Submitted:
12 September 2025
Posted:
15 September 2025
You are already at the latest version
Abstract
Idiopathic pulmonary fibrosis (IPF) is a chronic, rare and fatal disease that is the conse-quence of aberrant remodeling and defective repair mechanisms within the lung cul-minating in the loss of alveolar integrity. Although significant progress has been made in understanding the pathogenesis, it would be crucial to identify biomarkers for diagnosis, prognosis and prediction of therapy response to improve management of this challenging and debilitating disease. Omics technologies have profoundly advanced the under-standing of disease mechanisms, presenting considerable potential for the identification of clinically relevant biomarkers. To date, specific molecular pathways have been impli-cated in the onset and progression of idiopathic pulmonary fibrosis, including abnormal wounding, fibroblast proliferation, inflammation, deposition of extracellular matrix, oxidative stress, endoplasmic reticulum stress and coagulation system. This review highlights the role of proteomics in identifying key biomarkers for IPF, focusing on their clinical relevance, including diagnosis, prognosis, disease progression and the identifi-cation of new therapeutic options, in light of the most recent technological advancements in mass spectrometry.
Keywords:
mass spectrometry
; IPF
; biomarkers
; proteomic analysis
; machine learning
1. Introduction
Idiopathic pulmonary fibrosis (IPF) represents the most common form of idiopathic interstitial pneumonia (IIP) and is characterized by the progressive distortion of the lung parenchyma structure, aberrant extracellular matrix (ECM) deposition and irreversible scarring. It is a chronic and rare disease occurring in adults over 60 years old, with a median survival of 3 years after diagnosis, which is diagnosed according to multidisciplinary clinicopathological criteria, tipically displaying the so called usual interstitial pneumonia (UIP) pattern [1].
Although the cause of IPF onset is unknown by definition, several risk factors have been identified that might increase the likelihood of developing this disease [2], [3] such as environmental exposure, radiation therapy, occupational exposures (e.g., asbestosis), smoking cigarettes and exposure to allergens.
Prognosis of IPF might be unpredictable, since patients can be characterized by progressive respiratory failure, often precipitated by acute events, namely, acute exacerbations (AE)[4] Despite the introduction of drugs such as Pirfenidone and Nintedanib, which can slow respiratory functional decline, IPF has still a poor survival times (20-40% after 5-year from diagnosis).
One of the most crucial clinical needs in the diagnosis and prognosis of this disease is the search for biomarkers capable of detecting disease onset, as well as assessing progression in terms of stage and rate [5]. Nowadays, the introduction of different omics techniques which refer to high-throughput technologies that provide comprehensive data at different levels and with unprecedented high resolution, can support both biomarker discovery and the identification of molecular mechanisms of disease onset and progression [6]. Different omics techniques involve Genomics aimed to analyze the entire genome (e.g., whole genome sequencing, SNP genotyping), Transcriptomics with the study of RNA transcripts (e.g., RNA-Seq), Proteomics based on large-scale study of proteins (e.g., mass spectrometry, 2D gel electrophoresis), Metabolomics with the analysis of small-molecule metabolites, Epigenomics as the study of epigenetic modifications (e.g., DNA methylation arrays, ChIP-Seq), Glycomics with the study of glycan structures and functions, Microbiomics, which studies the microbial communities (e.g., 16S rRNA sequencing) and pharmacogenomics, which studies the individual responses to drugs. Although genomics and transcriptomics have been considered the most used techniques of the past 20 years for the molecular profiling studies, they might have significant limitations since they do not analyze changes in posttranslational modifications (PTMs), including phosphorylation and protein degradation [7]. Thus, mass spectrometry (MS)-based proteomics through the identification and quantification of amino-acid sequences allows the analysis of protein expression and modifications under different conditions that mirror the complexity at protein level [8]. Then, mass spectrometry (MS)-based proteomics represents a crucial integrative analysis of multi-omics data that will allow the discovery and functional studies of complex human diseases. This approach has enabled to enter the era of so-called precision medicine, thus centering the decision-making process to the molecular profiling of patients.
This Review aims to provide a broad overview of the current MS-based proteomics workflow and techniques that might be used to understand the mechanism underlying the onset, progression, and and AE of pulmonary fibrosis, to support personalized diagnostics and treatment. Here, we describe applications in single-cell and spatial analysis, as well as in human body fluid profiling and finally, we discuss the potential transition of proteomics from basic research to clinical practice.
2. Diagnostic, Prognostic and therapeutic challenges of IPF
IPF is diagnosed by clinicopathological criteria, including the radiographic and/or histopatological hallmark pattern of UIP [9]. Histologically, the UIP pattern is characterized by a heterogeneous phenotype with the presence of fibroblastic foci (FF) interspersed with regions of relatively preserved parenchyma, as well as areas of inflammation and honeycombing. The High-resolution computed tomography (HRCT) of IPF patients shows a peripheral, subpleural and basal distribution pattern characterized by reticular opacities, honeycombing and architectural distortion [10]. The definition of IPF requires the exclusion of other forms of interstitial pneumonia including other idiopathic interstitial pneumonias and ILD associated with environmental exposures, medications, or systemic diseases. Since the histological pattern is not exclusively associated with IPF but with other ILDs, a proven diagnosis of IPF may represent a challenge for physicians.
The pathogenesis of IPF is complex and far from being fully understood. Much evidence suggests that IPF is a consequence of multiple interacting genetic, and environmental risk factors such as cigarette smoking, metal and silica dust, or microbial agents, leading to repetitive local micro-injuries on alveolar epithelium and vascular endothelium, that cause aberrant epithelial–fibroblast functional activity, excessive ECM deposition, aberrant tissue repair process. These pathological mechanisms involve the activation, proliferation, and differentiation of fibroblasts into myofibroblasts and the secretion of pro-fibrotic, coagulant, and inflammatory cytokines, ultimately resulting in lung architectural distortion and progressive decline in respiratory function [11], [12]. In addition, some genetic variants have also been associated with IPF onset and progression such as variants in the mucin 5B (MUC5B) gene, which play a role in bronchoalveolar epithelial function, and in the TERT and TERC genes, which are involved in the telomere length maintenance [7]. Nevertheless, IPF remains a fatal disease, and the identification of different and new biomarkers indicative of progression represents an unmet clinical need that has to be addressed.
The natural course of IPF is variable and survival can be influenced by multiple clinical factors, including frequency and severity of AE, comorbidities (e.g., lung cancer, emphysema, pulmonary hypertension), and the rate of functional decline [4].
Therapeutic options remain a clinical challenge. Two antifibrotic agents, Pirfenidone and Nintedanib, have demonstrated efficacy in slowing FVC decline and disease progression, although neither has been shown to significantly reduce mortality.
Pirfenidone has antifibrotic, anti-inflammatory, and antioxidant effects, primarily through inhibition of transforming growth factor-beta (TGF-β) signalling. Common adverse effects include photosensitivity, gastrointestinal disturbances, and nausea. Nintedanib, a non-selective tyrosine kinase inhibitor targeting PDGF, VEGF, and FGF receptors, is also associated with gastrointestinal side effects, particularly diarrhea, nausea, and vomiting [13]. As a therapeutic strategy, the two antifibrotic drugs Pirfenidone and Nintedanib can slow the decline in the respiratory functional decline of IPF patients, and according to both the real-world data and randomized controlled trials show that mortality rates and survival times in IPF are highly heterogenous [14]. Emerging therapies offer potential hope. Recently, BI 1015550 (Nerandomilast), an oral selective phosphodiesterase 4B (PDE4B) inhibitor with antifibrotic and immunomodulatory properties, demonstrated efficacy in a Phase 2 trial by stabilizing lung function over a 12 weeks period. Furthermore, a Phase 3 randomized, double-blind, placebo-controlled trial reported that Nerandomilast significantly reduced the decline in FVC over 52 weeks compared to placebo [15].
Supportive measures, including long-term oxygen therapy in cases of hypoxemia and pulmonary rehabilitation in functionally disabled individuals, are essential components of comprehensive care.
3. Identification of the optimal biomarkers through mass spectrometry
3.1. Selection criteria of biomarkers and study design
The identification of sensitive and specific biomarkers should be based on a comprehensive comparative analysis of biological samples drawn from diagnosed cases and appropriately matched healthy controls. A proper comparison is essential to distinguish pathological changes from normal physiological variation, which depends on factors such as age, sex, lifestyle, genetic background, and environmental exposure [16], [17].
Typically, the biomarkers should be associated in a unique and definite manner with the disease under consideration, to minimize false positives and, in turn, enhance the overall diagnostic precision. An ideal biomarker should show a dynamic response both to the disease progression and therapeutic intervention [18]. In general, various biofluids and tissues, including plasma, serum, urine, bronchoalveolar lavage fluid, and tissue biopsies, provide unique information regarding biological processes that lead to disease onset and progression but differ in accessibility, invasiveness, and suitability for high-throughput analysis. For ensuring biomarker assay validity, preanalytical conditions such as sample collection process, storage, and processing must be strictly controlled to avoid degradation, contamination, or other undesirable modifications [19].
3.2. Protein analysis through different type of mass spectrometry
A robust proteomic study must be based on the accurate definition of study cohorts and meticulous design of the experimental workflow.
The selection of an appropriate analytical strategy depends on multiple factors, including the biochemical properties of the target protein, the depth of proteome coverage required, and the need for absolute versus relative quantification. Different analysis platforms are being used in mass spectrometry research in order to quantitatively determine and identify specific proteins or organic compounds. In particular, gas chromatography-mass spectrometry (GC-MS) and liquid chromatography-mass spectrometry (LC-MS) are two core techniques employed in proteomics on a daily basis to identify and distinguish complex mixtures of proteins [20]. Although both methods are highly sensitive and specific, they play different roles in proteomics. For example, GC-MS is particularly useful for analyzing volatile and thermally stable molecules, and while it is less commonly applied in large scale proteomics, is has been employed in complementary analyses [21] such as identifying metabolic changes through volatile organic compounds (VOCs) in exhaled breath. Plantier et al. used GC-MS to analyze VOC profiles in IPF patients, patients with different ILDs, and healthy controls. Their study identified significantly higher concentrations of acetone and other VOCs in IPF patients, suggesting metabolic alterations associated to oxidative stress and inflammation. The results demonstrated that specific VOC patterns could differentiate IPF from other respiratory diseases, highlighting their potential role as early biomarkers for disease detection and progression monitoring. These findings support the use of VOC analysis as a non-invasive diagnostic tool for IPF [22]. LC-MS is more suitable for analyzing a wider range of biomolecules, including peptides and intact proteins [23]. In proteomics, protein identification can be achieved through a bottom-up approach (Figure 1A), where peptides derived from enzymatic digestion are analyzed, or by a top-down approach (Figure 1B), where intact proteins are examined directly. This provides more detailed sequence coverage and information about post-translational modifications (PTMs) [24] [25]. Top-down proteomics typically involves separating proteins by one- or two-dimensional liquid chromatography, followed by identification using high-resolution mass spectrometry, facilitating the detection of proteoforms resulting from PTMs, genetic mutations, RNA and histone splicing, and other multisite heterogeneities.
The main limitation of the top-down proteomic approach for wider application is related to the lower solubility of intact proteins compared to their peptide counterparts [26]. In contrast, bottom-up proteomic approaches are widely applied because peptides are easier to separate and analyze by LC coupled to tandem mass spectrometry (MS/MS). Since bottom-up proteomics involves protein digestion, it handles higher sample complexity, although this complexity may require various chromatographic separations before final analysis. With bottom-up analysis, a sequence coverage equivalent to 100% of the original protein can only rarely be achieved. This may lead to incomplete or incorrect evaluation of protein isoforms and combinatorial PTMs. The data obtained from the two working approaches top-down and bottom-up are complementary and interdependent [27]. The combined approach enabled a detailed analysis of both intact proteins and peptide fragments, enhancing the identification of PTMs and isoforms.
