
Submitted:
02 September 2025
Posted:
04 September 2025
You are already at the latest version
Abstract
Pediatric tumors such as neuroblastoma are characterized by a genome-wide ‘transcrip-tional burden’ surmising the involvement of multiple alterations of gene expression. Search for master regulators of transcription whose inactivation is lethal for tumor cells identified the non-POU domain-containing octamer-binding protein (NONO), a member of the Drosophila Behavior/Human Splicing family known for the ability to form com-plexes with macromolecules. NONO emerges as an essential mechanism in normal neu-rogenesis as well as in tumor biology. In particular, NONO interactions with RNAs, largely with long non-coding MYCN transcripts, have been attributed to the aggressive-ness of neuroblastoma. Broadening its significance beyond MYCN regulation, NONO guards a subset of transcription factors that comprise a core regulatory circuit, a self-sustained loop that maintains transcription. Furthermore, as a component of pro-tein-protein complexes, NONO has been implicated in the control of cell cycle progres-sion, double strand DNA repair and, generally, cell survival. Altogether, the pro-oncogenic roles of NONO justify the need for its inactivation as a therapeutic strategy. However, con-sidering NONO as a therapeutic target, its druggability is a challenge. Recent advances in inactivation of NONO and downstream signaling with small molecular weight com-pounds make promising the development of pharmacological antagonists of NONO pathway(s) for neuroblastoma treatment.
Keywords:
gene transcription
; NONO
; MYCN
; lncRNAs
; therapeutic target
; neuroblastoma
; cancer
1. Introduction
In the growing list of molecular mechanisms critical in various aspects of cancer biology, gene transcription attracts increasing attention. The term ‘transcriptional burden’ coined by Comitani et al. as an alternative to the ‘mutational burden’ [1], highlights the significance of epigenetic regulation in specific malignancies, most importantly, in pediatric cancers. Along with a great amount of data on the pathogenetic role of transcriptional events in carcinogenesis, this mechanism is being considered a therapeutic target. Indeed, it is plausible to hypothesize that interference with this vital mechanism would be lethal for tumor cells.
Neuroblastoma, a malignancy that originates from embryonic neural crest cells, is one of the most common solid tumors in early childhood [2]. This aggressive cancer exhibits a broad spectrum of clinical manifestations, ranging from spontaneous regression to relentless progression despite multimodal therapies that include surgery, radiation, and high-dose chemotherapy [3,4]. Approximately 50% of cases are classified as high-risk with a 5-year survival rate below 50% [5], underscoring the urgent need for novel therapeutic approaches.
Biological heterogeneity, a hallmark of neuroblastoma, is driven by epigenetic and, to a lesser extent, genetic alterations. Amplification of the MYCN oncogene is an established marker of neuroblastoma aggressiveness [6,7]. Among other genetic factors are the activating mutations in the ALK (anaplastic lymphoma kinase) gene that drive proliferation and the hemizygous deletion of the 11q chromosome associated with the loss of tumor suppressor genes [8,9,10].
More complex and pathogenetically significant are the genome-wide changes of the transcriptional landscape. Two major subtypes of neuroblastoma differ in the arrays of individual transcriptional mechanisms. The adrenergic (ADRN) subtype is characterized by MYCN amplification along with deregulation of a set of genes mechanistically linked to MYCN network, including paired-like homeobox 2 (PHOX2), heart- and neural crest derivatives-expressed protein 2 (HAND2), GATA binding protein 3 (GATA3), insulin enhancer binding protein 1 (ISL1), T-box transcription factor 2 (TBX2) and Achaete-scute family bHLH transcription factor 1 (ASCL1) [11,12,13]. The transcription factors encoded by these genes comprise a core regulatory circuit (CRC), a self-sustainable loop that maintains transcription of specific genes [10,13]. As a result of constitutively active transcription of CRC-dependent genes, the ADRN subtype is known for its aggressiveness. Unlike ADRN, the mesenchymal (MES) subtype contains CD133-positive cells that overexpress Yes-associated protein/transcriptional coactivator with PDZ binding motif (YAP/TAZ), paired related homeobox 1 (PRRX1), and Snail family transcriptional repressor 1 (SNAI1) [9,14,15]. Resistance to chemotherapy is a characteristic trait of this immature (stem-like) phenotype.
Amid the intricate transcriptional landscape, the non-POU domain-containing octamer-binding protein (NONO) emerges as a pivotal regulator. This multifunctional protein orchestrates gene expression by interacting with long non-coding RNAs (lncRNAs, transcripts >200 bp) and transcription factors. Studies in normal cells and in tumor models including neuroblastoma position NONO as a bridge between genetic alterations and transcriptional reprogramming [16]. In neurogenesis, NONO overexpression during embryonic cortex development disrupts radial neuronal migration by impairing multipolar-to-bipolar transition — reducing uni/bipolar neurons and disorienting the leading processes relative to radial glia. This leads to persistent postnatal disorders such as shortened processes, simplified dendritic arborization, and Golgi misorientation. These findings implicate dysregulated NONO in neurodevelopmental defects involving neuronal mispositioning [17]. Since neuroblastoma is a transcription-dependent malignancy, one may hypothesize that NONO plays a substantial role in its pathogenesis. If so, can NONO be therapeutically druggable, alone and/or in combination with conventional agents?
2. NONO in Transcriptional Regulation
2.1. Physiological States
NONO belongs to the Drosophila behavior/human splicing (DBHS) family that also includes the paraspeckle component 1 (PSPC1) and the splicing factor, proline- and glutamine-rich (SFPQ) protein [18]. The NONO protein consists of four distinct regions: the RNA recognition motifs 1 and 2 (RRM1-2), the NONO/paraspeckle (NOPS) and the C-terminal coiled-coil domain. RRM1 and -2 are responsible for binding to RNAs, a function critically important for the role of NONO in RNA splicing and posttranscriptional regulation. These domains can dimerize upon protein-RNA complex formation [19]. The NOPS domain, being unique for DBHS family, is necessary for the formation of paraspeckles, the membraneless dynamic organelles that control nuclear RNA retention and functions (reviewed in [20]). This domain allows for SFPQ-NONO heterodimerization [21]. The spiral domain at the C-terminus also participates in di- or oligomerization. Both N- and C-terminal regions are involved in phase separation to form condensates that concentrate the reaction components, thereby optimizing the conditions for their efficient interaction [22,23,24].
NONO, being part of macromolecular (i.e., protein-protein or protein-nucleic acids) complexes, orchestrates major cellular processes, namely, maintaining genome stability and gene expression landscape [18]. Structural data strongly suggest that NONO acts not as much as a single molecule. Two (perhaps, not mutually excluding) modes of transcriptional control by NONO imply 1) formation of complexes with RNA, and 2) interactions with the transcription factors at the regulatory regions of target genes. First, NONO homodimers preferentially interact with RNAs in which adenosine is changed for inosine (A-to-I editing). This modification occurs in mRNAs, miRNAs or non-coding transcripts. A-to-I editing can lead to interference with RNA splicing resulting in nuclear accumulation of mRNAs [25,26]. The NONO/SFPQ/mat3 complex specifically binds to I-edited RNA thereby entrapping it in the nuclei [27]. A pseudo-symmetric structure of NONO/PSPC1 allows for the heterodimer’s interaction with lncRNAs as well as with hyper-edited RNA sequences [19]. LncRNAs are especially important in neuroblastoma being targets for MYCN oncoprotein (see section Relevance to neuroblastoma).
