1. Introduction
Global fisheries and aquaculture production reached a record high of 223.2 million tonnes in 2023, comprising 185.4 million tonnes of aquatic animals and 37.8 million tonnes of algae [
1]. China remains the world’s largest producer, accounting for over 60% of the total output, followed by other major Asian producers such as India, Indonesia, and Vietnam. Finfish account for 90.2% of global aquaculture production, with the majority being freshwater species. Although seaweed production is significantly lower in volume compared to finfish, it plays a critical role in aquaculture, as virtually all macroalgae are cultivated within this sector. Projections suggest that by 2030, aquaculture will supply 53% of global aquatic production, meeting approximately 59% of global demand for aquatic food products [
2]. The growing contribution of aquaculture to global food security highlights the need not only to maintain current production levels but also to invest in the quality, diversity, and environmental sustainability of aquaculture systems.
As the aquaculture industry expands, concerns about waste discharge, resource efficiency, and environmental sustainability are increasingly pressing [
3]. Intensive aquaculture operations have been associated with antibiotic contamination, eutrophication, land degradation, and various other ecological impacts [
4]. Several studies have evaluated the environmental consequences of different aquaculture systems, particularly intensive ones such as shrimp farming in mangrove areas [
5,
6], salmon farming in coastal zones [
7,
8], and carp farming in inland water bodies [
9]. Moreover, many aquaculture systems are highly sensitive to environmental fluctuations and are thus vulnerable to the effects of climate change.
In response to these challenges, Recirculating Aquaculture Systems (RAS) have emerged as a promising adaptive strategy. These systems continuously treat and reuse water, removing ammonia and particulate waste materials through mechanical and biological filtration. RAS technologies eliminate solid waste, convert ammonia and nitrite to nitrate, remove CO₂, regulate pH, control microbial populations, and maintain adequate oxygenation [
3,
10,
11]. Variations in RAS design consider factors such as species requirements, stocking densities, water exchange rates, and system components [
10,
12,
13].
One of the most promising and sustainable approaches within RAS is the implementation of Integrated Multi-Trophic Aquaculture (IMTA). In this system, fed organisms (e.g., fish or shrimp) are co-cultured with extractive species such as bivalves (which absorb organic particulates) and macroalgae (which assimilate dissolved nutrients), creating a more balanced and synergistic biological system [
14]. The waste produced by one trophic level becomes a resource for another, enhancing nutrient recycling, improving water quality, and increasing overall productivity. IMTA systems can generate multiple value-added products, reduce environmental impacts, and promote economic resilience for producers [
15].
Combining IMTA with RAS technology (multi-trophic RAS) further strengthens the sustainability and circularity of aquaculture. In this context, macroalgae not only serve as effective bioremediators but can also be used as feed ingredients for low-trophic level organisms such as sea urchins, which are highly valued for their edible gonads, particularly in Asian markets. The interest in aquaculture-based production of sea urchins has grown significantly in recent years [
1,
16,
17].
Macroalgae of the
Ulva genus has been identified as particularly suitable candidates for integration in multitrophic RAS. Ulva contributes to water quality by increasing pH and oxygen levels, lowering CO₂ concentrations, and aiding in the removal of ammonia and nitrates [
18]. Moreover, it provides a balanced nutritional profile for sea urchins, supporting good growth and survival rates throughout the cultivation period. Nutritionally, Ulva is notable for its high content of protein, minerals, dietary fiber, and omega-3 fatty acids [
19,
20,
21]. Additionally, its bioactive compounds, such as ulvans and polyphenols, have shown potential to improve immune function and reduce susceptibility to parasitic infections in fish, thus contributing positively to overall fish health [
22,
23,
24].
As aquaculture expands to meet rising seafood demand and reduce pressure on wild stocks, production systems have become increasingly intensive, often involving higher stocking densities, limited water renewal, and frequent handling of fish [
25]. These conditions elevate physiological stress in farmed fish, compromising their immune response and increasing vulnerability to infectious agents. Consequently, parasitic outbreaks have become a frequent and persistent challenge in intensive aquaculture environments [
25,
26].
Parasitic infections now rank among the most pressing concerns for fish health and farm productivity, with some parasites causing substantial and lasting impacts across aquaculture sectors worldwide [
27,
28]. Within this context, monogenea have emerged as particularly problematic, given their high transmission potential and significant pathogenic effects on high-value species such as gilthead seabream (
Sparus aurata) [
25,
29,
30].
Monogenea are ectoparasitic flatworms with a direct life cycle that includes three key stages: resistant (eggs), infective (free-swimming larvae), and pathogenic (attached juveniles/adults) [
29]. Adult monogeneans release eggs into the water column, which frequently become entangled on fibrous surfaces within aquaculture systems, allowing them to persist and maintain recurring infection cycles [
28,
31]. Once developed, the larvae hatches and must quickly locate and attach to a suitable host, typically targeting the gills, skin, or fins, where they cause direct tissue damage and facilitate secondary infections, particularly under intensive rearing conditions [
32,
33].
S. aurata, one of the most economically important marine species cultivated in Europe [
2], is especially susceptible to monogenean infections, which represent a significant health and welfare challenge in both hatchery and grow-out phases [
34,
35]. Environmental factors, especially water temperature, are known to influence infection dynamics. Warmer conditions accelerate parasite development and enhance transmission rates [
36] but outbreaks can also occur during colder months due to host immunosuppression [
37].
In the search for effective, sustainable parasite control strategies, macroalgae-derived bioactive compounds have gained increasing attention. These natural substances exhibit antimicrobial, anti-inflammatory, and antiparasitic properties, offering promising eco-friendly alternatives to conventional chemical treatments [
27,
28]. Their antiparasitic mechanisms may include inhibition of parasite adhesion to host tissues or direct toxicity to parasite larvae or adults [
27,
38]. When integrated into multitrophic RAS systems, macroalgae such as
U. ohnoi may contribute not only to water bioremediation and nutritional enrichment, but also to the health protection of co-cultured species, including gilthead seabream.
Recirculating Aquaculture Systems (RAS) coupled with Integrated Multi-Trophic Aquaculture (IMTA) demonstrate significant potential in alignment with the European Union’s strategic objectives for zero waste and circular economy
[39]. However, assessing their applicability and economic viability is essential to ensure these practices become financially sustainable.
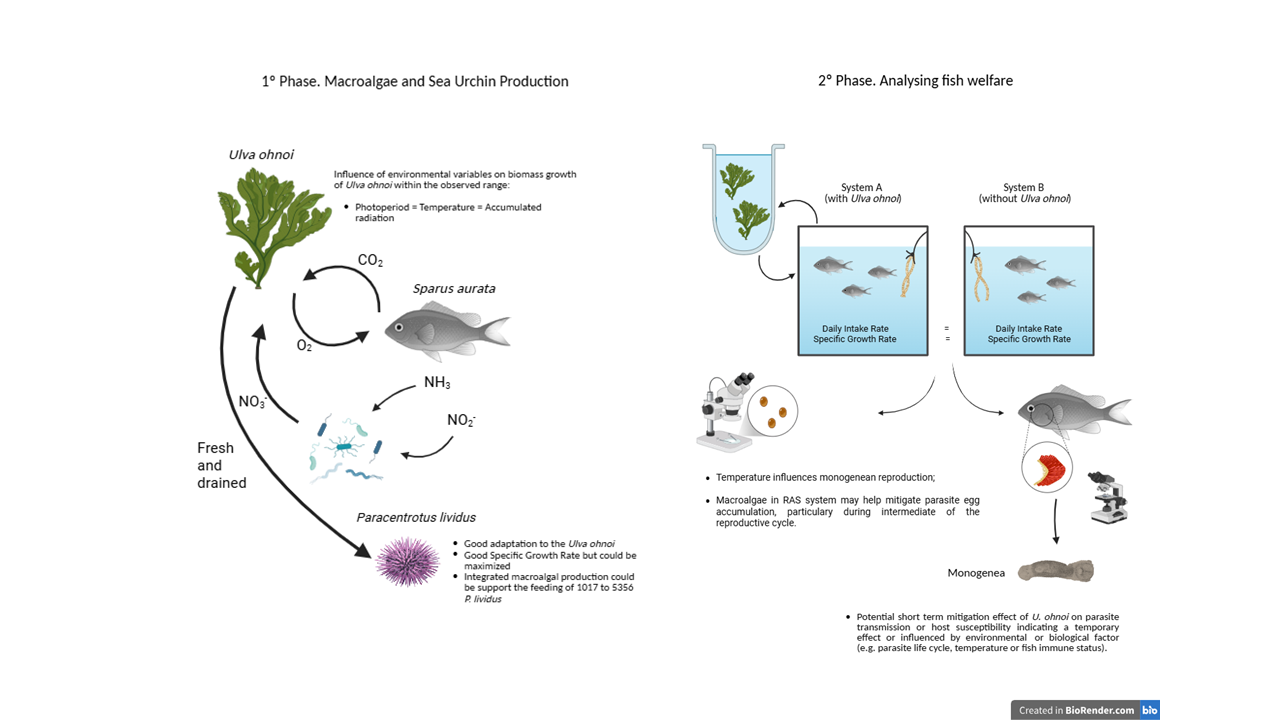
The aim of this study is to evaluate the performance of a multitrophic RAS system combining gilthead seabream (S. aurata) and macroalgae (Ulva ohnoi), focusing on three main aspects: (i) the productivity of Ulva within the system; (ii) the suitability of cultivated U. ohnoi as a feed component for sea urchins (Paracentrotus lividus); and (iii) the potential impact of this integrated system on the health condition of the fish.
