Submitted:
29 August 2025
Posted:
01 September 2025
You are already at the latest version
Abstract
The molecular mechanism behind the loss of neuro-melanin-containing dopaminergic neurons in the nigrostriatal sys-tem in idiopathic Parkinson’s disease remains unclear. However, research suggests that aminochrome, an endogenous neurotoxin, may trigger the degeneration of these neurons through a sin-gle-neuron degeneration model. In this model, aminochrome selec-tively destroys individual neurons without spreading to neighbor-ing cells. Aminochrome is produced during neuromelanin synthe-sis, a process that is normally harmless because protective enzymes like DT-diaphorase and glutathione transferase M2-2 neutralize aminochrome’s neurotoxic effects. Increasing the levels of these enzymes could offer neuroprotection. The KEAP1/NRF2 signaling pathway plays a crucial role in regulating antioxidant enzymes, including DT-diaphorase and glutathione transferase M2-2. Nota-bly, certain dietary bioactive compounds can activate the KEAP1/NRF2 pathway, enhancing the production of these protec-tive enzymes. For example, the omega-3 fatty acids eicosapentae-noic acid and docosahexaenoic acid, as well as astaxan-thin—found in cold-water fish like salmon—have been shown to boost enzyme expression. This raises an important question: Could dietary interventions help prevent idiopathic Parkinson’s disease?
Keywords:
neuroprotection
; neurodegeneration
; KEAP1/NRF2 pathway
; aminochrome
; dopamine
; neuromelanin
; natural compounds
; Omega-3
; DT-diaphorase
; glutathione transferase M2-2
1. Parkinson´s Disease
Parkinson’s disease is the second most common neurodegenerative disorder, characterized by the loss of dopamine-producing, neuromelanin-containing neurons in the nigrostriatal system. Most patients (70%) are diagnosed with idiopathic Parkinson’s disease, which primarily affects individuals aged 55–60 and older. Since 1967, the standard treatment has been L-dopa, which greatly improves motor function and helps patients maintain a near-normal life. However, after 4 to 6 years of treatment, side effects like dyskinesia often develop, significantly reducing patients' quality of life. Despite extensive research, no new drugs have successfully slowed or stopped disease progression. Preclinical studies on compounds such as coenzyme Q, mitoquinone, urate, deferiprone, TCH346, and neurturin have shown promise, but these benefits haven’t carried over into clinical trials [1,2,3,4,5]. The failure of these trials has been attributed to flaws in trial design and the lack of reliable biomarkers [6,7,8].
However, we believe the failure of these clinical trials can be attributed to two key factors:
(i) The use of preclinical models that poorly replicate the disease process. These models rely on exogenous neurotoxins, which induce an extremely rapid and widespread degenerative process [9,10]. This sudden, aggressive degeneration seen in preclinical neurotoxin models sharply contrasts with the slow progression of idiopathic Parkinson’s disease, both before and after motor symptoms appear. The most common preclinical models for testing Parkinson’s drugs—1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) and 6-hydroxydopamine—produce effects that are inconsistent with the natural course of the disease. For example, MPTP can induce severe parkinsonism in just three days in individuals exposed to contaminated drugs [11], whereas idiopathic Parkinson’s develops over many years, with neurodegeneration progressing gradually long before and after motor symptoms emerge. It is unlikely that a drug effective in these rapid, extreme neurotoxin models would translate to patients with idiopathic Parkinson’s, where degeneration occurs at a far slower pace.
(ii) The lack of a methodology capable of detecting subtle degenerative changes. Parkinson’s disease may follow a single-neuron degeneration model [12], consistent with its gradual progression before and after symptom onset. This raises the question of whether current tools, such as the Unified Parkinson’s Disease Rating Scale (UPDRS), are sensitive enough to measure these minute, incremental changes in neurodegeneration over time.
To this day, the exact triggers that initiate the loss of dopamine-producing neurons containing neuromelanin in the nigrostriatal system in idiopathic Parkinson’s disease remain unknown. However, scientists widely agree that multiple factors contribute to this process. These include mitochondrial dysfunction, oxidative stress, the formation of toxic alpha-synuclein clumps, disruptions in both proteasomal and lysosomal protein-clearing systems, endoplasmic reticulum stress, and neuroinflammation [13,14,15,16,17,18,19,20].
2. Single-Neuron Degeneration
Research suggests that the neurodegeneration affecting neuromelanin-containing dopaminergic neurons in the nigrostriatal system follows a single-neuron degeneration model [12]. According to this model, the neurotoxin responsible for triggering mechanisms like mitochondrial dysfunction, oxidative stress, neurotoxic alpha-synuclein oligomer formation, impaired proteasomal and lysosomal degradation, endoplasmic reticulum stress, and neuroinflammation originates within the neuron itself and does not spread to nearby cells. This model explains why the degenerative process is so slow, taking years before motor symptoms emerge and continuing gradually throughout the disease. Patients typically survive 10 to 20 years after diagnosis before succumbing to the condition. A recent study estimates that the total number of dopaminergic neurons in the substantia nigra (across both hemispheres) ranges from 800,000 to 1,000,000 [21]. For a patient who lives 15 years after motor symptoms begin—by which point 60% of neuromelanin-containing neurons are lost—this translates to a loss of 58 to 73 neurons per day. Such a slow progression can only occur if an endogenous neurotoxin selectively destroys neurons one at a time without affecting neighboring cells. Over time, the cumulative loss of these neurons eventually reaches a threshold where symptoms develop.
3. Endogenous Neurotoxins in Parkinson´s Disease
Potential endogenous neurotoxins in a single-neuron degeneration model include alpha-synuclein, 3,4-dihydroxyphenylacetaldehyde (DOPAL), and aminochrome. Mutations in alpha-synuclein that generate neurotoxic oligomers have been associated with familial Parkinson’s disease, accounting for roughly 10% of all Parkinson’s cases. However, in idiopathic Parkinson’s disease—which makes up 70% of cases—alpha-synuclein’s neurotoxicity depends on another neurotoxin to trigger the formation of these harmful oligomers. Studies indicate that aminochrome can induce the creation of such neurotoxic oligomers [20]. Furthermore, alpha-synuclein has been shown to cause widespread neurotoxicity, affecting nearby neurons [22,23,24,25,26,27]. Because of this, alpha-synuclein cannot be the sole endogenous neurotoxin responsible for initiating single-neuron degeneration.
During the oxidative deamination of dopamine—a reaction catalyzed by monoamine oxidase (MAO)—ammonia, hydrogen peroxide, and 3,4-dihydroxyphenylacetaldehyde (DOPAL) are produced. DOPAL is subsequently converted into 3,4-dihydroxyphenylacetic acid (DOPAC) by the enzyme aldehyde dehydrogenase-1 (ALDH1) [28]. Studies analyzing postmortem brain tissue from Parkinson’s disease (PD) patients found reduced ALDH1 expression, suggesting that DOPAL accumulation may contribute to the degeneration of the nigrostriatal system [29]. However, postmortem tissue does not exclusively reflect the vulnerable neurons where neurodegeneration occurs. Furthermore, DOPAL exhibits intercellular spread, as it has been shown to transfer from neurons to glial cells [30].
Aminochrome is an endogenous neurotoxin produced during neuromelanin synthesis. The catechol group of dopamine undergoes oxidation, generating three ortho-quinones in a sequential process: dopamine ortho-quinone, aminochrome, and 5,6-indolequinone. Among these, aminochrome is the most stable ortho-quinone and is known to induce mitochondrial dysfunction, neurotoxic oligomer formation, oxidative stress, disruption of proteasomal and lysosomal protein degradation systems, endoplasmic reticulum stress, and neuroinflammation. Although aminochrome is a transient neurotoxin, it remains stable for approximately 40 minutes in in vitro experiments before converting to 5,6-indolequinone [31]. When aminochrome forms in the cytosol of a neuron during neuromelanin synthesis, it is quickly neutralized by flavoenzymes that transfer one or two electrons or form protein adducts. As a result, it cannot be exported to affect nearby neurons, limiting its neurotoxic effects to the neuron where it originates. These properties suggest that aminochrome may drive a single-neuron degeneration model, in which dopaminergic neurons containing neuromelanin are gradually lost over many years.
