Submitted:
13 September 2025
Posted:
16 September 2025
You are already at the latest version
Abstract
Species are disappearing worldwide and the expectation is this will get worse in the fu-ture. The most important factors responsible for this are changes in climate and land use, with land use ranked the most important. Changes in the latter resulting in the destruc-tion of natural habitats are thought to be the primer driver. Climate change is, however, also often cited as one of the major driving forces. To reduce the effect of climate change on the decline in biodiversity, it should be moderated by adopting the so called “climate smart conservation”. For this, it is important to know, how climate change will affect the abundance and distribution of species in the future. While it is thought that changes in land use are now more important than climate change in the reduction of biodiversity, it is expected that the latter will play an increasingly important role in the future. On top of that, there exist many other contributing factors: e.g., overexploitation, eutrophication and the introduction of pathogens and herbivorous mammals that also play an important role.
Keywords:
climate change
; decline
; extinction
; habitat suitability
; land use change
; species
1. Introduction
Species are disappearing worldwide and the expectation is this will get worse in the future [1]. The most important factors responsible for this are assumed to be changes in climate and land use [2,3], with land use often ranked the most important [4]. Changes in land use resulting in the destruction of undisturbed or slightly disturbed habitats are thought to be driving the decline in biodiversity [5]. It is assumed that the global transformation of land cover will shortly increase the rate at which species go extinct [6,7,8,9]. Climate change is, however, also often cited as one of the major driving forces [1,10,11,12,13,14,15,16]. To reduce the effect of climate change on the decline in biodiversity, its effects should be moderated by adopting proactive actions, so called “climate smart conservation” [17]. For this, it is important to know how climate change will affect the abundance and distribution of species in the future [18,19].
The negative interactions between the effects of changes in climate and land use are often addressed [20,21,22,23], but attempts to include changes in climate and land use in predictions of future trends in biodiversity are still rudimentary [24,25]. Nevertheless, it is highly likely that the interaction between changes in climate and habitat loss will be disastrous [26]. While it is thought that changes in land use are more important than climate change in the reduction of biodiversity [8,27,28], it is expected that climate change will play an increasingly important role in the future [14,29,30,31,32]. In addition, there are many other contributing factors: e.g., overexploitation, eutrophication and the introduction of pathogens and herbivorous mammals [33,34,35,36].
Potential future distributions of species are usually presented on habitat suitability maps. These maps are generated by species distribution models (SDMs), whose inputs are locations of species in nature and climatic variables generated by climate models [37,38,39], abiotic factors such as geological substrate and soil [40,41], and in a few cases also biotic factors such as magnet species or potential distributions of pollinators [42,43,44]. The use of SDMs has recently become very popular. According to the Web of Science, more than 6,000 papers using SDMs were published over the last 25 years and this number is rapidly increasing [39]. Many of these studies predict how the distribution of species will be affected by climate change [43,45,46,47]. However, it is impossible to verify the accuracy of such predictions, because they refer to scenarios 30, 50, or more years from now and include lots of uncertainties, such as Tsiftsis et al. [48]: (i) future climatic scenarios, (ii) adaptation capacity, (iii) speed of n organisms’ spread to more favourable areas [49,50,51,52], (iv) effect of the spatial scale of the analyses [27], (v) extinction rates associated with increased temperature [49], (vi) life history and (meta)population dynamics of the species studied [53], etc.
Thus, it is worth questioning the reliability of these predictions, as according to Araújo et al. [54] it is estimated that the habitats of 63% of the species of European concern will become climatically unsuitable in the Natura 2000 network by 2080. But are the predicted shifts and losses of species realistic? Santini et al. [55] assessment of the reliability of species distribution projections associated with climate change concludes that the ability to predict future species distributions is on average low and the application and interpretation of such SDM projections should be treated with caution. Thus, there is a need to determine the reliability of SDM predictions.
A step forward was made by Valentin et al. [56] who used species distribution models (SDMs) and projected range changes for 31 orchid species in Estonia under moderate (SSP245) and high-emission (SSP585) scenarios, integrating land use change projections through the 21st century. They found that under both scenarios, many calcicole tuberous species are expected to shift eastward. However, significant losses in orchid richness are predicted as early as 2040, severely impacting western orchid hotspots (western islands and coastal areas) and large parts of central and eastern Estonia, with non-calcicole rhizomatous species inhabiting forest and wet forest habitats emerging as the most vulnerable taxa [56]. According to their analyses, the period up to 2040 emerges as a critical bottleneck, especially for rare and threatened species. While some recovery is projected after 2060 under high-emission scenarios, earlier population declines may lead to irreversible losses [56].
2. Climate Change (CC)
Climate change is an important threat to the world’s plant and animal species, including those on which humans depend. However, predicting how species will respond to future climate change is very difficult. Climate change has strong direct global effects and affects other drivers. Rises in atmospheric concentrations of GHGs (namely carbon dioxide, methane and nitrous oxide) over at least the last 800,000 years have probably been the dominant cause of the present trends in warming worldwide. Natural variations in global temperatures are low compared to human-induced warming [57].
Human-induced warming reached ~1°C (±0.2°C) above pre-industrial levels in 2017, with rises of 0.2°C (±0.1°C) per decade. Effects include thermal stress, coral bleaching, and melting of sea and land ice [57]. The highest changes in temperature are reported in flat landscapes and at high latitudes [57,58]. Most land is warming faster and oceans slower than the average [57]. Evidence of long-term geophysical and biological changes due to warming is now clearer in many parts of the world, such as, in the retreat of mountain glaciers, the earlier arrival of spring [57,59], changes in the phenological responses of vegetation [57,60] and in primary productivity [57,61]. Changes in precipitation have also occurred. Areas in tropical regions have experienced increased precipitation while areas in subtropical regions have experienced decreased precipitation [57,62]. Precipitation has decreased most in Northern and Central African Countries and Western Asia [57,63].
Climate models predict the effects of anthropogenic forcing described above on increases in the frequencies and intensities of extreme events [57,64], e.g., heat waves, droughts, heavy rainfall, storms and coastal flooding [57,65]. These events result from sporadic weather patterns [57,66] and can be intensified by climate variability (e.g., El Niño/Southern oscillation) [57,67,68,69,70]. The increase in the frequency and intensity of such extreme events has considerable effects on well-being, with losses of life, injuries, and other negative health effects, together with damage to property, infrastructure, livelihoods, service provision and environmental resources. In particular, increases in the frequency and intensity of hurricanes are predicted [57]. The cumulative effects of all these changes in temperature, precipitation, and frequency and intensity of extreme weather events are likely to interact and result in unexpected nonlinear changes, with irreversible effects on nature and nature’s contributions to people and society, including economic growth and food and water security [57,71]. Climate driven changes can interact with other direct drivers, at times exacerbating effects on nature and society. Interactions of climate with other factors could also initiate nonlinear climate responses, yielding more extreme and/or rapid effects of climate change [57].
2.1. CC as a Cause of Extinctions
Climate change may be an important threat to global biodiversity, potentially leading to the extinction of numerous species. Wiens and Zelinka´s [16] review of recent SDM studies, indicates ~17% loss of species due to climate change in worst case scenarios. However, this review shows that many SDM studies are biased in excluding the most vulnerable species (those recorded at a few localities), which may lead to underestimating global species loss. Their preliminary estimates tentatively forecast climate-related extinction of 14%–32% of macroscopic species in the next ~50 years, which for intermediate climate changes is potentially 3–6 million (or more) animal and plant species [16].
Conservation of biodiversity in a changing climate requires both limits on change and concentration on strategies that respond to the inevitable changes. Conservation at a scale and with objectives that explicitly address the potential effects of climate change are required. Here these are referred to as “climate change integrated conservation strategies” (CCS) although they must be tailored to individual regions; to be successful, each CCS needs to include the following [72]:
- regional modelling of the response of biodiversity to climate change;
- systematic selection of protected areas with climate change as an integral factor;
- management of diversity across regional landscapes, including protected areas and their surroundings, with climate change as an explicit parameter;
- mechanisms to support regional coordination of management, both across international borders and across the interface between park and non-park Conservation areas; and
- provision of resources from wealthy countries that have had a major role in generating climate change in countries where its effects on biodiversity are highest. To adequately respond to the uncertainties posed by climate change, the provision of resources will be required on a much larger scale than at present.
Hannah et al. [72] claim that no conservation system can succeed in the face of unlimited change. Temperatures predicted for the end of the century may already represent the warmest global climate in over 2 million years. Landmark events, such as the melting of all glaciers, will result in an environment different from that experienced in the evolutionary history of most modern species. Further changes may exceed the capacity of many species to survive.
Hannah et al. [72] also state that reductions in emissions is essential if loss of biodiversity is to be kept at a reasonable level for the survival of natural systems, which as a benchmark is already recognized in international agreements and the UN Framework Convention on Climate Change specifically recognizes the critical link between climate change and the capacity of ecosystems to continue functioning. Thus, it falls to biologists to advocate that the benchmark of functioning is the full complement of the world's species and not just a few, fast-dispersing subset. Present international greenhouse gas emissions will result in increases in temperature that will result in large scale shifts in vegetation and increase in the risk of the widespread extinction of species unable to spread elsewhere due to poor dispersal or disappearance of suitable habitats. It is also likely to increase the risk of a breakdown of human food production systems in ways that will encourage increased pressure on natural areas. Effective lobbying for more rapid reductions in emissions and stabilization of greenhouse gas concentrations closer to present levels could help avoid these changes. Essentially, conservation must extend beyond terrestrial and marine realms and include the atmosphere.
Climate change is expected to result in changes of the distribution and abundance of many species. Predictions of climate-induced population extinctions are supported by changes in geographic range that correspond to climatic warming, but few extinctions have been directly linked to climate change. Shifts in the abiotic environment are also likely to increase the risk of species becoming extinct [73], and references therein. These authors modelled checkerspot (Figure 1) populations and show that changes in precipitation amplified population fluctuations, leading to rapid extinctions. As populations of checkerspots and other species become further isolated by habitat loss, climate change is likely to cause more extinctions, threatening both species diversity and critical ecosystem services. They also report that checkerspot extinctions were caused by interactions among these factors. When distributions become insular because of habitat loss, populations become more vulnerable to climate change and other threats. Extinctions caused by the interaction between habitat loss and climate change are particularly likely in species like checkerspot, with limited dispersal ability [74]. As the interacting factors increasingly overlap, they are likely to cause extinctions in many other taxa.
Climate change may reduce and/or result in changes in the ranges of plants thereby increasing their vulnerability and need for targeted conservation to facilitate their survival. Summers et al. [75] quantified the vulnerability to climate change of plants based on exposure, sensitivity, and adaptive capacity and assessed the effects of including these components in complementarity-based spatial conservation prioritization. They conclude that whilst taking an integrated approach to mitigating vulnerability to climate change can ensure rare species are well-represented in a conservation network, this can come at the cost of a reduced representation of other species. They also stress that conservation decisions aimed at reducing species' vulnerability to climate change need also to consider the sensitivity of spatial conservation priorities to individual components of vulnerability and the trade-offs associated with focusing on rare species.
Climate change has already resulted in the local extinction of many plants and animals, based on surveys spanning many decades. As climate change accelerates, the pace of these extinctions may also accelerate, potentially leading to large-scale, species-level extinctions. Holzman et al. [76] tested this hypothesis for a montane lizard. Contrary to their expectations, populations at two of the hottest sites survived. They also report that genomic data helped predict which populations survived and which went extinct and that overall, the increasing risk to biodiversity posed by accelerating climate change is constrained by only having the opportunity to study its effects over very short timescales.
2.2. Species Richness Affected by CC
Global climate change is increasingly influencing ecosystems [77]. In the past few decades, there has been an increasing number of studies dealing with the responses of biological assemblages to climate change. They have concentrated on more integrated measures of change, such as species richness, community composition and ecosystem properties [78,79,80]. Although it is species that respond to climate change, the consequences of these changes are changes in communities and ecosystems [80]. Moreover, it is commonly believed that climate plays an important role in shaping the patterns of plant and animal species richness [81]. It is also expected that plants and animals with different lifeforms will respond differently to climate change [81].
Species-energy theory indicates that recent climate warming may cause an increase in species richness in temperate, cool and species-poor parts of the Northern Hemisphere [80,82] and a decrease in dry or tropical regions [81,83], which is confirmed by Menéndez et al. [80]. They provided the first assessment, at a geographical scale, of how species richness has changed in response to climate change and investigated changes in species richness using the well-monitored British butterfly fauna. They found that the average species richness of British butterflies has increased, but much more slowly than predicted, with only one-third of the predicted increase. The resultant species assemblages are increasingly dominated by generalist species that can respond quickly. The time lag is confirmed by the successful introduction of many species to climatically suitable areas beyond their ranges. These results imply that it may be decades or centuries before the species richness and composition of biological communities adjusts to the current climate. Another interesting study is that of Lemoine et al. [77], who tested whether changes in the composition of bird communities are associated with recent climate change. They studied migratory and resident bird species because they are likely to respond differently to climatic change and report that they do differ significantly, and that climate change has already influenced the species richness and composition of European bird communities.
On the other hand, plants may respond differently than animals to increasing temperatures. In drylands in China, plants in the mountainous regions and deserts respond differently to climate change. Plant species richness in mountain-basin systems is more associated with climate change than in other regions (meadows and steppes). Moreover, there are slight increases in the mean species richness of herbaceous, woody and all species [81]. They propose that both the mountain-basin regions and desert diversity hotspots should be given high priority for conservation, as many species may be at high risk of extinction as the climate becomes warmer and drier.
Forests are another environment very prone to changes. It is predicted that the composition of one-third of the planet’s forests could be altered markedly due to climate change [84,85]. A study in the eastern United States, by Iverson and Prasad [85] reports a potential for significant changes in forest communities associated with different climate change scenarios (Figure 2). The effect may be greater regionally and could strongly affect regional biodiversity. Iverson and Prasad [85] report that the distributions of the two types of forests most likely to increase are oak-hickory and oak-pine, whereas that of other types of forest will decline. Spruce-fir is likely to spread beyond the US border, aspen-birch is predicted to decline in the East and the loblolly-shortleaf and longleaf-slash pine forests are predicted to decrease in most scenarios [85]. Moreover, these increases and decreases in distribution will be associated with secondary effects on the flora and fauna depending on the type of forest. One example of the potential increase in oak is the conversion of some pine type to oak–pine type forests. Oak-hickory supports a variety of species that rely on heavy mast production, whereas pinelands provide valuable habitat for a different variety of mammals and birds [85], and this balance might be disrupted. This study indicates that communities are ad hoc mixtures of species that are unlikely to move together as intact communities if conditions change [85].
