
Submitted:
10 September 2025
Posted:
11 September 2025
You are already at the latest version
Abstract
Alzheimer's disease(AD) is a complex pathological process that incurs significant societal costs, yet effective treatments have not yet been developed. Novel compounds targeting β-amyloid, based on the amyloid cascade hypothesis, have failed to demonstrate clinical efficacy. Among natural products with diverse mechanisms, components contained in Astragali Radix (AR) have shown anti-dementia effects in various preclinical studies, including improved cognitive function, reduced β-amyloid levels, and decreased insulin resistance. This study administered a water-extracted roasted AR to 3-month-old female 5xFAD mice for 3 months, observing changes in cognitive behavior, blood glucose, and neural signaling. Roasted AR lowered glucose levels, improved working memory, fear avoidance memory, and spatial memory, and reduced anxiety behavior in 5xFAD mice. In the hippocampus, the protein expression of BDNF and p-CREB/CREB was increased, while p-JNK/JNK was decreased. The effects of roasted AR were similar to unroasted AR in 5xFAD mice, with some components being more abundant. Therefore, roasting AR enhances its taste and aroma, making it suitable for long-term consumption in the form of tea, which could be effective in preventing neurodegenerative diseases such as dementia.
Keywords:
dementia
; Alzheimer’s disease
; Astragalus membranaceus
; roasted astragali radix
; 5xFAD
1. Introduction
Alzheimer’s disease (AD) is a prevalent neurodegenerative disorder associated with aging and other factors, imposing a significant burden on medical resources and the economy in aging societies [1]. Various hypotheses exist regarding the pathogenesis of AD, including the amyloid cascade hypothesis, tau protein hypothesis, metal ion disorder hypothesis, oxidative stress hypothesis, dysfunctional glucose metabolism, and cholinergic hypothesis [2,3,4]. Based on this theory, many new drugs have been developed to reduce β-amyloid production and aggregation, but no drug has yet been developed that is both sufficiently effective and has minimal side effects [5]. Instead, methods to increase neuronal viability and reduce cell damage have been proposed, and in particular, substances from natural products with antioxidant effects that reduce cellular damage and improve neuronal survival are attracting attention [6].
Empirically used natural products tend to have fewer side effects and are rich in diverse bioactive compounds, enabling multi-component, multi-target effects. These properties provide significant benefits for diseases with complex pathophysiology, such as AD. In traditional oriental medicine, Astragali Radix (AR), or ’Huang gi,’ is the dried root of Astragalus membranaceus [7]. The Astragalus genus is widely used in folk medicine, dietary supplements, and cosmetics [7]. Over 200 compounds have been identified in Astragalus, known for their antioxidant, anti-inflammatory, anti-aging, cytoprotective, anti-tumor, anti-diabetic, antibiotic, and immune-enhancing properties [8,9]. Astragalus contains 170 saponins, with astragaloside IV known for its antioxidative, anti-aging, anti-inflammatory, anti-asthmatic, anti-diabetic, anti-atherosclerotic activities, and neuroprotective effects [10,11,12]. About 60 flavonoids, including calycosin, calycosin-7-O-β-D-glucopyranoside (CCGR), and formononetin, are the main active isoflavonoid compounds with anti-inflammatory, antibiotic, antioxidative, anti-cancer, anti-osteoporosis, anti-diabetic, neuroprotective, and cardioprotective effects [8,13]. Approximately 30 species of Astragalus polysaccharides (APS) have been reported with anti-tumor, anti-aging, anti-diabetic activities, and cardiovascular, neuroprotective, and hepatoprotective effects [14]. Additionally, APS is associated with antimicrobial activity due to immunopotentiation and a probiotic effect on normal intestinal flora [15].
As AR contains numerous beneficial constituents, an increasing number of studies have reported its effects on improving memory and cognitive function [16]. Bioactive compounds in AR have been demonstrated to inhibit β-amyloid production and aggregation, tau hyperphosphorylation, protect neurons against oxidative stress, inhibit neuroinflammation and apoptosis, promote neural stem cell proliferation and differentiation, and reduce mitochondrial dysfunction [17]. However, natural bioactive substances are sensitive to environmental factors, can easily degrade due to light, oxygen, and temperature, or may have low absorption rates. To address these issues, heat processing can increase the bioavailability and enhance the pharmacological effects of bioactive components [18]. Roasting AR deepens its flavor and texture, improves storage properties, and prevents oxidation. Roasting also breaks down cell walls, improving the release of isoflavones and saponins and as a result, the amount of saponin and isoflavone increases, showing enhanced antioxidant and anti-inflammatory effects [19].
Based on our preliminary tests comparing the effects of AR and roasted AR (RA), we decided to test the efficacy of water-extracted RA in an AD model using mice (data not shown). We anticipated that RA would inhibit cognitive symptoms related to β-amyloid accumulation in 5xFAD mice.
2. Materials and Methods
2.1. Materials
The RA extract was provided by the Department of Herbal Crop Research, National Institute of Horticultural and Herbal Science, and its extraction procedure and composition are described by Ji et al. [19]. Briefly, dried AR root was roasted at 260°C and extracted twice with 2.4 L of water at 98 ± 2°C for 7 hours. Scopolamine hydrobromide (Cat# S0929) and D-(+)-Glucose (Cat# G7021) were purchased from Sigma-Aldrich. The primary antibodies used were: APP (amyloid precursor protein, Abcam, Cat# ab32136, 1:5000), β-amyloid (Cell Signaling, Cat# 8243S, 1:1000), tau (Cell Signaling, Cat# 4019S, 1:2500), phospho-tau (Cell Signaling, Cat# 9632S, 1:5000), BDNF (Cell Signaling, Cat# 60071S, 1:2500), phospho-CREB (Cell Signaling, Cat# 9198S, 1:2500), CREB (Cell Signaling, Cat# 4820S, 1:2000), JNK (c-Jun N-terminal kinase, Abcam, Cat# ab179461, 1:2000), phospho-JNK (Abcam, Cat# ab124956, 1:2500), NeuN (Abcam, Cat# ab104225, 1:2000), synaptophysin (Abcam, Cat# ab14692, 1:5000), GAPDH (Santa Cruz, Cat# sc-32233, 1:5000), beta-actin (Santa Cruz, Cat# sc-47778, 1:5000). The secondary antibodies used were: anti-rabbit IgG (H + L) HRP conjugate (Promega, Madison, WI, USA, Cat# W4011, 1:5000), and anti-mouse IgG (H + L) HRP conjugate (Promega, Madison, WI, USA, Cat# W4021, 1:5000).
2.2. Animals and Experimental Groups
To maintain the 5xFAD line, male hemizygous 5xFAD mice aged 8–10 weeks (B6SJL-Tg(APPSwFlLon, PSEN1*M146L*L286V)6799Vas/Mmjax, Strain #034840, Jackson Laboratory, ME, USA) were mated with 8-week-old B6SJL F1 female mice (Jackson Laboratory, Stock No. 100012). The resulting offspring were weaned at 3 weeks of age, and genotyping was performed after the weaning period by polymerase chain reaction using DNA extracted from tail biopsies. The following primers were used: common forward (5′-ACC CCC ATG TCA GAG TTC CT-3′), wild type reverse (5′-TAT ACA ACC TTG GGG GAT GG-3′), and 5xFAD reverse (5′-CGG GCC TCT TCG CTA TTA C-3′).
Female wild-type (WT) and 5xFAD littermates, confirmed by genotyping, were group-housed (n = 6 per cage) and used for experiments at 3 months of age. Experimental groups consisted of wild-type and 5xFAD mice, which were further divided according to RA treatment as follows: WT, WT + RA 500 mg/kg, 5xFAD, and 5xFAD + RA 500 mg/kg (Figure 1).
Animals were maintained under controlled environmental conditions (24 ± 1 °C, 55 ± 5% relative humidity) with a 12 h light/dark cycle (lights on at 6:00 and off at 18:00). Food and water were available ad libitum. All animal studies were conducted in accordance with the NIH Guide for the Care and Use of Laboratory Animals, and the protocols were approved by the Institutional Animal Care and Use Committee of Kyung Hee University (KHGASP-24-155).
2.3. Metabolic Changes
Body weight and Food intake :Body weight and food intake of the mice were measured weekly at the same time of day. Food intake was monitored by weighing the chow provided to each cage, and the amount consumed was calculated by subtracting the remaining food from the initial amount. Food efficiency ratio (FER) was calculated as the ratio of body weight gain to food intake over the experimental period.
Blood glucose : Non-fasting blood glucose levels were measured from tail vein blood at 10:00 AM. On the same day, mice were fasted for 6 hours, and fasting blood glucose levels were measured again from tail vein blood using the same procedure.
Oral glucose tolerance test (oGTT) : To measure glucose tolerance, mice were fasted for 6 hours and orally administered a glucose solution at a dose of 1 g/kg. Blood glucose levels were measured from tail vein blood at 15, 30, 60, 90, and 120 minutes after administration, and these values were used to calculate the area under the curve (AUC).
2.4. Behavioral Measures
Mouse behavior was evaluated using the open field test(OFT), Y-maze, elevated Plus maze (EPM), active avoidance (AA), and Morris water maze (MWM) tasks. Behavioral assessments were conducted 8 weeks after the initiation of RA administration, at which time the mice were 5 months old. RA was administered orally 30 minutes prior to each behavioral test. The tests were performed sequentially in the following order: OFT, Y-maze, AA, and MWM. All behavioral experiments were conducted under controlled illumination (20 lux) and were automatically recorded and analyzed using a video tracking system (SMART v3.0 software and SMART video tracking system, Panlab Harvard Apparatus, Barcelona, Spain).
Open Field Test (OFT) : In a white opaque Plexiglas chamber (50 × 50 × 40 cm), mice were gently placed and their movements were filmed and analyzed for an hour. Total distance moved (cm) was compared between experimental groups.
Y maze : The Y-maze test was conducted to assess working memory and cognitive flexibility. The apparatus consisted of three identical arms (35 × 13 × 7 cm) arranged at 120° angles, forming a Y-shape. Visual cues were placed at the end of each arm to facilitate spatial orientation. Each mouse was placed at the end of one arm and allowed to freely explore the maze for 10 minutes. During the test, spontaneous exploratory behavior was monitored, and the total distance traveled as well as the number of arm entries were recorded to evaluate locomotor activity. Cognitive performance was assessed by measuring the number of spontaneous alternations, defined as successive entries into all three arms without repetition. The following formula was used to calculate of the percentage of spontaneous alternation: [Number of alternation triplets / (Total arm entries – 2)] × 100
Elevated Plus Maze (EPM) : Apparatus consisted of a cross-shaped maze made with a white opaque Plexiglas arms located 40 cm above the ground. Mice were placed in the center of the maze with their heads facing the open arm, and their movement was recorded for 5 minutes. The time they spent in the open arm was measured.
