Submitted:
08 September 2025
Posted:
09 September 2025
You are already at the latest version
Abstract
Pyometra is a frequent and life-threatening reproductive disorder in bitches, characterized by profound immune and inflammatory responses within the uterus. The endocannabinoid system (eCS) is a key modulator of immune regulation, tissue homeostasis, and inflammation; however, its role in canine uterine physiology and pathology remains unexplored. This study aimed to characterize the presence and regulation of eCS components in the uterus of healthy and pyometra-affected dogs. Twenty-eight bitches were categorized into four groups: closed-cervix pyometra (CP; n=7), open-cervix pyometra (OP; n=7), diestrus (DE; n=7), and anestrus (AE; n=7). Uterine tissues were obtained by ovariohysterectomy. Serum progesterone (P4), anandamide (AEA), and 2-arachidonylglycerol (2-AG) concentrations were quantified, while uterine expression of cannabinoid receptors (CB1 and CB2) was assessed using real-time PCR and localized by immunohistochemistry (IHC). Serum AEA levels were significantly reduced in CP compared with AE (p=0.017), whereas 2-AG differences did not reach significance (p=0.072). Both CB1 and CB2 were consistently expressed across all groups, with IHC revealing receptor-specific patterns within uterine compartments. Collectively, these findings demonstrate for the first time that the canine uterus possesses a functional eCS, and that its modulation is linked to reproductive physiology and pyometra-associated inflammatory processes.

Keywords:
dog
; endocannabinoid system
; pyometra
1. Introduction
The endocannabinoid system (eCS) is a major factor involved in homeostasis maintenance, inflammation, pain, and both male and female reproductive processes [1]. Therefore, eCS compounds are expressed in various cells in the brain and peripheral tissues, including the uterus, leukocytes, spleen, heart, adrenal glands, ovaries, endometrium and testes in humans and several other mammalian species [2,3]. The eCS is defined as a neuromodulator network, which includes endocannabinoids (ECBs), cannabinoid receptors (CBs), and ECB enzymes [1,2]. Endocannabinoids are bioactive lipids acting on CB1 and CB2, which are the best characterized receptors of the eCS [1]. The most extensively studied ECBs are N-arachidonoylethanolamine (anandamide; AEA) and 2-arachidonoylglycerol (2-AG). Anandamide mediates its biological effects predominantly via CB1, whereas 2-AG signals primarily through CB2 or through both CB1 and CB2, with the potential of both ECBs activating either receptor depending on the tissue-specific context [4]. While CB1 is localized predominantly to the nervous system and expressed in various types of neurons, including glutamatergic, cholinergic, glycinergic, and serotonergic neurons, CB2 is expressed predominantly in the immune system cells, particularly in microglia, lymphocytes, macrophages and neutrophils [1]. Endocannabinoids are synthesized exclusively on demand and are not stored [5]. Their synthesis, degradation, and transport processes are quite complex and have not yet been fully elucidated, but ECB enzymes are known to undertake important roles in the eCS: AEA is synthesized by N-acyl-phosphatidylethanolamine phospholipase D (NAPE-PLD) and hydrolyzed by fatty acid amide hydrolase (FAAH), whereas 2-AG is synthesized by diacylglycerol lipases (DAGL) and hydrolyzed by monoacylglycerol lipase (MAGL) (1). In addition, arachidonic acid, released during the degradation of ECBs, acts as a substrate for cyclooxygenase-2 (COX-2)-mediated oxidative metabolism, leading to the formation of prostaglandin (PG) compounds such as PG-ethanolamides or prostamides (PM). These PG-derived PMs play diverse roles in inflammation, pain, parturition, and reproductive processes [7].
The relationship between the eCS and reproduction is a subject that is still being studied in women and rodents. It has been determined that eCS components play important roles in the menstrual/estrus cycles, ovulation, spermatogenesis, fertilization, implantation, and pregnancy in humans and rodents [1,8,9]. Endocannabinoid components such as CBs and ECB and its enzymes are widely expressed in women and rodent ovarian and uterine tissues [2,3]. Furthermore, the presence of eCS components such as CB1 in the hypothalamus and adenohypophysis provides evidence for a close relationship between the eCS and hypothalamus-pituitary-ovary (HPO) triangle [3,8]. Indeed, it has been shown that a CB1 agonist completely suppresses the pulsatile release of GnRH [8], and that the direct administration of AEA into the brain ventricles similarly inhibits GnRH secretion [10]. The eCS is also related to ovarian hormones [1]. In women, FAAH activity is suppressed by estradiol and increased by P4, indicating that AEA degradation is affected by these hormones [11,12]. Therefore, serum AEA concentrations are high during the follicular phase and low during the luteal phase in women [11]. Anandamide concentrations typically rise during ovulation, peaking in both the follicular fluid and serum in association with LH surges, and then decline during the luteal phase [1]. In mice, abnormal ECB signaling disrupts the physiological sperm–oocyte interaction, thereby contributing to fertility impairments [1]. In the mouse, low AEA concentrations promote implantation, whereas high levels cause inhibition [13].
The eCS has also been shown to be involved in reproductive pathologies in women [14] and mice [15]. In women with endometriosis, CB2 signaling leads to a reduction in pro-inflammatory cytokine production by macrophages and dendritic cells [16]. In mouse models, CB2 deficiency has been associated with endometriosis-specific alterations in T cell function [15]. Elevated AEA levels may disrupt ECB signaling in the uterus via CB1, thereby increasing the risk of abortion, whereas low ECB levels support embryo implantation and the maintenance of pregnancy through CB1 and CB2 as shown in mouse models [1].
The eCS also emerges as an important regulator in the immune response to sepsis, which is usually triggered by bacterial infections [17,18,19]. This regulatory role is quite complex and has not been clearly elucidated yet. However, toll-like receptors (TLRs), lipopolysaccharide (LPS), PG synthesis and the eCS are known to be associated with each other [20]. Toll-like receptors are part of the innate immune system and are among the first receptors to maintain normal immune homeostasis. They are expressed across a broad spectrum of immune and non-immune cells, including macrophages, monocytes, dendritic cells, lymphocytes, as well as endothelial and epithelial cells. Suppressed or overactivated immune responses mediated by TLRs impair host defense against pathogens and play a key role in the pathogenesis of sepsis. Toll-like receptor-4 recognizes endotoxin, which is a cell wall component of Gram-negative bacteria. One of the best-known bacterial ligands is LPS, another cell wall component of Gram-negative bacteria [21]. Briefly, TLRs, particularly TLR-4, play a critical role in initiating the inflammatory response by recognizing LPS. In other words, LPS upregulates TLR-4 expression in the uterus. Lipopolysaccharides also induces PG synthesis by activating COX-2 via TLR-4 [22]. Toll-like receptor-4 stimulation induces firstly COX-2 transcription and then PG synthesis, with its expression undergoing rapid induction in response to a wide array of pro-inflammatory mediators and pathogenic challenges, reflecting its role as an early-response gene in inflammatory signaling pathways [23].
Since macrophages constitute the principal source of ECBs, TLR-mediated macrophage activation has the capacity to modulate both CB expression and ECB concentrations [20]. For instance, decreased circulating levels of AEA and 2-AG in patients with sepsis have been associated with prolonged hospitalization [18]. The pharmacological blockade of CB1 antagonists has been shown to elevate blood pressure in septic rats and to attenuate hypothermia in systemic infection models. These findings suggest that CB1 contributes to blood pressure regulation, thermoregulation, and nociceptive responses during sepsis [17,19]. In contrast, CB2 deficiency has been linked to reduced survival, exaggerated inflammatory responses, increased serum IL-6 concentrations, and pulmonary injury. Consistently, the administration of CB agonists has been reported to improve survival by reducing inflammation and bacteremia in experimental mouse sepsis models [19]. Lipopolysaccharide increases CB1 mRNA expression while decreasing CB2 mRNA expression in the mouse uterus [24]. Briefly, elevated concentrations of ECBs have been shown to decrease the LPS-induced release of pro-inflammatory mediators [25]. These effects are mediated through CB1/2-dependent mechanisms [26]. It has been proposed that AEA and 2-AG contribute to the maintenance of homeostasis by modulating CB1/2 expression, stimulating the production of anti-inflammatory cytokines, and suppressing pro-inflammatory cytokine levels [1,16,17,18,19].
Pyometra has been proposed as a clinical model for sepsis, as the genetic and pathological resemblance between humans and dogs suggests that its molecular features may reflect those of sepsis in humans [27]. It represents the most common and life-threatening reproductive disorder in dogs, typically developing during the luteal phase and affecting mostly older animals [28]. Despite its clinical relevance, the underlying etiology is still not fully understood [28,29]. Pyometra is primarily a bacterial disease in which the prolonged and repeated exposure of the uterus to P4 skews the balance between the uterine immune defenses and bacterial challenge towards bacterial persistence [28,30]. In the etiology of pyometra, both cystic endometrial hyperplasia (CEH) and pseudoplacentational endometrial hyperplasia (PEH) may play important roles, as both are P4-related phenomena. Cystic endometrial hyperplasia, an age-associated pathology, has been suggested to facilitate bacterial colonization [28]. However, the long-standing paradigm that CEH is a prerequisite for pyometra has been challenged, since both conditions may occur independently [28,31,32]. Pseudoplacentational endometrial hyperplasia refers to decidual changes within the superficial endometrial epithelium that histologically mimic changes observed in the placentation sites of normal pregnancy during canine pseudopregnancy [33,34]. In dogs, the non-pregnant diestrus phase is characterized by a state of pseudopregnancy, reflecting the species-specific physiological condition in which luteal function is maintained regardless of conception [35]. Similar to CEH, recurrent PEH may also create a microenvironment favorable for bacterial colonization [34].
Bacteria and their endotoxins act as potent stimuli, triggering a robust inflammatory response in the uterus. Transcriptomic studies have demonstrated that more than 800 genes are upregulated, whereas over 700 genes are downregulated during the inflammatory process [27]. The majority of the upregulated genes are associated with chemokine and cytokine signaling, inflammatory cell extravasation, antibacterial defense mechanisms, the complement cascade, and innate immunity. Notably, these include genes encoding TLR-4, ligands for LPS recognition, and key enzymes involved in PG synthesis [21,22,36]. Such gene expression patterns are consistent with the observation that Escherichia coli (E. coli) is the most frequently isolated bacterium in pyometra cases and serves as a major source of LPS [22].
In dogs, studies on the eCS are scarce, with existing research being limited to peripheral tissues, including the skin [37], gastrointestinal tract [38], hair follicles [39], salivary glands [40], and central nervous system [41], all confirming the presence of eCS components. Despite evidence from human and experimental animal studies implicating the involvement of the eCS in sepsis and menstrual/estrous cycle regulation, the role of this system in canine reproductive physiology and pathology remains unexplored. Building on this foundation, we hypothesize that the eCS is similarly expressed in the canine uterus and may play a functional role in both normal physiology and pathological states such as pyometra. Accordingly, the present study was designed to systematically characterize the expression and localization of key eCS components in canine uterine tissue from pyometric animals and healthy controls, providing the first comprehensive evaluation of this system in the canine reproductive tract.