Modelling the effects of climate change on species and communities on isolated islands is also very important [86], as the species there have limited space in which to move or escape. Results of Khalyani et al. [86] for the tropical island of Puerto Rico indicate that it is likely that the environmental suitability for wet climate species will decrease and for dry and warm climate species will increase in the future. This implies species will spread to high altitudes if unable to adapt to the new conditions, which will increase species richness in uplands and an increase in the competition with wet and cold climate species. Hence, the need for conservation of island biological diversity by protecting the wet climate species in uplands [86].
Associated with climate change, there are three potential consequences for species: adaptation in situ, tracking climate change spatially and/or temporally, and local extinction [87,88]. Short-term experiments in which the temperature of tundra was increased resulted in a decrease in species richness [89,90]. However, species that remain in situ and experience the consequences of climate change will meet in the future and interact with species tracking climate change [91]. One of the predicted consequences of climate warming is the spread of plant species to higher altitudes and latitudes [82 and citations therein], which could eventually result in many new communities [82,91]. The results of Wang et al. [82] in which plant communities are transplanted indicate that transferring to lower altitudes generally result in a net increase in species richness, whereas transfers to higher altitudes result in either net increases or decreases in species richness. Moreover, Wang et al. [82] show that ignoring gains in species in climate change experiments may result in inaccurate conclusions about the responses of plant diversity to climate change. Wang et al. [82] conclude that species that can track warming at low latitudes and altitudes are more likely to spread to and alter plant diversity at high latitudes and altitudes in the region. Moreover, their study highlights the importance of biotic factors, in addition, to the abiotic environment, when considering how climate change will affect plant diversity.
2.3. Species Richness Affected by CC
O’Neill et al. [92] in their model point to the need to consider population differences when modelling biotic responses to climate change and suggest that climate change will result in populations being maladapted in many areas. Their model also indicates a way to mitigate the effects of climate change by identifying populations expected to be best adapted in the next generation. Finally, their study highlights the value of wide-ranging long-term tests in addressing key issues in ecology and climate change.
Ahola et al. [93] concentrate on how climate change can alter competitive relationships between resident and migratory birds (Figure 3). They claim that climate change could affect competition for resources between resident and migratory species of birds by changing the time of onset of breeding or by altering their population densities. On this basis they conclude that climate change has a great potential to alter the competitive balance between the two species they studied [93].
Harrington et al. [94] mainly predict the effects of climate change on insect pests and beneficial insects of agriculture and forestry in the northern hemisphere's temperature zones, although the general principles may be applied more widely. They claim that both mean temperature and the extent and frequency of extreme events can have major effects on insect populations. They outline what one intuitively expects to happen to insects because of the general trend in temperature, and then they consider various complicating factors that have the potential to confound these expectations. They then give examples of changes that are already occurring and the consistent expectations. Finally, they consider some of the possible implications of these changes for insect management and insect conservation.
Many factors, including climate, resource availability and habitat diversity are proposed as determinants of global diversity, but the links between them are rarely studied. Menéndez et al. [95] studied the direct and indirect effects of climate and habitat factors on butterfly diversity and reported that climate variables have strong effects on habitat generalists, whereas host-plant richness and habitat diversity are more important for habitat specialists. Considering all these effects (direct and indirect together), climate has the strongest association with butterfly species richness for all groups of species. Their results indicate that the different mechanistic hypotheses concerning species richness may be appropriate for habitat generalists and specialists, with generalists hypothesized to show direct physiological limitations and that specialists are in addition constrained by trophic interactions (climate affecting host-plant richness).
Van der Putten [96] review a number of studies on trophic interactions in a changing world in relation to (i) the scale of ecosystem response to environmental change with emphasis on the soil subsystem, (ii) the linkage of above- and belowground subsystems and (iii) natural selection and the stability of community structure and ecosystem functioning. They discuss the need to bring together isolated sub-disciplines of ecology to better understand the implications of global changes for ecosystem processes.
2.4. Potential Effects of CC on Different Communities
Human-induced changes in the global environment have long been known to affect organisms by altering their physiology, distribution, or longevity [79,97]. However, the responses of species differ widely, making it difficult to predict how entire ecosystems will respond in the future [98]. This unpredictability is particularly evident when considering the complex interactions within food webs.
Tylianakis [99] emphasizes the importance of considering multitrophic interactions when predicting ecological responses to climate change, as they can have cascading effects on biodiversity and ecosystem services. Climate warming can modify interactions between predators and their prey, leading to changes in predation rates and subsequent shifts in species populations within ecological communities. In addition, increasing temperatures may disrupt mutualistic interactions, such as those between plants and their pollinators, potentially leading to declines in plant reproduction and alterations in community composition. Increase in temperature can also boost herbivore metabolism and consumption rates and increasing herbivore pressure on plant communities, which may affect plant diversity and ecosystem functioning.
In addition to changes in food web dynamics, climate change is now thought by some biologists to be the main threat to biodiversity, though uncertainties remain about which species are most at risk. Wilson and Maclean [100] report the threats posed by climate change to Lepidoptera (butterflies and moths) and other insects and conclude that their ecological responses are likely to increase risk of extinctions. Insects, particularly those with short life cycles, respond quickly to climate change, making them valuable indicators for monitoring the effects of climate change on biodiversity. However, extinction risks are not uniform globally. Certain regions, such as South America, Australia and New Zealand, face particularly high risks, thus there is a need for region-specific conservation strategies. Their research underscores the pressing threat that climate change poses for insect populations and highlights the importance of integrating climate considerations into conservation planning.
To predict changes in species distributions due to climate change, it is crucial to understand populations at the poleward edges of their ranges. Ecologists generally expect range shifts as edge populations spread into areas where conditions increasingly resemble their historical range. Hellmann et al. [101] report the effects of variations in climate on two butterfly species at the northern edge of their ranges to assess their potential for a poleward shift in range. Their findings suggest that individual species respond uniquely to climate change, complicating predictions of range shifts. This highlights the need for species-specific analyses to be included in conservation strategies, as general assumptions about range extension may not hold for all taxa.
Another key factor influencing species' responses to climate change is behavioural thermoregulation. Kearney et al. [102] explore how well ectothermic animals (e.g., reptiles, amphibians and insects) can use behaviour to cope with rising temperatures. In habitats with diverse microclimates, such as forests, animals can often mitigate the effects of climate change by seeking shade or by changing their activity patterns (Figure 4). However, in open and homogeneous landscapes, such as deserts, these options are limited, and species could reach thermal limits much faster. Species living near their upper thermal limits, particularly in tropical regions, have far less able to cope with additional warming. Kearney et al. [102] also highlight that behavioural buffering is only effective if microhabitats exist; in degraded landscapes, such as deforested areas and urban zones, thermoregulation may no longer be possible. This makes species in fragmented habitats particularly vulnerable. If generalist species with broad temperature tolerances can adapt more effectively than specialists, community structures may change and result in disruptions in food webs and ecosystem processes. To summarize, Kearney et al. [102] underscore the importance of preserving habitat complexity as a strategy for climate adaptation and highlight the unequal risks faced by species in different environments.
The effects of climate change on plant communities are also of great concern. Trivedi et al. [103] report potential effects of climate change on montane plant communities in three Scottish nature reserves using species distribution models. Their findings indicate that climate change could significantly alter the distribution of suitable habitats for many montane species of plants. While some species may experience reductions in suitable areas, for others new habitats become available. This variability in species responses underscores the complexity of predicting community-level effects. Conservation strategies may need to be adjusted to account for these changes, such as enhancing habitat connectivity to facilitate the spread of species or prioritizing areas predicted to remain suitable for threatened species.
To monitor shifts in plant diversity in response to climate change at a landscape scale, Stohlgren et al. [104] used a multi-scale vegetation monitoring method in the Rocky Mountain National Park, Colorado. Their research revealed that altitude and under-canopy solar radiation accounted for 64% of the variance in understory species distributions, highlighting the role of broad-scale climatic factors in shaping plant communities. However, small-scale environmental factors, such as light availability, soil fertility and soil moisture, also had significant roles. This study stressed the importance of monitoring ecotones, transitional zones between vegetation types, as important areas for tracking changes in plant diversity and patterns of spread under climate change. Their approach provides a framework for assessing climate change effects on plant communities by capturing both broad and fine-scale environmental influences.
Klanderud and Totland [105] also report how climate warming might influence alpine plant community diversity by considering the roles of dispersal ability and local biotic interactions. Their experimental warming and vegetation removal treatments in Norwegian alpine Dryas heath reveals that warming alone did not significantly affect species richness. However, when seeds from non-resident species were added, species richness increased, indicating that dispersal ability has a major role in shaping these communities. In addition, the removal of the natural vegetation facilitated the establishment of new species, suggesting that competition with existing plants restricts species establishment. These findings highlight dispersal ability and local biotic interactions as crucial determinants of the diversity of alpine plant communities and may have more immediate effects under climate warming than temperature increases alone.
Montane mammalian species are also highly vulnerable to climate change. McDonald and Brown [106] modelled the risk of extinction of these mammals in the Great Basin and predict that a 3°C increase in temperature could lead to the extinction of up to 62% of them. As suitable habitats shrink, smaller areas will support fewer species, leading to significant declines in biodiversity. Their findings indicate that species with restricted distributions on isolated mountain tops face particularly high risks of extinction because of climate change.
Similarly, Burns et al. [107] assess the potential effects of climate change on the diversity of mammals in U.S. national parks. Their study predicts a significant turnover of species due to climate-induced changes in their distributions, with some species disappearing from certain parks and new ones arriving. If atmospheric CO₂ levels double, then national parks may lose up to 20% of their mammal species. These findings highlight conservation challenges, as protected areas may no longer encompass the habitats needed to support their current biodiversity, underscoring the need for adaptive conservation strategies.
Levinsky et al. [108] extended the above analysis to European mammals and report how climate change might alter the distributions of 120 native species. Their bioclimatic envelope models predict that, assuming none spread to other areas, 5–9% of European mammals could face extinction, while 70–78% may be severely threatened due to reductions around the suitable range exceeding 30%. Endemic species are likely to be the most negatively affected, whereas widely distributed species less so. In addition, mammalian species richness is expected to decline dramatically in the Mediterranean area and increase in north-eastern Europe and at high altitudes. These findings highlight the need for conservation strategies that account for species' varying vulnerabilities and potential ability to spread into more suitable areas.
Finally, Emmerson et al. [109] explore how global changes in productivity might affect species interactions and the stability of ecological communities. Their results indicate that increased productivity could lead to a decline in the population sizes of the consumers due to the poor-quality of the food. Furthermore, the interactions between species may weaken, and population fluctuations could become more pronounced, making ecosystems more vulnerable to environmental disturbances. This study highlights the complex ways in which global changes can influence trophic interactions, with significant consequences for ecosystem stability and resilience.
Overall, these studies collectively highlight the far-reaching effects of climate change on ecological communities. Whether through changes in species distributions, altered food web dynamics, disruptions in mutualistic interactions, or increased risks of extinction, climate change poses a multifaceted threat to biodiversity. Addressing these challenges requires species-specific analyses, habitat conservation and adaptive management strategies to mitigate the effects of a warmer world.
2.5. Effect of CC on Predator–Prey and Plant–Pollinator Interactions
Despite much effort over the past decade on the ecological consequences of global warming, ecologists still have little understanding of the importance of interspecific interactions in species responses to environmental change. Models predict that predation should mitigate species responses to environmental change and that interspecific competition should aggravate species responses to environmental change. To test this prediction, Jiang and Kulczycki [110] studied how predation and competition affected the responses of two ciliates, Colpidium striatum and Paramecium tetraurelia, to changes in temperature in laboratory microcosms. They found that neither predation nor competition altered the responses of Colpidium striatum to changes in temperature and that competition but not predation altered the responses of Paramecium tetraurelia to changes in temperature. Asymmetric interactions and temperature-dependent interactions may have contributed to the disparity between model predictions and experimental results. Their results indicate that models that do not include the inherent complexities in ecological communities may be inadequate for predicting species responses to environmental change.
Anthropogenic climate change is widely predicted to increase the rate of extinction of species by hampering individual survival and reproduction, by reducing the amount and accessibility of suitable habitat, or by eliminating other organisms that are essential for the species in question [111]. They also stress that what is less well appreciated is the likelihood that climate change will directly disrupt or eliminate mutually beneficial (mutualistic) ecological interactions between species even before extinctions occur. Memmott et al. [111] studied the potential disruption of an ubiquitous mutualistic interaction in terrestrial habitats, that between plants and their pollinators. They used a network of interactions between 1420 different pollinators and 429 species of plants to simulate consequences of changes in phenology that can be expected with a doubling of atmospheric CO2. Depending on model assumptions, changes in phenology reduced the floral resources available to 17–50% of all pollinator species, which resulted in as much as half of the natural activity period of the animals to occur when no food plants were available. Reduced overlap between plants and pollinators also decreased the diet breadth of the pollinators. The predicted result of these disruptions is the extinction of the pollinators and the plants that depend on them.
2.6. CC Affects the Phenology of Organisms
Numerous studies indicate that currently birds are migrating to their breeding grounds earlier than previously, primarily due to the rise in average global temperatures e.g. [112,113,114,115,116]. Changes in timing of bird migration are among the best-documented biological responses to global warming [60,117]. Miller-Rushing et al. [118] report that currently many migratory birds are arriving at their breeding grounds earlier in spring than previously. This change in the timing of migration is directly linked to warmer springs. On average, birds have been arriving 2-4 days earlier per decade, demonstrating a clear effect of increasing temperature on patterns in migration.
Spottiswoode et al. [119] report that sexual selection can also influence changes in the timing of spring migration in birds in response to climate change. Species in which there is a strong sexual selection with early-arriving males having a reproductive advantage, have advanced their spring migration dates more significantly in recent decades than species with weak sexual selection. This indicates that sexual selection can result in more rapid changes in phenology in response to climate warming, highlighting the importance of considering mating systems when predicting species' responses to environmental changes.