Second, NONO interacts directly with transcription factors to regulate developmental and cell cycle related genes. In mouse embryonic stem cells, NONO binds Pax9, Tbx3, Cdx2 and Gata4 to control developmental loci. Besides transcription factors, NONO recruits and cooperates with chromatin modifiers. It physically associates with TET1 - sharing up to 85% of ChIP peaks - to guide TET1 to neurogenic gene promoters. NONO knockdown displaced TET1, elevated 5-hydroxymethylcytosine at Bmp1, Fgf8 and Wnt3a genes, and down-regulated their expression [28]. Furthermore, NONO co-occupied chromatin with Erk2 and phosphoSer5 RNA polymerase II; Erk2 knockdown abolished NONO-chromatin association [29]. The role in chromatin remodeling is further supported by interaction of NONO with the acetyltransferase CBP/p300 [30]. Moreover, NONO partners with TAZ to activate mechanoresponsive genes: following cell adhesion, NONO–TAZ complexes peak at 2–4 h, coinciding with TAZ dephosphorylation and driving CTGF/CYR61 expression. Over 80% of TAZ and ~50% of NONO ChIP-seq peaks are co-localized to active enhancers containing H3K4me1+, H3K4me3- or H3K27Ac+ epigenetic marks. Loss of NONO significantly reduced TAZ distribution across the enhancer regions [31].
Studies in primary mouse fibroblasts showed that, while circadian rhythms remained intact in NONO-deficient cells (via gene trap technology - NONO gt), the circadian control of G1 exit was lost: cells proliferated faster, p16Ink4A expression decreased, senescence markers were reduced, and the circadian gating of S-phase entry was suppressed. At the molecular level, NONO interacts with circadian PERIOD (Per1/Per2) proteins co-activating the p16Ink4A gene and enforcing temporal segregation of the cell cycle. Deletion of either NONO or PERIOD disrupted the rhythmic expression of p16Ink4A and abolished circadian gating of cell division. The absence of NONO resulted in wound healing defects similar to those observed in mice with disrupted circadian mechanisms [32].
Intriguingly, physical interaction of NONO with protein partners does not necessarily presume transcriptional activation. Dong et al. [33]. demonstrated that, in rat myometrium cells, NONO forms complexes with progesterone receptors via direct binding independently of the hormone. These complexes are recruited to the connexin 43 (Gja1) promoter to attenuate this gene. Accordingly, siRNA-mediated NONO knockdown elevated Gja1 transcripts 2.5-fold. Transcriptomic analysis further substantiated the significance of NONO as a transcriptional repressor of 150 genes associated with myometrial contractility. This study places NONO into the regulatory network that controls the timing of labor.
Together, the ability to interact with nucleic acids and proteins makes NONO a versatile regulator of gene expression in a variety of physiological situations. It is plausible to expect that this role is translated to (or reflected in) tumor biology.
2.2. Roles in Cancer
As in non-malignant cells, functions of NONO in cancer are implemented via RNA binding as well as protein-protein complex formation. NONO has been implicated in progression of colorectal cancer due to its interaction with GAPLINC (gastric adenocarcinoma associated, positive CD44 regulator, long intergenic non-coding RNA). Knockdown of NONO attenuated GAPLINC-activated invasion of HCT116 colon carcinoma cells in Transwell assays. Global analysis of gene expression profiles showed that NONO knockdown decreased the expression of 1,776 genes that overlapped with GAPLINC-induced SNAI2, LAMB3 and IGFBP7 genes whose functions are linked to metastasis [34]. Similarly, Cheng et al. showed that increased NONO levels correlated with the depth of invasion in esophageal squamous cell carcinoma (ESCC); NONO played a critical role in suppressing apoptosis through activation of Akt and Erk1/2 signaling pathways. NONO silencing by siRNA inhibited ESCC cell proliferation and invasion while inducing apoptosis, indicating its key contribution to ESCC tumorigenesis [35].
In hepatocellular carcinoma, NONO interacts with lncRNA DIO3OS, an anti-oncogenic transcript down-regulated in cancers. DIO3OS implements its suppressive role through down-regulation of zinc finger E-box binding homeobox 1 protein (ZEB1), a factor of tumor cell stemness. Binding of DIO3OS to NONO restricts nuclear export of ZEB1 mRNA. Therefore, the DIO3OS-NONO-ZEB1 axis emerges as a mechanism of control of malignant potential in hepatocellular carcinoma [36].
Functions of NONO as an RNA binder and a component of protein-protein complexes are not mutually exclusive but rather act in concert. In U2OS sarcoma cells, DNA damage triggered relocation of NONO from gene promoters to the nucleoli, an effect relevant to the attenuation of de novo pre-mRNA synthesis for the sake of rescuing the existing transcripts from degradation. In untreated cells NONO formed stable complexes with RNA polymerase II; these complexes were largely disrupted in response to topoisomerase II blocker etoposide. NONO inhibition led to an increase of γH2AX foci and acetylated histone (H2BK120ac) marks on the transcription start sites of actively expressing genes. NONO recruited X-ray repair cross-complementing protein 4 (XRCC4) for double strand DNA repair using a non-homologous end joining; NONO deficiency prevented the repair [37].
The response of NONO to genotoxic stress was further dissected by Li et al. The authors subjected mouse males to whole body ionizing radiation and found that NONO levels in testicles were elevated severalfold by 24-48 h post exposure to 4 Gy. NONO deficiency enhanced radiation-induced apoptosis in Sertoli cells, supporting the positive role of NONO in double strand DNA repair and rescue of testicular cells from genotoxic stress. Interestingly, SFPQ tended to compensate for NONO inactivation because irradiation of gt/0 males, but not wild type counterparts, led to an increase of SFPQ [38]. Furthermore, NONO silencing resulted in defective intra-S-phase checkpoint activation in UV-irradiated HeLa and melanoma cells evidenced by continued DNA synthesis, failure to block origin firing, impaired CHK1S345 phosphorylation, and compromised recruitment of key checkpoint proteins. NONO re-expression rescued checkpoint function [39]. These findings are in line with other reports that highlighted different modes of NONO participation in DNA damage repair [40,41].
In breast cancer cells, NONO acts as a component of chromatin-remodeling machinery in complexes with EGFR и STAT3. Accordingly, the pro-proliferative and pro-survival genes are activated: cyclin D1, Aurora A, B-myb, c-Myc (EGFR targets) and CCNB1 and CCND1 (STAT3 targets) [30,42]. Interaction of NONO with sterol regulatory element-binding proteins SREBP-1a and SREBP-2 increased the intracellular amounts of triglycerides >3-fold. Knockdown of NONO negatively regulated the abundance and accumulation of cholesterol [43].
Among post-translation mechanisms of NONO regulation, evidence is growing in support of proteolysis-associated events [44]. One may expect that NONO as a partner in protein-protein complexes, should be protected from proteolytic degradation. Feng et al. demonstrated that, in melanoma cells, the ubiquitin-specific protease 11 (USP11) interacted with nuclear NONO and prevented its subsequent proteolysis by hydrolyzing the polyubiquitinated chains. USP11 deficiency slowed down melanocyte proliferation; cell growth rate was restored with the exogenous NONO, suggesting that NONO is a pro-proliferation factor. A positive correlation between the amounts of USP11 and NONO was confirmed in clinical melanoma samples [45].
2.3. Relevance to Neuroblastoma
Thus, NONO emerges as a genome-wide guardian of transcription. This pivotal role is implemented via the formation of complexes with other proteins and lncRNAs to ensure the onset of transcription and message stabilization. High NONO expression in neuroblastoma correlates with elevated MYCN levels and poor patient survival [46]. In 88 out of 341 patient samples, MYCN amplification was associated with lncUSMYCN overexpression [47]. Interaction of lncUSMYCN RNA with NONO activates NCYM, an antisense transcript that prevents MYCN ubiquitination by inhibiting its GSK3β-mediated degradation [48]. Thus, NONO promotes MYCN-driven oncogenesis. Antisense lncUSMYCN oligonucleotides may therefore be considered as tools to abrogate NONO binding for limiting neuroblastoma progression in mouse models.
Along with lncMYCN, the MYCN protein binds other neuroblastoma-specific lncRNAs such as CASC15, LINC00511, ZRANB2-AS2 and PPP1R26-AS1; the former two transcripts have been characterized as risk factors [49]. The neuroblastoma associated transcript 1 (NBAT-1) lncRNA functions as a tumor suppressor by down-regulating the genes involved in proliferation and invasion, e.g., SOX9, whose overexpression impedes retinoic acid-induced neuronal differentiation. NBAT-1 lncRNA overexpression contributes to inhibition of neuroblastoma growth and metastasis; therefore, efforts have been put on activation of this transcript [50,51,52]. Accordingly, NBAT-1 lncRNA levels inversely correlated with MYCN, suggesting that NBAT-1 lncRNA can be down-regulated in the aggressive disease [53]. NONO binds to lncUSMYCN RNA, leading to up-regulation of MYCN mRNA and proliferation of neuroblastoma cells [47,54].
