Submitted:
15 September 2025
Posted:
16 September 2025
You are already at the latest version
Abstract
Blueberry mosaic virus (BlMaV) is a persistent pathogen that alters host physiology, however its impact on secondary metabolism in blueberry fruits remains poorly characterized. In this study, the phenolic profile of the cultivar ‘Duke’ was systematically examined in healthy and BlMaV-infected plants over two successive years. Using UHPLC Q-ToF MS, a total of 46 phenolic compounds were detected, spanning flavonols, phenolic acids, and anthocyanins. Comparative analyses revealed consistent shifts in metabolite abundance between healthy and infected samples. Several flavonol aglycones and phenolic acid derivatives accumulated in infected fruits, whereas multiple anthocyanins and glycosides were reduced. To further explore metabolic relationships, color correlation analysis highlighted distinct co-variation patterns among compound classes. Principal component analysis clearly separated infected and healthy fruits, confirming that viral infection was the dominant source of variation, surpassing the influence of harvest year or environmental factors. Nevertheless, the antioxidant capacity remained unchanged, regardless of the presence of the virus or the variation in environmental conditions. These results provide novel biochemical evidence that BlMaV infection reshapes the phenolic composition of blueberries and lays the groundwork for future studies on the metabolic consequences of viral stress in fruit crops.
Keywords:
highbush blueberry (Vaccinium corymbosum L.)
; blueberry mosaic associated ophiovirus
; mosaic disease
; PCR
; UHPLC Q-ToF MS
1. Introduction
Highbush blueberry (Vaccinium corymbosum L.) is an appreciated berry fruit whose cultivation is rapidly growing around the world. The popularity of blueberries among consumers increased with new information on their nutritional benefits [1]. The world production of blueberries is estimated at 1.2 million tons in 2023 [2], with the United States as the largest producer. Over the last two decades blueberry plantations in Serbia expanded rapidly, reaching more than 3,000 hectares today [3]. Plantations are highly intensive, with blueberries grown in the soil or substrate in pots, under anti-hail nets, and managed according to integrated or organic farming principles. Blueberries are becoming an important export commodity to diverse markets. Blueberry ‘Duke’ is one of the most known and economically important cultivars throughout the world. According to estimations, ‘Duke’ occupies about 80% of the orchards in Serbia. It was bred and released in the United States in 1986 [4]. ‘Duke’ is known for early ripening, high yields, and long shelf life. Fruits are medium to large-sized, uniform, firm, light blue, mild and sweet.
Highbush blueberry is a host of more than 15 viruses and virus-like agents [5]. Ophiovirus vaccinii (blueberry mosaic associated ophiovirus, BlMaV) has been recently characterized [6], even though the disease it causes has been known for decades [7]. Typical leaf symptoms include white, yellow, light green and pink mosaic and patterns. The symptoms may be localized on the leaves of one or two stems or cover the entire bush. Latent BlMaV infections are reported, and depending on the year, symptoms may be latent for some period and then reappear the following year. The symptoms of blueberry mosaic disease were described in many countries, but BlMaV was officially confirmed in the USA, Serbia, Slovenia, Turkey, Japan, Poland and Germany [8].
The information on the influence of BlMaV on blueberries is scarce. No quantitative data on its economic importance are available. The influence of BlMaV on pomological properties (fruit length, width and height, shape, fruit weight, and soluble solids content) of blueberry ‘Duke’ fruits was reported by Jevremović et al. [9]. The results confirmed no statistically significant impact of BlMaV of the examined fruit traits. The aim of our study was to evaluate the influence of blueberry mosaic disease caused by blueberry mosaic associated ophiovirus on polyphenolic profile and antioxidant capacity of blueberry ‘Duke’ fruits.
2. Materials and Methods
2.1. Leaf and Fruit Sampling
The material for this study was collected in 2019 and 2020 in the blueberry orchard in locality Budoželja, Ivanjica (43° 32.262'N 20° 16.568'E). The orchard was located at an altitude of 652 m on a parcel with a gentle slope. Blueberries were grown directly in the soil at 2.5 x 1.5 m spacing following the principles of integrated pest and disease management system. For the study, we randomly selected two groups of 20 plants from the orchard. The first group included 10 plants exhibiting clear mosaic symptoms, including light green mosaic patterns, yellowish and dark red mosaic patterns. The other group of 10 plants included plants without symptoms (Table 1). Leaves from each plant were collected in May each year to evaluate the virus presence. Fruits were picked each year at the optimal maturity stage (end June-early July).
During 2019 and 2020, average monthly and annual air temperature (°C) and precipitation (mm) data were obtained from automatic weather sensors located near the orchards (Table 2). These data were sourced from the official Republic Hydrometeorological Service of Serbia.
2.2. RT-PCR Analysis
The presence of BlMaV in selected plants was evaluated using reverse transcription polymerase chain reaction (RT-PCR). Total nucleic acids (TNA) from fresh leaf samples were extracted with a modified CTAB method [10]. Reverse transcription was performed with random hexamer primers and Maxima Reverse Transcriptase (Thermo Scientific, USA). PCR was done with specific BlMaV-specific primers amplifying a 756 bp fragment from RNA3 [6]. Amplified PCR products were analyzed by 1.5% agarose gel electrophoresis, ethidium bromide staining and visualized in Gel Doc EZ System with UV tray (Biorad laboratories, USA). All plants were further tested for the presence of other viruses infecting blueberries. For the presence of blueberry shock virus (Ilarvirus BSV, BlShV), blueberry leaf mottle virus (Nepovirus myrtilli, BLMoV), blueberry scorch virus (Carlavirus vaccinii, BlScV), blueberry shoestring virus (Sobemovirus BSSV, BSSV), tomato ringspot virus (Nepovirus lycopersici, ToRSV), tobacco ringspot virus (Nepovirus nicotianae, TRSV) and peach rosette mosaic virus (Nepovirus persicae, PRMV) plants were tested by ELISA test following manufacturers’ protocols (Agdia Inc., USA and BIOREBA AG, Switzerland). To analyze the plants for the presence of blueberry red ringspot virus (Soymovirus maculavaccinii, BRRV) we performed PCR test as described earlier [11].
2.3. Preparation of Blueberry Extracts
Ground blueberry samples (10 g) were extracted with 25 mL of methanol using ultrasonic treatment. After 30 minutes of extraction, the mixture was centrifuged twice sequentially for 15 minutes at 3500 rpm. The resulting supernatant was filtered through a 0.22 μm Minisart filter prior to the determination of flavonoid, phenolic contents, and antioxidant capacity. The obtained extract was also utilized for UHPLC Q-ToF-MS analysis. The same extraction procedure was conducted using 25 mL of methanol/HCl (85:15, v/v) to obtain an extract for determining the anthocyanin content. All analyses were performed in triplicate, and the results are expressed as the mean ± standard deviation of three measurements.
2.4. Determination of Total Anthocyanins, Flavonoids, Phenolics, and Antioxidant Capacity
Monomeric anthocyanin content in the methanolic extracts was quantified using the pH differential method, as previously described [12,13]. Results are expressed as mg of cyanidin-3-glucoside equivalents per 100 g of fresh weight (C3GE/100 g fw). Total flavonoid content was quantified using a colorimetric method [13,14], with results expressed as mg of catechin equivalents per 100 g fresh weight (mg CE/100 g fw). Total phenolic content was determined by a modified Folin–Ciocalteu colorimetric method, and values were reported as mg of gallic acid equivalents per 100 g of fresh weight (mg GAE/100 g fw) [13,15]. Antioxidant activity was determined using the DPPH method [16] and the results were expressed as μmol trolox equivalents per 100 g fresh weight (μmol TE/100 g fw).
2.5. UHPLC Q-ToF MS Analysis
High-performance liquid chromatography (Agilent 1290 Infinity Liquid Chromatography) coupled with a quadrupole and time-of-flight mass spectrometer (6530C Q-ToF-MS, Agilent Technologies, Inc., CA, USA) was used for the analysis. Chromatographic separation was performed at 40 °C on a Zorbax C18 column (2.1 × 50 mm, 1.8 µm, Agilent Technologies, Inc., CA, USA). The mobile phase consisted of (A) ultrapure water and (B) 98% acetonitrile (MS grade), with both phases containing 0.1% formic acid (MS grade). The flow rate was set to a constant 0.3 mL/min, and the injection volume was 5 µL. The gradient elution program was as follows: 0–2 min (98% A), 2–17 min (linear decrease to 2% A), followed by re-equilibration to the initial conditions (98% A) over the next 5 minutes to prepare the column for the subsequent injection.