2. Material and Methods
2.1. Integrated Recirculation System (RAS)
In this study, a recirculating aquaculture system (RAS) was tested for the integrated production of fish (Sparus aurata) and macroalgae (Ulva ohnoi), with the macroalgal biomass used as feed for sea urchins (Paracentrotus lividus) cultured independently from this system. To meet the objectives of the study, two identical and autonomous RAS units were assembled.
In the first phase of the study (macroalgae and sea urchin production), only one RAS unit was used to collect data on macroalgae productivity and its application in sea urchin diets. In the second phase (impact of integrated macroalgae production on fish health and welfare), the second RAS unit was used, in which the macroalgae cultivation circuit was deactivated.
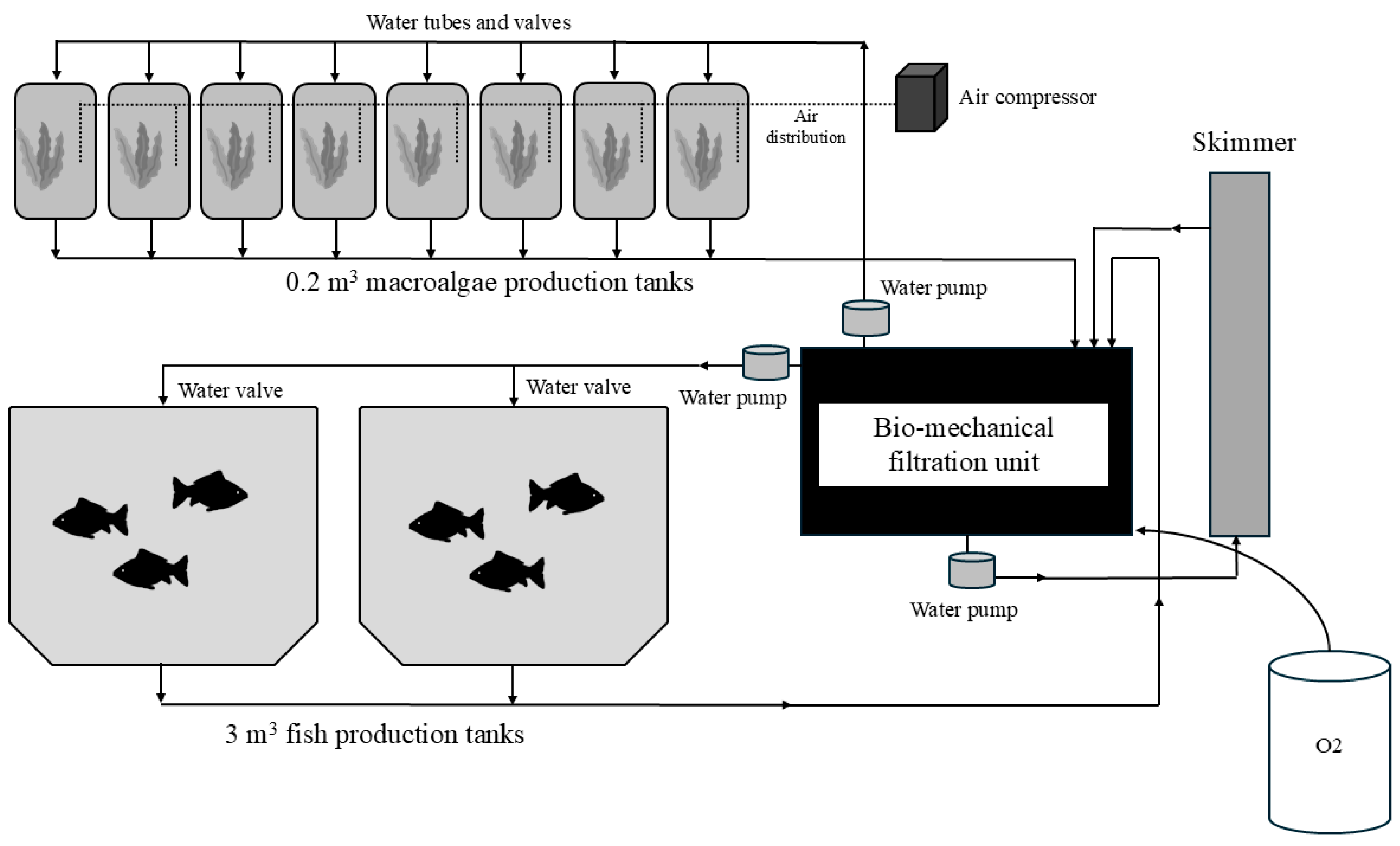
Each RAS comprised two 3 m³ conical-bottom fiberglass tanks for fish rearing, eight 0.2 m³ cylindrical plastic tanks (90 cm tall and 54 cm in diameter) for macroalgae cultivation a combined bio-mechanical filtration unit, and a foam fractionator (protein skimmer).
In each system, the two fish tanks received water from the filtration unit, with an average renewal rate of approximately 30% per hour. Water exited from the bottom of each tank and was regulated by a standpipe system controlling the water column height. Effluent from the fish tanks was directed through a network of PVC pipes into the filtration unit.
Upon entering the filtration tank, the water flowed through a cascade-based mechanical filtration stage composed of a poly-fiber filter pad and a layer of reticulated aquatic filter foam. Within the same reservoir, the water circulated through a biological filter consisting of bio-balls and was oxygenated via diffused aeration using air stones.
Each filtration tank was equipped with a water inlet valve connected to the main reservoir of the facility. The opening of this valve was controlled by a float switch installed inside the filtration tank, ensuring automatic compensation for water losses within the system.
Following filtration, the water was pumped into the macroalgae cultivation circuit, which also acted as a bioremediation stage for improving water quality. Each RAS included nine algae culture tanks with a water renewal rate of approximately 320% per hour. These tanks were strongly aerated by perforated PVC structures affixed to the bottom, allowing for uniform air bubble release. Water flow through the algae tanks occurred in series.
After passing through the algae tanks, the water was recirculated back to the bio-mechanical filtration unit, completing the loop. Finally, each system was equipped with a foam fractionator (protein skimmer), operating with an independent water pump, to remove dissolved organic compounds.
Figure 1.
Circuit diagram of the experimental Integrated Recirculation System (RAS).
Figure 1.
Circuit diagram of the experimental Integrated Recirculation System (RAS).
2.2. Macroalgae and Sea Urchin Production
In the first part of the work, the analysis focused on the production of macroalgae and their use as exclusive food for sea urchins. Only one RAS unit was used, with eight macroalgae production tanks and two fish production tanks. The sea urchins were cultivated in a system independent of the RAS. This first trial phase lasted 187 days.
2.2.1. Macroalgae
In the initial phase, two species of marine macroalgae were selected: Ulva ohnoi and Gracilaria gracilis. However, G. gracilis was found to be unsuitable for the developed system, and therefore only U. ohnoi was cultured. The initial biomass of U. ohnoi was collected from earthen ponds at EPPO (Aquaculture Research Center) and was subjected to a cleaning process to remove sediment and associated organisms. The identification of this macroalgae was previously carried out by the Molecular Biology Lab at S2AQUA CoLab (Portugal).
Each 200-liter tank was inoculated with 1 kg of U. ohnoi. Every ten days, a sampling procedure was conducted, consisting of harvesting and weighing the biomass from each tank. After tank cleaning, the tanks were reinoculated with 1 kg of fresh macroalgae biomass. The remaining macroalgae were used as feed for sea urchins. Water flow rates in each tank were periodically measured and adjusted as needed.
2.2.2. Sea Urchin Rearing and Feeding Protocol
A total of 210 sea urchins (Paracentrotus lividus) with an initial average weight of 14.82 ± 1.29 g and a test diameter of 36.84 ± 1.27 mm were selected for this study. All individuals were hatchery-born at EPPO facilities from F1-generation broodstock. The spawning induction and larval rearing protocols are described in Araujo et al. (2020).
The sea urchins were distributed into six plastic boxes (0.57 × 0.38 × 0.08 m), which were placed within two fiberglass raceway-type tanks containing approximately 300 L of water (working volume). These tanks were located indoors at the hatchery building and operated under continuous water flow, with aeration provided by air stones. Environmental parameters, including temperature and dissolved oxygen, were monitored twice daily (morning and afternoon) using a portable multiparameter probe.
Sea urchins were fed exclusively with macroalgae cultivated in the RAS developed for this study. Feeding was carried out ad libitum, with the availability of algae in each box checked daily. Whenever algae were scarce or absent, the boxes were temporarily removed for cleaning, and approximately 100 g of fresh macroalgae was added. This quantity was adjusted based on the consumption observed in the preceding days.
2.2.3. Sea Bream Cultivation Protocol
The RAS unit used in the first part of the work (macroalgae and sea urchin production) included the two sea bream tanks described in
Section 2.1. At this stage, 150 fish were placed in each tank, resulting in a stocking density of 16.33 kg/m3. The fish were reared at EPPO facilities and originated from two separate spawns, one 311 days old and the other 313 days old at the time of introduction into the RAS. During the trial, fish were fed a commercial diet formulated for sea bream at a daily feeding rate of 0.94%. Pellet size was adjusted over time according to fish growth.
