Submitted:
12 August 2025
Posted:
14 August 2025
You are already at the latest version
Abstract
Orthodontic tooth movement (OTM) is governed by a highly coordinated cascade of cellular and molecular events triggered by mechanical forces. At the heart of this process lies the dynamic interplay between osteoblasts, osteoclasts, periodontal ligament (PDL) cells, and components of the immune system. This review begins with an overview of the key cellular players in OTM and the biological foundations of force-induced bone remodeling, highlighting the critical roles of mechanotransduction, cytokine-mediated signaling, and extracellular matrix turnover. Building upon these principles, we explore the fundamentals of photobiomodulation and its broader clinical applications in dentistry, with particular emphasis on Low-Level Laser Therapy (LLLT). Recent systematic reviews have assessed its clinical value in orthodontics, reporting variable outcomes in terms of accelerating tooth movement and alleviating treatment-related discomfort. A central theme of this review is the emerging recognition of LLLT as a modulator of intercellular cross-talk during OTM. We analyze how photonic energy may interfere with or amplify mechanical stimuli at the cellular level, integrating into mechanosensitive pathways such as MAPK, Wnt/β-catenin, and PI3K/Akt. Special attention is given to the influence of LLLT on reciprocal interactions between osteoblasts, osteoclasts, and the PDL stromal environment, as well as on LLLT-mediated cross-talk between innate and adaptive immune mechanisms under mechanical stress, a dimension of OTM biology that has received limited attention in previous reviews. Additionally, we evaluate the role of LLLT in regulating apoptosis and autophagy, key processes for cellular adaptation in the mechanically active microenvironment. By bridging photonic and mechanical signaling, LLLT may offer a novel strategy for fine-tuning biological remodeling in orthodontics. Recognition of its immunomodulatory capacity could inform targeted, evidence-based protocols that optimize both treatment efficiency and patient outcomes.
Keywords:
orthodontic tooth movement
; low-level laser therapy
; photobiomodulation
; cellular cross-talk
; immunomodulation
; bone remodeling
1. Introduction
Orthodontic tooth movement (OTM) is initiated by the application of mechanical forces to teeth, triggering adaptive responses in the surrounding periodontal ligament (PDL) and alveolar bone. These responses involve structural remodeling that enables the gradual repositioning of teeth within the alveolar socket and represents the essential biological mechanism underlying orthodontic therapy. Achieving efficient and predictable OTM is a major clinical goal, as it contributes to reduced treatment duration, improved patient comfort, and minimization of complications such as root resorption or periodontal tissue damage [1,2].
The biomechanical process of OTM is characterized by distinct cellular activities on opposing sides of the moving tooth. Compression areas favor recruitment and activation of osteoclasts, promoting localized bone resorption, while tensile zones support osteoblastic activity and new bone deposition [3]. This dynamic remodeling is finely regulated by a network of molecular mediators, including various cytokines, growth factors, and lipid-derived molecules, which coordinate the inflammatory and regenerative phases of tissue adaptation [4].
Because the rate of physiologic tooth movement is inherently limitedvarious surgical and nonsurgical interventions have been investigated to accelerate the process. Surgical procedures such as corticotomy and piezocision induce localized bone injury to stimulate remodeling, whereas less invasive approaches aim to activate bone and periodontal cells through physical or biochemical stimulation [5,6]. Among the latter, Low-Level Laser Therapy (LLLT), also termed photobiomodulation, has emerged as a promising adjunct.
LLLT applies light of a specific wavelength that is absorbed by mitochondrial chromophores, enhancing adenosine triphosphate (ATP) synthesis, modulating reactive oxygen species (ROS) levels, and activating intracellular signaling pathways involved in cell proliferation, differentiation, and inflammatory modulation [7,8]. Its effects are influenced by parameters such as wavelength, energy density, and irradiation timing, which determine the balance between stimulatory and inhibitory responses. In orthodontics, LLLT has been proposed to promote the functional activity of both osteoblasts and osteoclasts, support the coupled processes of bone resorption and formation, and alleviate treatment-related discomfort, making it a potentially valuable tool for optimizing clinical outcomes [9].
2. Key Cellular Players in OTM
Osteoblasts, osteoclasts, periodontal ligament (PDL) cells, and immune cells are key participants in OTM, communicating through direct contact and signaling molecules.
2.1. Osteoblasts
Osteoblasts are bone-forming cells derived from mesenchymal stem cells (MSCs) located in bone marrow, periosteum, and endosteum. These cells commit to osteogenesis under mechanical and biochemical cues, instead of differentiating into adipocytes or chondrocytes [10,11]. Their differentiation is directed by Bone Morphogenetic Proteins (BMP-2, BMP-7) and Wingless/Int-1 (Wnt)/β-catenin pathways, which activate transcription factors like Runt-related transcription factor 2 (RUNX2) and osterix (Osx) [12,13,14]. Additional molecules such as fibroblast growth factors (FGFs), insulin-like growth factors (IGFs), and parathyroid hormone (PTH) enhance this osteogenic signaling [15]. Maturing osteoblasts secrete extracellular matrix proteins, primarily type I collagen, along with osteopontin, osteocalcin, and bone sialoprotein [16]. Matrix vesicles released from these cells initiate mineralization through alkaline phosphatase (ALP) and calcium-phosphate nucleation [17].
Importantly, osteoblasts regulate bone resorption by balancing receptor activator of nuclear factor kappa-Β ligand (RANKL) and osteoprotegerin (OPG) expression, tipping the scale between osteoclast activation and inhibition [18]. Mature osteoblasts either undergo apoptosis, become inactive lining cells, or embed in the matrix as osteocytes, mechanosensitive cells crucial for bone homeostasis [19]. These anabolic and regulatory roles underscore their central function in bone remodeling during OTM.
2.2. Osteoclasts
Osteoclasts are multinucleated cells derived from hematopoietic stem cells of the monocyte–macrophage lineage, sharing their origin with immune cells such as macrophages and dendritic cells (DCs) [20,21]. They play a crucial role in bone resorption, essential for skeletal remodeling and calcium homeostasis.
Their differentiation is primarily driven by Macrophage Colony-Stimulating Factor (M-CSF), which binds to c-Fms receptors to promote precursor proliferation and RANK expression, and RANK Ligand (RANKL), produced by osteoblasts, stromal cells, and T cells. RANKL–RANK interaction activates signaling pathways including nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB), Mitogen-Activated Protein Kinase (MAPK), and calcium signaling, leading to activation of nuclear factor of activated T cells cytoplasmic 1 (NFATc1) and the formation of mature, bone-resorbing osteoclasts [22,23,24,25].
Mature osteoclasts adhere to the bone matrix via integrins (e.g., αvβ3), forming a sealed resorption lacuna where they secrete hydrogen ions and proteolytic enzymes like cathepsin K to dissolve both mineral and organic components of bone [26,27]. Beyond resorption, osteoclasts also engage in crosstalk with osteoblasts, contributing to the regulation of bone remodeling [28].
2.3. Periodontal Ligament
The PDL is a specialized connective tissue that anchors the tooth root to alveolar bone, transmitting forces during chewing and orthodontic treatment. In OTM, the PDL senses mechanical stress and triggers cellular responses that drive bone remodeling [29]. Its extracellular matrix, rich in type I collagen, supports diverse cells, including fibroblasts, osteoblasts, osteoclasts, cementoblasts, mesenchymal stromal cells, immune cells, and periodontal ligament stem cells (PDLSCs) with regenerative potential [30]. Under orthodontic force, compressed PDL regions promote osteoclast-mediated resorption, while tension zones stimulate osteoblast-driven bone formation [31]. These localized responses involve changes in blood flow, cell recruitment, and signaling molecule release, allowing the PDL to dynamically adapt for tooth stability and effective orthodontic movement [31,32].
2.4. Immune Cells
The PDL harbors a diverse array of immune cells essential for maintaining tissue homeostasis and orchestrating responses to mechanical stress. Myeloid-derived innate immune cells include macrophages, neutrophils, DCs, and mast cells. These cells are involved in immune surveillance, phagocytosis, antigen presentation, and initiating inflammation via cytokines and alarmins released. Lymphoid innate cells like natural killer (NK) cells, innate lymphoid cells (ILCs), gamma delta T cells, and potentially invariant NK T (iNKT) cells contribute to early immune regulation and tissue remodeling. The adaptive immune compartment comprises CD4⁺ T cells, CD8⁺ cytotoxic T cells, and B cells, which participate in antigen-specific responses, immunoregulation, and bone remodeling through cytokine signaling and RANKL expression [33,34].
In homeostasis, these immune cells help maintain tissue integrity, support resident fibroblasts and stem cells, and ensure immunological balance in the face of constant mechanical challenges. During OTM, they contribute to all phases of bone remodeling and periodontal ligament regeneration [35].
3. Biological and Molecular Basis of Orthodontic Tooth Movement
OTM is a highly orchestrated biological process initiated by the application of mechanical forces to the dentition. These forces induce remodeling of the PDL and alveolar bone through a cascade of cellular and molecular responses [36]. OTM is a temporally dynamic biological process composed of interrelated and overlapping stages, rather than a continuous or linear progression. Each stage is marked by specific histological, molecular, and immunological events, the timing of which may vary depending on individual biological factors such as bone density, systemic health, and the characteristics of the applied mechanical forces [35,36].
The earliest biological responses occur within the first 48 hours following the application of orthodontic force. This initial period is characterized by a sterile inflammatory response triggered by mechanical deformation of the PDL, characterized by an increased blood flow (vasodilation), infiltration of white blood cells, and the release of inflammatory mediators and signaling molecules. These events initiate the early remodeling of periodontal tissues involving a minor displacement of the tooth within its socket [36,37].
The initial mechanical stimulus is sensed by local cells, such as fibroblasts, osteocytes, and endothelial cells, through mechanotransduction, whereby physical forces are translated into intracellular biochemical signals. Key mechanosensors include integrins (e.g., αvβ3 and α5β1), primary cilia, and stretch-activated ion channels such as Yes-associated protein 1 (YAP1), Crk-associated substrate lymphocyte type (CasL), and Piezo1. These receptors initiate downstream intracellular signaling cascades involving focal adhesion kinase (FAK), Src family kinases, MAPKs, and phosphatidylinositol 3-kinase/protein kinase B (PI3K/Akt), NFAT1 (also known as NFATC2), and Ca²⁺ influx, promoting cell survival, proliferation, and differentiation [37,38,39].
In the following days, tooth movement enters a temporary plateau, known as the lag period, as acute inflammation transitions into a chronic phase. Leukocyte migration persists, and structural remodeling begins in the surrounding periodontal tissues. This stage, lasting 20–30 days, may involve hyalinized, necrotic zones within the compressed PDL, which are cleared by macrophages and multinucleated phagocytes. Simultaneously, monocytes differentiate into osteoclast precursors under RANKL signaling. On the tension side, osteogenic differentiation begins with upregulation of RUNX2 and ALP, alongside vascular endothelial growth factor (VEGF)-driven angiogenesis [34,36,40].
Once necrotic zones are cleared, tooth movement accelerates, typically after the second or third week. A new inflammatory flare-up activates macrophages, fibroblasts, osteoblasts, and osteoclasts. On the pressure side, increased RANKL/OPG ratios and elevated matrix metalloproteinase (MMP) activity drive robust osteoclastogenesis. Acidic resorption lacunae enable matrix degradation, while the tension side supports osteoblast differentiation and matrix mineralization [34,36,40].
Compression and tension sides exhibit distinct osteobiological responses. On the pressure side, mechanical loading creates a pro-osteoclastogenic microenvironment. PDL fibroblasts, osteoblasts, and osteocytes upregulate RANKL and M-CSF, promoting osteoclast precursor recruitment and differentiation. Early secretion of IL-1β, TNF-α, and IL-6 amplifies osteoclastogenic signaling. As inflammation progresses, RANKL, primarily expressed by PDLSCs and PDL fibroblasts, osteoblasts, and activated T cells, becomes central to osteoclast maturation, peaking around 24–48 hours post-stimulus [40,41].
COX-2 induction leads to elevated prostaglandin E2 (PGE2), further enhancing RANKL expression and osteoclast activity. COX-2, encoded by PTGS2, promotes PGE2 production, contributing to vascular changes, nociception, and osteoclastic differentiation [42].
Local hypoxia from vascular compression stabilizes hypoxia-inducing factor (HIF)-1α, increasing VEGF to support angiogenesis and osteoclast recruitment. HIF-1α activation by hypoxia and mechanical stress upregulates VEGF, enabling nutrient delivery and precursor migration. This coordination of osteoclastic activity and angiogenesis facilitates timely resorption and repair [43]. ROS, generated via nicotinamide adenine dinucleotide phosphate (NADPH) oxidases, act as secondary messengers reinforcing osteoclastogenic signaling, though excessive ROS may cause tissue damage [44].
Matrix metalloproteinases like MMP-2 and MMP-9 contribute to matrix remodeling by breaking down type IV collagen and other extracellular matrix (ECM) elements, thereby facilitating cell recruitment to resorption sites [45]. The resorptive activity of osteoclasts is tightly regulated by the ratio of RANKL to OPG, a decoy receptor that blocks RANKL-RANK interaction and limits osteoclastogenesis [1,18,46].
Conversely, the tension side is marked by anabolic responses and osteogenesis. Mechanical stretching activates Wnt/β-catenin signaling, which stabilizes β-catenin and upregulates osteoblast-specific transcription factors such as RUNX2 and Osx. Activation by Wnt ligands inhibits GSK-3β, stabilizing β-catenin and promoting transcription of bone-related genes like RUNX2 and OPG [47]. Inhibitory proteins such as sclerostin and Dickkopf-related protein (DKK)-1 are typically downregulated, enabling Wnt-mediated bone formation [45].
Integrin-mediated signaling activates FAK and downstream extracellular signal-regulated kinase (ERK)/MAPK pathways, stimulating osteoblast proliferation and matrix deposition. Mechanical strain triggers ERK1/2 and p38 MAPK cascades in PDL cells, regulating MMP expression and inflammatory responses [48]. In osteoblasts, ERK signaling enhances RUNX2 activation, directly linking mechanotransduction with osteogenic differentiation [18].
Mechanosensitive ion channels increase intracellular calcium, triggering calmodulin-dependent kinases and transcription factors such as NFATc1. PI3K/Akt and calcium-dependent pathways are less characterized in OTM, but available data suggest they promote osteoblast viability and function. Calcium-sensing receptors (CaSR) may also translate ionic shifts into downstream gene activation through phospholipase C (PLC) and intracellular Ca²⁺ mobilization [49,50].
Growth factors such as BMP-2, BMP-7, and transforming growth factor-beta (TGF-β), released in response to mechanical loading and osteoclastic activity, promote osteoblast differentiation, collagen synthesis, and recruitment of osteoprogenitor cells. TGF-β/Smad signaling, activated by matrix-derived TGF-β, supports collagen production and cell proliferation via Smad2/3 phosphorylation, and intersects with MAPK and Wnt pathways to coordinate remodeling [51].
Mechanically induced IGF-1 further supports osteoblast survival and function. BMP-2 and IGF-1, often acting through Wnt signaling, enhance osteoblast proliferation and maturation [18]. On the tension side, gene expression shifts toward osteogenesis and matrix deposition. Transcription factors such as RUNX2 and Osx promote MSC commitment to the osteoblastic lineage. Upregulation of COL1A1 and ALP reflects active matrix production and early mineralization [18,45].
Bone remodeling during OTM is sustained by angiogenesis and increased vascular permeability, essential for nutrient delivery and progenitor cell recruitment. Hypoxia-driven VEGF expression supports vascular adaptation, while transient hypoxia in compressed regions activates HIF-1α, enhancing VEGF and CXCL12 expression to promote vascularization and progenitor cell migration to the remodeling site [52]. Redox balance is maintained by antioxidant enzymes such as superoxide dismutase (SOD) and glutathione peroxidase [53].
In the late phase, tooth movement becomes more stable and continuous. Remodeling is more organized, supported by elevated ALP activity and sustained involvement of bone and connective tissue cells. As the tooth nears its final position and mechanical force diminishes, inflammatory signaling subsides, bone turnover slows, and tissue homeostasis within the periodontium is gradually restored [32,34,36,41].
In summary, OTM is orchestrated by a complex interplay of mechanical stimuli, signaling pathways, cytokines, growth factors, and immune regulation. These spatially and temporally coordinated responses ensure effective and controlled remodeling of the alveolar bone and PDL, enabling successful orthodontic outcomes.
4. Photobiomodulation in Dentistry: Principles and Broader Clinical Utility
Photomodulation (PBM) has found significant application in OTM. PBM, also known as LLLT, involves the application of low-intensity red or near-infrared light to modulate biological responses without causing thermal damage. A variety of light sources have been employed in PBM, including Gallium-Aluminum-Arsenide (GaAlAs) lasers (780–980 nm), Gallium-Arsenide (GaAs) lasers (∼904 nm, pulsed), Helium-Neon (HeNe) lasers (632.8 nm), diode lasers (660–980 nm), and non-coherent Light Emitting Diodes (LEDs). These devices differ in tissue penetration depth, coherence, and clinical utility, making them suitable for a wide range of dental applications [54,55].
In dentistry, PBM is increasingly utilized for its anti-inflammatory, analgesic, and regenerative properties. Its clinical indications extend well beyond orthodontics, including the promotion of soft and hard tissue healing, management of temporomandibular disorders, treatment of oral mucositis, enhancement of periodontal regeneration, and relief of dentinal hypersensitivity [56,57]. PBM improves cellular functions such as osteoblast and fibroblast activity, angiogenesis, and collagen synthesis, while also modulating nociceptive pathways and inflammatory mediators [54,55,56,57,58].
