
Submitted:
15 August 2025
Posted:
18 August 2025
You are already at the latest version
Abstract
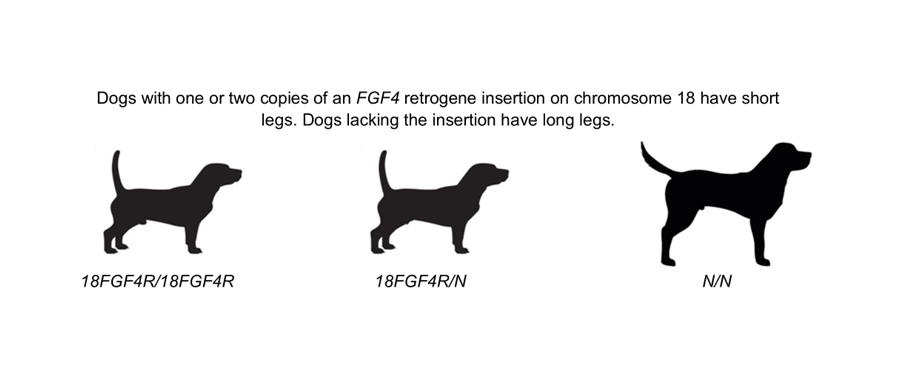
Every dog breed has characteristic physical features that have been fixed through numerous generations of selective breeding. For the Grand Basset Griffon Vendéen (GBGV) breed, short-leggedness has been established as the breed standard. Although breeders have been consistent in only breeding dogs that meet this standard, occasionally litters have included dogs that are significantly taller. In many dog breeds short leg length is associated with insertion of an FGF4 retrogene (FGF4R) at specific sites on either chromosome 12 (12-FGF4R) or 18 (12-FGF4R). A study was undertaken to determine whether FGF4R genotype is the basis of the height difference. Of 22 GBGVs that were evaluated, 18 had leg lengths within the breed standard and 4 were significantly taller. None of the dogs had the 12-FGF4R insertion. The short-legged dogs were either homozygous or heterozygous for the 18-FGF4R insertion. All 4 long-legged dogs were homozygous for the non-insertion allele. This indicates that selective breeding based on phenotype alone has not been sufficient to fix short-leggedness in the breed due to retention of the non-insertion allele. The finding that one copy of the insertion is sufficient to confer short-leggedness indicates that expression of the retrogene during development is likely to inhibit growth of the limbs.
Keywords:
canine
; retrogene
; leg length
; trait selection
; fibroblast growth factor
1. Introduction
Each recognized dog breed has a set of physical characteristics that have been designated as the breed standard by national and international breed organizations. Among these characteristics is height as determined primarily by leg length. A number of breeds are characterized by short stature (short-leggedness) including Dachshunds, Corgis, and Basset Hounds. Among these short-legged breeds is the Grand Basset Griffon Vendéen (GBGV). The American Kennel Club standard for this breed specifies a height of 15 to 18 inches (38 to 46 cm). The GBGV and a smaller Petit Basset Griffon Vendéen (PBGV) breed were derived from common long-legged ancestors. Litters of the ancestral breed included smaller offspring called Petit and larger dogs called Grande [1]. Subsequent selective breeding led to the establishment of the two separate breeds, both of which have short legs relative to the related Griffon Vendéen and Grande Griffon Vendéen breeds. Canine breed-specific traits, including those for the Griffon Vendéen group of dogs, have been developed over many generations of selection for desired phenotypic features, including size [2,3,4]. Although breed standards for physical attributes continue to be based primarily on selecting dogs for breeding that exhibit the desired phenotypic features, advances in canine molecular genetics are starting to expand knowledge of the linkages between DNA sequence variants and specific phenotypic characteristics [3]. This knowledge will enable selection for specific phenotypic features based on DNA sequence analyses. Despite many generations of breeding in which the short-legged phenotype was selected for, a number of GBGV dogs that are significantly taller than the breed standard have been born recently to parents that are within the breed standard for height. Short-leggedness in a number of other breeds has been associated with FGF4 retrogene insertions on chromosomes 12 (12-FGF4R) and 18 (18-FGF4R) [1,5,6,7]. In a 2009 survey of over 70 breeds, 3 of 4 GBGVs evaluated were heterozygous for the 18-FGF4R insertion and one was homozygous for the insertion. This suggested that the presence of this insertion may be a determining factor in leg length in GBGVs. Therefore, a study was undertaken to determine whether differences in the FGF4R insertion genotypes were responsible for the significant height differences among GBGVs.
