
Submitted:
07 September 2025
Posted:
09 September 2025
You are already at the latest version
Abstract
Aging is a complex biological process marked by gradual functional decline, reduced cellular resilience, and heightened vulnerability to chronic diseases. Maintaining cell health has become a crucial strategy for extending healthspan and promoting healthy aging. Nutrition at the cellular level plays a vital role in this effort, with bioactive compounds from functional foods increasingly recognized for their ability to influence pathways related to aging. Fruitflow®, a water-soluble tomato extract, contains a variety of bioactive compounds, including polyphenols, nucleosides, and phenolic conjugates, that support cardiovascular, metabolic, and immune health. Preclinical and clinical research demonstrate that Fruitflow® helps reduce platelet hyperactivity, lower blood pressure, and enhance blood vessel function. Moreover, emerging evidence suggests that it offers broader cellular benefits, including reducing oxidative stress, decreasing in-flammation, boosting mitochondrial activity, promoting cellular growth, and supporting a beneficial gut microbiota. These actions align with key aspects of aging, including oxidative imbalance, mitochondrial dysfunction, chronic inflammation, and impaired cellular communication. This review outlines current evidence on the cellular and sys-temic effects of Fruitflow®, explores the molecular mechanisms behind these effects, and assesses its potential as a dietary strategy to enhance cellular health and promote healthy aging.
Keywords:
Fruitflow ®
; Cellular aging
; Water soluble tomato concentrate
; Aging
; DNA repair
; DNA damage
; Cell proliferation
; Cell Damage
; Cell Senescence
; Mitochondria
; ROS
; Cell Cycle
; Polyphenols
; Flavonoids
; Cholorogenic acid
; Rutin
; Quercetin
; lifespan
; and Nucleosides
1. Introduction
Cellular health is essential for healthy aging. However, the aging process is highly complex, shaped by the interactions of numerous molecular mechanisms and cellular systems. From a phenotypic perspective, aging is characterized by a progressive decline in cellular function and widespread tissue degeneration, which contributes to the increased risk of age-associated disorders. While aging is unavoidable, optimizing cellular health can help support a healthy aging process and enhance longevity and quality of life [1,2]. Maintaining good cellular health is vital for overall health, vitality, and longevity, as healthy cells play a key role in energy production, metabolism, immune defense, and protection against chronic diseases [2,3].
Healthy aging, as defined by the World Health Organization (WHO), refers to the ability to develop and maintain the functional capacity required for well-being in older age [4]. This perspective extends beyond the absence of disease, emphasizing the preservation of physiological function, independence, and resilience throughout the aging process. From a geroscience standpoint, healthy aging involves slowing or counteracting the molecular and cellular processes that drive functional decline, thereby extending health span rather than just lifespan [5].
Cellular nutrition has become a transformative idea, based on the belief that supporting cells can significantly improve overall health, well-being, and longevity [3]. Supplying cells with the essential nutrients is the foundation of healthy living, optimal wellness, and healthy aging. It encompasses aspects such as how well cells produce energy, repair themselves, and communicate with one another. Among nutritional strategies, polyphenols are naturally occurring bioactive compounds abundant in fruits and vegetables, and they have attracted significant interest for their ability to modulate pathways associated with aging. Their protective actions include scavenging reactive oxygen species (ROS), attenuating inflammation, preserving mitochondrial activity, regulating nutrient-sensing mechanisms, and influencing epigenetic regulation[6,7,8]. Large-scale epidemiological studies consistently associate polyphenol-rich diets with a reduced risk of cardiovascular disease, diabetes, neurodegenerative disorders, and other conditions linked to aging [7,8,9]. Taken together, these findings identify polyphenols as important dietary factors with the potential to counteract the hallmarks of aging.
The progression of aging is influenced by several fundamental mechanisms, such as inflammatory activity, intracellular oxidative imbalance, loss of epigenetic fidelity, telomeric shortening, cellular senescence, DNA damage, protein homeostasis disruption, mitochondrial dysfunction, dysregulated nutrient sensing, defective intercellular signaling, and microbial dysbiosis, which together contribute to its acceleration [3]. Aging is also closely linked to decreased nicotinamide adenine dinucleotide (NAD+) levels, impaired DNA repair, defective macroautophagy, disruption of protein balance, impaired nutrient sensing, cell senescence, and stem cell exhaustion, all of which influence aging, body weight, glucose levels, and immune response [10]. In aging cells, mitochondria produce less energy, which increases cellular stress and raises the risk of cell senescence. The senescence-associated secretory phenotype (SASP) consists of inflammatory cytokines, lipid-based molecules, and other compounds that impair tissue function [11].
Several markers, consistently expressed across human cells and DNA, are linked to physiological aging. These markers are associated with increased chronic inflammation, immune system disruptions, and heightened oxidative stress, among other factors [12]. Strategies to counteract these effects include lifestyle modifications, such as regular exercise and a balanced diet, as well as emerging cellular rejuvenation therapies involving specific nutrients or functional foods [13]. Some dietary supplements and functional foods now market specialized ingredients as tools to reverse these markers [14]. Despite notable progress in medical science, maintaining a healthy diet remains the initial step toward wellness and the most effective defense against chronic and infectious diseases [15]. Multiple studies confirm that dietary supplements can be vital for maintaining health and preventing disease, provided they are selected based on scientific principles and properly tested [13].
Lifestyle and gene-targeted interventions, including calorie restriction, better sleep cycle, enhanced physical activity, and regulation of longevity-related genes, represent promising avenues for reducing age-associated decline. An in-depth exploration of the mechanisms underlying the reversibility of age-related phenotypic changes, along with an emphasis on the importance of cell viability and proliferation, mitochondrial function, cellular reactive species, senescence, and angiogenesis in cellular health, is crucial.
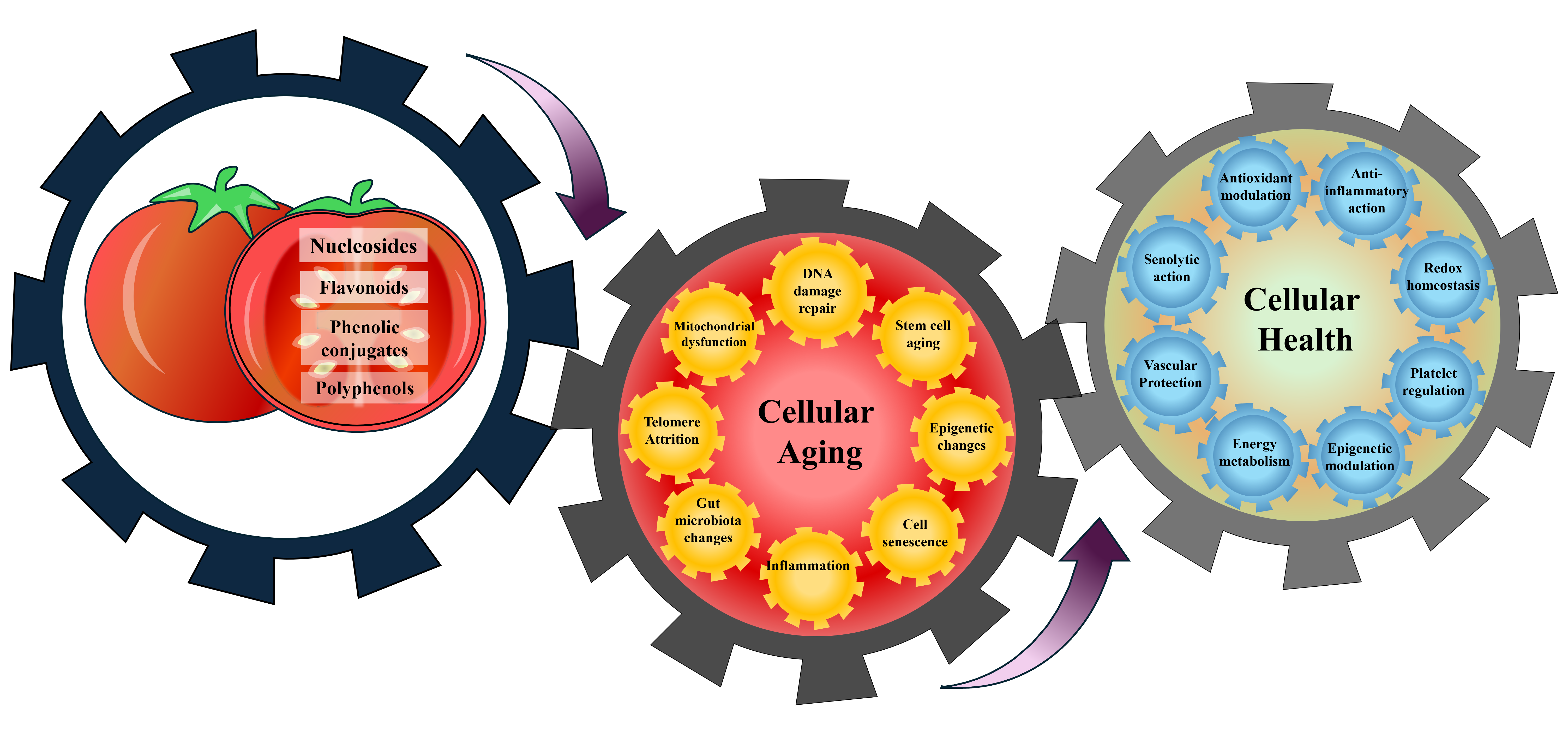
Fruitflow®, an aqueous tomato extract, is best known for its primary health benefit in promoting healthy blood flow by inhibiting platelet aggregation and maintaining blood pressure. However, recent studies using cells and animal models have shown that this extract can influence various cellular functions, including proliferation, mitochondrial activity, protection against oxidative damage, and anti-inflammatory effects, positioning it as a potential anti-aging agent.
Fruitflow can be utilized as a functional food to decrease platelet hyperactivity, as observed in conditions such as aging, diabetes, obesity, and other illnesses associated with smoking, pollution, and a high-fat diet [16].Fruitflow® contains a complex mixture of nucleosides, polyphenols, and phenolic conjugates that collectively exhibit antiplatelet, antihypertensive, antioxidant, and anti-inflammatory properties [16]. A standard 65 mg dose of Fruitflow® provides approximately 6–10% (up to 9 mg) nucleoside derivatives, 13–15% (up to 10 mg) phenolic conjugates, and 8–10% (up to 7 mg) flavonoid derivatives, including at least 2.4 mg of quercetin derivatives [16,17,18,19]. The various metabolic pathways of the Fruitflow® compounds lead to observable antiplatelet effects that occur 1.5 to 3 hours after ingestion and last (from a single dose) for approximately 18 hours [16]. The data suggest that the effective range for Fruitfow® lies between 75mg and 300mg, depending on the individual [17]. The antihypertensive effect of Fruitflow® has also been demonstrated in humans [17,18]. Fruitflow® reduced the levels of TXB2, 6-keto-PGF1α, and PF4 levels and inhibited platelet aggregation in ADP- and collagen-stimulated platelets [16,19]. This effect may occur through suppression of Akt/GSK3β, Syk/PLCγ2, and p38 MAPK phosphorylation in collagen-activated platelets [18]. Preclinical and translational research have highlighted the role of Fruitflow® in supporting cellular health by enhancing blood flow and reducing diseases associated with platelets, particularly those resulting from platelet hyperactivity [19]. Fruitflow® has garnered significant attention for its effects on immune function, mitochondrial health, and metabolic pathways [16,20,21]. Fruitflow® promotes these effects by modulating multiple mechanisms, including protection against ROS, oxidative stress, and cell death, as well as stimulating angiogenesis and signaling pathways [19,22,23].
Fruitflow® became the first ingredient to obtain an Article 13(5) health claim from the European Commission. The Commission Decision of December 17, 2009 (C(2009)10113), approved the claim that water-soluble tomato concentrate supports the reduction of platelet aggregation and afforded proprietary data protection in accordance with Regulation (EC) No. 1924/2006 https://eur-lex.europa.eu/eli/dec/2009/980/oj/eng.
Recent cellular and animal research also indicates protective effects against high-fat diet–induced pathology, chemotherapy-related cardiotoxicity, and trimethylamine-N-oxide (TMAO)-mediated metabolic disturbances [16,17,18,19,20,21]. These findings suggest that the combined bioactive compounds in Fruitflow® may act through multiple mechanisms, including suppression of oxidative stress, regulation of signaling pathways, and promotion of angiogenesis, to support cellular function and resilience [24,25,26,27]. The unique combination of diverse compounds in Fruitflow® makes it a distinctive and thoroughly researched functional dietary supplement that promotes cardiovascular health and cellular protection. This review emphasizes the potential role of bioactive compounds in Fruitflow® in supporting healthy aging and cellular health. This review examines the emerging evidence on Fruitflow® and its bioactive compounds, highlighting their potential to modulate key cellular processes and age-related pathways. It discusses how these effects may contribute to maintaining cellular health and promoting healthy aging.
2. Fruitflow® and the Dietary–Molecular Interplay in Aging
A complex interplay of genetic and environmental factors influences cellular aging. These can be broadly categorized into intrinsic factors, such as telomere shortening, DNA damage, and epigenetic changes, and extrinsic factors, including platelet hyperactivity, oxidative stress, gut microbiota, and lifestyle choices.
2.1. Dietary Bioactive Compounds of Fruitflow® and Telomere Attrition
Telomeres are short stretches of repetitive DNA–protein complexes that cap the ends of linear eukaryotic chromosomes, where they play a critical role in preserving genomic stability, regulating cell cycle dynamics, and supporting organismal longevity and reproductive capacity [24]. With aging, telomere shortening, mitochondrial impairment, and DNA damage compromise cellular metabolic homeostasis [25,26,27]. Given their limited reserves, telomere loss is closely linked to the decline in adaptive capacity during aging [28]. These telomere characteristics make them a key focus in the study of aging biology. Excessive telomere attrition triggers DNA damage responses, including cell cycle arrest, apoptosis, differentiation issues, and senescence [29]. As aging continues, the accumulation of ROS in hypoproliferative tissues, including the heart, brain, and liver, induces oxidative damage to telomeric sequences, thereby accelerating telomere attrition and ultimately resulting in telomere uncapping [29].
Nutrients such as vitamins C, D, E, polyphenols, omega-3 fatty acids, and fiber can help protect against telomere attrition [30]. Conversely, a diet high in processed or red meats and sugary drinks may be associated with shorter telomeres [30]. Studies involving telomere length in leukocytes have shown that omega-3 fatty acids are generally associated with longer telomeres [31]. However, findings for mono- or n-6 polyunsaturated fatty acids are less consistent, with some studies reporting no clear relationship[31,32] and others indicating a potential inverse association [33].
Diets rich in fruits, vegetables, legumes, tomatoes, and nuts are a significant source of polyphenols, unsaturated fatty acids, and fiber, and are linked to improving cardiovascular health and longer telomeres [34,35]. On the contrary, diets high in saturated fats, alcohol, and sugar are associated with inflammation, oxidative stress, and shorter telomeres. Polyphenols, through their antioxidant and anti-inflammatory effects, may slow telomere attrition, enhance telomerase activity, and support healthy aging [36]. Many studies suggest that polyphenols can influence telomere length and prevent its shortening, indicating that polyphenols may play a role in determining the state of telomere length.