Active Avoidance (AA) : Mice were placed in the active avoidance system chamber (Scitech Korea, Model No. PAAS) and left undisturbed for 2 minutes. Each session consisted of 20 trials, in which a conditioned stimulus (CS; 2000 Hz tone and light) was presented for 10 seconds, followed by an unconditioned stimulus (US; 0.3 mA foot shock) with an inter-trial interval of 2 seconds. Test was conducted over 4 consecutive days and for each trial, the latency to move to the opposite compartment was measured.
Morris Water Maze (MWM) : The test was performed in a round metal tank 180 cm in diameter and 45 cm in height filled with non-transparent water (22 degree Celsius, 30 cm in depth). A platform was submerged 0.5 cm under the water and was hidden to the animals sight by adding white-colored paint into the water. Four symbolic figures were attached to the wall in well-sighted places from the maze. The training lasted 5 days and was performed four times per day. During the 4 daily trials, the starting point differed on each trial. Training sessions continued until the mouse climbed up to find the platform. The time to reach the platform was measured. On the 6th day, the probe test was performed by removing the platform. The swimming trajectories were tracked for a total of 90 s and the time spent in the quadrant where the platform was previously placed was measured.
2.5. Western Blotting
Under deep anesthetization by isoflurane, mice were sacrificed and hippocampus tissues were dissected and stored at −80°C until use. For the analysis of protein expression, tissues were homogenized with lysis buffer containing 1% each of phosphatase inhibitor cocktails 2 and 3 (P5726, P044, Sigma- Aldrich) in protein extraction solution (PRO-PREP, iNtRON Biotechnology, Seongnam, South Korea) and centrifuged at 13,000 rpm for 15 minutes at 4°C. Protein concentrations were measured using DC™ Protein Assay Reagents A, B, and S (5000113, 5000114, and 5000115, BIO-RAD, Hercules, CA, USA). After electrophoresis using 10% SDS-polyacrylamide gel, the gel phase proteins were transferred to a PVDF membrane (BIO-RAD, Cat# 1620177). Membranes were incubated for 1 hour at room temperature in 5% skim milk (BD Difco™, Cat# 212100, USA) dissolved in TBS-T (0.1% tween 20 in TBS) for blocking. Primary antibodies were diluted in 5% skim milk in TBS-T and incubated overnight at 4°C. Next day, the membranes were washed thrice for 10 minutes with TBS-T. Secondary antibodies were diluted with 5% skim milk in TBS-T and incubated for 1 hour at room temperature. After washing twice for 10 minutes with TBS-T, luminescence was induced using a ECL solution (WSE-7120S, ATTO Co., Tokyo, Japan) and the samples were photographed with an EZ-Capture MG (ATTO, Tokyo, Japan). Band density was analyzed using ImageJ software (NIH, Bethesda, MD, USA).
2.6. Statistical Analysis
All results are expressed as the mean ± S.E.M. Statistical comparisons were performed using ANOVA (blood glucose, OGTT-AUC, OFT, EPM, Y-maze, MWM-probe, density analysis of western blot images), repeated measure ANOVA (body weight, OGTT, MWM-latency) followed by LSD or Tukey’s HSD post hoc multiple comparisons testing using SPSS 22 (SPSS Inc., Chicago, IL, USA).
3. Results
3.1. RA Increased Body Weight and Improved Food Efficiency Ratio of 5xFAD Mice
Three-month-old WT and WT+RA mice gained body weight, but not the 5xFAD mice during the 12-week experimental period. RA increased the weight gain in 5xFAD mice (Figure 2 A&B). In 5xFAD mice, the average weekly food intake (Figure 2C) and FER was less than WT mice, but RA improved the FER in 5xFAD mice (Figure 2D).
3.2. RA Reduced Fasting Blood Glucose and Improved Glucose Tolerance of 5xFAD Mice
RA decreased the non-fasting and fasting blood glucose concentration in wild type mice and decreased the fasting blood glucose concentration of 5xFAD mice (Figure 3. A&B). RA also reduced the peak blood glucose concentration in both wild type and 5xFAD mice, but the AUC was reduced only in 5xFAD mice by RA (Figure 3. C&D). There was no significant change in the insulin levels and HOMA-IR among experimental groups (Figure 3. E&F).
3.3. RA Improved Cognitive Deficits in 5xFAD Mice
Percent alternation in the Y-maze as a measure of working memory was improved by RA in 5xFAD mice (Figure 4A). The latency to escape in the active avoidance paradigm was prolonged in 5xFAD mice and RA treatment normalized the lag of latency in 5xFAD mice (Figure 4B). As a measure of spatial learning and memory, latency to find the platform and the percentage spent in the platform area in the MWM test was also improved by RA in 5xFAD mice (Figure 4. C&D). There was no significant difference of locomotor activity in the OFT (Figure 4E). 5xFAD mice showed anxiety-like response in the EPM and RA reduced anxiety-like behavior in both wild type and 5xFAD mice (Figure 4F).
3.4. RA Didn’t Affect the Expression of Hippocampal β-Amyloid and p-Tau in 5xFAD Mice
The expression of APP and β-amyloid increased in 5xFAD mice, but the expression of Tau and p-Tau remained unchanged in the hippocampus (Figure 5). RA treatment did not affect the amount of APP and β-amyloid accumulated in 5xFAD mice (Figure 5. A&B). Also, RA did not affect the amount of Tau or p-Tau (Figure 5. C&D).
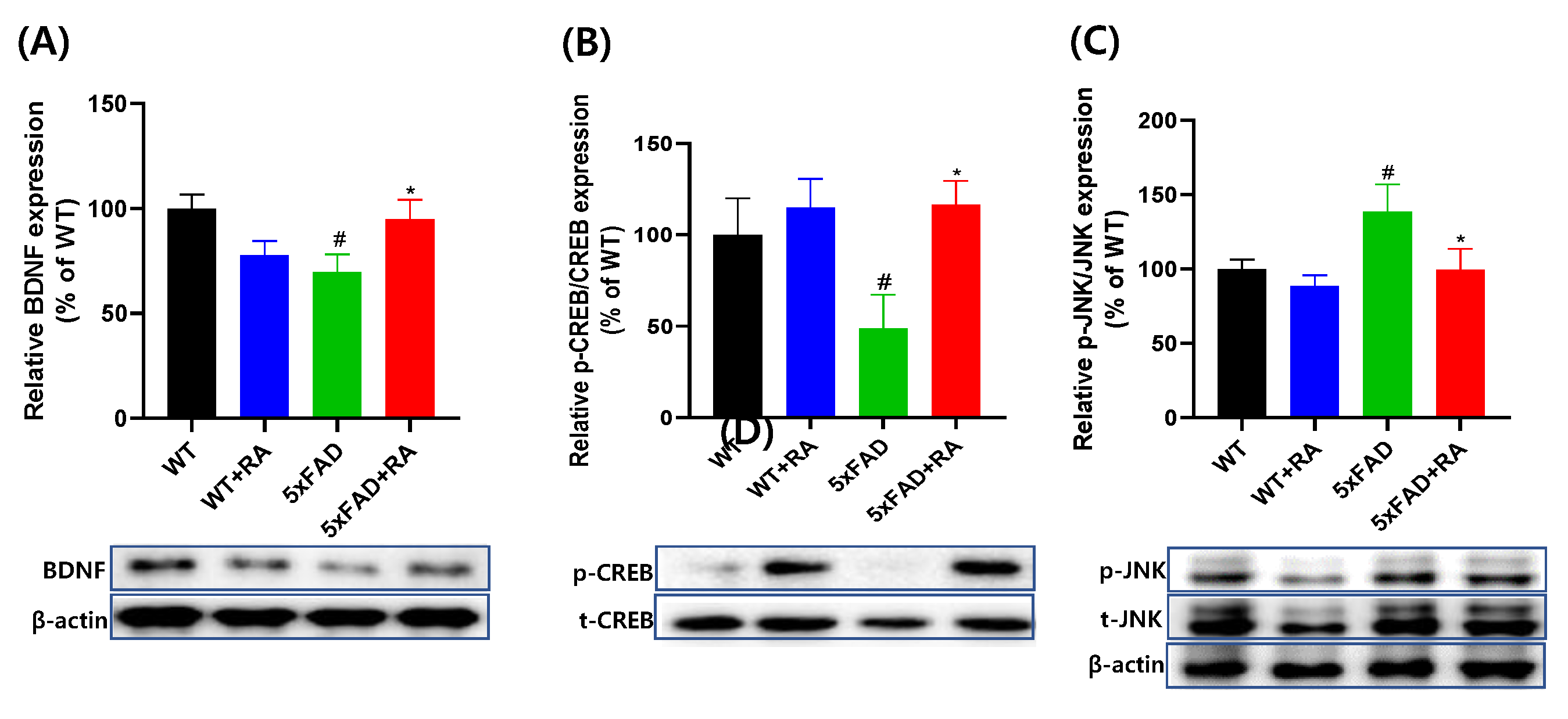
3.5. RA Normalized Altered Neurotrophic Signaling Pathways in 5xFAD Mice
In 5xFAD mice, the hippocampal expression of BDNF and p-CREB/CREB was decreased and p-JNK/JNK was increased (Figure 6). RA increased the expression of BDNF and the p-CREB/CREB and decreased the p-JNK/JNK.
4. Discussion
This study shows that RA can improve cognitive symptoms in an AD model mouse by normalizing the expression of BDNF, p-CREB, and p-JNK, and by lowering blood sugar, although it does not reduce the accumulation of β-amyloid.
Recently, many new substances aimed at reducing β-amyloid accumulation in brain tissue to treat AD have been tested in clinical trials, but most have not shown satisfactory effects. For example, Bapineuzumab, an anti-Aβ monoclonal antibody, failed to improve daily life and cognitive functions in patients with mild to moderate AD [20]. Semagacestat, a γ-secretase inhibitor, failed to improve cognitive function and had serious side effects [21]. Vrubecestat, a BACE1 inhibitor, was discontinued in Phase 2/3 clinical trials because it did not sufficiently improve cognitive function [22]. While the amyloid cascade hypothesis is recognized as the cause of AD, these cases suggest that treatments targeting only β-amyloid are insufficiently effective. It is becoming increasingly clear that AD is caused by complex etiologies, and treatments should be developed based on diverse mechanisms [23]. Therefore, natural products with multiple effects represent a useful approach for developing dementia treatments, particularly those targeting neurotrophic factors such as BDNF, which have proven effective in preclinical studies [24].
