
Submitted:
29 August 2025
Posted:
02 September 2025
You are already at the latest version
Abstract
Shelter availability is a critical factor influencing the behaviour, welfare, and productivity of crayfish in aquaculture systems. This study examined the preference for four types of shelters among 0+ marron (Cherax cainii) juveniles under two light intensities: 1200 lux and 20 lux. These four distinct shelters were a sheet composed of radially rounded mesh (A), vertically layered mesh (B), PVC pipe (C), and a two-layered flat sheet on top of two PVC pipes (D). Two sets of experiments, one using a single juvenile and the second using two (paired) juvenile marron in separate tanks under laboratory conditions, were conducted across a triplicated design at the Curtin Aquatic Research Laboratory. Three variables, including shelter preference (%), total resident time (seconds) in shelters, and total search time (seconds) before residing in shelters, were determined through video surveillance. The results demonstrated a consistent selection for the rounded shelter, accounting for 47% and 59% by an individual marron, and 58% and 60% by two marron under 1200 and 20 lux, respectively. Light intensity was a significant factor influencing the total search time, as darker conditions reduced the duration spent by the marron searching for shelters. Agonistic interactions were observed when the second marron was present in the tanks, resulting in a decrease in the marron’s overall search time during their daily activity patterns. However, the total resident time remained unchanged after the marron occupied their preferred shelters, even when social interactions occurred.
Keywords:
marron
; shelters
; light intensity
; behaviour
; resident time
; search time
1. Introduction
Crustaceans, including freshwater crayfish, have been in high demand, and hence their economic value has significantly increased. [1]. Among the crayfish species in high demand are marron, which are one of the highest-market-value freshwater crayfish species in Western Australia [2]. Marron are primarily cultured in the south-west of Western Australia and exported to various countries [3, 4]. With a wide range of habitats, including freshwater springs, lakes, ponds, and rivers, they exhibit diverse life histories with a broad spectrum of behaviour patterns, including their intense activity in darkness [5, 6], and have developed sensory organs for movement and communication [7]. With a complex visual system, as compound eyes, helping them to rapidly detect movement and transmit images to the brain [8], however, the transmission of visual signals over long distances is affected by terrain barriers, availability and quality of ambient light, and light transmission conditions as turbidity [8].
Light as an environmental parameter is considered a stressor affecting the behaviour and physiology of freshwater crayfish [9]. The adaptations induced by light intensity or photoperiods on the biological rhythms of animals are previously known [10], suggesting the importance of this abiotic parameter. The effects of light conditions were studied when freshwater crawfish (Procambarus spp.) were challenged with different photoperiods, including a normal light regime (12L: 12D) and longer daytime (20L: 4D) conditions. The study concluded that the metabolic and behavioural parameters, such as lactate, oxygen consumption of the crayfish, were decreased at longer daytime periods [11]. Authors also observed a significant decrease in motor activities and a high rate of mortality under laboratory conditions, indicating photoperiods can lead to changes in metabolic and behavioural parameters of this species. In addition, crayfish interactions can alter under different light intensities, with high intensity potentially leading to a significant increase in their social interactions [12] or decreasing their aggressive behaviour [13]. Kozák, Martín Gallardo [14] indicated red swamp crayfish (P. clarkii) exhibited a preference for high light intensity (1000 lux), although they are nocturnal animals. However, in noble crayfish (Astacus astacus), anxious behaviour was observed as they showed a trend to avoid a strong light condition of 761 lux. This is considered the earlier preliminary signs of pain perception and neophobia in this species [15].
Shelters, as one of the cues, cause visual communication and are often related to elaborate courtship behaviours in crustaceans [16]. In crayfish aquaculture, density, light conditions, and shelter availability are the most important factors related directly to the mortalities due to agonistic behaviour and cannibalism. Cannibalism may be reduced by increasing the availability of shelters [13, 17-20]. Studies have investigated the impact of shelters on freshwater crayfish behaviour and growth performance under different habitats, including ponds and lakes, and explored the introduction of predators, suggesting that shelters play a crucial role in crayfish aquaculture, particularly during vulnerable stages of pre and post-moult [21-25]. By providing suitable and preferred shelters, a wide range of habitats can be utilised, and simultaneously reducing stress in crayfish [26].
Crustacean shelter preferences are influenced by various factors, such as sex, body size, and the presence of predators [19, 27]. For instance, signal crayfish (Pacifastacus leniusculus) exhibited distinct shelter size preferences between males and females [28]. Similarly, predator risk and conspecific density significantly impacted shelter selection in spiny lobsters (Panulirus argus) [29]. In addition, the shelter availability was found to influence social behaviour and habitat choice in virile crayfish (Orconectes virilis). The study revealed that shelter use and habitat selection in O. virilis are influenced by social status, with lower-ranking crayfish occupying shelters less frequently and preferring shelter-poor habitats, especially during the day [30]. Furthermore, dominance rank significantly impacts shelter use, acquisition, and eviction, with lower-ranking rusty crayfish (O. rusticus) forced to select smaller shelters and leave larger ones for the dominant crayfish [31].
Understanding the shelter preferences of marron is beneficial in improving animal care, which is also crucial from the perspective of ecological studies to comprehend their competition for shelters within the population. However, there is no research on the shelter preferences of marron under various light intensities to comprehend their behaviour. In this study, the shelter preferences of marron were investigated, and their behaviours were also described under two distinct light intensities, namely 1200 and 20 lux.
2. Materials and Methods
2.1. Experimental Design
Juvenile marron (0+) were purchased from the Marron Farm, Capel, Western Australia (33.58014° S, 115.58624° E) and transferred to the Curtin Aquatic Research Laboratory at Technology Park, Bentley, Western Australia, 6102. In a first trial, marron weighing 4.10 ± 0.04 g were used, while in the second trial, two marron weighing 6.63 ± 0.16 g were stocked in a 300L (100 cm x 40 cm) freshwater tank with continuous aeration. All marron were male.
Before the commencement of the experiment, marron kept in tanks under the natural photoperiod of 12L : 12D in the dark laboratory were fed daily with a basal feed (Crude protein 25%, Lipid 4%, Ash 2%), at a room temperature of 24°C for 2 weeks. Both trials were conducted in six tanks using two treatments with three replications per treatment, and lasted for 3 days. In the first trial, marron were exposed to two different light intensities of 1200 and 20 lux under a 24L: 0D cycle regime with four shelters in each tank.
Four types of handmade shelters were used, as shown in Figure 1, including:
(i) Radially rounded mesh shelter (A): This shelter was made from plastic mesh, comprising four pieces of 18 cm x 12 cm each. Each piece was initially folded in half, then folded again lengthwise. Finally, four mesh pieces were assembled and secured together using a stapler.
(ii) Vertically layered mesh shelter (B): Using the same mesh piece dimensions as shelter A, this design was created by folding a single mesh piece into six vertical layers.
