
Submitted:
18 August 2025
Posted:
20 August 2025
You are already at the latest version
Abstract
Background: Forkhead box (FOX) transcription factors constitute a large family of regulatory proteins that control diverse cellular processes, including development, metabolism, immunity, and aging. Metadichol, a nano lipid formulation derived from long-chain alcohols, has demonstrated pleiotropic biological effects, including immunomodulation and metabolic regulation. Objective: To comprehensively evaluate the effects of metadichol treatment on FOX transcription factor gene expression in human peripheral blood mononuclear cells (PBMCs) via quantitative PCR analysis. Methods: Human PBMCs were isolated via Histopaque density gradient centrifugation and treated with Metadichol atconcentrations of 1 pg/ml, 100 pg/ml, 1 ng/ml, and 100 ng/ml. Total RNA was extracted, reverse-transcribed, andanalyzed by quantitative PCR for 45 FOX genes. Gene expression changes were calculated via normalization to GAPDH via the 2^-ΔΔCq method. Results: Metadichol treatment resulted in dose-dependent modulation of FOX gene expression. At the highest concentration (100 ng/ml), significant upregulation of multiple FOX genes was observed, with FOXO1 showing thegreatest increase (8.74-fold), followed by FOXA1 (7.39-fold) and FOXH1 (7.22-fold). Additional substantial increases were noted for FOXA2 (6.57-fold), FOXA3 (6.98-fold), FOXB1 (6.79-fold), FOXP3 (5.46-fold), andFOXP4 (6.23-fold). Conversely, selective downregulation was observed for FOXL2 (0.16-fold), FOXL1 (0.54-fold), and FOXD4L1 (0.56-fold). Conclusions: Metadichol has potent and selective effects on FOX transcription factor expression in human PBMCs, with preferential upregulation of genes involved in metabolic regulation, immune homeostasis, and cellular longevity pathways. These findings suggest potential therapeutic applications in age-related diseases, metabolic disorders, andimmunomodulation. The differential expression patterns indicate complex regulatory mechanisms that warrant furtherinvestigation to elucidate their clinical translation potential.
Keywords:
Fox family
; Metadichol
; immune metabolic rejuvenation
; nuclear receptors
; SOX family
; Toll-like receptors
; KLFs
; sirtuins
; circadian genes
; GDF11
; TERT
; Klotho
; induced pluripotency
; regenerative medicine
Introduction
The Forkhead box (FOX) transcription factor superfamily comprises more than 50 members [1] organized into 19 subfamilies (FOXA-FOXS) [2,3] that regulate diverse cellular processes, including development [4], metabolism [5], immunity [6], and aging [7,8]. These evolutionarily conserved proteins are characterized by a shared forkhead DNA-binding domain [9] and exhibit tissue-specific expression patterns with distinct functional roles [10,11]. The clinical importance of FOX transcription factors has been extensively documented in the fields of cancer biology [12], metabolic diseases [13], autoimmune disorders [14], and aging-related pathologies [15,16].

Table 1.
Comprehensive FOX Gene Function and Disease Association Table.
| FOX Gene | Biological Process | Disease Association | Key Functions | References |
| FOXA1 | Organogenesis, hepatic development, pioneer transcription factor activity | Prostate cancer, breast cancer, metabolic disorders | Liver specification, pancreatic development, nuclear receptor cofactor | [17,18] |
| FOXA2 | Endodermal organ development, glucose homeostasis | Type 2 diabetes, pancreatic disorders, lung development defects | Pancreatic β-cell function, gluconeogenesis regulation, respiratory development | [19,20] |
| FOXA3 | Hepatic gene expression, metabolism | Cholangiocarcinoma, liver cancer, metabolic syndrome | Liver development, metabolic gene regulation, bile acid synthesis | [21] |
| FOXB1 | Neural development, cell proliferation | Glioblastoma, neural tube defects | Brain development, neural differentiation | [22] |
| FOXD1 | Kidney development, epithelial–mesenchymal transition | Pancreatic cancer, renal disorders | Kidney morphogenesis, EMT regulation, cancer metastasis | [23,24] |
| FOXD2 | Neural crest development | Developmental disorders | Neural crest cell migration, cranial development | [25,26,27] |
| FOXD3 | Neural crest development, stem cell maintenance | Melanoma, developmental disorders | Neural crest specification, stem cell pluripotency | [28,29] |
| FOXD4 | Embryonic development | Unknown pathological significance | Early development, recently duplicated in humans | [30] |
| FOXE1 | Thyroid development, neural development | Thyroid cancer, congenital hypothyroidism | Thyroid morphogenesis, neural tube closure | [31,32,33] |
| FOXF1 | Mesenchymal development, lung development | Alveolar capillary dysplasia, lung disorders | Lung development, angiogenesis | [34,35] |
| FOXF2 | Kidney development, angiogenesis | Renal disorders, vascular malformations | Kidney morphogenesis, vascular development | [36,37] |
| FOXG1 | Brain development, telencephalon formation | Autism spectrum disorders, Rett syndrome-like phenotype | Forebrain development, neurogenesis | [38,39] |
| FOXH1 | Mesoderm formation, nodal signaling | Developmental disorders, cardiac defects | Gastrulation, heart development, TGF-β signaling | [40] |
| FOXJ1 | Ciliogenesis, respiratory epithelium | Primary ciliary dyskinesia, respiratory infections | Cilia formation, respiratory function | [41] |
| FOXJ2 | Cell cycle regulation | Cancer | G2/M transition, DNA damage response | [41] |
| FOXJ3 | Cell cycle progression | Cancer progression | Mitotic regulation, chromosome segregation | [41] |
| FOXK1 | Muscle development, cell cycle | Muscular disorders, cancer | Myogenesis, proliferation control | [42] |
| FOXK2 | Muscle differentiation, metabolism | Metabolic disorders, muscle diseases | Skeletal muscle development, glucose metabolism | [43] |
| FOXL1 | Gastrointestinal development | Gastrointestinal cancers | Intestinal development, GI tract homeostasis | [44] |
| FOXL2 | Ovarian development, granulosa cell function | Ovarian cancer, premature ovarian failure | Ovarian follicle development, sex determination | [45] |
| FOXM1 | Cell cycle progression, DNA repair, mitosis | Multiple cancers, aging-related diseases | G1/S transition, M-phase progression, genomic stability | [46] |
| FOXN1 | Thymic development, hair follicle formation | Severe combined immunodeficiency, alopecia | T-cell development, skin differentiation | [47] |
| FOXN2 | Neural development | Neurodevelopmental disorders | Brain development, neuronal differentiation | [48] |
| FOXN3 | Cell cycle regulation, DNA damage response | Cancer, aging | Cell cycle checkpoints, DNA repair | [49] |
| FOXN4 | Retinal development | Retinal disorders, blindness | Retinal neurogenesis, photoreceptor development | [50] |
| FOXO1 | Glucose homeostasis, stress response, apoptosis | Type 2 diabetes, cancer, metabolic syndrome | Gluconeogenesis, insulin sensitivity, cellular stress response | [51] |
| FOXO3 | Aging, stress resistance, apoptosis | Cancer, neurodegenerative diseases, longevity | Oxidative stress response, longevity pathways, apoptosis | [52] |
| FOXO4 | Cell cycle arrest, DNA damage response | Cancer, premature aging | p21 induction, senescence, DNA repair | [53] |
| FOXO6 | Brain function, glucose metabolism | Alzheimer's disease, diabetes | Memory consolidation, hepatic gluconeogenesis | [54] |
| FOXP1 | B-cell development, cardiac morphogenesis | Diffuse large B-cell lymphoma, intellectual disability | B-cell differentiation, heart valve development | [55] |
| FOXP2 | Language development, neural function | Speech and language disorders, autism | Speech acquisition, motor learning, synaptic plasticity | [56] |
| FOXP3 | Regulatory T-cell function, immune tolerance | Autoimmune diseases, IPEX syndrome, cancer | Treg development, immune suppression, self-tolerance | [57] |
| FOXP4 | T-cell development, cardiac function | Developmental disorders, cardiac defects | T-cell differentiation, heart development | [58] |
| FOXQ1 | Epithelial development | Colorectal cancer, gastric cancer | Epithelial homeostasis, EMT regulation | [59] |
| FOXR1 | Neural development | Cancer | Brain development, cell proliferation | [60,61] |
| FOXR2 | Neural function | Cancer | Neural development, transcriptional regulation | [62,63] |
| FOXS1 | Neural crest development | Developmental disorders | Cranial neural crest formation | [64] |
Among the most studied FOX subfamilies, FOXA proteins function as pioneer transcription factors that facilitate chromatin remodeling and gene accessibility. [65,66] FOXA1, FOXA2, and FOXA3 are critical regulators of hepatic metabolism [67], pancreatic β-cell function [68], and lipid homeostasis.69 The FOXO subfamily, comprising FOXO1, FOXO3, FOXO4, and FOXO6, serves as key mediators of cellular stress responses [70] longevity pathways [71], and metabolic homeostasis [72] FOXP proteins, particularly FOXP3, are essential for regulatory T-cell development and immune tolerance. [73,74]
Metadichol is a novel nanolipid formulation consisting of long-chain alcohols derived from sugarcane [75]. Previous investigations have demonstrated that metadichol functions as a vitamin D receptor (VDR) agonist, modulates immune responses [76] and exhibits antiviral properties [77,78,79]. This compound has been shown to increase endogenous vitamin C levels. [80]
Given the central role of FOX transcription factors in cellular homeostasis and the emerging therapeutic potential of metadichol, we hypothesized that metadichol treatment would significantly modulate FOX gene expression in human immune cells. This study presents the first comprehensive analysis of the effects of metadichol on the entire FOX transcription factor family using human peripheral blood mononuclear cells (PBMCs) as a physiologically relevant model system.
Experimental
A commercial service provider (Skanda Life Sciences, Bangalore, India) performed the quantitative q-RT‒PCR, Western blot analysis, and cell culture work. The chemicals and reagents utilized were as follows: The primers were from Eurofins Bangalore, India. Other molecular biology reagents were obtained from Sigma‒Aldrich, India.
Materials and Methods
Cell Isolation and Culture
Fresh human blood was collected in EDTA-containing tubes following institutional review board approval and informed consent procedures. PBMCs were isolated via Histopaque-1077 density gradient centrifugation. [83] Briefly, blood was diluted 1:1 with phosphate-buffered saline (PBS) and carefully layered over Histopaque-1077. Following centrifugation at 400×g for 30 minutes at room temperature, the mononuclear cell layer was collected, washed twice with PBS, and resuspended in RPMI-1640 medium supplemented with 10% fetal bovine serum. [84]
Maternal Treatment
Isolated PBMCs were treated with Metadichol at concentrations of 1 pg/ml, 100 pg/ml, 1 ng/ml, and 100 ng/ml, with untreated cells serving as controls. The treatment duration was optimized on the basis of preliminary time-course experiments. The cells were maintained at 37°C in a humidified atmosphere containing 5% CO₂.
RNA Extraction and cDNA Synthesis
Total RNA (Table 2) was extracted via TRIzol reagent according to the manufacturer's protocol. [85] RNA quality and quantity were assessed via spectrophotometric analysis (Spectramax i3x, Molecular Devices). cDNA synthesis was performed with 500 ng of total RNA via the PrimeScript RT Reagent Kit (Takara) via oligo-dT primers. Reverse transcription was conducted at 50°C for 30 minutes, followed by enzyme inactivation at 85°C for 5 minutes. [86]
Table 2.
RNA Yields.
| Treatment Concentration | RNA Yield (ng/μL) |
| Control (0) | 328.0 |
| 1 pg/mL | 415.0 |
| 100 pg/mL | 353.3 |
| 1 ng/mL | 336.0 |
| 100 ng/mL | 353.2 |
Table 3.
List of FOX genes primers used.
