
Submitted:
10 September 2025
Posted:
12 September 2025
You are already at the latest version
Abstract
The open ocean cage aquaculture system is facing considerable challenges with disease outbreaks resulting from over-farming and the rise of resistance to antimicrobial treatment. Although antibiotics can deal with some health issues, they have other implications for the environment, so more judicious options are warranted. Lipopeptides, which are a compound that can be produced by marine bacteria such as Bacillus amyloliquefaciens or Bacillus subtilis, could represent a new solution, and this review will assess their feasibility. Lipopeptides, including surfactins, fengycins, iturins, and the clinically-used daptomycin, have notable antiviral, antifungal, and antimicrobial properties, and can have positive effects on the immune system. Notably, lipopeptides have a remarkable antioxidant profile, and have excellent free radical scavenging ability, making them interesting candidates for improving disease resistance in fish relating to oxidative stress. The surfactins and iturins have amphiphilic structure and can destabilize pathogen cell membranes, inhibit biofilm formation and elicit host immune responses. This completely changes the game with regards to multi targeting pathogens of aquaculture like Vibrio spp. and Aeromonas spp. Surfactins and iturins show broad-spectrum activity while fengycins are selectively active against fungal threats. Daptomycin, which is primarily derived from Streptomyces, demonstrates the potential of the lipopeptide class to develop therapeutically, which is something that tends to be overlooked. Unlike synthetic antibiotics, they are also biodegradable therefore there is much less environmental impact from lipopeptides. The complexity of the structure may have also some impact on the rate of development of resistance, if any. Their commercialization is possible however, the main hurdles that need to be solved to improve aquaculture are the biologically scalable production, the economically viable purification, and the stability for practical application at sea. Fast-evolving fermentation techniques, genetic manipulation and the use of nanodrug systems have the potential to enhance aquaculture processes and productivity, and treatment. Implementing lipopeptides into disease management systems could also ensure the sustainability of open ocean cage aquaculture and reduce unnecessary antibiotic application. This review has drawn attention to an area that has immense value, the marine lipopeptides, and as it is clear from this review that we need multiple disciplines to work create user-friendly pathways to transition research into action to address global aquaculture sustainability.
Keywords:
Aquaculture
; Disease outbreaks
; antimicrobial resistance
; Lipopeptides
; Aquaculture pathogens
; Sustainable aquaculture
1. Introduction
Aquaculture, defined as the processed and controlled development of aquatic organisms (fish, mollusks, shrimp, and plants) is the fastest-growing food production sector in the world, two additional goals are to support higher demand for seafood globally, promote food sovereignty, import replacement, and promote the development of economies in several developing countries and consumption feeding 3 billion people worldwide [1,2]. The process can aid with food security, creation of livelihood and economic development. Aquaculture also has some ecological gains if sustainability practices are established [3,4]. The global supply of aquatic food has risen according to the FAO FishStat Database [5] from 32.4 million metric tonnes in 2000 to 132.4 million metric tonnes in 2013, with roughly 17% of global animal protein provided by aquaculture. Aquaculture growth in aquaculture is anticipated to trend upward and provide 66% of the total fish consumption by the year 2030 accounted for by a global population climbing to 9.3 billion in 2050 and requiring 60% food production [6]. Due to the associated emerging issues related to mass production, expansion and growth of aquaculture can introduce tremendous risks associated with environmental pollution, habitat degradation, and infection control [7,8]. Open-sea cage aquaculture is directly associated with infection risk due to the genetic homogeneity and high-density stocking of fish populations lived in open sea environments exposed directly to ocean. The open-sea and open-aquaculture settings create unique conditions to exacerbate the rate of transmission and spread of viral, bacterial, and parasitic infections. Furthermore, cumulative environmental stressors found in open aquaculture, such as temperature, salinity, dissolved oxygen, environmental quality, stress, will amplify transmission risk rates and increase risk for management or treatment failure causing significant economic loss due to mortalities, lower growth, and/or increased treatment costs. Common diseases in open-water cage aquaculture include bacterial diseases such as vibriosis and aeromonasis; viral diseases, with infectious pancreatic necrosis (IPN) and viral hemorrhagic septicemia (VHS) being many of the prominent closing; and parasitic infestations like sea lice. Several of these diseases can lead to high mortality rates and in the case of vibriosis is capable of causing 100 percent mortality rate if the impacted fish are not treated. The losses are substantial, with various estimates of global losses due to disease in aquaculture reaching billions of dollars annually. Annual global losses due to diseases take into account lost fish due to mortality, but diseases also lead to reduced growth rates, increased feed conversion ratios, and increased production costs because of treatment or other management measures to reduce the effects of diseases [9,10].
In the past, disease management followed an effective but not environmentally friendly way, traditionally employing antibiotics or chemical treatments. Antibiotic and chemical resistance have developed from overuse of these materials which places health risk to both human and fish health and ecology. The impact of chemicals on the environment is concerning ecosystem health in terms of pollution and habitat destruction. Hence, more sustainable and environmentally-friendly approaches are badly required for disease control in aquaculture. Marine bacteria are responsible for the synthesis of different lipopeptides, such as antimicrobial compounds (Bacillus amyloliquefaciens and Bacillus subtilis) with antioxidant activity or that influence the immune system. Another alternative to the traditional antibiotics is lipopeptides, which have also been recognized as an excellent source of antimicrobials in most cases given that they are biodegradable and less prone to resistance. Lipopeptides are formed by a peptide ring and a fatty acid chain, resulting in amphiphilic properties that allow them to have interactions with biological membranes as they disrupt cells membranes of pathogens while also disrupting the biofilm formation of many pathogens. Marine bacteria (especially those from the genus Bacillus) produce a variety of lipopeptides surfactins, iturins, and fengycins all possess various antimicrobial spectra as surfactins have broad-spectrum antibacterial activity, iturins have pronounced antifungal activity, while fengycins has activity against filamentous fungi. They have also demonstrated significant antioxidant properties, which perhaps has the greatest benefit through the mitigation of oxidative stress in fish and ultimately help increase the fish’s resilience to diseases. The long history of aquaculture, dating back to 1000 BC carp aquaculture in Chinese texts, has experienced rapid modern growth as an industry since the mid-20th century buoyed by advancing technology and increasing seafood demand [11]. Presently, aquaculture is an international industry, 90% of global production occurs in Asia, which produces many fish, crustaceans, and mollusks such as carp, tilapia, and shrimp [5]. Countries like India, the second largest producer in the world, and others have seen tremendous growth across developing countries, with states like Andhra Pradesh and Gujarat as leaders in production, demonstrating its importance for food security, and sustainable economic development in developing countries. This review will consider the suitability of the incorporation of lipopeptide-based antibiotics and lipopeptide-based antioxidants in the management of disease in open-sea cage aquaculture to manage the disease. In addition, we will discuss the mode of action, applications, and issues concerning commercialization. By summarizing some literature and primary literature on lipopeptides from marine bacteria, we hope to inform future research and development efforts to improve more sustainable and resilient aquaculture practices to further address challenges facing open-sea cage systems and food security globally.
Data Analysis
A comprehensive literature search was conducted across SCOPUS, Google Scholar, PubMed, and Web of Science to identify studies published between 1996 and 2025 relevant to the application of lipopeptides in aquaculture, particularly concerning antimicrobial resistance and sustainable practices. The search utilized a combination of keywords such as “aquaculture,” “lipopeptides,” “Bacillus subtilis,” “antimicrobial resistance,” “marine bacteria,” “surfactins,” “fengycins,” “iturins,” “daptomycin,” and “sustainable aquaculture,” employing Boolean operators (AND, OR) to refine the results. A total of approximately 600 citations were retrieved from these databases. After removing duplicates and excluding studies that did not meet the inclusion criteria based on titles and abstracts, 106 unique publications were selected. To ensure the inclusion of the most recent research, additional key publications from 2024 and 2025 were incorporated into the review. This selection provides a solid foundation for assessing the role of lipopeptides in mitigating antimicrobial resistance and advancing sustainable aquaculture practices.
2. Open Sea Cage and Disease Management
Aquaculture includes inland, coastal and mariculture, Coastal aquaculture and mariculture include finfish, shellfish, mollusks and seaweed, which are produced in a marine environment, while inland aquaculture produces in freshwater systems such as ponds, tanks and reservoirs (uses mostly groundwater). Inland aquaculture accounted for approximately 77% of total edible aquatic products, comprised of carp, tilapia, catfish and shrimp obtained between 2010 and 2022 [12,13]. Inland saline aquaculture systems are found in arid areas where the availability of freshwater is scarce [12]. Pond culture contributes approximately 70% to the global freshwater output and ranges from extensive to intensive systems, and includes systems that can be at risk of eutrophication [14]. Recirculating Aquaculture Systems (RAS) reuse water (90–99%) for production with better control and high-density production [15]. Integrated systems like rice–fish farming as an example of integrated systems are found to improve sustainability, consuming only 3,000 m3 of water in producing one kg of fish, with rice–fish farming producing 850 kg/ha/year (Figure 1).
Coastal aquaculture occurs on ponds or cages near the coasts, resulting in significant contributions to economies and ecosystems, [16,17]. Mariculture (e.g., fish and shellfish farming) is responsible for 37.5% of aquaculture production and 97% of the total world seaweed harvest, while mariculture has sustained losses due to disease outbreaks, with total cumulative losses ranging up to USD 800 million between 2015 and 2023. [18] (Figure 2). In terms of shellfish culture, they provide nutrients, ecological and environmental benefits such as water cleansing [19]. Finfish culture in cage-net structures has been established for certain species such as groupers, seabass and others [20] but there are also different substances for finfish energy capture, storage, and use in additional consumption as fish, like pen culture as an adaptation of artificial culture systems cosigning the natural waters for species such as tilapia or carp [21,22], there are also floating near-shore cages provided by the environmental system that are dependent on the current flow [23,24], there is increasing interest in offshore cage culture provided by deeper water currents of species such as tuna or cobia given the conflicts with coastal communities [25,26]. Open-sea cage culture involves the use floating or submerged net cages which are robust and suitable for finfish like sea bass and milkfish, in exposed water conditions in marine waters. Open-sea cage systems may also embrace polyculture, such as fin fish and seaweed, as a more sustainable approach but nutrient enrichment and seabed impacts must be monitored to manage use appropriately [21,27,28,29].
Open sea cage culture is an aquaculture system that transports floating cages with anchors into open waters for farming fish in near natural environmental conditions. The open-cage system has several important structural components. The main component would be the structure support such as high density polyethylene (HDPE) plastic or steel structure that are constructed specifically to be flexible or durable according to the environment it will be installed. The nylon or polythene used to make the nets is one which allows adequate water movement in an appropriate design yet which blocks entry by predators and mesh size is important in this regard (Figure 6). Support facilities are moorings to which the cages are tied, floating and buoying devices, and feeding plants which can be manual or automatic. Modern cage systems also incorporate tracking and monitoring techniques to observe water-quality variables and fish activity for good health and potential for production [30,31,32,33,34]. The selection of the site is very important in open sea cage aquaculture practice, since the environmental and operational constraints affect the productivity greatly. To choose the optimal location, technics of spatial modeling (e.g., GIS) are developed and useful. A study in India also found 134 potential sites for utilizing marine resources through multi-criteria evaluation [35]. With respect to cage design, the fully enclosed pile-net pens (FPAPs) have potential to be used for the offshore farming, as they are able to imitate the natural habitats more realistically and protect the marine stock from exacerbated stress [36,37]. The quality of fish seed is yet another factor for successful cage culture. Good-quality seed stock is healthy, uniformly sized and of great genetic variability. In comparing wild fish seed versus hatchery fish seed, fish caught from the wild appeared to have better survivorship characteristics as compared to fish from hatcheries. The acclimation process is the most important step in putting young fish into a cage. Acclimation allows for the fish to become familiar with their new environment, and therefore there will be less initial stress and mortality in the fish [38,39]. Feeding strategies for open-sea cages can be a combination of approaches that include commercially produced feeds that are formulated protein-dense feeds and local natural feed types in the form of trash fish. It is possible to effectively manage feeds with feeding frequency and feast or famine rations with the support of rapid technological advancements in automated feeders and drones [40,41]. While one of the main challenges to open-sea cage aquaculture is susceptibility to disease and disease outbreaks. The density of fish stocked creates suitable density and conditions to ensure an outbreak of pathogens is possibly based upon the surrounding environment and the stability of the environmental conditions [42] (Figure 3). Additionally, fish are susceptible to numerous bacterial and parasitic diseases. Vibriosis infections from other Vibrio species are troublesome infections that have a diverse host or bacterial species. Parasitic fish diseases that typically infect brackish aquaculture barramundi fish are diseases such as trypanosomiasis caused by Trypanosoma epinepheli and sea lice (also found in wild fish) (Figure 4). An example of a parasite affecting European seabass is Diplectanum aequans [43,44], or tenacibaculosis (Tenacibaculum maritimum) had its own considerations as the summer jellyfish blooms move along coastal areas toward the management areas of aquaculture [28,45].
