Submitted:
18 August 2025
Posted:
20 August 2025
Read the latest preprint version here
Abstract
Fungi and oomycetes have four large intracellular membrane systems consisting of tubes, lamellae, or a combination thereof. Such elongated membrane structures can extend for tens of micrometers in hyphal cells, branching and connecting into bundles and networks. These systems are represented by tubular vacuoles, elongated mitochondria, endoplasmic reticulum, and macroinvaginations of the plasma membrane. They are most developed in xylotrophic and mycorrhiza-forming basidiomycetes. This review examines three membrane systems, excluding the macroinvagination system. It proposes that, at least in apical hyphae, tubular vacuoles, elongated mitochondria, and the endoplasmic reticulum form a single intrahyphal complex with shared physiological functions. The main function is the transport of dissolved substances, independent of intrahyphal mass flow. In basidiomycetes, such a complex plays an important role, ensuring the independence of resource exchange or accumulation with the host plant or dead wood from the growth rate of apical hyphae and variable turgor gradients within them.
Keywords:
basidiomycetes
; tubular vacuoles
; super-elongated mitochondria
; endoplasmic reticulum
1. Introduction
The elongated form of fungal hyphae suggests that certain intracellular structures can also be extended. For example, tubulin microtubules or actin cables, which perform force-support, transport, and cytokinetic functions, can extend along the hyphae for considerable distances (Berepiki et al. 2011; Bera, Gupta 2022; Mazheika, Kamzolkina 2025). Many fungal intracellular membrane structures have an elongated shape (e.g., endocytic tubes like FEME, Golgi cisternae, Rab6-dependent retrograde transport tubes from the Golgi to the endoplasmic reticulum, and others; Rossanese et al. 1999; Hinze, Boucrot 2018; Dornan, Simpson 2023). However, their sizes usually do not exceed several micrometers. Of particular interest are large-scale membrane structures that have the form of tubes or lamellae (plates, shields, flattened cisterns) with lumens, or that combine both tubes and lamellae, and which, by themselves or when combined into bundles and networks, are capable of reaching tens of micrometers in length. They can be metaphorically spoken of as hyphae within hypha. Four such tubular and tubular-lamellar membrane systems are known in fungi and fungus-like organisms. These include a system of tubular vacuoles, a mitochondrial system, an endoplasmic reticulum, and a system of macroinvaginations of the plasma membrane. Tubular vacuoles and mitochondria consist primarily of tubes of varying diameter, length, and branching. The other two systems may contain both tubes and lamellae. All systems, except for the cell membrane’s macroinvagination system, are endomembrane. The macroinvagination system is not considered endomembrane because it consists of large, simple or complex invaginations of the plasma membrane into the fungal cell (Mazheika et al. 2022; Mazheika, Kamzolkina 2025). This review will not address the macroinvagination system due to its non-endomembrane origin and the limited overlap of its physiological functions with those of other tubular and tubular-lamellar systems. On the contrary, the presence of common functions and coupled work, not merely the morphological similarity of the tubular vacuolar, mitochondrial, and endoplasmic reticulum systems, led to the joining of all three systems into a single review paper. This work’s purpose extends beyond describing large-scale tubular and tubular-lamellar systems in fungi and oomycetes; it proposes the concept of integrating these systems into a single intrahyphal structural-physiological complex. The greatest development, complexity, and extent of tubular vacuoles and, especially, mitochondria are achieved in xylotrophic and mycorrhiza-forming basidiomycetes. Therefore, basidiomycetes are the focus of attention in this review.
2. Tubular Vacuolar System
At the morphological-cytochemical level, tubular vacuoles are the most studied among the three tubular and tubular-lamellar fungal systems. As early as 1941, the renowned French researcher Alexandre Guilliermond studied in detail the reticulum of thin tubular vacuoles in the apical cells of the oomycete Saprolegnia (Guilliermond 1941). Later, Armentrout and colleagues demonstrated, using DIC microscopy, the presence of reticulate tubular vacuoles in the zygomycete Mycotypha microspora (Armentrout et al. 1968). The greatest contribution to the study of tubular vacuoles in fungi was made by the research group led by Professor Anne E. Ashford. Using fluorescent probes, they studied in detail the morphology, dynamics, and topology of the vacuoles of the gasteromycete Pisolithus tinctorius and screened other filamentous fungi and oomycetes (Shepherd et al., 1993a, b; Rees et al. 1994; Rost et al. 1995; Ashford 1997, 1998; Cole et al. 1998; Allaway, Ashford 2001; Hyde et al. 2002; Darrah et al. 2006; Tuszynska et al. 2006; Hyde, Allaway et al. 2007; Zhuang et al. 2009). In collaboration with other researchers, they studied the vacuolar system of the basidiomycete xylotroph Phanerochaete velutina (Darrah et al. 2006; Fricker et al. 2008; Zhuang et al. 2009).
At present, the tubular vacuolar system has been described in numerous species of fungi and fungus-like organisms from various taxonomic groups (Inselman et al. 1999; Ohneda et al. 2002; Uetake et al. 2002; Weber 2002; Lilje, Lilje 2006; Saito et al. 2006; Shoji et al. 2006a, b; Tuszyńska 2006; Bowman et al. 2009; Richards et al. 2010; Shoji, Craven. 2011; Shoji et al. 2014; Funamoto et al. 2015; Higuchi 2021; Groth et al. 2022). This allows concluding that tubular vacuoles are an attribute of most filamentous organisms, from oomycetes to basidiomycetes.
2.1. Methods of Labeling and Microscopic Visualization of the Fungal Vacuolar System
Guilliermond (1941) used the chromophore probe Neutral Red to label vacuoles, including tubular ones, in Saprolegnia. Weber et al. (2001, 2002) also used Neutral Red to study the tubular and spherical vacuoles of Botrytis cinerea and Magnaporthe grisea. Fluorescent probes are widely used to label the vacuoles of fungi and oomycetes. The most commonly used fluorescents are those based on fluorescein in diacetate form (FDA). The diacetate group prevents fluorescence, providing a low fluorescent background and allowing the probe to enter the cells. Intracellular esterases deacetylate the probe, preventing its rapid release from the cell and triggering fluorescence in the green light spectrum (Ashford et al., 2001). In the vacuole lumen, probes accumulate differently depending on the probe modification. It is assumed that 5-chloromethylfluorescein diacetate (CMFDA), due to its chloromethyl group, forms bonds with glutathione and is pumped into the vacuoles by a carrier of glutathione derivatives. 6-Carboxyfluorescein diacetate (CFDA) and Oregon Green 488 carboxylic acid diacetate (carboxy-DFFDA) enter the vacuoles with the help of anion transporters due to their negatively charged carboxyl group (Ashford et al., 2001). The power and stability of the fluorescent signal from these probes are likely influenced to some extent by the pH of the vacuole lumen, which is most often below seven due to the activity of the H+ ATPase in the tonoplast (Rost et al. 1995; Weber 2002; Vesel et al. 2008; Richards et al. 2010; Funamoto et al. 2015). On one hand, the positive charge of the lumen facilitates the influx and retention of the negatively charged probe; on the other hand, the fluorescence of some fluorescein derivatives decreases with decreasing pH (Marchetti et al. 2009). This is why many researchers prefer carboxy-DFFDA over CFDA, as it not only fades more slowly under the microscope’s light beam but also has fluorescence that is less dependent on pH (Cole et al., 1997; Cole et al., 1998; Ashford et al., 2001; Hickey et al. 2004; Hickey, Read 2009).
It has been shown that in slow-growing fungi with a more hydrophobic hyphal surface, labeling with fluorescein probes is more efficient and stable than in fast-growing fungi (Rees et al. 1994). According to our observations, 200 nM CFDA without washing, when the specimen is mounted in a liquid nutrient medium for mycelial growth, provides stable labeling that persists in the dark for tens of minutes in the slower-growing humus coprotroph basidiomycete Coprinus cinereus. In contrast, for labeling vacuoles of the faster-growing xylotroph basidiomycete Stereum hirsutum, a higher concentration of CFDA is more suitable, and photographs should be taken as quickly as possible. After 10-20 minutes, CFDA will begin to leave the vacuoles for the cytoplasm.
The coumarin probe 7-amino-4-chloromethylcoumarin (CMAC) is also used to label the vacuolar system of fungi (Cole et al., 1997; Ashford et al., 2001; Richards et al. 2012). This probe is well-suited for labeling vacuoles in model yeasts, where vacuoles can appear as short tubes, and in filamentous ascomycete fungi (Shoji et al. 2006a; Ohneda et al. 2002). According to our observations, in the hyphae of xylotrophic basidiomycetes, some intracellular structures and inclusions (such as lipid droplets and possibly some vacuoles) exhibit strong blue light autofluorescence. This may complicate CMAC’s use. Other fluorescent probes and methods may be applicable to label fungal vacuoles (Butt et al. 1989; Inselman et al. 1999; Richards et al. 2012).
In TEM samples, tubular vacuoles in fungi are rarely found (Shepherd et al. 1993a, b; Rees et al. 1994; Lilje, Lilje 2006; Saito et al. 2006; Allaway et al. 2007). Although, Rees et al. (1994) cite eight papers from the 1970s to the 1990s that provide TEM images of long vacuoles. The difficulty of studying tubular vacuoles at the ultrastructural level is due to their high mobility, localization in a limited area of apical hyphae, and their tubularity (it is always challenging to identify a tube in a single focal section). In addition, Ashford and colleagues believe that tubular vacuoles do not withstand chemical fixation, fragment rapidly, and can only be studied using freeze-substitution methods (Orlovich, Ashford 1993; Rees et al. 1994; Ashford et al., 2001).
The modern standard for fluorescent labeling of intracellular structures is genetic labeling, which includes the expression of FP (fluorescent protein) in the same ORF as the target protein. Fungal vacuoles are visualized through genetic labeling using various vacuolar target proteins: CPY in Aspergillus nidulans and A. oryzae (Ohneda et al. 2002; Hickey, Read 2009; Higuchi 2021); AoVam3p in A. oryzae (Shoji et al. 2006a, b; Higuchi 2021); VAM-3 and VMA-1 in Neurospora crassa (Bowman et al. 2009); Vac14, Vam1, and Rab7 in Sordaria macrospora (Groth et al. 2022), and others. However, genetic vacuolar labeling, especially for basidiomycetes, is still less commonly used than cytochemical labeling.
2.2. Morphotypes of the Tubular Vacuolar System and Their Variants
Various morphotypes of the tubular vacuolar system exist in the apical hyphae of fungi and fungus-like organisms. Each morphotype may correspond to a large taxonomic or ecotrophic group, or, most likely due to limited data, be an attribute of a distinct fungal genus or species. Figure 1 shows examples of basic variants of four morphotypes. The basic variant is the most characteristic or frequently encountered morphological variant. In addition to the basic variant, in most cases, different variants of the same vacuolar morphotype can be found in the apical hyphae, even in the same colony (see, for example, the variants of morphotype P; Figure 2). Additionally, within one or similar morphotypes, the tubular vacuolar system (on average, tube length, number, and cellular volume occupied) may vary among different representatives. For instance, our observations indicate that among three basidiomycetes from diverse ecotrophic groups – Rhizoctonia solani, C. comatus, and S. hirsutum – the tubular vacuolar system is most prominent in the xylotroph S. hirsutum. In the other two fungi, it is less pronounced (Figure 3a-c), though individual apical hyphae may exhibit comparable development to S. hirsutum (Figure 3d and Figure S1). It can be assumed that among basidiomycetes, the tubular vacuolar system reaches its greatest development in mycorrhiza-forming soil basidiomycetes and wood-decaying fungi (Shepherd et al. 1993b; Rees et al. 1994; Hyde et al. 2002; Darrah et al. 2006; Zhuang et al. 2009). Variability also arises under the influence of external conditions. For example, Hyde and Ashford (1997) indicate that the addition of fresh nutrient medium increases the number and motility of tubular vacuoles in P. tinctorius. There is evidence that tubular vacuoles in A. oryzae are most developed in hyphae isolated from the nutrient substrate, such as hyphae growing on the wall of a Petri dish (Shoji et al. 2006a, b). Well-developed tubular vacuoles are often seen in bent hyphae at the bending point (Figure S2; Video S1).
The diversity of vacuolar morphotypes in fungi and fungus-like organisms is not limited to the four morphotypes described herein. As knowledge accumulates, other morphotypes and their variations will likely be described, and the four morphotypes presented here will expand the lists of species containing them.
Morphotype S (Saprolegnia-type). In oomycetes, the vacuolar system in the apical domain of hyphae is a low-mobility reticulum of frequently branching vacuolar tubes. These tubes fill the entire cellular space and are closely adjacent to the plasma membrane. Tens of microns from the apex, the tubular reticulum gradually transforms into larger vacuoles, which can then merge into a single vacuolar canal running along the center of the hypha (Figure 1a; Guilliermond 1941; Rees et al. 1994; Lilje, Lilje 2006; Allaway et al. 2007). Non-tubular vacuoles are rarely smooth, spherical, or oval; they are more often amorphous. In variants differing from the basic morphotype, the transition from a tubular reticulum to larger vacuoles can occur at varying distances from the apex (Guilliermond 1941). Armentrout et al. (1968) argue that the zygomycete Mycotypha microspora has a vacuolar system in its apical hyphae similar to that of oomycetes.
Morphotype G (Gigaspora-type). This morphotype of the vacuolar system has been described for glomeromycete Gigaspora margarita (Ashford 2002; Uetake et al. 2002; Funamoto et al. 2015). In some hyphae of this fungus, the tubular system of vacuoles resembles the vacuolar reticulum of oomycetes, but the tubules are thinner and more intricately entangled. In other hyphae, thin tubular vacuoles are combined into bundles that fill the space of the hypha and run along its long axis. Bundles of tubular vacuoles appear to replace solid vacuolar canals in oomycetes. Spherical vacuoles in G. margarita are usually small and rarely connected to tubular vacuoles (Uetake et al. 2002; Funamoto et al. 2015).
Morphotype P (Pisolithus-type). Ashford’s group proposed a base scheme for the topology of vacuoles in the hyphae of P. tinctorius (Hyde et al. 2002) that has become canonical because, with some modifications, it is suitable for many ascomycetes and basidiomycetes, and some studied zygomycetes (Figure 1c and Figure 3e; Video S2; Armentrout et al. 1968; Rees et al. 1994; Ashford 1997; Hyde, Darrah et al. 2006; Tuszynska et al. 2006; Hickey, Read 2009; Zhuang et al. 2009). When a new branch of a hypha is formed, all the vacuolar zones of the mother hypha are usually replicated in it (Darrah et al. 2006). According to the canonical scheme, there are no vacuoles at the very tip of the apical cell. Following this, there is a zone approximately 5-50 µm from the tip containing very small spherical vacuoles and short vacuole tubes. Here and further up to the zone of spherical vacuoles, PVCs (prevacuolar compartments) are found. They are described in P. tinctorius, P. velutina, A. oryzae, Neurospora crassa, and budding yeasts (Ohneda et al. 2002; Darrah et al. 2006; Shoji et al. 2006b; Zhuang et al. 2009; Bowman et al. 2015; Day et al. 2018; Bowman 2023). They primarily attracted attention as ring-like structures, but they can also take the form of semi-rings, wavy tubes, and Y-tubes on a single focal section, changing shape and joining together. According to some data, PVCs can migrate between the tip of the hypha and subapical regions (Zhuang et al. 2009). Other data suggest they reside in the subapical zone and receive vesicles from the apex and tubular vacuoles, functioning as vesicle-sorting centers (Bowman et al. 2015; Bowman 2023). PVCs have been studied most thoroughly in N. crassa. They appear to be plates that often close into a sac-like structure, hence the ring shape seen in regular photographs (Bowman et al. 2015; Bowman 2023). It is believed that the PVC plate is not a membranous flattened cistern but consists of vesicles joined together. PVCs carry many vacuolar markers, including Rab7, but they lack some vacuolar lumen proteins (Bowman et al. 2015; Bowman 2023). Probably, PVCs are structurally and physiologically closest to late endosomes, specific to the apical cells of fungi, and carry out specific functions.
After the zone of very small vacuoles and PVCs, there is a zone of tubular vacuoles (approximately 50-200 µm from the apex, but within the apical cell). In this zone, small spherical vacuoles are present, but tubular vacuoles dominate to varying degrees (Figure 1c; Figure 3e; Video S3). Tubular vacuoles can be well-developed, long, and branching. They can unite into powerful networks, but these are not as dense as those of oomycetes. In addition, they are extremely dynamic here – constantly lengthening, shortening, moving, branching, merging, and fragmenting (Videos S4-6; Rees et al. 1994; Hyde, Ashford 1997; Tuszynska et al. 2006). Several authors note the peristaltic movement of the tonoplast in tubular vacuoles (Shepherd et al. 1993b; Rees et al. 1994; Cole et al., 1998; Weber 2002; Tuszynska et al. 2006). Although other works deny it (Darrah et al. 2006). Tubular vacuoles can enter the clamp connections in basidiomycetes (Video S7). In the basal part of the apical cell, as well as in the subapical and several subsequent cells, spherical vacuoles begin to dominate (Video S8; Boenisch et al. 2017). Their size increases with distance from the apical cell, while their motility decreases. In several cells following the apical one, spherical vacuoles actively form outgrowths, move with the help of tubular outgrowths, or relocate without changing shape (Figure 3f; Videos S9-11). They connect with each other through tubular outgrowths (Figure 3g; Video S12). In contrast, large spherical vacuoles in the basal cells of the hyphae are typically slightly motile and do not form tubular outgrowths (Figure 3h; Video S13; Rees et al. 1994; Hyde, Ashford 1997; Hyde et al. 2002; Tuszynska et al. 2006; Hickey, Read 2009; Zhuang et al. 2009).
The tubularity of fungal vacuoles can be expressed in different ways. There are real vacuole tubes; they are relatively thin, more or less uniform in thickness, and round in cross-section (Figure 3a-e; Video S14). Such tubular vacuoles usually occupy the middle zone of the apical hyphal cell. There are vacuoles, which are essentially elongated spherical vacuoles or their outgrowths. They are thicker than true tubular vacuoles, uneven in thickness, and can be somewhat flattened (Figure 3f,g; Videos S15-17).
Morphotype R (Rhizoctonia-type). According to our observations, in R. solani, CFDA usually marks only weakly developed tubular vacuoles and small spherical vacuoles. Although there are also rare hyphae with a network of tubular vacuoles (Figure 3d and Figure S1). R. solani does not have large spherical vacuoles that occupy most of the cell on standard nutrient media (Figure S3; Video S18). At the same time, R. solani has another feature, along with the absence of large spherical vacuoles, which determines the original vacuolar morphotype of this fungus. In the subapex of some apical cell of R. solani, a dense vacuolar spindle is localized, accumulating CFDA, from which short dynamic tubular vacuoles emanate both towards the tip of the hypha and basipitally (Figure 3i and Figure S4; ).