2.2.4. Biometric Sampling
For the biometric sampling of sea urchins, weight and test diameter were considered. Weighing was done with a KERN PRS / PRJ precision and analytical balance. To measure the test-diameter the sea urchins were photographed next to a ruler, and the pixel-centimeter conversion was performed using the ImageJ software. An analytical scale with an accuracy of 0.1 g was used to weigh the fish and macroalgae. The length of the fish was measured using an ichthyometer ruler.
2.2.5. Environmental Parameters and Sampling Procedures
Environmental parameters were monitored daily in the RAS and sea urchin rearing systems. Temperature (°C), dissolved oxygen (mg/L), and oxygen saturation (%) were measured twice daily using a HANNA HI9147 multiparameter probe. Global radiation data were obtained from the IPMA meteorological station located at Faro Airport, approximately 12 km from the EPPO facilities. Raw data were processed by technical staff to generate daily total global radiation values (MJ/m2/day). Assuming that around 45–50% of total solar radiation falls within the 400–700 nm range, global radiation values were converted to photosynthetically active radiation (PAR). The daily photon flux available for photosynthesis was estimated by calculating the daily light integral (DLI, mol photons m⁻² d⁻¹) with a conversion factor of 2.1 mol MJ⁻¹.
Supplementary Table S1 displays the entire dataset of estimated PAR values. For each algae sample, pH (Fisher Scientific) and ammonia (NH3) concentrations were measured using Salifert colorimetric test kits to ensure values remained below 0.05 mg/L. Due to the nature of the test, ammonia measurements were primarily used to verify compliance with maximum recommended levels for gilthead sea bream culture. However, theoretical models were used to estimate the excretion of phosphorus and nitrogen and thus assess the relationship between nutrient production and macroalgae production.
2.3. Impact of Integrated Macroalgae Production on Fish Health and Welfare
The second part of the work focused on the impact of integrating macroalgae into an RAS system. Two RAS units were used, differing in whether they integrated a macroalgae production circuit. System A consisted of 2 tanks with fish (S. aurata), 8 macroalgae production tanks (U. onhoi), a filtration unit and skimmer. System B consisted only of 2 fish tanks, a filtration unit and a skimmer. System B therefore did not include a macroalgae circuit. In each system, 110 sea breams were placed in each tank, giving an initial density of 16.9 Kg/m3 for system A and 17.2 Kg/m3 for system B. The average initial weight of the fish was 232.1 ± 0.04 grams. The feeding protocol was like the previous trial. For this work, the periodic collection and weighing of the U. onhoi macroalgae was maintained.
The protocol for monitoring environmental parameters such as temperature, dissolved oxygen and oxygen saturation in the water was also maintained.
2.3.1. Fish Welfare: Ectoparasite Analysis
Seven sampling points were established to evaluate the presence of parasite eggs in the system with and without macroalgae. Cotton strips, with an area of 12 cm², were placed fixed to the water outlet for five days in exactly same position in the four tanks, to facilitate the collection of parasites eggs. After this period, the strips were removed and stored in seawater to preserve the eggs until quantification analysis [
25]. The eggs attached to the strips were observed under a stereomicroscope (Nikon SMZ1000) and counted manually. The fish’s parasite charge was also analysed three times during the experiment. At each sampling point (from systems A and B), 3 fish were randomly caught from each tank and sacrificed by an incision in the spinal column. After the fish died, the two gill arches on the left side were removed and observed under an optical microscope (Nikon ECLIPSE Ci) to quantify the parasites. The level of infection was determined based on three parameters described by Bush et al. (1997).
2.4. Data Processing
2.4.1. Macroalgae and Sea Urchin Production
In order to statistically compare the production of macroalgae biomass at different times of the experiment, Kruskal-Wallis ANOVA analyses and Dunn’s test for multiple comparisons were carried out.) These analyses were carried out using SigmaPlot software.
To evaluate the influence of environmental factors on algal biomass production, two complementary statistical approaches were applied: Generalized Additive Models (GAMs) and Principal Component Analysis (PCA).
The GAM was used to model the relationship between biomass gain (%) as the response variable and three environmental predictors: accumulated radiation, mean photoperiod, and mean water temperature. A GAM with smoothing splines was fitted using the pyGAM library in Python, allowing for flexible, non-linear relationships between predictors and the response. Model performance was assessed using the pseudo-R2 and Akaike Information Criterion (AIC), and the smooth functions were visualized to interpret the individual effects of each predictor [
41].
To explore patterns of covariation among environmental variables and their association with biomass, a PCA was performed using the scikit-learn package. The variables were first standardized (zero mean, unit variance) before PCA computation. The first two principal components (PC1 and PC2) were retained for interpretation and visualization. A biplot was used to project the original variables, and biomass values were overlaid as a color gradient to assess whether any patterns aligned with principal environmental gradients [
42].
All GAM and PCA statistical analyses were performed in Python using JupyterLab.
Based on the nutrient input through feed and the retention of nutrients in fish biomass, a mass balance approach was used to estimate the amount of nitrogen and phosphorus excreted by fish in the RAS system [
43]. According to this concept, the amount of nutrients expelled into the water is equal to the difference between the amount consumed through feed and the amount retained during fish growth. The amount of crude protein and phosphorus in the feed, as well as the total amount delivered throughout each session, were noted.
Nutrient excretion was calculated as follows:
Nitrogen and phosphorus retentions were calculated according to the following equations:
To calculate excretion, protein and phosphorus content values (16% and 1.3%) obtained by [
44] were used.
In aquaculture studies, this method has been frequently utilized to assess the potential for nutrient recycling and to quantify nutrient loading in land-based systems. To analyze possible correlations between nutrient excretion and macroalgae production, scatter plots and Person analysis were generated through Python using JupyterLab.
To understand how sea urchins growth fed by integrated macroalgae
U. onhoi raised into the RAS system and, also how this macroalgae diet influenced the growth and quality of their gonads, the Daily Intake Rate (DIR) (%), Feed Conversion Rate (FCR), Specific Growth Rate (SGR) (% day ˉ¹), and Gonadossomatic Index (GSI) (%) data was obtained using formulas described below and based on Zhao et al. (2024), Lourenço et al. (2020), Araújo et al. (2023) and Araújo et al. (2024), respectively. In these formulas, ‘Total Daily Feed given’ correspond of total macroalga given daily in considered time (period of days) (76, 150 and 175 days). As well as the Initial Wet weight corresponds to 0 days, and the Final Wet weight corresponds to 76, 150 or 175 days.
In addiction to analyse how sea urchin growth is influenced by fed with U. onhoi producted in conditions of this study, was also obtained graphics that ilustrate how wet weigth and test diameter of sea urchin varied with the water temperature.
2.4.2. Fish Production and Ectoparasite Analysis
To analyse how the presence or absence of integrated macroalgae into an RAS system affects the growth and part of health and wellness of studied fishes (
S. aurata), the Daily Intake Rate (DIR) (%), Feed Conversion Rate (FCR), Specific Growth Rate (SGR) (% day ˉ¹) and Condition Factor (CF) (g cmˉ³) (following Fulton’s K-index) were calculated in the presence and absence of macroalgae, whose formulas based on Bonaldo et al. (2010) are described below. Considering that the number of days and the final weight concerns to period of time (days) that has passed since the data of first biometric sampling (0 days), corresponding to at 43 and 65 days.
Statistical analysis on parasite analysis was performed with a significance level of p<0.05, where the normality test (Shapiro-Wilk) was checked and significant differences between legion stage levels were analyzed by one-way ANOVA with Tukey’s post-hoc test. All statistical analyses were performed using GraphPad Software.
2.5. Use of Artificial Intelligence Assistance
To support the statistical analysis, particularly in the implementation of complex procedures such as Generalized Additive Models (GAM) and Principal Component Analysis (PCA), the JupyterLab environment was used in combination with Python libraries. During this process, generative Artificial Intelligence (AI) tools (ChatGPT, OpenAI, 2024) were consulted to assist in writing, adapting, and troubleshooting Python code. All AI-generated code was reviewed and validated by the authors before application to ensure methodological rigor and relevance to the study objectives.
4. Discussion
In this study, a multitrophic RAS was developed for the production of fish (
Sparus aurata), integrated with macroalgae (
Ulva ohnoi) cultivation for water bioremediation. The choice of a macroalgae of the genus
Ulva is due to its known ease of adaptation to IMTA systems for biomass production and bioremediation, being especially abundant in land-based aquaculture production systems in the Algarve region [
49,
50].
4.1. Macroalgae Production
The evaluation of
U. ohnoi biomass production was monitored over 180 days, covering the autumn and winter seasons. Atmospheric and quality conditions are the parameters that most limit the production of marine macroalgae. Their growth generally increases with radiation, temperature, nutrient concentration and pCO2 level [
51,
52,
53], and these factors may also have a significant impact on the biochemical composition of macroalgae, this influence being notable in species of the genus
Ulva [
54].