The Q-ToF-MS system was equipped with an Agilent Jet Stream electrospray ionization (ESI) source operating in both negative and positive ionization modes. Blueberry extracts were analyzed in both ionization modes. The ion source operating parameters were as follows: nebulizer pressure 45 psi, drying gas temperature 225 °C with a flow rate of 8 L/min, sheath gas temperature 300 °C with a flow rate of 10 L/min, capillary voltage 2500 V, fragmentor voltage 175 V, skimmer voltage 65 V, and RF peak voltage on the octopole 750 V. The Q-ToF-MS system acquired spectra in the m/z range of 100–1700, at a scan rate of 2 Hz. Extracts were analyzed in Auto MS/MS mode to detect phenolic compounds in the blueberry extracts. The parameters for the Auto MS/MS mode were: m/z range = 100–1700, scan rate of 1 spectrum/sec, and a fixed collision energy of 30 eV.
Agilent MassHunter software was used for data acquisition, instrument control, and spectral evaluation. Bioactive compounds were identified based on their accurate m/z (monoisotopic mass), MS fragmentation patterns, and available literature data [17,18,19,20]. Exact masses of the compounds were calculated using ChemDraw software (version 12.0, CambridgeSoft, Cambridge, MA, USA). CAS SciFinder (https://scifinder.cas.org) was used for the structural and formula-based search of bioactive compounds.
2.6. Statistical Analysis
A two-factorial experimental design using ANOVA and Tukey’s multiple comparison tests was used to analyze the data, using Statistica 7 (StatSoft, Inc., Tulsa, OK, USA). The viral status of the plant (BlMaV‒infected, BlMaV‒free), and harvest year (2019, 2020) were taken as the factors of variation. Principal component analysis (PCA), based on the content of 11 individual phenolic compounds and 2 groups of compounds, was performed, and PCA was designed. The correlations between observed compounds were graphically presented using color correlation analysis performed in R software version 4.0.3 (64-bit).
3. Results
3.1. BlMaV Detection
A total of 20 blueberry ‘Duke’ plants were analyzed for BlMaV presence by RT-PCR. The analysis confirmed the presence of BlMaV in all samples with mosaic symptoms (10 samples) (Figure 1) and confirmed BlMaV-free status in asymptomatic samples (10 samples). The ELISA and PCR analysis confirmed the absence of all other tested viruses (BlShV, BLMoV, BlScV, BSSV, ToRSV, TRSV, PRMV, and BRRV), excluding their influence on the investigated properties.
3.2. Quantification of Total Anthocyanins, Flavonoids, and Phenolics and Determination of Antioxidatove Capacity
The total amounts of certain phenolic groups, and antioxidative capacity in healthy and viral blueberry samples over two successive years are presented in Table 3. These results are comparable with previously reported data for ‘Duke’ blueberry cultivar [21,22,23]. Viral infection had no influence on total phenolic content and, consequently on the antioxidative capacity. Nevertheless, BlMaV infection exerted a detectable influence on the flavonoid content, restricted to 2019. The influence of the harvest year was significantly more pronounced, most likely due to the different environmental conditions (temperature and rainfall) during 2019 and 2020. For instance, the total amounts of anthocyanins in B19− and B20− were 112.06 and 129.57, respectively, which is an evident influence of harvest year on healthy blueberry samples.
Taking into account only these results, one can conclude that BlMaV infection had very restricted influence on the phenolic profile of ‘Duke’ blueberry samples, which may even suggest that the ‘Duke’ cultivar is resistant to BlMaV infection. However, in order to validate or refute such a claim, we found it essential to conduct a detailed polyphenolic analysis of the fruit samples.
3.3. Identification of Phenolic Compounds
Using the UHPLC Q-ToF MS technique, 42 phenolic compounds were detected in all blueberry samples, including 12 phenolic acids and their derivatives, 7 flavonol aglycones, 10 flavonol glycosides and acyl derivatives, one flavanone (naringenin), one other phenolic compound (vanilloloside), one proanthocyanidin (procyanidin B2), and 10 anthocyanins (Table 4). All of these compounds were previously identified in blueberry fruits of various cultivars [24,25,26,27,28]. In addition to phenolic compounds, three terpenoids (compounds 43‒45) and one plant hormone (abscisic acid; compound 46) were identified among the non-phenolic constituents (Table 4).
In this study, total of eight caffeic acid derivatives (compounds 2, 4‒7, 10‒12), three derivatives of gallic acid (compounds 1, 3, 8), and diferulic acid (compound 9) were identified in the observed samples. Ancillotti et al. [29] reported a very similar acidic profile across different Vaccinium berry species, highlighting the predominance of caffeic acid derivatives. The same authors detected nearly identical set of flavonol aglycones (compounds 13‒16, 18‒19) and flavonol glycosides and acyl derivatives (compounds 20‒29), with the exception of patuletin (compounds 17). To our knowledge, this is the first report of patuletin detected in blueberry fruits.
Anthocyanins carry a positive charge at pH values below 3 and are therefore typically detected as molecular ions [M]⁺. Consequently, these polyphenols were identified using positive ionization mode. In blueberries, anthocyanins are predominantly present as glycosides of cyanidin (m/z 287), peonidin (m/z 301), delphinidin (m/z 303), petunidin (m/z 317), and malvidin (m/z 331). These molecules typically contain a sugar moiety—galactose and glucose (as hexoses) or arabinose (or pentose) —linked to the 3-hydroxyl position of the aglycone, and are often acylated with acetic acid at the 6′′-hydroxyl position of the sugar residue. Kim et al. [30] identified 22 anthocyanin derivatives, consisting of mono-glycosides and acetyl-glycosides attached to previously mentioned aglycones, in nine cultivars of highbush blueberries. In all blueberry samples, four species of cyanidin derivatives (cyanidin 3-O-galactoside, cyanidin 3-O-glucoside, cyanidin 3-O-arabinoside, cyanidin 3-O-(6′′-O-acetyl)glucoside) were detected. Wang et al. [25] determined the anthocyanin compositions of 62 blueberry cultivars using the UPLC–MS technique, and identified 30 anthocyanins derived from five anthocyanidins, including five xylosides. The most comprehensive analysis of the anthocyanin profile was conducted by Ancillotti et al. [29], who identified as many as 64 derivatives. In our fruit samples, pentosides and hexosides of delphinidin, petunidin and malvidin were identified (compounds 33‒38), along with acetyl-hexosides of delphinidin, petunidin, malvidin, and peonidin (compounds 39‒42). However, no glycosides of cyanidin were traced.
The presence of pentacyclic terpenoid (compounds 43‒45) in berry fruits is well-documented [31,32]. Abscisic acid, a plant growth regulator whose increased content is associated with the onset of ripening, likely plays a key role in the ripening process and may contribute to the regulation of flavonoid biosynthesis in blueberries [33].
3.4. Color Correlation Analysis
Correlation analysis was conducted to explore potential relationships among the 46 detected compounds in the blueberry samples (Table 4). Figure 2 provides a visual representation of these correlations, with circle color indicating the direction (red: negative; blue: positive) and circle size reflecting the magnitude of the correlation (larger circles correspond to stronger correlations) [34]. This visualization facilitates the identification of compound clusters and underlying patterns in the dataset. By highlighting strong positive or negative correlations, this approach can help to reveal which groups of bioactive compounds may be biosynthetically or functionally related, providing insights into the phytochemical composition of the samples.