Beyond accelerating tooth movement, PBM offers several additional clinical benefits in orthodontics. It significantly reduces pain, most pronounced within the first 72 hours following force application, by downregulating inflammatory mediators such as PGE2, bradykinin, and substance P. Its analgesic effect and capacity to lower PGE2 levels have been confirmed in multiple studies [59,60,61]. PBM may also help prevent root resorption by suppressing osteoclastic activity, as shown in animal models using LLLT [62].
Following surgical procedures such as corticotomy or rapid maxillary expansion, PBM has been reported to enhance bone regeneration and increase VEGF expression, though findings differ between animal and clinical studies [63,64]. It also improves the stability and osseointegration of TADs, with studies demonstrating greater bone-to-implant contact at laser-treated miniscrew sites [65]. Although much of the current evidence comes from animal models, early human data support these observations, indicating improved temporary anchorage device (TAD) stability and enhanced periodontal healing, particularly in patients with compromised tissue conditions.
5. Summary of Recent Systematic Reviews on the Effects of Low-Level Laser Therapy in Orthodontics
LLLT has gained interest in orthodontics for accelerating tooth movement, shortening treatment time, and reducing pain. Over the past 20–30 years, multiple randomised clinical trials (RCTs) and meta-analyses have explored its efficacy and safety. Doshi-Mehta and Bhad-Patil (2012) reported 30–58% faster canine retraction with LLLT, without increased root or bone resorption [67]. Elgadi et al. (2023) consolidated evidence from systematic reviews and confirmed that LLLT significantly reduces orthodontic treatment time. However, authors emphasize that further histological studies are needed to gain a deeper understanding of the biological effects of the therapy [68].
Zheng et al. (2023) found LLLT shortens leveling and alignment phases by about 30 days, though with moderate study heterogeneity [69]. Bakdach and Haddad (2020) observed significant acceleration but rated evidence as low to very low quality, emphasizing the need for standardized laser dosage reporting and more rigorous trials [70]. Isola et al. (2019) demonstrated, in a split-mouth RCT with diode laser (810 nm, 1 W, 66.7 J/cm²), significant acceleration of maxillary canine distalization and pain reduction [71].
A systematic review and meta-analysis by Yavagal et al. (2021) evaluated PBM’s efficacy in accelerating OTM in children. Among 14 studies reviewed (9 included in meta-analysis), pooled results showed a significant increase in tooth movement rate in PBM groups compared to controls. However, substantial heterogeneity limited the strength of conclusions. Still, PBM was considered a promising, safe, and non-invasive adjunct, with further trials needed to define optimal protocols [72]. Similarly, Inchingolo et al. (2022) reviewed 41 studies on LLLT in orthodontics over the past decade. LLLT was found to promote osteoblast activity in the PDL, potentially enhancing tooth movement, bone regeneration, miniscrew stability, and reducing root resorption. However, evidence on its effectiveness in pain reduction remained inconsistent [73].
Beyond accelerating tooth movement, LLLT has been shown to reduce pain during OTM, likely due to its anti-inflammatory and analgesic effects. A clinical trial conducted by Wu et al. (2018) showed that applying LLLT decreased tooth and gingival pain and sensitivity related to orthodontic treatment and might also produce effects on the opposite side within the trigeminal system [74]. Similar results were observed during the initial stage of OTM [75]. However, a Cochrane review found that while LLLT shortened the alignment phase and reduced appointments, it did not significantly increase tooth movement early on. Minor improvements during space closure and canine retraction were noted but lacked clear clinical significance. Secondary outcomes like root resorption, periodontal health, and pain showed no significant differences, with overall evidence rated low to very low quality [76]. Li et al. (2015) also highlighted methodological flaws and bias in pain management studies, concluding that current evidence is insufficient to recommend LLLT routinely for orthodontic pain relief, emphasizing the need for further high-quality research [77].
LLLT has been investigated for its effects on orthodontically-induced inflammatory root resorption (OIIRR), a common adverse outcome of OTM. A systematic review by Michelogiannakis et al. (2019) analyzed nine studies with conflicting results. Some studies showed LLLT reduced root resorption and had reparative effects post-OTM, while others suggested that LLLT might increase OIIRR under certain conditions. Although study quality ranged from moderate to high, the overall impact of LLLT on OIIRR remains inconclusive, necessitating further research [78].
Despite promising clinical outcomes, LLLT’s clinical adoption in orthodontics faces challenges. Treatment protocols vary widely in laser parameters such as wavelength, power, energy density, exposure time, and frequency, hindering comparison and standardization. Many trials have small samples, short follow-ups, and lack blinding, limiting robustness and generalizability. Methods for assessing tooth movement differ, affecting accuracy. Most mechanistic data come from animal or in vitro studies, with uncertain applicability to humans. LLLT shows a favorable safety profile with minimal adverse effects due to its non-ionizing, low-energy nature, but limited long-term safety data warrant ongoing monitoring [70,73,76].
6. Molecular Aspects of Low-Level Laser Therapy and the Integration of Mechanical and Photon-Induced Signals during Orthodontic Tooth Movement
Mitochondria, particularly the electron transport chain enzyme cytochrome c oxidase (COX or complex IV), are recognized as the primary intracellular targets of LLLT. When red or near-infrared (NIR) light is absorbed by COX, it enhances mitochondrial membrane potential and stimulates ATP synthesis. This is accompanied by a transient increase in ROS, which serve as signaling molecules that can influence various downstream cellular pathways involved in inflammation, differentiation, and tissue remodeling [79,80].
PBM has also been shown to alter mitochondrial morphology and distribution in osteoblastic cell cultures, supporting the idea that mitochondria respond dynamically to light exposure [81]. Beyond COX, PBM may upregulate other key components of the mitochondrial respiratory chain, such as complex I and ATP synthase, thereby further boosting cellular energy metabolism, especially under stress conditions like ischemia or high glucose levels [80,82].
A key aspect of this mechanism involves the dissociation of nitric oxide (NO) from COX. Under physiological stress, excess NO can bind to COX and inhibit respiration; however, PBM can reverse this effect, restoring electron flow and ATP production [83,84]. These mitochondrial changes can trigger broader cellular responses through a mechanism known as retrograde mitochondrial signaling, which can impact gene expression, cytokine production, and cell survival [85,86].
Beyond mitochondrial effects, PBM therapy appears to influence calcium signaling and ion channel activity. Increases in intracellular calcium and cAMP have been observed following PBM, suggesting activation of calcium-permeable channels [85]. In osteoblast-like Saos-2 cells, PBM-induced expression of osteogenic markers such as ALP, osteopontin, and RUNX2 was suppressed when stretch-activated channels (SACs), including TRPC1, were blocked [87]. These findings indicate that calcium influx through mechanosensitive channels is essential for PBM-mediated osteogenic responses, and similar mechanisms have been linked to Akt phosphorylation in MSCs exposed to 635 nm light [88]. PBM also activates TRPV1 channels in mast cells, promoting calcium entry and ATP production, with potential neuroprotective applications [89,90]. While not fully explored, calcium-sensing receptors (CaSR) may also participate in detecting PBM-induced calcium shifts and modulating downstream signaling [91]. Additionally, PBM may exert direct effects on extracellular molecules, including activation of TGF-β1 and SOD production, contributing to stem cell differentiation and tissue regeneration [79,85,92].
The integration of mechanical and photonic stimuli represents a central mechanism for coordinating the biological responses necessary for efficient OTM. Mechanical forces and LLLT activate a spectrum of overlapping intracellular signaling cascades that converge on shared molecular effectors, highlighting a complex cross-talk between mechanotransduction and photobiomodulation.
Mechanical stimuli such as cyclic stretch, fluid shear, or compression are detected via mechanosensitive elements, including integrins, focal adhesion complexes, and ion channels like Piezo1 and TRPC1. These structures initiate downstream pathways, most notably focal adhesion kinase (FAK), Src, MAPK/ERK, and PI3K/Akt, that regulate cellular responses such as proliferation, migration, and differentiation [37,38,39]. Similarly, as already elaborated, LLLT modulates intracellular metabolism and redox state through mitochondrial activation and ROS/Ca²⁺ generation, activating some of the same cascades: MAPK/ERK, PI3K/Akt, and NF-κB [79,85].
The convergence of these pathways is especially evident at the level of ERK1/2 and Akt kinases, which serve as integrators of proliferative and pro-survival signals in both fibroblasts and osteoblasts. Transcriptional modulators such as NF-κB and AP-1 are activated by both mechanical stress and photobiomodulatory inputs, regulating inflammatory mediators (e.g., IL-1β, TNF-α, COX-2) and matrix remodeling enzymes necessary for controlled tissue adaptation during OTM [37,39,79,85,92].
ROS and intracellular calcium (Ca²⁺), which are modulated by both mechanical loading and light exposure, act as key second messengers driving further activation of calcium/calmodulin-dependent protein kinases (CaMKs), calcineurin-NFAT, and redox-sensitive transcription factors [93]. These mechanisms contribute to fine-tuning the signaling dynamics and to synchronizing the temporal sequence of events such as osteoclastogenesis, fibroblast proliferation, and extracellular matrix remodeling [37,39]. Ultimately, this dual activation fosters a harmonized biological response to orthodontic stimuli. Through molecular convergence, cells are equipped to interpret and integrate both mechanical and photonic signals, enhancing tissue regeneration, accelerating bone remodeling, and potentially reducing adverse events like inflammation or root resorption. Therefore, this integrated network of signaling pathways underpins the rationale for using adjunctive LLLT in orthodontics, not merely as a metabolic booster, but as a molecular modulator of biomechanical responses essential for optimal treatment outcomes.
7. Cross-talk Between Osteoblasts, Osteoclasts, and Periodontal Stromal Cells During Orthodontic Tooth Movement: Influence of Low-Level Laser Therapy
During OTM, a dynamic cross-talk between osteoblasts, osteoclasts, osteocytes, PDLSCs, and PDL fibroblasts orchestrates bone remodeling. This cellular interplay differs significantly between the compression and tension sides of the PDL. As already emphasized, PDLSCs upregulate RANKL and downregulate OPG on the compression side, shifting the RANKL/OPG ratio in favor of osteoclastogenesis. In contrast, on the tension side, PDLSCs and fibroblasts promote osteogenic differentiation by activating signaling pathways involving RUNX2, Osx, and BMP-2, leading to osteoblast maturation and extracellular matrix production. Osteoblasts then deposit type I collagen, ALP, and osteocalcin, facilitating new bone formation to accommodate the repositioned tooth [94,95,96].
PDLSCs are positioned at the crossroads of these interactions. Under tensile forces, they differentiate into osteoblasts, while under compressive forces, they promote osteoclastogenesis via RANKL expression. Moreover, their immunomodulatory effects on T cells and macrophages indirectly influence the bone remodeling process [97].
Osteoblasts and osteoclasts interact either through direct physical contact or via the secretion of various signaling molecules. Direct interactions involve receptor–ligand binding and the subsequent activation of intracellular signaling pathways. For instance, ephrin B2 (EFNB2), expressed on the surface of osteoclasts, binds to the EPHB4 receptor on osteoblasts. This interaction activates EPHB4, promoting osteoblast differentiation and inhibiting apoptosis [28,98].
Semaphorin 3A (SEMA3A), produced by osteoblasts, binds to neuropilin-1 (NRP1) on osteoclasts. This binding inhibits RANKL-induced osteoclast differentiation and simultaneously promotes osteoblast differentiation through activation of the WNT/β-catenin pathway. The interaction between the FAS receptor on osteoclasts and its ligand (FASL) on osteoblasts induces osteoclast apoptosis [28,99].
Osteoblasts also secrete several factors that regulate osteoclast development, including M-CSF, RANKL, and WNT5A. M-CSF and RANKL act synergistically to promote osteoclast differentiation, fusion, and activation. RANKL binds to RANK receptors on monocyte-derived precursors, promoting their fusion into multinucleated osteoclasts that express cathepsin K, TRAP, and carbonic anhydrase II, enabling bone resorption [28,40]. WNT5A, which is highly expressed in osteoblast-lineage cells, binds to the receptor tyrosine kinase-like orphan receptor 2 (ROR2) on osteoclasts to further influence their behavior. In contrast, osteoblast-derived OPG and WNT16 inhibit osteoclast activity. OPG acts as a decoy receptor by binding to RANKL, thereby preventing its interaction with RANK and suppressing osteoclastogenesis apoptosis [28]. Conversely, osteoclasts secrete signaling molecules such as sphingosine-1-phosphate (S1P), collagen triple helix repeat containing 1 (CTHRC1), and complement component C3, all of which enhance osteoblast differentiation. Additionally, TGF-β and insulin-like growth factor 1 (IGF-1), released from the bone matrix during osteoclastic bone resorption, stimulate osteoblast-mediated bone formation. In contrast, semaphorin 4D (SEMA4D) suppresses osteoblast differentiation [28].
Beyond these interactions, other bone-related cells also contribute to this regulatory network. Osteocytes produce RANKL, sclerostin (SOST), and prostaglandin E2 (PGE2), while PDLSCs secrete RANKL, PGE2, and IL-6. SOST, a glycoprotein encoded by the SOST gene, binds to low-density lipoprotein receptor-related proteins LRP5 and LRP6, as well as frizzled receptors, to inhibit the WNT signaling pathway. Through this mechanism, SOST acts as a negative regulator of osteoblast differentiation in both osteoblasts and PDLSCs [40].
PGE2 stimulates osteoclastogenesis in areas of compression primarily through the EP4 receptor, which is its dominant receptor subtype in bone. This mechanism is largely mediated by the induction of RANKL expression in both PDLSCs and osteoblasts [40].
MicroRNAs (miRNAs) also play a key role in mediating communication between osteoblasts and osteoclasts during OTM. In general, miRNAs regulate stemness, proliferation, differentiation, and apoptosis by binding to the 3′-untranslated regions (3′-UTRs) of target genes, leading to either mRNA degradation or inhibition of protein translation. Through these mechanisms, miRNAs modulate multiple signaling pathways and transcription factors involved in osteogenic differentiation. These include the TGF-β/BMP pathway, the Wnt/β-catenin pathway, as well as Notch and Hedgehog signaling. Additionally, they influence key transcription factors such as RUNX2 and Osx [100]. It has been shown that miR-3198 is upregulated under compressive force and downregulated under tension, suggesting that it suppresses OPG expression in response to mechanical stress [101]. MiR-223 functions as a negative regulator of osteogenic differentiation in PDL-derived cells by targeting two essential growth factor receptor genes, such as TGFβR2 and fibroblast growth factor receptor 2 (FGFR2). Similarly, Zhang et al. (2021) reported that miR-93-5p enhances cell proliferation while inhibiting osteogenic differentiation in human bone marrow-derived mesenchymal stem cells (BMSCs) [102].
PDLSCs exposed to compressive stress release extracellular vesicles (EVs) containing miRNAs such as miR-28, which directly promote osteoclast differentiation and bone resorption in OTM models [103]. They also secrete exosomes enriched with mechano-sensitive miR-21-5p, which appears to be involved in both osteogenic and osteoclastogenic regulation, although its direct effect on osteoclasts remains to be fully elucidated [104]. Under inflammatory conditions (e.g., stimulation with LPS), PDLSC-derived exosomes exhibit increased levels of miR-155-5p, which may influence the balance between T helper 17 (Th17) cells and regulatory T cells (Tregs), thereby contributing to alveolar bone remodeling [105].
Collectively, these findings suggest that the release of specific miRNA-containing EVs by stromal cells under mechanical and inflammatory stimuli plays a significant role in promoting osteoclastogenesis and bone remodeling during OTM.
LLLT promotes proliferation and osteogenic differentiation of PDLSCs mainly through BMP/Smad signaling. Irradiation with Nd:YAG lasers (1064 nm, 2–6 J/cm²) significantly increased proliferation, ALP activity, and mineralization. These effects were diminished by BMP/Smad inhibition [106]. Although direct evidence for Wnt/β-catenin activation in PDLSCs is limited, BMP–Wnt cross-talk during osteogenesis is well established in MSCs [107]. Another study demonstrated that PBM increases VEGF and BMP expression in PDLSCs, along with enhanced mineralization [108]. Based on a systematic review of in vitro studies, Mylona et al. (2024) reported that PBM within the 630–830 nm wavelength range can enhance both the stemness and differentiation potential of PDLSCs. Regarding fluence, doses should not exceed 4 J/cm² when using laser-based systems [109]. A systematic review by Berni et al. (2023), analyzing in vitro and in vivo models of bone regeneration, concluded that LLLT facilitates angiogenesis, supports fracture repair, and induces osteogenic differentiation in MSCs [110]. Yet, the molecular mechanisms driving these effects during bone formation remain poorly characterized.
LLLT stimulates osteoclastogenesis on the compression side by upregulating RANKL and osteoclast markers. In vitro, Ga–As–Al laser (810 nm) exposure increased NFATc1, TRAP, cathepsin K (CTSK), and phosphorylation of NF-κB and MAPKs in RAW264.7 cells [111]. Similar results have recently been confirmed by Huang et al. (2024) [112]. In vivo rodent OTM models also show increased osteoclast numbers post-LLLT, linked to elevated RANKL and M-CSF from PDL cells [113]. Regarding cytokine production, the results vary. Wang et al. (2022) demonstrated that LLLT stimulates the secretion of TNF-α and IL-1β by PDLSCs, supporting the hypothesis that PBM may accelerate OTM by enhancing the early inflammatory response in the compressed PDL [106]. However, in a rat model of medication-related osteonecrosis of the jaw (MRONJ), LLLT reduced levels of TNF-α and IL-1β, while increasing the expression of IL-1 receptor antagonist (IL-1RA), thereby supporting tissue repair 114 [114]. These effects may contribute to the reparative phase of OTM.