2. Materials and Methods
2.1. Subject Dogs
GBGVs born between 2010 and 2023 were evaluated in this study. Dog stature, pedigree information, and DNA samples from purebred GBGV dogs were provided by the dog owners and breeders. Among the GBGVs included in the study were littermates of both short and tall stature, as well as their parents. Samples from additional unrelated tall and short GBGVs were also obtained. Stature of the subject GBGVs (height measured at the withers) was provided by the owners. Dogs with heights of 38 to 46 cm were classified as short-legged (the breed standard) and dogs taller than 56 cm were classified as long-legged. All dogs fell into one of these two size classes. DNA samples from unrelated Dachshunds with short legs characteristic of the breed were also utilized as well as samples and phenotype information on dogs of other breeds. All samples and phenotype information were provided by the owners with informed consent. The study protocols were approved by the University of Missouri Animal Care and Use Committee.
2.2. Molecular Genetics
DNA from long-legged and short-legged Grand Basset Griffon Vendéen dogs, as well as from dogs of three additional breeds used as controls, was isolated from EDTA anticoagulated blood or FTA Elute cards as previously described [8,9]. A total of 22 GBGVs were evaluated for the presence of the FGF4R insertions. Eighteen of the dogs were within the breed standard 38 to 46 cm height and four were at least 58 cm tall (Figure 1). All four of the long-legged dogs were born to parents that had the short legs characteristic of the breed. Litters born to these parents included both tall and short-legged dogs (Figure 1 and Figure 2). The long-legged dogs were from 3 different sets of parents. DNA samples were genotyped for the FGF4R chromosome 12 and 18 insertions using 3-Primer PCR amplification. Flanking primers were used in separate reactions for each insertion site as previously described [7]. In each assay, one pair of the primers flanking the insertion site amplified the non-insertion allele. A third FGF4R internal primer was paired with one of the flanking primers to amplify a segment of the FGF4R insertion allele. This 3-primer assay generated different product sizes depending on whether the retrogene insertion was absent or present. The PCR products were visualized using a microcapillary system (QIAxcel, QIAGEN N.V., Venlo, The Netherlands). Sanger sequencing was used to confirm the identities of the amplicons.
Figure 1.
Littermate GBGVs. A dog that had the leg length typical of the breed standard (left) was homozygous for the 18-FGFR insertion (I/I). Its much taller littermate (right) was homozygous for the allele lacking the insertion (N/N).
Figure 1.
Littermate GBGVs. A dog that had the leg length typical of the breed standard (left) was homozygous for the 18-FGFR insertion (I/I). Its much taller littermate (right) was homozygous for the allele lacking the insertion (N/N).