The correlation analysis (Figure 2) revealed numerous statistically significant associations (p < 0.05) among the 46 detected metabolites, particularly within biosynthetically related groups such as phenolic acids, flavonol aglycones, glycosides, anthocyanins, and terpenoids. A strong negative correlation was observed between methyl gallate and vanilloloside (r = − 0.955, p < 0.01) as well as malvidin 3-O-pentoside (r = − 0.963, p < 0.01). Caffeic acid hexoside showed strong positive correlation with and tetramethyl-digallate (r = 0.977, p < 0.01) and laricitrin (r = 0.959, p < 0.01), while on the other hand showing negative correlation with quercetin (r = − 0.990, p < 0.001), myricetin 3-O-pentoside (r = − 0.988, p < 0.01), quercetin 3-O-(6"-acetyl)hexoside (r = − 0.963, p < 0.01), naringenin (r = − 0.981, p < 0.01), petunidin 3-O-(6"-acetyl)hexoside (r = − 0.983, p < 0.01). Dimethyl-digallate showed a strong positive association with caffeoylquinic acid is. I (r = 0.978, p < 0.01) and patuletin (r = 0.980, p < 0.01), while showing a notable inverse correlation with caffeoylquinic acid is. II (r = − 0.953, p < 0,01), delphinidin 3-O-pentoside (r = − 0.951, p < 0.01), and delhinidin 3-O-hexoside (r = − 0.953, p < 0.01). Caffeoylquinic acid is. III is positively correlated with kaempferol (r = 0.979, p < 0.01) and pentacyclic terpenoid II (r = 0.979, p < 0.01). Caffeoylquinic acid methyl ester strongly negatively correlated with isorhamnetin (r = − 0.970, p < 0.01), while positively correlating with myricetin 3-O-hexoside (r = 0.995, p < 0.01), laricitrin 3-O-hexoside (r = 0.966, p < 0.01), quercetin 3-O-(6"-acetyl)hexoside (r = 0.955, p < 0.01), malvidin 3-O-hexoside (r = 0.956, p < 0.01), and abscisic acid (r = 0.974, p < 0.01). Tetramethyl-digallate is negatively correlated with quercetin (r = − 0.975, p < 0.01) and petunidin 3-O-(6"-acetyl)hexoside (r = − 0.953, p < 0.01), while at the same time positively correlated with laricitrin (r = 0.997, p < 0.01) and vanilloloside (r = 0.989, p < 0.01). Diferulic acid is statistically significant positively corelated with syringetin (r = 0.981, p < 0.01), isorhamnetin 3-O-(6"-O-rhamnosyl)hexoside (r = 0.975, p < 0.01), procyanidin B-type dimer (r = 0.970, p < 0.01). Caffeoyl coumaroylquinic acid is negatively correlated with peonidin 3-O-(6"-acetyl)hexoside (r = − 0.974, p < 0.01), while caffeoylquinic acid dimer is negatively correlated with petunidin 3-O-hexoside (r = − 0.997, p < 0.01). Kaempferol is positively correlated with petunidin 3-O-hexoside (r = 0.951, p < 0.01). Quercetin is negatively correlated with laricitrin (r = − 0.962, p < 0.01), while positively correlating with myricetin 3-O-pentoside (r = 0.959, p < 0.01), naringenin (r = 0.992, p < 0.01), and petunidin 3-O-(6"-acetyl)hexoside (r = 0.996, p < 0.01). Isorhamnetin is in high negative correlation with myricetin 3-O-pentoside (r = − 0.971, p < 0.01), myricetin 3-O-hexoside (r = − 0.988, p < 0.01), laricitrin 3-O-hexoside (r = − 0.998, p < 0.01), quercetin 3-O-(6"-acetyl)hexoside (r = − 0.996, p < 0.01), and abscisic acid (r = − 0.987, p < 0.01). Myricetin showed strong positive correlation with quercetin 3-O-hexoside (r = 0.986, p < 0.01), on the other hand, patuletin exhibited strong negative correlation with delphinidin 3-O-pentoside (r = − 0.979, p < 0.01) and delhinidin 3-O-hexoside (r = − 0.964, p < 0.01). Laricitrin was negatively correlated with quercetin 3-O-pentoside (r = − 0.966, p < 0.01), while at the same time showed positive correlation with vanilloloside (r = 0.997, p < 0.01) and malvidin 3-O-pentoside (r = 0.954, p < 0.01). Syringetin showed statistically significant positive correlation with isorhamnetin 3-O-(6"-O-rhamnosyl)hexoside (r = 0.968, p < 0.01) and pentacyclic terpenoid (r = 0.981, p < 0.01), while the opposite correlation is noticed between quercetin 3-O-pentoside and vanilloloside (r = − 0.977, p < 0.01), and malvidin 3-O-pentoside (r = − 0.994, p<0.01). On the other hand, myricetin 3-O-pentoside showed positive correlation with laricitrin 3-O-hexoside (r = 0.956, p < 0.01), quercetin 3-O-(6"-acetyl)hexoside (r = 0.988, p < 0.01), naringenin (r = 0.954, p < 0.01), petunidin 3-O-(6"-acetyl)hexoside (r = 0.953, p < 0.01), and abscisic acid (r = 0.985, p < 0.01). Furthermore, myricetin 3-O-hexoside showed strong positive correlation with laricitrin 3-O-hexoside (r = 0.987, p < 0.01), quercetin 3-O-(6"-acetyl)hexoside (r = 0.974, p < 0.01), and abscisic acid (r = 0.980, p < 0.01). There is a strong positive correlation between abscisic acid and laricitrin 3-O-hexoside and quercetin 3-O-(6"-acetyl)hexoside (r = 0.975, p < 0.01 and r = 0.991, p < 0.01, respectively). Laricitrin 3-O-hexoside is also positively correlated with quercetin 3-O-(6"-acetyl)hexoside (r = 0.989, p < 0.01). A significant negative correlation can be observed between quercetin 3-O-(6"-O-rhamnosyl)hexoside and naringenin (r = − 0.976, p < 0.01), delphinidin 3-O-(6"-acetyl)hexoside (r = − 0.992, p < 0.01), and petunidin 3-O-(6"-acetyl)hexoside (r = − 0.966, p < 0.01). Isorhamnetin 3-O-(6"-O-rhamnosyl)hexoside showed positive correlation with malvidin 3-O-hexoside (r = 0.969, p < 0.01) and pentacyclic terpenoid (r = 0.975, p < 0.01). There is also a high positive correlation between the following compounds: naringenin and petunidin 3-O-(6"-acetyl)hexoside (r = 0.999, p < 0.01), vanilloloside and delphinidin 3-O-pentoside (r = 0.972, p < 0.01), procyanidin B-type dimer and pentacyclic terpenoid (r = 0.970, p < 0.01), delphinidin 3-O-pentoside and delhinidin 3-O-hexoside (r = 0.993, p < 0.01), malvidin 3-O-pentoside and vanilloloside (r = 0.972, p < 0.01), petunidin 3-O-hexoside and pentacyclic terpenoid II (r = 0.951, p < 0.01).
3.5. Principal Component Analysis
In order to present the results in a more illustrative manner, principal component analysis (PCA) was applied to determine how the infected and healthy blueberry ‘Duke’ samples, over two consecutive harvest years were grouped based on the relative concentrations of 46 detected individual compounds. In this work the actual concentrations of detected compounds were not determined. Therefore, the peak areas were calculated, and for each compound the highest peak among the four samples (B19−, B19+, B20−, B20+) was set as 100%. The peak areas of the remaining three samples were then expressed as percentages relative to the highest peak. The results of these calculations were presented in Table S1. Based on the normalized peak areas (expressed as percentages relative to the highest peak for each compound), a PCA was performed in order to evaluate the similarities and differences among the healthy and infected blueberry samples.
The PCA results are presented in Figure 3. The PCA biplot illustrates the relationships among observed compounds and different blueberry samples. It was found that the first two principal components accounted for 81.49% of the total variance (55.81% and 25.68%, respectively) in the observed parameters.