8. Matrix Remodeling and Vascular Response in Orthodontic Tooth Movement Under the Influence of Low-Level Laser Therapy
Orthodontic force application triggers a coordinated reorganization of the periodontal ECM and associated vascular adaptations, both crucial for enabling cellular migration, immune infiltration, bone resorption, and tissue regeneration. These processes are under tight molecular control and differ spatially between tension and compression zones [115].
The ECM of the PDL, composed mainly of type I and III collagen, proteoglycans, and glycoproteins, preserves tissue structure and resilience. During OTM, ECM remodeling facilitates the migration of osteoclasts, fibroblasts, endothelial cells, and immune cells to sites of active bone turnover. This remodeling is driven by MMPs, a family of zinc-dependent enzymes that degrade ECM components. Mechanical stimuli upregulate MMPs such as MMP-1 and MMP-8 (targeting fibrillar collagens), while MMP-2 and MMP-9 degrade gelatin, elastin, and denatured collagens, clearing pathways for cell migration [116].
In vascular smooth muscle cells, LLLT enhances MMP-1 and MMP-2 expression and collagen synthesis [117]. In contrast, during OTM with cortical punching in rats, LLLT upregulated MMP-1, MMP-8, and MMP-13 while decreasing collagen expression in the PDL [118].
MMPs are produced by PDL fibroblasts, osteoclasts, and endothelial cells, with their expression rising rapidly, especially in compression zones, after force application. Their activity is modulated by mechanosensitive signaling pathways like MAPK and NF-κB and counterbalanced by tissue inhibitors of metalloproteinases (TIMPs), particularly TIMP-1 and TIMP-2. The MMP/TIMP ratio governs ECM degradation, and compressive forces have been shown to increase MMP-9 expression while transiently suppressing TIMPs, thereby promoting early ECM breakdown [119].
Zang et al. (2023) [120] showed that LLLT applied under tension stress stimulated PDL cell proliferation and promoted periodontal remodeling. It suppressed osteogenic markers while increasing osteoclast-related factors, collagen, MMPs, and TIMPs. The authors hypothesized that LLLT may more strongly stimulate TIMPs than MMPs, potentially limiting collagen degradation and enabling net collagen accumulation. However, the dynamic balance of MMPs/TIMPs during OTM remains poorly defined and warrants further investigation. Persistent disruption of this balance may lead to complications like root resorption or impaired healing, underscoring the importance of precise temporal regulation.
In parallel, vascular remodeling is initiated to meet the elevated metabolic and cellular demands of PDL tissues during OTM. Compression-induced hypoxia stabilizes HIF-1α, a key regulator that activates angiogenic mediators such as VEGF. VEGF expression in PDL fibroblasts and osteoblasts increases within 24–48 hours of force application, stimulating endothelial cell proliferation, vascular permeability, and neovascularization in bone resorption areas [121]. Angiopoietins (Ang-1 and Ang-2) further regulate vascular architecture. Ang-1 promotes vessel stabilization and pericyte recruitment, whereas Ang-2 facilitates endothelial sprouting by loosening junctions [122].
Vascular and ECM remodeling are closely linked. VEGF promotes both angiogenesis and MMP expression, facilitating osteoclast recruitment and ECM degradation. MMPs, in turn, remodel the basement membrane to support endothelial migration and capillary growth. These processes are orchestrated by cytokines such as IL-1β, TNF-α, and TGF-β, coordinating ECM turnover with angiogenesis during the shift from inflammation to repair [123]. LLLT enhances these effects by upregulating VEGF and accelerating neovascularization in tension-stressed periodontal tissues, likely via HIF-1α and ERK pathway activation [124,125]. These responses are most evident in the areas of active remodeling and are essential for maintaining tissue viability during OTM.
Mechanical strain also influences endothelial function by inducing Intercellular Adhesion Molecule 1 (ICAM-1) and Vascular Cell Adhesion Molecule 1 (VCAM-1), promoting leukocyte transmigration, while eNOS activation and NO production contribute to vasodilation and regulation of osteoclastic and immune activity [126,127]. LLLT further stimulates Human Umbilical Vein Endothelial Cells (HUVEC) proliferation, migration, and angiogenesis through increased HIF-1α, eNOS, and VEGF expression, mediated via the PI3K/Akt/mTOR pathway, as shown by Li et al. (2020) [128]. Szymczyszyn et al. (2016) [129] demonstrated that transdermal LLLT improves endothelial function through enhanced antioxidant and angiogenic responses, though NO metabolite levels remained unchanged within 24 hours.
Based on these findings, LLLT appears to modulate both extracellular matrix and vascular remodeling during OTM by balancing MMP/TIMP expression and enhancing angiogenic signaling. This dual regulation may facilitate controlled tissue remodeling, reduce adverse effects, and accelerate orthodontic outcomes. However, the precise dynamics and long-term effects of these interactions require further investigation.
9. Apoptosis and Autophagy in Orthodontic Tooth Movement: Mechanosensitive Responses and Cellular Interplay under Low-Level Laser Therapy
Apoptosis and autophagy are interrelated but often antagonistic processes that play central roles during OTM, particularly in regulating the behavior of osteocytes, osteoblasts, PDLSCs, and immune cells within the alveolar bone environment. While apoptosis removes damaged or overstimulated cells, autophagy promotes survival by eliminating dysfunctional organelles, controlling ROS levels, and modulating inflammatory signaling. The balance between these pathways is critical for proper coordination of bone resorption and formation during OTM [130].
Mechanical stress has been shown to trigger apoptosis in multiple periodontal cell types. In a mouse OTM model, TUNEL staining revealed a significant increase in osteocyte apoptosis on the compression side as early as 12 h, with a peak at 24 h [131]. Similarly, a human biopsy study reported a rise in the apoptotic index beginning on day 3, peaking on day 7 at both the compression and tension sites, followed by a decline reaching minimal levels by day 21 [132].
In vitro experiments support these findings. Cyclic or static mechanical loading induces caspase-3 and caspase-5 activation in PDLSCs and MG 63 osteoblast-like cells, indicating force-dependent apoptosis [133]. Yu et al. (2017) further confirmed this in MG 63 cells subjected to 15% cyclic stretch at 0.1 Hz, demonstrating increased Cleaved Poly (ADP-ribose) Polymerase (cPARP) and caspase-3 expression. Interestingly, periostin exerted a protective effect, significantly reducing stretch-induced apoptosis [134]. Li et al. (2021) reviewed a wide range of in vitro studies involving PDLCs and MG 63 cells, concluding that mechanical loading consistently triggers programmed cell death, primarily through caspase activation, thus reinforcing the concept of mechanically induced apoptosis in these cells [135].
Orthodontic force application has been directly linked to early osteocyte apoptosis in alveolar bone. Kaplan et al. (2021) proposed that this apoptosis is not merely a passive consequence of mechanical stress but an active process that facilitates the recruitment and differentiation of osteoclasts and macrophages, supporting bone remodeling. Notably, it does not appear to significantly affect the overall rate or magnitude of tooth movement [136].
Recent findings highlight the functional relevance of apoptosis in bone regeneration. Liu et al. () demonstrated that apoptosis of PDLSCs activates Lepr⁺ osteoprogenitors, particularly on the compression side of the PDL. This is mediated through apoptotic vesicles derived from PDLSCs, which serve as signaling cues for Lepr⁺ cells, revealing a novel link between force-induced apoptosis and osteogenesis during OTM [137]. Shen et al. showed that inhibition of Piezo1 reduced force-induced apoptosis in periodontal tissues and fibroblasts via downregulation of the p38/ERK1/2 pathway, indicating that modulation of mechanotransduction can influence cell survival under orthodontic load [138]. However, some studies suggest that PDLSCs may adapt to compressive stress without undergoing significant apoptosis, indicating variability in cell-type or context-specific responses [139].
Although no studies have directly assessed the effects of LLLT on apoptosis in OTM models, evidence from other systems offers important insight. LLLT exhibits biphasic effects depending on energy dose and cell type. At lower doses, it can protect cells, such as endothelial or muscle cells, from apoptosis triggered by inflammatory or oxidative stress. Conversely, higher doses or different cellular contexts, such as in colorectal cancer models, can activate apoptotic pathways and promote cell death. This dose-dependent duality underscores the complexity of LLLT-induced cellular responses [140,141,142].
Autophagy, alongside apoptosis, is a key cellular stress response during OTM. These two processes are often reciprocally regulated. In a rat OTM model, autophagy-related gene expression in PDLSCs rose shortly after force application but declined by day 7, while pro-apoptotic proteins followed the opposite trend [6]. This inverse relationship is supported by findings that caspase-3 can inhibit autophagy through the cleavage of Beclin 1, a central autophagic regulator [130,143].
Experimental studies confirm rapid autophagy activation post-force. In rats, Beclin 1 and LC3 II peaked within 1 h of loading, while p62 decreased, indicating early autophagic flux. Apoptotic markers increased later, suggesting autophagy precedes apoptosis in mechanically stressed PDL cells [143]. In GFP-LC3 reporter mice, autophagosome accumulation was observed in PDL and osteoclast precursors following force application. Notably, rapamycin-induced autophagy reduced both osteoclast numbers and tooth movement, implying a suppressive role in bone turnover and inflammation [144].
On the tension side, human PDLSCs exposed to stretch showed concurrent upregulation of osteogenic and autophagy-related markers. Silencing ATG7 suppressed osteogenesis, while pharmacologic autophagy enhancement amplified it, suggesting autophagy supports bone formation under tensile strain [145]. A 2025 narrative review [146] synthesized these insights, reporting that compressive force activates autophagy in osteocytes and PDLSCs but inhibits it in cementoblasts, while tensile force and fluid shear stress broadly stimulate autophagy across periodontal tissues. The review emphasized the involvement of PI3K/Akt, Hippo, and MAPK pathways, and identified periostin and non-coding RNAs as potential targets for modulating force-induced autophagy.
Clinically, autophagy plays both protective and regulatory roles. Its enhancement (e.g., via rapamycin) may mitigate inflammation, promote bone deposition, and reduce excessive tooth movement, benefiting cases prone to root resorption. Conversely, short-term autophagy inhibition could accelerate movement when needed, although increased inflammation must be carefully weighed [144].
LLLT is a known modulator of autophagy. In MC3T3-E1 pre-osteoblasts exposed to H₂O₂, 808 nm LLLT enhanced autophagic flux (an increase in LC3B, and a decrease in p62) by inhibiting the PI3K/Akt/mTOR pathway, promoting osteogenic differentiation and matrix formation. Blocking autophagy reversed these effects, underscoring its functional role [147]. Similar results were reported in osteoporotic bone marrow-derived stem cells treated with 650 nm PBM, where autophagy activation improved ALP activity and mineral deposition [148]. Fernandes et al. (2024) further demonstrated that PBM upregulates autophagy markers (SQSTM1/p62, Beclin, Parkin) and activates AMPK and TGF-β signaling in a muscle degeneration model [149].
Collectively, these findings emphasize apoptosis and autophagy as interconnected, force-sensitive processes central to periodontal remodeling during orthodontic treatment. Their time-dependent regulation, mechanosensitive pathways, and influence on progenitor cell activation provide mechanistic insight into tissue adaptation. By modulating these pathways, LLLT may support regenerative responses while limiting excessive cell loss, offering therapeutic potential to optimize OTM outcomes and reduce tissue damage.
10. Immune Dynamics in Orthodontic Tooth Movement: Unresolved Mechanisms of Low-Level Laser Therapy
10.1. Innate Immunity
Neutrophils and monocytes are key innate immune cells, comprising approximately 70% of leukocytes in the murine PDL [150]. Neutrophils act as early responders to orthodontic force, rapidly infiltrating the PDL to perform phagocytosis, degrade ECM, and recruit monocytes and macrophages before undergoing apoptosis [33,150,151]. They also release pro-inflammatory mediators, including IL-1, IL-6, TNF-α, CCL2, CXCL1α, and CCL3, which collectively drive osteoclastogenesis and bone resorption [152]. In humans, increased neutrophil numbers in gingival crevicular fluid (GCF) and saliva are observed within 2 h after force application [153], and murine models consistently show elevated levels by day 3 [151].
Monocytes serve dual roles. They clear apoptotic cells and act as precursors for osteoclasts, macrophages, and DCs [151,154]. Their infiltration peaks around day 3 post-force application [150], followed by osteoclastic differentiation on the compression side, contributing to bone resorption [154]. Experimental depletion of monocytes/macrophages reduces osteoclast formation, lowers TNF-α and iNOS expression, and markedly impairs tooth movement in mice [155].
Macrophages, derived from monocytes, modulate inflammation and bone remodeling. M1-type macrophages (CD68⁺/iNOS⁺), generated via the JAK/STAT3 pathway, produce IL-1, IL-6, IL-12, TNF-α, and iNOS, thereby supporting matrix degradation, debris clearance, and osteoclast activation on the compression side [150,156,157]. Their numbers increase by day 3, peak at day 7, and remain elevated through day 14 [150,151,155]. Around day 7, macrophages exhibit enhanced chemokine signaling and inflammatory gene expression [158]. CCR2⁺ macrophages are predominant during both homeostasis and OTM. Inhibition of CCR2 suppresses inflammatory signaling and delays tooth movement, indicating a regulatory role [158]. Increased phagocytic activity and monocyte pathway activation further support their involvement [159]. In vitro, mechanically stimulated macrophages release cytokines that influence PDL cell behavior and promote osteoclastogenesis [160].
M2 macrophages, generated via the NF-κB pathway in the presence of Th2 cytokines IL-4 and IL-13, contribute to the resolution of inflammation by suppressing osteoclastogenesis and secreting BMP2, which promotes bone formation [161]. In rat OTM models, M2 macrophages become more prevalent during later phases, aligning with inflammation decline and the onset of tissue remodeling [162]. In co-culture with human osteoblasts, M2 macrophages enhance osteogenesis, whereas M1 macrophages inhibit it, reinforcing their opposing roles in bone metabolism [150,151].
DCs, present in the PDL, help maintain tissue homeostasis and are increasingly recognized as players in OTM. Following orthodontic force, CD11c⁺ DCs increase in number in rat PDL, suggesting force-induced activation [163]. Transcriptomic profiling in mice revealed peak DC maturation and interaction with NK cells around day 3 of OTM [159]. However, direct evidence regarding DC phenotype, migration, and impact on bone remodeling remains limited and requires further investigation.
Among innate lymphocytes, γδ T cells are critical in early OTM. Although sparse, their deletion in mice reduces IL-17A and RANKL expression, impairs neutrophil and monocyte recruitment, diminishes osteoclastogenesis, and delays tooth movement [164]. In mice, γδ T cells are the main IL-17A source, whereas in humans, this function is primarily carried out by Th17 cells.
NK cells also contribute to the early inflammatory phase. Their numbers rise in the PDL post-force, where they secrete TNF-α and IFN-γ [151,159]. Depletion of NK cells via Ncr1 receptor impairs tooth movement, suggesting a role in osteoclast regulation [151].
Innate lymphoid cells (ILCs) have emerged as important mediators of sterile inflammation in OTM. Unlike adaptive lymphocytes, ILCs respond rapidly and without antigen specificity, exhibiting phenotypic plasticity influenced by local cues [165]. Wang et al. (2024) reported that ILC1s migrate toward force-stressed PDL cells, while ILC2s and ILC3s expand in co-culture with such cells, indicating distinct, subtype-specific responses to mechanical stress [165].
LLLT significantly modulates innate immunity during OTM, though much of the evidence is indirect, derived from non-orthodontic models such as wound healing, autoimmune diseases, periodontitis, and in vitro systems. This contributes to heterogeneity and occasional contradictions in reported outcomes.
Given the pro-inflammatory nature of early OTM, it is plausible that LLLT facilitates tooth movement by enhancing early pro-inflammatory signaling. Clinical studies report elevated pro-inflammatory cytokines, including IL-1β, IL-6, IL-8, and RANKL in GCF of patients undergoing OTM with laser treatment [166,167,168]. A systematic review by Reis et al. (2022), covering LLLT wavelengths from 618 to 980 nm, found increased IL-1β, IL-8, OPN, and PGE2 in GCF, but no significant changes in TNF-α or TGF-β. Levels of IL-6, RANKL, and OPG varied depending on timing and laser parameters [169]. Kapoor et al. (2014) showed rapid release of IL-1β and TNF-α into GCF immediately after orthodontic force application, with IL-1β rising within 1 minute and peaking at 24 hours, and TNF-α increasing between 1 hour and 1 day. This early inflammatory surge declines over time, especially under continuous force, highlighting the importance of precise sampling timing [170]. While multiple PDL cell types produce these cytokines, infiltrating innate immune cells are the primary source.
Regarding neutrophils, Cerdeira et al. (2016) showed that LLLT stimulates oxidative burst and enhances fungicidal activity in human neutrophils [171]. Wasik et al. (2007) demonstrated increased chemiluminescence in fMLP-stimulated granulocytes from irradiated peripheral blood (632.8 nm, 0.6 J/cm²) compared to controls [172]. PBM has also been shown to increase IL-1β, CCL2, and CCL3 levels in PBMC cultures, indicating a dose-dependent pro-inflammatory response [173].