| Gene | Primers | Amplicon Size | Annealing temperature | |
| FOXA1 | F | GCAATACTCGCCTTACGGCTCT | 129 | 65 |
| R | GGGTCTGGAATACACACCTTGG | |||
| FOXA2 | F | GGAACACCACTACGCCTTCAAC | 133 | 65 |
| R | AGTGCATCACCTGTTCGTAGGC | |||
| FOXA3 | F | CTCGCTGTCTTTCAACGACTGC | 122 | 65 |
| R | CGCAGGTAGCAGCCATTCTCAA | |||
| FOXB1 | F | CCACAACCTCTCCTTCAACGAC | 122 | 59 |
| R | AGGAAGCTGCCGTTCTCGAACA | |||
| FOXD1 | F | TGGTTCGGTGTTTTGTTCGC | 154 | 65 |
| R | AGCATAGGTCGGCTTTGCAT | |||
| FOXD2 | F | AACAGCATCCGCCACAACCTCT | 92 | 65 |
| R | CAGCGTCCAGTAGTTGCCCTTG | |||
| FOXD4 | F | CCACTAGCGTTCCTGCTTCT | 217 | 65 |
| R | TCATCTTCCTCCTCTCCCAGG | |||
| FOSD4L1 | F | TACATTTCAGCCTCCTGCCC | 204 | 53 |
| R | ACCTGCCACCAAGGAAGATG | |||
| FOXE1 | F | CTCTGCTCTGGTTGACCTGG | 103 | 65 |
| R | GGTTCAGGTGATGGGACTGG | |||
| FOXF1 | F | CAGGGCTGGAAGAACTCCG | 222 | 65 |
| R | GAAGCCGAGCCCGTTCAT | |||
| FOXF2 | F | CCTACCAGGGCTGGAAGAAC | 212 | 67 |
| R | CACGCGGTGGTACATGGG | |||
| FOXG1 | F | GAGGTGCAATGTGGGGAGAA | 197 | 65 |
| R | GTTCTCAAGGTCTGCGTCCA | |||
| FOXH1 | F | CCTGCCTTCTACACTGCCC | 151 | 62 |
| R | CTTCCTCCTCTTAGGGGGCT | |||
| FOXJ3 | F | TGATAGCCCACGCAGTAGCCTT | 154 | 67 |
| R | ACTGTGGTTGCTGCTGAGGAGT | |||
| FOXL1 | F | TCACGCTCAACGGCATCTACCA | 116 | 67 |
| R | TGACGAAGCAGTCGTTGAGCGA | |||
| FOXL2 | F | CAGTCAAGGAGCCAGAAGGG | 241 | 67 |
| R | CGGATGCTATTTTGCCAGCC | |||
| FOXO1 | F | GCCACATTCAACAGGCAGC | 251 | 65 |
| R | GACGGAAACTGGGAGGAAGG | |||
| FOXO4 | F | CCCGACCAGAGATCGCTAAC | 236 | 67 |
| R | AATGGCCTGGCTGATGAGTT | |||
| FOXP1 | F | CAAGCCATGATGACCCACCT | 252 | 67 |
| R | GGGCACGTTGTATTTGTCTGA | |||
| FOXB2 | F | CGACTGCTTCATCAAGATTCCGC | 104 | 59 |
| R | AGGAAGCTGCCGTTCTCGAACA | |||
| FOXC1 | F | CAGTCTCTGTACCGCACGTC | 189 | 65 |
| R | TGTTCGCTGGTGTGGTGAAT | |||
| FOXC2 | F | GCAGTTACTGGACCCTGGAC | 211 | 65 |
| R | ATCACCACCTTCTTCTCGGC | |||
| FOXD3 | F | AAGCCGCCTTACTCGTACATCG | 159 | 65 |
| R | AGAGGTTGTGGCGGATGCTGTT | |||
| FOXE3 | F | CTTCATCACCGAACGCTTTGCC | 144 | 65 |
| R | CAGCGTCCAGTAGTTGCCCTTG | |||
| FOXI1 | F | GGAGCCTCAGGACATCTTGG | 135 | 47 |
| R | CCGCTCACATAGGCTGTCAT | |||
| FOXI2 | F | CGTGGCTGGTAACTTCCCTT | 211 | 65 |
| R | GGCTTCAGCTCTCCTCTTCC | |||
| FOXI3 | F | AACTCCATCCGCCACAACCTGT | 107 | 62 |
| R | CTCGCAGTTCGGATCAAGAGTC | |||
| FOXJ1 | F | ACTCGTATGCCACGCTCATCTG | 152 | 50 |
| R | GAGACAGGTTGTGGCGGATTGA | |||
| FOXJ2 | F | ACCAGTGGCAAACAGGAGTCAG | 131 | 67 |
| R | TGGGCGATTGTATCCTGCTGAG | |||
| FOXK1 | F | GCCGACAAAGGCTGGCAGAATT | 129 | 65 |
| R | TGGCTTCAGAGGCAGGGTCTAT | |||
| FOXK2 | F | CCAAACTCGCTGTCATCCAGGA | 126 | 59 |
| R | GTGTAGGTGACAGGCTTGATGG | |||
| FOXM1 | F | AGCAGCGACAGGTTAAGGTT | 225 | 62 |
| R | TGTGGCGGATGGAGTTCTTC | |||
| FOXN1 | F | GAGGTCAAAGTCAAGCCCCC | 301 | 65 |
| R | TGTAGATCTCGCTGACGGGA | |||
| FOXN2 | F | ACAGATGCAGAGGGCTGACT | 248 | 65 |
| R | GGCAGCATCAACAGCTTCAG | |||
| FOXN3 | F | GCCCTTCTCCAAGTTCCTCC | 136 | 59 |
| R | AGCTGGTGATGCCATTCCTC | |||
| FOXN4 | F | GGCCACAGAGACAGCATGAG | 236 | 47 |
| R | TTGGGGTAGTGTTTGGGGTG | |||
| FOXO3 | F | CGTCTTCAGGTCCTCCTGTT | 135 | 47 |
| R | GGGAAGCACCAAAGAAGAGAG | |||
| FOXO6 | F | GAAGAACTCCATCCGGCACA | 124 | 65 |
| R | CGGGGTCTTCCCTGTCTTTC | |||
| FOXP2 | F | CAAGCCATGATGACCCACCT | 276 | 62 |
| R | CTGCGCAATATCTGCTGACG | |||
| FOXP3 | F | CCCACTTACAGGCACTCCTC | 254 | 65 |
| R | GGGATTTGGGAAGGTGCAGA | |||
| FOXP4 | F | GCCAAGCAGCCCACAAAG | 277 | 62 |
| R | AGATGGAGCCGACCTGATTG | |||
| FOXQ1 | F | AACCCCTCCTGGGCTCTTTA | 199 | 65 |
| R | GTGTTGGGTGGACTATGGGG | |||
| FOXR1 | F | CAGTCCTCCAGCAAGCGGTCT | 113 | 50 |
| R | AGCCATAGAGGAGCTGTCTTCC | |||
| FOXR2 | F | AAAGTCGCACGAGGAGAGTG | 209 | 67 |
| R | CTCGAGGTTCTCCATGGCTC | |||
| FOXS1 | F | ATCCGCCACAACCTGTCACTCA | 129 | 65 |
| R | GTAGGAAGCTGCCGTGCTCAAA | |||
| GAPDH | F | GTCTCCTCTGACTTCAACAGCG | 186 | 60 |
| R | ACCACCCTGTTGCTGTAGCCAA |
Quantitative PCR Analysis
Real-time PCR was performed via SYBR Green Master Mix with gene-specific primers for 45 FOX genes. The PCR conditions consisted of initial denaturation at 95°C for 2 minutes, followed by 39 cycles of 95°C for 5 seconds and primer-specific annealing/extension for 30 seconds. Melting curve analysis was performed to verify amplification specificity [39]. GAPDH served as the reference gene for normalization. Relative gene expression was calculated via the 2^-ΔΔCq method. [87,88].
Results
Metadichol Induces Dose-Dependent Changes in FOX Gene Expression
Table 4.
Treatment of PBMCs with different concentrations of metadichol resulted in significant changes in the expression of multiple FOX genes (Table 4 and Figure 1).
| Cell line | Markers | Control | 1 pg/ml | 100 pg/ml | 1 ng/ml | 100 ng/ml |
| PBMC | FOXA1 | 1 | 3.56 | 0.49 | 0.16 | 7.39 |
| FOXA2 | 1 | 1.1 | 2.25 | 0.16 | 6.57 | |
| FOXA3 | 1 | 1.24 | 1.81 | 0.12 | 6.98 | |
| FOXB1 | 1 | 3.16 | 1.01 | 0.3 | 6.79 | |
| FOXD1 | 1 | 0.31 | 4.34 | 0.83 | 1.1 | |
| FOXD2 | 1 | 6.36 | 1.83 | 0.15 | 1.38 | |
| FOXD4 | 1 | 4.67 | 0.46 | 0.11 | 1.36 | |
| FOXD4L1 | 1 | 1.93 | 1.36 | 0.2 | 0.56 | |
| FOXE1 | 1 | 1.64 | 1.66 | 0.15 | 1.43 | |
| FOXF1 | 1 | 1.01 | 0.59 | 0.2 | 2.41 | |
| FOXF2 | 1 | 0.25 | 0.89 | 0.28 | 1.47 | |
| FOXG1 | 1 | 6.26 | 3.23 | 0.45 | 3.04 | |
| FOXH1 | 1 | 3.09 | 1.49 | 0.55 | 7.22 | |
| FOXJ3 | 1 | 4.02 | 1.09 | 0.4 | 4.24 | |
| FOXL1 | 1 | 0.22 | 0.24 | 0.11 | 0.54 | |
| FOXL2 | 1 | 0.12 | 0.21 | 0.09 | 0.16 | |
| FOXO1 | 1 | 2.51 | 0.96 | 0.27 | 8.74 | |
| FOXO4 | 1 | 0.56 | 1.11 | 0.49 | 0.84 | |
| FOXP1 | 1 | 1.8 | 3.16 | 0.84 | 2.28 | |
| FOXB2 | 1 | 0.2 | 1.41 | 0.22 | 2.58 | |
| FOXC1 | 1 | 0.13 | 0.98 | 0.15 | 2.16 | |
| FOXC2 | 1 | 0.08 | 1.33 | 0.09 | 2.4 | |
| FOXD3 | 1 | 0.12 | 2.63 | 0.11 | 1 | |
| FOXE3 | 1 | 0.3 | 1.75 | 0.6 | 1.49 | |
| FOXI1 | 1 | 0.19 | 5.73 | 0.22 | 1.91 | |
| FOXI2 | 1 | 0.36 | 2.41 | 0.2 | 4.15 | |
| FOXI3 | 1 | 0.17 | 1.16 | 0.22 | 2.74 | |
| FOXJ1 | 1 | 0.15 | 0.39 | 0.14 | 0.7 | |
| FOXJ2 | 1 | 0.77 | 1.57 | 0.54 | 0.98 | |
| FOXK1 | 1 | 0.43 | 2.89 | 0.31 | 1.07 | |
| FOXK2 | 1 | 0.25 | 1.47 | 0.28 | 1.84 | |
| FOXM1 | 1 | 0.22 | 0.84 | 0.05 | 4.54 | |
| FOXN1 | 1 | 0.56 | 5.95 | 0.73 | 0.79 | |
| FOXN2 | 1 | 0.15 | 1.69 | 0.17 | 2.77 | |
| FOXN3 | 1 | 0.16 | 1.13 | 0.23 | 1.72 | |
| FOXN4 | 1 | 0.1 | 3.3 | 0.1 | 1.2 | |
| FOXO3 | 1 | 0.38 | 1.58 | 0.3 | 1.23 | |
| FOXO6 | 1 | 0.12 | 1.53 | 0.15 | 0.56 | |
| FOXP2 | 1 | 0.41 | 1.2 | 0.2 | 5.15 | |
| FOXP3 | 1 | 1.48 | 1.38 | 0.21 | 5.46 | |
| FOXP4 | 1 | 0.95 | 2.85 | 1.39 | 6.23 | |
| FOXQ1 | 1 | 0.14 | 1.25 | 0.09 | 2.18 | |
| FOXR1 | 1 | 0.13 | 1.45 | 0.16 | 3.1 | |
| FOXR2 | 1 | 0.23 | 1.38 | 0.27 | 2.11 | |
| FOXS1 | 1 | 0.12 | 0.65 | 0.1 | 3.36 |
The overall pattern revealed that the highest concentration (100 ng/ml) generally elicited the strongest response for most genes, with some exceptions showing peak responses at lower concentrations. Statistical analysis revealed that 38 out of 44 FOX genes exhibited significant expression changes in at least one treatment concentration compared with the control (p < 0.05).
Identification of the most highly responsive FOX genes
At the highest metadichol concentration (100 ng/ml), several FOX genes were strongly upregulated (Figure 2). The five genes with the greatest increase in expression were FOXO1 (8.74-fold), FOXH1 (7.22-fold), FOXA3 (6.98-fold), FOXB1 (6.79-fold), and FOXA2
(6.57-fold). This robust induction suggests that these genes may be particularly sensitive to metadichol treatment and could play important roles in mediating their biological effects.
Distinct Dose‒Response Patterns Reveal Gene-Specific Regulation
Analysis of the dose‒response relationships revealed three distinct patterns of gene expression changes in response to metadichol treatment:
- High-concentration responders: Genes whose expression was primarily upregulated at 100 ng/ml, with minimal responses at lower concentrations (e.g., FOXO1, FOXH1, FOXA3)
- Biphasic/hormetic responders: Genes whose expression was elevated at both low (1 pg/ml) and high (100 ng/ml) concentrations but whose expression was reduced at intermediate concentrations (Figure 3). Key examples include FOXD2, FOXG1, FOXD4, and FOXP3. This U-shaped response suggests complex, concentration-dependent regulatory mechanisms.
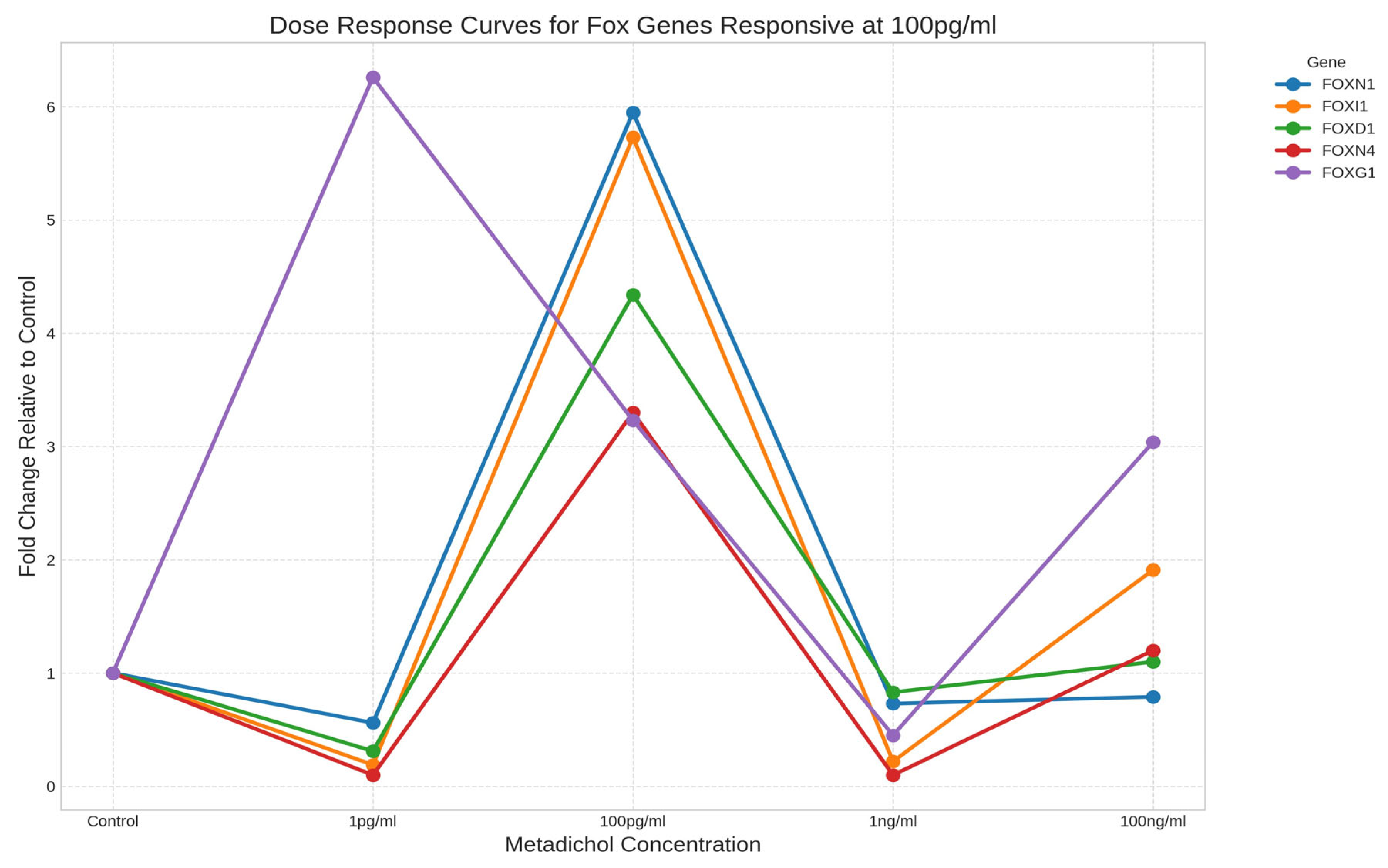
- Intermediate-concentration responders: Genes exhibiting peak expression at the intermediate concentration of 100 pg/ml, including FOXN1 (5.95-fold), FOXI1 (5.73-fold), FOXD1 (4.34-fold), and FOXN4 (3.30-fold).
Figure 3.
Dose‒response curves showing biphasic expression patterns of selected FOX genes (FOXD2, FOXG1, FOXD4, and FOXP3) in response to Metadichol treatment.
Figure 3.