2. Materials and Methods
2.1. Animals and Groups
The study was carried out on the uterine tissue and blood samples of 28 dogs. The dogs included in the study were of different breeds and body weights (average 13.75 kg; 7.3-32.5 kg) and of an average age of 5.35 years (1-12 years) and were admitted to the Animal Hospital of Ondokuz Mayıs University Veterinary Faculty for pyometra treatment and elective ovariohysterectomy (OHE). All procedures were approved by the Local Ethics Committee for Animal Experiments of Ondokuz Mayıs University (HADYEK: 2022/47).
Animals were assigned to groups based on exfoliative vaginal cytology, serum P4 measurements, ultrasonographic examination and uterine histomorphology. Pyometra was diagnosed using ultrasonography in conjunction with elevated serum P4 concentrations (>10 ng/mL), and animals were further classified according to vaginal discharge into those with closed cervix pyometra (CP; n=7) and open cervix pyometra (OP; n=7). Healthy animals were identified based on their general and reproductive health, exfoliative vaginal cytology, and serum P4 concentrations (>10 ng/mL for diestrus and <1 ng/mL for anestrus), and were assigned to the diestrus (DE; n=7) or anestrus (AE; n=7) groups. Ovariohysterectomy was performed in all animals, and the histomorphological examination of the uterine tissues collected post-surgery confirmed the clinical diagnoses, thereby validating the group assignments.
2.2. Blood Sampling and Measurements of Progesterone and Endocannabinoids
Blood samples were collected from the cephalic antebrachial vein of each animal into an EDTA tube and a gel separator tube (BD Vacutainer®, Franklin Lakes, NJ, USA) just before surgery. The samples were centrifuged at 5000 rpm for 10 min (M615M, Elektro-mag, İstanbul, Türkiye) and the obtained sera were stored at -20◦C until P4, AEA and 2-AG measurements were performed.
Serum P4 measurements were performed by the electrochemiluminescence immunoassay using a Cobas Modular E170 fully automatic analyzer at the laboratory of the TÜRKAK-accredited (TS EN ISO/IEC 17025:2005) Düzen Laboratories Group, Ankara, Türkiye.
Anandamide and 2-AG were analyzed by liquid chromatography-electrospray ionization-tandem mass spectrometry (LC-ESI-MS/MS) using a Shimadzu LC-20AXR system coupled with a tandem mass spectrometer (Shimadzu 8030 MS/MS). The mass spectrometric detection was operated in the positive electrospray ionization and multiple reaction monitoring (MRM) mode.
Separations were carried out using GLS-ODS-4 C18 (50 x 3.0 mm; 2.1 μm) at a flow rate of 0.3 mL/min and at 40 °C. A gradient elution program with mobile phase A [0.1% (v/v) formic acid in water] and B [0.1% (v/v) formic acid in acetonitrile] was performed. The sample injection volume was 20 μL and the analytical run time was 10 min. The ESI-MS/MS conditions were as follows: interface voltage, 4.5 kV; Q1 pre-rod bias voltage, −13 V; Q3 pre-rod bias voltage, −23 V; collision energy, −14 eV; nebulizer gas flow rate, 3 mL/min; drying gas flow rate, 15 L/min; desolvation line temperature, 250 °C; and heat block temperature, 400 °C. The transitions 344.30 → 203.10 m/z and 379.00 → 287.20 m/z were used to monitor AEA and 2-AG, respectively.
Polymeric (Strata X, 30 mg/1mL) solid phase cartridges were used. The 20 µL serum samples were diluted to 1 mL with ultrapure water and prepared for analysis by following the solid phase extraction (SPE) procedure using acetonitrile and methanol. Calibration solutions containing AEA and 2-AG standards at increasing concentrations (1-1000 ng/mL) and an internal standard (ACPA) at a constant concentration were prepared under the same SPE conditions as the samples and analyzed by the LC-MS/MS method. As a result of the analyses, calibration curves were created and the amounts of AEA and 2-AG in the serum samples were calculated using the equations of these calibration curves.
2.4. Uterine Tissue Sampling and Tissue Processing
Uterine tissues were collected by a ventral midline ovariohysterectomy performed under general anesthesia, a protocol commonly used in veterinary practice. After premedication with propofol (1% Propofol, 200 mg/20 mL, IV injection/infusion emulsion, Polifarma, Türkiye), inhalation anesthesia was initiated with isoflurane (Isoflurane USP®, Adeka, Primal, USA). Intravenous fluid (Lactated Ringer's Solution, 10 mL/kg/hour, Polifarma® 500 mL, Ankara, Türkiye) and constant rate infusion (12 mg morphine, 30 mg ketamine and 150 mg lidocaine diluted in 500 mL saline solution, 10 mL/kg/hour) were administered during the surgery. Post-operative care was provided, including intravenous fluid treatment, antimicrobial treatment (cefazolin sodium; 30 mg/kg/day, IM, Iespor® 500 mg vial; I.E. Ulugay, Türkiye) and supportive treatments tailored to the needs of the pyometric and healthy animals. All dogs received analgesic treatment (meloxicam; 0.2 mg/kg, SC, Maxicam® 5 mg/mL; Sanovel, Türkiye) for three days following OHE.
Immediately after surgery, a swab sample was taken under sterile conditions from the uterine lumen of each dog for microbiological evaluation and those showing bacterial growth were evaluated as cases of pyometra. Two tissue samples (1 x 1 cm) were taken from the middle part of the left uterine horn: One was fixed in 4% buffered formal overnight at +4◦C for subsequent paraffin embedding [42], histomorphological evaluation and further immunohistochemistry procedures; and the other was frozen at −196◦C and stored at −80°C for further analysis by quantitative PCR (RT-qPCR).
2.5. RT-qPCR Analysis
The extraction of total RNA was performed using the TRIzol reagent (Invitrogen, Carlsbad, CA, USA) following the manufacturer’s protocol. The quality and quantity of the isolated total RNA were measured with a NanoDropTM 2000C spectrophotometer (Thermo Fisher Scientific, Wilmington, DE, USA). The concentrations were fixed at 1000 ng for use in the next steps of the procedures. The RNA samples were reverse transcribed (RT) into cDNA according to the manufacturer’s instructions (İScriptTM cDNA synthesis kit, Bio-Rad Laboratories, USA) and stored at -20°C until PCR analysis.
The primers for the two target genes (CB1 and CB2) and three reference genes (KDM4A, PTK2 and EIF4H) were designed using Primer Express Software version 3.0 (Applied Biosystems by Thermo Fischer) (Table 1), and were synthesized by the Macrogen-Oligo Company (Geumcheon-gu, Seoul, Korea).
The mRNA transcript levels of the target and reference genes were determined using the SsoAdvancedTM Universal SYBR® Green Supermix kit (BIO-RAD Laboratories, USA) in accordance with the protocol of the manufacturing company. Subsequently, cDNA expression in the samples, including the negative control (NTC), was determined by triplicate measurements with the optimized RT-qPCR method, according to the manufacturer's instructions and depending on the primer annealing temperature, using the CFX®96 Real-Time PCR system of BIO-RAD. In the final step, the target genes of CB1 and CB2 were evaluated by the comparative CT method (ΔΔCT method or Livak method). This method involves the calculation of the ΔΔCt value using an arithmetic formula, comparing the CT value of the target gene with the result of the endogenous reference gene, ultimately expressed as 2^-(ΔΔCt).
2.6. Immunohistochemical Staining
The standard immunoperoxidase method was used as described by Kowalewski et al. (2010) [42]. Accordingly, all uterine tissue samples taken from the bitches with pyometra during DE and AE, were fixed in 4% formalin and embedded in paraffin so that cross-sections (2–3 µm thick/sections) were taken (Leica Rm 2125Rt, Leica Biosystems, USA), stained and mounted on SuperFrost Plus microscope slides (Menzel-Glaeser, Braunschweig, Germany).
The sections were examined under an Olympus BX51 microscope (Tokyo, Japan). The immunolocalization of CB1 and CB2 was detected by the streptavidin-biotin immunoperoxidase complex technique. Accordingly, the tissue sections were deparaffinized in xylene, rehydrated through a graded series of ethanol, treated with 10 mM sodium citrate (pH 6.0) at 600W in a microwave for 15 min, washed several times in phosphate buffered saline (PBS), each time for 5 min, treated with 0.3% hydrogen peroxide in methanol for 20 min to quench endogenous peroxidase activity, incubated with Ultra V Block (Thermo Fisher Scientific Lab Vision Corporation, Fremont, CA, USA; TA-125UB) at room temperature for 5 min to block non-specific antibody binding, and finally incubated with primer antibodies (Table 2) overnight at +4°C. For the negative controls, the slides were incubated with nonimmunized IgGs (isotype controls) of the same species and protein concentration as that of the primary antibody. Furthermore, tissues incubated with PBS without primary antibodies were also used as negative controls. After incubation the slides were washed in PBS for 20 min, incubated with secondary anti-rabbit goat IgG antibodies for 20 min, and washed in PBS for another 20 min. The slides were incubated with streptavidin (Thermo Scientific, Ultravision kit, TM-125-HL, USA) for 20 min, and 3,3-diaminobenzidine (DAB; Thermo Fisher Scientific Lab Vision Corporation) was used to detect peroxidase activity in the sections. Finally, the slides were counterstained with Gill's hematoxylin for 5 min, washed under running tap water, and mounted in Entellan (Entellan®, Merck, 1079610500, Darmstadt, Germany).
The observation of brown precipitation in the tissue sections was considered as positive immunostaining. The description and quantitative evaluation of immunostaining were made blindly by three different investigators, and the final results for immunoreactivity at high magnification (bar: 20μm-X40) and low magnification (bar: 50/100μm-X10/X20) were obtained by calculating the average of these investigators’ results. The following criteria were used for the evaluation of immunoreactivity: the absence of cellular immunoreactivity at high magnification indicated a negative (-) reaction. The presence of cellular immunoreactivity at high magnification was categorized as a weak reaction (+/-), and at low magnification was considered as a strong reaction (+).The clear observation of cellular immunoreactivity at low magnification was considered to be a very strong reaction (++).
