Submitted:
25 August 2025
Posted:
26 August 2025
You are already at the latest version
Abstract
Developing sustainable and eco-friendly approaches to plant propagation, development, and protection is a common goal for the scientific community. Plant cell culturing enables us to obtain plant clones and produce biomolecules under controlled conditions. The same principle can be applied to the harvesting of extracellular vesicles (EVs). These nanosized structures are key players in cell communication and stress response by carrying, protecting and delivering important biomolecules. Raising interest in the scientific community, EVs have been tested successfully as nanocarriers in therapeutics and biotechnology. However, despite their potential, there remains a gap in research regarding scalable and reliable sources for EV production. Our goals were to optimize EV production and isolation from induced poplar callus cell lines (Populus tremula x P. alba) and load these with RNA to validate their functionality as nanocarriers. We were able to isolate 2.5 x 1010 EVs/g, highlighting the potential of these lines to mass produce them. Furthermore, RNA loaded into EVs through electroporation was internalized into Botrytis cinerea hyphae, reassuring their potential in protecting and delivering cargo. Our findings contribute to EV characterization and demonstrate that RNA-delivery through EV transport could be a safe and effective method for future EV-based technologies in plant protection.
Keywords:
callus
; extracellular vesicles
; RNA-loading
; RNA-delivery
; nanocarrier
1. Introduction
Callus induction in vitro, or callogenesis, represents a well-established technique in plant biotechnology for generating undifferentiated plant tissue under controlled conditions [1]. This method not only enables the continuous propagation of plant material independent of seasonal or environmental constraints, when the induced callus has morphogenic potential, as in indirect organogenesis [2] or somatic embryogenesis processes [3], but also serves as a powerful platform for the large-scale production of valuable metabolites and biologically active compounds [4].
In recent years, plant callus cultures have also emerged as a promising source of extracellular vesicles (EVs) [5,6,7,8,9], which are structures with an elevated biotechnological potential due to their capacity for drug delivery [10]. Most current studies rely on mammalian or limited plant systems, which often face challenges in yield, consistency, and cost-effectiveness [11]. Identifying alternative biological platforms capable of producing EVs at a larger scale is therefore critical to unlocking their full translational potential [12,13].
EVs are nano-sized structures, with an average diameter of 80-150 nm, with a bilipid membrane that confers on them the ability to carry and protect their contents [14]. They are known to transport a myriad of biomolecules like RNA, proteins and secondary metabolites [15]. These stand out due to their intrinsic value in molecular studies and applications [16]. Currently, EVs are a hot topic in the creation of novel methods in medicine [17], mainly due to their ability to naturally transport biomolecules [18] (e.g., RNA) and lower toxicity due to their natural genesis [19]. Some works have already focused on the impact that callus-produced EVs could have in therapeutics [7].
Given the scalability, reproducibility, and manipulability of in vitro systems, callus-derived EVs offer exciting potential for applications in plant science, biotechnology, and nanomedicine [5,9]. However, the use of callus cultures, particularly from woody species such as Populus, for EV production remains underexplored and warrants further investigation.
Currently, directives of the European authorities aim to reduce the use of chemical pesticides in crop protection and increase the use of more sustainable and environmentally friendly ones [20]. A current technique being explored in crop protection is the use of interfering RNA (RNAi) as a biocontrol agent, namely small interfering RNAs (siRNAs) [21]. siRNAs can regulate gene expression by silencing specific target genes in the plant or of invading pathogens [22]. Ledprona is an example of an RNAi-based commercial product in the US, where a double-stranded RNA is used against the potato beetle [23]. Although RNAi is an upcoming and potentially highly effective technique to help in plant protection, it still has some issues, such as its delivery and instability, namely the rapid degradation of RNA under environmental conditions [24]. Therefore, their effectiveness could be greatly improved using nanocarriers like EVs, as they could contribute to a more targeted delivery, RNA stability and uptake efficiency [25]. Although not EV-based, Bioclay, a formulation of a clay nanostructure to deliver and protect RNA, is an example of what EVs could improve in RNAi-based techniques [26].
Botrytis cinerea, commonly known as grey mold, infects a wide range of plant hosts, from fruits to vegetables [27]. Previous works showed that B. cinerea spores easily take up RNA during spore germination [28], making them an ideal choice for RNA uptake assays.
In this work, we characterize EVs in poplar callus and show that this system is a viable, clean and valuable source of extracellular vesicles, with the capacity to deliver RNA into fungal cells, filling the gap as a scalable and reliable system for EV production for future EV-based technologies.
2. Materials and Methods
2.1. Callus Induction and Maintenance
For callus induction, poplar (Populus tremula x P. alba) leaves of in vitro established plantlets were used. Leaf segments were cut within the midrib (1 cm x 1 cm) and punctured. These were placed in petri dishes with MS medium [29] supplemented with 30 g/L of sucrose, 7 g/L agar, 1 mg/L 2,4-D (2,4-dichlorophenoxyacetic acid) and 0.05 mg/L BAP (6-Benzylaminopurine) and the pH was adjusted to 5.7. The leaf segments were incubated at 24 °C in the dark for 17 weeks, being subcultured 3 times with intervals of 4-6 weeks until the proliferating callus was formed. Afterwards, the induced callus was harvested and placed in fresh medium.
2.2. Extracellular Vesicles Extraction
Ten grams of callus were harvested and placed on a needleless syringe, filled with VIB (Vesicle Isolation Buffer) [30]. The buffer was carefully infiltrated by slowly pulling and releasing the plunger. Then the callus was transferred into a pierced syringe inside a 50 mL centrifuge tube. The samples were then run at 900 x g for 10 minutes (4°C) and the apoplastic washing fluid (AWF) was obtained. To pellet the EVs, the differential centrifugation method was used [31]. The AWF was centrifuged at 2000 x g for 20 minutes, followed by 10,000 x g for 30 minutes, always at 4°C. The pellet was discarded, and the supernatant filtered through a 0.45 μm Nylon filter (Sterile PES Syringe Filter, Fisherbrand™, New Hampshire, USA). Then, the supernatant was moved into ultracentrifuge tubes (13.2 mL open-top thinwall, Beckman Coulter, Brea, USA) and centrifuged at 100,000 x g for 80 minutes (swinging-bucket rotor SW 41 Ti, L-100 XP centrifuge, Beckman Coulter, Brea, USA). The resulting pellet was washed with fresh buffer and ultracentrifuged a second time at 100,000 x g for 80 minutes, at 4 °C. The pellet was resuspended as follows: in 20 μL of VIB for Transmission Electron Microscopy (TEM); in 1 mL of VIB for Nanoparticle Tracker Analysis (NTA) and; in 100 μL of VIB for the loading assays.
2.3. Transmission Electron Microscopy
For Transmission electron microscopy (TEM), paraformaldehyde was added to the EV resuspended samples to a final concentration of 4% (v/v) and kept in the fridge for up to a week. Samples were transferred to Formvar- carbon coated grids for 5 min and contrasted with uranyl acetate (2%; w/v) for 1 min. Observations were carried out using a Tecnai G2 Spirit BioTwin electron microscope (FEI, Oregon, USA) at 100 kV.
2.4. Nanoparticle Track Analysis
The samples for Nanoparticle track analysis (NTA) were kept at -80 °C until analysis. Samples were analysed using Nanosight (Malvern Pananalytical, Malvern, England). EV concentration was adjusted to the initial volume and weight for each sample. The EV size average, mode, standard deviation and D10/50/90 (percent of the particles are equal to or smaller than) were included in the results.