Dose‒response curves showing biphasic expression patterns of selected FOX genes (FOXD2, FOXG1, FOXD4, and FOXP3) in response to Metadichol treatment.
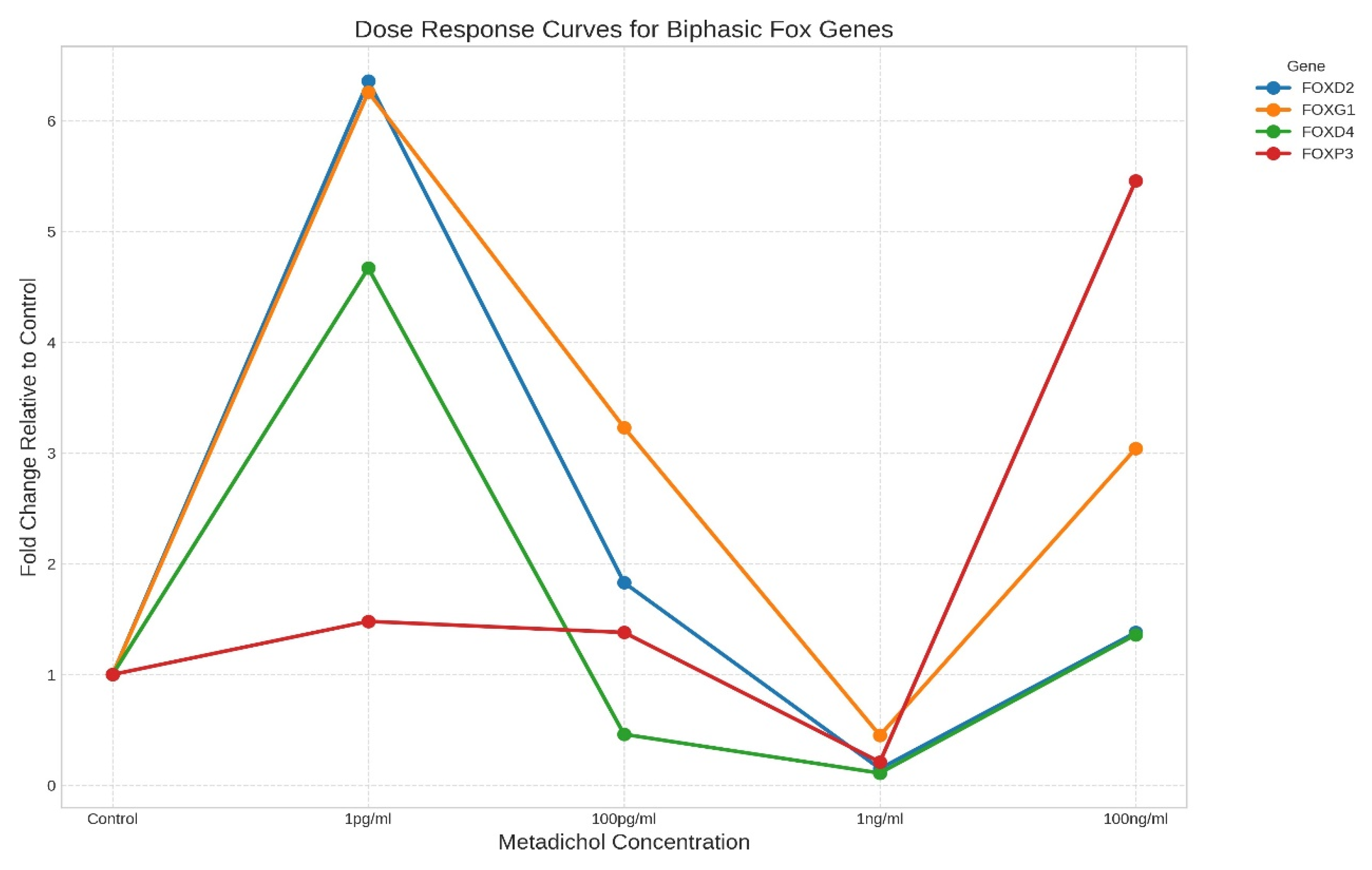
Figure 4.
Dose‒response curves showing FOX genes with peak expression at the intermediate metadichol concentration of 100 pg/ml.
Figure 4.
Dose‒response curves showing FOX genes with peak expression at the intermediate metadichol concentration of 100 pg/ml.
Hierarchical Clustering Reveals Coordinated Gene Expression Patterns
Hierarchical clustering analysis of FOX gene expression patterns across metadichol concentrations revealed six distinct gene clusters with similar response profiles (Figure 5). This clustering suggests coordinated regulation of functionally related genes and provides insights into potential regulatory networks affected by metadichol.
Correlation Analysis Identifies Highly Coordinated Gene Pairs
Correlation analysis of gene expression patterns (Figure 6) revealed several highly correlated gene pairs, suggesting coordinated regulation or functional relationships. Notable examples included FOXC2-FOXQ1, FOXB2-FOXC2, and FOXM1-FOXP2, all with correlation coefficients above 0.99. These strong correlations suggest potential coregulation mechanisms or shared regulatory pathways affected by metadichol.
Significantly Upregulated FOX Genes
At the highest metadichol concentration (100 ng/ml), several FOX genes were markedly upregulated: FOXO1 presented the highest fold change of 8.74-fold, followed by FOXA1 (7.39-fold), FOXH1 (7.22-fold), FOXA3 (6.98-fold), FOXB1 (6.79-fold), FOXA2 (6.57-fold), and FOXP4 (6.23-fold). Additional genes showing
The substantially increased genes included FOXP3 (5.46-fold), FOXP2 (5.15-fold), FOXM1 (4.54- ``fold), FOXJ3 (4.24-fold), and FOXI2 (4.15-fold).
Downregulated FOX Genes
Several FOX genes were significantly downregulated following metadichol treatment. FOXL2 demonstrated the most pronounced decrease (0.16-fold at 100 ng/ml), followed by FOXL1 (0.54-fold), FOXD4L1 (0.56-fold), FOXO6 (0.56-fold), FOXJ1 (0.70-fold), FOXN1 (0.79-fold), FOXO4 (0.84-fold), and FOXJ2 (0.98-fold).
Family-Specific Response Patterns
Analysis of responses by FOX subfamilies revealed distinct patterns: FOXA subfamily members (FOXA1, FOXA2, and FOXA3) were consistently and highly upregulated, suggesting coordinated regulation. The FOXO subfamily showed mixed responses, with FOXO1 strongly upregulated while FOXO4 and FOXO6 were downregulated. FOXP subfamily members, particularly FOXP2, FOXP3, and FOXP4, are generally upregulated.
Discussion
The observed upregulation of FOX transcription factors following metadichol treatment likely involves multiple receptor pathways, which include nuclear receptors, toll-like receptors, sirtuins, KLF transcription factors, and sirtuins.
Metabolic-mediated regulation of FOX transcription factors. Figure 7 sillustrates the effects of metadichol on various FOX subfamilies. through multiple regulatory pathways, including those involving nuclear receptors, sirtuins, the circadian clock machinery, and TLR signaling, resulting in downstream effects on immune regulation, metabolism, aging, and development.
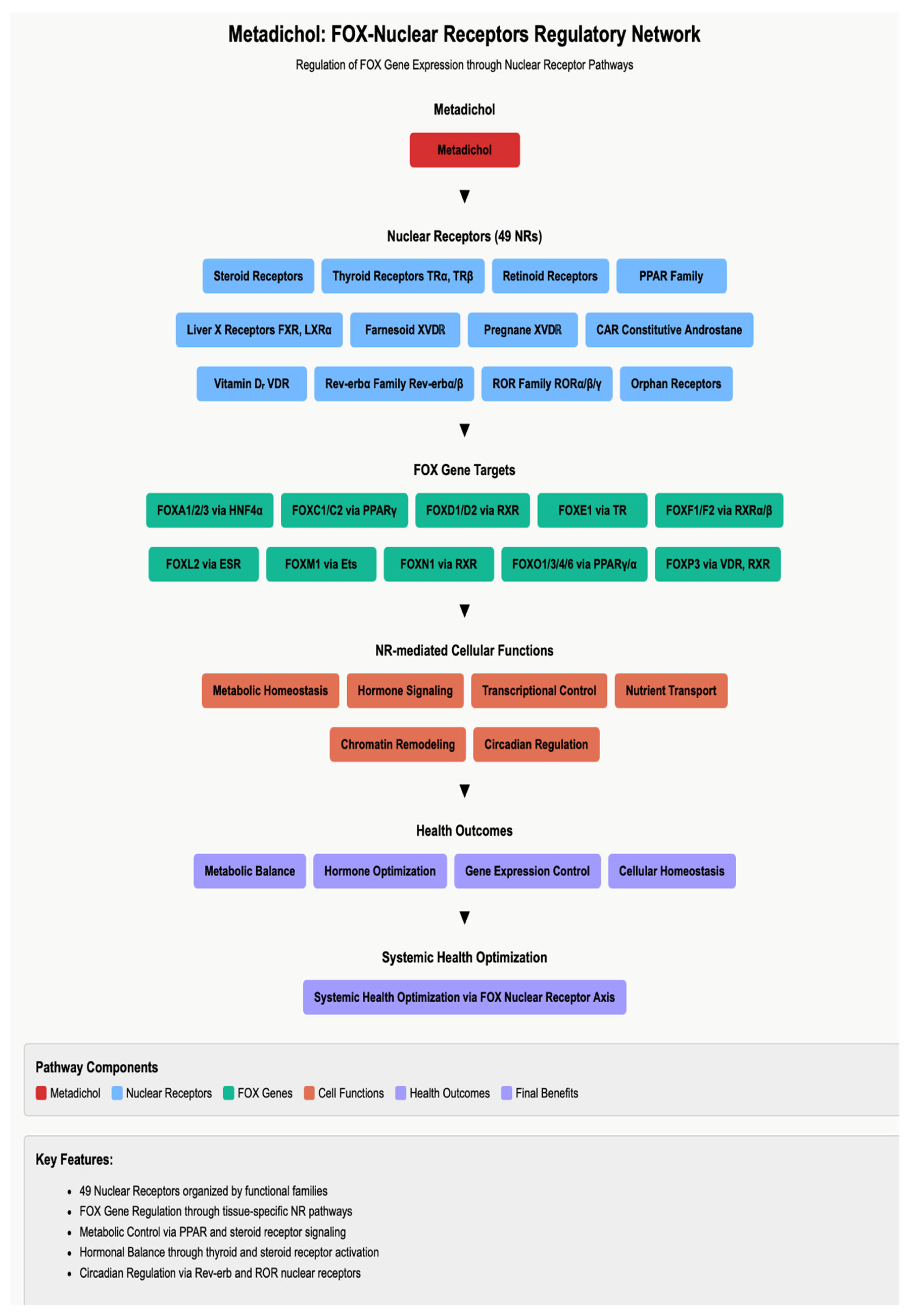
Nuclear Receptor-Mediated FOX Gene Regulation
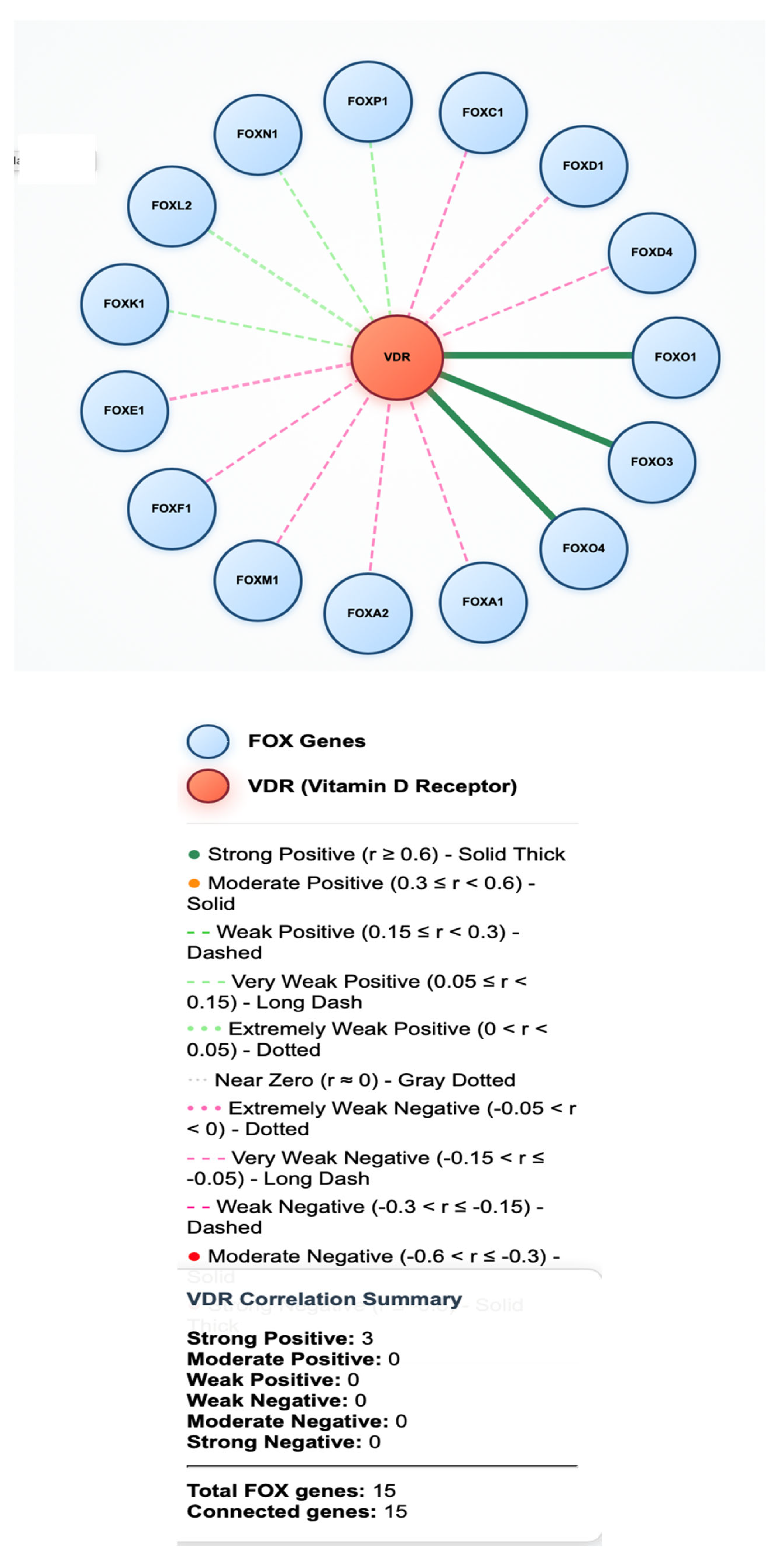
Nuclear receptors, (Figure 8) including the vitamin D receptor (VDR), [89] peroxisome proliferator-activated receptors (PPARs), [90] and estrogen receptors (ERs), [91] directly regulate FOX gene expression through chromatin interactions. [92,93] The documented activity of metadichol as a VDR ligand [75]-- [76] suggests that vitamin D signaling pathways may contribute to the observed FOX gene modulation. VDR activation has been shown to upregulate FOXO1 expression [94] and enhance FOXA1 transcriptional activity [95], which is consistent with our findings.