Myricetin, a flavonoid, has been shown to interact with telomeres and telomerase, potentially affecting the growth of cancer cells and the aging process. Studies suggest that myricetin can inhibit telomerase activity in cancer cells by binding to telomeric G-quadruplex structures, thereby influencing cell proliferation. However, the effect of myricetin on overall telomere length in normal cells and its role in healthy aging still needs further research [31].
Dietary administration of polyphenols and curcumin in mice induced the presence of longer telomeres [34]. Similarly, Fruitflow® intake was associated with modifications in microbial β-diversity, while α-diversity remained unchanged. Significant differences in Jaccard evidenced this distance-based Principal Component Analysis between groups. Fruitflow® treatment was linked to reduced abundance of Bacteroides, Ruminococcus, and Hungatella, and an increased presence of Alistipes-associated OTUs [37]. Therefore, polyphenols present in Fruitflow® may help preserve the telomere length; however, no direct data are available.
Polyphenols exert several beneficial effects, including delaying telomere attrition, through mechanisms associated with the gut microbiota. Upon reaching the colon, many polyphenols are fermented by resident microbes, yielding smaller bioactive metabolites that contribute to their physiological activity [35]. For example, cocoa-derived polyphenols largely reach the colon, where they are metabolized by gut microbiota into smaller bioactive derivatives. In conjunction with their metabolites, these compounds contribute to the modulation of gut microbial communities. Evidence from preclinical and clinical studies suggests that cocoa has prebiotic effects, favoring the proliferation of beneficial bacteria, including Lactobacillus and Bifidobacterium, while suppressing the abundance of potentially harmful species, notably certain members of the Clostridium genus [35].
Further prospective studies and randomized controlled trials are needed to establish a causal link between food or dietary patterns and telomere attrition. A survey of 1,743 individuals from the WHICAP study found a significant association between the Mediterranean diet and telomere length in a non-Hispanic white population. However, no such association was observed in the overall population studied [36]. This positive link between adherence to the Mediterranean diet at baseline and telomere length was also observed in a group of women at high cardiovascular risk in the Prevention with Mediterranean Diet and the NHANES studies [38,39]. However, current evidence on dietary intake and telomere length remains limited, and inconsistencies persist. Further research is needed better to understand the relationship between diet and telomere length.
2.2. Modulatory Effects of Fruitflow® on Oxidative DNA Damage and Repair Pathways:
DNA damage results in epigenetic modifications, genomic instability, mitochondrial dysfunction, and telomere shortening [40]. The progressive accumulation of DNA-damaged cells promotes apoptosis and senescence, which in turn drive chronic inflammation, functional deterioration, tissue atrophy, and the onset of disease [41]. Maintaining DNA integrity involves continuously repairing the complex DNA repair and DNA damage response (DDR) systems that oppose the gradual loss and destruction of genetic information. Telomere shortening is another key factor contributing to DNA damage, which in turn accelerates aging [42]. DNA damage represents a key driver of age-associated epigenetic alterations. The epigenome, encompassing processes such as DNA methylation and histone modifications, exhibits progressive instability across the lifespan of somatic cells [43]. Damage to DNA causes lasting chromatin changes that promote the accumulation of aging-related DNA fragments in senescent cells [44]. Persistent DNA damage can leave epigenetic marks, alter transcription, and contribute to cellular heterogeneity and functional decline. Although mammalian cells experience ~10⁵ DNA damage events daily, most are repaired; however, some DNA escapes detection or is mis-repaired, linking DNA damage closely to aging [45,46,47]. Aging is characterized by a gradual decline in DNA repair proficiency, contributing to genomic instability as a central hallmark. This is reflected by the accumulation of DNA damage markers, which are prevalent in age-associated diseases, particularly cardiovascular disease, Alzheimer’s disease, and cancer. Acquired defects in DNA repair proteins can cause premature aging across organs [48,49].
Additionally, mitochondrial defects are another underlying factor that contributes to multi-organ damage. Low concentrations of polyphenols are often found to be protective against DNA damage, likely due to their antioxidant properties. They can neutralize harmful ROS that can damage DNA. However, high concentrations of polyphenols can have the opposite effect.
The polyphenols, primarily phenolic acids such as caffeic, ferulic, and chlorogenic acids, along with flavonoids like quercetin derivatives, protect against DNA damage through multiple complementary mechanisms [50]. Previous studies highlight the crucial role of dietary polyphenols in protecting DNA from oxidative damage. These bioactive compounds possess hydroxyl-rich structures that enable them to effectively neutralize reactive oxygen species and prevent oxidative modifications of DNA [51]. Experimental studies have demonstrated that polyphenols can significantly reduce DNA fragmentation and other forms of damage induced by oxidative stress. They directly scavenge reactive oxygen and nitrogen species, preventing oxidative base lesions and strand breaks, and chelate transition metals like iron and copper to inhibit Fenton reaction–driven hydroxyl radical formation near DNA [52]. Under cellular conditions, the Fenton reaction and inflammatory processes can generate carbonate radical anion, a strong oxidant that produces base radical cations in DNA. These radicals migrate to redox-sensitive sequences such as GGG, leading to the formation of 8-oxo-7,8-dihydroguanine. This enables G-quadruplex regions to function as long-range sensors of oxidative stress, influencing the DNA repair mechanism and the removal of oxidized bases [53]. The bioactive compounds present in Fruitflow® activate the Nrf2 pathway, boosting endogenous antioxidant enzymes such as superoxide dismutase, catalase, and glutathione peroxidase, while simultaneously suppressing NF-κB–mediated inflammation to reduce ROS and RNS generated during chronic inflammatory responses [54,55]. In addition, quercetin and related polyphenols may enhance DNA repair pathways, including base and nucleotide excision repair, and protect against lipid peroxidation–derived genotoxins such as 4-HNE and MDA. Oxidative stress, a key driver of aging, damages macromolecules, in part, through lipid peroxidation, producing reactive aldehydes such as 4-hydroxynonenal (HNE) and malondialdehyde (MDA). These aldehydes form long-lived adducts with critical proteins, impairing their function and accelerating molecular turnover. In aging-related disorders such as osteopenia, sarcopenia, immune senescence, and myelodysplastic syndromes, they play significant pathogenic roles [50,56,57]. Redox proteomic studies targeting aldehyde-protein modifications may aid in the development of therapies to reduce morbidity and mortality in these conditions. Through their combined antioxidant, anti-inflammatory, and DNA repair–supportive activity, Fruitflow® polyphenols help maintain genomic integrity and reduce mutation risk under oxidative stress conditions [Patent ID: CN120130651A].
2.3. Countering Diet- and Age-Related Mitochondrial Stress with Fruitflow®
A healthy mitochondrial network produces ATP through the TCA cycle and oxidative phosphorylation, supporting the cell’s essential energy conversion and information exchange vital for life. Conversely, disruptions in mitochondrial activity, including protein-folding stress within mitochondria, reduced energy availability, and excessive ROS generation, activate nuclear gene reprogramming as a compensatory metabolic response [57]. The decline in mitochondrial function is a central feature of aging and contributes significantly to metabolic disorders. Small molecules, including AMP, NAD⁺, oxygen, ROS, and TCA cycle intermediates, produced through mitochondrial activity, serve as signaling mediators that influence mitochondrial performance, nuclear function, and inter-organelle communication [58]. ATP serves as a sensitive marker of mitochondrial health. A sustained decline in intracellular ATP elevates AMP levels, thereby activating the AMPK signaling pathway [59]. Once activated, AMPK phosphorylates and regulates key metabolic enzymes, influencing lipid and glucose metabolism, mitochondrial dynamics, autophagy, and protein synthesis, thereby contributing to the restoration of cellular energy homeostasis. Disruption of this mechanism can lead to various mitochondrial diseases. Aging results from many biological processes, with factors such as mitochondrial dysfunction, DNA damage, telomere attrition, NAD+ loss, autophagy problems, and stem cell exhaustion contributing to it.
Furthermore, toxic byproducts, such as ROS, generated within mitochondria can interact with mitochondrial permeability pores when calcium (Ca2+) levels are high, leading to oxidative damage and cellular swelling. This process promotes inflammation and impairs mitochondrial function [60]. Optimal mitochondrial function represents a cornerstone of longevity, with the stability of mtDNA and nuclear DNA serving as critical determinants of cellular energy homeostasis and tissue maintenance during aging [21,61]. Unlike nuclear DNA, mitochondrial DNA (mtDNA) can replicate continuously, independent of the cell cycle. Due to the low repair efficiency of mtDNA, mutated copies tend to accumulate over time. As organisms age, mutations in nuclear DNA and mtDNA can surpass a threshold, causing harmful effects such as disrupted glycolipid metabolism, cognitive decline, and a shortened lifespan [62]. mtDNA methylation is a key epigenetic modification influenced by environmental and metabolic factors. Hypomethylation of the D-loop region is associated with increased risk of neurodegenerative diseases such as amyotrophic lateral sclerosis and Parkinson’s disease. Mitochondrial dysfunction results from transcriptional and epigenetic changes triggered by stress responses, including mtDNA mutations, defective clearance, lysosomal impairment, ER stress, and cytoplasmic alterations. Due to their sensitivity to the microenvironment, mitochondria act as central drivers of aging and metabolic disorders. However, further studies are needed to clarify the mechanistic links, which may lead to new targets for anti-aging therapies. Similarly, NAD⁺ is an essential cofactor that participates in numerous metabolic reactions and plays a pivotal role in sensing and transmitting the mitochondrial metabolic state to other organelles. Under normoxic conditions, cellular homeostasis is maintained through the stabilization of prolyl hydroxylase domain enzymes (PHDs), which regulate hypoxia-inducible factors (HIF-1 and HIF-2). This process safeguards mitochondrial integrity and minimizes damage associated with fluctuations in oxygen levels. Alterations in NAD⁺ metabolism disrupt cellular redox balance and energy homeostasis, thereby impairing processes such as DNA repair, organelle function, immune cell survival, and overall cellular integrity. Abnormal NAD⁺ dynamics are strongly implicated in aging, with evidence from both rodent and human studies showing a direct association between NAD⁺ levels, health status, and longevity.
As we age, NAD+ levels decline, which disrupts energy metabolism and increases disease risk. During aging, degradation of NAD+ exceeds its synthesis, and excess nicotinamide (NAM), the amide form of vitamin B3 and an essential precursor in the NAD+ salvage pathway, is diverted into alternative routes. By midlife, NAD+ levels fall to nearly half of those in youth, accelerating functional decline and the development of age-related disorders [63]. The precise mechanisms remain unclear. Recent research suggests that aging triggers inflammation and oxidative stress, which impair the rate-limiting enzyme for NAD+ production and impact the activity of downstream NAD+-dependent enzymes, such as Sirtuins, PARPs, and CD38 [64]. Senescent cells exhibit altered mitochondrial morphology, reduced membrane potential, increased proton leak, and elevated ROS levels, which impair fatty acid oxidation and overall metabolism. Mitochondrial dysfunction arises from factors such as low ATP, NAD+, and oxygen levels, accumulated ROS, and disrupted TCA cycle signaling, leading to stress responses, mtDNA mutations, and secondary effects on lysosomes, the ER, and cytoplasmic homeostasis. As key regulators of redox balance and energy metabolism, mitochondria and NAD+ play a central role in the aging process. A decline in NAD+ levels, a hallmark of aging, compromises DNA repair, organelle function, immune activity, and metabolic stability, thus promoting age-related diseases. Although NAD+ is usually synthesized, recycled, and degraded to maintain balance, this regulation becomes impaired with age [65,66]. NAD+ cofactors such as quercetin and apigenin, found in Fruitflow® [15], may help restore cellular function during aging.
A mitochondrial function diet emphasizes whole foods rich in antioxidants, B vitamins, and healthy fats while limiting processed foods and refined sugars [67]. High-fat diets impair mitochondrial function, whereas calorie restriction, fasting, ketogenic diets, and the Mediterranean diet generally enhance mitochondrial biogenesis, mitophagy, and renewal. By sustaining mitochondrial health, these dietary patterns help slow cellular aging and support stem cell function, thereby promoting tissue repair and healthy aging.
In contrast, the Mediterranean diet has the opposite effect on mitochondrial dynamics and autophagic processes. A high-fat diet (HFD) promotes chronic, low-grade systemic inflammation, thereby accelerating the progression of atherosclerosis and compromising cardiovascular function [68]. Experimental evidence in rat models demonstrates that HFD consumption elevates key inflammatory markers, including leptin, tumor necrosis factor-α (TNF-α), intercellular adhesion molecule-1 (ICAM-1/1/CD54), and von Willebrand factor (vWF) [69], indicating increased inflammation, endothelial dysfunction, and cardiovascular risk. Treatment with Fruitflow® effectively lowers these markers in HFD-fed rats[69]. These findings suggest that Fruitflow® may confer protective effects against HFD-induced inflammation, endothelial dysfunction, and pro-thrombotic events, effects that are likely attributable to its anti-inflammatory and antiplatelet properties. The evidence suggests that Fruitflow® effectively attenuates HFD-induced dyslipidemia, inflammation, and oxidative stress, reflected by marked reductions in cholesterol, triglycerides, inflammatory mediators, ROS, and lipid peroxidation, together with enhanced catalase activity.
Moreover, Fruitflow® modulates the expression of critical genes associated with cardiac structure and function, thereby mitigating HFD-induced alterations and promoting cellular repair and recovery. Additionally, Fruitflow® further reduced fibrosis, fat cell infiltration, collagen buildup, and α-smooth muscle actin levels. Fruitflow® protects against cardiotoxicity and HFD-induced cardiac dysfunction through multiple mechanisms [69]. These findings suggest that Fruitflow® can safeguard cells from stress caused by diet and drugs.
2.4. Role of Rutin, Quercetin, and CGA in Modulating Senescence and SASP-Related Aging
Cell senescence involves the irreversible arrest of the cell cycle, development of secretory features, damage to macromolecules, and changes in metabolism, which influence each other in the process of aging. It may act as a warning mechanism against harmful stimuli or uncontrolled proliferation, leading to outcomes such as cell cycle arrest, quiescence, or terminal differentiation. Senescent cells are characterized by the accumulation of the CDK2 inhibitor p21WAF1/CIP1 (CDKN1A) and the CDK4/6 inhibitor p16INK4A (CDKN2A) [70]. Senescent cells release a wide range of factors, including proinflammatory cytokines, chemokines, growth modulators, angiogenic molecules, and matrix metalloproteinases, collectively known as the senescence-associated secretory phenotype (SASP). The onset of cellular senescence can be triggered by multiple stresses, including genomic damage, telomere attrition, epigenetic alterations, protein and lipid modifications, mitochondrial and lysosomal dysfunction, excessive ROS, and chronic inflammation [71]. Cells undergoing senescence typically display significant damage, ranging from DNA and telomere disruption to mitochondrial dysfunction and oxidative stress caused by ROS accumulation.