Moreover, when the goal is to achieve a more comprehensive understanding of protein structure and metabolic activity, nuclear magnetic resonance (NMR) spectroscopy becomes an invaluable tool. NMR provides a powerful and non-destructive approach for structural elucidation and metabolic profiling. It enables detailed insights into molecular conformations and dynamic interactions without requiring extensive sample preparation.
In quantitative proteomics, isobaric tags for relative and absolute quantitation (iTRAQ) enable simultaneous comparison of multiple biological samples by labeling peptides with distinct isotopic tags, thereby allowing relative abundance measurements across experimental conditions [28]. In addition to MS methods, gel electrophoresis techniques such as two-dimensional gel electrophoresis (2D) and differential gel electrophoresis (DIGE) remain valuable for assessing differential protein expression patterns. These methods are able to resolve complex protein mixtures based on isoelectric point and molecular weight, with DIGE offering higher quantitative precision via fluorescent labeling, enabling direct comparison of protein samples on the same gel [29].
Furthermore, high-resolution mass spectrometry (HRMS) can employ electrospray ionization (ESI) or matrix-assisted laser desorption ionization (MALDI). ESI is particularly suitable for analyzing proteins in solution due to its ability to generate high-quality ionized peptides with better fragmentation, while MALDI allows for rapid analysis of complex tissue samples, making it a valuable tool in pulmonary research [30]. MALDI, as a diagnostic and prognostic tool, might enable direct spatial localization of ECM alterations in different tissues. The ability to detect molecular changes at a histopathological level reinforces the importance of mass spectrometry-based approaches in understanding the mechanisms underlying progressive fibrosis and in identifying potential biomarkers for disease stratification and therapeutic targeting.
Thus, the growing spread of MS technique has changed the method of analysis in the proteomic field, increasing resolution, accuracy, and sensitivity. Combined use of advanced proteomic technologies, high-resolution mass spectrometry, quantitative proteomics, and bioinformatics analysis has enabled the characterization of the complex protein networks involved in rare lung diseases like IPF from different biological samples like blood serum and plasma, bronchoalveolar lavage fluid (BALF), fresh lung tissue, cells, and Formalin-fixed paraffin-embedded tissue (FFPE). In the next sections, we will summarize the biomarkers of IPF onset and progression identified through mass spectrometry from different biological samples.
4. Mass spectrometry-based serum and plasma biomarkers in idiopathic pulmonary fibrosis
Since the diagnosis of IPF is often complex and invasive, identifying non-invasive markers could simplify the process and improve access to diagnosis for patients [31]. The identification of circulating biomarkers in the serum or plasma of patients affected by IPF offers noninvasive tools for diagnosis, outcome prediction, and treatment monitoring, aiming to prevent significant clinical and functional decline[32]. One of the most consistent and reproducible circulating biomarkers is matrix metalloproteinase-7 (MMP-7) [33]. MMP-7 belongs to the zinc-dependent endopeptidases and exerts a pivotal role in degrading ECM components, such as collagens, fibronectin, and laminin [34]. The key role of the protein in fibrosis has been demonstrated in MMP-7 knockout mice, revealing a strong resistance to developing a fibrotic phenotype following intratracheal injection of bleomycin [35]. The key proteolytic activity in lung fibrosis and the elevated levels of MMP-7 in the plasma of patients affected by IPF, suggest MMP-7 as a valid marker capable to discriminate between healthy and diseased individuals and, at the same time, to enable differential diagnosis between IPF and other lung diseases such as chronic obstructive pulmonary disease (COPD), sarcoidosis and hypersensitivity pneumonitis [36].
Like MMP-7, other matrix remodeling proteins (such as MMP-1 and MMP-10) have been investigated and proposed as markers of disease [37]. Collagen production and the abnormal ECM deposition, typical events in fibrosis, are also linked to CC-chemokine 18 (CCL18), a chemokine mediating chemotactic and immunoregulatory functions. Literature reports evidence of elevated serum levels of CCL18 in patients with pulmonary fibrosis and a relationship between serum concentrations and the level of pulmonary fibrotic activity. CCL18 is associated with organ failure and, as predictor of survival and disease progression, it is positioned as a prognostic marker for IPF [38], [39]. Furthermore, high levels of periostin, involved in the remodeling of the ECM and linked to pulmonary function loss, were found in the serum of IPF patients [40]. Once secreted by fibroblast, epithelial and endothelial cells, it interacts with integrin receptor after stimulation by IL-4, IL-13, and TGF-β, sustaining fibrosis [41], [42]. Several studies have demonstrated that IPF-affected patients exhibit elevated surfactant proteins SP-A and SP-D expression profile in comparison with healthy individuals [32]. The elevated levels of these peculiar proteins, involved in maintaining tissue homeostasis and immune response in the serum of affected patients help the differential diagnosis and prognosis prediction [39]. Krebs von den Lungen-6 (KL-6) is a high-molecular-weight mucin-like glycoprotein produced by damaged and regenerating alveolar type II cells in the lungs[43]. KL-6 was proposed as a diagnostic tool for interstitial lung diseases (ILDs) and as a predictor of response to antifibrotic therapy.
In recent years, thanks to advanced protein-based techniques, new emerging blood-based biomarker panels for IPF have been proposed. For example, the new proteomic platform slow-off rate-modified aptamer (SOMAmer) SOMAscan panel was used to analyze plasma isolated from 60 IPF affected patients in 80 weeks of follow-up. The study identified a panel of six analytes associated with the disease. Specifically, disease progression was associated with sub-threshold levels of soluble vascular endothelial growth factor receptor 2, ficolin-2, legumain, and cathepsin, along with elevated levels of inducible T cell co-stimulator and trypsin-3 [44].
In addition, iTRAQ, which is able to simultaneously identify and quantify the peak intensities of reporter ions using tandem mass spectrometry (MS/MS), revealed the presence of four analytes associated with IPF. In the cohort of patients enrolled in the study, elevated levels of CRP and fibrinogen-α, along with decreased levels of haptoglobin and kininogen-1 were observed compared to healthy individuals. These biomarkers, identified through proteomic studies, provide insights into IPF pathogenesis and hold potential for improving diagnosis, prognosis, and treatment monitoring. However, further validation is needed before they can be routinely used in clinical practice [45]. Furthermore, Oldham et al. analyzed plasma samples from IPF patients using quantitative proteomics, found significant alterations in the expression of specific plasma proteins in progressive IPF compared to stable cases. Key biomarkers included increased levels of serum amyloid A1 (SAA1), haptoglobin (HP), and hemopexin (HPX), which were involved in inflammatory response, oxidative stress, and ECM remodeling—processes critical in IPF pathogenesis [46]. Blood biomarkers offer potential for diagnosing and managing IPF, but challenges such as lack of specificity and the need for standardization persist. Using biomarker panels may improve accuracy by capturing multiple disease aspects. Long-term monitoring is crucial to assess their predictive value, while integration with advanced study techniques could further enhance diagnosis and monitoring.
5. Biomarkers of IPF identified in BALF fluid through proteomics
BALF has been increasingly used to identify disease-associated protein signatures in IPF, reflecting the lung microenvironment offering insights into the local immune response and tissue remodeling processes, through minimal invasiveness compared to surgical biopsy. Advances in proteomic technologies and MS have enabled the identification of multiple proteins in BALF that are differentially expressed in IPF patients compared to healthy individuals or those with other interstitial lung diseases. Elevated levels of mediators of pulmonary fibrosis (osteopontin, MMP7, CXCL7, CCL18), eosinophil- and neutrophil-derived proteins, and proteins associated with fibroblast foci have been observed in BALF from IPF patients using gel-free quantitative proteomics (HDMSE) and targeted multiple reaction monitoring (MRM). Additionally, this study describes, for the first time, the upregulation of the pro-fibrotic cytokine, CCL24, in BALF of IPF patients [47]. Furthermore, the comparison of BALF protein profiles of IPF patients, never-smoker healthy control (nonsmoker control) and smoker control subjects using comparative two-dimensional gel electrophoresis (2D-PAGE) analysis, revealed transcriptional factors NF-kB, PPARγ, and c-MYC as a group of functional hubs, indicating the principal pathways involved in the progression and pathogenesis of IPF [48]. Comparative 2D-PAGE and MALDI analysis uncovered overexpressed proteins in BALF of patients with IPF, such as S100A9 compared to BALF of patients with other fibrotic diseases the importance of proteomics in identifying biomarkers that might discriminate different types of fibrotic interstitial pneumonia [49]. Then, the study of BALF protein composition from patients with IPF, sarcoidosis and pulmonary fibrosis associated with systemic sclerosis (SSc) analyzed by 2-DE showed that there were quantitative rather than qualitative differences in the protein profile between the three diseases, where IPF was characterized by a greater abundance in low molecular weight proteins compared to sarcoidosis and SSc. Additionally, cytokine-focused proteomic study further highlighted elevated levels of IL-1, IL-6, IL-8, TNF-α, IL-17A/F, alongside increased cellular counts of neutrophils, macrophages, and eosinophils in BALF of IPF patients. Furthermore, macrophage migration inhibitory factor (MIF) along with p23 and Calgranulin B, two calcium binding proteins, were also significantly increased in IPF [50]. Finally, proteomic profiling of BALF conducted using label-free quantitative methods (LFQ), through unsupervised cluster analyses identified protein expression profiles that were then analyzed to predict survival in patients with IIP, reinforcing the prognostic relevance of BALF biomarkers [51].
In summary, a wide array of BALF-derived biomarkers—identified through high-throughput proteomics and MS techniques including HDMSE, 2D-PAGE, MALDI-ToF/ToF, LC-MS/MS, and MRM—has elucidated key proteins involved in inflammation, ECM remodeling, cell signaling, and immune modulation in IPF. These findings underscore the potential of BALF proteomics to inform disease monitoring, prognostication, and development of personalized therapeutic strategies—pending validation in larger, multicenter studies.
6. Biomarkers of IPF identified in lung tissues through proteomics
Over the last few years, various attempts have been made to understand the molecular mechanism of IPF. Among them, tissue-based proteomics has had a particular impact, giving a comprehensive overview of molecular pathways active in the onset and progression of the disease. Given the rarity of the disease and the challenges associated with obtaining fresh samples such as surgical biopsies or cryobiopsies, FFPE tissue specimens from pathology archives offer a reliable and accessible alternative for proteomic investigations and biomarker discovery[52]. However, formalin fixation and paraffin embedding can affect protein quality, limiting the efficiency of proteomic analysis [53]. Despite these difficulties with FFPE samples, they have been successfully used for advanced proteomic analysis. In a study by Samarelli et al., FFPE lung tissue samples from patients with IPF were analyzed through LC-MS/MS and label-free quantitative analysis, leading to the identification of differentially expressed proteins, including those involved in the ECM signaling pathways, focal adhesion and transforming growth factor β (TGF-β) signaling pathways strongly associated with IPF onset and progression, compared with non-fibrotic lung as control samples. Moreover, five proteins were significantly overexpressed in the lung of IPF patients with either advanced disease stage (Stage II) or impaired pulmonary function (FVC <75%, Diffusion Lung Carbon Monoxide DLCO <55%) compared to controls; these were lymphocyte cytosolic protein 1 (LCP1), peroxiredoxin-2 (PRDX2), transgelin 2 (TAGLN2), lumican (LUM) and mimecan (OGN) that might play a key role in the fibrogenic processes, representing potential biomarkers of disease progression [54]. Thus, the analysis of FFPE samples has the potential to identify key proteins that might be crucial for the IPF pathogenesis.