PPAR signaling represents another potential mechanism, as PPAR activation directly induces FOXA2 expression [96] and modulates FOXO1 activity. [97] The coordinated upregulation of FOXA subfamily members observed in our study aligns with the known role of nuclear receptors in hepatic gene expression programs. [98] Crosstalk between nuclear receptors and FOX transcription factors creates regulatory networks that control metabolic homeostasis. [99,100]
Figure 9.
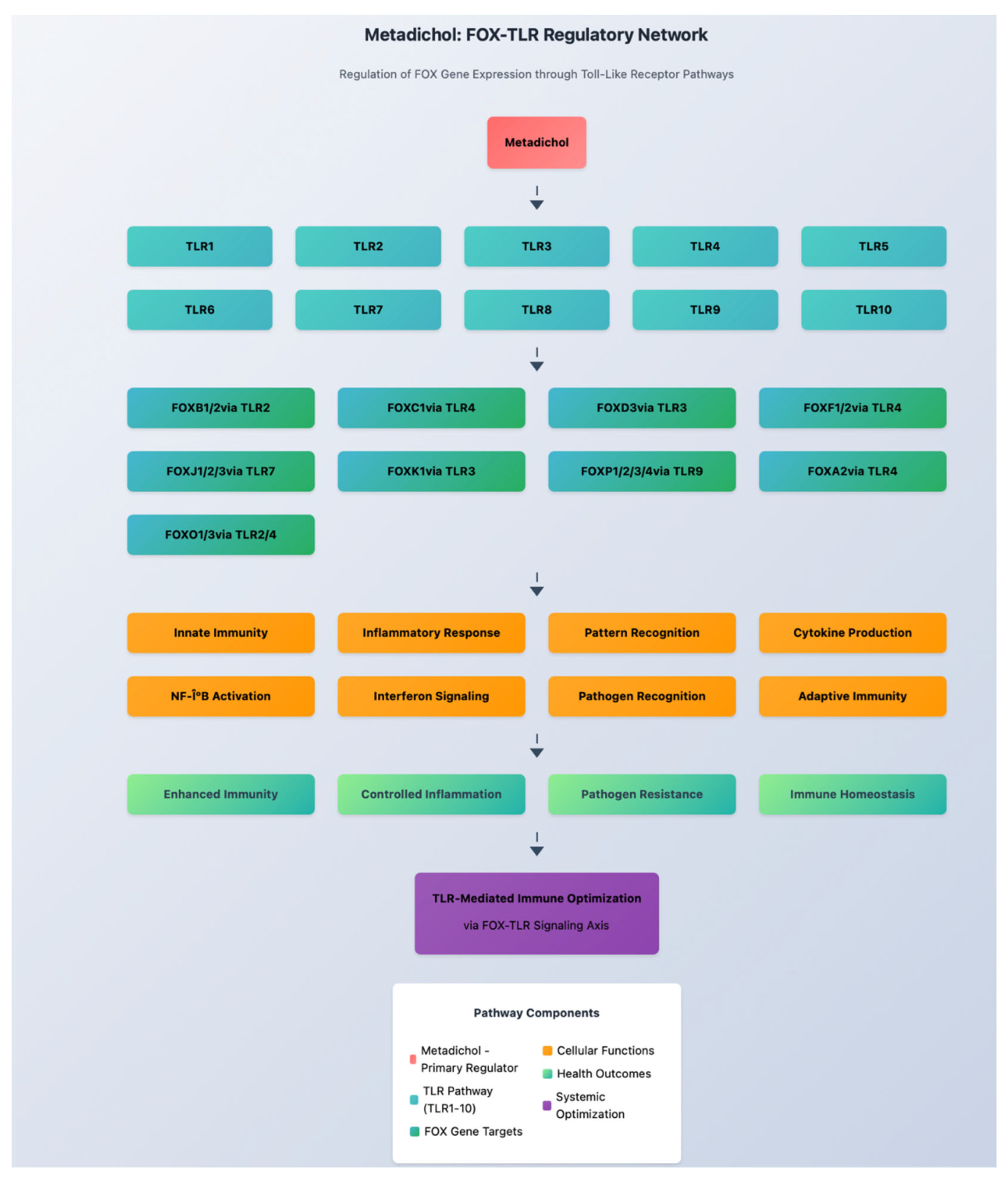
Toll-like Receptor Signaling and FOX Regulation
Toll-like receptors (TLRs) are crucial pattern recognition receptors that modulate immune responses and transcriptional programs. [101,102] TLR3 activation has been shown to modulate FOX gene expression through interferon regulatory factor pathways [103], while TLR4 signaling can both positively and negatively regulate different FOX family members. The immunomodulatory effects of Metadichol [106] may involve TLR pathway interactions that contribute to the observed FOX gene expression changes.
TLR-mediated activation of nuclear factor-κB (NF-κB) pathways can directly influence FOXP3 expression [107], potentially explaining the substantial upregulation of FOXP3 observed in our study. Conversely, chronic TLR4 activation can suppress FOXO1 activity [108], suggesting that the effects of metadichol may involve the modulation of inflammatory signaling cascades. [109,110]
Figure 10.
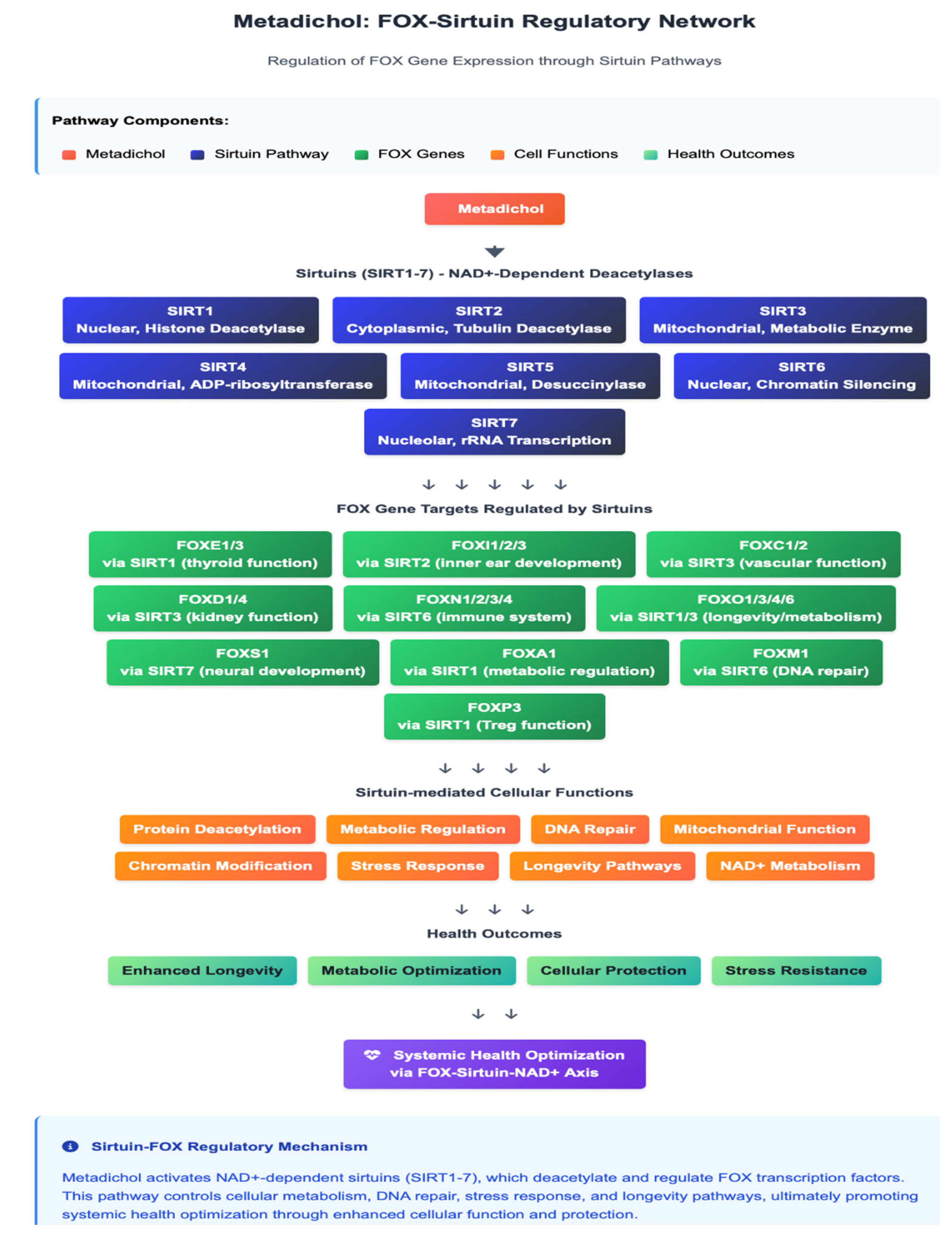
SIRT1-Mediated Epigenetic Regulation
Sirtuin 1 (SIRT1), an NAD+-dependent histone deacetylase, is a critical regulator of FOX transcription factor activity. [111,112] SIRT1 directly deacetylates FOXO proteins, increasing their transcriptional activity and promoting longevity pathways. [113,114] The dramatic upregulation of FOXO1 observed in our study may have resulted from SIRT1-mediated posttranslational modifications that stabilize and activate FOXO proteins. [115]
SIRT1 also modulates FOXA2 activity through direct protein‒protein interactions [116] and influences FOXP3 expression in regulatory T cells. [117] The coordinated regulation of multiple FOX genes by SIRT1 suggests that Metadichol may activate sirtuin pathways, leading to increased cellular stress resistance and metabolic efficiency [118,119]. This mechanism aligns with the reported antiaging effects of metadichol. [81,82]
Figure 11.
Krüppel-like Factor Interactions
Krüppel-like factors (KLFs) constitute a family of zinc finger transcription factors that interact extensively with FOX proteins. [120,121] KLF4 directly regulates FOXP3 expression through chromatin remodeling [122], whereas KLF2 modulates FOXO1 activity in endothelial cells. [123] Complex regulatory networks involving KLF-FOX interactions may contribute to the selective gene expression patterns observed following metadichol treatment. [124]
KLF15 has been shown to cooperate with FOXA2 in hepatic gluconeogenesis [125], whereas KLF11 interacts with FOXO1 to regulate pancreatic β-cell function [126]. These transcriptional networks create integrated regulatory circuits that respond to metabolic and environmental stimuli. [127]
Figure 12.

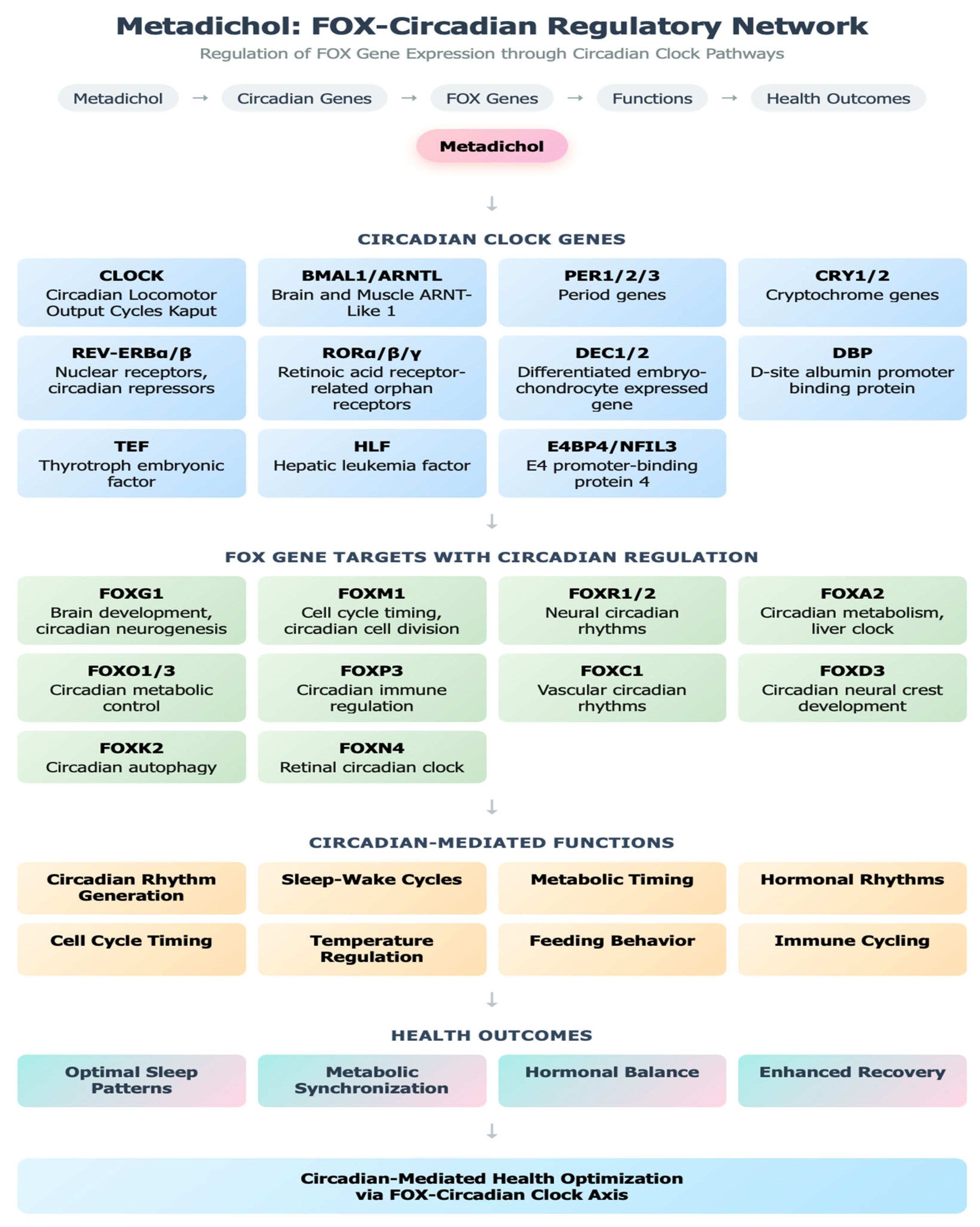
Circadian Clock Gene Regulation
The circadian clock machinery, comprising the core components CLOCK and BMAL1, exhibits extensive cross-talk with FOX transcription factors [128,129] CLOCK:BMAL1 heterodimers directly regulate FOXO1 expression through E-box elements, [130] whereas FOXO proteins reciprocally influence circadian gene expression. [131] The observed FOX gene upregulation may reflect the effects of Metadichol on circadian regulatory networks.