2.5. In Vitro Transcription of Fluorescent RNA
A double-stranded RNA complementary to exons 13 and 14 of wheat Phytoene Desaturase PDS1 (dsPDS) was selected and generated through in vitro transcription (IVT). cDNA from wheat cv. Morocco was synthesized using the SuperScript™ IV One-Step RT-PCR System (Thermo Scientific, Massachusetts, USA), and two pairs of gene-specific primers (Table 1) were used to amplify a 219 bp segment containing a T7 RNA polymerase promoter at either the 5’ or the 3’ end, respectively. Each segment was cloned into E. coli DH5α and sequenced (outsourced to STAB VIDA, Caparica, Portugal). Plasmids containing each segment were purified using NZYMiniprep (NZYTech, Lisbon, Portugal), DNA templates for IVT were amplified by PCR with Phusion High-Fidelity DNA polymerase (Thermo Scientific, Massachusetts, USA) and purified using NZYGelPure (NZYTech, Lisbon, Portugal). To produce fluorescently labelled dsPDS (dsPDS-F), Fluorescein RNA Labelling Mix (Roche, Basel, Switzerland) was used according to the manufacturer's protocol and Hamby et al, 2020 and stored at -80 °C until further use [32]. Integrity of dsPDS-F was visualized by running on a 1% TAE gel with commercial bleach at 1.5% v/v [33].
2.6. Loading Assays
The samples for loading assays were kept at -80 °C. Several methods from previous known experiments were used to try and load the obtained EVs with the fluorescent RNA (100 ng/µl), but electroporation, sonication, co-incubation and freeze-thaw were the ones chosen. Before electroporation, a buffer was freshly prepared containing 1.15 mM potassium phosphate (pH 7.2). 25 mM potassium chloride and 21% (v/v) OptiPrepTM (Iodixanol) (STEMCELL Technologies Inc., Vancouver, Canada). EV solution and electroporation buffer were mixed in a 1:1 ratio and transferred to 2 mm electroporation cuvettes. The samples were electroporated at 400, 200 and 100 Volts (Bio-Rad GenePulser XCell Electroporation System, Bio-Rad Laboratories Inc., California, USA) for 5 ms and incubated in the dark at 37 °C for 30 minutes [34]. Sonication was performed following the protocol described in Kim and colleagues (2016) [31]. Six cycles of 30 seconds on/off for 3 minutes were applied, with 2 minutes on ice incubation after each cycle, and then incubated in the dark at 37 °C for 30 minutes. For co-incubation, samples were left for 4 hours at room temperature with mild shaking. For the freeze-thaw method, 3 cycles of -80 °C to RT, 30 minutes each, were applied and then incubated in the dark at 37 °C for 30 minutes. To remove the unloaded RNA, 50 U of Micrococcal Nuclease (MNase, New England Biolabs, Massachusetts, USA) was used. To confirm that the EVs were loaded with RNA, samples were observed under a Zeiss LSM 710 Confocal Microscope (Carl Zeiss AG, Oberkochen, Germany).
2.7. Uptake Assays
RNA-loaded vesicles were used for this assay. Botrytis cinerea was grown in V8 media [35] at 24 °C in a 16/8 h light cycle until sporulation (≈ 10 days). The mycelia were harvested using sterile distilled water and a cell scraper. This solution was filtered through a cell strainer (70 µm) to obtain a solution containing mostly spores, which was diluted to a 1 x 105 spores/mL. To 10 µL of spore solution, 10 µL of loaded EVs were added. The solution was carefully mixed and placed onto microscope slides, which were kept in a petri dish with a wet filter paper and wrapped in aluminium foil to keep a humid and dark environment. Samples were left overnight and imaged the following morning. For observation, a Nikon Ni-E fluorescence microscope (Nikon Instruments Inc., New York, USA) was used. NIS-Elements (Nikon Instruments Inc., New York, USA) was used to acquire brightfield and darkfield-illuminated micrographs.
3. Results
3.1. Callus Induction and Mass Propagation
According to what has been described, callus was successfully induced in poplar leaf explants [36]. Punctured leaf explants were subcultured 3 times in the same media with intervals of 4 to 6 weeks. Over 17 weeks, these subcultures gradually transitioned through a dedifferentiation phase, during which the tissue progressively lost its original structure and began forming unorganized cell masses (Figure 1a). By the end of this period, a proliferating callus was successfully established (Figure 1b). The resulting callus line was highly homogeneous, friable, and exhibited a distinct yellow coloration, characteristic of active, undifferentiated cells (Figure 1c,d). This line demonstrated a vigorous proliferation rate, requiring transfer to fresh medium every 4 weeks to maintain optimal growth. On average, each culture cycle yielded approximately 500-600 mg of fresh callus tissue, starting from 100 mg, enabling the production of sufficient biomass to proceed with extracellular vesicle (EV) isolation.
3.2. EV Isolation and Characterization
Differential centrifugation is a common method for EV isolation, and it was used in this work to isolate EVs from callus. This method allows for a wide array of vesicles to be isolated, ranging from ≈ 80 to ≈ 250 nm in width. Transmission electron microscopy is a powerful tool to confirm the presence of EVs (Figure 2a). Under TEM, the EVs can often be observed with a distinct bilipid layer and are round-shaped; however, the method also causes some EVs to collapse and attain new shapes. We can observe that some become cup-shaped, and others seem broken. We observed that the population of EVs in callus is heterogeneous, as different-sized EVs are observed, ranging from 70 to 220 nm. Aggregation of EVs was present in most of our images, as it seems that EVs tend to aggregate between themselves and form bigger blobs. Some smaller structures are sometimes present, although it is difficult to pinpoint what they are exactly due to their erroneous shapes. Overall, differential centrifugation worked well to isolate our EVs with minimal to no contamination from other sources.
To further characterize our EV population, Nanoparticle track analysis was used. Both TEM and NTA are gold standards in EV characterization, thus, both were employed in this study. NTA allows us to quantify our EV sample and observe the size distribution of the EV population. Two distinct peaks can be observed on the graph (Figure 2b) at 121 nm and 211 nm. The peak at 121 nm is more prominent than the one at 211 nm, making these smaller-sized vesicles the more common type of EVs in our samples. The final concentration of total EVs in our samples was 2.5 x 1010 particles/mL. The mean size was 142 nm, the mode,121 nm and the standard deviation,48 nm. D10, D50 and D90 were also calculated. The particle size below which 10% of the total particles is contained was 101 nm, below 50%,132 nm and below 90%,191 nm. This means that the graph area represented until 200 nm constitutes almost all the EVs population in our sample. Some peaks also appear at higher sizes than 211 nm, which may be EVs aggregates, or machine-induced errors like out-of-focus particles.
3.2. Loading and Uptake Assays
Different types of loading assays were performed to test the loading of fluorescently tagged RNA (dsPDS-F) into isolated EVs- electroporation, sonication, co-incubation and freeze-thaw. To confirm the effectiveness of these methods, EVs samples (Figure 3a) and RNA-loaded EVs (Figure 3b), were treated with MNase to remove any dsPDS-F attached to the outer membrane of the EVs or located outside of the EVs. MNase degraded any nucleotides that are outside of the EVs, while the ones inside them are protected. This confirms that the EVs protected the RNA from MNase Fluorescence was detected throughout all methods (Figure 3, and data not shown). We observed some dispersed dots, but also some agglomerates of fluorescence, probably due to EV aggregation, as seen in Figure 1b.