During the study period, the water temperature in the RAS fluctuated normally for the time of year, starting with a maximum peak of 20.7 ºC, then decreasing to 14.3 ºC in the last days of December, and then rising to 20.3 ºC in mid-March, the final period of the trial. The optimum temperature for
Ulva ohnoi is between 20 and 25ºC, with temperatures below 10ºC and above 30ºC being detrimental to growth [
55]. Solar radiation also underwent natural fluctuation. At the beginning of October, radiation was around 18 MJ. m¯², falling to around 9 MJ. m¯² in December. At the end of the test, in mid-March, the average radiation was around 13 MJ. m¯². There were daily fluctuations caused by unstable weather conditions typical of the period in question. In the southern region of Portugal, the average global solar radiation is between 16 and 22 MJ. m¯² in the spring and autumn months and between 8 and 12 MJ. m¯² [
56]. The values recorded in this work are within expected values for the region of work. Estimated daily light integrals (DLI) in Faro ranged from approximately 2 to 53 mol photons m⁻² d⁻¹ (see
Supplementary Table S1), largely exceeding the reported light saturation points (Iₖ) for
Ulva [
55]. This indicates that light availability was unlikely to limit algal growth during the study period.
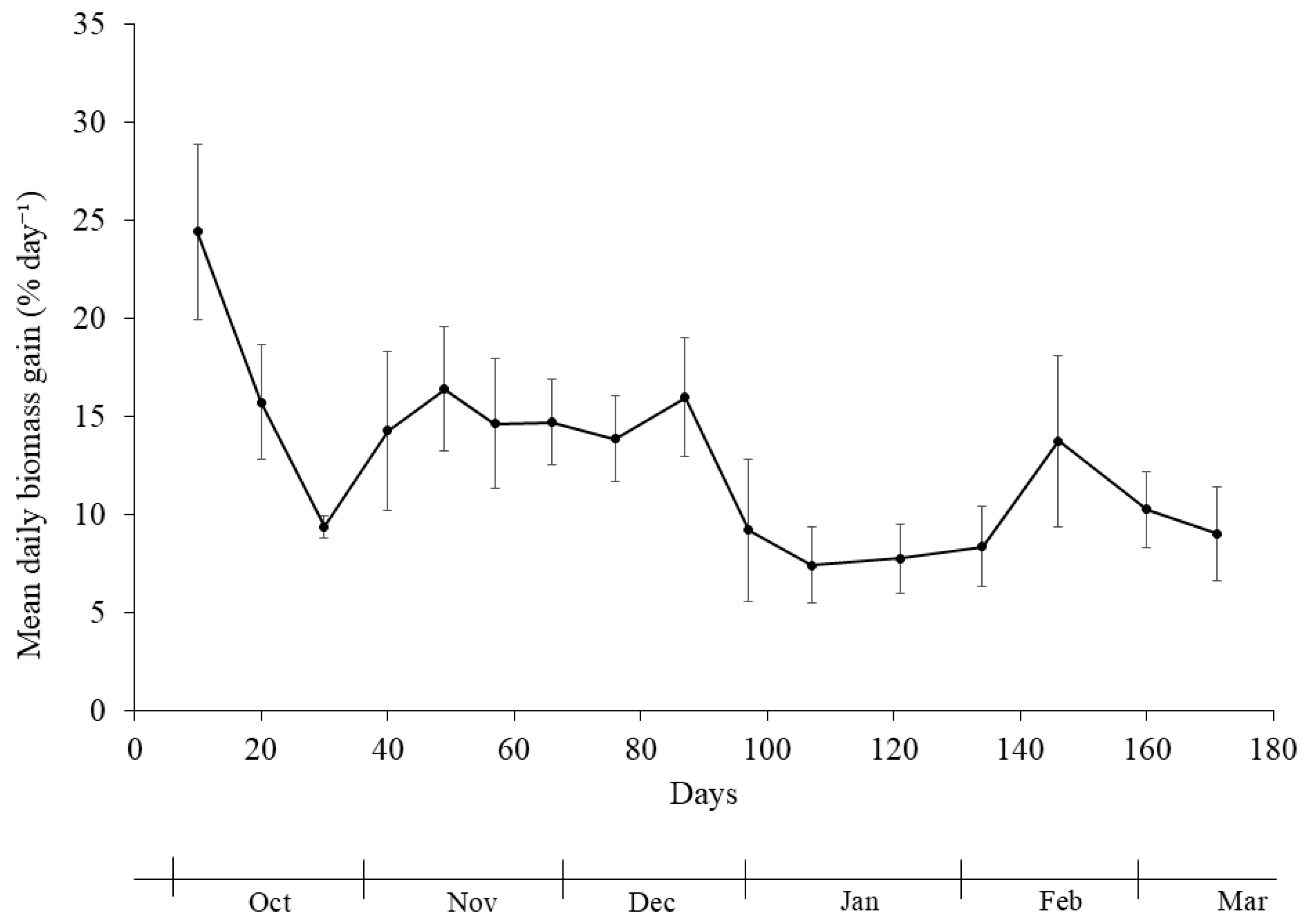
The production of
U. ohnoi exhibited moderate variation across sampling periods, with biomass yield remaining stable despite seasonal fluctuations in environmental conditions. Its daily growth varied between 7.4 and 24.4%, with the highest value recorded at the beginning of autumn and the lowest in January. In the work of Lawton et al. (2012) [
57] a daily growth rate that varied between 10 and 35% was recorded to
U.ohnoi, in cultivation with wastewater at temperatures between 17.5 and 28.8ºC. Other studies report growth rates between 7 and 14% d-1 for
Ulva lactuca in laboratory environment [
58], 15.6 and 17.8% d-1 for the same species cultivated in the ocean [
59] 12 to 16% d-1 for
Ulva rigida in outdoor tanks [
60], and 25% d-1 for
Ulva rotundata cultivated directly in the ocean [
61]. The results obtained in this work are therefore within the expected values for macroalgae of the genus
Ulva.
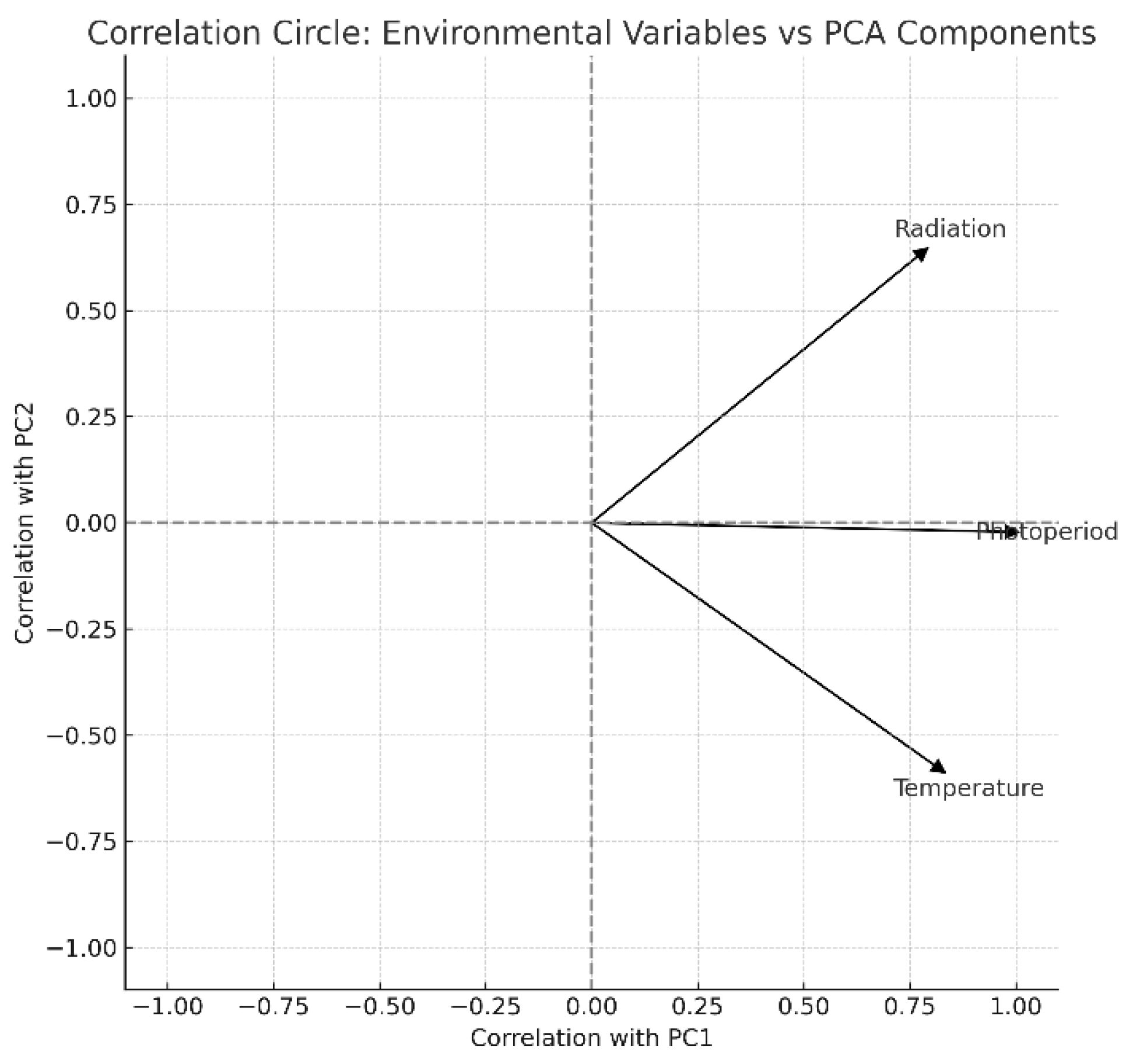
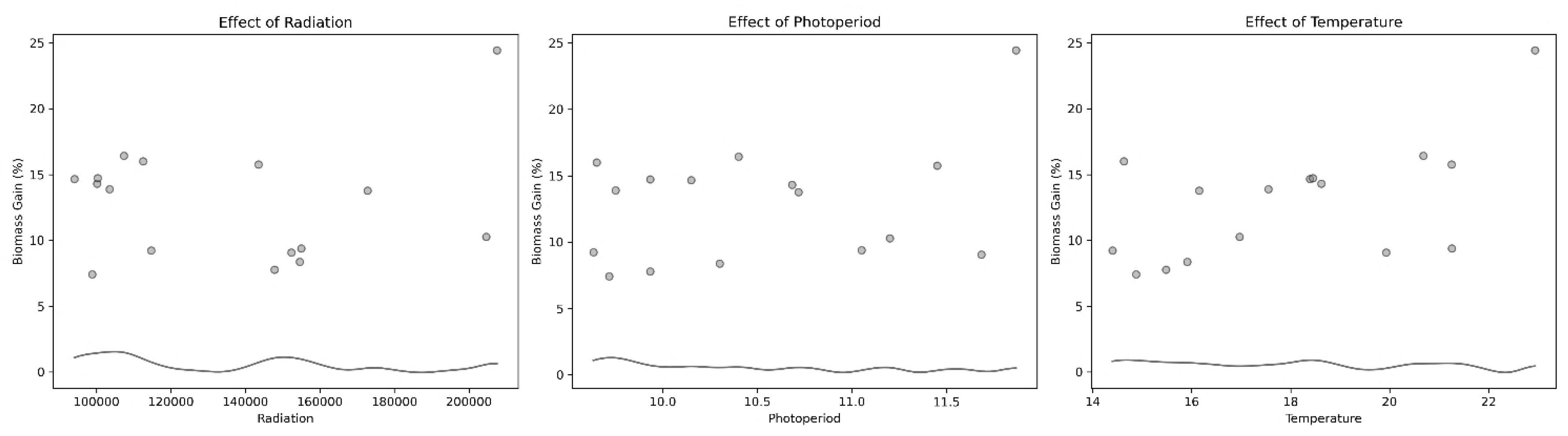
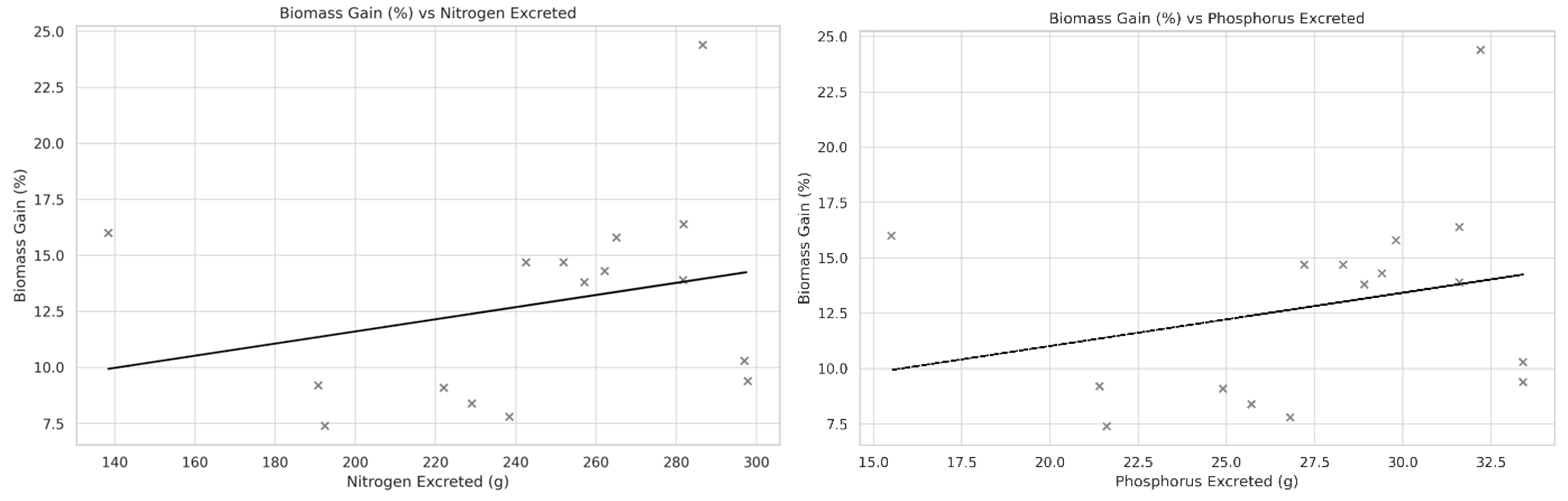
Principal component analysis (PCA) revealed a strong seasonal gradient driven primarily by photoperiod and temperature, whereas accumulated radiation contributed independently. Generalized additive model (GAM) analysis, however, detected no significant non-linear relationships between biomass yield and the tested environmental variables, indicating resilience of U. ohnoi production to environmental variation within the observed range. The nitrogen and phosphorus excretion values calculated through analysis of nutrient flows models also did not reveal any correlation with biomass production, leading us to suspect that the variation in nutrient concentration was not sufficient to have an impact on U. ohnoi production.
These findings align with previous studies demonstrating
U. ohnoi’s broad ecological plasticity and tolerance to fluctuations in temperature and irradiance [
57]. Although, in this work, it was not possible to carry out a continuous and rigorous characterization of the concentration of nutrients, the consistently high availability in recirculating aquaculture system (RAS) water likely further mitigated the effects of radiation and photoperiod variability. This high availability in recirculating aquaculture system (RAS) water as well as the high water flow rates in the macroalgae tanks allowed create a greater flow of nutrients [
62] even as to create some movement, like the aerators, on the tank for the particulates were in suspension and consequently and, possibly for this reason, there was a greater and better algae biomass [