(iii) PVC pipe (C): This shelter was made by a PVC pipe segment measuring 11 cm in length and 4.5 cm in diameter.
(iv) Two-layer flat sheet on top of two PVC pipes (D): This shelter was adapted from elevated sheets, but in a smaller size [22]. Each consisted of two flat plastic boards, 11 cm x 11 cm, attached to two PVC pipes, 11 cm in length and 3 cm in diameter each.
Smart light bulbs (Vanance by Zengge Co., Ltd., China) were positioned at the top of each tank, and the intensity was adjusted using a mobile application (Surplife, China). Before experimenting, the light intensity was measured using a handheld digital lux meter (Domary, China) positioned at the bottom of the tanks.
In the second trial, immediately following the first trial, two marron were used in six other tanks with two treatments with three replications per treatment. In this trial, similar to the first trial, marron were also exposed to two light intensities and eight shelters of four shelter types, two of each per tank. Surveillance cameras (3.0 MP Security camera, Taishixing, China) were installed on top of the tanks to record the occurrence of marron and then saved as MP4 extension files every 24 hours.
2.2. Data Collection and Analysis
Marron's behaviour was analysed by examining recorded videos. Three variables were analysed, including:
(i) Shelter preferences (%): For achieving this, all observations from day one to day three were counted by the number of times marron selected a shelter and then converted into percentages.
Percentage (%) = () x 100
(ii) The resident time (s): The time in seconds, marron spending in each shelter type, starting from the moment that two-thirds of their bodies were inside shelters.
(iii) The search time (s): The time in seconds taken to decide a shelter type, starting from the moment that marron were outside or left any shelters until one-third of their bodies were inside another shelter.
Both the resident and search time were presented as total, minimum, and maximum values with the mean ± standard errors (SE). The pattern of daily activity was characterised by adding up the total resident time and the total search time of each marron over 3 days.
Statistical analysis was conducted using the IBM SPSS Statistics version 30.0 (IBM Corporation, Chicago, IL, USA). All data showed a normal distribution by Sharpiro – Wilk test. The interactions between light intensities and shelter types were determined using a two-way ANOVA followed by a Tukey post hoc test. An independent t-test was performed to determine the differences between any two groups of variables if no interaction was found. All analyses were conducted at a significance level of 0.05.
3. Results
3.1. Shelter Preference
3.1.1. Trial 1 - One Marron with Four Shelters
There were interactions between light intensity and shelter types in marron preference (F=10.360, df=3, p<0.001). At 1200 lux, shelter A and B were the most preferred, accounting for 47.48 ± 6.05% and 39.60 ± 5.12% respectively, while at 20 lux, shelter A was the most preferred over the other three shelters, as shown in Figure 2. Marron rarely selected shelter C and D, which were a maximum of 16.43 ± 4.46% under dark conditions.
3.1.2. Trial 2 - Two Marron with Eight Shelters
There were no interactions between light intensity and shelter types in marron 1 (F=2.287, df=3, p=0.118) and marron 2 (F=1.357, df=3, p=0.291), but their preference was mainly affected by shelter types in marron 1 (F=12.602, df=3, p<0.001) and marron 2 (F=12.937, df=3, p<0.001), as shown in Figure 3. In this trial, shelter A was the most preferred and significantly different from shelters C and D, but not from shelter B. Shelter C and D were still the least preferred under both light conditions.
Besides, there was a significant difference in percentages at shelter D between marron 1 and 2 (t= -5.336, df=2, P=0.033).
At shelter C, combined data from three marron from both trials showed the shelter preferences of two marron were statistically different from those of a single marron (t=-3.714, df=7, P=0.008). Also at shelter D, percentages in all three marron were significantly different (F=10.844, df=2, p=0.010).
3.2. The Resident Time
3.2.1. Trial 1 - One Marron with Four Shelters
No interaction between light intensity and shelter types was found (F=0.551, df=3, p=0.654), but the type of shelters was the main factor affecting total resident time (F=6.724, df=3, p=0.004). The longest time was in shelter A by 119,448 seconds (33.18 hours) at 1200 lux and 183,021 seconds (50.84 hours) at 20 lux. Shelter A was a significant choice, which was statistically different than other shelters, shown in Table 1.
At 1200 lux, marron stayed longer in shelter A than others, with the longest time of 31,296 seconds (8.69 hours). The longest time for shelter B was 5,230 seconds (1.45 hours), shelter C was 9,010 seconds (2.50 hours), and shelter D was 8,115 seconds (2.25 hours). At 20 lux, the longest time marron stayed was also in shelter A as 183,021 seconds (50.84 hours), followed by shelter D as 55,971 seconds (15.55 hours), shelter C as 437.667 seconds (0.12 hours), and shelter B as 210 seconds (0.06 hours).
3.2.2. Trial 2 - Two Marron with Eight Shelters
There were no interactions between light intensity and shelter types in total resident time in marron 1 (F=0.404, df=3, p=0.752) and marron 2 (F=3.300, df=3, p=0.050) but shelter types played a main role in affecting marron 1 (F=15.765, df=3, p<0.001) and marron 2 (F=13.364, df=3, p<0.001). The longest total time (p<0.05) was in shelter A. The longest time in shelter A was 61,753 seconds (17.15 hours) by marron 1 and 80,891 seconds (22.47 hours) by marron 2, as described in Table 2, followed by shelter D as 38,579 seconds (10.72 hours) by marron 2, shelter B as 18,374 seconds (5.10 hours) by marron 1, and shelter C as from 1,114 to 1,203 seconds (0.31 to 0.33 hour).
When combining the data from two trials, the total resident time of a group of two marron was significantly different from that of a single marron in the first trial (t=-3.664, df=7, P=0.008) at shelter B only.
3.3. The search time
3.3.1. Trial 1 - One Marron with Four Shelters
There was an interaction between light intensity and shelter types in total search time (F=8.668, df=3, p=0.001). Total search time before choosing shelter A and B at 1200 lux was significantly different from the others, as shown in Table 3. At 1200 lux, the longest time for marron searching for shelter A was 6,406 seconds (1.78 hours), followed by shelter B as 4,056 seconds (1.12 hours), while at 20 lux, the longest time for all shelters was less than 310 seconds (0.09 hours).
3.3.2. Trial 2 - Two Marron with Eight Shelters
There were no interactions between light intensity and shelter types in total search time in marron 1 (F=1.110, df=3, p=0.374) and marron 2 (F=1.071, df=3, p=0.389). However, light intensity was found as the only main factor affecting this variable in marron 1 (F=6.550, df=3, p=0.021) and marron 2 (F=7.063, df=3, p=0.017). At 20 lux, total search time was significantly higher than that of both marron at 1200 lux (t=-2.109, df=21.942, p=0.047), as described in Table 4.