2.7. Statistical Analyses
Data analyses were performed using the IBM SPSS 25.0 package program (SPSS®, IL, USA). Graphs were generated using Microsoft Office Excel 2019 (Microsoft Corporation, Redmond, WA, USA). The Shapiro-Wilk test was used to assess the normality of data distribution. The Student t and Mann-Whitney U tests were performed for two-group comparisons. When comparing four groups, one-way analysis of variance (One-Way ANOVA) was performed for normally distributed data, and the Kruskal-Wallis test was performed for data that did not show a normal distribution. Pearson’s and Spearman’s correlation tests were used to identify potential correlations between the parameters. Tukey’s honestly significant difference (HSD) test, Tamhane’s test and the Mann-Whitney U test with the Bonferroni correction were used for pairwise comparisons (post-hoc). The results are given as mean ± standard error of mean (SEM). Values of p<0.05 were considered statistically significant.
3. Results
3.1. Mean Ages and Body Weights of the Animals
There were statistically significant differences between the mean ages and body weights of the groups (Table 3).
3.2. Histomorphology of the Uterine Tissues and Microbiological Identification
All the tissue samples collected from the dogs with pyometra underwent histomorphological examination and bacterial identification. Accordingly, cystic endometrial hyperplasia (CEH) was detected in all the samples collected from the pyometric dogs (Figure 14/14; 100%). Furthermore, E. coli was isolated from all the samples (14/14; 100%) belonging to the pyometra cases, but no bacteria were isolated from Groups DE and AE.
3.3. Serum Progesterone Concentrations
While the lowest serum P4 concentration was detected in the AE group (0.52±0.05 ng/mL), the highest concentration was measured in the DE group (16.63±7.30 ng/mL). In brief, the serum P4 concentrations of the CP, OP and DE groups were found to be significantly higher than those of the AE group (between p = 0.004 and p = 0.044) (Figure 1).
3.4. Serum Anandamide and 2-AG Concentrations
Serum AEA concentrations were highest in the OP group and lowest in the CP group, but statistically significant differences were observed only between the CP and AE groups (p=0.010) (Figure 2).
The 2-AG concentrations were found to be lower from Group CP to Group AE, yet no statistically significant difference was determined between the groups (p=0.072) (Figure 2).
3.5. Gene Expressions of CB1 and CB2
It was determined that all of the target genes were expressed at different levels in all of the pyometra cases (CP and OP) and cyclic uterine tissues (DE and AE). The expression of CB1 significantly differed between the groups, such that it was significantly higher in the pyometra groups, and in the DE group compared to the AE group (Figure 3). On the other hand, no statistically significant difference was determined for the expression of the CB2 genes.
3.6. Localization of CB1 and CB2
Varying degrees of immunoreactivity for CB1 and CB2 were observed in the different layers of the uterus in all groups (Figure 4).
It was observed that CB1 immunoreactivity decreased from the surface to the depths of the uterine tissue in the CP, DE and AE groups, but not in the OP group. Immunoreactivity for CB1 was very strong in the luminal epithelial cells and endometrial gland cells of the uterus in the CP, DE and AE groups. It was also very strong in the immune cells and myometrium of the OP group. While CB1 immunoreactivity in the deep uterine glands was weak in Group CP, it was strong or very strong in the other groups. On the other hand, there was similar CB2 immunoreactivity in all the uterine layers in all of the groups (Figure 5).
4. Discussion
To the best of our knowledge, this is the first study to demonstrate the presence of the eCS in the canine uterus. The majority of previous investigations on the relationship between reproductive physiology or pathology and the eCS have focused on women or experimental animal models. In women, marked increases in glandular epithelium and endometrial vascular endothelial cells occur during the luteal phase of the menstrual cycle [43], which corresponds to the DE period in dogs [44]. Although endometriosis and pyometra represent two different conditions, both share the involvement of E2 and P4 in their etiology [45]. Owing to their shared environment, comparable medical care, and similar aging patterns, dogs also serve as a valuable translational model for several human diseases, including reproductive disorders [46]. Accordingly, comparisons of eCS alterations between women and dogs appear well justified.
Serum ECB concentrations are influenced by multiple factors, including nutrition, physical activity, sex hormones, age, inflammation, and pregnancy [47]. In the present study, nutrition and physical activity were negligible, as all animals were companion dogs maintained on commercial diets. Thus, our discussion focuses primarily on the influence of hormonal status, age, inflammation, and decidualization.
4.1. Effect of Hormonal Status
Serum P4 concentrations were within the expected physiological range in all groups, being elevated in the pyometra cases and during DE and at basal levels during AE [48]. The influence of sex hormones on the eCS remains controversial. For example, in women with endometriosis, luteal-phase AEA levels are higher compared to both the follicular phase of endometriotic women and the luteal phase of healthy controls [14]. Higher CB1 expression has been reported in the fallopian tubes of women during the luteal phase, with P4 treatment inducing further upregulation [9,49]. However, other reports found no difference in AEA or 2-AG concentrations between the follicular and luteal phases of healthy women [14,50] and no effect of P4 on circulating ECB concentrations [11]. Uterine CB1 expression in women has been reported to be at low levels during the follicular phase [51].
The present study also highlights this complexity, as serum 2-AG concentrations were determined to have increased in the P4-dominant groups, while AEA concentrations had not. This discrepancy may be explained by two factors: (i) differences in the biosynthetic and degradation pathways of these mediators (1) and (ii) the effects of pre-analytical handling, including blood collection, storage, and processing [52]. Although the validity of the measurement method used here was previously confirmed [52], it has been reported that AEA and 2-AG concentrations may increase during the interval between blood collection and serum separation, even when samples are kept on ice. Moreover, prolonged storage at –80°C appears to preserve 2-AG but not AEA, which continues to increase over time. Therefore, serum AEA and 2-AG are unlikely to serve as reliable clinical biomarkers [53]. Additionally, serum levels do not necessarily reflect tissue concentrations, as shown in reproductive tissues [54].
In our study, CB1/2 mRNA expression was numerically or statistically higher in the P4-dominant groups, suggesting that P4 may upregulate CB1/2 expression despite the absence of a direct correlation.
In this context, decidualization requires consideration, as it is a P4-driven process critical for the establishment of pregnancy [55]. Unlike in humans, decidualization in rats and dogs is known to be embryo-dependent [56]. Nonetheless, decidual-like changes, such as PEH, are observed during canine diestrus/pseudopregnancy [33]. Moreover, some authors have described CEH as a deciduoma, reflecting a proliferative response that mimics decidual implantation sites [33,34,57].
The expression of eCS components has been demonstrated at both the mRNA and protein levels during decidualization and tissue remodeling in women and rats, although its precise role remains unclear [9,58]. The upregulation of CB1 mRNA has been shown in human [9] and rat decidua [54,59], and immunoreactivity for CB1/2 and other eCS proteins has been observed in human decidua [60]. These findings suggest a potential link between CB1 upregulation and peak decidual development [59]. This could explain the CB1/2 mRNA expression patterns observed in our DE group. Given that all nonpregnant bitches in DE are physiologically pseudopregnant, regardless of clinical signs [35], we propose that the increased CB1/2 expression detected in this study may be related to pseudoplacentational changes occurring during pseudopregnancy. We recognize that our interpretation above is speculative because we did not conduct a histological evaluation of PEH in the DE group. However, we believe that the increasing number of publications on PEH and their supporting data strengthen our speculation.
4.2. Effect of Age
In this study, the dogs in the pyometra groups were significantly older and heavier than those in the control groups. This finding is not unexpected, as pyometra is an age-associated pathology, whereas dogs in the control groups had been electively neutered at a young age. Nevertheless, age itself should be considered a critical contributing factor for the results obtained in the pyometra groups.
Throughout life, endometrial tissue undergoes continuous cycles of degeneration and regeneration under the influence of the HPO axis, referred to as endometrial plasticity [61]. A fundamental component of this plasticity is endometrial cell migration, which plays a central role in wound closure, prevention of excessive fibrosis, and facilitation of embryonic trophoblast invasion [62].
Stromal aging, defined as the irreversible arrest of the cell cycle [63], has been documented in canine CEH [64] and disrupts endometrial elasticity, thereby increasing susceptibility to pathological changes [65]. The high recurrence rate of CEH despite medical management [66] further supports the concept of irreversible alterations in stromal cell cycling. In the present study, CEH was observed in all pyometra cases, underscoring its relevance in this context.
Components of the eCS exhibit dynamic, phase-specific regulation throughout the estrous cycle. In mice, eCS expression varies across the prepubertal, adult, late reproductive, and post-reproductive stages, reflecting both age- and organ-specific modulation in the endometrium [67]. The metabolic enzymes NAPE-PLD, FAAH, and DAGL-β exhibit the highest expression levels in the ovarian, oviductal, and uterine tissues, and these levels gradually increase with advancing age [67].
No significant differences were observed in CB1 mRNA expression in the uterine tissue between aged animals with pyometra and young animals in the DE and AE groups. These findings suggest that age alone may not directly influence CB1 or CB2 expression. Nevertheless, the potential impact of cellular senescence cannot be overlooked, as senescent cells persist within the uterus. The physiological and pathophysiological factors driving uterine cellular senescence, and their relationship with the endocannabinoid system, need to be elucidated.
4.3. Inflammatory Status
Previous studies have demonstrated that TLR-2 and TLR-4 mRNA are involved in the uterine immune response to E. coli [21,22]. Specifically, uterine TLR-4 transcription is upregulated 2.4-fold in cases of pyometra compared to the DE phase of the estrus cycle [21]. Crosstalk between TLR-4 and ECB signaling has also been reported in the uterine tissue. In the mouse uterus, LPS increases CB1 mRNA expression while decreasing CB2 expression [24], and the pro-apoptotic effects of LPS are abolished by a CB1 agonist in pregnant mice [68]. Endocannabinoid system components are integral to macrophage-mediated inflammatory responses to TLR-4 stimulation [69], with macrophages being the main producers of ECBs [20]. Consequently, TLR-mediated macrophage activity modulates CB and ECB expression, as evidenced by the LPS-induced upregulation of CB1 in mouse macrophages [69]. In contrast, LPS-induced CB2 expression in rat macrophages is controversial, with studies reporting both increases [70] and decreases [71]. Collectively, CB1 appears to inhibit TLR-4-mediated cytokine production [17], while CB2 may antagonize TLR-4 signaling [72]. These mechanisms likely underlie the CB1/2 mRNA expression observed in the pyometra groups, whereas the expression patterns in the DE and AE groups cannot be fully explained by LPS stimulation alone.
Toll-like receptor expression is also present during the DE and AE phases, with higher levels being observed during DE [22]. Two potential factors may contribute. Firstly, TLR-4 can be activated by sterile inflammation [73], which may explain its increased expression during DE [74]. During DE, stromal leukocyte numbers are elevated [75], likely contributing to TLR-4 expression and suggesting coordinated endometrial defense against microbial invasion [76]. Secondly, TLR signaling regulates apoptosis via anti-apoptotic proteins or inhibitors, consistent with apoptosis occurring during DE [22]. Nonetheless, these factors alone do not explain CB1/2 expression during AE, when CB1 is low and CB2 is high. The elevated CB2 expression may reflect leukocyte activity, as CB2 is strongly expressed in both human and canine leukocytes, despite their low numbers in the uterus during AE [77,78,79].