Two lncRNA isoforms, NEAT1_1 and NEAT1_2, are also being investigated as NONO partners in neuroblastoma. Elevated NEAT1_2 promotes the formation of NONO-containing paraspeckles. The increase of NEAT1_2 with anti-NEAT1_1 oligonucleotides led to sequestration of NONO in paraspeckles. This effect can limit the accessibility of NONO to pro-oncogenic lncRNAs, e.g., lncUSMYCN, thereby decreasing tumor cell viability and/or activating differentiation. Consequently, the anti-oncogenic NONO-lncNEAT1 interaction may attenuate neuroblastoma aggressiveness [20,55]. The study by Simko et al. [56] revealed that G-quadruplex (G4) structures in lncRNA NEAT1 serve as a conserved structural motif for recruiting NONO, a critical step in paraspeckle assembly. The authors demonstrated that NONO exhibits structural specificity for G4 conformations — initially identified in C9orf72 repeat RNA — and that NEAT1 lncRNA harbors abundant, evolutionarily conserved G4 motifs. The persisted G4 enrichment positions the RNA quadruplexes as primary structural elements mediating NONO recruitment and paraspeckle biogenesis.
Yet another level of complexity is provided by recent insights into NONO-dependent gene regulation in neuroblastoma via the formation of nucleic acid-containing condensates across the nuclei. Phase separation allows for NONO involvement in super-enhancer (SE)-regulated HAND2 and GATA2 genes whose products modulate neural cell differentiation and migration [57,58,59].
Furthermore, in neuroblastoma cells NONO acts as a partner with transcription factors. NONO and SOX2 form a bidirectional regulatory network that influences neuroblastoma heterogeneity, stem cell-like properties, plasticity, and resistance to therapy [60]. NONO acts as a transcriptional silencer of the SOX2 gene in neuroblastoma cells. NONO binds to the SOX2 promoter, repressing its transcription and contributing to the maintenance of stemness [61]. In turn, SOX2 protein represses NONO gene expression through a feedback loop, potentially limiting NONO-mediated oncogenic transcription [62].
Along with SOX2, the SOX11 transcription factor emerges as a pivotal regulator in ADRN neuroblastoma, where its amplification stabilizes a lineage-specific CRC essential for tumorigenesis. This effect is mediated by physical interactions with DBHS proteins (NONO, SFPQ, PSPC1) enabling SOX11 to modulate alternative splicing while co-regulating the transcription of splicing factors SRSF2, RBMX and RBM4. Partnership of NONO with SOX11 enhances mRNA processing of SE-driven oncogenes, including CRC members GATA2 and HAND2. The SOX11-NONO axis links chromatin remodeling and RNA splicing, creating a coherent epigenetic RNA operon that confers ADRN identity and represents a therapeutic vulnerability in MYCN-amplified tumors [63]. This reciprocal regulation suggests that the NONO-SOX2/SOX11 axes are critical for neuroblastoma progression via the balance between stemness and differentiation.
In complexes with YY1 transcription factor, NONO regulates the expression of the PARP1 gene whose product covalently adds poly(ADP-riboso) moieties to a variety of targets including NONO. PARylation stabilizes NONO by protecting from ubiquitination. The YY1-PARP1-NONO axis maintains the maturation of ADAM8 and TEX14 transcripts associated with aggressive disease and poor survival [64]. This mechanism, together with YY1-mediated activation of glycolysis genes [65], further supports the negative role of this transcription factor and implicates NONO into YY1-dependent phenomena in neuroblastoma.
Overall, NONO maintains the pro-oncogenic phenotypes in the ‘transcriptional’ tumor such as neuroblastoma via the formation of complexes with nucleic acids and transcription-related proteins. These remarkable abilities validate NONO as a therapeutic target. Figure 1 shows the main functions of NONO.
3. Therapeutic Potential of NONO Targeting: Not Only Neuroblastoma
Roles of NONO in anticancer drug response, as well as the opportunities to pharmacologically inactivate this mechanism, deserve a thorough investigation. At present the reports on these potentially exciting problems are scarce. Tsofack et al. have found that NONO interacts with YB-1 transcription factor in SW480 and HT29 colon carcinoma cell lines. Knockdown of NONO decreased the levels of YB-1 mRNA and protein twofold, in parallel with cell sensitization to oxaliplatin. In turn, YB-1 knockdown attenuated NONO abundance. Overexpression of YB-1 decreased the response to oxaliplatin 2.5–4.4-fold whereas NONO deficiency sensitized YB-1-overexpressing cells to this drug up to 4.4-fold as determined by cell count after 16 h exposure. These results provide strong evidence in favor of the NONO-YB-1 partnership in transcriptional control of survival of colon cancer cells treated with the platinum-containing drug [66].
Cytosine arabinoside is an essential component of therapeutic regiments in acute myelogenous leukemia (AML). NONO stabilized the deoxynucleoside triphosphate triphosphohydrolase SAMHD1 by preventing its ubiquitination. Elevated levels of SAMHD1 were mechanistically attributed to the resistance of AML cells to cytosine arabinoside; down-regulation of NONO sensitized cells to this drug via decreasing SAMHD1. Interestingly, DNA damaging drugs DDP and adriamycin were able to decrease NONO and SAMHD1 proteins, thereby increasing the efficacy of cytosine arabinoside against otherwise resistant AML cells [67,68].
In triple negative breast cancer cell lines (Hs 578T, MDA-MB-231, MDA-MB-468) NONO knockdown (shRNA) suppressed proliferation and colony formation. In MDA-MB-231-derived xenotransplants, siRNA NONO attenuated Ki-67 expression and tumor growth. Li et al. [69] reported that NONO, together with PSPC1 and SFPQ, participates in double strand DNA repair. Knockout of NONO was tolerated by mouse embryo fibroblasts irradiated with γ-photons. However, radiosensitivity was potentiated by knocking out the PSPC1 gene coupled with the NONO knockout. These results substantiated the search for pharmacological blockers of NONO.
Screening of FDA-approved drugs identified auranofin as a NONO antagonist. This agent caused a decrease of NONO mRNA and protein in MDA-MB-231 cells [42]. Floros et al. demonstrated that auranofin (0.33–3.3 μM) was cytotoxic for MYCN-amplified cell lines (IMR-5, SK-N-DZ, Kelly) but not for SK-N-SH and CHLA20 cells that carry single MYCN gene copy. Ferrostatin-1 attenuated the cytotoxicity of auranofin in Kelly and SK-N-DZ cell lines, supporting a role for ferroptosis in auranofin-induced cell death. Administration of auranofin (10 mg/kg i.p. daily) abrogated the growth of IMR-5 transplants as well as patient-derived xenografts in NCG mice This compound elevated ferroptosis markers TfR1 and MDA in IMR-5 tumors [70]. In our experiments the combinations of siRNA-mediated NONO knockdown with auranofin potentiated the reactive oxygen species (ROS) burst and death of MYCN-amplified Kelly cell line [71]. If NONO is indeed a redox sensor, its inhibition justifies the strategy of enhancing the efficacy of ROS-generating drug regimens. This mechanism might be particularly important for eradication of cells that acquired pleiotropic resistance in the course of consecutive rounds of chemotherapy [72].
In MYCN-amplified tumors, dual inhibition of PARP and CHK1 intensified replication stress and blocked S-phase checkpoint control, causing unresolved DNA damage and mitotic catastrophe. PARP blockade stalls replication forks, while CHK1 inhibition prevents their stabilization and homologous recombination repair. This synthetic lethality is selectively potent for MYCN-driven neuroblastoma cell lines (IMR-32 LAN-5, SH-EP). In xenograft models, a low-dose PARP inhibitor olaparib plus MK-8776 markedly suppressed tumor growth and prolonged survival without significant systemic toxicity [73]. Given that NONO is a part of DNA repair machinery, it is worth testing whether NONO depletion would further decrease the viability of aggressive neuroblastoma cells in combination with PARP1/CHK1 inhibition.