4. Why Can Aminochrome be Neurotoxic During Neuromelanin Synthesis?
There seems to be a contradiction regarding the neurotoxic effects of aminochrome, which is produced during neuromelanin synthesis. Normally, neuromelanin synthesis is a harmless process—healthy elderly individuals often retain intact, neuromelanin-containing dopaminergic neurons at death [32,33,34]. This raises a key question: Why don’t healthy people experience aminochrome’s neurotoxicity during neuromelanin synthesis? The answer lies in two critical enzymes that neutralize aminochrome’s harmful effects.
- (i)
- DT-Diaphorase – DT-diaphorase (NAD(P)H:quinone oxidoreductase; NQO1; EC 1.6.99.2) is a distinct flavoenzyme that catalyzes the two-electron reduction of quinones to hydroquinones [35]. Unlike other flavoenzymes—which use NADH or NADPH as electron donors to drive the one-electron reduction of quinones, producing highly reactive semiquinones—DT-diaphorase avoids semiquinone formation entirely. Semiquinones react with oxygen, generating superoxide and contributing to oxidative stress. For instance, NADPH-cytochrome P450 reductase catalyzes the reduction of aminochrome to leukoaminochrome radicals, which are highly reactive with oxygen [36]. In contrast, DT-diaphorase facilitates the two-electron reduction of aminochrome directly to leukoaminochrome [37]. Inhibition of DT-diaphorase via siRNA has been demonstrated to trigger cell death in catecholaminergic cell cultures [38]. DT-diaphorase is expressed in multiple organs, including the brain, with notable activity in the substantia nigra, striatum, hypothalamus, hippocampus, and cerebral cortex. It represents 97% of total quinone reductase activity (which includes other NADH/NADPH-dependent flavoenzymes) and is present in both dopaminergic neurons and astrocytes [39]. DT-diaphorase provides protection against: aminochrome-induced cell death, formation of neurotoxic α-synuclein oligomers, mitochondrial dysfunction, oxidative stress, autophagy and lysosomal dysfunction, disruption of cytoskeletal architecture [38,40,41,42,43,44,45,46,47,48,49,50,51,52].
- (ii)
- Glutathione transferase M2-2 – This enzyme catalyzes the conjugation of aminochrome with glutathione, forming 4-S-glutathionyl-5,6-dihydroxyindoline, a compound resistant to biological oxidizing agents such as superoxide, hydrogen peroxide, and dioxygen [53,54]. Glutathione transferase M2-2 also conjugates dopamine ortho-quinone (a precursor of aminochrome) to produce 5-glutathionyldopamine, which is typically metabolized into 5-cysteinyldopamine [55]. The detection of 5-cysteinyldopamine in human cerebrospinal fluid and neuromelanin suggests it is a stable end product, supporting its potential neuroprotective role. Notably, while glutathione transferase M2-2 is predominantly expressed in astrocytes, these cells secrete exosomes containing the enzyme, which then enter dopaminergic neurons and release the enzyme into their cytosol. This mechanism implies that astrocytes contribute to neuroprotection by boosting the defensive capacity of DT-diaphorase in neuromelanin-containing dopaminergic neurons [56,57,58,59].
The combined neuroprotective effects of DT-diaphorase and glutathione transferase M2-2 play a key role in preventing aminochrome-induced neurotoxicity during neuromelanin synthesis. However, decreased expression of these enzymes—along with excessive dopamine production and a resulting rise in aminochrome levels that overwhelms their protective capacity—may explain why neuromelanin-containing dopaminergic neurons are lost in Parkinson’s disease (Figure 1).
5. Natural Bioactive Compounds that Trigger Neuroprotection in Dopaminergic Neurons Containing Neuromelanin
The proposed neuroprotective role of DT-diaphorase and glutathione transferase M2-2 suggests that higher levels of these enzymes may be key in preventing neurotoxic effects on neuromelanin synthesis in idiopathic Parkinson’s disease. The increased expression of these antioxidant enzymes is controlled by the KEAP1/NRF2 signaling pathway, which includes DT-diaphorase and glutathione transferase M2-2 [60]. Many natural compounds—such as curcumin, hyperoside, resveratrol, quercetin, sulforaphane, and safranal—have been shown to activate the KEAP1/NRF2 pathway [61]. However, not all studies on KEAP1/NRF2 activation demonstrate increased expression of DT-diaphorase and glutathione transferase M2-2. Since this pathway regulates multiple antioxidant enzymes, researchers often focus on measuring just one or two specific enzymes relevant to their study goals (Figure 2).
6. Bioactive Compounds in Food
Natural foods not only provide proteins, fats, and glucose to produce the energy the body needs to function, but they also contain a variety of essential minerals and vitamins that support critical physiological processes. For example, iron is crucial for oxygen transport, while many enzymes and transporters depend on small molecules like magnesium, selenium, copper, and iron as cofactors. Animal-based foods deliver key nutrients such as vitamin B12, vitamin D, iron, zinc, and calcium, while seafood supplies essential fatty acids like eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA). In contrast, vegetables and fruits are rich in vitamins A, B, C, and K, folate, and minerals such as magnesium, potassium, and non-heme iron. They also contain beneficial plant compounds, including phytochemicals, flavonoids, phenolic acids, and carotenoids [62,63].
Research suggests that plant-derived neuroprotective compounds could help prevent neurodegenerative diseases. For example, nobiletin—a polymethoxylated flavone from Citrus depressa peel—has been shown to improve cognitive and motor deficits in preclinical Parkinson’s disease models [64]. Similarly, tangeretin, a citrus flavonoid found in the peel and other parts of Citrus L. plants, exhibits neuroprotective effects in MPTP- and MPP+-induced Parkinson’s models [65]. Other compounds, such as iridoids (geniposide, harpagoside, catalpol, and 10-O-trans-p-coumaroylcatalpol), have demonstrated neuroprotective activity in Parkinson’s models by boosting antioxidant enzymes (e.g., glutathione peroxidase and superoxide dismutase) and increasing tyrosine hydroxylase-positive neurons [66]. Polydatin, a natural compound in peanuts, grapes, and red wine, has shown neuroprotective effects by suppressing microglia activation and reducing pro-inflammatory factors in a lipopolysaccharide-induced Parkinson’s model [67]. Additionally, caffeic acid—a natural phenol in argan oil—has been found to protect dopaminergic neurons, enhance autophagy, and reduce alpha-synuclein aggregation in the substantia nigra of A53T alpha-synuclein transgenic models [68].
Chicoric acid, a polyphenol found in chicory and purple coneflower, has been shown to prevent MPTP-induced motor dysfunction, overactivation of glial cells, and the loss of dopaminergic neurons [69]. Morin, a flavonol present in wine and fruits, has been found to reduce motor dysfunction, protect dopaminergic neurons in the substantia nigra and striatum, and decrease astrocyte activation in an MPTP-induced mouse model. In primary cultures treated with MPP+, Morin demonstrated neuroprotective effects by lowering reactive oxygen species (ROS) production, preserving mitochondrial membrane potential, and inhibiting astroglial activation [70]. Wolfberry (the fruit of Lycium barbarum L.) has shown neuroprotective properties in multiple preclinical Parkinson’s disease models, including 6-hydroxydopamine-treated rats, MPTP-treated mice, and α-synuclein A53T mice. It helped alleviate motor deficits and prevented dopaminergic neuron loss by regulating iron metabolism [71]. In another study, extracts from Vicia faba L. sprouts increased dopamine levels in the striatum, improved motor function, reduced inflammatory markers, and lowered malondialdehyde levels in a rotenone-treated mouse model [72]. Additionally, research using MPTP-treated animal models and cell cultures found that the alkaloid N-methylene-(5,7,4-trihydroxy)-isoflavone, derived from Sophora alopecuroides L. fruits, reduced motor deficits, oxidative stress, neuroinflammation, and dopaminergic neuron loss in both the striatum and substantia nigra [73].