Following confirmation that all assays resulted in dsPDS-F-loaded EVs, these were used to perform uptake assays in Botrytis cinerea spores. This is a reliable system to observe RNA uptake of single and double-stranded RNAs of different sizes. dsPDS-F-loaded EVs were incubated with germinating spores, and fluorescence was observed with an epifluorescence microscope (Figure 4). Germinating B. cinerea spores incubated with EVs electroporated at 400 V (EP400) with dsPDS-F, showed fluorescence after 16 hours (Figure 4 a/b), indicating that the RNA-loaded EVs delivered their cargo into the hyphae. Despite being able to load dsPDS-F in the EVs by different methods, we only observed dsPDS-F uptake by the B. cinerea hyphae when loaded by electroporation (400 V). When dsPDS-F were loaded into EVs through sonication, co-incubation, freeze-thaw and electroporation at 200 and 100 V, we did not observe uptake by the hyphae (Figure 4 c/d). It is also interesting to note that fluorescence was only detected in the hyphae and never inside the spores.
4. Discussion
Poplar cell lines are ideal to study not only somatic embryogenesis [37], but also to explore how they can be a tool in current and future systems [38,39]. The possibility to produce these in bioreactors allows us to mass-produce desired biomolecules like secondary metabolites and enzymes [40] in a safe environment. The same could be applied to the mass production of EVs. This would allow us to obtain safe and clean EVs to be used in biotechnological and medical applications. To further demonstrate that these could be a desirable system for EV production, it is possible to genetically engineer these cell lines to produce EVs with the desired characteristics [41]. In sum, combining mass production and bioengineering of plant cell lines, we could obtain desired EVs in a clean and safe environment.
Studies of plant cell line-derived EVs are recent, but they have already shown some potential [5,6,7,8,9,42]. Most of these aimed to characterize EVs from different types of cell lines. EVs from a Norway spruce cell line were suggested to be involved in the transport of enzymes responsible for lignin formation [5]. EVs from an Arabidopsis thaliana cell line contained proteins important for cell wall biogenesis and pathogen defense mechanisms [8]. EV characterization and their uptake by plant and rat cells was achieved in a tobacco cell line [9]. This study was followed by the characterization and loading of EVs from different sources, namely, callus, cell liquid culture and leaf apoplast [42]. Kırbaş and colleagues (2024) isolated EVs from Stevia rebaudiana and Vaccaria hispanica cell suspensions. These were characterized and an uptake assay performed, where these were successfully taken up by T lymphocyte cell lines after 2-hour incubation [6]. EVs from ginseng showed potential by enhancing skin regeneration, and in addition, it was suggested that they could be potential candidates for drug delivery in therapeutics [7]. From all these studies, one common conclusion was that EVs from cell lines can potentially be up-scaled and mass-produced for biotechnological applications.
As observed in TEM imaging (Figure 2a), these are nano-sized structures with a pronounced bilipid layer and are very similar to those found in other plant isolated EVs. [8,30]. Sometimes artefacts might appear due to TEM sample preparation or EV isolation method, like the cup shape present in some of the imaged EVs, instead of its original round shape [14]. We also noticed that sometimes other structures were present, which could potentially be caused by debris, image noise, dye remains, or the presence of co-isolated small protein complexes or lipoproteins [43,44]. EVs are also known to aggregate in big clumps, as observed in Figure 2b. This could lead to erroneous readings during NTA, hence the need to characterize EVs through TEM. As shown by Kocholatá and colleagues (2022), to avoid these artefacts, trehalose can be added to the EV sample to separate the EVs [9]. As far as size is concerned, our EVs have a mean size of 142 nm, which is similar to that observed in Norway spruce cell lines, where most of the EVs were below the 150 nm threshold [5]. Moreover, Arabidopsis thaliana [8], ginseng [7], and tomato [9] mean sizes were closer to 200 nm, and the populations seem highly heterogeneous. These differences may be caused either by the methodology employed in the studies or by the researched organisms that can have differences in their physiology and EV secretion [7].
In the present work, we observed that our callus produced 2.5 x 1010 particles/mL. In tobacco, it was seen that isolated callus EVs amounted to 1.95 × 1010 [42]. They also observed that suspension cultures had 5.70 × 109 and apoplastic fluid from leaves, 1.24 × 109 [42], which are way below the ones obtained from callus cultures. Another example is from EVs isolated from Arabidopsis thaliana callus, which obtained 1.8 x 1010 particles/g [8]. These results are very similar to the ones we obtained, confirming that this is a good system for EV isolation from a safe and clean source.
Calli are known for their high proliferation and enhanced metabolic activity [45,46,47,48], which can be good indicators of high EV production. Furthermore, callus induction and growth are subjected to stress, either by plant growth regulators or abiotic stress [3], which are known to induce the production of EV [30]. In sum, we believe that the high proliferation, enhanced metabolic activity, and stress exposure are great mechanisms to achieve high EVs production, making poplar callus a great system for EV production.
Although EV production from poplar cell lines offers considerable advantages, scaling this process for larger-scale applications remains a critical challenge. In the present work, EVs were isolated from callus cultured on solid media; however, adoption of liquid-culture systems could similarly facilitate EV recovery at increased scale [6,8,9].
In the future, it would be important to test the growth of poplar cell lines in liquid media (e.g., bioreactor), followed by EV extraction, as it would be much easier to handle and harvest from a bioreactor with litres of media growing cell suspensions. Considering space, feasibility, handling and time [42], we think growing cell suspensions in bioreactors is the most objective way for upscaling.
The loading methods are very dependent on the size of the RNA or DNA molecules [49]. siRNAs and microRNAs (miRNAs) were shown to be easily loaded through sonication, co-incubation, electroporation, and other methods [50]. Larger molecules, of sizes between 100 to 250 bp, seem to be loaded through electroporation and specialized methods, although their efficiency is not great [51]. Bigger RNAs like mRNAs need more specialized methods and advanced loading techniques [52]. In our case, the RNA is around 200 bp, which could explain the reason why sonication, co-incubation and freeze-thaw had low loading, as no RNA from these EVs was taken up by the hyphae. Despite that, optimization of these methods could yield better results and should be pursued.
Electroporation at 400 V produced one of the best results in both loading and uptaking by fungal hyphae. This further corroborates that the hyphae are responsible for the RNA internalization from the previously loaded EVs [28]. The RNA uptake by the fungal cell is very interesting, as it could mean that EVs from poplar cell lines could be used to protect, transport and deliver RNA molecules of such size (≤220 bp) and probably siRNAs and miRNAs that are smaller, and easier to load. It was demonstrated that smaller RNAs could be loaded into EVs from a tomato cell line, through long incubation and sonication [42] whereas electroporation was the least effective method.
Overall, the results show some promise that poplar EVs can be loaded with RNA, protect their contents and deliver their cargos into fungi hyphae. Despite the results, further experiments should include loading of smaller RNAs into poplar EVs and verify if these can be taken up by fungal cells. Going even further, the effectiveness of such RNAs should be assessed when loaded into EVs, since it could be thwarted by either the loading method or by the fact that it could remain inside the EVs and not be delivered into its targets [51,53].