Treatments that modulate cellular senescence are essential for aging interventions. While eliminating senescent cells has been shown to lower the incidence of age-related diseases, it does not necessarily slow the overall aging process. Clearing senescent cells also creates space for new cells, depending on the proliferation of stem cells or other resident cells. Cellular senescence helps maintain tissue function, but it also risks the accumulation of damage. Therefore, in the short term, intermittent removal of senescent cells may be the most effective approach to balance these factors. Reduced NAD⁺ during senescence is linked to inflammation, genomic instability, and neurodegeneration, ultimately exacerbating senescence, apoptosis, cancer risk, mitochondrial dysfunction, oxidative stress, and impaired proteostasis. Throughout their lifespan, stem cells detect signals from damaged and aging cells, migrate to repair and regeneration sites, and differentiate into the necessary cell types to support bodily functions. With advancing age, the proportion of stem cells relative to the total number of cells progressively declines. Numerous studies have demonstrated that aging introduces a range of cellular and tissue alterations, including enhanced DNA damage, replication stress, loss of polarity, mitochondrial defects, impaired autophagy, and epigenetic alterations, all of which drive stem cell aging and depletion. With aging, alterations in the stem cell niche can lead to the accumulation of stem cells, and the disparities observed between “young” and “old” stem cells may be driven more by mechanical changes in their microenvironment than by intrinsic cellular aging [72,73,74,75,76].
A cell senescence diet is a strategy designed to reduce cellular aging, which is linked to aging and age-related diseases. This involves consuming foods rich in antioxidants and senolytic compounds, while considering the impact of macronutrients and overall dietary patterns on cellular aging. Clearing senescent cells through a transgenic method helps delay the onset of several age-related diseases and extends the lifespan of progeroid and naturally aged mice [77]. Small molecules called senolytics can selectively remove senescent cells or their SASP, which are suppressed by senomorphics drugs [78]. Several senolytics have been shown to reduce senescence-related traits in preclinical models [79]. Some polyphenols have shown potential in influencing cellular senescence, a process involved in aging and related diseases [80]. Polyphenols exert diverse therapeutic effects through various mechanisms, including the stimulation of SIRT1, suppression of NF-κB activity, and modulation of autophagy. They are capable of maintaining the viability of healthy cells while promoting the clearance of senescent ones through apoptosis.
Experimental studies suggest that polyphenols can mitigate SASP-associated inflammation, enhance tissue function, and reduce the accumulation of senescent cells. Some members of this group also act as senolytics, selectively eliminating dysfunctional cells and thereby supporting tissue regeneration and healthy aging. Resveratrol, in particular, is recognized for enhancing SIRT1 activity, an NAD⁺-dependent deacetylase involved in numerous aging-related signaling and transcriptional pathways. Since SIRT1 levels decline with advancing age, strategies that increase its activity have been associated with delayed cellular senescence and extended lifespan in various model organisms [81]. Quercetin, kaempferol, apigenin, naringenin, epigallocatechin gallate, procyanidin C1, curcumin, rutin, oleuropein aglycone, hydroxytyrosol, fisetin, and luteolin all have senolytic activity and can influence multiple pathways related to cellular aging. These molecules contribute to limiting the accumulation of senescent cells and lowering SASP secretion, which may enhance tissue function and alleviate age-related inflammation. Nonetheless, despite encouraging preclinical findings suggesting that polyphenols could counteract cellular senescence, considerable challenges must be addressed before they can be translated into clinical applications.
The combination of bioactive compounds and a specific food matrix can also alter the bioavailability of these compounds [81]. In the context of senescence, several studies have highlighted the limitations of relying solely on individual phytochemicals. On the other hand, plant extracts have been investigated and often display stronger efficacy. For instance, recent work with Voghera pepper extract showed protective activity against senescence in aged normal human diploid fibroblasts [82]. Future research should prioritize the identification of synergistic phytochemical combinations that can serve as novel nutritional strategies for the safe and effective management of aging and age-related diseases, while also promoting healthier dietary practices. Further investigations are also needed to establish clinical evidence and assess the long-term outcomes of these compounds, supporting their potential advancement as therapeutic options in geriatric care.
Bioactive compounds with senolytic or senomorphic properties can influence multiple signaling cascades involved in cellular senescence, thereby reducing the accumulation of senescent cells and attenuating SASP secretion, which preserves tissue homeostasis and limits systemic inflammation during aging [83]. Rutin, a major polyphenolic flavonoid in Fruitflow®, has been identified as a novel senomorphic agent targeting the early stages of SASP development. In bleomycin-induced senescent PSC27 cells, rutin inhibited ATM kinase activation and its downstream mediators—HIF1α, a master regulator of homeostasis during senescence, and TRAF6, a key SASP driver—thereby suppressing the acute stress-associated phenotype (ASAP) and preventing SASP progression [84]. In diabetic mouse models, rutin improved glucose and lipid metabolism, reduced atherosclerotic plaque burden, limited senescent cell accumulation, and restored vascular smooth muscle cell (VSMC) content in aortic root lesions. In vitro studies have demonstrated that rutin protects VSMCs from H₂O₂-induced premature senescence by attenuating oxidative stress and preserving telomeres [84]. Other such compounds present in Fruitflow® include chlorogenic acid (CGA) and quercetin, which are known to exert anti-aging effects by modulating oxidative stress and senescence-associated pathways. CGA exhibited vascular anti-aging effects by upregulating Sirt1 and eNOS, key regulators of vascular senescence. Despite eNOS being downstream of Akt activation, CGA inhibited H₂O₂-induced Akt phosphorylation, potentially preventing excessive ROS generation and HO-1 overexpression. By modulating AKT1 phosphorylation, CGA may restore intracellular redox balance and delay senescence onset [85]. Quercetin similarly exerts senomorphic effects. In H₂O₂-induced senescent pre-adipocytes and adipocytes, quercetin reduced SA-β-gal positivity, ROS levels, and pro-inflammatory cytokine expression, accompanied by decreased miR-155-5p levels, p65 downregulation, and a trend toward increased SIRT1 expression [86]. In atherosclerotic models, it also inhibited macrophage senescence and inflammation by suppressing the p38 MAPK/p16 pathway [87]. Collectively, these findings suggest that the bioactive compounds present in Fruitflow® may also synergistically target oxidative stress, redox imbalance, and pro-inflammatory signaling, thereby modulating senescence pathways and mitigating vascular and metabolic aging.
2.5. Cell Proliferation, Ageing and Fruitflow®
Aging and cell proliferation are closely connected processes that reflect the balance between cellular renewal and decline. Strong proliferation, supported by active stem cells and effective DNA repair systems, maintains tissue integrity in young individuals. However, telomere shortening, accumulated DNA damage, and increased oxidative stress caused by aging disrupt this balance, leading to cellular senescence and a reduction in proliferative capacity [88]. Senescent cells also impair tissue regeneration and promote aging-related disorders by releasing matrix-degrading enzymes and inflammatory cytokines [89]. Changes in the tissue microenvironment, such as increased rigidity of the extracellular matrix, cause endothelial and stem cell senescence, further limiting their ability to proliferate [90].
Additionally, mitochondrial dysfunction impairs pro-senescent signaling and bioenergetics in aging cells [91]. Understanding the molecular mechanisms that control proliferation in aging cells, such as the CXCR4/CXCL12 signaling pathway and the p53/Δ133p53α axis, may help identify new therapeutic targets to improve tissue repair and delay age-related decline [92,93,94]. Adenosine is a nucleoside that is a key component of Fruitflow®. It plays a vital role in regulating vascular function and cellular energy metabolism [95,96]. Adenosine encourages endothelial proliferation and angiogenesis by activating its A2A and A2B receptors, both of which are essential for tissue regeneration and repair. This has been demonstrated in vitro, where adenosine stimulated the migration of endothelial cells and the formation of capillary tubes, indicating proliferative support in vascular cells [97].
The flavonoid glycoside rutin, which includes quercetin and the disaccharide rutinose, has effects on cell proliferation that depend on the context. In normal cells, it has been shown to promote growth primarily due to its antioxidant and protective properties. Ugusman et al. (2014) demonstrated that rutin significantly increased the expression of endothelial nitric oxide synthase (eNOS) and nitric oxide (NO), a key signaling molecule involved in maintaining vascular health and repairing the endothelium, in human umbilical vein endothelial cells (HUVECs)[98]. Although the study did not directly measure proliferation markers, it is known that higher NO levels and increased eNOS activity indirectly promote endothelial growth and new blood vessel formation, especially during tissue repair. In wound healing models, Almeleebia et al. (2025) showed that rutin-loaded nanocrystals significantly boosted fibroblast migration, growth, and collagen production [99]. Rutin has been shown to promote tissue remodeling and skin regeneration, key indicators of cellular growth in skin systems, as demonstrated in studies with fibroblast cultures and animal models. In contrast, it displays anti-proliferative effects in cancer, inducing cell cycle arrest, often at the G2/M phase, and triggering apoptosis through the regulation of p53, Bax, and caspase pathways [100,101]. Rutin selectively influences growth based on cell type and physiological condition, demonstrating dual roles: promoting growth during healing and regeneration, and inhibiting growth in cancer cells.
Fruitflow® contains high levels of CGA, a phenolic compound with potent antioxidant properties. It helps prevent oxidative DNA damage, a significant obstacle to cell growth, by neutralizing ROS. However, like rutin, CGA can both inhibit and stimulate cell proliferation depending on the cellular environment. A recent study demonstrated that CGA enhanced the growth of human embryonic stem cells (hESCs) by increasing fatty acid β-oxidation and lipid production—two key metabolic processes crucial for rapidly dividing cells [102]. Conversely, another study found that CGA could suppress cell migration, invasion, and proliferation by inducing ferroptosis through the PTGS2/AKR1C3/GPX4 pathway in hepatocellular carcinoma [103]. Similar to rutin, CGA has two distinct effects on cell proliferation: it promotes growth in antioxidant and regenerative settings but inhibits growth in tumor models. These effects are mediated through apoptosis, regulation of oxidative stress, and cell cycle arrest.
Using H9c2 cardiomyoblasts treated with the chemotherapy drug doxorubicin, Fruitflow® dramatically improved cell survival, decreased oxidative stress indicators, and restored mitochondrial membrane potential in extensive in vitro research [69]. These results suggest that its constituents collaborate to prevent cell death and revive the systems involved in proliferation. In summary, Fruitflow®’s constituents influence cell proliferation through several overlapping pathways, including growth factor signaling, mitochondrial support, and antioxidant defense. This makes it a promising therapeutic option for applications requiring tissue regeneration and protection from oxidative cellular damage, in addition to being an effective cardiovascular nutraceutical.
2.6. Fruitflow® Constituents in Regulating Cell Proliferation and Tissue Regeneration
Inflammation is a key factor in both the onset and progression of aging. It triggers the recruitment of immune cells from the circulation, which then move across the endothelial barrier, causing vascular injury, arterial narrowing, and stiffening [90]. Low-grade inflammation is associated with various diseases, including diabetes and atherosclerosis. Activating inflammatory pathways in macrophages is crucial for both initiating and exacerbating endothelial dysfunction, ultimately contributing to the development of atherosclerosis. This condition may worsen in patients with hypertension, cardiovascular disease, and obesity, all of which are linked to baseline platelet hyperreactivity. Eating tomatoes and tomato products has been linked to a lower risk of chronic inflammatory diseases, including atherosclerosis, and is associated with a healthy Mediterranean diet [90,91].
Increased oxidative stress and platelet apoptosis are significant risk factors for atherosclerotic cardiovascular disease. Fruitflow® has been shown to increase serum total antioxidant capacity levels and decrease serum malondialdehyde levels after four weeks of supplementation in healthy middle-aged and elderly adults. The production of endogenous ROS in platelets, mitochondrial membrane potential loss, and phosphatidylserine exposure was decreased. Fruitflow® can inhibit oxidative stress and related platelet apoptosis, potentially supporting its primary prevention effects against aging [69].
Nuclear factor-κB (NF-κB) is a major pro-inflammatory transcription factor, and its activation is closely linked to the progression of vascular inflammatory conditions. Research on gene expression demonstrated that Fruitflow® can suppress NF-κB levels, suggesting it helps regulate inflammatory mediators via the NF-κB signaling pathway. Similar findings were reported by Navarrete et al., who showed that an aqueous tomato extract reduced pro-inflammatory cytokines (TNF-α, IL-1β) and inhibited NF-κB activation in LPS-stimulated THP-1 macrophages [104]. Moreover, ICAM-1 and VCAM-1, which are early markers of endothelial dysfunction and atherosclerosis, were significantly downregulated in both production and gene expression in activated HUVECs after Fruitflow® treatment, as reported by Schwager et al. (2016) [15,105]. These outcomes suggest that Fruitflow® may offer protective effects against disorders associated with endothelial dysfunction. Adenosine in Fruitflow® is suggested to play a role in regulating immune activity and inflammation, potentially protecting against certain inflammatory conditions. In LPS-stimulated macrophages, Fruitflow® was shown to modulate cytokine and interleukin levels by reducing the release of pro-inflammatory mediators such as TNF-α, IL-1β, and IL-12, while enhancing the production of the anti-inflammatory cytokine IL-10. This shift contributes to reducing inflammatory factors commonly associated with chronic inflammation. Another key compound, chlorogenic acid, is a polyphenol with anti-inflammatory properties that suppresses LPS-induced NF-κB activation and the downstream inflammatory response. Through such mechanisms, Fruitflow® may impact signaling pathways associated with vascular health, the formation of atherosclerotic lesions, and cardiovascular disease (CVD) risk. However, the degree to which these effects occur in humans after Fruitflow® consumption remains to be fully established. A wealth of evidence has long demonstrated the influence of platelets on physiological systems beyond hemostasis [20]. Platelets serve as key mediators in both initiating and regulating inflammation. Recent studies have examined the effects of Fruitflow® on the inflammatory response of macrophages and endothelial dysfunction in HUVECs [105]. This suggests that Fruitflow® may affect specific pathologies often linked to endothelial dysfunction. Platelets release numerous inflammatory mediators that are not involved in hemostasis. These mediators influence how leukocytes and endothelial cells respond to various inflammatory stimuli [106]. Platelet-leukocyte aggregates are now recognized as essential components in atherosclerosis. Platelet P-selectin primarily facilitates the connections between leukocytes and the endothelium [106]. Platelets play a key role in regulating inflammation through their interactions with monocytes, neutrophils, lymphocytes, and endothelial cells. Following injury or disease, their ability to rapidly respond and recruit leukocytes plays a vital role in initiating inflammatory processes. While this reactivity is advantageous for host defense, excessive or prolonged platelet activity can contribute to heightened inflammation, promoting tissue damage and disease progression in disorders such as atherosclerosis, diabetes, and inflammatory bowel disease [107]. Antiplatelet therapy is often used to reduce platelet hyperactivation and manage inflammation.