Global warming may also influence the number of generations per year in certain species. Gomi et al. [120] report that climate change has affected the life cycle of the fall webworm (Hyphantria cunea) in Fukui, Japan. Historically, this population was bivoltine (two generations per year). However, within a span of seven years, the population became trivoltine (three generations per year). This change is attributed to a shorter critical photoperiod for diapause induction and an increased tendency for some individuals to not diapause, even under short photoperiods. These adaptations that are likely the result of recent climate warming, enable this species to exploit longer favourable seasons.
Climate change also significantly affects plants, which are now flowering markedly earlier than in the past. Plants are highly attuned to the seasonality of their environment and shifts in the timing of plant activity (i.e., phenology) provide some of the most compelling evidence that species and ecosystems are being influenced by global environmental change. Researchers report changes in phenology at many scales, including earlier spring flowering in individual plants and an earlier "spring green-up" of the land surface as revealed by satellite images. Cleland et al. [121] report how global environmental changes, particularly climate warming, are altering the timing of events in the life cycle plants. They emphasize that changes in phenology associated with global change are diverse and vary among species and in ecosystems. Understanding these varied responses is crucial for predicting future effects on ecosystem functions and services.
Miller-Rushing and Primack [122] analysed historical data on plant phenology from Concord, Massachusetts, originally recorded by Henry David Thoreau in the 1850s. They report that many plants in Concord over the past century and a half have flowered earlier in the spring due to rising temperatures. However, not all species adjusted their flowering times to the same extent, which resulted in changes in plant community composition. Species that were unable to change their time of flowering have declined in abundance or disappeared from the area, while those that were able to do so have persisted or even thrived (Figure 5). This study highlights how climate change can disrupt ecological relationships and alter community structure by affecting species differently based on their phenological plasticity.
Historical data was also studied by Post [123], who analysed 50 years of flowering data for three plant species in 26 populations in Norway. This study revealed that temporal fluctuations in flowering times were highly correlated both within and among species in these populations. Moreover, large-scale climatic fluctuations, particularly the Arctic Oscillation, synchronized flowering times of these three species over distances up to 500 km. These findings demonstrate that large-scale climate events can link populations by synchronizing their phenologies, which indicates that climatic changes can simultaneously influence many species over wide geographic areas.
Investigating the effect of climate change on phenology requires not only historical data but also an experimental approach. Dunne et al. [124] studied how climate change affects the flowering times of 11 subalpine meadow species in Gunnison County, Colorado. By combining experimental manipulations, such as advancing snowmelt dates and warming the soil, and observations along natural environmental gradients, they found that earlier snowmelt and warming the soil resulted in earlier flowering in most species. In addition, these conditions extended the flowering duration, particularly for early-flowering species. This study highlights the importance of integrating the results of many research approaches to understand the responses of species-specific phenology to climate change, which can have significant implications for ecosystem dynamics and species interactions.
Visser and Both [125] examine how climate change affects the timing of seasonal activities in various species. They emphasized that to assess whether changes in phenology are adequate, it is essential to compare them with an estimate of the optimum timing. Their analysis revealed that in many species there are mismatches, with the change in their phenology either too little or too much relative to the optimum. This misalignment has significant fitness consequences, potentially affecting survival and reproduction. The authors advocate the need for long-term data on phenology to link them to the optimum as this approach is crucial for evaluating the full effects of climate change on ecosystems.
When considering the effects of climate change on species, it is also necessary to consider what is happening in the soil. Perry et al. [126] report how climate-induced shifts in plant species distributions affect ecosystem stability, by focusing on interactions with what is happening in the soil. They emphasize that the compatibility between incoming plant species and existing soil mutualists, such as mycorrhizal fungi, is crucial for successful plant establishment. A lack of overlap between the soil mutualists of resident and spreading species could lead to ecosystem instability, potentially resulting in the proliferation of opportunistic species and soil degradation. The authors highlight the importance of considering belowground ecological processes in conservation strategies to maintain ecosystem stability during changes in climate.
2.7. Range Shifts Associated with CC
Current climate change may be a major threat to global biodiversity, but the extent of species loss will depend on how species respond to changing climates. Wiens [12] use data on climate-related changes in range to test the frequency of local extinctions related to recent climate change. The results indicate that climate-related local extinctions have already occurred in hundreds of species, including 47% of the 976 species surveyed. This frequency of local extinctions is similar in different climatic zones, clades and habitats, but significantly higher for tropical than temperate species (55% versus 39%), animals than plants (50% versus 39%), and in freshwater relative to terrestrial and marine habitats (74% versus 46% versus 51%). Overall, these results indicate that local extinctions related to climate change are already widespread, even though levels of climate change so far are modest relative to those predicted in the next 100 years. These extinctions will presumably become much more prevalent as global warming increases further by roughly 2-fold to 5-fold over the coming decades. According to Hampe and Petit [127], modern climate change is producing poleward range shifts of numerous taxa, communities and ecosystems worldwide. The response of species to changing environments is likely to be determined largely by population responses at the margins of a species range. In contrast to the poleward edge, the low-latitude limit (rear edge) of species ranges remains understudied, and the critical importance of rear edge populations as long-term stores of species genetic diversity and foci of speciation has been little acknowledged. Hampe and Petit [127] review recent findings from the fossil records, phylogeography and ecology to illustrate that rear edge populations are often disproportionately important for the survival and evolution of biota. Hampe and Petit [127] conclude that their ecological features, dynamics and conservation requirements differ from those of populations in other parts of the range, and some commonly recommended conservation practices might therefore be of little use or even counterproductive for rear edge populations.
Brooker et al. [128] emphasize the crucial role of biotic interactions (competition, predation and mutualism) in influencing species' ability to shift their ranges in response to climate change. These interactions can either facilitate or hinder the spread of species into new areas. The ability of species to disperse over long distances is a key factor determining whether they can track changes in climate zones. Species with limited dispersal abilities are unlikely to survive under the current pace of climate change, resulting in range contractions or local extinctions. Rapid climate change can outpace species' ability to adapt or spread elsewhere, resulting in changes in biodiversity and ecosystem composition. Brooker et al. [128] highlight that accurate models of changes in species' range should incorporate not only temperature and precipitation, but also biotic interactions and constraints on dispersal. This allows for more realistic predictions of future patterns in biodiversity under climate change. They also suggest that predicting species' range shifts in a changing climate is complicated by the interactions between environmental, biological and dispersal factors. There is no one-size-fits-all model, and predictions must consider local ecological contexts. In overall, this article stresses the importance of incorporating ecological complexity into models of species distribution due to climate change to better predict how biodiversity will be affected.
Gao et al. [129] identify climate change as the primary factor driving the expansion and contraction of alpine grasslands in the Mt. Qomolangma (Everest) National Nature Preserve on the southern Tibetan Plateau. Increasing temperatures in the region have facilitated the upward spread of alpine grasslands to higher altitudes, where warming allows plants to colonize previously inhospitable areas. Gao et al. [129] also highlight that aridification (decrease in precipitation and increase in evapotranspiration) can lead to contraction and degradation of grasslands at low altitudes. Changes in grassland dynamics are altitude dependent. Grassland has spread at high altitudes due to temperature increases, while at low altitudes they have contracted and become degraded due to lack of moisture. This study reveals that variability in regional climate, such as warming and different patterns in precipitation, has a significant influence on ecosystem responses, especially in fragile alpine environments like the Tibetan Plateau. Gao et al. [129] indicate there is a need for improved management strategies to mitigate the effects of climate change on alpine ecosystems and ensuring the preservation of the biodiversity and ecosystem functions in this sensitive region.
Harley and Paine [130] state that changes in a species’ distribution are often due to rare extreme events acting in combination with gradual warming, highlighting the complexity and unpredictability of species’ distributions. This study reveals that changes in distribution do not always occur gradually in response to climate change. Instead, sudden changes in distribution can occur when gradual warming is accompanied by rare major perturbations, such as heat waves or other environmental stressors. These rare, extreme events act as triggers, subjecting species to conditions outside their tolerance thresholds. Such events can have a disproportionate effect on ecosystems, often facilitating sudden range increases or contractions. The combination of gradual climate change and occasional extreme events destabilizes ecosystems, which results in the ranges of species changing much faster than expected under gradual warming alone. In addition, ecological contingencies, including interactions with other species, such as competition and predation have a significant role in determining how and when changes in range occur.
These interactions can either amplify or mitigate the effects of climate-related perturbations. The study by Harley and Paine [130] also highlights that there are thresholds in their responses to environmental change. This means that slow and steady warming may not cause a change in distribution until a critical threshold is reached. As predictions of species' responses to climate change must consider the role of extreme events, ecological contingencies and gradual warming, otherwise relying solely on average changes in temperature or precipitation risks underestimating the likelihood of sudden and dramatic changes in range.
Hill et al. [131] reveal that there has been a northward shift in the distributions of butterflies during the 20th century in response to climate warming. These shifts are associated with rising temperatures, which create opportunities for species to colonize areas that were previously unsuitable. The extent of these changes in range varies depending on the species and their ecological traits. Butterflies with high dispersal abilities and broad habitat preferences are more likely to successfully extend their ranges in response to warming. Many species have extended the northern boundary of their ranges, some are also experiencing range contractions at their southern or low-altitude boundaries where the climate has become too warm or dry. Hill et al. [131] report that continued climate warming will drive further changes in range, but the success of these extensions will depend on habitat availability, connectivity and the species' specific capacity for dispersal. Habitat fragmentation poses a significant challenge, as it can limit the ability of some species to track climate changes, even when suitable climatic conditions exist further north. Without connected habitats, colonization of new areas may be hindered. Butterflies serve as valuable indicators of the effects of climate change on biodiversity. Their changes in range are closely associated with changes in temperature and offering insights into broader ecological responses. To summarize, this study demonstrates that northward changes in range of butterflies are associated with climate warming during the 20th century, with future shifts contingent on ecological constraints, habitat availability and dispersal ability.
Pelini et al. [132] report that poleward extension of butterfly populations, although expected to occur due to climate warming, faces significant ecological and physiological limits that constrain their spread into newly suitable habitats. Even when climatic conditions there are favourable, other factors such as habitat quality, resource availability and biotic interactions, including competition and predation, may restrict their establishment and survival. At the margins of their northern range, butterfly populations often experience increased physiological stress, which can reduce survival and reproduction, further limiting their ability to spread polewards. Biotic interactions, such as competition with existing species or mismatches with host plants and resources hinder range extension at newly colonized poleward sites. The study by Pelini et al. [132] emphasizes the value of translocation experiments as a tool for testing species’ responses to climate change. These experiments show that range extensions are not driven solely by climatic factors - they are also influenced by ecological constraints. Efforts to facilitate changes in range, such as assisted spread, must therefore consider habitat suitability, resource availability and biotic interactions along with climatic factors to ensure the success of translocated populations. While climate warming creates new opportunities for poleward spread, the ability of species, such as butterflies, to establish populations in new regions is often restricted by physiological limits, habitat constraints and biotic interactions.
Parolo and Rossi [133] indicate that plants in the Alps have spread to higher altitudes in response to recent climate warming. Rising temperatures have created more favourable conditions at high altitudes and on average, plants have spread upwards by approximately 29 meters per decade, that is, there is a strong association between increases in temperature and changes in the distribution of species. However, not all species respond to climate warming in the same way, with some spreading more rapidly than others. These variations are likely to be influenced by ecological traits, such as species' ability to disperse (Figure 6) and its level of habitat specialization. The rate of upward spread reported by Parolo and Rossi [133] is faster than the historical trend, suggesting an accelerated response to the recent, more rapid warming in the Alps. The plants naturally growing at high altitudes face a heightened risk of range contraction or even local extinction, as the extent of suitable habitats at these altitudes decreases, leaving them with nowhere else to go. This upward spread of plants is a clear biological indicator of climate change and highlights the sensitivity of alpine ecosystems to rising temperatures. Such shifts may also result in changes in community composition, introducing new dynamics and potential competition among species that could alter the structure and function of alpine ecosystems. In conclusion, the study by Parolo and Rossi [133] highlights that climate warming is driving the upward spread of plants in the Alps, which has important implications for species persistence, biodiversity and the overall structure of high-altitude ecosystems.
Intraspecific competition is important in shaping how species shift their ranges in response to climate change [134]. At the edges of their ranges, competition can hinder further spread. However, density-dependent dispersal can alleviate the effect of competition and facilitate the colonization of new habitats. The ability of species to track climate change is strongly associated with their ecological traits, such as dispersal ability and sensitivity to competition. Species with strong density-dependent dispersal are better able to extend their ranges than those with poor dispersal ability. The dynamics at the edges of ranges is particularly important, as it reflects the interplay of dispersal, competition and climate-driven shifts in habitat, which together determine the speed and success of changes in range. Balancing the trade-off between competition and dispersal is important for species adapting to climate change. While competition can slow spread at edges of ranges, density-dependent dispersal acts as a compensatory mechanism, allowing species to overcome barriers and move into newly suitable areas. This balance is essential for those species that extend their ranges under changing environmental conditions. Accurate predictions of species' responses to climate change must include intraspecific competition and dispersal dynamics. These factors are critical for understanding which species are likely to adapt and thrive as the climate continues to warm. Ultimately, species with dispersal strategies that mitigate the effects of competition are likely to be better able to extend their ranges in response to changes in climate.
2.8. Future Perspectives of CC
It is one thing to agree on an international policy, quite another to achieve it [135]. The 192 signatories of the 1992 United Nations Framework Convention on Climate Change have committed themselves to reducing the emissions of carbon dioxide and other greenhouse gases. But policymakers around the world are still trying to work out how to achieve it. Unless emissions begin to decline very soon, severe disruption of the climate system will entail expensive remedial actions [135].