According to the PCA results, caffeic acid hexoside, dimethyl-digallate, caffeoylquinic acid is. I, tetramethyl-digallate, isorhamnetin, patuletin, laricitrin, quercetin 3-O-(6"-O-rhamnosyl)hexoside, vanilloloside, peonidin 3-O-(6"-acetyl)hexoside, which contributed 3.78%, 2.17%, 1.46%, 3.38%, 3.59%, 2.74%, 3.17%, 3.42%, 2.93% and 1.05% of the total variance respectively, demonstrated a positive impact on PC1. Additionally, caffeoylquinic acid is. III (7.16%), kaempferol (6.60%), myricetin (2.67%), quercetin 3-O-pentoside (2.49%), quercetin 3-O-hexoside (1.97%), isorhamnetin 3-O-(6"-acetyl)hexoside (2.02%), petunidin 3-O-hexoside (4.39%), malvidin 3-O-(6"-acetyl)hexoside (5.67%) and pentacyclic terpenoid II (6.59%) positively influenced the calculation of PC2, as presented in Figure 3.
PCA revealed that the primary factor driving variation in the phenolic profiles was the infection status of the plants. The first principal component (PC1), explaining over 55% of the variance, effectively separated virus-infected samples (B19+, B20+) from healthy controls (B19−, B20−), underscoring the profound metabolic impact of BlMaV infection. All BlMaV-infected blueberry samples (B19+, B20+) were grouped on the positive side of PC1, while healthy samples (B19−, B20−) are positioned on the negative side of the same axis (Figure 2). In contrast, the year of harvest (2019, 2020) contributed to a secondary source of variation, indicating that environmental factors linked to harvest year played a lesser, though still detectable, role. Variations between 2019 and 2020 may reflect environmental factors, such as temperature and rainfall (Table 2). However, these changes did not overshadow the biochemical perturbations induced by BlMaV. These detailed chemical/statistical analyses clearly demonstrate that viral infection is a dominant factor affecting the phenolic composition of blueberry fruits, leading to a general alterations in key phenolic compounds.
Specifically, phenolic acid derivatives such as methyl gallate (1), caffeic acid hexoside (2), tetramethyl digallate (9), and caffeoylquinic acid dimer (12) exhibited significantly altered concentrations in infected samples. For example, the levels of tetramethyl digallate (9), and caffeoylquinic acid dimer (12) were generally increased in virus-infected plants compared to healthy counterparts. Caffeic acid hexoside (2) was not even detected in healthy blueberry samples. On the other hand, level of methyl gallate (1) was reduced in infected samples. Among the other compound groups, increased amounts of isorhamnetin (15), patuletin (17), laricitrin (18), quercetin 3-O-(6"-O-rhamnosyl)hexoside (28), vanilloloside (31), and malvidin 3-O-pentoside (35) were detected in infected blueberry samples, whilst quercetin (14), myricetin (16), myricetin 3-O-pentoside (21), quercetin 3-O-hexoside (22), myricetin 3-O-hexoside (23), laricitrin 3-O-hexoside (24), quercetin 3-O-(6"-acetyl)hexoside (25), isorhamnetin 3-O-(6"-acetyl)hexoside (27), naringenin (30), procyanidin B-type dimer (32), delphinidin 3-O-pentoside (33), delhinidin 3-O-hexoside (36), petunidin 3-O-hexoside (37), malvidin 3-O-hexoside (38), delphinidin 3-O-(6"-acetyl)hexoside (40), petunidin 3-O-(6"-acetyl)hexoside (41), and abscisic acid (46) are dominant in healthy samples. All these alterations could suggest impaired biosynthesis or enhanced degradation under viral stress conditions. It seems that anthocyanin contents, a key determinant of blueberry pigmentation, and flavonol derivatives were the most influenced by the viral infection, leading to diminished levels in infected berries. For the remaining 19 compounds, no clear trend was observed.
The PCA score plots revealed a clear separation between BlMaV-infected and healthy blueberry samples, indicating that the infection status was the dominant factor explaining the observed variance in phenolic composition. In contrast, neither harvest year (i.e. environmental variation) resulted in distinct clustering, suggesting that the virus-induced changes outweighed seasonal effects. Along PC1, infected samples consistently grouped together, reflecting a shared pattern of altered accumulation of certain phenolic acids and flavonol derivatives, while reductions in specific anthocyanins contributed to the observed separation. These findings highlight the significant metabolic shift triggered by BlMaV infection and provide further support for the notion that viral stress strongly impacts the phenolic balance in ‘Duke’ blueberries.
4. Conclusions
In this study, comprehensive profiling of phenolic compounds in ‘Duke’ blueberries revealed distinct metabolic alterations associated with BlMaV infection. A total of 46 compounds were detected, with several flavonols, phenolic acids, and anthocyanins showing virus-related changes. Color correlation analysis highlighted clear patterns of co-variation among metabolites, while PCA demonstrated a consistent separation of healthy and infected samples, confirming that BlMaV infection was the dominant factor shaping phenolic composition, exceeding the influence of harvest year or environmental conditions. The infection induced profound metabolic perturbations, including the accumulation of certain phenolic acid derivatives and flavonol aglycones, along with the depletion of several anthocyanins and related glycosides, thereby disrupting the balance of secondary metabolism. These findings suggest that, despite its widespread cultivation, the cultivar ‘Duke’ displays measurable sensitivity to BlMaV infection at the biochemical level. Consequently, the observed changes provide valuable biochemical markers for monitoring viral stress in blueberries, with broader implications for fruit quality, nutritional value, and plant defense mechanisms. Future studies should further investigate the mechanistic basis of these shifts and their potential impact on consumer-relevant traits.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Peak areas and relative values (%) of detected compounds in BlMaV-infected and healthy blueberry samples, using UHPLC Q-ToF MS.
Author Contributions
Conceptualization, N.M. and D.J.; methodology, N.M. and D.J.; software, M.P. and B.L.; validation, M.P., B.V. and D.J.; formal analysis, N.M.; investigation, B.V. and D.J.; resources, N.M. and D.J.; data curation, B.L.; writing—original draft preparation, N.M. and D.J.; writing—review and editing, M.P., B.L. and B.V.; visualization, M.P. and B.L.; supervision, B.V.; project administration, N.M. and D.J.; funding acquisition, N.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by The Ministry of Education, Science and Technological Development of the Republic of Serbia, grant number 451-03-136/2025-03/200088, 451-03-136/2025-03/200134, and 451-03-136/2025-03/200215.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Dataset available on request from the authors.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| BlMaV | Blueberry mosaic associated ophiovirus |
| RT-PCR | Reverse transcription polymerase chain reaction |
| BSV, BlShV | Blueberry shock virus |
| BLMoV | Blueberry leaf mottle virus |
| BlScV | Blueberry scorch virus |
| BSSV | Blueberry shoestring virus |
| ToRSV | Tomato ringspot virus |
| TRSV | Tobacco ringspot virus |
| PRMV | Peach rosette mosaic virus |
| C3GE | Cyanidin-3-glucoside equivalents |
| CE | Catechin equivalents |
| GAE | Gallic acid equivalents |
| DPPH | 2,2-diphenyl-1-picrylhydrazyl |
| UHPLC Q-ToF MS | Ultra-high-performance liquid chromatography-quadrupole time-of-flight mass spectrometry |
| PCA | Principal component analysis |
References
- Ma, L.; Sun, Z.; Zeng, Y.; Luo, M.; Yang, J. Molecular mechanism and health role of functional ingredients in blueberry for chronic disease in human beings. Int. J. Mol. Sci. 2018, 19, 2785. [Google Scholar] [CrossRef]
- FAOSTAT (2025). Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 7 July 2025).
- Leposavić, A.; Jevremović, D. Borovnica - Tehnologije gajenja, zaštite i prerade. Naučno voćarsko društvo Srbije: Čačak, Serbia, 2020.
- Retamales, J.B.; Hancock, J.F. Blueberries, 2nd ed.; Cabi; Glasgow, UK, 2018.
- Saad, N.; Olmstead, J.W.; Jones, J.B.; Varsani, A.; Harmon, P.F. Known and new emerging viruses infecting blueberry. Plants 2021, 10, 2172. [Google Scholar] [CrossRef]
- Thekke-Veetil, T.; Ho, T.; Keller, K.E.; Martin, R.R.; Tzanetakis, I.E. A new ophiovirus is associated with blueberry mosaic disease. Virus Res. 2014, 189, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Ramsdell, D.C.; Stretch, A.W. Blueberry mosaic. In Virus Diseases of Small Fruits; Converse, R.H., Ed.; US Government Printing Office, US Department of Agriculture, Agricultural Research Service: Washington, D.C., USA, 1987; Agricultural Handbook (USDA), number 631, pp. 119−120.