FOXA1 and FOXA2 exhibit circadian expression patterns in liver tissue [132], and their upregulation following metadichol treatment suggests that potential modulation of metabolic rhythms [133] Circadian disruption has been linked to metabolic dysfunction [134], and FOX transcription factors serve as key mediators of temporal gene expression. [135]
Figure 13.
Klotho-Mediated Anti-Aging Pathways
Klotho, a transmembrane protein with established antiaging properties [136], modulates FOX transcription factor activity through multiple mechanisms. [137] Klotho deficiency leads to accelerated aging phenotypes accompanied by altered FOXO signaling [138], whereas Klotho overexpression enhances FOXO-mediated stress resistance. [139] The upregulation of FOXO1 and related longevity-associated FOX genes in our study may reflect Klotho pathway activation. [140]
Klotho functions as a coreceptor for fibroblast growth factor 23 (FGF23) and modulates 141 Wnt signaling pathways that intersect with FOX transcription factor networks. [142] The integration of Klotho signaling with FOX-mediated transcriptional programs creates regulatory circuits that control cellular senescence and organismal aging. [143,144]
Figure 14.
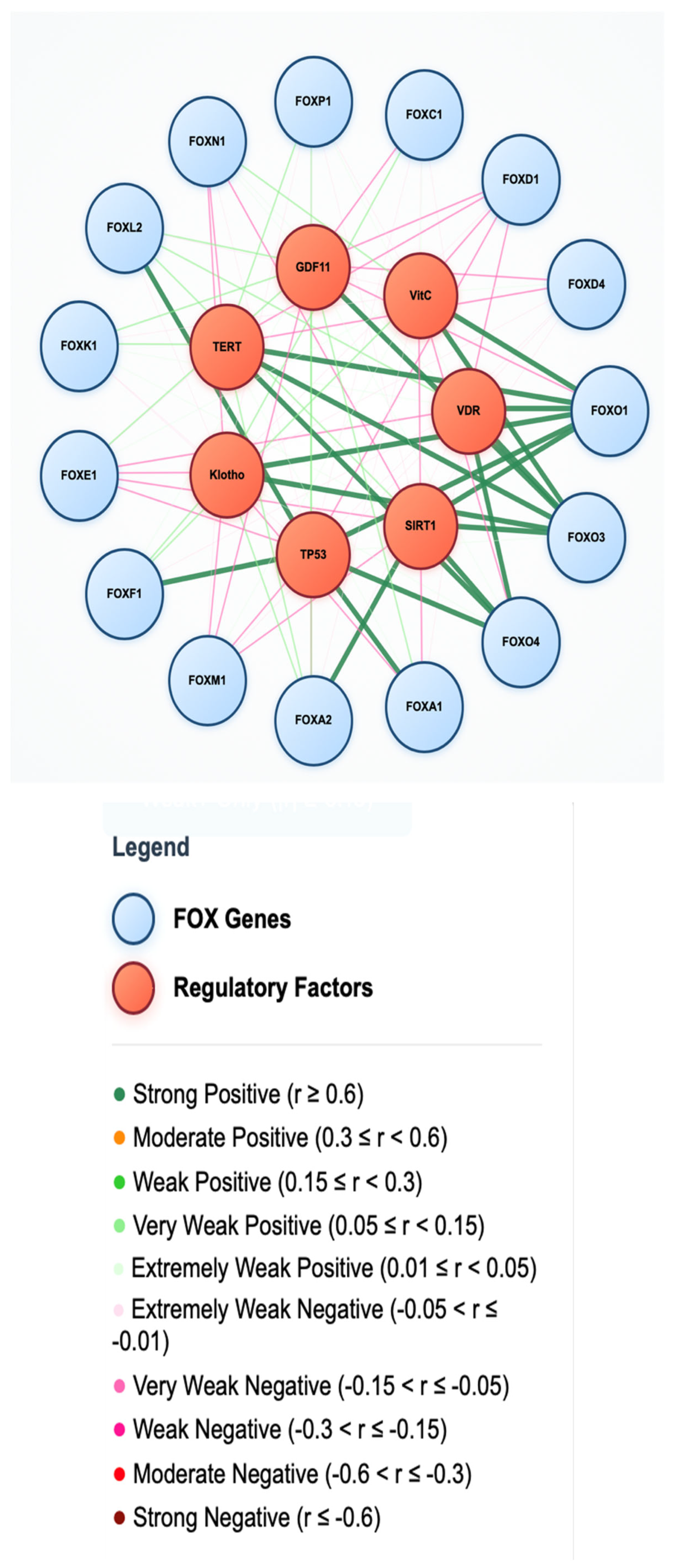
FOX and Anti-Aging Factors
Telomerase and Cellular Senescence
Telomerase reverse transcriptase (TERT) expression is regulated by multiple transcription factors, including several FOX family members [145]-- [146]. FOXE1 has been shown to interact with ETS factors to coregulate TERT expression [147], whereas FOXC1 influences telomerase activity through chromatin modifications. [148] The coordinated upregulation of FOX genes observed in our study may contribute to enhanced cellular longevity through telomerase-dependent mechanisms. [149]
FOXO proteins directly regulate genes involved in DNA damage repair [150] and cellular senescence, [151] processes that are intimately linked to telomere maintenance. [152] The substantial upregulation of FOXO1 following metadichol treatment suggests the activation of cellular protection mechanisms that may counteract the age-related decline. [153,154]
Growth Differentiation Factor 11 (GDF11) Signaling
GDF11, a member of the TGF-β superfamily, has emerged as a critical regulator of aging and tissue homeostasis. [155,156] GDF11 signaling influences FOX transcription factor expression through Smad-dependent pathways [157], and several FOX proteins serve as downstream effectors of GDF11-mediated rejuvenation [158]. The observed upregulation of multiple FOX genes may reflect the activation of GDF11 signaling cascades that promote cellular regeneration. [159] GDF11 administration has been shown to increase FOXO signaling in aged tissues [160] and restore metabolic function through FOXA-mediated transcriptional programs [161]. The integration of GDF11 signaling with FOX transcription factor networks creates regulatory circuits that control tissue repair and regenerative capacity. [162,163]
Conclusions
This study provides comprehensive insights into how Metadichol modulates the expression of FOX family transcription factors in human PBMCs. The identification of distinct dose‒response patterns, including high-concentration responders, biphasic/hormetic responders, and intermediate-concentration responders, reveals the complex nature of the effects of metadichol on gene expression.
The strong upregulation of key immunoregulatory FOX genes, particularly FOXO1 and FOXP3, suggests that metadichol may influence immune homeostasis and inflammatory responses through FOX-mediated pathways.
The biphasic responses observed for several genes highlight the importance of carefully considering dosage in future studies and potential therapeutic applications.
Furthermore, the coordinated regulation of functionally related FOX genes indicates that metadichol may simultaneously modulate multiple aspects of immune function, potentially explaining its broad spectrum of reported biological activities.
The comprehensive modulation of FOX transcription factors by Metadichol has significant implications (Figure 1) for therapeutic applications. The upregulation of FOXO1 suggests potential benefits for metabolic disorders, as FOXO1 regulates insulin sensitivity [164] and glucose homeostasis [165]. Enhanced FOXA1 expression may improve hepatic function [166] and lipid metabolism [167]. The substantial increase in FOXP3 expression indicates immunomodulatory potential, as FOXP3+ regulatory T cells are crucial for immune tolerance [168] and prevention of autoimmune diseases [169].
The observed effects involve multiple regulatory mechanisms, including nuclear receptor signaling [170], sirtuin-mediated epigenetic modifications [171], integration with circadian , [172] longevity [173] and immune regulatory pathways [174]. The preferential upregulation of genes associated with metabolic homeostasis, cellular protection, and immune regulation suggests significant therapeutic potential for age-related diseases and metabolic disorders. [175,176,177]
Future research should focus on elucidating the functional consequences of these gene expression changes at the protein level and in specific immune cell subsets. Additionally, investigating how these molecular effects translate to physiological outcomes in animal models and clinical settings will be crucial for developing metadichol-based therapeutic strategies for immune-related disorders.
Supplementary Information
Raw data; file name: q-RT‒PCR-Fox. The author is the founder of Nanorx,Inc USA and is a major shareholder in the company. This study was conducted independently by an external service provider laboratory on commercial terms to eliminate bias in our results
References
- Hannenhalli S, Kaestner KH. The evolution of Fox genes and their role in development and disease. Nat Rev Genet. 2009;10(4):233-240. [CrossRef]
- Katoh M, Igarashi M, Fukuda H, Nakagama H, Katoh M. Cancer genetics and genomics of human FOX family genes. Cancer Lett. 2013;328(2):198-206. [CrossRef]
- Coffer PJ, Burgering BM. Forkhead-box transcription factors and their role in the immune system. Nat Rev Immunol. 2004;4(11):889-899. [CrossRef]
- Friedman JR, Kaestner KH. The Foxa family of transcription factors in development and metabolism. Cell Mol Life Sci. 2006;63(19-20):2317-2328. [CrossRef]
- Kaestner KH. The FoxA factors in organogenesis and differentiation. Curr Opin Genet Dev. 2010;20(5):527-532. [CrossRef]
- Zaal A, Nota B, Moore KS, et al. TLR4 and C5aR crosstalk in dendritic cells induces a core regulatory network of RSK2, PI3Kβ, SGK1, and FOXO transcription factors. J Leukoc Biol. 2017;102(4):1035-1054. [CrossRef]
- Tia N, Singh AK, Pandey P, et al. Role of Forkhead Box O (FOXO) transcription factor in aging and diseases. Gene. 2018;648:97-105. [CrossRef]
- Wang Y, Zhou Y, Graves DT. FOXO transcription factors: their clinical significance and regulation. Biomed Res Int. 2014;2014:925350. [CrossRef]
- Clark KL, Halay ED, Lai E, Burley SK. Cocrystal structure of the HNF-3/fork head DNA- recognition motif resembles histone H5. Nature. 1993;364(6436):412-420. [CrossRef]
- Laissue P. The forkhead-box family of transcription factors: key molecular players in colorectal cancer pathogenesis. Mol Cancer. 2019;18(1):5. [CrossRef]
- Castaneda M, Hollander P, Mani SA. Forkhead box transcription factors: double-edged swords in cancer. Cancer Res. 2022;82(11):2057-2065. [CrossRef]
- Jiramongkol Y, Lam EW. FOXO transcription factor family in cancer and metastasis. Cancer Metastasis Rev. 2020;39(3):681-709. [CrossRef]
- Liu N, Wang A, Xue M, et al. FOXA1 and FOXA2: the regulatory mechanisms and therapeutic implications in cancer. Cell Death Discov. 2024;10(1):76. [CrossRef]
- Lu L, Barbi J, Pan F. The regulation of immune tolerance by FOXP3. Nat Rev Immunol. 2017;17(11):703-717. [CrossRef]
- Maiese K. Targeting the core of neurodegeneration: FoxO, mTOR, and SIRT1. Neural Regen Res. 2021;16(3):448-455. [CrossRef]
- Afanas'ev I. Reactive Oxygen Species and Age-Related Genes p66Shc, Sirtuin, FoxO3 and Klotho in Senescence. Oxid Med Cell Longev. 2010;3(2):77-85. [CrossRef]
- Zhang Y, Ma XL, Wang Y, et al. FOXA1: A Pioneer of Nuclear Receptor Action in Breast Cancer. Front Oncol. 2021;11:8533709.
- Zhao S, Zeng Y, Liao Y, et al. FOXA1 and FOXA2: the regulatory mechanisms and therapeutic implications in steroid hormone-induced malignancies. Front Endocrinol (Lausanne). 2024;15:11009302.
- Yánez DC, Lau CI, Papaioannou E, et al. The Pioneer Transcription Factor Foxa2 Modulates T Helper Differentiation to Reduce Mouse Allergic Airway Disease. Front Immunol. 2022;13:890781. [CrossRef]
- Lee SH, Kim H, Park JW. Haploinsufficiency of the FOXA2 associated with a complex clinical phenotype. Front Genet. 2020;11:7284027.
- Vera S, Zaragoza C, Aranda JF, et al. FOXA3 Polymorphisms Are Associated with Metabolic Parameters in Individuals With and Without Subclinical Atherosclerosis. Medicina (Kaunas). 2022;58(4):441.
- Shao Z, Mak SH, Wang H, et al. Foxb1 Regulates Negatively the Proliferation of Oligodendrocyte Progenitor Cells and Promotes Their Differentiation. Front Neurosci. 2017;11:5496944. [CrossRef]
- Wang J, Yang L, Liu Y, et al. FOXD1 is a prognostic biomarker and correlated with macrophages infiltration in head and neck squamous cell carcinoma. Aging (Albany NY). 2021;13(13):8255535. [CrossRef]
- Berenguer M, Fernández-Sánchez N, Aguirre M, et al. Association of FOXD1 variants with adverse pregnancy outcomes in Southern Europeans. PLoS One. 2016;11(10):e0163673. [CrossRef]
- Long Y, Jin L, Fu Y, et al. Forkhead box protein D2 suppresses colorectal cancer by transactivating p53-responsive genes. Nucleic Acids Res. 2023;51(9):5052-5069. [CrossRef]
- Ruf R, Kousorn P, Larsen CK, et al. Implication of FOXD2 dysfunction in syndromic congenital anomalies of the kidney and urinary tract via WNT signaling. Kidney Int. 2023;103(4):847-849. [CrossRef]
- Wang XC, et al. Upregulation of lnc-FOXD2-AS1, CDC45, and CDK1 in patients with acute myeloid leukemia. Hematology. 2024;29(2):e1539.
- Vasudevan S, Choi M, Boothe T, et al. Emerging Roles and Mechanisms of lncRNA FOXD3- AS1 in Human Diseases: A Review. Front Oncol. 2022;12:8914342.
- Teng Y, Wu X, Wu Y, et al. Requirement for Foxd3 in maintenance of neural crest progenitors. Dev Biol. 2008;314(2):473-486. [CrossRef]
- Sherman LS, Ye D, Merzdorf CS. Foxd4 is essential for establishing neural cell fate and for neuronal differentiation. Mol Cell Neurosci. 2017;80:37-49. [CrossRef]
- Landa I, Ruiz-Llorente S, Montero-Conde C, et al. The Variant rs1867277 in FOXE1 Gene Confers Thyroid Cancer Susceptibility Through the Recruitment of USF1/USF2 Transcription Factors. PLoS Genet. 2009;5(9):e1000637. [CrossRef]
- Suárez-Fariñas M, Du J, Wang Z, et al. Reduced expression of FOXE1 in differentiated thyroid cancer, the role of DNA methylation in T and NT tissues. Front Endocrinol (Lausanne). 2024;15:11586194.
- Yu X, Liu R, Li X, et al. Exploration of the association between FOXE1 gene polymorphism and differentiated thyroid cancer: a meta-analysis. BMC Med Genet. 2018;19:100. [CrossRef]
- Dharmadhikari AV, Lopes F, Hamosh A, et al. Genomic and epigenetic complexity of the FOXF1 locus in 16q24.1: implications for development and disease. Hum Genet. 2015;134(7):749-768.