As new techniques are emerging in the RNAi field in crop protection [28], it is also time to think about how we can increase their stability and effectiveness. siRNAs and RNAs in general are prone to degradation in the environment and foreign organisms [24], and if we wish to employ them in crop protection, their stability must be improved. EVs are reliable nanocarriers for efficient protection and delivery of RNA, with low toxicity, and should be considered as strong contenders for the improvement of RNAi techniques.
With this work, we showed that EVs from poplar cell lines can be easily isolated in relatively high amounts. Coupling mass production, for instance, in bioreactors, and genetic transformation, we could create an ideal system to produce specialized EVs. These nano-sized structures are known for their reliability in biomolecule transport and could be key in future RNAi-based techniques or drug delivery. We successfully loaded our EVs with fluorescently labelled dsRNAs through electroporation, although other methods could work by further optimizing the processes or using smaller RNAs. In the future, it is something that we should focus on since, given our results, the RNA from the loaded EVs were successfully taken up by the fungal cells, and it would be interesting to test if the same would happen when using smaller RNAs or other molecules. Furthermore, testing functional RNAs loaded into EVs should be focused on, as we believe that EVs not only could improve RNA stability but also their availability and delivery. To finalize, with this work, we hope it can contribute to the advancement of EV characterization and application in modern and future biotechnological techniques.
Author Contributions
Conceptualization, M.R. and S.C. (Sandra Correia); methodology, M.R. P.R. and S.C. (Sandra Correia); investigation, M.R., S.C. (Sandra Caeiro) and P.R. (Pedro Rosa); data analysis, M.R. and P.R.; resources, C.A. and S.C. (Sandra Correia); writing—original draft preparation, M.R.; writing—review and editing, all authors. All authors have read and agreed to the published version of the manuscript.”.
Funding
This research was supported by FCT - Fundação para a Ciência e Tecnologia, I.P., in the framework of the Project UIDB/04004/2025 - Centre for Functional Ecology - Science for the People & the Planet, and Associated Laboratory TERRA LA/P/0092/2020 financed by FCT/MCTES through national funds (PIDDAC), and by PRR Missão Interface (01/C05-i02/2022.P242) granted to InnovPlantProtect. Miguel Rito is the recipient of the fellowship SFRH/BD/09312/2020 from the FCT/MCTES.
Data Availability Statement
The data presented in this study are available in the article and from the corresponding author.
Acknowledgments
We would like to acknowledge the help of Teresa Rodrigues, Teresa Ribeiro and Professor Mónica Zuzarte from the Coimbra Institute for Clinical and Biomedical Research (iCBR) for the data acquired through Nanoparticle Track Analysis and Transmission Electron Microscopy. For the confocal imaging, we would like to thank Professor Luísa Cortes from CNC-UC - Center for Neuroscience and Cell Biology of the University of Coimbra.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| RNAi | RNA interference |
| siRNA | small interfering RNAs |
| miRNA | micro RNAs |
| 2,4-D | 2,4-dichlorophenoxyacetic acid |
| BAP | 6-Benzylaminopurine |
| VIB | Vesicle Isolation Buffer |
| AWF | Apoplastic Washing Fluid |
| NTA | Nanoparticle Track analysis |
| TEM | Transmission Electron Microscopy |
| MNase | Micrococcal Nuclease |
References
- Efferth, T. Biotechnology applications of plant callus cultures. Engineering 2019, 5, 50-59. [CrossRef]
- Long, Y.; Yang, Y.; Pan, G.; Shen, Y. New insights into tissue culture plant-regeneration mechanisms. Front. Plant Sci. 2022, 13, 926752. [CrossRef]
- Nic-Can, G.I.; Avilez-Montalvo, J.R.; Aviles-Montalvo, R.N.; Márquez-López, R.E.; Mellado-Mojica, E.; Galaz-Ávalos, R.M.; Loyola-Vargas, V.M. The relationship between stress and somatic embryogenesis. In Somatic embryogenesis: fundamental aspects and applications, 1st ed.; Loyola-Vargas, V., Ochoa-Alejo, N. Eds.; Springer: Cham, 2016; pp. 151-170. [CrossRef]
- Wawrosch, C.; Zotchev, S.B. Production of bioactive plant secondary metabolites through in vitro technologies—status and outlook. Appl. Microbiol. Biotechnol. 2021, 105, 6649-6668. [CrossRef]
- Kankaanpää, S.; Väisänen, E.; Goeminne, G.; Soliymani, R.; Desmet, S.; Samoylenko, A.; Vainio, S.; Wingsle, G.; Boerjan, W.; Vanholme, R.; Kärkönen, A. Extracellular vesicles of Norway spruce contain precursors and enzymes for lignin formation and salicylic acid. Plant Physiol. 2024, 196, 788–809. [CrossRef]
- Kırbaş, O.K.; Sağraç, D.; Çiftçi, Ö.C.; Özdemir, G.; Öztürkoğlu, D.; Bozkurt, B.T.; Derman, Ü.C.; Taşkan, E.; Taşlı, P.N.; Özdemir, B.S; Şahin, F. Unveiling the potential: Extracellular vesicles from plant cell suspension cultures as a promising source. BioFactors 2025, 51, e2090. [CrossRef]
- Park, H. Y.; Kang, M. H.; Lee, G.; Kim, J. W. Enhancement of skin regeneration through activation of TGF-β/SMAD signaling pathway by Panax ginseng meyer non-edible callus-derived extracellular vesicles. J. Ginseng Res. 2024, 49, 34-41. [CrossRef]