- Jevremović, D.; Vasilijević, B.; Leposavić, A.; Katanić, V. (2024) Molecular Characterization of Blueberry Mosaic-Associated Virus in Highbush Blueberries in Serbia. In Proceedings of the 6th International Scientific Conference: Modern Trends in Agricultural Production, Rural Development, Agroeconomy, Cooperatives and Environmental Protection, Vrnjačka Banja, Serbia, 27-28 June 2024, pp. 89−94.
- Jevremović, D.; Paunović, A. S.; Leposavić, A. Influence of blueberry mosaic associated virus on some fruit traits of highbush blueberry ‘Duke’. J. Mt. Agric. Balk. 2020, 23, 195–203. [Google Scholar]
- Li, R.; Mock, R.; Huang, Q.; Abad, J.; Hartung, J. , Kinard, G. A reliable and inexpensive method of nucleic acid extraction for the PCR-based detection of diverse plant pathogens. J. Virol. Methods 2008, 154, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Jevremović, D.; Leposavić, A.; Paunović, S. Incidence of viruses in highbush blueberry (Vaccinium corymbosum L.) in Serbia. Pestic. Fitomed. 2016, 31, 45–50. [Google Scholar] [CrossRef]
- Prior, R.L.; Cao, G. , Martin, A., Sofic, E., McEwen, J., O’Brien, C., et al. Antioxidant capacity as influenced by total phenolic and anthocyanin content, maturity, and variety of Vaccinium species. J. Agric. Food Chem. 1998, 46, 2686–2693. [Google Scholar] [CrossRef]
- Liu, M. , Li, X.Q., Weber, C., Lee, C.Y., Brown, J., Liu, R.H. Antioxidant and antiproliferative activities of raspber ries. J. Agric. Food Chem. 2002, 50, 2926–2930. [Google Scholar] [CrossRef]
- Zhishen, J. , Mengcheng, T., Jianming, W. The de termination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Singleton, V.L. , Orthofer, R., Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation sub strates and antioxidants by means of Folin-Ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [CrossRef]
- Vrhovsek, U.; Masuero, D.; Palmieri, L.; Mattivi, F. Identification and quantification of flavonol glycosides in cultivated blueberry cultivars. J. Food Compos. Anal. 2012, 25, 9–16. [Google Scholar] [CrossRef]
- Tu, P.C.; Liang, Y.C.; Huang, G.J.; Lin, M.K.; Kao, M.C.; Lu, T.L.; Sung, P.J.; Kuo, Y.H. Cytotoxic and anti-inflammatory terpenoids from the whole plant of Vaccinium emarginatum. Planta Med. 2020, 86, 1313–1322. [Google Scholar] [CrossRef] [PubMed]
- Baenas, N.; Ruales, J.; Moreno, D.A.; Barrio, D.A.; Stinco, C:M.; Martínez-Cifuentes, G.; Meléndez-Martínez, A.J.; García-Ruiz, A. Characterization of Andean blueberry in bioactive compounds, evaluation of biological properties, and in vitro bioaccessibility. Foods 2020, 9, 1483. [CrossRef]
- Barnes, J.S.; Nguyen, H.P.; Shen, S.; Schug, K.A. General method for extraction of blueberry anthocyanins and identification using high performance liquid chromatography–electrospray ionization-ion trap-time of flight-mass spectrometry. J. Chromatogr. A 2009, 1216, 4728–4735. [Google Scholar] [CrossRef] [PubMed]
- Dragović-Uzelac, V.; Savić, Z.; Brala, A.; Levaj, B.; Bursać Kovačević, D.; Biško, A. Evaluation of phenolic content and antioxidant capacity of blueberry cultivars (Vaccinium corymbosum L.) grown in the Northwest Croatia. Food Technol. Biotechnol. 2010, 48, 214–221.
- Okan, O.T.; Deniz, I.; Yayli, N.; Şat, I.G.; Öz, M.; Hatipoglu Serdar, G. Antioxidant activity, sugar content and phenolic profiling of blueberries cultivars: A comprehensive comparison. Not. Bot. Horti. Agrobo. 2018, 46, 639–652. [Google Scholar] [CrossRef]
- Shibata, Y.; Ohara, K.; Matsumoto, K.; Hasegawa, T.; Akimoto, M. Total anthocyanin content, total phenolic content, and antioxidant activity of various blueberry cultivars grown in Togane, Chiba Prefecture, Japan. J. Nutr. Sci. Vitaminol. 2021, 67, 201–209. [Google Scholar] [CrossRef]
- Subbiah, V.; Zhong, B.; Nawaz, M.A.; Barrow, C.J.; Dunshea, F.R.; Suleria, H.A.R. Screening of phenolic compounds in Australian grown berries by LC-ESI-QTOF-MS/MS and determination of their antioxidant potential. Antioxidants 2021, 10, 26. [CrossRef]
- Wang, S.; Wang, B.; Dong, K.; Li, J.; Li, Y.; Sun, H. Identification and quantification of anthocyanins of 62 blueberry cultivars via UPLC-MS. Biotechnol. Biotec. Eq. 2022, 36, 587–597. [Google Scholar] [CrossRef]
- Bergmann, A.R.; Siebeneichler, T.J.; Oliveira Fischer, L.; Holz, Í.R.; Rombaldi, C.V.; Santos Oliveira, B.A.; Oliveira Fischer, D.L.; Silva, C.S.; Helbig, E. Physicochemical characterization, phenolic composition and antioxidant activity of genotypes and commercial cultivars of blueberry fruits. Cienc. Rural 2023, 53, e20220450. [Google Scholar] [CrossRef]
- Araniti, F.; Baron, G.; Ferrario, G.; Pesenti, M.; Vedova, L.D.; Prinsi, B.; Sacchi, G.A.; Aldini, G.; Espen, L. Chemical profiling and antioxidant potential of berries from six blueberry genotypes harvested in the Italian Alps in 2020: A comparative biochemical pilot study. Agronomy 2025, 15, 262. [Google Scholar] [CrossRef]
- Zhang, Q.; Zang, H.; Guo, X.; Li, S.; Xin, X.; Li, Y. A systematic study on composition and antioxidant of 6 varieties of highbush blueberries by 3 soil matrixes in China. Food Chem. 2025, 472, 142974. [Google Scholar] [CrossRef]
- Ancillotti, C.; Ciofi, L.; Rossini, D.; Chiuminatto, U.; Stahl-Zeng, J.; Orlandini, S.; Furlanetto, S.; Bubba, M.D. Liquid chromatographic/electrospray ionization quadrupole/time of flight tandem mass spectrometric study of polyphenolic composition of different Vaccinium berry species and their comparative evaluation. Anal. Bioanal. Chem. 2017, 409, 1347–1368. [Google Scholar] [CrossRef]
- Kim, J.H.; Kwon, R.H.; Kim, S.A.; Na, H.; Cho, J.Y.; Kim, H.W. Characterization of anthocyanins including acetylated glycosides from highbush blueberry (Vaccinium corymbosum L.) cultivated in Korea based on UPLC-DAD-QToF/MS and UPLC-Qtrap-MS/MS. Foods 2025, 14, 188. [Google Scholar] [CrossRef]
- Neto, C.C. Ursolic acid and other pentacyclic triterpenoids: Anticancer activities and occurrence in berries. In Berries and Cancer Prevention; Stoner, G.D., Seeram, N.P., Eds.; Springer Nature: Heidelberg, Germany, 2011; pp. 41–49. [Google Scholar] [CrossRef]
- Similie, D.; Minda, D.; Bora, L.; Kroškins, V.; Lugiņina, J.; Turks, M.; Dehelean, C.A.; Danciu, C. An update on pentacyclic triterpenoids ursolic and oleanolic acids and related derivatives as anticancer candidates. Antioxidants 2024, 13, 952. [Google Scholar] [CrossRef]
- Zifkin, M.; Jin, A.; Ozga, J.A.; Zaharia, L.I.; Schernthaner, J.P.; Gesell, A.; Abrams, S.R.; Kennedy, J.A.; Constabel, C.P. Gene expression and metabolite profiling of developing highbush blueberry fruit indicates transcriptional regulation of flavonoid metabolism and activation of abscisic acid metabolism. Plant Physiol. 2012, 158, 200–224. [Google Scholar] [CrossRef]
- Petković, M.; Lukyanov, A.; Đurović, I.; Miletić, N. A novel method for analyzing the kinetics of convective/IR bread drying (CIRD) with sensor technology. Applied Sciences 2025, 15, 4964. [Google Scholar] [CrossRef]
Figure 1.