Cyclooxygenase-2 expression is inducible by inflammatory stimuli, growth factors, and pathological conditions [80]. It is overexpressed in pyometric uteri, compared to the healthy uterus [30,81,82,83,84], but this does not lead to increased myometrial contractions, likely due to elevated serum P4 concentrations. This suggests that high PG expression in pyometra is linked primarily to inflammation [21,82,83]. Additionally, COX-2 may interact with the ECB system, as COX-2 has substrate affinity for ECBs [85], and ECBs exert potent relaxant effects via CB1, and to a lesser extent CB2, in both women [86] and mice [87]. Cyclooxygenase-2 is also highly expressed in the luteal endometrium of women [43], rats [88], and dogs [44]; contributing to physiological endometrial adaptation and tissue homeostasis. Notably, the upregulation of COX-2 and PGE synthase (PGES) in pyometra cases, and during the DE and AE stages coincides with TLR-2/4 upregulation, and LPS stimulation synchronously increases COX-2 and mPGES-1 expression, enhancing PGE2 synthesis [23]. Prostaglandin E2 exerts immunosuppressive effects on lymphocytes, monocytes/macrophages, and polymorphonuclear cells [89].
Immunohistochemically, TLR-4 localizes to different endometrial compartments in association with leukocyte populations, cytokines, and sex hormones. In cases of pyometra, the surface epithelium exhibits higher TLR-4 expression than the stroma, glandular epithelium, or healthy uterus. During DE and AE, the glandular epithelium expresses TLR-4 more intensely than the other compartments, with lower expression during AE than DE [22]. Cyclooxygenase-2 shows a similar distribution, suggesting parallel activation by LPS [44,81]. In this study, CB1/2 were immunolocalized to similar endometrial regions across all groups, mirroring TLR-4 and COX-2 localization and intensity, implying the cooperative regulation of the inflammatory response by ECBs, TLR-4, and COX-2 [90].
Interestingly, in the OP group, CB1 and CB2 were more intensely expressed in the deep endometrial cells, inflammatory cells, and myometrium. This likely reflects OP-specific gene expression changes. Voorwald et al. (2015) reported the overexpression of fatty acid-binding protein 3 (FABP3) in OP, a protein abundant in myocytes and vascular endothelium that is upregulated by LPS and can exacerbate inflammation [29,91]. Fatty acid-binding proteins also function as chaperones for ECBs, with ECBs exhibiting pronounced affinity for FABP3 [92].
5. Conclusions
In conclusion, this study provides the first direct evidence of ECB presence in both pyometra-affected and healthy canine uterine tissue, unequivocally supporting our initial hypothesis. The differential expression of CB1 and CB2 appears to result from the integrated effects of P4, decidualization, stromal aging, LPS, TLR signaling, and COX activity, reflecting the intricate crosstalk between reproductive physiology, uterine pathology, and the eCS. These findings not only advance our understanding of ECB regulation in the canine uterus, but also emphasize the need for further comprehensive investigations of additional components, including FAAH, NAPE-PLD, MAGL, and DAGL, at both tissue-specific and molecular levels to unravel their precise roles in uterine homeostasis and pathophysiology.
Author Contributions
S.A.A. and A.G.A. designed and planned the project, analyzed the data; S.A.A., A.GA., and V.F. drafted and wrote the initial manuscript. F.B. performed the experiments and ovariohysterectomy operations, S.A.A. and M.F. provided data control. A.G.A and A.G. performed IHC analyses, and I.O. provided analysis control. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by Ondokuz Mayıs University (Türkiye) under the project number PYO.VET.1904.23.027.
Institutional Review Board Statement
Both the clinical part of the animal study and the tissue examination protocols were approved by Ondokuz Mayıs University Animal Experiments Local Ethics Committee (HADYEK: 2022/47).
Data Availability Statement
The datasets used during the current study are available from the corresponding author on reasonable request.
Conflicts of Interest
The authors declare no conflicts of interest. All coauthors have seen and agree with the contents of the manuscript.
References
- Maccarrone, M. Endocannabinoids: friends and foes of reproduction. Prog. Lipid Res. 2009, 48, 344–354. [Google Scholar] [CrossRef] [PubMed]
- Howlett, A.C.; Barth, F.; Bonner, T.I.; Cabral, G.; Casellas, P.; Devane, W.A.; Pertwee, R.G. International Union of Pharmacology. XXVII. Classification of cannabinoid receptors. Pharmacol. Rev. 2002, 54, 161–202. [Google Scholar] [CrossRef] [PubMed]
- El-Talatini, M.R.; Taylor, A.H.; Elson, J.C.; Brown, L.; Davidson, A.C.; Konje, J.C. Localisation and function of the endocannabinoid system in the human ovary. PLoS One 2009, 4, e4579. [Google Scholar] [CrossRef] [PubMed]
- Reggio, P.H. Endocannabinoid binding to the cannabinoid receptors: what is known and what remains unknown. Curr. Med. Chem. 2010, 17, 1468–1486. [Google Scholar] [CrossRef]
- Maccarrone, M.; Di Marzo, V.; Gertsch, J.; Grether, U.; Howlett, A.C.; Hua, T.; van der Stelt, M. Goods and bads of the endocannabinoid system as a therapeutic target: Lessons learned after 30 years. Pharmacol. Rev. 2023, 75, 885–958. [Google Scholar] [CrossRef]
- Correa, F.; Wolfson, M.L.; Valchi, P.; Aisemberg, J.; Franchi, A.M. Endocannabinoid system and pregnancy. Reproduction 2016, 152, R191–R200. [Google Scholar] [CrossRef]
- Peiris, H.N.; Vaswani, K.; Holland, O.; Koh, Y.Q.; Almughlliq, F.B.; Reed, S.; Mitchell, M.D. Altered productions of prostaglandins and prostamides by human amnion in response to infectious and inflammatory stimuli identified by multiplex mass spectrometry. Prostaglandins Leukot. Essent. Fatty Acids 2020, 154, 102059. [Google Scholar] [CrossRef]
- Gammon, C.M.; Freeman, G.M.; Xie, W.; Petersen, S.L.; Wetsel, W.C. Regulation of gonadotropin-releasing hormone secretion by cannabinoids. Endocrinology 2005, 146, 4491–4499. [Google Scholar] [CrossRef]
- Horne, A.W.; Phillips, J.A. III; Kane, N.; Lourenco, P.C.; McDonald, S.E.; Williams, A.R.; Critchley, H.O. CB1 expression is attenuated in Fallopian tube and decidua of women with ectopic pregnancy. PLoS One 2008, 3, e3969. [Google Scholar] [CrossRef]
- Scorticati, C.; Fernández-Solari, J.; De Laurentiis, A.; Mohn, C.; Prestifilippo, J.P.; Lasaga, M.; Rettori, V. The inhibitory effect of anandamide on luteinizing hormone-releasing hormone secretion is reversed by estrogen. Proc. Natl. Acad. Sci. U.S.A. 2004, 101, 11891–11896. [Google Scholar] [CrossRef]
- El-Talatini, M.R.; Taylor, A.H.; Konje, J.C. The relationship between plasma levels of the endocannabinoid, anandamide, sex steroids, and gonadotrophins during the menstrual cycle. Fertil. Steril. 2010, 93, 1989–1996. [Google Scholar] [CrossRef]
- Maccarrone, M.; Bari, M.; Battista, N.; Finazzi-Agrò, A. Estrogen stimulates arachidonoylethanolamide release from human endothelial cells and platelet activation. The J. Biol. Chem. 2002, 277, 23954–23960. [Google Scholar] [CrossRef]
- Wang, H.; Xie, H.; Guo, Y.; Zhang, H.; Takahashi, T.; Kingsley, P.J.; Marnett, L.J.; Das, S.K.; Cravatt, B.F.; Dey, S.K. Fatty acid amide hydrolase deficiency limits early pregnancy events. J. Clin. Invest. 2006, 116, 2122–2131. [Google Scholar] [CrossRef]
- Sanchez, A.M.; Cioffi, R.; Viganò, P.; Candiani, M.; Verde, R.; Piscitelli, F.; Panina-Bordignon, P. Elevated systemic levels of endocannabinoids and related mediators across the menstrual cycle in women with endometriosis. Reprod. Sci. 2016, 23, 1071–1079. [Google Scholar] [CrossRef]
- Lingegowda, H.; Zutautas, K.B.; Wei, Y.; Yolmo, P.; Sisnett, D.J.; McCallion, A.; Tayade, C. Endocannabinoids and their receptors modulate endometriosis pathogenesis and immune response. eLife 2024, 13, RP96523. [Google Scholar] [CrossRef]