3.4. Daily Activity Pattern
3.4.1. Trial 1 - One Marron with Four Shelters
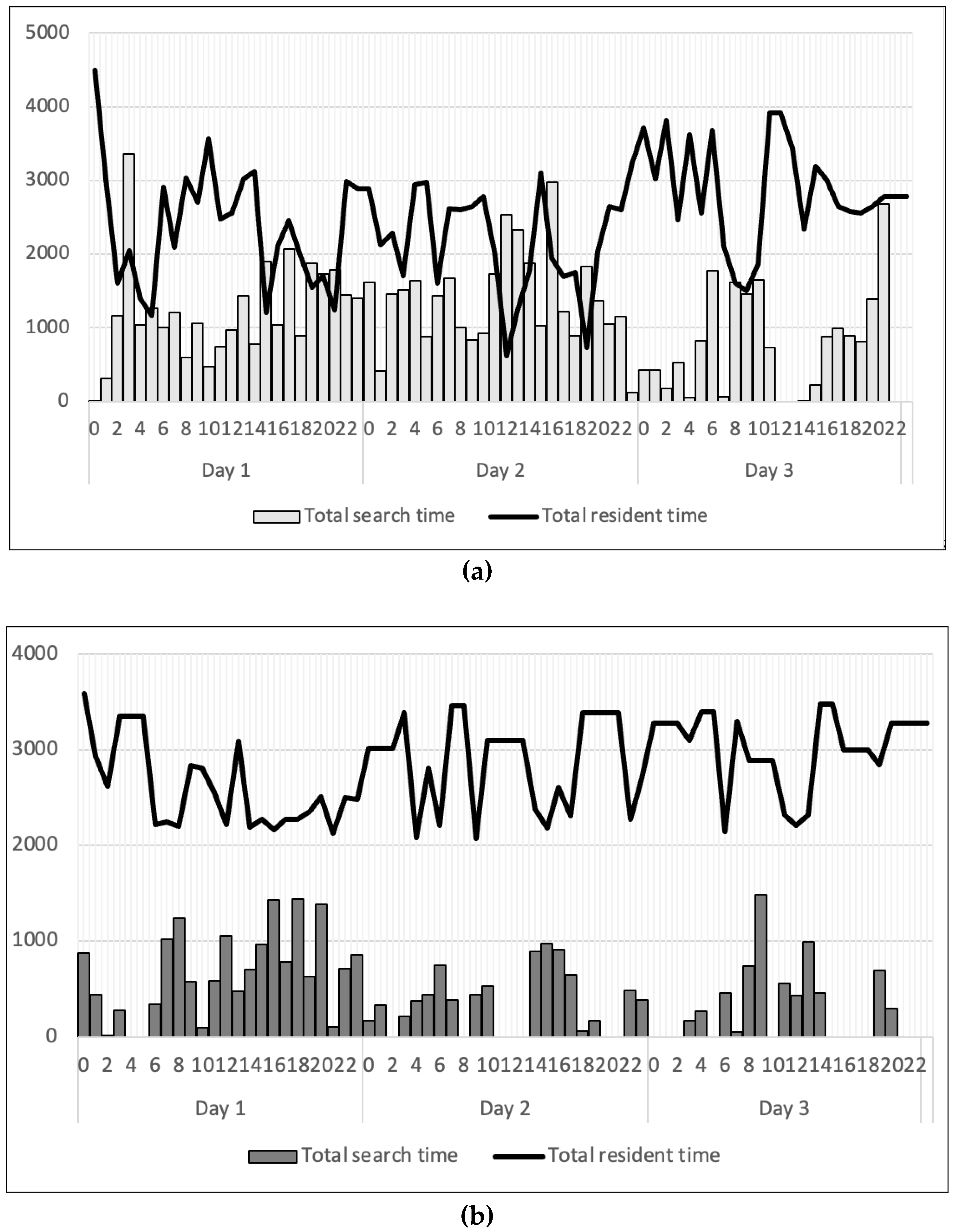
After exposure to light conditions, locomotion activities were observed, but most of their time was spent inside shelters. Under strong light conditions, marron actively walked around and explored all shelters with the longest total search time of 30 min. In the first hour, marron spent time inside shelters, then as hours passed, the time spent outside shelters increased gradually until 2 days. Marron became less active on day 3 as they spent more time on their refuges.
In darker conditions, their behaviour changed by decreasing total search time, as they also started to explore by walking around every shelter, staying inside for a few seconds, then moving to other shelters, but mostly they stayed inside a particular shelter for hours. The longest duration marron spent searching for shelters was only 14 min, which was half of those in brighter conditions, but the total resident time at 20 lux was higher than that at 1200 lux.
Figure 4.
Daily activity pattern of one marron under two different light intensities, including 1200 lux (a) and 20 lux (b).
Figure 4.
Daily activity pattern of one marron under two different light intensities, including 1200 lux (a) and 20 lux (b).
3.4.2. Trial 2 - Two Marron with Eight Shelters
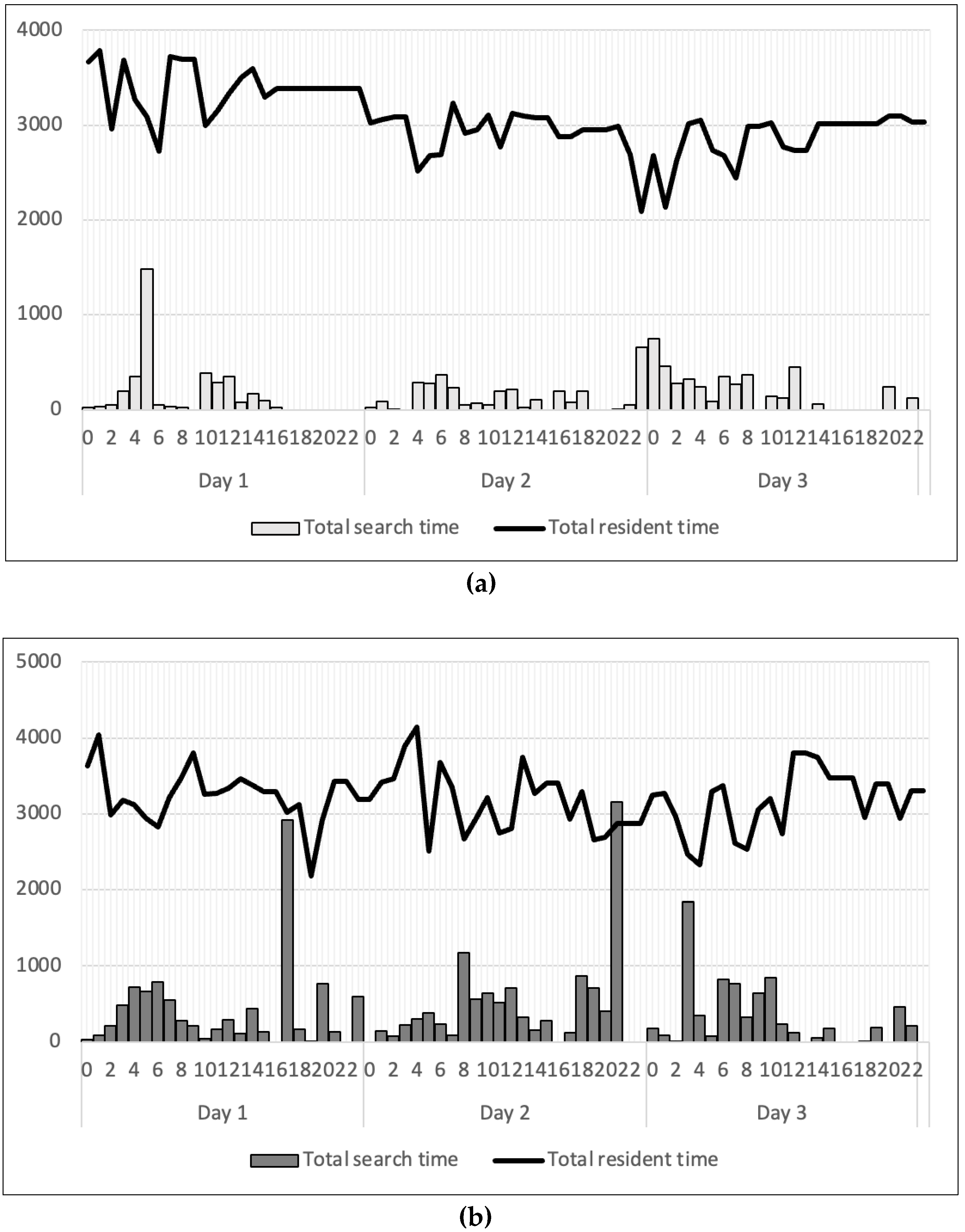
For the two-marron trial, social interactions occurred in the tanks under both light conditions. They chased each other around and fought to occupy their preferred shelters. Afterward, they remained in their dominated shelters. The longest search time was found after 5 hours of exposure under strong light. However, at weak light, there were instances of heightened activity, although they predominantly remained in protective shelters. No discernible pattern emerged when comparing the impact of different light intensity levels and shelter types on marron’s daily pattern under two light conditions.
Figure 5.
Daily activity pattern of two marron under two different light intensities, including 1200 lux (a) and 20 lux (b).
Figure 5.
Daily activity pattern of two marron under two different light intensities, including 1200 lux (a) and 20 lux (b).
4. Discussion
This is one of the first behavioural studies on the shelter preferences of marron under different light intensities. This study revealed that marron behaviour was influenced by two light intensities and shelter types. The marron preference was primarily dependent on shelter types, and then on the influence of light intensity. A single stand-alone marron showed its preferred choice for a radially rounded mesh shelter (shelter A) in two levels of light intensities. This type of shelter consisted of eight equally spaced compartments arranged around a central point and offered more space in terms of surface area, both at the top and bottom surfaces, so that marron had more flexibility in choosing the preferred area and easily concealing itself in any corner to evade light. Whereas, with three other shelters, they could expose themselves to any light conditions. The vertically layered mesh shelter (shelter B) was secondarily preferred in conditions of strong light, but not in darker conditions. This shelter shared the same material as the radially rounded mesh, but was vertically folded, allowing marron to conceal or hang in multiple angles over the folded areas, making it one of the top choices. PVC pipe (shelter C) and two-layered flat on top of two PVC pipes (shelter D) were constructed from PVC material and had a white colour, making it easy for marron to pass through them. It is considered that, in terms of labour cost for making these shelters, shelter D would be the most expensive compared to others, followed by shelter A, shelter B, and shelter C.
