.; Glabasnia, A.; Hofmann, T.; Dawid, C. Screening of a Microbial Culture Collection: Empowering Selection of Starters for Enhanced Sensory Attributes of Pea-Protein-Based Beverages. J. Agric. Food Chem. 2024, 72, 15890–15905. [CrossRef]
- de Ullivarri, M.F.; Merín, M.G.; Raya, R.R.; de Ambrosini, V.I.M.; Mendoza, L.M. Killer yeasts used as starter cultures to modulate the behavior of potential spoilage non-Saccharomyces yeasts during Malbec wine fermentation. Food Biosci. 2023, 57. [CrossRef]
- Kerr, B., Riley, M. A., Feldman, M. W. & Bohannan, B. J. M. Local dispersal promotes biodiversity in a real-life game of rock–paper–scissors. Nature 418, 171–174 (2002).
- Liao, M.J.; Miano, A.; Nguyen, C.B.; Chao, L.; Hasty, J. Survival of the weakest in non-transitive asymmetric interactions among strains of E. coli. Nat. Commun. 2020, 11, 1–8. [CrossRef]
- Chen, J.; Cai, Y.; Wang, Z.; Xu, Z.; Li, J.; Ma, X.; Zhuang, W.; Liu, D.; Wang, S.; Song, A.; et al. Construction of a synthetic microbial community based on multiomics linkage technology and analysis of the mechanism of lignocellulose degradation. Bioresour. Technol. 2023, 389, 129799. [CrossRef]
- Sui, M., Rong, J., Zhang, Y., Xie, H. & Zhang, C. Screening of Cellulose Degrading Bacteria and Construction of Complex Microflora. IOP Conf Ser Earth Environ Sci 632, 032021 (2021). [CrossRef]
Figure 1.
Conceptual roadmap for engineering synchronized synthetic consortia with predictable functionality. Schematic representation of the four foundational pillars of rational synthetic consortia design: (i) interspecies communication via finely tuned quorum sensing networks; (ii) temporal synchronization through oscillatory genetic modules; (iii) ecological stability achieved through niche partitioning, resource sharing, and keystone species; and (iv) predictive computational modelling that integrates metabolic and ecological parameters. Icons represent microbial cells, molecular signals, rhythmic oscillations, ecological interaction networks, and in silico modelling interfaces.
Figure 1.
Conceptual roadmap for engineering synchronized synthetic consortia with predictable functionality. Schematic representation of the four foundational pillars of rational synthetic consortia design: (i) interspecies communication via finely tuned quorum sensing networks; (ii) temporal synchronization through oscillatory genetic modules; (iii) ecological stability achieved through niche partitioning, resource sharing, and keystone species; and (iv) predictive computational modelling that integrates metabolic and ecological parameters. Icons represent microbial cells, molecular signals, rhythmic oscillations, ecological interaction networks, and in silico modelling interfaces.