Figure 2a-f shows different morphotype P variants in S. hirsutum. Zhuang et al. (2009) show some of these variants in P. velutina and suggest a relationship between hyphal growth rate and morphotype variant. We support this hypothesis and believe that, in some cases, the development of the tubular vacuole system in S. hirsutum is directly related to the hyphal growth rate. In a rapidly growing hypha, the apical cell elongates because the formation of clamp connections and septa lags behind the rapid growth of the hyphal tip. Such a long cell, from the subapex to the basal part, is filled with a well-developed network of tubular vacuoles (Figure 2a and Figure 3a). On VideoTrack S1, it is visible that developed tubular vacuoles, starting near the apex, occupy almost 1 mm along the hypha. Spherical vacuoles begin to appear approximately 800 µm from the tip of the hypha, whereas in the canonical version, they appear after about 200 µm and are practically immotile after 600 µm (Hyde, Ashford 1997; Hyde et al. 2002; Hickey, Read 2009). Individual tubular vacuoles can quickly stretch out to reach the apical body zone and be pulled back, which deviates from the basic variant (Figure 2g, left; Figure 3a; Videos S20-22). A similar phenomenon is described in the work of Zhuang et al. (2009) for P. velutina. In R. solani, the contact of tubular vacuoles with the hyphal tip may occur differently (Figure 2g, right). Video S23 shows how a spherical vacuole located several tens of microns from the apex forms an outgrowth reaching the hyphal apex (but not the very tip here).
In slow-growing hyphae of S. hirsutum, the tubular vacuole system in the apical cell is replaced by a system of small, spherical vacuoles (Figure 2c and Figure 3j). Zhuang et al. (2009) indicate that the apical cell of P. velutina is also filled with clusters of slow-moving, small, spherical vacuoles during slow hyphal growth. It can be assumed that the hyphae, the apical cells of which contain relatively large, spherical vacuoles distributed throughout the cell and lack tubular vacuoles, have completely stopped growing and have been in this state for some time (Figure 2d and Figure 3k). Such a cell in C. comatus can be seen in Figure S5. Tuszynska et al. (2006) showed that in Paxillus involutus, under the influence of zinc sulfate, apical cells lose tubular vacuoles and contain large and small spherical ones.
In microscopic samples, many apical hyphae exhibit no vacuolar signal (Figure 2e). However, it remains unclear whether these hyphae lack vacuoles or if the signal’s absence is due to low label accumulation within the vacuolar lumen. This could result from various factors, including low cell permeability, robust pumping of the probe out of the cell, low esterase activity, extreme vacuolar lumen pH (high or low), or low tonoplast permeability.
Inverted hyphae, as we have called them, are often observed in S. hirsutum (Figure 2f and Figure 3l). The apical cells of these hyphae contain spherical, sometimes large, vacuoles in the subapical zone, while the more basal part contains usually a robust network of tubular vacuoles (Figures S6 and S7). Tubular vacuoles in inverted hyphae can be localized quite far from the apex (Figure S8), as well as being poorly developed (Figures S8 and S9).
Other vacuolar morphotypes also have variants that do not coincide with the basic one. For example, morphotype S can transition from variants with a developed tubular reticulum extending far from the apex to the basal part, to variants with amorphous non-tubular vacuoles filling the entire apical part of the hypha (see illustrations in Guilliermond 1941; Rees et al., 1994; Allaway et al., 1997).
2.3. Molecular Mechanisms of Vacuole Morphogenesis, Influence of the Cytoskeleton on the Vacuolar System
The molecular mechanisms that ensure the dynamics of the vacuolar system are relatively well-studied in S. cerevisiae and other model yeasts (Richards et al. 2010; Chen et al. 2020; Gokbayrak et al. 2022). Various stimuli, such as osmotic and oxidative stress, endoplasmic reticulum stress, environmental acidification with acetate, TOR signaling, autophagy, and others, either activate Fab1 kinase or Fis4 phosphatase, altering the level of PI(3,5)P2 or act through Rab signaling (Gokbayrak et al. 2022). As a result, either the dynamin-like GTPases Vps1 or Dnm1 is activated, leading to vacuole fragmentation or tubulation. Conversely, homotypic fusion of vacuoles into larger ones occurs with the participation of HOPS and other factors (Richards et al. 2010). The pH level inside the vacuoles is important for the fusion and fragmentation process.
The mechanisms of fusion, fragmentation, and tubulation of vacuoles in filamentous fungi are poorly understood. They are assumed to be similar to the mechanisms in yeast. In A. nidulans, the participation of Rab7, HOPs, and Vps1 homologues in vacuolar dynamics has been demonstrated (Tarutani et al. 2001; Ohsumi et al. 2002; Oka et al. 2004; Richards et al. 2010). In P. tinctorius, treatment of mycelium with GTPγ-S (a GTP analogue that inhibits GTPases – when GTPγ-S is bound to dynamin, it irreversibly contracts around the neck of the membrane tube, causing permanent tubulation; Takei et al. 1995) results in the formation of tubular vacuoles in the first five cells from the apex (Hyde et al. 2002). This confirms the involvement of dynamin-like proteins in forming tubular vacuoles. GTPy-S also impacts the oomycete S. felax: the tubes of the vacuolar reticulum become thicker, and the reticulum spreads further from the apex, replacing the spherical vacuoles (Lilje, Lilje 2006).
In model ascomycete yeasts, actin is responsible for the movement and morphology of vacuoles (Richards et al. 2010). In filamentous fungi, microtubules probably play a major role here (Ashford 1998; Darrah et al. 2006; Tuszyńska 2006; Richards et al. 2010). For P. tinctorius, nocodazole (a microtubule assembly inhibitor) has been shown to disrupt tubular vacuoles and tubular outgrowths from spherical vacuoles, and many ultrastructural studies have demonstrated that microtubules run along long vacuoles (Ashford 1998; Hyde et al. 1999; Roberson et al. 2010). Conversely, evidence suggests that F-actin depolymerization in some glomeromycetes disrupts dissolved substance transport through hyphae. However, the disruption mechanism remains unknown. (Ashford 2002).
2.4. Physiological Functions of the Vacuolar System of Fungi
2.4.1. General Functions of Different Types of Vacuoles
Fungal vacuoles perform various functions (Weber 2002; Lilje, Lilje 2006; Shoji et al. 2006a; Veses et al. 2008; Richards et al. 2010; Roberson et al. 2010; Shoji et al. 2014; Chen et al. 2020). First, the hypothesis of “cytoplasmic economy” is interesting, which refers to large spherical vacuoles or vacuolar canals (Richards et al. 2010). In mature thick hyphae, which may not carry trunk transport functions and are low-active, as well as in search hyphae, hyphae of starving mycelium, some germ tubes, and other, large spherical vacuoles often fill almost the entire cell space. These vacuoles replace the cytoplasm, which is either unnecessary in such quantity or lacks sufficient resources for its creation.
Another important property of vacuoles is the concentration of substances. Proton V-ATPases create a potential difference across the vacuole membrane, acidifying the lumen. The energy from this potential difference is used to pump ions and various substances into the vacuole (Rost et al. 1995; Weber 2002; Veses et al. 2008; Richards et al. 2010; Funamoto et al. 2015). Vacuoles, especially spherical ones, contain negatively charged polyphosphates that act as cation traps, while tubular ones have fewer (Ashford 1998; Weber 2002; Saito et al. 2006; Funamoto et al. 2015). Vacuoles use the ability to concentrate substances for various purposes. Firstly, for the reservation of substances. It is known that polyphosphate traps bind amino acids, primarily arginine. In this form, fungi store nitrogen. Polyphosphates also trap and store ions of salts and various metabolites (Venables, Watkinson 1989; Griffin 1994; Ashford 1998; Cole et al. 1998; Weber 2002; Watkinson et al. 2005). Secondly, vacuoles act as cytoplasm detoxifiers: by pumping heavy metal ions and Ca2+ into the lumen and binding them, vacuoles not only store these substances but also reduce their toxic effects on the cell (Weber 2002). Thirdly, by binding osmotically active substances to lumen polyphosphates, vacuoles deprive them of osmotic activity, which is one of the mechanisms for regulating turgor pressure in the cell. Vacuoles are involved in the regulation of cellular osmosis in various ways. Rapid fragmentation of vacuoles under hyperosmotic conditions or fusion in a hypotonic environment (Richards et al. 2010), as well as in sudden cell swelling, such as in the trapping rings of Drechslerella dactyloides (Chen et al., 2022), is a mechanism for regulating the sudden entry or exit of water into or from the cell. The volume of a large vacuole, and therefore its capacity for water, is greater than the sum of the volumes of small vacuoles in the absence of de novo vacuole membrane synthesis. It is also suggested that vacuoles, where glycerol and other osmolytes are synthesized, are involved in creating powerful turgor in the appressoria of M. oryzae or the sporangia of Basidiobolus ranarum (Weber 2002).
Vacuoles can participate in the synthesis of some secondary metabolites (Richards et al. 2010; Shoji et al. 2014). Another important function of fungal vacuoles is their role in the degradation of cellular structures and biological molecules entering from the endocytic pathway, Golgi cisternae, through autophagy and other pathways. Such vacuoles are called vacuole-lysosomes (Ashford 1998; Richards et al. 2010; Shoji et al. 2014).
Finally, vacuoles, particularly tubular vacuoles (discussed in the next section), participate in the compartmentalized transport of dissolved substances in fungal hyphae (Cox et al. 1980; Ashford 1998; Allaway, Ashford 2001; Ashford 2002; Weber 2002; Watkinson et al. 2005; Darrah et al. 2006; Lilje, Lilje 2006; Shoji et al. 2006a; Fricker et al. 2008; Zhuang et al. 2009; Shoji, Craven. 2011; Funamoto et al. 2015).
Weber (2002) proposed a hypothesis regarding the functions of fungal vacuoles: in a fungal cell, different vacuoles can perform distinct functions. This hypothesis addresses the issue Ashford (1998) discussed: how vacuoles can simultaneously manage such diverse and incompatible functions as lysis, storage, and transport of substances. The results of our work can confirm these hypotheses (Mazheika et al. 2022). When the cell membrane and endocytic pathway were labeled with AM4-64 (analogous to FM4-64) and vacuoles with CFDA in S. hirsutum, it was found that the tonoplasts of only a small proportion of spherical vacuoles included the AM4-64 label. This indicates that, at least in a complete nutrient medium, only a small proportion of vacuoles function as vacuole-lysosomes, while the remaining spherical vacuoles perform other functions.
2.4.2. Proposed Functions of Tubular Vacuoles
Tubular vacuoles can perform functions common to all vacuoles, but they are most often assigned a transport function (Ashford 1998; Ashford 2002; Watkinson et al. 2005; Darrah et al. 2006; Zhuang et al. 2009; Abadeh, Lew, 2013; Funamoto et al. 2015;). The tubular vacuolar system can rapidly, and perhaps selectively, transport dissolved substances along the cell or hyphae simultaneously in different directions. It has been shown that tubular vacuoles can pass through septa (Shepherd et al. 1993a, b), ensuring continuous intercellular transport in the hypha. This transport, due to membrane compartmentalization, is not dependent on the mass flow and cytoplasmic streaming of the hyphae. It is probably not as energy-consuming as the active transport of substances in vesicles along the cytoskeleton elements. It does not require mandatory membrane movement within the hyphae (Ashford 1998; Cole et al. 1998, Ashford 2002; Fricker et al. 2008). Ashford (1998) suggests that substances are transported in tubular vacuoles in complex with polyphosphates. She and her colleagues believe that the electron-dense polyphosphate inclusions familiar in TEM samples are artifacts of the preparation of microscopic specimens (Orlovich, Ashford, 1993). Under native conditions, polyphosphates do not aggregate into granules. Ashford and other authors suggest that the intrahyphal transport of substances through tubular vacuoles may be accomplished by the peristaltic movement of their membranes, ensuring high-speed transport (Shepherd et al. 1993a, b; Rees et al. 1994; Weber 2002; Tuszynska et al. 2006). On the other hand, studies on P. velutipes showed that the spread of the fluorescent probe in tubular vacuoles occurs only at the rate of diffusion in an aqueous environment (Darrah et al. 2006; Fricker et al. 2008).
Long-distance transport of dissolved substances (sugars, polyols, amino acids, phosphates, potassium, etc.) in cord- and rhizomorph-forming basidiomycetes may occur over hundreds of meters and at high speed (Brownlee, Jennings 1982; Jennings 1987; Venables, Watkinson 1989; Cairney 1992; Timonen et al. 1996; Ashford 1998; Cole et al. 1998; Olsson, Gray 1998; Boddy 1999; Ashford et al. 2001; Weber 2002; Watkinson et al. 2005; Darrah et al. 2006; Fricker et al. 2008; Tlalka et al. 2008; Funamoto et al. 2015; Schmieder et al. 2019; Herman, Bleichrodt. 2022). Such transport occurs simultaneously in different directions, most likely along different conducting cords or distinct bundles of hyphae within a single cord (Granlund et al. 1985; Watkinson et al. 2005; Schmieder et al. 2019), and oscillizes (Tlalka et al. 2003; Watkinson et al. 2005; Fricker et al. 2008; Schmieder et al. 2019). The oscillation of the isotope-labeled, non-metabolizing amino acid analogue (14С-AIB; α-aminoisobutyric acid) in P. velutina occurs with a period of 12-16 hours (Tlalka et al. 2003; Watkinson et al. 2005; Fricker et al. 2008). However, it may be associated not with the pulsation of mass flow or other transport mechanisms, but with the accumulation and release of 14C-AIB in vacuoles (Tlalka et al. 2003). Transport oscillations in non-cord-forming C. cinerea occur in marginal trunk hyphae with a period of 4-6 hours and represents a real oscillation of mass flow with a periodic change in direction. The transport reverses direction asynchronously between adjacent trunk hyphae every 2-3 hours (Schmieder et al. 2019). The modern paradigm attributes the primary role of mass flow to the long-distance transport of fungi, including both cord-forming (Brownlee, Jennings 1982; Thompson et al. 1985; Fricker et al. 2008; Tlalka et al. 2008; Schmieder et al. 2019) and non-cord-forming species (Abadeh, Lew 2013; Schmieder et al. 2019). Mass flow in hyphae is caused by the movement of water due to differences in turgor pressure (Thompson et al. 1985; Fricker et al. 2008; Steinberg et al. 2017; Lew 2019; Schmieder et al. 2019; Herman, Bleichrodt. 2022). The actin cytoskeleton (both as a contractile force and as a transport mechanism) and turbulence from active organelle transport may also contribute to mass flow generation (Brownlee, Jennings 1982; Heath, Steinberg 1999; Reynaga-Peña, Bartnicki-García 2005; Steinberg et al. 2017; Mazheika, Kamzolkina, 2025). Mass flow not only participates in the transport of substances dissolved in the cytoplasm but also provides cytoplasmic streaming – the movement of unattached organelles and particles – which can be considered a special case of long-distance transport of resources (Fricker et al. 2008; Panstruga et al. 2023; Schuster et al. 2025). It has been hypothesized that tubular systems may also participate in long-distance transport, providing rapid, to some extent selective and multidirectional transport of dissolved substances, independent of mass flow. To a lesser extent, such participation was attributed to mitochondria (Zhuang et al. 2009) and, to a greater extent, to tubular vacuoles (Ashford 1998; Allaway, Ashford 2001; Weber 2002; Lilje, Lilje 2006; Fricker et al. 2008; Veses et al. 2008; Zhuang et al. 2009; Shoji, Craven. 2011; Abadeh, Lew, 2013; Funamoto et al. 2015). However, tubular vacuoles have been described only in the apical hyphae of fungal colonies. Our knowledge about fungal vacuoles is limited to a maximum area of several millimeters, located basipetally from the apical cell, and is most often studied in laboratory cultures of fungi. What happens to the vacuolar system beyond these several millimeters, especially in natural conditions, as well as in the hyphae of cords and rhizomorphs, is technically difficult to demonstrate. There have been separate attempts to study vacuoles remote from the apex, experimentally or through modeling, but the issue requires further investigation (Allaway, Ashford 2001; Darrah et al. 2006; Fricker et al. 2008).
Moreover, tubular vacuoles represent an unstable, motile, and rapidly changing system, which is hardly compatible with the rapid and stable transfer of resources over long distances, despite oscillating with a certain period (Tlalka et al. 2003; Watkinson et al. 2005; Fricker et al. 2008; Schmieder et al. 2019). Therefore, it is most likely to assume that tubular vacuoles perform a local transport function in individual parts of the colonies, particularly in the apical hyphae. Some variants of such a local transport function will be proposed below.
(i) It has been suggested that in oomycetes, the apical vacuolar reticulum replaces or supplements the system of secretory vesicles (Lilje, Lilje 2006). In true fungi, these vesicles are delivered to the tip of the hypha first by microtubules, then by actin cables, sorted in the apical body, and deliver material and enzymes to the growing apex (Takeshita et al. 2014; Peñalva et al. 2017; Steinberg et al. 2017; Takeshita 2019). The oomycete vacuolar reticulum can also function as endosomes by merging with endocytic vesicles in the subapex. This hypothesis is supported by the observation that the vacuolar reticulum in oomycetes can fill the entire apical part of the hypha and be nearly adjacent to the cell membrane (Guilliermond 1941; Rees et al. 1994; Lilje, Lilje 2006; Allaway et al. 2007). It has been hypothesized that the tubular vacuolar system may function as endosomes also in true fungi (Ashford, Orlovich, 1995; Ashford 1998; Cole et al. 1998). It is impossible to exclude the presence of this type of apical transport in true fungi, but it is more likely to be of a temporary auxiliary nature. This is confirmed by the facts presented above: in xylotrophic fungi, individual tubular vacuoles are briefly introduced into the very tip of the hypha (Zhuang et al. 2009; Figure 3a; Videos S20-22), or in R. solani, where the outgrowth of a spherical vacuole extends to the tip of the hypha (Video S23). In addition, the vacuolar spindle described above in R. solani (Figure 3i and Figure S4; Video S19) may function as a local pump that moves dissolved substances into and out of the apex.
(ii) The vacuolar system of G. margarita may function similarly to that of oomycetes, as it can fill most of the hypha’s internal space. However, as an endomycorrhizal fungus, G. margarita exchanges large amounts of resources with its host, resulting in a more complex vacuolar system with broader transport functions than that of oomycetes (Smith, Read, 1997; Uetake et al. 2002; Funamoto et al. 2015). Unlike the single vacuolar channel in oomycetes, where the vacuolar reticulum extends basipetally from the apex, G. margarita forms bundles of thin tubular vacuoles. This structural difference ensures rapid, selective, and multidirectional transport of dissolved substances, not only in the apex but also in more basipetal regions of the hypha (Ashford 2002).