2.7. Cell cycle, Oxidative Stress, and Fruitflow®
Fruitflow® was found to influence the cell cycle, boost mitochondrial activity, and lower oxidative stress. It also led to notable improvements in lipid metabolism and enzyme function, thereby reducing both inflammation and oxidative damage in rats maintained on a high-fat diet [69]. Fruitflow® has been reported to affect the expression of genes linked to cardiac remodeling, mitochondrial biogenesis, vascular health, and inflammatory pathways. In animals fed a high-fat diet, this treatment helped improve lipid balance and mitigate inflammation and oxidative stress, as evidenced by decreases in cholesterol, triglycerides, ROS, LPO, and inflammatory mediators, accompanied by an increase in catalase activity [69].
2.8. Age-Related Platelet Hyperactivity and Fruitflow®
Platelet hyperactivity, commonly associated with conditions such as diabetes, cardiovascular disease, or exposure to high levels of pollution, plays a significant role in increasing health risks. Its significance is highlighted by the elevated likelihood of thrombosis observed in these situations [20]. Increased platelet activity is often observed in obesity, diabetes, high blood pressure, smoking, pollution exposure, and a sedentary lifestyle. Reactive oxygen species (ROS) enhance this process by serving as secondary messengers or by forming oxidized proteins that are essential for platelet aggregation. [108,109]. This could help clarify why older individuals are more susceptible to thrombosis. With aging, platelet activity rises, contributing to a higher risk of vascular and thrombotic disorders. Platelets in elderly individuals differ from those in younger people in terms of count, function, and structure, and these age-related variations are strongly linked to the prothrombotic condition commonly observed in aging. Platelets, upon activation, can recruit leukocytes and progenitor cells to vascular sites [68]. Platelets secrete a range of proinflammatory, anti-inflammatory, and angiogenic molecules, along with releasing microparticles into the bloodstream, which contribute to pathological processes. These secreted factors also help maintain platelet activation, further influencing disease progression. With aging, platelet hyperactivity—marked by heightened aggregation and reactivity—becomes more prevalent. Current research continues to investigate the molecular pathways underlying this age-related increase in platelet activity. Researchers are investigating therapeutic strategies to regulate platelet activity and reduce the likelihood of thrombosis and cardiovascular complications in the elderly. Evidence suggests that platelets in older adults show functional and structural differences, making them more easily activated and less responsive to inhibitory signals. Such age-related alterations may drive platelet hyperactivity and contribute to a prothrombotic condition later in life. While the exact mechanisms are still unclear, various stimuli appear to trigger heightened platelet responsiveness through early activation pathways. Moreover, the wide range of platelet functions allows them to interact intricately with other vascular cells, extending their role beyond hemostasis [20]. As research advances, it is increasingly recognized that platelet-targeted therapies may be useful in managing inflammatory and infectious diseases beyond their traditional roles. While the exact contribution of platelet hyperactivity to these conditions is not fully understood, preventive antiplatelet approaches hold promise. Early control of platelet activation may play a crucial role in slowing disease progression. Nonetheless, in people without cardiovascular disease, routine use of antiplatelet drugs is not advised because of potential complications, including internal bleeding, resistance, and other side effects.
A major challenge in treating these conditions is identifying agents that can selectively block the pathways linked to platelet activity without completely impairing their essential clotting function. Since common antiplatelet drugs such as aspirin are not advised for primary prevention, there is a need for safer alternatives with minimal side effects. Over the last two decades, numerous studies have demonstrated that water-soluble compounds derived from tomatoes are capable of reducing platelet aggregation in both experimental and in vivo settings [15]. This water-soluble tomato extract (trade name Fruitflow®) was the first food ingredient to receive an EC-approved health claim, after evaluation by EFSA in 2009 (EFSA Journal 2009, 1101, 1-15; EFSA Journal 2010; 8(7), 1689). The approved claim was ‘Helps to maintain normal platelet aggregation, which contributes to healthy blood flow’. Fruitflow® is now a well-established, naturally derived functional food ingredient, marketed worldwide. Several human studies have indicated that, generally, consuming Fruitflow® can reduce platelet activity by up to 25% compared to baseline levels [15].
3. Modulatory Role of Fruitflow® in Stem Cell Aging
The aging of adult stem cells significantly contributes to the decline in tissue regeneration, repair, and overall homeostasis. This stem cell aging results from a complex interaction of cellular and molecular hallmarks, including genomic instability, telomere shortening, and epigenetic changes that weaken genomic integrity and gene regulation. Additionally, the accumulation of damaged or misfolded proteins (loss of proteostasis), impaired autophagy, and mitochondrial dysfunction all contribute to the deterioration of cellular stability. Dysregulated nutrient-sensing pathways, increased oxidative stress, and chronic low-grade inflammation also promote cellular senescence and disrupt intercellular communication. Collectively, the hallmarks of stem cell aging ultimately lead to stem cell exhaustion and microbiome imbalance, further accelerating the aging process [110]. Currently, slowing cellular aging involves activating sirtuins, reactivating telomeres, reducing inflammation, or using bioactive compounds, along with other therapeutic strategies [111]. Research is increasingly focused on identifying natural compounds that can counteract these processes, which is a growing area in regenerative medicine and healthy aging. Emerging evidence suggests that various dietary polyphenols and bioactive compounds can enhance cellular proliferation, inhibit tumor development, mitigate inflammatory responses, and modulate
multiple cellular signaling pathways. Considering such options, compounds rich in polyphenols and bioactive molecules may have a potential role in anti-aging therapy. Polyphenols may have a positive impact by regulating the redox state, modulating cell signaling, and protecting cellular and biological molecules, such as lipids, proteins, and nucleic acids [110]. Studies have shown that diets high in polyphenols may offer potent anti-aging benefits and mitigate the effects of various health disorders.
Fruitflow® is primarily recognized for its cardiovascular protective and antiplatelet effects. Although its direct impact on stem cell aging has not been explored, its bioactive compounds have been analyzed separately for their antioxidant, anti-inflammatory, metabolic, and epigenetic modulating activity, which are consistent with the cellular features of aging [110].
Genomic instability resulting from cumulative DNA damage and defective repair processes is a key driving force in stem cell senescence. Quercetin and rutin are highly effective antioxidants that modulate ROS levels, thus mitigating oxidative DNA damage. These polyphenols have been demonstrated to maintain genome integrity by inhibiting ROS-induced strand breaks and stimulating DNA repair enzyme activity [111,112]. Progressive telomere shortening restricts stem cells’ proliferative capacity, contributing to cellular aging. Although there is currently no direct evidence linking tomato-derived polyphenols to telomerase activation, compounds such as quercetin have been shown to help preserve telomere integrity indirectly by mitigating oxidative stress, which is one of the main drivers of telomeric DNA damage. Due to their extensive biological activities, polyphenols remain promising candidates in pharmaceutical and medical research for promoting health, preventing, or managing a wide range of age-related disorders [113]. Epigenetic modifications, including DNA methylation, histone alterations, and non-coding RNA, are not only central features of aging-related diseases but also fundamental molecular mechanisms that drive the aging process itself. Bioactive compounds, such as quercetin and chlorogenic acid, have been reported to modulate key epigenetic regulators, including DNA methyltransferases (DNMTs) and histone deacetylases (HDACs), thereby potentially restoring youthful epigenetic patterns and supporting the preservation of stem cell identity [114]. Autophagy plays a crucial role in removing damaged organelles and maintaining cellular health. Aging stem cells often exhibit diminished autophagic activity, resulting in the accumulation of damaged organelles and protein aggregates. The AMPK–SIRT1 axis plays a crucial role in regulating autophagy activation. AMPK phosphorylates ULK1 to trigger autophagy and suppresses mTORC1, which functions as a negative regulator for the process. Autophagy is further enhanced by SIRT1, a NAD⁺-dependent deacetylase, which modulates autophagy-related proteins and stress-response pathways. Quercetin has been shown to promote autophagy through the AMPK–SIRT1 pathway, which helps eliminate damaged mitochondria and restore cellular homeostasis in aging cells [115]. Aging disrupts proteostasis, leading to the accumulation of misfolded proteins. Tomato-derived polyphenols, such as rutin and quercetin, help prevent protein oxidation and enhance chaperone activity [116]. Age-related disruption in nutrient-sensing pathways, such as the mTOR and AMPK, impairs metabolic homeostasis. Bioactive compounds, like quercetin and adenosine, have been shown to activate AMPK and inhibit mTOR signaling, thereby mimicking the effects of caloric restriction and promoting cellular metabolic balance [117].
Figure 1.
Schematic representation of the potential effects of polyphenols and nucleosides on stem cell aging and its associated hallmarks.
Figure 1.
Schematic representation of the potential effects of polyphenols and nucleosides on stem cell aging and its associated hallmarks.
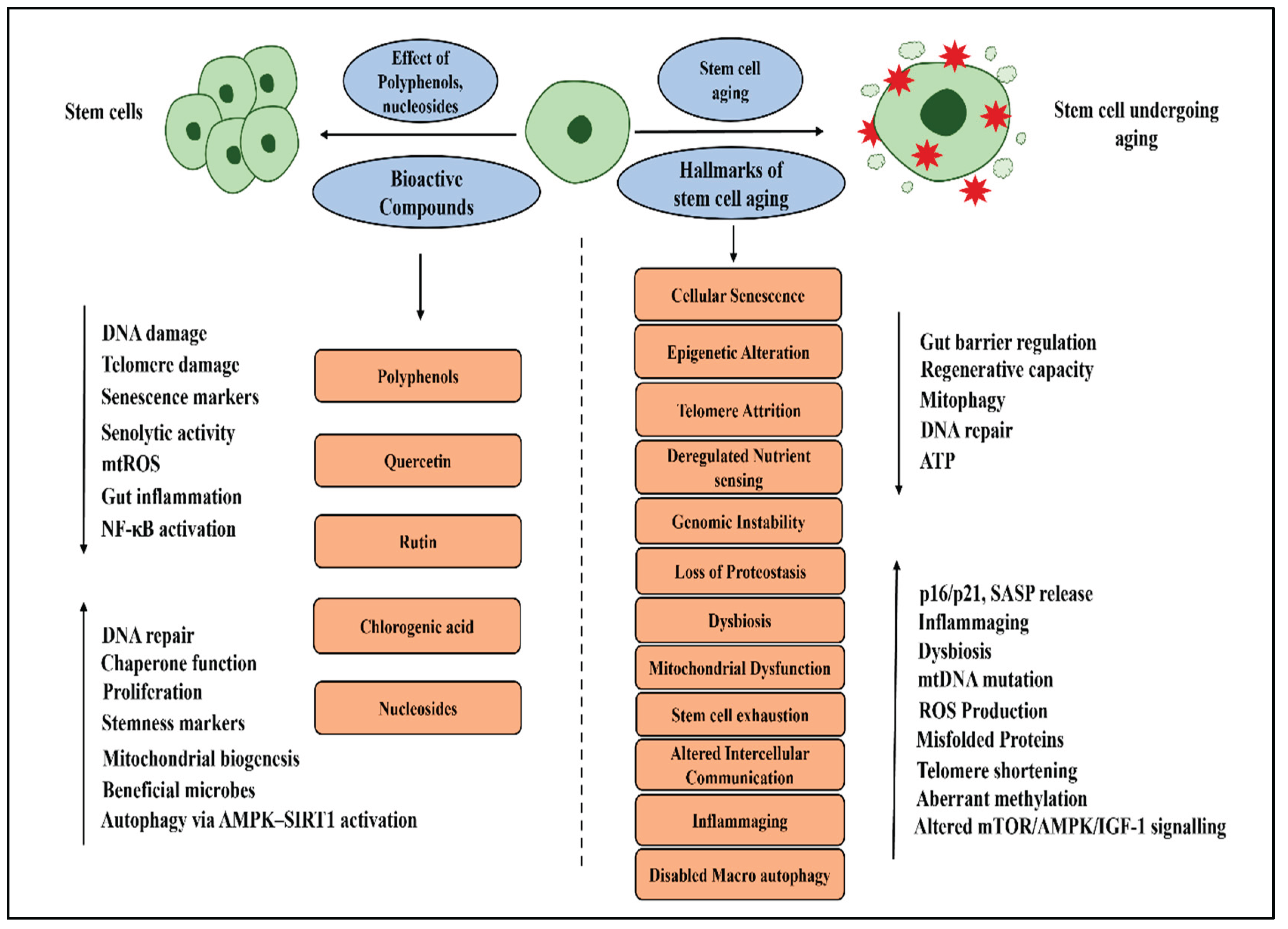
Mitochondrial dysfunction is another hallmark of aging, leading to excess ROS and decreased energy production. Studies have shown that quercetin, rutin, and chlorogenic acid promote mitochondrial biogenesis and lower ROS levels [118]. Adenosine and quercetin have been shown to reduce the release of pro-inflammatory cytokines and inhibit NF-κB activation. Quercetin, rutin, and chlorogenic acid also reduce inflammation by inhibiting the MAPK and TLR signaling pathways. Additionally, rutin and chlorogenic acid support gut microbiota and strengthen the intestinal barrier, indirectly enhancing the function and health of intestinal stem cells [111]. Although the connection between polyphenols and aging is not yet fully understood, growing research suggests that these bioactive compounds and nucleosides may have a positive influence on several aging-related processes. While polyphenols show promise in impacting aging, the exact mechanisms and their direct links to aging still need further clarification.
4. Regulation of Angiogenesis in the Context of Aging: Effects of Bioactive Compounds of Fruitflow®
Angiogenesis is the process of forming new blood vessels from existing vasculature and is essential for tissue oxygenation and nutrient delivery [119]. In aging, a decline in pro-angiogenic signaling, particularly reductions in VEGF (vascular endothelial growth factor) causes impaired wound healing, reduced neurovascular integrity, and decreased skeletal muscle perfusion [120]. These changes are linked to common age-related conditions such as sarcopenia, cognitive decline, and ischemic cardiovascular diseases [121,122]. On the other hand, abnormal or excessive angiogenesis in older adults can lead to pathological conditions like cancer progression and age-related macular degeneration. Maintaining a proper balance of angiogenesis is therefore crucial for enhancing longevity and preserving physiological function during aging Field [123].
Figure 2.
Schematic representation of the proangiogenic effect of Adenosine, AMP, and Quinic Acid. .
Figure 2.
Schematic representation of the proangiogenic effect of Adenosine, AMP, and Quinic Acid. .