Thinking about worst-case scenarios is nothing new as climate scientists have been doing it for many years. Schneider [136] explores what a world with 1,000 p.p.m. of CO2 in its atmosphere might look like and report that many unique or rare systems would probably be lost, including Arctic Sea ice, mountain-top glaciers, most threatened and endangered species, coral-reef communities, and many high-latitude and high-altitude indigenous human cultures. People would be vulnerable in other ways too: Asian mega-delta cities would face rising sea levels and severe tropical cyclones, creating hundreds of millions of refugees; valuable infrastructure, such as the London or New York underground systems could be damaged or lost; the elderly would be at risk from unprecedented heat waves; and children, who are especially vulnerable to malnutrition in poor areas, would face food shortages [136]. The economic outlook is no better. With warming of just 1–3°C, it is predicted there will be a mixture of benefit and loss. More than a few degrees of warming, however, and aggregate monetary effects will become negative virtually everywhere; and in a 1,000 p.p.m. scenario, current literature suggests the outcomes would be almost universally negative and could amount to a substantial loss of gross domestic product. Millions of people who are at risk of flooding and water supply problems would provide further economic challenges, tropical rainforests would become more vulnerable to wildfires and a rise in sea levels of up to 10 metres is likely after the melting of the Greenland and West Antarctic ice sheets [137]. It is obvious that there is an urgent need to keep the CO2 level well below 1,000 p.p.m. A whole range of policies will be needed, with international cooperation, and this should not be delayed by having to wait for the outcome of the current political negotiations [136].
The ranges of plants and animals are changing in response to recent changes in climate [138]. With rising temperatures, ecosystems with ‘nowhere to go’ (e.g. mountains) are considered to be the most threatened [139,140]. Loarie et al. [58] present a new index of the rate of change of temperature derived from spatial gradients and multi-model ensemble of predictions of rates of temperature increase in the twenty-first century. Loarie et al. [58] report that the rate of change of temperature is most likely to be lowest in mountainous biomes, such as tropical and subtropical coniferous forests, temperate coniferous forest and montane grasslands and highest in flooded grasslands, mangroves and deserts.
Small protected areas exacerbate the problem in Mediterranean-type and temperate coniferous forest biomes, whereas large protected areas may mitigate the problem in desert biomes. These results indicate a need for management strategies for minimizing biodiversity loss due to climate change. Montane landscapes may ensure the survival of many species into the next century [58]. Elsewhere, further steps must be taken. These include slowing the temporal gradient in climate change by reducing emissions, increasing the ability of plants and animals to disperse by managed relocation [141], or increasing the size of protected areas by means of habitat corridors and new reserves [142].
2.9. Projections of CC and Their Shortcomings
Dorman [143] examines how global climate change influences species distributions and the challenges of predicting these changes. Their paper reviews methods used to model species distributions under different climate scenarios and assesses the reliability of these predictions and discusses the use of species distribution models (SDMs) to predict how the geographic range of species might change in response to climate change. These models often link current species distributions to environmental variables (temperature and precipitation) to predict future distributions based on predicted climate scenarios and highlight the limitations and uncertainties of using SDMs for predicting future distributions. Some of the key challenges include limited data on species ecological requirements, the assumption that current species-climate relationships will be the same in the future and the complex interactions between species that are often not included in models. A significant concern raised by this author is the potential for increased risk of extinctions, particularly of species with small ranges, low dispersal ability, or those living in very specific habitats. If species cannot spread elsewhere or adapt quickly enough to changing conditions, they may face local or global extinction and climate is just one of many factors affecting species distributions. For more accurate predictions, models need to include other elements like land use changes, biotic interactions (competition and predation) and adaptive capacity of species to new conditions.
2.10. Ecological Consequences of CC
McCarty [144] focuses on the observable effects of climate change on ecological systems and reports that geographic ranges of many species are changing in response to increasing temperatures. This often involves spreading into higher latitudes or altitudes where the climate is more favourable. Such shifts can disrupt established ecological communities and interactions. Also, the timing of key biological events like breeding, migration and flowering are altered due to climate change. McCarty [144] points out that many species are experiencing earlier onset of these events as temperatures warm. These shifts can lead to mismatches, such as between the availability of food resources and the needs of migratory species. Climate change is a significant driver of ecological change, with wide-ranging effects that require urgent attention from conservationists to mitigate effects and facilitate adaptation.
3. Land Use and Habitat Loss
To date humans have changed the Earth in ways that will be detectable in a millennium from now [145]. The problem of species extinction has received much attention globally [146]. Global biodiversity will be affected by both climate change and changes in land use [6,21], the latter of which is assumed to be the primary driver of species extinctions and reduction in biodiversity [2,9,21,28,146]. Around three-quarters of the land have been modified by human activities [147], resulting in major losses and fragmentation of natural habitats and an increase in the risk of species' extinction [5,148]. Many currently threatened plant species will become extinct [6] and losses of local species richness exceeding 20% are likely to substantially impair the functioning of ecosystems and their services, and thus human well-being [149]. However, biodiversity is not evenly distributed, and as a result, extinction rates may not be the same in all regions [150].
In countries overlapping Biodiversity Hotspots, plant species are endangered by increases in climate change-driven habitat loss [6]. Le Roux et al. [150] reports more and faster accrual of absolute numbers of extinctions in biodiversity hotspots compared to elsewhere. Another interesting result is that higher levels of taxonomic uniqueness are being lost in areas outside biodiversity hotspots. There are distinct temporal patterns in plant extinctions with agriculture, invasions and urbanization being significant drivers in hotspots, whereas outside hotspots hydrological disturbance is an important driver [150]. The work of Jantz et al. [21] on assessing the potential effect of future climate policy on the loss of suitable habitats in biodiversity hotspots due to associated land use changes revealed that future land use changes are likely to reduce natural vegetative cover by 26-58% in hotspots. These estimates of potential future extinctions are based only on changes in land use, and it is likely they would have been higher if the direct effects of climate change had been considered [21]. On the other hand, direct effects of climate change may even increase local diversity in some regions [5]. How local ecological assemblages are affected is less clear, which is of concern given their importance for the functioning and services of ecosystems [5]. Moreover, quantifying its effects on global plant diversity remains a challenge due to the limited data on their distributions and how they are affected by land use [9].
Many assessments of biodiversity have focused on global metrics, such as rates of species extinction, but ecosystem functions and services are more likely to depend on local diversity [5]. The assessment of plant extinctions in Spain indicates that habitat loss has been the main cause of extinction, and the effect of collecting plants is irrelevant [146]. Newbold et al. [5] report the results of an analysis of a terrestrial assemblage database of unprecedented geographic and taxonomic coverage focused on quantifying local biodiversity responses to land use and related changes. Their models indicate land use changes and associated pressures strongly reduce local terrestrial biodiversity, with global average reductions of 13.6% in within-sample species richness, 10.7% in total abundance and 8.1% in rarefaction-based species richness. Rapid further losses are predicted if current land use remains unchanged with losses concentrated in biodiverse but economically poor countries. Their predictions indicate that such widespread large losses are not inevitable. Results presented by Moreira et al. [9], based on a novel approach, indicate that 11% (about 42 010) of plants are threatened with extinction globally. They report that global plant species extinction risks are not homogeneously distributed but concentrated in several regions in Venezuela, Colombia, Central Eastern Brazil and Eastern Asia. This is likely to be because these regions have a high relative endemism combined with a high degree of land conversion [9]. Because poverty and poor governance can compromise conservation, Giam et al. [6] consider the economic condition and quality of governance along with the degree of plant species endangerment and future habitat loss to prioritize countries based on need for conservation. They identified Angola, Cuba, Democratic Republic of Congo, Ethiopia, Kenya, Laos, Madagascar, Myanmar, Nepal, Tajikistan and Tanzania as the countries in greatest need of assistance with conservation. Moreover, croplands of all levels of intensity contribute most to the estimated extinction threat to plants [9]. Their estimate of 11% is slightly higher than the 7-10% and 4% extinctions predicted by the models of Van Vuuren et al. [151] and Di Marco et al. [152], respectively.
The only way to better understand the magnitude and processes determining the extinction crisis faced by biodiversity in general, is to urgently initiate a regional- or at least country-level biodiversity assessments and to update existing ones. Such assessments will be vital in more accurately inferring the main drivers and trends in plant extinctions during the Anthropocene and for making informed predictions about the future and preservation of Earth’s flora [150]. Future extinctions could potentially be reduced by incorporating the preservation of habitats to reduce projected future land use changes in hotspots or by lessening the effect of future land use activities on biodiversity within hotspots as they are disproportionately located in the tropics, where most of the future land use change is likely to occur [10]. There is the possibility of a positive relationship between the level of climate-change mitigation and habitat preservation in biodiversity hotspots, which indicates that the implementation of measures to prevent climate change could simultaneously prevent habitat loss driven by land use change in highly biodiverse regions [21]. Taking into consideration all aspects of changes in land use, with concerted action and the right societal choices, global sustainability of local biodiversity may be an achievable goal [5]. To reduce biodiversity loss, it is important to take conservation measures, particularly in areas identified as the most threatened by risk of extinction. The most positive conservation steps include restoring and re-using croplands, protecting the remaining wilderness areas, improving the management of existing protected areas, and targeting the demands and supply chains [153,154,155] in countries with high need of conservation [6].
4. Environmental Factors
Agriculture and climate change are the major drivers of biodiversity change in the UK: Burns et al. [156] report that changes in the populations of species (~1970–2012) is most strongly associated with an intensification of agriculture and climate change. The effect of the former was primarily deleterious, whereas that of climatic change to date has been more mixed. Findings were similar for the three major taxonomic groups assessed (insects, vascular plants and vertebrates). In general, the way a habitat was managed had a greater effect than changes in its extent, which accords with the relatively small changes in the areas occupied by different habitats during the period of this study, compared to changes in habitat management. Of the drivers, conservation, management of agricultural land and habitat creation had the greatest effect. Contrary to conventional predictions of conservation and population biology, yearly fluctuations in climate may be essential for the persistence of rare species in invaded habitats [157].
Devi et al. [158] predict habitat suitability and species richness of plants in the Great Himalayan National Park under different climate change scenarios. Their model identifies precipitation in the driest month followed by altitude and annual mean temperature as the most important variables determining the distribution of plants. They report that the overall extent of suitable habitat increased for nine species, decreased for thirty-four and remained unchanged for one species. The species richness is currently higher in the western and southwestern parts of Great Himalayan National Park in the current scenario; however, it could change under future climatic scenarios.
Willard et al. [159] report the response of trees on islands in the Everglades to environmental change. They used an analysis of pollen in sediment cores from the islands in the greater Everglades ecosystem to reconstruct the timing of tree island formation, patterns in their development and response to specific climatic and environmental stressors. This data provides evidence of the rapidity of tree island response to specific hydrologic changes and facilitates prediction of the response to future changes associated with the restoration of the Everglades.
5. Effect of Invasive Non-Native Species
The spread of plant species is a natural and continuous process [160]. However, nowadays the rate of human – assisted spread (i.e. invasions sensu [161]) of plants is much higher than that by natural processes. The Galápagos Islands are an example of how humans accelerated the slow process of the natural spread of plants. It is estimated that the natural colonization rate (e.g. carried by birds or sea currents) of these islands was approximately one species of vascular plant every 10,000 years, whereas, over the past c. 500 years, the human assisted colonization increased to 1.2 species per year [162].
In contrast to native species, which evolved in a region or arrived there naturally without human intervention, there are many so called alien species, which are also known as "exotic," "introduced," or "non-native", which exist in an area due to direct or indirect human activities. Most of these species are temporary colonists and cannot become established without human assistance. Naturalized species, on the other hand, establish stable populations without human support, though they may not always spread. Those capable of spreading are classified as invasive species [160]. Thus, invasive alien species (IAS) are non-native organisms that have no natural predators in areas where they invade and thus can spread rapidly and affect native plants and animals, harming the natural environment. These plants pose a threat to biodiversity and ecosystems [163,164,165] as they can displace native plants and crops by competing for essential resources like space, nutrients, water, and light [166,167,168]. Moreover, it has been found that IAS can compete with native species for pollinators, resulting in the reduced seed production of native species [169,170,171,172]. Their social and economic effects are significant, as they affect human health, agriculture, fisheries and food production [173,174]. It is worth noting that the former US President Bill Clinton pointed out that the economic losses caused by the spread of alien (c.f. non-native, non-indigenous, foreign, exotic) species in the United States were up to $123 billion annually [175].
Despite ongoing conservation efforts, biodiversity loss is occurring at rates 100–10,000 times higher than that recorded in the Cenozoic era [176]. Alongside habitat loss due to land use changes, direct exploitation, pollution and climate change, the spread of IAS is also a major reason for this decline (Figure 7) [10]. In Europe, although most alien species are in human made habitats (industrial areas, arable land, parks and gardens), 37% of the naturalized alien species are in grasslands, 32% in woodlands, whereas the percentage in mires, bogs and fens is c. 10% [177,178,179]. However, the risk of invasion is not uniformly distributed globally (Sala et al. 2000). For example, areas with a Mediterranean climate and grassland ecosystems are at higher risk than others (e.g. northern temperate ecosystems) [10,180].
Because of their usually higher levels of physiological, phenotypic and ecological flexibility, IAS are often characterized by a greater ability to adapt to environmental changes than native species [181,182,183]. Thus, they are less vulnerable than the native species to changes in temperature or CO2 concentrations and will likely thrive under future climatic scenarios [184,185,186,187].
The negative effect of the spread of IAS into natural and semi-natural habitats on the diversity of local flora is well illustrated by their effect on orchids (both epiphytic and terrestrial orchids) inhabiting these habitats in different ways. However, orchid survival is negatively affected when IAS cover exceeds 20%, below which orchids are unaffected [188]. Moreover, natural and semi-natural grasslands in Europe, especially those in lowlands with a high number of orchids, are threatened by the invasion of alien species [189]. Thus, according to Scramoncin et al. [190], the spread of IAS in Italy was the most significant threat for four grassland orchids, especially those in non-protected areas. Orchids could be also negatively affected by the spread of IAS because they can affect interactions between orchids and fungi or/and pollinators. Specifically, such plants can adversely affect fungal communities [191] and pollination of orchids [192,193,194].