Bacterial diseases significantly impact aquaculture productivity and pose a risk to sustainability (Figures 4). These include the hemorrahgic septicemiás and skin lesions caused by Vibrio anguillarum and V. parahaemolyticus; streptococcosis (neurological signs and exophthalmia) caused by Streptococcus iniae and S. agalactiae; pasteurellosis caused by Photobacterium damselae subsp. piscicida with an acute pathogenesis and mortality >50%; furunculosis caused by Aeromonas salmonicida with skin ulvercation and organ dysfunction; Yersinia ruckeri with the infection known as enteric redmouth disease, which manifests itself as hemorrhaging; bacterial kidney disease (BKD) caused by Renibacterium salmoninarum which persists as a chronic kidney infection; septicemia caused by, Pseudomonas aeruginosa/P. anguilliseptica; nocardiosis which is caused by granulomatous lesions produced by Nocardia seriolae [46,47,48,49,50,51,52,53,54]. New strategies for disease control are proposed, such as prevention measures: biorsecurity measures, vaccination, ideal nutrition and food combined with best management practices and data of control of the environmental [55,56,57,58] and management procedures with the goal of more restricted targeted control of disease: e.g., antibiotics, antiparasitic drugs, delousing procedures, selective breeding for disease resistance, and (biotherapeutics) supplements (e.g., probiotics, prebiotics, synbiotics) [59,60,61,62,63]. Over the past decades excessive use of antibiotics has led to antibiotic resistant bacteria (ARB) and genes (ARGs) transferring to food, farm products and water, which required the exploration of alternatives, especially non-pharmacological alternatives [64]. Antimicrobial lipopeptides appear as an interesting biotherapeutic technology, as they exhibit activity in vitro against resistant bacterial strains, and propose a sustainable disease management option for marine reared farmed fish.
3. Structural Features and Therapeutic Potential of Marine-Derived Lipopeptides
Lipopeptides are versatile amphiphilic molecules that are primarily derived from microbial sources, while marine bacteria are also an important source of lipopeptides due to their evolutionary adaptations to harsh oceans. Lipopeptides are a potential solution for disease control, including in sustainable aquaculture, where any disease unexpectedly arises and potential antimicrobial resistance develops, certainly merits a degree of exploratory perspective. Within this wide range of compounds there have been a relatively high number of studies conducted in association with the Bacillus genus including many different species, and Bacillus amyloliquefaciens and Bacillus subtilis specifically have been explicitly studied in connection with the therapeutic aspect of lipopeptides [65]. While Pseudomonas and Streptomyces are also recognized producers of lipopeptides, focusing on marine Bacillus species will be important in applications in the marine environment. There are several types of lipopeptides recognized for their biological activity. Surfactins and iturins have seen the most study due to their antimicrobial activity, demonstrated by their ability to disrupt pathogen cell membranes, inhibit biofilm formation, and stimulate the host’s immune response. Fengycins, on the other hand, have been shown to have specific efficacy against fungal threats and have antifungal effects while inhibiting plant pathogens [66]. Mycosubtilin, also a lipopeptide from marine Bacillus species, has been studied for its strong synergy against a broad spectrum of pathogens causing fish disease. While daptomycin is a clinically used lipopeptide primarily derived from Streptomyces and is not derived from marine bacteria, it serves as an example for the substantial therapeutic potential offered by this class of compounds but is often overlooked in a wider scope. Collectively these compounds demonstrate good antiviral, antifungal, and antibacterial properties and other beneficial effects on the immune system, and have a notable antioxidant capacity as well by scavenging free radicals, and providing fish with better resistance to disease. The unique biological activities of lipopeptides are due to their unique amphiphilic structural features which means that they possess both hydrophobic (water fearing) and hydrophilic (water loving) regions. The molecule consists of a fatty acid chain (lipid moiety) and a peptide segment [67]. The hydrophobic tail is typically a β-hydroxy fatty acid and usually made up of 10 to 16 carbons, linked to the peptide through either esters like that which occurs in surfactin’s lactone rings or amides like that which occurs in daptomycin. The lipid chain is important in activity as the C10–C12 lipid yields higher bactericidal activity and C14 or C16 lipid chains are broader in antibacterial and antifungal activity [68]. The hydrophilic peptide fragment is a short chain, typically 7 to 10 amino acid residues and may be cyclic or linear (Figure 5) [69]. This peptide is synthesized through a NRPS–non ribosomal peptide synthetase, allowing the incorporation of non-proteinogenic amino acids which is likely the source of the structural diversity of the lipopeptides.
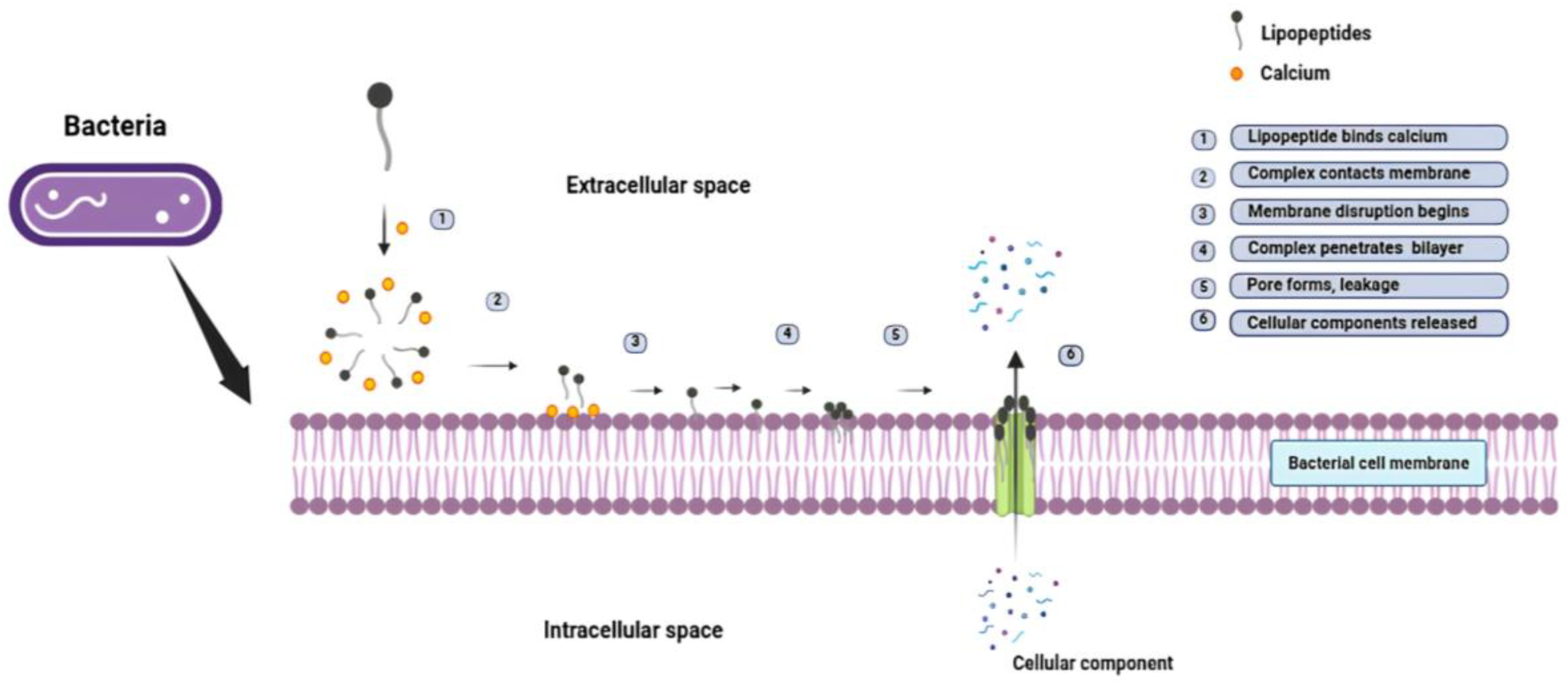
This unique amphipathic nature enables lipopeptides to reduce surface and interfacial tension to a remarkable degree, which makes them remarkable biosurfactants [67]. Additionally, their structurally amphiphilic character also gives them the ability to self-assemble into a variety of nanostructures which can be micelles, vesicles, bilayer sheets, and nanotubes, as shown in Figure 5. They exhibit strong antimicrobial activity primarily by disrupting the cell membranes of microbial cells. The hydrophobic lipid tail inserts into the pathogen’s membrane, while the peptide part form pores or causes depolarization of the membrane, causing the cellular contents to leak out of the cell and therefore cell death [68] as illustrated in Figure 6.
Figure 6.
Mechanism of bacterial cell membrane disruption by lipopeptides: The process occurs in six steps: (1) Lipopeptides bind calcium, (2) Complex forms, (3) Complex contacts membrane, (4) Disruption starts, (5) Pores form, leakage occurs, (6) Cellular components released. In the extracellular space, the amphipathic lipopeptide associate with calcium, allowing its hydrophobic lipid tail to insert into the bacterial membrane while the hydrophilic peptide remains outside. This forms a channel through which cellular components are released, leading to lysis and bacterial death. Created using Inscape software.
Figure 6.
Mechanism of bacterial cell membrane disruption by lipopeptides: The process occurs in six steps: (1) Lipopeptides bind calcium, (2) Complex forms, (3) Complex contacts membrane, (4) Disruption starts, (5) Pores form, leakage occurs, (6) Cellular components released. In the extracellular space, the amphipathic lipopeptide associate with calcium, allowing its hydrophobic lipid tail to insert into the bacterial membrane while the hydrophilic peptide remains outside. This forms a channel through which cellular components are released, leading to lysis and bacterial death. Created using Inscape software.
Iturin, for example disrupts cell wall biosynthesis and disrupts cell shape and leads to structural failure and cell lysis, in addition to potency against biofilms, this multi-target activity makes them effective against antibiotic-resistant strains that are emerging in aquaculture, depending on the region and species of aquaculture there are many different mortality rates depicted. As a result, it is prudent to explore alternatives such as these lipopeptides in light of the increasing challenges of disease outbreaks in aquaculture. Specific strains of marine bacteria have produced remarkably high yields of lipopeptides and efficacy in aquaculture settings. For example, Bacillus amyloliquefaciens isolated from marine sediment demonstrated a significant antimicrobial activity against Vibrio anguillarum, a common aquaculture pathogen, by affecting cell membrane and biofilms of pathogens. Similarly, surfactin and iturin produced by Bacillus subtilis of marine origin can not only break down cell membranes of pathogens, but also dissolve biofilm, protective shielding that pathogens such as Vibrios use to increase infection. Both B. amyloliquefaciens and B. subtilis produce different lipopeptides surfactins, iturins, fengycins, and mycosubtilin, that together effectively and synergistically protected against a wide range of fish disease pathogens [70]. Thus, the use of lipopeptides in aquaculture compounds provides an environmentally friendly alternative to conventional antibiotics for disease management in aquaculture and supports efforts to improve sustainable methodologies in worldwide food production.