Currently, there is no available research on the effects of shelter colour on the marron behaviour, but some studies in other species, such as freshwater prawn (Macrobrachium nobilii), giant prawn (M. rosenbergii), and adult crayfish (O. rusticus), confirmed that dark shelters are preferred over light-coloured shelters [32, 33]. This might explain why shelters C and D were the least popular choices due to brighter colours. This remains unchanged when there was one additional marron in the tank in the second trial.
Due to the increased number of shelters, two marrons could independently select their preferred shelters, resulting in shelter A still being the most preferred for both, even in the same tank. A previous study of Verhoef and Austin [34] observed no interaction between density, shelter types, and availability in Australian freshwater crayfish, yabbies (Cherax destructor), revealing that even in a more intensive rearing system, the addition of shelters was not necessary. Shelters were reported to reduce aggressive activity then enhance the growth and survival rate of cultured crayfish [26, 35-39], but some farmers previously have recommended that shelters are of limited value and unnecessarily complicate harvesting [40]. In red claw crayfish (C. quadricarinatus), shelter types were independent of the survival rates of this species [41], which is consistent with Verhoef and Austin [34], who strongly supported that shelters had no effects or an inverse relationship between density, growth, and survival rates of C. destructor. In this study, shelter preferences were unchanged by the same size of two marron fed with same diet under laboratory conditions. This is in agreement with a study of Verhoef and Austin [34] when they concluded that under controlled indoor conditions, with a cohort of juvenile C. destructor of the same size and appropriate feed provided, shelter types and provisions are of little or no value. Also, dominance, but not residency status, was a factor in shelter use in O. rusticus [42], supporting these current findings, revealing that when suitable shelters are provided and sufficient, marron can select their preferred refuges, eventually leading to reduced aggressive activities.
The current finding demonstrates that at higher light intensity, behavioural response can be negated by providing shelters. Light conditions were previously confirmed to alter crayfish activities [38, 43], such as locomotion and agonistic interactions, which were modified after being challenged by a long period of light regime. Crayfish choose shelters via visual cues in light and tactile cues in dark conditions to assess shelter adequacy [27]. Under two light intensities from the current study, marron explored their tanks by walking around and selecting their preferred shelter, but they spent more of their daily time searching for shelters in stronger light. Jackson and Moore [12] discovered crayfish (Faxonious virilis and F. rusticus) exposed to higher light intensities exhibited different behavioural trends, supporting this current result that increased light intensity may lead to an increase in marron activities. Previously, Franke and Hörstgen-Schwark [44] pointed out the ‘light-on’ signal, causing increased locomotor activities of unsheltered crayfish, then they stopped moving as soon as a suitable shelter was found and occupied. The current finding corresponds with observations made by Franke and Hörstgen-Schwark [44] and Flint [45] when A. astacus and P. leniusculus became more disposed to search and stayed secluded inside shelters under low nocturnal light due to a significant inhibitory effect on the animals’ behavioural activities. This might explain the heightened total search time at stronger light and total resident time at weaker light in the current study. Although crayfish are classified as nocturnal animals, it does not mean that they are continuously active throughout the night, also not each and every night [44, 46]. However, there is limited available information regarding the daily activity of freshwater crayfish under various shelters and light conditions, making this research hard to compare with.