- Yugay, Y.; Tsydeneshieva, Z.; Rusapetova, T.; Grischenko, O.; Mironova, A.; Bulgakov, D.; Silant’ev, V.; Tchernoded, G.; Bulgakov, V.; Shkryl, Y. Isolation and Characterization of Extracellular Vesicles from Arabidopsis thaliana Cell Culture and Investigation of the Specificities of Their Biogenesis. Plants 2023, 12, 3604. [CrossRef]
- Kocholatá, M.; Prusova, M.; Malinska, H. A.; Maly, J.; Janouskova, O. Comparison of two isolation methods of tobacco-derived extracellular vesicles, their characterization and uptake by plant and rat cells. Sci. Rep. 2022, 12, 19896. [CrossRef]
- Vader, P.; Mol, E.A.; Pasterkamp, G.; Schiffelers, R.M. Extracellular vesicles for drug delivery. Adv. Drug Deliv. Rev. 2016, 106, 148-156. [CrossRef]
- Paganini, C.; Capasso Palmiero, U.; Pocsfalvi, G.; Touzet, N.; Bongiovanni, A.; Arosio, P. Scalable production and isolation of extracellular vesicles: available sources and lessons from current industrial bioprocesses. Biotechnol. J. 2019, 14, 1800528. [CrossRef]
- Giancaterino, S.; Boi, C. Alternative biological sources for extracellular vesicles production and purification strategies for process scale-up. Biotechnol. Adv. 2023, 63, 108092. [CrossRef]
- Rome, S. Biological properties of plant-derived extracellular vesicles. Food Funct. 2019, 10, 529-538. [CrossRef]
- van Niel, G.; d'Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213-228. [CrossRef]
- Cui, Y.; Gao, J.; He, Y.; Jiang, L. Plant extracellular vesicles. Protoplasma 2020, 257, 3–12. [CrossRef]
- Lian, M.Q.; Chng, W.H.; Liang, J.; Yeo, H.Q.; Lee, C.K.; Belaid, M.; Tollemeto, M.; Wacker, M.G.; Czarny, B.; Pastorin, G. Plant-derived extracellular vesicles: Recent advancements and current challenges on their use for biomedical applications. J. Extracell. Vesicles 2022, 11, 12283. [CrossRef]
- Nemati, M.; Singh, B.; Mir, R.A.; Nemati, M.; Babaei, A.; Ahmadi, M.; Rasmi, Y.; Golezani, A.G.; Rezaie, J. Plant-derived extracellular vesicles: a novel nanomedicine approach with advantages and challenges. Cell Commun. Signal 2022, 20, 69. [CrossRef]
- Woith, E.; Guerriero, G.; Hausman, J.F.; Renaut, J.; Leclercq, C.C.; Weise, C.; Legay, S.; Weng, A.; Melzig, M.F. Plant extracellular vesicles and nanovesicles: focus on secondary metabolites, proteins and lipids with perspectives on their potential and sources. Int. J. Mol. Sci. 2021, 22, 3719. [CrossRef]
- Qiang, W.; Li, J.; Ruan, R.; Li, Q.; Zhang, X.; Yan, A.; Zhu, H. Plant-derived extracellular vesicles as a promising anti-tumor approach: a comprehensive assessment of effectiveness, safety, and mechanisms. Phytomedicine 2024, 130, 155750. [CrossRef]
- Lykogianni, M.; Bempelou, E.; Karamaouna, F.; Aliferis, K.A. Do pesticides promote or hinder sustainability in agriculture? The challenge of sustainable use of pesticides in modern agriculture. Sci. Total Environ. 2021, 795, 148625. [CrossRef]
- Niu, D.; Hamby, R.; Sanchez, J.N.; Cai, Q.; Yan, Q.; Jin, H. RNAs—a new frontier in crop protection. Curr. Opin. Biotechnol. 2021, 70, 204-212. [CrossRef]
- Kong, X.; Yang, M.; Le, B.H.; He, W.; Hou, Y. The master role of siRNAs in plant immunity. Mol. Plant Pathol. 2022, 23, 1565-1574. [CrossRef]
- Rodrigues, T.B.; Mishra, S.K.; Sridharan, K.; Barnes, E.R.; Alyokhin, A.; Tuttle, R.; Kokulapalan, W.; Garby, D.; Skizim, N.J.; Tang, Y.W.; Manley, B. First sprayable double-stranded RNA-based biopesticide product targets proteasome subunit beta type-5 in Colorado potato beetle (Leptinotarsa decemlineata). Front. Plant Sci. 2021, 12, 728652. [CrossRef]
- Zhao, J.H.; Liu, Q.Y.; Xie, Z.M.; Guo, H.S. Exploring the challenges of RNAi-based strategies for crop protection. Adv. Biotechnol. 2024, 2, 23. [CrossRef]
- Zhao, Y.; Zhou, Y.; Xu, J.; Fan, S.; Zhu, N.; Meng, Q.; Dai, S.; Yuan, X. Cross-Kingdom RNA Transport Based on Extracellular Vesicles Provides Innovative Tools for Plant Protection. Plants 2024, 13, 2712. [CrossRef]
- Niño-Sánchez, J.; Sambasivam, P.T.; Sawyer, A.; Hamby, R.; Chen, A.; Czislowski, E.; Li, P.; Manzie, N.; Gardiner, D.M.; Ford, R.; Xu, Z.P. BioClay™ prolongs RNA interference-mediated crop protection against Botrytis cinerea. J. Integr. Plant Biol. 2022, 64, 2187-2198. [CrossRef]
- Bi, K.; Liang, Y.; Mengiste, T.; Sharon, A. Killing softly: a roadmap of Botrytis cinerea pathogenicity. Trends Plant Sci. 2023, 28, 211-222. 10.1016/j.tplants.2022.08.024.
- Wang, M.; Weiberg, A.; Lin, F.M.; Thomma, B.P.; Huang, H.D.; Jin, H. Bidirectional cross-kingdom RNAi and fungal uptake of external RNAs confer plant protection. Nat. plants 2016, 2, 1-10. [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant 1962, 15, 473-497. 10.1111/j.1399-30541962tb08052.x.
- Rutter, B. D.; Innes, R. W. Extracellular vesicles isolated from the leaf apoplast carry stress-response proteins. Plant Physiol. 2017, 173, 728–741. [CrossRef]
- Théry, C.; Amigorena, S.; Raposo, G.; Clayton, A. Isolation and characterization of exosomes from cell culture supernatants and biological fluids. Curr. Protoc. Cell Biol. 2006, 30, 3-22. [CrossRef]