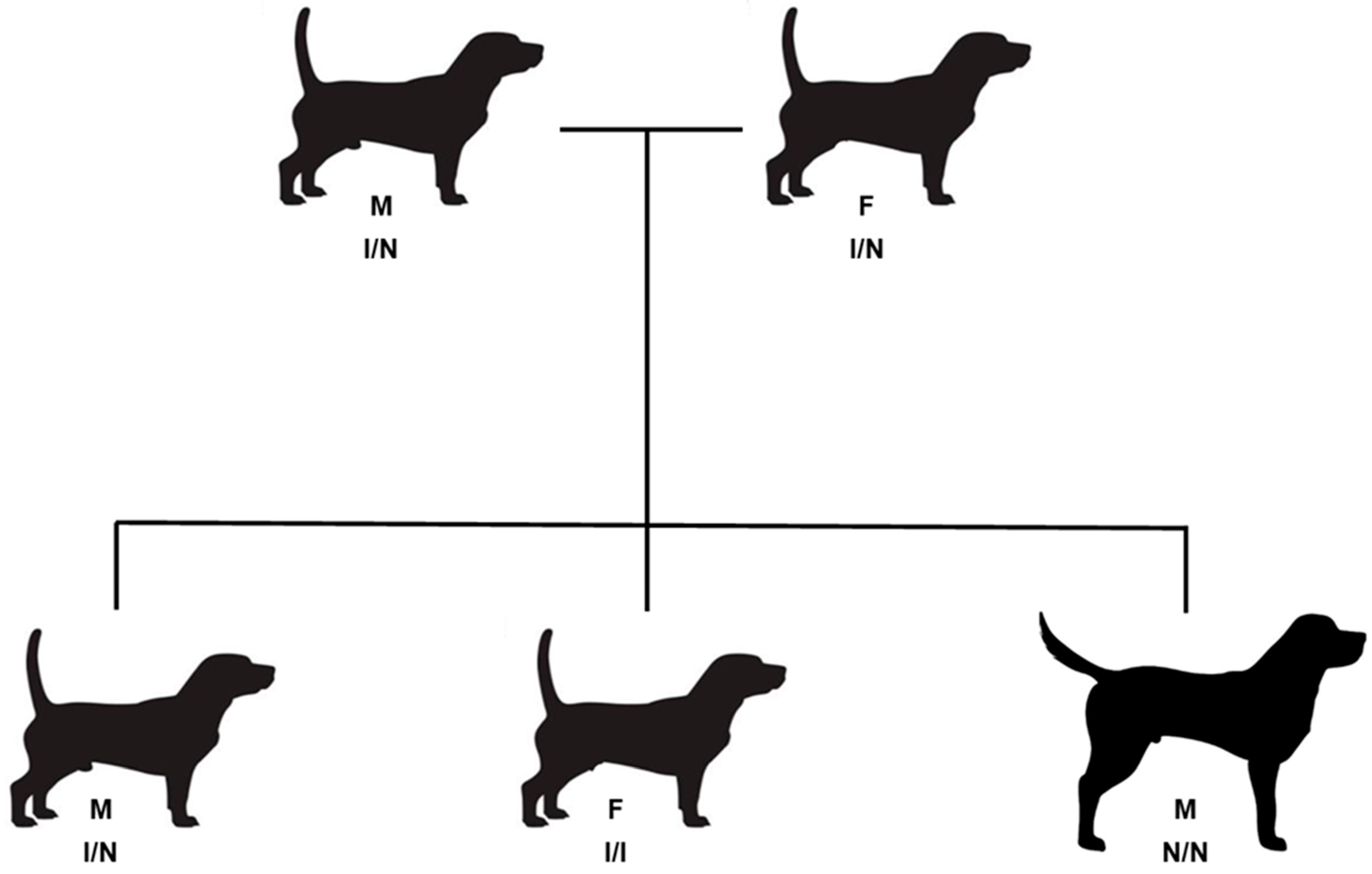
Figure 2.
Partial GBGV pedigree that included littermates with both long and short legs. Both the female (F) and male (M) parents had the short legs characteristic of the breed and were heterozygous for the 18-FGF4R insertion (I/N). Offspring included puppies that were short-legged and long-legged. The short-legged dogs were either homozygous for the 18-FGF4R insertion (I/I) or heterozygous (I/N). The long-legged dog was homozygous for the allele lacking the insertion. All four short-legged dogs were between 39 and 45 cm tall at the withers, whereas the long-legged dog was 60 cm tall.
Figure 2.
Partial GBGV pedigree that included littermates with both long and short legs. Both the female (F) and male (M) parents had the short legs characteristic of the breed and were heterozygous for the 18-FGF4R insertion (I/N). Offspring included puppies that were short-legged and long-legged. The short-legged dogs were either homozygous for the 18-FGF4R insertion (I/I) or heterozygous (I/N). The long-legged dog was homozygous for the allele lacking the insertion. All four short-legged dogs were between 39 and 45 cm tall at the withers, whereas the long-legged dog was 60 cm tall.