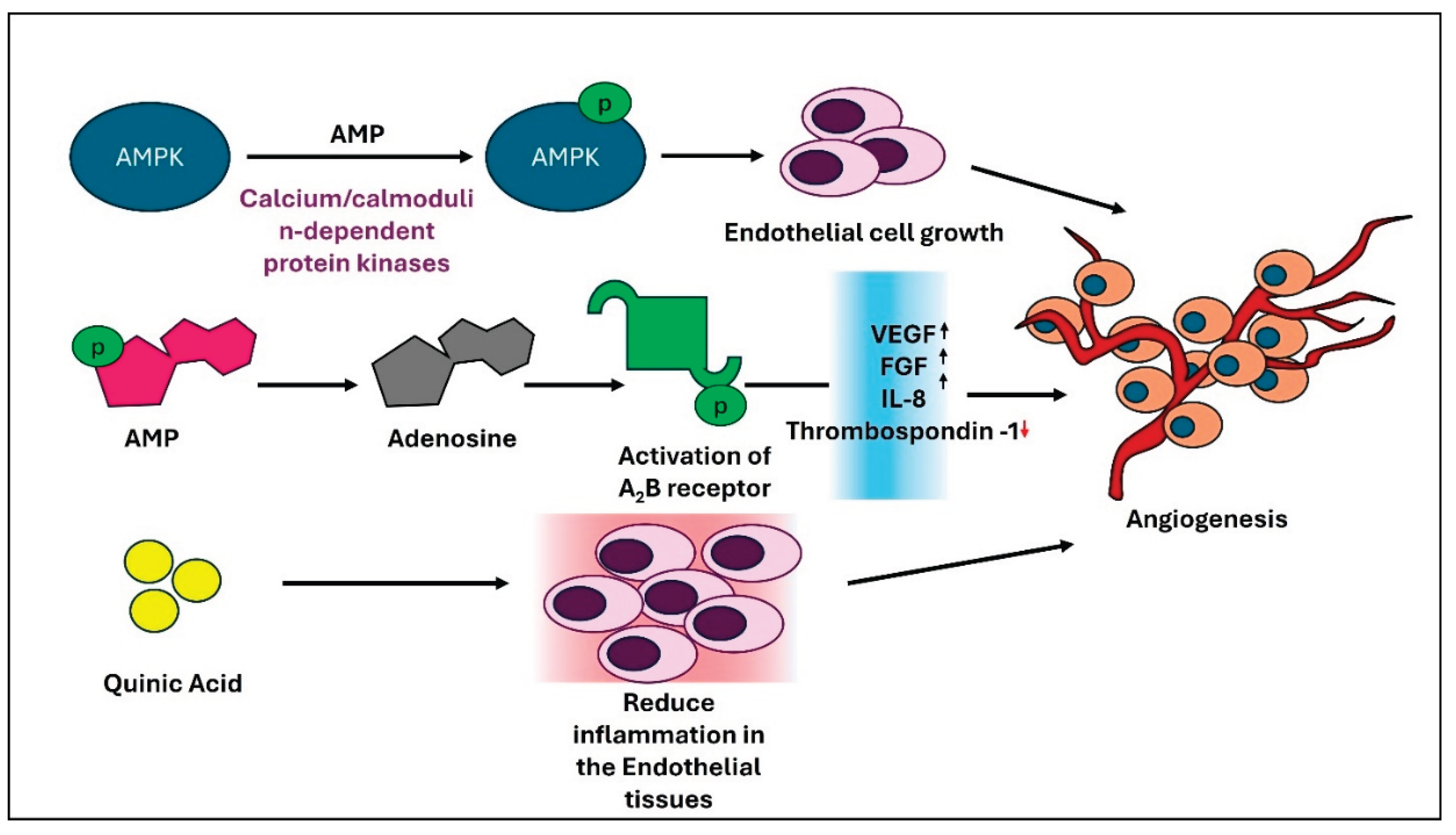
Fruitflow® contains a variety of bioactive compounds, including nucleosides like adenosine and AMP, flavonoids such as quercetin and kaempferol, and phenolic acids like caffeic acid, ferulic acid, and p-coumaric acid [15]. Among these, adenosine significantly promotes angiogenesis by stimulating endothelial cell proliferation and the release of VEGF [124]. AMP activates AMP-activated protein kinase (AMPK), a serine/threonine protein kinase that serves as a sensor of cellular energy status, thereby helping to maintain energy balance and regulate various cellular functions. AMPK is phosphorylated and activated by upstream kinases (AMPKKs), such as calcium/calmodulin-dependent protein kinase-β (CAMKII). CAMKII, activated by intracellular calcium, is expressed only in specific cell types, including endothelial cells. Activation of AMPK in endothelial cells encourages angiogenesis [125]. AMP also indirectly supports angiogenesis by converting to adenosine. Adenosine promotes the release of pro-angiogenic factors, such as interleukin-8 (IL-8), basic fibroblast growth factor (bFGF), and VEGF from endothelial cells through the activation of A2B receptors, which are coupled to Gs and Gq proteins. Simultaneously, adenosine reduces the production of the anti-angiogenic molecule thrombospondin-1, likely via the Gs protein-coupled A2A receptor. Additionally, adenosine promotes the release of VEGF, IL-8, and angiopoietin-1 from mast cells by stimulating A2B and A3 receptors [124,125,126,127]. Lebovich and colleagues identified a different mechanism involving the A2A receptor in macrophages. When activated, this receptor shifts macrophages from a pro-inflammatory state—characterized by high levels of tumor necrosis factor-alpha (TNF-α) and interleukin-6 (IL-6) to a pro-angiogenic phenotype that mainly produces VEGF [124]. This pathway is believed to play an important role in tissue regeneration after injury or infection. Quinic acid (QA), another major compound of Fruitflow®, is also reported to play a key role in reducing inflammation in vascular smooth muscle cells (VSMCs). QA decreases vascular cell adhesion molecule-1 (VCAM-1) expression by inhibiting the MAP kinase and NF-κB signaling pathways in TNF-α-stimulated VSMCs. This reduction in VCAM-1 caused by QA results in less monocyte adhesion to the stimulated VSMCs [128]. Conjugates of caffeic acid, such as caffeic acid phenethyl ester (CAPE), have been shown to induce VEGF production in human dental pulp cells (HDPCs) and also decrease the production of CXCL10. This indicates a positive response to angiogenesis [129]. However, several studies have also documented the anti-angiogenic properties of caffeic acid, especially in cancer environments [130,131]. A recent in vivo study reported that increasing NAD+ production stimulates pro-angiogenic effects, such as VEGF165, and inhibits anti-angiogenic effects[document_number_1b] by regulating the alternative splicing of VEGF. This promotes angiogenesis in diabetic mice with myocardial infarction [132].
In contrast, other constituents of Fruitflow®, such as quercetin and kaempferol, exhibit anti-angiogenic activities mostly in cancerous conditions [133,134]. Other components may have limited or neutral roles in angiogenic activity. Overall, most Fruitflow® constituents regulate angiogenesis through multiple pathways, promoting vascular health and tissue regeneration. While some components enhance pro-angiogenic signaling, others exhibit anti-angiogenic effects, especially in pathological settings. This balanced profile may support vascular integrity during aging, highlighting the potential of Fruitflow® in promoting healthy aging.
5. Epigenetic Regulation of Bioactive Compounds of Fruitflow® in Aging
Losing epigenetic information is a key factor in mammal aging. In yeast, epigenetic information deteriorates over time because chromatin-modifying proteins accumulate at DNA breaks, leading to a loss of cellular identity —a major feature of yeast aging [135]. Reports suggest that efficient DNA repair speeds up physiological, cognitive, and molecular aging. This process involves the deterioration of the epigenetic landscape, cellular dedifferentiation, senescence, and the progression of the DNA methylation clock, which can be reversed through OSK-mediated rejuvenation [136]. A decline in the activity of the protein-folding chaperone network and a loss of intracellular protein balance are essential markers of aging [29]. Aging also involves ongoing changes in cell-to-cell communication, including factors in the blood system that either promote aging or extend lifespan, interactions among different communication pathways between cells, and disruptions in two-way communication between the extracellular matrix and cells [25].
An epigenetic diet can influence gene expression by modifying the epigenome, a chemical tag that attaches to DNA and controls which genes are turned on or off. These diets include foods with bioactive compounds that can alter epigenetic modifications, potentially leading to health benefits and disease prevention. Dietary polyphenols play a significant role in influencing changes, which can help delay aging and increase lifespan. Recent research reviews have explored links between epigenetics, polyphenols, and aging using various dietary polyphenols found in different fruits and vegetables [137]. Evidence suggests that compounds such as apigenin, curcumin, epigallocatechin-3-gallate, genistein, pterostilbene, resveratrol, and quercetin may promote cell growth, reduce tumor incidence, delay tumor development, and modulate multiple signaling pathways [137]. Although the epigenetic changes associated with aging are not yet fully understood, numerous studies have been conducted to elucidate these mechanisms and their relationship to aging.
6. Gut Microbiota, Aging, and Fruitflow®
The gut microbiota plays a role in many physiological processes, including digesting and absorbing nutrients, defending against pathogens, and producing essential metabolites such as vitamins, amino acid derivatives, secondary bile acids, and short-chain fatty acids [138]. It also communicates with the peripheral and central nervous system organs, as well as other distant organs, significantly affecting overall host health [139]. Disrupting this two-way communication between bacteria and the host can lead to various biological disorders, resulting in conditions like obesity, liver disease, type 2 diabetes, ulcerative colitis, neurological diseases, cardiovascular diseases, and cancer [140]. The gut microbiota undergoes significant changes with aging, influencing overall health and potentially contributing to the development of age-related diseases. These changes, often characterized by a decrease in microbial diversity and an increase in potentially harmful bacteria, can affect the immune system, intestinal permeability, and inflammation, thereby accelerating the aging process.
The gut microbiome and its metabolites play a crucial role in the development and progression of liver disease. Various liver disorders, such as nonalcoholic fatty liver disease, alcoholic liver disease, hepatitis C, and hepatocellular carcinoma, are worsened by aging and have poorer prognoses [58]. This occurs when the microbiota and host balance are disrupted, negatively affecting health. Liver disease is associated with both qualitative and quantitative changes, including an increase in harmful bacteria and a decrease in beneficial bacteria, as well as alterations in the overall composition of the bacterial population. Intestinal gut microbiota and their metabolites may contribute to developing chronic liver disease through multiple mechanisms, such as increasing gut permeability, sustaining systemic inflammation, producing SCFA, bile acids, and altering metabolism [138]. As people age, a gradual decline in the ability to maintain homeostasis due to structural changes and gut dysbiosis leads to the progression of end-stage liver disease. The gut microbiome evolves in patients due to changes in pathophysiology during the preoperative phase. A diet rich in plant-based foods, fiber, and fermented products can promote the growth of beneficial bacteria. Conversely, a diet high in processed foods, saturated fats, and certain artificial sweeteners may negatively impact the gut microbiome.
Numerous animal and human studies have shown that the composition of gut microbiota changes with advancing age. In humans, these age-associated alterations in the intestinal microbiota are often linked to negative impacts on health [141,142,143]. In adult humans, the gut microbiota is primarily composed of Firmicutes and Bacteroidetes, with lower abundances of Proteobacteria, Actinobacteria, and Verrucomicrobia. [144]. Research has indicated that an increase in Bacteroidetes and Proteobacteria often accompanies aging, while the levels of Firmicutes and Bifidobacteria tend to decline [141,142,143,144,145]. Recent findings suggest the existence of a distinct core microbiota, primarily comprising the families Ruminococcaceae, Lachnospiraceae, and Bacteroidaceae, whose abundance declines with advancing age [142]. Substantial inter-individual differences and environmental factors likely influence these variations. Interestingly, older individuals tend to harbor a distinct gut microbiota, characterized by greater diversity and an enrichment of beneficial taxa, such as Akkermansia, Bifidobacterium, and Christensenellaceae [142,143]. Although the microbiota of long-lived individuals undergoes age-related alterations, it generally retains high diversity and a stable presence of beneficial microbes that contribute to healthy aging [146]. The gut microbial composition changes with a decline in overall diversity, marked by an increase in potentially proinflammatory species and a reduction in beneficial ones. After four weeks of Fruitflow® supplementation, notable reductions were observed in fasting plasma and urine TMAO levels, along with decreased plasma LPS compared to baseline values [37]. These alterations are significant only for urinary TMAO when comparing between groups. Since plasma TMAO and LPS are considered markers of cardiometabolic risk and low-grade inflammation, the observed outcomes may be regarded as beneficial for host health. Corresponding shifts in microbial beta diversity, but not alpha diversity, were also detected, with a significant difference in Jaccard distance-based Principal Component between groups. Furthermore, Fruitflow® intake influenced microbial composition, showing reductions in Bacteroides, Ruminococcus, and Hungatella, along with increases in Alistipes-related operational taxonomic units, all of which are associated with TMA/TMAO metabolism [147].
7. Conclusion and Future Perspective:
Aging is a gradual process marked by a decline in physiological functions. The biology of aging is complex because many cellular processes are affected. As people age, cells become dysfunctional, their ability to renew and repair tissues decreases, and chronic inflammation rises, making the body more susceptible to diseases. As a result, aging often involves multiple chronic conditions in one person, known as multimorbidity. Aging is actually the primary risk factor for cancer, dementia, and cardiovascular disease, far surpassing other common risk factors currently targeted. The scientific challenge of slowing aging and extending healthy lifespan faces several obstacles, including insufficient regulation, difficulties in clinical validation, challenges in identifying additional biomarkers of human aging, and uncertainties related to introducing new interventions to the market. Fruitflow® carries an European Commission-approved health claim, following EFSA’s positive scientific opinion, for its antiplatelet and cardio-protective effects, and emerges as a promising dietary intervention with broader implications for cellular health and aging. Its bioactive constituents, particularly polyphenols and nucleosides, have demonstrated potential in preserving mitochondrial function, maintaining DNA integrity, promoting autophagy, and mitigating oxidative stress and inflammation. These effects are associated with key aging-related pathways, including DNA methylation, telomere shortening, senescence, and NAD+ metabolism. Moving forward, comprehensive preclinical and clinical investigations are required to elucidate how Fruitflow® modulates the hallmarks of aging and to confirm its long-term benefits in humans. Addressing this gap requires carefully designed studies to validate its efficacy in humans, identify relevant biomarkers, and establish its role within integrative dietary and lifestyle strategies for longevity. Thus, Fruitflow® can be regarded as an emerging candidate in the expanding field of nutrition-based approaches to delay age-related decline and promote a healthy lifespan.
To fully establish Fruitflow®’s role in healthy aging, further preclinical and clinical research is needed to clarify how its individual bioactives contribute to these mechanisms, identify robust biomarkers of efficacy, and evaluate long-term outcomes in humans. Integrating approaches such as multi-omics profiling and biomarker-driven trials will be key to understanding its impact on the hallmarks of aging. With such evidence, Fruitflow® could move beyond its validated role in platelet regulation and emerge as a natural dietary intervention capable of supporting cellular resilience, slowing age-related decline, and promoting health span across multiple organ systems.
Author Contributions
Asim K. Duttaroy wrote the original draft of the manuscript. Surajit Pathak wrote and edited the manuscript. Ruedi Duss critically reviewed the manuscript. Asim K. Duttaroy, Surajit Pathak, Ruedi Duss, Amit Dey, and Keerthi N conducted the wide-ranging aspects of manuscript preparation and pictorial representations. Antara Banerjee, Surajit Pathak, Asim K. Duttaroy, and Ruedi Duss critically reviewed the draft manuscript and provided feedback. All authors read and approved the final manuscript.