- Adhikary, S.; Kocieda, V.P.; Yen, J.H.; Tuma, R.F.; Ganea, D. Signaling through cannabinoid receptor 2 suppresses murine dendritic cell migration by inhibiting matrix metalloproteinase 9 expression. Blood 2012, 120, 3741–3749. [Google Scholar] [CrossRef]
- Duncan, M.; Galic, M.A.; Wang, A.; Chambers, A.P.; McCafferty, D.M.; McKay, D.M.; Pittman, Q.J. Cannabinoid 1 receptors are critical for the innate immune response to TLR4 stimulation. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305, R224–R231. [Google Scholar] [CrossRef]
- Šahinović, I.; Mandić, S.; Mihić, D.; Duvnjak, M.; Loinjak, D.; Sabadi, D.; Šerić, V. Endocannabinoids, anandamide and 2-arachidonoylglycerol, as prognostic markers of sepsis outcome and complications. Cannabis Cannabinoid Res. 2023, 8, 802–811. [Google Scholar] [CrossRef] [PubMed]
- Barker, H.; Ferraro, M.J. Exploring the versatile roles of the endocannabinoid system and phytocannabinoids in modulating bacterial infections. Infect. Immun. 2024, 92, e00020-24. [Google Scholar] [CrossRef] [PubMed]
- McCoy, K.L. Interaction between cannabinoid system and toll-like receptors controls inflammation. Mediators of Inflamm. 2016, 2016, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Silva, E.; Leitão, S.; Henriques, S.; Kowalewski, M.P.; Hoffmann, B.; Ferreira-Dias, G.; Mateus, L. Gene transcription of TLR2, TLR4, LPS ligands and prostaglandin synthesis enzymes are up-regulated in canine uteri with cystic endometrial hyperplasia–pyometra complex. J. Reprod. Immunol. 2010, 84, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Chotimanukul, S.; Sirivaidyapong, S. Differential expression of Toll-like receptor 4 (TLR4) in healthy and infected canine endometrium. Theriogenology 2011, 76, 1152–1161. [Google Scholar] [CrossRef] [PubMed]
- Helliwell, R.J.; Adams, L.F.; Mitchell, M.D. Prostaglandin synthases: recent developments and a novel hypothesis. Prostaglandins Leukot. Essent. Fatty Acids 2004, 70, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Bariani, M.V.; Domínguez Rubio, A.P.; Cella, M.; Burdet, J.; Franchi, A.M.; Aisemberg, J. Role for the endocannabinoid system in the mechanisms involved in the LPS-induced preterm labor. Reproduction 2015, 150, 293–302. [Google Scholar] [CrossRef]
- Correa, F.; Hernangómez, M.; Mestre, L.; Loría, F.; Spagnolo, A.; Docagne, F.; Guaza, C. Anandamide enhances IL-10 production in activated microglia by targeting CB2 receptors: Roles of ERK1/2, JNK, and NF-κB. Glia 2010, 58, 135–147. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, C. Endocannabinoid 2-arachidonoylglycerol protects neurons by limiting COX-2 elevation. J. Biol. Chem. 2008, 283, 22601–22611. [Google Scholar] [CrossRef]
- Hagman, R.; Rönnberg, E.; Pejler, G. Canine uterine bacterial infection induces upregulation of proteolysis-related genes and downregulation of homeobox and zinc finger factors. PLoS One 2009, 4, e8039. [Google Scholar] [CrossRef]
- Wallace, G.B.; Casal, M.L. A review of pyometra in small animal medicine: incidence, pathophysiology, clinical diagnosis, and medical management. Clin. Theriogenol. 2018, 10, 435–452. [Google Scholar] [CrossRef]
- Voorwald, F.A.; Marchi, F.A.; Villacis, R.A.R.; Alves, C.E.F.; Toniollo, G.H.; Amorim, R.L.; Rogatto, S.R. Molecular expression profile reveals potential biomarkers and therapeutic targets in canine endometrial lesions. PLoS One 2015, 10, e0133894. [Google Scholar] [CrossRef]
- Foster, R.F. Bases da Patologia em Veterinária. In Sistema Reprodutivo da Fêmea, 4th ed.; McGavin, M.D., Zachary, J.F., Eds.; Elsevier: Rio de Janeiro, Brazil, 2009; pp. 1263–1315. [Google Scholar]
- De Bosschere, H.; Ducatelle, R.; Vermeirsch, H.; Van Den Broeck, W.; Coryn, M. Cystic endometrial hyperplasia-pyometra complex in the bitch: Should the two entities be disconnected? Theriogenology 2001, 55, 1509–1519. [Google Scholar] [CrossRef]
- Feldman, E.C.; Nelson, R.W. Ovarian cycle and vaginal cytology. In Canine and Feline Endocrinology and Reproduction; Feldman, E.C., Nelson, R.W., Eds.; Saunders: Philadelphia, PA, USA, 2004; pp. 752–774. [Google Scholar]
- Nomura, K.; Funahashi, H. Histological characteristics of canine deciduoma induced by intrauterine inoculation of E. coli suspension. J. Vet. Med. Sci. 1999, 61, 433–438. [Google Scholar] [CrossRef]
- Schlafer, D.H. Diseases of the canine uterus. Reprod. Domest. Anim. 2012, 47, 318–322. [Google Scholar] [CrossRef] [PubMed]
- Concannon, P.W. Endocrinologic control of normal canine ovarian function. Reprod. Domest. Anim. 2009, 44, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Chu, P.Y.; Salamonsen, L.A.; Lee, C.S.; Wright, P.J. Matrix metalloproteinases (MMPs) in the endometrium of bitches. Reproduction 2002, 123, 467–477. [Google Scholar] [CrossRef]
- Campora, L.; Miragliotta, V.; Ricci, E.; Cristino, L.; Di Marzo, V.; Albanese, F.; Abramo, F. Cannabinoid receptor type 1 and 2 expression in the skin of healthy dogs and dogs with atopic dermatitis. Am. J. Vet. Res. 2012, 73, 988–995. [Google Scholar] [CrossRef]
- Galiazzo, G.; Giancola, F.; Stanzani, A.; Fracassi, F.; Bernardini, C.; Forni, M.; Chiocchetti, R. Localization of cannabinoid receptors CB1, CB2, GPR55, and PPARα in the canine gastrointestinal tract. Histochem. Cell Biol. 2018, 150, 187–205. [Google Scholar] [CrossRef]
- Mercati, F.; Dall’Aglio, C.; Pascucci, L.; Boiti, C.; Ceccarelli, P. Identification of cannabinoid type 1 receptor in dog hair follicles. Acta Histochem. 2012, 114, 68–71. [Google Scholar] [CrossRef]
- Dall’Aglio, C.; Mercati, F.; Pascucci, L.; Boiti, C.; Pedini, V.; Ceccarelli, P. Immunohistochemical localization of CB1 receptor in canine salivary glands. Vet. Res. Commun. 2010, 34, 9–12. [Google Scholar] [CrossRef]
- Freundt-Revilla, J.; Kegler, K.; Baumgärtner, W.; Tipold, A. Spatial distribution of cannabinoid receptor type 1 (CB1) in normal canine central and peripheral nervous system. PLoS One 2017, 12, e0181064. [Google Scholar] [CrossRef]
- Kowalewski, M.P.; Beceriklisoy, H.B.; Pfarrer, C.; Aslan, S.; Kindahl, H.; Kücükaslan, I.; Hoffmann, B. Canine placenta: a source of prepartal prostaglandins during normal and antiprogestin-induced parturition. Reproduction 2010, 139, 655–664. [Google Scholar] [CrossRef]
- Stavreus-Evers, A.; Koraen, L.; Scott, J.E.; Zhang, P.; Westlund, P. Distribution of cyclooxygenase-1, cyclooxygenase-2, and cytosolic phospholipase A2 in the luteal phase human endometrium and ovary. Fertil. Steril. 2005, 83, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Veiga, G.A.L.; Miziara, R.H.; Angrimani, D.S.R.; Papa, P.C.; Cogliati, B.; Vannucchi, C.I. Cystic endometrial hyperplasia–pyometra syndrome in bitches: identification of hemodynamic, inflammatory, and cell proliferation changes. Biol. Reprod. 2017, 96, 58–69. [Google Scholar] [PubMed]
- Osteen, K.G.; Bruner-Tran, K.L.; Eisenberg, E. Reduced progesterone action during endometrial maturation: a potential risk factor for the development of endometriosis. Fertil. Steril. 2005, 83, 529–537. [Google Scholar] [CrossRef] [PubMed]
- Penell, J.C.; Morgan, D.M.; Watson, P.; Carmichael, S.; Adams, V.J. Body weight at 10 years of age and change in body composition between 8 and 10 years of age were related to survival in a longitudinal study of 39 Labrador retriever dogs. Acta Vet. Scand. 2019, 61, 1–16. [Google Scholar] [CrossRef]
- Maia, J.; Fonseca, B.M.; Teixeira, N.; Correia-da-Silva, G. The fundamental role of the endocannabinoid system in endometrium and placenta: implications in pathophysiological aspects of uterine and pregnancy disorders. Hum. Reprod. Update 2020, 26, 586–602. [Google Scholar] [CrossRef]
- Chinnu, V.P. Efficacy of medical management and surgical trans-cervical catheterisation for canine cystic endometrial hyperplasia. Ph.D. Thesis, College of Veterinary and Animal Science, Mannuthy, 2016. Available online: http://krishikosh.egranth.ac.in/handle/1/5810110584 (accessed on 28 August 2025).