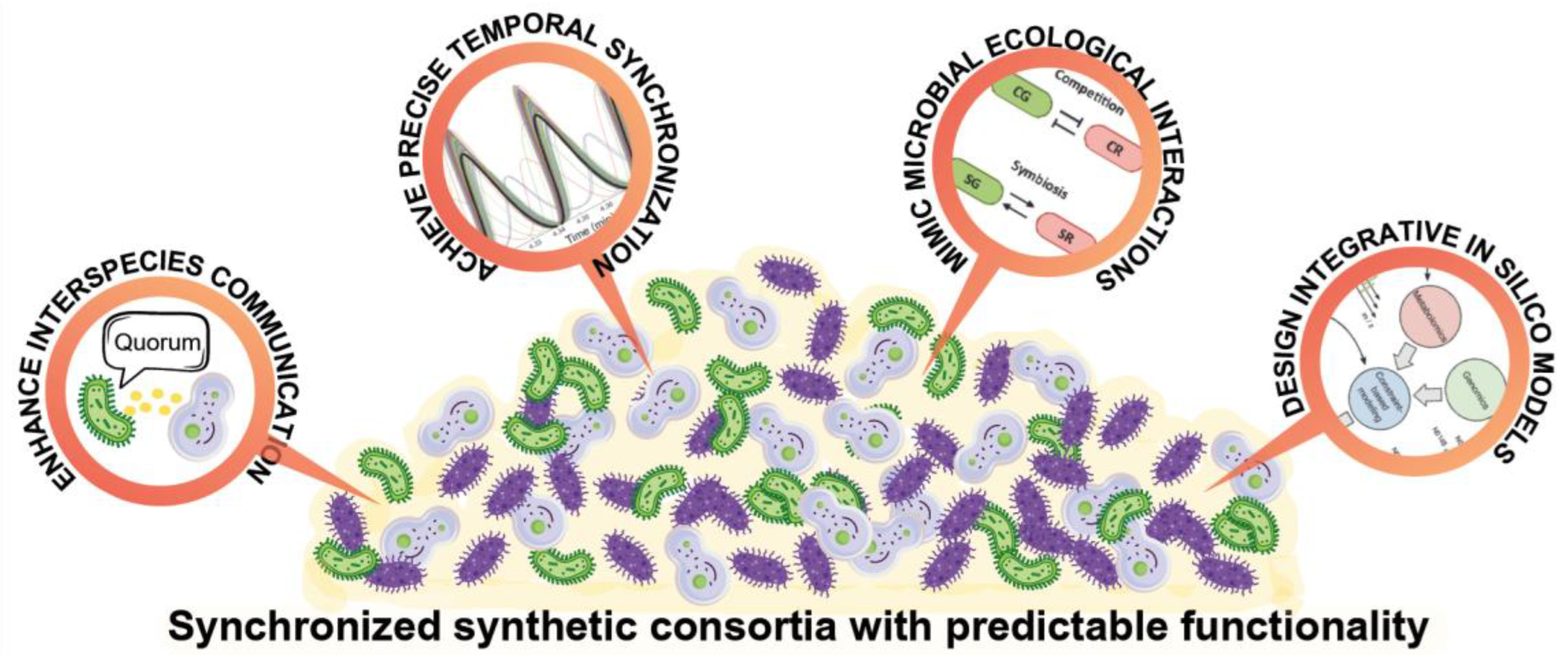
Figure 2.
From circuits to symphonies: a paradigm shift in synthetic biology. Conceptual illustration contrasting the reductionist “circuit” metaphor with the proposed “symphony” paradigm. Left: the circuit view emphasizes isolated, linear processes, where electronic components (switches, gates, and modules) are mapped onto single-cell functions such as proteins, biochemical reactions, and metabolic pathways within a microbial chassis. Right: the symphony view highlights coordinated, multi-species design, where microbes function as an ensemble. Visual metaphors represent key principles: Microbial communication as “notes,” genetic oscillators as rhythmic synchronization, division of labor as modular arrangement, and ecological dynamics as harmonic balance between cooperation and competition. A conductor figure symbolizes integrative modelling frameworks, guiding the system through an in silico–experimental iterative cycle. The figure illustrates the conceptual transition from single-species, circuit-based engineering toward ensemble-level, ecologically informed synthetic design.
Figure 2.
From circuits to symphonies: a paradigm shift in synthetic biology. Conceptual illustration contrasting the reductionist “circuit” metaphor with the proposed “symphony” paradigm. Left: the circuit view emphasizes isolated, linear processes, where electronic components (switches, gates, and modules) are mapped onto single-cell functions such as proteins, biochemical reactions, and metabolic pathways within a microbial chassis. Right: the symphony view highlights coordinated, multi-species design, where microbes function as an ensemble. Visual metaphors represent key principles: Microbial communication as “notes,” genetic oscillators as rhythmic synchronization, division of labor as modular arrangement, and ecological dynamics as harmonic balance between cooperation and competition. A conductor figure symbolizes integrative modelling frameworks, guiding the system through an in silico–experimental iterative cycle. The figure illustrates the conceptual transition from single-species, circuit-based engineering toward ensemble-level, ecologically informed synthetic design.