Several recent studies employed proteomic analysis on fresh or fresh-frozen lung tissue from IPF patients, identifying a more complex proteomic profiling compared to that from FFPE samples. Among these, iTRAQ labeling combined with LC-MS/MS to IPF fresh-frozen lung tissues revealed over 600 differentially expressed proteins, including both well-studied ECM components—such as COL1A1, SCGB1A1, TAGLN, CTSB, SERPINB3—and novel ECM-associated proteins such as LGALS7, ASPN, and HSP90AA1 and HSP90AB1 [55].
One large-scale quantitative proteomic study detected more than 1,500 differentially expressed proteins in IPF lungs compared to controls [56]. This study showed that although transcriptional signatures of IPF were mainly enriched in pathways associated with immune-mediated process and inflammatory responses, protein expression profiles of IPF lung tissues were related to production, deposition and remodeling of ECM. Intriguingly, two candidate proteins, BTNL9 crucial in the process of antigen presentation, and PLLP involved in epithelial cell development and differentiation, were down-regulated at the mRNA and protein levels in IPF lung tissue and bleomycin-induced mice, indicating that they have a protective effect by inhibiting ECM production and promoting wound repair in alveolar epithelial cells [56].
6.1. New insight in the biomarker’s identification with spatial proteomic
Spatial proteomics has emerged recently as a powerful approach for dissecting the molecular complexity of IPF by mapping protein expression within histologically distinct regions of lung tissue. In particular, the spatially resolved proteomic studies have contributed to the identification of niche-specific biomarkers in IPF, providing novel insights into disease progression and potential region-targeted therapies.
Laser capture microdissection combined with mass spectrometry (LCM-MS) has been used to isolate specific fibrotic niches—such as fibroblast foci, fibrotic alveoli, and scarred regions—from IPF lungs. A study by Herrera et al. [57] used LCM-MS to analyze FF, adjacent mature scar, and adjacent alveoli in 6 fibrotic (UIP/IPF) specimens plus 6 nonfibrotic alveolar specimens as controls, founding that among 3000 proteins identified, the fibrotic alveoli protein signature was characterized by immune deregulation. Furthermore, FF were positive for both transforming growth factor beta 1 TGFβ1 and TGFβ3, whereas the aberrant basaloid cell lining of FF was predominantly positive for TGFβ2. A recent integrative study combining spatial proteomics with single-cell RNA sequencing (scRNA-seq) revealed that fibroblast foci are enriched in profibrotic proteins such as LTBP1 and fibronectin (FN1), confirming their active role in TGF-β signaling and ECM remodeling [58]. Furthermore, spatial proteomics of the basement membrane from distal bronchioles of IPF patients compared to control patients revealed the downregulation of cell junctional proteins, upregulation of epithelial-mesenchymal transition factors and dysregulated basement membrane matrix that induced epithelial desquamation in IPF distal bronchioles [59]. Finally, the study from Griesser et al. applied quantitative proteomics to whole-lung homogenates from an AAV-DTR mouse model of acute epithelial lung injury that might lead to IPF in patients, comparing bulk tissue data with spatially resolved proteomes from epithelial regions from the same animals. Injury induced upregulation in pathways related to interferon response, proliferation, DNA replication, and ECM deposition. Conversely, proteins associated with epithelial markers (SP-A, SP-C, Scgb1a1), cilium assembly, lipid metabolism, and redox balance were downregulated. This study based on a mouse model showed that the comparison between two proteomic methods, revealed both extensive overlap and notable differences, highlighting that while bulk proteomics is broadly informative, region-specific analyses add valuable resolution [60].
Thus, the advancement of the proteomic techniques has allowed the employment of mass spectrometry techniques for a broad range of tissue samples from FFPE to fresh or fresh-frozen lung tissue, to integrate the complex landscape of biomarkers that might be associated to the progression of IPF, identified through BALF and blood.
7. Biomarkers of IPF identified in pulmonary cell line and primary cells from IPF patients
Proteome analysis reflects functional protein expression, post-translational modifications, and secretion dynamics—especially at cellular resolution levels. Mass spectrometry–based proteomic studies of IPF lung fibroblast cell lines, LL97A and LL29, have revealed distinct biomarkers and molecular pathways implicated in fibrosis. In a label-free LC-MS/MS study comparing these IPF fibroblast cell lines with the normal lung fibroblast cell line (CCD19Lu), 80 proteins were found exclusively expressed in IPF cells, with additional 19 upregulated in LL97A and 10 in LL29. Furthermore, hub proteins identified via STRING network analysis were enriched in cell adhesion, integrin binding and hematopoietic cell lineage pathways, suggesting key roles in fibroblast activation and ECM remodeling [61].
Primary IPF patient-derived fibroblast proteomic profiling has provided detailed information on disease protein alterations that are potentially lost in non-disease or immortal models. Among these studies, comparative proteomics of IPF and systemic sclerosis (SSc) patients' primary fibroblasts, through the LC-MS on sequential extraction of cellular and ECM proteins processed with Tandem Mass Tag (TMT) reagents, revealed a shared ECM (matrisome) signature with overexpression of proteins such as PLOD2, LUM, POSTN, IGFBP5, GREM1, and SPARC [62], showing similar “fibrotic signatures” for IPF and SSc of activated myofibroblasts, mirroring the results of transcriptomic and miRNA data. Here, the matrisome proteomic profiles in IPF and SSc lung fibroblasts together with analyses of mRNA and miRNA, gave a crucial contribution to the comprehensive analysis of fibrotic signatures at both the gene and protein expression levels of IPF and SSc.
In a mouse model of IPF, LC-MS/MS and iTRAQ were applied to characterize proteomic changes in murine primary fibroblasts after fibrotic stimulation with bleomycin. In their study, Della Latta et al; identified key proteins involved in the transition from physiological to fibrotic states, including MMPs, OPN, CHI3L1, and CD44, with profound differences in soluble protein secretion and extracellular vesicle-derived factors compared to untreated fibroblasts [63].
To this purpose, in conjunction with cell and tissue-based proteomic approaches, extracellular vesicles (EVs) proteomics has also become a useful technology to study intercellular communication in IPF. Mass spectrometry-based proteomic profiling has recently been applied to IPF-derived fibroblast-secreted EVs for the identification and characterization of their protein cargo within the IPF lung microenvironment [64]. Proteomic analysis of EVs may serve as a valuable tool for both diagnostic and prognostic applications, while also reducing the need for invasive procedures and providing insight into the mechanisms driving IPF progression. The first shotgun proteomic investigation of EVs isolated from BALF of IPF patients characterized the proteome of the vesicular component of BALF. To do this, LC-MS/MS was performed on the total proteins isolated from EVs BALF compared to that of whole BALF in IPF patients, showing considerable differences between them. Analysis of protein content and protein functions evidenced that EV proteins are predominantly involved in cytoskeleton remodeling, adenosine signaling, adrenergic signaling, C-peptide signaling and lipid metabolism. Finally, their findings suggests the importance of the application of mass spectrometry as high sensitive technique for elucidating the pathogenesis of IPF analyzing the low-abundant proteins-mediated pathways. [65]. Moreover, proteomic analysis was performed through LC-MS/MS on plasma EVs in patients diagnosed with IPF, chronic hypersensitivity pneumonitis, nonspecific interstitial pneumonitis, and healthy subjects. Thus, five-protein signature was identified and validated in an independent cohort using ELISA, building a biomarker panel for differential diagnosis of IPF [66]. Finally, label-free proteomics of bronchoalveolar lavage fluid EVs (BALF-EVs) collected from mice treated with bleomycin, identified 107 proteins enriched in fibrotic vesicles compared to that in control mice. Moreover, integrative omics analysis revealed fibroblasts as a major cellular source of BALF-EV cargo, which was enriched in secreted frizzled related protein 1 (SFRP1) that led to increased transitional cell markers, such as keratin 8, and WNT/β-catenin signaling in primary alveolar type 2 cells. Then, Sfrp1 deficiency in fibroblast-derived EVs attenuates lung fibrosis in vivo. These findings suggest the role for SFRP1-EVs as both biomarker of IPF and therapeutic target [67].
7.1. New insights in biomarker discovery with single cell proteomic
Currently, several antibody-based methods for quantifying proteins in single cells have been introduced, including Cytometry by time of flight (CyTOF) [68], single-cell Western blots [69] and simultaneous multiplexed measurement of proteins in single cells using immune-assay coupled with PCR [70]. These techniques are able to quantify a few dozen of endogenous proteins recognized by specific antibodies. Of particular interest is the application of single-cell proteomics, an emerging frontier that enables protein quantification at near-single-cell resolution, aiming at a multi-layered characterization of the proteome signature compared to bulk MS. Analysis of protein content from individual cells can reveal cellular heterogeneity, providing insights into both normal and pathological physiology, but is currently limited to expression proteomics.
Single-cell proteomic is a challenging technique considering both the small quantity of protein in a single cell [7], [71], [8] and the evidence that proteomics cannot amplify the proteins quantity measured, although there is typically more than 100-fold more protein than transcript for every expressed gene. Nowadays, single-cell proteomics technology is based on advances in sample preparation, MS instrumentation and data acquisition that lead to an ultrasensitive single cell resolution. Specifically, it consists of lysis and proteolytic digestion of limited number of cells in nanoliter droplets, followed by direct loading onto the analytical column [72].
In particular, in a study using the Single Cell Proteomics by Mass Spectrometry (SCoPE) method, individually picked live cells were lysed by sonication and after the digestion with trypsin of the proteins in the lysates, the resulting peptides were labeled with TMT labels, combined and analyzed by LC-MS/MS, giving as readout about 1000 proteins for 100 analyzed tumor cells [73].
Nowadays, single-cell proteomics primarily exists as an emerging methodological technology and has not yet been directly applied to IPF tissues at single-cell resolution. However, the use of robust scRNA-seq and spatial proteomics provides highly relevant insights into the pathological mechanisms and target cell populations in IPF lungs. Then, integrating bulk tissue proteomics, single-cell platforms, EV profiling, and spatial proteomic are helping to delineate layered protein dysregulation and paracrine signaling networks that remain undetectable through transcriptomics alone. These high-resolution datasets are transforming our understanding of IPF pathogenesis from a generalized tissue response to a spatially resolved, functionally specialized process and are increasingly being leveraged to identify precision therapeutic targets.
8. AI-Powered Proteomics: A New Era in Translational Medicine
In complex and high-dimensional datasets of mass spectrometry–based proteomics, Artificial Intelligence (AI), particularly machine learning and deep learning, can improve peptide and protein identification enhancing quantification accuracy. AI-driven computational and predictive models can improve the proteomic workflows in biomarkers discovery identifying the protein expression profile that specifically discriminate between healthy and disease states in different medical fields such as cardiovascular diseases, cancer and neurodegenerative diseases [74], [75]. Deep learning can forecast experimental peptide measurements from amino acid sequences across huge proteomic datasets, resulting in higher reliability and consistency in MS workflows. Moreover, an AI technology named federated deep learning (e.g., ProCanFDL), allows the distribution of large datasets across several institutions without sharing confidential patient data [76]. AI facilitates the integration of proteomics with other omics techniques—such as genomics, transcriptomics, and metabolomics, which is critical for identifying novel therapeutic targets and developing personalized treatment strategies [74].