2.3. Statistical Analysis
Dog heights measured at the withers were compared between the long-legged and short-legged groups with a t-test. Pearson chi square analysis was performed to assess the association between FGF4R genotypes and leg length phenotype. For these analyses, dogs of both sexes were pooled. Dogs were each assigned to one of six groups for the statistical analyses: short-legged/homozygous for the insertion allele; short-legged/heterozygous the insertion allele; short-legged/homozygous for the non-insertion allele; long-legged/homozygous for the insertion allele; long-legged/heterozygous the insertion allele; long-legged/homozygous for the non-insertion allele.
3. Results
Dogs evaluated in the study fell into two height classes; short-legged dogs were an average of 41 + 2 cm tall and long-legged dogs were an average of 59 + 1 cm tall (difference p<0.0001). Of the 18 GBGVs that had short legs, 5 were homozygous for the chromosome 18 non-insertion allele and 13 were heterozygous for the 18-FGF4R insertion. There was no difference in the average height between the heterozygous and homozygous dogs. All 4 of the tall dogs were homozygous for the for the non-insertion allele (Table 1). The association between 18-FGF4R genotype and leg length phenotype was statistically significant (p<0.01). None of the GBGVs that were evaluated had the 12-FGF4 insertion that is common in other breeds with short legs. A short-legged Dachshund was homozygous for the 12-FGF4 insertion, a Basset Hound was heterozygous, and a Great Dane was homozygous for the chromosome 12 non-insertion allele.

Table 1.
Association between 18-FGF4R insertion genotype and height phenotype in GBGVs.
| 18-FGF4R Insertion | Short | Tall |
|---|---|---|
| Insertion/Insertion | 5 | 0 |
| Insertion/No Insertion | 13 | 0 |
| No Insertion/No Insertion | 0 | 4 |
4. Discussion
The domestication of dogs began over 30,000 years ago with the association between wolves and human hunter-gatherers [10,11]. Early breeding selection was based primarily on behavioral traits that fostered compatibility between ancient dogs and their human companions. Natural variation and selective breeding led to the development of distinct breeds at least as long as 9,500 years ago [11,12]. Over the millennia since their first domestication, numerous other dog breeds were established by selectively breeding dogs with distinct phenotypic features that appeared as a result of natural variation. In the modern era breed standards based primarily on physical features have become formalized by dog breed organizations. Although variation between breeds is considered desirable, uniformity within a breed is considered the ideal. Among dog fanciers, dogs that most closely meet the breed physical standards are used most widely as breeders. Modern breeding practices still rely primarily on phenotypic selection. However, with advances in molecular genetic analyses, DNA sequence variants associated with many specific phenotypic traits have been identified, enabling the use of DNA sequence analysis to guide breeding selection.
In an individual dog with generally desirable phenotypic features, an undesired natural phenotypic variant of a particular trait may occur. In such instances, a breeder may wish to preserve the desirable phenotypic features but select against the single undesired variant feature. For GBGVs, breeding selection is performed to maintain the short-legged phenotype. If selective breeding is done based on phenotype alone, breeders may elect to discontinue breeding dogs that have produced long-legged offspring. This would constrict the breeding population resulting in increased risk of homozygosity for deleterious genetic variants. Now, with the ability to screen for the 18-FGF4R insertion, breeders can avoid producing long-legged puppies by breeding dogs that are heterozygous to dogs that are homozygous for the insertion or by breeding long-legged dogs to dogs that are homozygous for the insertion. By increasing the options for selecting breeding pairs, this will enable maintaining the short-legged phenotype while reducing the risk of producing puppies with recessive genetic disorders.