4. Multifunctional Lipopeptides for Health Management in Open-Sea Cage Aquaculture
Lipopeptides cyclic peptides covalently bonded to fatty acid chains, produced by marine bacteria including Bacillus amyloliquefaciens and Bacillus subtilis possess many bio activities making them ideal candidates for disease control in open-sea cage aquaculture [70]. Lipopeptides include a wide range of bios under persistent attack from a wide variety of pathogens including e.g., antimicrobial, antioxidant, and immune-modulating properties, lipopeptides offer a sacrificial degree of protection to aquatic organisms, and their biodegradable features and low resistant development make them useful sustainable alternatives to synthetic antibiotics. Lipopeptides have broad-spectrum inhibition across bacteria, fungi, and viruses addressing the major micro threats to aquaculture developed by novel sources highlighting their formidable antifungal and antiviral protection. Lipopeptide can be classified in three categories surfactins, iturins, (including bacillomycins and mycosubtilin) and fengycins. each group has varied efficacy profiles. Surfactins, the smallest heptapeptides, are known to act as biosurfactants, but they can also cause membrane disruption in Gram+ bacteria while having some inhibitory effects on Gram negative bacterial fish pathogens, including the Vibrio and Aeromonas species. For example, B. amyloliquefaciens M1 lipopeptides demonstrated a 34 mm inhibition zone against multidrug-resistant Vibrio anguillarum in agar plates [71,72]. Surfactins can also damage enveloped viruses, such as herpes simplex by disrupting viral envelopes, which indicates the potential for these agents as viral disease prevention and control [73]. Iturins and mycosubtilin have very strong antifungal activity through binding to and forming an ion-conducting pore through sterols in fungal membranes. They are also effective against pathogens, including Fusarium and Alternaria respectively, that pose a threat to both plants and fish [74]. Fengycins are decapeptides with a lactone ring near the middle of the sequence that inhibit filamentous fungi: during biocontrol studies, they inhibited Botrytis cinerea and other agronomic pathogens [75]. Interestingly, they were able to also exhibit low levels of activity against some Gram-negative bacteria, including Xanthomonas spp. which suggests that fengycins may be able to promote control not only of fungi but of specific bacteria as well [76]. By amalgamating families within one bacterial strain or formulations, one can provide a synergistically, multi-targeted antimicrobial effect which can be advantageous in the complex microbial communities of sea cages. Oxidative stress encompasses an imbalance between reactive oxygen species (ROS) and antioxidant defenses and can damage fish immunity and growth when fish are kept at high stocking densities with fluctuating environmental conditions. However, lipopeptides are known to approach oxidative stress and ROS through free radical scavenging, inhibition of lipid-peroxidation and chelation of metal ions. In a study, DCS1 lipopeptides from Bacillus methylotrophicus DCS1 exhibited 80.6% DPPH radical scavenging at 1 mg/mL, 80.8% inhibition of β-carotene bleaching at the same concentration, and ferrous-ion chelation [77]. The delayed consumption of the antioxidant assays is an indication that lipopeptides possess strong antioxidant activity and could be a way to protect the tissue of fish from oxidative damage to preserve cellular intactness and improve growth rate in stressful times. We acknowledge the absence of direct trials in fish, however, the microbial and in vitro studies published support that lipopeptides are natural antioxidants and could be beneficial in aquaculture feed additives or water treatments.
Lipopeptides serve as immunomodulators in addition to disinfecting pathogens and neutralizing ROS by activating innate immune receptors. Toll-like receptors (TLRs) in vertebrates identify pathogen-associated molecular patterns (PAMPs), such as tri-acylated lipopeptides, stimulate signal cascades initiating phagocytosis and cytokine production. While many immunology studies have focused on research in mammals, fish possess a complex with an important set of innate immune receptors including TLR1 and TLR2 that can still form heterodimers to detect bacterial lipopeptides [77]. In zebrafish, Bricknell and Dalmo [78] have shown through work in models that tri-acylated lipopeptides are robust adjuvants and indicate they may play a similar role in aquaculture species. They may be able to improve early pathogen clearance by priming macrophages and neutrophils and therefore reduce the number of times antibiotics would need to be used therapeutically. Since marine Bacillus strains can co-produce multiple families of lipopeptides, they can produce intrinsic synergy, surfactins can allow access to membranes, along with iturins permitting pore formation and fengycins preferably affecting the integrity of fungal cell walls. We would recommend formulating lipopeptides in combination, using drug cocktail formulations, or using a controlled-release matrix to improve field application. Nanoparticle encapsulation provides a new benefit to lipopeptides by preserving their potency from rapid dilution and enzymatic degradation in seawater, while allowing sustained released at the target sites [79]. As a further bonus, the use of a delivery system assists in resolving challenges associated with stability in rapidly changing ocean environmental conditions and maximizing bioavailability to inhibit established biofilms and chronic pathogens. Although their biological potential is established, their use in cage aquaculture systems requires successful navigation of a number of dependent factors that influence efficacy, from molecular structure and environmental persistence, to innovation in delivery, and regulatory uptake. These are summarized in Table 1 to guide future development efforts. While lipopeptides share efficacy with synthetic antibiotics, they present lower ecotoxicological risks. Biodegradation processes of lipopeptides occur quickly in the marine environment, which can reduce accumulations, which can eliminate downstream impacts without promoting resistance, while synthetic drugs can last longer in the environment, promote resistance, and add antibiotic residues to the human food chain [79]. Additionally, the main action of surfactins and iturins act by disrupting membranes which targets non-specific essential cellular structures; this means that developing mutational resistance is likely less attainable compared with antibiotics that target single sites. Additionally, many lipopeptides derived from Bacillus, have been given GRAS status (generally regarded as safe), which proved their safety to aquatic animals and humans as consumers [75,76,77]. The synergistic antimicrobial, antioxidant, and immune-stimulatory properties of lipopeptides derived from marine-associated bacteria address the three fundamental aspects of disease management in open-sea cage aquaculture: pathogen inhibition, tolerance to stressors, and host-mediated defense. Because lipopeptides have multiple target mechanisms, including cell membrane disruption and immune stimulation through PAMPs, lipopeptides can be effective against numerous fish pathogens, including Vibrio, Aeromonas, Streptococcus, and other opportunistic fungi. Moreover, because lipopeptides are environmentally friendly and have low resistance potential, they have shown to be a high potential to be sustainable alternatives to traditional antibiotics. Suitable concentrations, better co-formulations, better organized shipping forms, and batch production processes for lipopeptides should be developed next. Future efforts in aquaculture should also be focused on in vivo trials confirming efficacy, safe dosage regimens, safety for non-target species, and potential economic feasibility in commercial aquaculture [75,76,77,78,79].
6. Antimicrobial and Immunomodulatory Roles of Marine Bacterial Lipopeptides in Sustainable Aquaculture
Lipopeptides are amphiphilic compounds composed of a hydrophobic lipid tail connected to a hydrophilic peptide that are just beginning to develop as multi-action antimicrobial agents in aquaculture targeting, in particular, drug-resistant pathogens. Although some lipopeptides can be found in higher plants, they are mostly characterized in marine Bacillus species including Bacillus subtilis, B. amyloliquefaciens, and other relatives There lipopeptides are, surfactin, iturin, fengycin, and mycosubtilin. They are primarily defined due to their antimicrobial activity, targeting microbial cell membranes and biofilms. The amphipathic structure allows lipopeptides to insert into lipid bilayers through hydrophobic forces that form transient pores that cause intracellular leakage, while some lipopeptide isoforms slow cell-wall biosynthesis [85]. Considering the information above we can move forward to the application of lipopeptides, and recent studies have demonstrated broad-spectrum activity against significant fish pathogens. For example, B. amyloliquefaciens isolates from marine sediment showed significant antagonism towards Vibrio anguillarum, a prevalent aquaculture pathogen, by damaging its cellular membrane and dissolving protective biofilm matrices which normally protect the pathogen from standard drug effects [86]. In a similar vein, Pathak and Keharia (2014) identified the surfactin-rich lipopeptide, “N3” from Bacillus amyloliquefaciens M1, which produced a zone of inhibition on agar plates of 34 mm against multidrug resistant Vibrio anguillarum, showing its impressive efficacy towards vibriosis, the most common disease facing marine cage systems [87]. There is also evidence that lipopeptides have activity against pathogens other than vibrios. In shrimp aquaculture, a gelatinized lipopeptide biosurfactant (MSA31), was added to feed for Penaeus vannamei [88]. In a 60-day trial, treated shrimp demonstrated increased growth performance, weight gain, and 50% survival rate in relation to a Vibrio parahaemolyticus SF 14 challenge compared to the 100% mortality in the untreated controls within 12 h. Additionally, the effects of lipopeptide on growth performance and immune responses were associated with increased digestive enzyme activity as well. The lipopeptide(s) are likely to not only kill the pathogens but also live in a symbiotic manner with the natural gut microbial community that help to improve the innate defence of the host.
In finfish, Kim et al. [89] fed Bacillus subtilis E20 (another lipopeptide producing strain) and this provided similar benefits. The authors reported improvement in growth and disease resistance for parrot fish (Oplegnathus fasciatus) against Vibrio alginolyticus with higher survival rates and increased immune sensitization for treated fish [89]. The authors conclude that Bacillus-derived lipopeptides have potential benefits for various species in aquaculture from crustaceans to teleosts. Different delivery mechanisms have been designed to transition research success into the application phase. A number of recent studies have quantified the efficacy of marine-derived lipopeptides against major aquaculture pathogens under different delivery strategies. The addition of feed is the more scalable option for prophylactic use: lipopeptides can be gelatinized or microencapsulated, to be added to feed, providing an oral method that consistently delivers lipopeptides to modify the gut microbiota and MOD or enhance the systemic or mucosal immune response [88,89]. Immersion bath systems provide direct contact to lipopeptides on the gill and skin surfaces is also a very effective delivery system, particularly as a treatment for ectoparasites, for studies with sea lice for example, can provide lipopeptide solutions to is rearing water, thus pathogens on surfaces encounter membrane disrupting molecules upon contact. Injection, whether IP or IM, of lipopeptides is more laborious, but it would provide the most control for therapeutic use of known volumes of a known concentration in order to control acute outbreaks, directly to the fish, compared to other delivery mechanisms. There may be an opportunity to add Bacillus probiotics live to culture water as a control measure, due to the outcompeting live nature of Bacillus and to reduce the total environmental load of Vibrio and Aeromonas spp. pathogenic bacteria. In description, pathogenic bacteria compete with Bacillus for nutrients and attachment to surfaces [90]. Besides antibacterial capabilities, lipopeptides also have antioxidant mechanisms that contribute to fish health. The DPPH (2,2-diphenyl-1-picrylhydrazyl) assay, which is a widely employed method for assessing free radical scavenging capacity, showed that a few lipopeptide isolates quenched DPPH free radicals and had inhibition percentages that depended on molecular characteristics including hydrophobic domain length and functional groups that donates electrons [91]. Also, a typical positive control, ascorbic acid, approaches complete scavenging activity, so this adds rigor to the assay protocols and provides a basis of comparison against lipopeptides.