AI and machine learning for analyzing mass spectrometry–derived proteomic data, have been applied in the research of IPF, for advancement in both biomarker identification and disease classification. To date, serum proteomics analyzed by DIA together with machine learning stratified IPF patients into distinct molecular subgroups associated with clinical outcomes, identifying age-related pathways and proteins such as LDHA and CCT6A. A machine-learned combinatorial biomarker panel distinguished IPF patients from healthy individuals [77]. Furthermore, plasma proteomes together with 1,600 patients with either IPF or connective tissue disease-associated interstitial lung disease employed recursive feature elimination and multiple classifiers to train classifiers such as support vector machine (SVM) and Least Absolute Shrinkage and Selection Operator (LASSO), which are suitable and reliable for high-dimensional proteomic data. This gave as outcome 37-protein classifier (PC37) that with high sensitivity and specificity were able to distinguish IPF from other ILDs [78]. Additionally, integrated analyses of lung transcriptomic and plasma proteomic supported by an AI-based analytic approach identified a set of 34 differentially expressed analytes in IPF samples compared to healthy controls. Indeed, IPF samples showed strong enrichment of chemotaxis, tumor infiltration and mast cell migration pathways and downregulated ECM degradation together with mucosal (CCL25 and CCL28) and Th2 (CCL17 and CCL22) chemokines upregulated in IPF and highly correlated within the subjects [79]. Finally, direct application of deep learning models to raw MS spectra—for proteomics—offer the potential for automated feature discovery and improved interpretability in IPF proteomic research facilitating the translation to the clinic.
9. Clinical Translation of proteomics from bench to bedside in IPF
Biological markers that might be identified with aforementioned techniques of mass spectrometry are essential elements for forecasting the course and fatality of IPF.
Biomarkers, when paired with medical evaluations, allow for the creation of predictive models that estimate individual risks related to disease progression and mortality[32]. Patients with IPF have different rates of disease progression and treatment response, making the condition inherently heterogeneous. Clinicians can use biomarkers to detect progressive phenotypes, which is essential for adjusting therapeutic interventions and disease management to each patient's needs [5]. In addition to helping with risk stratification, biomarkers may be early warning signs for acute exacerbations, a dangerous and frequently fatal side effect of IPF [4], [80]. A more precise and personalised prognosis is made possible by a multifaceted approach layered risk assessment in IPF, which integrates imaging results and omics data with clinician-based evaluations. This integrative approach can help determine whether a more conservative treatment plan or more aggressive therapeutic approaches are necessary based on individual risk profiles [81]. The effectiveness of treatment and patient standard of care in IPF can be thus improved by combining this information to create a full, tailored treatment programme according to the collected data [82].
Notwithstanding, rigorous validation is necessary for biomarkers to cross the research-to-clinical divide [83]. Research into clinical utility examines the effects of biomarkers on patient management and therapeutic decisions; confirmatory studies evaluate the efficacy and reliability of biomarkers in wider patient cohorts; initial exploratory studies identify potential biomarkers; and, lastly, biomarkers must gain legislative endorsement before being included in standard clinical protocols for the diagnosis, prognosis and treatment of IPF [84].
To guarantee uniformity and comparability across various studies and research facilities, standardized protocols for the collection and storage of breath samples must be established. For widespread clinical adoption, it is essential to develop analytical methods for measuring breath biomarkers that are sensitive, specific, and economical. Taken together, extensive and multicenter studies are necessary to prove the accuracy and consistency of respiration-based biomarkers as tools for diagnosis or prognosis [85].
To guarantee the scalability and feasibility for clinical application of biomarkers and the improvement of patient prognosis in such conditions as idiopathic pulmonary fibrosis, cooperation between academic, clinical, and industry partners is essential, as is adherence to regulatory guidelines. The use of biomarkers in IPF management appears to have a bright future with the introduction of new technologies in the fields of single-cell sequencing and proteomic, genomic profiling, and spatial transcriptomics and proteomic that will advance our knowledge of the illness and help identify new biomarkers. For IPF patients, combining several biomarkers with clinical data could significantly advance personalized medicine by revolutionizing risk assessment and treatment selection.
9.1. Current limitations to the clinical translation of proteomics
Cost remains an important issue for the widespread use of proteomics in clinical protocols and settings. Thus, it is critical to advance technologies that are both effective and economically sustainable with the implementation of biochemical protocols that will allow high-throughput analyses of several samples adapted to the demands of routine clinical practice. Another concern is related to the limited awareness and integration of mass spectrometry technologies within the clinical community, as their use has been confined to basic and preclinical research contexts rather than certified clinical diagnostic laboratories, whose validation is mandatory before their implementation in the routine patient care [86].
Thus, the transition of LC-MS techniques from research laboratories into clinical laboratories requires the alignment with Clinical Laboratory Improvement Amendments (CLIA) that implies adequate laboratory space and infrastructure, staff training, competency and quality management [87]. For this reason, a limited number of FDA-approved diagnostic assays, such as MALDI-TOF MS, are available. Nowadays, MS-based diagnostic tests and device that either measure or identify biomarkers are not currently used in clinical practice [88], whilst few biomarkers identified through mass spectrometry and validated with orthogonal assays in one or several independent cohorts, have been translated into clinical settings [89], [90]. Then, if a panel of different biomarkers identified needs to be validated, a subsequent computational model based on AI, [91] must to be employed for precise quantification with additional costs, staff training and competency.
10. Conclusion
Different MS approaches—from GC-MS analysis of volatile organic compounds in exhaled breath to innovative LC-MS/MS applications on plasma, bronchoalveolar lavage fluid, lung tissue, cell line, primary cells from lung niche and extracellular vesicles—have uncovered a broad spectrum of IPF biomarkers associated with crucial molecular pathways such as ECM remodeling, immune regulation, oxidative stress, inflammation, and fibrotic signaling. Furthermore, several key proteins such as MMPs, CCL18, periostin, and surfactant proteins, summarized in Table 1, consistently emerged across multiple sample types and studies, underscoring their potential as reliable biomarkers of disease progression and therapeutic targets. Finally, emerging spatial proteomics and single cell-based techniques have enhanced resolution at tissue and cellular levels, providing novel insights into IPF pathogenesis. Developing minimally invasive biomarker assays from accessible samples like blood or exhaled breath will facilitate early diagnosis and real-time disease monitoring.
Taken together, these findings reveal the potential of proteomic mass spectrometry to deepen our understanding of IPF for the development of diagnostic and prognostic tools along with new therapeutic target therapy. The integration of advanced mass spectrometry technologies with multi-omics data and large-scale validation studies is fundamental for translating proteomic biomarkers into clinical practice for IPF. Emerging approaches such as single-cell and spatial proteomics offer new opportunities to characterize cellular heterogeneity and microenvironmental factors driving fibrosis, potentially unveiling new therapeutic targets. Standardization of sample collection, processing, and analytical protocols across studies will be critical to ensure reproducibility and comparability of results. Additionally, interdisciplinary collaboration among researchers, clinicians, and industry will accelerate the translation of these proteomic insights into personalized medicine frameworks, ultimately improving patient outcomes and quality of life.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, G.R, AVS, SC. and R.T.; resources, E.C.; data curation, G.R, AVS, F.G, G.B, T.P, F.A, writing—original draft preparation, G.R,A V.S,R.T, E.T, V.M, B.A, T.P, G.B, F.G, E.T, A.E, F.A,.; writing—review and editing, A.E, M.D, E.C.; visualization G.R,A V.S,R.T, E.T, V.M, B.A, T.P, G.B, F.G, E.T, A.E, F.A, A.E, M.D, E.C; supervision, E.C, M.D, A.E.; project administration, S.C, E.C.; funding acquisition, E.C. All authors have read and agreed to the published version of the manuscript.” Please turn to the CRediT taxonomy for the term explanation. Authorship must be limited to those who have contributed substantially to the work reported.
Funding
The authors declare financial support was received for the research, authorship, and/or publication of this article. The realization of this project produced as part of the research program at Experimental Pneumology, University of Modena and Reggio Emilia (www.experimentalpneumology.unimore.it) “This research was funded by Fondazione Modena (FOMO) within the project: CLINI_FAR2023_INTER DISCIPLINARE_FOMO - UA.20 - CUP E53C23001690007 - Progetto “Single cell and spatial biology for the precise molecular profile of primary and secondary lung fibrotic diseases” - FAR 2023 INTERDISCIPLINARE Linea FOMO, The research leading to these results has received funding from the European Union-NextGenerationEU through the Italian Ministry of University and Research under PNRR-M4C2-l1.3 Project PE_00000019“HEAL ITALIA” to prof. Enrico Clini, CUP E93C22001860006. The views and opinions expressed are those of the authors only and do not necessarily select those of the European Union or the European Commission. Neither the European Union nor the European Commission can be held responsible for them.
Data Availability Statement
We encourage all authors of articles published in MDPI journals to share their research data. In this section, please provide details regarding where data supporting reported results can be found, including links to publicly archived datasets analyzed or generated during the study. Where no new data were created, or where data is unavailable due to privacy or ethical restrictions, a statement is still required. Suggested Data Availability Statements are available in section “MDPI Research Data Policies” at https://www.mdpi.com/ethics.
Acknowledgments
We thank for the valuable contribution “Federazione Italiana Malattie dell’Apparato Respiratorio (FIMARP), Modena Golf Country Club, AMMI (Associazione Mogli di Medici Italiani B-PER Banca, Felicia Kingsley and Massimo Borri for the support in the figures preparation.
Conflicts of Interest
“The authors declare no conflicts of interest. ”The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results”.