18-FGF4R retrogene insertion is associated with short stature (short-leggedness) in multiple other dog breeds [1]. In most of the breeds characterized by short legs, all of the dogs evaluated were homozygous for the 18-FGF4R insertion. On the other hand, the insertion allele was not present in tall (long-legged) breeds. In one publication, among the short-legged breeds in which all dogs evaluated were homozygous for the insertion allele were Basset Hound, Cardigan and Pembroke Welsh Corgi, Dachshund, Pekingese, Petit Basset Griffon Vendéen (PBGV), Shi Tzu, Skye Terrier, Swedish Valhund, Tibetan Spaniel, and West Highland Terrier (1). Homozygosity for the 18-FGF4R insertion has also been reported in other short-legged breeds including Cairn Terrier and Coton de Tulear, as well as in short-legged mixed breed dogs [7]. Although there is an association of 18-FGF4R insertion genotype and leg length among many breeds, this alone does not establish a cause and effect relationship. Of four Grande Basset Griffon Vendéen (GBGV) that were evaluated previously, three were heterozygous for the 18-FGF4R insertion, and one was homozygous for the insertion [1]. This suggested that if the 18-FGF4R insertion was responsible for the short-legged phenotype, one copy of the retrogene insertion was sufficient to confer short-leggedness. However, because no dogs that were either long-legged or lacking the insertion were identified, the data did not establish that the presence of the insertion was responsible for short-leggedness in this breed. We found that among 22 GBGVs born between 2010 and 2023, all 4 that were long-legged lacked the 18-FGF4R insertion, whereas all of the dogs with short legs had either one or two copies of the insertion. This makes it highly probable that the presence of either one or two copies of the insertion conferred the short-legged phenotype.
The finding that only 5 of the 22 GBGVs that were evaluated were homozygous for the 18-FGF4R insertion indicates that despite many generations of breeding in which the short-legged phenotype was selected for, the genetic variant responsible for long-leggedness was still common in the breed. This is due in large part to the fact that both dogs homozygous and heterozygous for the insertion exhibit the short-legged phenotype. Now that it has been established that the long-legged phenotype in GBGVs is due to lack of the 18-FGF4R insertion, breeders can utilize genotyping for the insertion allele to minimize the frequency with which long-legged puppies are produced.
Long-legged variants associated with the absence of the 18-FGF4R insertion may also appear in other breeds that normally have short legs. For example, it was recently reported that two Wirehaired Dachshund littermates with abnormally long legs were born to parents with the short-legged phenotype typical of the breed [6]. Both long-legged dogs lacked the 18-FGF4R insertion, whereas both parents and a short-legged littermate were heterozygous for the insertion. The Dachshund group of breeds have been established for centuries, and the short-legged phenotype associated with the 18-FGF4R insertion appears to be fixed in the breed. Thus, the appearance of Dachshunds lacking the insertion suggests that retrogene can be lost even in breeds where it has been maintained for many generations. Therefore, in many breeds characterized by short-leggedness, the rare appearance of long-legged puppies is likely to be due to loss of the 18-FGF4R insertion in prior generations.
In addition to insertion of the FGF4R retrogene on chromosome 18, many dog breeds have an FGF4R retrogene insertion on chromosome 12 [7,13,14]. The 12-FGF4R insertion is associated with intervertebral disc disease [13] and in at least some of breeds, it has also been found to be associated with height (leg length). For example, among Nova Scotia Duck Tolling Retrievers dogs homozygous for the 12-FGF4R insertion were significantly shorter than dogs lacking the insertion, whereas heterozygous dogs were of intermediate height [7]. None of the GBGVs evaluated for the current study harbored the 12-FGF4R insertion, so it appears that the height differences were due exclusively to differences in 18-FGF4R genotype.