63]. Another reason why a linear relationship was not observed between the environmental parameters analyzed and the production of macroalgae may also be due to the impact that these had on the growth of other algae on the walls of the cultivation tanks. That is, with the improvement of the cultivation conditions, namely with the increase in temperature, photoperiod and solar radiation, there was a parallel growth of other marine organisms on the walls of the tanks whose presence significantly reduced the access of the cultivated macroalgae to solar radiation, thus acting as a brake on the growth of
U. ohnoi. This fact suggests that the cultivation of macroalgae should be carried out in tanks where the water surface area is prioritized over the height of the water column. In this way, the photosynthetic capacity of the macroalgae can be optimized, without needing to increase the work effort in cleaning the walls of the tanks.
4.2. Sea Urchin Production
Feeding sea urchins with macroalgae of the genus
Ulva is a common practice, with a large collection of studies describing its potential as food for these echinoderms. Its application has been tested in its fresh form [
20,
64] or as an ingredient in inert food formulations [65 ,20,66]. This macroalgae is characterized by a nutritional profile rich in protein, minerals, fibers and active compounds, being relatively low in fats [
19]. In a previous study with the same alga [
20], protein values were determined in the order of 22.13% (dry matter) and fats in 1.63%, with the fatty acid profile characterized by a PUFA content of 26.8%, with a good contribution from 20:5 ω3 (EPA), which reached 1.6% of the total fatty acid content. Through the same study it was found that the high presence of this fatty acid in
Ulva contributed to the enrichment of this same fatty acid in the gonads of the sea urchin. It is therefore possible to state that feeding sea urchins with this type of algae favors the nutritional qualities of the gonads, for human consumption [
20,
65]
In this study, sea urchins were fed with
Ulva ohnoi produced in RAS supplied directly, fresh and drained. The sea urchins adapted perfectly to this alga, and were generally ingested within a few days, with the excretion of feces typical of feeding on this alga being observed. The daily consumption rate (DIR) was relatively stable for all replicates, ranging from 1.77 to 2.04% of their body weight per day. These values were relatively low when compared to similar studies [
63,
66,
67]. In this study, food was supplied to sea urchins periodically, with the amount supplied at that time varying according to previous consumption, and for this purpose the amount of macroalgae remaining in the tanks was observed. This strategy may not have optimized consumption for higher DIR values. Sea-urchin growth is a parameter that varies greatly depending on the life stage of sea urchins, with a significant slowdown being observed after 4 years of age [
68]. The growth rate is also obviously dependent on other environmental factors such as food availability and quality, and water temperature. The maturation of the gonads also influences the growth rate [
69,
70]. In this work, growth rates between 0.27 and 0.33% day-1 were observed, values higher than those obtained by Araújo et al. (2023) [
20], Loureiro (2021) [
69] but lower than those obtained by Candeias-Mendes et al. (2020) [
64]. Spiegel et al. (2018) [
71] analyzed the growth of
P. lividus in an IMTA system where it was fed with
Ulva lactuca, obtaining a maximum SGR of 0.34 day-1, which reduced to a minimum of 0.34 day-1. We can therefore verify that the growth of sea urchins in the present work presented values considered normal considering the diet applied. These values can be maximized using inert foods such as those tested by Loureiro (2021) [
69], which despite containing a 20%
Ulva sp. content, were enriched with other macroalgae, plant proteins and carbohydrates, as well as microalgae biomass rich in DHA. These inert foods have clear nutritional advantages, but are much more expensive, especially considering that the macroalgae biomass used in this work was produced in bioremediation tanks of a RAS system.