The antagonistic interaction between the two marron, which might modify their daily patterns, particularly a reduction in total search time during dark conditions, was evident in the current study. Dominance easily occurs in crayfish communities and is mainly dependent on the availability of a certain resource, including food or shelter [47]. Competition over a resource can lead to increased aggressive motivation and agonistic interaction [48]. Current findings have indicated that when adequate shelter resources were available, light intensities mainly contributed to modify marron’s total search time, supported by a study of Franke and Hörstgen-Schwark [44], wherein it was shown that the activity of noble crayfish (A. astacus) in a crowded culture system was mainly manipulated directly by light while their circadian system played a subordinate role, revealing that light can affect animals’ decision on being out or staying in their shelters [44]. Previously, Martin III and Moore [31] also demonstrated that dominance or hierarchy in crayfish plays a role in the acquisition of shelters. In comparison to this research, our study observed that the presence of a second marron can induce higher avoidance activities to get away from each other. Dominant marrons typically occupied their preferred shelters and evicted subdominant marrons through combat. After securing a shelter, dominant marrons spent most of their time inside, which can explain the reduced total search time during the dark conditions. Mathews [49] hypothesized that after the sheltering contest, the winning F. virilis would spend less time in shelters and explore more in the environment, while losers would spend more time inside shelters. However, the author’s outcome supported this current finding that after fighting, marron would engage in less risky behaviour by spending more time in their shelters instead of exploring the tanks more. Taken together, social interaction, in conjunction with light conditions, may be a contributing factor affecting the total search time of marron.
5. Conclusions
The marron exhibited a preference for the radially rounded mesh over vertically layered mesh, PVC pipe, and two-layer flat sheet on top of two PVC pipes. Marron's daily activities were influenced by light intensities only when they were searching for their preferred shelters. After occupying a particular shelter, marron spent most of their daily time inside their most preferred shelters, even under strong or weak light intensities. Agonistic interaction occurred in the presence of an additional marron in tanks, which contributed to altering their daily pattern by decreasing their search time but not changing their resident time.
Author Contributions
Conceptualization, R.F. and L.D.; methodology, R.F. and L.D.; software, L.D and M.R.; validation, R.F. and L.D.; formal analysis, L.D.; investigation, R.F. and L.D.; resources, R.F. and R.K.; data curation, R.F. and L.D.; writing—original draft preparation, L.D.; writing—review and editing, R.F.; visualization, R.F. and L.D.; supervision, R.F.; project administration, R.F. and R.K.; funding acquisition, R.F. All authors have read and agreed to the published version of the manuscript.
Funding
The described work is part of a doctorate of Luc Dang and was supported through the Vietnamese Government (Project 89) and a Top-up scholarship by Curtin University, Australia.
Institutional Review Board Statement
Ethical review and approval were waived for this study due to invertebrates (crustacean).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data are contained in this article.
Acknowledgments
We express our sincere gratitude to the Marron Growers Association (MGA) and Scott Bell, Vice President of the MGA, for generously providing marron juveniles for the experiment. Their insightful discussions on various challenges faced in contemporary aquaculture production have been invaluable to our research.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- FAO. The state of world fisheries and aquaculture 2024 - Blue transformation in action; FAO: Rome, Italy, 2024; p. 264. [Google Scholar]
- Alonso, A.D. Marron growing: a Western Australian rural niche market? Asia Pac. J. Mark. Logist. 2009, 21, 433–446. [Google Scholar] [CrossRef]
- Lawrence, C. , Improved performance of marron using genetic and pond management strategies. Fisheries Research Contract Report 2007, 17, 1–178. [Google Scholar]
- McClain, W.R. Crayfish aquaculture. Fish. Aquac 2020, 9, 259. [Google Scholar]
- Souty-Grosset, C.; et al. Atlas of crayfish in Europe; Muséum national d'Histoire naturelle Paris, 2006. [Google Scholar]
- McMahon, A.; Patullo, B.W.; Macmillan, D.L. Exploration in a T-Maze by the CrayfishCherax destructorSuggests Bilateral Comparison of Antennal Tactile Information. Biol. Bull. 2005, 208, 183–188. [Google Scholar] [CrossRef]
- Basil, J.; Sandeman, D. Crayfish (Cherax destructor) use Tactile Cues to Detect and Learn Topographical Changes in Their Environment. Ethology 2000, 106, 247–259. [Google Scholar] [CrossRef]
- Bradbury, J.W.; Vehrencamp, S.L. Principles of animal communication, Sinauer Associates: Sunderland, MA, 1998; Vol. 132.
- Aschoff, J. Freerunning and entrained circadian rhythms. In Biological rhythms; Springer, 1981; pp. 81–93. [Google Scholar]
- Prosser, C.L. Physiological variation in animals. Biological Reviews 1955, 30, 229–261. [Google Scholar] [CrossRef]
- Fanjul-Moles, M.a.L. , et al. Effect of variation in photoperiod and light intensity on oxygen consumption, lactate concentration and behavior in crayfish Procambarus clarkii and Procambarus digueti. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology 1998, 119, 263–269. [Google Scholar]
- Jackson, K.M.; Moore, P.A. The intensity and spectrum of artificial light at night alters crayfish interactions. Mar. Freshw. Behav. Physiol. 2019, 52, 131–150. [Google Scholar] [CrossRef]
- González, R.; Celada, J.D.; García, V.; Carral, J.M.; González, Á.; Sáez-Royuela, M. Shelter and lighting in the intensive rearing of juvenile crayfish (Pacifastacus leniusculus, Astacidae) from the onset of exogenous feeding. Aquac. Res. 2010, 42, 450–456. [Google Scholar] [CrossRef]
- Kozák, P.; Gallardo, J.M.; García, J.C.E. Light preferences of red swamp crayfish (Procambarus clarkii). Hydrobiologia 2009, 636, 499–503. [Google Scholar] [CrossRef]
- Abeel, T.; Vervaecke, H.; Roelant, E.; Platteaux, I.; Adriaen, J.; Durinck, G.; Meeus, W.; van de Perre, L.; Aerts, S. Evaluation of the influence of light conditions on crayfish welfare in intensive aquaculture. In Food futures: ethics, science and culture; Wageningen Academic: Leiden, The Netherlands, 2016; pp. 244–250. [Google Scholar]
- Díaz, E.R.; Thiel, M. Chemical and Visual Communication During Mate Searching in Rock Shrimp. Biol. Bull. 2004, 206, 134–143. [Google Scholar] [CrossRef]
- Mason, J.C. , Effects of temperature, photoperiod, substrate, and shelter on survival, growth, and biomass accumulation of juvenile Pacifastacus leniusculus in culture. Freshwater crayfish 1981, 4, 73–82. [Google Scholar]
- Savolainen, R.; Ruohonen, K.; Tulonen, J. Effects of bottom substrate and presence of shelter in experimental tanks on growth and survival of signal crayfish,Pacifastacus leniusculus(Dana) juveniles. Aquac. Res. 2003, 34, 289–297. [Google Scholar] [CrossRef]
- Figler, M.H.; Cheverton, H.M.; Blank, G.S. Shelter competition in juvenile red swamp crayfish (Procambarus clarkii): the influences of sex differences, relative size, and prior residence. Aquaculture 1999, 178, 63–75. [Google Scholar] [CrossRef]
- Wangpen, P. The role of shelter in Australian freshwater crayfish (Cherax spp.) polysystems. 2007, [Curtin University.