Symptoms of the blueberry mosaic disease on ‘Duke’ leaves.

Figure 2.
Color correlation diagram between observed compounds for blueberry samples.
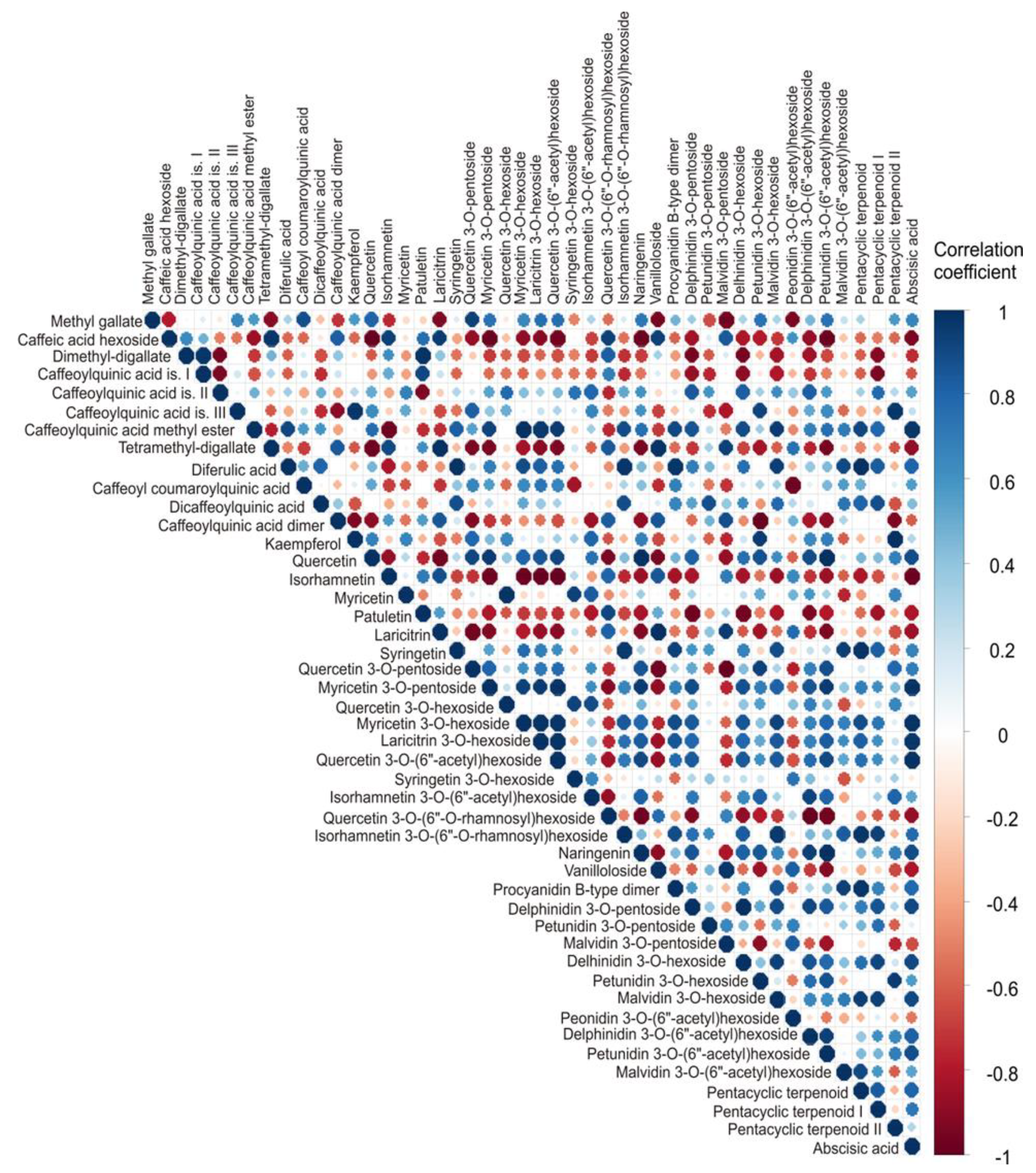
Figure 3.
Segregation of four blueberry samples based on detected compounds (42 phenolic compounds, three terpenoids, and one plant hormone) determined by principal component analysis (PCA). (A) Variable loadings; (B) sample scores. The first letter abbreviates the class of compound (P ‒ phenolic acids and derivatives; F (dark) ‒ flavonol aglycones; F (light) ‒ flavonol glycosides and acyl derivatives: O ‒ other phenolic compounds; A ‒ anthocyanins; T ‒ terpenoids; H ‒ plant hormone), while the number corresponds to the specific compound listed in Table 4.
Figure 3.
Segregation of four blueberry samples based on detected compounds (42 phenolic compounds, three terpenoids, and one plant hormone) determined by principal component analysis (PCA). (A) Variable loadings; (B) sample scores. The first letter abbreviates the class of compound (P ‒ phenolic acids and derivatives; F (dark) ‒ flavonol aglycones; F (light) ‒ flavonol glycosides and acyl derivatives: O ‒ other phenolic compounds; A ‒ anthocyanins; T ‒ terpenoids; H ‒ plant hormone), while the number corresponds to the specific compound listed in Table 4.
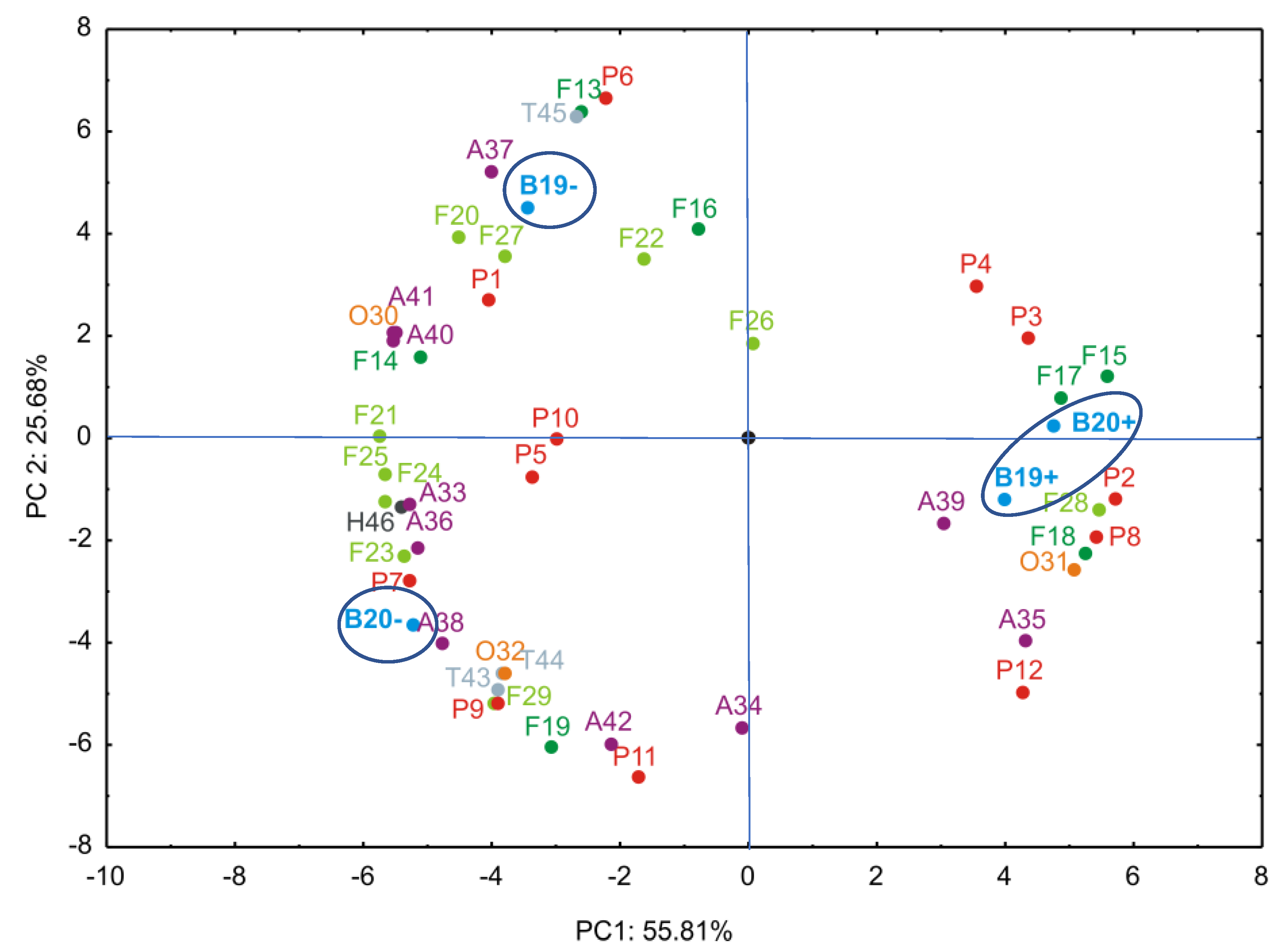

Table 1.