4.3. Sea Bream Production and Welfare
In this study, the condition of the fish (
Sparus aurata) was monitored during testing of the experimental RAS system, comparing the growth and health of fish reared with and without macroalgae integration (
U. ohnoi). Regarding growth and nutritional parameters, no significant differences were observed between the two systems and fish performance was below the expected standards for the species under optimal RAS conditions. The FCR (feed conversion ratio) values ranged from 2.13 to 2.58 and 2.50 to 2.53 for the system with and without algae, respectively. These values are considered high compared to those reported in the literature, where FCR values for
S. aurata fed under controlled conditions in RAS systems typically range between 1.42 and 1.70 in Parma et al. (2020) [
72] and around 1.89 in Zohar et al. (2005) [
73]. Likewise, the specific growth rate (SGR) varied from 0.33 ± 0.14 to 0.28 ± 0.24%·day⁻¹ in the presence of algae, and from 0.31 ± 0.15 to 0.29 ± 0.48%·day⁻¹ in the absence of algae. These values are considerably low for this type of optimized system, as reported by Parma et al. (2020) [
72], who recorded SGR values between 0.97 and 1.22%·day⁻¹, and by Petridis & Rogdakis (2008) [
74], who observed SGR values typically between 1.0 and 2.2%·day⁻¹, with values below 1.0%·day⁻¹ occurring only during colder months. Although this experimental RAS system had a low water flow rate in the fishes tanks compared to other studies, like Parma et al. (2020), the density of gilthead seabream in the tanks didn’t justify a larger water flow rate and the ammonium concentrations never reached up a critical level, remaining from a low to moderate concentration. However, these conclusions are based on calculated theoretical values and regular measurements using a colorimetric test kit. Additionally, despite the low SGR, the fishes never were in undernutrition presently an average weight for the system with and without algae, respectively, of 265.7 ± 40.8 g and 267.1 ± 40.5 g in the 43 days as well as 276.8 ± 45.1 g and 282.1 ± 51.8 g in the 65 days. For these reasons, no major problems were recorded in water quality, fish health or tank load. So the FCR and SGR values may be associated with failures in the physical process of feeding (manual). In general, it can consequently be stated that the gilthead seabream farming system under these conditions was not optimized to maximize production and, therefore this study highlights the importance of putting automatic feeders in future RAS systems for a better and controlled management of feeding.
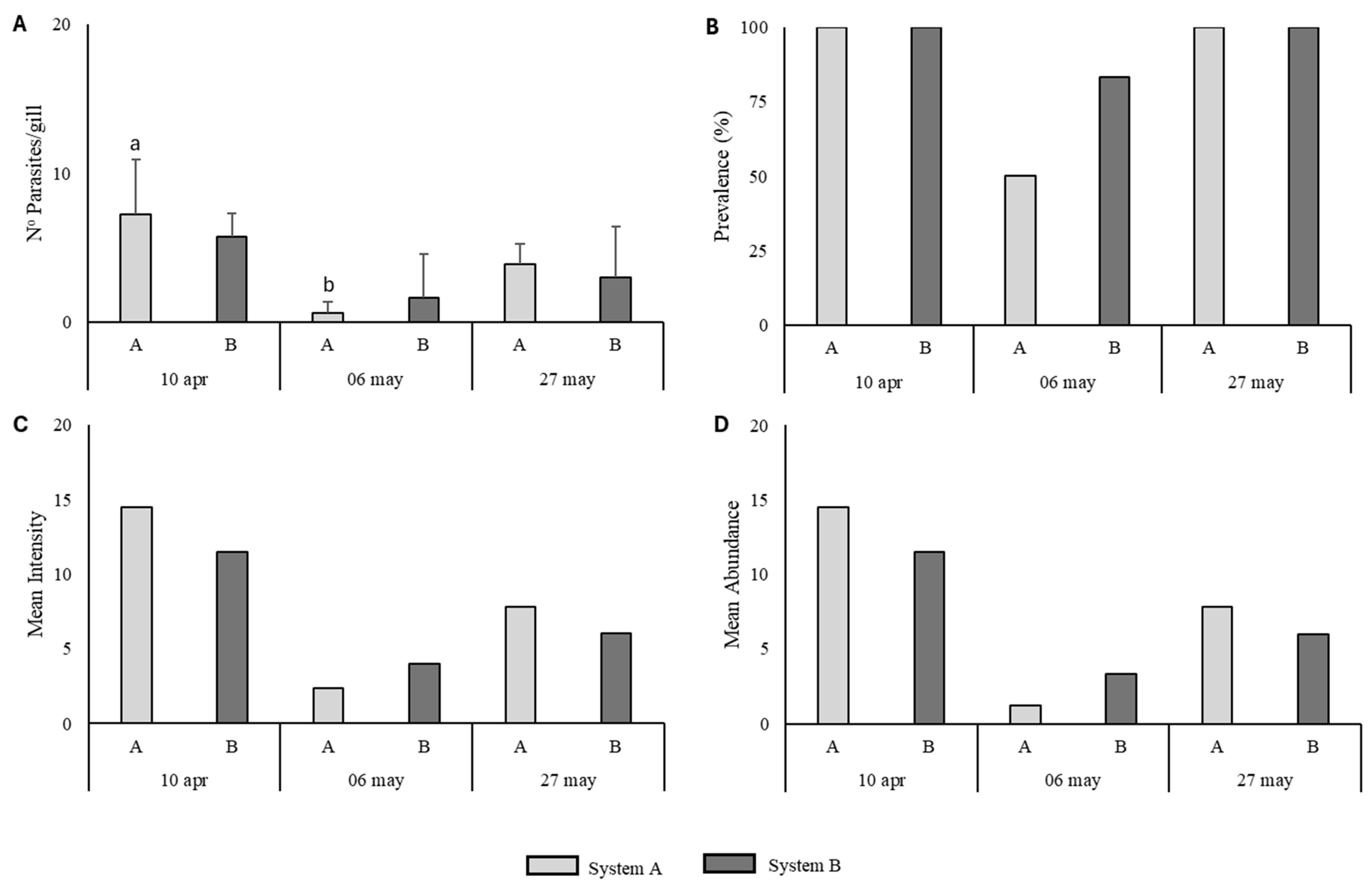
The parasitological monitoring revealed distinct patterns between the two systems over time, with system A (macroalgae) showing reduced parasite loads during the second sampling point. This coincided with the lowest prevalence (50%) and abundance values, suggesting a potential short-term mitigating effect of U. ohnoi on parasite transmission or host susceptibility. However, at the final sampling date, infection levels increased again in both systems, returning to 100% prevalence, indicating that the effect may be temporary or influenced by environmental or biological factors (e.g., parasite life cycle, temperature, or fish immune status).
Although parasite intensity and abundance were slightly higher in system A at the final sampling, no significant differences were observed, highlighting the need for longer-term studies to determine the consistency and mechanisms of macroalgae-related parasite control in multitrophic RAS systems. Hutson et al. (2012) [
28] also observed no significant differences in the number of
Neobenedenia sp. parasites attached to the gills of
Lates calcarifer after 24 hours of exposure to several macroalgal extracts, including
Ulva sp.
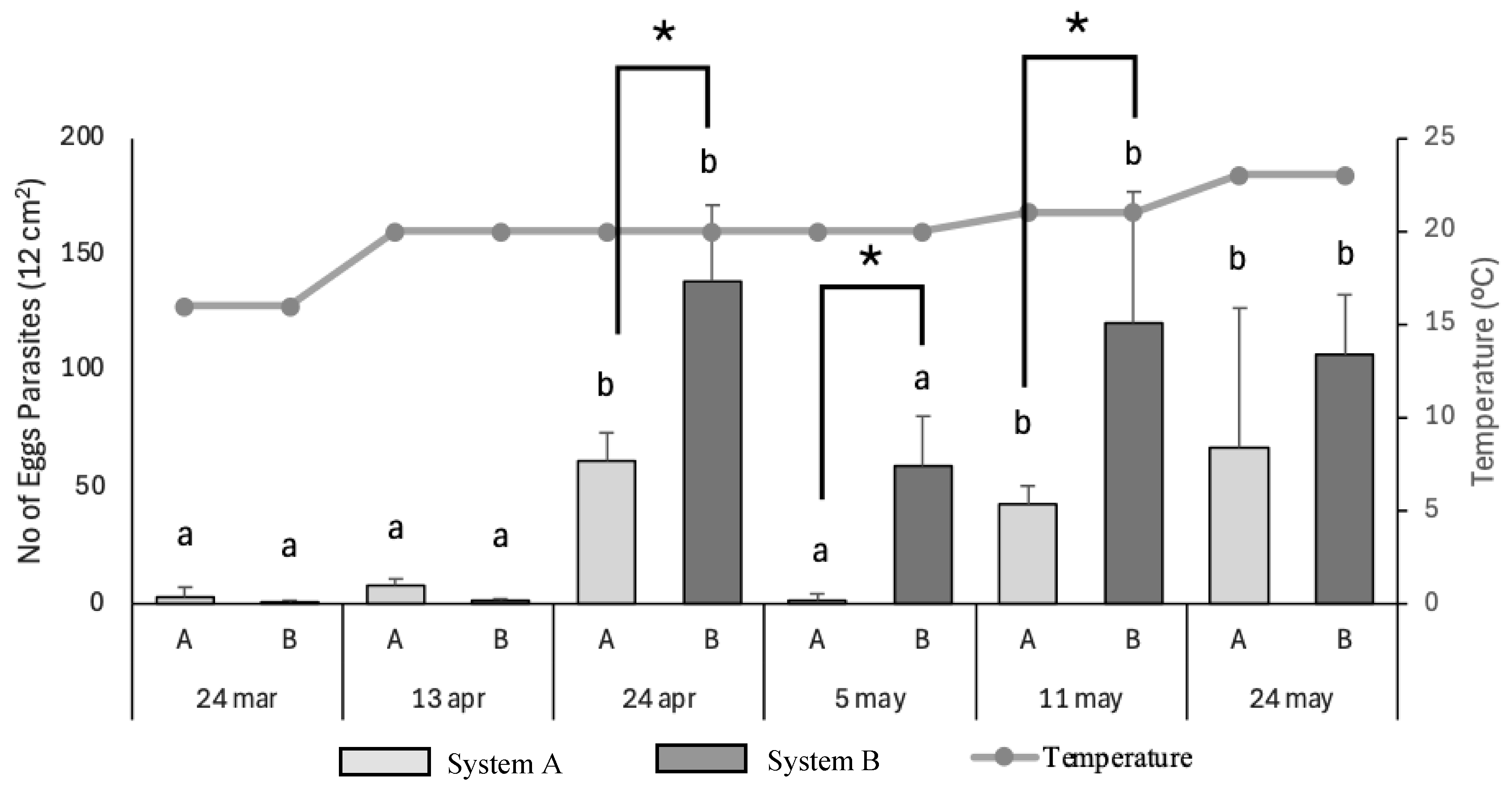
The number of monogenean eggs deposited on cotton substrate varied substantially throughout the trial period and appeared to be partially influenced by water temperature and system type.