Acknowledgments
The authors are thankful to Chettinad Academy of Research and Education (CARE) for providing the infrastructural support to complete this piece of work
Conflicts of Interest
Professor Asim K. Duttaroy is the inventor of Fruitflow® and serves on the scientific advisory board of Provexis PLC, UK. Ruedi Duss is employed by the DSM-Firmenich. All other authors have no conflicts of interest.
Abbreviations
Acute stress-associated phenotype, ASAP; AMP-activated protein kinase, AMPK; European
Food Standard Agency, EFSA; Fibroblast growth factor, bFGF; Intercellular adhesion molecule-1, ICAM-
1/1/CD54; Caffeic acid phenethyl ester, CAPE; Human dental pulp cells, HDPCs; Malondialdehyde, MDA;
Mitochondrial DNA, mtDNA ; Nicotinamide adenine dinucleotide, NAD+; Reactive oxygen species, Quinic acid,
QA; Reactive oxygen species, ROS; Senescence-associated secretory phenotype, SASP: 4-hydroxynonenal, Shortchain
fatty acids, SCFA; High-fat diet, HFD; Tumor necrosis factor-α, TNF-α; von Willebrand factor, vWF;
Vascular endothelium growth factor, VEGF; Vascular smooth muscle cells, VSMCs., Vascular cell adhesion
molecule-1;VCAM-1
References
- Vijg, J.; Dong, X. Pathogenic Mechanisms of Somatic Mutation and Genome Mosaicism in Aging. Cell 2020, 182, 12–23. [CrossRef]
- Navarro Negredo, P.; Yeo, R.W.; Brunet, A. Aging and Rejuvenation of Neural Stem Cells and Their Niches. Cell Stem Cell 2020, 27, 202–223. [CrossRef]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. Hallmarks of Aging: An Expanding Universe. Cell 2023, 186, 243–278. [CrossRef]
- Rudnicka, E.; Napierała, P.; Podfigurna, A.; Męczekalski, B.; Smolarczyk, R.; Grymowicz, M. The World Health Organization (WHO) Approach to Healthy Ageing. Maturitas 2020, 139, 6–11. [CrossRef]
- Kennedy, B.K.; Berger, S.L.; Brunet, A.; Campisi, J.; Cuervo, A.M.; Epel, E.S.; Franceschi, C.; Lithgow, G.J.; Morimoto, R.I.; Pessin, J.E.; et al. Geroscience: Linking Aging to Chronic Disease. Cell 2014, 159, 709–713. [CrossRef]
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The Role of Polyphenols in Human Health and Food Systems: A Mini-Review. Front Nutr 2018, 5. [CrossRef]
- Baur, J.A.; Sinclair, D.A. Therapeutic Potential of Resveratrol: The in Vivo Evidence. Nat Rev Drug Discov 2006, 5, 493–506. [CrossRef]
- Vauzour, D.; Rodriguez-Mateos, A.; Corona, G.; Oruna-Concha, M.J.; Spencer, J.P.E. Polyphenols and Human Health: Prevention of Disease and Mechanisms of Action. Nutrients 2010, 2, 1106–1131. [CrossRef]
- Del Rio, D.; Rodriguez-Mateos, A.; Spencer, J.P.E.; Tognolini, M.; Borges, G.; Crozier, A. Dietary (Poly)Phenolics in Human Health: Structures, Bioavailability, and Evidence of Protective Effects Against Chronic Diseases. Antioxid Redox Signal 2013, 18, 1818–1892. [CrossRef]
- Diwan, B.; Sharma, R. Nutritional Components as Mitigators of Cellular Senescence in Organismal Aging: A Comprehensive Review. Food Sci Biotechnol 2022, 31, 1089–1109. [CrossRef]
- Huang, W.; Hickson, L.J.; Eirin, A.; Kirkland, J.L.; Lerman, L.O. Cellular Senescence: The Good, the Bad and the Unknown. Nat Rev Nephrol 2022, 18, 611–627. [CrossRef]
- Meirow, Y.; Baniyash, M. Immune Biomarkers for Chronic Inflammation Related Complications in Non-Cancerous and Cancerous Diseases. Cancer Immunology, Immunotherapy 2017, 66, 1089–1101. [CrossRef]
- Fekete, M.; Szarvas, Z.; Fazekas-Pongor, V.; Feher, A.; Csipo, T.; Forrai, J.; Dosa, N.; Peterfi, A.; Lehoczki, A.; Tarantini, S.; et al. Nutrition Strategies Promoting Healthy Aging: From Improvement of Cardiovascular and Brain Health to Prevention of Age-Associated Diseases. Nutrients 2022, 15, 47. [CrossRef]
- AlAli, M.; Alqubaisy, M.; Aljaafari, M.N.; AlAli, A.O.; Baqais, L.; Molouki, A.; Abushelaibi, A.; Lai, K.-S.; Lim, S.-H.E. Nutraceuticals: Transformation of Conventional Foods into Health Promoters/Disease Preventers and Safety Considerations. Molecules 2021, 26, 2540. [CrossRef]
- O’Kennedy, N.; Raederstorff, D.; Duttaroy, A.K. Fruitflow®: The First European Food Safety Authority-Approved Natural Cardio-Protective Functional Ingredient. Eur J Nutr 2017, 56, 461–482. [CrossRef]
- Das, R.K.; Datta, T.; Biswas, D.; Duss, R.; O’Kennedy, N.; Duttaroy, A.K. Evaluation of the Equivalence of Different Intakes of Fruitflow in Affecting Platelet Aggregation and Thrombin Generation Capacity in a Randomized, Double-Blinded Pilot Study in Male Subjects. BMC Nutr 2021, 7, 80. [CrossRef]
- Uddin, M.; Biswas, D.; Ghosh, A.; O’Kennedy, N.; Duttaroy, A.K. Consumption of Fruitflow ® Lowers Blood Pressure in Pre-Hypertensive Males: A Randomised, Placebo Controlled, Double Blind, Cross-over Study. Int J Food Sci Nutr 2018, 69, 494–502. [CrossRef]
- Biswas, D.; Uddin, Md.M.; Dizdarevic, L.L.; Jørgensen, A.; Duttaroy, A.K. Inhibition of Angiotensin-Converting Enzyme by Aqueous Extract of Tomato. Eur J Nutr 2014, 53, 1699–1706. [CrossRef]
- Chen, H.; Zhang, S.; Wang, H.; Bao, L.; Wu, W.; Qi, R. Fruitflow Inhibits Platelet Function by Suppressing Akt/GSK3β, Syk/PLCγ2 and P38 MAPK Phosphorylation in Collagen-Stimulated Platelets. BMC Complement Med Ther 2022, 22, 75. [CrossRef]
- Duttaroy, A.K. Functional Foods in Preventing Human Blood Platelet Hyperactivity-Mediated Diseases—An Updated Review. Nutrients 2024, 16, 3717. [CrossRef]
- Somasundaram, I.; Jain, S.M.; Blot-Chabaud, M.; Pathak, S.; Banerjee, A.; Rawat, S.; Sharma, N.R.; Duttaroy, A.K. Mitochondrial Dysfunction and Its Association with Age-Related Disorders. Front Physiol 2024, 15. [CrossRef]
- Tian, Z.; Li, K.; Fan, D.; Gao, X.; Ma, X.; Zhao, Y.; Zhao, D.; Liang, Y.; Ji, Q.; Chen, Y.; et al. Water-Soluble Tomato Concentrate, a Potential Antioxidant Supplement, Can Attenuate Platelet Apoptosis and Oxidative Stress in Healthy Middle-Aged and Elderly Adults: A Randomized, Double-Blinded, Crossover Clinical Trial. Nutrients 2022, 14, 3374. [CrossRef]
- Das, D.; Adhikary, S.; Das, R.K.; Banerjee, A.; Radhakrishnan, A.K.; Paul, S.; Pathak, S.; Duttaroy, A.K. Bioactive Food Components and Their Inhibitory Actions in Multiple Platelet Pathways. J Food Biochem 2022, 46. [CrossRef]
- Keefe, D.L. Telomeres and Genomic Instability during Early Development. Eur J Med Genet 2020, 63, 103638. [CrossRef]
- Hughes, C.E.; Coody, T.K.; Jeong, M.-Y.; Berg, J.A.; Winge, D.R.; Hughes, A.L. Cysteine Toxicity Drives Age-Related Mitochondrial Decline by Altering Iron Homeostasis. Cell 2020, 180, 296-310.e18. [CrossRef]
- Heintz, C.; Doktor, T.K.; Lanjuin, A.; Escoubas, C.C.; Zhang, Y.; Weir, H.J.; Dutta, S.; Silva-García, C.G.; Bruun, G.H.; Morantte, I.; et al. Splicing Factor 1 Modulates Dietary Restriction and TORC1 Pathway Longevity in C. Elegans. Nature 2017, 541, 102–106. [CrossRef]
- Guo, L.; Karpac, J.; Tran, S.L.; Jasper, H. PGRP-SC2 Promotes Gut Immune Homeostasis to Limit Commensal Dysbiosis and Extend Lifespan. Cell 2014, 156, 109–122. [CrossRef]
- Pousa, P.A.; Souza, R.M.; Melo, P.H.M.; Correa, B.H.M.; Mendonça, T.S.C.; Simões-e-Silva, A.C.; Miranda, D.M. Telomere Shortening and Psychiatric Disorders: A Systematic Review. Cells 2021, 10, 1423. [CrossRef]
- Li, Y.; Tian, X.; Luo, J.; Bao, T.; Wang, S.; Wu, X. Molecular Mechanisms of Aging and Anti-Aging Strategies. Cell Commun Signal 2024, 22, 285. [CrossRef]
- Galiè, S.; Canudas, S.; Muralidharan, J.; García-Gavilán, J.; Bulló, M.; Salas-Salvadó, J. Impact of Nutrition on Telomere Health: Systematic Review of Observational Cohort Studies and Randomized Clinical Trials. Advances in Nutrition 2020, 11, 576–601. [CrossRef]
- Kark, J.D.; Goldberger, N.; Kimura, M.; Sinnreich, R.; Aviv, A. Energy Intake and Leukocyte Telomere Length in Young Adults. Am J Clin Nutr 2012, 95, 479–487. [CrossRef]
- Mondal, S.; Jana, J.; Sengupta, P.; Jana, S.; Chatterjee, S. Myricetin Arrests Human Telomeric G-Quadruplex Structure: A New Mechanistic Approach as an Anticancer Agent. Mol Biosyst 2016, 12, 2506–2518. [CrossRef]
- Parekh, N.; Garg, A.; Choudhary, R.; Gupta, M.; Kaur, G.; Ramniwas, S.; Shahwan, M.; Tuli, H.S.; Sethi, G. The Role of Natural Flavonoids as Telomerase Inhibitors in Suppressing Cancer Growth. Pharmaceuticals 2023, 16, 605. [CrossRef]
- Thomas, P.; Wang, Y.-J.; Zhong, J.-H.; Kosaraju, S.; O’Callaghan, N.J.; Zhou, X.-F.; Fenech, M. Grape Seed Polyphenols and Curcumin Reduce Genomic Instability Events in a Transgenic Mouse Model for Alzheimer’s Disease. Mutation Research/Fundamental and Molecular Mechanisms of Mutagenesis 2009, 661, 25–34. [CrossRef]
- Sorrenti, V.; Ali, S.; Mancin, L.; Davinelli, S.; Paoli, A.; Scapagnini, G. Cocoa Polyphenols and Gut Microbiota Interplay: Bioavailability, Prebiotic Effect, and Impact on Human Health. Nutrients 2020, 12, 1908. [CrossRef]
- Gu, Y.; Honig, L.S.; Schupf, N.; Lee, J.H.; Luchsinger, J.A.; Stern, Y.; Scarmeas, N. Mediterranean Diet and Leukocyte Telomere Length in a Multi-Ethnic Elderly Population. Age (Omaha) 2015, 37, 24. [CrossRef]
- Rehman, A.; Tyree, S.M.; Fehlbaum, S.; DunnGalvin, G.; Panagos, C.G.; Guy, B.; Patel, S.; Dinan, T.G.; Duttaroy, A.K.; Duss, R.; et al. A Water-Soluble Tomato Extract Rich in Secondary Plant Metabolites Lowers Trimethylamine-n-Oxide and Modulates Gut Microbiota: A Randomized, Double-Blind, Placebo-Controlled Cross-over Study in Overweight and Obese Adults. J Nutr 2023, 153, 96–105. [CrossRef]
- García-Calzón, S.; Martínez-González, M.A.; Razquin, C.; Arós, F.; Lapetra, J.; Martínez, J.A.; Zalba, G.; Marti, A. Mediterranean Diet and Telomere Length in High Cardiovascular Risk Subjects from the PREDIMED-NAVARRA Study. Clinical Nutrition 2016, 35, 1399–1405. [CrossRef]
- Leung, C.W.; Fung, T.T.; McEvoy, C.T.; Lin, J.; Epel, E.S. Diet Quality Indices and Leukocyte Telomere Length Among Healthy US Adults: Data From the National Health and Nutrition Examination Survey, 1999–2002. Am J Epidemiol 2018, 187, 2192–2201. [CrossRef]
- Hao, L.-Y.; Armanios, M.; Strong, M.A.; Karim, B.; Feldser, D.M.; Huso, D.; Greider, C.W. Short Telomeres, Even in the Presence of Telomerase, Limit Tissue Renewal Capacity. Cell 2005, 123, 1121–1131. [CrossRef]
- Gorbunova, V.; Seluanov, A.; Mita, P.; McKerrow, W.; Fenyö, D.; Boeke, J.D.; Linker, S.B.; Gage, F.H.; Kreiling, J.A.; Petrashen, A.P.; et al. The Role of Retrotransposable Elements in Ageing and Age-Associated Diseases. Nature 2021, 596, 43–53. [CrossRef]
- Coluzzi, E.; Leone, S.; Sgura, A. Oxidative Stress Induces Telomere Dysfunction and Senescence by Replication Fork Arrest. Cells 2019, 8, 19. [CrossRef]
- Tiwari, M.; Parvez, S.; Agrawala, P.K. Role of Some Epigenetic Factors in DNA Damage Response Pathway. AIMS Genet 2017, 04, 069–083. [CrossRef]
- Zheng, G.; Fu, Y.; He, C. Nucleic Acid Oxidation in DNA Damage Repair and Epigenetics. Chem Rev 2014, 114, 4602–4620. [CrossRef]
- Fernandez, A.; O’Leary, C.; O’Byrne, K.J.; Burgess, J.; Richard, D.J.; Suraweera, A. Epigenetic Mechanisms in DNA Double Strand Break Repair: A Clinical Review. Front Mol Biosci 2021, 8. [CrossRef]
- Li, W.; Jones, K.; Burke, T.J.; Hossain, M.A.; Lariscy, L. Epigenetic Regulation of Nucleotide Excision Repair. Front Cell Dev Biol 2022, 10. [CrossRef]
- Mondal, A.; Sarkar, A.; Das, D.; Sengupta, A.; Kabiraj, A.; Mondal, P.; Nag, R.; Mukherjee, S.; Das, C. Epigenetic Orchestration of the DNA Damage Response: Insights into the Regulatory Mechanisms. In; 2024; pp. 99–141.