However, IAS also occur in tropical and subtropical areas where there are many epiphytic orchids. It is reported that in native forests invaded by IAS the native host trees are decreasing dramatically [195]. The increased mortality of native trees means that fewer and fewer suitable host plants are available for epiphytic orchids. Competition for light is only one of the direct threats that IAS pose to native orchids. As invasion by alien plants increases over time [195], the density of native trees continues to decline [196], leading to a gradual, long-term displacement of native trees by alien species in the invaded forests. Invasive plants also slow the growth rate of native trees [197], which is likely to result in smaller trees that are not suitable for epiphytic orchids. This reduction in tree size also limits seed production and regeneration, as smaller trees generally bear less fruit.
Like most of the other IAS that invade natural or semi-natural habitats and threaten native plants, there are also a few orchid species that are environmental weeds – species that invade native ecosystems and cause alterations and problems [198]. Studies indicate that the spread of certain orchids, considered invasive, will be facilitated by climate change, enabling them to spread into areas they do not currently inhabit (e.g. Disa bracteata [199]; Eulophia graminea [200]). Plant species, such as the orchid Spathoglottis plicata, is expected to negatively affect native plants [201].
6. Conclusions
As diversity continues to grow and expands its scope, we believe it is time to publish a comprehensive review that covers interesting aspects of the scope and highlights of multidisciplinary research in the field of ecology and diversity, which is this paper, a review of the relative importance of various environmental drivers that may affect the habitat suitability and survival of endangered species in the coming decades. This is an important issue that should be publicized and broadly discussed, and the hope is that this review will support and initiate this discussion.
Of course, as evident from the text of this review, the topic is so diverse that it is impossible to cover all its aspects. Therefore, (subjectively) only those topics that are of interest or important for us were selected. Of course, other authors might present a completely different comprehensive review in the future.
There is also another side to this story. Future species distributions are usually presented on habitat suitability maps [48], which are generated by species distribution models (SDMs), whose inputs are locations of species in nature and climatic variables generated by climate models [37,38,39], abiotic factors such as geological substrate and soil [40,41], and in a few cases also biotic factors such as key species or distributions of pollinators [42,43,44]. The use of SDMs is currently very popular [48] as according to the Web of Science, more than 6,000 papers using SDMs were published over the last 25 years, and this number is rapidly increasing [39]. Many of these studies predict how the distribution of species will be affected by climate change [e.g., 43,45–47]. However, it is impossible to verify the accuracy of such predictions, because they refer to scenarios 30, 50, or more years from now and include lots of uncertainties [48]: (i) future climatic scenarios, (ii) adaptation capacity, (iii) speed of organisms’ spread into more favourable areas [49,50,51,52], (iv) effect of the spatial scale on the analyses [27], (v) extinction rates associated with increased temperature [49], (vi) life history and population dynamics of the studied species, etc. [48].
Thus, it is worth questioning the reliability of these predictions. The answers are very important as according to Araújo et al. [54], it is estimated the habitats of 63% of the species of concern in Europe will become climatically unsuitable in the Natura 2000 network by 2080. But are all the predicted changes in distribution and losses of these species realistic? Santini et al. [55] assessed the reliability of species distribution projections in climate change research. They conclude that our ability to predict future distributions of species is low on average and they advise caution in the application and interpretation of SDM predictions under different climates. This further highlights the necessity for determining the reliability of SDM predictions [48].
Author Contributions
Conceptualization, P.K. and Z.Š.; methodology, P.K. and Z.Š.; validation, P.K., S.T. and Z.Š.; resources, P.K., S.T. and Z.Š.; writing—original draft preparation, P.K., S.T., I.T. and Z.Š.; writing—review and editing, P.K. and Z.Š.; visualization, L.B.; supervision, P.K. and Z.Š.; project administration, P.K. and Z.Š. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
MDPI Research Data Policies.
Acknowledgements
We are greatly indebted to Tony Dixon for uncountable helpful hints as to how to improve the style of English in this paper.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Román-Palacios, C.; Wiens, J.J. Recent responses to climate change reveal the drivers of species extinction and survival. PNAS 2020, 117(8), 4211–4217. [CrossRef]
- Antonelli, A.; Fry, C.; Smith, R.J.; Eden, J.; Govaerts, R.H.A.; Kersey, P.; Nic Lughadha, E.; Onstein, R.E.; Simmonds, M.S.J.; Zizka, A.; et al. State of the World’s Plants and Fungi, 2023; Royal Botanic Gardens: Kew, UK, 2023.
- WWF. Living Planet Report. WWF, 2020.
- Jaureguiberry, P.; Titeux, N.; Wiemers, M.; Bowler, D.E.; Coscieme, L.; Golden, A.S.; Guerra, C.A.; Jacob, U.; Takahashi, Y.; Settele, J.; et al. The direct drivers of recent global anthropogenic biodiversity loss. Sci. Adv. 2022, 8, eabm9982. [CrossRef]
- Newbold, T.; Hudson, L.N.; Hill, S.L.L.; Contu, S.; Lysenko, I.; Senior, R.A.; Börger, L.; Bennett, D.J.; Chiomes, A.; Collen, B. Global effects of land use on local terrestrial biodiversity. Nature 2015, 520, 45–70.
- Giam, X.; Bradshaw, C.J.A.; Tan, H.T.W.; Sodhi, N.S. Future habitat loss and the conservation of plant biodiversity. Biol. Conserv. 2010, 143, 1594–1602.
- Rejmánek, M. Vascular plant extinctions in California: A critical assessment. Divers. Distrib. 2018, 24, 129–136.
- Rejmánek, M.; Krahulec, F.; Grulich, V. Jak rychle a proč vymírají rostliny v antropocénu. Živa 2021, 5, 219–223.
- Moreira, H.; Kuipers, K.J.J.; Posthuma, L.; Zijp, M.C.; Hauck, M.; Huijbregts, M.A.J.; Schipper, A.M. Threats of land use to the global diversity of vascular plants. Divers. Distrib. 2023, 29, 688–697.
- Sala, O.E.; Stuart Chapin, F.; Armesto, J.J.; Berlow, E.; Bloomfield, J.; Davis, F.; Dirzo, R.; Froydis, I.; Huber-Sanwald, E.; Huenneke, L.F.; et al. Global biodiversity scenarios for the Year 2100. Science 2000, 287, 1770–1774.
- Urban, M.C. Accelerating extinction risk from climate change. Science 2015, 348, 571–573.
- Wiens, J.J. Climate-related local extinctions are already widespread among plant and animal species. PLOS Biol. 2016, 14(12), e2001104. [CrossRef]
- Warren, R.; Price, J.; Graham, E.; Forstenhaeusler, N.; Vanderwal, J. The projected effect on insects, vertebrates, and plants of limiting global warming to 1.5°C rather than 2°C. Science 2018, 360, 791–795.
- Pigot A.L.; Merow, C.; Wilson, A.; Trisos, C.H. Abrupt expansion of climate change risk for species globally. Nature Ecol. Evol. 2023, 7, 1060–1071.
- Mancini, G.; Santini, L.; Gazalis, V.; Akcakaya, H.R.; Lucas, P.M.; Brooks, T.M.; Foden, W.; Di Marco, M. A standard approach for including climate change responses in IUCN Red List assessments. Conserv. Biol. 2024, 38(3), e14227. [CrossRef]
- Wiens, J.J.; Zelinka, J. How many species will Earth lose to climate change? Global Change Biol. 2024, 30, e17125. [CrossRef]
- Stein, B.; Glick, P.; Edelson, N.; Staudt, A. Climate-smart conservation: Putting adaptation principles into practice. National Wildlife Federation: Washington D.C., USA, 2014.
- Tsiftsis, S.; Djordjević, V.; Tsiripidis, I. Neottia cordata (Orchidaceae) at its southernmost distribution border in Europe: threat status and effectiveness of Natura 2000 Network for its conservation. J. Nat. Conserv. 2019, 48, 27–35.
- Pearson, R.G. Species’ distribution modeling for conservation educators and practitioners. Lessons Conserv. 2010, 3, 54–89.
- Mantyka-Pringle, C.S.; Martin, T.G.; Rhodes, J.R. Interactions between climate and habitat loss effects on biodiversity: a systematic review and meta-analysis. Global Change Biol. 2012, 18, 1239–1252.
- Jantz, S.M.; Barker, B.; Brooks, T.M.; Chini, L.P.; Huang, Q.; Moore, R.M.; Noel, J.; Hurtt, G.C. Future habitat loss and extinctions driven by land use change in biodiversity hotspots under four scenarios of climate-change mitigation. Conserv. Biol. 2015, 29(4), 1122–1131.
- Nunez, S.; Alkemande, R. Exploring interaction effects from mechanisms between climate and land–use changes and the projected consequences on biodiversity. Biodivers. Conserv. 2021, 30, 3685–3696.
- Suppula, M.; Hällfors, M.H.; Apala, K.; Aalto, J.; Kemppainen, E.; Leikola, N.; Pirinen, P.; Heikkinen, R.K. Climate and landscape-use change drive population decline in a red-listed plant species. Global Ecol. Conserv. 2023, 45, e02526. [CrossRef]
- Oliver, T.H.; Morecroft, M.D. Interactions between climate change and land use change on biodiversity: attribution problems, risks, and opportunities. WIREs Clim. Change 2014, 5(3), 317–335. [CrossRef]
- Cabral, J.S.; Mendoza-Poce, A.; Silva, A.P.; Oberpriller, J.; Mimet, A.; Kieslinger, J.; Berger, T.; Blechschmidt, J.; Broenner, M.; Classen, A.; et al. The road to integrate climate change projections with regional land use-biodiversity models. People and Nature 2023, 6(5), 1716–1741. [CrossRef]
- Travis, J.M.J. Climate change and habitat destruction: a deadly anthropogenic cocktail. P. Roy. Soc. B-Biol. Sci. 2003, 270, 467–473.
- Štípková, Z.; Tsiftsis, S.; Kindlmann, P. How did the agricultural policy during the communist period affect the decline in orchid biodiversity in central and eastern Europe? Global Ecol. Conserv. 2021, 26, e01498. [CrossRef]
- Gallego-Zamorano, J.; Huijbregts, M.A.J.; Schipper, A.M. Changes in plant species richness due to land use and nitrogen deposition across the globe. Divers. Distrib. 2022, 28, 745–755.
- Harrison, S. Plant community diversity will decline more than increase under climate warming. Philos. T. Roy. Soc. B 2020, 375, 20190106.
- Miniere, A.; von Schuckman, K.; Sallée, J.-B.; Vogt, L. Robust acceleration of Earth system heating observed over the past six decades. Sci. Rep. 2023, 13, 22975. [CrossRef]
- Wudu, K.; Abegaz, A.; Ayele, L.; Ybabe, M. The impact of climate change on biodiversity loss and its remedial using nature-based conservation approach: a global perspective. Biodivers. Conserv. 2023, 32, 3681–3701.
- Rejmánek, M. Jsme na počátku zrychleného oteplování? Vesmír 2024, 103, 110–113.
- Moss, B.; Kosten, S.; Meerhoff, M.; Battarbee, R.W.; Jeppesen, E.; Mazzeo, N.; Havens, K.; Lacerot, G.; Liu, Z.W.; De Meester, L.; et al. Allied attack: climate change and eutrophication. Inland Waters 2011, 1(2), 101–105. [CrossRef]
- Nazari-Sharabian, M.; Ahmad, S.; Karakouzian, M. Climate change and eutrophication: A short review. ETASR 2018, 8(6), 3668–3672.
- Wei, Y.Q.; Zhang, L.; Wang, J.N.; Wang, W.W.; Niyati, N.; Guo, Y.L.; Wang, X.F. Chinese caterpillar fungus (Ophiocordyceps sinensis) in China: Current distribution, trading, and futures under climate change and overexploitation. Sci. Total Environ. 2021, 755, 1. [CrossRef]
- Raza, M.M.; Bebber, D.P. Climate change and plant pathogens. Curr. Opin. Microbiol. 2022, 70, 102233. [CrossRef]
- Pearson, R.G.; Raxworthy, C.; Nakamura, M.; Peterson, A. Predicting species distributions from small numbers of occurrence records: A test case using cryptic geckos in Madagascar. J. Biogeogr. 2006, 34, 102–117.
- Elith, J.; Leathwick, J. The contribution of species distribution modelling to conservation prioritization. In Spatial conservation prioritization. Quantitative methods & computational tools; Moilanen, A., Wilson, A.K., Possingham, H.P., Eds.; Oxford University Press Inc.: New York, USA, 2009; pp. 70–93.
- Araújo, M.B.; Anderson, R.P.; Barbosa, A.M.; Beale, C.M.; Dormann, C.F.; Early, R.; Garcia, R.A.; Guisan, A.; Maiorano, L.; Naimi, B.; et al. Standards for distribution models in biodiversity assessments. Sci. Adv. 2019, 5(1), eaat4858. [CrossRef]
- Djordjević, V.; Tsiftsis, S.; Lakušić, D.; Jovanović, S.; Stevanović, V. Factors affecting the distribution and abundance of orchids in grasslands and herbaceous wetlands. Syst. Biodivers. 2016, 14(4), 355–370. [CrossRef]
- Štípková, Z.; Kindlmann, P. Factors determining the distribution of orchids – a review with examples from the Czech Republic. Eur. J. Environ. Sci. 2021, 11, 21–30.
- Giannini, T.C.; Chapman, D.S.; Saraiva, A.M.; Alves-dos-Santos, I.; Biesmeijer, J.C. Improving species distribution models using biotic interactions: a case study of parasites, pollinators and plants. Ecography 2013, 36(6), 649–656. [CrossRef]
- Tsiftsis, S.; Djordjević, V. Modelling sexually deceptive orchid species distributions under future climates: the importance of plant–pollinator interactions. Sci. Rep. 2020, 10, 10623. [CrossRef]
- Kolanowska, M. The future of a montane orchid species and the impact of climate change on the distribution of its pollinators and magnet species. Global Ecol. Conserv. 2021, 32, e01939. [CrossRef]
- Abdelaal, M.; Fois, M.; Dakhil, M.A.; Bacchetta, G.; El-Sherbeny, G.A. Predicting the potential, current and future distribution of the endangered endemic vascular plant Primula boveana Decne. ex Duby in Egypt. Plants 2020, 9(8), 957. [CrossRef]
- Pinto-Ledezma, J.N.; Cavender-Bares, J. Predicting species distributions and community composition using satellite remote sensing predictors. Sci. Rep. 2021, 11, 16448. [CrossRef]
- Zhang, Y.; Tang, J.; Ren, G.; Zhao, K.; Wang, X. Global potential distribution prediction of Xanthium italicum based on Maxent model. Sci. Rep. 2021, 11, 16545. [CrossRef]
- Tsiftsis, S.; Štípková, Z.; Rejmánek, M.; Kindlmann, P. Predictions of species distributions based only on models estimating future climate change are not reliable. Sci. Rep. 2024, 14(1), 25778.