During the initial sampling dates (24 March and 13 April), parasite egg counts remained low in both systems, coinciding with relatively lower water temperatures (16–20 °C). This suggests that early in the reproductive season, environmental conditions were not yet optimal for monogenean oviposition.
On 24 April, egg deposition increased significantly in both systems, despite stable temperatures, indicating that parasite reproduction had intensified. Notably, egg counts were significantly higher in system B, suggesting that the presence of Ulva ohnoi in system A may have exerted a limiting effect on parasite reproduction or egg retention.
By 5 May, a significant reduction in egg numbers was observed in system A, while egg counts in system B remained elevated, albeit without statistical significance. This divergence supports the hypothesis that macroalgae may contribute to interrupting the parasite’s reproductive cycle, possibly by altering water chemistry or microbial composition.
On 11 May, with a temperature increase to 21 °C, egg production rose again in both systems. However, system B consistently exhibited higher egg densities, reinforcing the pattern observed earlier. At the final sampling point (24 May, 23 °C), both systems displayed increased egg counts, although differences were no longer statistically significant. Similar findings from Hutson et al. (2012) [
28] demonstrated that extracts of
Ulva sp. and
Asparagopsis taxiformis had significant inhibitory effects on the embryonic development and hatching success of the monogenean
Neobenedenia sp. infecting
Lates calcarifer, resulting in prolonged embryonation and a reduction in overall hatching success, more pronounced in
A.
taxiformis.
Overall, the results suggest that while temperature clearly influences monogenean reproduction, the integration of macroalgae in RAS systems may help mitigate parasite egg accumulation, particularly during intermediate stages of the reproductive cycle. Further investigation is warranted to determine whether this effect is due to physical interference, chemical interactions, or shifts in microbial communities associated with the algae.
5. Conclusions
This study demonstrates the technical feasibility and ecological potential of integrating Ulva ohnoi cultivation and Paracentrotus lividus rearing into a multitrophic recirculating aquaculture system (RAS) with Sparus aurata. The macroalgae showed consistent biomass production across seasons, while effectively supporting sea urchin growth and providing indications of parasite mitigation in the fish component of the system. Although no strong environmental drivers of Ulva biomass were detected, its resilience and adaptability confirm its suitability for land-based integrated systems. However limited environmental variation observed during autumn and winter may have reduced the ability to detect clear dependencies between environmental parameters and macroalgal growth. We suggest that future experiments should include periods of greater seasonal variability—such as spring and summer—or be complemented by controlled indoor trials. These approaches would allow better isolation of the effects of temperature, light intensity, and photoperiod on Ulva ohnoi growth, and further validate the species’ ecological plasticity under a broader range of conditions.
Based on the algal growth rates observed in this study and the feeding behavior of sea urchins, it is estimated that the integrated macroalgal production could support the feeding of approximately 1,017 to 5,356 P. lividus individuals, depending on the season and corresponding algal productivity. Considering the average algal biomass produced, the system can sustainably feed around 3,000 sea urchins under the given dimensions and a daily intake rate (DIR) of 5%. In terms of final biomass at the time of sale, using this exact system, this can correspond to between 150 and 360 kg of whole sea urchins, depending on their diameter and condition. This estimation highlights the potential of this RAS-IMTA configuration not only for water bioremediation but also as a reliable biomass source for valuable echinoderm species, contributing to circularity and economic diversification in aquaculture.
Further optimization and longer-term assessments will be essential to fully understand the dynamics of species interactions and improve the productivity and resilience of such systems under commercial conditions.
Figure 2.
Mean daily macroalgae (Ulva ohnoi) biomass gain (% day⁻¹) during the first phase of the experiment.
Figure 2.
Mean daily macroalgae (Ulva ohnoi) biomass gain (% day⁻¹) during the first phase of the experiment.
Figure 3.
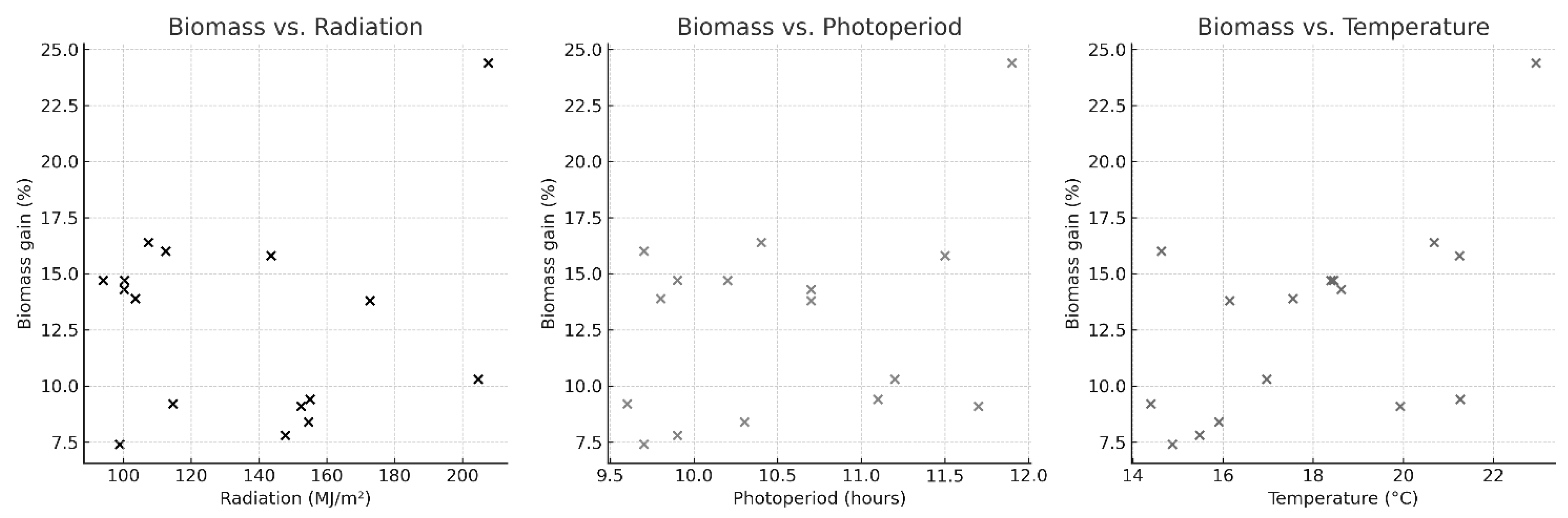
Scatter plots showing the relationship between daily Ulva ohnoi biomass gained and each environmental variable (accumulated global radiation, photoperiod and water temperature).
Figure 3.
Scatter plots showing the relationship between daily Ulva ohnoi biomass gained and each environmental variable (accumulated global radiation, photoperiod and water temperature).
Figure 4.
Principal Component Analysis (PCA) for the three environmental variables (accumulated global radiation, photoperiod and water temperature) and macroalgae biomass production (Ulva ohnoi).
Figure 4.
Principal Component Analysis (PCA) for the three environmental variables (accumulated global radiation, photoperiod and water temperature) and macroalgae biomass production (Ulva ohnoi).
Figure 5.
Relationship between Ulva ohnoi daily biomass growth (%) and selected environmental parameters (temperature, radiation, and photoperiod) assessed using Generalized Additive Models (GAM). The markers represent observed values at each sampling date, while the black curve shows the non-linear relationship estimated by the GAM. The shaded area represents the 95% confidence interval around the fitted curve.
Figure 5.
Relationship between Ulva ohnoi daily biomass growth (%) and selected environmental parameters (temperature, radiation, and photoperiod) assessed using Generalized Additive Models (GAM). The markers represent observed values at each sampling date, while the black curve shows the non-linear relationship estimated by the GAM. The shaded area represents the 95% confidence interval around the fitted curve.
Figure 6.
Correlation between nitrogen (left) and phosphorus (right) excretion and the average macroalgal (Ulva ohnoi) biomass gain in the different periods of the experiment.
Figure 6.