- Hamby, R.; Wang, M.; Qiao, L.; Jin, H. Synthesizing Fluorescently Labeled dsRNAs and sRNAs to Visualize Fungal RNA Uptake. In RNA Tagging: Methods and Protocols, 1st ed.; Heinlein, M., Eds.; Springer US: NY, USA, 2020; Vol. 2166, pp . 215–225. [CrossRef]
- Aranda, P. S.; LaJoie, D. M.; Jorcyk, C. L. Bleach Gel: A Simple Agarose Gel for Analyzing RNA Quality. Electrophoresis 2012, 33, 366–369. [CrossRef]
- Kim, M.S.; Haney, M.J.; Zhao, Y.; Mahajan, V.; Deygen, I.; Klyachko, N.L.; Inskoe, E.; Piroyan, A.; Sokolsky, M.; Okolie, O.; Hingtgen, S.D. Development of exosome-encapsulated paclitaxel to overcome MDR in cancer cells. Nanomed. Nanotechnol. Biol. Med. 2016, 12, 655-664. [CrossRef]
- Yang, P.; Zhao, Z.; Virag, A.; Becker, T.; Zhao, L.; Liu, W.; Xia, Y. Botrytis cinerea in vivo Inoculation Assays for Early-, Middle-and Late-stage Strawberries. Bio-protocol 2023, 13, e4859. [CrossRef]
- Michler, C.H.; and Bauer, E.O. High frequency somatic embryogenesis from leaf tissue of Populus spp. Plant Sci. 1991, 77, 111-118. [CrossRef]
- Lelu-Walter, M.A.; Thompson, D.; Harvengt, L.; Sanchez, L.; Toribio, M.; Pâques, L.E. Somatic embryogenesis in forestry with a focus on Europe: state-of-the-art, benefits, challenges and future direction. Tree Genet. Genomes 2013, 9, 883-899. [CrossRef]
- Arencibia, A.D.; Gómez, A.; Poblete, M.; Vergara, C. High-performance micropropagation of dendroenergetic poplar hybrids in photomixotrophic Temporary Immersion Bioreactors (TIBs). Ind. Crops Prod. 2017, 96, 102-109. [CrossRef]
- Kim, S.; Kim, Y.O.; Lee, Y.; Choi, I.; Joshi, C.P.; Lee, K.; Bae, H.J. The transgenic poplar as an efficient bioreactor system for the production of xylanase. Biosci. Biotechnol. Biochem. 2012, 76, 1140-1145. [CrossRef]
- Verdú-Navarro, F.; Moreno-Cid, J.A.; Weiss, J.; Egea-Cortines, M. The advent of plant cells in bioreactors. Front. Plant Sci. 2023, 14, 1310405. [CrossRef]
- Martínez, M.; Corredoira, E. Recent Advances in Plant Somatic Embryogenesis: Where We Stand and Where to Go? Int. J. Mol. Sci. 2024, 25, 8912. [CrossRef]
- Kocholatá, M.; Malý, J.; Kříženecká, S.; Janoušková, O. Diversity of extracellular vesicles derived from calli, cell culture and apoplastic fluid of tobacco. Sci. Rep. 2024, 14, 1-13. [CrossRef]
- Rutter, B. D.; Innes, R.W. Growing pains: addressing the pitfalls of plant extracellular vesicle research. New Phytol. 2020, 228, 1505-1510. [CrossRef]
- Rikkert, L.G.; Nieuwland, R.; Terstappen, L.W.M.M.; Coumans, F.A.W. Quality of extracellular vesicle images by transmission electron microscopy is operator and protocol dependent. J. Extracell. Vesicles 2019, 8, 1555419. [CrossRef]
- Duan, X.; Chen, L.; Liu, Y.; Chen, H.; Wang, F.; Hu, Y. Integrated physicochemical, hormonal, and transcriptomic analysis reveals the underlying mechanism of callus formation in Pinellia ternata hydroponic cuttings. Front. Plant Sci. 2023, 14, p.1189499. [CrossRef]
- Wen, S.S.; Ge, X.L.; Wang, R.; Yang, H.F.; Bai, Y.E.; Guo, Y.H.; Zhang, J.; Lu, M.Z.; Zhao, S.T.; Wang, L.Q. An efficient Agrobacterium-mediated transformation method for hybrid poplar 84K (Populus alba× P. glandulosa) using calli as explants. Int. J. Mol. Sci. 2022. 23, 2216. [CrossRef]
- Ge, F.; Hu, H.; Huang, X.; Zhang, Y.; Wang, Y.; Li, Z.; Zou, C.; Peng, H.; Li, L.; Gao, S.; Pan, G. Metabolomic and proteomic analysis of maize embryonic callus induced from immature embryo. Sci. Rep. 2017, 7, 1004. [CrossRef]
- Kumari, A.; Ray, K.; Sadhna, S.; Pandey, A. K.; Sreelakshmi, Y.; Sharma, R. Metabolomic homeostasis shifts after callus formation and shoot regeneration in tomato. PloS One 2017, 12, e0176978. 10.1371/journal.pone.0176978.
- Amiri, A.; Bagherifar, R.; Ansari Dezfouli, E.; Kiaie, S.H.; Jafari, R.; Ramezani, R. Exosomes as bio-inspired nanocarriers for RNA delivery: preparation and applications. J. Transl. Med. 2022, 20, 125. [CrossRef]
- Donoso-Quezada, J.; Ayala-Mar, S.; González-Valdez, J. State-of-the-art exosome loading and functionalization techniques for enhanced therapeutics: a review. Crit. Rev. Biotechnol. 2020, 40, 804-820. [CrossRef]
- Nordmeier, S.; Ke, W.; Afonin, K. A.; Portnoy, V. Exosome mediated delivery of functional nucleic acid nanoparticles (NANPs). Nanomedicine 2020, 30, 102285.
- Iqbal, Z.; Rehman, K.; Mahmood, A.; Shabbir, M.; Liang, Y.; Duan, L.; Zeng, H. Exosome for mRNA delivery: strategies and therapeutic applications. J. Nanobiotechnology 2024, 22, 395. [CrossRef]
- de Voogt, W.S.; Tanenbaum, M.E.; Vader, P. Illuminating RNA trafficking and functional delivery by extracellular vesicles. Adv. Drug Deliv. Rev. 2021, 174, 250-264. [CrossRef]
Figure 1.
Induced callus from poplar. (a) Leaf tissues dedifferentiation in the induction medium after 4 weeks of culture. (b) Isolated and proliferating cell line after a 4-week subculture, before EV isolation. (c)/(d) Cell line close-up plans, showing the homogenous and friable aspect of the callus with a yellowish color.
Figure 1.
Induced callus from poplar. (a) Leaf tissues dedifferentiation in the induction medium after 4 weeks of culture. (b) Isolated and proliferating cell line after a 4-week subculture, before EV isolation. (c)/(d) Cell line close-up plans, showing the homogenous and friable aspect of the callus with a yellowish color.
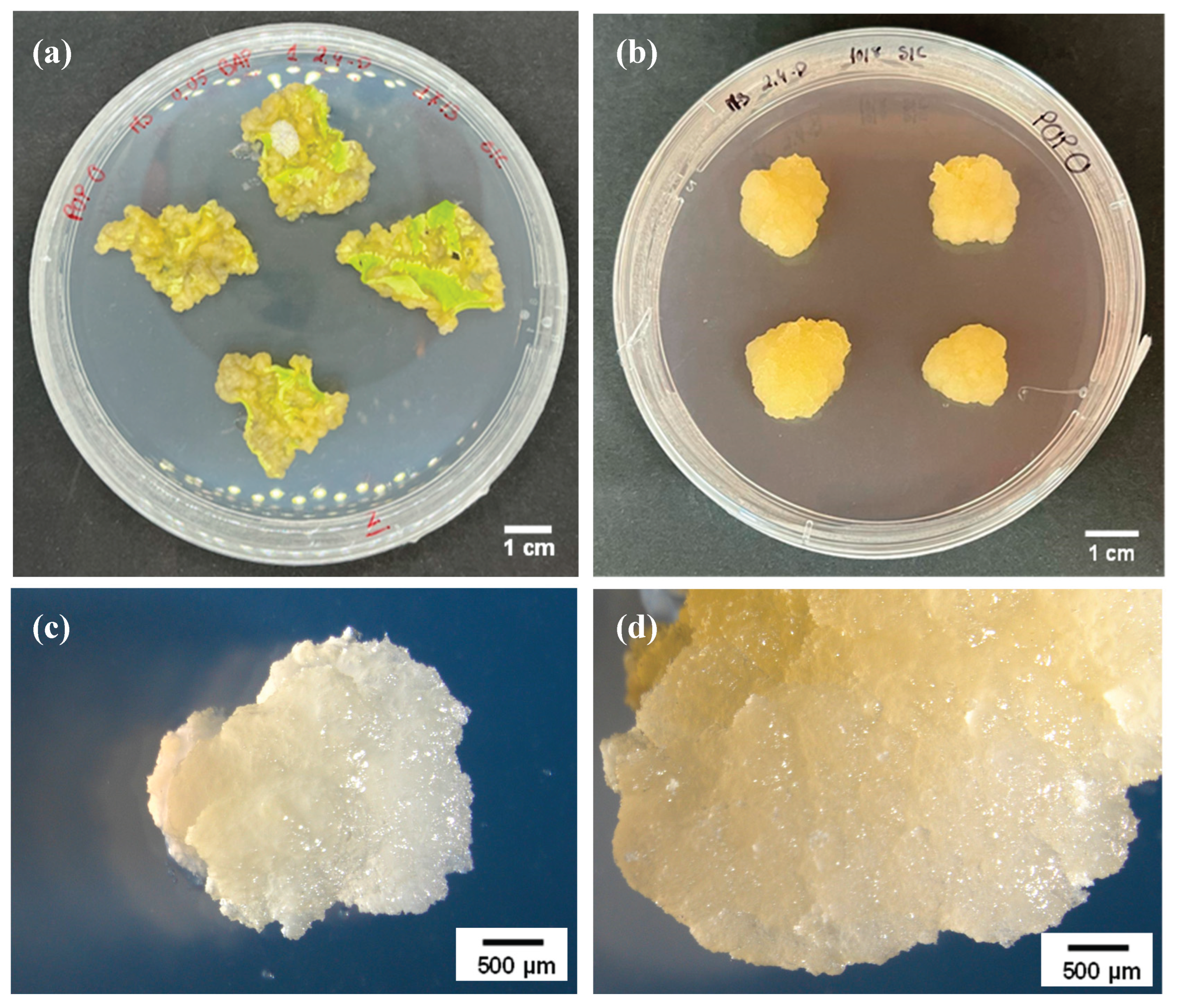
Figure 2.