In addition to FGF4R, numerous other retrogene insertions have been identified in the canine genome. For example, 3,892 candidate retrocopies from 1,316 parental genes were identified in the CanFam4 genome assembly [15], and 3,025 retrocopies including 476 intact retrogenes, 2,518 retropseudogenes, and 31 chimerical retrogenes were identified in the CanFam3.1 genome assembly [16]. Unlike the FGF4 retrogenes, the potential phenotypic effects of most of these retrogenes, if any, are unknown. However, in human subjects retrogene expression has been associated with a number of hereditary disorders [17,18]. Thus, it seems likely that at least some of the other retrogenes in the canine genome are expressed and have phenotypic effects. Over 90% of the retrocopy insertions in the CanFam4 were also present in the wolf genome assembly, suggesting that they arose long ago. Based on the number of generations since genome divergence, it was estimated that new retrocopy insertions appear, on average, in 1 out of 3,514 births. Like the FGF4 retrogene, formation of these retrogenes can lead to new phenotypic features that can be benign, beneficial, or pathogenic (16).
The mechanism by which the presence of the 18-FGF4R insertion confers short-leggedness has not been elucidated, but it has been established in other dog breeds that this retrogene is expressed, and it has been hypothesized that abnormal production of fibroblast growth factor 4 (FGF4) in chondrocytes during development impairs limb elongation [1]. Consistent with this hypothesis, it has been demonstrated that FGF4 plays a role in axial elongation during development in mice [19]. Because the FGF4 retrogene does not contain the introns of the parent gene and because its location in the genome is different from that of the parent gene, it is unlikely that expression of the retrogene is under the same regulatory control as the parent gene. Aberrantly regulated expression of FGF4 from the 18-FGF4R retrogene during development is therefore likely to account for the short-leggedness in dogs. It is interesting that no other phenotypic effects of the retrogene expression have been recognized. In contrast to the 18-FGF4R insertion, an FGF4R retrogene insertion on canine chromosome 12 is associated not only with leg length, but has also been associated with intervertebral disc disease [7,13,14]. This suggests that regulation of expression of the retrogene varies depending on its location within the genome. It should be possible to investigate this possibility in a cell culture model.
Retrogene insertions can have phenotypic effects even if the retrogene is not expressed. For example, it was found that insertion of a SNNL1 retrocopy on canine chromosome 18 likely disrupted regulation of a nearby gene involved in coat color determination in Poodles [20]. Although the roles of most retrogenes, if any, remain to be elucidated, analysis of retrogenes across multiple species indicates that they are subject to natural selection, and those that survive over the course of evolution are likely to be involved in adaptation [21]. The situation with 18-FGF4R is a case where retrogene survival is determined primarily by human-directed selection rather than natural selection. In the future, identification of the effects of other retrogene insertions on canine phenotypes can be employed to select for desired traits or eliminate undesired traits, including some hereditary disorders. This applies to other species, including humans.
5. Conclusions
In summary, the 18-FGF4R is the major determinant of height (leg length) in GBGVs as it is in other short-legged dog breeds. Expression of the retrogene during development is likely responsible for the short-legged phenotype in dogs with one or two copies of the insertion. The appearance of long-legged variants in many short-legged breeds is most likely due to the absence or altered expression of the retrogene in these dogs. In order to avoid generating dogs taller than the breed standard, dogs can be genotyped for the insertion allele prior to breeding rather than relying on phenotypic selection alone, which does not distinguish between dogs heterozygous or homozygous for the insertion allele. For other short-legged breeds with the 18-FGF4R insertion, general screening is not necessary if the appearance of long-legged variants within those breeds is a rare occurrence. However, screening of close relatives of any rare long-legged variants would be advisable to minimize propagation of the long-legged trait. Numerous other retrogenes have been identified in dogs that may influence other phenotypic traits. The findings suggest that the potential role of retrogene insertions in modulating many phenotypic traits, not only in dogs, but in other species, warrants greater attention than it has received.
Author Contributions
Conceptualization, M.L.K.; methodology,T.M..; validation, M.L.K. and T.M. formal analysis,. M.L.K. and T.M.; investigation, M.L.K. and L.H. resources, M.L.K..; data curation, T.M. and L.H.; writing—original draft preparation, M.L.K. writing—review and editing, M.L.K.. T.M. and L.H.; visualization, M.L.K.; supervision, M.L.K.; project administration, M.L.K.; funding acquisition, M.L.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded in part by grants from the Grand Basset Griffon Vendéen Club of America and the Orthopedic Foundation for Animals.