Hericium erinaceus, a medicinal mushroom, has demonstrated neuroprotective effects in neurodegenerative diseases like Parkinson’s. Its benefits are tied to boosting the production of neurotrophic factors [74,75]. Studies in rats have shown that Cinnamomum osmophloeum Kanehira extract increases dopamine and tyrosine hydroxylase levels while reducing alpha-synuclein buildup in the striatum. In the midbrain, it also enhances antioxidant enzymes like superoxide dismutase, catalase, and glutathione peroxidase [76]. Additionally, two neuroactive β-carbolines in coffee provide neuroprotective, antioxidant, and anti-inflammatory effects, potentially lowering Parkinson’s risk [77]. Another compound, nobiletin—a polymethoxylated flavone found in Citrus depressa peel—has been shown in animal models to improve both motor and cognitive deficits linked to Parkinson’s [78].
Resveratrol glucoside (also called polydatin), found in red wine, peanuts, and other foods, acts as a neuroprotectant in a preclinical model of lipopolysaccharide-induced Parkinson’s disease. It protects dopaminergic neurons from degeneration and improves motor dysfunction. Additionally, polydatin suppresses microglia activation and blocks the release of pro-inflammatory factors [79]. Caffeic acid, present in fruits, vegetables, coffee beans, and other dietary sources, reduces neurotoxicity caused by A53T alpha-synuclein overexpression in SH-SY5Y cells by activating the Nrf2/Bcl-2-mediated autophagy pathway [80]. Studies suggest that phytochemicals can prevent α-synuclein from forming neurotoxic oligomers and may even help break down existing aggregates [81]. Curcumin, a polyphenol in turmeric (Curcuma longa) used as a spice and food coloring, provides neuroprotection by modulating the brain-derived neurotrophic factor (BDNF) and PI3K/Akt signaling pathways [82]. Quercetin, abundant in apples, citrus fruits, onions, tea, and red wine, has been shown to inhibit alpha-synuclein aggregation into toxic oligomers [83]. Finally, L-theanine, found in green and black tea as well as certain mushrooms, exhibits neuroprotective effects in MPTP-treated SH-SY5Y cells. It boosts tyrosine hydroxylase-positive cells while decreasing alpha-synuclein clumping and Lewy body formation [84].
Omega-3 fatty acids, which are highly concentrated in salmon, have demonstrated neuroprotective effects in preclinical models of Parkinson’s disease [85,86,87,88]. These benefits are linked to multiple mechanisms, including: reducing endoplasmic reticulum stress, inhibiting microglial activation and the release of pro-inflammatory factors, decreasing mitochondrial dysfunction, promoting the expression of neurotrophic factors, maintaining calcium homeostasis and alpha-synuclein proteostasis [89]. In studies using unilaterally 6-hydroxydopamine-lesioned animals, treatment with fish oil for 50 days reduced neuronal loss in the substantia nigra pars compacta and their terminals in the striatum. The neuroprotection from fish oil was associated with fewer iNOS-immunoreactive cells and reduced microglial and astrocyte reactivity [90]. Additionally, omega-3 polyunsaturated fatty acids improved motor symptoms in 6-hydroxydopamine-treated animals, further confirming their neuroprotective role [91].
DHA, an omega-3 fatty acid, has been shown to restore tyrosine hydroxylase-positive neurons and decrease lipid peroxidation in rotenone-treated animals. It also boosts the production of antioxidant enzymes like catalase and superoxide dismutase [92]. In a rat model of 6-OHDA-induced Parkinson’s disease, DHA exhibited neuroprotective benefits by enhancing tyrosine hydroxylase levels and improving motor function, including gait and posture [93]. Additionally, DHA suppresses microgliosis and astrogliosis in both the substantia nigra and striatum in partial 6-OHDA lesion models, further supporting its neuroprotective effects [94].
The EPA has shown neuroprotective effects in differentiated human SH-SY5Y cells and primary mesencephalic cells exposed to MPP+ by countering the neurotoxin’s effects through the suppression of pro-inflammatory factor release [95]. Studies also indicate that EPA may help prevent Parkinson’s disease by reducing the neurotoxic effects of 6-hydroxydopamine in vitro. It helps restore mitochondrial function and boosts the expression of glial cell line-derived neurotrophic factor (GDNF) and brain-derived neurotrophic factor (BDNF), both of which are essential for neuronal survival, differentiation, and synapse formation [96]. Additionally, EPA has demonstrated protective effects in an MPTP-probenecid animal model, decreasing pro-inflammatory factor production and improving memory deficits [95]. A systematic review of 39 published studies further confirms the neuroprotective role of omega-3 fatty acids in Parkinson’s disease, noting improvements in behavior, pathological markers, and antioxidant, anti-inflammatory, and anti-apoptotic effects, along with higher omega-3 levels in the brain [97].
Astaxanthin (AST), a red dietary carotenoid found in foods such as salmon, krill, shrimp, crayfish, trout, yeast, and algae, has neuroprotective, antioxidant, and anti-inflammatory properties. Its antioxidant effects come from its ability to increase the expression of DT-diaphorase and glutathione transferase M2-2 [98]. Notably, AST—along with DHA and EPA—activates the KEAP1/NRF2 signaling pathway, which boosts the production of these enzymes. These mechanisms are thought to play a key role in AST’s potential neuroprotective effects against Parkinson’s disease.
7. Conclusions
In our view, the degenerative process in Parkinson’s disease follows the single-neuron degeneration model, where degeneration occurs through the loss of one neuron at a time during each degenerative event. The non-expansive nature of this model suggests an exceptionally slow progression of degeneration, unfolding over many years. According to this model, the neurotoxin responsible for triggering the degeneration of neuromelanin-containing dopaminergic neurons is believed to be aminochrome, which forms inside these neurons and lacks an expansive effect. However, the enzymes DT-diaphorase and glutathione transferase M2-2 counteract aminochrome’s neurotoxic effects, protecting these neurons from degeneration. Thus, increasing the expression of these enzymes could enhance neuroprotection in dopaminergic neurons when aminochrome is produced. The KEAP1/NRF2 signaling pathway plays a key role by activating the expression of antioxidant enzymes, including DT-diaphorase and glutathione transferase M2-2 [60,61]. Notably, bioactive compounds in certain foods—such as omega-3 fatty acids found in salmon, herring, pollock, chia oil, and others (Table 1)—activate this pathway. Among these neuroprotective foods, salmon stands out due to its high levels of DHA, EPA, and astaxanthin, all of which enhance the expression of protective enzymes via KEAP1/NRF2 activation. Therefore, increasing dietary intake of these bioactive-rich foods may help strengthen neuroprotection in dopaminergic neurons against aminochrome’s toxicity during neuromelanin synthesis.