- Ratchford, S.G.; Eggleston, D.B. Temporal shift in the presence of a chemical cue contributes to a diel shift in sociality. Anim. Behav. 2000, 59, 793–799. [Google Scholar] [CrossRef]
- Jones, C.M.; Ruscoe, I.M. Assessment of Five Shelter Types in the Production of Redclaw Crayfish Cherax quadricarinatus (Decapoda: Parastacidae) Under Earthen Pond Conditions. J. World Aquac. Soc. 2001, 32, 41–52. [Google Scholar] [CrossRef]
- Taylor, P.; Elwood, R. The mismeasure of animal contests. Anim. Behav. 2003, 65, 1195–1202. [Google Scholar] [CrossRef]
- Tews, J.; Brose, U.; Grimm, V.; Tielbörger, K.; Wichmann, M.C.; Schwager, M.; Jeltsch, F. Animal species diversity driven by habitat heterogeneity/diversity: the importance of keystone structures. J. Biogeogr. 2003, 31, 79–92. [Google Scholar] [CrossRef]
- Meager, J.; Williamson, I.; Loneragan, N.; Vance, D. Habitat selection of juvenile banana prawns, Penaeus merguiensis de Man: Testing the roles of habitat structure, predators, light phase and prawn size. J. Exp. Mar. Biol. Ecol. 2005, 324, 89–98. [Google Scholar] [CrossRef]
- Steele, C.; Skinner, C.; Alberstadt, P.; Antonelli, J. Short communication: Importance of adequate shelters for crayfishes maintained in aquaria. Aquar. Sci. Conserv. 1997, 1, 189–192. [Google Scholar] [CrossRef]
- Takahashi, K.; Nagayama, T. Shelter preference in the Marmorkrebs (marbled crayfish). Behaviour 2016, 153, 1913–1930. [Google Scholar] [CrossRef]
- Ranta, E.; Lindström, K. Body size and shelter possession in mature signal crayfish, Pacifastacus leniusculus. Annales Zoologici Fennici 1993, 30, 125–132. [Google Scholar]
- Eggleston, D.B.; Lipcius, R.N. Shelter Selection by Spiny Lobster Under Variable Predation Risk, Social Conditions, and Shelter Size. Ecology 1992, 73, 992–1011. [Google Scholar] [CrossRef]
- Fero, K.C.; Moore, P.A. Shelter availability influences social behavior and habitat choice in crayfish, Orconectes virilis. Behaviour 2014, 151, 103–123. [Google Scholar] [CrossRef]
- Martin, A.L.; Moore, P.A. The Influence of Dominance on Shelter Preference and Eviction Rates in the Crayfish, Orconectes rusticus. Ethology 2008, 114, 351–360. [Google Scholar] [CrossRef]
- Mariappan, P.; Balasundaram, C. Sheltering behaviour of Macrobrachium nobilii (Henderson and Matthai, 1910). Acta Ethologica 2003, 5, 89–94. [Google Scholar] [CrossRef]
- Kawamura, G.; Bagarinao, T.; Yong, A.S.K.; Fen, T.C.; Lim, L.S. Shelter colour preference of the postlarvae of the giant freshwater prawn Macrobrachium rosenbergii. Fish. Sci. 2017, 83, 259–264. [Google Scholar] [CrossRef]
- Verhoef, G.; Austin, C. Combined effects of shelter and density on the growth and survival of juveniles of the Australian freshwater crayfish, Cherax destructor Clark. Aquaculture 1999, 170, 49–57. [Google Scholar] [CrossRef]
- He, J.; Shi, H.; Xu, W.; Su, Z. Research progress on the cannibalistic behavior of Aquatic Animals and The Screening of Cannibalism-Preventing Shelters (Review). Israeli Journal of Aquaculture-Bamidgeh 2020, 72, 21014. [Google Scholar] [CrossRef]
- Capelli, G.M.; Hamilton, P.A. Effects of Food and Shelter on Aggressive Activity in the Crayfish Orconectes rusticus (Girard). J. Crustac. Biol. 1984, 4, 252–260. [Google Scholar] [CrossRef]
- Westin, L.; Gydemo, R. The locomotor activity patterns of juvenile noble crayfish (Astacus astacus) and the effect of shelter availability. Aquaculture 1988, 68, 361–367. [Google Scholar] [CrossRef]
- Franke, R.; Hörstgen-Schwark, G. Influence of social factors on the nocturnal activity pattern of the noble crayfish,Astacus astacus(Crustacea, Decapoda) in recirculating aquaculture systems. Aquac. Res. 2014, 46, 2929–2937. [Google Scholar] [CrossRef]
- Song, G. , et al. Effects of broodstock sizes, shelter, illumination and stocking density on breeding in red swamp crayfish Procambarus clarkii. Fisheries Science (Dalian) 2012, 31, 549–553. [Google Scholar]
- Fielder, D.; Thorne, M. Are shelters really necessary; Australian fisheries: aquaculture special: redclaw, Macreadie, M. (ed); Australian Government Publishing Service: Canberra, 1990; pp. 26–28. [Google Scholar]
- Jones, C.M. Production of juvenile redclaw crayfish, Cherax quadricarinatus (von Martens) (Decapoda, Parastacidae) I. Development of hatchery and nursery procedures. Aquaculture 1995, 138, 221–238. [Google Scholar] [CrossRef]
- Klar, N.M.; Crowley, P.H. Shelter Availability, Occupancy, and Residency in Size-Asymmetric Contests Between Rusty Crayfish,Orconectes rusticus. Ethology 2011, 118, 118–126. [Google Scholar] [CrossRef]
- Luna, A.J.F.; Hurtado-Zavala, J.I.; Reischig, T.; Heinrich, R. Circadian Regulation of Agonistic Behavior in Groups of Parthenogenetic Marbled Crayfish, Procambarus sp. J. Biol. Rhythm. 2009, 24, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Franke, R.; Hörstgen-Schwark, G. Control of activity patterns in crowded groups of male noble crayfish Astacus astacus (Crustacea, Astacidea) by light regimes: A way to increase the efficiency of crayfish production? Aquaculture 2015, 446, 103–110. [Google Scholar] [CrossRef]
- Flint, R.W. Seasonal Activity, Migration and Distribution of the Crayfish, Pacifastacus Ieniusculus, in Lake Tahoe. Am. Midl. Nat. 1977, 97, 280–292. [Google Scholar] [CrossRef]
- Hazlett, B.; Rittschof, D.; Ameyaw-Akumfi, C. Factors affecting the daily movements of the crayfish Orconectes virilis (Hagen, 1870) (Decapoda, Cambaridae), in Studies on Decapoda. 1979, Brill. pp. 121-130.