The above considerations make NONO inhibition therapeutically meaningful. Indeed, its down-regulation would be beneficial for simultaneous attenuation of several mechanisms. However, the structure of NONO provides a serious challenge for its druggability. A significant step forward has been made by Kathman et al. [74]. The authors screened a broad library of small molecular weight compounds in search for an agent capable of countering the function of the androgen receptor (both wild type and splice variants) in prostate carcinoma cells. The compound termed (R)-SKBG-1 covalently interacted with the cysteine residue at the position 145 of NONO. This site appeared to be critical since NONO disruption or cysteine-to-serine substitution at this position abrogated cell sensitivity to (R)-SKBG-1. Furthermore, NONO inactivation was stereoselective because (S)-SKBG-1 isomer was inefficient. Importantly, (R)-SKBG-1 did not cause NONO depletion but instead trapped the protein to RNA, thereby stabilizing the complex. As a net effect of (R)-SKBG-1, the abundance of androgen receptor mRNA substantially decreased. This elegant study convincingly demonstrated that: 1) NONO is an upstream regulator of the gene pathogenetically involved in carcinogenesis, and 2) NONO is a druggable target.
It is plausible to hypothesize that pharmacological inactivation of NONO can be therapeutically relevant in neuroblastoma. With the pioneer chemical instrument (R)-SKBG-1 and its derivatives being designed on the basis of the crystal structure of ligand-NONO complexes [75,76], it becomes possible to address the question whether NONO inhibition would down-regulate a single or a couple of NONO-dependent CRC genes. In turn, the combinations of conventional chemotherapeutics with (R)-SKBG-1 are expected to reveal the potential of NONO inactivation as a component of treatment regimens if inactivation of NONO alone is insufficient for killing tumor cells.
Furthermore, an imidazolium-based compound sepantronium bromide (YM155) has been recently reported to prevent the formation of complexes between the interleukin enhancer binding factor 3 (ILF3) and NONO (p54nrb), a mechanism involved in the survivin gene regulation. In HEK293 and PC-3 cell lines, YM155 down-regulated survivin via destabilization of ILF3/p54nrb complexes. In co-immunoprecipitation experiments this compound decreased IFL3-p54nrb interaction by ~80% without affecting the abundance of each protein. ChIP analysis revealed that p54nrb and ILF3 were enriched on the survivin gene promoter whereas YM155 attenuated these interactions. The siRNA-assisted p54nrb knockdown led to down-regulation of the survivin gene [77]. YM155 entered clinical trials in several intractable tumors including castration-resistant prostate cancer. Thus, this agent could be useful for the design of therapeutic regiments in MYCN/NONO overexpressing neuroblastoma. One may expect that YM155 will be efficient in combinations with another chemotherapeutics. However, chemical structures of R(SKBG-1) and YM155 are strikingly different, surmising that each compound acts via an individual mechanism. On the one hand, this diversity expands pharmacological opportunities for NONO inactivation. On the other hand, the precise mechanisms remain elusive due to manifold modes of NONO regulation.
Finally, regulation of NONO ubiquitination should be considered therapeutically relevant. As mentioned in section Roles in cancer, targeting USP can be an option for the control of NONO stability to proteasomal degradation. Inhibition of USP7 and -28 destabilized MYCN in SK-N-BE(2) and IMR32 cell lines [78,79]. USP11 in complexes with the transcription elongation factor A-like 1 has been found to be essential for the expression program of mesenchymal genes [80]. Lone et al. reported an indirect way of NONO destabilization by inhibiting its partner peptidyl-prolyl cis-trans isomerase NIMA-interacting 1 (Pin1) protein with 5-hydroxy-1,4-naphthalenedione (Juglone) [44]. This effect was reversible by the proteasomal blocker MG-132. Since NONO regulation by small molecular weight compounds is a developing area, the selectivity and the mechanisms warrant a cautious interpretation. Table 1 summarizes information on NONO antagonists applicable in neuroblastoma and other tumors.
4. Conclusions
It is the genome-wide disordering of transcription that underlies a remarkable biological heterogeneity of neuroblastoma. The current classification determines transcriptional mechanisms unified by common regulation. According to this analysis, the ADRN and MES subtypes are characterized by the sets of 18 and 20 transcription factors, respectively [15]. Some of these factors comprise a specific autoregulatory loop termed CRC, a hallmark of tumors with ‘transcriptional’ (rather than ‘mutational’) burden [1]. This machinery forms a network of intergenic interactions that are ultimately translated into specific clinical manifestations [81].
It is logical to suggest that this complex machinery is governed by the hierarchy of mechanisms. Among other candidates, the NONO protein emerged as a major master regulator of specific genes including CRC. Not surprisingly, NONO has been implicated in a plethora of functions in normal development as well as in tumor biology, particularly in the neural tissue. Now the question arises as to whether such a multifunctional mechanism should – or can – be considered a therapeutic target?
Several difficulties stay on the road to the positive answer. First, as a protein that lacks enzymatic function, NONO is expected to be hardly druggable especially keeping in mind its propensity to interact with macromolecules. Moreover, if NONO acts not as much as a homodimer but in partnership with other DBHS family proteins and specific RNAs, its druggability becomes even more problematic. The discovery of the small molecular weight covalent NONO blocker (R)-SKBG-1 set the stage for systematic investigation of mechanisms of NONO inactivation. Intriguingly, in (R)-SKBG-1-treated cells the abundance of NONO remained unaltered whereas NONO-dependent expression of the androgen receptor gene was abolished [72]. The proposed mechanism suggests that (R)-SKBG-1 immobilizes NONO-containing complexes. Detailed structural investigation of ligand-NONO interaction requires molecular modeling and X-ray studies of crystallized complexes; also, dissection of structure-activity relationship by analyzing individual chemical moieties in (R)-SKBG-1, e.g., the reactive chloroacetyl group, is worthy. Can NONO be immobilized by molecular glues, a developing class of compounds aimed at sticking to and fixing the non-enzymatic proteins (see [82] for recent review)?
Next, pharmacological inactivation of NONO alone, even if the target genes are attenuated, may or may not be sufficient for cell death. The net effect of NONO inhibition on cell survival would depend on the significance of individual NONO-dependent genes for a particular cell type in a specific context. This notion holds true for relapsed neuroblastoma after the repeated rounds of chemo- and radiotherapy. What happens to NONO in the course of prolonged treatment and what is an additional mechanism(s) suitable for drug combinations? The former question needs special investigation; for the latter opportunity, literature data points at DNA repair proteins as tentative candidates. Availability and efficacy of DNA damage response pathway inhibitors make their combinations with the NONO blockers straightforward.
In summary, the strategies of targeting the master regulators in ‘transcriptional’ tumors including neuroblastoma are founded on the growing knowledge of molecular pathogenesis of gene expression alterations. One may expect that inactivation of one druggable mechanism would cause a ramification effect, that is, inhibit a number of downstream pathways. Trials of the current NONO antagonists in monotherapy and in combinations with conventional and targeted drugs will demonstrate whether the antitumor efficacy is achievable within a reasonable therapeutic ‘window’.
Author Contributions
SP and OM contributed to data acquisition, analysis and interpretation, and drafted the manuscript. SP and OK designed the figures and table. OK and AS generated the concept; designed and revised the manuscript. NA and AS performed editing, proofreading, and critical evaluation of the study. All authors read and approved the final manuscript.
Funding
This research was supported by the Russian Science Foundation, grant 24-15-00097.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable (only appropriate if no new data is generated or the article describes entirely theoretical research).