- Lin, X.; Kapoor, A.; Gu, Y.; Chow, M.J.; Peng, J.; Zhao, K.; Tang, D. Contributions of DNA Damage to Alzheimer’s Disease. Int J Mol Sci 2020, 21, 1666. [CrossRef]
- Shukla, P.C.; Singh, K.K.; Yanagawa, B.; Teoh, H.; Verma, S. DNA Damage Repair and Cardiovascular Diseases. Can J Cardiol 2010, 26 Suppl A, 13A-16A. [CrossRef]
- Rudrapal, M.; Rakshit, G.; Singh, R.P.; Garse, S.; Khan, J.; Chakraborty, S. Dietary Polyphenols: Review on Chemistry/Sources, Bioavailability/Metabolism, Antioxidant Effects, and Their Role in Disease Management. Antioxidants 2024, 13, 429. [CrossRef]
- Rudrapal, M.; Khairnar, S.J.; Khan, J.; Dukhyil, A. Bin; Ansari, M.A.; Alomary, M.N.; Alshabrmi, F.M.; Palai, S.; Deb, P.K.; Devi, R. Dietary Polyphenols and Their Role in Oxidative Stress-Induced Human Diseases: Insights Into Protective Effects, Antioxidant Potentials and Mechanism(s) of Action. Front Pharmacol 2022, 13. [CrossRef]
- Truong, V.-L.; Jeong, W.-S. Cellular Defensive Mechanisms of Tea Polyphenols: Structure-Activity Relationship. Int J Mol Sci 2021, 22, 9109. [CrossRef]
- Fleming, A.M.; Burrows, C.J. On the Irrelevancy of Hydroxyl Radical to DNA Damage from Oxidative Stress and Implications for Epigenetics. Chem Soc Rev 2020, 49, 6524–6528. [CrossRef]
- Ngo, V.; Duennwald, M.L. Nrf2 and Oxidative Stress: A General Overview of Mechanisms and Implications in Human Disease. Antioxidants 2022, 11, 2345. [CrossRef]
- Jomova, K.; Raptova, R.; Alomar, S.Y.; Alwasel, S.H.; Nepovimova, E.; Kuca, K.; Valko, M. Reactive Oxygen Species, Toxicity, Oxidative Stress, and Antioxidants: Chronic Diseases and Aging. Arch Toxicol 2023, 97, 2499–2574. [CrossRef]
- Darband, S.G.; Sadighparvar, S.; Yousefi, B.; Kaviani, M.; Ghaderi-Pakdel, F.; Mihanfar, A.; Rahimi, Y.; Mobaraki, K.; Majidinia, M. Quercetin Attenuated Oxidative DNA Damage through NRF2 Signaling Pathway in Rats with DMH Induced Colon Carcinogenesis. Life Sci 2020, 253, 117584. [CrossRef]
- Weindel, C.G.; Martinez, E.L.; Zhao, X.; Mabry, C.J.; Bell, S.L.; Vail, K.J.; Coleman, A.K.; VanPortfliet, J.J.; Zhao, B.; Wagner, A.R.; et al. Mitochondrial ROS Promotes Susceptibility to Infection via Gasdermin D-Mediated Necroptosis. Cell 2022, 185, 3214-3231.e23. [CrossRef]
- Zhu, D.; Li, X.; Tian, Y. Mitochondrial-to-Nuclear Communication in Aging: An Epigenetic Perspective. Trends Biochem Sci 2022, 47, 645–659. [CrossRef]
- Traba, J.; Satrústegui, J.; del Arco, A. Transport of Adenine Nucleotides in the Mitochondria of Saccharomyces Cerevisiae: Interactions between the ADP/ATP Carriers and the ATP-Mg/Pi Carrier. Mitochondrion 2009, 9, 79–85. [CrossRef]
- Alfatni, A.; Riou, M.; Charles, A.-L.; Meyer, A.; Barnig, C.; Andres, E.; Lejay, A.; Talha, S.; Geny, B. Peripheral Blood Mononuclear Cells and Platelets Mitochondrial Dysfunction, Oxidative Stress, and Circulating MtDNA in Cardiovascular Diseases. J Clin Med 2020, 9, 311. [CrossRef]
- Mottis, A.; Herzig, S.; Auwerx, J. Mitocellular Communication: Shaping Health and Disease. Science (1979) 2019, 366, 827–832. [CrossRef]
- Lima, T.; Li, T.Y.; Mottis, A.; Auwerx, J. Pleiotropic Effects of Mitochondria in Aging. Nat Aging 2022, 2, 199–213. [CrossRef]
- Schumacher, B.; Pothof, J.; Vijg, J.; Hoeijmakers, J.H.J. The Central Role of DNA Damage in the Ageing Process. Nature 2021, 592, 695–703. [CrossRef]
- Furlan, L.; Catala, A. Lipid Modifications of Microsomes Isolated from Villus and Crypt Zones of Bovine Intestinal Mucosa: Relationship with Fatty Acid Binding Protein. Arch Physiol Biochem 1997, 105, 86–91. [CrossRef]
- Belenky, P.; Bogan, K.L.; Brenner, C. NAD+ Metabolism in Health and Disease. Trends Biochem Sci 2007, 32, 12–19. [CrossRef]
- Gerner, R.R.; Klepsch, V.; Macheiner, S.; Arnhard, K.; Adolph, T.E.; Grander, C.; Wieser, V.; Pfister, A.; Moser, P.; Hermann-Kleiter, N.; et al. NAD Metabolism Fuels Human and Mouse Intestinal Inflammation. Gut 2018, 67, 1813–1823. [CrossRef]
- Kyriazis, I.; Vassi, E.; Alvanou, M.; Angelakis, C.; Skaperda, Z.; Tekos, F.; Garikipati, V.; Spandidos, D.; Kouretas, D. The Impact of Diet upon Mitochondrial Physiology (Review). Int J Mol Med 2022, 50, 135. [CrossRef]
- Das, D.; Shruthi, N.R.; Banerjee, A.; Jothimani, G.; Duttaroy, A.K.; Pathak, S. Endothelial Dysfunction, Platelet Hyperactivity, Hypertension, and the Metabolic Syndrome: Molecular Insights and Combating Strategies. Front Nutr 2023, 10. [CrossRef]
- Das, D.; Jothimani, G.; Banerjee, A.; Duttaroy, A.K.; Pathak, S. The Cardioprotective Effects of Fruitflow® against Doxorubicin-Induced Toxicity in Rat Cardiomyoblast Cells H9c2 (2−1) and High-Fat Diet-Induced Dyslipidemia and Pathological Alteration in Cardiac Tissue of Wistar Albino Rats. Biomedicine & Pharmacotherapy 2024, 180, 117607. [CrossRef]
- Degirmenci, U.; Lei, S. Role of LncRNAs in Cellular Aging. Front Endocrinol (Lausanne) 2016, 7. [CrossRef]
- Gasek, N.S.; Kuchel, G.A.; Kirkland, J.L.; Xu, M. Strategies for Targeting Senescent Cells in Human Disease. Nat Aging 2021, 1, 870–879. [CrossRef]
- Schneider, J.L.; Rowe, J.H.; Garcia-de-Alba, C.; Kim, C.F.; Sharpe, A.H.; Haigis, M.C. The Aging Lung: Physiology, Disease, and Immunity. Cell 2021, 184, 1990–2019. [CrossRef]
- Xie, N.; Zhang, L.; Gao, W.; Huang, C.; Huber, P.E.; Zhou, X.; Li, C.; Shen, G.; Zou, B. NAD+ Metabolism: Pathophysiologic Mechanisms and Therapeutic Potential. Signal Transduct Target Ther 2020, 5, 227. [CrossRef]
- Schultz, M.B.; Sinclair, D.A. When Stem Cells Grow Old: Phenotypes and Mechanisms of Stem Cell Aging. Development 2016, 143, 3–14. [CrossRef]
- Bigot, A.; Duddy, W.J.; Ouandaogo, Z.G.; Negroni, E.; Mariot, V.; Ghimbovschi, S.; Harmon, B.; Wielgosik, A.; Loiseau, C.; Devaney, J.; et al. Age-Associated Methylation Suppresses SPRY1 , Leading to a Failure of Re-Quiescence and Loss of the Reserve Stem Cell Pool in Elderly Muscle. Cell Rep 2015, 13, 1172–1182. [CrossRef]
- Lukjanenko, L.; Karaz, S.; Stuelsatz, P.; Gurriaran-Rodriguez, U.; Michaud, J.; Dammone, G.; Sizzano, F.; Mashinchian, O.; Ancel, S.; Migliavacca, E.; et al. Aging Disrupts Muscle Stem Cell Function by Impairing Matricellular WISP1 Secretion from Fibro-Adipogenic Progenitors. Cell Stem Cell 2019, 24, 433-446.e7. [CrossRef]
- Baker, D.J.; Childs, B.G.; Durik, M.; Wijers, M.E.; Sieben, C.J.; Zhong, J.; A. Saltness, R.; Jeganathan, K.B.; Verzosa, G.C.; Pezeshki, A.; et al. Naturally Occurring P16Ink4a-Positive Cells Shorten Healthy Lifespan. Nature 2016, 530, 184–189. [CrossRef]
- Calubag, M.F.; Robbins, P.D.; Lamming, D.W. A Nutrigeroscience Approach: Dietary Macronutrients and Cellular Senescence. Cell Metab 2024, 36, 1914–1944. [CrossRef]
- Della Vedova, L.; Baron, G.; Morazzoni, P.; Aldini, G.; Gado, F. The Potential of Polyphenols in Modulating the Cellular Senescence Process: Implications and Mechanism of Action. Pharmaceuticals 2025, 18, 138. [CrossRef]
- Chen, C.; Zhou, M.; Ge, Y.; Wang, X. SIRT1 and Aging Related Signaling Pathways. Mech Ageing Dev 2020, 187, 111215. [CrossRef]
- Shahidi, F.; Pan, Y. Influence of Food Matrix and Food Processing on the Chemical Interaction and Bioaccessibility of Dietary Phytochemicals: A Review. Crit Rev Food Sci Nutr 2022, 62, 6421–6445. [CrossRef]
- De Luca, F.; Gola, F.; Azzalin, A.; Casali, C.; Gaiaschi, L.; Milanesi, G.; Vicini, R.; Rossi, P.; Bottone, M.G. A Lombard Variety of Sweet Pepper Regulating Senescence and Proliferation: The Voghera Pepper. Nutrients 2024, 16, 1681. [CrossRef]
- Della Vedova, L.; Baron, G.; Morazzoni, P.; Aldini, G.; Gado, F. The Potential of Polyphenols in Modulating the Cellular Senescence Process: Implications and Mechanism of Action. Pharmaceuticals 2025, 18, 138. [CrossRef]
- Liu, H.; Xu, Q.; Wufuer, H.; Li, Z.; Sun, R.; Jiang, Z.; Dou, X.; Fu, Q.; Campisi, J.; Sun, Y. Rutin Is a Potent Senomorphic Agent to Target Senescent Cells and Can Improve Chemotherapeutic Efficacy. Aging Cell 2024, 23. [CrossRef]
- Hada, Y.; Uchida, H.A.; Otaka, N.; Onishi, Y.; Okamoto, S.; Nishiwaki, M.; Takemoto, R.; Takeuchi, H.; Wada, J. The Protective Effect of Chlorogenic Acid on Vascular Senescence via the Nrf2/HO-1 Pathway. Int J Mol Sci 2020, 21, 4527. [CrossRef]
- Zoico, E.; Nori, N.; Darra, E.; Tebon, M.; Rizzatti, V.; Policastro, G.; De Caro, A.; Rossi, A.P.; Fantin, F.; Zamboni, M. Senolytic Effects of Quercetin in an in Vitro Model of Pre-Adipocytes and Adipocytes Induced Senescence. Sci Rep 2021, 11, 23237. [CrossRef]
- Luo, G.; Xiang, L.; Xiao, L. Quercetin Alleviates Atherosclerosis by Suppressing Oxidized LDL-Induced Senescence in Plaque Macrophage via Inhibiting the P38MAPK/P16 Pathway. J Nutr Biochem 2023, 116, 109314. [CrossRef]
- Ragonnaud, E.; Biragyn, A. Gut Microbiota as the Key Controllers of “Healthy” Aging of Elderly People. Immunity & Ageing 2021, 18, 2. [CrossRef]
- Hoyles, L.; Jiménez-Pranteda, M.L.; Chilloux, J.; Brial, F.; Myridakis, A.; Aranias, T.; Magnan, C.; Gibson, G.R.; Sanderson, J.D.; Nicholson, J.K.; et al. Metabolic Retroconversion of Trimethylamine N-Oxide and the Gut Microbiota. Microbiome 2018, 6, 73. [CrossRef]
- Viola, J.; Soehnlein, O. Atherosclerosis – A Matter of Unresolved Inflammation. Semin Immunol 2015, 27, 184–193. [CrossRef]
- Landi, A.; Schoenhuber, R.; Funicello, R.; Rasio, G.; Esposito, M. Compartment Syndrome of the Scapula. Definition on Clinical, Neurophysiological and Magnetic Resonance Data. Ann Chir Main Memb Super 1992, 11, 383–388. [CrossRef]
- Raiola, A.; Rigano, M.M.; Calafiore, R.; Frusciante, L.; Barone, A. Enhancing the Health-Promoting Effects of Tomato Fruit for Biofortified Food. Mediators Inflamm 2014, 2014, 1–16. [CrossRef]
- Rayes, J.; Jenne, C.N. Platelets: Bridging Thrombosis and Inflammation. Platelets 2021, 32, 293–294. [CrossRef]
- Lievens, D.; Hundelshausen, P. von Platelets in Atherosclerosis. Thromb Haemost 2011, 106, 827–838. [CrossRef]
- Zhang, Y.; Wernly, B.; Cao, X.; Mustafa, S.J.; Tang, Y.; Zhou, Z. Adenosine and Adenosine Receptor-Mediated Action in Coronary Microcirculation. Basic Res Cardiol 2021, 116, 22. [CrossRef]
- Porkka-Heiskanen, T.; Kalinchuk, A. V. Adenosine, Energy Metabolism and Sleep Homeostasis. Sleep Med Rev 2011, 15, 123–135. [CrossRef]
- Meininger, C.J.; Schelling, M.E.; Granger, H.J. Adenosine and Hypoxia Stimulate Proliferation and Migration of Endothelial Cells. American Journal of Physiology-Heart and Circulatory Physiology 1988, 255, H554–H562. [CrossRef]
- Ugusman, A.; Zakaria, Z.; Chua, K.H.; Megat Mohd Nordin, N.A.; Abdullah Mahdy, Z. Role of Rutin on Nitric Oxide Synthesis in Human Umbilical Vein Endothelial Cells. The Scientific World Journal 2014, 2014, 1–9. [CrossRef]