In monocytes and macrophages, Chen et al. (2014) reported that 660 nm LLLT enhanced M1-associated cytokines and chemokines at mRNA and protein levels, accompanied by histone modifications at TNF-α and IP-10 gene loci [174]. In RAW264.7 macrophages, LLLT dose-dependently increased M1 markers (iNOS, IL-12, TNF-α, IL-1β, IL-6) in naïve M0 cells, while downregulating M1 and upregulating M2 markers (Arg-1, IL-10) in pre-polarized M1 cells via PI3K/AKT/mTOR signaling. This suggests that LLLT promotes M1 polarization early on but can shift polarized M1 macrophages toward an M2 phenotype, aiding inflammation resolution during later OTM stages [175]. Fernandes et al. (2015) observed IL-6 upregulation at 660 nm LLLT in M1 macrophages despite general suppression of other pro-inflammatory cytokines [176].
Similarly, in a rat spinal cord injury model, 810 nm LLLT reduced M1 markers and increased M2 cytokines (IL-4, IL-13), reprogramming microglia/macrophages toward an M2 phenotype that promoted neuronal survival and recovery [177]. Zhang et al. (2019) confirmed that 810 nm LLLT reduced M1 markers, enhanced M2 polarization, and increased neurotrophic factor secretion via PKA-CREB signaling in mouse bone marrow-derived macrophages [178]. These findings are consistent with reports that LLLT suppresses pro-inflammatory responses and impairs neutrophil and monocyte function under inflammatory conditions such as periodontitis [179] and RA [180], potentially indicating a stage-dependent immunomodulatory effect during OTM.
The precise mechanisms of LLLT’s immunomodulatory effects on innate immune cells during early OTM remain unclear. These effects may occur directly, through light-responsive photoreceptors expressed by various cells, including innate immune cells, or indirectly, via other PDL cell types such as PDLSCs. PDLSCs are central in initiating mechanical force-induced sterile inflammation by secreting pro-inflammatory mediators that recruit and activate immune cells. Notably, all studies to date report anti-inflammatory effects of LLLT on PDLSCs, suggesting that LLLT’s modulation of PDLSCs is more relevant during later OTM stages when inflammation resolution is critical [181,182].
10.2. Adaptive Immunity
While innate immunity has been extensively studied in the context of OTM, the role of adaptive immunity remains comparatively underexplored. T lymphocytes infiltrate the PDL early during OTM, guided by pro-inflammatory mediators released from PDL cells, neutrophils, and M1 macrophages [2,183]. DCs in regional mucosal lymphoid tissues are primarily responsible for initiating T-cell responses. However, the mechanisms by which they migrate from the PDL to lymphoid tissues under mechanical stress, as well as the nature of their maturational stimuli, remain unclear. It is postulated that damage-associated molecular patterns (DAMPs) and alarmins released from compressed PDL structures may trigger this process [184].
Rodent models highlight adaptive immunity as a key regulator of tissue remodeling. Among CD4⁺ T helper subsets, Th1 and Th17 cells dominate early responses. Th1 cells, induced by IL-12 and IFN-γ via T-bet, produce IFN-γ, which enhances macrophage function and antigen presentation. Th17 differentiation, driven by IL-6, IL-1β, and TGF-β via Retinoic acid receptor-related orphan receptor (ROR) γt, results in IL-17A, IL-17F, and IL-22 production, cytokines with pro-inflammatory and osteoclastogenic roles [185].
Th17 cells promote RANKL-mediated osteoclastogenesis more potently than other T cell subsets [186]. IL-17 stimulates fibroblasts, PDLSCs, and epithelial cells to secrete chemokines (e.g., CCL2, CXCL8) and MMPs, aiding ECM remodeling and immune cell recruitment [187]. It also induces IL-6, IL-8, and PGE2 release from epithelial, endothelial, and stromal cells, and enhances osteoblast RANKL expression, driving periodontal bone resorption [188]. Elevated IL-17 in GCF may indicate root resorption during OTM [189].
Th1 cells also express RANKL and contribute to osteoclastogenesis, often indirectly via IFN-γ–mediated modulation of macrophages and DCs [190]. IFN-γ, produced by Th1, cytotoxic T cells, DCs, and NK cells, activates macrophages through NO production and MHC upregulation. In OTM, IFN-γ’s role is complex. It localizes compression sites, correlating with increased trabecular bone volume and reduced separation, suggesting bone-preserving effects. Conversely, it may suppress osteoclastogenesis and limit tooth movement, highlighting context-dependent effects influenced by timing and local environment [191].
Th2 cells’ involvement in OTM is less defined. IL-4, a key Th2 cytokine, inhibits RANKL- and TNF-α–induced osteoclastogenesis by blocking NF-κB and MAPK pathways, specifically suppressing NFATc1 activation via Signal Transducer and Activator of Transcription 6 (STAT6)-mediated NF-κB inhibition. IL-4 also indirectly reduces pro-inflammatory mediators such as TNF-α, IL-1, and IL-6 [192,193].
CD220⁺ B lymphocytes increase during the initial and linear tooth movement phases. Though a known RANKL source, their precise role is unclear. Only one study reported that hPDLSCs can suppress B cell activity via direct contact [194,195]. B cells exhibit dual functions. They promote osteoclastogenesis by secreting IL-6, GM-CSF, and RANKL, and indirectly stimulate bone resorption through IL-12–mediated Th1 activation. Simultaneously, they produce anti-inflammatory factors such as OPG, IL-10, and TGF-β, suppressing RANKL and osteoclast differentiation [196,197]. Despite these regulatory capabilities, direct evidence of B cells as major modulators in orthodontic bone remodeling is limited.
In the later phase of OTM, a shift in the Th cell subset composition occurs, characterized by a predominance of Tregs [182,191,194]. Tregs are a specialized subset of CD4⁺ T cells that play a pivotal role in maintaining immune homeostasis and controlling excessive inflammatory responses. They are defined by the expression of the transcription factor Foxp3 and typically express high levels of CD25 (IL-2 receptor α chain), while exhibiting low expression of CD127. Tregs exert their immunosuppressive function through both cell contact-dependent mechanisms and the secretion of anti-inflammatory cytokines, including IL-10 and TGF-β [198].
During OTM, Tregs contribute to the resolution of inflammation and support tissue remodeling by inhibiting osteoclastogenesis and limiting the pro-inflammatory activity of effector Th subsets, especially Th17 cells. Notably, the balance between Th17 and Treg populations is crucial for the regulation of bone resorption, tissue integrity, and the pathogenesis of gingivitis and periodontitis [199]. While Th17 cells are osteoclastogenic through the production of RANKL and IL-17, Tregs counteract these effects by downregulating RANKL and promoting OPG expression, thereby suppressing osteoclast differentiation. Tregs generally suppress osteoclast production by inhibiting RANKL and M-CSF expression, secreting IL-10 and TGF-β, and directly binding to osteoclast precursors. This mechanism helps maintain bone homeostasis and prevents excessive bone resorption [200].
In experimental models, orthodontic force has been shown to elevate Treg levels. For example, in a rat model of OTM with concurrent periodontitis, the proportion of peripheral and gingival Foxp3⁺ Tregs peaked early (around day 3), dipped by day 7, and then rose again during tooth movement, suggesting a dynamic Treg response to mechanical loading under inflammatory conditions [201]. Similarly, in both in vitro and in vivo models, heavy orthodontic force combined with surgical corticotomy led to enhanced Foxp3 expression and increased Tregs in periodontal tissues, correlating with faster tooth movement and reduced inflammatory bone injury [202].
There is a cross-talk between Tregs and PDLSCs. Tregs interact with PDLSCs by enhancing their osteogenic differentiation via the Jagged1–Notch2 signaling axis and promoting mineralization through direct cell contact [203]. PDLSCs, in turn, modulate T-cell responses by secreting immunoregulatory factors such as indoleamine 2,3-dioxygenase (IDO), TGF-β1, and PGE₂, which suppress effector T cells and induce Treg differentiation [204]. Additionally, PDLSCs downregulate CD1b expression on DCs, limiting their antigen-presenting capacity and favoring Treg expansion [205].
The effect of LLLT on adaptive immunity during OTM is still poorly understood. Given the known impact of mechanical stress on mobilization, activation, and differentiation of effector T cells in early OTM, it is hypothesized that LLLT may synergistically modulate these processes. Some in vitro studies support this, showing that LLLT enhances T cell proliferation in the presence of PHA, a T cell mitogen [206,207], and B cell proliferation with SAC, a B cell mitogen [172].
However, other studies show inconsistent results. Musawi et al. (2017) found that irradiation of whole blood at 589 nm (72 J/cm²) increased total CD45⁺ lymphocytes and NK cells but did not alter T or B cell subsets [208]. The stimulatory effect of PBM on T cells has been supported by findings from other experimental models. In a murine neuroinflammation model, PBM activated the JAK2/STAT4/STAT5 pathway, promoting IFN-γ and IL-10 secretion from CD4⁺ T cells in vivo and in vitro [209]. Fractional CO₂ laser treatment in a tumor model increased both CD4⁺ and CD8⁺ T cell subsets locally [210].
Conversely, more consistent evidence shows that LLLT promotes Treg development, demonstrated in models of asthma [211], RA [212], and stroke [213]. PDLSCs, similar to other MSCs, induce Tregs potentially via conversion of pro-inflammatory Th17 cells into Tregs, a shift likely occurring in later OTM stages and contributing to inflammation resolution [214,215]. However, the mechanisms driving this immunological switch and how LLLT influences these regulatory pathways remain unclear.
A summary of the involvement of immune mechanisms in OTM and their modulation by LLLT is presented in Figure 1.
11. Conclusions
OTM involves a finely regulated sequence of mechanobiological events mediated by osteoblasts, osteoclasts, PDL cells, and immune system components. LLLT, a noninvasive photobiomodulatory technique, has emerged as a promising adjunct in orthodontics by influencing these processes at both cellular and molecular levels.
Beyond its traditional effects on osteoblast and osteoclast activity, evidence suggests that LLLT modulates intercellular communication during OTM by interfacing with mechanotransduction pathways such as MAPK, PI3K/Akt, and Wnt/β-catenin. This suggests that photonic energy may act synergistically with orthodontic mechanical forces, altering intracellular signaling cascades and thereby fine-tuning bone remodeling dynamics. Moreover, LLLT may affect key adaptive responses in the orthodontic microenvironment, including apoptosis, autophagy, and cytokine-mediated cross-talk between skeletal and immune cells.
Preclinical and clinical evidence point to LLLT’s potential to accelerate tooth movement and alleviate treatment discomfort. However, variability in outcomes underscores the importance of standardizing irradiation protocols and gaining deeper insight into the mechanistic basis of its effects. Future research should focus on controlled in vitro studies using relevant human cell types under force application, alongside well-designed clinical trials incorporating molecular biomarkers and imaging tools for real-time evaluation of tissue remodeling.
Overall, LLLT provides a biologically plausible, mechanism-based approach to enhancing orthodontic treatment by incorporating photonic cues into the natural framework of force-driven tissue adaptation. With further validation, this strategy could help optimize treatment timelines, improve patient comfort, and minimize adverse effects, positioning LLLT as a valuable component of evidence-based orthodontic practice.
12. Future Perspectives
To strengthen the clinical integration of LLLT in orthodontics, future studies should prioritize mechanistic investigations that combine advanced in vitro modeling with clear translational endpoints. Emerging technologies now offer powerful tools for dissecting how PBM influences the interplay between mechanical forces, intracellular signaling, and immune regulation during OTM.
Single-cell RNA sequencing (scRNA-seq) provides unmatched resolution in mapping cell-specific transcriptional responses to mechanical stress and PBM. Applying this approach to human PDL tissues or cultured PDL-derived cells could reveal novel signaling pathways and specialized cell subsets involved in PBM-mediated remodeling.
Mechanically active in vitro models, such as 2D stretch systems, 3D compression platforms, and force-controlled bioreactors, allow precise simulation of orthodontic forces under standardized conditions. When paired with PBM, these platforms can clarify how specific wavelengths and energy doses affect mechanosensitive pathways.
Another promising avenue is the use of co-culture systems involving PDL cells, osteoclast/osteoblast precursors, and immune cells such as macrophages, dendritic cells, or T lymphocytes. These models are ideally suited for studying PBM-driven changes in cytokine signaling, osteoimmunologic crosstalk, and programmed cell responses such as apoptosis and autophagy. Likewise, organoids, spheroids, and organ-on-a-chip platforms offer physiologically relevant 3D microenvironments that mimic the in vivo periodontal milieu, enabling high-content analysis of PBM effects on angiogenesis, ECM remodeling, and spatiotemporal immune regulation.
Integrating multi-omics profiling, live-cell imaging, and quantitative biomarker analysis will further advance understanding of PBM’s dynamic impact on tissue adaptation. Establishing validated in vitro systems that use human primary cells exposed to controlled orthodontic forces and standardized PBM protocols will be essential for translating laboratory findings into clinical protocols. Such efforts could ultimately guide the development of precision PBM strategies tailored to optimize orthodontic treatment outcomes.