Lipopeptides that have hydrophobic areas optimized or that have hydroxyl-bearing side chains, associate more closely with DPPH free radicals while those that have less favorable conformations to react have less activity. The antioxidant capacity of these biomolecules suggests potential application beyond disease control that could enter into food preservation, cosmetics, and pharmaceutical areas, as natural radical scavengers are favored products in these industries. Yet to fully exploit these potentials there is still work to be done, including comparisons with synthetic antioxidants, further toxicity studies, and mechanistic studies such as additional assays (ABTS, FRAP) [91,92]. These studies will provide insight into the structure–activity relationships for antioxidant efficacy and enable molecular design and/or fermentation improvements to produce lipopeptides of superior efficacy. Providing lipopeptides to aquaculture can therefore be multifaceted; they not only replace a range of bacteria and fungi and lessen reliance on conventional antibiotics, and the development of resistance, but enhance growth, immunity and oxidative stress in the host [93,94]. They have also exhibited low toxicity and biodegradability, make them a desirable contributor to sustainable aquaculture by promoting healthier fish stocks, maximizing yields and minimizing environmental impacts. With research underway into optimal production and delivery systems, marine bacterial lipopeptides are a revolutionary approach to fish-disease management and in an age of emerging pathogens and drug resistance, their potential impact may be profound [95,96].
7. Challenges and Future Directions
In open-sea cage aquaculture, lipopeptides have huge potential as natural antimicrobial products for disease management; however, there are multiple substantial barriers that need to be addressed before commercialization can take place. Foremost among them is the need for biologically scalable fermentation processes for the production of lipopeptides (current production is limited to expensive laboratory scale experiments) and the need for improved fermentation technologies that have the capacity to generate larger volumes cost-effectively. The success of the industry will depend on advances in fermentation technology that will make large scale production feasible and economically profitable. In addition, economically viable, effective methods of purification to obtain pure lipopeptides from complex microbial broths is still a major hurdle, and the inability to isolate lipopeptides contributes to high production costs. Most countries have very stringent food safety regulations that add barriers to commercialization inside marine cage farming systems and are further complicated in disease management in the open ocean where it is more difficult to manage disease outbreaks.
Open water cage aquaculture faces considerable sustainability challenges associated with the release of fish waste and excess feed disposal releasing nutrients into the open ocean, leading to phytoplankton blooms that can consume oxygen and disrupt marine ecosystems. Extreme weather events can damage cages resulting in escaped fish, the variable temperatures where fish are grown can introduce further stress on the fish which negatively affects growth performance and fish health. Increased stocking density can facilitate disease outbreaks compared to the contained systems. Vaccine protocols that are developed do not always have a high success rate when spread over diverse species of fish, and it is logistically hard to vaccinate fish in commercial aquacultural operations, especially broodstock.
Biosecurity practices implemented during aquaculture to prevent and mitigate risks of disease are less effective in open water due to continual introduction of uncontrolled water outside into the system, and also due to introduction of potential vectors or pathogens being re-introduced into the aquaculture system as wild migratory fish travel by. Consequently, high use of antibiotic medicine contributes to development of marine pathogens to build resistance to such antibiotics which threatens health at the larger scale, especially considering studies done in regions like southeast Asia. The context makes it an absolute priority to establish stability and delivery of alternatives such as lipopeptides in such an extremely dynamic environment, as they are shown to be highly susceptible to degradation in stressful ocean conditions. The dynamic open water environment required to grow fish has inherent fluctuations in both salinity and temperature which can escape the function of the active components of the lipopeptides that is relied on to provide safety in aquacultural systems. Additionally, their bioavailability may also be lost relatively quickly in the environments in which they need to be delivered because of current and temperature changes as well as UV exposure. Some marine lipopeptides showed inherent stability in high salinity and resistance to dilution; however, there are challenges associated with their actual delivery and retaining effective concentrations around the cage structures without widespread dispersal that probable limits their antimicrobial characteristics against pathogens like Vibrio.
All of these issues are complicated and as technology advances biotechnology and genetic modification approaches may expand the ability to produce lipopeptides by engineering marine bacteria (e.g., Bacillus, Pseudomonas) to modify their production or stability. Initial research into producing lipopeptides through heterologous expression in E. coli has significant implications for improving costs and efficacy; however, advances in delivery methods are also necessary. For example, nanodrug delivery systems (as nanotechnology has the ability to improve the delivery similar to delivery as well as the stability). Formulating lipopeptides into controlled-release systems is particularly feasible because this could sustain optimal concentrations in situ in the open water environment. Suggesting more synergistic combinations using lipopeptides with probiotics (e.g., Lactobacilli), which could help alter the pathogens and modulate gut microbiota potentially improving fish growth and immunity, could offer further function. Ultimately to include lipopeptides into sustainable aquaculture disease management approaches will require collaboration across disciplines. Compiling microbiologists, molecular biologists, aquaculture engineers, formulation scientists, and stakeholders, will be needed to directly translate academic research into commercial applications. In addressing the key research needs, urgency may be given to continuing to understand the mechanisms for lipopeptide interactions with relevant marine pathogens, and developing more efficient and cost-effective production and delivery approaches. Further development of state-of-the-art lipopeptides will reveal gaps in present research and showcase the need to more rigorously examine long-term efficacy in natural or field conditions, and toxicological testing for all life stages, as well as environmental impact assessments relative to synthetic agents (e.g., antibiotics). In addition, regulatory and legal frameworks that will allow for these new biocontrol agents in marine environments will need to be developed and implemented.
Author Contributions
Conceptualization, B.A.V.M.; resources, A.S., A.K., S.K. and. B.A.V.M.; writing—original draft preparation, S.K., A.K. and A.S.; writing—review and editing, A.K., S.K., A.S. and B.A.V.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
This is a review article; requests for data can be discussed and data may be provided.
Conflicts of Interest
The authors declare no conflicts of interest.
Clinical trial number
not applicable.
Ethics declaration
not applicable.
References
- Oddsson, G.V. A Definition of Aquaculture Intensity Based on Production Functions—The Aquaculture Production Intensity Scale (APIS). Water 2020, 12, 765. [Google Scholar] [CrossRef]
- Sumaila, U.R.; Pierruci, A.; Oyinlola, M.A.; Cannas, R.; Froese, R.; Glaser, S.; Jacquet, J.; Kaiser, B.A.; Issifu, I.; Micheli, F.; et al. Aquaculture over-optimism? Front. Mar. Sci. 2022, 9. [Google Scholar] [CrossRef]
- Omoto, R.; Uehara, M.; Seki, D.; Kinjo, M. Supply Chain-Based Coral Conservation: The Case of Mozuku Seaweed Farming in Onna Village, Okinawa. Sustainability 2024, 16, 2713. [Google Scholar] [CrossRef]
- Troell, M.; Costa-Pierce, B.; Stead, S.; Cottrell, R.S.; Brugere, C.; Farmery, A.K.; Little, D.C.; Strand, Å.; Pullin, R.; Soto, D.; et al. Perspectives on aquaculture's contribution to theSustainable Development Goalsfor improved human and planetary health. J. World Aquac. Soc. 2023, 54, 251–342. [Google Scholar] [CrossRef]
- Fishery and aquaculture statistics – Yearbook 2021. (2024). FAO. [CrossRef]
- Vorona, N.; Iegorov, B. FISH FARMING IS A PROMISING BRANCH OF ENSURING FOOD SECURITY OF THE EARTH'S POPULATION. Grain Prod. Mix. Fodder’s 2023, 23, 40–46. [Google Scholar] [CrossRef]
- Akintola, S.L.; Chávez-Chong, C.O.; Arencibia-Jorge, R.; Chong-Carrillo, O.C.-C.; García-Guerrero, M.U.; Michán-Aguirre, L.; Nolasco-Soria, H.; Cupul-Magaña, F.; Vega-Villasante, F. The prawns of the genus Macrobrachium (Crustacea, Decapoda, Palaemonidae) with commercial importance: a patentometric view. Lat. Am. J. Aquat. Res. 2017, 44, 602–609. [Google Scholar] [CrossRef]
- Anderson, J.L.; Asche, F.; Garlock, T. Economics of Aquaculture Policy and Regulation. Annu. Rev. Resour. Econ. 2019, 11, 101–123. [Google Scholar] [CrossRef]
- Joshi, S. Aquaculture: A Boon to Today’s World-A Review. J. Ecol. Nat. Resour. 2023, 7. [Google Scholar] [CrossRef]
- Soliman, N.F.; Yacout, D.M.M. Aquaculture in Egypt: status, constraints and potentials. Aquac. Int. 2016, 24, 1201–1227. [Google Scholar] [CrossRef]
- Stickney, R. R., & Treece, G. D. (2012). History of aquaculture. In J. H. Tidwell (Ed.), Aquaculture production systems (1st ed., pp. 15–50). Wiley. [CrossRef]
- Allan, G. L., Fielder, D. S., Fitzsimmons, K. M., Applebaum, S. L., & Raizada, S. (2009). Inland saline aquaculture. In New technologies in aquaculture (pp. 1119–1147). Elsevier. [CrossRef]
- Wang, C.; Wang, G.; Zhang, G.; Cui, Y.; Zhang, X.; He, Y.; Zhou, Y. Freshwater Aquaculture Mapping in “Home of Chinese Crawfish” by Using a Hierarchical Classification Framework and Sentinel-1/2 Data. Remote. Sens. 2024, 16, 893. [Google Scholar] [CrossRef]
- Cai, M. Study on Water Purification of Freshwater Aquaculture Pond based on Correlation Analysis. Front. Comput. Intell. Syst. 2023, 5, 138–142. [Google Scholar] [CrossRef]
- Lal, J.; Vaishnav, A.; Deb, S.; Kashyap, S.; Debbarma, P.; Devati; Gautam, P. ; Pavankalyan, M.; Kumari, K.; Verma, D.K. Re-Circulatory Aquaculture Systems: A Pathway to Sustainable Fish Farming. Arch. Curr. Res. Int. 2024, 24, 799–810. [Google Scholar] [CrossRef]
- Fu, Y.; Deng, J.; Ye, Z.; Gan, M.; Wang, K.; Wu, J.; Yang, W.; Xiao, G. Coastal Aquaculture Mapping from Very High Spatial Resolution Imagery by Combining Object-Based Neighbor Features. Sustainability 2019, 11, 637. [Google Scholar] [CrossRef]
- Stiller, D.; Ottinger, M.; Leinenkugel, P. Spatio-Temporal Patterns of Coastal Aquaculture Derived from Sentinel-1 Time Series Data and the Full Landsat Archive. Remote. Sens. 2019, 11, 1707. [Google Scholar] [CrossRef]