- Oliveira, E.G. de L.; de Oliveira, H.P. β-Cyclodextrin-Silver Nanoparticles Inclusion Complexes: Insights into Applications in Trace Level Detection (Light-Driven and Electrochemical Assays) and Antibacterial Activity. ACS Omega 2025, 10, 23943–23956. [CrossRef]
- Satari, A.; Ghasemi, S.; Habtemariam, S.; Asgharian, S.; Lorigooini, Z. Rutin: A Flavonoid as an Effective Sensitizer for Anticancer Therapy; Insights into Multifaceted Mechanisms and Applicability for Combination Therapy. Evidence-Based Complementary and Alternative Medicine 2021, 2021, 1–10. [CrossRef]
- Nouri, Z.; Fakhri, S.; Nouri, K.; Wallace, C.E.; Farzaei, M.H.; Bishayee, A. Targeting Multiple Signaling Pathways in Cancer: The Rutin Therapeutic Approach. Cancers (Basel) 2020, 12, 2276. [CrossRef]
- Zong, M.; Ji, J.; Wang, Q.; Cai, Y.; Chen, L.; Zhang, L.; Hou, W.; Li, X.; Kong, Q.; Zheng, C.; et al. Chlorogenic Acid Promotes Fatty Acid Beta-Oxidation to Increase HESCs Proliferation and Lipid Synthesis. Sci Rep 2025, 15, 7095. [CrossRef]
- Dai, C.; Cheng, J.; Li, H.; Zhao, W.; Li, B.; Fu, Y.; Wang, X.; Liao, C.; Chen, Y.; Yan, J. Chlorogenic Acid Alleviates Deoxynivalenol-Induced Damage in Porcine Trophectoderm Cells by Regulating the PI3K/AKT Signaling Pathway. Biol Reprod 2025, 113, 466–477. [CrossRef]
- Navarrete, S.; Alarcón, M.; Palomo, I. Aqueous Extract of Tomato (Solanum Lycopersicum L.) and Ferulic Acid Reduce the Expression of TNF-α and IL-1β in LPS-Activated Macrophages. Molecules 2015, 20, 15319–15329. [CrossRef]
- Schwager, J.; Richard, N.; Mussler, B.; Raederstorff, D. Tomato Aqueous Extract Modulates the Inflammatory Profile of Immune Cells and Endothelial Cells. Molecules 2016, 21, 168. [CrossRef]
- Rayes, J.; Jenne, C.N. Platelets: Bridging Thrombosis and Inflammation. Platelets 2021, 32, 293–294. [CrossRef]
- Lievens, D.; Hundelshausen, P. von Platelets in Atherosclerosis. Thromb Haemost 2011, 106, 827–838. [CrossRef]
- Fuentes, F.; Palomo, I.; Fuentes, E. Platelet Oxidative Stress as a Novel Target of Cardiovascular Risk in Frail Older People. Vascul Pharmacol 2017, 93–95, 14–19. [CrossRef]
- Jang, J.Y.; Min, J.H.; Chae, Y.H.; Baek, J.Y.; Wang, S. Bin; Park, S.J.; Oh, G.T.; Lee, S.-H.; Ho, Y.-S.; Chang, T.-S. Reactive Oxygen Species Play a Critical Role in Collagen-Induced Platelet Activation via SHP-2 Oxidation. Antioxid Redox Signal 2014, 20, 2528–2540. [CrossRef]
- Pereira, Q.C.; dos Santos, T.W.; Fortunato, I.M.; Ribeiro, M.L. The Molecular Mechanism of Polyphenols in the Regulation of Ageing Hallmarks. Int J Mol Sci 2023, 24, 5508. [CrossRef]
- Liu, Y.; Fang, M.; Tu, X.; Mo, X.; Zhang, L.; Yang, B.; Wang, F.; Kim, Y.-B.; Huang, C.; Chen, L.; et al. Dietary Polyphenols as Anti-Aging Agents: Targeting the Hallmarks of Aging. Nutrients 2024, 16, 3305. [CrossRef]
- Zhong, J.; Fang, J.; Wang, Y.; Lin, P.; Wan, B.; Wang, M.; Deng, L.; Tang, X. Dietary Flavonoid Intake Is Negatively Associated with Accelerating Aging: An American Population-Based Cross-Sectional Study. Nutr J 2024, 23, 158. [CrossRef]
- D’Angelo, S. Diet and Aging: The Role of Polyphenol-Rich Diets in Slow Down the Shortening of Telomeres: A Review. Antioxidants 2023, 12, 2086. [CrossRef]
- Nair, R.R.; Madiwale, S. V.; Saini, D.K. Clampdown of Inflammation in Aging and Anticancer Therapies by Limiting Upregulation and Activation of GPCR, CXCR4. NPJ Aging Mech Dis 2018, 4, 9. [CrossRef]
- Pallauf, K.; Rimbach, G. Autophagy, Polyphenols and Healthy Ageing. Ageing Res Rev 2013, 12, 237–252. [CrossRef]
- Wedel, S.; Manola, M.; Cavinato, M.; Trougakos, I.P.; Jansen-Dürr, P. Targeting Protein Quality Control Mechanisms by Natural Products to Promote Healthy Ageing. Molecules 2018, 23, 1219. [CrossRef]
- Xu, W.; Luo, Y.; Yin, J.; Huang, M.; Luo, F. Targeting AMPK Signaling by Polyphenols: A Novel Strategy for Tackling Aging. Food Funct 2023, 14, 56–73. [CrossRef]
- Sandoval-Acuña, C.; Ferreira, J.; Speisky, H. Polyphenols and Mitochondria: An Update on Their Increasingly Emerging ROS-Scavenging Independent Actions. Arch Biochem Biophys 2014, 559, 75–90. [CrossRef]
- Johnson, K.E.; Wilgus, T.A. Vascular Endothelial Growth Factor and Angiogenesis in the Regulation of Cutaneous Wound Repair. Adv Wound Care (New Rochelle) 2014, 3, 647–661. [CrossRef]
- Lee, C.; Kim, M.-J.; Kumar, A.; Lee, H.-W.; Yang, Y.; Kim, Y. Vascular Endothelial Growth Factor Signaling in Health and Disease: From Molecular Mechanisms to Therapeutic Perspectives. Signal Transduct Target Ther 2025, 10, 170. [CrossRef]
- Callahan, C.M.; Apostolova, L.G.; Gao, S.; Risacher, S.L.; Case, J.; Saykin, A.J.; Lane, K.A.; Swinford, C.G.; Yoder, M.C. Novel Markers of Angiogenesis in the Setting of Cognitive Impairment and Dementia. Journal of Alzheimer’s Disease 2020, 75, 959–969. [CrossRef]
- Damluji, A.A.; Alfaraidhy, M.; AlHajri, N.; Rohant, N.N.; Kumar, M.; Al Malouf, C.; Bahrainy, S.; Ji Kwak, M.; Batchelor, W.B.; Forman, D.E.; et al. Sarcopenia and Cardiovascular Diseases. Circulation 2023, 147, 1534–1553. [CrossRef]
- Ionescu, C.; Oprea, B.; Ciobanu, G.; Georgescu, M.; Bică, R.; Mateescu, G.-O.; Huseynova, F.; Barragan-Montero, V. The Angiogenic Balance and Its Implications in Cancer and Cardiovascular Diseases: An Overview. Medicina (B Aires) 2022, 58, 903. [CrossRef]
- Auchampach, J.A. Adenosine Receptors and Angiogenesis. Circ Res 2007, 101, 1075–1077. [CrossRef]
- Teng, R.-J.; Du, J.; Afolayan, A.J.; Eis, A.; Shi, Y.; Konduri, G.G. AMP Kinase Activation Improves Angiogenesis in Pulmonary Artery Endothelial Cells with in Utero Pulmonary Hypertension. American Journal of Physiology-Lung Cellular and Molecular Physiology 2013, 304, L29–L42. [CrossRef]
- Feoktistov, I.; Goldstein, A.E.; Ryzhov, S.; Zeng, D.; Belardinelli, L.; Voyno-Yasenetskaya, T.; Biaggioni, I. Differential Expression of Adenosine Receptors in Human Endothelial Cells. Circ Res 2002, 90, 531–538. [CrossRef]
- Clark, A.N.; Youkey, R.; Liu, X.; Jia, L.; Blatt, R.; Day, Y.-J.; Sullivan, G.W.; Linden, J.; Tucker, A.L. A 1 Adenosine Receptor Activation Promotes Angiogenesis and Release of VEGF From Monocytes. Circ Res 2007, 101, 1130–1138. [CrossRef]
- Jang, S.-A.; Park, D.W.; Kwon, J.E.; Song, H.S.; Park, B.; Jeon, H.; Sohn, E.-H.; Koo, H.J.; Kang, S.C. Quinic Acid Inhibits Vascular Inflammation in TNF-α-Stimulated Vascular Smooth Muscle Cells. Biomedicine & Pharmacotherapy 2017, 96, 563–571. [CrossRef]
- Kuramoto, H.; Nakanishi, T.; Takegawa, D.; Mieda, K.; Hosaka, K. Caffeic Acid Phenethyl Ester Induces Vascular Endothelial Growth Factor Production and Inhibits CXCL10 Production in Human Dental Pulp Cells. Curr Issues Mol Biol 2022, 44, 5691–5699. [CrossRef]
- Kim, J.H.; Lee, B.J.; Kim, J.H.; Yu, Y.S.; Kim, K.-W. Anti-Angiogenic Effect of Caffeic Acid on Retinal Neovascularization. Vascul Pharmacol 2009, 51, 262–267. [CrossRef]
- Alam, M.; Ashraf, G.M.; Sheikh, K.; Khan, A.; Ali, S.; Ansari, Md.M.; Adnan, M.; Pasupuleti, V.R.; Hassan, Md.I. Potential Therapeutic Implications of Caffeic Acid in Cancer Signaling: Past, Present, and Future. Front Pharmacol 2022, 13. [CrossRef]
- Jiao, L.; Gong, M.; Yang, X.; Li, M.; Shao, Y.; Wang, Y.; Li, H.; Yu, Q.; Sun, L.; Xuan, L.; et al. NAD+ Attenuates Cardiac Injury after Myocardial Infarction in Diabetic Mice through Regulating Alternative Splicing of VEGF in Macrophages. Vascul Pharmacol 2022, 147, 107126. [CrossRef]
- Tan, W.; Lin, L.; Li, M.; Zhang, Y.-X.; Tong, Y.; Xiao, D.; Ding, J. Quercetin, a Dietary-Derived Flavonoid, Possesses Antiangiogenic Potential. Eur J Pharmacol 2003, 459, 255–262. [CrossRef]
- Luo, H.; Rankin, G.O.; Liu, L.; Daddysman, M.K.; Jiang, B.-H.; Chen, Y.C. Kaempferol Inhibits Angiogenesis and VEGF Expression Through Both HIF Dependent and Independent Pathways in Human Ovarian Cancer Cells. Nutr Cancer 2009, 61, 554–563. [CrossRef]
- Zhou, Z.; Liu, Y.; Feng, Y.; Klepin, S.; Tsimring, L.S.; Pillus, L.; Hasty, J.; Hao, N. Engineering Longevity—Design of a Synthetic Gene Oscillator to Slow Cellular Aging. Science (1979) 2023, 380, 376–381. [CrossRef]
- Yang, J.-H.; Hayano, M.; Griffin, P.T.; Amorim, J.A.; Bonkowski, M.S.; Apostolides, J.K.; Salfati, E.L.; Blanchette, M.; Munding, E.M.; Bhakta, M.; et al. Loss of Epigenetic Information as a Cause of Mammalian Aging. Cell 2023, 186, 305-326.e27. [CrossRef]
- Arora, I.; Sharma, M.; Sun, L.Y.; Tollefsbol, T.O. The Epigenetic Link between Polyphenols, Aging and Age-Related Diseases. Genes (Basel) 2020, 11, 1094. [CrossRef]
- Ghosh, T.S.; Shanahan, F.; O’Toole, P.W. The Gut Microbiome as a Modulator of Healthy Ageing. Nat Rev Gastroenterol Hepatol 2022, 19, 565–584. [CrossRef]
- Mallick, R.; Basak, S.; Das, R.K.; Banerjee, A.; Paul, S.; Pathak, S.; Duttaroy, A.K. Roles of the Gut Microbiota in Human Neurodevelopment and Adult Brain Disorders. Front Neurosci 2024, 18. [CrossRef]
- Adhikary, S.; Esmeeta, A.; Dey, A.; Banerjee, A.; Saha, B.; Gopan, P.; Duttaroy, A.K.; Pathak, S. Impacts of Gut Microbiota Alteration on Age-Related Chronic Liver Diseases. Digestive and Liver Disease 2024, 56, 112–122. [CrossRef]
- Odamaki, T.; Kato, K.; Sugahara, H.; Hashikura, N.; Takahashi, S.; Xiao, J.-Z.; Abe, F.; Osawa, R. Age-Related Changes in Gut Microbiota Composition from Newborn to Centenarian: A Cross-Sectional Study. BMC Microbiol 2016, 16, 90. [CrossRef]
- Biagi, E.; Franceschi, C.; Rampelli, S.; Severgnini, M.; Ostan, R.; Turroni, S.; Consolandi, C.; Quercia, S.; Scurti, M.; Monti, D.; et al. Gut Microbiota and Extreme Longevity. Current Biology 2016, 26, 1480–1485. [CrossRef]
- Kong, F.; Hua, Y.; Zeng, B.; Ning, R.; Li, Y.; Zhao, J. Gut Microbiota Signatures of Longevity. Current Biology 2016, 26, R832–R833. [CrossRef]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D.A. Diversity of the Human Intestinal Microbial Flora. Science (1979) 2005, 308, 1635–1638. [CrossRef]
- Claesson, M.J.; Cusack, S.; O’Sullivan, O.; Greene-Diniz, R.; de Weerd, H.; Flannery, E.; Marchesi, J.R.; Falush, D.; Dinan, T.; Fitzgerald, G.; et al. Composition, Variability, and Temporal Stability of the Intestinal Microbiota of the Elderly. Proceedings of the National Academy of Sciences 2011, 108, 4586–4591. [CrossRef]
- Ragonnaud, E.; Biragyn, A. Gut Microbiota as the Key Controllers of “Healthy” Aging of Elderly People. Immunity & Ageing 2021, 18, 2. [CrossRef]
- Hoyles, L.; Jiménez-Pranteda, M.L.; Chilloux, J.; Brial, F.; Myridakis, A.; Aranias, T.; Magnan, C.; Gibson, G.R.; Sanderson, J.D.; Nicholson, J.K.; et al. Metabolic Retroconversion of Trimethylamine N-Oxide and the Gut Microbiota. Microbiome 2018, 6, 73. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