- Kougioumoutzis, K.; Kokkoris, I.P.; Panitsa, M.; Trigas, P.; Strid, A.; Dimopoulos, P. Plant diversity patterns and conservation implications under climate-change scenarios in the Mediterranean: The case of Crete (Aegean, Greece). Diversity 2020, 12(7), 270. [CrossRef]
- Fedorov, N.; Kutueva, A.; Muldashev, A.; Mikhaylenko, O.; Martynenko, V.; Fedorova, Y. Prediction of habitat suitability for Patrinia sibirica Juss. in the Southern Urals. Sci. Rep. 2021, 11, 19606. [CrossRef]
- Shrestha, B.; Tsiftsis, S.; Chapagain, D.-J.; Khadka, C.; Bhattarai, P.; Kayastha, N.; Kolanowska, M.; Kindlmann, P. Dactylorhiza hatagirea in Nepal: Distribution prediction under current and future climate change context. Plants 2021, 10(3), 467.
- Evans, A.; Jacquemyn, H. Range size and niche breadth as predictors of climate–induced habitat change in Epipactis (Orchidaceae). Front. Ecol. Evol. 2022, 10, 894616. [CrossRef]
- Švecová, M.; Štípková, Z.; Traxmandlová, I.; Kindlmann, P. Difficulties in determining distribution of population sizes within different orchid metapopulations. Eur. J. Environ. Sci. 2023, 13, 96–109. [CrossRef]
- Araújo, M.B.; Alagador, D.; Cabeza, M.; Nogués-Bravo, D.; Thuiller, W. Climate change threatens European conservation areas. Ecol. Lett. 2011, 14(5), 484–492.
- Santini, L.; Benítez-López, A.; Maiorano, L.; Čengić, M.; Huijbregts, M.A.J. Assessing the reliability of species distribution projections in climate change research. Divers. Distrib. 2021, 27, 1035–1050. [CrossRef]
- Valentin, D.N.; Adamo, M.; Richiardi, C.; Mammola, S.; Kull, T. Impact of climate and land use change on the distribution of orchids in Estonia. Plant Ecol. 2025, 226, 831–844. [CrossRef]
- Balvanera, P., Pfaff, A., Viña, A., García-Frapolli, E., Merino, L., Minang, P.A., Nagabhatla, N., Hussain, S A.; Sidorovich, A.A. Chapter 2.1. Status and Trends – Drivers of Change. In Global assessment report of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services; Brondízio, E.S., Settele, J., Díaz, S., Ngo, H.T., Eds.; IPBES secretariat: Bonn, Germany, 2019; [CrossRef]
- Loarie, S.R.; Duffy, P.B.; Hamilton, H.; Asner, G.P.; Field, C.B.; Ackerly, D.D. The velocity of climate change. Nature 2009, 462, 1052–1055. [CrossRef]
- Smit, B.; Pilifosova, O.; Burton, I.; Challenger, B.; Huq, S.; Klein, R.J.T.; Yohe, G. Adaptation to climate change in the context of sustainable development and equity. In Climate Change 2001—Impacts, Adaptation, and Vulnerability; McCarthy, J.J., Canziani, O.F., Leary, N.A., Dokken, D.J., White, K.S., Eds.; Cambridge University Press: Cambridge, UK, 2001; pp. 877–912.
- Root, T.L.; Price, J.T.; Hall, K.R.; Schneider, S.H.; Rosenzweig, C.; Pounds, J.A. Fingerprints of global warming on wild animals and plants. Nature 2003, 421, 57–60.
- Lucht, W., Prentice, I.C.; Myneni, R.B.; Sitch, S.; Friedlingstein, P.; Cramer, W.; Bousquet, P.; Buermann, W.; Smith, B. Climatic control of the high-latitude vegetation greening trend and Pinatubo effect. Science 2002, 296, 1687–1689. [CrossRef]
- Rummukainen, M. Changes in climate and weather extremes in the 21st century. WIREs Clim. Change 2023, 3, 115–129. [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [CrossRef]
- King, A.D.; Donat, M.G.; Fischer, E.M.; Hawkins, E.; Alexander, L.V.; Karoly, D.J.; Dittus, A.J.; Lewis, S.C.; Perkins, S.E. The timing of anthropogenic emergence in simulated climate extremes. Environ. Res. Lett. 2015, 10, 094015.
- Mitchell, J.F.B.; Lowe, J.; Wood, R.A.; Vellinga, M. Extreme events due to human-induced climate change. Philos. T. Roy. Soc. A 2006, 364, 2117–2133.
- Luber, G.; McGeehin, M. Climate Change and Extreme Heat Events. Am. J. Prev. Med. 2008, 35, 429–435. [CrossRef]
- Cai, W.; Santoso, A.; Wang, G.; Yeh, S.-W.; An, S.-I.; Cobb, K.M.; Collins, M.; Guilyardi, E.; Jin, F.-F.; Kug, J.-S.; et al. ENSO and greenhouse warming. Nature Clim Change 2015, 5, 849–859. [CrossRef]
- L'Heureux, M.; Takahashi, K.; Watkins, A.; Barnston, A.; Becker, E.; Liberto, T.; Gamble, F.; Gottschalck, J.; Halpert, M.; Huang, B.; et al. Observing and Predicting the 2015–16 El Niño. B. Am. Meteorol. Soc. 2017, 98, 1363–1382. [CrossRef]
- Weller, T.; Castle, K.; Liechti, F.; Hein, C.D.; Schirmacher, M.R.; Cryan P.M. First direct evidence of long-distance seasonal movements and hibernation in a migratory bat. Sci. Rep. 2016, 6, 34585. [CrossRef]
- Newman, M.; Wittenberg, A.T.; Cheng, L.; Compo, G.P.; Smith, C.A. The Extreme 2015/16 El Niño, in the Context of Historical Climate Variability and Change. B. Am. Meteorol. Soc. 2018, 99, S16–S20. [CrossRef]
- Schneider, S.H. Abrupt non-linear climate change, irreversibility and surprise. Global Environ. Change 2004, 14(3), 245–258. [CrossRef]
- Hannah, L.; Midgley, G.F.; Lovejoy, T.; Bond, W.J.; Bush, M.; Lovett, J.C.; Scott, D.; Woodwards, F.I. Conservation of Biodiversity in a Changing Climate. Conserv. Biol. 2002, 16(1), 264–268.
- McLaughlin, J.F.; Hellmann, J.J.; Boggs, C.L.; Ehrlich, P.R. Climate change hastens population extinctions. PNAS 2002, 99(9), 6070–6074. [CrossRef]
- Warren, M.S.; Hill, J.K.; Thomas, J.A.; Asher, J.; Fox, R.; Huntley, B.; Roy, D.B.; Telfer, M.G.; Jeffcoate, S.; Harding, P.; et al. Rapid responses of British butterflies to opposing forces of climate and habitat change. Nature 2001, 414, 65–69.
- Summers, D.M.; Bryan B.A.; Crossman, N.D.; Meyer, W.S. Species vulnerability to climate change: impacts on spatial conservation priorities and species representation. Global Change Biol. 2012, 18, 2335–2348. [CrossRef]
- Holzman, K.L.; Walls, R.L.; Wiens, J.J. Accelerating local extinction associated with very recent climate change. Ecol. Lett. 2023, 26, 1877–1886. [CrossRef]
- Lemoine, N.; Schaefer, H.-C.; Böhning-Gaese, K. Species richness of migratory birds is influenced by global climate change. Global Ecol. Biogeogr. 2006, 16(1), 55–64. [CrossRef]
- Lemoine, N.; Böhning-Gaese, K. Potential impact of global climate change on species richness of long-distance migrants. Conserv. Biol. 2002, 17, 577–586. [CrossRef]
- Walther, G.R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.C.; Fromentin, J.M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [CrossRef]
- Menéndez, R.; Gonzáles-Megías, A.; Hill, J.K.; Braschler, B.; Willis, S.G.; Collingham, Y.; Fox, R.; Roy, D.B.; Thomas, C.D. Species richness changes lag behind climate change. P. Roy. Soc. B-Biol. Sci. 2006, 273, 1465–1470. [CrossRef]
- Sun, Y.; Sun, Y.; Yao, S.; Akram, M.A.; Hu, W.; Dong, L.; Li, H.; Wei, M.; Gong, H.; Xie, S.; et al. Impact of climate change on plant species richness across drylands in China: From past to present and into the future. Ecol. Indicators 2021, 132, 108288. [CrossRef]
- Wang, Q.; Zhang, Z.; Du, R.; Wang, S.; Duan, J.; Iler, A.M.; Piao, S.; Luo, C.; Jiang, L.; Lv, W.; et al. Richness of plant communities plays a larger role than climate in determining responses of species richness to climate change. J. Ecol. 2019, 107, 1944–1955. [CrossRef]
- Sommer, J.H.; Kreft, H.; Kier, G.; Jetz, W.; Mutke, J.; Barthlott, W. Projected impacts of climate change on regional capacities for global plant species richness. P. Roy. Soc. B-Biol. Sci. 2010, 277, 2271–2280. [CrossRef]
- Melillo, J.M. Warm, warm on the range. Science 1999, 283, 183–184.
- Iverson, L.R.; Prasad, A.M. Potential changes in tree species richness and forest community types following climate change. Ecosystems 2001, 4, 186–199. [CrossRef]
- Khalyani, A.H.; Gould, W.A.; Falkowski, M.J.; Muscarella, R.; Uriarte, M.; Yousef, F. Climate change increases potential plant species richness on Puerto Rican uplands. Clim. Change 2019, 156, 15–30. [CrossRef]
- Urban, M.C.; Tewksbury, J.J.; Sheldon, K.S. On a collision course: Competition and dispersal differences create no-analogue communities and cause extinctions during climate change. P. Roy. Soc. B-Biol. Sci. 2012, 279, 2072–2080. [CrossRef]
- Blois, J.L.; Zarnetske, P.L.; Fitzpatrick, M.C.; Finnegan, S. Climate change and the past, present, and future of biotic interactions. Science 2013, 341, 499–504. [CrossRef]
- Walker, M.D.; Wahren, C.H.; Hollister, R.; Henry, G.H.R.; Ahlquist, L.E.; Alatalo, J.M.; Bret-Harte, M.S.; Calef, M.P.; Callaghan, T.V.; Carroll, A.B. Plant community responses to experimental warming across the tundra biome. PNAS 2006, 103, 1342–1346. [CrossRef]
- Elmendorf, S.C.; Henry, G.H.R.; Hollister, R.D.; Björk, R.G.; Bjorkman, A.D.; Callaghan, T.V.; Collier, L.S.; Cooper, E.J.; Cornelissen, J.H.C.; Day, T.A.; et al. Global assessment of experimental climate warming on tundra vegetation: Heterogeneity over space and time. Ecol. Lett. 2012, 15, 164–175. [CrossRef]
- Alexander, J.M.; Diez, J.M.; Levine, J.M. Novel competitors shape species’ responses to climate change. Nature 2015, 525, 515–518. [CrossRef]
- O´Neill, G.A.; Hamann, A.; Wang, T. Accounting for population variation improves estimates of the impact of climate change on species’ growth and distribution. J. Appl. Ecol. 2008, 45, 1040–1049. [CrossRef]
- Ahola, M.P.; Laaksonen, T.; Eeva, T.; Lehikoinen, E.; Blackwell Publishing Ltd. Climate change can alter competitive relationships between resident and migratory birds. J. Anim. Ecol. 2007, 76, 1045–1052. [CrossRef]
- Harrington, R.; Fleming, R.A.; Wolwod, I.P. Climate change impacts on insect management and conservation in temperate regions: can they be predicted? Agr. Forest Entomol. 2001, 3, 233–240.
- Menéndez, R.; Gonzáles-Megías, A.; Collingham, Y.; Fox, R.; Roy, D.B.; Ohlemüller, R.; Thomas, C.D. Direct and indirect effects of climate and habitat factors on butterfly diversity. Ecology 2007, 88, 605–611.
- Van der Putten, W.H.; de Ruiter, P.C.; Bezemer, T.M.; Harvey, J.A.; Wassen, M.; Wolters, V. Trophic interactions in a changing world. Basic Appl. Ecol. 2004, 5, 487–494. [CrossRef]
- Parmesan, C. Ecological and evolutionary responses to recent climate change. Ann. Rev. Ecol. Evol. Syst. 2006, 37, 637–669. [CrossRef]
- Tylianakis, J.M.; Didham, R.K.; Bascompte, J.; Wardle, D.A. Global change and species interactions in terrestrial ecosystems. Ecol. Lett. 2008, 11, 1351–1363. [CrossRef]
- Tylianakis, J.M. Warming up food webs. Ecology 2009, 323, 1300–1301. [CrossRef]
- Wilson, R.J.; Maclean, I.M.D. Recent evidence for the climate change threat to Lepidoptera and other insects. J. Insect. Conserv. 2011, 15, 259–268.
- Hellmann, J.J.; Pelini, S.L.; Prior, K.M.; Dzurisin, J.D.K. The response of two butterfly species to climatic variation at the edge of their range and the implications for poleward range shifts. Oecologia 2008, 157, 583–592. [CrossRef]
- Kearney, M.; Shine, R.; Porter, W.P. The potential for behavioral thermoregulation to buffer “cold-blooded” animals against climate warming. PNAS 2009, 106, 3835–3840. [CrossRef]
- Trivedi, M.R.; Morercroft, M.D.; Berry, P.M.; Dawson, T.P. Potential effects of climate change on plant communities in three montane nature reserves in Scotland, UK. Biol. Conserv. 2008, 141, 1665–1675.
- Stohlgren, T.J.; Owen, A.J.; Lee, M. Monitoring shifts in plant diversity in response to climate change: a method for landscapes. Biodivers. Conserv. 2000, 9, 65–86.