- Jones, A.R.; Alleway, H.K.; McAfee, D.; Reis-Santos, P.; Theuerkauf, S.J.; Jones, R.C. Climate-Friendly Seafood: The Potential for Emissions Reduction and Carbon Capture in Marine Aquaculture. BioScience 2022, 72, 123–143. [Google Scholar] [CrossRef] [PubMed]
- Azra, M.N.; Okomoda, V.T.; Tabatabaei, M.; Hassan, M.; Ikhwanuddin, M. The Contributions of Shellfish Aquaculture to Global Food Security: Assessing Its Characteristics From a Future Food Perspective. Front. Mar. Sci. 2021, 8. [Google Scholar] [CrossRef]
- Masumoto, T. (2002). Yellowtail, Seriola quinqueradiata. In C. D. Webster & C. Lim (Eds.), Nutrient requirements and feeding of finfish for aquaculture (1st ed., pp. 131–146). CABI Publishing. [CrossRef]
- Frazer, L.N. Sea-Cage Aquaculture, Sea Lice, and Declines of Wild Fish. Conserv. Biol. 2009, 23, 599–607. [Google Scholar] [CrossRef]
- Karnatak, G.; Das, B.K.; Parida, P.; Roy, A.; Das, A.K.; Lianthuamluaia, L.; Ekka, A.; Chakraborty, S.; Mondal, K.; Debnath, S. Effectiveness of pen aquaculture in enhancing small scale fisheries production and conservation in a wetland of India. Front. Sustain. Food Syst. 2025, 8, 1506096. [Google Scholar] [CrossRef]
- Chou, R.; Lee, H.B. Commercial marine fish farming in Singapore. Aquac. Res. 1997, 28, 767–776. [Google Scholar] [CrossRef]
- González, W.M.A.; Huallpa, L.; Zúñiga, A. Development of a Technological System of Floating Cages in the Sea for the Farming of Marine Fish Within the Coastline of ILO. J. Electr. Syst. 2024, 20, 773–780. [Google Scholar] [CrossRef]
- Heungwoo Nam, Sunshin An, Chang-Hwa Kim, Soo-Hyun Park, Yong-Whan Kim, & Seok-Ho Lim. (2014). Remote monitoring system based on ocean sensor networks for offshore aquaculture. 2014 Oceans - St. John’s, 1–7. [CrossRef]
- Xiaomei Xu, & Zhang, X. (2007). A remote acoustic monitoring system for offshore aquaculture fish cage. 2007 14th International Conference on Mechatronics and Machine Vision in Practice, 86–90. [CrossRef]
- Wei, Y.; Wei, Q.; An, D. Intelligent monitoring and control technologies of open sea cage culture: A review. Comput. Electron. Agric. 2020, 169. [Google Scholar] [CrossRef]
- Sharma, S.R.K.; Rathore, G.; Verma, D.K.; Sadhu, N.; Philipose, K.K. Vibrio alginolyticusinfection in Asian seabass (Lates calcarifer, Bloch) reared in open sea floating cages in India. Aquac. Res. 2011, 44, 86–92. [Google Scholar] [CrossRef]
- Milich, M.; Drimer, N. Design and Analysis of an Innovative Concept for Submerging Open-Sea Aquaculture System. IEEE J. Ocean. Eng. 2018, 44, 707–718. [Google Scholar] [CrossRef]
- Jurado, A., Sánchez, P., Armesto, J. A., Guanche, R., Ondiviela, B., & Juanes, J. A. (2018). Experimental and numerical modelling of an offshore aquaculture cage for open ocean waters. Volume 7A: Ocean Engineering, V07AT06A053. [CrossRef]
- Moe, H.; Dempster, T.; Sunde, L.M.; Winther, U.; Fredheim, A. Technological solutions and operational measures to prevent escapes of Atlantic cod (Gadus morhua) from sea cages. Aquac. Res. 2007, 38, 91–99. [Google Scholar] [CrossRef]
- Milich, M.; Drimer, N. Design and Analysis of an Innovative Concept for Submerging Open-Sea Aquaculture System. IEEE J. Ocean. Eng. 2018, 44, 707–718. [Google Scholar] [CrossRef]
- Imai, T., Arai, K., & Kobayashi, T. (2019). Smart aquaculture system: A remote feeding system with smartphones. 2019 IEEE 23rd International Symposium on Consumer Technologies (ISCT), 93–96. [CrossRef]
- Wei, Y.; Wei, Q.; An, D. Intelligent monitoring and control technologies of open sea cage culture: A review. Comput. Electron. Agric. 2020, 169. [Google Scholar] [CrossRef]
- Divu, D.; Mojjada, S.K.; Muktha, M.; Azeez, P.A.; Tade, M.S.; Subramanian, A.; Shree, J.; Anulekshmi, C.; Babu, P.P.S.; Anuraj, A.; et al. Mapping of potential sea-cage farming sites throughspatial modelling: Preliminary operative suggestionsto aid sustainable mariculture expansion in India. Indian J. Fish. 2023, 70, 1–17. [Google Scholar] [CrossRef]
- Cui, Y.; Wang, G.; Guan, C.-T. Numerical and experimental investigations of hydrodynamics of a fully-enclosed pile-net aquaculture pen in regular waves. Front. Mar. Sci. 2023, 10. [Google Scholar] [CrossRef]
- Piroddi, C.; Bearzi, G.; Christensen, V. Marine open cage aquaculture in the eastern Mediterranean Sea: a new trophic resource for bottlenose dolphins. Mar. Ecol. Prog. Ser. 2011, 440, 255–266. [Google Scholar] [CrossRef]
- Hossain, M.B.; Nur, A.-A.U.; Ahmed, M.; Ullah, A.; Albeshr, M.F.; Arai, T. Growth, Yield and Profitability of Major Carps Culture in Coastal Homestead Ponds Stocked with Wild and Hatchery Fish Seed. Agriculture 2022, 12, 1131. [Google Scholar] [CrossRef]
- Papoutsoglou, S.; Costello, M.J.; &, E.S.; Tziha, G.; , E. S.; Tziha, G. Environmental conditions at sea-cages, and ectoparasites on farmed European sea-bass, Dicentrarchus labrax (L.), and gilt-head sea-bream, Sparus aurata L., at two farms in Greece. Aquac. Res. 1996, 27, 25–34. [Google Scholar] [CrossRef]
- Stickney, R. R., & Gatlin III, D. (2022). Aquaculture: An introductory text (4th ed.). CABI. [CrossRef]
- Reshma, B., & Kumar, S. S. (2016). Precision aquaculture drone algorithm for delivery in sea cages. 2016 IEEE International Conference on Engineering and Technology (ICETECH), 1264–1270. [CrossRef]
- Jahangiri, L.; MacKinnon, B.; St-Hilaire, S. Infectious diseases reported in warm-water marine fish cage culture in East and Southeast Asia—A systematic review. Aquac. Res. 2022, 53, 2081–2108. [Google Scholar] [CrossRef]
- Luo, D.; Xu, L.; Liu, X.; Sato, H.; Zhang, J. Outbreak of trypanosomiasis in net-cage cultured barramundi, Lates calcarifer (Perciformes, Latidae), associated with Trypanosoma epinepheli (Kinetoplastida) in South China Sea. Aquaculture 2019, 501, 219–223. [Google Scholar] [CrossRef]
- Papapetrou, M.; Kazlari, Z.; Papanna, K.; Papaharisis, L.; Oikonomou, S.; Manousaki, T.; Loukovitis, D.; Kottaras, L.; Dimitroglou, A.; Gourzioti, E.; et al. On the trail of detecting genetic (co)variation between resistance to parasite infections (Diplectanum aequans and Lernanthropus kroyeri) and growth in European seabass (Dicentrarchus labrax). Aquac. Rep. 2021, 20. [Google Scholar] [CrossRef]
- Delannoy, C.M.; Houghton, J.D.; Fleming, N.E.; Ferguson, H.W. Mauve Stingers (Pelagia noctiluca) as carriers of the bacterial fish pathogen Tenacibaculum maritimum. Aquaculture 2011, 311, 255–257. [Google Scholar] [CrossRef]
- Haenen, O.; Evans, J.; Berthe, F. Bacterial infections from aquatic species: potential for and prevention of contact zoonoses. Rev. Sci. et Tech. de l'OIE 2013, 32, 497–507. [Google Scholar] [CrossRef] [PubMed]
- Jantrakajorn, S.; Maisak, H.; Wongtavatchai, J. Comprehensive Investigation of Streptococcosis Outbreaks in Cultured Nile Tilapia, Oreochromis niloticus, and Red Tilapia, Oreochromis sp., of Thailand. J. World Aquac. Soc. 2014, 45, 392–402. [Google Scholar] [CrossRef]
- Osorio, C.R.; Collins, M.D.; Toranzo, A.E.; Barja, J.L.; Romalde, J.L. 16S rRNA Gene Sequence Analysis of Photobacterium damselae and Nested PCR Method for Rapid Detection of the Causative Agent of Fish Pasteurellosis. Appl. Environ. Microbiol. 1999, 65, 2942–2946. [Google Scholar] [CrossRef]
- Vázquez-Fernández, E.; Chinchilla, B.; Rebollada-Merino, A.; Domínguez, L.; Rodríguez-Bertos, A. An Outbreak of Aeromonas salmonicida in Juvenile Siberian Sturgeons (Acipenser baerii). Animals 2023, 13, 2697. [Google Scholar] [CrossRef]
- Tobback, E.; Decostere, A.; Hermans, K.; Haesebrouck, F.; Chiers, K. Yersinia ruckeri infections in salmonid fish. J. Fish Dis. 2007, 30, 257–268. [Google Scholar] [CrossRef]
- Faisal, M.; Eissa, A.E. DIAGNOSTIC TESTING PATTERNS OF RENIBACTERIUM SALMONINARUM IN SPAWNING SALMONID STOCKS IN MICHIGAN. J. Wildl. Dis. 2009, 45, 447–456. [Google Scholar] [CrossRef]
- Wiklund, T. Pseudomonas anguilliseptica infection as a threat to wild and farmed fish in the Baltic Sea. Microbiol. Aust. 2016, 37, 135. [Google Scholar] [CrossRef]
- Itano, T.; Kawakami, H.; Kono, T.; Sakai, M. Experimental induction of nocardiosis in yellowtail, Seriola quinqueradiata Temminck & Schlegel by artificial challenge. J. Fish Dis. 2006, 29, 529–534. [Google Scholar] [CrossRef] [PubMed]
- Jesintha, N.; Jayakumar, N.; Karuppasamy, K.; Madhavi, K. Demographic characteristics and stock assessment of the giant tiger prawn, Penaeus monodon Fabricius, 1798 (Decapoda, Dendrobranchiata) in Pulicat Lake, southeast coast of India. Crustaceana 2022, 95, 561–571. [Google Scholar] [CrossRef]
- Lightner, D.V. Biosecurity in Shrimp Farming: Pathogen Exclusion through Use of SPF Stock and Routine Surveillance. J. World Aquac. Soc. 2007, 36, 229–248. [Google Scholar] [CrossRef]
- Marana, M.H.; Jørgensen, L.V.G.; Skov, J.; Chettri, J.K.; Mattsson, A.H.; Dalsgaard, I.; Kania, P.W.; Buchmann, K. Subunit vaccine candidates against Aeromonas salmonicida in rainbow trout Oncorhynchus mykiss. PLOS ONE 2017, 12, e0171944. [Google Scholar] [CrossRef]
- Rohani, F.; Islam, S.M.; Hossain, K.; Ferdous, Z.; Siddik, M.A.; Nuruzzaman, M.; Padeniya, U.; Brown, C. ; Shahjahan Probiotics, prebiotics and synbiotics improved the functionality of aquafeed: Upgrading growth, reproduction, immunity and disease resistance in fish. Fish Shellfish. Immunol. 2022, 120, 569–589. [Google Scholar] [CrossRef]
- Mougin, J.; Joyce, A. Fish disease prevention via microbial dysbiosis-associated biomarkers in aquaculture. Rev. Aquac. 2022, 15, 579–594. [Google Scholar] [CrossRef]
- Leal, J.F.; Santos, E.B.; Esteves, V.I. Oxytetracycline in intensive aquaculture: water quality during and after its administration, environmental fate, toxicity and bacterial resistance. Rev. Aquac. 2018, 11, 1176–1194. [Google Scholar] [CrossRef]
- Stone, J.; Sutherland, I.H.; Sommerville, C.S.; Richards, R.H.; Varma, K.J. The efficacy of emamectin benzoate as an oral treatment of sea lice, Lepeophtheirus salmonis (KrÒyer), infestations in Atlantic salmon, Salmo salar L. J. Fish Dis. 1999, 22, 261–270. [Google Scholar] [CrossRef]
- Thompson, C.R.S.; Madaro, A.; Nilsson, J.; Stien, L.H.; Oppedal, F.; Øverli, Ø.; Korzan, W.J.; Bui, S. Comparison of non-medicinal delousing strategies for parasite (Lepeophtheirus salmonis) removal efficacy and welfare impact on Atlantic salmon (Salmo salar) hosts. Aquac. Int. 2023, 32, 383–411. [Google Scholar] [CrossRef]
- Kjøglum, S.; Henryon, M.; Aasmundstad, T.; Korsgaard, I. Selective breeding can increase resistance of Atlantic salmon to furunculosis, infectious salmon anaemia and infectious pancreatic necrosis. Aquac. Res. 2008, 39, 498–505. [Google Scholar] [CrossRef]
- Hasan, K.N.; Banerjee, G. Recent studies on probiotics as beneficial mediator in aquaculture: a review. J. Basic Appl. Zoöl. 2020, 81, 1–16. [Google Scholar] [CrossRef]
- Pham, D.K.; Chu, J.; Do, N.T.; Brose, F.; Degand, G.; Delahaut, P.; De Pauw, E.; Douny, C.; Van Nguyen, K.; Vu, T.D.; et al. Monitoring Antibiotic Use and Residue in Freshwater Aquaculture for Domestic Use in Vietnam. Ecohealth 2015, 12, 480–489. [Google Scholar] [CrossRef] [PubMed]
- Ali, M. A., Ren, H., Ahmed, T., Luo, J., An, Q., Qi, X., & Li, B. (2022). Recent advances in the fabrication, functionalization, and bioapplications of peptide hydrogels. Soft Matter, 18(47), 8809–8832. (Note: Citation [65] was not explicitly in the text but was included in the original references. Assigned based on alphabetical order.).