Abbreviations
The following abbreviations are used in this manuscript:
2D Two-Dimensional
2D-PAGE Two-Dimensional Polyacrylamide Gel Electrophoresis
16S rRNA 16S Ribosomal Ribonucleic Acid
AAV DTR Adeno-Associated Virus Diphtheria Toxin Receptor
AI Artificial Intelligence
ASPN Asporin
ATS American Thoracic Society
BALF Bronchoalveolar Lavage Fluid
BALF-EVs Extracellular Vesicles Isolated from Bronchoalveolar Lavage Fluid
BTNL9 Butyrophilin Like 9
CCL CC-chemokine Ligand
CCL17, 22, 25, 28 Chemokine (C-C motif) Ligands
CCL18 Chemokine (C-C motif) Ligand 18
CCD19Lu Normal Human Lung Fibroblast Cell Line
CD44 Cluster of Differentiation 44
ChIP-Seq Chromatin Immunoprecipitation Sequencing
CLIA Clinical Laboratory Improvement Amendments
COL1A1 Collagen Type I Alpha 1 Chain
COPD Chronic Obstructive Pulmonary Disease
CRP C-Reactive Protein
CyTOF Cytometry by Time of Flight
CCT6A Chaperonin Containing TCP1 Subunit 6A
DIA Data-Independent Acquisition
DIGE Differential Gel Electrophoresis
DLCO Diffusion Lung Carbon Monoxide
ECM Extracellular Matrix
ELISA Enzyme-Linked Immunosorbent Assay
ERS European Respiratory Society
ESI Electrospray Ionization
FDA Food and Drug Administration
FF Fibroblastic Foci
FFPE Formalin-Fixed Paraffin-Embedded (Tissue)
FGF Fibroblast Growth Factor
FVC Forced Vital Capacity
GC-MS Gas Chromatography–Mass Spectrometry
GREM1 Gremlin 1
HDMSE High Definition Mass Spectrometry with Data-Dependent Acquisition
HP Haptoglobin
HPX Hemopexin
HRCT High-Resolution Computed Tomography
HRMS High-Resolution Mass Spectrometry
HSP90AA1 Heat Shock Protein 90 Alpha Family Class A Member 1
HSP90AB1 Heat Shock Protein 90 Alpha Family Class B Member 1
IIP Idiopathic Interstitial Pneumonia
IGFBP5 Insulin-Like Growth Factor Binding Protein 5
IL Interleukin
ILD Interstitial Lung Disease
IPF Idiopathic Pulmonary Fibrosis
iTRAQ Isobaric Tags for Relative and Absolute Quantitation
KL-6 Krebs von den Lungen-6
LASSO Least Absolute Shrinkage and Selection Operator
LC-MS Liquid Chromatography–Mass Spectrometry
LC-MS/MS Liquid Chromatography–Tandem Mass Spectrometry
LCP1 Lymphocyte Cytosolic Protein 1
LDHA Lactate Dehydrogenase A
LGALS7 Galectin-7
LL29, LL97A IPF-derived Human Lung Fibroblast Cell Lines
LFQ Label-Free Quantification
LTBP1 Latent Transforming Growth Factor Beta Binding Protein 1
LUM Lumican
MALDI Matrix-Assisted Laser Desorption/Ionization
MALDI-TOF MS Matrix-Assisted Laser Desorption Ionization–Time of Flight Mass Spectrometry
MALDI-ToF/ToF Matrix-Assisted Laser Desorption/Ionization Time-of-Flight/Time-of-Flight Mass Spectrometry
MIF Macrophage Migration Inhibitory Factor
MIMECAN (OGN) Mimecan
MMP Matrix Metalloproteinase
MMPs Matrix Metalloproteinases
MS Mass Spectrometry
MS/MS Tandem Mass Spectrometry
MUC5B Mucin 5B
NF-kB Nuclear Factor Kappa-light-chain-enhancer of Activated B Cells
NMR Nuclear Magnetic Resonance
OGN Mimecan
OPN Osteopontin
PC37 37-Protein Classifier
PCR Polymerase Chain Reaction
PDE4B Phosphodiesterase 4B
PLOD2 Procollagen-Lysine, 2-Oxoglutarate 5-Dioxygenase 2
POSTN Periostin
PPARγ Peroxisome Proliferator-Activated Receptor Gamma
PRDX2 Peroxiredoxin-2
ProCanFDL Proteomics Cancer Federated Deep Learning
PTMs Post-Translational Modifications
RNA-Seq RNA Sequencing
RCT Randomized Controlled Trial
SAA1 Serum Amyloid A1
SCGB1A1 Secretoglobin Family 1A Member 1
SCoPE Single Cell Proteomics by Mass Spectrometry
SERPINB3 Serpin Family B Member 3
SFRP1 Secreted Frizzled-Related Protein 1
SNP Single Nucleotide Polymorphism
SOMAmer Slow-Off Rate-Modified Aptamer
SP-A Surfactant Protein A
SP-D Surfactant Protein D
SPARC Secreted Protein Acidic and Rich in Cysteine
STRING Search Tool for the Retrieval of Interacting Genes/Proteins
SSc Systemic Sclerosis
SVM Support Vector Machine
TAGLN2 Transgelin 2
TERC Telomerase RNA Component
TERT Telomerase Reverse Transcriptase
TGF-β Transforming Growth Factor Beta
Th2 T Helper Type 2
TMT Tandem Mass Tag
TNF-α Tumor Necrosis Factor Alpha
UIP Usual Interstitial Pneumonia
VEGF Vascular Endothelial Growth Factor
VOCs Volatile Organic Compounds
References
- Moss, B.J.; Ryter, S.W.; Rosas, I.O. Pathogenic Mechanisms Underlying Idiopathic Pulmonary Fibrosis. Annu Rev Pathol 2022, 17, 515–546. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, S.; Tonelli, R.; Murray, M.; Samarelli, A.V.; Spagnolo, P. Environmental Causes of Idiopathic Pulmonary Fibrosis. Int J Mol Sci 2023, 24, 16481. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela, C.; Cottin, V. Epidemiology and Real-Life Experience in Progressive Pulmonary Fibrosis. Curr Opin Pulm Med 2022, 28, 407–413. [Google Scholar] [CrossRef]
- Biondini, D.; Balestro, E.; Sverzellati, N.; Cocconcelli, E.; Bernardinello, N.; Ryerson, C.J.; Spagnolo, P. Acute Exacerbations of Idiopathic Pulmonary Fibrosis (AE-IPF): An Overview of Current and Future Therapeutic Strategies. Expert Rev Respir Med 2020, 14, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Clynick, B.; Corte, T.J.; Jo, H.E.; Stewart, I.; Glaspole, I.N.; Grainge, C.; Maher, T.M.; Navaratnam, V.; Hubbard, R.; Hopkins, P.M.A.; et al. Biomarker Signatures for Progressive Idiopathic Pulmonary Fibrosis. Eur Respir J 2022, 59, 2101181. [Google Scholar] [CrossRef]
- Zheng, P.; Sun, S.; Wang, J.; Cheng, Z.J.; Lei, K.C.; Xue, M.; Zhang, T.; Huang, H.; Zhang, X.D.; Sun, B. Integrative Omics Analysis Identifies Biomarkers of Idiopathic Pulmonary Fibrosis. Cell. Mol. Life Sci. 2022, 79, 66. [Google Scholar] [CrossRef]
- Guo, T.; Steen, J.A.; Mann, M. Mass-Spectrometry-Based Proteomics: From Single Cells to Clinical Applications. Nature 2025, 638, 901–911. [Google Scholar] [CrossRef]
- Aebersold, R.; Mann, M. Mass-Spectrometric Exploration of Proteome Structure and Function. Nature 2016, 537, 347–355. [Google Scholar] [CrossRef]
- Raghu, G.; Collard, H.R.; Egan, J.J.; Martinez, F.J.; Behr, J.; Brown, K.K.; Colby, T.V.; Cordier, J.-F.; Flaherty, K.R.; Lasky, J.A.; et al. An Official ATS/ERS/JRS/ALAT Statement: Idiopathic Pulmonary Fibrosis: Evidence-Based Guidelines for Diagnosis and Management. Am J Respir Crit Care Med 2011, 183, 788–824. [Google Scholar] [CrossRef]
- Lynch, D.A.; Travis, W.D.; Müller, N.L.; Galvin, J.R.; Hansell, D.M.; Grenier, P.A.; King, J. , Talmadge E. Idiopathic Interstitial Pneumonias: CT Features. Radiology 2005, 236, 10–21. [Google Scholar] [CrossRef]
- Selman, M.; Pardo, A. The Leading Role of Epithelial Cells in the Pathogenesis of Idiopathic Pulmonary Fibrosis. Cell Signal 2020, 66, 109482. [Google Scholar] [CrossRef]
- Samarelli, A.V.; Tonelli, R.; Heijink, I.; Martin Medina, A.; Marchioni, A.; Bruzzi, G.; Castaniere, I.; Andrisani, D.; Gozzi, F.; Manicardi, L.; et al. Dissecting the Role of Mesenchymal Stem Cells in Idiopathic Pulmonary Fibrosis: Cause or Solution. Front Pharmacol 2021, 12, 692551. [Google Scholar] [CrossRef] [PubMed]
- Cerri, S.; Monari, M.; Guerrieri, A.; Donatelli, P.; Bassi, I.; Garuti, M.; Luppi, F.; Betti, S.; Bandelli, G.; Carpano, M.; et al. Real-Life Comparison of Pirfenidone and Nintedanib in Patients with Idiopathic Pulmonary Fibrosis: A 24-Month Assessment. Respir Med 2019, 159, 105803. [Google Scholar] [CrossRef]
- Noble, P.W.; Albera, C.; Bradford, W.Z.; Costabel, U.; Glassberg, M.K.; Kardatzke, D.; King, T.E.; Lancaster, L.; Sahn, S.A.; Szwarcberg, J.; et al. Pirfenidone in Patients with Idiopathic Pulmonary Fibrosis (CAPACITY): Two Randomised Trials. Lancet 2011, 377, 1760–1769. [Google Scholar] [CrossRef]
- Richeldi, L.; Azuma, A.; Cottin, V.; Kreuter, M.; Maher, T.M.; Martinez, F.J.; Oldham, J.M.; Valenzuela, C.; Clerisme-Beaty, E.; Gordat, M.; et al. Nerandomilast in Patients with Idiopathic Pulmonary Fibrosis. N Engl J Med 2025, 392, 2193–2202. [Google Scholar] [CrossRef]
- Cassidy, L.; Kaulich, P.T.; Maaß, S.; Bartel, J.; Becher, D.; Tholey, A. Bottom-up and Top-down Proteomic Approaches for the Identification, Characterization, and Quantification of the Low Molecular Weight Proteome with Focus on Short Open Reading Frame-Encoded Peptides. Proteomics 2021, 21, e2100008. [Google Scholar] [CrossRef]
- Eckhardt, C.M.; Wu, H. Environmental Exposures and Lung Aging: Molecular Mechanisms and Implications for Improving Respiratory Health. Curr Environ Health Rep 2021, 8, 281–293. [Google Scholar] [CrossRef] [PubMed]
- Lau, A.T.Y.; Chiu, J.-F. Biomarkers of Lung-Related Diseases: Current Knowledge by Proteomic Approaches. Journal of Cellular Physiology 2009, 221, 535–543. [Google Scholar] [CrossRef]
- Agrawal, L.; Engel, K.B.; Greytak, S.R.; Moore, H.M. Understanding Preanalytical Variables and Their Effects on Clinical Biomarkers of Oncology and Immunotherapy. Semin Cancer Biol 2018, 52, 26–38. [Google Scholar] [CrossRef] [PubMed]
- Chapman, E.A.; Baker, J.; Aggarwal, P.; Hughes, D.M.; Nwosu, A.C.; Boyd, M.T.; Mayland, C.R.; Mason, S.; Ellershaw, J.; Probert, C.S.; et al. GC-MS Techniques Investigating Potential Biomarkers of Dying in the Last Weeks with Lung Cancer. International Journal of Molecular Sciences 2023, 24, 1591. [Google Scholar] [CrossRef]
- Graham, D.R.M.; Elliott, S.T.; Van Eyk, J.E. Broad-Based Proteomic Strategies: A Practical Guide to Proteomics and Functional Screening. J Physiol 2005, 563, 1–9. [Google Scholar] [CrossRef]
- Plantier, L.; Smolinska, A.; Fijten, R.; Flamant, M.; Dallinga, J.; Mercadier, J.J.; Pachen, D.; d’Ortho, M.P.; van Schooten, F.J.; Crestani, B.; et al. The Use of Exhaled Air Analysis in Discriminating Interstitial Lung Diseases: A Pilot Study. Respir Res 2022, 23, 12. [Google Scholar] [CrossRef]
- Velázquez-Enríquez, J.M.; Ramírez-Hernández, A.A.; Navarro, L.M.S.; Reyes-Avendaño, I.; González-García, K.; Jiménez-Martínez, C.; Castro-Sánchez, L.; Sánchez-Chino, X.M.; Vásquez-Garzón, V.R.; Baltiérrez-Hoyos, R. Proteomic Analysis Reveals Differential Expression Profiles in Idiopathic Pulmonary Fibrosis Cell Lines. Int J Mol Sci 2022, 23, 5032. [Google Scholar] [CrossRef]
- Brodbelt, J.S. Deciphering Combinatorial Post-Translational Modifications by Top-down Mass Spectrometry. Curr Opin Chem Biol 2022, 70, 102180. [Google Scholar] [CrossRef]
- Moradian, A.; Kalli, A.; Sweredoski, M.J.; Hess, S. The Top-down, Middle-down, and Bottom-up Mass Spectrometry Approaches for Characterization of Histone Variants and Their Post-Translational Modifications. Proteomics 2014, 14, 489–497. [Google Scholar] [CrossRef]
- Po, A.; Eyers, C.E. Top-Down Proteomics and the Challenges of True Proteoform Characterization. J Proteome Res 2023, 22, 3663–3675. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.M.; Smith, L.M. Overview and Considerations in Bottom-up Proteomics. Analyst 2023, 148, 475–486. [Google Scholar] [CrossRef]