Institutional Review Board Statement
The animal study protocol was approved by the Animal Care and Use Committee of the University of Missouri (protocol number 44562, approved 8 January, 2025).
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
Our thanks to the GBGV owners who provided us with DNA samples and height information on their dogs for these studies.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| GBGV | Grand Basset Griffon Vendéen |
| PBGV | Petit Basset Griffon Vendéen |
| EDTA | Ethylene diamine tetra acetic acid |
| PCR | Polymerase chain reaction |
| I | Insertion |
| N | Non-insertion |
| FGF4 | fibroblast growth factor 4 |
References
- Parker, H.G.; VonHoldt, B.M.; Quignon, P.; Margulies, E.H.; Shao, S.; Mosher, D.S.; Spady, T.C.; Elkahloun, A.; Cargill, M.; Jones, P.G.; et al. An Expressed Fgf4 Retrogene Is Associated with Breed-Defining Chondrodysplasia in Domestic Dogs. Science 2009, 325, 995–998. [Google Scholar] [CrossRef] [PubMed]
- Helmink, S.K.; Shanks, R.D.; Leighton, E.A. Investigation of Breeding Strategies to Increase the Probability That German Shepherd Dog and Labrador Retriever Dog Guides Would Attain Optimum Size. J Anim Sci 2003, 81, 2950–2958. [Google Scholar] [CrossRef]
- Chen, F.L.; Zimmermann, M.; Hekman, J.P.; Lord, K.A.; Logan, B.; Russenberger, J.; Leighton, E.A.; Karlsson, E.K. Advancing Genetic Selection and Behavioral Genomics of Working Dogs Through Collaborative Science. Front Vet Sci 2021, 8, 662429. [Google Scholar] [CrossRef]
- Hecht, E.E.; Smaers, J.B.; Dunn, W.D.; Kent, M.; Preuss, T.M.; Gutman, D.A. Significant Neuroanatomical Variation Among Domestic Dog Breeds. J Neurosci 2019, 39, 7748–7758. [Google Scholar] [CrossRef]
- Bannasch, D.; Batcher, K.; Leuthard, F.; Bannasch, M.; Hug, P.; Marcellin-Little, D.J.; Dickinson, P.J.; Drogemuller, M.; Drogemuller, C.; Leeb, T. The Effects of <ovid:I>FGF4</Ovid:I> Retrogenes on Canine Morphology. Genes (Basel) 2022, 13. [Google Scholar] [CrossRef]
- Sullivan, S.; Szeremeta, K.J.; Kutzler, M. Case Report: FGF4L1 Retrogene Insertion Is Lacking in the Tall Dachshund Phenotype. Front Vet Sci 2024, 11, 1522745. [Google Scholar] [CrossRef]
- Brown, E.A.; Dickinson, P.J.; Mansour, T.; Sturges, B.K.; Aguilar, M.; Young, A.E.; Korff, C.; Lind, J.; Ettinger, C.L.; Varon, S.; et al. FGF4 Retrogene on CFA12 Is Responsible for Chondrodystrophy and Intervertebral Disc Disease in Dogs. Proc Natl Acad Sci U S A 2017, 114, 11476–11481. [Google Scholar] [CrossRef]
- Katz, M.L.; Khan, S.; Awano, T.; Shahid, S.A.; Siakotos, A.N.; Johnson, G.S. A Mutation in the CLN8 Gene in English Setter Dogs with Neuronal Ceroid-Lipofuscinosis. Biochem Biophys Res Commun 2005, 327. [Google Scholar] [CrossRef]
- Zeng, R.; Coates, J.R.; Johnson, G.C.; Hansen, L.; Awano, T.; Kolicheski, A.; Ivansson, E.; Perloski, M.; Lindblad-Toh, K.; O’Brien, D.P.; et al. Breed Distribution of SOD1 Alleles Previously Associated with Canine Degenerative Myelopathy. J Vet Intern Med 2014, 28. [Google Scholar] [CrossRef]