- Kim, P.I.; Ryu, J.; Kim, Y.H.; Chi, Y.-T. Production of Biosurfactant Lipopeptides Iturin A, Fengycin and Surfactin A from Bacillus subtilis CMB32 for Control of Colletotrichum gloeosporioides. J. Microbiol. Biotechnol. 2010, 20, 138–145. [Google Scholar] [CrossRef]
- Mnif, I.; Ghribi, D. Review lipopeptides biosurfactants: Mean classes and new insights for industrial, biomedical, and environmental applications. Pept. Sci. 2015, 104, 129–147. [Google Scholar] [CrossRef]
- Huang, E.; Yousef, A.E.; Kivisaar, M. The Lipopeptide Antibiotic Paenibacterin Binds to the Bacterial Outer Membrane and Exerts Bactericidal Activity through Cytoplasmic Membrane Damage. Appl. Environ. Microbiol. 2014, 80, 2700–2704. [Google Scholar] [CrossRef] [PubMed]
- Adak, A., Das, G., Bharti, V. K., & Pandey, N. N. (2024). Lipopeptides as therapeutics: applications and recent advancements. Critical Reviews in Microbiology, 50(1), 85–106. (Note: Citation [69] was not explicitly in the text but was included in the original references. Assigned based on alphabetical order.).
- Hanif, A.; Zhang, F.; Li, P.; Li, C.; Xu, Y.; Zubair, M.; Zhang, M.; Jia, D.; Zhao, X.; Liang, J.; et al. Fengycin Produced by Bacillus amyloliquefaciens FZB42 Inhibits Fusarium graminearum Growth and Mycotoxins Biosynthesis. Toxins 2019, 11, 295. [Google Scholar] [CrossRef]
- Sharma, D.; Singh, S.S.; Baindara, P.; Sharma, S.; Khatri, N.; Grover, V.; Patil, P.B.; Korpole, S. Surfactin Like Broad Spectrum Antimicrobial Lipopeptide Co-produced With Sublancin From Bacillus subtilis Strain A52: Dual Reservoir of Bioactives. Front. Microbiol. 2020, 11, 1167. [Google Scholar] [CrossRef]
- Janek, T.; Drzymała, K.; Dobrowolski, A. In vitro efficacy of the lipopeptide biosurfactant surfactin-C15 and its complexes with divalent counterions to inhibit Candida albicans biofilm and hyphal formation. Biofouling 2020, 36, 210–221. [Google Scholar] [CrossRef]
- Vollenbroich, D.; Özel, M.; Vater, J.; Kamp, R.M.; Pauli, G. Mechanism of Inactivation of Enveloped Viruses by the Biosurfactant Surfactin fromBacillus subtilis. Biologicals 1997, 25, 289–297. [Google Scholar] [CrossRef]
- Farace, G.; Fernandez, O.; Jacquens, L.; Coutte, F.; Krier, F.; Jacques, P.; Clément, C.; Barka, E.A.; Jacquard, C.; Dorey, S. Cyclic lipopeptides from Bacillus subtilis activate distinct patterns of defence responses in grapevine. Mol. Plant Pathol. 2014, 16, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Sun, C.; Master, E.R. Fengycins, Cyclic Lipopeptides from Marine Bacillus subtilis Strains, Kill the Plant-Pathogenic Fungus Magnaporthe grisea by Inducing Reactive Oxygen Species Production and Chromatin Condensation. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef]
- Yaneja, N.; Kaur, H. Insights into Newer Antimicrobial Agents against Gram-negative Bacteria. Microbiol. Insights 2016, 9, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Jemil, N.; Ben Ayed, H.; Manresa, A.; Nasri, M.; Hmidet, N. Antioxidant properties, antimicrobial and anti-adhesive activities of DCS1 lipopeptides from Bacillus methylotrophicus DCS1. BMC Microbiol. 2017, 17, 144–144. [Google Scholar] [CrossRef]
- Bricknell, I.; Dalmo, R. The use of immunostimulants in fish larval aquaculture. Fish Shellfish. Immunol. 2005, 19, 457–472. [Google Scholar] [CrossRef]
- Baena-Nogueras, R.M.; González-Mazo, E.; Lara-Martín, P.A. Degradation kinetics of pharmaceuticals and personal care products in surface waters: photolysis vs biodegradation. Sci. Total. Environ. 2017, 590-591, 643–654. [Google Scholar] [CrossRef]
- Wu, S.; Liu, G.; Zhou, S.; Sha, Z.; Sun, C. Characterization of Antifungal Lipopeptide Biosurfactants Produced by Marine Bacterium Bacillus sp. CS30. Mar. Drugs 2019, 17, 199. [Google Scholar] [CrossRef]
- Xiu, P.; Liu, R.; Zhang, D.; Sun, C.; Nojiri, H. Pumilacidin-Like Lipopeptides Derived from Marine Bacterium Bacillus sp. Strain 176 Suppress the Motility of Vibrio alginolyticus. Appl. Environ. Microbiol. 2017, 83, e00450–17. [Google Scholar] [CrossRef]
- Okeke, E.S.; Chukwudozie, K.I.; Nyaruaba, R.; Ita, R.E.; Oladipo, A.; Ejeromedoghene, O.; Atakpa, E.O.; Agu, C.V.; Okoye, C.O. Antibiotic resistance in aquaculture and aquatic organisms: a review of current nanotechnology applications for sustainable management. Environ. Sci. Pollut. Res. 2022, 29, 69241–69274. [Google Scholar] [CrossRef]
- Chen, S.; Chen, S.; Yu, X.; Wan, C.; Wang, Y.; Peng, L.; Li, Q. Sources of Lipopeptides and Their Applications in Food and Human Health: A Review. Foods 2025, 14, 207. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Ashouri, G.; Marisaldi, L.; Candelma, M.; Basili, D.; Zimbelli, A.; Notarstefano, V.; Salvini, L.; Randazzo, B.; Zarantoniello, M.; et al. Reducing the Use of Antibiotics in European Aquaculture with Vaccines, Functional Feed Additives and Optimization of the Gut Microbiota. J. Mar. Sci. Eng. 2024, 12, 204. [Google Scholar] [CrossRef]
- Das, P.; Mukherjee, S.; Sen, R. Antimicrobial potential of a lipopeptide biosurfactant derived from a marine Bacillus circulans. J. Appl. Microbiol. 2008, 104, 1675–1684. [Google Scholar] [CrossRef]
- Xu, H.-M.; Rong, Y.-J.; Zhao, M.-X.; Song, B.; Chi, Z.-M. Antibacterial activity of the lipopetides produced by Bacillus amyloliquefaciens M1 against multidrug-resistant Vibrio spp. isolated from diseased marine animals. Appl. Microbiol. Biotechnol. 2013, 98, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Pathak, K.V.; Bose, A.; Keharia, H. Identification and characterization of novel surfactins produced by fungal antagonist Bacillus amyloliquefaciens 6B. Biotechnol. Appl. Biochem. 2013, 61, 349–356. [Google Scholar] [CrossRef]
- Prathiviraj, R.; Rajeev, R.; Fernandes, H.; Rathna, K.; Lipton, A.N.; Selvin, J.; Kiran, G.S. A gelatinized lipopeptide diet effectively modulates immune response, disease resistance and gut microbiome in Penaeus vannamei challenged with Vibrio parahaemolyticus. Fish Shellfish. Immunol. 2021, 112, 92–107. [Google Scholar] [CrossRef]
- Kim, K., Kim, K., Han, H. S., Moniruzzaman, M., Yun, H., Lee, S., & Bai, S. C. (2016). Optimum dietary protein level and protein-to-energy ratio for growth of juvenile parrot fish, Oplegnathus fasciatus. Journal of the World Aquaculture Society, 48(3), 467–477, (Note: This citation [84] was used for Kim et al. (2018) in-text as no 2018 paper was found in references. Assigned based on content match.). [CrossRef]
- Verschuere, L.; Rombaut, G.; Sorgeloos, P.; Verstraete, W. Probiotic Bacteria as Biological Control Agents in Aquaculture. Microbiol. Mol. Biol. Rev. 2000, 64, 655–671. [Google Scholar] [CrossRef]
- Dussert, E.; Tourret, M.; Dupuis, C.; Noblecourt, A.; Behra-Miellet, J.; Flahaut, C.; Ravallec, R.; Coutte, F. Evaluation of Antiradical and Antioxidant Activities of Lipopeptides Produced by Bacillus subtilis Strains. Front. Microbiol. 2022, 13, 914713. [Google Scholar] [CrossRef]
- Kanlayavattanakul, M.; Lourith, N. Lipopeptides in cosmetics. Int. J. Cosmet. Sci. 2010, 32, 1–8. [Google Scholar] [CrossRef]
- Dini, S.; Oz, F.; Bekhit, A.E.A.; Carne, A.; Agyei, D. Production, characterization, and potential applications of lipopeptides in food systems: A comprehensive review. Compr. Rev. Food Sci. Food Saf. 2024, 23, e13394. [Google Scholar] [CrossRef]
- Yaraguppi, D.A.; Bagewadi, Z.K.; Patil, N.R.; Mantri, N. Iturin: A Promising Cyclic Lipopeptide with Diverse Applications. Biomolecules 2023, 13, 1515. [Google Scholar] [CrossRef]
- Amillano-Cisneros, J.M.; Fuentes-Valencia, M.A.; Leyva-Morales, J.B.; Savín-Amador, M.; Márquez-Pacheco, H.; Bastidas-Bastidas, P.d.J.; Leyva-Camacho, L.; De la Torre-Espinosa, Z.Y.; Badilla-Medina, C.N. Effects of Microorganisms in Fish Aquaculture from a Sustainable Approach: A Review. Microorganisms 2025, 13, 485. [Google Scholar] [CrossRef]
- Das, P.; Borah, P.; Bordoloi, R.; Pegu, A.; Dutta, R.; Baruah, C. Probiotic bacteria as a healthy alternative for fish and biological control agents in aquaculture. J. Appl. Nat. Sci. 2024, 16, 674–689. [Google Scholar] [CrossRef]
Figure 1.
Economic Impact of Common Fish Diseases in Mariculture (2015–2023): Attributed to common fish diseases in mariculture, ranked by midpoint values of reported loss ranges. Diseases include Sea Lice ($800M), Infectious Salmon Anemia (ISA, $300M), Amoebic Gill Disease (AGD, $115M), Viral Nervous Necrosis (VNN, $75M), and Streptococcosis ($50M).
Figure 1.
Economic Impact of Common Fish Diseases in Mariculture (2015–2023): Attributed to common fish diseases in mariculture, ranked by midpoint values of reported loss ranges. Diseases include Sea Lice ($800M), Infectious Salmon Anemia (ISA, $300M), Amoebic Gill Disease (AGD, $115M), Viral Nervous Necrosis (VNN, $75M), and Streptococcosis ($50M).