- Griffin, T.J.; Xie, H.; Bandhakavi, S.; Popko, J.; Mohan, A.; Carlis, J.V.; Higgins, L. iTRAQ Reagent-Based Quantitative Proteomic Analysis on a Linear Ion Trap Mass Spectrometer. J Proteome Res 2007, 6, 4200–4209. [Google Scholar] [CrossRef] [PubMed]
- Principi, L.; Ferrini, E.; Ciccimarra, R.; Pagani, L.; Chinello, C.; Previtali, P.; Smith, A.; Villetti, G.; Zoboli, M.; Ravanetti, F.; et al. Proteomic Fingerprint of Lung Fibrosis Progression and Response to Therapy in Bleomycin-Induced Mouse Model. Int J Mol Sci 2023, 24, 4410. [Google Scholar] [CrossRef] [PubMed]
- Aichler, M.; Kunzke, T.; Buck, A.; Sun, N.; Ackermann, M.; Jonigk, D.; Gaumann, A.; Walch, A. Molecular Similarities and Differences from Human Pulmonary Fibrosis and Corresponding Mouse Model: MALDI Imaging Mass Spectrometry in Comparative Medicine. Lab Invest 2018, 98, 141–149. [Google Scholar] [CrossRef]
- Stainer, A.; Faverio, P.; Busnelli, S.; Catalano, M.; Della Zoppa, M.; Marruchella, A.; Pesci, A.; Luppi, F. Molecular Biomarkers in Idiopathic Pulmonary Fibrosis: State of the Art and Future Directions. Int J Mol Sci 2021, 22, 6255. [Google Scholar] [CrossRef]
- Zheng, Z.; Peng, F.; Zhou, Y. Biomarkers in Idiopathic Pulmonary Fibrosis: Current Insight and Future Direction. Chin Med J Pulm Crit Care Med 2024, 2, 72–79. [Google Scholar] [CrossRef]
- Richards, T.J.; Kaminski, N.; Baribaud, F.; Flavin, S.; Brodmerkel, C.; Horowitz, D.; Li, K.; Choi, J.; Vuga, L.J.; Lindell, K.O.; et al. Peripheral Blood Proteins Predict Mortality in Idiopathic Pulmonary Fibrosis. Am J Respir Crit Care Med 2012, 185, 67–76. [Google Scholar] [CrossRef]
- Cui, N.; Hu, M.; Khalil, R.A. Biochemical and Biological Attributes of Matrix Metalloproteinases. Prog Mol Biol Transl Sci 2017, 147, 1–73. [Google Scholar] [CrossRef]
- Zuo, F.; Kaminski, N.; Eugui, E.; Allard, J.; Yakhini, Z.; Ben-Dor, A.; Lollini, L.; Morris, D.; Kim, Y.; DeLustro, B.; et al. Gene Expression Analysis Reveals Matrilysin as a Key Regulator of Pulmonary Fibrosis in Mice and Humans. Proc Natl Acad Sci U S A 2002, 99, 6292–6297. [Google Scholar] [CrossRef]
- Rosas, I.O.; Richards, T.J.; Konishi, K.; Zhang, Y.; Gibson, K.; Lokshin, A.E.; Lindell, K.O.; Cisneros, J.; Macdonald, S.D.; Pardo, A.; et al. MMP1 and MMP7 as Potential Peripheral Blood Biomarkers in Idiopathic Pulmonary Fibrosis. PLoS Med 2008, 5, e93. [Google Scholar] [CrossRef] [PubMed]
- Chuliá-Peris, L.; Carreres-Rey, C.; Gabasa, M.; Alcaraz, J.; Carretero, J.; Pereda, J. Matrix Metalloproteinases and Their Inhibitors in Pulmonary Fibrosis: EMMPRIN/CD147 Comes into Play. Int J Mol Sci 2022, 23, 6894. [Google Scholar] [CrossRef] [PubMed]
- Prasse, A.; Probst, C.; Bargagli, E.; Zissel, G.; Toews, G.B.; Flaherty, K.R.; Olschewski, M.; Rottoli, P.; Müller-Quernheim, J. Serum CC-Chemokine Ligand 18 Concentration Predicts Outcome in Idiopathic Pulmonary Fibrosis. Am J Respir Crit Care Med 2009, 179, 717–723. [Google Scholar] [CrossRef] [PubMed]
- Hamai, K.; Iwamoto, H.; Ishikawa, N.; Horimasu, Y.; Masuda, T.; Miyamoto, S.; Nakashima, T.; Ohshimo, S.; Fujitaka, K.; Hamada, H.; et al. Comparative Study of Circulating MMP-7, CCL18, KL-6, SP-A, and SP-D as Disease Markers of Idiopathic Pulmonary Fibrosis. Dis Markers 2016, 2016, 4759040. [Google Scholar] [CrossRef]
- Okamoto, M.; Izuhara, K.; Ohta, S.; Ono, J.; Hoshino, T. Ability of Periostin as a New Biomarker of Idiopathic Pulmonary Fibrosis. Adv Exp Med Biol 2019, 1132, 79–87. [Google Scholar] [CrossRef]
- Takayama, G.; Arima, K.; Kanaji, T.; Toda, S.; Tanaka, H.; Shoji, S.; McKenzie, A.N.J.; Nagai, H.; Hotokebuchi, T.; Izuhara, K. Periostin: A Novel Component of Subepithelial Fibrosis of Bronchial Asthma Downstream of IL-4 and IL-13 Signals. J Allergy Clin Immunol 2006, 118, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, M.; Hoshino, T.; Kitasato, Y.; Sakazaki, Y.; Kawayama, T.; Fujimoto, K.; Ohshima, K.; Shiraishi, H.; Uchida, M.; Ono, J.; et al. Periostin, a Matrix Protein, Is a Novel Biomarker for Idiopathic Interstitial Pneumonias. Eur Respir J 2011, 37, 1119–1127. [Google Scholar] [CrossRef]
- d’Alessandro, M.; Bergantini, L.; Cameli, P.; Pieroni, M.; Refini, R.M.; Sestini, P.; Bargagli, E. Serum Concentrations of KL-6 in Patients with IPF and Lung Cancer and Serial Measurements of KL-6 in IPF Patients Treated with Antifibrotic Therapy. Cancers (Basel) 2021, 13, 689. [Google Scholar] [CrossRef] [PubMed]
- Ashley, S.L.; Xia, M.; Murray, S.; O’Dwyer, D.N.; Grant, E.; White, E.S.; Flaherty, K.R.; Martinez, F.J.; Moore, B.B. Six-SOMAmer Index Relating to Immune, Protease and Angiogenic Functions Predicts Progression in IPF. PLoS One 2016, 11, e0159878. [Google Scholar] [CrossRef]
- Zhang, Y.; Xin, Q.; Wu, Z.; Wang, C.; Wang, Y.; Wu, Q.; Niu, R. Application of Isobaric Tags for Relative and Absolute Quantification (iTRAQ) Coupled with Two-Dimensional Liquid Chromatography/Tandem Mass Spectrometry in Quantitative Proteomic Analysis for Discovery of Serum Biomarkers for Idiopathic Pulmonary Fibrosis. Med Sci Monit 2018, 24, 4146–4153. [Google Scholar] [CrossRef]
- Oldham, J.M.; Huang, Y.; Bose, S.; Ma, S.-F.; Kim, J.S.; Schwab, A.; Ting, C.; Mou, K.; Lee, C.T.; Adegunsoye, A.; et al. Proteomic Biomarkers of Survival in Idiopathic Pulmonary Fibrosis. Am J Respir Crit Care Med 2024, 209, 1111–1120. [Google Scholar] [CrossRef] [PubMed]
- Foster, M.W.; Morrison, L.D.; Todd, J.L.; Snyder, L.D.; Thompson, J.W.; Soderblom, E.J.; Plonk, K.; Weinhold, K.J.; Townsend, R.; Minnich, A.; et al. Quantitative Proteomics of Bronchoalveolar Lavage Fluid in Idiopathic Pulmonary Fibrosis. J Proteome Res 2015, 14, 1238–1249. [Google Scholar] [CrossRef]
- Bargagli, E.; Carleo, A.; Bianchi, L.; Gagliardi, A.; Prasse, A.; Perari, M.G.; Refini, R.M.; Bini, L.; Rottoli, P. A System Biology Study of BALF from Patients Affected by Idiopathic Pulmonary Fibrosis (IPF) and Healthy Controls. Proteomics Clin Appl 2014, 8, 932–950. [Google Scholar] [CrossRef]
- Hara, A.; Sakamoto, N.; Ishimatsu, Y.; Kakugawa, T.; Nakashima, S.; Hara, S.; Adachi, M.; Fujita, H.; Mukae, H.; Kohno, S. S100A9 in BALF Is a Candidate Biomarker of Idiopathic Pulmonary Fibrosis. Respir Med 2012, 106, 571–580. [Google Scholar] [CrossRef]
- Rottoli, P.; Magi, B.; Perari, M.G.; Liberatori, S.; Nikiforakis, N.; Bargagli, E.; Cianti, R.; Bini, L.; Pallini, V. Cytokine Profile and Proteome Analysis in Bronchoalveolar Lavage of Patients with Sarcoidosis, Pulmonary Fibrosis Associated with Systemic Sclerosis and Idiopathic Pulmonary Fibrosis. Proteomics 2005, 5, 1423–1430. [Google Scholar] [CrossRef] [PubMed]
- Ngo, L.T.; Rekowski, M.J.; Koestler, D.C.; Yorozuya, T.; Saito, A.; Azeem, I.; Harrison, A.; Demoruelle, M.K.; Boomer, J.; England, B.R.; et al. Proteomic Profiling of Bronchoalveolar Lavage Fluid Uncovers Protein Clusters Linked to Survival in Idiopathic Forms of Interstitial Lung Disease. ERJ Open Research 2024, 10. [Google Scholar] [CrossRef]
- Darville, L.N.F.; Lockhart, J.H.; Putty Reddy, S.; Fang, B.; Izumi, V.; Boyle, T.A.; Haura, E.B.; Flores, E.R.; Koomen, J.M. A Fast-Tracking Sample Preparation Protocol for Proteomics of Formalin-Fixed Paraffin-Embedded Tumor Tissues. Methods Mol Biol 2024, 2823, 193–223. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Liu, W.; Ju, Z.; Tamboli, P.; Jonasch, E.; Mills, G.B.; Lu, Y.; Hennessy, B.T.; Tsavachidou, D. An Efficient Procedure for Protein Extraction from Formalin-Fixed, Paraffin-Embedded Tissues for Reverse Phase Protein Arrays. Proteome Science 2012, 10, 56. [Google Scholar] [CrossRef] [PubMed]
- Samarelli, A.V.; Tonelli, R.; Raineri, G.; Bruzzi, G.; Andrisani, D.; Gozzi, F.; Marchioni, A.; Costantini, M.; Fabbiani, L.; Genovese, F.; et al. Proteomic Profiling of Formalin-Fixed Paraffine-Embedded Tissue Reveals Key Proteins Related to Lung Dysfunction in Idiopathic Pulmonary Fibrosis. Front Oncol 2023, 13, 1275346. [Google Scholar] [CrossRef]
- Tian, Y.; Li, H.; Gao, Y.; Liu, C.; Qiu, T.; Wu, H.; Cao, M.; Zhang, Y.; Ding, H.; Chen, J.; et al. Quantitative Proteomic Characterization of Lung Tissue in Idiopathic Pulmonary Fibrosis. Clin Proteom 2019, 16, 6. [Google Scholar] [CrossRef]
- Zheng, P.; Sun, S.; Wang, J.; Cheng, Z.J.; Lei, K.C.; Xue, M.; Zhang, T.; Huang, H.; Zhang, X.D.; Sun, B. Integrative Omics Analysis Identifies Biomarkers of Idiopathic Pulmonary Fibrosis. Cell. Mol. Life Sci. 2022, 79, 66. [Google Scholar] [CrossRef]
- Herrera, J.A.; Dingle, L.; Montero, M.A.; Venkateswaran, R.V.; Blaikley, J.F.; Lawless, C.; Schwartz, M.A. The UIP/IPF Fibroblastic Focus Is a Collagen Biosynthesis Factory Embedded in a Distinct Extracellular Matrix. JCI Insight 7. [CrossRef]
- Wang, F.; Jin, L.; Wang, X.; Cui, B.; Yang, Y.; Duggan, L.; Schwartz Sterman, A.; Lloyd, S.M.; Hazelwood, L.A.; Chaudhary, N.; et al. Novel Integration of Spatial and Single-Cell Omics Data Sets Enables Deeper Insights into IPF Pathogenesis. Proteomes 2025, 13, 3. [Google Scholar] [CrossRef]
- Chandran, R.R.; Vijayaraj, P.; Garcia-Milian, R.; King, J.; Castillo, K.; Chen, L.; Kwon, Y.; William, S.; Rickabaugh, T.M.; Langerman, J.; et al. Loss of Cell Junctional Components and Matrix Alterations Drive Cell Desquamation and Fibrotic Changes in Idiopathic Pulmonary Fibrosis. bioRxiv 2024, 2024.06.17.599411. [CrossRef]
- Griesser, E.; Gesell, M.; Veyel, D.; Lamla, T.; Geillinger-Kästle, K.; Rist, W. Whole Lung Proteome of an Acute Epithelial Injury Mouse Model in Comparison to Spatially Resolved Proteomes. PROTEOMICS 2023, 23, 2100414. [Google Scholar] [CrossRef] [PubMed]
- Velázquez-Enríquez, J.M.; Ramírez-Hernández, A.A.; Navarro, L.M.S.; Reyes-Avendaño, I.; González-García, K.; Jiménez-Martínez, C.; Castro-Sánchez, L.; Sánchez-Chino, X.M.; Vásquez-Garzón, V.R.; Baltiérrez-Hoyos, R. Proteomic Analysis Reveals Differential Expression Profiles in Idiopathic Pulmonary Fibrosis Cell Lines. Int J Mol Sci 2022, 23, 5032. [Google Scholar] [CrossRef]
- Mullenbrock, S.; Liu, F.; Szak, S.; Hronowski, X.; Gao, B.; Juhasz, P.; Sun, C.; Liu, M.; McLaughlin, H.; Xiao, Q.; et al. Systems Analysis of Transcriptomic and Proteomic Profiles Identifies Novel Regulation of Fibrotic Programs by miRNAs in Pulmonary Fibrosis Fibroblasts. Genes 2018, 9, 588. [Google Scholar] [CrossRef]
- Latta V, D.; A, C.; L, C.; N, U.; C, D.P.; M, T.; S, B.; G, P.; S, R.; Ma, M. A Proteomics Approach to the Study of Bleomycin- Induced Lung Fibrosis. J Transl Sci 2018, 5. [Google Scholar] [CrossRef]
- Kadota, T.; Kosaka, N.; Fujita, Y.; Araya, J.; Kuwano, K.; Ochiya, T. Chapter 13 - Extracellular Vesicles in Fibrotic Diseases: New Applications for Fibrosis Diagnosis and Treatment. In Exosomes; Edelstein, L., Smythies, J., Quesenberry, P., Noble, D., Eds.; Academic Press, 2020; pp. 307–323 ISBN 978-0-12-816053-4.