5xFAD transgenic mice are widely used as a preclinical model of AD, developing neuroinflammation from 2 months of age, with a decrease in neuron numbers from 4 months, and significant reduction in cognitive function by 9 months [25]. In this study, the effects of feeding RA for 3 months to 3-month-old female WT and 5xFAD mice were analyzed both behaviorally and biochemically. 5xFAD mice did not gain weight during this period and showed less average food intake than WT mice. Avoidance memory and spatial memory-related behavior decreased, and anxiety-like behavior increased. The amount of APP, Aβ, and p-JNK expression increased in the hippocampus, while the expression of BDNF and p-CREB decreased. However, Tau and p-Tau expression did not change in the hippocampus.
Previous studies have shown that AR has anti-oxidative, anti-inflammatory, anti-dementia and anti-diabetic effects in various preclinical models [17]. In a study using roasted AR, referred to as RA, it activated the AKT/CREB/BDNF pathway in vitro and activated antioxidant enzymes via the Nrf2 pathway, exhibiting neuroprotective effects. In this study, in vivo RA increased body weight, decreased fasting blood glucose, and improved glucose tolerance in 5xFAD mice. Behaviorally, RA improved working memory (Y-maze), avoidance memory (active avoidance), and spatial memory (Morris water maze), and reduced anxiety-like behavior (elevated plus maze). In the hippocampus, RA increased the expression of BDNF and p-CREB and suppressed the expression of p-JNK in 5xFAD mice. However, it had no effect on the expression of APP, β-amyloid, Tau, and p-Tau. Therefore, RA may be related to the effect of protecting cell damage caused by β-amyloid accumulation, such as neurotrophic effect and protection from oxidative stress [26].
AR contains many ingredients with antioxidant effect, such as flavonoids, saponins, and APS components [10]. Among these components, isoflavonoids such as formononetin, ononin, calycosin-7-β-glucoside, and calycosin were detected in plasma after oral administration in rodents [27,28]. After roasting the AR, the amounts of calycosin, formononetin, and ononin increased, while calycosin 7-glucoside decreased [19]. Calycosin and calycosin-7-glucoside, which have similar known effects, are the components with the highest content in AR and are well absorbed even after oral administration. They reduced β-amyloid, tau, IL-1β, TNF-α, and AChE activities in the hippocampus and increased acetylcholine and glutathione activities in APP/PS1 transgenic mice [29]. Formononetin is involved in APP protein processing and inhibits the production of amyloid proteins, increases amyloid protein clearance through LRP1, and reduces the influx of amyloid proteins into neurons through RAGE inhibition [30]. Ononin is known to have free radical scavenging and HIF-1 regulation effects [17]. AR also contains quercetin, which is known to inhibit β-amyloid production, aggregation, and tau phosphorylation [31]; inhibit AChE activity [32]; and attenuate oxidative stress and neuroinflammation [33]. Despite these previous findings, we did not observe a reduction in β-amyloid after treating with RA 500 mg/kg for 3 months.
Astragaloside IV, a representative saponin component of AR, is a PPARγ agonist that inhibits BACE1, thereby suppressing the production and aggregation of β-amyloid [34]. It is also known to inhibit tau protein hyperphosphorylation [35], have anti-apoptotic [36] and anti-inflammatory effects [37], promote the proliferation and differentiation of neural stem cells [38], and induce BDNF [39]. BDNF was increased by RA in 5xFAD mice, but since there was no reduction in β-amyloid, our results do not align with the known effects of astragaloside IV.
APS is a component that dissolves in water but not well in alcohol, and is found in large quantities in water-extracted AR [40]. APS is known for various bioactivities, including immunomodulatory, antioxidant, antitumor, antidiabetic, antiviral, hepatoprotective, anti-inflammatory, anti-atherosclerotic, hematopoietic, and neuroprotective effects [26]. In APP/PS1 transgenic mice, a dementia model, APS improved cognitive behavior and reduced the effects of metabolic stress, but did not reduce β-amyloid accumulation [41]. Instead, it lowered blood sugar and improved insulin resistance [26]. In streptozotocin-induced diabetic rats, APS improved spatial memory, increased hippocampal neurons in the CA1 region, and increased p-CREB, p-NMDA, and p-CaMKII. APS and AR can also inhibit neuroinflammation. Our study using water-extracted RA lowered blood glucose and improved cognitive behavior but did not reduce β-amyloid accumulation, reflecting the known effects of APS.
5xFAD mice exhibit various phenotypes in addition to cognitive dysfunction. According to Forner et al. (2021), 5xFAD male and female mice at 12 and 18 months had lower body weight than wild-type mice, showed less activity in the open field at 18 months, spent more time in the open arm in the EPM at 4, 8, and 12 months, and lasted longer on the rotarod at 4 months [42]. According to Oblak et al. (2021), cholesterol, LDL, and HDL lipoproteins were reduced in 12-month-old male 5xFAD mice compared to wild-type mice, and blood glucose was reduced in 12-month-old female 5xFAD mice [43]. According to our findings, 5xFAD females gained less weight after 3 months of age. At this time, 5xFAD had lower food intake and a lower food efficiency ratio than other groups, indicating low body weight gain compared to food consumption. In response, RA increased body weight, lowered fasting blood glucose, and improved glucose tolerance in 5xFAD during this period. However, there was no change in insulin resistance as indicated by the HOMA-IR index. In a previous study, AR also improved hyperlipidemia by promoting lipolysis [44]. APS can reduce hyperglycemia, insulin resistance, and inhibit myostatin secretion from skeletal muscle, thereby inhibiting bone muscle loss [45].
RA showed an anti-anxiety effect in the EPM. Regarding the anxiolytic effect, AR reduced anxiety-like behavior and stress hormones in immobilized mice [46,47]. In particular, astragaloside IV improved anxiety and depressive symptoms induced by bisphenol A [48], and showed an anti-depressant effect on chronic unpredictable mild stress [49]. In the OFT, the average locomotor activity of 5xFAD mice was similar to other groups, but variation increased. This may relate to variable activity levels in 5xFAD mice depending on age [25,42]. However, the average activity level did not differ significantly from other groups, indicating that anxiety-like behavior in the elevated plus maze and behavioral changes in latency and active avoidance in the MWM were not confounded by activity level differences, but rather resulted from anxiety and cognitive behavior.
BDNF plays an important role in neuroprotection by reducing β-amyloid toxicity in AD [50]. BDNF expression in the hippocampus was reduced in female 5xFAD mice [51], and increasing BDNF secretion improved cognitive function [52]. Increasing BDNF secretion through astrocytes in 5xFAD mice improved cognitive function not by reducing β-amyloid accumulation or stimulating neuronal regeneration, but by restoring cognitive function through improving the structure and function of the hippocampal neural circuit [53]. Astragaloside IV, formononetin, formononetin-7-O-glucoside, and calycosin interact with TrkB [54]. In a previous study inducing neuroinflammation with lipopolysaccharide, AR reduced microglial activation, BBB dysfunction, and cognitive dysfunction associated with BDNF/TrkB/CREB signaling pathway activation [55]. In our study, BDNF and the downstream effector of BDNF/TrK signaling, p-CREB/CREB, increased with RA in 5xFAD mice but not in WT mice.
JNK plays an important regulatory role in oxidative damage and programmed cell death [56]. The degree of JNK activity correlated with cognitive decline [57], and JNK inhibitors suppressed neuronal cell death, improved cognitive function, and at high doses, reduced β-amyloid in 5xFAD [58]. Flavonoids from AR inhibited AKT and JNK signaling in LPS-activated BV-2 microglia cells [59]. APS inhibited LPS-induced JNK activation in cardiomyocytes [60]. Astragaloside IV reduced apoptosis by inhibiting the p-JNK/JNK pathway in oxygen glucose deprivation/reoxygenation-induced damage [61]. In vivo, AR inhibited JNK and p-JNK expression in the hippocampus of an LPS-induced depression model [62] and showed neuroprotective effects by inhibiting JNK3 expression in an MCAO model [63]. As shown by Ji et al. (2024), RA was enriched with calycosin, formononetin, and ononin, which may have inhibited p-JNK expression, but other ingredients may also have this effect [19].
Although some studies have shown that AR directly inhibits β-amyloid and p-tau accumulation [17,64], we could not reproduce this effect in our experimental setup. This may be due to insufficient dose or administration period of RA used. Despite failing to reduce β-amyloid levels, RA may contribute to improving dementia symptoms through reducing hyperglycemia, inducing BDNF, and suppressing the JNK pathway. As roasted AR increased the concentration of isoflavone components related to antioxidant protein induction and decrease of ROS production [19]. Especially, caspase-9, caspase-3 and cytochrome C release was inhibited and increased Bcl-2 and decreased Bax expression effectively suppressed oxidative stress induced by Aβ accumulation [19]. These findings suggest that anti-apoptotic and antioxidant effects play a vital role in cognitive improvement of RA. Furthermore, AR appears non-toxic with long-term administration. Studies in SD rats and Beagle dogs have shown that AR is low in toxicity and does not produce adverse effects even with long-term use [65]. APS was also evaluated as safe for chronic neuropathy due to low toxicity at conventional doses, with no side effects observed even with long-term use [66].
5. Conclusions
This study tested the efficacy of roasted AR in 5xFAD transgenic mice. It improved cognitive function despite not preventing pathological β-amyloid accumulation in these mice. This may be related to the neurotrophic and antioxidant effects protecting cell damage caused by β-amyloid accumulation. Furthermore, RA improved glucose tolerance and BDNF expression, and suppressed the JNK pathway, suggesting its potential use in sporadic AD. RA is a roasted version of AR, enhancing its flavor and aroma. While there were some compositional changes in RA, many effects were similar to known effects of AR. Furthermore, AR has minimal side effects, suggesting that long-term consumption of RA may help prevent neurodegenerative diseases such as dementia.
Author Contributions
Conceptualization, S.M. and J-H.Y.; methodology, J.S. G.K. and J-S.S.; validation, S.J. and S.I.; formal analysis, S.I. and S.J,; investigation, J-H.Y., J.M. and Y.K.; resources, G-Y.J.; data curation, J.S.; writing—original draft preparation, S.M.; writing—review and editing, G.K. and J-S.S.; visualization, J-H.Y., J.M. and Y.K.; supervision, S-S.K.; project administration, S-S.K .; funding acquisition, S-S.K . All authors have read and agreed to the published version of the manuscript.
Funding
This work was carried out with the support of "Research Program for Agriculture Science and Technology Development (Project No. RS-2022-RD010294)" Rural Development Administration, Republic of Korea and BK21 plus program "AgeTech-Service Convergence Major" through the National Research Foundation (NRF) funded by the Ministry of Education of Korea [5120200313836].