Author Contributions
Conceptualization, J.M. and M.Č.; data curation, J.M.; writing—original draft preparation, J.M. and M.Č.; writing—review and editing, M.Č.; supervision, M.Č. Both authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
This work was supported by the Serbian Academy of Sciences and Arts (Project number: F135). The authors gratefully acknowledge Professor Dragana Vučević for her help with figure drawing.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Krishnan, V.; Davidovitch, Z. Cellular, molecular, and tissue-level reactions to orthodontic force. Am. J. Orthod. Dentofacial Orthop. 2006, 129, 469.e1–469.e32. [Google Scholar] [CrossRef] [PubMed]
- Meikle, M.C. The tissue, cellular, and molecular regulation of orthodontic tooth movement: 100 years after Carl Sandstedt. Eur. J. Orthod. 2006, 28, 221–240. [Google Scholar] [CrossRef]
- Ren, Y.; Maltha, J.C.; Kuijpers-Jagtman, A.M. Optimum force magnitude for orthodontic tooth movement: a systematic literature review. Angle Orthod. 2003, 73, 86–92. [Google Scholar]
- d’Apuzzo, F.; Cappabianca, S.; Ciavarella, D.; Monsurrò, A.; Silvestrini-Biavati, A.; Perillo, L.; Perillo, L. Biomarkers of periodontal tissue remodeling during orthodontic tooth movement in mice and men: Overview and clinical relevance. Sci. World J. 2013, 2013, 105873. [Google Scholar] [CrossRef]
- Al-Naoum, F.; Hajeer, M.Y.; Al-Jundi, A. Does alveolar corticotomy accelerate orthodontic tooth movement when retracting upper canines? A split-mouth design randomized controlled trial. J. Oral Maxillofac. Surg. 2014, 72, 1880–1889. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Kim, S.J.; Yoon, H.J.; Lee, P.J.; Moon, W.; Park, Y.G. Effect of piezopuncture on tooth movement and bone remodeling in dogs. Am. J. Orthod. Dentofac. Orthop. 2013, 144, 23–31. [Google Scholar] [CrossRef]
- Hamblin, M.R. Mechanisms and applications of the anti-inflammatory effects of photobiomodulation. AIMS Biophys. 2017, 4, 337–361. [Google Scholar] [CrossRef]
- Khadra, M.; Lyngstadaas, S.P.; Haanaes, H.R.; Mustafa, K. Effect of laser therapy on attachment, proliferation and differentiation of human osteoblast-like cells cultured on titanium implant material. Biomaterials 2005, 26, 3503–3509. [Google Scholar] [CrossRef]
- Alzahrani, A.M.; Aljibrin, F.J.; Alqahtani, A.M.; Saklou, R.; Alhassan, I.A.; Alamer, A.H.; Al Ameer, M.H.; Hatami, M.S.; Dahhas, F.Y. Photobiomodulation in Orthodontics: Mechanisms and Clinical Efficacy for Faster Tooth Movement. Cureus 2024, 16, e59061. [Google Scholar] [CrossRef]
- Bianco, P.; Cao, X.; Frenette, P.S.; Mao, J.J.; Robey, P.G.; Simmons, P.J.; Wang, C.Y. The meaning, the sense and the significance: Translating the science of mesenchymal stem cells into medicine. Nat. Med. 2013, 19, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef]
- Chen, D.; Zhao, M.; Mundy, G.R. Bone morphogenetic proteins. Growth Factors 2004, 22, 233–241. [Google Scholar] [CrossRef]
- Baron, R.; Kneissel, M. WNT signaling in bone homeostasis and disease: From human mutations to treatments. Nat. Med. 2013, 19, 179–192. [Google Scholar] [CrossRef]
- Komori, T. Regulation of osteoblast differentiation by Runx2. Adv. Exp. Med. Biol. 2010, 658, 43–49. [Google Scholar] [CrossRef]
- Canalis, E. Growth factor control of bone mass. J. Cell Biochem. 2009, 108, 769–777. [Google Scholar] [CrossRef]
- Florencio-Silva, R.; Sasso, S.; Sasso-Cerri, E.; Simões, M.J.; Cerri, P.S. Biology of bone tissue: Structure, function, and factors that influence bone cells. BioMed Res. Int. 2015, 2015, 421746. [Google Scholar] [CrossRef]
- Golub, E.E. Role of matrix vesicles in biomineralization. Biochim. Biophys. Acta 2009, 1790, 1592–1598. [Google Scholar] [CrossRef]
- Hofbauer, L.C.; Heufelder, A.E. Role of receptor activator of nuclear factor-kappaB ligand and osteoprotegerin in bone cell biology. J. Mol. Med. 2001, 79, 243–253. [Google Scholar] [CrossRef]
- Dallas, S.L.; Bonewald, L.F. Dynamics of the transition from osteoblast to osteocyte. Ann. N. Y. Acad. Sci. 2010, 1192, 437–443. [Google Scholar] [CrossRef]
- Teitelbaum, S.L. Bone resorption by osteoclasts. Science 2000, 289, 1504–1508. [Google Scholar] [CrossRef]
- Boyle, W.J.; Simonet, W.S.; Lacey, D.L. Osteoclast Differentiation and Activation. Nature 2003, 423, 337–342. [Google Scholar] [CrossRef]
- Tsurukai, T.; Udagawa, N.; Matsuzaki, K.; Takahashi, N.; Suda, T. Roles of Macrophage-Colony Stimulating Factor and Osteoclast Differentiation Factor in Osteoclastogenesis. J. Bone Miner. Metab. 2000, 18, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Takayanagi, H. Osteoimmunology: Shared Mechanisms and Crosstalk between the Immune and Bone Systems. Nat. Rev. Immunol. 2007, 7, 292–304. [Google Scholar] [CrossRef]
- Takayanagi, H.; Kim, S.; Koga, T.; Nishina, H.; Isshiki, M.; Yoshida, H.; Saiura, A.; Isobe, M.; Yokochi, T.; Inoue, J.; Wagner, E.F.; Mak, T.W.; Kodama, T.; Taniguchi, T. Induction and Activation of the Transcription Factor NFATc1 (NFAT2) Integrate RANKL Signaling in Terminal Differentiation of Osteoclasts. Dev. Cell 2002, 3, 889–901. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Lee, N.K.; Lee, S.Y. Current Understanding of RANK Signaling in Osteoclast Differentiation and Maturation. Mol. Cells 2017, 40, 706–713. [Google Scholar] [CrossRef]
- Xu, F.; Teitelbaum, S.L. Osteoclasts: New Insights. Bone Res. 2013, 1, 11–26. [Google Scholar] [CrossRef]
- Drake, M.T.; Clarke, B.L.; Khosla, S. Bisphosphonates: Mechanism of Action and Role in Clinical Practice. Mayo Clin. Proc. 2008, 83, 1032–1045. [Google Scholar] [CrossRef]
- Kim, J.M.; Lin, C.; Stavre, Z.; Greenblatt, M.B.; Shim, J.H. Osteoblast–Osteoclast Communication and Bone Homeostasis. Cells 2020, 9, 2073. [Google Scholar] [CrossRef]
- Wen, X.; Pei, F.; Jin, Y.; Zhao, Z. Exploring the Mechanical and Biological Interplay in the Periodontal Ligament. Int. J. Oral Sci. 2025, 17, 1–12. [Google Scholar] [CrossRef]
- Lei, T.; Wang, J.; Liu, Y.; Zhang, X.; Du, H. Comparative Proteomics Analysis of Human Stem Cells from Dental Gingival and Periodontal Ligament. Proteomics 2022, 22, e2200027. [Google Scholar] [CrossRef] [PubMed]
- Rizk, M.; Niederau, C.; Florea, A.; Kiessling, F.; Morgenroth, A.; Mottaghy, F.M.; Schneider, R.K.; Wolf, M.; Craveiro, R.B. Periodontal Ligament and Alveolar Bone Remodeling during Long Orthodontic Tooth Movement Analyzed by a Novel User-Independent 3D-Methodology. Sci. Rep. 2023, 13, 19919. [Google Scholar] [CrossRef] [PubMed]
- Danz, J.C.; Degen, M. Selective Modulation of the Bone Remodeling Regulatory System through Orthodontic Tooth Movement—A Review. Front. Oral Health 2025, 6, 1472711. [Google Scholar] [CrossRef] [PubMed]
- Chaushu, S.; Klein, Y.; Mandelboim, O.; Barenholz, Y.; Fleissig, O. Immune Changes Induced by Orthodontic Forces: A Critical Review. J. Dent. Res. 2022, 101, 11–20. [Google Scholar] [CrossRef]
- Jiang, N.; Guo, W.; Chen, M.; Zheng, Y.; Zhou, J.; Kim, S.G.; Embree, M.C.; Song, K.S.; Marao, H.F.; Mao, J.J. Periodontal Ligament and Alveolar Bone in Health and Adaptation: Tooth Movement. Front. Oral Biol. 2016, 18, 1–8. [Google Scholar] [CrossRef]
- Klein, Y.; David, E.; Pinto, N.; Khoury, Y.; Barenholz, Y.; Chaushu, S. Breaking a Dogma: Orthodontic Tooth Movement Alters Systemic Immunity. Prog. Orthod. 2024, 25, 38. [Google Scholar] [CrossRef]
- Henneman, S.; Von den Hoff, J.W.; Maltha, J.C. Mechanobiology of Tooth Movement. Eur. J. Orthod. 2008, 30, 299–306. [Google Scholar] [CrossRef]
- Robling, A.G.; Turner, C.H. Mechanical Signaling for Bone Modeling and Remodeling. Crit. Rev. Eukaryot. Gene Expr. 2009, 19, 319–338. [Google Scholar] [CrossRef]
- Li, X.; Han, L.; Nookaew, I.; Mannen, E.; Silva, M.J.; Almeida, M.; Xiong, J. Stimulation of Piezo1 by Mechanical Signals Promotes Bone Anabolism. Elife 2019, 8, e49631. [Google Scholar] [CrossRef]
- Goldmann, W.H. Mechanotransduction and Focal Adhesions. Cell Biol. Int. 2012, 36, 649–652. [Google Scholar] [CrossRef]
- Nakai, Y.; Praneetpong, N.; Ono, W.; Ono, N. Mechanisms of Osteoclastogenesis in Orthodontic Tooth Movement and Orthodontically Induced Tooth Root Resorption. J. Bone Metab. 2023, 30, 297–310. [Google Scholar] [CrossRef] [PubMed]
- Ubuzima, P.; Nshimiyimana, E.; Mukeshimana, C.; Mazimpaka, P.; Mugabo, E.; Mbyayingabo, D.; Mohamed, A.S.; Habumugisha, J. Exploring Biological Mechanisms in Orthodontic Tooth Movement: Bridging the Gap between Basic Research Experiments and Clinical Applications – A Comprehensive Review. Ann. Anat. 2024, 255, 152286. [Google Scholar] [CrossRef]
- Lisowska, B.; Kosson, D.; Domaracka, K. Lights and Shadows of NSAIDs in Bone Healing: The Role of Prostaglandins in Bone Metabolism. Drug Des. Devel. Ther. 2018, 12, 1753–1758. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Wu, P.; Yu, F.; Luo, G.; Qing, L.; Tang, J. HIF-1α Regulates Bone Homeostasis and Angiogenesis, Participating in the Occurrence of Bone Metabolic Diseases. Cells 2022, 11, 3552. [Google Scholar] [CrossRef] [PubMed]
- Marques-Carvalho, A.; Kim, H.N.; Almeida, M. The Role of Reactive Oxygen Species in Bone Cell Physiology and Pathophysiology. Bone Rep. 2023, 19, 101664. [Google Scholar] [CrossRef] [PubMed]
- Behm, C.; Nemec, M.; Blufstein, A.; Schubert, M.; Rausch-Fan, X.; Andrukhov, O.; Jonke, E. Interleukin-1β Induced Matrix Metalloproteinase Expression in Human Periodontal Ligament-Derived Mesenchymal Stromal Cells under In Vitro Simulated Static Orthodontic Forces. Int. J. Mol. Sci. 2021, 22, 1027. [Google Scholar] [CrossRef]
- Tobeiha, M.; Moghadasian, M.H.; Amin, N.; Jafarnejad, S. RANKL/RANK/OPG Pathway: A Mechanism Involved in Exercise-Induced Bone Remodeling. Biomed. Res. Int. 2020, 2020, 6910312. [Google Scholar] [CrossRef]
- Hu, L.; Chen, W.; Qian, A.; Li, Y.P. Wnt/β-Catenin Signaling Components and Mechanisms in Bone Formation, Homeostasis, and Disease. Bone Res. 2024, 12, 39. [Google Scholar] [CrossRef]
- Liu, X.; Zhao, W.; Peng, Y.; Liu, N.; Liu, Q. The Relationship between MAPK Signaling Pathways and Osteogenic Differentiation of Periodontal Ligament Stem Cells: A Literature Review. PeerJ 2025, 13, e19193. [Google Scholar] [CrossRef]
- Maycas, M.; Esbrit, P.; Gortázar, A.R. Molecular Mechanisms in Bone Mechanotransduction. Histol. Histopathol. 2017, 32, 751–760. [Google Scholar] [CrossRef]
- Hao, Y.; Yang, N.; Sun, M.; Yang, S.; Chen, X. The Role of Calcium Channels in Osteoporosis and Their Therapeutic Potential. Front. Endocrinol. 2024, 15, 1450328. [Google Scholar] [CrossRef]
- Zhang, Y.E. Non-Smad Signaling Pathways of the TGF-β Family. Cold Spring Harb. Perspect. Biol. 2017, 9, a022129. [Google Scholar] [CrossRef]
- Kirschneck, C.; Thuy, M.; Leikam, A.; Memmert, S.; Deschner, J.; Damanaki, A.; Spanier, G.; Proff, P.; Jantsch, J.; Schröder, A. Role and Regulation of Mechanotransductive HIF-1α Stabilisation in Periodontal Ligament Fibroblasts. Int. J. Mol. Sci. 2020, 21, 9530. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Y.; Tang, Q.; Zhang, Y.; Yin, Y.; Chen, L. Application of Antioxidant Compounds in Bone Defect Repair. Antioxidants 2024, 13, 789. [Google Scholar] [CrossRef]
- Heiskanen, V.; Hamblin, M.R. Photobiomodulation: Lasers vs. Light Emitting Diodes? Photochem. Photobiol. Sci. 2018, 17, 1003–1017. [Google Scholar] [CrossRef]
- Di Palma, G.; Inchingolo, A.M.; Patano, A.; Palumbo, I.; Guglielmo, M.; Trilli, I.; Netti, A.; Ferrara, I.; Viapiano, F.; Inchingolo, A.D.; Favia, G.; Dongiovanni, L.; Palermo, A.; Inchingolo, F.; Limongelli, L. Photobiomodulation and Growth Factors in Dentistry: A Systematic Review. Photonics 2023, 10, 1095. [Google Scholar] [CrossRef]
- Cunha, M.J.; Esper, L.A.; Sbrana, M.C.; de Oliveira, P.G.; do Valle, A.L.; de Almeida, A.L. Effect of Low-Level Laser on Bone Defects Treated with Bovine or Autogenous Bone Grafts: In Vivo Study in Rat Calvaria. Biomed. Res. Int. 2014, 2014, 104230. [Google Scholar] [CrossRef]
- Cronshaw, M.; Parker, S.; Anagnostaki, E.; Mylona, V.; Lynch, E.; Grootveld, M. Photobiomodulation and Oral Mucositis: A Systematic Review. Dent. J. 2020, 8, 87. [Google Scholar] [CrossRef]
- Ahmad, S.A.; Hasan, S.; Saeed, S.; Khan, A.; Khan, M. Low-Level Laser Therapy in Temporomandibular Joint Disorders: A Systematic Review. J. Med. Life 2021, 14, 148–164. [Google Scholar] [CrossRef] [PubMed]
- Dervisbegovic, S.; Lettner, S.; Tur, D.; Laky, M.; Georgopoulos, A.; Moritz, A.; Sculean, A.; Rausch-Fan, X. Adjunctive Low-Level Laser Therapy in Periodontal Treatment—A Randomized Clinical Split-Mouth Trial. Clin. Oral Investig. 2025, 29, 273. [Google Scholar] [CrossRef] [PubMed]
- Turhani, D.; Scheriau, M.; Kapral, D.; Benesch, T.; Jonke, E.; Bantleon, H.P. Pain Relief by Single Low-Level Laser Irradiation in Orthodontic Patients Undergoing Fixed Appliance Therapy. Am. J. Orthod. Dentofacial Orthop. 2006, 130, 371–377. [Google Scholar] [CrossRef] [PubMed]
- Furquim, R.D.; Pascotto, R.C.; Rino Neto, J.; Cardoso, J.R.; Ramos, A.L. Low-Level Laser Therapy Effects on Pain Perception Related to the Use of Orthodontic Elastomeric Separators. Dental Press J. Orthod. 2015, 20, 37–42. [Google Scholar] [CrossRef]
- Yassin, A.M.; Shehata, F.I.; Al Sawa, A.A.; Karam, S.S. Effect of Low Level Laser Therapy on Orthodontic Induced Inflammatory Root Resorption in Rats. Alexandria Dent. J. 2020, 45, 62–67. [Google Scholar] [CrossRef]
- Dompe, C.; Moncrieff, L.; Matys, J.; Grzech-Leśniak, K.; Kocherova, I.; Bryja, A.; Bruska, M.; Dominiak, M.; Mozdziak, P.; Skiba, T.H.I.; Shibli, J.A.; Angelova Volponi, A.; Kempisty, B.; Dyszkiewicz-Konwińska, M. Photobiomodulation—Underlying Mechanism and Clinical Applications. J. Clin. Med. 2020, 9, 1724. [Google Scholar] [CrossRef] [PubMed]