Virus-presence indication for ‘Duke’ blueberry fruits collection over two successive harvest years.
Table 1.
Virus-presence indication for ‘Duke’ blueberry fruits collection over two successive harvest years.
| Viral status | Harvest year | |
| 2019 | 2020 | |
| BlMaV− | B19− | B20− |
| BlMaV+ | B19+ | B20+ |
* BlMaV=B: blueberry mosaic associated ophiovirus. Signs (+) and (−) stand for BlMaV‒infected and BlMaV‒free blueberry samples, respectively.
Table 2.
Mean monthly air temperature and precipitation values recorded during the study period (2019–2020).
Table 2.
Mean monthly air temperature and precipitation values recorded during the study period (2019–2020).
| Month | Average Air Temperature (°C) |
Average Precipitation (mm) |
||
| 2019 | 2020 | 2019 | 2020 | |
| April | 11.7 | 10.5 | 166.8 | 70.0 |
| May | 13.1 | 14.1 | 110.2 | 197.2 |
| June | 20.4 | 17.7 | 328.4 | 231.0 |
| July | 20.1 | 19.4 | 107.2 | 17.4 |
| August | 21.3 | 20.6 | 71.2 | 46.6 |
| September | 16.5 | 17.2 | 2.4 | 49.6 |
| October | 12.4 | 11.8 | 5.4 | 152.6 |
Table 3.
Changes in total anthocyanin, flavonoid, and phenolic contents, and antioxidant capacity (DPPH), as influenced by BlMaV, over two consecutive years.
Table 3.
Changes in total anthocyanin, flavonoid, and phenolic contents, and antioxidant capacity (DPPH), as influenced by BlMaV, over two consecutive years.
| ANOVA | Blueberry samples | ||||
| B19− | B19+ | B20− | B20+ | ||
| Anthocyanins (mg/100 g fw) | ** | 112.06 ± 5.02b | 125.05 ± 7.09ab | 129.57 ± 5.51a | 134.34 ± 2.89a |
| Flavonoids (mg/100 g fw) | *** | 106.73 ± 2.45b | 101.45 ± 1.60c | 112.05 ± 0.52a | 108.68 ± 1.81ab |
| Phenols (mg/100 g fw) | ns | 325.26 ± 4.16 | 322.61 ± 6.49 | 324.64 ± 9.67 | 328.15 ± 4.49 |
| Antioxidativity capacity (μmol TE/100 g fw) |
ns | 77.03 ± 4.23 | 74.14 ± 3.99 | 73.34 ± 2.78 | 70.51 ± 6.28 |
* Values with different letter in the same row denote statistically significant difference (Tukey’s test, p < 0.05) among blueberry fruits of different viral status within both harvest years. ns, *, **, ***: not significant or significant at p < 0.05, 0.01, 0.001, respectively.
Table 4.
Identification of phenolic compounds and its derivatives, and some non-phenolic compounds in BlMaV-infected and virus-free blueberry samples, using UHPLC Q-ToF MS.
Table 4.
Identification of phenolic compounds and its derivatives, and some non-phenolic compounds in BlMaV-infected and virus-free blueberry samples, using UHPLC Q-ToF MS.
| No. | Compounds name |
RT (min) |
Formula |
Calculated mass |
m/z exact mass |
mDa | Fragments (MS2) |
| Phenolic acids and derivatives | |||||||
| 1 | Methyl gallate | 5.64 | C8H7O5¯ | 183.0293 | 183.0295 | 0.15 | 124.0159(100), 125.019, 106.0054 |
| 2 | Caffeic acid hexoside | 5.50 | C15H17O9¯ | 341.0873 | 341.0865 | – 0.76 | 135.0444(100), 179.0338, 161.0228 |
| 3 | Dimethyl-digallate | 7.39 | C16H13O9¯ | 349.056 | 349.056 | 0.04 | 165.0183(100), 137.0236, 123.0081, 151.0028, 183.0297, 197.0445 |
| 4 | Caffeoylquinic acid is. I | 4.81 | C16H17O9¯ | 353.0873 | 353.0866 | – 0.66 | 191.0549(100), 135.0443, 179.0339, 161.0231, 173.0450, 127.0398, 111.0441 |
| 5 | Caffeoylquinic acid is. II | 6.34 | C16H17O9¯ | 353.0873 | 353.0866 | – 0.66 | 191.0550(100), 173.0446, 161.0235, 135.0444, 127.0395 |
| 6 | Caffeoylquinic acid is. III | 7.05 | C16H17O9¯ | 353.0873 | 353.0866 | – 0.66 | 191.0550(100), 173.0435, 161.0235, 127.0395, 111.0442 |
| 7 | Caffeoylquinic acid methyl ester | 7.65 | C17H19O9¯ | 367.1029 | 367.1026 | – 0.31 | 135.0446(100), 179.0342, 161.0234, 191.0551 |
| 8 | Tetramethyl-digallate | 9.07 | C18H17O9¯ | 377.0873 | 377.087 | – 0.26 | 165.0184(100), 137.0231, 121.0186, 151.0034, 190.9980, 315.0131, 330.0345, 166.0219 |
| 9 | Diferulic acid | 11.56 | C20H17O8¯ | 385.0923 | 385.092 | – 0.34 | 193.0491(100), 134.0366, 133.0302, 161.0230, 178.0253, 317.0350 |
| 10 | Caffeoyl coumaroylquinic acid | 8.77 | C25H23O11¯ | 499.124 | 499.1231 | – 0.94 | 163.0392(100), 191.0550, 173.0442, 155.0341, 135.0446, 119.0495, 337.0907, 179.0337 |
| 11 | Dicaffeoylquinic acid | 8.18 | C25H23O12¯ | 515.119 | 515.1183 | – 0.65 | 179.0339(100), 191.0550, 173.0445, 161.0233, 135.0444, 335.0766, 353.0860 |
| 12 | Caffeoylquinic acid dimer | 7.19 | C32H33O18¯ | 705.1667 | 705.1652 | – 1.49 | 513.1014(100), 339.0483, 191.0545, 321.0375, 495.0926 |
| Flavonol aglycones | |||||||
| 13 | Kaempferol | 10.22 | C15H9O6¯ | 285.0399 | 285.0397 | – 0.21 | 285.0390(100), 257.0425, 229.0488, 211.0404, 185.0587, 143.0528, 151.0064 |
| 14 | Quercetin | 9.50 | C15H9O7¯ | 301.0348 | 301.0345 | – 0.33 | 151.0029(100), 121.0290, 107.0135, 164.0109, 178.9975, 229.0487, 245.0438, 271.0234 |
| 15 | Isorhamnetin | 10.42 | C16H11O7¯ | 315.0505 | 315.0497 | – 0.78 | 300.0261(100), 151.0035, 107.0141, 137.0233, 164.0108, 178.9993, 203.0324, 227.0339, 259.0225 |
| 16 | Myricetin | 8.69 | C15H9O8¯ | 317.0297 | 317.0296 | – 0.14 | 151.0029(100), 137.0237, 107.0135, 125.0239, 165.0182, 178.9977, 227.0338, 243.0280, 271.0233 |