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Review Board of Kyung Hee Univeristy (KHGASP-24-155, April-25, 2024) for studies involving animals in accordance with the NIH Guide for the Care and Use of Laboratory Animals.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Conflicts of Interest
“The authors declare no conflicts of interest.”
Abbreviations
The following abbreviations are used in this manuscript:
| AChE | Acetylcholinesterase |
| AD | Alzheimer’s disease |
| APP | Amyloid precursor protein |
| APS | Astragalus polysaccharides |
| AR | Astragali radix |
| BDNF | Brain-derived neurotrophic factor |
| CREB | cAMP-responsive element binding protein |
| EPM | Elevated plus maze |
| JNK | c-Jun NH2-terminal kinase |
| MWM | Morris water maze task |
| OFT | Open field task |
| RA | Roasted astragali radix |
| WT | Wild type |
| 5xFAD | Five familial Alzheimer’s disease mutations |
References
- Scheltens, P.; Blennow, K.; Breteler, M.M.; de Strooper, B.; Frisoni, G.B.; Salloway, S.; Van der Flier, W.M. Alzheimer’s disease. Lancet 2016, 388, 505–517. [Google Scholar] [CrossRef] [PubMed]
- Savelieff, M.G.; Lee, S.; Liu, Y.; Lim, M.H. Untangling amyloid-beta, tau, and metals in Alzheimer’s disease. ACS Chem Biol 2013, 8, 856–865. [Google Scholar] [CrossRef]
- Butterfield, D.A.; Halliwell, B. Oxidative stress, dysfunctional glucose metabolism and Alzheimer disease. Nat Rev Neurosci 2019, 20, 148–160. [Google Scholar] [CrossRef] [PubMed]
- Hampel, H.; Mesulam, M.M.; Cuello, A.C.; Farlow, M.R.; Giacobini, E.; Grossberg, G.T.; Khachaturian, A.S.; Vergallo, A.; Cavedo, E.; Snyder, P.J.; et al. The cholinergic system in the pathophysiology and treatment of Alzheimer’s disease. Brain 2018, 141, 1917–1933. [Google Scholar] [CrossRef] [PubMed]
- Breijyeh, Z.; Karaman, R. Comprehensive Review on Alzheimer’s Disease: Causes and Treatment. Molecules 2020, 25. [Google Scholar] [CrossRef]
- Chu, C.T.; Levinthal, D.J.; Kulich, S.M.; Chalovich, E.M.; DeFranco, D.B. Oxidative neuronal injury. The dark side of ERK1/2. Eur J Biochem 2004, 271, 2060–2066. [Google Scholar] [CrossRef]
- Li, X.; Qu, L.; Dong, Y.; Han, L.; Liu, E.; Fang, S.; Zhang, Y.; Wang, T. A review of recent research progress on the astragalus genus. Molecules 2014, 19, 18850–18880. [Google Scholar] [CrossRef]
- Bratkov, V.M.; Shkondrov, A.M.; Zdraveva, P.K.; Krasteva, I.N. Flavonoids from the Genus Astragalus: Phytochemistry and Biological Activity. Pharmacogn Rev 2016, 10, 11–32. [Google Scholar] [CrossRef]
- Yang, L.P.; Shen, J.G.; Xu, W.C.; Li, J.; Jiang, J.Q. Secondary metabolites of the genus Astragalus: structure and biological-activity update. Chem Biodivers 2013, 10, 1004–1054. [Google Scholar] [CrossRef]
- Costa, I.M.; Lima, F.O.V.; Fernandes, L.C.B.; Norrara, B.; Neta, F.I.; Alves, R.D.; Cavalcanti, J.; Lucena, E.E.S.; Cavalcante, J.S.; Rego, A.C.M.; et al. Astragaloside IV Supplementation Promotes A Neuroprotective Effect in Experimental Models of Neurological Disorders: A Systematic Review. Curr Neuropharmacol 2019, 17, 648–665. [Google Scholar] [CrossRef]
- Gong, L.; Chang, H.; Zhang, J.; Guo, G.; Shi, J.; Xu, H. Astragaloside IV Protects Rat Cardiomyocytes from Hypoxia-Induced Injury by Down-Regulation of miR-23a and miR-92a. Cell Physiol Biochem 2018, 49, 2240–2253. [Google Scholar] [CrossRef]
- Li, L.; Hou, X.; Xu, R.; Liu, C.; Tu, M. Research review on the pharmacological effects of astragaloside IV. Fundam Clin Pharmacol 2017, 31, 17–36. [Google Scholar] [CrossRef] [PubMed]
- Ong, S.K.L.; Shanmugam, M.K.; Fan, L.; Fraser, S.E.; Arfuso, F.; Ahn, K.S.; Sethi, G.; Bishayee, A. Focus on Formononetin: Anticancer Potential and Molecular Targets. Cancers (Basel) 2019, 11. [Google Scholar] [CrossRef]
- Jin, M.; Zhao, K.; Huang, Q.; Shang, P. Structural features and biological activities of the polysaccharides from Astragalus membranaceus. Int J Biol Macromol 2014, 64, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Huang, G. The antiviral activity of polysaccharides and their derivatives. Int J Biol Macromol 2018, 115, 77–82. [Google Scholar] [CrossRef]
- Sun, L.; Zhang, H.; Wang, W.; Chen, Z.; Wang, S.; Li, J.; Li, G.; Gao, C.; Sun, X. Astragaloside IV Exerts Cognitive Benefits and Promotes Hippocampal Neurogenesis in Stroke Mice by Downregulating Interleukin-17 Expression via Wnt Pathway. Front Pharmacol 2020, 11, 421. [Google Scholar] [CrossRef] [PubMed]
- Dong, Q.; Li, Z.; Zhang, Q.; Hu, Y.; Liang, H.; Xiong, L. Astragalus mongholicus Bunge (Fabaceae): Bioactive Compounds and Potential Therapeutic Mechanisms Against Alzheimer’s Disease. Front Pharmacol 2022, 13, 924429. [Google Scholar] [CrossRef]
- Sruthi, N.U.; Premjit, Y.; Pandiselvam, R.; Kothakota, A.; Ramesh, S.V. An overview of conventional and emerging techniques of roasting: Effect on food bioactive signatures. Food Chem 2021, 348, 129088. [Google Scholar] [CrossRef]
- Ji, Y.J.; Kang, M.H.; Han, S.H.; Kim, G.S.; Kim, H.D.; Jang, G.Y. Roasted Astragalus membranaceus Inhibits Abeta25-35-Induced Oxidative Stress in Neuronal Cells by Activating the Nrf2/HO-1 and AKT/CREB/BDNF Pathways. Antioxidants (Basel) 2024, 13. [Google Scholar] [CrossRef]
- Ivanoiu, A.; Pariente, J.; Booth, K.; Lobello, K.; Luscan, G.; Hua, L.; Lucas, P.; Styren, S.; Yang, L.; Li, D.; et al. Long-term safety and tolerability of bapineuzumab in patients with Alzheimer’s disease in two phase 3 extension studies. Alzheimers Res Ther 2016, 8, 24. [Google Scholar] [CrossRef]
- Henley, D.B.; Sundell, K.L.; Sethuraman, G.; Dowsett, S.A.; May, P.C. Safety profile of semagacestat, a gamma-secretase inhibitor: IDENTITY trial findings. Curr Med Res Opin 2014, 30, 2021–2032. [Google Scholar] [CrossRef] [PubMed]
- Egan, M.F.; Mukai, Y.; Voss, T.; Kost, J.; Stone, J.; Furtek, C.; Mahoney, E.; Cummings, J.L.; Tariot, P.N.; Aisen, P.S.; et al. Further analyses of the safety of verubecestat in the phase 3 EPOCH trial of mild-to-moderate Alzheimer’s disease. Alzheimers Res Ther 2019, 11, 68. [Google Scholar] [CrossRef]
- Decourt, B.; D’Souza, G.X.; Shi, J.; Ritter, A.; Suazo, J.; Sabbagh, M.N. The Cause of Alzheimer’s Disease: The Theory of Multipathology Convergence to Chronic Neuronal Stress. Aging Dis 2022, 13, 37–60. [Google Scholar] [CrossRef]
- Habtemariam, S. Natural Products in Alzheimer’s Disease Therapy: Would Old Therapeutic Approaches Fix the Broken Promise of Modern Medicines? Molecules 2019, 24. [Google Scholar] [CrossRef]
- Padua, M.S.; Guil-Guerrero, J.L.; Lopes, P.A. Behaviour Hallmarks in Alzheimer’s Disease 5xFAD Mouse Model. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef]
- Shi, Y.; Ma, P. Pharmacological effects of Astragalus polysaccharides in treating neurodegenerative diseases. Front Pharmacol 2024, 15, 1449101. [Google Scholar] [CrossRef]
- Shi, J.; Zheng, L.; Lin, Z.; Hou, C.; Liu, W.; Yan, T.; Zhu, L.; Wang, Y.; Lu, L.; Liu, Z. Study of pharmacokinetic profiles and characteristics of active components and their metabolites in rat plasma following oral administration of the water extract of Astragali radix using UPLC-MS/MS. J Ethnopharmacol 2015, 169, 183–194. [Google Scholar] [CrossRef]
- Liu, X.H.; Zhao, J.B.; Guo, L.; Yang, Y.L.; Hu, F.; Zhu, R.J.; Feng, S.L. Simultaneous determination of calycosin-7-O-beta-D-glucoside, ononin, calycosin, formononetin, astragaloside IV, and astragaloside II in rat plasma after oral administration of Radix Astragali extraction for their pharmacokinetic studies by ultra-pressure liquid chromatography with tandem mass spectrometry. Cell Biochem Biophys 2014, 70, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Li, X.; Bai, X.X.; Gao, J.; Wang, C.Y. Calycosin improves cognitive function in a transgenic mouse model of Alzheimer’s disease by activating the protein kinase C pathway. Neural Regen Res 2017, 12, 1870–1876. [Google Scholar] [CrossRef] [PubMed]
- Fei, H.X.; Zhang, Y.B.; Liu, T.; Zhang, X.J.; Wu, S.L. Neuroprotective effect of formononetin in ameliorating learning and memory impairment in mouse model of Alzheimer’s disease. Biosci Biotechnol Biochem 2018, 82, 57–64. [Google Scholar] [CrossRef]
- Paris, D.; Mathura, V.; Ait-Ghezala, G.; Beaulieu-Abdelahad, D.; Patel, N.; Bachmeier, C.; Mullan, M. Flavonoids lower Alzheimer’s Abeta production via an NFkappaB dependent mechanism. Bioinformation 2011, 6, 229–236. [Google Scholar] [CrossRef]