- Farid, K.A.; Eid, A.A.; Kaddah, M.A.; Elsharaby, F.A. The Effect of Combined Corticotomy and Low Level Laser Therapy on the Rate of Orthodontic Tooth Movement: Split Mouth Randomized Clinical Trial. Laser Ther. 2019, 28, 275–283. [Google Scholar] [CrossRef]
- Costa, A.C.F.; Maia, T.A.C.; de Barros Silva, P.G.; Abreu, L.G.; Gondim, D.V.; Santos, P.C.F. Effects of Low-Level Laser Therapy on the Orthodontic Mini-Implants Stability: A Systematic Review and Meta-Analysis. Prog. Orthod. 2021, 22, 6. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.; McGrath, C.; Jin, L.; Zhang, C.; Yang, Y. The Effectiveness of Low-Level Laser Therapy as an Adjunct to Non-Surgical Periodontal Treatment: A Meta-Analysis. J. Periodontal Res. 2017, 52, 8–20. [Google Scholar] [CrossRef] [PubMed]
- Doshi-Mehta, G.; Bhad-Patil, W.A. Efficacy of Low-Intensity Laser Therapy in Reducing Treatment Time and Orthodontic Pain: A Clinical Investigation. Am. J. Orthod. Dentofacial Orthop. 2012, 141, 289–297. [Google Scholar] [CrossRef]
- Elgadi, R.; Sedky, Y.; Franzen, R. The Effectiveness of Low-Level Laser Therapy on Orthodontic Tooth Movement: A Systematic Review. Lasers Dent. Sci. 2023, 7, 1–9. [Google Scholar] [CrossRef]
- Zheng, D.H.; Du, Y.Q.; Zhang, Q.Q.; Hou, F.C.; Niu, S.Q.; Zang, Y.J.; Li, B. Effect of low-level laser therapy on orthodontic dental alignment: A systematic review and meta-analysis. Lasers Med. Sci. 2023, 38, 184. [Google Scholar] [CrossRef]
- Bakdach, W.M.M.; Hadad, R. Effectiveness of low-level laser therapy in accelerating the orthodontic tooth movement: A systematic review and meta-analysis. Dent. Med. Probl. 2020, 57, 73–94. [Google Scholar] [CrossRef]
- Isola, G.; Matarese, M.; Briguglio, F.; Grassia, V.; Picciolo, G.; Fiorillo, L.; Matarese, G. Effectiveness of Low-Level Laser Therapy during Tooth Movement: A Randomized Clinical Trial. Materials 2019, 12, 2187. [Google Scholar] [CrossRef]
- Yavagal, C.M.; Matondkar, S.P.; Yavagal, P.C. Efficacy of Laser Photobiomodulation in Accelerating Orthodontic Tooth Movement in Children: A Systematic Review with Meta-Analysis. Int. J. Clin. Pediatr. Dent. 2021, 14 (Suppl 1), S94–S100. [Google Scholar] [CrossRef]
- Inchingolo, F.; Inchingolo, A.M.; Latini, G.; Del Vecchio, G.; Trilli, I.; Ferrante, L.; Dipalma, G.; Palermo, A.; Inchingolo, A.D. Low-Level Light Therapy in Orthodontic Treatment: A Systematic Review. Appl. Sci. 2023, 13, 10393. [Google Scholar] [CrossRef]
- Wu, S.; Chen, Y.; Zhang, J.; Chen, W.; Shao, S.; Shen, H.; Zhu, L.; Ye, P.; Svensson, P.; Wang, K. Effect of Low-Level Laser Therapy on Tooth-Related Pain and Somatosensory Function Evoked by Orthodontic Treatment. Int. J. Oral Sci. 2018, 10, 22. [Google Scholar] [CrossRef]
- Brito, M.H.; Nogueira, C.Q.; Cotrin, P.; Fialho, T.; Oliveira, R.C.; Oliveira, R.G.; Salmeron, S.; Valarelli, F.P.; Freitas, K.M.S.; Cançado, R.H. Efficacy of Low-Level Laser Therapy in Reducing Pain in the Initial Stages of Orthodontic Treatment. Int. J. Dent. 2022, 2022, 3934900. [Google Scholar] [CrossRef] [PubMed]
- El-Angbawi, A.; McIntyre, G.; Fleming, P.S.; Bearn, D. Non-Surgical Adjunctive Interventions for Accelerating Tooth Movement in Patients Undergoing Orthodontic Treatment. Cochrane Database Syst. Rev. 2023, 6, CD010887. [Google Scholar] [CrossRef] [PubMed]
- Li, F.J.; Zhang, J.Y.; Zeng, X.T.; Guo, Y. Low-Level Laser Therapy for Orthodontic Pain: A Systematic Review. Lasers Med. Sci. 2015, 30, 1789–1803. [Google Scholar] [CrossRef] [PubMed]
- Michelogiannakis, D.; Al-Shammery, D.; Akram, Z.; Rossouw, P.E.; Javed, F.; Romanos, G.E. Influence of Low-Level Laser Therapy on Orthodontically-Induced Inflammatory Root Resorption: A Systematic Review. Arch. Oral Biol. 2019, 100, 1–13. [Google Scholar] [CrossRef]
- Wang, X.; Liu, Q.; Peng, J.; Song, W.; Zhao, J.; Chen, L. The Effects and Mechanisms of PBM Therapy in Accelerating Orthodontic Tooth Movement. Biomolecules 2023, 13, 1140. [Google Scholar] [CrossRef]
- Masha, R.T.; Houreld, N.N.; Abrahamse, H. Low-Intensity Laser Irradiation at 660 nm Stimulates Transcription of Genes Involved in the Electron Transport Chain. Photomed. Laser Surg. 2013, 31, 47–53. [Google Scholar] [CrossRef]
- Pires Oliveira, D.A.; de Oliveira, R.F.; Zangaro, R.A.; Soares, C.P. Evaluation of Low-Level Laser Therapy of Osteoblastic Cells. Photomed. Laser Surg. 2008, 26, 401–404. [Google Scholar] [CrossRef]
- González, P.; Lozano, P.; Ros, G.; Solano, F. Hyperglycemia and Oxidative Stress: An Integral, Updated and Critical Overview of Their Metabolic Interconnections. Int. J. Mol. Sci. 2023, 24, 9352. [Google Scholar] [CrossRef]
- Sarti, P.; Forte, E.; Mastronicola, D.; Giuffrè, A.; Arese, M. Cytochrome c Oxidase and Nitric Oxide in Action: Molecular Mechanisms and Pathophysiological Implications. Biochim. Biophys. Acta 2012, 1817, 610–619. [Google Scholar] [CrossRef]
- Gao, W.; Huang, M.; Chen, X.; Chen, J.; Zou, Z.; Li, L.; Ji, K.; Nie, Z.; Yang, B.; Wei, Z.; Xu, P.; Jia, J.; Zhang, Q.; Shen, H.; Wang, Q.; Li, K.; Zhu, L.; Wang, M.; Ye, S.; Liu, Q. The Role of S-Nitrosylation of PFKM in Regulation of Glycolysis in Ovarian Cancer Cells. Cell Death Dis. 2021, 12, 1–14. [Google Scholar] [CrossRef]
- de Freitas, L.F.; Hamblin, M.R. Proposed Mechanisms of Photobiomodulation or Low-Level Light Therapy. IEEE J. Sel. Top. Quantum Electron. 2016, 22, 7000417. [Google Scholar] [CrossRef]
- Karu, T.I. Mitochondrial Signaling in Mammalian Cells Activated by Red and Near-IR Radiation. Photochem. Photobiol. 2008, 84, 1091–1099. [Google Scholar] [CrossRef]
- Chellini, F.; Sassoli, C.; Nosi, D.; Deledda, C.; Tonelli, P.; Zecchi-Orlandini, S.; Formigli, L.; Giannelli, M. Low Pulse Energy Nd:YAG Laser Irradiation Exerts a Biostimulative Effect on Different Cells of the Oral Microenvironment: An In Vitro Study. Lasers Surg. Med. 2010, 42, 527–539. [Google Scholar] [CrossRef]
- Tani, A.; Chellini, F.; Giannelli, M.; Nosi, D.; Zecchi-Orlandini, S.; Sassoli, C. Red (635 nm), Near-Infrared (808 nm) and Violet-Blue (405 nm) Photobiomodulation Potentiality on Human Osteoblasts and Mesenchymal Stromal Cells: A Morphological and Molecular In Vitro Study. Int. J. Mol. Sci. 2018, 19, 1946. [Google Scholar] [CrossRef] [PubMed]
- Solís-López, A.; Kriebs, U.; Marx, A.; Mannebach, S.; Liedtke, W.B.; Caterina, M.J.; Freichel, M.; Tsvilovskyy, V.V. Analysis of TRPV Channel Activation by Stimulation of FCεRI and MRGPR Receptors in Mouse Peritoneal Mast Cells. PLoS ONE 2017, 12, e0171366. [Google Scholar] [CrossRef] [PubMed]
- Salehpour, F.; Khademi, M.; Hamblin, M.R. Photobiomodulation Therapy for Dementia: A Systematic Review of Pre-Clinical and Clinical Studies. J. Alzheimers Dis. 2021, 83, 1431–1452. [Google Scholar] [CrossRef] [PubMed]
- Cianferotti, L.; Gomes, A.R.; Fabbri, S.; Tanini, A.; Brandi, M.L. The Calcium-Sensing Receptor in Bone Metabolism: From Bench to Bedside and Back. Osteoporos. Int. 2015, 26, 2055–2071. [Google Scholar] [CrossRef]
- Arany, P.R.; Cho, A.; Hunt, T.D.; Sidhu, G.; Shin, K.; Hahm, E.; Huang, G.X.; Weaver, J.; Chen, A.C.; Padwa, B.L.; Hamblin, M.R.; Barcellos-Hoff, M.H.; Kulkarni, A.B.; Mooney, D.J. Photoactivation of Endogenous Latent Transforming Growth Factor-β1 Directs Dental Stem Cell Differentiation for Regeneration. Sci. Transl. Med. 2014, 6, 238ra69. [Google Scholar] [CrossRef]
- Görlach, A.; Bertram, K.; Hudecova, S.; Krizanova, O. Calcium and ROS: A Mutual Interplay. Redox Biol. 2015, 6, 260–271. [Google Scholar] [CrossRef]
- Asiry, M.A. Biological Aspects of Orthodontic Tooth Movement: A Review of Literature. Saudi J. Biol. Sci. 2018, 25, 1027–1032. [Google Scholar] [CrossRef] [PubMed]
- Jeon, H.H.; Teixeira, H.; Tsai, A. Mechanistic Insight into Orthodontic Tooth Movement Based on Animal Studies: A Critical Review. J. Clin. Med. 2021, 10, 1733. [Google Scholar] [CrossRef]
- Jin, Y.; Ding, L.; Ding, Z.; Fu, Y.; Song, Y.; Jing, Y.; Li, Q.; Zhang, J.; Ni, Y.; Hu, Q. Tensile Force-Induced PDGF-BB/PDGFRβ Signals in Periodontal Ligament Fibroblasts Activate JAK2/STAT3 for Orthodontic Tooth Movement. Sci. Rep. 2020, 10, 11269. [Google Scholar] [CrossRef] [PubMed]
- Li, P.P.; Poblano Pérez, L.; Zhang, X.; Shi, S.; Liu, Y. Mesenchymal Stromal Cells Derived from Dental Tissues: Immunomodulatory Actions in Periodontal Regeneration. Int. J. Mol. Sci. 2024, 25, 1986. [Google Scholar] [CrossRef]
- Zhao, C.; Irie, N.; Takada, Y.; Shimoda, K.; Miyamoto, T.; Nishiwaki, T.; Suda, T.; Matsuo, K. Bidirectional EphrinB2-EphB4 Signaling Controls Bone Homeostasis. Cell Metab. 2006, 4, 111–121. [Google Scholar] [CrossRef]
- Lontos, K.; Adamik, J.; Tsagianni, A.; Galson, D.L.; Chirgwin, J.M.; Suvannasankha, A. The Role of Semaphorin 4D in Bone Remodeling and Cancer Metastasis. Front. Endocrinol. 2018, 9, 322. [Google Scholar] [CrossRef]
- Loh, H.; Norman, B.P.; Lai, K.; Cheng, W.; Afizan, N.M.; Mohamed Alitheen, N.B.; Osman, M.A. Post-Transcriptional Regulatory Crosstalk between MicroRNAs and Canonical TGF-β/BMP Signalling Cascades on Osteoblast Lineage: A Comprehensive Review. Int. J. Mol. Sci. 2023, 24, 6423. [Google Scholar] [CrossRef]
- Kanzaki, H.; Wada, S.; Yamaguchi, Y.; et al. Compression and Tension Variably Alter Osteoprotegerin Expression via miR-3198 in Periodontal Ligament Cells. BMC Mol. Cell Biol. 2019, 20, 6. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhuang, Z.; Wei, Q.; Li, P.; Li, J.; Fan, Y.; Zhang, L.; Hong, Z.; He, W.; Wang, H.; Liu, Y.; Li, W. Inhibition of miR-93-5p Promotes Osteogenic Differentiation in a Rabbit Model of Trauma-Induced Osteonecrosis of the Femoral Head. FEBS Open Bio 2021, 11, 2152–2165. [Google Scholar] [CrossRef]
- Zhang, X.; Zhao, Z.; Chen, Y.; Han, X.; Jie, Y. Extracellular Vesicles Secreted by Human Periodontal Ligament Induce Osteoclast Differentiation by Transporting miR-28 to Osteoclast Precursor Cells and Further Promote Orthodontic Tooth Movement. Int. Immunopharmacol. 2022, 113, 109388. [Google Scholar] [CrossRef]
- Huang, X.; Zhao, Z.; Zhan, W.; Deng, M.; Wu, X.; Chen, Z.; Xie, J.; Ye, W.; Zhao, M.; Chu, J. miR-21-5p Enriched Exosomes from Human Embryonic Stem Cells Promote Osteogenesis via YAP1 Modulation. Int. J. Nanomedicine 2024, 19, 13095–13112. [Google Scholar] [CrossRef]
- Zheng, Y.; Dong, C.; Yang, J.; Jin, Y.; Zheng, W.; Zhou, Q.; Liang, Y.; Bao, L.; Feng, G.; Ji, J.; Feng, X.; Gu, Z. Exosomal microRNA-155-5p from PDLSCs Regulated Th17/Treg Balance by Targeting Sirtuin-1 in Chronic Periodontitis. J. Cell. Physiol. 2019, 234, 20662–20674. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, C.; Song, Y.; Wu, F. Low-Level Laser Therapy Promotes Proliferation and Osteogenic Differentiation of PDLSCs via BMP/Smad Activation in a Dose-Dependent Manner. Lasers Med. Sci. 2022, 37, 3591–3599. [Google Scholar] [CrossRef] [PubMed]
- Arya, P.N.; Saranya, I.; Selvamurugan, N. Crosstalk between Wnt and Bone Morphogenetic Protein Signaling during Osteogenic Differentiation. World J. Stem Cells 2024, 16, 102–113. [Google Scholar] [CrossRef]
- Gholami, L.; Parsamanesh, G.; Shahabi, S.; Jazaeri, M.; Baghaei, K.; Fekrazad, R. The Effect of Laser Photobiomodulation on Periodontal Ligament Stem Cells. Photochem. Photobiol. 2021, 97, 851–859. [Google Scholar] [CrossRef] [PubMed]
- Mylona, V.; Anagnostaki, E.; Chiniforush, N.; Barikani, H.; Lynch, E.; Grootveld, M. Photobiomodulation Effects on Periodontal Ligament Stem Cells: A Systematic Review of In Vitro Studies. Curr. Stem Cell Res. Ther. 2024, 19, 544–558. [Google Scholar] [CrossRef]
- Berni, M.; Brancato, A.M.; Torriani, C.; Bina, V.; Annunziata, S.; Cornella, E.; Trucchi, M.; Jannelli, E.; Mosconi, M.; Gastaldi, G.; Caliogna, L.; Grassi, F.A.; Pasta, G. The Role of Low-Level Laser Therapy in Bone Healing: Systematic Review. Int. J. Mol. Sci. 2023, 24, 7094. [Google Scholar] [CrossRef]
- Song, J.; Park, B.; Shin, S.; Kim, I. Low-Level Laser Irradiation Stimulates RANKL-Induced Osteoclastogenesis via the MAPK Pathway in RAW 264.7 Cells. Appl. Sci. 2021, 11, 5360. [Google Scholar] [CrossRef]
- Huang, C.-Y.; Le, H.H.T.; Tsai, H.-C.; Tang, C.-H.; Yu, J.-H. The Effect of Low-Level Laser Therapy on Osteoclast Differentiation: Clinical Implications for Tooth Movement and Bone Density. J. Dent. Sci. 2024, 19, 1452–1460. [Google Scholar] [CrossRef]
- Yassaei, S.; Fekrazad, R.; Shahraki, N. Effect of Low-Level Laser Therapy on Orthodontic Tooth Movement: A Review Article. J. Dent. (Tehran) 2013, 10, 264–272. [Google Scholar] [PubMed] [PubMed Central]
- Zheng, Y.; Zhang, X.; Li, C.; Wang, L.; Liu, D.; Guo, H. Low-Level Laser Therapy (LLLT) Reduces IL-1β and TNF-α and Increases IL-1RA in Gingival and Bone Tissue, Enhancing Wound Healing in MRONJ Models. Bone Rep. 2023, 18, 101079. [Google Scholar] [CrossRef]
- Li, Y.; Jacox, L.A.; Little, S.H.; Ko, C.C. Orthodontic Tooth Movement: The Biology and Clinical Implications. Kaohsiung J. Med. Sci. 2018, 34, 207–214. [Google Scholar] [CrossRef]
- Behm, C.; Nemec, M.; Weissinger, F.; Rausch, M.A.; Andrukhov, O.; Jonke, E. MMPs and TIMPs Expression Levels in the Periodontal Ligament during Orthodontic Tooth Movement: A Systematic Review of In Vitro and In Vivo Studies. Int. J. Mol. Sci. 2021, 22, 6967. [Google Scholar] [CrossRef]
- Gavish, L.; Perez, L.; Gertz, S.D. Low-Level Laser Irradiation Modulates Matrix Metalloproteinase Activity and Gene Expression in Porcine Aortic Smooth Muscle Cells. Lasers Surg. Med. 2006, 38, 779–786. [Google Scholar] [CrossRef]
- Gwack, C.; Kim, S.-S.; Park, S.B.; Park, M.-H. The Expression of MMP-1, -8, and -13 mRNA in the Periodontal Ligament of Rats during Tooth Movement with Cortical Punching. Korean J. Orthod. 2008, 38, 187–196. [Google Scholar] [CrossRef]