Characterization of the EV population from poplar callus (a) – Isolated EVs seen through transmission electron microscopy. *EVs are usually round-shaped with a bilipid layer. ** EVs can collapse and acquire different shapes, such as the common cup shape. (b) – Nanoparticle track analysis. The NTA graph shows a big peak at 121 nm and a smaller at 211 nm. Most of the EVs analysed were smaller than 200 nm, and the total concentration of EVs in our sample was 2.5 x 1010 particles/mL. CS – cup shape; BL – bilipid layer.
Figure 2.
Characterization of the EV population from poplar callus (a) – Isolated EVs seen through transmission electron microscopy. *EVs are usually round-shaped with a bilipid layer. ** EVs can collapse and acquire different shapes, such as the common cup shape. (b) – Nanoparticle track analysis. The NTA graph shows a big peak at 121 nm and a smaller at 211 nm. Most of the EVs analysed were smaller than 200 nm, and the total concentration of EVs in our sample was 2.5 x 1010 particles/mL. CS – cup shape; BL – bilipid layer.
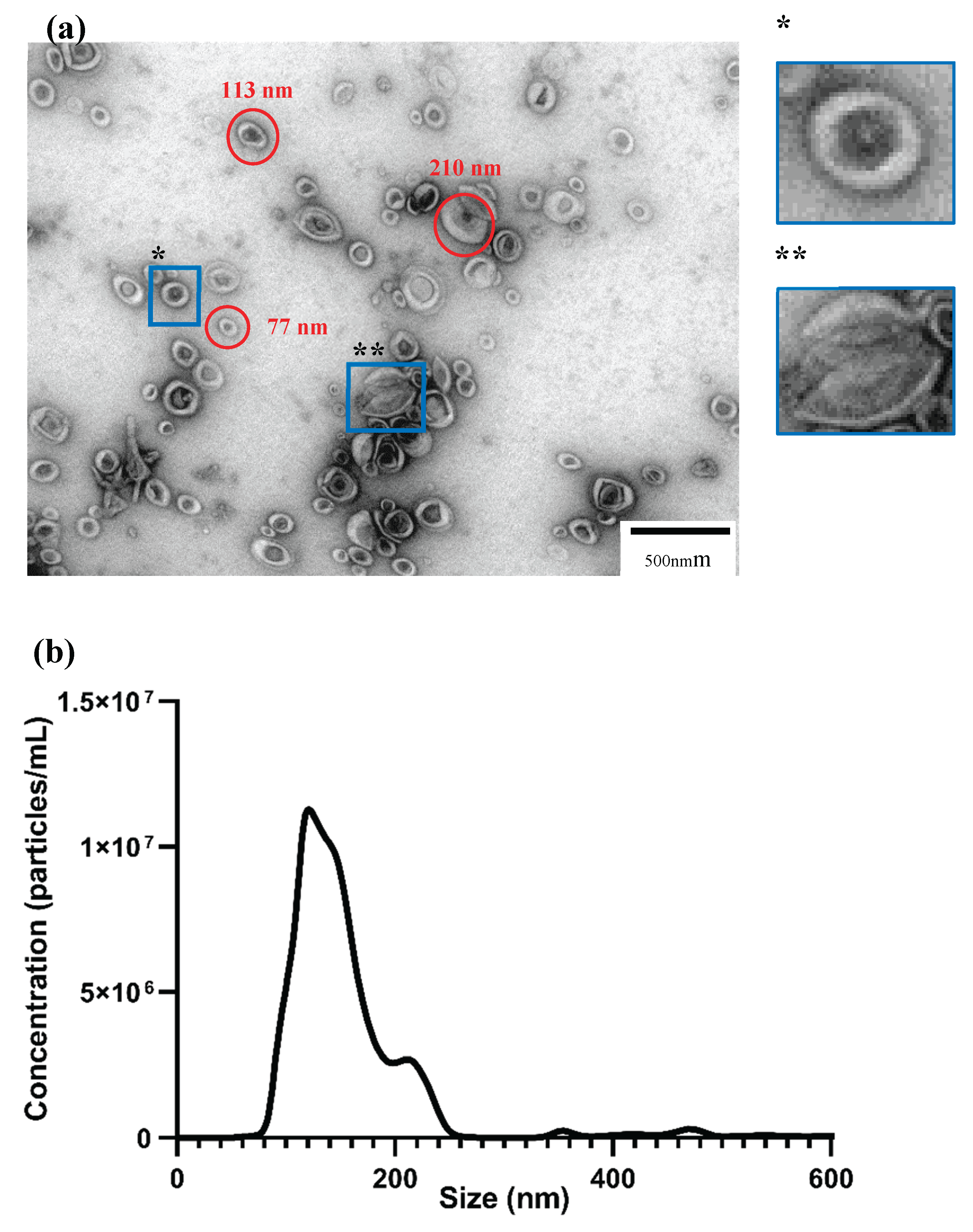
Figure 3.
Loading of dsPDS-F into EVs by electroporation (400V) and uptake by Botrytis cinerea spores. (a) Confocal overlapped bright field and fluorescent image of unloaded EVs. (b) Confocal overlapped bright field and fluorescent images of dsPDS-F-loaded EVs, showing clumps* and dispersed** loaded EVs.
Figure 3.
Loading of dsPDS-F into EVs by electroporation (400V) and uptake by Botrytis cinerea spores. (a) Confocal overlapped bright field and fluorescent image of unloaded EVs. (b) Confocal overlapped bright field and fluorescent images of dsPDS-F-loaded EVs, showing clumps* and dispersed** loaded EVs.