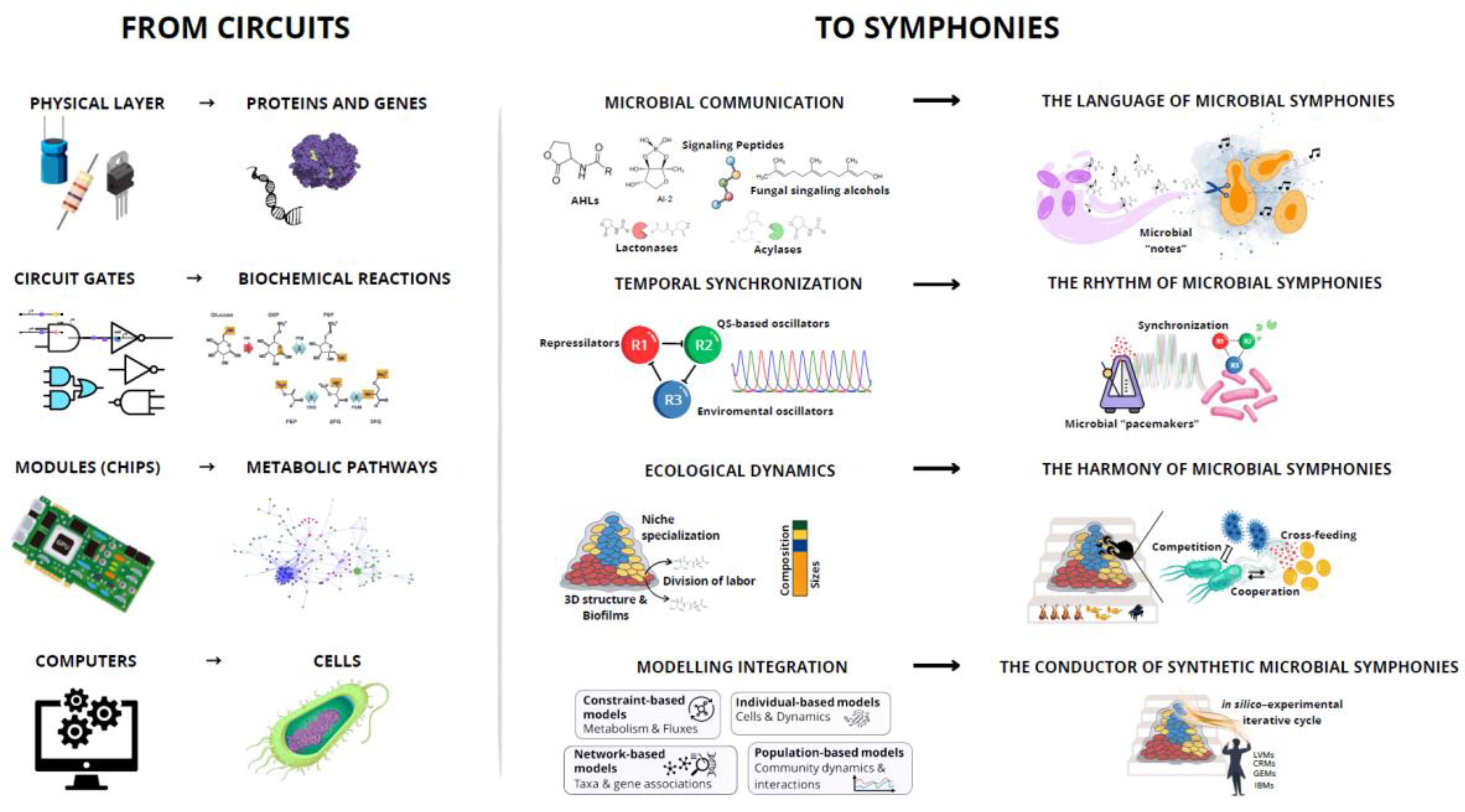
Figure 3.
Microbial communication: the language of microbial symphonies. Infographic and comparative table of quorum sensing (QS) and quorum quenching (QQ) systems relevant to synthetic consortia. Top panel: representative QS systems (AHL-based, AI-2-based, peptide-based, fungal QS, and synthetic analogs) are presented along with their key attributes (orthogonality, stability, diffusion range). Bottom panel: comparative summary of natural producers, engineering applications, advantages, and limitations. The figure highlights QS and QQ channels most suitable for the design of microbial consortia.
Figure 3.
Microbial communication: the language of microbial symphonies. Infographic and comparative table of quorum sensing (QS) and quorum quenching (QQ) systems relevant to synthetic consortia. Top panel: representative QS systems (AHL-based, AI-2-based, peptide-based, fungal QS, and synthetic analogs) are presented along with their key attributes (orthogonality, stability, diffusion range). Bottom panel: comparative summary of natural producers, engineering applications, advantages, and limitations. The figure highlights QS and QQ channels most suitable for the design of microbial consortia.