- Feller, L.; Khammissa, R.A.; Schechter, I.; Thomadakis, G.; Fourie, J.; Lemmer, J. Biological Events in Periodontal Ligament and Alveolar Bone Associated with Application of Orthodontic Forces. Sci. World J. 2015, 2015, 876509. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, X.; Han, K.; Wang, X.; Guo, Z.; Deng, Q.; Li, J.; Lv, S.; Yu, W. Effects of Low Level Laser on Periodontal Tissue Remodeling in hPDLCs under Tensile Stress. Lasers Med. Sci. 2023, 38, 232. [Google Scholar] [CrossRef]
- Miyagawa, A.; Chiba, M.; Hayashi, H.; Igarashi, K. Compressive Force Induces VEGF Production in Periodontal Tissues. J. Dent. Res. 2009, 88, 752–756. [Google Scholar] [CrossRef]
- Li, G.; Gao, J.; Ding, P.; Gao, Y. The Role of Endothelial Cell–Pericyte Interactions in Vascularization and Diseases. J. Adv. Res. 2025, 67, 269–288. [Google Scholar] [CrossRef]
- Firth, F.A.; Farrar, R.; Farella, M. Investigating Orthodontic Tooth Movement: Challenges and Future Directions. J. R. Soc. N. Z. 2020, 50, 67–79. [Google Scholar] [CrossRef]
- Cury, V.; Moretti, A.I.; Assis, L.; Bossini, P.; Crusca, J. de S.; Neto, C.B.; Fangel, R.; de Souza, H.P.; Hamblin, M.R.; Parizotto, N.A. Low Level Laser Therapy Increases Angiogenesis in a Model of Ischemic Skin Flap in Rats Mediated by VEGF, HIF-1α and MMP-2. J. Photochem. Photobiol. B 2013, 125, 164–170. [Google Scholar] [CrossRef]
- Bai, J.; Li, L.; Kou, N.; Bai, Y.; Zhang, Y.; Lu, Y.; Gao, L.; Wang, F. Low Level Laser Therapy Promotes Bone Regeneration by Coupling Angiogenesis and Osteogenesis. Stem Cell Res. Ther. 2021, 12, 432. [Google Scholar] [CrossRef] [PubMed]
- Bloemen, V.; Schoenmaker, T.; de Vries, T.J.; Everts, V. Direct Cell–Cell Contact between Periodontal Ligament Fibroblasts and Osteoclast Precursors Synergistically Increases the Expression of Genes Related to Osteoclastogenesis. J. Cell. Physiol. 2010, 222, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Klein-Nulend, J.; van Oers, R.F.; Bakker, A.D.; Bacabac, R.G. Nitric Oxide Signaling in Mechanical Adaptation of Bone. Osteoporos. Int. 2014, 25, 1427–1437. [Google Scholar] [CrossRef]
- Li, Y.; Xu, Q.; Shi, M.; Gan, P.; Huang, Q.; Wang, A.; Tan, G.; Fang, Y.; Liao, H. Low-Level Laser Therapy Induces Human Umbilical Vascular Endothelial Cell Proliferation, Migration and Tube Formation through Activating the PI3K/Akt Signaling Pathway. Microvasc. Res. 2020, 129, 103959. [Google Scholar] [CrossRef]
- Szymczyszyn, A.; Doroszko, A.; Szahidewicz-Krupska, E.; Rola, P.; Gutherc, R.; Jasiczek, J.; Mazur, G.; Derkacz, A. Effect of the Transdermal Low-Level Laser Therapy on Endothelial Function. Lasers Med. Sci. 2016, 31, 1301–1307. [Google Scholar] [CrossRef]
- Mariño, G.; Niso-Santano, M.; Baehrecke, E.H.; Kroemer, G. Self-consumption: the interplay of autophagy and apoptosis. Nat. Rev. Mol. Cell Biol. 2014, 15, 81–94. [Google Scholar] [CrossRef]
- Moin, S.; Kalajzic, Z.; Utreja, A.; Nihara, J.; Wadhwa, S.; Uribe, F.; Nanda, R. Osteocyte death during orthodontic tooth movement in mice. Angle Orthod. 2014, 84, 1086–1092. [Google Scholar] [CrossRef]
- Duggal, R.; Singh, N. Detection of apoptosis in human periodontal ligament during orthodontic tooth movement. J. Dent. Spec. 2015, 3, 217. [Google Scholar] [CrossRef]
- Wu, Y.; Zhuang, J.; Zhao, D.; Xu, C. Interaction between caspase-3 and caspase-5 in the stretch-induced programmed cell death in the human periodontal ligament cells. J. Cell Physiol. 2019, 234, 13571–13581. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.W.; Yao, C.C.; Jeng, J.H.; Shieh, H.Y.; Chen, Y.J. Periostin inhibits mechanical stretch-induced apoptosis in osteoblast-like MG-63 cells. J. Formos. Med. Assoc. 2018, 117, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhan, Q.; Bao, M.; Yi, J.; Li, Y. Biomechanical and biological responses of periodontium in orthodontic tooth movement: up-date in a new decade. Int. J. Oral Sci. 2021, 13, 20. [Google Scholar] [CrossRef]
- Kaplan, M.; Kalajzic, Z.; Choi, T.; Maleeh, I.; Ricupero, C.L.; Skelton, M.N.; Daily, M.L.; Chen, J.; Wadhwa, S. The role of inhibition of osteocyte apoptosis in mediating orthodontic tooth movement and periodontal remodeling: A pilot study. Prog. Orthod. 2021, 22, 21. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, Y.; Zhang, Y.; Huang, Y.; Yang, Y.; Zhao, Y.; Chen, S.; Deng, J.; Li, W.; Han, B. Periodontal ligament cell apoptosis activates Lepr⁺ osteoprogenitors in orthodontics. J. Dent. Res. 2024, 103, 937–947. [Google Scholar] [CrossRef]
- Shen, X.; Wu, W.; Ying, Y.; Zhou, L.; Zhu, H. A regulatory role of Piezo1 in apoptosis of periodontal tissue and periodontal ligament fibroblasts during orthodontic tooth movement. Aust. Endod. J. 2023, 49 (Suppl. 1), 228–237. [Google Scholar] [CrossRef]
- Brockhaus, J.; Craveiro, R.B.; Azraq, I.; Niederau, C.; Schröder, S.K.; Weiskirchen, R.; Jankowski, J.; Wolf, M. In vitro compression model for orthodontic tooth movement modulates human periodontal ligament fibroblast proliferation, apoptosis and cell cycle. Biomolecules 2021, 11, 932. [Google Scholar] [CrossRef]
- Pansani, T.N.; Basso, F.G.; Turirioni, A.P.; Kurachi, C.; Hebling, J.; de Souza Costa, C.A. Effects of low-level laser therapy on the proliferation and apoptosis of gingival fibroblasts treated with zoledronic acid. Int. J. Oral Maxillofac. Surg. 2014, 43, 1030–1034. [Google Scholar] [CrossRef]
- Chu, Y.H.; Chen, S.Y.; Hsieh, Y.L.; Teng, Y.H.; Cheng, Y.J. Low-level laser therapy prevents endothelial cells from TNF-α/cycloheximide-induced apoptosis. Lasers Med. Sci. 2018, 33, 279–286. [Google Scholar] [CrossRef] [PubMed]
- de Souza Oliveira, C.; de Oliveira, H.A.; Teixeira, I.L.A.; Antonio, E.L.; Silva, F.A.; Sunemi, S.; Leal-Junior, E.C.; de Tarso Camillo de Carvalho, P.; Tucci, P.J.F.; Serra, A.J. Low-level laser therapy prevents muscle apoptosis induced by a high-intensity resistance exercise in a dose-dependent manner. Lasers Med. Sci. 2020, 35, 1867–1870. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, L.; Lin, F.; Zheng, Q.; Xu, X.; Mei, L. Dynamic study into autophagy and apoptosis during orthodontic tooth movement. Exp. Ther. Med. 2021, 21, 430. [Google Scholar] [CrossRef]
- Li, Y.; Jacox, L.A.; Coats, S.; Kwon, J.; Xue, P.; Tang, N.; Rui, Z.; Wang, X.; Kim, Y.I.; Wu, T.J.; Lee, Y.T.; Wong, S.W.; Chien, C.H.; Cheng, C.W.; Gross, R.; Lin, F.C.; Tseng, H.; Martinez, J.; Ko, C.C. Roles of autophagy in orthodontic tooth movement. Am. J. Orthod. Dentofacial Orthop. 2021, 159, 582–593. [Google Scholar] [CrossRef]
- Zheng, J.; Xu, B.; Yang, K. Autophagy regulates osteogenic differentiation of human periodontal ligament stem cells induced by orthodontic tension. Stem Cells Int. 2022, 2022, 2983862. [Google Scholar] [CrossRef]
- Li, B.; Wang, L.; He, H. Autophagy in orthodontic tooth movement: advances, challenges, and future perspectives. Mol. Med. 2025, 31, 245. [Google Scholar] [CrossRef] [PubMed]
- Zuo, X.; Wei, X.; Ju, C.; Wang, X.; Zhang, Z.; Ma, Y.; Zhu, Z.; Li, X.; Song, Z.; Luo, L.; Hu, X.; Wang, Z. Protective effect of photobiomodulation against hydrogen peroxide-induced oxidative damage by promoting autophagy through inhibition of PI3K/AKT/mTOR pathway in MC3T3-E1 cells. Oxid. Med. Cell Longev. 2022, 2022, 7223353. [Google Scholar] [CrossRef]
- Li, H.; Yang, W.; Zhu, B.; Li, M.; Zhang, X. Photobiomodulation therapy at 650 nm enhances osteogenic differentiation of osteoporotic bone marrow mesenchymal stem cells through modulating autophagy. Photodiagnosis Photodyn. Ther. 2024, 50, 104389. [Google Scholar] [CrossRef]
- Silva, H.N.M.D.; Fernandes, E.M.; Pereira, V.A.; Mizobuti, D.S.; Covatti, C.; Rocha, G.L.D.; Minatel, E. LEDT and idebenone treatment modulate autophagy and improve regenerative capacity in the dystrophic muscle through an AMPK-pathway. PLoS ONE 2024, 19, e0300006. [Google Scholar] [CrossRef] [PubMed]
- Jeon, H.H.; Huang, X.; Rojas Cortez, L.; Sripinun, P.; Lee, J.M.; Hong, J.J.; Graves, D.T. Inflammation and mechanical force-induced bone remodeling. Periodontol. 2000 2024, in press. [Google Scholar] [CrossRef]
- Wan Hassan, W.N.; Waddington, R.J. Immunology of tooth movement and root resorption in orthodontics. In Title of Book or Book Series; Publisher: Place, Year; pp. 134–155. [CrossRef]
- Hajishengallis, G. New developments in neutrophil biology and periodontitis. Periodontol 2000. 2020, 82, 78–92. [Google Scholar] [CrossRef] [PubMed]
- Marcaccini, A.M.; Amato, P.A.; Leão, F.V.; Gerlach, R.F.; Ferreira, J.T. Myeloperoxidase activity is increased in gingival crevicular fluid and whole saliva after fixed orthodontic appliance activation. Am. J. Orthod. Dentofacial Orthop. 2010, 138, 613–616. [Google Scholar] [CrossRef] [PubMed]
- Zeng, M.; Kou, X.; Yang, R.; Liu, D.; Wang, X.; Song, Y.; Zhang, J.; Yan, Y.; Liu, F.; He, D.; Gan, Y.; Zhou, Y. Orthodontic force induces systemic inflammatory monocyte responses. J. Dent. Res. 2015, 94, 1295–1302. [Google Scholar] [CrossRef]
- He, D.; Kou, X.; Yang, R.; Liu, D.; Wang, X.; Luo, Q.; Song, Y.; Liu, F.; Yan, Y.; Gan, Y.; Zhou, Y. M1-like macrophage polarization promotes orthodontic tooth movement. J. Dent. Res. 2015, 94, 1286–1294. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Cai, X.; Ren, F.; Ye, Y.; Wang, F.; Zheng, C.; Qian, Y.; Zhang, M. The macrophage-osteoclast axis in osteoimmunity and osteo-related diseases. Front. Immunol. 2021, 12, 664871. [Google Scholar] [CrossRef] [PubMed]
- Mills, C.D. M1 and M2 macrophages: oracles of health and disease. Crit. Rev. Immunol. 2012, 32, 463–488. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Zhang, S.; Sathe, A.A.; Lin, Y.; Zhang, Q.; Xu, L.; Yang, P.; Zhong, W.; Wang, Y.; Qian, Y. CCR2⁺ macrophages promote orthodontic tooth movement and alveolar bone remodeling. Front. Immunol. 2022, 13, 835986. [Google Scholar] [CrossRef] [PubMed]
- Klein, Y.; Fleissig, O.; Polak, D.; Barenholz, Y.; Mandelboim, O.; Chaushu, S. Immunorthodontics: in vivo gene expression of orthodontic tooth movement. Sci. Rep. 2020, 10, 8172. [Google Scholar] [CrossRef]
- Schroder, A.; Kappler, P.; Nazet, U.; Jantsch, J.; Proff, P.; Cieplik, F.; Deschner, J.; Kirschneck, C. Effects of compressive and tensile strain on macrophages during simulated orthodontic tooth movement. Mediators Inflamm. 2020, 2020, 2814015. [Google Scholar] [CrossRef]
- Gu, Q.; Yang, H.; Shi, Q. Macrophages and bone inflammation. J. Orthop. Translat. 2017, 10, 86–93. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, H.; Sun, W.; Wang, Y.; Ren, L.; Wei, W.; Bai, D. Macrophages mediate corticotomy-accelerated orthodontic tooth movement. Sci. Rep. 2018, 8, 16788. [Google Scholar] [CrossRef] [PubMed]
- Vandevska-Radunovic, V.; Kvinnsland, I.H.; Kvinnsland, S.; Jonsson, R. Immunocompetent cells in rat periodontal ligament and their recruitment incident to experimental orthodontic tooth movement. Eur. J. Oral Sci. 1997, 105, 36–44. [Google Scholar] [CrossRef]
- Wald, S.; Leibowitz, A.; Aizenbud, Y.; Chaushu, S.; Reuven, Y.; Shapira, Y.; Dardick, A.; Redlich, M. γδT cells are essential for orthodontic tooth movement. J. Dent. Res. 2021, 100, 731–738. [Google Scholar] [CrossRef]
- Pastille, E.; Konermann, A. Exploring the Role of Innate Lymphoid Cells in the Periodontium: Insights into Immunological Dynamics during Orthodontic Tooth Movement. Front. Immunol. 2024, 15, 1428059. [Google Scholar] [CrossRef]
- Prathapan Santhakumari, P.; Varma Raja, V.; Joseph, J.; Devaraj, A.; John, E.; Oommen Thomas, N. Impact of low-level laser therapy on orthodontic tooth movement and various cytokines in gingival crevicular fluid: a split-mouth randomized study. Cureus 2023, 15, e42809. [Google Scholar] [CrossRef]
- Zheng, J.; Yang, K. Clinical research: low-level laser therapy in accelerating orthodontic tooth movement. BMC Oral Health 2021, 21, 324. [Google Scholar] [CrossRef]
- Fernandes, M.R.U.; Suzuki, S.S.; Suzuki, H.; Martinez, E.F.; Garcez, A.S. Photobiomodulation increases intrusion tooth movement and modulates IL-6, IL-8 and IL-1β expression during orthodontically bone remodeling. J. Biophotonics 2019, 12, e201800311. [Google Scholar] [CrossRef]
- Reis, C.L.B.; de Souza Furtado, T.C.; Mendes, W.D.; Matsumoto, M.A.N.; Alves, S.Y.F.; Stuani, M.B.S.; Borsatto, M.C.; Corona, S.A.M. Photobiomodulation impacts the levels of inflammatory mediators during orthodontic tooth movement? A systematic review with meta-analysis. Lasers Med. Sci. 2022, 37, 771–787. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, P.; Kharbanda, O.P.; Monga, N.; Miglani, R.; Kapila, S. Effect of orthodontic forces on cytokine and receptor levels in gingival crevicular fluid: a systematic review. Prog. Orthod. 2014, 15, 65. [Google Scholar] [CrossRef]
- Cerdeira, C.D.; Lima Brigagão, M.R.; Carli, M.L.; de Souza Ferreira, C.; de Oliveira Isac Moraes, G.; Hadad, H.; Costa Hanemann, J.A.; Hamblin, M.R.; Sperandio, F.F. Low-level laser therapy stimulates the oxidative burst in human neutrophils and increases their fungicidal capacity. J. Biophotonics 2016, 9, 1180–1188. [Google Scholar] [CrossRef] [PubMed]
- Wasik, M.; Gorska, E.; Modzelewska, M.; Nowicki, K.; Jakubczak, B.; Demkow, U. The influence of low-power helium-neon laser irradiation on function of selected peripheral blood cells. J. Physiol. Pharmacol. 2007, 58 Pt 2 (Suppl. 5), 729–737. [Google Scholar] [PubMed]
- Kujawa, J.; Agier, J.; Kozłowska, E. Impact of photobiomodulation therapy on pro-inflammation functionality of human peripheral blood mononuclear cells – a preliminary study. Sci. Rep. 2024, 14, 1–11. [Google Scholar] [CrossRef]