- Klanderud, K.; Totland, O. The relative role of dispersal and local interactions for alpine plant community diversity under simulated climate warming. Oikos 2007, 116, 1279–1288. [CrossRef]
- McDonald, K.A.; Brown, J.H. Using montane mammals to model extinctions due to global change. Conserv. Biol. 1992, 6, 409–415.
- Burns, C.E.; Johnston, K.M.; Schmitz, O.J. Global climate change and mammalian species diversity in U.S. national parks. PNAS 2003, 100, 11474–11477. [CrossRef]
- Levinsky, I.; Skov, F.; Svenning, J.-C.; Rahbek, C. Potential impacts of climate change on the distributions and diversity patterns of European mammals. Biodivers. Conserv. 2007, 16, 3803–3816.
- Emmerson, M.; Bezemer, T.M.; Hunter, M.D.; Jones, T.H.; Masters, G.J.; van Dam, N.M. How does global change affect the strength of trophic interactions? Basic Appl. Ecol. 2004, 5, 505–514. [CrossRef]
- Jiang, L.; Kulczycki, A. Competition, predation and species responses to environmental change. Oikos 2004, 106, 217–224.
- Memmott, J.; Craze, P.G.; Waser, N.M.; Price, M.V. Global warming and the disruption of plant-pollinator interactions. Ecol. Lett. 2007, 10, 710–717. [CrossRef]
- Cotton, P.A. Avian migration phenology and global climate change. PNAS 2003, 100, 12219–12222.
- Hüppop, O., Hüppop, K. North Atlantic Oscillation and timing of spring migration in birds. P. Roy. Soc. B-Biol. Sci. 2003, 270, 233–240.
- Marra, P.P.; Francis, C.M.; Mulvihill, R.S.; Moore, F.R. The influence of climate on the timing and rate of spring bird migration. Oecologia 2005, 142, 307–315.
- Mills, A.M. Changes in the timing of spring and autumn migration in North American migrant passerines during a period of global warming. Ibis 2005, 147, 259–269.
- Sparks, T.H.; Bairlein, F.; Bojarinova, J.G.; Hüppop, O.; Lehikoinen, E.A.; Rainio, K.; Sokolov, L.V.; Walker, D. Examining the total arrival distribution of migratory birds. Global Change Biol. 2005, 11, 22–30.
- Parmesan, C. Influences of species, latitudes and methodologies on estimates of phenological response to global warming. Global Change Biol. 2007, 13, 1860–1872.
- Miller-Rushing, A.J.; Lloyd-Evans, T.L.; Primack, R.B.; Satzinger P. Bird migration times, climate change, and changing population sizes. Glob. Change Biol. 2008, 14, 1–14.
- Spottiswoode, C.N.; Tottrup, A.P.; Coppack, T. Sexual selection predicts advancement of avian spring migration in response to climate change. P. Roy. Soc. B-Biol. Sci. 2006, 273, 3023–3029.
- Gomi, T.; Nagasaka, M.; Fukunda, T.; Hagihara, H. Shifting of the life cycle and life-history traits of the fall webworm in relation to climate change. Entomol. Exp. Appl. 2007, 125, 179–184. [CrossRef]
- Cleland, E.E.; Chuine, I; Menzel, A.; Mooney, H.A.; Schwartz, M.D. Shifting plant phenology in response to global change. Trends Ecol. Evol. 2007, 22. [CrossRef]
- Miller-Rushing, A.J.; Primack, R.B. Global warming and flowering times in Thereau´s Concord: A community perspective. Ecology 2008, 89, 332–341.
- Post, E. Large-scale climate synchronizes the timing of flowering by multiple species. Ecology 2003, 84, 277–281.
- Dunne, J.A.; Harte, J.; Taylor, K.J. Subalpine meadow flowering phenology responses to climate change: integrating experimental and gradient methods. Ecol. Monogr. 2003, 73, 69–86.
- Visser, M.E.; Both, Ch. Shifts in phenology due to global climate change: the need for a yardstick. P. Roy. Soc. B-Biol. Sci. 2005, 272, 2561–2569.
- Perry, D.A.; Borchers, J.G.; Borchers, S.L.; Amaranthus, M.P. Species migrations and ecosystem stability during climate change: the belowground connection. Conserv. Biol. 1990, 4, 266–274.
- Hampe, A.; Petit, R.J. Conserving biodiversity under climate change: the rear edge matters. Ecol. Lett. 2005, 8(5), 461–467. [CrossRef]
- Brooker, R.W.; Travis, J.M.J.; Clark, E.J.; Dytham, C. Modelling species’ range shifts in a changing climate: The impacts of biotic interactions, dispersal distance and the rate of climate change. J. Theor. Biol. 2007, 245, 59–65.
- Gao, J.-G.; Zhang, Y.-L.; Liu, L.-S.; Wang, Z.-F. Climate change as the major driver of alpine grasslands expansion and contraction: A case study in the Mt. Qomolangma (Everest) National Nature Preserve, southern Tibetan Plateau. Quatern. Int. 2014, 336, 108–116.
- Harley, C.D.G.; Paine, R.T. Contingencies and compounded rare perturbations dictate sudden distributional shifts during periods of gradual climate change. PNAS 2009, 106, 11172–11176.
- Hill, J.K.; Thomas, C.D.; Fox, R.; Tefler, M.G.; Willis, S.G.; Asher, J.; Huntley, B. Responses of butterflies to twentieth century climate warming: implications for future ranges. P. Roy. Soc. B-Biol. Sci. 2002, 269, 2163–2171. [CrossRef]
- Pelini, S.L.; Dzurisin, J.D.K.; Prior, K.M.; Williams, C.M.; Marsico, T.D.; Sinclair, B.J.; Hellmann, J.J. Translocation experiments with butterflies reveal limits to enhancement of poleward populations under climate change. PNAS 2009, 106, 11160–11165.
- Parolo, G.; Rossi, G. Upward migration of vascular plants following a climate warming trend in the Alps. Basic Appl. Biol. 2008, 9, 100–107.
- Best, A.S.; Johst, K.; Münkemüller, T.; Travis, J.M.J. Which species will successfully track climate change? The influence of intraspecific competition and density dependent dispersal on range shifting dynamics. Oikos 2007, 116, 1531–1539.
- Schmidt, G.; Archer, D. Too much of a bad thing. Nature 2009, 458, 1117–1118.
- Schneider, S.H. The worst-case scenario. Nature 2009, 458, 1104–1105.
- IPCC. Climate Change 2007: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press: Cambridge, UK, 2007; 976pp.
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–42.
- Nogués-Bravo, D.; Araujo, M.B.; Errea, M.P.; Martinez-Rica, J.P. Exposure of global mountain systems to climate warming during the 21st Century. Global. Environ. Change 2007, 17, 420–428.
- Colwell, R.K.; Brehm, G.; Cardelus, C.L.; Gilman, A.C.; Longino, J.T. Global warming, elevational range shifts, and lowland biotic attrition in the wet tropics. Science 2008, 322, 258–261.
- Hoegh-Guldberg, O.; Hughes, L.; Mcintyre, S.; Lindenmayer D.B.; Parmesan, C.; Possingham, H.P.; Thomas, C.D. Assisted colonization and rapid climate change. Science 2008, 321, 345–346.
- Hannah, L. Protected areas and climate change. Ann. NY Acad. Sci. 2008, 1134, 201–212.
- Dormann, C.F. Promising the future? Global change projections of species distributions. Basic Appl. Ecol. 2007, 8, 387–397.
- McCarty, J.P. Ecological Consequences of Recent Climate Change. Conserv. Biol. 2001, 15, 320–331.
- Waters, C.N.; Zalasiewicz, J.; Summerhayes, C.; Barnosky, A.D.; Poirier, C.; Ga1uszka, A.; Cearreta, A.; Edgeworth, M.; Ellis, E.C.; Ellis, M.; et al. The Anthropocene is functionally and stratigraphically distinct from the Holocene. Science 2016, 351, aad2622.
- Aedo, C.; Medina, L.; Barberá, P.; Fernández-Albert, M. Extinctions of vascular plants in Spain. Nord. J. Bot. 2015, 33, 83–100.
- Watson, J.E.M.; Venter, O.; Lee, J.; Jones, K.R.; Robinson, J.G.; Possingham, H.P.; Allan, J.R. Protect the last of the wild. Nature 2018, 563, 27–30. [CrossRef]
- Maxwell, S.L.; Fuller, R.A.; Brooks, T.M.; Watson, J.E.M. Biodiversity: The ravages of guns, nets and bulldozers. Nature 2016, 536, 143–145. [CrossRef]
- Hooper, D.U., Adair, E.C., Cardinale, B.J.; Byrnes, J.E.K.; Hungate, B.A.; Matulich, K.L.; Gonzales, A.; Duffy, J.E.; Gamfeldt, L.; O´Connor, M.I. A global synthesis reveals biodiversity loss as a major driver of ecosystem change. Nature 2012, 486, 105–108.
- Le Roux, J.J.; Hui, C.; Castillo, M.L.; Iriondo, J.M.; Keet, J.-H.; Khapugin, A.A.; Médail, F.; Rejmánek, M.; Theron, G.; Yannelli, F.A.; et al. Recent anthropogenic plant extinctions differ in biodiversity hotspots and coldspots. Curr. Biol. 2019, 29, 2912–2918. [CrossRef]
- Van Vuuren, D.P.; Sala, O.E.; Pereira, H.M. The future of vascular plant diversity under four global scenarios. Ecol. Soc. 2006, 11(2), 25. [CrossRef]
- Di Marco, M.; Harwood, T.D.; Hoskins, A.J.; Ware, C.; Hill, S.L.L.; Ferrier, S. Projecting impacts of global climate and land use scenarios on plant biodiversity using compositional-turnover modelling. Global Change Biol. 2019, 25(8), 2763–2778. [CrossRef]
- Knoke, T.; Bendix, J.; Pohle, P.; Hamer, U.; Hildebrandt, P.; Roos, K.; Gerique, A.; Sandoval, M.L.; Breuer, L.; Tischer, A.; et al. Afforestation or intense pasturing improve the ecological and economic value of abandoned tropical farmlands. Nat. Commun. 2014, 5(1), 5612. [CrossRef]
- Maxwell, S.L.; Cazalis, V.; Dudley, N.; Hoffmann, M.; Rodrigues, A.S.L.; Stolton, S.; Visconti, P.; Woodley, S.; Kingston, N.; Lewis, E. Area-based conservation in the twenty-first century. Nature 2020, 586, 217–227. [CrossRef]
- Winkler, K.; Fuchs, R.; Rounsevell, M.; Herold, M. Global land use changes are four times greater than previously estimated. Nat. Commun. 2021, 12, 2501. [CrossRef]
- Burns, F.; Eaton, M.A.; Barlow, K.E.; Beckmann, B.C.; Brereton, T.; Brooks, D.R.; Brown, P.M.J.; Fulaij, N.A.; Gent, T.; Henderson, I.; et al. Agricultural management and climatic change are the major drivers of biodiversity change in the UK. PloS ONE 2016, 11, e0151595. [CrossRef]
- Levine, J.M.; Rees, M. Effects of temporal variability on rare plant persistence in annual systems. Am. Nat. 2004, 164, 350–363.
- Devi, V.; Fulekar, M.H.; Charles, B.; Reddy, C.S.; Pathak, B. Predicting the habitat suitability and species richness of plants of Great Himalayan National Park under different climate change scenarios. Environ. Monit. Assess. 2024, 196, 1136. [CrossRef]
- Willard, D.A.; Bernhardt, C.E.; Holmes, C.W.; Landcare, B.; Marot, M. Response of everglades tree islands to environmental change. Ecol. Monogr. 2006, 76, 565–583.
- Rejmánek, M.; Richardson, D.M.; Pyšek, P. Plant invasions and invasibility of plant communities. In Vegetation Ecology, 2nd Edition; van der Maarel, E., Franklin, J., Eds.; John Wiley & Sons: Chichester, UK, 2013; pp. 387–424.
- Pyšek, P.; Richardson, D.M.; Rejmánek, M.; Webster, G.L.; Williamson, M.; Kirschner, J. Alien plants in checklists and floras: towards better communication between taxonomists and ecologists. Taxon 2004, 53, 131–143.
- Tye, A. Can we infer island introduction and naturalization rates from inventory data? Evidence from introduced plants in Galapagos. Biol. Invasions 2006, 8, 201–215.
- Pimentel, D.; McNair, S.; Janecka, J.; Wightman, J.; Simmonds, C.; O'Connell, C.; Wong, E.; Russel, L.; Zern, J.; Aquino, T.; et al. Economic and environmental threats of alien plant, animal, and microbe invasions. Agric. Ecosyst. Environ. 2001, 84, 1–20.
- Traveset, A.; Richardson, D.M. Biological invasions as disruptors of plant reproductive mutualisms. Trends Ecol. Evol. 2006, 21, 208–216.
- Bjerknes, A.L.; Totland, Ø.; Hegland, S.J.; Nielsen, A. Do alien plant invasions really affect pollination success in native plant species? Biol. Conserv. 2007, 138, 1–12.
- Wardle, D.A.; Nicholson, K.S.; Ahmed, M.; Rahman, A. Interference effects of the invasive plant Carduus nutans L. against the nitrogen fixation ability of Trifolium repens L. Plant Soil 1994, 163, 287–297.
- Weihe, P.E.; Neely, R.K. The effects of shading on competition between Purple Loosestrife and Broadleaved Cattail. Aquat. Bot. 1997, 59, 127–138.
- Mack, R.N.; D’Antonio, C.M. Impacts of biological invasions on disturbance regimes. Trends Ecol. Evol. 1998, 13, 195–198.
- Chittka, L.; Schürkens, S. Metrology: Successful invasion of a floral market. Nature 2001, 411, 653.
- Brown, B.J.; Mitchell, R.J.; Graham, S.A. Competition for pollination between an invasive species (Purple Loosestrife) and a native congener. Ecology 2002, 83, 2328–2336.
- Moragues, E.; Traveset, A. Effect of Carpobrotus spp. on the pollination success of native plant species of the Balearic Islands. Biol. Conserv. 2005, 122, 611–619.