| 17 | Patuletin | 9.06 | C16H11O8¯ | 331.0454 | 331.0453 | – 0.09 | 243.0285(100), 299.0176, 271.0230, 255.0273, 227.0341, 215.0335, 199.0389, 183.0447, 171.0443, 143.0498 |
| 18 | Laricitrin | 9.58 | C16H11O8¯ | 331.0454 | 331.0453 | – 0.09 | 151.0044(100), 316.0206, 299.0171, 271.0230, 259.0236, 178.9978, 164.0104, 287.0184, 136.0160, 107.0132 |
| 19 | Syringetin | 10.38 | C17H13O8¯ | 345.061 | 345.0602 | – 0.84 | 315.0134(100), 287.0184, 330.0364, 345.0603, 301.0340, 259.0235, 271.0237, 203.0336, 151.0029 |
|
Flavonol glycosides and acyl derivatives |
|||||||
| 20 | Quercetin 3-O-pentoside | 8.16 | C20H17O11¯ | 433.0771 | 433.0767 | – 0.39 | 300.0259(100), 271.0234, 255.0289, 178.9989, 151.0032 |
| 21 | Myricetin 3-O-pentoside | 5.97 | C20H17O12¯ | 449.072 | 449.0714 | – 0.60 | 449.0702(100), 299.0172, 317.0280, 271.0215, 190.9972 |
| 22 | Quercetin 3-O-hexoside | 7.89 | C21H19O12¯ | 463.0877 | 463.0871 | – 0.55 | 300.0260(100), 301.0323, 463.0864, 271.0234, 255.0284, 151.0030, 178.9987 |
| 23 | Myricetin 3-O-hexoside | 7.44 | C21H19O13¯ | 479.0826 | 479.084 | 1.43 | 316.0207(100), 317.0261, 271.0234, 187.0184, 479.0810, 178.9980 |
| 24 | Laricitrin 3-O-hexoside | 8.06 | C22H21O13¯ | 493.0982 | 493.0977 | – 0.52 | 330.0365(100), 331.0419, 315.0133, 300.0260, 287.0514, 178.9973, 151.0039, 433.0758 |
| 25 | Quercetin 3-O-(6"-acetyl)hexoside | 8.49 | C23H21O13¯ | 505.0982 | 505.0978 | – 0.42 | 300.0262(100), 344.0518, 271.0234, 178.9974, 151.0025, 463.0861 |
| 26 | Syringetin 3-O-hexoside | 8.39 | C23H23O13¯ | 507.1139 | 507.1125 | – 1.37 | 344.0521(100), 507.1112, 345.0574, 387.0699, 329.0300, 316.0569, 301.0403, 273.0381, 151.0031 |
| 27 | Isorhamnetin 3-O-(6"-acetyl)hexoside | 8.99 | C24H23O13¯ | 519.1139 | 519.113 | – 0.87 | 314.0418(100), 519.1125, 315.0462, 299.0203, 285.0393, 271.0241, 257.0443, 243.0289, 151.0025, 357.0595 |
| 28 | Quercetin 3-O-(6"-O-rhamnosyl)hexoside (like Rutin) | 7.74 | C27H29O16¯ | 609.1456 | 609.145 | – 0.56 | 300.0261(100), 609.1435, 301.0329, 271.0235, 151.003, 178.9975, 343.0431 |
| 29 | Isorhamnetin 3-O-(6"-O-rhamnosyl)hexoside (like Narcissin) | 8.22 | C28H31O16¯ | 623.1612 | 623.1608 | – 0.41 | 315.049(100), 623.1592, 314.0416, 300.0249, 271.0241, 151.0022, 357.0595 |
| Other phenolic compounds | |||||||
| 30 | Naringenin | 10.04 | C15H11O5¯ | 271.0606 | 271.0603 | – 0.35 | 119.0499(100), 107.0132, 151.0024, 161.0590, 187.0388, 229.0458, 245.0477 |
| 31 | Vanilloloside | 3.95 | C14H19O8¯ | 315.108 | 315.108 | 0.01 | 123.0445(100), 153.0547, 124.0478 |
| 32 | Procyanidin B-type dimer (like Procyanidin B2) | 6.04 | C30H25O12¯ | 577.1346 | 577.1335 | – 1.10 | 289.0700(100), 407.0752, 125.0237, 137.0243, 161.0241, 245.0796, 273.0388, 339.0842, 381.0951, 425.0876, 451.1001 |
| Anthocyanins | |||||||
| 33 | Delphinidin 3-O-pentoside | 6.47 | C20H19O11⁺ | 435.0927 | 435.0919 | – 0.84 | 303.0488(100), 304.0523, 305.0543 |
| 34 | Petunidin 3-O-pentoside | 6.74 | C21H21O11⁺ | 449.1084 | 449.1075 | – 0.89 | 317.0644(100), 318.068, 287.0535, 302.0409 |
| 35 | Malvidin 3-O-pentoside | 7.07 | C22H23O11⁺ | 463.124 | 463.1232 | – 0.84 | 331.0802(100), 332.0835, 301.0695, 315.0488, 287.0534 |
| 36 | Delhinidin 3-O-hexoside | 6.19 | C21H21O12⁺ | 465.1033 | 465.1025 | – 0.80 | 303.0488(100), 304.0522, 305.0543 |
| 37 | Petunidin 3-O-hexoside | 6.6 | C22H23O12⁺ | 479.119 | 479.1182 | – 0.75 | 317.0645(100), 318.0677, 302.0409 |
| 38 | Malvidin 3-O-hexoside | 6.93 | C23H25O12⁺ | 493.1346 | 493.1338 | – 0.8 | 331.0802(100), 332.0835, 315.0486, 287.0536 |
| 39 | Peonidin 3-O-(6"-acetyl)hexoside | 7.8 | C24H25O12⁺ | 505.1346 | 505.1343 | – 0.30 | 301.0692(100), 302.0731, 303.0702, 286.0457 |
| 40 | Delphinidin 3-O-(6"-acetyl)hexoside | 7.32 | C23H23O13⁺ | 507.1139 | 507.1131 | – 0.77 | 303.0487(100), 304.0521, 305.0547 |
| 41 | Petunidin 3-O-(6"-acetyl)hexoside | 7.48 | C24H25O13⁺ | 521.1295 | 521.1286 | – 0.92 | 317.0643(100), 318.068, 302.0406 |
| 42 | Malvidin 3-O-(6"-acetyl)hexoside | 7.68 | C25H27O13⁺ | 535.1452 | 535.1444 | – 0.77 | 331.0801(100), 332.0835, 315.0486 |
| Other compounds (Terpenoids) | |||||||
| 43 | Pentacyclic terpenoid (like Maslinic or Pomolic acid) | 14.79 | C30H47O4¯ | 471.3474 | 471.3472 | -0.23 | 471.3460(100), 453.3342, 427.3566, 409.3457 |
| 44 | Pentacyclic terpenoid I (like Arjunolic, Euscaphic or Rotundic acid) | 12.58 | C30H47O5¯ | 487.3423 | 487.3415 | -0.85 | 487.3396(100), 469.3305, 437.3053, 425.3392, 405.3107, 393.3127, 443.3483 |
| 45 | Pentacyclic terpenoid II (like Arjunolic, Euscaphic or Rotundic acid) | 12.92 | C30H47O5¯ | 487.3423 | 487.3415 | -0.85 | 487.3405(100), 469.3288, 425.3394, 407.3258, 443.3536, 393.3111 |
| Other compounds (Plant hormone) | |||||||
| 46 | Abscisic acid | 9.31 | C15H19O4¯ | 263.1283 | 263.1283 | -0.03 | 203.1068(100), 219.1372, 289.0910, 153.0911, 136.0521, 122.0367, 125.0605, 148.0525 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