- Shaba, E.; Landi, C.; Carleo, A.; Vantaggiato, L.; Paccagnini, E.; Gentile, M.; Bianchi, L.; Lupetti, P.; Bargagli, E.; Prasse, A.; et al. Proteome Characterization of BALF Extracellular Vesicles in Idiopathic Pulmonary Fibrosis: Unveiling Undercover Molecular Pathways. International Journal of Molecular Sciences 2021, 22, 5696. [Google Scholar] [CrossRef] [PubMed]
- Adduri, R.S.R.; Cai, K.; Velasco-Alzate, K.; Vasireddy, R.; Miller, J.W.; de Frías, S.P.; de Frías, F.P.; Horimasu, Y.; Iwamoto, H.; Hattori, N.; et al. Plasma Extracellular Vesicle Proteins as Promising Noninvasive Biomarkers for Diagnosis of Idiopathic Pulmonary Fibrosis. J Extracell Biol 2023, 2, e98. [Google Scholar] [CrossRef] [PubMed]
- Burgy, O.; Mayr, C.H.; Schenesse, D.; Papakonstantinou, E.F.; Ballester, B.; Sengupta, A.; She, Y.; Hu, Q.; Melo-Narvaéz, M.C.; Jain, E.; et al. Fibroblast-Derived Extracellular Vesicles Contain SFRP1 and Mediate Pulmonary Fibrosis. JCI Insight 2024, 9. [Google Scholar] [CrossRef]
- Bandura, D.R.; Baranov, V.I.; Ornatsky, O.I.; Antonov, A.; Kinach, R.; Lou, X.; Pavlov, S.; Vorobiev, S.; Dick, J.E.; Tanner, S.D. Mass Cytometry: Technique for Real Time Single Cell Multitarget Immunoassay Based on Inductively Coupled Plasma Time-of-Flight Mass Spectrometry. Anal. Chem. 2009, 81, 6813–6822. [Google Scholar] [CrossRef]
- Hughes, A.J.; Spelke, D.P.; Xu, Z.; Kang, C.-C.; Schaffer, D.V.; Herr, A.E. Single-Cell Western Blotting. Nat Methods 2014, 11, 749–755. [Google Scholar] [CrossRef]
- Darmanis, S.; Gallant, C.J.; Marinescu, V.D.; Niklasson, M.; Segerman, A.; Flamourakis, G.; Fredriksson, S.; Assarsson, E.; Lundberg, M.; Nelander, S.; et al. Simultaneous Multiplexed Measurement of RNA and Proteins in Single Cells. Cell Reports 2016, 14, 380–389. [Google Scholar] [CrossRef]
- Blume, J.E.; Manning, W.C.; Troiano, G.; Hornburg, D.; Figa, M.; Hesterberg, L.; Platt, T.L.; Zhao, X.; Cuaresma, R.A.; Everley, P.A.; et al. Rapid, Deep and Precise Profiling of the Plasma Proteome with Multi-Nanoparticle Protein Corona. Nat Commun 2020, 11, 3662. [Google Scholar] [CrossRef]
- Li, Z.-Y.; Huang, M.; Wang, X.-K.; Zhu, Y.; Li, J.-S.; Wong, C.C.L.; Fang, Q. Nanoliter-Scale Oil-Air-Droplet Chip-Based Single Cell Proteomic Analysis. Anal Chem 2018, 90, 5430–5438. [Google Scholar] [CrossRef]
- Budnik, B.; Levy, E.; Harmange, G.; Slavov, N. SCoPE-MS: Mass Spectrometry of Single Mammalian Cells Quantifies Proteome Heterogeneity during Cell Differentiation. Genome Biology 2018, 19, 161. [Google Scholar] [CrossRef]
- Mann, M.; Kumar, C.; Zeng, W.-F.; Strauss, M.T. Artificial Intelligence for Proteomics and Biomarker Discovery. Cell Syst 2021, 12, 759–770. [Google Scholar] [CrossRef]
- Gessulat, S.; Schmidt, T.; Zolg, D.P.; Samaras, P.; Schnatbaum, K.; Zerweck, J.; Knaute, T.; Rechenberger, J.; Delanghe, B.; Huhmer, A.; et al. Prosit: Proteome-Wide Prediction of Peptide Tandem Mass Spectra by Deep Learning. Nat Methods 2019, 16, 509–518. [Google Scholar] [CrossRef]
- How Is AI Shaping Proteomics and Multiomics? Available online: http://www.technologynetworks.com/immunology/articles/how-is-ai-shaping-proteomics-and-multiomics-395403 (accessed on 24 July 2025).
- Wang, L.; Zhu, M.; Li, Y.; Yan, P.; Li, Z.; Chen, X.; Yang, J.; Pan, X.; Zhao, H.; Wang, S.; et al. Serum Proteomics Identifies Biomarkers Associated With the Pathogenesis of Idiopathic Pulmonary Fibrosis. Mol Cell Proteomics 2023, 22, 100524. [Google Scholar] [CrossRef]
- Huang, Y.; Ma, S.-F.; Oldham, J.M.; Adegunsoye, A.; Zhu, D.; Murray, S.; Kim, J.S.; Bonham, C.; Strickland, E.; Linderholm, A.L.; et al. Machine Learning of Plasma Proteomics Classifies Diagnosis of Interstitial Lung Disease. Am J Respir Crit Care Med 2024, 210, 444–454. [Google Scholar] [CrossRef]
- Sivakumar, P.; Ammar, R.; Thompson, J.R.; Luo, Y.; Streltsov, D.; Porteous, M.; McCoubrey, C.; Cantu, E.; Beers, M.F.; Jarai, G.; et al. Integrated Plasma Proteomics and Lung Transcriptomics Reveal Novel Biomarkers in Idiopathic Pulmonary Fibrosis. Respir Res 2021, 22, 273. [Google Scholar] [CrossRef]
- Tsubouchi, K.; Hamada, N.; Tokunaga, S.; Ichiki, K.; Takata, S.; Ishii, H.; Kitasato, Y.; Okamoto, M.; Kawakami, S.; Yatera, K.; et al. Survival and Acute Exacerbation for Patients with Idiopathic Pulmonary Fibrosis (IPF) or Non-IPF Idiopathic Interstitial Pneumonias: 5-Year Follow-up Analysis of a Prospective Multi-Institutional Patient Registry. BMJ Open Respir Res 2023, 10, e001864. [Google Scholar] [CrossRef] [PubMed]
- Ruan, P.; Todd, J.L.; Zhao, H.; Liu, Y.; Vinisko, R.; Soellner, J.F.; Schmid, R.; Kaner, R.J.; Luckhardt, T.R.; Neely, M.L.; et al. Integrative Multi-Omics Analysis Reveals Novel Idiopathic Pulmonary Fibrosis Endotypes Associated with Disease Progression. Respir Res 2023, 24, 141. [Google Scholar] [CrossRef] [PubMed]
- Salisbury, M.L.; Wijsenbeek, M.S. Management of Idiopathic Pulmonary Fibrosis. Clin Chest Med 2021, 42, 275–285. [Google Scholar] [CrossRef]
- Aronson, J.K.; Ferner, R.E. Biomarkers—A General Review. Current Protocols in Pharmacology 2017, 76, 9.23–1. [Google Scholar] [CrossRef] [PubMed]
- Bartold, K.; Iskierko, Z.; Sharma, P.S.; Lin, H.-Y.; Kutner, W. Idiopathic Pulmonary Fibrosis (IPF): Diagnostic Routes Using Novel Biomarkers. Biomed J 2024, 47, 100729. [Google Scholar] [CrossRef]
- Karampitsakos, T.; Juan-Guardela, B.M.; Tzouvelekis, A.; Herazo-Maya, J.D. Precision Medicine Advances in Idiopathic Pulmonary Fibrosis. EBioMedicine 2023, 95, 104766. [Google Scholar] [CrossRef]
- Kosack, C.S.; Page, A.-L.; Klatser, P.R. A Guide to Aid the Selection of Diagnostic Tests. Bull World Health Organ 2017, 95, 639–645. [Google Scholar] [CrossRef]
- Han, C.-L.; Lai, C.-T.; Reyes, A.J.; Yang, H.-C.; Lu, J.-Y.; Shih, S.-R.; Chen, K.-Y.; Hoofnagle, A.N.; Yu, S.-L.; Bocik, W.; et al. Lessons Learned: Establishing a CLIA-Equivalent Laboratory for Targeted Mass Spectrometry Assays – Navigating the Transition from Research to Clinical Practice. Clin Proteomics 2024, 21, 12. [Google Scholar] [CrossRef] [PubMed]
- Lathrop, J.T.; Jeffery, D.A.; Shea, Y.R.; Scholl, P.F.; Chan, M.M. US Food and Drug Administration Perspectives on Clinical Mass Spectrometry. Clinical Chemistry 2016, 62, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Thomas, S.N. Impact of VALID Act Implementation on Mass Spectrometry-Based Clinical Proteomic Laboratory Developed Tests. J Mass Spectrom Adv Clin Lab 2023, 28, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Molina-Pinelo, S.; Pastor, M.D.; Paz-Ares, L. VeriStrat: A Prognostic and/or Predictive Biomarker for Advanced Lung Cancer Patients? Expert Review of Respiratory Medicine 2014, 8, 1–4. [Google Scholar] [CrossRef]
- Sun, Y.; Selvarajan, S.; Zang, Z.; Liu, W.; Zhu, Y.; Zhang, H.; Chen, W.; Chen, H.; Li, L.; Cai, X.; et al. Artificial Intelligence Defines Protein-Based Classification of Thyroid Nodules. Cell Discov 2022, 8, 85. [Google Scholar] [CrossRef]
Figure 1.