- Totland, Ø.; Nielsen, A.; Bjerknes, A.L.; Ohlson, M. Effects of an exotic plant and habitat disturbance on pollinator visitation and reproduction in a boreal forest herb. Am. J. Bot. 2006, 93, 868–873.
- Wittenberg, R.; Cock, M.J.W. Invasive alien species: a toolkit for best prevention and management practices. CABI: Wallingford, UK, 2001.
- McDowell, W.G.; Byers, J.E. High abundance of an invasive species gives it an outsized ecological role. Freshw. Biol. 2019, 64, 577e586.
- Lodge, D.M.; Williams, S.; MacIsaac, H.J.; Hayes, K.R.; Leung, B.; Reichard, S.; Mack, R.N.; Moyle, P.B.; Smith, M.; Andow, D.A.; et al. Biological invasions: recommendations for U.S. Policy and management. Ecol. Appl. 2006, 16, 2035e2054.
- May, R.M.; Lawton, J.H.; Stork, N.E. Assessing extinction rates. In Extinction rates; Lawton, H., May, R.M., Eds.; Oxford University Press: Oxford, UK, 1995; pp 1–24.
- Chytrý, M.; Pyšek, P.; Tichý, L.; Knollová, I.; Danihelka, J. Invasions by alien plants in the Czech Republic: a quantitative assessment across habitats. Preslia 2005, 77, 339–354.
- Chytrý, M.; Jarošík, V.; Pyšek, P.; Hájek, O.; Knollová, I.; Tichý, L.; Danihelka, J. Separating habitat invasibility by alien plants from the actual level of invasion. Ecology 2008a, 89, 1541–1553.
- Chytrý, M.; Maskell, L.; Pino, J.; Pyšek, P.; Vilà, M.; Font, X.; Smart, S. Habitat invasions by alien plants: a quantitative comparison between Mediterranean, subcontinental and oceanic regions of Europe. J. Appl. Ecol. 2008b, 45, 448–458.
- Hulme, P.E.; Brundu, G.; Camarda, I.; Dalias, P.; Llambdon, P.; Lloret, F.; Medail, F.; Moragues, E.; Suehs, C.; Traveset, A.; et al. Assessing the risks of alien plant invasions on Mediterranean islands. In Plant invasions? Human perception, ecological impacts and management; Tokarska-Guzik, B., Brundu, G., Brock, J.H., Child, L.E., Pyšek, P., Daehler, C., Eds.; Backhuys: Leiden, The Netherlands, 2008; pp 39–56.
- Davidson, A.M.; Jennions, M.; Nicotra, A.B. Do invasive species show higher phenotypic plasticity than native species and, if so, is it adaptive? A meta-analysis. Ecol. Lett. 2011, 14, 419–431.
- Godoy, O.; Valladares, F.; Castro-Díez, P. Multispecies comparison reveals that invasive and native plants differ in their traits but not in their plasticity. Funct. Ecol. 2011, 25, 1248–1259.
- Van Kleunen, M.; Schlaepfer, D.R.; Glaettli, M.; Fischer, M. Preadapted for invasiveness: do species traits or their plastic response to shading differ between invasive and non-invasive plant species in their native range? J. Biogeogr. 2011, 38, 1294–1304.
- Hellmann, J.J.; Byers, J.E.; Bierwagen, B.G.; Dukes, J.S. Five potential consequences of climate change for invasive species. Conserv. Biol. 2008, 22, 534–543.
- Diez, J.M.; D’Antonio, C.M.; Dukes, J.S.; Grosholz, E.D.; Olden, J.D.; Sorte, C.J.B.; Blumenthal, D.M.; Bradley, B.A.; Early, R.; Ibáñez, I.; et al. Will extreme climatic events facilitate biological invasions? Front. Ecol. Environ. 2012, 10, 249–257.
- Sorte, C.J.B.; Ibáñez, I.; Blumenthal, D.M.; Molinari, N.A.; Miller, L.P.; Grosholz, E.D.; Diez, J.M.; D’Antonio, C.M.; Olden, J.D.; Jones, S.J.; et al. Poised to prosper? A cross-system comparison of climate change effects on native and nonnative species performance. Ecol. Lett. 2013, 16, 261–270.
- Liu, Y.; Oduor, A.M.O.; Zhang, Z.; Manea, A.; Tooth, I.M.; Leishman, M.R.; Xu, X.; Van Kleunen, M. Do invasive alien plants benefit more from global environmental change than native plants? Global Change Biol. 2017, 23, 3363–3370.
- Martín-Forés, I.; Bywaters, S.L.; Sparrow, B.; Guerin, G.R. Simultaneous effect of habitat remnancy, exotic species, and anthropogenic disturbance on orchid diversity in South Australia. Conserv. Sci. Pract. 2022, 4, e12652.
- Axmanová, I.; Kalusová, V.; Danihelka, J.; Dengler, J.; Pergl, J.; Pyšek, P.; Večeřa, M.; Attorre, F.; Biurrun, I.; Boch, S.; et al. Neophyte invasions in European grasslands. J. Veget. Sci. 2021, 32, 1–17. [CrossRef]
- Scramoncin, L.; Gerdol, R.; Brancaleoni, L. How effective is environmental protection for ensuring the vitality of wild orchid species? A case study of a protected area in Italy. Plants 2024, 13, 610. [CrossRef]
- Anthony, M.A.; Stinson, K.A.; Moore, J.A.; Frey, S.D. Plant invasion impacts on fungal community structure and function depend on soil warming and nitrogen enrichment. Oecologia 2020, 194, 659–672.
- Morales, C.L.; Traveset, A. A meta-analysis of impacts of alien vs. native plants on pollinator visitation and reproductive success of co-flowering native plants. Ecol. Lett. 2009, 12, 716–728.
- Pyšek, P.; Jarošík, V.; Chytrý, M.; Danihelka, J.; Kühn, I.; Pergl, J.; Tichý, L.; Biesmeijer, J.; Ellis, W.N.; Kunin, W.; et al. Successful invaders co-opt pollinators of native flora and accumulate insect pollinators with increasing residence time. Ecol. Monogr. 2011, 81(2), 277–293.
- Steffelová, M.; Traxmandlová, I.; Štípková, Z.; Kindlmann, P. Pollination strategies of deceptive orchids – a review. Eur. J. Environ. Sci. 2023, 13, 110–116. [CrossRef]
- Florens, F.B.V.; Baider, C.; Martin, G.M.; Seegoolam, N.B.; Zmanay, Z.; Strasberg, D. Invasive alien plants progress to dominate protected and best-preserved wet forests of an oceanic island. J. Nat. Conserv. 2016, 34, 93–100.
- Florens, F.B.V.; Baider, C.; Seegoolam, N.B.; Zmanay, Z.; Strasberg, D. Long-term declines of native trees in an oceanic island’s forests invaded by alien plants. Appl. Veg. Sci. 2017, 20, 94–105.
- Baider, C.; Florens, F.B.V. Current decline of the ‘Dodo-tree’: a case of broken-down interactions with extinct species or the result of new interactions with alien invaders? In Emerging threats to tropical forests; Laurance, W., Peres, C., Eds.; Chicago University Press: Chicago, USA, 2006; pp. 199–214.
- Ackerman, J.D. Invasive orchids: weeds we hate to love? Lankesteriana 2007, 7, 19–21.
- Konowalik, K.; Kolanowska, M. Climatic niche shift and possible future spread of the invasive South African Orchid Disa bracteata in Australia and adjacent areas. PeerJ 2018, 6, e6107.
- Kolanowska, M.; Rewicz, A.; Ackerman, J.D. Climate change will likely facilitate invasion of Asian orchid Eulophia graminea into new areas. Biol Invasions 2024, 26, 1969–1983.
- Recart, W.; Ackerman, J.D.; Cuevas, A.A. There goes the neighbourhood: apparent competition between invasive and native orchids mediated by a specialist florivorous weevil. Biol. Invasions 2013, 15, 283–293. [CrossRef]
Figure 1.
Checkerspot (Euphydryas editha bayensis), one of many species studied for its vulnerability to effects of climate change.
Figure 1.
Checkerspot (Euphydryas editha bayensis), one of many species studied for its vulnerability to effects of climate change.

Figure 2.
Changing composition of forests, represented by changes in distribution of six forest classes in eastern United States due to climate change.
Figure 2.
Changing composition of forests, represented by changes in distribution of six forest classes in eastern United States due to climate change.
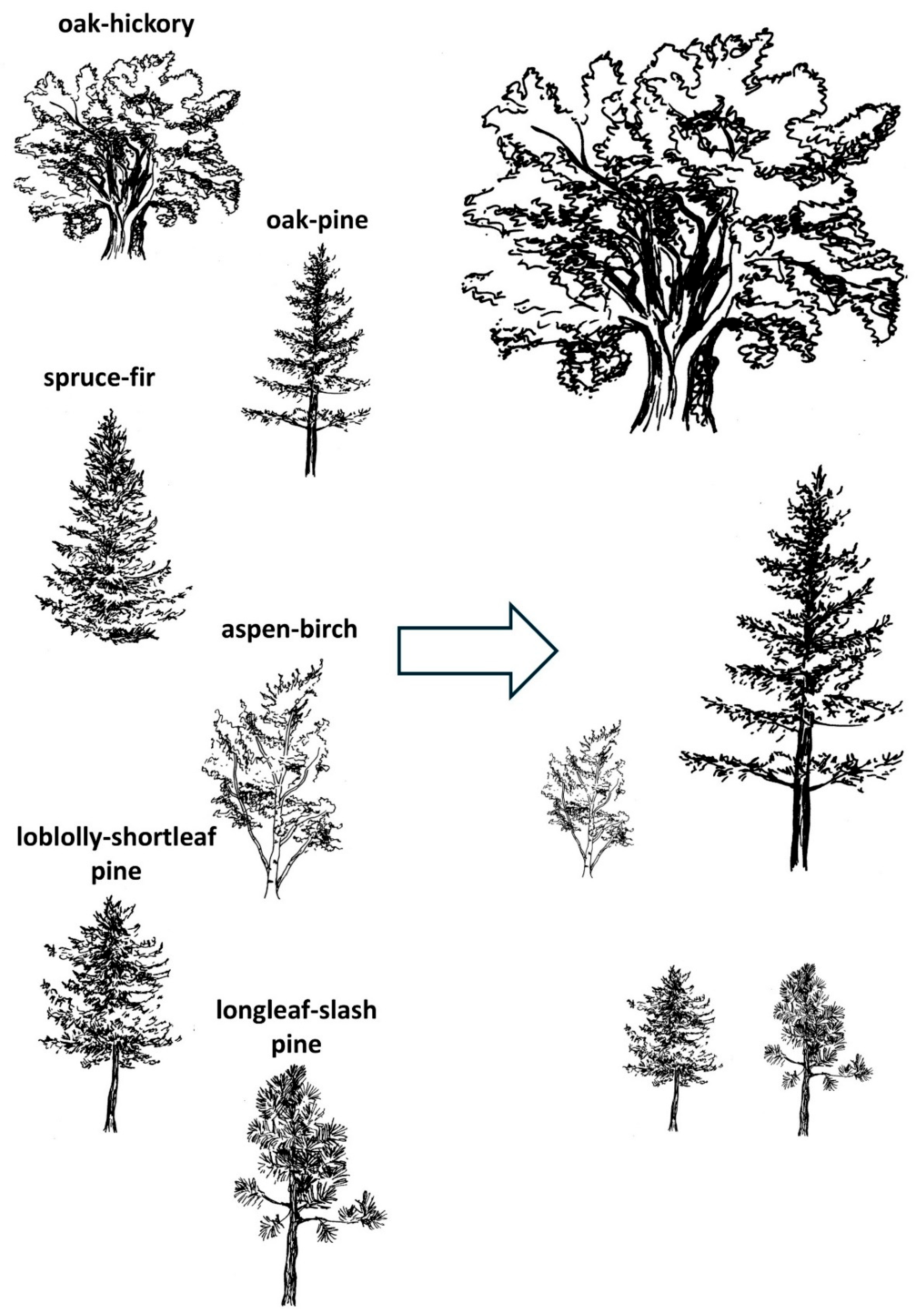
Figure 3.
The competitive relationship between pied flycatcher (Ficedula hypoleuca) and great tit (Parus major) may be one example of the effect of climate change on changing balance of their respective populations.
Figure 3.
The competitive relationship between pied flycatcher (Ficedula hypoleuca) and great tit (Parus major) may be one example of the effect of climate change on changing balance of their respective populations.
Figure 4.
Blotched blue-tongued skink (Tiliqua nigroglutea), one of the Australian skinks which the findings in Kearney et al. [102] refer to.
Figure 4.
Blotched blue-tongued skink (Tiliqua nigroglutea), one of the Australian skinks which the findings in Kearney et al. [102] refer to.
Figure 5.
Highbush blueberry (Vaccinium corymbosum) is one of the species which are particularly responsive to changes in climate.
Figure 5.
Highbush blueberry (Vaccinium corymbosum) is one of the species which are particularly responsive to changes in climate.
Figure 6.
Categories of diaspore morphology of plants studied by Parolo and Rossi. a) diaspores with no apparent structures for dispersal (as in Saxifraga sp.); b) diaspores with wings, fringes, hooks or calices (as those on Trisetum spicatum seeds); c) parachutists with plumes, pappi or similar structures (Leontodon sp.); d) spores.
Figure 6.
Categories of diaspore morphology of plants studied by Parolo and Rossi. a) diaspores with no apparent structures for dispersal (as in Saxifraga sp.); b) diaspores with wings, fringes, hooks or calices (as those on Trisetum spicatum seeds); c) parachutists with plumes, pappi or similar structures (Leontodon sp.); d) spores.
Figure 7.
Five profoundly impactful mechanisms by which the world’s biodiversity is threatened. a) land use changes (e.g. agriculture); b) direct overexploitation (e.g. fishery); c) pollution (e.g. carcinogenic PFAS); d) climate change; e) invasive alien species.
Figure 7.
Five profoundly impactful mechanisms by which the world’s biodiversity is threatened. a) land use changes (e.g. agriculture); b) direct overexploitation (e.g. fishery); c) pollution (e.g. carcinogenic PFAS); d) climate change; e) invasive alien species.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