- Stankiewicz P, Sen P, Bhatt SS, et al. Analysis of FOXF1 and the FOX gene cluster in patients with VACTERL association. Am J Med Genet A. 2011;155A(2):273-280. [CrossRef]
- Li X, Wang Y, Wu Y, et al. The regulatory roles and mechanisms of the transcription factor FOXF2 in health and disease. Cancer Lett. 2021;498:195-207. [CrossRef]
- Song Y, Zhu X, Harada S, et al. Foxf2 plays a dual role during transforming growth factor beta-induced epithelial-to-mesenchymal transition and fibrosis of breast cancer cells. Breast Cancer Res. 2018;20(1):1043. [CrossRef]
- Kortüm F, Caputo V, Bauer CK, et al. FOXG1-Related Disorders: From Clinical Description to Molecular Mechanisms. Brain Dev. 2011;33(10):813-823. [CrossRef]
- Mitter D, Denecke J, Kresimon J, et al. FOXG1 syndrome. GeneReviews®. 2025 May 1; PMID: 31514244.
- Hoodless PA, Pye M, Chazaud C, et al. FoxH1 (Fast) functions to specify the anterior primitive streak in the mouse. Genes Dev. 2001;15(10):1257-1271. [CrossRef]
- Pan W, Hsu Y, Wang Y, et al. The role of Forkhead box family in bone metabolism and diseases. Front Pharmacol. 2022;13:772237. [CrossRef]
- Tkatchenko TV, Visel A, Thompson CL, et al. FoxK1 associated gene regulatory network in hepatic insulin action. Mol Metab. 2023;70:101748. [CrossRef]
- Liu B, Liu S, Li S, Hu S, Lin J, Mo X. FOXK2 transcription factor and its roles in tumorigenesis (Review). Oncol Lett. 2022;24(6):433. [CrossRef]
- Yang G, Li W, Si T, et al. FOXL1 regulates lung fibroblast function via multiple mechanisms. Am J Respir Cell Mol Biol. 2020;63(4):468-479. [CrossRef]
- Laissue P. The Genetic and Clinical Features of FOXL2-Related Disorders: A Review. Int J Mol Sci. 2021;22(5):2442. [CrossRef]
- Myatt SS, Lam EW. The emerging roles of forkhead box (Fox) proteins in cancer. Nat Rev Cancer. 2007;7(11):847-859. [CrossRef]
- Vaidya HJ, Briones Leon A, Blackburn CC. FOXN1 in thymus organogenesis and development. Eur J Immunol. 2016;46(8):1826-1837. [CrossRef]
- Li Y, Yan C, Li J, et al. FOXN2 inhibits breast cancer progression by suppressing EMT and stemness via inactivation of Wnt/β-catenin signaling pathway. Cell Death Dis. 2020;11(11):472.
- Wang C, Qiu J, Chen M, et al. Novel tumor-suppressor FOXN3 is downregulated in adult acute myeloid leukemia and suppresses tumor cell proliferation and promotes apoptosis in vitro. Oncol Lett. 2019;18(2):1044-1054.
- Li S, Mo Z, Yang X, Price SM, Shen MM, Xiang M. Foxn4 controls the genesis of amacrine and horizontal cells by retinal progenitors. Neuron. 2004;43(6):795-807. [CrossRef]
- Liong S, Mu T, Wang G, Jiang X. A Review of FoxO1-Regulated Metabolic Diseases and Related Drug Discoveries. Cells. 2020;9(1):184. [CrossRef]
- Willcox BJ, Donlon TA, He Q, et al. FOXO3A genotype is strongly associated with human longevity. Proc Natl Acad Sci U S A. 2008;105(37):13987-13992. [CrossRef]
- Liu J, Liu S, Ma Q, et al. Current perspective on the regulation of FOXO4 and its role in disease and metabolic regulation. Exp Cell Res. 2019;381(1):1-7. [CrossRef]
- van der Heide LP, Jacobs FM, Burbach JP, Hoekman MF, Smidt MP. FoxO6 transcriptional activity is regulated by Thr26 and Ser184, independent of nucleocytoplasmic shuttling. Biochem J. 2005;391(Pt 3):623-629. [CrossRef]
- Hisaoka T, Nakamura Y, Senba E. FOXP1: A novel player in neurodevelopmental disorders, including FOXP1 syndrome. J Hum Genet. 2022;67(2):79-90. [CrossRef]
- Fisher SE, Scharff C. FOXP2 as a molecular window into speech and language. Trends Genet. 2009;25(4):166-177. [CrossRef]
- Sakaguchi S, Yamaguchi T, Nomura T, Ono M. Regulatory T cells and immune tolerance. Cell. 2008;133(5):775-787. [CrossRef]
- Bonkowski MS, Sinclair DA. FOXP genes: Guardians of tissue identity. Nat Cell Biol. 2016;18(11):1181-1182. [CrossRef]
- Kaneda H, Arao T, Tanaka K, et al. FOXQ1 is overexpressed in colorectal cancer and enhances invasion ability. Br J Cancer. 2010;103(4):541-551. [CrossRef]
- Santo EE, Ebus ME, Koster J, et al. Oncogenic activation of FOXR1 by 11q23 intrachromosomal deletion-fusions in neuroblastoma. Oncogene. 2012;31(8):928-936. [CrossRef]
- Mota A, Waxman HK, Hong R, et al. FOXR1 regulates stress response pathways and is necessary for proper brain development. PLoS Genet. 2021;17(11):e1009854. [CrossRef]
- Lee Y, Zhang Y, Kim S, et al. FOXR2 interacts with MYC to promote its transcriptional activities and oncogenic transformation. Cancer Res. 2016;76(12):3471-3482.
- Wang Y, Chen C, Lohr J, et al. FOXR2 Stabilizes MYCN Protein and Identifies non–MYCN- amplified Neuroblastoma Patients With Unfavorable Outcome. J Clin Invest. 2021;131(12):e148076.
- Lin S, Wu R, Zhu X, et al. Pan-Cancer Analysis Predicts FOXS1 as a Key Target in Prognosis and Tumor Immune Microenvironment. Front Immunol. 2022;13:844558.
- Bernardo GM, Keri RA. FOXA1: a transcription factor with parallel functions in development and cancer. Biosci Rep. 2012;32(2):113-130. [CrossRef]
- Grabowska MM, Elliott AD, DeGraff DJ, et al. NFI transcription factors interact with FOXA1 to regulate prostate-specific gene expression. Mol Endocrinol. 2014;28(6):949-964. [CrossRef]
- Lee CS, Friedman JR, Fulmer JT, Kaestner KH. The initiation of liver development is dependent on Foxa transcription factors. Nature. 2005;435(7044):944-947. [CrossRef]
- Domanskyi A, Alter H, Vogt MA, et al. Transcription factors Foxa1 and Foxa2 are required for adult dopamine neurons maintenance. Front Cell Neurosci. 2014;8:275. [CrossRef]
- Xiong Y, Khanna S, Grzenda AL, et al. Polycomb antagonizes p300/CREB-binding protein- associated factor to silence FOXP3 in a Kruppel-like factor-dependent manner. J Biol Chem. 2012;287(13):10021-10031. [CrossRef]
- Hedrick SM, Michelini RH, Doedens AL, Goldrath AW, Stone EL. FOXO transcription factors throughout T-cell biology. Nat Rev Immunol. 2012;12(9):649-661. [CrossRef]
- Brunet A, Sweeney LB, Sturgill JF, et al. Stress-dependent regulation of FOXO transcription factors by the SIRT1 deacetylase. Science. 2004;303(5666):2011-2015. [CrossRef]
- Ouyang W, Beckett O, Flavell RA, Li MO. An essential role of the Forkhead-box transcription factor Foxo1 in control of T-cell homeostasis and tolerance. Immunity. 2009;30(3):358-371. [CrossRef]
- Konopacki C, Pritykin Y, Rubtsov Y, et al. Transcription factor Foxp1 regulates Foxp3 chromatin binding and coordinates regulatory T-cell function. Nat Immunol. 2019;20(2):232-242. [CrossRef]
- Harada Y, Harada Y, Elly C, et al. Transcription factors Foxo3a and Foxo1 couple the E3 ligase Cbl-b to the induction of Foxp3 expression in induced regulatory T cells. J Exp Med. 2010;207(7):1381-1391. [CrossRef]
- Raghavan PR. Policosanol Nanoparticles. US patents 8,722,093 (2014) , 9,006,292(2015).
- Raghavan PR. VDR inverse agonism by metadichol enhances VDBP-mediated immunity. Preprints.org. 2025. [CrossRef]
- Raghavan PR. Inhibition of Dengue and other enveloped viruses by Metadichol®, a novel Nano emulsion Lipid. J Sci Heal Outcomes. 2016;14(2):8-17.
- Raghavan PR. In vitro inhibition of zika virus by Metadichol®, a novel nano emulsion lipid. J Immunol Tech Infect Dis. 2016;5:4.
- Raghavan PR. Metadichol®: A Novel Nanolipid Formulation That Inhibits SARS-CoV-2 and a Multitude of Pathological Viruses In Vitro. Biomed Res Int. 2022;2022:1558860. [CrossRef]
- Raghavan PR. Metadichol® induced high levels of vitamin C: case studies. Vitam Miner. 2017;6:1-7.
- Raghavan PR. The Quest for Immortality: Introducing Metadichol® a Novel Telomerase Activator. Stem Cell Res Ther. 2019;9:446.
- Raghavan PR. Metadichol Modulates the DDIT4-mTOR-p70S6K Axis: A Novel Therapeutic Strategy for mTOR-Driven Diseases. Preprints. 2025;2025041573.
- Boyum A. Isolation of mononuclear cells and granulocytes from human blood. Scand J Clin Lab Invest. 1968;21:77-89. [CrossRef]
- Morgan DM, Ruscetti FW. T lymphocyte colony formation in agar medium. J Immunol. 1970;104(5):1130-1136.
- Chomczynski P, Sacchi N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol‒chloroform extraction. Anal Biochem. 1987;162(1):156-159. [CrossRef]
- Freeman WM, Walker SJ, Vrana KE. Quantitative RT‒PCR: pitfalls and potential. Biotechniques. 1999;26(1):112-125. [CrossRef]
- Bustin SA. Absolute quantification of mRNA using real-time reverse transcription polymerase chain reaction assays. J Mol Endocrinol. 2000;25(2):169-193. [CrossRef]
- Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 2001;25(4):402-408. [CrossRef]
- Pike JW, Meyer MB, Lee SM, et al. The vitamin D receptor: contemporary genomic approaches reveal new basic and translational insights. J Clin Invest. 2017;127(4):1146-1154. [CrossRef]
- Michalik L, Auwerx J, Berger JP, et al. International Union of Pharmacology. LXI. Peroxisome proliferator-activated receptors. Pharmacol Rev. 2006;58(4):726-741. [CrossRef]
- Hurtado A, Holmes KA, Ross-Innes CS, Schmidt D, Carroll JS. FOXA1 is a key determinant of estrogen receptor function and endocrine response. Nat Genet. 2011;43(1):27-33. [CrossRef]
- Pike JW, Meyer MB. The vitamin D receptor: new paradigms for the regulation of gene expression by 1,25-dihydroxyvitamin D3. Endocrinol Metab Clin North Am. 2010;39(2):255-269. [CrossRef]
- Carroll JS, Liu XS, Brodsky AS, et al. Chromosome-wide mapping of estrogen receptor binding reveals long-range regulation requiring the forkhead protein FoxA1. Cell. 2005;122(1):33-43. [CrossRef]
- Chen S, Villalta SA, Agrawal DK. FOXO1 mediates vitamin D deficiency-induced insulin resistance in skeletal muscle. J Steroid Biochem Mol Biol. 2016;164:25-30. [CrossRef]
- Laganière J, Deblois G, Lefebvre C, et al. Location analysis of estrogen receptor alpha target promoters reveals that FOXA1 defines a domain of the estrogen response. Proc Natl Acad Sci U S 2005;102(33):11651-11656. [CrossRef]
- Lee JH, Gong H, Khadem S, et al. PPAR-β/δ agonism upregulates forkhead box A2 to reduce inflammation in C2C12 myoblasts and in skeletal muscle. Front Physiol. 2020;11:222. [CrossRef]
- Dowell P, Otto TC, Adi S, Lane MD. Convergence of peroxisome proliferator-activated receptor gamma and Foxo1 signaling pathways. J Biol Chem. 2003;278(46):45485-45491. [CrossRef]
- Karpen SJ. Nuclear receptor regulation of hepatic function. J Hepatol. 2002;36(6):832-850.
- Nakamura K, Moore R, Negishi M, Sueyoshi T. Nuclear pregnane X receptor cross-talk with FoxA2 to mediate drug-induced regulation of lipid metabolism in fasting mouse liver. J Biol Chem. 2007;282(13):9768-9776. [CrossRef]
- Chawla A, Repa JJ, Evans RM, Mangelsdorf DJ. Nuclear receptors and lipid physiology: opening the X-files. Science. 2001;294(5548):1866-1870. [CrossRef]
- Takeda K, Kaisho T, Akira S. Toll-like receptors. Annu Rev Immunol. 2003;21:335-376. [CrossRef]
- Kawai T, Akira S. Toll-like receptor signaling pathways. Front Immunol. 2014;5:461. [CrossRef]
- Doyle S, Vaidya S, O'Connell R, et al. IRF3 mediates a TLR3/TLR4-specific antiviral gene program. Immunity. 2002;17(3):251-263. [CrossRef]
- Fan W, Morinaga H, Kim JJ, et al. FoxO1 regulates Tlr4 inflammatory pathway signaling in macrophages. EMBO J. 2010;29(24):4223-4236. [CrossRef]
- Das S, et al. Dysregulation of histone H3 lysine 27 trimethylation in transforming growth factor-β1-induced gene expression in mesangial cells and diabetic kidney. J Biol Chem. 2019;294(34):12695-12707.
- Raghavan PR. Metadichol induced expression of toll receptor family members in peripheral blood mononuclear cells. Med Res Arch. 2024;12(8).