- Taylor, A.H.; Abbas, M.S.; Habiba, M.A.; Konje, J.C. Histomorphometric evaluation of cannabinoid receptor and anandamide modulating enzyme expression in the human endometrium through the menstrual cycle. Histochem. Cell Biol. 2010, 133, 557–565. [Google Scholar] [CrossRef]
- Tanaka, K.; Mayne, L.; Khalil, A.; Baartz, D.; Eriksson, L.; Mortlock, S.A.; Amoako, A.A. The role of the endocannabinoid system in aetiopathogenesis of endometriosis: A potential therapeutic target. Eur. J. Obstet. Gynecol. Reprod. Biol. 2020, 244, 87–94. [Google Scholar] [CrossRef]
- Resuehr, D.; Glore, D.R.; Taylor, H.S.; Bruner-Tran, K.L.; Osteen, K.G. Progesterone-dependent regulation of endometrial cannabinoid receptor type 1 (CB1-R) expression is disrupted in women with endometriosis and in isolated stromal cells exposed to 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD). Fertil. Steril. 2012, 98, 948–956. [Google Scholar] [CrossRef]
- Bayazeid, O.; Eylem, C.C.; Reçber, T.; Yalçın, F.N.; Kır, S.; Nemutlu, E. An LC-ESI-MS/MS method for the simultaneous determination of pronuciferine and roemerine in some Papaver species. J. Chromatogr. B 2018, 1096, 223–227. [Google Scholar] [CrossRef]
- Lanz, C.; Mattsson, J.; Stickel, F.; Dufour, J.F.; Brenneisen, R. Determination of the endocannabinoids anandamide and 2-arachidonoyl glycerol with gas chromatography-mass spectrometry: analytical and preanalytical challenges and pitfalls. Med. Cannabis Cannabinoids 2018, 1, 9–18. [Google Scholar]
- Fonseca, B.M.; Correia-da-Silva, G.; Taylor, A.H.; Lam, P.M.W.; Marczylo, T.H.; Bell, S.C.; Teixeira, N.A. The endocannabinoid 2-arachidonoylglycerol (2-AG) and metabolizing enzymes during rat fetoplacental development: a role in uterine remodelling. Int. J. Biochem. Cell Biol. 2010, 42, 1884–1892. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, H.C.; Dhakal, P.; Behura, S.K.; Schust, D.J.; Spencer, T.E. Self-renewing endometrial epithelial organoids of the human uterus. Proc. Natl. Acad. Sci. 2019, 116, 23132–23142. [Google Scholar] [CrossRef] [PubMed]
- Tavares Pereira, M.; Kazemian, A.; Rehrauer, H.; Kowalewski, M.P. Transcriptomic profiling of canine decidualization and effects of antigestagens on decidualized dog uterine stromal cells. Sci. Rep. 2022, 12, 21890–10.1038. [Google Scholar] [CrossRef] [PubMed]
- Nomura, K. Induction of canine deciduoma in some reproductive stages with the different conditions of corpora lutea. J. Vet. Med. Sci. 1997, 59, 185–190. [Google Scholar] [CrossRef]
- Fonseca, B.M.; Correia-da-Silva, G.; Almada, M.; Costa, M.A.; Teixeira, N.A. The endocannabinoid system in the postimplantation period: a role during decidualization and placentation. Int. J. Endocrinol. 2013, 2013, 510540. [Google Scholar] [CrossRef]
- Fonseca, B.M.; Correia-da-Silva, G.; Taylor, A.H.; Konje, J.C.; Bell, S.C.; Teixeira, N.A. Spatio-temporal expression patterns of anandamide-binding receptors in rat implantation sites: evidence for a role of the endocannabinoid system during the period of placental development. Reprod. Biol. Endocrin. 2009, 7, 1–13. [Google Scholar] [CrossRef]
- Taylor, A.H.; Finney, M.; Lam, P.M.; Konje, J.C. Modulation of the endocannabinoid system in viable and non-viable first trimester pregnancies by pregnancy-related hormones. Reprod. Biol. Endocrinol. 2011, 9, 1–18. [Google Scholar] [CrossRef]
- Albertini, D.F. Commentary: Paying a price for plasticity—what the endometrium has to do with it! J. Assist. Reprod. Genet. 2019, 36, 809–810. [Google Scholar] [CrossRef]
- Gentilini, D.; Besana, A.; Vigano, P.; Dalino, P.; Vignali, M.; Melandri, M.; Di Blasio, A.M. Endocannabinoid system regulates migration of endometrial stromal cells via cannabinoid receptor 1 through the activation of PI3K and ERK1/2 pathways. Fertil. Steril. 2010, 93, 2588–2593. [Google Scholar] [CrossRef]
- Stojiljković, A.; Gaschen, V.; Forterre, F.; Rytz, U.; Stoffel, M.H.; Bluteau, J. Novel immortalization approach defers senescence of cultured canine adipose-derived mesenchymal stromal cells. GeroScience 2022, 44, 1–23. [Google Scholar] [CrossRef]
- De Cesaris, V.; Grolli, S.; Bresciani, C.; Conti, V.; Basini, G.; Parmigiani, E.; Bigliardi, E. Isolation, proliferation and characterization of endometrial canine stem cells. Reprod. Domest. Anim. 2017, 52, 235–242. [Google Scholar] [CrossRef]
- Lucas, E.S.; Dyer, N.P.; Murakami, K.; Hou Lee, Y.; Chan, Y.W.; Grimaldi, G.; Brosens, J.J. Loss of endometrial plasticity in recurrent pregnancy loss. Stem Cells 2016, 34, 346–356. [Google Scholar] [CrossRef]
- Verstegen, J.; Dhaliwal, G.; Verstegen-Onclin, K. Mucometra, cystic endometrial hyperplasia, and pyometra in the bitch: advances in treatment and assessment of future reproductive success. Theriogenology 2008, 70, 364–374. [Google Scholar] [CrossRef]
- Rossi, G.; Di Nisio, V.; Chiominto, A.; Cecconi, S.; Maccarrone, M. Endocannabinoid system components of the female mouse reproductive tract are modulated during reproductive aging. Int. J. Mol. Sci. 2023, 24, 7542. [Google Scholar] [CrossRef]
- Salazar, A.I.; Vercelli, C.; Schiariti, V.; Davio, C.; Correa, F.; Franchi, A.M. Heparin exerts anti-apoptotic effects on uterine explants by targeting the endocannabinoid system. Apoptosis 2016, 21, 965–976. [Google Scholar] [CrossRef] [PubMed]
- Romano, B.; Borrelli, F.; Fasolino, I.; Capasso, R.; Piscitelli, F.; Cascio, M.G.; Izzo, A.A. The cannabinoid TRPA1 agonist cannabichromene inhibits nitric oxide production in macrophages and ameliorates murine colitis. Br J. Pharmacol. 2013, 169, 213–229. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.P.; Yuan, Q.H.; Zhang, B.; Yang, L.; He, Q.W.; Chen, K.; Zhan, J. Cannabinoid receptor 2 activation alleviates septic lung injury by promoting autophagy via inhibition of inflammatory mediator release. Cell. Signal. 2020, 69, 109556. [Google Scholar] [CrossRef] [PubMed]
- Tanikawa, T.; Kitamura, M.; Hayashi, Y.; Yokogawa, T.; Inoue, Y. Curcumae Longae Rhizoma and Saussureae Radix inhibit nitric oxide production and cannabinoid receptor 2 down-regulation. In Vivo 2022, 36, 227–232. [Google Scholar] [CrossRef]
- Xiang, W.; Shi, R.; Kang, X.; Zhang, X.; Chen, P.; Zhang, L.; Miao, H. Monoacylglycerol lipase regulates cannabinoid receptor 2-dependent macrophage activation and cancer progression. Nat. Commun. 2018, 9, 2574. [Google Scholar] [CrossRef]
- Takeda, K.; Akira, S. Toll-like receptors in innate immunity. Int. Immunol. 2005, 17, 1–14. [Google Scholar] [CrossRef]
- Allhorn, S.; Böing, C.; Koch, A.A.; Kimmig, R.; Gashaw, I. TLR3 and TLR4 expression in healthy and diseased human endometrium. Reprod. Biol. Endocrinol. 2008, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Chu, P.Y.; Lee, C.S.; Wright, P.J. Degeneration and apoptosis of endometrial cells in the bitch. Theriogenology 2006, 66, 1545–1549. [Google Scholar] [CrossRef] [PubMed]
- Hirata, T.; Osuga, Y.; Hamasaki, K.; Hirota, Y.; Nose, E.; Morimoto, C.; Taketani, Y. Expression of toll-like receptors 2, 3, 4, and 9 genes in the human endometrium during the menstrual cycle. J. Reprod. Immunol. 2007, 74, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Freundt-Revilla, J.; Heinrich, F.; Zoerner, A.; Gesell, F.; Beyerbach, M.; Shamir, M.; Tipold, A. The endocannabinoid system in canine steroid-responsive meningitis-arteritis and intraspinal spirocercosis. PLoS One 2018, 13, e0187197. [Google Scholar] [CrossRef]
- Simard, M.; Rakotoarivelo, V.; Di Marzo, V.; Flamand, N. Expression and functions of the CB2 receptor in human leukocytes. Front. Pharmacol. 2022, 13, 826400. [Google Scholar] [CrossRef]
- Pires, M.A.; Payan-Carreira, R. Resident macrophages and lymphocytes in the canine endometrium. Reprod. Domest. Anim. 2015, 50, 740–749. [Google Scholar] [CrossRef]
- Minghetti, L. Cyclooxygenase-2 (COX-2) in inflammatory and degenerative brain diseases. J. Neuropathol. Exp. Neurol. 2004, 63, 901–910. [Google Scholar] [CrossRef]
- Beg, A.A. Endogenous ligands of Toll-like receptors: Implications for regulating inflammatory and immune responses. Trends Immunol. 2002, 23, 509–512. [Google Scholar] [CrossRef]
- Silva, E.; Leitao, S.; Ferreira-Dias, G.; Lopes da Costa, L.; Mateus, L. Prostaglandin synthesis genes are differentially transcripted in normal and pyometra endometria of bitches. Reprod. Domest. Anim. 2009, 44, 200–203. [Google Scholar] [CrossRef]
- Kuleš, J.; Horvatić, A.; Guillemin, N.; Ferreira, R.F.; Mischke, R.; Mrljak, V.; Eckersall, P.D. The plasma proteome and the acute phase protein response in canine pyometra. J. Proteomics 2020, 223, 103817. [Google Scholar] [CrossRef]
- Hagman, R.; Kindahl, H.; Fransson, B.A.; Bergström, A.; Holst, B.S.; Lagerstedt, A.S. Differentiation between pyometra and cystic endometrial hyperplasia/mucometra in bitches by prostaglandin F2α metabolite analysis. Theriogenology 2006, 66, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Watkins, B.A. Cannabinoid receptor antagonists and fatty acids alter endocannabinoid system gene expression and COX activity. J. Nutr. Biochem. 2014, 25, 815–823. [Google Scholar] [CrossRef] [PubMed]
- Dennedy, M.C.; Friel, A.M.; Houlihan, D.D.; Broderick, V.M.; Smith, T.; Morrison, J.J. Cannabinoids and the human uterus during pregnancy. Am. J. Obstet. Gynecol. 2004, 190, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Pagano, E.; Orlando, P.; Finizio, S.; Rossi, A.; Buono, L.; Iannotti, F.A.; Borrelli, F. Role of the endocannabinoid system in the control of mouse myometrium contractility during the menstrual cycle. Biochem. Pharmacol. 2017, 124, 83–93. [Google Scholar] [CrossRef]
- Dong, Y.L.; Gangula, P.R.; Fang, L.; Yallampalli, C. Differential expression of cyclooxygenase-1 and -2 proteins in rat uterus and cervix during the estrous cycle, pregnancy, labor and in myometrial cells. Prostaglandins 1996, 52, 13–34. [Google Scholar] [CrossRef]
- Rocca, B.; FitzGerald, G.A. Cyclooxygenases and prostaglandins: shaping up the immune response. Int. Immunopharmacol. 2002, 2, 603–630. [Google Scholar] [CrossRef]
- Vercelli, C.A.; Aisemberg, J.; Billi, S.; Cervini, M.; Ribeiro, M.L.; Farina, M.; Franchi, A.M. Anandamide regulates lipopolysaccharide-induced nitric oxide synthesis and tissue damage in the murine uterus. Reprod. Biomed. Online 2009, 18, 824–831. [Google Scholar] [CrossRef]
- Nguyen, H.C.; Bu, S.; Nikfarjam, S.; Rasheed, B.; Michels, D.C.; Singh, A.; Singh, K.K. Loss of fatty acid binding protein 3 ameliorates lipopolysaccharide-induced inflammation and endothelial dysfunction. J. Biol. Chem. 2023, 299, 102921. [Google Scholar] [CrossRef]
- Elmes, M.W.; Kaczocha, M.; Berger, W.T.; Leung, K.; Ralph, B.P.; Wang, L.; Deutsch, D.G. Fatty acid-binding proteins (FABPs) are intracellular carriers for Δ9-tetrahydrocannabinol (THC) and cannabidiol (CBD). J. Biol. Chem. 2015, 290, 8711–8721. [Google Scholar] [CrossRef]
Figure 1.
Serum P4 concentrations in pyometric and healthy dog uterus. A non-parametric Kruskal Wallis test was applied followed by the Mann Whitney U tests with Bonferroni correction. Results are shown as mean ± SEM. p<0.05 were considered significant. CP: Close cervix pyometra, OP: Open cervix pyometra, DE: Diestrus, AE: Anestrus.
Figure 1.
Serum P4 concentrations in pyometric and healthy dog uterus. A non-parametric Kruskal Wallis test was applied followed by the Mann Whitney U tests with Bonferroni correction. Results are shown as mean ± SEM. p<0.05 were considered significant. CP: Close cervix pyometra, OP: Open cervix pyometra, DE: Diestrus, AE: Anestrus.