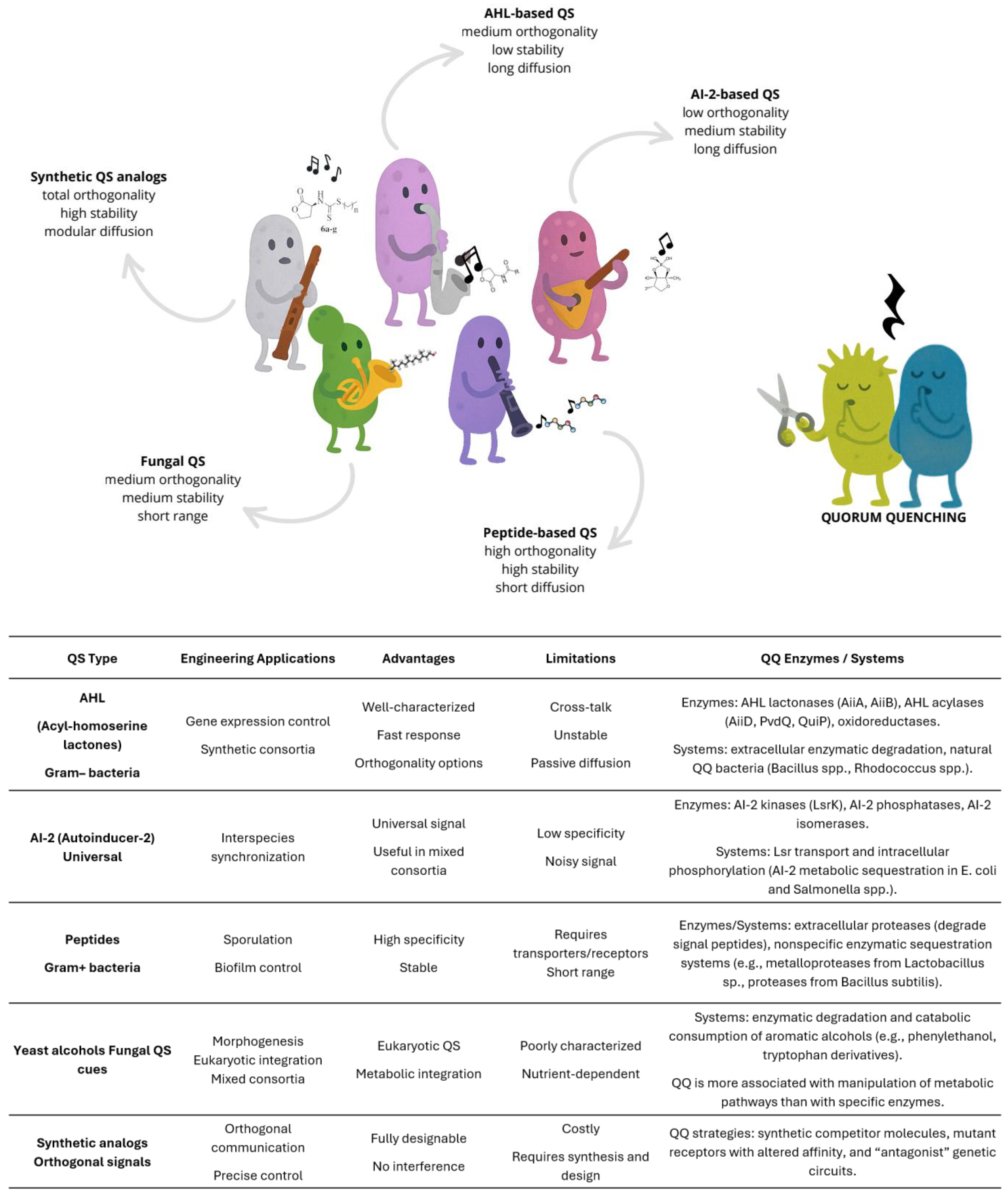
Figure 4.
Temporal synchronization: setting the rhythm of microbial symphonies. Conceptual representation of oscillatory genetic modules as temporal regulators in synthetic consortia. Multiple microbial populations align to a common metronome and harmonized waveforms, symbolizing synchronized gene expression cycles. Representative oscillator types include repressilator loops, quorum sensing–based oscillators, and modules gated by environmental cues. A side panel lists key applications: just-in-time division of labor, sequential activation of biosynthetic pathways, and synchronization of stress responses.