- Chen, H.; Wang, C.Z.; Wang, Y.H.; Liao, W.T.; Chen, Y.J.; Kuo, C.H.; Kuo, H.F.; Hung, C.H. Effects of low-level laser therapy on M1-related cytokine expression in monocytes via histone modification. Mediators Inflamm. 2014, 2014, 625048. [Google Scholar] [CrossRef]
- Tian, T.; Wang, Z.; Chen, L.; Xu, W.; Wu, B. Photobiomodulation activates undifferentiated macrophages and promotes M1/M2 macrophage polarization via PI3K/AKT/mTOR signaling pathway. Lasers Med. Sci. 2023, 38, 86. [Google Scholar] [CrossRef]
- Fernandes, K.P.S.; Souza, N.H.C.; Mesquita-Ferrari, R.A.; Silva, D.d.F.T.d.; Rocha, L.A.; Alves, A.N.; et al. Photobiomodulation with 660-nm and 780-nm laser on activated J774 macrophage-like cells: effect on M1 inflammatory markers. J. Photochem. Photobiol. B Biol. 2015, 153, 344–351. [Google Scholar] [CrossRef]
- Song, J.W.; Li, K.; Liang, Z.W.; Dai, C.; Shen, X.F.; Gong, Y.Z.; Wang, S.; Hu, X.Y.; Wang, Z. Low-level laser facilitates alternatively activated macrophage/microglia polarization and promotes functional recovery after crush spinal cord injury in rats. Sci. Rep. 2017, 7, 620. [Google Scholar] [CrossRef]
- Zhang, J.; Sun, J.; Zheng, Q.; Hu, X.; Wang, Z.; Liang, Z.; Li, K.; Song, J.; Ding, T.; Shen, X.; Zhang, J.; Qiao, L. Low-level laser therapy 810-nm up-regulates macrophage secretion of neurotrophic factors via PKA-CREB and promotes neuronal axon regeneration in vitro. J. Cell. Mol. Med. 2020, 24, 476–486. [Google Scholar] [CrossRef]
- Ng, M.Y.; Lin, T.; Chen, S.H.; Liao, Y.W.; Liu, C.M.; Yu, C.C. Er:YAG laser suppresses pro-inflammatory cytokines expression and inflammasome in human periodontal ligament fibroblasts with Porphyromonas gingivalis-lipopolysaccharide stimulation. J. Dent. Sci. 2024, 19, 1135–1142. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.H.; Park, J.; Kim, B.Y.; Kim, Y.; Kim, N.G.; Shin, Y.I. Photobiomodulation ameliorates inflammatory parameters in fibroblast-like synoviocytes and experimental animal models of rheumatoid arthritis. Front. Immunol. 2023, 14, 1122581. [Google Scholar] [CrossRef]
- Lee, J.H.; Chiang, M.H.; Chen, P.H.; Ho, M.L.; Lee, H.E.; Wang, Y.H. Anti-inflammatory effects of low-level laser therapy on human periodontal ligament cells: in vitro study. Lasers Med. Sci. 2018, 33, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Wada, N.; Tomokiyo, A.; Gronthos, S.; et al. Immunomodulatory properties of PDLSC and relevance to periodontal regeneration. Curr. Oral Health Rep. 2015, 2, 245–251. [Google Scholar] [CrossRef]
- Oner, F.; Kantarci, A. Periodontal Response to Nonsurgical Accelerated Orthodontic Tooth Movement. Periodontol. 2000, Epub ahead of print. 2025. [Google Scholar] [CrossRef]
- Nugraha, A.P.; Khoswanto, C.; Kriswandini, I.L. The involvement of damage associated molecular pattern and resolution associated molecular pattern in alveolar bone remodeling during orthodontic tooth movement: narrative review. Teikyo Med. J. 2022, 45, 1. [Google Scholar]
- Miron, R.J.; Bohner, M.; Zhang, Y.; Bosshardt, D.D. Osteoinduction and Osteoimmunology: Emerging Concepts. Periodontology 2000 2024, 94, 9–26. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Wong, P.; Li, J.; Lv, Z.; Xu, L.; Zhu, G.; He, M.; Luo, Y. Osteoimmunology: the correlation between osteoclasts and the Th17/Treg balance in osteoporosis. J. Cell. Mol. Med. 2022, 26, 3591–3597. [Google Scholar] [CrossRef]
- Lin, T.; Yang, L.; Zheng, W.; Zhang, B. Matrix metalloproteinases and Th17 cytokines in the gingival crevicular fluid during orthodontic tooth movement. Eur. J. Paediatr. Dent. 2021, 22, 135–138. [Google Scholar]
- Tang M, Lu L, Yu X. Interleukin-17A Interweaves the Skeletal and Immune Systems. Front Immunol. 2021, 11, 625034. [CrossRef] [PubMed]
- Zhang, H.; Wang, L.; Liu, A.; Zhou, H.; Liang, X.; Kang, N. The IL-17 level in gingival crevicular fluid as an indicator of orthodontically induced inflammatory root resorption. J. Orofac. Orthop. Epub ahead of print]. 2025. [Google Scholar] [CrossRef]
- Gao, Y.; Grassi, F.; Ryan, M.R.; Terauchi, M.; Page, K.; Yang, X.; Weitzmann, M.N.; Pacifici, R. IFN-gamma stimulates osteoclast formation and bone loss in vivo via antigen-driven T cell activation. J. Clin. Invest. 2007, 117, 122–132. [Google Scholar] [CrossRef]
- Jiang, C.; Li, Z.; Quan, H.; Xiao, L.; Zhao, J.; Jiang, C.; Wang, Y.; Liu, J.; Gou, Y.; An, S.; Huang, Y.; Yu, W.; Zhang, Y.; He, W.; Yi, Y.; Chen, Y.; Wang, J. Osteoimmunology in orthodontic tooth movement. Oral Dis. 2015, 21, 694–704. [Google Scholar] [CrossRef]
- Cheng, J.; Liu, J.; Shi, Z.; Xu, D.; Luo, S.; Siegal, G.P.; Feng, X.; Wei, S. Interleukin-4 inhibits RANKL-induced NFATc1 expression via STAT6: a novel mechanism mediating its blockade of osteoclastogenesis. J. Cell. Biochem. 2011, 112, 3385–3392. [Google Scholar] [CrossRef] [PubMed]
- te Velde, A.A.; Huijbens, R.J.; Heije, K.; de Vries, J.E.; Figdor, C.G. Interleukin-4 (IL-4) inhibits secretion of IL-1 beta, tumor necrosis factor alpha, and IL-6 by human monocytes. Blood 1990, 76, 1392–1397. [Google Scholar] [CrossRef] [PubMed]
- Behm, C.; Zhao, Z.; Andrukhov, O. Immunomodulatory activities of periodontal ligament stem cells in orthodontic forces-induced inflammatory processes: current views and future perspectives. Front. Oral Health 2022, 3, 877348. [Google Scholar] [CrossRef]
- Settem, R.P.; Honma, K.; Chinthamani, S.; Kawai, T.; Sharma, A. B-cell RANKL contributes to pathogen-induced alveolar bone loss in an experimental periodontitis mouse model. Front. Physiol. 2021, 12, 722859. [Google Scholar] [CrossRef]
- Harada, Y.; Han, X.; Yamashita, K.; Kitami, M.; Owaki, T.; Huang, G.T.; Sasaki, K.; Kurihara, H. Effect of adoptive transfer of antigen-specific B cells on periodontal bone resorption. J. Periodontal Res. 2006, 41, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Terauchi, M.; Vikulina, T.; Roser-Page, S.; Weitzmann, M.N. B Cell Production of Both OPG and RANKL Is Significantly Increased in Aged Mice. Open Bone J. 2014, 6, 8–17. [Google Scholar] [CrossRef]
- Sumida, T.S.; Cheru, N.T.; Hafler, D.A. The Regulation and Differentiation of Regulatory T Cells and Their Dysfunction in Autoimmune Diseases. Nat. Rev. Immunol. 2024, 24, 503–517. [Google Scholar] [CrossRef]
- Wang, W.; Wang, X.; Lu, S.; Lv, H. Metabolic Disturbance and Th17/Treg Imbalance Are Associated with Progression of Gingivitis. Front. Immunol. 2021, 12, 670178. [Google Scholar] [CrossRef]
- Yang, M.; Zhu, L. Osteoimmunology: The Crosstalk between T Cells, B Cells, and Osteoclasts in Rheumatoid Arthritis. Int. J. Mol. Sci. 2023, 25, 2688. [Google Scholar] [CrossRef]
- Ge, N.; Peng, J.; Yu, L.; Huang, S.; Xu, L.; Su, Y.; Chen, L. Orthodontic Treatment Induces Th17/Treg Cells to Regulate Tooth Movement in Rats with Periodontitis. Iran J. Basic Med. Sci. 2020, 23, 1315–1322. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Huang, J.; Zhang, Z.; Yu, X.; Cai, X.; Liu, C. Periodontal Ligament Cells under Mechanical Force Regulate Local Immune Homeostasis by Modulating Th17/Treg Cell Differentiation. Clin. Oral Investig. 2022, 26, 3747–3764. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Xia, Y.; Gao, H.; Wang, C.; Jiang, Y.; Song, W.; Yu, J.F.; Liang, L. Regulatory T Cells Promote Osteogenic Differentiation of Periodontal Ligament Stem Cells through the Jagged1-Notch2 Signaling Axis. J. Dent. 2025, 158, 105772. [Google Scholar] [CrossRef]
- Chansaenroj, J.; Suwittayarak, R.; Egusa, H.; Samaranayake, L.P.; Osathanon, T. Mechanical Force Modulates Inflammation and Immunomodulation in Periodontal Ligament Cells. Med. Rev. 2024, 4, 544–548. [Google Scholar] [CrossRef]
- Shin, C.; Kim, M.; Han, J.A.; Choi, B.; Hwang, D.; Do, Y.; Yun, J.H. Human Periodontal Ligament Stem Cells Suppress T-Cell Proliferation via Down-Regulation of Non-Classical Major Histocompatibility Complex-Like Glycoprotein CD1b on Dendritic Cells. J. Periodontal Res. 2017, 52, 135–146. [Google Scholar] [CrossRef]
- Stadler, I.; Evans, R.; Kolb, B.; Naim, J.O.; Narayan, V.; Buehner, N.; Lanzafame, R.J. In vitro effects of low-level laser irradiation at 660 nm on peripheral blood lymphocytes. Lasers Surg. Med. 2000, 27, 255–261. [Google Scholar] [CrossRef]
- Gulsoy, M.; Ozer, G.H.; Bozkulak, O.; Tabakoglu, H.O.; Aktas, E.; Deniz, G.; Ertan, C. The biological effects of 632.8-nm low energy He-Ne laser on peripheral blood mononuclear cells in vitro. J. Photochem. Photobiol. B 2006, 82, 199–202. [Google Scholar] [CrossRef]
- Al Musawi, M.S.; Jaafar, M.S.; Al-Gailani, B.; Ahmed, N.M.; Suhaimi, F.M.; Suardi, N. Effects of low-level laser irradiation on human blood lymphocytes in vitro. Lasers Med. Sci. 2017, 32, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Shen, Q.; Chang, H.; Li, J.; Xing, D. Promoted CD4+ T cell-derived IFN-γ/IL-10 by photobiomodulation therapy modulates neurogenesis to ameliorate cognitive deficits in APP/PS1 and 3xTg-AD mice. J. Neuroinflammation 2022, 19, 253. [Google Scholar] [CrossRef]
- Kawakubo, M.; Cunningham, T.J.; Demehri, S.; et al. Fractional laser releases tumor-associated antigens in poorly immunogenic tumor and induces systemic immunity. Sci. Rep. 2017, 7, 12751. [Google Scholar] [CrossRef] [PubMed]
- de Brito, A.A.; Gonçalves Santos, T.; Herculano, K.Z.; Estefano-Alves, C.; de Alvarenga Nascimento, C.R.; Rigonato-Oliveira, N.C.; Chavantes, M.C.; Aimbire, F.; da Palma, R.K.; Ligeiro de Oliveira, A.P. Photobiomodulation therapy restores IL-10 secretion in a murine model of chronic asthma: relevance to the population of CD4+CD25+Foxp3+ cells in lung. Front. Immunol. 2022, 12, 789426. [Google Scholar] [CrossRef] [PubMed]
- Dos Anjos, L.M.J.; Salvador, P.A.; de Souza, Á.C.; de Souza da Fonseca, A.; de Paoli, F.; Gameiro, J. Modulation of immune response to induced-arthritis by low-level laser therapy. J. Biophotonics 2019, 12, e201800120. [Google Scholar] [CrossRef]
- Choi, D.H.; Lim, J.H.; Lee, K.H.; Kim, M.Y.; Kim, H.Y.; Shin, C.Y.; Han, S.H.; Lee, J. Effect of 710-nm visible light irradiation on neuroprotection and immune function after stroke. Neuroimmunomodulation 2012, 19, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, A.; Albuquerque-Souza, E.; Gasparoni, L.M.; de França, B.N.; Pelissari, C.; Trierveiler, M.; Holzhausen, M. Therapeutic potential of periodontal ligament stem cells. World J. Stem Cells 2021, 13, 605–618. [Google Scholar] [CrossRef] [PubMed]
- Weiss, A.R.R.; Dahlke, M.H. Immunomodulation by mesenchymal stem cells (MSCs): mechanisms of action of living, apoptotic, and dead MSCs. Front. Immunol. 2019, 10, 1191. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Interactions between innate and adaptive immune cells, stromal components of the periodontal ligament and alveolar bone during orthodontic tooth movement (OTM), and the modulatory effects of photobiomodulation. Early and late OTM phases very often exhibit opposite immune–stromal interaction patterns, both in the type and intensity of signaling. Blue lines indicate mechanical force effects; red lines indicate low-level laser therapy (LLLT) effects. Thick red lines mark known LLLT target cells. Abbreviations: IL – Interleukin; ILC – Innate lymphoid cell; DC – Dendritic cell; PDLSC – Periodontal ligament stromal/stem cell; RANKL – Receptor activator of nuclear factor kappa-B ligand; NF-κB – Nuclear factor kappa-light-chain-enhancer of activated B cells; MAPK – Mitogen-activated protein kinase; Treg – Regulatory T cell; NO – Nitric oxide; TNF-α – Tumor necrosis factor alpha; MMP9 – Matrix metalloproteinase-9; PGE₂ – Prostaglandin E₂; IFN-γ – Interferon gamma; TGF-β – Transforming growth factor beta; GM-CSF – Granulocyte–macrophage colony-stimulating factor; JAK/STAT3 – Janus kinase/signal transducer and activator of transcription 3; RUNX2 – Runt-related transcription factor 2; Osx – Osterix; ALP – Alkaline phosphatase; BMP2 – Bone morphogenetic protein 2; Smad – Mothers against decapentaplegic homolog; IDO – Indoleamine 2,3-dioxygenase; Arg-1 – Arginase-1; Jagged-1 – Jagged canonical Notch ligand 1; VEGF – Vascular endothelial growth factor; DAMPs – Damage-associated molecular patterns; OPG – Osteoprotegerin.
Figure 1.
Interactions between innate and adaptive immune cells, stromal components of the periodontal ligament and alveolar bone during orthodontic tooth movement (OTM), and the modulatory effects of photobiomodulation. Early and late OTM phases very often exhibit opposite immune–stromal interaction patterns, both in the type and intensity of signaling. Blue lines indicate mechanical force effects; red lines indicate low-level laser therapy (LLLT) effects. Thick red lines mark known LLLT target cells. Abbreviations: IL – Interleukin; ILC – Innate lymphoid cell; DC – Dendritic cell; PDLSC – Periodontal ligament stromal/stem cell; RANKL – Receptor activator of nuclear factor kappa-B ligand; NF-κB – Nuclear factor kappa-light-chain-enhancer of activated B cells; MAPK – Mitogen-activated protein kinase; Treg – Regulatory T cell; NO – Nitric oxide; TNF-α – Tumor necrosis factor alpha; MMP9 – Matrix metalloproteinase-9; PGE₂ – Prostaglandin E₂; IFN-γ – Interferon gamma; TGF-β – Transforming growth factor beta; GM-CSF – Granulocyte–macrophage colony-stimulating factor; JAK/STAT3 – Janus kinase/signal transducer and activator of transcription 3; RUNX2 – Runt-related transcription factor 2; Osx – Osterix; ALP – Alkaline phosphatase; BMP2 – Bone morphogenetic protein 2; Smad – Mothers against decapentaplegic homolog; IDO – Indoleamine 2,3-dioxygenase; Arg-1 – Arginase-1; Jagged-1 – Jagged canonical Notch ligand 1; VEGF – Vascular endothelial growth factor; DAMPs – Damage-associated molecular patterns; OPG – Osteoprotegerin.
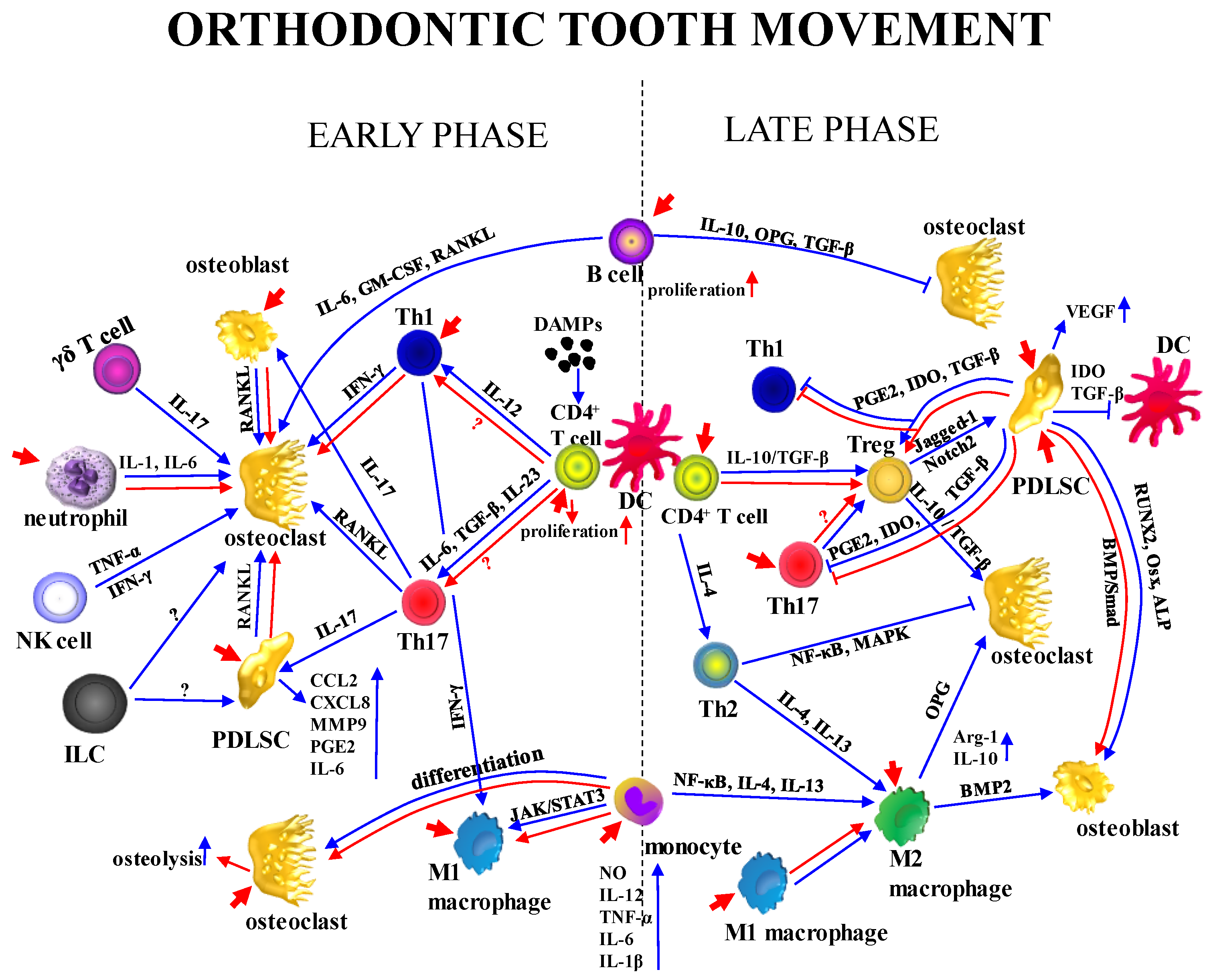
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