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ADRN | adrenergic |
| CRC | core regulatory circuit |
| DBHS | Drosophila Behavior/Human Splicing |
| lncRNA | long non-coding RNA |
| MES | mesenchymal |
| SE | super-enhancer |
References
- Comitani, F.; Nash, J. O.; Cohen-Gogo, S.; Chang, A. I.; Wen, T. T.; Maheshwari, A.; Goyal, B.; Tio, E. S.; Tabatabaei, K.; Mayoh, C.; et al. Diagnostic Classification of Childhood Cancer Using Multiscale Transcriptomics. Nat. Med. 2023, 29 (3), 656–666. [CrossRef]
- Cheung, N.-K. V.; Dyer, M. A. Neuroblastoma: Developmental Biology, Cancer Genomics and Immunotherapy. Nat. Rev. Cancer 2013, 13 (6), 397–411. [CrossRef]
- Matthay, K. K.; Maris, J. M.; Schleiermacher, G.; Nakagawara, A.; Mackall, C. L.; Diller, L.; Weiss, W. A. Neuroblastoma. Nat. Rev. Dis. Primer 2016, 2 (1), 16078. [CrossRef]
- Tolbert, V. P.; Matthay, K. K. Neuroblastoma: Clinical and Biological Approach to Risk Stratification and Treatment. Cell Tissue Res. 2018, 372 (2), 195–209. [CrossRef]
- Park, J. R.; Bagatell, R.; London, W. B.; Maris, J. M.; Cohn, S. L.; Mattay, K. M.; Hogarty, M.; Committee, on behalf of the C. N. Children’s Oncology Group’s 2013 Blueprint for Research: Neuroblastoma. Pediatr. Blood Cancer 2013, 60 (6), 985–993. [CrossRef]
- Lundberg, K. I.; Treis, D.; Johnsen, J. I. Neuroblastoma Heterogeneity, Plasticity, and Emerging Therapies. Curr. Oncol. Rep. 2022, 24 (8), 1053–1062. [CrossRef]
- Irwin, M. S.; Naranjo, A.; Zhang, F. F.; Cohn, S. L.; London, W. B.; Gastier-Foster, J. M.; Ramirez, N. C.; Pfau, R.; Reshmi, S.; Wagner, E.; et al. Revised Neuroblastoma Risk Classification System: A Report From the Children’s Oncology Group. J. Clin. Oncol. 2021, 39 (29), 3229–3241. [CrossRef]
- Pugh, T. J.; Morozova, O.; Attiyeh, E. F.; Asgharzadeh, S.; Wei, J. S.; Auclair, D.; Carter, S. L.; Cibulskis, K.; Hanna, M.; Kiezun, A.; et al. The Genetic Landscape of High-Risk Neuroblastoma. Nat. Genet. 2013, 45 (3), 279–284. [CrossRef]
- Rajbhandari, P.; Lopez, G.; Capdevila, C.; Salvatori, B.; Yu, J.; Rodriguez-Barrueco, R.; Martinez, D.; Yarmarkovich, M.; Weichert-Leahey, N.; Abraham, B. J.; et al. Cross-Cohort Analysis Identifies a TEAD4–MYCN Positive Feedback Loop as the Core Regulatory Element of High-Risk Neuroblastoma. Cancer Discov. 2018, 8 (5), 582–599. [CrossRef]
- Kuchur, O. A.; Pogodaeva, S. S.; Zhdankina, V. I.; Scherbakova, A. V.; Shtil, A. A.; Antipova, N. V. Targeting Transcription in Neuroblastoma: Focus on the Core Regulatory Circuit. Expert Opin. Ther. Targets. 2025, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Boeva, V.; Louis-Brennetot, C.; Peltier, A.; Durand, S.; Pierre-Eugène, C.; Raynal, V.; Etchevers, H. C.; Thomas, S.; Lermine, A.; Daudigeos-Dubus, E.; et al. Heterogeneity of Neuroblastoma Cell Identity Defined by Transcriptional Circuitries. Nat. Genet. 2017, 49 (9), 1408–1413. [CrossRef]
- Wang, L.; Tan, T. K.; Durbin, A. D.; Zimmerman, M. W.; Abraham, B. J.; Tan, S. H.; Ngoc, P. C. T.; Weichert-Leahey, N.; Akahane, K.; Lawton, L. N.; et al. ASCL1 Is a MYCN- and LMO1-Dependent Member of the Adrenergic Neuroblastoma Core Regulatory Circuitry. Nat. Commun. 2019, 10 (1), 5622. [CrossRef]
- Durbin, A. D.; Zimmerman, M. W.; Dharia, N. V.; Abraham, B. J.; Iniguez, A. B.; Weichert-Leahey, N.; He, S.; Krill-Burger, J. M.; Root, D. E.; Vazquez, F.; et al. Selective Gene Dependencies in MYCN-Amplified Neuroblastoma Include the Core Transcriptional Regulatory Circuitry. Nat. Genet. 2018, 50 (9), 1240–1246. [CrossRef]
- Gautier, M.; Thirant, C.; Delattre, O.; Janoueix-Lerosey, I. Plasticity in Neuroblastoma Cell Identity Defines a Noradrenergic-to-Mesenchymal Transition (NMT). Cancers 2021, 13 (12), 2904. [CrossRef]
- van Groningen, T.; Koster, J.; Valentijn, L. J.; Zwijnenburg, D. A.; Akogul, N.; Hasselt, N. E.; Broekmans, M.; Haneveld, F.; Nowakowska, N. E.; Bras, J.; et al. Neuroblastoma Is Composed of Two Super-Enhancer-Associated Differentiation States. Nat. Genet. 2017, 49 (8), 1261–1266. [CrossRef]
- Ronchetti, D.; Traini, V.; Silvestris, I.; Fabbiano, G.; Passamonti, F.; Bolli, N.; Taiana, E. The Pleiotropic Nature of NONO, a Master Regulator of Essential Biological Pathways in Cancers. Cancer Gene Ther. 2024, 31 (7), 984–994. [CrossRef]
- Liu, X.; Zheng, J.; Qi, S.; Shen, Q. NONO Regulates Cortical Neuronal Migration and Postnatal Neuronal Maturation. Neurosci. Bull. 2019, 35 (6), 1097–1101. [CrossRef]
- Yu, D.; Huang, C.-J.; Tucker, H. O. Established and Evolving Roles of the Multifunctional Non-POU Domain-Containing Octamer-Binding Protein (NonO) and Splicing Factor Proline- and Glutamine-Rich (SFPQ). J. Dev. Biol. 2024, 12 (1), 3. [CrossRef]
- Passon, D. M.; Lee, M.; Rackham, O.; Stanley, W. A.; Sadowska, A.; Filipovska, A.; Fox, A. H.; Bond, C. S. Structure of the Heterodimer of Human NONO and Paraspeckle Protein Component 1 and Analysis of Its Role in Subnuclear Body Formation. Proc. Natl. Acad. Sci. 2012, 109 (13), 4846–4850. [CrossRef]
- Taiana, E.; Ronchetti, D.; Todoerti, K.; Nobili, L.; Tassone, P.; Amodio, N.; Neri, A. LncRNA NEAT1 in Paraspeckles: A Structural Scaffold for Cellular DNA Damage Response Systems? Non-Coding RNA 2020, 6 (3), 26. [CrossRef]
- Schell, B.; Legrand, P.; Fribourg, S. Crystal Structure of SFPQ-NONO Heterodimer. Biochimie 2022, 198, 1–7. [Google Scholar] [CrossRef]
- Zhang, H.; Ji, X.; Li, P.; Liu, C.; Lou, J.; Wang, Z.; Wen, W.; Xiao, Y.; Zhang, M.; Zhu, X. Liquid-Liquid Phase Separation in Biology: Mechanisms, Physiological Functions and Human Diseases. Sci. China Life Sci. 2020, 63 (7), 953–985. [CrossRef]
- Knott, G. J.; Lee, M.; Passon, D. M.; Fox, A. H.; Bond, C. S. Caenorhabditis Elegans NONO-1: Insights into DBHS Protein Structure, Architecture, and Function. [CrossRef]
- Lee, M.; Sadowska, A.; Bekere, I.; Ho, D.; Gully, B. S.; Lu, Y.; Iyer, K. S.; Trewhella, J.; Fox, A. H.; Bond, C. S. The Structure of Human SFPQ Reveals a Coiled-Coil Mediated Polymer Essential for Functional Aggregation in Gene Regulation. Nucleic Acids Res. 2015, 43 (7), 3826–3840. [CrossRef]