- Costa, L.G.; Garrick, J.M.; Roque, P.J.; Pellacani, C. Mechanisms of Neuroprotection by Quercetin: Counteracting Oxidative Stress and More. Oxid Med Cell Longev 2016, 2016, 2986796. [Google Scholar] [CrossRef]
- Qureshi, A.A.; Tan, X.; Reis, J.C.; Badr, M.Z.; Papasian, C.J.; Morrison, D.C.; Qureshi, N. Inhibition of nitric oxide in LPS-stimulated macrophages of young and senescent mice by delta-tocotrienol and quercetin. Lipids Health Dis 2011, 10, 239. [Google Scholar] [CrossRef]
- Wang, X.; Wang, Y.; Hu, J.P.; Yu, S.; Li, B.K.; Cui, Y.; Ren, L.; Zhang, L.D. Astragaloside IV, a Natural PPARgamma Agonist, Reduces Abeta Production in Alzheimer’s Disease Through Inhibition of BACE1. Mol Neurobiol 2017, 54, 2939–2949. [Google Scholar] [CrossRef]
- Chang, C.P.; Liu, Y.F.; Lin, H.J.; Hsu, C.C.; Cheng, B.C.; Liu, W.P.; Lin, M.T.; Hsu, S.F.; Chang, L.S.; Lin, K.C. Beneficial Effect of Astragaloside on Alzheimer’s Disease Condition Using Cultured Primary Cortical Cells Under beta-amyloid Exposure. Mol Neurobiol 2016, 53, 7329–7340. [Google Scholar] [CrossRef]
- Sun, Q.; Jia, N.; Wang, W.; Jin, H.; Xu, J.; Hu, H. Protective effects of astragaloside IV against amyloid beta1-42 neurotoxicity by inhibiting the mitochondrial permeability transition pore opening. PLoS One 2014, 9, e98866. [Google Scholar] [CrossRef]
- Chen, F.; Yang, D.; Cheng, X.Y.; Yang, H.; Yang, X.H.; Liu, H.T.; Wang, R.; Zheng, P.; Yao, Y.; Li, J. Astragaloside IV Ameliorates Cognitive Impairment and Neuroinflammation in an Oligomeric Abeta Induced Alzheimer’s Disease Mouse Model via Inhibition of Microglial Activation and NADPH Oxidase Expression. Biol Pharm Bull 2021, 44, 1688–1696. [Google Scholar] [CrossRef] [PubMed]
- Haiyan, H.; Rensong, Y.; Guoqin, J.; Xueli, Z.; Huaying, X.; Yanwu, X. Effect of Astragaloside IV on Neural Stem Cell Transplantation in Alzheimer’s Disease Rat Models. Evid Based Complement Alternat Med 2016, 2016, 3106980. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xu, W.; Chen, H.; Li, W.; Li, W.; Zhu, G. Astragaloside IV prevents Abeta(1-42) oligomers-induced memory impairment and hippocampal cell apoptosis by promoting PPARgamma/BDNF signaling pathway. Brain Res 2020, 1747, 147041. [Google Scholar] [CrossRef]
- Li, R.; Chen, W.-c.; Wang, W.-p.; Tian, W.-y.; Zhang, X.-g. Optimization of extraction technology of Astragalus polysaccharides by response surface methodology and its effect on CD40. Carbohydrate Polymers 2009, 78, 784–788. [Google Scholar] [CrossRef]
- Huang, Y.C.; Tsay, H.J.; Lu, M.K.; Lin, C.H.; Yeh, C.W.; Liu, H.K.; Shiao, Y.J. Astragalus membranaceus-Polysaccharides Ameliorates Obesity, Hepatic Steatosis, Neuroinflammation and Cognition Impairment without Affecting Amyloid Deposition in Metabolically Stressed APPswe/PS1dE9 Mice. Int J Mol Sci 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Forner, S.; Kawauchi, S.; Balderrama-Gutierrez, G.; Kramar, E.A.; Matheos, D.P.; Phan, J.; Javonillo, D.I.; Tran, K.M.; Hingco, E.; da Cunha, C.; et al. Systematic phenotyping and characterization of the 5xFAD mouse model of Alzheimer’s disease. Sci Data 2021, 8, 270. [Google Scholar] [CrossRef]
- Oblak, A.L.; Lin, P.B.; Kotredes, K.P.; Pandey, R.S.; Garceau, D.; Williams, H.M.; Uyar, A.; O’Rourke, R.; O’Rourke, S.; Ingraham, C.; et al. Comprehensive Evaluation of the 5XFAD Mouse Model for Preclinical Testing Applications: A MODEL-AD Study. Front Aging Neurosci 2021, 13, 713726. [Google Scholar] [CrossRef]
- Wang, L.; Zheng, W.; Yang, J.; Ali, A.; Qin, H. Mechanism of Astragalus membranaceus Alleviating Acquired Hyperlipidemia Induced by High-Fat Diet through Regulating Lipid Metabolism. Nutrients 2022, 14. [Google Scholar] [CrossRef]
- Liu, M.; Qin, J.; Hao, Y.; Liu, M.; Luo, J.; Luo, T.; Wei, L. Astragalus polysaccharide suppresses skeletal muscle myostatin expression in diabetes: involvement of ROS-ERK and NF-kappaB pathways. Oxid Med Cell Longev 2013, 2013, 782497. [Google Scholar] [CrossRef]
- Oh, H.A.; Choi, H.J.; Kim, N.J.; Kim, D.H. Anti-stress effect of astragaloside IV in immobilized mice. J Ethnopharmacol 2014, 153, 928–932. [Google Scholar] [CrossRef]
- Park, H.J.; Kim, H.Y.; Yoon, K.H.; Kim, K.S.; Shim, I. The Effects of Astragalus Membranaceus on Repeated Restraint Stress-induced Biochemical and Behavioral Responses. Korean J Physiol Pharmacol 2009, 13, 315–319. [Google Scholar] [CrossRef]
- Abd Elkader, H.A.E.; Abdou, H.M.; Khamiss, O.A.; Essawy, A.E. Anti-anxiety and antidepressant-like effects of astragaloside IV and saponins extracted from Astragalus spinosus against the bisphenol A-induced motor and cognitive impairments in a postnatal rat model of schizophrenia. Environ Sci Pollut Res Int 2021, 28, 35171–35187. [Google Scholar] [CrossRef] [PubMed]
- Liu. X; Xu. W; Zhao. Z; Wu. X; Wang. S; Jian. C; Li. M; Li. K; Wang. Y; X., Q. Prophylactic Effects of Astragalus Polysaccharides on Depression-Like Behaviors in Rats With Chronic Unpredictable Mild Stress: A Role of Gut-Microbiota–Brain Axis. Food Frontiers 2024, 6, 1343–1361. [Google Scholar] [CrossRef]
- Zhao, H.; Alam, A.; San, C.Y.; Eguchi, S.; Chen, Q.; Lian, Q.; Ma, D. Molecular mechanisms of brain-derived neurotrophic factor in neuro-protection: Recent developments. Brain Res 2017, 1665, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, Y.; Nakasone, H.; Kasahara, R.; Fukuchi, M. Expression Profiles of Brain-Derived Neurotrophic Factor Splice Variants in the Hippocampus of Alzheimer’s Disease Model Mouse. Biol Pharm Bull 2024, 47, 1858–1867. [Google Scholar] [CrossRef]
- Patel, D.; Roy, A.; Raha, S.; Kundu, M.; Gonzalez, F.J.; Pahan, K. Upregulation of BDNF and hippocampal functions by a hippocampal ligand of PPARalpha. JCI Insight 2020, 5. [Google Scholar] [CrossRef]
- de Pins, B.; Cifuentes-Diaz, C.; Farah, A.T.; Lopez-Molina, L.; Montalban, E.; Sancho-Balsells, A.; Lopez, A.; Gines, S.; Delgado-Garcia, J.M.; Alberch, J.; et al. Conditional BDNF Delivery from Astrocytes Rescues Memory Deficits, Spine Density, and Synaptic Properties in the 5xFAD Mouse Model of Alzheimer Disease. J Neurosci 2019, 39, 2441–2458. [Google Scholar] [CrossRef]
- Yao, H.; Gu, L.J.; Guo, J.Y. [Study on effect of astragali radix polysaccharides in improving learning and memory functions in aged rats and its mechanism]. Zhongguo Zhong Yao Za Zhi 2014, 39, 2071–2075. [Google Scholar] [PubMed]
- Liu, K.; Wan, G.; Jiang, R.; Zou, L.; Wan, D.; Zhu, H.; Feng, S. Astragalus injection ameliorates lipopolysaccharide-induced cognitive decline via relieving acute neuroinflammation and BBB damage and upregulating the BDNF-CREB pathway in mice. Pharm Biol 2022, 60, 825–839. [Google Scholar] [CrossRef] [PubMed]
- Ginet, V.; Puyal, J.; Magnin, G.; Clarke, P.G.; Truttmann, A.C. Limited role of the c-Jun N-terminal kinase pathway in a neonatal rat model of cerebral hypoxia-ischemia. J Neurochem 2009, 108, 552–562. [Google Scholar] [CrossRef]
- Priori, E.C.; Musi, C.A.; Giani, A.; Colnaghi, L.; Milic, I.; Devitt, A.; Borsello, T.; Repici, M. JNK Activation Correlates with Cognitive Impairment and Alteration of the Post-Synaptic Element in the 5xFAD AD Mouse Model. Cells-Basel 2023, 12. [Google Scholar] [CrossRef]
- Porte, B.; Marguerit, G.; Thomasseau, S.; Paquet, C.; Hugon, J. Dose-dependent neuroprotective effect of the JNK inhibitor Brimapitide in 5xFAD transgenic mice. Brain Res 2020, 1727, 146587. [Google Scholar] [CrossRef]
- Yang, L.; Han, X.; Xing, F.; Wu, H.; Shi, H.; Huang, F.; Xu, Q.; Wu, X. Total flavonoids of astragalus attenuates experimental autoimmune encephalomyelitis by suppressing the activation and inflammatory responses of microglia via JNK/AKT/NFkappaB signaling pathway. Phytomedicine 2021, 80, 153385. [Google Scholar] [CrossRef]
- Ren, Q.; Zhao, S.; Ren, C.; Ma, Z. Astragalus polysaccharide alleviates LPS-induced inflammation injury by regulating miR-127 in H9c2 cardiomyoblasts. Int J Immunopathol Pharmacol 2018, 32, 2058738418759180. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Shao, C.; Li, W.; Wan, H.; He, Y.; Yang, J. Protective effects of Astragaloside IV against oxidative injury and apoptosis in cultured astrocytes by regulating Nrf2/JNK signaling. Exp Brain Res 2021, 239, 1827–1840. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wang, Y.; Qu, J.; Chen, B.; Dai, G.; Zhang, X.; Nie, K.; Mao, S.; Sun, H. Astragalus polysaccharide inhibits lipopolysaccharide-induced depressive-like behaviors and inflammatory response through regulating NF-κB and MAPK signaling pathways in rats. Int J Clin Exp Med 11 2361–2370.