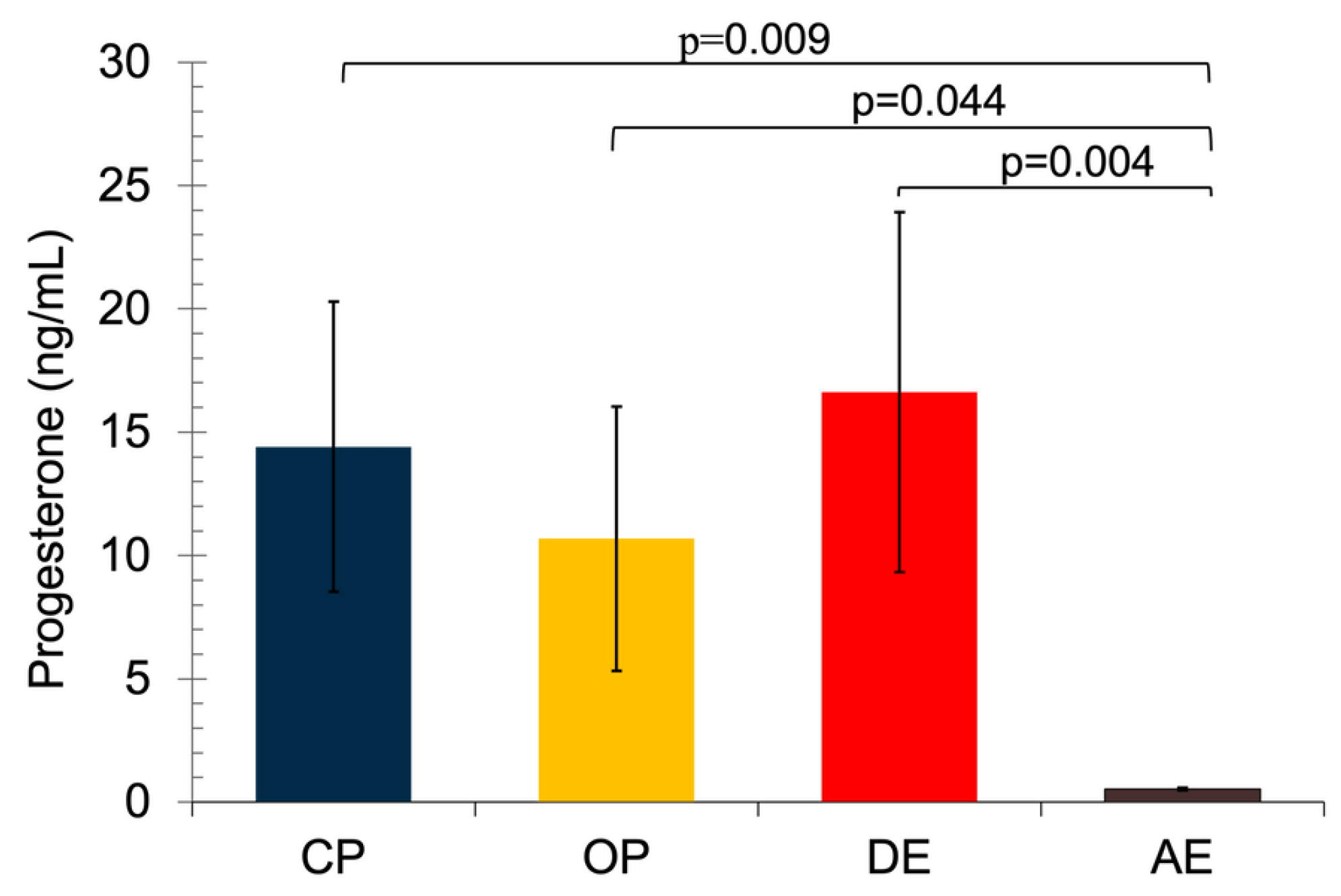
Figure 2.
Serum ECB concentrations in pyometric and healthy dog uterus (a) Serum AEA concentrations. A non-parametric Kruskal Wallis test was applied followed by the Mann Whitney U tests with Bonferroni correction. (b) 2-AG concentrations. A non-parametric Kruskal Wallis test was applied. Results are shown as mean ± SEM. p< 0.05 were considered significant. CP: Close cervix pyometra, OP: Open cervix pyometra, DE: Diestrus, AE: Anestrus.
Figure 2.
Serum ECB concentrations in pyometric and healthy dog uterus (a) Serum AEA concentrations. A non-parametric Kruskal Wallis test was applied followed by the Mann Whitney U tests with Bonferroni correction. (b) 2-AG concentrations. A non-parametric Kruskal Wallis test was applied. Results are shown as mean ± SEM. p< 0.05 were considered significant. CP: Close cervix pyometra, OP: Open cervix pyometra, DE: Diestrus, AE: Anestrus.
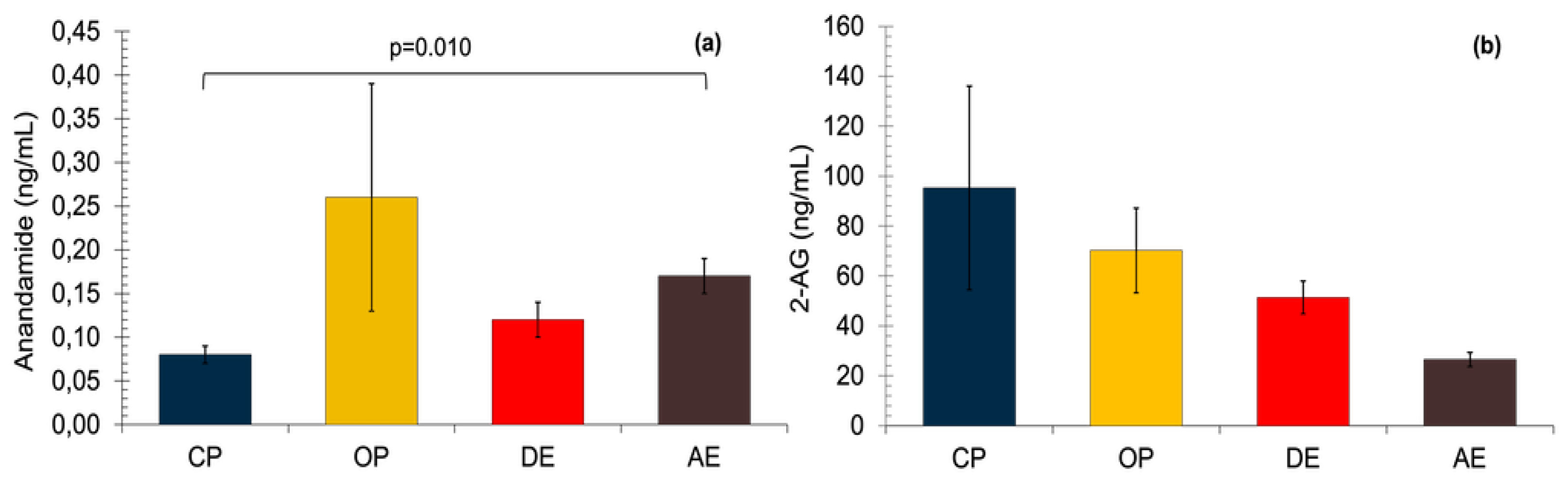
Figure 3.
The mRNA expression of CBs in pyometric and healthy dog uterus. (a) Expression of CB1 gene. A parametric one-way ANOVA was applied followed by the Tukey HSD post-hoc test. (b) Expression of CB2 gene. A parametric one-way ANOVA was applied. Results are shown as mean ± SEM. p< 0.05 were considered significant. CP: Close cervix pyometra, OP: Open cervix pyometra, DE: Diestrus, AE: Anestrus.
Figure 3.
The mRNA expression of CBs in pyometric and healthy dog uterus. (a) Expression of CB1 gene. A parametric one-way ANOVA was applied followed by the Tukey HSD post-hoc test. (b) Expression of CB2 gene. A parametric one-way ANOVA was applied. Results are shown as mean ± SEM. p< 0.05 were considered significant. CP: Close cervix pyometra, OP: Open cervix pyometra, DE: Diestrus, AE: Anestrus.
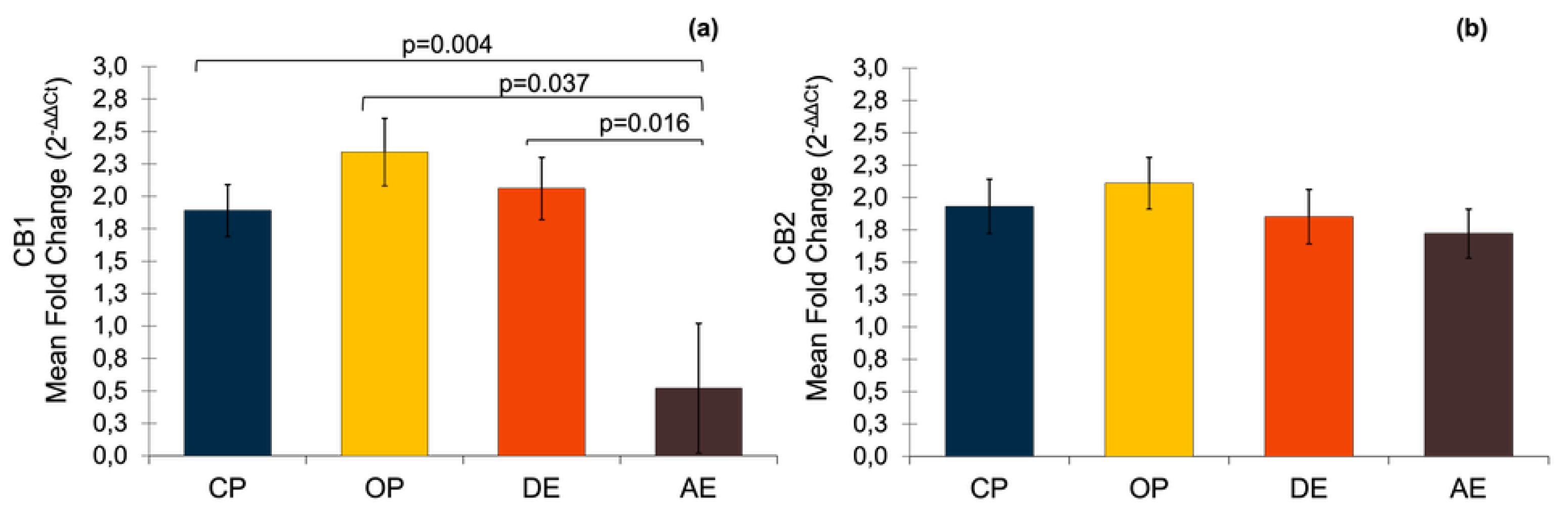
Figure 4.