(iii) A simple model of apical growth in fungi suggests that in a growing apical hypha, the turgor gradient is created by the elongation of the hyphal tip. The pressure gradient creates a mass flow directed from the basal to the apical part of the hypha, which, by increasing pressure at the tip, contributes to the tip elongation mechanism (Money 2025). Schuster et al. (2025) confirm with modern equipment that in Trichoderma reesei the first three cells from the tip have a small cytoplasmic streaming directed towards the apex. From the fourth cell onwards it is absent. However, given the above-described change in the direction of mass flow in C. cinereus every 2-3 hours (Schmieder et al. 2019) and the strictly regulated, phase-shifted oscillation of calcium influx, F-actin assembly, and turgor pressure shown at the apex (Weber 2002; Takeshita et al. 2017; Takeshita 2019), more complex models of apical growth can be assumed. The pressure in the apical cell, or its apical part, may be created by its own influx of water from the outside. In this case, mass flow directed towards the apex does not reach the hyphal tip (Herman, Bleichrodt 2022). Counter-directed mass flows are even possible, in certain periods, colliding in the subapical zone. In other words, even regardless of the choice of apical growth model, this indicates that an actively growing hypha must possess a local transport system. This system would supply the growing apex and carry out retrograde transport of resources, irrespective of the turgor gradient and the direction or oscillation of mass flow. Schuster et al. (2025) showed that in T. reesei at least five subapical cells perform active transport into the apical cell via the cytoskeleton and vesicles. However, in xylotrophic and mycorrhizal basidiomycetes, this mechanism is insufficient, necessitating a robust system of tubular vacuoles for local transport. This hypothesis aligns with the idea that the development of the tubular vacuolar system directly correlates with the hypha’s growth rate and the apical cell’s length in xylotrophs (Figure 2a; Zhuang et al. 2009).
(iv) Inverted hyphae manifest another local transport function of tubular vacuoles. Large spherical vacuoles in the apical part of the hypha likely indicate autolysis of this hypha. In this scenario, tubular vacuoles powerfully pump resources from the dying hypha without involving mass flow, which could non-selectively pump autolysis factors (signal molecules, enzymes, autophagosomes, etc.) into the still-living parts of the hypha.
(v) Another variant of the local transport function of tubular vacuoles is the reparative-transport function. As noted above, powerful tubular vacuoles can be found in strongly curved, broken hyphae (Figure S2; Video S1). Tubular vacuoles pass through the hyphal fracture, possibly restoring resource transport to the post-fracture hyphal region.
3. Filamentous, Tubular, Network and Super-Elongated Mitochondria
One of the first to describe fungal mitochondria at the beginning of the twentieth century was Guilliermond (1911). Mitochondria are visualized by DIC microscopy, are well-preserved in electron microscopic samples using various specimen preparation methods, and have extensive possibilities for fluorescent labeling. Therefore, it is difficult to claim that fungal mitochondria are poorly studied. However, a striking example of a gap in our knowledge is the super-elongated mitochondria, which represent the most powerful tubular mitochondrial system in fungi and are the focus of this review. Super-elongated mitochondria are characteristic primarily of xylotrophic and mycorrhizal basidiomycetes (Armentrout et al. 1968; Weber et al. 1998; Tuszynska et al. 2006; Tuszyńska 2006; Mazheika et al., 2020), but may also be present in other groups of fungi (Weber et al. 1998; Faoro et al. 2022). It is difficult to determine who first described super-elongated mitochondria in fungi. Hatch (1935) described “long slender chondrioconts” in Allomyces arbusculus, and Guilliermond, in his 1941 book, wrote that “in almost all fungi, the chondriosomes predominate in the form of chondrioconts, generally very elongated” (chondriosomes and chondrioconts are obsolete names for mitochondria). One of the first descriptions of super-elongated mitochondria in xylotrophic basidiomycetes was in the previously mentioned work of Armentrout et al. (1968). Using DIC, the authors demonstrated the presence of very elongated mitochondria at a certain distance from the hyphal apices in Fomes annosus (now Heterobasidion annosum). Weber et al. (1998) studied the mitochondria of seven species of fungi and oomycetes using TEM and light microscopy. He showed that in Schizophyllum commune, very long mitochondria spiral along the periphery of the cell. Paxillus involutus also exhibits “very long mitochondrial filaments” at a certain distance from the hyphal apex (Tuszynska et al. 2006; Tuszyńska 2006). In our work (Mazheika et al., 2020), we studied super-elongated mitochondria in several species of xylotrophic basidiomycetes using fluorescence microscopy and TEM. In that study, we introduced the term “super-elongated mitochondria”.
3.1. Methods of Labeling and Microscopic Visualization of the Fungal Mitochondrial System
In classical studies, fungal mitochondria were labeled with various chromophore dyes, among which the vital dye Janus Green is notable (Guilliermond 1941). In his 1998 work, Weber et al. labeled mitochondria chromophorically using iodotetrazolium. As with vacuoles, fluorescent labeling of fungal mitochondria is widespread (Hickey et al. 2004; Zhuang et al. 2009). All fluorescent probes that are specific to mitochondria can be divided into potential-dependent and potential-independent ones. The accumulation of potential-dependent probes in mitochondria is associated with the potential difference across the inner mitochondrial membrane. In an actively respiring mitochondrion, H+ ATPases of the inner membrane pump protons into the intermembrane space, resulting in a negatively charged matrix in a polarized mitochondrion. This contributes to the fact that certain lipophilic cationic fluorescent probes, each with specific properties, can accumulate in the matrix of polarized mitochondria. In this case, different probes will have varying affinities for the molecules of the mitochondrial matrix or membranes. Some probes, such as TMRM and TMRE (tetramethylrhodamine methyl and ethyl esters), will quickly exit the mitochondria for the cytoplasm when depolarization occurs. In contrast, other probes that bind to mitochondrial proteins, phospholipids, and other molecules will exit the mitochondria more slowly when they lose their potential. Some potential-dependent probes, such as orange-red MitoTrackers, can label mitochondria after depolarization and after aldehyde fixation due to the chloromethyl groups introduced into them. However, unlike potential-independent probes, they cannot accumulate in initially depolarized mitochondria.
The direct dependence of the fluorescent signal on the degree of mitochondrial polarization occurs only when using probes at low concentrations (usually not higher than 100 nM for animal cells). When there is an excess of the probe, it begins to form conglomerates in the mitochondrial matrix, and the signal drops. In this case, the relationship between the signal and mitochondrial polarization becomes inverse – the lower the mitochondrial potential, the less probe aggregation and the stronger the fluorescent signal (Perry et al. 2011).
Rhodamine 123 is often used to label fungal mitochondria. Some studies also use rosamine MitoTrackers (Thermo Fisher Scientific, USA; Hickey et al. 2004; Hickey, Read, 2009; Potapova et al., 2014; Stodulkova et al., 2015). Styryl probes such as DASPMI, FM1-43, and FM4-64 are also applied (Fischer-Parton et al., 2000; Hickey et al. 2004; Hickey, Read 2009). However, styryls require significant time to accumulate in mitochondria (20-40 minutes) and, according to our observations, they label mitochondria only in some individual cells. The carbocyanine probe DiOC6(3) (3,3’-dihexyloxacarbocyanine iodide) is frequently used (Cole et al. 1997).
Another type of probe is the potential-independent probe. These probes concentrate in mitochondria regardless of their potential, binding to mitochondrial residents to varying degrees. An example of such a probe is MitoTracker™ Green FM and its analogs from other manufacturers, which, like DiOC6(3), is a carbocyanine derivative (Hickey et al. 2004; Besserer et al. 2008; Walter et al. 2010; Scheckhuber 2015).
Based on our experience, the best fluorescent probe for labeling mitochondria of basidiomycetes, including super-elongated mitochondria, is rhodamine 6G (R6G) among potential-dependent probes. Among potential-independent probes, the inexpensive yet effective nonyl acridine orange (NAO) stands out (Mazheika et al. 2020). R6G labels mitochondria well in orange-red light in slides filled with mycelial growth medium, without washing, at concentrations of 100 nM and below. R6G is possibly the only drug that qualitatively labels fungal mitochondria at such low concentrations. The disadvantages of the probe include penetration into the green light spectrum and relatively fast fading under a microscope’s light beam. R6G is undeservedly forgotten by mycologists, since it is generally considered to be a respiratory inhibitor or uncoupler, and its signal may depend on the concentration of active oxygen forms in the mitochondria (Gear 1974). However, at nanomolar concentrations, we did not detect any significant effect of R6G on the growth of xylotrophic basidiomycetes or on mitochondrial morphology. We compared two probes in the same experiments on S. hirsutum: the popular rhodamine 123 and R6G. Rhodamine 123, unlike R6G, labels mitochondria at concentrations an order of magnitude higher, gives a stronger background, and requires washing the slide.
In our experience, NAO labels many basidiomycetes well without washing, at concentrations below 100 nM (though higher concentrations may sometimes be necessary). NAO is known to bind to cardiolipin on the inner mitochondrial membrane in many eukaryotes (Mileykovskaya et al. 2001). The disadvantage of this fluorescent dye is that it fades faster than R6G under the microscope light beam. Therefore, like CDFA, it is poorly suited for long time-lapse or Z-stack videos. Working with mitochondrial probes requires a higher speed of work than with vacuolar probes. R6G and NAO leave the mitochondria 10-30 minutes after slide preparation, and super-elongated mitochondria are best observed in the first 10 minutes. After leaving the mitochondria, NAO may begin to label the vacuole tonoplasts.
Genetic transformation is also used for fluorescent labeling of fungal mitochondria (Suelmann, Fischer 2000; Bowman et al. 2009; Hickey, Read 2009; Gibeaux et al. 2013; Jagernath et al. 2020).
3.2. Mitochondrial Morphology and Diversity, Morphotypes of the Fungal Mitochondrial System
Most fungal species have mitochondria with lamellar cristae, while oomycetes and some true fungi exhibit tubular cristae (Moore, McAlear 1963; Weber et al. 1998). The diameter of fungal mitochondria in cross-section, according to different sources, ranges from 250 nm to 2 µm (Armentrout et al. 1968; Zhuang et al. 2009; Chatre, Ricchetti 2014).
Mitochondria are extremely pleomorphic organelles. In the same cells, or in cells along the same hyphae, mitochondria with completely different morphologies can be found. They can be small, granular (Video S24) or spherical; in the form of short (Videos S24, 25) or long rods (Figure 5a); thin filamentous (filiform; Figure 5b,c; Video S26), branched, intertwined, collected in bundles or a network (Video S24); tubular (thicker than filamentous and longer than rod-shaped ones; Figure 5a,d; Videos S27 and 28), also branched; can swell into a vesicle, close into a ring (Video S29); and finally, be super-elongated mitochondria (very long, up to 100 or more micrometers in length), both individual (Figure 5e; Video S30) and forming a powerful network (Figure 5f,g; Videos S31 and 32; Moore, McAlear 1963; Armentrout et al. 1968; Tuszyńska 2006; Scheckhuber et al. 2007; Zhuang et al. 2009; Oettmeier, Döbereiner 2019; Mazheika et al., 2020).
Fungal mitochondria are extremely dynamic organelles. Small mitochondria can move rapidly within fungal cells, often in jumps and back and forth along the long axis of the hypha or across. Small mitochondria can also move along long mitochondria (Video S33; Guilliermond 1941; Honda et al. 1964; Armentrout et al. 1968; Tuszynska et al. 2006; Tuszyńska 2006; Zhuang et al. 2009). Larger, longer mitochondria also move constantly within the cell (Video S34); however, a relatively stationary picture can be observed in some cases (Video S35). Another important dynamic feature of mitochondria is that they undergo constant cycles of fusion and fragmentation (Videos S36, 37). Smaller mitochondria fuse into larger ones, and then break up into smaller ones again. This can involve either complete end-to-end fusion or concatenation through MCSs (membrane contact sites; Girbardt 1955; Hawley, Wagner 1967; Armentrout et al. 1968; Suelmann, Fischer 2000; Skulachev 2001; Okamoto, Shaw 2005; Tuszyńska 2006; Hickey, Read 2009; Zhuang et al. 2009; Scheckhuber et al. 2012; Osiewacz, Bernhardt 2013; Chatre, Ricchetti 2014; Koch, Traven 2019; Navarro-Espíndola et al. 2020). The speed of these cycles, as well as the degree of fragmentation and fusion, can vary. There are descriptions of a complete cycle of mitochondrial fusion and fission occurring in about one minute (Armentrout et al. 1968; Hickey, Read 2009). However, the equilibrium can also shift towards highly fragmented mitochondria (often at the tip of the hypha) or, conversely, towards long/network mitochondria. In such cases, fusion or fragmentation will be rarer and may not occur completely. For example, a tubular mitochondrion can only divide into two parts, not into short fragments.
In fungi, there are mitochondrial morphotypes corresponding to certain taxonomic or ecotrophic groups, similar to the vacuolar system. For example, each of the three ecotrophically diverse basidiomycetes – S. hirsutum, C. comatus, and R. solani – has a different mitochondrial morphotype (Figure 4; Mazheika et al. 2020). C. comatus does not have long mitochondria; it contains either fragmented and rod-shaped mitochondria or short tubular ones (Figure 4a,b). Moreover, the short mitochondria are usually hyperpolarized, while some of the longer ones are depolarized (Mazheika et al. 2020). A typical basic variant of the Coprinus-morphotype is presented in the track collage, Figure S10 (see also Figure 4e). It shows a hypha, labeled with R6G, more than 5 mm long, and consisting of 11 cells. In the apical cell, the longest at approximately 1 mm (the other ten cells are 350 to 500 μm long), the hyphal tip is devoid of mitochondria. Large, granular, hyperpolarized mitochondria are concentrated in the subapex. The central region of the apical cell contains thin, short rods and granules (see also Videos S38, 39). In the basal part of the apical cell, rod-shaped mitochondria become thicker and brighter. The second, third, and fourth cells contain fragmented, short, rod-shaped mitochondria in their anterior part; no labeled mitochondria are present in the basal part. In the fifth through tenth cells, no mitochondrial signal is detected, or only single parietal mitochondria are found. In the last cell, located closest to the hypha, a branch of which is the described hypha, there are many mitochondria. These are rod-shaped and short tubular (up to 5 µm), hyperpolarized, parietal mitochondria. Figure 5a shows that NAO labeling can reveal tubular mitochondria 10 µm or longer. Video S40 and Figure 4b show a variant of the Coprinus-morphotype in which the apical cell is filled only with granular, hyperpolarized mitochondria. Similar variants of the Coprinus-morphotype are observed in representatives of the genus Agaricus, which may indicate that this morphotype is inherent in various fungi within the copro-humus ecotrophic group (Matrosova et al. 2009).
Figure 4.
Examples of mitochondrial morphotypes in basidiomycetes. a, b. Morphotype of C. comatus: mitochondria are granular, rod-shaped and short tubular. a. Basic variant with fragmented mitochondria in the subapex, thin rod-shaped mitochondria in the center, and thicker rod-shaped and short-tubular mitochondria in the basal region. The second cell exhibits fragmented and rod-shaped mitochondria in its acropetal section. b. A variant of the Coprinus-morphotype with completely fragmented mitochondria in the apical cell. c, d. Rhizoctonia-morphotype: characterized by a high density of thin, twisted, possibly branched, or very small, fragmented, or filiform mitochondria, collected in complex interweavings. e-g. Morphotype of S. hirsutum and other basidiomycetes: the majority of the mitochondriome is represented by long mitochondria – filiform, tubular and super-elongated, both single and collected in bundles and networks. e. Basic variant of xylotrophic morphotype: the apical region of the hypha contains short, filiform mitochondria. More basally, filiform and tubular mitochondria combine into bundles. Super-elongated mitochondria appear several cells from the apical cell. Septa are not depicted. f, g. Variants of the xylotrophic morphotype with fragmented mitochondria (f) and with powerful tubular mitochondria (g) in the apex.
Figure 4.
Examples of mitochondrial morphotypes in basidiomycetes. a, b. Morphotype of C. comatus: mitochondria are granular, rod-shaped and short tubular. a. Basic variant with fragmented mitochondria in the subapex, thin rod-shaped mitochondria in the center, and thicker rod-shaped and short-tubular mitochondria in the basal region. The second cell exhibits fragmented and rod-shaped mitochondria in its acropetal section. b. A variant of the Coprinus-morphotype with completely fragmented mitochondria in the apical cell. c, d. Rhizoctonia-morphotype: characterized by a high density of thin, twisted, possibly branched, or very small, fragmented, or filiform mitochondria, collected in complex interweavings. e-g. Morphotype of S. hirsutum and other basidiomycetes: the majority of the mitochondriome is represented by long mitochondria – filiform, tubular and super-elongated, both single and collected in bundles and networks. e. Basic variant of xylotrophic morphotype: the apical region of the hypha contains short, filiform mitochondria. More basally, filiform and tubular mitochondria combine into bundles. Super-elongated mitochondria appear several cells from the apical cell. Septa are not depicted. f, g. Variants of the xylotrophic morphotype with fragmented mitochondria (f) and with powerful tubular mitochondria (g) in the apex.
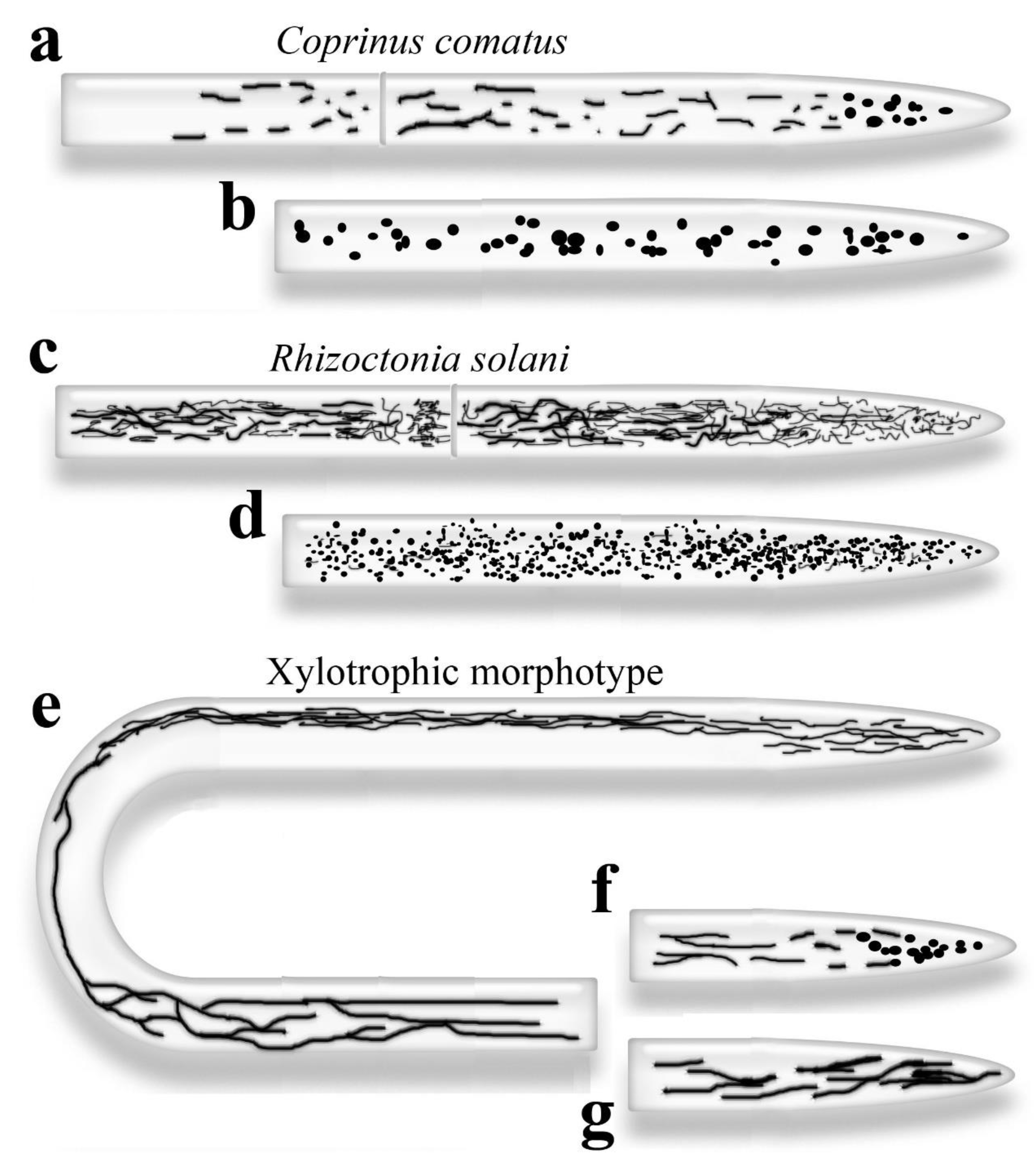
Figure 5.