References
- Athauda, D.; Foltynie, T. The ongoing pursuit of neuroprotective therapies in Parkinson disease. Nat. Rev. Neurol. 2015, 11, 25–40. [Google Scholar] [CrossRef]
- Parkinson Study Group QE3 Investigators Beal, M.F.; Oakes, D.; Shoulson, I.; Henchcliffe, C.; Galpern, W.R.; Haas, R.; Juncos, J.L.; Nutt, J.G.; Voss, T.S.; et al. Randomized clinical trial of high-dosage coenzyme Q10 in early Parkinson disease: No evidence of benefit. JAMA Neurol. 2014, 71, 543–552. [Google Scholar] [CrossRef]
- Snow, B.J.; Rolfe, F.L.; Lockhart, M.M.; Frampton, C.M.; O’Sullivan, J.D.; Fung, V.; Smith, R.A.; Murphy, M.P.; Taylor, K.M. ; Protect Study Group A double-blind, placebo-controlled study to assess the mitochondria-targeted antioxidant MitoQ as a disease-modifying therapy in Parkinson’s disease. Mov. Disord. 2010, 25, 1670–1674. [Google Scholar] [CrossRef] [PubMed]
- Parkinson Study Group SURE-PD3 Investigators Schwarzschild, M.A.; Ascherio, A.; Casaceli, C.; Curhan, G.C.; Fitzgerald, R.; Kamp, C.; Lungu, C.; Macklin, E.A.; Marek, K.; et al. Effect of Urate-Elevating Inosine on Early Parkinson Disease Progression: The SURE-PD3 Randomized Clinical Trial. JAMA. 2021, 326, 926–939. [CrossRef]
- Warren Olanow, C.; Bartus, R.T.; Baumann, T.L.; Factor, S.; Boulis, N.; Stacy, M.; Turner, D.A.; Marks, W.; Larson, P.; Starr, P.A.; Jankovic, J.; Simpson, R.; Watts, R.; Guthrie, B.; Poston, K.; Henderson, J.M.; Stern, M.; Baltuch, G.; Goetz, C.G.; Herzog, C.; Kordower, J.H.; Alterman, R.; Lozano, A.M.; Lang, A.E. Gene delivery of neurturin to putamen and substantia nigra in Parkinson disease: A double-blind, randomized, controlled trial. Ann Neurol. 2015, 78, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Olanow, C.W.; Bartus, R.T.; Volpicelli-Daley, L.A.; Kordower, J.H. Trophic factors for Parkinson's disease: To live or let die. Mov Disord. 2015, 30, 1715–1724. [Google Scholar] [CrossRef] [PubMed]
- Lenka, A.; Jankovic, J. How should future clinical trials be designed in the search for disease-modifying therapies for Parkinson's disease? Expert Rev Neurother. 2023, 23, 107–122. [Google Scholar] [CrossRef]
- Fabbri, M.; Rascol, O.; Foltynie, T.; Carroll, C.; Postuma, R.B.; Porcher, R.; Corvol, J.C. Advantages and Challenges of Platform Trials for Disease Modifying Therapies in Parkinson's Disease. Mov Disord. 2024, 39, 1468–1477. [Google Scholar] [CrossRef]
- Filograna, R.; Beltramini, M.; Bubacco, L.; Bisaglia, M. Anti-Oxidants in Parkinson's Disease Therapy: A Critical Point of View. Curr Neuropharmacol. 2016, 14, 260–271. [Google Scholar] [CrossRef]
- Segura-Aguilar, J. The importance of choosing a preclinical model that reflects what happens in Parkinson's disease. Neurochem Int. 2019, 126, 203–209. [Google Scholar] [CrossRef]
- Williams, A. MPTP parkinsonism. Br Med J. 1984, 289, 1401–1402. [Google Scholar] [CrossRef]
- Huenchuguala, S.; Segura-Aguilar, J. Single-neuron neurodegeneration as a degenerative model for Parkinson's disease. Neural Regen Res. 2024, 19, 529–535. [Google Scholar] [CrossRef]
- Ordonez, D.G.; Lee, M.K.; Feany, M.B. α-synuclein Induces Mitochondrial Dysfunction through Spectrin and the Actin Cy-toskeleton. Neuron. 2018, 97, 108–124. [Google Scholar] [CrossRef] [PubMed]
- Morales-Martínez, A.; Martínez-Gómez, P.A.; Martinez-Fong, D.; Villegas-Rojas, M.M.; Pérez-Severiano, F.; Del Toro-Colín, M.A.; Delgado-Minjares, K.M.; Blanco-Alvarez, V.M.; Leon-Chavez, B.A.; Aparicio-Trejo, O.E.; et al. Oxidative Stress and Mitochondrial Complex I Dysfunction Correlate with Neurodegeneration in an α-Synucleinopathy Animal Model. Int. J. Mol. Sci. 2022, 23, 11394. [Google Scholar] [CrossRef] [PubMed]
- Rocha, E.M.; De Miranda, B.; Sanders, L.H. Alpha-synuclein: Pathology, mitochondrial dysfunction and neuroinflammation in Parkinson’s disease. Neurobiol. Dis. 2018, 109 Pt B, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, A.C.; Erustes, A.G.; Reckziegel, P.; Bincoletto, C.; Ureshino, R.P.; Pereira, G.J.S.; Smaili, S.S. α-Synuclein Overexpression Induces Lysosomal Dysfunction and Autophagy Impairment in Human Neuroblastoma SH-SY5Y. Neurochem. Res. 2020, 45, 2749–2761. [Google Scholar] [CrossRef]
- Popova, B.; Galka, D.; Häffner, N.; Wang, D.; Schmitt, K.; Valerius, O.; Knop, M.; Braus, G.H. α-Synuclein Decreases the Abundance of Proteasome Subunits and Alters Ubiquitin Conjugates in Yeast. Cells. 2021, 10, 2229. [Google Scholar] [CrossRef]
- Nguyen, M.; Wong, Y.C.; Ysselstein, D.; Severino, A.; Krainc, D. Synaptic, Mitochondrial, and Lysosomal Dysfunction in Parkinson’s Disease. Trends Neurosci. 2019, 42, 140–149. [Google Scholar] [CrossRef]
- Chen, L.; Thiruchelvam, M.J.; Madura, K.; Richfield, E.K. Proteasome dysfunction in aged human alpha-synuclein transgenic mice. Neurobiol. Dis. 2006, 23, 120–126. [Google Scholar] [CrossRef]
- Muñoz, P.; Cardenas, S.; Huenchuguala, S.; Briceño, A.; Couve, E.; Paris, I.; Segura-Aguilar, J. DT-Diaphorase Prevents Aminochrome-Induced Alpha-Synuclein Oligomer Formation and Neurotoxicity. Toxicol Sci. 2015, 145, 37–47. [Google Scholar] [CrossRef]
- Ni, A.; Ernst, C. Evidence That Substantia Nigra Pars Compacta Dopaminergic Neurons Are Selectively Vulnerable to Oxidative Stress Because They Are Highly Metabolically Active. Front. Cell. Neurosci. 2022, 16, 826193. [Google Scholar] [CrossRef]
- Pérez-Acuña, D.; Shin, S.J.; Rhee, K.H.; Kim, S.J.; Lee, S.J. α-Synuclein propagation leads to synaptic abnormalities in the cortex through microglial synapse phagocytosis. Mol. Brain. 2023, 16, 72. [Google Scholar] [CrossRef] [PubMed]
- Henderson, M.X.; Cornblath, E.J.; Darwich, A.; Zhang, B.; Brown, H.; Gathagan, R.J.; Sandler, R.M.; Bassett, D.S.; Trojanowski, J.Q.; Lee, V.M.Y. Spread of α-synuclein pathology through the brain connectome is modulated by selective vulnerability and predicted by network analysis. Nat. Neurosci. 2019, 22, 1248–1257. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kwon, S.H.; Kam, T.I.; Panicker, N.; Karuppagounder, S.S.; Lee, S.; Lee, J.H.; Kim, W.R.; Kook, M.; Foss, C.A.; et al. Transneuronal Propagation of Pathologic α-Synuclein from the Gut to the Brain Models Parkinson’s Disease. Neuron. 2019, 103, 627–641. [Google Scholar] [CrossRef] [PubMed]
- Okuzumi, A.; Hatano, T.; Matsumoto, G.; Nojiri, S.; Ueno, S.I.; Imamichi-Tatano, Y.; Kimura, H.; Kakuta, S.; Kondo, A.; Fukuhara, T.; et al. Propagative α-synuclein seeds as serum biomarkers for synucleinopathies. Nat. Med. 2023, 29, 1448–1455. [Google Scholar] [CrossRef]
- Bigi, A.; Cascella, R.; Cecchi, C. α-Synuclein oligomers and fibrils: Partners in crime in synucleinopathies. Neural Regen. Res. 2023, 18, 2332–2342. [Google Scholar] [CrossRef]
- Desplats, P.; Lee, H.J.; Bae, E.J.; Patrick, C.; Rockenstein, E.; Crews, L.; Spencer, B.; Masliah, E.; Lee, S.J. Inclusion formation and neuronal cell death through neuron-to-neuron transmission of alpha-synuclein. Proc. Natl. Acad. Sci. USA. 2009, 106, 13010–13015. [Google Scholar] [CrossRef]
- Segura-Aguilar. Dopamine oxidative deamination. In Segura-Aguilar, editor, Clinical Studies and Therapies in Parkinson's Disease-Translations from Preclinical Models, Elsevier; Cambridge, MA, USA: 2021, pp 203-207.