- Liu, G.; Song, J.; Guo, Y.; Wang, T.; Zhou, Z. Astragalus injection protects cerebral ischemic injury by inhibiting neuronal apoptosis and the expression of JNK3 after cerebral ischemia reperfusion in rats. Behav Brain Funct 2013, 9, 36. [Google Scholar] [CrossRef]
- Yan, H.; Feng, L.; Li, M. The Role of Traditional Chinese Medicine Natural Products in beta-Amyloid Deposition and Tau Protein Hyperphosphorylation in Alzheimer’s Disease. Drug Des Devel Ther 2023, 17, 3295–3323. [Google Scholar] [CrossRef]
- Yu, S.Y.; Ouyang, H.T.; Yang, J.Y.; Huang, X.L.; Yang, T.; Duan, J.P.; Cheng, J.P.; Chen, Y.X.; Yang, Y.J.; Qiong, P. Subchronic toxicity studies of Radix Astragali extract in rats and dogs. J Ethnopharmacol 2007, 110, 352–355. [Google Scholar] [CrossRef]
- Chen, Z.; Liu, L.; Gao, C.; Chen, W.; Vong, C.T.; Yao, P.; Yang, Y.; Li, X.; Tang, X.; Wang, S.; et al. Astragali Radix (Huangqi): A promising edible immunomodulatory herbal medicine. J Ethnopharmacol 2020, 258, 112895. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
outline of experimental settings.
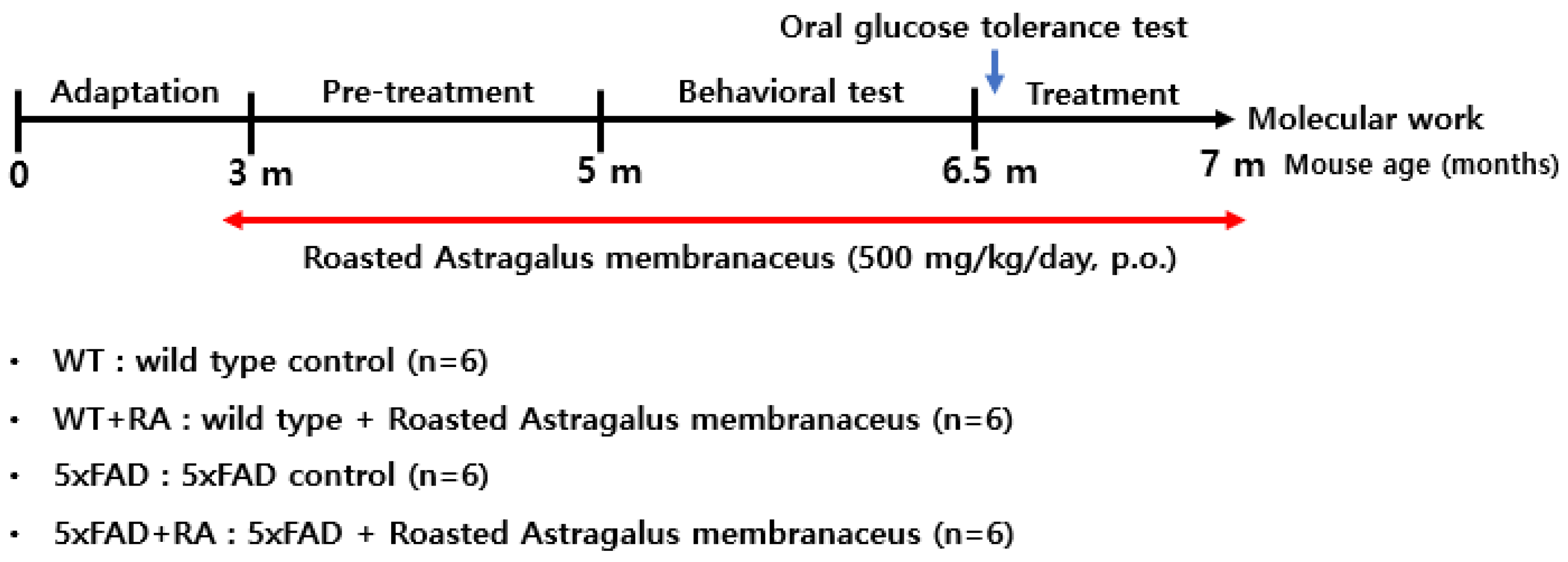
Figure 2.
Body weight and food intake during the experimental period. (A) Body weight changes over 12 weeks after RA administration (3 to 6 months of age). Repeated measure ANOVA, within group F(2.6, 52.5) = 14, p < 0.001, between group F(3, 20) = 0.5, p = 0.67, within-between interaction F(7.9, 52.5) = 1.5, p = 0.18. (B) Body weight change before (week 0) and after (12 weeks) RA administration. 5xFAD+RA initial vs final ; Paired t-test (t(5) = -4.6, p = 0.006. (C) Mean food intake during the experimental period. F(3, 44) = 3.3, p = 0.03; WT vs 5xFAD, p = 0.015. (D) Mean food efficiency ratio (weight gain divided by food intake) during the experimental period. F(3, 20) = 3.6, p = 0.03; WT vs 5xFAD, p = 0.012; 5xFAD vs 5xFAD+RA, p = 0.027. All values are mean ± S.E.M. WT: DW fed in wild type females, WT+RA: RA 500 mg/kg fed in wild type females, 5xFAD: DW fed 5xFAD females, 5xFAD+RA: RA 500 mg/kg fed 5xFAD females. * p<0.05, ** p<0.01, *** p<0.001 vs initial body weight of each group in (B), # p<0.05 vs WT, * p<0.05 vs 5xFAD in (C) and (D). ANOVA, LSD post-hoc test.
Figure 2.
Body weight and food intake during the experimental period. (A) Body weight changes over 12 weeks after RA administration (3 to 6 months of age). Repeated measure ANOVA, within group F(2.6, 52.5) = 14, p < 0.001, between group F(3, 20) = 0.5, p = 0.67, within-between interaction F(7.9, 52.5) = 1.5, p = 0.18. (B) Body weight change before (week 0) and after (12 weeks) RA administration. 5xFAD+RA initial vs final ; Paired t-test (t(5) = -4.6, p = 0.006. (C) Mean food intake during the experimental period. F(3, 44) = 3.3, p = 0.03; WT vs 5xFAD, p = 0.015. (D) Mean food efficiency ratio (weight gain divided by food intake) during the experimental period. F(3, 20) = 3.6, p = 0.03; WT vs 5xFAD, p = 0.012; 5xFAD vs 5xFAD+RA, p = 0.027. All values are mean ± S.E.M. WT: DW fed in wild type females, WT+RA: RA 500 mg/kg fed in wild type females, 5xFAD: DW fed 5xFAD females, 5xFAD+RA: RA 500 mg/kg fed 5xFAD females. * p<0.05, ** p<0.01, *** p<0.001 vs initial body weight of each group in (B), # p<0.05 vs WT, * p<0.05 vs 5xFAD in (C) and (D). ANOVA, LSD post-hoc test.
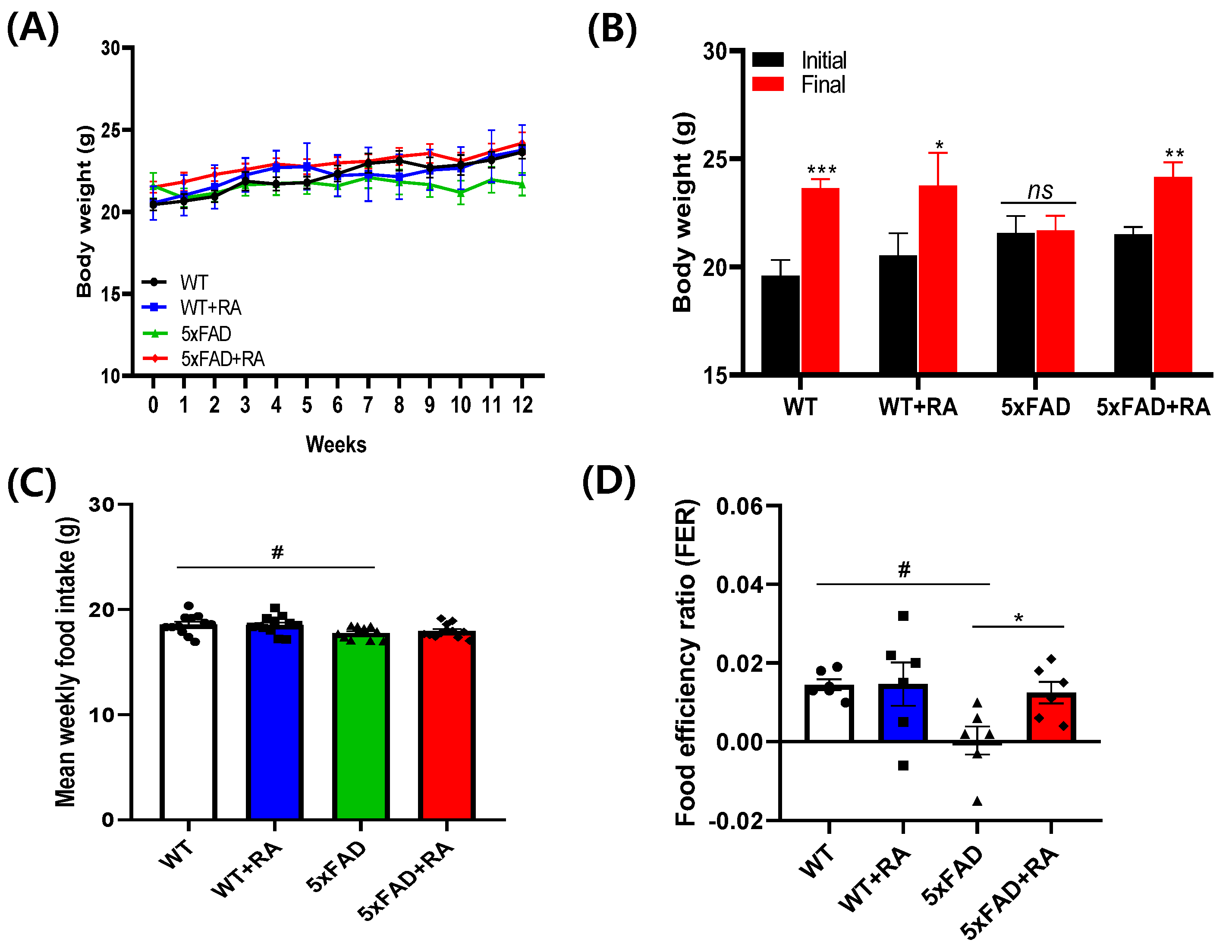
Figure 3.