Heat map of CBs immunoreactivity at the cellular level in uterine tissue. CP: Close cervix pyometra, OP: Open cervix pyometra, DE: Diestrus, AE: Anestrus.
Figure 4.
Heat map of CBs immunoreactivity at the cellular level in uterine tissue. CP: Close cervix pyometra, OP: Open cervix pyometra, DE: Diestrus, AE: Anestrus.
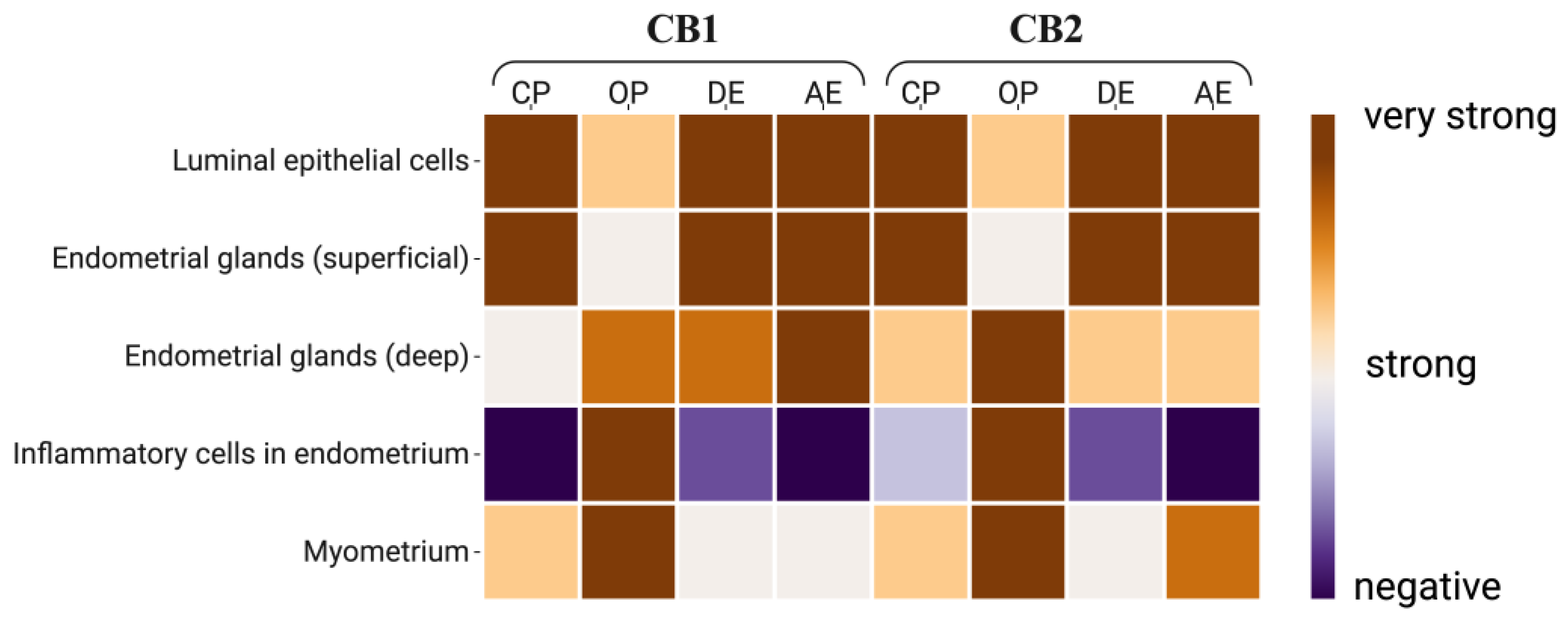
Figure 5.
Immunolocalization of CBs in uterine tissue (bar: 50μm-X20). Immunoreactivity of CB1 in different areas of the uterus by groups; CP (A), OP (D), DE (G), AE (J). Immunoreactivity of CB2 in different areas of the uterus by groups; CP (B), OP (E), DE (H), AE (H). No immunoreactivity in isotype controls (M, N). (white arrows: luminal epithelial cells, black arrows: endometrial gland cells, *: immuncells).
Figure 5.
Immunolocalization of CBs in uterine tissue (bar: 50μm-X20). Immunoreactivity of CB1 in different areas of the uterus by groups; CP (A), OP (D), DE (G), AE (J). Immunoreactivity of CB2 in different areas of the uterus by groups; CP (B), OP (E), DE (H), AE (H). No immunoreactivity in isotype controls (M, N). (white arrows: luminal epithelial cells, black arrows: endometrial gland cells, *: immuncells).
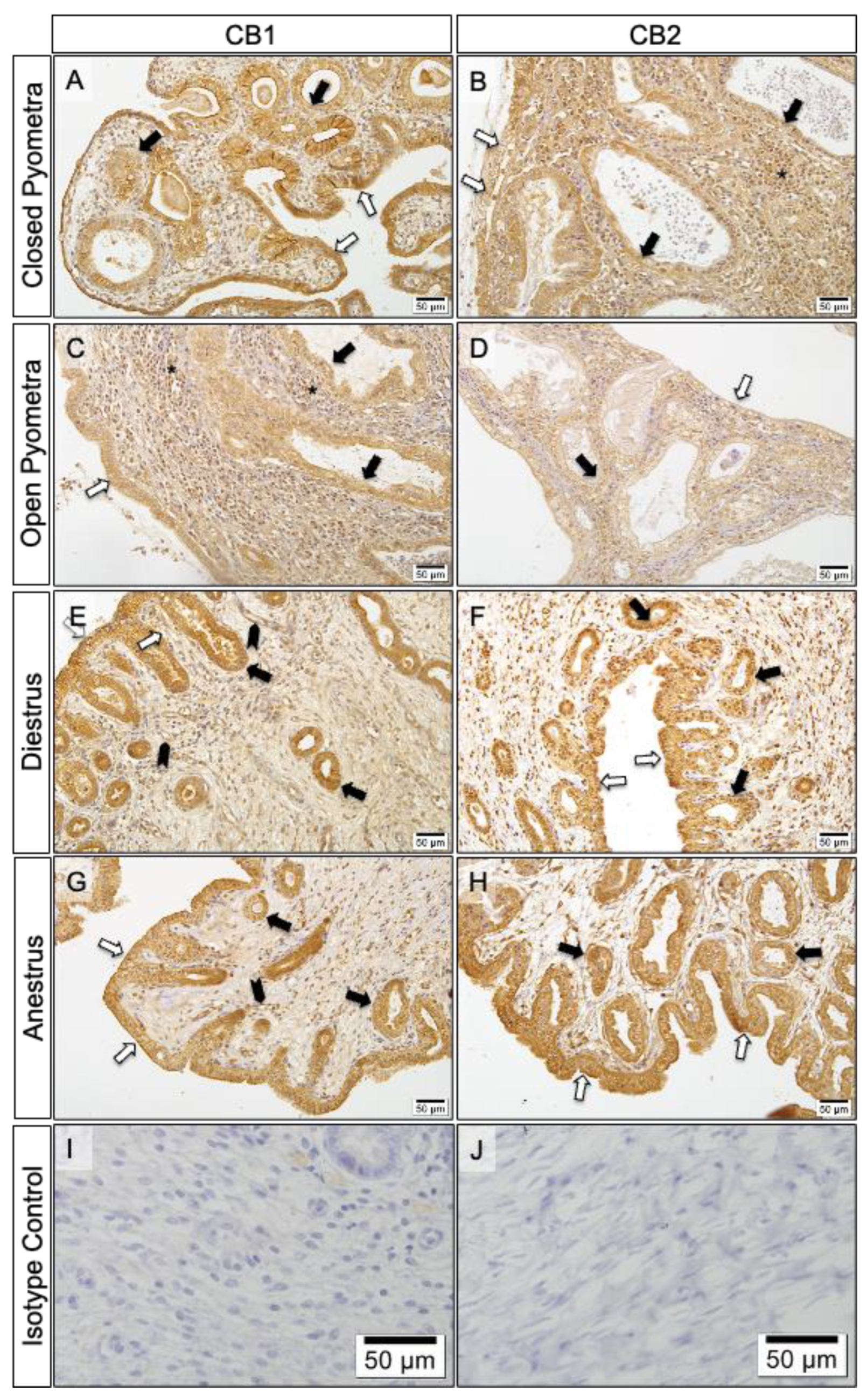

Table 1.
List of primers used for the RT-qPCR.
| Gene | Primer sequence | Product length (bp) | Accession numbers |
|---|---|---|---|
| CNR1 | F: TCCTGGGGAGCGTCATATTT | 99 | AY011618.1 |
| R: TGACCCCACCCAGTTTGAAA | |||
| CNR2 | F: TCCTGGCCAGTGTGATCTTT | 119 | NM_001284480.1 |
| R: AGAGGCTGTGAAGGTCATGG | |||
| KDM4A | F: TCACAGAGAAGGAAGTTAAG | 82 | XM_005629107 |
| R: TCACAGAGAAGGAAGTTAAG | |||
| PTK2 | F: ACCTGGCTGACTTCAATC | 85 | XM_005627993 |
| R: ATCTTCAACTGTAGCATTCCT | |||
| EIF4H | F: TAAGGTCTCAGCAATTAC | 101 | XM_014114129 |
| R: TAAGGTCTCAGCAATTAC |
Table 2.
Conditions employed for all antibodies during immunohistochemical procedures.
| Antibody | Reference (Company) |
Biological source | Antibody Dilution |
Antibody Incubtion |
Epitope Revelation (DAB) |
|---|---|---|---|---|---|
| CNR1/CB1 | bs-1683R (Bioss, Woburn, USA) | Rabbit (Polyclonal) |
1:200 | Overnight (17 h), 4°C |
Thermo Fisher Scientific Lab Vision Corporation |
| CNR2/CB2 | LS-A34-50 (Life Span, Lynnwood, USA) | ||||
|
Isotype control |
IgG (Vector Laboratories, Burlingame, USA |
Rabbit | Same protein concentration as primary antibody | - | - |
Table 3.
Average age and body weight of the groups. The data given as Mean ± SEM.
| Groups | n | Ages (year) |
Body weight (kg) |
|---|---|---|---|
| CP | 7 | 8.1 ± 1.1a | 18.4 ± 1.2a |
| OP | 7 | 10.0 ± 1.0a | 15.5 ± 0.1a |
| DE | 7 | 1.8 ± 0.1b | 10.1 ± 0.9b |
| AE | 7 | 1.5 ± 0.2b | 11.2 ± 1.0b |
| p | <0.001 | <0.001 | |
CP: Close cervix pyometra, OP: Open cervix pyometra, DE: Diestrus, AE: Anestrus. Values within the same column followed by different letters (a, b) are significantly different.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