- Goldstein, D.S. The “Sick-but-not-Dead” phenomenon applied to catecholamine deficiency in neurodegenerative diseases. Semin. Neurol. 2020, 40, 502–514. [Google Scholar] [CrossRef]
- Jinsmaa, Y.; Sullivan, P.; Sharabi, Y.; Goldstein, D.S. DOPAL is transmissible to and oligomerizes alpha-synuclein in human glial cells. Auton. Neurosci. 2016, 194, 46–51. [Google Scholar] [CrossRef]
- Bisaglia, M.; Mammi, S.; Bubacco, L. Kinetic and structural analysis of the early oxidation products of dopamine: Analysis of the interactions with alpha-synuclein. J. Biol. Chem. 2007, 282, 15597–15605. [Google Scholar] [CrossRef]
- Zecca, L.; Fariello, R.; Riederer, P.; Sulzer, D.; Gatti, A.; Tampellini, D. The absolute concentration of nigral neuromelanin, assayed by a new sensitive method, increases throughout the life and is dramatically decreased in Parkinson’s disease. FEBS Lett. 2002, 510, 216–220. [Google Scholar] [CrossRef]
- Zucca, F.A.; Basso, E.; Cupaioli, F.A.; Ferrari, E.; Sulzer, D.; Casella, L.; Zecca, L. Neuromelanin of the human substantia nigra: an update. Neurotox Res. 2014, 25, 13–23. [Google Scholar] [CrossRef]
- Zucca, F.A.; Capucciati, A.; Bellei, C.; Sarna, M.; Sarna, T.; Monzani, E.; Casella, L.; Zecca, L. Neuromelanins in brain aging and Parkinson's disease: synthesis, structure, neuroinflammatory, and neurodegenerative role. IUBMB Life. 2023, 75, 55–65. [Google Scholar] [CrossRef]
- Segura-Aguilar. Neuroprotective mechanisms against dopamine oxidation-dependent neurotoxicity. In Segura-Aguilar, editor, Clinical Studies and Therapies in Parkinson's Disease-Translations from Preclinical Models, Elsevier; Cambridge, MA, USA: 2021, Pages 229-240.
- Segura-Aguilar, J.; Metodiewa, D.; Welch, C.J. Metabolic activation of dopamine o-quinones to o-semiquinones by NADPH cytochrome P450 reductase may play an important role in oxidative stress and apoptotic effects. Biochim Biophys Acta. 1998, 1381, 1–6. [Google Scholar] [CrossRef]
- Segura-Aguilar, J.; Lind, C. On the mechanism of the Mn3(+)-induced neurotoxicity of dopamine:prevention of quinone-derived oxygen toxicity by DT diaphorase and superoxide dismutase. Chem Biol Interact. 1989, 72, 309–324. [Google Scholar] [CrossRef]
- Lozano, J.; Muñoz, P.; Nore, B.F.; Ledoux, S.; Segura-Aguilar, J. Stable expression of short interfering RNA for DT-diaphorase induces neurotoxicity. Chem Res Toxicol. 2010, 23, 1492–1496. [Google Scholar] [CrossRef] [PubMed]
- Schultzberg, M.; Segura-Aguilar, J.; Lind, C. Distribution of DT diaphorase in the rat brain: biochemical and immunohistochemical studies. Neuroscience. 1988, 27, 763–776. [Google Scholar] [CrossRef] [PubMed]
- Arriagada, C.; Paris, I.; Sanchez de las Matas, M.J.; Martinez-Alvarado, P.; Cardenas, S.; Castañeda, P.; Graumann, R.; Perez-Pastene, C.; Olea-Azar, C.; Couve, E.; et al. On the neurotoxicity mechanism of leukoaminochrome o-semiquinone radical derived from dopamine oxidation: Mitochondria damage, necrosis, and hydroxyl radical formation. Neurobiol. Dis. 2004, 16, 468–477. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, P.; Cardenas, S.; Huenchuguala, S.; Briceño, A.; Couve, E.; Paris, I.; Segura-Aguilar, J. DT-Diaphorase Prevents Aminochrome-Induced Alpha-Synuclein Oligomer Formation and Neurotoxicity. Toxicol. Sci. 2015, 145, 37–47. [Google Scholar] [CrossRef]
- Muñoz, P.; Huenchuguala, S.; Paris, I.; Segura-Aguilar, J. Dopamine oxidation and autophagy. Parkinsons. Dis. 2012, 2012, 920953. [Google Scholar] [CrossRef]
- Paris, I.; Perez-Pastene, C.; Cardenas, S.; Iturriaga-Vasquez, P.; Muñoz, P.; Couve, E.; Caviedes, P.; Segura-Aguilar, J. Aminochrome induces disruption of actin, alpha-, and beta-tubulin cytoskeleton networks in substantia-nigra-derived cell line. Neurotox. Res. 2010, 18, 82–92. [Google Scholar] [CrossRef]
- Meléndez, C.; Muñoz, P.; Segura-Aguilar, J. DT-Diaphorase Prevents Aminochrome-Induced Lysosome Dysfunction in SH-SY5Y Cells. Neurotox. Res. 2019, 35, 255–259. [Google Scholar] [CrossRef]
- Paris, I.; Muñoz, P.; Huenchuguala, S.; Couve, E.; Sanders, L.H.; Greenamyre, J.T.; Caviedes, P.; Segura-Aguilar, J. Autophagy protects against aminochrome-induced cell death in substantia nigra-derived cell line. Toxicol Sci. 2011, 121, 376–388. [Google Scholar] [CrossRef] [PubMed]
- Zafar, K.S.; Inayat-Hussain, S.H.; Siegel, D.; Bao, A.; Shieh, B.; Ross, D. Overexpression of NQO1 protects human SK-N-MC neuroblastoma cells against dopamine-induced cell death. Toxicol. Lett. 2006, 166, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Huenchuguala, S.; Muñoz, P.; Segura-Aguilar, J. The Importance of Mitophagy in Maintaining Mitochondrial Function in U373MG Cells. Bafilomycin A1 Restores Aminochrome-Induced Mitochondrial Damage. ACS Chem Neurosci. 2017, 8, 2247–2253. [Google Scholar] [CrossRef]
- Segura-Aguilar, J. On the Role of Aminochrome in Mitochondrial Dysfunction and Endoplasmic Reticulum Stress in Parkinson's Disease. Front Neurosci. 2019, 13, 271. [Google Scholar] [CrossRef]
- Segura-Aguilar, J.; Baez, S.; Widersten, M.; Welch, C.J.; Mannervik, B. Human class Mu glutathione transferases, in particular isoenzyme M2-2, catalyze detoxication of the dopamine metabolite aminochrome. J Biol Chem. 1997, 272, 5727–5731. [Google Scholar] [CrossRef]
- Baez, S.; Segura-Aguilar, J.; Widersten, M.; Johansson, A.S.; Mannervik, B. Glutathione transferases catalyse the detoxication of oxidized metabolites (o-quinones) of catecholamines and may serve as an antioxidant system preventing degenerative cellular processes. Biochem J. 1997, 324 ( Pt 1)(Pt 1), 25–28. [Google Scholar] [CrossRef]
- Huenchuguala, S.; Muñoz, P.; Zavala, P.; Villa, M.; Cuevas, C.; Ahumada, U.; Graumann, R.; Nore, B.; Couve, E.; Mannervik, B.; et al. Glutathione transferase mu 2 protects glioblastoma cells against aminochrome toxicity by preventing autophagy and lysosome dysfunction. Autophagy. 2014, 10, 618–630. [Google Scholar] [CrossRef]
- Dagnino-Subiabre, A.; Cassels, B.K.; Baez, S.; Johansson, A.S.; Mannervik, B.; Segura-Aguilar, J. Glutathione transferase M2-2 catalyzes conjugation of dopamine and dopa o-quinones. Biochem Biophys Res Commun. 2000, 274, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Cuevas, C.; Huenchuguala, S.; Muñoz, P.; Villa, M.; Paris, I.; Mannervik, B.; Segura-Aguilar, J. Glutathione transferase-M2-2 secreted from glioblastoma cell protects SH-SY5Y cells from aminochrome neurotoxicity. Neurotox Res. 2015, 27, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Valdes, R.; Armijo, A.; Muñoz, P.; Hultenby, K.; Hagg, A.; Inzunza, J.; Nalvarte, I.; Varshney, M.; Mannervik, B.; Segura-Aguilar, J. Cellular Trafficking of Glutathione Transferase M2-2 Between U373MG and SHSY-S7 Cells is Mediated by Exosomes. Neurotox Res. 2021, 39, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Segura-Aguilar, J.; Muñoz, P.; Inzunza, J.; Varshney, M.; Nalvarte, I.; Mannervik, B. Neuroprotection against Aminochrome Neurotoxicity: Glutathione Transferase M2-2 and DT-Diaphorase. Antioxidants (Basel). 2022, 11, 296. [Google Scholar] [CrossRef] [PubMed]