Effects of RA on glucose metabolism on 5xFAD mice. (A) Non-fasting blood glucose after 12 weeks of RA treatment. F(3, 20) = 4.5, p = 0.015. (B) Six hour fasting blood glucose after 12 weeks of RA treatment. F(3, 20) = 9.7, p < 0.001. (C) Blood glucose concentration changes after 1g/kg of glucose injection in 6 hour fasted mice. Within group F(3.2, 64.6) = 198, p < 0.001, between group F(3, 20) = 8.3, p < 0.001, within-between interaction F(9.7, 64.6) = 2.9, p = 0.005. (D) Area under curve of oGTT. F(3, 20) = 5.8, p = 0.005. (E) fasting insulin level after 12 weeks of RA treatment. (F) HOMA-IR after 12 weeks of RA treatment. All values are mean ± S.E.M. WT vs 5xFAD, p = 0.012; 5xFAD vs 5xFAD+RA, p = 0.027. All values are mean ± S.E.M. WT: DW fed in wild type females, WT+RA: RA 500 mg/kg fed in wild type females, 5xFAD: DW fed 5xFAD females, 5xFAD+RA: RA 500 mg/kg fed 5xFAD females. # p<0.05, ### p<0.001 vs WT, ** p<0.01, *** p<0.001 vs 5xFAD. ANOVA and repeated measure ANOVA, LSD post-hoc test.
Figure 3.
Effects of RA on glucose metabolism on 5xFAD mice. (A) Non-fasting blood glucose after 12 weeks of RA treatment. F(3, 20) = 4.5, p = 0.015. (B) Six hour fasting blood glucose after 12 weeks of RA treatment. F(3, 20) = 9.7, p < 0.001. (C) Blood glucose concentration changes after 1g/kg of glucose injection in 6 hour fasted mice. Within group F(3.2, 64.6) = 198, p < 0.001, between group F(3, 20) = 8.3, p < 0.001, within-between interaction F(9.7, 64.6) = 2.9, p = 0.005. (D) Area under curve of oGTT. F(3, 20) = 5.8, p = 0.005. (E) fasting insulin level after 12 weeks of RA treatment. (F) HOMA-IR after 12 weeks of RA treatment. All values are mean ± S.E.M. WT vs 5xFAD, p = 0.012; 5xFAD vs 5xFAD+RA, p = 0.027. All values are mean ± S.E.M. WT: DW fed in wild type females, WT+RA: RA 500 mg/kg fed in wild type females, 5xFAD: DW fed 5xFAD females, 5xFAD+RA: RA 500 mg/kg fed 5xFAD females. # p<0.05, ### p<0.001 vs WT, ** p<0.01, *** p<0.001 vs 5xFAD. ANOVA and repeated measure ANOVA, LSD post-hoc test.
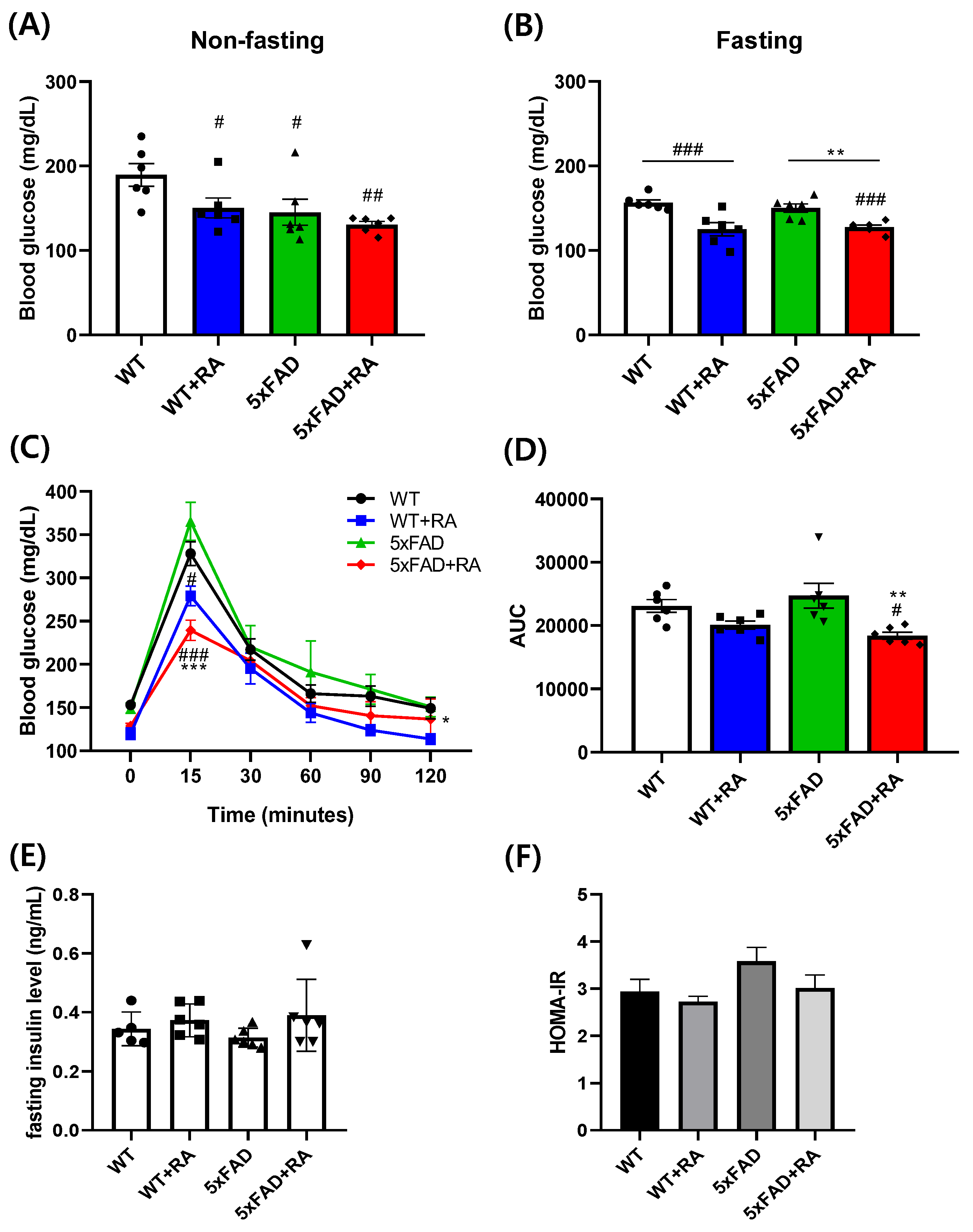
Figure 4.
Effects of RA on cognitive behaviors on 5xFAD mice. (A) Percent alternation in the Y-maze test. F(3,20) = 3.2, p = 0.04. (B) Latency to escape in the active avoidance test. Repeated measure ANOVA, within group F(3, 60) = 56, p < 0.001; between group F(3, 20) = 4.5, p = 0.014; within-between interaction F(9, 60) = 2.9, p = 0.007; 5xFAD vs 5xFAD+RA, p = 0.004. (C) Latency to find hidden platform in MWM test. Repeated measure ANOVA, within group F(2, 40) = 11.7, p < 0.001; between group F(3, 20) = 16.3, p < 0.001; within-between interaction F(6, 40) = 1.04, p = 0.41; WT vs 5xFAD, p <0.001; 5xFAD vs 5xFAD+RA, p<0.001. (D) Percent time in the platform area in MWM probe trial. F(3, 20) = 4.9, p = 0.01. (E) Distance moved in the open field test. (F) Percent time spent in the open arms of the EPM. F(3, 20) = 11.7, p <0.001. All values are mean ± S.E.M. WT: DW fed in wild type females, WT+RA: RA 500 mg/kg fed in wild type females, 5xFAD: DW fed 5xFAD females, 5xFAD+RA: RA 500 mg/kg fed 5xFAD females. # p<0.05, ## p<0.01, ### p<0.001 vs WT, * p<0.05, *** p<0.001 vs 5xFAD. ANOVA and repeated measure ANOVA, LSD post-hoc test.
Figure 4.
Effects of RA on cognitive behaviors on 5xFAD mice. (A) Percent alternation in the Y-maze test. F(3,20) = 3.2, p = 0.04. (B) Latency to escape in the active avoidance test. Repeated measure ANOVA, within group F(3, 60) = 56, p < 0.001; between group F(3, 20) = 4.5, p = 0.014; within-between interaction F(9, 60) = 2.9, p = 0.007; 5xFAD vs 5xFAD+RA, p = 0.004. (C) Latency to find hidden platform in MWM test. Repeated measure ANOVA, within group F(2, 40) = 11.7, p < 0.001; between group F(3, 20) = 16.3, p < 0.001; within-between interaction F(6, 40) = 1.04, p = 0.41; WT vs 5xFAD, p <0.001; 5xFAD vs 5xFAD+RA, p<0.001. (D) Percent time in the platform area in MWM probe trial. F(3, 20) = 4.9, p = 0.01. (E) Distance moved in the open field test. (F) Percent time spent in the open arms of the EPM. F(3, 20) = 11.7, p <0.001. All values are mean ± S.E.M. WT: DW fed in wild type females, WT+RA: RA 500 mg/kg fed in wild type females, 5xFAD: DW fed 5xFAD females, 5xFAD+RA: RA 500 mg/kg fed 5xFAD females. # p<0.05, ## p<0.01, ### p<0.001 vs WT, * p<0.05, *** p<0.001 vs 5xFAD. ANOVA and repeated measure ANOVA, LSD post-hoc test.