Figure 4.
Temporal synchronization: setting the rhythm of microbial symphonies. Conceptual representation of oscillatory genetic modules as temporal regulators in synthetic consortia. Multiple microbial populations align to a common metronome and harmonized waveforms, symbolizing synchronized gene expression cycles. Representative oscillator types include repressilator loops, quorum sensing–based oscillators, and modules gated by environmental cues. A side panel lists key applications: just-in-time division of labor, sequential activation of biosynthetic pathways, and synchronization of stress responses.
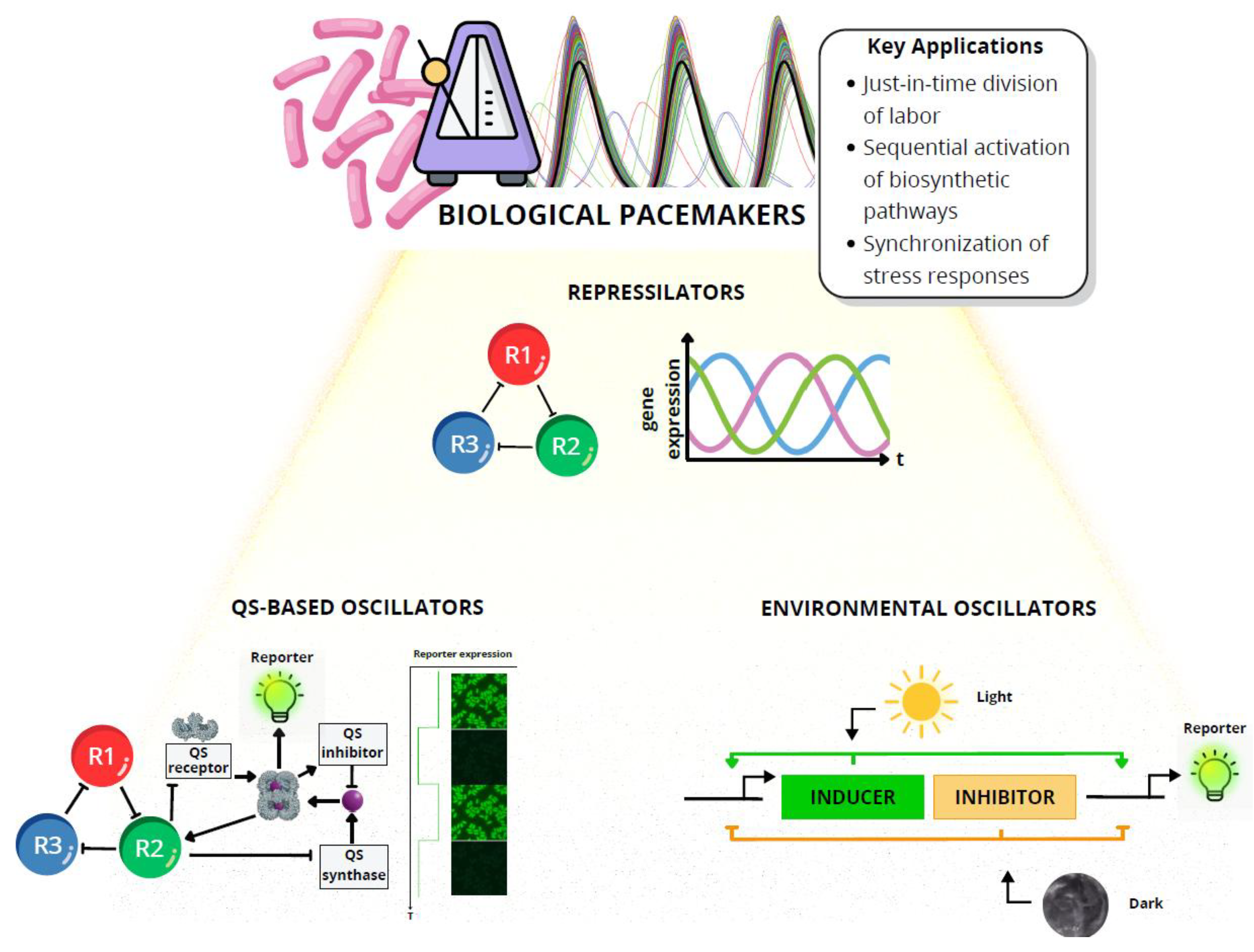
Figure 5.