- Segura-Aguilar, J.; Mannervik, B.; Inzunza, J.; Varshney, M.; Nalvarte, I.; Muñoz, P. Astrocytes protect dopaminergic neurons against aminochrome neurotoxicity. Neural Regen Res. 2022, 17, 1861–1866. [Google Scholar] [CrossRef]
- Segura-Aguilar, J.; Mannervik, B. A Preclinical Model for Parkinson's Disease Based on Transcriptional Gene Activation via KEAP1/NRF2 to Develop New Antioxidant Therapies. Antioxidants (Basel). 2023, 12, 673. [Google Scholar] [CrossRef]
- Huenchuguala, S.; Segura-Aguilar, J. Natural Compounds That Activate the KEAP1/Nrf2 Signaling Pathway as Potential New Drugs in the Treatment of Idiopathic Parkinson's Disease. Antioxidants (Basel). 2024, 13, 1125. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.H. Health-promoting components of fruits and vegetables in the diet. Adv Nutr. 2013, 4, 384S–92S. [Google Scholar] [CrossRef]
- Neufingerl, N.; Eilander, A. Nutrient Intake and Status in Adults Consuming Plant-Based Diets Compared to Meat-Eaters: A Systematic Review. Nutrients. 2021, 14, 29. [Google Scholar] [CrossRef]
- Nakajima, A.; Ohizumi, Y. Potential Benefits of Nobiletin, A Citrus Flavonoid, against Alzheimer's Disease and Parkinson's Disease. Int J Mol Sci. 2019, 20, 3380. [Google Scholar] [CrossRef]
- Braidy, N.; Behzad, S.; Habtemariam, S.; Ahmed, T.; Daglia, M.; Nabavi, S.M.; Sobarzo-Sanchez, E.; Nabavi, S.F. Neuroprotective Effects of Citrus Fruit-Derived Flavonoids, Nobiletin and Tangeretin in Alzheimer's and Parkinson's Disease. CNS Neurol Disord Drug Targets. 2017, 16, 387–397. [Google Scholar] [CrossRef]
- Dinda, B.; Dinda, M.; Kulsi, G.; Chakraborty, A.; Dinda, S. Therapeutic potentials of plant iridoids in Alzheimer's and Parkinson's diseases: A review. Eur J Med Chem. 2019, 169, 185–199. [Google Scholar] [CrossRef]
- Huang, B.; Liu, J.; Meng, T.; Li, Y.; He, D.; Ran, X.; Chen, G.; Guo, W.; Kan, X.; Fu, S.; Wang, W.; Liu, D. Polydatin Prevents Lipopolysaccharide (LPS)-Induced Parkinson's Disease via Regulation of the AKT/GSK3β-Nrf2/NF-κB Signaling Axis. Front Immunol. 2018, 9, 2527. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, Q.; Zhang, L.; Wang, Q.; Yang, Z.; Liu, J.; Feng, L. Caffeic acid reduces A53T α-synuclein by activating JNK/Bcl-2-mediated autophagy in vitro and improves behaviour and protects dopaminergic neurons in a mouse model of Parkinson's disease. Pharmacol Res. 2019, 150, 104538. [Google Scholar] [CrossRef]
- Wang, N.; Feng, B.N.; Hu, B.; Cheng, Y.L.; Guo, Y.H.; Qian, H. Neuroprotection of chicoric acid in a mouse model of Parkinson's disease involves gut microbiota and TLR4 signaling pathway. Food Funct. 2022, 13, 2019–2032. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.M.; Lee, Y.; Chun, H.J.; Kim, A.H.; Kim, J.Y.; Lee, J.Y.; Ishigami, A.; Lee, J. Neuroprotective and anti-inflammatory effects of morin in a murine model of Parkinson's disease. J Neurosci Res. 2016, 94, 865–878. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.M.; Wang, M.; Xiao, X.; Shi, Y.L.; Zheng, Y.S.; Huang, Z.H.; Cheng, Y.T.; Huang, R.T.; Huang, F.; Li, K.; Sun, J.; Sun, W.Y.; Kurihara, H.; Li, Y.F.; Duan, W.J.; He, R.R. Wolfberry (Lycium barbarum) glycopeptide attenuates dopaminergic neurons loss by inhibiting lipid peroxidation in Parkinson's disease. Phytomedicine. 2025, 136, 156275. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Sattar, E.; Mahrous, E.A.; Thabet, M.M.; Elnaggar, D.M.Y.; Youssef, A.M.; Elhawary, R.; Zaitone, S.A.; Celia Rodríguez-Pérez Segura-Carretero, A.; Mekky, R.H. Methanolic extracts of a selected Egyptian Vicia faba cultivar mitigate the oxidative/inflammatory burden and afford neuroprotection in a mouse model of Parkinson's disease. Inflammopharmacology. 2021, 29, 221–235. [Google Scholar] [CrossRef]
- Sun, T.; Chen, L.; Liu, R.; Liu, Q.S.; Cheng, Y. Sophora alopecuroides Alleviates Neuroinflammation and Oxidative Damage of Parkinson's Disease In Vitro and In Vivo. Am J Chin Med. 2023, 51, 309–328. [Google Scholar] [CrossRef]
- Brandalise, F.; Roda, E.; Ratto, D.; Goppa, L.; Gargano, M.L.; Cirlincione, F.; Priori, E.C.; Venuti, M.T.; Pastorelli, E.; Savino, E.; Rossi, P. Hericium erinaceus in Neurodegenerative Diseases: From Bench to Bedside and Beyond, How Far from the Shoreline? J Fungi (Basel). 2023, 9, 551. [Google Scholar] [CrossRef]
- Li, I.C.; Lee, L.Y.; Tzeng, T.T.; Chen, W.P.; Chen, Y.P.; Shiao, Y.J.; Chen, C.C. Neurohealth Properties of Hericium erinaceus Mycelia Enriched with Erinacines. Behav Neurol. 2018, 2018, 5802634. [Google Scholar] [CrossRef]
- Wang, Y.C.; Wang, V.; Chen, B.H. Analysis of bioactive compounds in cinnamon leaves and preparation of nanoemulsion and byproducts for improving Parkinson's disease in rats. Front Nutr. 2023, 10, 1229192. [Google Scholar] [CrossRef]
- Piechowska, P.; Zawirska-Wojtasiak, R.; Mildner-Szkudlarz, S. Bioactive β-Carbolines in Food: A Review. Nutrients. 2019, 11, 814. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, A.; Ohizumi, Y. Potential Benefits of Nobiletin, A Citrus Flavonoid, against Alzheimer's Disease and Parkinson's Disease. Int J Mol Sci. 2019, 20, 3380. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Liu, J.; Meng, T.; Li, Y.; He, D.; Ran, X.; Chen, G.; Guo, W.; Kan, X.; Fu, S.; Wang, W.; Liu, D. Polydatin Prevents Lipopolysaccharide (LPS)-Induced Parkinson's Disease via Regulation of the AKT/GSK3β-Nrf2/NF-κB Signaling Axis. Front Immunol. 2018, 9, 2527. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wu, Q.; Zhang, L.; Wang, Q.; Yang, Z.; Liu, J.; Feng, L. Caffeic acid reduces A53T α-synuclein by activating JNK/Bcl-2-mediated autophagy in vitro and improves behaviour and protects dopaminergic neurons in a mouse model of Parkinson's disease. Pharmacol Res. 2019, 150, 104538. [Google Scholar] [CrossRef]
- Naoi, M.; Maruyama, W.; Shamoto-Nagai, M. Disease-modifying treatment of Parkinson's disease by phytochemicals: targeting multiple pathogenic factors. J Neural Transm (Vienna). 2022, 129, 737–753. [Google Scholar] [CrossRef]
- Jin, T.; Zhang, Y.; Botchway, B.O.A.; Zhang, J.; Fan, R.; Zhang, Y.; Liu, X. Curcumin can improve Parkinson's disease via activating BDNF/PI3k/Akt signaling pathways. Food Chem Toxicol. 2022, 164, 113091. [Google Scholar] [CrossRef]