Fluorescently labeled basidiomycete mitochondria and corresponding hyphal regions in transmitted light. a. C. comatus, NAO labeling. Rod-shaped and short tubular mitochondria. The arrow points to a tubular mitochondrion about 10 μm long; many of these mitochondria are not detected with R6G labeling. b. Apical region of R. solani hypha, NAO labeling. The cell is filled with short, twisted, and intertwined filiform mitochondria. Individual mitochondria can be seen touching the very tip of the cell. c. S. hirsutum, R6G labeling. The apical part of the hypha exhibits a typical xylotrophic morphotype: the apex and subapex are filled with filiform mitochondria, in some places collected in bundles. d. S. hirsutum, NAO labeling. One of the variants of the xylotrophic morphotype: the apical part of the hypha contains tubular mitochondria (arrow). This variant probably corresponds to hyphae that have stopped growing. e. S. hirsutum, NAO labeling. Super-elongated mitochondrion, possibly not solid but consisting of individual tubular mitochondria joined together (arrows). f, g. S. hirsutum, NAO labeling: A robust network of super-elongated mitochondria. h. S. hirsutum, co-labeling of NAO (green) and R6G (red). In the apical part of the hypha, the probe signals mostly coincide (yellow signal). In the basal cells (arrow), mitochondria carry only the NAO signal. All photographs were obtained by the authors. The scale bar is 5 μm.
Figure 5.
Fluorescently labeled basidiomycete mitochondria and corresponding hyphal regions in transmitted light. a. C. comatus, NAO labeling. Rod-shaped and short tubular mitochondria. The arrow points to a tubular mitochondrion about 10 μm long; many of these mitochondria are not detected with R6G labeling. b. Apical region of R. solani hypha, NAO labeling. The cell is filled with short, twisted, and intertwined filiform mitochondria. Individual mitochondria can be seen touching the very tip of the cell. c. S. hirsutum, R6G labeling. The apical part of the hypha exhibits a typical xylotrophic morphotype: the apex and subapex are filled with filiform mitochondria, in some places collected in bundles. d. S. hirsutum, NAO labeling. One of the variants of the xylotrophic morphotype: the apical part of the hypha contains tubular mitochondria (arrow). This variant probably corresponds to hyphae that have stopped growing. e. S. hirsutum, NAO labeling. Super-elongated mitochondrion, possibly not solid but consisting of individual tubular mitochondria joined together (arrows). f, g. S. hirsutum, NAO labeling: A robust network of super-elongated mitochondria. h. S. hirsutum, co-labeling of NAO (green) and R6G (red). In the apical part of the hypha, the probe signals mostly coincide (yellow signal). In the basal cells (arrow), mitochondria carry only the NAO signal. All photographs were obtained by the authors. The scale bar is 5 μm.
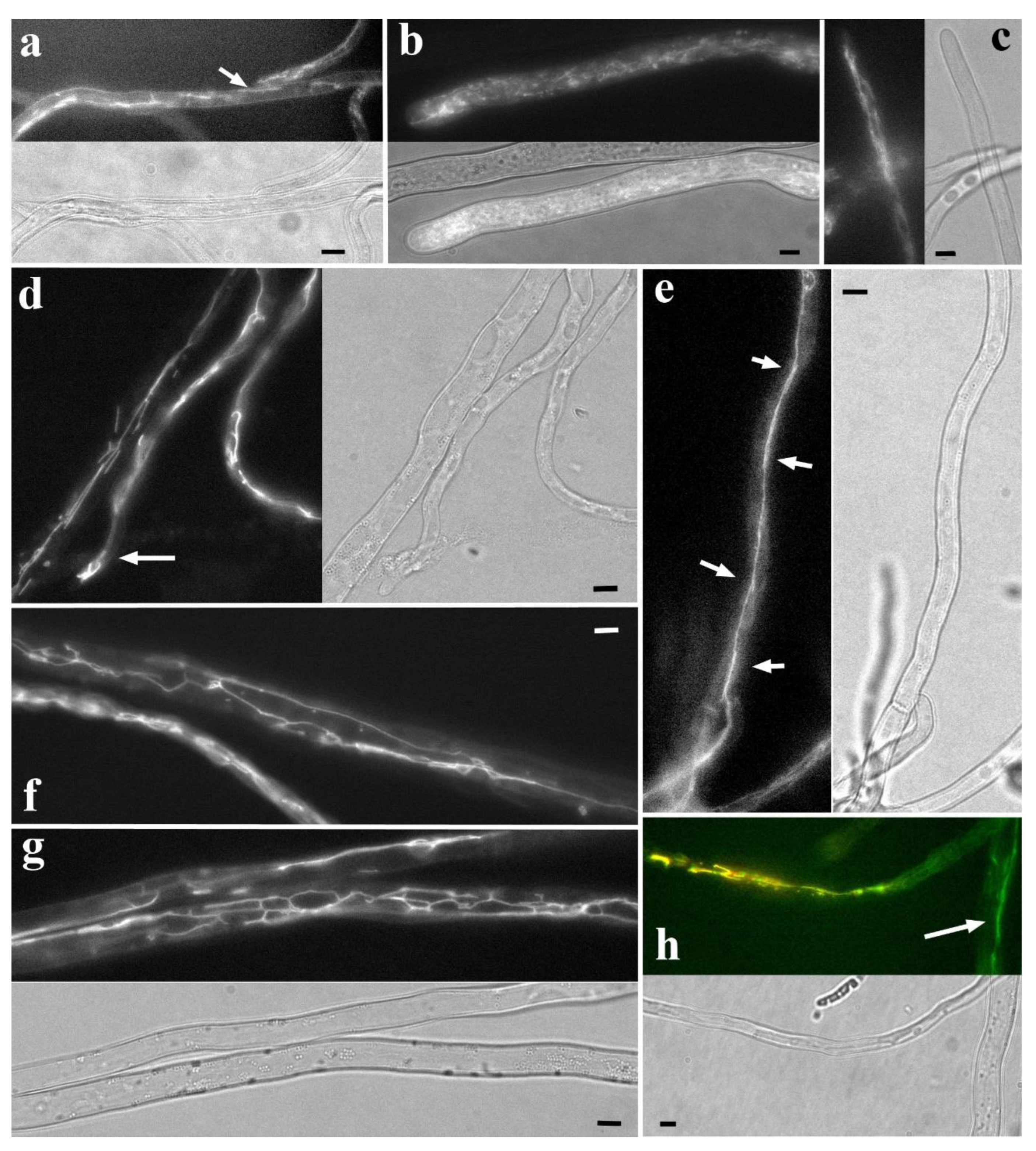
A distinctive feature of the R. solani morphotype is the abundance of mitochondria, which fill the entire cell (Figure 4c,d and Figure 5b). The mitochondria are either small granular (Videos S41-43), or thin, short, and convoluted, possibly branched, or thin and longer, uniting into an amorphous network (Videos S44-46). Track collage (Figure S11) shows the basic variant of the Rhizoctonia-morphotype. Closer to the apex, the mitochondria are shorter; further from the apex, they become longer. In both cases, they combine into a complex, tangled network, making it almost impossible to visualize individual mitochondria (see also Figure 4c,d). NAO and R6G often label relatively large ring structures in R. solani (Video S47). The saccharomycete filamentous fungus Ashbya gossypii may possess a mitochondrial morphotype similar to the Rhizoctonia-morphotype (Gibeaux et al. 2013). This morphotype may characterize a specific category of phytopathogenic or endophytic fungi, but further research is needed.
S. hirsutum, like many wood-decaying basidiomycetes and possibly ectomycorrhizal fungi, has a morphotype with highly developed and elongated mitochondria (Armentrout et al. 1968; Zhuang et al. 2009; Mazheika et al. 2020). The basic variant of the morphotype is presented in the track collage (Figure S12; see also Figure 4e). The apical part of the first cell is usually filled with short or long filiform mitochondria (Figure 4e and Figure 5c; Videos S26, S48 and S49). Further, starting from the middle of the apical cell and extending over several basal cells, the mitochondrion is represented by bundles or a dense network of filiform mitochondria, stretching along the hyphae, often pressed to one side of the cell (Figure 4e; Videos S50 and S51). In such bundles or networks, individual mitochondria are difficult to distinguish. As the distance from the apical cell increases, the mitochondria within the bundles/network become thicker and more discernible. Gradually, super-elongated mitochondria appear in the cells (Figure 4e ad Figure 5e-g; Video S32, S52 and S53). In Figure S12 they appear starting from the fifth cell from the apex. In even more basal cells the mitochondria are fragmented into rod-shaped or tubular ones. Often, one can observe a sharp transition: in neighboring cells, mitochondrial morphology is completely different – fragmented in one cell, and super-elongated and networked in another (Video S24). As with the bundles of filiform ones, the super-elongated mitochondria and their networks may be confined preferentially to one side of the cell (Video S54). In distant basal cells, the signal from the mitochondria usually disappears (Armentrout et al. 1968; Tuszyńska 2006; Zhuang et al. 2009; Mazheika et al., 2020).
Variants of the xylotrophic morphotype are possible, most often concerning the apical zone of the hypha. The apical part of the first cell in the basic variant is filled with short, filiform mitochondria (Figure 4e and Figure 5c). Variants with granular, hyperpolarized mitochondria in the apex and subapex are also possible (Figure 4f). Another possible variant is tubular mitochondria, collected in small bundles, starting from the subapex (Figure 4g and Figure 5d). As with the vacuolar system, morphological variations of mitochondria in the apical hypha are probably associated with the hypha’s growth rate. In a rapidly growing hypha, mitochondria are maximally fragmented in the apex and subapex; when growth slows, they lengthen, and when growth stops, the mitochondria thicken.
Super-elongated mitochondria are a distinctive feature of xylotrophic and mycorrhiza-forming basidiomycetes. They can exhibit different morphologies: stretching along the cell as a slightly or frequently branched tube, or forming a large network (Figure 4e and Figure 5e-g; Mazheika et al., 2020). It is not always obvious whether a particular super-elongated mitochondrion is a single unit or consists of shorter tubular mitochondria concatenated at their ends (Figure 5e). Weber et al. (1998) noted that long mitochondria in S. commune cells, where large spherical vacuoles are localized, contain few cristae. A similar result was obtained in our work: based on TEM, we concluded that in S. hirsutum, younger cells have long mitochondria with cristae, while in more mature cells, long/super-elongated mitochondria lose cristae and transform into long double-membrane tubes (Mazheika et al., 2020). This transformation must be accompanied by mitochondrial depolarization (Figure 5h).
3.3. Molecular Mechanisms of Mitochondrial Morphogenesis and Dynamic, Influence of the Cytoskeleton on the Mitochondrial System
The conserved protein complex MICOS is responsible for the formation of cristae on the inner mitochondrial membrane (Muñoz-Gómez et al. 2015).
As with many endomembrane structures, mitochondrial fragmentation and elongation are carried out by the dynamin-like proteins Dnm1 and Fis1 (Okamoto, Shaw 2005; Westermann 2010; Chang, Doering 2018; Garrido-Bazán et al. 2020; Navarro-Espíndola et al. 2020; Hernández-Sánchez, Peraza-Reyes 2022). In mutants of these proteins, mitochondrial fragmentation is disrupted, and mitochondria become elongated or swell into large spherical structures (however, Dnm\Fis-independent fragmentation also occurs; Scheckhuber et al. 2007; Westermann 2010; Gibeaux et al. 2013; Navarro-Espíndola et al. 2020; Hernández-Sánchez, Peraza-Reyes 2022). Mitofusins (Mfn1/2/Fzo1 and OPA1/Mgm1) are responsible for the fusion of mitochondria (Navarro-Espíndola et al. 2020). Mitochondrial morphology is also influenced by mitochondrial import proteins and ERMESes (ER-mitochondrial encounter structures – MCSs between endoplasmic reticulum and mitochondria; Kornmann et al. 2009; Westermann 2010; Koch, Traven 2019; Navarro-Espíndola et al. 2020; Cheema et al. 2021).
The tubular morphology of mitochondria in many fungi is also due to the tubulin cytoskeleton (Westermann 2010). As with tubular vacuoles, microtubules can be observed in TEM photographs along elongated mitochondria (Roberson et al. 2010; Faoro et al. 2022). However, it is generally accepted that in some fungi, actin is responsible for the movement of mitochondria, as seen in A. nidulans and yeast (Westermann 2010).
3.4. Physiological Functions of the Mitochondrial System in Fungi, Division of Labor, the Meaning of Fragmentation and Fusion Cycles
One of the primary functions of mitochondria is oxidative phosphorylation. However, like any other cell organelle, mitochondria are multifunctional (Osiewacz, Bernhardt 2013; Chatre, Ricchetti 2014; Chang, Doering 2018; Koch, Traven 2019; Cheema et al. 2021). Having their own genome, mitochondria synthesize proteins, and it turns out that some yeast proteins previously considered nuclear are produced by mitochondria (Koch, Traven 2019). In mitochondria, there is active synthesis of various metabolites, such as certain amino acids, the synthesis of Fe-S clusters and heme. Mitochondria are important producers of phospholipids, including phosphatidylethanolamine, which they supply to the entire cell (Verma et al. 2018; Koch, Traven 2019). Mitochondria are the key link in triggering apoptosis (Lu 2006).
Depolarized mitochondria are excluded from the respiratory process but often retain high transcriptional and synthetic activities. Cryptococcus neoformans populations contain VNBCs (viable but non-culturable cells), which, like in many microorganisms, help the population survive adverse conditions and antibiotic therapy. VNBCs contain many depolarized mitochondria but exhibit high transcriptional activity, apparently performing protective and detoxification functions (Black et al. 2021). Budding yeasts in microaerobic conditions switch to glycolysis. However, they can maintain the potential difference on the mitochondrial membrane by expending ATP to sustain the necessary physiological functions of mitochondria (Jazwinski 2004). Examples of the division of labor between mitochondria or fungal cells carrying functional different mitochondria are interesting. The human pathogen C. gattii has two cell populations: in the first population, the mitochondria are elongated; these cells divide slowly but are resistant to oxidative stress and survive inside macrophages. In the second population, the mitochondria are fragmented; these cells divide quickly but are susceptible to oxidative stress and are easily killed by macrophages. The cells of the first population can stimulate the cells of the second population to rapidly divide. Such two-population strains are highly pathogenic (Voelz et al. 2014). The presence of two populations of mitochondria in C. comatus cells, short polarized and longer depolarized, as we have shown, may also be a manifestation of the division of labor (Mazheika et al., 2020). Short mitochondria provide fungal cells with energy, while long ones perform a synthetic function, possibly a protective one. In particular, longer depolarized mitochondria may serve as a reserve pool for active hyperpolarized mitochondria.
The cycle of fragmentation of longer mitochondria into shorter ones, followed by their reassembly into long ones, is fundamentally important for the physiology of the cell and mitochondria. Skulachev, in the 1960s (Skulachev 2001), proposed that rapid fragmentation of long mitochondria or their clusters serves as a protective mechanism; when a long mitochondrion is damaged, it disintegrates to maintain the functionality of undamaged sections. Later, this hypothesis evolved into a modern concept: the constant cycle of fragmentation and fusion of mitochondria is a powerful checkpoint system. Damaged, fragmented mitochondria are subject to mitophagy, thereby preventing the loss of the entire long mitochondrion through the elimination of individual fragments (Scheckhuber et al. 2012). In cells with high activity, such as the growing apex of a hypha, increased respiratory activity and hyperpolarization of mitochondria raise the probability of oxidative shock. Additionally, the cell can produce reactive oxygen species (ROS) for specific purposes. In such cells, the equilibrium of the mitochondrial cycle is usually shifted towards fragmentation to effectively protect the cellular mitochondrial pool from damage. Experiments on fungal mutants with impaired mitochondrial fragmentation, characterized by long or large swollen mitochondria, have shown that these mutants typically exhibit reduced levels of respiration and ROS synthesis, and increased sensitivity to oxidative stress (Scheckhuber et al. 2007; Westermann 2010; Navarro-Espíndola et al. 2020). Although such a scenario is not obligatory: in A. nidulans mutants with elongated mitochondria, ROS synthesis, on the contrary, can increase (Navarro-Espíndola et al. 2020); in C. neoformans and C. gattii cells with fragmented mitochondria are more sensitive to oxidative stress (Voelz et al. 2014; Chang, Doering 2018). It follows the size of mitochondria does not determine their respiratory and oxidative activity. In most fungal cells, elongated mitochondria, ranging from rod-shaped to super-elongated, predominate, indicating the biological importance of elongated mitochondria for the cell/mycelium.
What is the functional significance of mitochondrial elongation, and why does a fungal cell without stress tend to shift the equilibrium towards mitochondrial fusion? Here, we can also discuss multifunctionality. For example, in budding yeast, it has been shown that lengthening mitochondria is important for their proper segregation during meiosis. In the second division of meiosis, mitochondria transition from a fragmented to an elongated state, attach to the nuclear envelope, and are transported to ascospores (Hernández-Sánchez, Peraza-Reyes 2022). Long mitochondria, similar to tubular vacuoles, can perform a transport function. Skulachev (2001) suggests that in animal cells, elongated mitochondria transport protons from active, fragmented peripheral mitochondria to the center of the cell, where fragmented mitochondria are experiencing an oxygen deficiency. The transport function of fungi is expanded: it is assumed that mitochondria can transport various metabolites, similar to vacuoles (Zhuang et al. 2009). The loss of cristae, as shown by Weber et al. and us in long and super-elongated mitochondria (Weber et al. 1998, Mazheika et al. 2020), also supports the transport hypothesis – mitochondria transform into long hollow pipes. However, this transformation occurs sufficiently far from the active apical cell to assume that thick, super-elongated mitochondria, both depolarized and polarized, undertake the function of mass-flow-independent generalized transport in the basal cells of the apical hypha. In the apical cell and several subapical ones, where xylotrophic basidiomycetes typically exhibit bundles or amorphous networks of filiform mitochondria, transport likely assumes a more specialized form. Here, the primary function of the mitochondria is respiration and the transfer of energy, in the form of macroergs, to the growing apex of the hypha.