A Bottom-up proteomics is based on the indirect identification of proteins through their constituent peptides. The workflow starts with the acquisition of a biological sample (1) and extraction of total proteins (2), followed by enzymatic digestion, commonly with trypsin, to cleave proteins at defined sites and generate peptides suitable for MS analysis (3). Peptides are purified and, when needed, fractionated using strong cation exchange (SCX), high-pH reversed-phase LC, or other orthogonal techniques to reduce complexity (4). Separation by nanoLC or CE, and subsequent ionization by electrospray (5), enable MS1 analysis for peptide profiling and quantification (6). Selected precursor ions undergo fragmentation, typically via HCD or CID, to produce fragment ion spectra (7), which are analyzed by tandem MS (MS/MS) to determine peptide sequences (8). The resulting spectra are interpreted using database search engines and bioinformatics pipelines to identify peptides and infer the presence, abundance, and PTMs of the corresponding proteins (9). Although widely applicable and highly scalable, this approach does not preserve proteoform-level information and may obscure isoform- and PTM-specific distinctions. B Top-down proteomics enables the direct analysis of intact proteoforms without enzymatic digestion. The workflow begins with the collection of a biological sample (1), followed by the extraction of intact proteins under native or denaturing conditions (2). After protein separation, typically via reversed-phase liquid chromatography (RPLC) or capillary electrophoresis (CE), and electrospray ionization (3), intact protein ions are introduced into the mass spectrometer for high-resolution MS1 analysis to determine precursor masses and charge states (4). Gas-phase fragmentation techniques, such as electron capture dissociation (ECD), electron transfer dissociation (ETD), or higher-energy collisional dissociation (HCD), are then applied to generate fragment ions (5), which are analyzed by tandem MS to obtain sequence information (6). Computational tools subsequently perform spectral deconvolution, proteoform identification, and localization of post-translational modifications (PTMs) and sequence variants (7). This approach provides direct, site-resolved characterization of proteoforms, although it is currently limited by dynamic range and proteome coverage.
Figure 1.
A Bottom-up proteomics is based on the indirect identification of proteins through their constituent peptides. The workflow starts with the acquisition of a biological sample (1) and extraction of total proteins (2), followed by enzymatic digestion, commonly with trypsin, to cleave proteins at defined sites and generate peptides suitable for MS analysis (3). Peptides are purified and, when needed, fractionated using strong cation exchange (SCX), high-pH reversed-phase LC, or other orthogonal techniques to reduce complexity (4). Separation by nanoLC or CE, and subsequent ionization by electrospray (5), enable MS1 analysis for peptide profiling and quantification (6). Selected precursor ions undergo fragmentation, typically via HCD or CID, to produce fragment ion spectra (7), which are analyzed by tandem MS (MS/MS) to determine peptide sequences (8). The resulting spectra are interpreted using database search engines and bioinformatics pipelines to identify peptides and infer the presence, abundance, and PTMs of the corresponding proteins (9). Although widely applicable and highly scalable, this approach does not preserve proteoform-level information and may obscure isoform- and PTM-specific distinctions. B Top-down proteomics enables the direct analysis of intact proteoforms without enzymatic digestion. The workflow begins with the collection of a biological sample (1), followed by the extraction of intact proteins under native or denaturing conditions (2). After protein separation, typically via reversed-phase liquid chromatography (RPLC) or capillary electrophoresis (CE), and electrospray ionization (3), intact protein ions are introduced into the mass spectrometer for high-resolution MS1 analysis to determine precursor masses and charge states (4). Gas-phase fragmentation techniques, such as electron capture dissociation (ECD), electron transfer dissociation (ETD), or higher-energy collisional dissociation (HCD), are then applied to generate fragment ions (5), which are analyzed by tandem MS to obtain sequence information (6). Computational tools subsequently perform spectral deconvolution, proteoform identification, and localization of post-translational modifications (PTMs) and sequence variants (7). This approach provides direct, site-resolved characterization of proteoforms, although it is currently limited by dynamic range and proteome coverage.
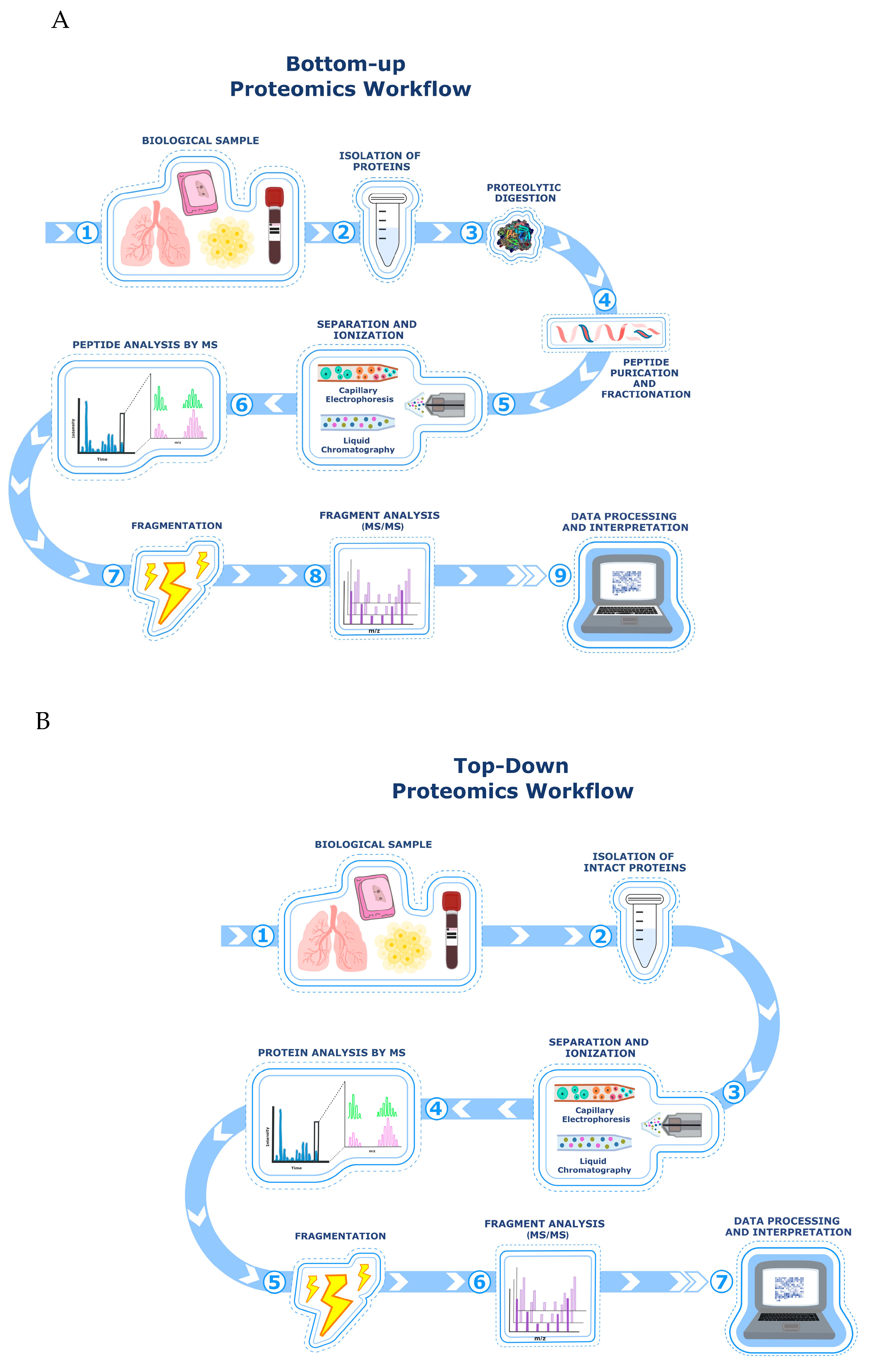

Table 1.
Mass Spectrometry-Based Biomarker Discovery in IPF and Related Pathways.
| Mass Spectrometry Technique | Identified Biomarkers | Activated Molecular Pathway | Sample Type | Reference(s) |
| GC-MS | Acetone, other VOCs | Oxidative stress, inflammation | Exhaled breath (VOCs) | [23] |
| LC-MS/MS | MMP-7, MMP-1, MMP-10 | ECM remodeling | Plasma/Serum | [34,35,36,37,38] |
| LC-MS/MS | CCL18 | Inflammation, immunoregulation | Plasma/Serum | [39,40] |
| LC-MS/MS | Periostin | TGF-β, IL-4, IL-13 signaling | Plasma/Serum | [41,42,43] |
| LC-MS/MS | SP-A, SP-D, KL-6 | Immune response, alveolar regeneration | Serum | [33, 40, 44] |
| SOMAscan (aptamer-based) | VEGFR2, Ficolin-2, Legumain, Cathepsin, ICOS, Trypsin-3 | IPF progression (not specified) | Plasma | [45] |
| iTRAQ + LC-MS/MS | CRP, Fibrinogen-α, Haptoglobin, Kininogen-1 | Systemic inflammation | Plasma | [46] |
| Quantitative proteomics | SAA1, Haptoglobin, Hemopexin | Inflammation, oxidative stress, ECM | Plasma | [47] |
| HDMSE, MRM, 2D-PAGE, MALDI-ToF | MMP-7, CXCL7, CCL18, S100A9, ILs, MIF, Calgranulin B, CCL24 | Inflammation, ECM remodeling, cellular signaling | BALF | [48,49,50,51,52] |
| LC-MS/MS (FFPE tissue) | LCP1, PRDX2, TAGLN2, LUM, OGN | TGF-β, cell adhesion, ECM | Lung tissue (FFPE) | [55] |
| iTRAQ + LC-MS/MS | COL1A1, SCGB1A1, HSP90AA1/AB1, LGALS7, ASPN | ECM production and remodeling | Fresh lung tissue | [56] |
| LCM-MS, Spatial Proteomics | TGF-β1/2/3, LTBP1, FN1, SFRP1 | TGF-β signaling, ECM remodeling, EMT | Laser-captured lung tissue sections | [58,59,60,61] |
| Label-free LC-MS/MS | >80 proteins (e.g., POSTN, IGFBP5, SPARC) | Matrisome, cell adhesion, ECM signaling | Primary cell lines (fibroblasts) | [62,63] |
| LC-MS/MS (EVs from BALF & plasma) | SFRP1, signaling proteins, cytokines, cytoskeletal proteins | WNT/β-catenin, cell–cell communication | EVs from BALF and plasma | [64,65,66,67,68] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