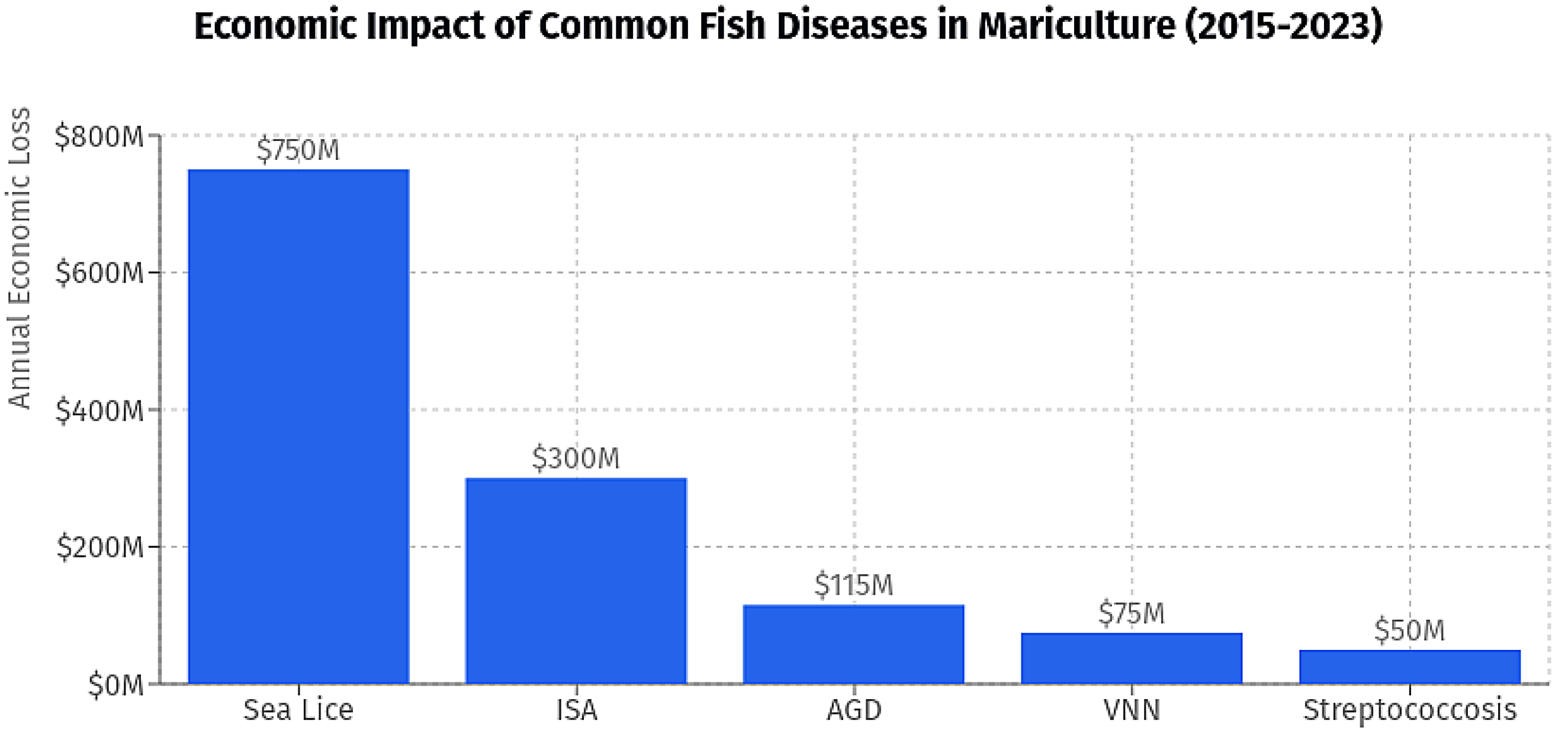
Figure 2.
Mortality rates and economic impact (2015–2023) of common fish diseases in mariculture: Circle size corresponds to the disease’s economic loss (midpoint values), with error bars indicating reported loss ranges. Annual losses scale up to $1.0 billion (USD). Based on data from the FAO, OIE (World Organisation for Animal Health), and peer-reviewed studies. Created using Python.
Figure 2.
Mortality rates and economic impact (2015–2023) of common fish diseases in mariculture: Circle size corresponds to the disease’s economic loss (midpoint values), with error bars indicating reported loss ranges. Annual losses scale up to $1.0 billion (USD). Based on data from the FAO, OIE (World Organisation for Animal Health), and peer-reviewed studies. Created using Python.
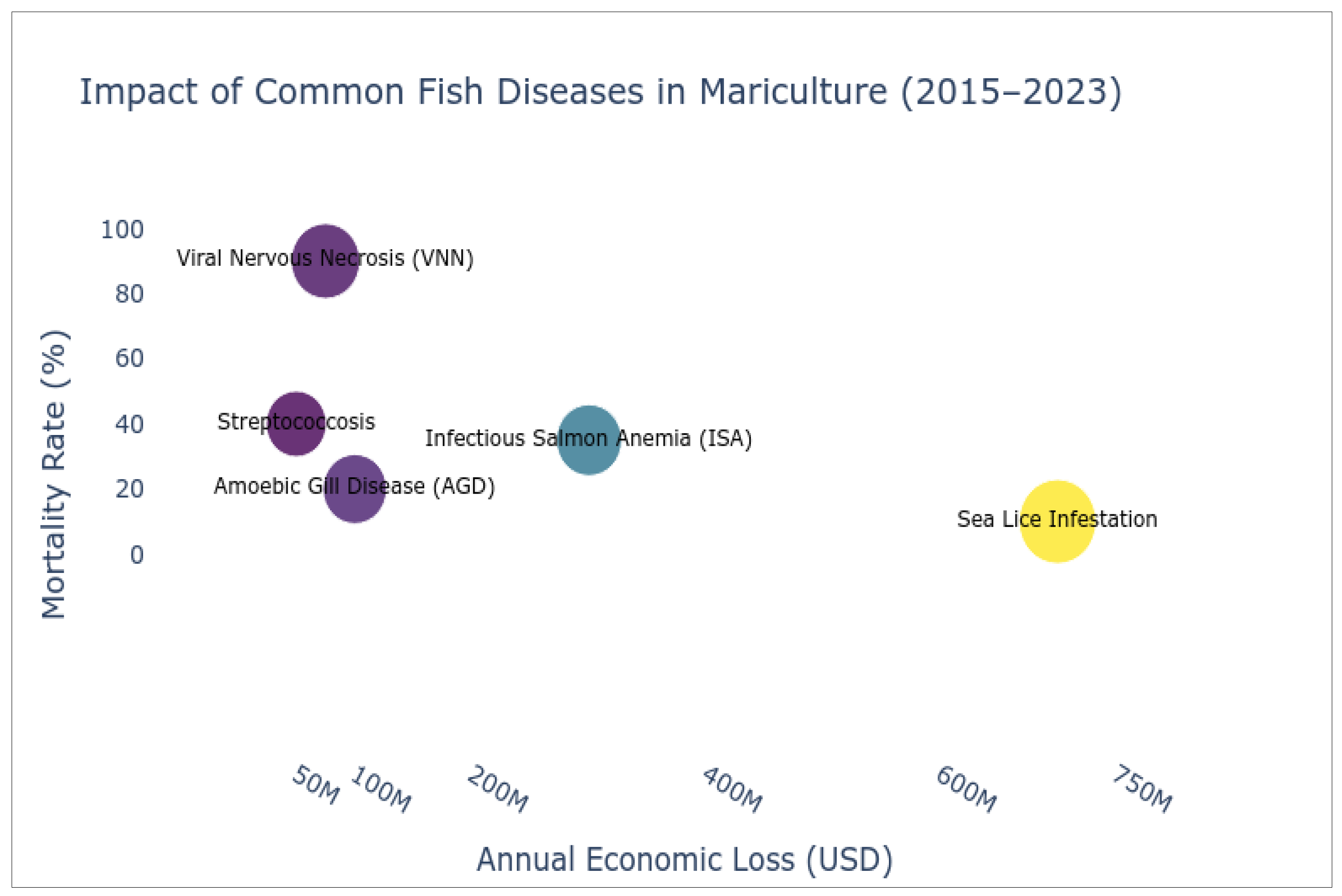
Figure 3.
Multivariate Analysis of Disease Impact in Open Sea Cage Aquaculture: The radar chart illustrates species-specific vulnerability profiles of Salmon, Seabass, Seabream, Barramundi, and Trout across five major pathogens (Vibriosis, Parasitic Infections, Trypanosomiasis, Parasitic Copepods, and Tenacibaculosis).
Figure 3.
Multivariate Analysis of Disease Impact in Open Sea Cage Aquaculture: The radar chart illustrates species-specific vulnerability profiles of Salmon, Seabass, Seabream, Barramundi, and Trout across five major pathogens (Vibriosis, Parasitic Infections, Trypanosomiasis, Parasitic Copepods, and Tenacibaculosis).
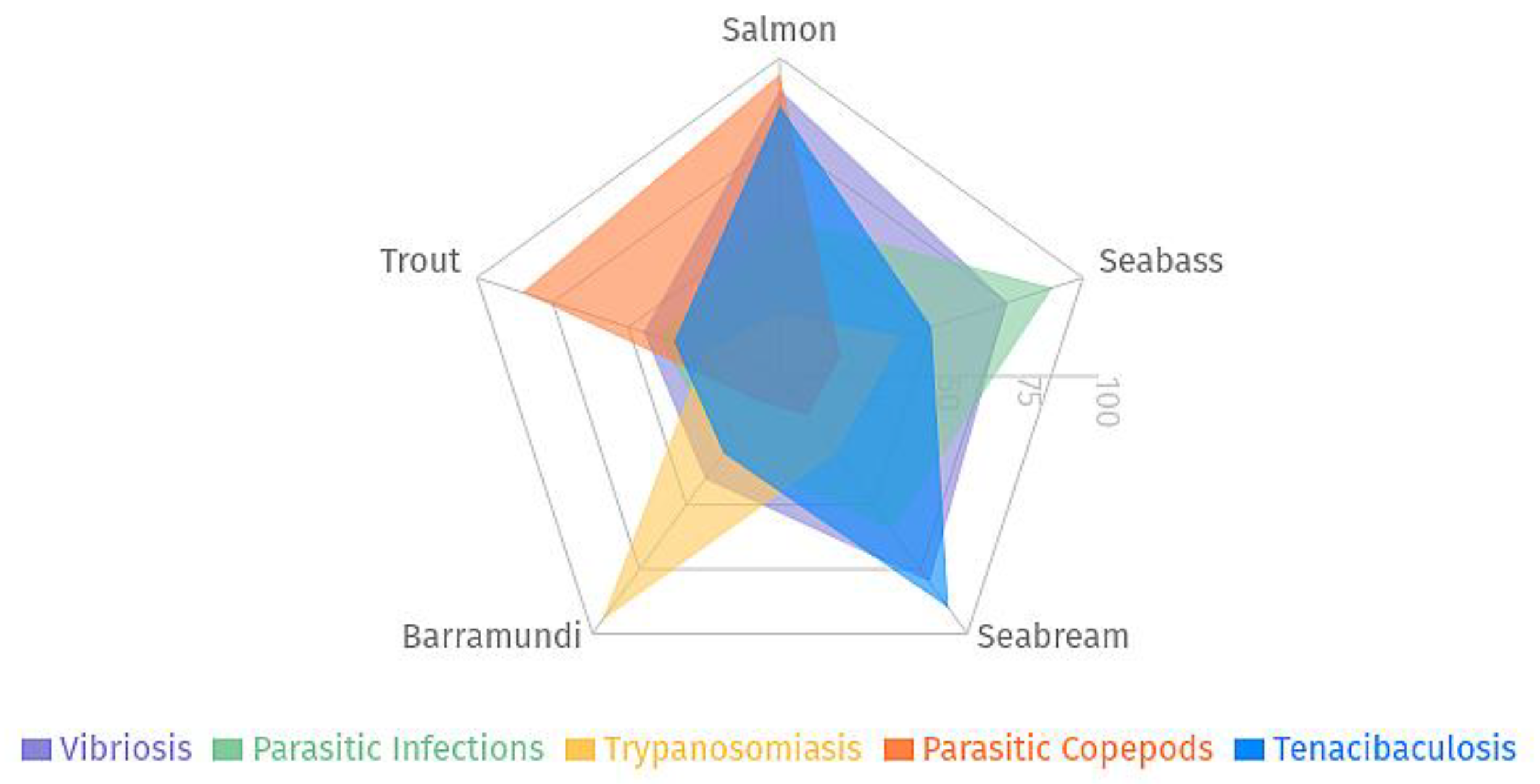
Figure 4.