4. Fungal Endoplasmic Reticulum
The endoplasmic reticulum (ER) has been relatively well studied in fungi, but unevenly among different taxonomic groups. In yeast, the ER has been studied more thoroughly than in basidiomycetes, for example. A review dedicated to the ER of filamentous fungi was recently published, covering the main modern achievements in the study of fungal ER (Martínez-Andrade et al. 2024).
Unlike the two previous tubular systems, fluorescent labeling of the ER is mainly achieved through genetic transformation. This is because there are few specific fluorescent probes for the ER. In fact, only one experimentally proven probe is suitable for studying the ER of fungi – ER-tracker Blue-White DPX (Cole et al. 2000a, b; Benhamou et al. 2018; Martínez-Andrade et al. 2024). It relatively specifically marks the ER in some species of fungi from different taxonomic groups, which suggests that it can be used for a wide range of fungal species. However, this probe emits in a wide spectral range, which does not allow its use for colocalization of the ER with other tubular systems (Cole et al. 2000a). Brefeldin A (BFA), conjugated with fluorochromes, also marks fungal ER. BFA is an inhibitor of the retrograde traffic of COPI vesicles from the Golgi to the ER (Dornan, Simpson 2023). However, BFA in fungi can disrupt the native state of the ER. In P. tinсtorius, it has been shown that BFA increases the lumen of the ER cisternae, shifts the cisternae to the center of the cells, and destroys the system of tubular vacuoles (Cole et al. 2000a, b). Fluorescents based on some azoles are promising, but so far they have been tested only on certain species of Candida (Benhamou et al. 2018). Some fluorescent probes that label mitochondria (for example, DiOC6(3) and FM4-64) can also accumulate in the ER, especially when used in high concentrations. This is known for animal cells and has been shown for some fungi (Butt et al. 1989; Cole et al. 2000a; Benhamou et al. 2018; Martínez-Andrade et al. 2024). In rare cases, we observed cells of xylotrophic basidiomycetes, labeled by R6G, with an ER-like thin threaded network filling the cell.
At the ultrastructural level, ER is well preserved even with aldehyde fixation (Shepherd et al. 1993b; Ashford 1998; Cole et al. 2000a, b; Boenisch et al. 2017; Faoro et al. 2022). However, due to the specificity of its morphology (thin lamellae and thin tubes), without the reconstruction of serial sections, TEM is not always informative in studying the ER.
Various targets are used for genetic fluorescent labeling of fungal ER. In the simplest case, an FP with an ER retention signal HDEL/KDEL at the C-terminus is expressed in the fungus (Fernandez-Abalos et al. 1998; Wedlich-Soldner et al. 2002; Hickey, Read 2009; Kilaru et al. 2017; Schuster et al. 2025). A wide range of ER resident proteins is also used, as detailed in the review article by Martinez-Andrade et al. (2024).
4.1. Morphology, Diversity and Intrahyphal Topology of Fungal Endoplasmic Reticulum
Among the three endomembrane systems reviewed, the ER has a tubular-lamellar structure, whereas the other two systems rarely exhibit lamellar morphology. The ER of fungi consists of lamellae (also known as cisternae, plates, or sheets) and tubes. The ER lamellae are rarely solid but are through perforated (like a slice of Maasdam cheese). The width of the lamellae lumen varies among fungi but is usually around 50 nm (Uetake et al. 2002; West et al. 2011; Martínez-Andrade et al. 2024). Thus, the ER is the thinnest of the three endomembrane systems discussed. ER tubes connect ER lamellae to each other and also form MCSs with other organelles (West et al. 2011). Some authors identify ER lamellae with rough ER (rER, ER regions that carry ribosomes on the cytoplasmic side) and ER tubules with smooth ER (sER, Martínez-Andrade et al. 2024). However, this is not entirely true. In many basidiomycetes, we observed in TEM preparations ER lamellae extending along the hyphae. These are easily distinguished from tubes, as thin tubes cannot remain in the plane of the ultrathin section for a long distance. These lamellae do not carry ribosomes on their surface. Interestingly, the outer membrane of fungal mitochondria can also be densely covered with ribosomes (Matrosova et al. 2009).
The intracellular topology of the ER has been well-studied in budding yeast (Rossanese et al. 1999; West et al. 2011; Stradalova et al. 2012). The ER in yeast consists of three zones. The first zone is the ER lamellae, which surround the nucleus and are connected to the outer nuclear membrane. Many researchers understand the term nuclear envelope (NE) to mean a single membrane structure consisting of nuclear membranes and perinuclear cisternae of the ER (see, for example, the figure in Groth et al. 2021). The NE is connected to the cortical ER by 6-8 ER tubes and one central ER plate; this is the second zone. The third zone: the cortical or peripheral ER, which in yeast consists predominantly of perforated lamellae lining the plasma membrane from the inside at a distance of 15-60 nm. The cortical ER occupies 20% to 45% of the inner surface of the yeast cell. It is never found under the eisosomes, and endocytosis occurs only in areas free of cortical ER (Stradalova et al. 2012).
In filamentous fungi, the ER topology differs from that of yeast. For many filamentous fungi the presence of perinuclear ER cisterns has been demonstrated (Kilaru et al. 2017). However, filamentous fungi usually do not have a cortical ER. Instead, fungal cells are permeated by a network of ER, most of which consists of lamellae running parallel to the long axis of the cell but can also be located across the cell (Ashford 1998; Cole et al. 2000a, b; Maruyama et al. 2006; Maruyama, Kitamoto 2007; Shoji et al. 2014; Faoro et al. 2022). The ER is likely the least mobile of the three tubular and tubular-lamellar systems. In P. tinctorius, for example, the ER is described as having low mobility (Cole et al. 2000a, b). Although there is evidence that in A. oryzae, at least in apical cells, the ER forms a fairly mobile network (Maruyama et al. 2006; Higuchi 2021).
For A. oryzae, it was shown that the ER in cells has a topological gradient: in the anterior part of the cell (closest to the apex), the ER network is denser, while in the posterior part, it is sparser (Maruyama et al. 2006; Maruyama, Kitamoto 2007; Shoji et al. 2014; Higuchi 2021). tER sites (sites of COPII vesicle detachment from the ER) are genetical labeled as foci of a point fluorescent signal and also exhibit a gradient of location, with a higher concentration in the frontal part of the cell (Kimura et al. 2010). In contrast, spherical vacuoles in A. oryzae are more concentrated in the basal part of the cell (Maruyama et al. 2006; Maruyama, Kitamoto 2007).
There is evidence that in some species of fungi, the ER does not penetrate the tip of the hyphae but is localized in the subapex, no further than the subapical ring. According to other data, ER tubules are found at the tip and in Spitzenkörper (see references in Martínez-Andrade et al. 2024).
Under certain conditions, such as when a fungus needs to synthesize a large number of secondary metabolites, the ER can form OSER (organized sER). OSER has been described in budding yeast and Fusarium graminearum (Wright et al. 1988; Boenisch et al. 2017). OSER is a large stack of ER lamellae, both flat and concentric. Additionally, some fungi have ER in the form of a polygonal network (Martínez-Andrade et al. 2024).
4.2. Molecular Mechanisms of Morphogenesis, Influence of the Cytoskeleton, and Physiological Functions of the Fungal Endoplasmic Reticulum
In budding yeast, ER morphology depends on reticulon proteins (Rtns), BP1/Yop1, and microtubules (West et al. 2011). With the overexpression of Rtns/Yop1, the entire ER of yeast is transformed into tubes. With the deletion of the genes for these proteins, the equilibrium shifts toward cisternae, transforming the ER into a single cisterna without perforations and tubes. Microtubule depolymerization also shifts equilibrium toward cisternae but does not affect the cortical ER in yeast. The membrane-anchored dynamin-like protein Sey1 has also been shown to influence fungal ER morphology (Martínez-Andrade et al. 2024).
Like all eukaryotes, the fungal ER has fundamental functions for the cell. First, the ER performs primary sorting of proteins: folding of newly synthesized proteins, initiating the degradation of misfolded proteins, and glycosylation of proteins, which involves the attachment of a transport signal. Lipid synthesis and their glycosylation also occur in the ER. The ER is one of the calcium storage pools (Rossanese et al. 1999; Askew 2014; Shoji et al. 2014).
In addition, the ER regulates the metabolism of phosphatidylinositol and phosphatidylcholine. In yeast, it determines the sites of endocytosis. It is known that the ER in fungi controls the morphology and sites of fragmentation of mitochondria (Koch, Traven 2019; Navarro-Espíndola et al. 2020; Cheema et al. 2021).
5. Interaction of Tubular and Tubular-Lamellar Systems with Each Other, and Their Functions as a Single Endomembrane Complex in Fungi
5.1. Vacuolar and Mitochondrial Tubular Systems
Xylotrophic basidiomycetes exhibit a pattern: thick, super-elongated tubular mitochondria form exclusively in hyphal regions devoid of tubular or motile, outgrowth-forming spherical vacuoles. Conversely, in hyphal regions dominated by tubular vacuoles or spherical vacuoles with tubular outgrowths, mitochondria are typically thin, filiform, and clustered in bundles, less frequently fragmented. This suggests a topological division along the length of the apical hypha between the tubular vacuolar system and the developed tubular mitochondrial system. The division may be not only spatial but also conditional. Thus, the faster the hypha grows, the more pronounced the tubular vacuolar system is in its apical cell, and the more fragmented the mitochondrial system is. Conversely, in the inhibited and autolytic hypha, the apical cell contains only spherical vacuoles, and the mitochondria are represented by more powerful tubular structures, uncharacteristic of the active fungal apex (Figure 4g; Figure 5d).
Spatial division of the hypha between the vacuolar and mitochondrial systems in xylotrophs occurs not only along the hyphal length but also in the cell’s cross-section. In any cell of R. solani (Figure S13) and in the apical part of the S. hirsutum hypha, mitochondria and vacuoles are usually clearly labeled separately but do not have isolated compartments in the hyphal cross-section. However, with distance from the apex in S. hirsutum, the situation changes. In photographs and videos of co-labeled mitochondria and vacuoles, the two systems often divide the cell in cross-section, with each system occupying its own half (Figure 6; Video S55). This compartmentalization is also supported by the previously presented 3D reconstructions, which show that mitochondrial bundles or networks of super-elongated mitochondria occupy a parietal position predominantly on one side of the cell (Videos S51, 54). However, such a division of space is characteristic of thick hyphae; in thin hyphae, if it exists, it is more difficult to visualize.
The spatial or condition-dependent division of the apical hypha’s internal volume between two powerful tubular membrane systems, most pronounced in xylotrophic and mycorrhizal basidiomycetes, likely holds significant biological meaning. Apical hyphae are zones of active nutrient absorption from the substrate, substance exchange with host plant cells, and resource-intensive hyphal growth (Fricker et al. 2008; Steinberg et al. 2017; Lew 2019; Schmieder et al. 2019; Herman, Bleichrodt. 2022). Besides, the apical hyphae are zones of constantly changing turgor pressure gradients, alterations in the direction of mass flow, and even areas of counter-directed mass flow (Takeshita et al. 2014; Peñalva et al. 2017; Steinberg et al. 2017; Takeshita 2019). Therefore, the two tubular systems ensure the transfer of substances from the trunk hyphae along the apical hypha to its tip and back, independent of turgor changes and mass flow directions. It can be assumed that these two systems complement each other in this single transport function. Such complementation is important because neither system can fully implement this transport function independently. The multifunctionality of these systems is the reason. Large spherical vacuoles are necessary for turgor regulation, nutrient storage, cytoplasm detoxification, etc (Weber 2002; Lilje, Lilje 2006; Shoji et al. 2006a; Veses et al. 2008; Richards et al. 2010; Shoji et al. 2014; Chen et al. 2020). Their isolation from each other is probably important for the proper functioning of the hyphae. Therefore, in the basal part of the apical hypha, where the cells are forced to form only large spherical vacuoles, the super-elongated mitochondria assume the transport function. In the apical part of the hypha, within the zone of active growth, a substantial amount of energy is required. The mitochondria’s function transitions from transport to respiratory. This transition is likely incompatible with the rapid transport of fluids within the mitochondrial matrix or intermembrane space. In addition, in active mitochondria, the equilibrium shifts toward fragmentation, resulting in granular mitochondria at the apical part of the rapidly growing hypha. This further impedes the performance of the local transport function. Therefore, in subapical cells, this function is assumed by motile spherical vacuoles connected by tubular outgrowths, and in the apical cell, by tubular vacuoles.
5.2. The Endoplasmic Reticulum Accompanies, Isolates, and Controls the Vacuolar and Mitochondrial Tubular Systems
The patterns of mutual arrangement of ER and vacuoles/mitochondria in apical hyphae are not obvious. In TEM photographs, it is often possible to observe that ER cisternae, located along the longitudinal axis of the hypha, separate mitochondria and vacuoles. Additionally, in some cases, if mitochondria are not located too close to the cell’s periphery, then ER cisternae extend between mitochondria or vacuoles and the plasma membrane (Roberson, Fuller 1988; Shepherd et al. 1993a; Ashford 1998; Cole et al. 2000a; Faoro et al. 2022). It can be assumed that the ER creates a type of braid for the mitochondrial and vacuolar (especially tubular vacuolar) systems, separating the mitochondria and vacuoles from each other. This may also create a common braid, separating both systems from the cell membrane. This is analogous to an electric cable, where individual multi-core wires are covered with their own insulating braid, and all are enclosed in a common braid. This mutual topology may indicate an isolating function of the ER in the tubular-lamellar complex and/or an accompanying function – tubular systems require the transfer and/or exchange of substances with the ER at discrete sites or continuously along their length.
In budding yeast, ERMES includes three lipid transfer proteins (Mmm1, Mdm12/34) and the mitochondrial protein Mdm10. Mmm1 is embedded in the ER membrane, and Mdm10 is embedded in the outer mitochondrial membrane (Cheema et al. 2021). ERMESes are known to be responsible for lipid transport between the ER and mitochondria (Kawano et al. 2018). Mutants in ERMES proteins have disrupted mitochondrial tubular morphology (Koch, Traven 2019; Cheema et al. 2021). ERMESes identify sites of mitochondrial fragmentation or fusion (Navarro-Espíndola et al. 2020). All this suggests that the ER can not only have accompanying and isolating functions in relation to the mitochondrial system, but also a control function. Unfortunately, similar data for the interaction of the ER and the vacuolar system in fungi are unavailable. However, the Vam6 protein, a component of vCLAMP (an MCS between mitochondria and vacuoles; Cheema et al. 2021), is known to affect the morphology and function of vacuoles in several fungi (Liu et al. 2024). This suggests a stepwise control: ER–mitochondria–vacuoles.
6. Conclusions
It is highly probable that the tubular endomembrane systems of fungi facilitate localized transport in physiologically compromised regions of the mycelium. Such regions include, but are not limited to, areas of hyphal curving, breakage, or mycelial autolysis. However, tubular systems are most frequently and fully used by fungi and oomycetes in apical hyphae. Tubular systems either replace or supplement the exocytotic-endosomal system in the apex and subapex, or provide mass-flow-independent transfer of dissolved substances to and from the apex. Tubular vacuoles and mitochondria are most developed in mycorrhizal and xylotrophic basidiomycetes. This development facilitates the rapid and diverse exchange of resources with host plants or the powerful pumping function of apical hyphae, which concentrate nitrogen and other compounds from wood. These fungi contain a dynamic, developed system of tubular and motile spherical vacuoles that works in close connection with a powerful system of super-elongated mitochondria. These systems complement each other along the length of the apical hypha, but divide it into compartments. Together with the endoplasmic reticulum, the cisternae of which appear to braid both tubular systems, accompany them, and isolate and control them, all three systems form a single intrahyphal complex, united by common physiological functions.
Despite a century of accumulating data on fungal tubular and tubular-lamellar systems, understanding these unusual intracellular structures is still in its nascent stages. The functional differences between bundles of filiform mitochondria in apical hyphae’s anterior cells and the powerful, super-elongated mitochondria of basal cells remain to be determined. It is necessary to determine whether the polarized, elongated mitochondria of subapical cells can transport energy, in the form of high-energy molecules or protons, along the hypha, and precisely what is transported by the thick, cristae-free mitochondria of the posterior cells of the apical hypha. Understanding the biological function of vacuole and mitochondria compartmentalization, and the significance of their isolation by endoplasmic reticulum cisternae, is essential. A detailed study of the interaction between the endoplasmic reticulum and tubular systems at structural and biochemical levels is necessary. This will help elucidate the endoplasmic reticulum’s controlling role concerning vacuoles and mitochondria. It is necessary to study the transport of dissolved substances through tubular vacuoles directly, not through theoretical models. This includes examining its mechanisms, speed, selectivity, and energy consumption. This is a small part of the problems mycologists will have to solve in the study of tubular and tubular-lamellar fungal systems. Studies of this kind are of fundamental importance for mycology in general and the structural physiology of fungi in particular, and indicate directions for research into large, elongated endomembrane structures in other organisms.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Acknowledgements
The research was carried out as part of the Scientific Project of the State Order of the Government of Russian Federation to Lomonosov Moscow State University No 121032300079–4 and VIGG RAS No 0092-2022-0002. The Moscow University Development Program (UDP-10) provided part of the equipment for this study.
Abbreviations
CFDA – carboxyfluorescein diacetate; ER – endoplasmic reticulum; FP – fluorescent protein; MCS – membrane contact site; NAO – nonyl acridine orange; PVC – prevacuolar compartment; R6G – rhodamine 6G; TEM – transmission electron microscopy.
References
- Abadeh A, Lew RR. 2013. Mass flow and velocity profiles in Neurospora hyphae: partial plug flow dominates intrahyphal transport. Microbiol 159:2386–2394. [CrossRef]
- Allaway WG, Ashford AE, Heath IB, Hardham AR. 1997. Vacuolar reticulum in oomycete hyphal tips: an additional component of the Ca2+ regulatory system? Fungal Genet Biol 22:209–20. [CrossRef]
- Allaway WG, Ashford AE. 2001. Motile tubular vacuoles in extramatrical mycelium and sheath hyphae of ectomycorrhizal systems. Protoplasma 215:218–25. [CrossRef]
- Armentrout VN, Smith GG, Wilson CL. 1968. Spherosomes and mitochondria in the living fungal cell. Am J Bot 55:1062–7. [CrossRef]
- Ashford AE, Orlovich DA. 1995. Vacuoles, phosphorus and endosomes in fungal hyphae. In Current Topics of Plant Physiology. Pollen–Pistil Interactions and Pollen Tube Growth. Kao TH, Stephenson AG, Eds. pp. 135–149. American Society of Plant Physiologists, Rockville, MD.