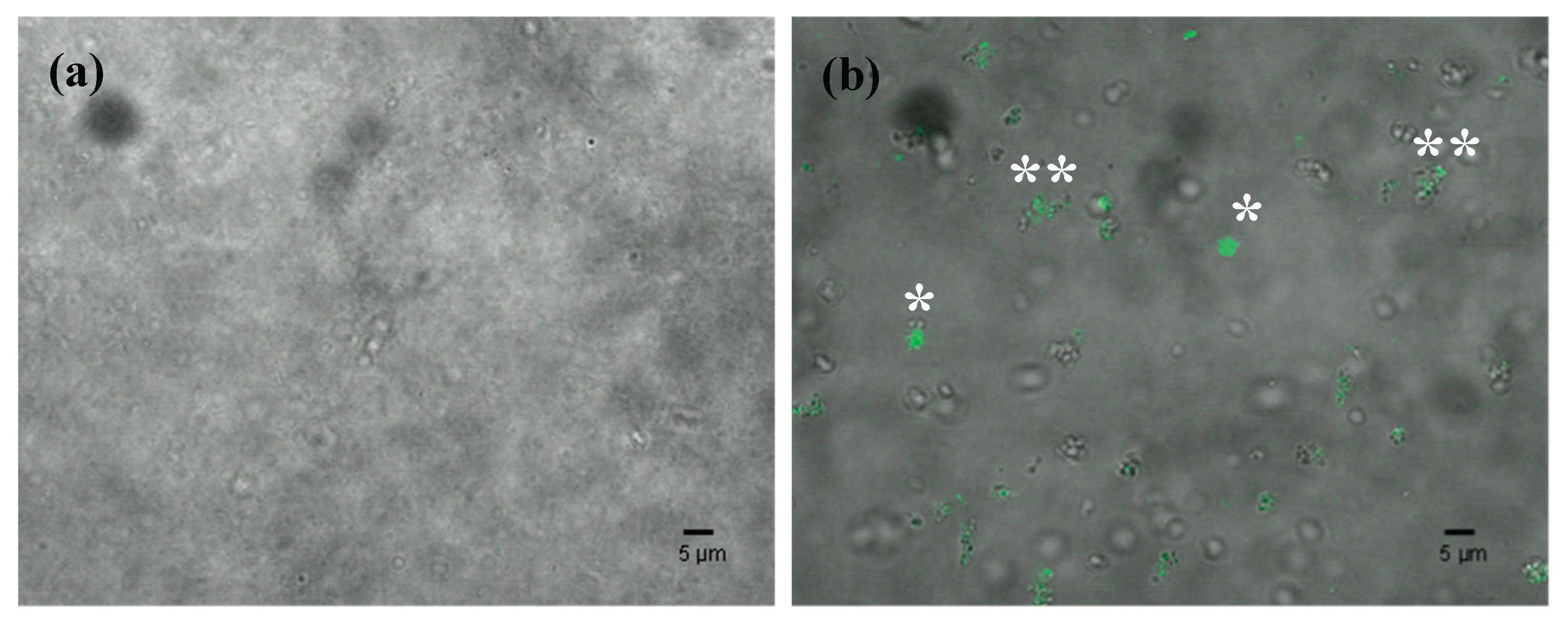
Figure 4.
dsPDS-F uptake by Botrytis cinerea spores. (a) Overlapped bright field and fluorescent images of EP400 EVs incubated with B. cinerea spores. (b) Fluorescent images of EP400 EVs incubated with B. cinerea spores. (c) Overlapped bright field and fluorescent images of sonicated EVs incubated with B. cinerea spores. (d) Fluorescent images of sonicated EVs incubated with B. cinerea spores. Example of no uptake observed.
Figure 4.
dsPDS-F uptake by Botrytis cinerea spores. (a) Overlapped bright field and fluorescent images of EP400 EVs incubated with B. cinerea spores. (b) Fluorescent images of EP400 EVs incubated with B. cinerea spores. (c) Overlapped bright field and fluorescent images of sonicated EVs incubated with B. cinerea spores. (d) Fluorescent images of sonicated EVs incubated with B. cinerea spores. Example of no uptake observed.
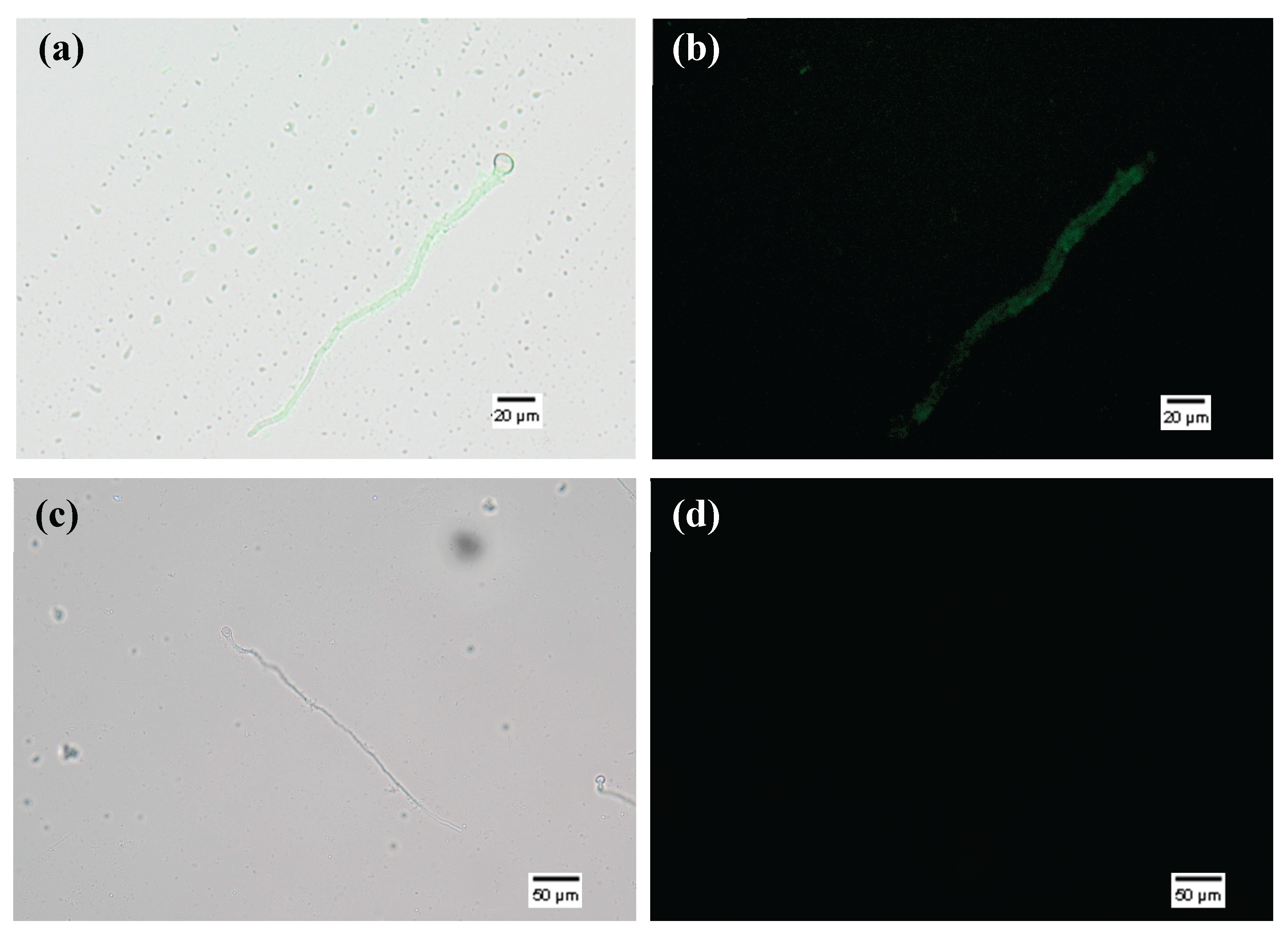

Table 1.
Primers used for IVT (underlined bases: T7 RNA Polymerase promoter).
| Primer Name | Sequence |
|---|---|
| TaPDS Frw | TTTGCTCCAGCAGAGGAATGG |
| TaPDS Frw T7pol | TAATACGACTCACTATAGGTTTGCTCCAGCAGAGGAATGG |
| TaPDS Rev | AAACCCTTCGATCGGTGATCG |
| TaPDS Rev T7pol | TAATACGACTCACTATAGGAAACCCTTCGATCGGTGATCG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