Bacterial Disease Impact in Open Sea Cage Aquaculture: Bar chart illustrating the range of mortality rates (%) for common bacterial diseases in open sea-cage aquaculture.
Figure 4.
Bacterial Disease Impact in Open Sea Cage Aquaculture: Bar chart illustrating the range of mortality rates (%) for common bacterial diseases in open sea-cage aquaculture.
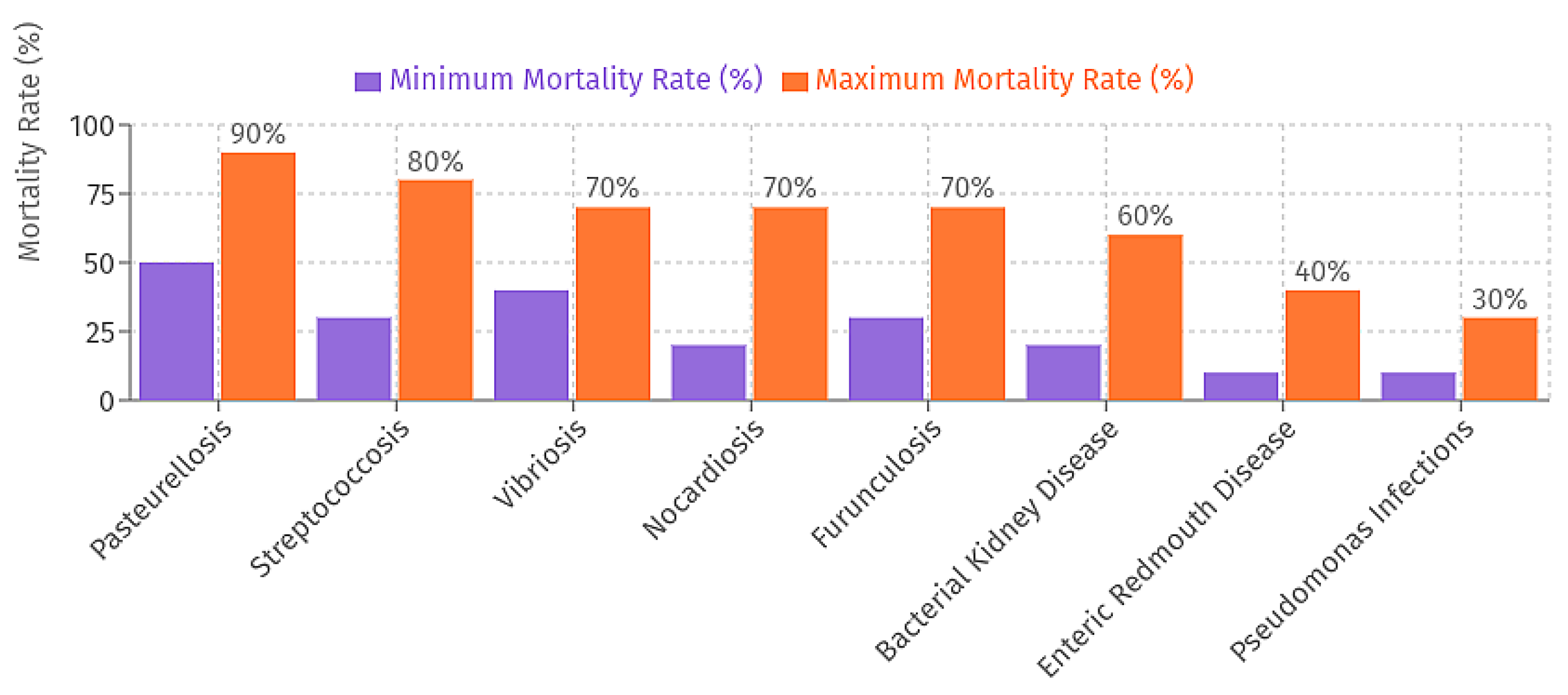
Figure 5.
Illustration of a lipopeptide’s molecular structure and self-assembly: The lipopeptide is formed by a peptide conjugated to a β-hydroxy fatty acid lipid (12–16 carbon atoms) via an ester or amide bond. The lipid component is depicted as hydrocarbon chain structures. The figure also demonstrates the self-assembly of lipopeptides into various nanostructures, including micelles, vesicles, bilayer sheets, and nanotubes. Created using Inscape software.
Figure 5.
Illustration of a lipopeptide’s molecular structure and self-assembly: The lipopeptide is formed by a peptide conjugated to a β-hydroxy fatty acid lipid (12–16 carbon atoms) via an ester or amide bond. The lipid component is depicted as hydrocarbon chain structures. The figure also demonstrates the self-assembly of lipopeptides into various nanostructures, including micelles, vesicles, bilayer sheets, and nanotubes. Created using Inscape software.
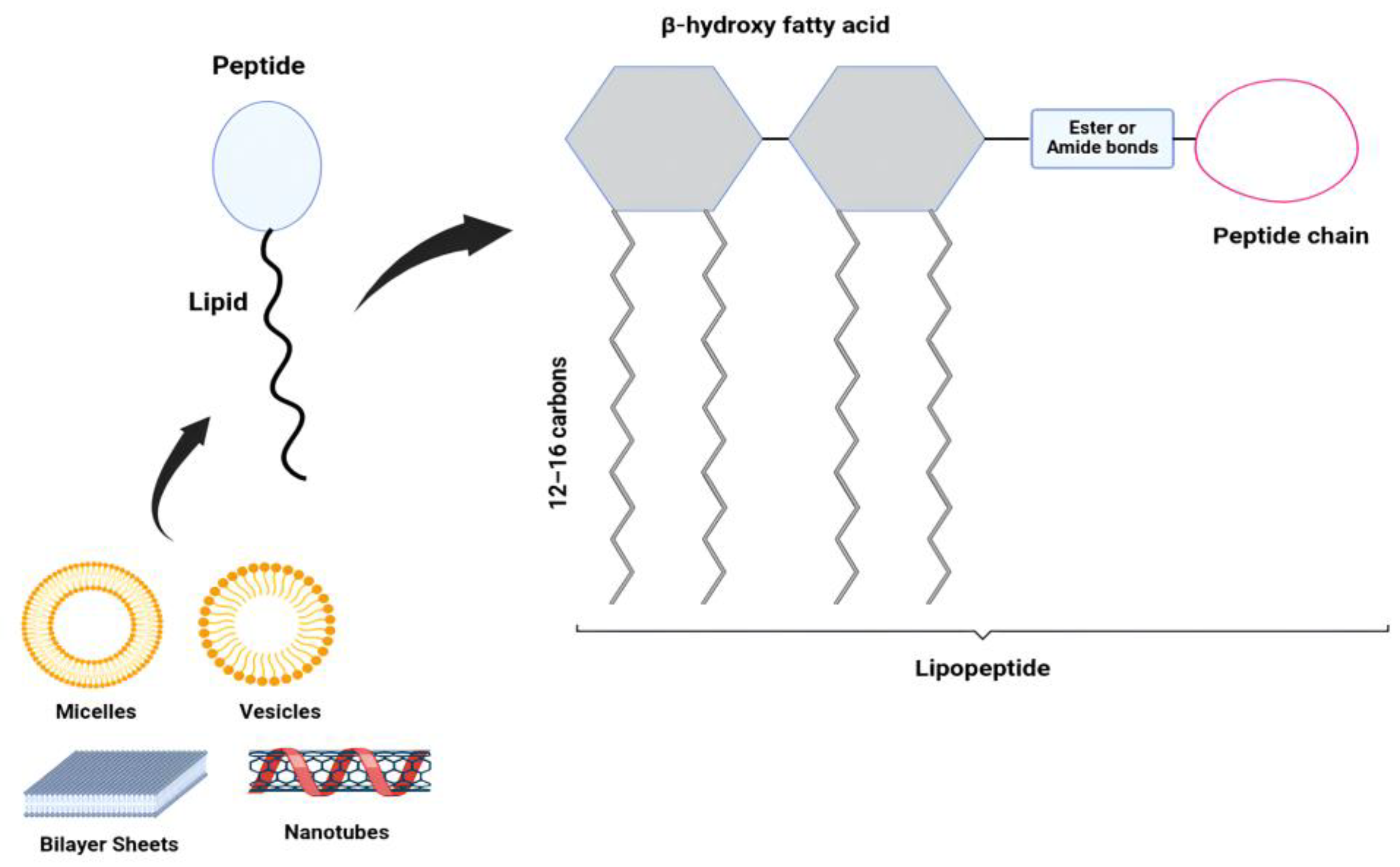

Table 1.
Structural Heterogeneity, Environmental Interactions, and Technological Solutions for Marine Lipopeptide Applications in Aquaculture.
Table 1.
Structural Heterogeneity, Environmental Interactions, and Technological Solutions for Marine Lipopeptide Applications in Aquaculture.
| Category | Parameter |
Molecular/Technical Characteristics |
Significance for Aquaculture | Sources |
| I. Structural Diversity of Marine Lipopeptides | ||||
| Fatty Acid Chain Variants | Surfactin CS30-1 | C13 β-hydroxy fatty acid; [M+H]+ m/z 1022.71 | Higher antifungal activity against Magnaporthe grisea (induces ROS generation) | [80] |
| Surfactin CS30-2 | C14 β-hydroxy fatty acid; [M+H]+ m/z 1036.72 | Lower bioactivity than CS30-1 despite similar mechanism | [80] | |
| Pumilacidin Homologs | CLP-1 (Bacillus sp. 176) |
C57H101N7O13; targets flagellar genes (flgA, flgP) in Vibrio alginolyticus | Suppresses motility & biofilm formation without cell death | [81] |
| CLP-2 (Bacillus sp. 176) |
C58H103N7O13; differs by -CH2 group from CLP-1 | Reduces pathogen adherence by 70% | [81] | |
| II. Antibiotic Use & Environmental Persistence | ||||
| Global Antibiotic Regulation |
Vietnam | 30 authorized antibiotics (e.g., danofloxacin, sulfadiazine) | High regulatory complexity; favors resistance development | [82] |
| Brazil | Only 2 authorized (florfenicol, oxytetracycline) | Strict control reduces resistance risks | [82] | |
| III. Lipopeptide Delivery Innovations | ||||
| Nano-Encapsulation | Chitosan Nanoparticles | Enhance surfactin stability in seawater by 40%; sustain release >72 hrs | Prevents rapid dilution in open-sea cages | [83] |
| Surface Functionalization | Dopamine-AMPs Coatings | Antibacterial peptides bound to 304 SS/nylon; inhibit S. aureus biofilms by 88.68% | Anti-fouling for cage nets; reduces pathogen colonization | [83] |
| V. Economic & Regulatory Landscape | ||||
| Production Costs | Surfactin Purification | Yield recovery: 3–9% after HPLC; $120–150/kg production cost | Scalability barrier for commercial use | [83] |
| EU Regulatory Status | Lipopeptide Biosurfactants | Classified as “Advanced Bioagents” under EC No 1107/2009 | Fast-track approval for aquaculture biologics | [84] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