- Das, S.S.; Jha, N.K.; Jha, S.K.; Verma, P.R.P.; Ashraf, G.M.; Singh, S.K. Neuroprotective Role of Quercetin against Alpha-Synuclein-Associated Hallmarks in Parkinson's Disease. Curr Neuropharmacol. 2023, 21, 1464–1466. [Google Scholar] [CrossRef]
- Ratih, K.; Lee, Y.R.; Chung, K.H.; Song, D.H.; Lee, K.J.; Kim, D.H.; An, J.H. L-Theanine alleviates MPTP-induced Parkinson's disease by targeting Wnt/β-catenin signaling mediated by the MAPK signaling pathway. Int J Biol Macromol. 2023, 226, 90–101. [Google Scholar] [CrossRef]
- Cholewski, M.; Tomczykowa, M.; Tomczyk, M. A Comprehensive Review of Chemistry, Sources and Bioavailability of Omega-3 Fatty Acids. Nutrients. 2018, 10, 1662. [Google Scholar] [CrossRef]
- Bandarra, N.M.; Marçalo, A.; Cordeiro, A.R.; Pousão-Ferreira, P. Sardine (Sardina pilchardus) lipid composition: Does it change after one year in captivity? Food Chem. 2018, 244, 408–413. [Google Scholar] [CrossRef] [PubMed]
- Jin, F.; Nieman, D.C.; Sha, W.; Xie, G.; Qiu, Y.; Jia, W. Supplementation of milled chia seeds increases plasma ALA and EPA in postmenopausal women. Plant Foods Hum Nutr. 2012, 67, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Santos, H.O.; Price, J.C.; Bueno, A.A. Beyond Fish Oil Supplementation: The Effects of Alternative Plant Sources of Omega-3 Polyunsaturated Fatty Acids upon Lipid Indexes and Cardiometabolic Biomarkers-An Overview. Nutrients. 2020, 12, 3159. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Song, C. Potential treatment of Parkinson's disease with omega-3 polyunsaturated fatty acids. Nutr Neurosci. 2022, 25, 180–191. [Google Scholar] [CrossRef]
- Mori, M.A.; Delattre, A.M.; Carabelli, B.; Pudell, C.; Bortolanza, M.; Staziaki, P.V.; Visentainer, J.V.; Montanher, P.F.; Del Bel, E.A.; Ferraz, A.C. Neuroprotective effect of omega-3 polyunsaturated fatty acids in the 6-OHDA model of Parkinson's disease is mediated by a reduction of inducible nitric oxide synthase. Nutr Neurosci. 2018, 21, 341–351. [Google Scholar] [CrossRef]
- Delattre, A.M.; Kiss, A.; Szawka, R.E.; Anselmo-Franci, J.A.; Bagatini, P.B.; Xavier, L.L.; Rigon, P.; Achaval, M.; Iagher, F.; de David, C.; Marroni, N.A.; Ferraz, A.C. Evaluation of chronic omega-3 fatty acids supplementation on behavioral and neurochemical alterations in 6-hydroxydopamine-lesion model of Parkinson's disease. Neurosci Res. 2010, 66, 256–264. [Google Scholar] [CrossRef]
- Oguro, A.; Ishihara, Y.; Siswanto, F.M.; Yamazaki, T.; Ishida, A.; Imaishi, H.; Imaoka, S. Contribution of DHA diols (19,20-DHDP) produced by cytochrome P450s and soluble epoxide hydrolase to the beneficial effects of DHA supplementation in the brains of rotenone-induced rat models of Parkinson's disease. Biochim Biophys Acta Mol Cell Biol Lipids. 2021, 1866, 158858. [Google Scholar] [CrossRef]
- Chitre, N.M.; Wood, B.J.; Ray, A.; Moniri, N.H.; Murnane, K.S. Docosahexaenoic acid protects motor function and increases dopamine synthesis in a rat model of Parkinson's disease via mechanisms associated with increased protein kinase activity in the striatum. Neuropharmacology. 2020, 167, 107976. [Google Scholar] [CrossRef]
- Hernando, S.; Requejo, C.; Herran, E.; Ruiz-Ortega, J.A.; Morera-Herreras, T.; Lafuente, J.V.; Ugedo, L.; Gainza, E.; Pedraz, J.L.; Igartua, M.; Hernandez, R.M. Beneficial effects of n-3 polyunsaturated fatty acids administration in a partial lesion model of Parkinson's disease: The role of glia and NRf2 regulation. Neurobiol Dis. 2019, 121, 252–262. [Google Scholar] [CrossRef]
- Luchtman, D.W.; Meng, Q.; Wang, X.; Shao, D.; Song, C. ω-3 fatty acid eicosapentaenoic acid attenuates MPP+-induced neurodegeneration in fully differentiated human SH-SY5Y and primary mesencephalic cells. J Neurochem. 2013, 124, 855–868. [Google Scholar] [CrossRef] [PubMed]
- Ceccarini, M.R.; Ceccarelli, V.; Codini, M.; Fettucciari, K.; Calvitti, M.; Cataldi, S.; Albi, E.; Vecchini, A.; Beccari, T. The Polyunsaturated Fatty Acid EPA, but Not DHA, Enhances Neurotrophic Factor Expression through Epigenetic Mechanisms and Protects against Parkinsonian Neuronal Cell Death. Int J Mol Sci. 2022, 23, 16176. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Alves, B.D.S.; Schimith, L.E.; da Cunha, A.B.; Dora, C.L.; Hort, M.A. Omega-3 polyunsaturated fatty acids and Parkinson's disease: A systematic review of animal studies. J Neurochem. 2024, 168, 1655–1683. [Google Scholar] [CrossRef] [PubMed]
- Saw, C.L.; Yang, A.Y.; Guo, Y.; Kong, A.N. Astaxanthin and omega-3 fatty acids individually and in combination protect against oxidative stress via the Nrf2-ARE pathway. Food Chem Toxicol. 2013, 62, 869–875. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Aminochrome-induced single-neuron degeneration is a key process in Parkinson's disease. The synthesis of neuromelanin within dopaminergic neurons requires the oxidation of dopamine's catechol group. This reaction generates three ortho-quinones: dopamine o-quinone, aminochrome, and 5,6-indolequinone. Among these, aminochrome is the most stable and neurotoxic intermediate. It triggers a cascade of detrimental effects, including mitochondrial dysfunction, impaired protein degradation, the formation of neurotoxic alpha-synuclein oligomers, endoplasmic reticulum stress, oxidative stress, and neuroinflammation. Crucially, aminochrome's neurotoxicity is highly focused, selectively damaging the neuron it forms in while sparing adjacent cells, resulting in single-neuron death. The slow, cumulative loss of these individual neurons over many years is what ultimately initiates the motor symptoms and drives the progression of idiopathic Parkinson's disease.
Figure 1.
Aminochrome-induced single-neuron degeneration is a key process in Parkinson's disease. The synthesis of neuromelanin within dopaminergic neurons requires the oxidation of dopamine's catechol group. This reaction generates three ortho-quinones: dopamine o-quinone, aminochrome, and 5,6-indolequinone. Among these, aminochrome is the most stable and neurotoxic intermediate. It triggers a cascade of detrimental effects, including mitochondrial dysfunction, impaired protein degradation, the formation of neurotoxic alpha-synuclein oligomers, endoplasmic reticulum stress, oxidative stress, and neuroinflammation. Crucially, aminochrome's neurotoxicity is highly focused, selectively damaging the neuron it forms in while sparing adjacent cells, resulting in single-neuron death. The slow, cumulative loss of these individual neurons over many years is what ultimately initiates the motor symptoms and drives the progression of idiopathic Parkinson's disease.