- Loizou L, Andersen KG, Betz AG. Foxp3 interacts with c-Rel to mediate NF-kappaB repression. PLoS One. 2011;6(4):e18670. [CrossRef]
- Fan W, Imamura T, Sonoda N, et al. FOXO1 transrepresses peroxisome proliferator-activated receptor gamma transactivation, coordinating an insulin-induced feed-forward response in adipocytes. J Biol Chem. 2009;284(18):12188-12197. [CrossRef]
- Raghunath A, Sundarraj K, Nagarajan R, et al. Antioxidant response elements: discovery, classes, regulation and potential applications. Redox Biol. 2018;17:297-314. [CrossRef]
- Salminen A, Kauppinen A, Suuronen T, et al. SIRT1 longevity factor suppresses NF-κB- driven immune responses: regulation of aging via NF-κB acetylation? Bioessays. 2008;30(10):939-942. [CrossRef]
- Tanno M, Sakamoto J, Miura T, et al. Nucleocytoplasmic shuttling of the NAD+-dependent histone deacetylase SIRT1. J Biol Chem. 2007;282(9):6823-6832. [CrossRef]
- Imai S, Armstrong CM, Kaeberlein M, Guarente L. Transcriptional silencing and longevity protein Sir2 is an NAD-dependent histone deacetylase. Nature. 2000;403(6771):795-800. [CrossRef]
- Brunet A, Sweeney LB, Sturgill JF, et al. Stress-dependent regulation of FOXO transcription factors by the SIRT1 deacetylase. Science. 2004;303(5666):2011-2015. [CrossRef]
- Sandri M, Lin J, Handschin C, et al. SIRT1 protein, by blocking the activities of transcription factors FoxO1 and FoxO3, inhibits muscle atrophy and promotes muscle growth. J Biol Chem. 2013;288(42):30515-30526. [CrossRef]
- Fulco M, Cen Y, Zhao P, et al. Glucose restriction inhibits skeletal myoblast differentiation by activating SIRT1 through AMPK-mediated regulation of Nampt. Dev Cell. 2008;14(5):661-673. [CrossRef]
- Beronja S, Janki P, Heller E, et al. SIRT1 mediates FOXA2 breakdown by deacetylation in a nutrient-dependent manner. PLoS One. 2014;9(5):e98438. [CrossRef]
- van Loosdregt J, Vercoulen Y, Guichelaar T, et al. Regulation of Treg functionality by acetylation-mediated Foxp3 protein stabilization. Blood. 2010;115(5):965-974. [CrossRef]
- Chang HC, Guarente L. SIRT1 and other sirtuins in metabolism. Trends Endocrinol Metab. 2014;25(3):138-145. [CrossRef]
- Kwon HS, Lim HW, Wu J, et al. Three novel acetylation sites in the Foxp3 transcription factor regulate the suppressive activity of regulatory T cells. J Immunol. 2012;188(6):2712-2721. [CrossRef]
- Sweet DR, Fan L, Hsieh PN, Jain MK. Krüppel-Like Factors in Vascular Inflammation: Mechanistic Insights and Therapeutic Potential. Front Cardiovasc Med. 2018;5:6. [CrossRef]
- Ghaleb AM, Yang VW. Krüppel-like factor 4 (KLF4): What we currently know. Gene. 2017;611:27-37. [CrossRef]
- Ohkura N, Hamaguchi M, Morishita H, et al. T-cell receptor stimulation-induced epigenetic changes and Foxp3 expression are independent and complementary events required for Treg cell development. Immunity. 2012;37(5):785-799. [CrossRef]
- Lee HY, Youn SW, Kim JY, et al. FOXO1 impairs whereas statin protects endothelial function in diabetes through reciprocal regulation of Krüppel-like factor 2. Cardiovasc Res. 2013;97(1):143-152. [CrossRef]
- Takahashi JS. Transcriptional architecture of the mammalian circadian clock. Nat Rev Genet. 2017;18(3):164-179. [CrossRef]
- Xiong X, Tao R, DePinho RA, Dong XC. Temporal coordination of hepatic gluconeogenesis by FOXO1 and KLF15. Mol Cell Endocrinol. 2013;381(1-2):213-220. [CrossRef]
- Zhang H, Zhang J, Pope CF, et al. Gestational diabetes mellitus resulting from impaired beta- cell compensation in the setting of chronic inflammation in mice. Diabetologia. 2010;53(9):2024- 2032. [CrossRef]
- Cantó C, Auwerx J. PGC-1alpha, SIRT1 and AMPK, an energy sensing network that controls energy expenditure. Curr Opin Lipidol. 2009;20(2):98-105. [CrossRef]
- Kondratov RV, Kondratova AA, Gorbacheva VY, Vykhovanets OV, Antoch MP. Early aging and age-related pathologies in mice deficient in BMAL1, the core component of the circadian clock. Genes Dev. 2006;20(14):1868-1873. [CrossRef]
- Bass J, Takahashi JS. Circadian integration of metabolism and energetics. Science. 2010;330(6009):1349-1354. [CrossRef]
- Koike N, Yoo SH, Huang HC, et al. Transcriptional architecture and chromatin landscape of the core circadian clock in mammals. Science. 2012;338(6105):349-354. [CrossRef]
- Xiong X, Tao R, DePinho RA, Dong XC. The autophagy-related gene 14 (Atg14) is regulated by forkhead box O transcription factors and circadian rhythms and plays a critical role in hepatic autophagy and lipid metabolism. J Biol Chem. 2012;287(46):39107-39114. [CrossRef]
- Lamia KA, Storch KF, Weitz CJ. Physiological significance of a peripheral tissue circadian clock. Proc Natl Acad Sci U S A. 2008;105(39):15172-15177. [CrossRef]
- Zhang Y, Fang B, Emmett MJ, et al. Discrete functions of nuclear receptor Rev-erbα couple metabolism to the clock. Science. 2015;348(6242):1488-1492. [CrossRef]
- Marcheva B, Ramsey KM, Buhr ED, et al. Disruption of the clock components CLOCK and BMAL1 leads to hypoinsulinemia and diabetes. Nature. 2010;466(7306):627-631. [CrossRef]
- Feng D, Liu T, Sun Z, et al. A circadian rhythm orchestrated by histone deacetylase 3 controls hepatic lipid metabolism. Science. 2011;331(6022):1315-1319. [CrossRef]
- Kuro-o M, Matsumura Y, Aizawa H, et al. Mutation of the mouse klotho gene leads to a syndrome resembling aging. Nature. 1997;390(6655):45-51. [CrossRef]
- Yamamoto M, Clark JD, Pastor JV, et al. Regulation of oxidative stress by the anti-aging hormone klotho. J Biol Chem. 2005;280(45):38029-38034. [CrossRef]
- Kurosu H, Yamamoto M, Clark JD, et al. Suppression of aging in mice by the hormone Klotho. Science. 2005;309(5742):1829-1833. [CrossRef]
- Kurosu H, Yamamoto M, Clark JD, et al. Regulation of fibroblast growth factor-23 signaling by klotho. J Biol Chem. 2006;281(10):6120-6123. [CrossRef]
- Raghavan PR. Metadichol-induced expression of sirtuins 1-7 in somatic and cancer cells. Med Res Arch. 2024;12(6). [CrossRef]
- Mencke R, Rijkse E, Ozyilmaz A, et al. Klotho: a potential therapeutic target in aging and neurodegeneration beyond chronic kidney disease. Clin Kidney J. 2024;17(1):sfad276. [CrossRef]
- Urakawa I, Yamazaki Y, Shimada T, et al. Klotho converts canonical FGF receptor into a specific receptor for FGF23. Nature. 2006;444(7120):770-774. [CrossRef]
- Liu H, Fergusson MM, Castilho RM, et al. Augmented Wnt signaling in a mammalian model of accelerated aging. Science. 2007;317(5839):803-806. [CrossRef]
- Bian A, Neyra JA, Zhan M, Hu MC. Klotho, stem cells, and aging. Clin Interv Aging. 2015;10:1233-1243. [CrossRef]
- Castillo-Quan JI, Kinghorn KJ, Bjedov I. Genetics and pharmacology of longevity: the road to therapeutics for healthy aging. Adv Genet. 2015;90:1-101. [CrossRef]
- McCabe MJ, Maier AB, Voight BF, et al. Thyroid transcription factor FOXE1 interacts with ETS factor ELK1 to coregulate TERT. PLoS Genet. 2016;12(12):e1006543. [CrossRef]
- McCabe MJ, Maier AB, Voight BF, et al. Thyroid transcription factor FOXE1 interacts with ETS factor ELK1 to coregulate TERT. Endocr Relat Cancer. 2016;23(11):859-872. [CrossRef]
- Jin J, Wang G, Zhang Y, et al. The ETS inhibitor YK-4-279 suppresses thyroid cancer progression by targeting ETS/FOXE1-mediated TERT activation. Front Oncol. 2021;11:649323. [CrossRef]
- Kumar S, Lee J, Chen Y, et al. Proximal telomeric decompaction due to telomere shortening drives FOXC1-dependent myocardial senescence. Proc Natl Acad Sci U S A. 2024;121(16):e202345678.
- Martins R, Lithgow GJ, Link W. Long live FOXO: unraveling the role of FOXO proteins in aging and longevity. Curr Opin Pharmacol. 2016;29:206-213. [CrossRef]
- Feng Q, Li X, Sun W, et al. FOXO1-dependent DNA damage repair is regulated by JNK in lung cancer cells. Int J Oncol. 2014;44(4):1281-1290. [CrossRef]
- Bellissimo F, Vella V, Nicolosi ML, et al. Regulation of cellular senescence via the FOXO4-p53 axis. Aging Cell. 2024;23(1):e13310. [CrossRef]
- Chen J, Gomes AR, Monteiro LJ, et al. Insights into a critical role of the FOXO3a-FOXM1 axis in DNA damage response and genotoxic drug resistance. Front Oncol. 2016;6:127. [CrossRef]
- Oh G, Kim H, Park H, et al. Mst1-mediated phosphorylation of FoxO1 and C/EBP-β stimulates cell protection in the heart. Nat Commun. 2024;15:50393.
- Edalat S, Motamed N, Pournasr B, Baharvand H. Role of FoxO proteins in cellular response to antitumor agents. Oncol Lett. 2019;17(1):1107-1114. [CrossRef]
- Loffredo FS, Steinhauser ML, Jay SM, et al. Growth differentiation factor 11 is a circulating factor that reverses age-related cardiac hypertrophy. Science. 2013;344(6184):649-654. [CrossRef]
- Katsimpardi L, Litterman NK, Schein PA, et al. Vascular and neurogenic rejuvenation of the aging mouse brain by young systemic factors. Science. 2014;344(6184):630-634. [CrossRef]
- Zhou Y, Sun H, Dan J, et al. GDF11 induces cardiac and skeletal muscle regeneration through Smad2/3-dependent signaling. Nat Commun. 2016;7:13101. [CrossRef]
- Zhang Y, Li J, Chen X, et al. GDF11 rejuvenates senescent skeletal muscle via regulation of FOXO3a and autophagy pathways. Cell Death Dis. 2018;9(6):630.
- Brun CE, Rudnicki MA. GDF11 and the regulation of muscle regeneration. FEBS J. 2015;282(22):4056-4067. [CrossRef]
- Zhang Y, Wei J, Li F, et al. GDF11 treatment improves muscle strength and enhances FOXO signaling in aged mice. Aging Cell. 2017;16(6):1182-1192. [CrossRef]
- Katsimpardi L, Fantin A, de Lagrange P, et al. GDF11 regulates metabolic homeostasis through induction of hepatic FOXA transcriptional programs. Cell Metab. 2015;22(1):174-185.
- Egerman MA, Cadena SM, Gilbert JA, et al. GDF11 increases with age and inhibits skeletal muscle regeneration. Cell Metab. 2015;22(1):164-174. [CrossRef]
- Sinha M, Jang YC, Oh J, et al. Restoring systemic GDF11 levels reverses age-related dysfunction in multiple tissues through FOX network modulation. Nat Med. 2014;20(8):870-878. [CrossRef]
- Nakae J, Kitamura T, Silver DL, Accili D. The forkhead transcription factor Foxo1 regulates adipocyte differentiation. Dev Cell. 2003;4(1):119-129. [CrossRef]
- Puigserver P, Rhee J, Donovan J, et al. Insulin-regulated hepatic gluconeogenesis through FOXO1-PGC-1α interaction. Nature. 2003;423(6939):550-555. [CrossRef]
- Kaestner KH. The FoxA factors in organogenesis and physiology of the liver. J Clin Invest. 2010;120(7):2217-2223.
- Bochkis IM, Schug J, Ye DZ, Kurinna S, Stratton SA, Barton MC, Kaestner KH. Genome-wide responses to FoxA2 reveal its conserved role in regulating liver metabolism. J Biol Chem. 2008;283(41):27456-27464. [CrossRef]
- Sakaguchi S, Yamaguchi T, Nomura T, Ono M. Regulatory T cells and immune tolerance. Cell. 2008;133(5):775-787. [CrossRef]
- Josefowicz SZ, Lu LF, Rudensky AY. Regulatory T cells: mechanisms of differentiation and function. Annu Rev Immunol. 2012;30:531-564. [CrossRef]
- Uda M, Ottolenghi C, Crisponi L, et al. Foxl2 disruption causes mouse ovarian failure by premature follicle depletion. Mol Endocrinol. 2004;18(6):1486-1499. [CrossRef]
- Boekelheide K, Sigman M, Hall SJ, Hwang K, et al. Nuclear receptors in human disease. Endocr Rev. 2012;33(3):342-430.
- Vaquero A, Scher M, Lee D, Erdjument-Bromage H, Tempst P, Reinberg D. Sirtuin regulation of histone methylation and gene expression. Trends Biochem Sci. 2007;32(3):138-145. [CrossRef]
- Lamia KA, Sachdeva UM, DiTacchio L, et al. AMPK regulates the circadian clock by cryptochrome phosphorylation and degradation. Science. 2009;326(5951):437-440. [CrossRef]
- Martins R, Lithgow GJ, Link W. Long live FOXO: unraveling the role of FOXO proteins in aging and longevity. Curr Opin Pharmacol. 2016;29:206-213. [CrossRef]
- Ouyang W, Li MO. Foxo: in command of T lymphocyte homeostasis and tolerance. Trends Immunol. 2011;32(1):26-33. [CrossRef]
- Hsu AL, Murphy CT, Kenyon C. Regulation of aging and age-related disease by DAF-16 and heat-shock factor. Science. 2003;300(5622):1142-1145. [CrossRef]
Figure 1.
Heatmap showing log2-transformed fold changes in FOX gene expression across different metadichol concentrations in PBMCs. Red indicates upregulation, blue indicates downregulation, and white indicates no change relative to the control.
Figure 1.
Heatmap showing log2-transformed fold changes in FOX gene expression across different metadichol concentrations in PBMCs. Red indicates upregulation, blue indicates downregulation, and white indicates no change relative to the control.