Modelling integration as the conductor of synthetic microbial symphonies. Conceptual illustration of modelling approaches used to design synthetic microbial consortia. Each model captures a distinct layer of community dynamics: Lotka–Volterra models (populations and interactions), consumer–resource models (resources and consumption), constraint-based models (networks and fluxes), and individual-based models (cells and dynamics). The conductor symbolizes the iterative in silico–experimental design cycle, where predictions and data are continuously refined to orchestrate stable, productive communities. The accompanying table summarizes the focus, scale, strengths, and limitations of each modelling framework, highlighting how their integration enables a global perspective on ecological and metabolic design.
Figure 5.
Modelling integration as the conductor of synthetic microbial symphonies. Conceptual illustration of modelling approaches used to design synthetic microbial consortia. Each model captures a distinct layer of community dynamics: Lotka–Volterra models (populations and interactions), consumer–resource models (resources and consumption), constraint-based models (networks and fluxes), and individual-based models (cells and dynamics). The conductor symbolizes the iterative in silico–experimental design cycle, where predictions and data are continuously refined to orchestrate stable, productive communities. The accompanying table summarizes the focus, scale, strengths, and limitations of each modelling framework, highlighting how their integration enables a global perspective on ecological and metabolic design.
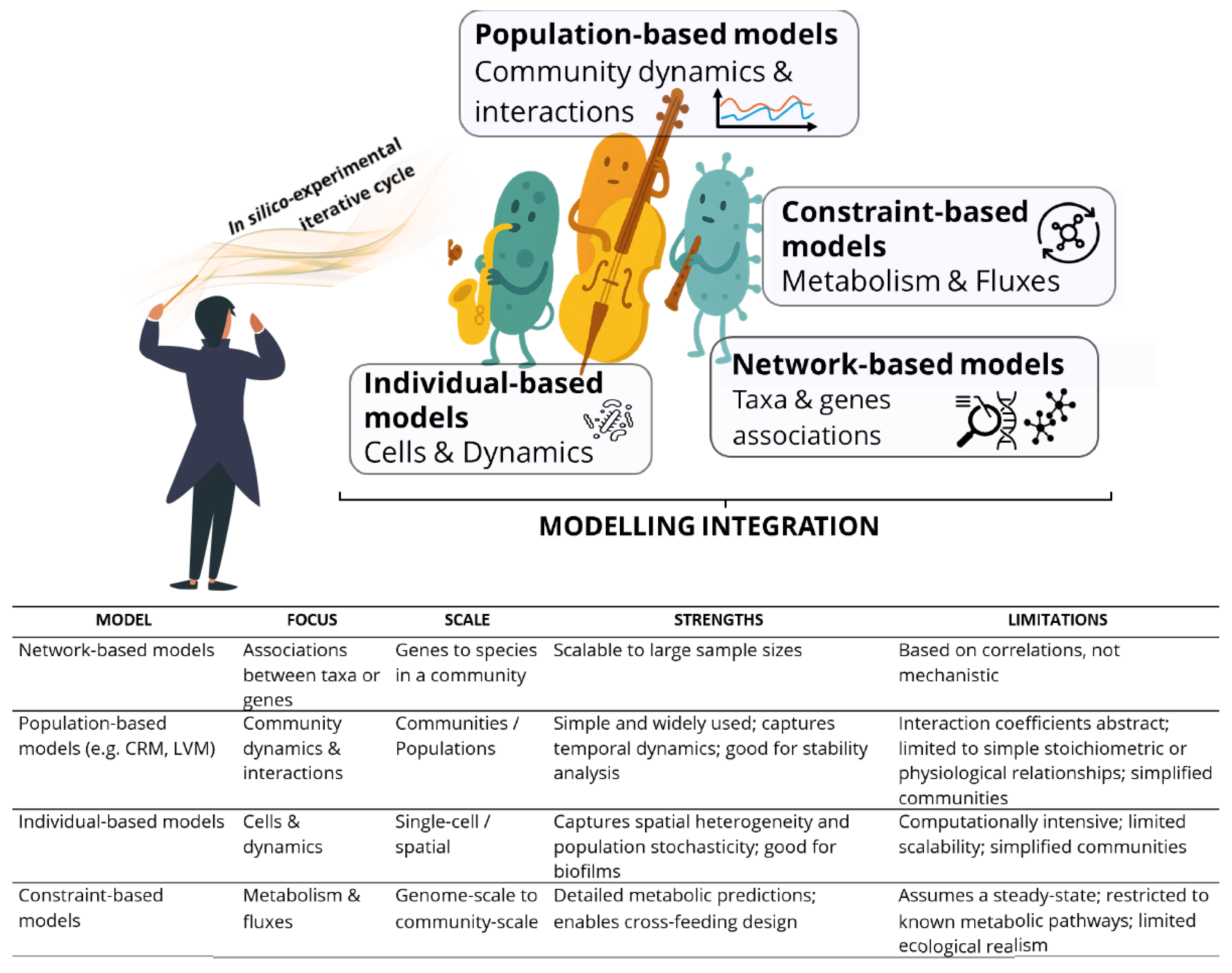

Table 1.