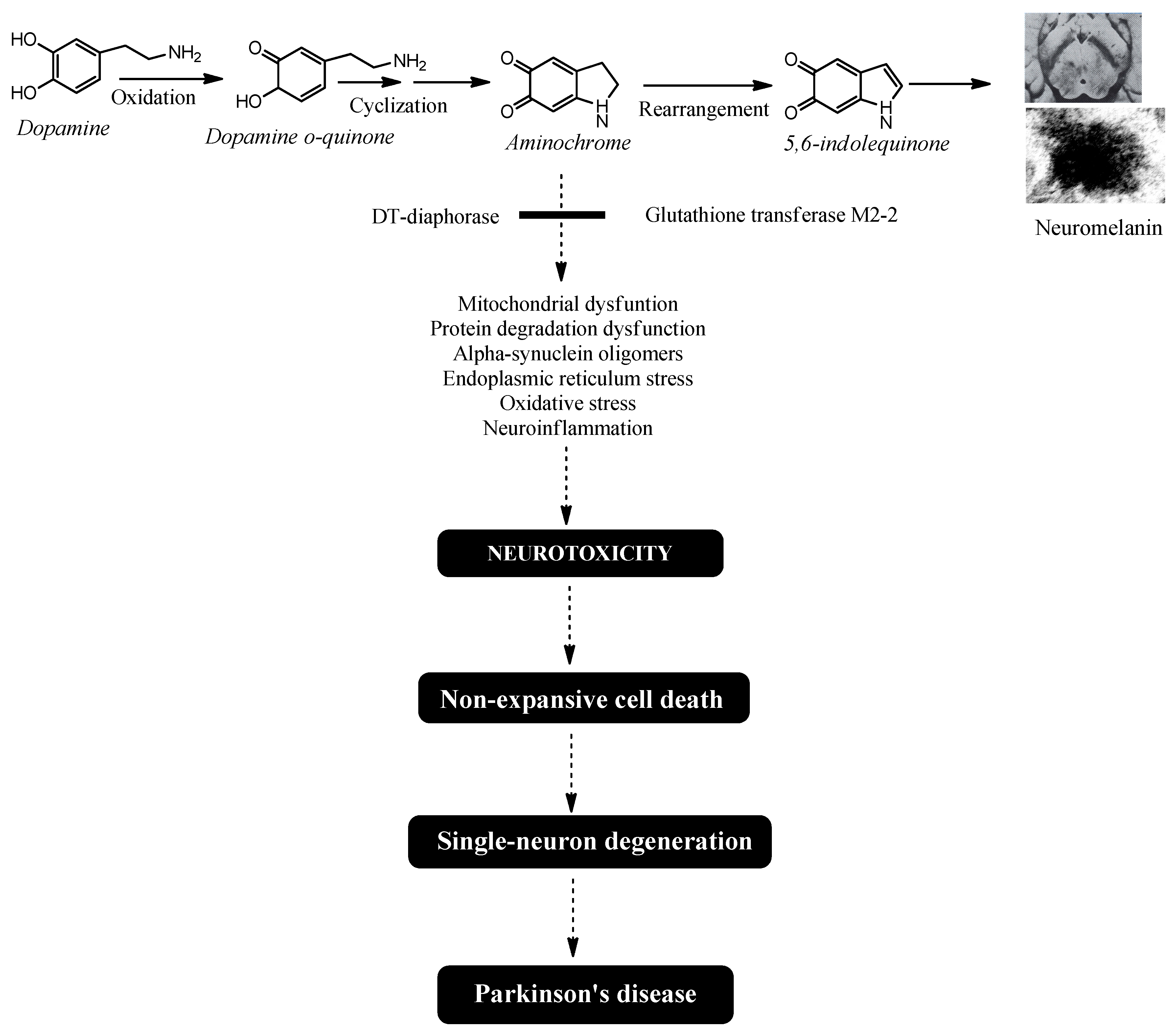
Figure 2.
Possible neuroprotective mechanism of docosahexaenoic acid (DHA), eicosapentaenoic acid (EPA), and astaxanthin (AST) in idiopathic Parkinson's disease. DHA, EPA, and AST activate the KEAP1/Nrf2 signaling pathway, increasing the expression of DT-diaphorase and glutathione transferase M2-2, which prevent the neurotoxic effects of aminochrome.
Figure 2.
Possible neuroprotective mechanism of docosahexaenoic acid (DHA), eicosapentaenoic acid (EPA), and astaxanthin (AST) in idiopathic Parkinson's disease. DHA, EPA, and AST activate the KEAP1/Nrf2 signaling pathway, increasing the expression of DT-diaphorase and glutathione transferase M2-2, which prevent the neurotoxic effects of aminochrome.
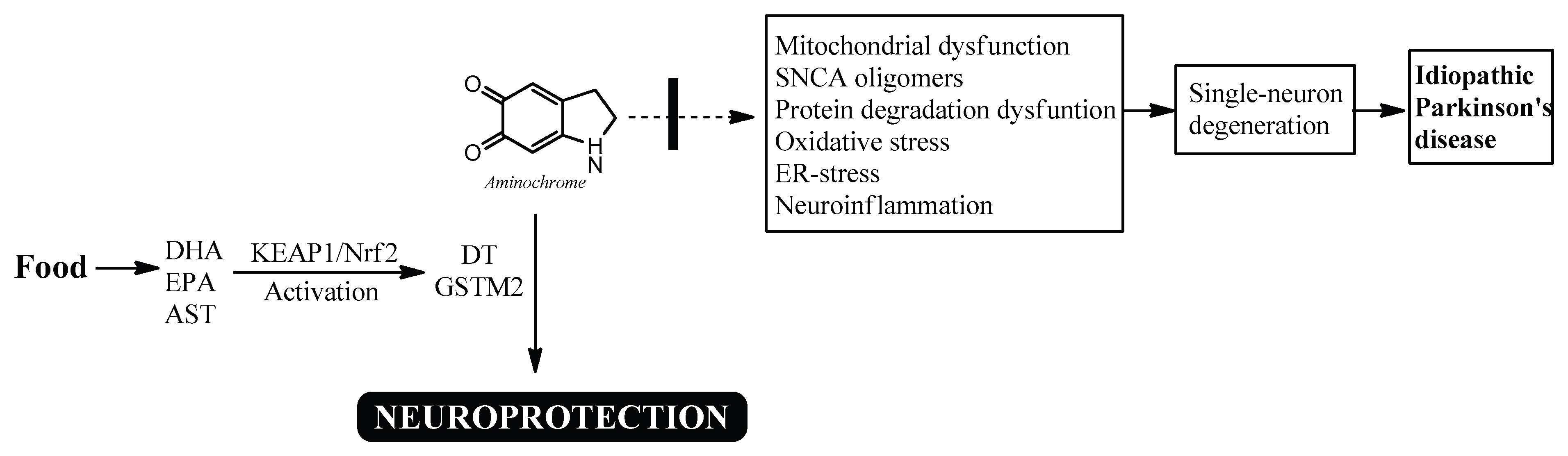

Table 1.
Natural sources of omega-3 compounds and alpha-linolenic acid.
| Source | Amount of total lipids | Reference | |
| Eicosapentaenoic acid (EPA) |
Herring | 15 % | [85] |
| Wild sardine | 13.6 % in muscle | [86] | |
| Pollock roe | 18.8 | [85] | |
| Undaria pinnatifida | 13 % of essential oil composition) | [85] | |
| Rhododendron sochadzeae | 2 % of leaf extract |
[85] | |
| Docosahexaenoic acid (DHA) | Flyingfish | 27.9 % | [85] |
| Herring | 22.6 % | [85] | |
| Pollock | 22.2 % | [85] | |
| Salmon roe | 17.4 % | [85] | |
| Cirrhinus mrigata | 18.07 g/ 100 g muscle | [85] | |
| Catla catla | 17.98 g/100 g muscle | [85] | |
| Jackalberry | 4.54 g/ 100 g oil | [85] | |
| Alpha-linolenic acid | Chia (Salvia hispanica L.) seed | 64.04% of seed oil fatty acids | [85] |
| Trichosanthes kirilowii | 33.77–38.66% of seed oils | [85] | |
| Paprika Capsicum annuum | 29.93% of fresh pericarp fatty acids in the Jaranda variety and 30.27% in the Jariza variety | [85] | |
| Sardine (Sardina pilchardus) |
1.1 | [86] | |
| Linum usitatissimum | 1.1 to 65.2 % | [85] | |
| Rapeseed oil | 9.1 % | [88] | |
| Olive oil | 0.76 % | [88] | |
| Flaxseed oil | 53.4 % | [88] | |
| Soybean oil | 6.7 % | [88] | |
| Corn oil | 1.2 % | [88] | |
| Walnut oil | 10.4 % | [88] | |
| Walnuts seed | 9.0 % of the total seed weight | [88] | |
| Flaxseed seed | 22.8 % of the total seed weight | [88] | |
| Hemp seed | 10 % of the total seed weight | [88] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