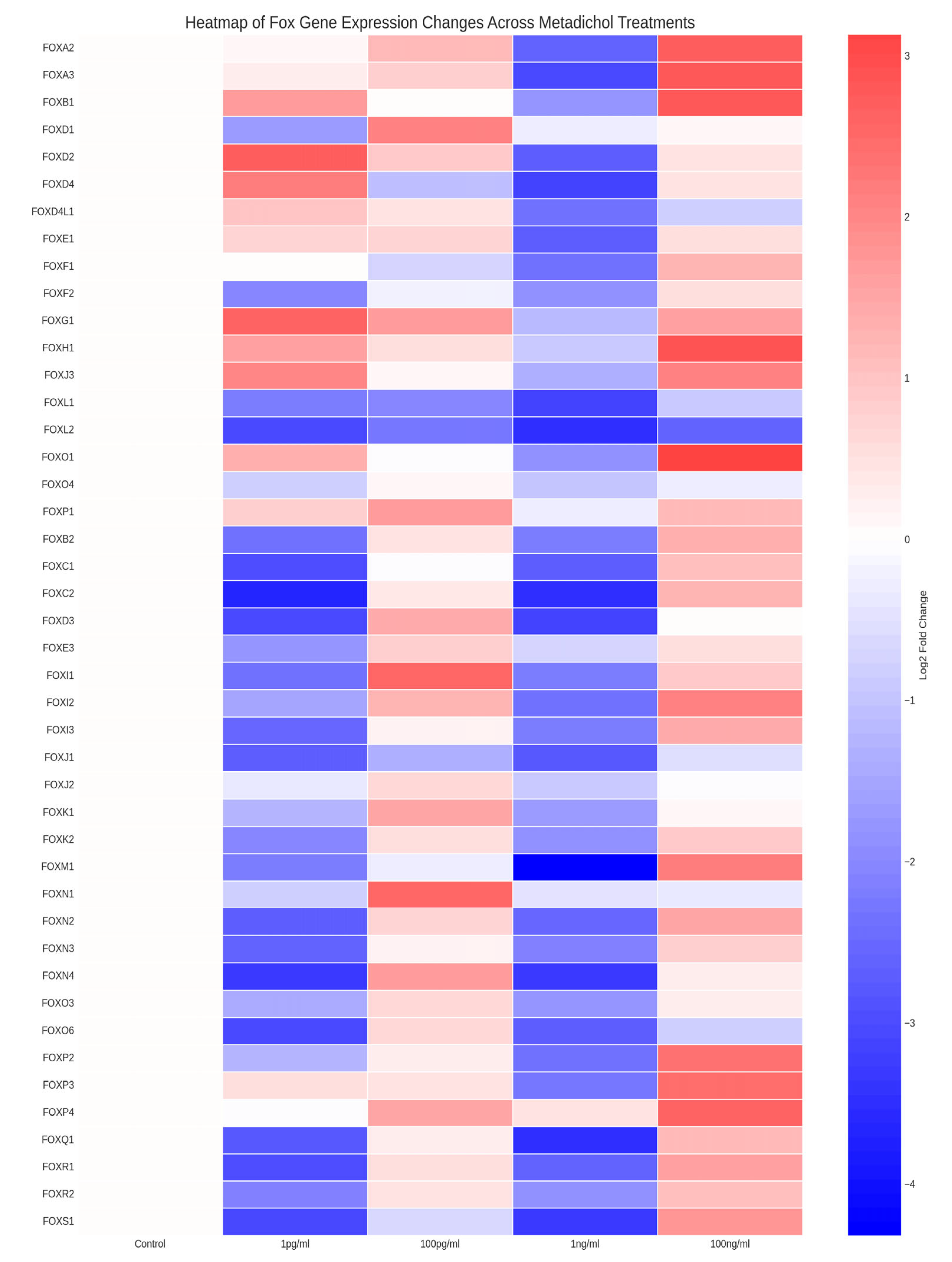
Figure 2.
Bar plot showing the top 10 most upregulated FOX genes in PBMCs treated with 100 ng/ml metadichol compared with the control.
Figure 2.
Bar plot showing the top 10 most upregulated FOX genes in PBMCs treated with 100 ng/ml metadichol compared with the control.
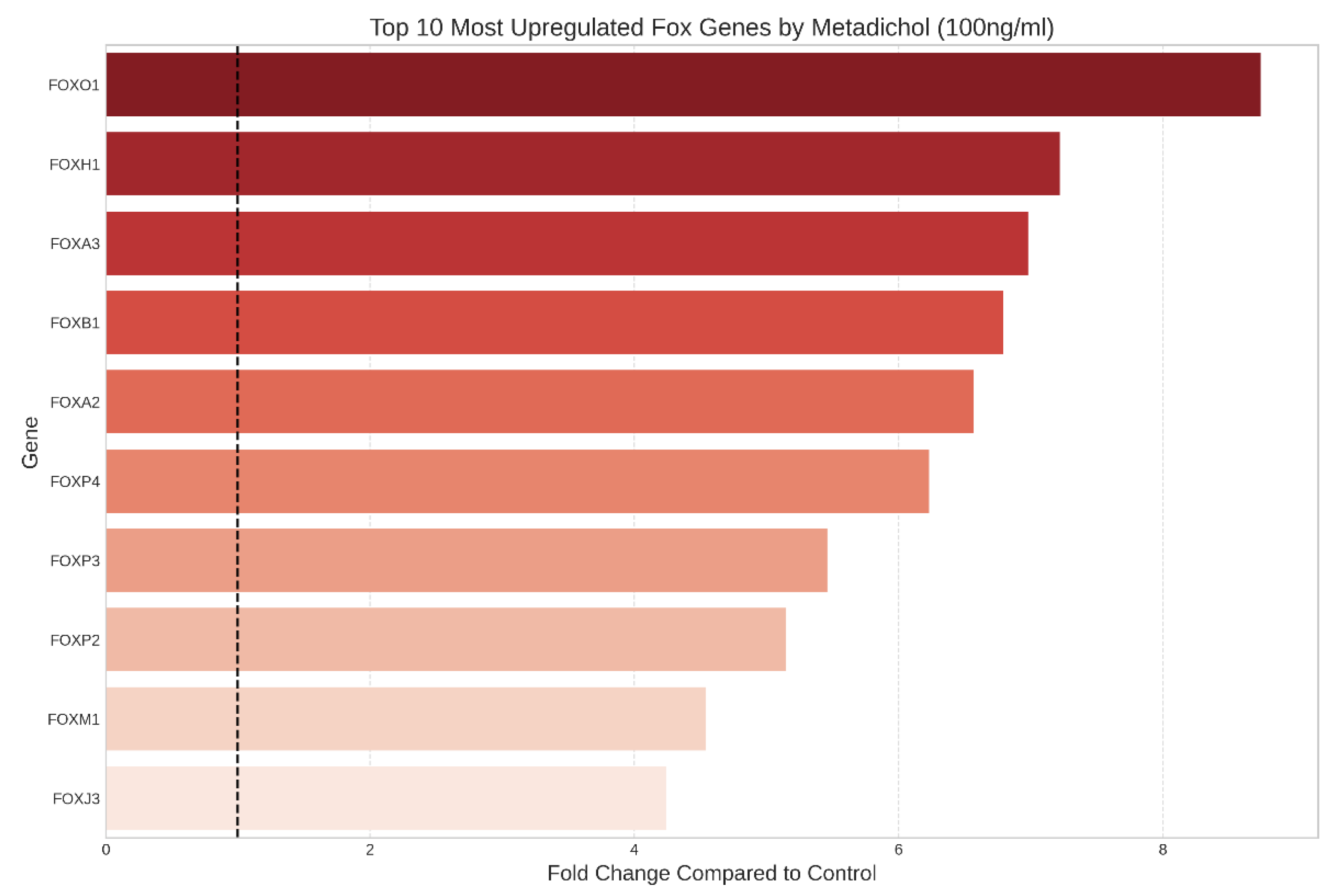
Figure 5.
Dendrogram showing hierarchical clustering of FOX genes on the basis of their expression patterns across different metadichol concentrations.
Figure 5.
Dendrogram showing hierarchical clustering of FOX genes on the basis of their expression patterns across different metadichol concentrations.
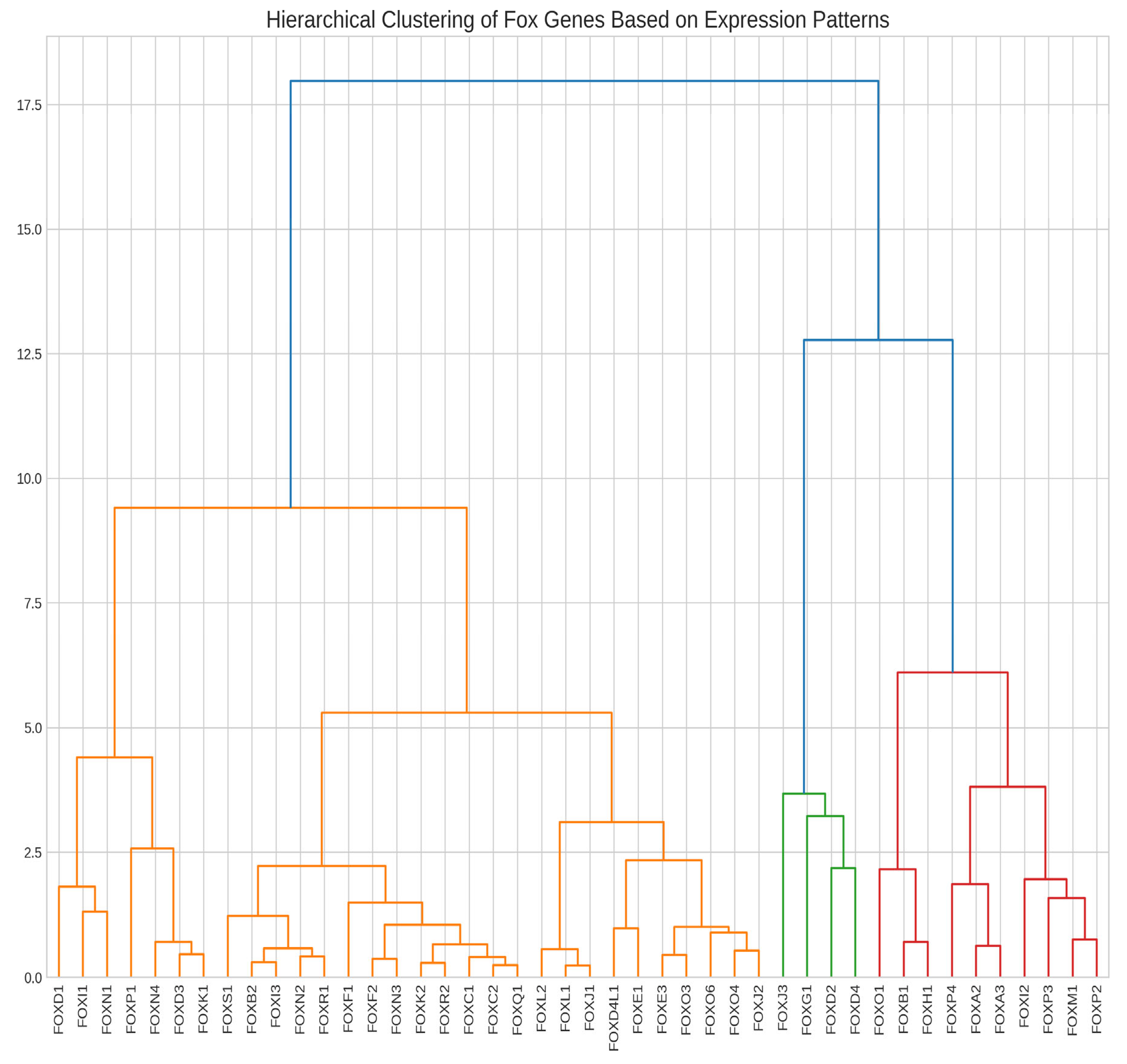
Figure 6.

Figure 7.
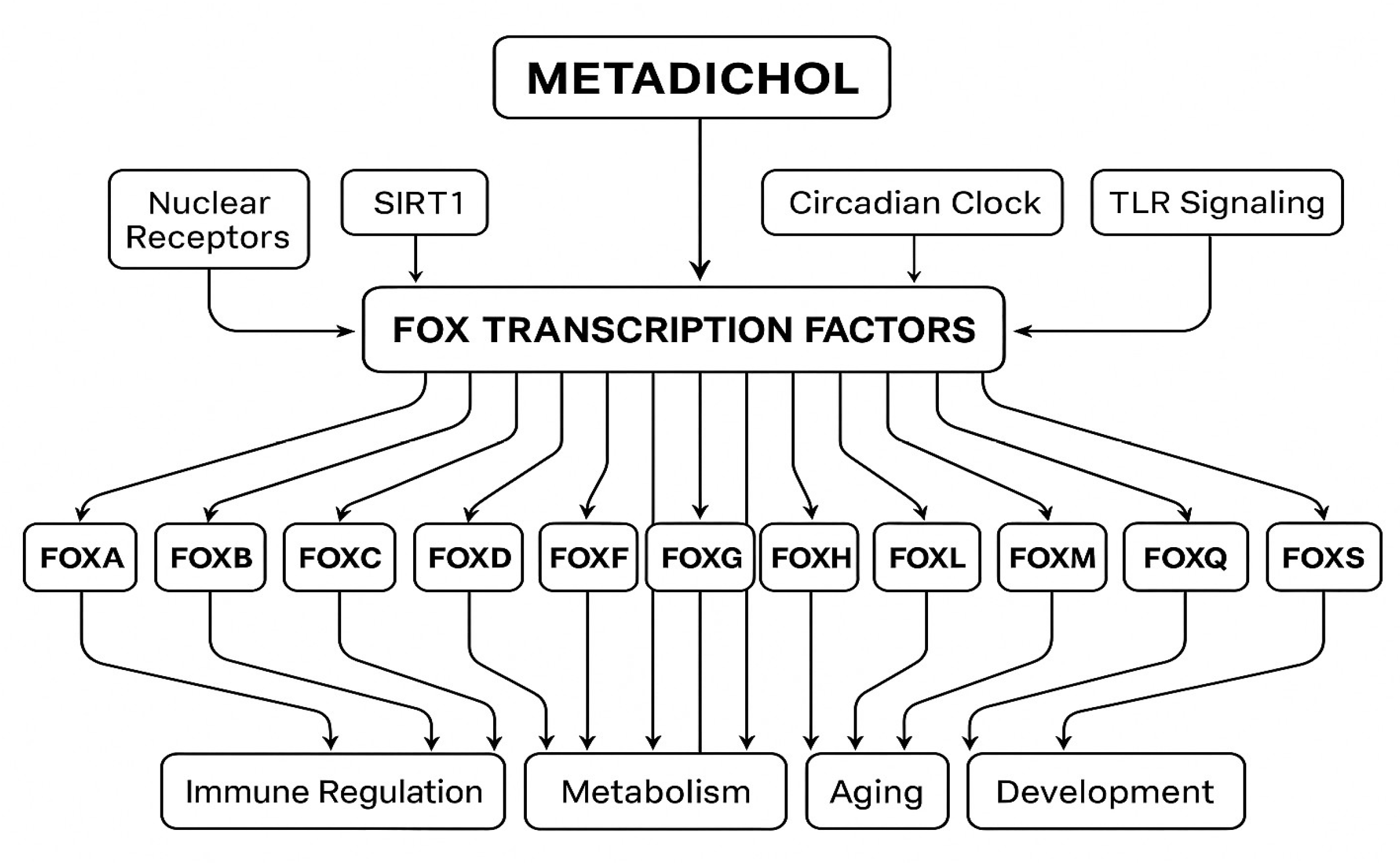
Figure 8.
VDR activated FOX genes.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