- Francis, R.C. On the Relationship between Aggression and Social Dominance. Ethology 1988, 78, 223–237. [Google Scholar] [CrossRef]
- Cioni, A.; Gherardi, F. Agonism and interference competition in freshwater decapods. Behaviour 2004, 141, 1297–1324. [Google Scholar] [CrossRef]
- Mathews, L. Outcomes of agonistic interactions alter sheltering behavior in crayfish. Behav. Process. 2021, 184, 104337. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Four types of shelters in the experiment, including Radially rounded mesh (A), Vertical layered mesh (B), PVC pipe (C), and Two-layer flat sheet on top of two PVC pipes (D).
Figure 1.
Four types of shelters in the experiment, including Radially rounded mesh (A), Vertical layered mesh (B), PVC pipe (C), and Two-layer flat sheet on top of two PVC pipes (D).

Figure 2.
Shelter preferences in percentages of single marron at two different light intensities: (a) – 1200 lux and (b) – 20 lux.
Figure 2.
Shelter preferences in percentages of single marron at two different light intensities: (a) – 1200 lux and (b) – 20 lux.
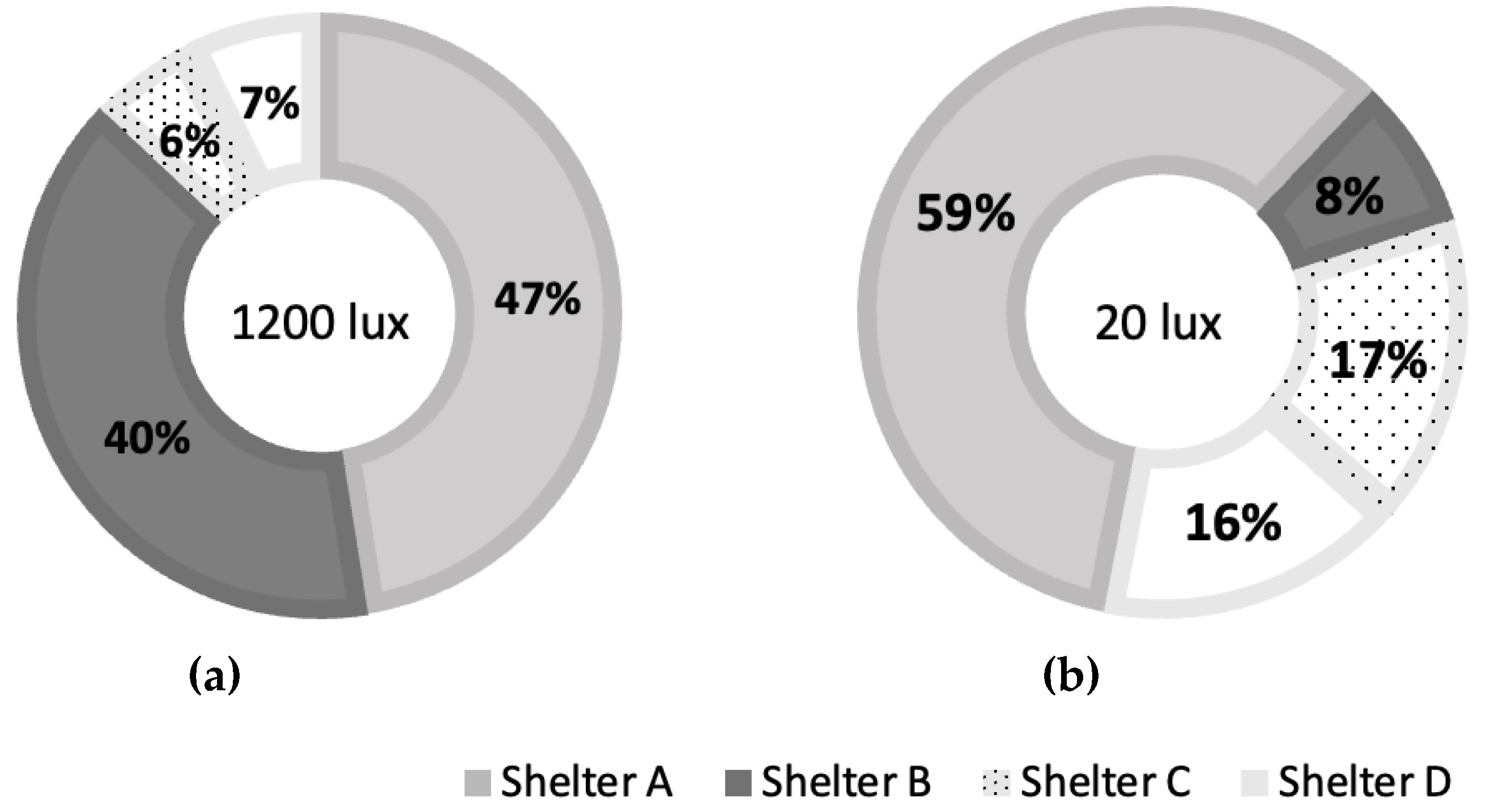
Figure 3.
Shelter preferences in percentages of two marron at two different light intensities; (a) - Marron 1 and (b) - Marron 2.
Figure 3.
Shelter preferences in percentages of two marron at two different light intensities; (a) - Marron 1 and (b) - Marron 2.