- Wang, H.; Chen, S.; Wei, J.; Song, G.; Zhao, Y. A-to-I RNA Editing in Cancer: From Evaluating the Editing Level to Exploring the Editing Effects. Front. Oncol. 2021, 10. [Google Scholar] [CrossRef]
- Feng, P.; Li, L.; Deng, T.; Liu, Y.; Ling, N.; Qiu, S.; Zhang, L.; Peng, B.; Xiong, W.; Cao, L.; et al. NONO and Tumorigenesis: More than Splicing. [CrossRef]
- Zhang, Z.; Carmichael, G. G. The Fate of dsRNA in the Nucleus: A P54nrb-Containing Complex Mediates the Nuclear Retention of Promiscuously A-to-I Edited RNAs. Cell 2001, 106 (4), 465–476. [CrossRef]
- Li, W.; Karwacki-Neisius, V.; Ma, C.; Tan, L.; Shi, Y.; Wu, F.; Shi, Y. G. Nono Deficiency Compromises TET1 Chromatin Association and Impedes Neuronal Differentiation of Mouse Embryonic Stem Cells. Nucleic Acids Res. 2020, 48 (9), 4827–4838. [CrossRef]
- Ma, C.; Karwacki-Neisius, V.; Tang, H.; Li, W.; Shi, Z.; Hu, H.; Xu, W.; Wang, Z.; Kong, L.; Lv, R.; et al. Nono, a Bivalent Domain Factor, Regulates Erk Signaling and Mouse Embryonic Stem Cell Pluripotency. Cell Rep. 2016, 17 (4), 997–1007. [CrossRef]
- Shen, M.; Zhang, R.; Jia, W.; Zhu, Z.; Zhao, L.; Huang, G.; Liu, J. RNA-Binding Protein P54nrb/NONO Potentiates Nuclear EGFR-Mediated Tumorigenesis of Triple-Negative Breast Cancer. Cell Death Dis. 2022, 13 (1), 42. [CrossRef]
- Wei, Y.; Luo, H.; Yee, P. P.; Zhang, L.; Liu, Z.; Zheng, H.; Zhang, L.; Anderson, B.; Tang, M.; Huang, S.; et al. Paraspeckle Protein NONO Promotes TAZ Phase Separation in the Nucleus to Drive the Oncogenic Transcriptional Program. [CrossRef]
- Kowalska, E.; Ripperger, J. A.; Hoegger, D. C.; Bruegger, P.; Buch, T.; Birchler, T.; Mueller, A.; Albrecht, U.; Contaldo, C.; Brown, S. A. NONO Couples the Circadian Clock to the Cell Cycle. Proc. Natl. Acad. Sci. 2013, 110 (5), 1592–1599. [CrossRef]
- Dong, X.; Yu, C.; Shynlova, O.; Challis, J. R. G.; Rennie, P. S.; Lye, S. J. P54nrb Is a Transcriptional Corepressor of the Progesterone Receptor That Modulates Transcription of the Labor-Associated Gene, Connexin 43 (Gja1). Mol. Endocrinol. 2009, 23 (8), 1147–1160. [CrossRef]
- Yang, P.; Chen, T.; Xu, Z.; Zhu, H.; Wang, J.; He, Z. Long Noncoding RNA GAPLINC Promotes Invasion in Colorectal Cancer by Targeting SNAI2 through Binding with PSF and NONO. Oncotarget 2016, 7 (27), 42183–42194. [CrossRef]
- Cheng, R.; Zhu, S.; Guo, S.; Min, L.; Xing, J.; Guo, Q.; Li, P.; Zhang, S. Downregulation of NONO Induces Apoptosis, Suppressing Growth and Invasion in Esophageal Squamous Cell Carcinoma. Oncol. Rep. 2018, 39 (6), 2575–2583. [CrossRef]
- Hou, Y.-R.; Diao, L.-T.; Hu, Y.-X.; Zhang, Q.-Q.; Lv, G.; Tao, S.; Xu, W.-Y.; Xie, S.-J.; Zhang, Q.; Xiao, Z.-D. The Conserved LncRNA DIO3OS Restricts Hepatocellular Carcinoma Stemness by Interfering with NONO-Mediated Nuclear Export of ZEB1 mRNA. [CrossRef]
- Trifault, B.; Mamontova, V.; Cossa, G.; Ganskih, S.; Wei, Y.; Hofstetter, J.; Bhandare, P.; Baluapuri, A.; Nieto, B.; Solvie, D.; et al. Nucleolar Detention of NONO Shields DNA Double-Strand Breaks from Aberrant Transcripts. Nucleic Acids Res. 2024, 52 (6), 3050–3068. [CrossRef]
- Li, S.; Shu, F.; Li, Z.; Jaafar, L.; Zhao, S.; Dynan, W. S. Cell-Type Specific Role of the RNA-Binding Protein, NONO, in the DNA Double-Strand Break Response in the Mouse Testes. DNA Repair 2017, 51, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Alfano, L.; Costa, C.; Caporaso, A.; Altieri, A.; Indovina, P.; Macaluso, M.; Giordano, A.; Pentimalli, F. NONO Regulates the Intra-S-Phase Checkpoint in Response to UV Radiation. Oncogene 2016, 35 (5), 567–576. [CrossRef]
- Fan, X.-J.; Wang, Y.-L.; Zhao, W.-W.; Bai, S.-M.; Ma, Y.; Yin, X.-K.; Feng, L.-L.; Feng, W.-X.; Wang, Y.-N.; Liu, Q.; et al. NONO Phase Separation Enhances DNA Damage Repair by Accelerating Nuclear EGFR-Induced DNA-PK Activation. Am. J. Cancer Res. 2021, 11(6), 2838–2852. [Google Scholar]
- Mamontova, V.; Trifault, B.; Burger, K. Nono Induces Gadd45b to Mediate DNA Repair. Life Sci. Alliance 2024, 7(8). [Google Scholar] [CrossRef]
- Kim, S.-J.; Ju, J.-S.; Kang, M.-H.; Won, J. E.; Kim, Y. H.; Raninga, P. V.; Khanna, K. K.; Győrffy, B.; Pack, C.-G.; Han, H.-D.; Lee, H. J.; et al. RNA-Binding Protein NONO Contributes to Cancer Cell Growth and Confers Drug Resistance as a Theranostic Target in TNBC. Theranostics 2020, 10(18), 7974–7992. [Google Scholar] [CrossRef]
- Zhu, Z.; Zhao, X.; Zhao, L.; Yang, H.; Liu, L.; Li, J.; Wu, J.; Yang, F.; Huang, G.; Liu, J. P54nrb/NONO Regulates Lipid Metabolism and Breast Cancer Growth through SREBP-1A. Oncogene 2016, 35(11), 1399–1410. [Google Scholar] [CrossRef]
- Lone, B. A.; Siraj, F.; Sharma, I.; Verma, S.; Karna, S. K. L.; Ahmad, F.; Nagar, P.; Sachidanandan, C.; Pokharel, Y. R. Non-POU Domain-Containing Octomer-Binding (NONO) Protein Expression and Stability Promotes the Tumorigenicity and Activation of Akt/MAPK/β-Catenin Pathways in Human Breast Cancer Cells. Cell Commun. Signal. 2023, 21(1), 157. [Google Scholar] [CrossRef]
- Feng, P.; Li, L.; Dai, J.; Zhou, L.; Liu, J.; Zhao, J.; Li, X.; Ling, N.; Qiu, S.; Zhang, L.; et al. The Regulation of NONO by USP11 via Deubiquitination Is Linked to the Proliferation of Melanoma Cells. [CrossRef]
- Wang, X.; Han, M.; Wang, S.; Sun, Y.; Zhao, W.; Xue, Z.; Liang, X.; Huang, B.; Li, G.; Chen, A.; et al. Targeting the Splicing Factor NONO Inhibits GBM Progression through GPX1 Intron Retention. Theranostics 2022, 12(12), 5451–5469. [Google Scholar] [CrossRef] [PubMed]
- Liu, P. Y.; Erriquez, D.; Marshall, G. M.; Tee, A. E.; Polly, P.; Wong, M.; Liu, B.; Bell, J. L.; Zhang, X. D.; Milazzo, G.; et al. Effects of a Novel Long Noncoding RNA, lncUSMycN, on N-Myc Expression and Neuroblastoma Progression. JNCI J. Natl. Cancer Inst. 2014, 106(7), dju113. [Google Scholar] [CrossRef]
- Liu, P. Y.; Atmadibrata, B.; Mondal, S.; Tee, A. E.; Liu, T. NCYM Is Upregulated by lncUSMycN and Modulates N-Myc Expression. Int. J. Oncol. 2016, 49(6), 2464–2470. [Google Scholar] [CrossRef] [PubMed]
- Prajapati, B.; Fatma, M.; Fatima, M.; Khan, M. T.; Sinha, S.; Seth, P. K. Identification of lncRNAs Associated With Neuroblastoma in Cross-Sectional Databases: Potential Biomarkers. Front. Mol. Neurosci. 2019, 12. [Google Scholar] [CrossRef] [PubMed]