- Ashford, AE. 1998. Dynamic pleiomorphic vacuole systems: are they endosomes and transport compartments in fungal hyphae? Adv Bot Res 28:119–59. [CrossRef]
- Ashford AE, Cole L, Hyde GJ. 2001. Motile Tubular Vacuole Systems. In: Howard RJ, Gow NAR. (eds) Biology of the Fungal Cell. The Mycota 8. Springer, Berlin, Heidelberg. [CrossRef]
- Ashford, A. Tubular vacuoles in arbuscular mycorrhizas. 2002. New Phytol 154:545–547. [CrossRef]
- Askew, DS. 2014. Endoplasmic reticulum stress and fungal pathogenesis converge. Virulence 5:331–3. [CrossRef]
- Benhamou RI, Jaber QZ, Herzog IM, Roichman Y, Fridman M. 2018. Fluorescent tracking of the endoplasmic reticulum in live pathogenic fungal cells. ACS Chem Biol 13:3325–32. [CrossRef]
- Bera A, Gupta Jr ML. 2022. Microtubules in microorganisms: how tubulin isotypes contribute to diverse cytoskeletal functions. Front Cell Develop Biol 10:913809. [CrossRef]
- Berepiki A, Lichius A and Read ND. 2011. Actin organization and dynamics in filamentous fungi. Nat Rev Microbiol 9:876–887. [CrossRef]
- Besserer A, Bécard G, Jauneau A, Roux C, Séjalon-Delmas N. 2008. GR24, a synthetic analog of strigolactones, stimulates the mitosis and growth of the arbuscular mycorrhizal fungus Gigaspora rosea by boosting its energy metabolism. Plant Physiol 148:402–13. [CrossRef]
- Black B, Lee C, Horianopoulos LC, Jung WH, Kronstad JW. 2021. Respiring to infect: Emerging links between mitochondria, the electron transport chain, and fungal pathogenesis. PLoS Pathogens 17:e1009661. [CrossRef]
- Boddy, L. 1999. Saprotrophic cord-forming fungi: meeting the challenge of heterogeneous environments. Mycologia 91:13–32. [CrossRef]
- Boenisch MJ, Broz KL, Purvine SO, Chrisler WB, Nicora CD, Connolly LR, Freitag M, Baker SE, Kistler HC. 2017. Structural reorganization of the fungal endoplasmic reticulum upon induction of mycotoxin biosynthesis. Sci Rep 7:44296. [CrossRef]
- Bowman BJ, Draskovic M, Freitag M, Bowman EJ. 2009. Structure and distribution of organelles and cellular location of calcium transporters in Neurospora crassa. Eukaryot Cell 8:1845–55. [CrossRef]
- Bowman BJ, Draskovic M, Schnittker RR, El-Mellouki T, Plamann MD, Sánchez-León E, Riquelme M, Bowman EJ. 2015. Characterization of a novel prevacuolar compartment in Neurospora crassa. Eukaryot Cell 14:1253–63. [CrossRef]
- Bowman, BJ. 2023. The structure of prevacuolar compartments in Neurospora crassa as observed with super-resolution microscopy. PLOS One18:e0282989. [CrossRef]
- Brownlee C, Jennings DH. 1982. Long distance translocation in Serpula lacrimans: velocity estimates and the continuous monitoring of induced perturbations. Trans Brit Mycol Soc 79:143–8. [CrossRef]
- Butt TM, Hoch HC, Staples RC, Leger RYS. 1989. Use of Fluorochromes in the Study of Fungal Cytology and Differentiation. Exp Mycol 13:303–313. [CrossRef]
- Cairney JWG. 1992. Translocation of solutes in ectomycorrhizal and saprotrophic rhizomorphs. Mycol Res 96:135–141. [CrossRef]
- Chang AL, Doering TL. 2018. Maintenance of mitochondrial morphology in Cryptococcus neoformans is critical for stress resistance and virulence. MBio 9:10–128. [CrossRef]
- Chatre L, Ricchetti M. 2014. Are mitochondria the Achilles’ heel of the Kingdom Fungi?. Curr Opin Microbiol Aug 20:49–54. [CrossRef]
- Cheema JY, He J, Wei W, Fu C. 2021. The endoplasmic reticulum-mitochondria encounter structure and its regulatory proteins. Contact 4:25152564211064491. [CrossRef]
- Chen L, Wei X, Liu GL, Hu Z, Chi ZM, Chi Z. 2020. Glycerol, trehalose and vacuoles had relations to pullulan synthesis and osmotic tolerance by the whole genome duplicated strain Aureobasidium melanogenum TN3-1 isolated from natural honey. Int J Biol Macromol 165:131–40. [CrossRef]
- Chen Y, Liu J, Fan Y, Xiang M, Kang S, Wei D. and Liu X. 2022. SNARE protein DdVam7 of the nematode-trapping fungus Drechslerella dactyloides regulates vegetative growth, conidiation, and the predatory process via vacuole assembly. Microbiol Spectr 10:e01872–22. [CrossRef]
- Cole L, Hyde GJ, Ashford AE. 1997. Uptake and compartmentalisation of fluorescent probes by Pisolithus tinctorius hyphae: evidence for an anion transport mechanism at the tonoplast but not for fluid-phase endocytosis. Protoplasma 199:18–29. [CrossRef]
- Cole L, Orlovich DA and Ashford AF. 1998. Structure, function, and motility of vacuoles in filamentous fungi. Fungal Genet Biol 24:86–100. [CrossRef]
- Cole L, Davies D, Hyde GJ, Ashford AE. 2000a. ER-Tracker dye and BODIPY-brefeldin A differentiate the endoplasmic reticulum and Golgi bodies from the tubular-vacuole system in living hyphae of Pisolithus tinctorius. J Microsc 197:239–49. [CrossRef]
- Cole L, Davies D, Hyde GJ, Ashford AE. 2000b. Brefeldin A affects growth, endoplasmic reticulum, Golgi bodies, tubular vacuole system, and secretory pathway in Pisolithus tinctorius. Fungal Genet Biol 29:95–106. [CrossRef]
- Cox G, Moran KJ, Sanders F, Nockolds C, Tinker PB. 1980. Translocation and transfer of nutrients in vesicular-arbuscular mycorrhizas. III. Polyphosphate granules and phosphorus translocation. New Phytol 84:649–659. [CrossRef]
- Darrah PR, Tlalka M, Ashford A, Watkinson SC, Fricker MD. 2006. The vacuole system is a significant intracellular pathway for longitudinal solute transport in basidiomycete fungi. Eukaryot Cell 5:1111–25. [CrossRef]
- Day KJ, Casler JC, Glick BS. 2018. Budding yeast has a minimal endomembrane system. Developmental Cell 44:56–72. [CrossRef]
- Dornan LG, Simpson JC. 2023. Rab6-mediated retrograde trafficking from the Golgi: the trouble with tubules. Small GTPases 14:26–44. [CrossRef]
- Faoro F, Faccio A, Balestrini R. 2022. Contributions of ultrastructural studies to the knowledge of filamentous fungi biology and fungi-plant interactions. Front Fungal Biol 2:805739. [CrossRef]
- Fernandez-Abalos JM, Fox H, Pitt C, Wells B, Doonan JH. 1998. Plant – adapted green fluorescent protein is a versatile vital reporter for gene expression, protein localization and mitosis in the filamentous fungus, Aspergillus nidulans. Mol Microbiol 27:121–30. [CrossRef]
- Fischer-Parton S, Parton RM, Hickey PC, Dijksterhuis J, Atkinson HA, Read ND. 2000. Confocal microscopy of FM4-64 as a tool for analysing endocytosis and vesicle trafficking in living fungal hyphae. J Microsc 198:246e259. [CrossRef]
- Fricker MD, Lee JA, Bebber DP, Tlalka M, Hynes J, Darrah PR, Watkinson SC, Boddy L. 2008. Imaging complex nutrient dynamics in mycelial networks. J Microsc 231:317–31. [CrossRef]
- Funamoto R, Saito K, Oyaizu H, Aono T, Saito M. 2015. pH measurement of tubular vacuoles of an arbuscular mycorrhizal fungus, Gigaspora margarita. Mycorrhiza 25:55–60. [CrossRef]
- Garrido-Bazán V, Pardo JP, Aguirre J. 2020. DnmA and FisA mediate mitochondria and peroxisome fission, and regulate mitochondrial function, ROS production and development in Aspergillus nidulans. Front Microbiol 11:837. [CrossRef]
- Gear, AR. 1974. Rhodamine 6G: a potent inhibitor of mitochondrial oxidative phosphorylation. J Biol Chem 249:3628–37. [CrossRef]
- Gibeaux R, Hoepfner D, Schlatter I, Antony C, Philippsen P. 2013. Organization of organelles within hyphae of Ashbya gossypii revealed by electron tomography. Eukaryotic Cell 12:1423–32. [CrossRef]
- Girbardt, M. 1955. Lebendbeobachtungen an Polystictus versicolor (L). Flora 142:540–563.
- Gokbayrak ZD, Patel D, Brett CL. 2022. Acetate and hypertonic stress stimulate vacuole membrane fission using distinct mechanisms. Plos One 17:e0271199. [CrossRef]
- Granlund HI, Jennings DH, Thompson W. 1985. Translocation of solutes along rhizomorphs of Armillaria mellea. Trans Br Mycol Soc 84:111–119. [CrossRef]
- Griffin, DH. 1994. Fungal Physiology. 2nd edn. Wiley-Liss, New York. p.
- Groth A, Ahlmann S, Werner A, Pöggeler S. 2022. The vacuolar morphology protein VAC14 plays an important role in sexual development in the filamentous ascomycete Sordaria macrospora. Curr Genet 68:407–27. [CrossRef]
- Groth A, Schmitt K, Valerius O, Herzog B, Pöggeler S. 2021. Analysis of the putative nucleoporin POM33 in the filamentous fungus Sordaria macrospora. J Fungi 7:682. [CrossRef]
- Guilliermond, A. 1911. Sur les mitochondries des cellules végétales. Compt Rend Acad Sci 153:199–201.
- Guilliermond, A. 1941. The cytoplasm of the plant cell. Chronica Botanica Co. Waltham, Mass. USA. p. 247.
- Hatch, WR. 1935. Gametogenesis in Allomyces arbuscula. Ann Bot 49:623–649.
- Hawley ES, Wagner RP. 1967. Synchronous mitochondrial division in Neurospora crassa. J Cell Biol 35:489–99. [CrossRef]
- Heath I, Steinberg G. 1999. Mechanisms of hyphal tip growth: tube dwelling amebae revisited. Fungal Genet Biol 28:79–93. [CrossRef]
- Herman KC, Bleichrodt R. 2022. Go with the flow: mechanisms driving water transport during vegetative growth and fruiting. Fungal Biol Rev 41:10–23. [CrossRef]
- Hernández-Sánchez F, Peraza-Reyes L. 2022. Spatiotemporal dynamic regulation of organelles during meiotic development, insights from fungi. Front Cell Dev Biol 10:886710. [CrossRef]
- Hickey PC, Swift SR, Roca MG, Read ND. 2004. Live-cell imaging of filamentous fungi using vital fluorescent dyes and confocal microscopy. Methods Microbiol 34:63–87. [CrossRef]
- Hickey PC, Read ND. 2009. Imaging living cells of Aspergillus in vitro. Med Mycol 47:S110–9. [CrossRef]
- Higuchi, Y. 2021. Membrane traffic in Aspergillus oryzae and related filamentous fungi. J Fungi 7:534. [CrossRef]
- Hinze C, Boucrot E. 2018. Local actin polymerization during endocytic carrier formation. Biochem Soc Trans 46:565–576. [CrossRef]
- Honda SI, Hongladarom T, Wildman SG. 1964. Characteristic movements of organelles in streaming cytoplasm of plant cells, p. 485-502. In Allen RD, Kamlya N. [ed.], Primitive motile systems in cell biology. Academic Press, New York.
- Hyde GJ, Ashford AE. 1997. Vacuole motility and tubule-forming activity in Pisolithus tinctorius hyphae are modified by environmental conditions. Protoplasma 198:85–92. [CrossRef]
- Hyde GJ, Davies D, Perasso L, Cole L, Ashford AE. 1999. Microtubules, but not actin microfilaments, regulate vacuole motility and morphology in hyphae of Pisolithus tinctorius. Cell Motil Cytoskeleton 42:114–124. [CrossRef]
- Hyde GJ, Davies D, Cole L, Ashford AE. 2002. Regulators of GTP-binding proteins cause morphological changes in the vacuole system of the filamentous fungus, Pisolithus tinctorius. Cell Motil Cytoskeleton 51:133–46. [CrossRef]
- Inselman AL, Gathman AC, Lilly WW. 1999. Two fluorescent markers identify the vacuolar system of Schizophyllum commune. Curr Microbiol 38:295–9. [CrossRef]
- Jagernath JS, Meng S, Qiu J, Shi H, Kou Y. 2020. Selective degradation of mitochondria by mitophagy in pathogenic fungi. Am J Mol Biol 11:15–27. [CrossRef]
- Jazwinski, SM. 2004. Mitochondria, metabolism, and aging in yeast. Topics in Current Genetics: Model Systems in Aging. Eds Nystrőm T, Osiewacz HD. Berlin; Heidelberg; N.Y.: Springer-Verlag. p. 39–55. [CrossRef]
- Jennings, DH. 1987. The translocation of solutes in fungi. Biol Rev 62:215–143. [CrossRef]
- Kawano S, Tamura Y, Kojima R, Bala S, Asai E, Michel AH, Kornmann B, Riezman I, Riezman H, Sakae Y et al. 2018. Structure-function insights into direct lipid transfer between membranes by Mmm1-Mdm12 of ERMES. J Cell Biol 217:959–974. [CrossRef]
- Kilaru S, Schuster M, Ma W, Steinberg G. 2017. Fluorescent markers of various organelles in the wheat pathogen Zymoseptoria tritici. Fungal Genet Biol 105:16–27. [CrossRef]
- Kimura S, Maruyama JI, Watanabe T, Ito Y, Arioka M, Kitamoto K. 2010. In vivo imaging of endoplasmic reticulum and distribution of mutant α-amylase in Aspergillus oryzae. Fungal Genet Biol 47:1044–54. [CrossRef]
- Koch B, Traven A. 2019. Mitochondrial control of fungal cell walls: models and relevance in fungal pathogens. In Latgé JP. (eds) The Fungal Cell Wall. Current Topics in Microbiology and Immunology 425. Springer, Cham. [CrossRef]
- Kornmann B, Currie E, Collins SR, Schuldiner M, Nunnari J, Weissman JS, Walter P. 2009. An ER-mitochondria tethering complex revealed by a synthetic biology screen. Science 325:477–481. [CrossRef]
- Lew, RR. 2019. Biomechanics of hyphal growth. In: Hoffmeister, D., Gressler, M. (eds) Biology of the Fungal Cell. The Mycota, Springer, Cham. 8:83–94. [CrossRef]
- Lilje O, Lilje E. Comparative imaging of the vacuolar reticulum of Saprolegnia ferax. 2006. In Imaging, Manipulation, and Analysis of Biomolecules, Cells, and Tissues IV 6088:401–411 SPIE. USA. [CrossRef]
- Liu J, Zhang J, Yan H, Yi T, Shim WB, Zhou Z. 2024. FvVam6 is associated with fungal development and fumonisin biosynthesis via vacuole morphology regulation in Fusarium verticillioides. J Integr Agric In Press. [CrossRef]
- Lu, BC. 2006. Programmed cell death in fungi. The Mycota I: Growth, Differentiation and Sexuality. Eds Kües U, Fischer R. Berlin; Heidelberg: Springer-Verlag. p. 167–187. [CrossRef]
- Marchetti A, Lelong E, Cosson P. 2009. A measure of endosomal pH by flow cytometry in Dictyostelium. BMC Res Notes 2:7. [CrossRef]
- Martínez-Andrade JM, Roberson RW, Riquelme M. 2024. A bird’s-eye view of the endoplasmic reticulum in filamentous fungi. MMBR 88:e00027–23. [CrossRef]
- Maruyama JI, Kikuchi S, Kitamoto K. 2006. Differential distribution of the endoplasmic reticulum network as visualized by the BipA–EGFP fusion protein in hyphal compartments across the septum of the filamentous fungus, Aspergillus oryzae. Fungal Genet Biol 43:642–54. [CrossRef]
- Maruyama JI, Kitamoto K. 2007. Differential distribution of the endoplasmic reticulum network in filamentous fungi. FEMS Microbiol Lett 272:1–7. [CrossRef]
- Matrosova EV, Masheyka IS, Kudryavtseva OA, Kamzolkina OV. 2009. Mitochondrial morphogenesis and ultrastructure of basidiomycetes from genera Agaricus and Pleurotus. Cell Tiss Biol 3:369–380. [CrossRef]
- Mazheika I, Voronko O, Kudryavtseva O, Novoselova D, Pozdnyakov L, Mukhin V, Kolomiets O, Kamzolkina O. 2020. Nitrogen-obtaining and -conserving strategies in xylotrophic basidiomycetes. Mycologia 112:455–473. [CrossRef]
- Mazheika IS, Psurtseva NV, Kamzolkina OV. 2022. Lomasomes and other fungal plasma membrane macroinvaginations have a tubular and lamellar genesis. J Fungi 8:1316. [CrossRef]
- Mazheika IS, Kamzolkina OV. 2025. The curtain model as an alternative and complementary to the classic turgor concept of filamentous fungi. Archiv Microbiol 207:65. [CrossRef]
- Mileykovskaya E, Dowhan W, Birke RL, Zheng D, Lutterodt L, Haines TH. 2001. Cardiolipin binds nonyl acridine orange by aggregating the dye at exposed hydrophobic domains on bilayer surfaces. FEBS Lett 507:187–90. [CrossRef]
- Money, NP. 2025. Physical forces supporting hyphal growth. Fungal Genet Biol 177:103961. [CrossRef]