- Thalmann, O.; Shapiro, B.; Cui, P.; Schuenemann, V.J.; Sawyer, S.K.; Greenfield, D.L.; Germonpre, M.B.; Sablin, M.V.; Lopez-Giraldez, F.; Domingo-Roura, X.; et al. Complete Mitochondrial Genomes of Ancient Canids Suggest a European Origin of Domestic Dogs. Science 2013, 342, 871–874. [Google Scholar] [CrossRef]
- Ni Leathlobhair, M.; Perri, A.R.; Irving-Pease, E.K.; Witt, K.E.; Linderholm, A.; Haile, J.; Lebrasseur, O.; Ameen, C.; Blick, J.; Boyko, A.R.; et al. The Evolutionary History of Dogs in the Americas. Science 2018, 361, 81–85. [Google Scholar] [CrossRef]
- Sinding, M.-H.S.; Gopalakrishnan, S.; Ramos-Madrigal, J.; de Manuel, M.; Pitulko, V.V.; Kuderna, L.; Feuerborn, T.R.; Frantz, L.A.F.; Vieira, F.G.; Niemann, J.; et al. Arctic-Adapted Dogs Emerged at the Pleistocene-Holocene Transition. Science 2020, 368, 1495–1499. [Google Scholar] [CrossRef]
- Batcher, K.; Dickinson, P.; Giuffrida, M.; Sturges, B.; Vernau, K.; Knipe, M.; Rasouliha, S.H.; Drögemüller, C.; Leeb, T.; Maciejczyk, K.; et al. Phenotypic Effects of FGF4 Retrogenes on Intervertebral Disc Disease in Dogs. Genes (Basel) 2019, 10. [Google Scholar] [CrossRef]
- Reunanen, V.L.J.; Jokinen, T.S.; Lilja-Maula, L.; Hytonen, M.K.; Lappalainen, A.K. Allelic Frequency of 12-FGF4RG and the Association between the Genotype with Number of Calcified Intervertebral Discs Visible on Radiographs in Coton de Tulear and French Bulldog Breeds. BMC Vet Res 2025, 21, 140. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, A.K.; Blacksmith, M.S.; Kidd, J.M. Duplications and Retrogenes Are Numerous and Widespread in Modern Canine Genomic Assemblies. Genome Biol Evol 2024, 16. [Google Scholar] [CrossRef]
- Gao, X.; Li, Y.; Adetula, A.A.; Wu, Y.; Chen, H. Analysis of New Retrogenes Provides Insight into Dog Adaptive Evolution. Ecol Evol 2019, 9, 11185–11197. [Google Scholar] [CrossRef]
- Ciomborowska-Basheer, J.; Staszak, K.; Kubiak, M.R.; Makalowska, I. Not So Dead Genes-Retrocopies as Regulators of Their Disease-Related Progenitors and Hosts. Cells 2021, 10. [Google Scholar] [CrossRef]
- Vinckenbosch, N.; Dupanloup, I.; Kaessmann, H. Evolutionary Fate of Retroposed Gene Copies in the Human Genome. Proc Natl Acad Sci U S A 2006, 103, 3220–3225. [Google Scholar] [CrossRef]
- Boulet, A.M.; Capecchi, M.R. Signaling by FGF4 and FGF8 Is Required for Axial Elongation of the Mouse Embryo. Dev Biol 2012, 371, 235–245. [Google Scholar] [CrossRef]
- Batcher, K.; Varney, S.; Affolter, V.K.; Friedenberg, S.G.; Bannasch, D. An SNN Retrocopy Insertion Upstream of GPR22 Is Associated with Dark Red Coat Color in Poodles. G3 (Bethesda) 2022, 12. [Google Scholar] [CrossRef]
- Machado, J.P.; Antunes, A. The Genomic Context of Retrocopies Increases Their Chance of Functional Relevancy in Mammals. Genomics 2020, 112, 2410–2417. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