- Pandey, G. K.; Mitra, S.; Subhash, S.; Hertwig, F.; Kanduri, M.; Mishra, K.; Fransson, S.; Ganeshram, A.; Mondal, T.; Bandaru, S.; et al. The Risk-Associated Long Noncoding RNA NBAT-1 Controls Neuroblastoma Progression by Regulating Cell Proliferation and Neuronal Differentiation. Cancer Cell 2014, 26(5), 722–737. [Google Scholar] [CrossRef]
- Yang, C.-L.; Serra-Roma, A.; Gualandi, M.; Bodmer, N.; Niggli, F.; Schulte, J. H.; Bode, P. K.; Shakhova, O. Lineage-Restricted Sympathoadrenal Progenitors Confer Neuroblastoma Origin and Its Tumorigenicity. Oncotarget 2020, 11(24), 2357–2371. [Google Scholar] [CrossRef] [PubMed]
- Mondal, T.; Juvvuna, P. K.; Kirkeby, A.; Mitra, S.; Kosalai, S. T.; Traxler, L.; Hertwig, F.; Wernig-Zorc, S.; Miranda, C.; Deland, L.; et al. Sense-Antisense lncRNA Pair Encoded by Locus 6p22.3 Determines Neuroblastoma Susceptibility via the USP36-CHD7-SOX9 Regulatory Axis. Cancer Cell 2018, 33(3), 417–434.e7. [Google Scholar] [CrossRef]
- Juvvuna, P. K.; Mondal, T.; Di Marco, M.; Kosalai, S. T.; Kanduri, M.; Kanduri, C. NBAT1/CASC15-003/USP36 Control MYCN Expression and Its Downstream Pathway Genes in Neuroblastoma. Neuro-Oncol. Adv. 2021, 3(1), vdab056. [Google Scholar] [CrossRef]
- Pichler, M.; Calin, G. A. Long Noncoding RNA in Neuroblastoma: New Light on the (Old) N-Myc Story. JNCI J. Natl. Cancer Inst. 2014, 106(7), dju150. [Google Scholar] [CrossRef]
- Naveed, A.; Cooper, J. A.; Li, R.; Hubbard, A.; Chen, J.; Liu, T.; Wilton, S. D.; Fletcher, S.; Fox, A. H. NEAT1 polyA-Modulating Antisense Oligonucleotides Reveal Opposing Functions for Both Long Non-Coding RNA Isoforms in Neuroblastoma. Cell. Mol. Life Sci. 2021, 78(5), 2213–2230. [Google Scholar] [CrossRef]
- Simko, E. A. J.; Liu, H.; Zhang, T.; Velasquez, A.; Teli, S.; Haeusler, A. R.; Wang, J. G-Quadruplexes Offer a Conserved Structural Motif for NONO Recruitment to NEAT1 Architectural lncRNA. Nucleic Acids Res. 2020, 48(13), 7421–7438. [Google Scholar] [CrossRef]
- Zhang, S.; Cooper, J. A.; Chong, Y. S.; Naveed, A.; Mayoh, C.; Jayatilleke, N.; Liu, T.; Amos, S.; Kobelke, S.; Marshall, A. C.; et al. NONO Enhances mRNA Processing of Super-enhancer-associated GATA2 and HAND2 Genes in Neuroblastoma. EMBO Rep. 2023, 24(2), e54977. [Google Scholar] [CrossRef]
- Voth, H.; Oberthuer, A.; Simon, T.; Kahlert, Y.; Berthold, F.; Fischer, M. Co-Regulated Expression of HAND2 and DEIN by a Bidirectional Promoter with Asymmetrical Activity in Neuroblastoma. BMC Mol. Biol. 2009, 10(1), 28. [Google Scholar] [CrossRef]
- Willett, R. T.; Greene, L. A. Gata2 Is Required for Migration and Differentiation of Retinorecipient Neurons in the Superior Colliculus. J. Neurosci. 2011, 31(12), 4444–4455. [Google Scholar] [CrossRef]
- Yang, S.; Zheng, J.; Xiao, X.; Xu, T.; Tang, W.; Zhu, H.; Yang, L.; Zheng, S.; Dong, K.; Zhou, G.; et al. SOX2 Promotes Tumorigenicity and Inhibits the Differentiation of I-Type Neuroblastoma Cells. Int. J. Oncol. 2015, 46(1), 317–323. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Takahashi, H.; Hirose, T.; Kuramitsu, Y.; Hatakeyama, S.; Yoshiyama, H.; Wang, R.; Hamada, J.-I.; Iizasa, H. NONO Is a Negative Regulator of SOX2 Promoter. Cancer Genomics Proteomics 2020, 17(4), 359–367. [Google Scholar] [CrossRef]
- Bao, J.; Qin, L.; Cui, L.; Wang, X.; Meng, Q.; Zhu, L.; Zhang, S. Microarray Data Analysis of Neuroblastoma: Expression of SOX2 Downregulates the Expression of MYCN. Mol. Med. Rep. 2015, 12(5), 6867–6872. [Google Scholar] [CrossRef]
- Louwagie, A. Molecular Characterization of SOX11 as Multifaceted Epigenetic Regulator in Neuroblastoma. dissertation, Ghent University, 2023. http://hdl.handle.net/1854/LU-01HCYBJTB9H842EN2QH2V4R5FR (accessed 2025-08-19).
- Yang, C.; Qu, J.; Cheng, Y.; Tian, M.; Wang, Z.; Wang, X.; Li, X.; Zhou, S.; Zhao, B.; Guo, Y.; et al. YY1 Drives PARP1 Expression Essential for PARylation of NONO in mRNA Maturation during Neuroblastoma Progression. J. Transl. Med. 2024, 22(1), 1153. [Google Scholar] [CrossRef] [PubMed]
- Fang, E.; Wang, X.; Wang, J.; Hu, A.; Song, H.; Yang, F.; Li, D.; Xiao, W.; Chen, Y.; Guo, Y.; et al. Therapeutic Targeting of YY1/MZF1 Axis by MZF1-uPEP Inhibits Aerobic Glycolysis and Neuroblastoma Progression. Theranostics 2020, 10(4), 1555–1571. [Google Scholar] [CrossRef]
- Tsofack, S. P.; Garand, C.; Sereduk, C.; Chow, D.; Aziz, M.; Guay, D.; Yin, H. H.; Lebel, M. NONO and RALY Proteins Are Required for YB-1 Oxaliplatin Induced Resistance in Colon Adenocarcinoma Cell Lines. Mol. Cancer 2011, 10(1), 145. [Google Scholar] [CrossRef] [PubMed]
- Mohty, M.; Malard, F.; Blaise, D.; Milpied, N.; Socié, G.; Huynh, A.; Reman, O.; Yakoub-Agha, I.; Furst, S.; Guillaume, T.; et al. Sequential Regimen of Clofarabine, Cytosine Arabinoside and Reduced-Intensity Conditioned Transplantation for Primary Refractory Acute Myeloid Leukemia. Haematologica 2017, 102(1), 184–191. [Google Scholar] [CrossRef]
- Zhang, F.; Sun, J.; Tang, X.; Liang, Y.; Jiao, Q.; Yu, B.; Dai, Z.; Yuan, X.; Li, J.; Yan, J.; et al. Stabilization of SAMHD1 by NONO Is Crucial for Ara-C Resistance in AML. Cell Death Dis. 2022, 13(7), 590. [Google Scholar] [CrossRef]
- Li, S.; Li, Z.; Shu, F.-J.; Xiong, H.; Phillips, A. C.; Dynan, W. S. Double-Strand Break Repair Deficiency in NONO Knockout Murine Embryonic Fibroblasts and Compensation by Spontaneous Upregulation of the PSPC1 Paralog. Nucleic Acids Res. 2014, 42(15), 9771–9780. [Google Scholar] [CrossRef] [PubMed]
- Floros, K. V.; Cai, J.; Jacob, S.; Kurupi, R.; Fairchild, C. K.; Shende, M.; Coon, C. M.; Powell, K. M.; Belvin, B. R.; Hu, B.; et al. MYCN-Amplified Neuroblastoma Is Addicted to Iron and Vulnerable to Inhibition of the System Xc-/Glutathione Axis. Cancer Res. 2021, 81(7), 1896–1908. [Google Scholar] [CrossRef] [PubMed]
- Kuchur, O.A.; Pogodaeva, S.S., et al. Method for enhancing cell death of MYCN-amplified neuroblastoma cells. Russian Federation patent pending, № 2025119605, Rospatent, 2025.