Design principles and challenges for engineering ecological stability. Key strategies for stabilizing synthetic microbial consortia, their main limitations, and representative examples.
Table 1.
Design principles and challenges for engineering ecological stability. Key strategies for stabilizing synthetic microbial consortia, their main limitations, and representative examples.
| Design principle | Challenges | Examples |
|---|---|---|
| Feedback circuits regulate predator–prey oscillations and prevent collapse | Population crashes; partner dominance | Predator–prey E. coli with toxin–antitoxin kill switches [185,186] |
| Mutualism and division of labor enforce metabolic interdependence and reduce competition | Escape mutants; social cheaters | E. coli–S. cerevisiae acetate cross-feeding; amino acid auxotrophies [187,188,189] |
| Competitive balance via negative feedback avoids exclusion and stabilizes coexistence | Ecological drift; unintended dominance | Synthetic bacteriocin modules; programmed toxin–antitoxin systems[190,191] |
| Spatial partitioning and niche specialization buffer direct competition and enhance robustness | Loss of redundancy; stochastic fluctuations | Microfluidic consortia; structured starter cultures in wine/cheese [192,193,194] |
| Higher-order motifs (e.g. rock–paper–scissors) promote coexistence and emergent stability | Context dependence; collapse under fluctuation. | Non-transitive E. coli consortia [195,196] |
| Functional redundancy distributes key tasks across partners, increasing resilience | Keystone loss; cascading failures | Multi-strain cellulose degradation; trait-based lignocellulose consortia [197,198] |
Table 2.
Outstanding challenges and priorities in multimicrobial synthetic biology. Key domains where progress is required to move from conceptual consortia to robust, scalable applications.
Table 2.
Outstanding challenges and priorities in multimicrobial synthetic biology. Key domains where progress is required to move from conceptual consortia to robust, scalable applications.
| Domain | Current challenge | Gap / limitation | Bullet points |
|---|---|---|---|
|
Microbial communication (language) |
Crosstalk Environmental noise Undermine orthogonality |
Signal instability; lack of insulated channels | Expand chemical space (engineered autoinducers, peptide libraries, bioprospecting); standardize orthogonal systems; implement real-time monitoring |
|
Temporal synchronization (rhythm) |
Oscillators drift under fluctuations and scale-up | No robust frameworks for multi-signal coupling | Engineer noise-resistant, feedback-stabilized oscillators; embedded within ecological contexts |
|
Ecological stability (harmony) |
Evolutionary escape, cheaters, shifting keystones | Fragile reliance on keystones; limited redundancy | Design adaptive scaffolds; incorporate redundancy and distributed resilience; monitor higher-order motifs dynamically |
|
Modelling integration (conductor) |
Fragmented tools (GEMsa, CRMsb, IBMsc) rarely connected | No unified platform across genetic–ecological–process scales | Develop hybrid frameworks; create dedicated software; link directly to design–build–test–learn cycles |
|
Process-scale translation (bridge) |
Scale-up introduces heterogeneity, mixing and oxygen limits | Few validated lab-to-industry workflows | Couple bioprocess engineering with synthetic design from the outset; establish microfluidics-to-fermenter pipelines |
aGEMs, genome-scale metabolic models; bCRMs, consumer–resource models; cIBMs, individual-based models.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