- Moore RT, McAlear JH. 1963. Fine structure of mycota: 9. Fungal mitochondria. J Ultrastruct Res 8:144–53. [CrossRef]
- Muñoz-Gómez SA, Slamovits CH, Dacks JB, Baier KA, Spencer KD, Wideman JG. 2015. Ancient homology of the mitochondrial contact site and cristae organizing system points to an endosymbiotic origin of mitochondrial cristae. Curr Biol 25:1489–95. [CrossRef]
- Navarro-Espíndola R, Suaste-Olmos F, Peraza-Reyes L. 2020. Dynamic regulation of peroxisomes and mitochondria during fungal development. J Fungi 6:302. [CrossRef]
- Oettmeier C, Döbereiner HG. 2019. Mitochondrial numbers increase during glucose deprivation in the slime mold Physarum polycephalum. Protoplasma 256:1647–55. [CrossRef]
- Ohneda M, Arioka M, Nakajima H, Kitamoto K. 2002. Visualization of vacuoles in Aspergillus oryzae by expression of CPY–EGFP. Fungal Genet Biol 37:29–38. [CrossRef]
- Ohsumi K, Arioka M, Nakajima H, Kitamoto K. 2002. Cloning and characterization of a gene (avaA) from Aspergillus nidulans encoding a small GTPase involved in vacuolar biogenesis. Gene 291:77e84. [CrossRef]
- Oka M, Maruyama J, Arioka M, Nakajima H, Kitamoto K. 2004. Molecular cloning and functional characterization of avaB, a gene encoding Vam6p/Vps39p-like protein in Aspergillus nidulans. FEMS Microbiol Lett 232:113e121. [CrossRef]
- Okamoto K, Shaw JM. 2005. Mitochondrial morphology and dynamics in yeast and multicellular eukaryotes. Annu Rev Genet 39:503–36. [CrossRef]
- Olsson S, Gray SN. 1998. Patterns and dynamics of 32P-phosphate and labelled 2-aminoisobutyric acid (14C-AIB) translocation in intact basidiomycete mycelia. FEMS Microbiol Ecology 26:109–120. [CrossRef]
- Orlovich DA, Ashford AE. 1993. Polyphosphate granules are an artefact of specimen preparation in the ecto-mycorrhizal fungus Pisolithus tinctorius. Protoplasma 173:91–105. [CrossRef]
- Osiewacz HD, Bernhardt D. 2013. Mitochondrial quality control: impact on aging and life span-a mini-review. Gerontology 59:413–20. [CrossRef]
- Peñalva MA, Zhang J, Xiang X and Pantazopoulou A. 2017. Transport of fungal RAB11 secretory vesicles involves myosin-5, dynein/dynactin/p25, and kinesin-1 and is independent of kinesin-3. Mol Biol Cell 28:947–961. [CrossRef]
- Perry SW, Norman JP, Barbieri J, Brown EB, Gelbard HA. 2011. Mitochondrial membrane potential probes and the proton gradient: a practical usage guide. BioTechniques 50:98–115. [CrossRef]
- Potapova TV, Boitzova LY, Golyshev SA, Popinako AV. 2014. The organization of mitochondria in growing hyphae of Neurospora crassa. Cell Tiss Biol 8:166–74. [CrossRef]
- Rees B, Shepherd VA, Ashford AE. 1994. Presence of a motile tubular vacuole system in different phyla of fungi. Mycol Res 98:985–92. [CrossRef]
- Reynaga-Peña CG, Bartnicki-García S. 2005. Cytoplasmic contractions in growing fungal hyphae and their morphogenetic consequences. Arch Microbiol 183:292–300. [CrossRef]
- Richards A, Veses V, Gow NA. 2010. Vacuole dynamics in fungi. Fungal Biol Rev 24:93–105. [CrossRef]
- Richards A, Gow NA, Veses V. 2012. Identification of vacuole defects in fungi. J Microbiol Methods 91:155–63. [CrossRef]
- Roberson RW, Fuller MS. 1988. Ultrastructural aspects of the hyphal tip of Sclerotium rolfsii preserved by freeze substitution. Protoplasma 146:143–149. [CrossRef]
- Roberson RW, Abril M, Blackwell M, Letcher P, McLaughlin DJ, Mouriño-Pérez RR, Riquelme M, Uchida M. 2010. Hyphal structure. In Cellular and molecular biology of filamentous fungi. Eds. Borkovich KA, Ebbole DJ. ASM Press, Washington, DC. 8–24. [CrossRef]
- Rossanese OW, Soderholm J, Bevis BJ, Sears IB, O’Connor J, Williamson EK, Glick BS. 1999. Golgi structure correlates with transitional endoplasmic reticulum organization in Pichia pastoris and Saccharomyces cerevisiae. J Cell Biol 145:69–81. [CrossRef]
- Rost FW, Shepherd VA, Ashford AE. 1995. Estimation of vacuolar pH in actively growing hyphae of the fungus Pisolithus tinctorius. Mycol Res 99:549–53. [CrossRef]
- Saito K, Kuga-Uetake Y, Saito M, Peterson RL. 2006. Vacuolar localization of phosphorus in hyphae of Phialocephala fortinii, a dark septate fungal root endophyte. Can J Microbiol 52:643–50. [CrossRef]
- Scheckhuber CQ, Erjavec N, Tinazli A, Hamann A, Nyström T, Osiewacz HD. 2007. Reducing mitochondrial fission results in increased life span and fitness of two fungal ageing models. Nat Cell Biol 9:99–105. [CrossRef]
- Scheckhuber CQ, Brust D, Osiewacz HD. 2012. Cellular homeostasis in fungi: impact on the aging process. Subcell Biochem 57:233–250. [CrossRef]
- Scheckhuber, CQ. 2015. Penicillium chrysogenum, 1: system for studying cellular effects of methylglyoxal. BMC Microbiol 15. [CrossRef]
- Schmieder SS, Stanley CE, Rzepiela A, van Swaay D, Sabotič J, Nørrelykke SF, deMello AJ, Aebi M, Künzler M. 2019. Bidirectional propagation of signals and nutrients in fungal networks via specialized hyphae. Curr Biol 29:217–28. [CrossRef]
- Schuster M, Kilaru S, Wösten HAB, Steinberg G. 2025. Secretion and endocytosis in subapical cells support hyphal tip growth in the fungus Trichoderma reesei. Nat Commun 16:4402. [CrossRef]
- Shepherd VA, Orlovich DA, Ashford AE. 1993a. A dynamic continuum of pleiomorphic tubules and vacuoles in growing hyphae of a fungus. J Cell Sci 104:495–507. [CrossRef]
- Shepherd VA, Orlovich DA, Ashford AE. 1993b. Cell-to-cell transport via motile tubules in growing hyphae of a fungus. J Cell Sci 105:1173–8. [CrossRef]
- Shoji JY, Arioka M, Kitamoto K. 2006a. Possible involvement of pleiomorphic vacuolar networks in nutrient recycling in filamentous fungi. Autophagy 2:226–7. [CrossRef]
- Shoji JY, Arioka M, Kitamoto K. 2006b. Vacuolar membrane dynamics in the filamentous fungus Aspergillus oryzae. Eukaryot Cell 5:411–21. [CrossRef]
- Shoji JY, Craven KD. 2011. Autophagy in basal hyphal compartments: A green strategy of great recyclers. Fungal Biol Rev 25:79–83. [CrossRef]
- Shoji JY, Kikuma T, Kitamoto K. 2014. Vesicle trafficking, organelle functions, and unconventional secretion in fungal physiology and pathogenicity. Curr Opin Microbiol 20:1–9. [CrossRef]
- Skulachev, VP. 2001. Mitochondrial filaments and clusters as intracellular power-transmitting cables. Trends Biochem Sci 26:23–9. [CrossRef]
- Stodulkova E, Císařová I, Kolařík M, Chudíčková M, Novak P, Man P, Kuzma M, Pavlů B, Černý J, Flieger M. 2015. Biologically active metabolites produced by the basidiomycete Quambalaria cyanescens. PLoS One 10:e0118913. [CrossRef]
- Smith SE, Read DJ. 1997. Mycorrhizal symbiosis, 2nd edn. London, UK: Academic Press.
- Steinberg G, Penalva MA, Riquelme M, Wosten HA, Harris SD. 2017. Cell biology of hyphal growth. Microbiol Spectr 5:1–34. [CrossRef]
- Stradalova V, Blazikova M, Grossmann G, Opekarová M, Tanner W, Malinsky J. 2012. Distribution of cortical endoplasmic reticulum determines positioning of endocytic events in yeast plasma membrane. PloS One 7:e35132. [CrossRef]
- Suelmann R, Fischer R. 2000. Mitochondrial movement and morphology depend on an intact actin cytoskeleton in Aspergillus nidulans. Cell Motil Cytoskeleton 45:42–50. [CrossRef]
- Takei K, McPherson PS, Schmid SL, Camilli PD. 1995. Tubular membrane invaginations coated by dynamin rings are induced by GTP-γ S in nerve terminals. Nature 374:186–190. [CrossRef]
- Takeshita N, Manck R, Grün N, de Vega SH and Fischer R. 2014. Interdependence of the actin and the microtubule cytoskeleton during fungal growth. Curr Opin Microb 20:34–41. [CrossRef]
- Takeshita N, Evangelinos M, Zhou L, Serizawa T, Somera-Fajardo RA, Lu L, Takayab N, Nienhaus GU, Fischer R. 2017. Pulses of Ca2+ coordinate actin assembly and exocytosis for stepwise cell extension. PNAS 114:5701–5706. [CrossRef]
- Takeshita, N. 2019. Control of actin and calcium for chitin synthase delivery to the hyphal tip of Aspergillus. In: Latgé, JP. (eds) The Fungal Cell Wall. Current Topics in Microbiology and Immunology. Springer, Cham. 425. [CrossRef]
- Tarutani Y, Ohsumi K, Arioka M, Nakajima H, Kitamoto K. 2001. Cloning and characterization of Aspergillus nidulans vpsA gene which is involved in vacuolar biogenesis. Gene 268:23e30. [CrossRef]
- Thompson W, Eamus D, Jennings DH. 1985. Water flux through mycelium of Serpula lacrimans. Trans Brit Mycol Soc 84:601–8. [CrossRef]
- Timonen S, Finlay RD, Olsson S, Söderström B. 1996. Dynamics of phosphorus translocation in intact ectomycorrhizal systems: non-destructive monitoring using a ß-scanner. FEMS Microbiol Ecol 19:171–180. [CrossRef]
- Tlalka M, Hensman D, Darrah PR, Watkinson SC, Fricker MD. 2003. Noncircadian oscillations in amino acid transport have complementary profiles in assimilatory and foraging hyphae of Phanerochaete velutina. New Phytol 158:325–35. [CrossRef]
- Tlalka M, Bebber DP, Darrah PR, Watkinson SC, Fricker MD. 2008. Quantifying dynamic resource allocation illuminates foraging strategy in Phanerochaete velutina. Fungal Genet Biol 45:1111–1121. [CrossRef]
- Tuszynska S, Davies D, Turnau K, Ashford AE. 2006. Changes in vacuolar and mitochondrial motility and tubularity in response to zinc in a Paxillus involutus isolate from a zinc-rich soil. Fungal Gen Biol 43:155–63. [CrossRef]
- Tuszyńska, S. 2006. Ni2+ induces changes in the morphology of vacuoles, mitochondria and microtubules in Paxillus involutus cells. New Phytologist 169:819–28. [CrossRef]
- Uetake Y, Kojima T, Ezawa T, Saito M. 2002. Extensive tubular vacuole system in an arbuscular mycorrhizal fungus, Gigaspora margarita. New Phytologist 154:761–8. [CrossRef]
- Venables CE, Watkinson SC. 1989. Medium-induced changes in patterns of free and combined amino acids in mycelium of Serpula lacrymans. J General Microbiol 135:1369–1374. [CrossRef]
- Verma S, Shakya VP, Idnurm A. 2018. Exploring and exploiting the connection between mitochondria and the virulence of human pathogenic fungi. Virulence 9:426–46. [CrossRef]
- Veses V, Richards A, Gow NA. 2008. Vacuoles and fungal biology. Curr Opin Microbiol 11:503–10. [CrossRef]
- Voelz K, Johnston SA, Smith LM, Hall RA, Idnurm A, May RC. 2014. ‘Division of labour’in response to host oxidative burst drives a fatal Cryptococcus gattii outbreak. Nat Comm 5:5194. [CrossRef]
- Walter A, Erdmann S, Bocklitz T, Jung EM, Vogler N, Akimov D, Dietzek B, Rösch P, Kothe E, Popp J. 2010. Analysis of the cytochrome distribution via linear and nonlinear Raman spectroscopy. Analyst 135:908–17. [CrossRef]
- Watkinson SC, Boddy L, Burton K, Darrah PR, Eastwood D, Fricker MD, Tlalka M. 2005. New approaches to investigating the function of mycelial networks. Mycologist 19:11–7. [CrossRef]
- Weber RW, Wakley GE, Pitt D. 1998. Histochemical and ultrastructural characterization of fungal mitochondria. Mycologist 12:174–9. [CrossRef]
- Weber RW, Wakley GE, Thines E, Talbot NJ. 2001. The vacuole as central element of the lytic system and sink for lipid droplets in maturing appressoria of Magnaporthe grisea. Protoplasma 216:101–12. [CrossRef]
- Weber, RW. 2002. Vacuoles and the fungal lifestyle. Mycologist 16:10–20. [CrossRef]
- Wedlich-Soldner R, Schulz I, Straube A, Steinberg G. 2002. Dynein supports motility of endoplasmic reticulum in the fungus Ustilago maydis. Mol Biol Cell 13:965–77. [CrossRef]
- West M, Zurek N, Hoenger A, Voeltz GK. 2011. A 3D analysis of yeast ER structure reveals how ER domains are organized by membrane curvature. J Cell Biol 193:333–46. [CrossRef]
- Westermann, B. 2010. Mitochondrial dynamics in model organisms: what yeasts, worms and flies have taught us about fusion and fission of mitochondria. Semin Cell Dev Biol 21:542–9.
- Wright R, Basson M, D’Ari L, Rine J. 1988. Increased amounts of HMG-CoA reductase induce “karmellae”: a proliferation of stacked membrane pairs surrounding the yeast nucleus. J Cell Biol 107:101–114. [CrossRef]
- Zhuang X, Tlalka M, Davies DS, Allaway WG, Watkinson SC, Ashford AE. 2009. Spitzenkörper, vacuoles, ring-like structures, and mitochondria of Phanerochaete velutina hyphal tips visualized with carboxy-DFFDA, CMAC and DiOC6 (3). Mycol Res 113:417–31. [CrossRef]
Figure 1.
Basic variants of four morphotypes of the vacuolar system of apical hyphae in fungi and oomycetes. a. Morphotype S, observed in various oomycetes, is characterized by a slightly mobile tubular vacuolar reticulum that transforms into larger amorphous vacuoles as it moves away from the apex. b. Morphotype G – described in the glomeromycete fungus Gigaspora margarita, is represented by bundles of thin tubular vacuoles and small spherical vacuoles. c. Morphotype P, exemplified by the gasteromycete Pisolithus tinctorius, is common in true fungi. The apical cell contains a zone of highly mobile, well-developed, long, branching tubular vacuoles. Basipetal tubular vacuoles transition into spherical ones, often interconnected by tubular outgrowths. In the more mature region of the hypha, large, low-mobility spherical vacuoles, lacking tubular outgrowths, predominate. d. Morphotype R, found in R. solani, differs from morphotype P by the absence of large spherical vacuoles, a less developed tubular vacuole system in the apical cell, and the presence of an apical vacuolar spindle (a dense bundle of thin tubular vacuoles located in the subapex) in some hyphae. The proportions of hyphae and vacuolar structures in the figures are not strictly observed; the septa are shown schematically, without pores. Clamp connections, where present, are not shown.
Figure 1.
Basic variants of four morphotypes of the vacuolar system of apical hyphae in fungi and oomycetes. a. Morphotype S, observed in various oomycetes, is characterized by a slightly mobile tubular vacuolar reticulum that transforms into larger amorphous vacuoles as it moves away from the apex. b. Morphotype G – described in the glomeromycete fungus Gigaspora margarita, is represented by bundles of thin tubular vacuoles and small spherical vacuoles. c. Morphotype P, exemplified by the gasteromycete Pisolithus tinctorius, is common in true fungi. The apical cell contains a zone of highly mobile, well-developed, long, branching tubular vacuoles. Basipetal tubular vacuoles transition into spherical ones, often interconnected by tubular outgrowths. In the more mature region of the hypha, large, low-mobility spherical vacuoles, lacking tubular outgrowths, predominate. d. Morphotype R, found in R. solani, differs from morphotype P by the absence of large spherical vacuoles, a less developed tubular vacuole system in the apical cell, and the presence of an apical vacuolar spindle (a dense bundle of thin tubular vacuoles located in the subapex) in some hyphae. The proportions of hyphae and vacuolar structures in the figures are not strictly observed; the septa are shown schematically, without pores. Clamp connections, where present, are not shown.
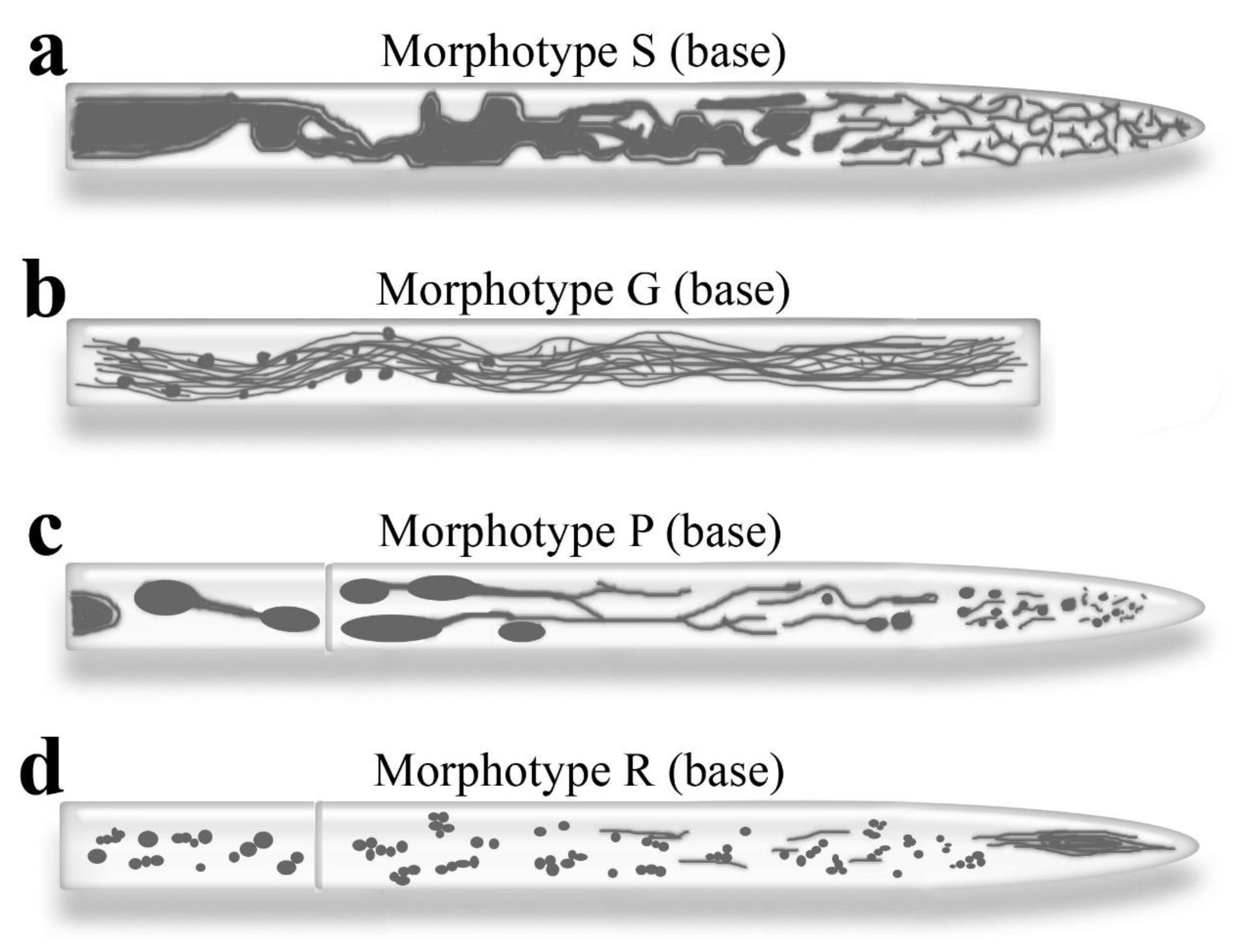
Figure 2.