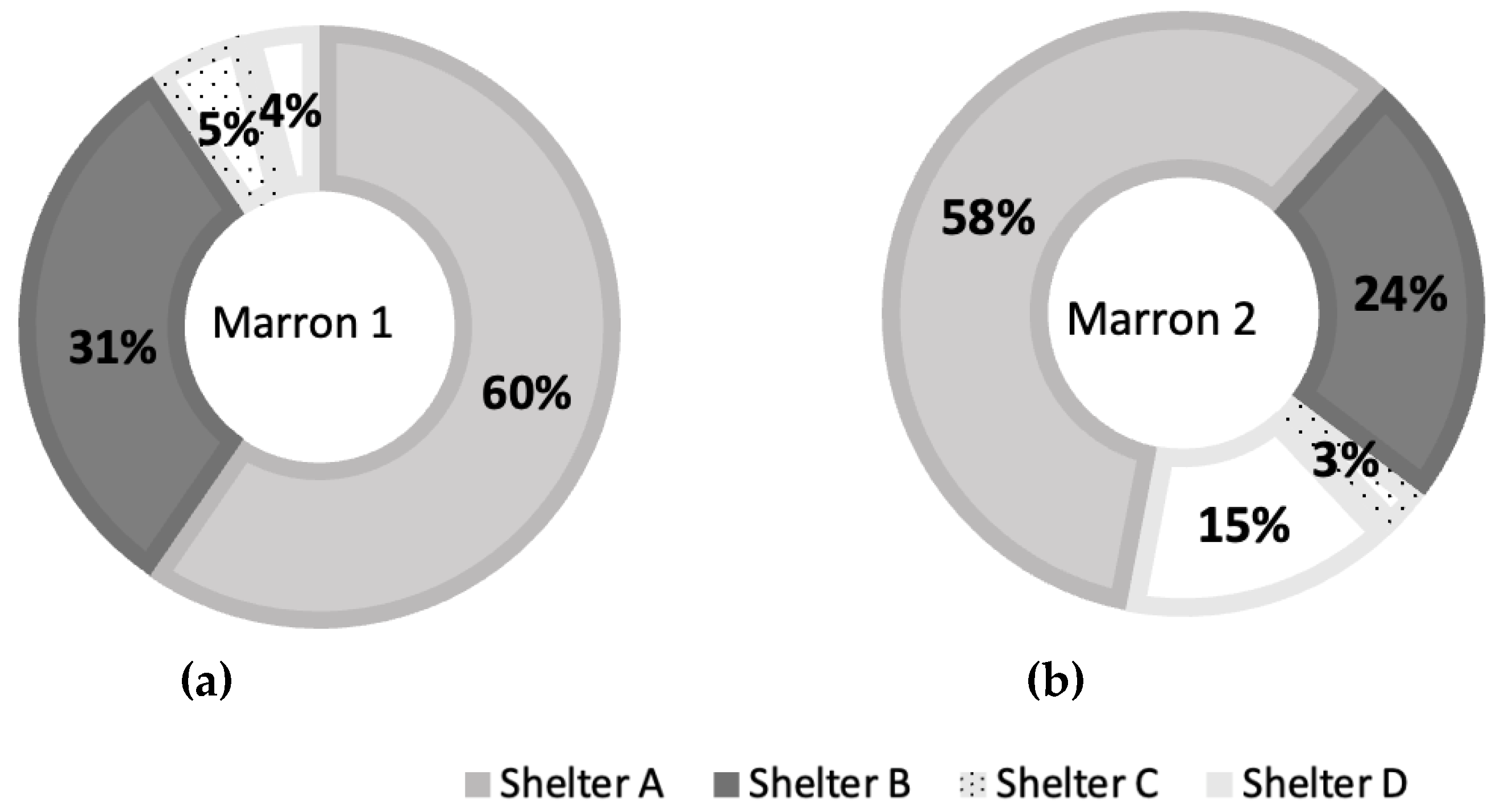

Table 1.
The resident time (Mean ± SE) in shelters of one marron under different light intensities.
| The resident time (s) | Shelter types | |||
|---|---|---|---|---|
| A | B | C | D | |
| 1200 lux | ||||
| Total time | 119,448 ± 11,251 | 23,342 ± 12,133 | 9,867 ± 8,556 | 13,273 ± 6,660 |
| Min | 42.67 ± 14.38 | 52.00 ± 17.04 | 83.33 ± 38.63 | 70.00 ± 25.17 |
| Max | 31,296 ± 11,251 | 5,230 ± 3,542 | 9,010 ± 8,462 | 8,115 ± 4,479 |
| 20 lux | ||||
| Total time | 183,021 ± 88,760 | 210.00 ± 152.27 | 437.67 ± 314.05 | 55,971 ± 54,121 |
| Min | 5,351 ± 5,258 | 190.67 ± 158.81 | ||
| Max | 83,121 ± 43,339 | |||
| Grouped by statistical difference | * |
 ** |
||
Table 2.
The resident time (Mean ± SE) in shelters of two marron.
| The resident time (s) | Shelter types | |||
|---|---|---|---|---|
| A | B | C | D | |
| Marron 1 | ||||
| Total time | 182,714 ± 24,558 | 40,174 ± 34,390 | 1,594 ± 1,028 | 5,247 ± 4,939 |
| Min | 2,994 ± 2,921 | 53.83 ± 14.90 | 473.67 ± 252.76 | 192.83 ± 117.18 |
| Max | 61,753 ± 11,305 | 18,374 ± 17,169 | 1,203 ± 752.43 | 1,669 ± 1,556 |
| Marron 2 | ||||
| Total time | 152,269 ± 23,272 | 23,896 ± 15,372 | 1,577 ± 472.46 | 62,853 ± 20,027 |
| Min | 7,019 ± 4,694 | 672.17 ± 413.70 | 578.50 ± 493.50 | 118.17 ± 15.21 |
| Max | 80,891 ± 19,584 | 18.37 ± 14,534 | 1,114 ± 400.25 | 38,579 ± 22,285 |
| Grouped by statistical difference | * |
** |
||
Table 3.
The search time (Mean ± SE) for shelters of one marron under different light intensities.
| Shelter | The search time (s) | ||
|---|---|---|---|
| Total time | Min | Max | |
| 1200 lux | |||
| A | 47,092 ± 14,025a | 4.67 ± 0.33 | 6,406 ± 726.670 |
| B | 26,974 ± 2,054a | 20.67 ± 7.33 | 4,056 ± 1,275 |
| C | 2,243 ± 485.35b | 23.67 ± 9.96 | 1,446 ± 402.78 |
| D | 5,500 ± 5,318b | 17.67 ± 5.61 | 2,693 ± 2,613 |
| 20 lux | |||
| A | 308.67 ± 178.91b | 13.00 ± 3.79 | 152.00 ± 71.65 |
| B | 52.00 ± 26.91b | 52.00 ± 26.91 | |
| C | 57.00 ± 28.58b | 57.00 ± 28.58 | |
| D | 306.00 ± 255.32b | 306.00 ± 255.32 | |
ab Different superscripts represent a significant difference between four shelters and two light intensities (p<0.05).
Table 4.
Total search time (Mean ± SE) for shelters of two marron under different light intensities.
Table 4.
Total search time (Mean ± SE) for shelters of two marron under different light intensities.
| The search time (s) | Light intensity | |||
|---|---|---|---|---|
| 1200 lux | 20 lux | |||
| Marron 1 | Marron 2 | Marron 1 | Marron 2 | |
| Total | 4,315 ± 4,103a | 1,646 ± 1,522* | 10,727 ± 2,788b | 7,198 ± 3,403** |
| Min | 16.75 ± 11.16 | 10.67 ± 4.69 | 25.17 ± 6.81 | 20.75 ± 2.43 |
| Max | 1,300 ± 1,198 | 424.42 ± 361.81 | 2,733 ± 405.63 | 1,868 ± 283.81 |
ab,* ** Different superscripts represent a significant difference between two groups of marron (p <0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