- Tsymbal, S. A.; Moiseeva, A. A.; Agadzhanian, N. A.; Efimova, S. S.; Markova, A. A.; Guk, D. A.; Krasnovskaya, O. O.; Alpatova, V. M.; Zaitsev, A. V.; Shibaeva, A. V.; et al. A. A. Copper-Containing Nanoparticles and Organic Complexes: Metal Reduction Triggers Rapid Cell Death via Oxidative Burst. Int. J. Mol. Sci. 2021, 22(20), 11065. [Google Scholar] [CrossRef]
- Di Giulio, S.; Colicchia, V.; Pastorino, F.; Pedretti, F.; Fabretti, F.; Nicolis di Robilant, V.; Ramponi, V.; Scafetta, G.; Moretti, M.; Licursi, V.; et al. A Combination of PARP and CHK1 Inhibitors Efficiently Antagonizes MYCN-Driven Tumors. Oncogene 2021, 40(43), 6143–6152. [Google Scholar] [CrossRef] [PubMed]
- Kathman, S. G.; Koo, S. J.; Lindsey, G. L.; Her, H.-L.; Blue, S. M.; Li, H.; Jaensch, S.; Remsberg, J. R.; Ahn, K.; Yeo, G. W.; et al. Remodeling Oncogenic Transcriptomes by Small Molecules Targeting NONO. Nat. Chem. Biol. 2023, 19(7), 825–836. [Google Scholar] [CrossRef] [PubMed]
- Lindsey, G. L.; Hockley, T. K.; Gomez, A. V.; Marshall, A. C.; Brothers, W. R.; Finney, C. T.; Gross, J.; Fox, A. H.; Yeo, G. W.; Melillo, B.; et al. Structural and Mechanistic Analysis of Covalent Ligands Targeting the RNA-Binding Protein NONO. [preprint] bioRxiv June 11, 2025, p 2025.06.10.658857. [CrossRef]
- Florio, A. V.; Buré, C.; Fribourg, S. Structural Basis for NONO Specific Modification by the α-Chloroacetamide Compound (R)-SKBG-1. [preprint] bioRxiv June 12, 2025, p 2025.06.10.658868. [CrossRef]
- Yamauchi, T.; Nakamura, N.; Hiramoto, M.; Yuri, M.; Yokota, H.; Naitou, M.; Takeuchi, M.; Yamanaka, K.; Kita, A.; Nakahara, T.; et al. Sepantronium Bromide (YM155) Induces Disruption of the ILF3/P54nrb Complex, Which Is Required for Survivin Expression. Biochem. Biophys. Res. Commun. 2012, 425(4), 711–716. [Google Scholar] [CrossRef]
- Li, J.; Peng, J.; Wu, L.; Shen, X.; Zhen, X.; Zhang, Y.; Ma, H.; Xu, Y.; Xiong, Q.; Zhu, Q.; et al. The Deubiquitinase USP28 Maintains the Expression of the Transcription Factor MYCN and Is Essential in Neuroblastoma Cells. J. Biol. Chem. 2023, 299(7). [Google Scholar] [CrossRef]
- Tavana, O.; Li, D.; Dai, C.; Lopez, G.; Banerjee, D.; Kon, N.; Chen, C.; Califano, A.; Yamashiro, D. J.; Sun, H.; et al. HAUSP Deubiquitinates and Stabilizes N-Myc in Neuroblastoma. Nat. Med. 2016, 22(10), 1180–1186. [Google Scholar] [CrossRef]
- Dehmer, M.; Trunk, K.; Gallant, P.; Fleischhauer, D.; Müller, M.; Herold, S.; Cossa, G.; Conte, F.; Koster, J.; Sauer, F.; et al. The USP11/TCEAL1 Complex Promotes Transcription Elongation to Sustain Oncogenic Gene Expression in Neuroblastoma. Genes Dev. 2025, 39(11–12), 751–769. [Google Scholar] [CrossRef]
- Jahangiri, L.; Pucci, P.; Ishola, T.; Pereira, J.; Cavanagh, M. L.; Turner, S. D. Deep Analysis of Neuroblastoma Core Regulatory Circuitries Using Online Databases and Integrated Bioinformatics Shows Their Pan-Cancer Roles as Prognostic Predictors. Discov. Oncol. 2021, 12(1), 56. [Google Scholar] [CrossRef] [PubMed]
- Jungfleisch, J.; Gebauer, F. RNA-Binding Proteins as Therapeutic Targets in Cancer. RNA Biol. 2025, 22(1), 1–8. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Roles of the multifunctional protein NONO. Shown are NONO functions in the context of protein-protein and protein-RNA complexes, as well as molecular mechanisms underlying each function.
Figure 1.
Roles of the multifunctional protein NONO. Shown are NONO functions in the context of protein-protein and protein-RNA complexes, as well as molecular mechanisms underlying each function.

Table 1.
Small molecular weight compounds for NONO inhibition.
| Compound | Mechanism of action/Effects | Trial | Refs |
|---|---|---|---|
R-SKBG-1
|
Covalently binds Cys145 to lock NONO in an RNA-bound state for inactivation of androgen receptor gene regulation | Preclinical | [74,75,76] |
Auranofin
|
Suppresses NONO (possibly through thioredoxin reductase inhibition) to enhance ROS-driven ferroptosis in MYCN-amplified cells | Phase II: chronic lymphocytic leukemia (NCT01419691) Phase I: non-small cell lung carcinoma or small cell lung carcinoma (NCT01737502), ovarian, primary peritoneal, or fallopian tube cancer (NCT01747798) |
[42,71] |
Sepantronium bromide (YM155)
|
Disrupts NONO–ILF3 complex at the survivin promoter, thereby down-regulating this gene and inducing apoptosis. | Phase I/II: solid tumors, NSCLC (NCT01100931), Phase II: melanoma (NCT01009775) | [77] |
Olaparib + MK-8776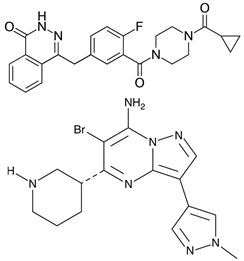
|
Olaparib inhibits PARP1-mediated DNA repair, intensifying replicative stress in cells with active NONO. The CHK1 blocker MK-8776 prevents DNA repair. | Olaparib - Phase II: BRCA-mutated tumors (NCT04041128, NCT02677038) MK-8776 - Phase I: solid tumors, lymphoma (NCT00779584) |
[73] |
Juglone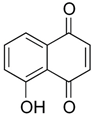
|
Inhibits NONO stabilization via prevention of Pin-1-dependent ubiquitination and proteasomal degradation. | Preclinical | [44] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