Different variants of vacuolar morphotypes P and R. а-f. Variants of morphotype P, exemplified by S. hirsutum, differ from the basic variant (b). The differences between some variants are presumably related to the rate of hyphal growth, but other factors may also contribute. a. With a significant growth rate, the apical cell elongates up to 1 mm, and the network of tubular vacuoles develops, occupying almost the entire cell. b. At an average growth rate, vacuole topology corresponds to the canonical Ashford scheme. c. When growth slows, small spherical vacuoles dominate the apical cell. d. A non-growing hypha’s apical cell may contain only non-small, spherical vacuoles. e. The apical hypha may exhibit a complete absence or a small number of labeled vacuoles. f. The inverted hypha is characterized by large, spherical vacuoles at the apex, followed by often well-developed, tubular vacuoles. g. Variants of deviation from the basic variant at the very tip of the hypha. The apex of a xylotrophic basidiomycete (S. hirsutum, P. velutina) is shown on the left. A network of tubular vacuoles fills the hypha’s subapex; individual vacuoles dynamically protrude and reach the apical body zone. The apex of R. solani is shown on the right. A spherical vacuole, located a relative distance from the apex, produces a tubular outgrowth that dynamically grows to the hyphal tip.
Figure 2.
Different variants of vacuolar morphotypes P and R. а-f. Variants of morphotype P, exemplified by S. hirsutum, differ from the basic variant (b). The differences between some variants are presumably related to the rate of hyphal growth, but other factors may also contribute. a. With a significant growth rate, the apical cell elongates up to 1 mm, and the network of tubular vacuoles develops, occupying almost the entire cell. b. At an average growth rate, vacuole topology corresponds to the canonical Ashford scheme. c. When growth slows, small spherical vacuoles dominate the apical cell. d. A non-growing hypha’s apical cell may contain only non-small, spherical vacuoles. e. The apical hypha may exhibit a complete absence or a small number of labeled vacuoles. f. The inverted hypha is characterized by large, spherical vacuoles at the apex, followed by often well-developed, tubular vacuoles. g. Variants of deviation from the basic variant at the very tip of the hypha. The apex of a xylotrophic basidiomycete (S. hirsutum, P. velutina) is shown on the left. A network of tubular vacuoles fills the hypha’s subapex; individual vacuoles dynamically protrude and reach the apical body zone. The apex of R. solani is shown on the right. A spherical vacuole, located a relative distance from the apex, produces a tubular outgrowth that dynamically grows to the hyphal tip.
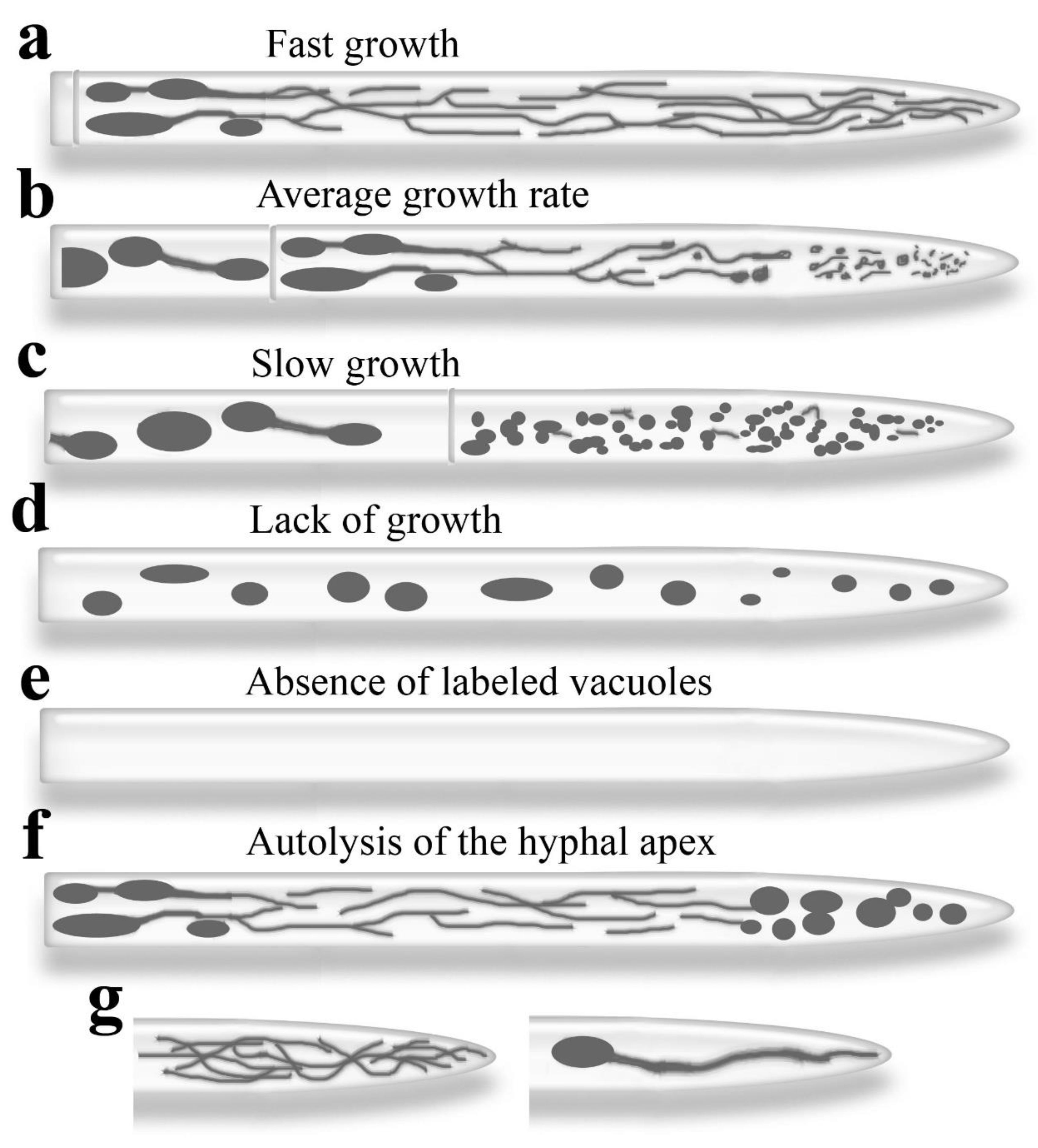
Figure 3.
Vacuolar system of apical hyphae of basidiomycetes, labeled with CFDA. a-c. Apices of three basidiomycete species, S. hirsutum, C. comatus, and R. solani, are shown. The images display the varied expression of the tubular vacuole system, with frequent variants provided for each species. An arrow indicates a dynamically elongated tubular vacuole extending to the hyphal tip. d. Apical cell of R. solani hyphae shows that although tubular vacuoles are less developed in this species than in S. hirsutum, the network of tubular vacuoles can be quite robust in individual hyphae. e. An example of the basic variant of morphotype P in S. hirsutum. f-g. Motile spherical vacuoles in S. hirsutum give rise to long tubular outgrowths and tubular connections between vacuoles. h. Non-motile, spherical vacuoles are present in cells distant from the apex in S. hirsutum. i. Apical cell of R. solani. The vacuolar spindle, characteristic of the morphotype R, is shown (arrow). j. Middle zone of the apical cell of S. hirsutum slow-growing hypha. The zone is filled with small spherical vacuoles, with no tubular vacuoles present. k. Apical cell of S. hirsutum hypha that has stopped growing. There are few tubular vacuoles; the cell contains relatively large spherical vacuoles. l. Inverted hypha of S. hirsutum. The apical part of the hypha is filled with large spherical vacuoles, while tubular vacuoles are visible basipetally. All photographs were obtained by the authors. Scale bar 5 µm.
Figure 3.
Vacuolar system of apical hyphae of basidiomycetes, labeled with CFDA. a-c. Apices of three basidiomycete species, S. hirsutum, C. comatus, and R. solani, are shown. The images display the varied expression of the tubular vacuole system, with frequent variants provided for each species. An arrow indicates a dynamically elongated tubular vacuole extending to the hyphal tip. d. Apical cell of R. solani hyphae shows that although tubular vacuoles are less developed in this species than in S. hirsutum, the network of tubular vacuoles can be quite robust in individual hyphae. e. An example of the basic variant of morphotype P in S. hirsutum. f-g. Motile spherical vacuoles in S. hirsutum give rise to long tubular outgrowths and tubular connections between vacuoles. h. Non-motile, spherical vacuoles are present in cells distant from the apex in S. hirsutum. i. Apical cell of R. solani. The vacuolar spindle, characteristic of the morphotype R, is shown (arrow). j. Middle zone of the apical cell of S. hirsutum slow-growing hypha. The zone is filled with small spherical vacuoles, with no tubular vacuoles present. k. Apical cell of S. hirsutum hypha that has stopped growing. There are few tubular vacuoles; the cell contains relatively large spherical vacuoles. l. Inverted hypha of S. hirsutum. The apical part of the hypha is filled with large spherical vacuoles, while tubular vacuoles are visible basipetally. All photographs were obtained by the authors. Scale bar 5 µm.
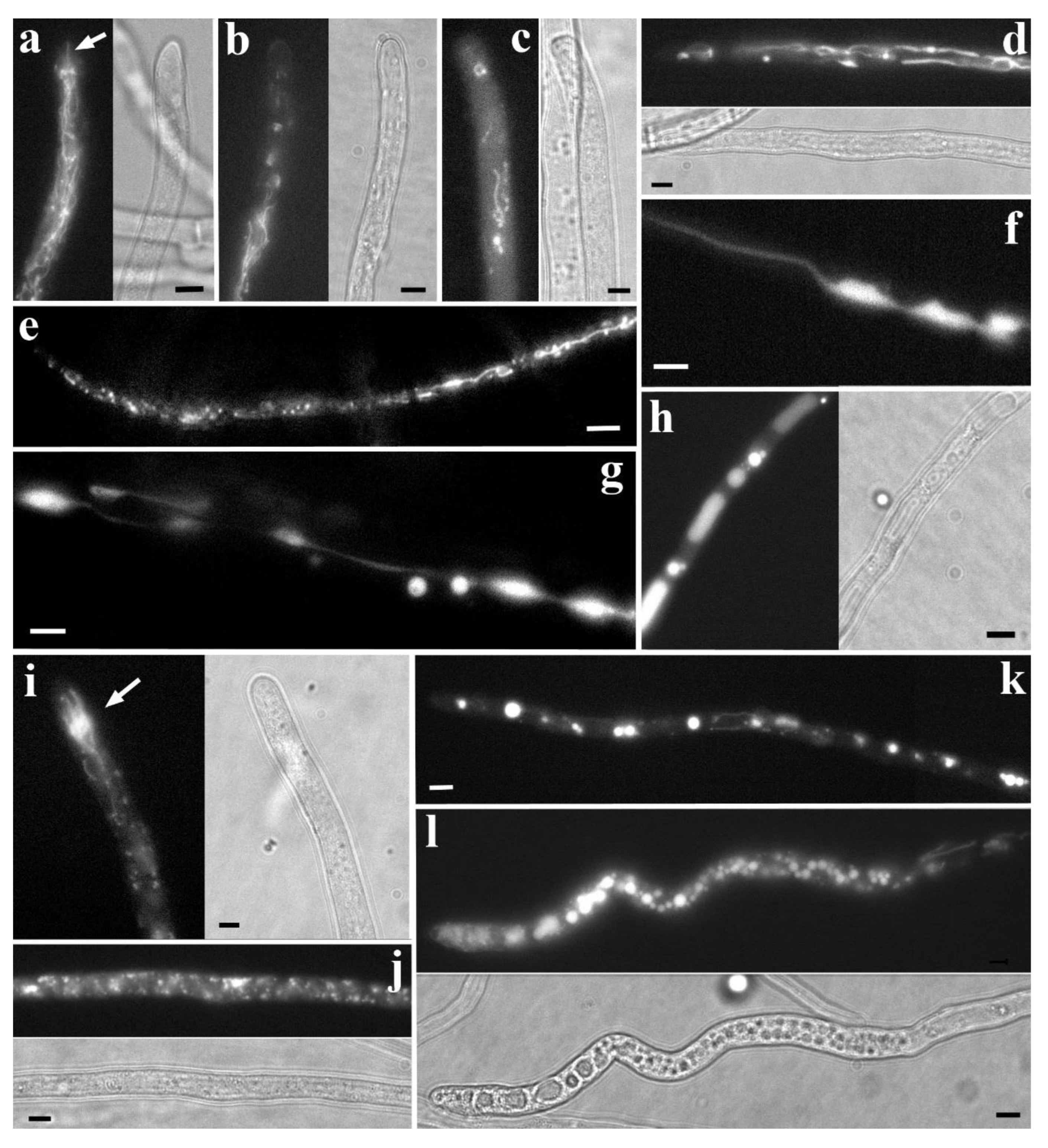
Figure 6.
Three tubular-lamellar endomembrane systems of xylotrophic basidiomycetes as a single intrahyphal complex. a. Schematic representation of the first five cells of the apical hypha of a xylotrophic (mycorrhiza-forming) basidiomycete. The main patterns can be traced: the vacuolar and mitochondrial systems have distinct territories along the hypha’s length. The super-elongated and powerful tubular mitochondria are localized where tubular vacuoles are absent. The endoplasmic reticulum is shown as thin blue threads; unlike mitochondria and vacuoles, however, these are mostly lamellae rather than tubes. Pores and clamp connections are not shown. b, c. Fluorescently labeled mitochondria (R6G, red) and vacuoles (CFDA, green) are visible in subapical cells of S. hirsutum. The two systems are distinct, each occupying a specific side of the hypha. Photographs by the authors. Scale bar is 5 μm.
Figure 6.
Three tubular-lamellar endomembrane systems of xylotrophic basidiomycetes as a single intrahyphal complex. a. Schematic representation of the first five cells of the apical hypha of a xylotrophic (mycorrhiza-forming) basidiomycete. The main patterns can be traced: the vacuolar and mitochondrial systems have distinct territories along the hypha’s length. The super-elongated and powerful tubular mitochondria are localized where tubular vacuoles are absent. The endoplasmic reticulum is shown as thin blue threads; unlike mitochondria and vacuoles, however, these are mostly lamellae rather than tubes. Pores and clamp connections are not shown. b, c. Fluorescently labeled mitochondria (R6G, red) and vacuoles (CFDA, green) are visible in subapical cells of S. hirsutum. The two systems are distinct, each occupying a specific side of the hypha. Photographs by the authors. Scale bar is 5 μm.
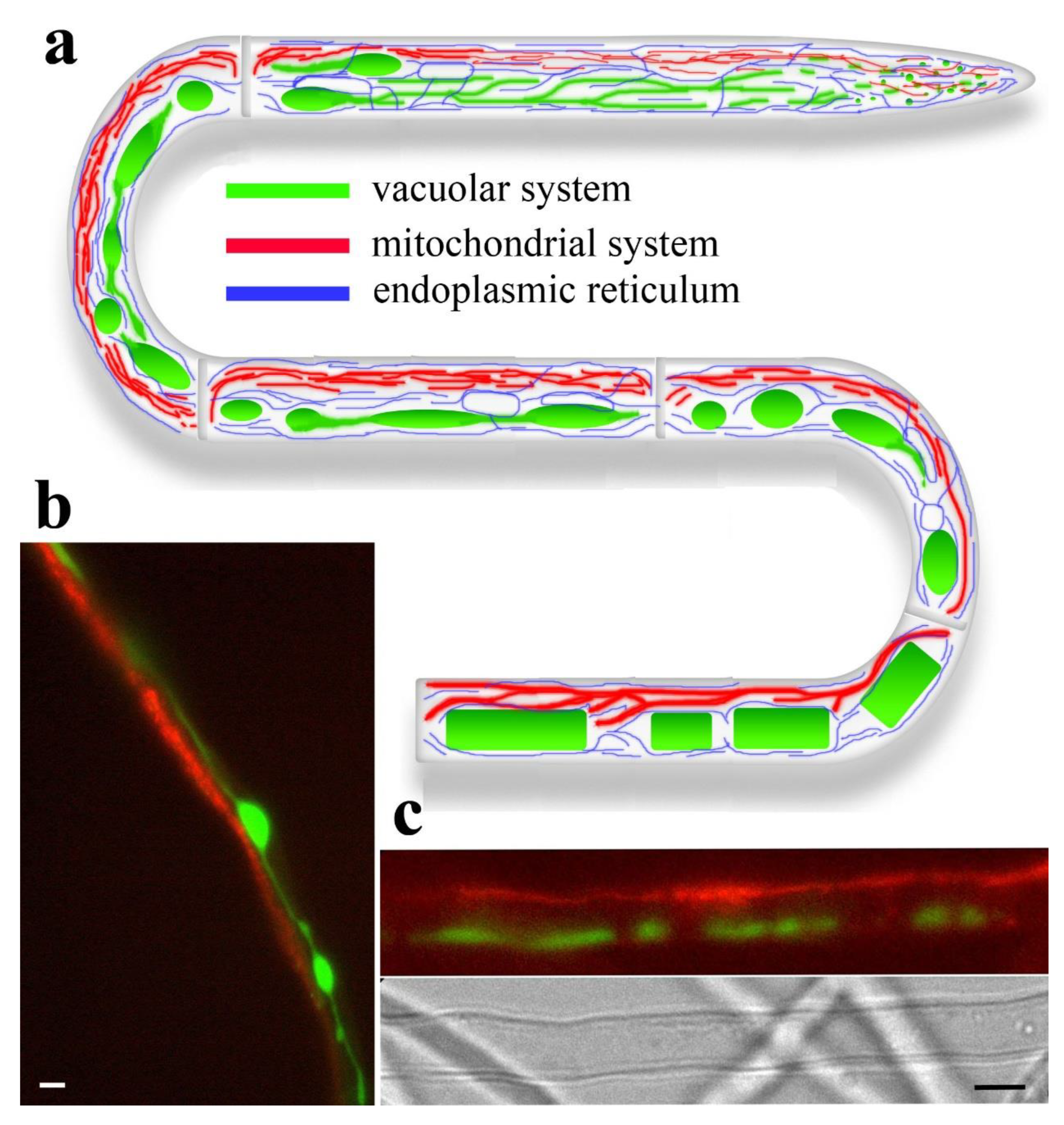
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