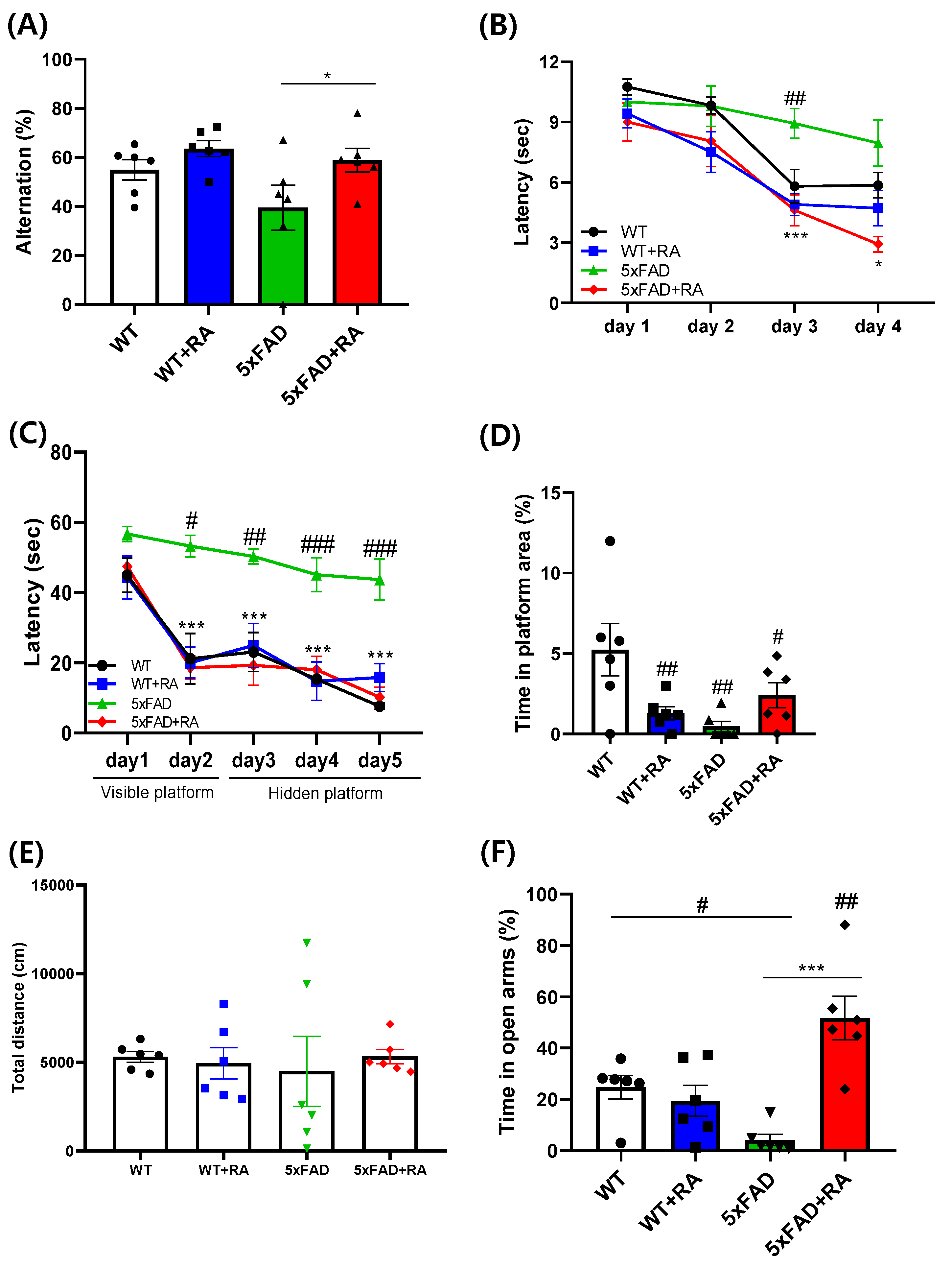
Figure 5.
Effect of RA on hippocampal expression on amyloid and Tau proteins in 5xFAD mice. (A) Normalized relative expression and representative image of APP. F(3,16) = 36.2 , p < 0.001. (B) Normalized relative expression and representative image of β-amyloid. No detectable band was observed in WT mice. F(3,16) = 0.73, p = 0.49. (C) Normalized relative expression of Tau. F(3,16) = 1.8, p = 0.18. (D) Normalized relative expression of p-Tau. F(3, 16) = 0.27, p = 0.85. All values are mean ± S.E.M. WT: DW fed in wild type females, WT+RA: RA 500 mg/kg fed in wild type females, 5xFAD: DW fed 5xFAD females, 5xFAD+RA: RA 500 mg/kg fed 5xFAD females. ### p<0.001 vs WT. ANOVA and LSD post-hoc test.
Figure 5.
Effect of RA on hippocampal expression on amyloid and Tau proteins in 5xFAD mice. (A) Normalized relative expression and representative image of APP. F(3,16) = 36.2 , p < 0.001. (B) Normalized relative expression and representative image of β-amyloid. No detectable band was observed in WT mice. F(3,16) = 0.73, p = 0.49. (C) Normalized relative expression of Tau. F(3,16) = 1.8, p = 0.18. (D) Normalized relative expression of p-Tau. F(3, 16) = 0.27, p = 0.85. All values are mean ± S.E.M. WT: DW fed in wild type females, WT+RA: RA 500 mg/kg fed in wild type females, 5xFAD: DW fed 5xFAD females, 5xFAD+RA: RA 500 mg/kg fed 5xFAD females. ### p<0.001 vs WT. ANOVA and LSD post-hoc test.
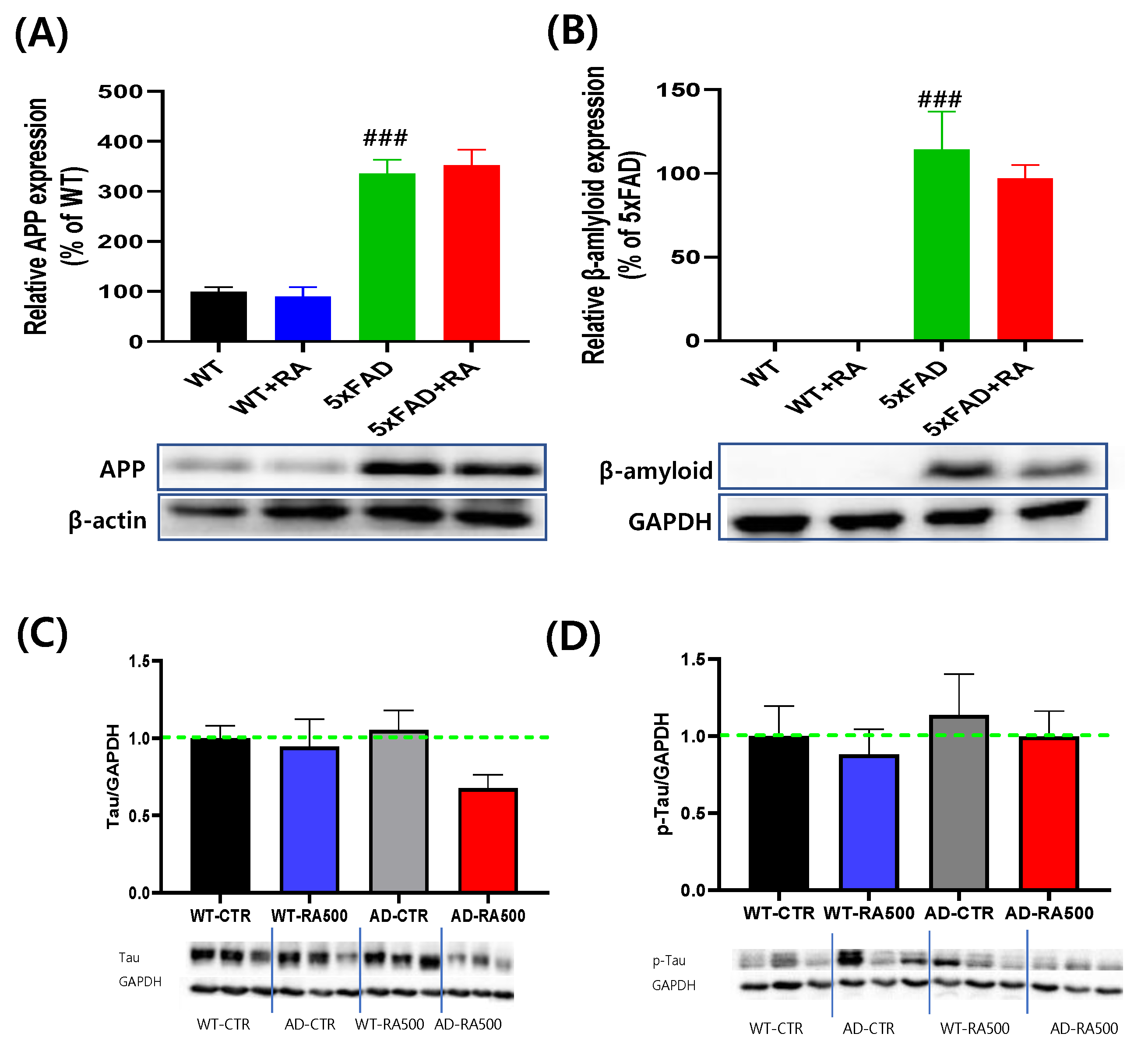
Figure 6.
Effect of RA on hippocampal expression of neurotrophic signaling pathways in 5xFAD mice. (A) Normalized relative expression and representative image of BDNF. F(3, 20) = 3.3, p = 0.04. (B) Relative expression and representative image of p-CREB/CREB F(3, 20) = 3.5 , p = 0.034. (C) Relative expression and representative image of p-JNK/JNK. F(3, 20) = 3.1 , p = 0.049. All values are mean ± S.E.M. WT: DW fed in wild type females, WT+RA: RA 500 mg/kg fed in wild type females, 5xFAD: DW fed 5xFAD females, 5xFAD+RA: RA 500 mg/kg fed 5xFAD females. # p<0.05 vs WT, * p<0.05 vs 5xFAD. ANOVA and repeated measure ANOVA, LSD post-hoc test.
Figure 6.
Effect of RA on hippocampal expression of neurotrophic signaling pathways in 5xFAD mice. (A) Normalized relative expression and representative image of BDNF. F(3, 20) = 3.3, p = 0.04. (B) Relative expression and representative image of p-CREB/CREB F(3, 20) = 3.5 , p = 0.034. (C) Relative expression and representative image of p-JNK/JNK. F(3, 20) = 3.1 , p = 0.049. All values are mean ± S.E.M. WT: DW fed in wild type females, WT+RA: RA 500 mg/kg fed in wild type females, 5xFAD: DW fed 5xFAD females, 5xFAD+RA: RA 500 mg/kg fed 5xFAD females. # p<0.05 vs WT, * p<0.05 vs 5xFAD. ANOVA and repeated measure ANOVA, LSD post-hoc test.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