Correlation between nitrogen (left) and phosphorus (right) excretion and the average macroalgal (Ulva ohnoi) biomass gain in the different periods of the experiment.
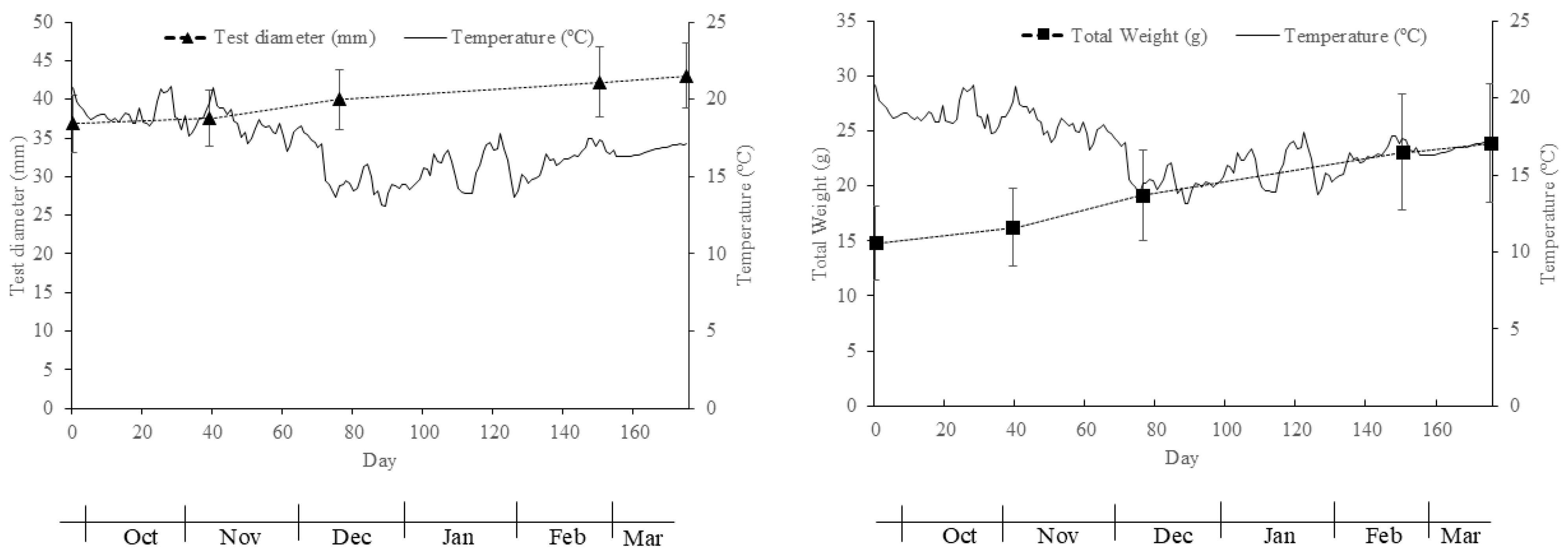
Figure 7.
Variation in water temperature (ºC), the total weight (g) and test diameter (mm) of sea urchins (Paracentrotus lividus) during the first phase of the test.
Figure 7.
Variation in water temperature (ºC), the total weight (g) and test diameter (mm) of sea urchins (Paracentrotus lividus) during the first phase of the test.
Figure 8.
Parasitological parameters of monogenean infection in gilthead seabream (Sparus aurata) reared in two different RAS systems (A: with macroalgae Ulva ohnoi; B: control) at three sampling points (10 April, 6 May, and 27 May). (A) Mean number of parasites per gill arch (± SD); (B) Prevalence of infection (% of infected fish); (C) Mean intensity (parasites per infected fish); (D) Mean abundance (parasites per total number of fish sampled). Different letters above bars indicate statistically significant differences between groups (p < 0.05).
Figure 8.
Parasitological parameters of monogenean infection in gilthead seabream (Sparus aurata) reared in two different RAS systems (A: with macroalgae Ulva ohnoi; B: control) at three sampling points (10 April, 6 May, and 27 May). (A) Mean number of parasites per gill arch (± SD); (B) Prevalence of infection (% of infected fish); (C) Mean intensity (parasites per infected fish); (D) Mean abundance (parasites per total number of fish sampled). Different letters above bars indicate statistically significant differences between groups (p < 0.05).
Table 1.
All pairwise multiple comparison procedures (Dunn’s Method) analysis of the daily biomass gain (% day⁻¹) of Ulva ohnoi for the sampling pairs. Shows only the results on pairs with significant differences (P<0.005).
Table 1.
All pairwise multiple comparison procedures (Dunn’s Method) analysis of the daily biomass gain (% day⁻¹) of Ulva ohnoi for the sampling pairs. Shows only the results on pairs with significant differences (P<0.005).
| Pair of samples |
Q value |
P value |
| 4 October / 30 December |
3.854 |
0.014 |
| 4 October / 9 January |
4.589 |
<0.001 |
| 4 October / 23 January |
4.480 |
<0.001 |
| 4 October / 05 February |
4.240 |
0.003 |
| 4 October / 14 March |
3.900 |
0.012 |
| 12 Nov / 9 January |
4.149 |
0.004 |
| 12 Nov / 23 January |
4.016 |
0.007 |
| 12 Nov / 05 February |
3.721 |
0.024 |
| 20 December / 09 January |
4.028 |
0.007 |
| 20 December / 23 January |
3.895 |
0.012 |
| 20 December / 05 February |
3.600 |
0.038 |
Table 2.
Daily Intake Rate (%), Feed Conversion Ratio (FCR), Specific Growth Rate (SGR) (% day ˉ¹) and Gonadossomatic Index (GI) (%) data of Sea Urchins Paracentrotus lividus production fed by Ulva ohnoi.
Table 2.
Daily Intake Rate (%), Feed Conversion Ratio (FCR), Specific Growth Rate (SGR) (% day ˉ¹) and Gonadossomatic Index (GI) (%) data of Sea Urchins Paracentrotus lividus production fed by Ulva ohnoi.
| Sea Urchins |
At 0 days
T (ºC) = 20,8 ± 0,99
|
At 76 days
T (ºC) = 17,7 ± 1,59
|
At 150 days
T (ºC) = 15,3 ± 1,16
|
At 175 days
T (ºC) = 16,4 ± 0,61
|
| Biomass (Kg) |
3,11 ± 0,35 |
4,01 ± 0,81 |
4,82 ± 1,05 |
4,99 ± 1,23 |
| DIR (%) |
- |
2,04 ± 0,06 |
1,83 ± 0,02 |
1,77 ± 0,02 |
| FCR |
- |
5,97 ± 0,27 |
6,36 ± 0,10 |
6,60 ± 0,06 |
| SGR (% day ˉ¹) |
- |
0,33 ± 0,23 |
0,29 ± 0,29 |
0,27 ± 0,34 |
| GSI (%) |
7,34 ± 1,72 |
- |
- |
2,88 ± 1,23 |
Table 3.
Biomass (Kg), density (Kg m3−1), Daily Intake Rate (DIR) (%), Feed Conversion Ratio (FCR), Specific Growth Rate (SGR) (% day−1) and Condition Factor (CF) (g cm−3) of gilt-head seabream (Sparus aurata) produced in the presence and absence of macroalgae (Ulva ohnoi).
Table 3.
Biomass (Kg), density (Kg m3−1), Daily Intake Rate (DIR) (%), Feed Conversion Ratio (FCR), Specific Growth Rate (SGR) (% day−1) and Condition Factor (CF) (g cm−3) of gilt-head seabream (Sparus aurata) produced in the presence and absence of macroalgae (Ulva ohnoi).
| |
|
|
At 0 days
T (ºC) = 17.2 ± 0.17
|
At 43 days
T (ºC) = 20.0 ± 1.41
|
At 65 days
T (ºC) = 21.8 ± 1.98
|
| Gilthead Seabream (S. aurata) Production |
In the presence of algae |
Number of Fishes |
220 |
213 |
207 |
| Biomass (Kg) |
50.66 ± 8.46 |
56.58 ± 8.69 |
57.30 ± 9.33 |
| Density (Kg m³ ˉ¹) |
16.89 ± 2.82 |
18.86 ± 2.90 |
19.10 ± 3.11 |
| DIR (%) |
- |
0.70 ± 0.02 |
0.71 ± 0.01 |
| FCR |
- |
2.13 ± 0.13 |
2.58 ± 0.05 |
| SGR (% day ˉ¹) |
- |
0.33 ± 0.14 |
0.28 ± 0.24 |
| |
CF (g cmˉ³) |
1.66 ± 0.13 |
1.72 ± 0.25 |
1.65 ± 0.10 |
| In the absence of algae |
Number of Fishes |
220 |
214 |
208 |
| Biomass (Kg) |
51.47 ± 8.36 |
57.16 ± 8.67 |
58.68 ± 10.78 |
| Density (Kg m³ ˉ¹) |
17.16 ± 2.79 |
19.05 ± 2.89 |
19.56 ± 3.59 |
| DIR (%) |
- |
0.71 ± 0.02 |
0.71 ± 0.01 |
| FCR |
- |
2.50 ± 0.12 |
2.53 ± 0.03 |
| SGR (% day ˉ¹) |
- |
0.31± 0.15 |
0.29 ± 0.48 |
| |
|
CF (g cmˉ³) |
1.64 ± 0.14 |
1.78 ± 0.21 |
1.64 ± 0.10 |