
Submitted:
11 August 2025
Posted:
12 August 2025
You are already at the latest version
Abstract
Exosomes are lipid bilayer vesicles measuring 30–150 nm in diameter that serve as crucial mediators of intercellular communication. By transporting bioactive molecules such as proteins and nucleic acids, they play a pivotal role in cancer progression. Among their cargo, exosomal microRNAs (miRNAs) are central to epigenetic regulation and signal transduction. Cancer stem cells (CSCs), a distinct subpopulation with self-renewal and multi-lineage differentiation capabilities, drive tumor initiation, metastasis, and recurrence, making them critical therapeutic targets. This review systematically summarizes the regulatory mechanisms of exosomal miRNAs in tumorigenesis, metastasis, and drug resistance, with a particular focus on their roles in maintaining CSC properties and promoting therapy resistance. We further discuss their clinical potential as liquid biopsy biomarkers and therapeutic targets. Finally, we outline current research limitations and future directions to advance precision oncology.
Keywords:
exosomes
; miRNA
; cancer
; CSCs
1. Introduction
Cancer remains one of the most formidable threats to global public health. According to the latest global cancer burden report (2022), approximately 20 million new cancer cases were diagnosed worldwide, with cancer-related deaths reaching 9.7 million [1]. While significant advancements in cancer therapeutics have improved patient survival rates and quality of life, clinical outcomes continue to be hampered by tumor recurrence, metastasis, and drug resistance [2]. Emerging evidence highlights the pivotal role of CSCs in tumor progression and therapy failure. Characterized by their self-renewal capacity, multilineage differentiation potential, and intrinsic resistance to conventional therapies, CSCs are now recognized as key drivers of tumor initiation, metastatic dissemination, and post-treatment relapse [3]. The dynamic interplay between CSCs and their tumor microenvironment (TME), particularly through intercellular communication, has recently emerged as a critical area of cancer research. Among the mediators of cell-cell crosstalk, exosomes (30-150 nm extracellular vesicles) have gained considerable attention as essential information carriers [4]. These nanoscale vesicles transport bioactive molecules, including miRNAs, which post-transcriptionally regulate gene expression in recipient cells. Notably, exosomal miRNAs have been implicated in various hallmarks of cancer, from tumorigenesis to metastatic colonization [5]. Of particular interest, recent studies reveal that exosome-derived miRNAs participate in sophisticated regulatory networks that govern CSC stemness maintenance, therapy resistance, and metastatic competence [6]. Elucidating the molecular mechanisms by which exosomal miRNAs regulate CSCs and cancer progression holds dual significance: it provides novel insights into tumor heterogeneity and recurrence while offering promising avenues for developing CSC-targeted therapeutic strategies. This research direction may ultimately overcome current clinical challenges in cancer treatment, particularly in addressing therapeutic resistance and preventing metastatic spread.
2. Biogenesis and miRNA Sorting of Exosomes
Exosomes are extracellular vesicles containing various components such as proteins, nucleic acids, and lipids. They are widely present in biological fluids and participate in functions including intercellular information transmission, immune responses, and tissue repair. Additionally, they can serve as carriers for targeted tumor delivery, enhancing therapeutic efficacy. Notably, due to their natural biocompatibility and targeting ability, exosomes have emerged as ideal carriers for novel drug delivery systems, demonstrating significant advantages in targeted tumor therapy [7]. The biogenesis of exosomes is a highly programmed and precisely regulated multi-step process (Figure 1). This process begins with the internalization of the plasma membrane: under clathrin-mediated endocytosis, the plasma membrane invaginates to form early endosomes. Subsequently, under the precise regulation of the endosomal sorting complex required for transport (ESCRT) system, the early endosomal membrane buds inward to form multiple intraluminal vesicles (ILVs), gradually developing into mature multivesicular bodies (MVBs) [8,9]. The ESCRT system, through a cascade reaction of its core components (ESCRT-0 to ESCRT-III) in collaboration with ATPases such as VPS4, accomplishes the selective sorting of cargo molecules (e.g., transmembrane proteins, nucleic acids) and vesicle formation [10,11,12]. The fate determination of MVBs is a critical regulatory node in exosome generation: on one hand, MVBs can fuse with lysosomes and be degraded by hydrolytic enzymes such as cathepsins, a process regulated by GTPases like Rab7; on the other hand, mediated by GTPases such as Rab27a/b and Rab35, as well as SNARE proteins (e.g., VAMP7, Syntaxin-4), MVBs migrate directionally to the plasma membrane and fuse with it, releasing ILVs into the extracellular space via exocytosis, thereby forming exosomes [13]. The release of exosomes is a tightly regulated process involving the coordinated action of multiple Rab GTPases, SNARE proteins, and cytoskeletal components. This ensures their spatiotemporally precise secretion, enabling exosomes to mediate critical biological functions, including intercellular communication, material transport, and regulatory signaling [14].
The sorting of miRNAs into exosomes is a highly sophisticated and precisely regulated process involving multiple mechanisms: (1) Lipid microdomain-dependent sorting mediated by membrane components like cholesterol and sphingolipids, with nSMase2 playing a key regulatory role as demonstrated by Kosaka et al. [15] and subsequent breast cancer studies [16]; (2) RBP-mediated sorting through specific recognition of miRNA motifs by proteins including hnRNPA2B1 (binding GGAG/GGCU sequences) [17], YBX1 (interacting with cold shock domains) [18,19], and Ago2 (affecting miR-142-3p, miR-150 and miR-451 levels) [20]; and (3) 3'-end modification-dependent sorting evidenced by uridylation patterns in urinary and B-cell exosomal miRNAs [21]. While these mechanisms are known to be cell type- and microenvironment-dependent, current understanding remains incomplete due to technical limitations in exosome isolation and miRNA detection, necessitating future multi-omics approaches to fully elucidate the molecular mechanisms and biological significance of exosomal miRNA.
3. Cancer and Cancer Stem Cells
Cancer stem cells (CSCs) represent a distinct subpopulation within tumor tissues that possess stem cell-like properties including self-renewal capacity, multi-lineage differentiation potential, and tumorigenic ability, thereby being regarded as the "seed cells" responsible for tumor initiation and progression [22]. The seminal discovery in CSC research occurred in 1997 when Bonnet and Dick's research team first successfully identified and isolated leukemia stem cells (LSCs) from acute myeloid leukemia using CD34+/CD38- surface markers, marking a groundbreaking advancement in the field [23]. Subsequent progress in tumor biology has enabled researchers to identify CSC populations in various solid tumors (including breast cancer, glioma, hepatocellular carcinoma, gastric cancer, colorectal cancer, and ovarian cancer) through specific biomarkers such as CD44, CD133, and ALDH1 [24]. Notably, the expression profiles of CSC surface markers exhibit remarkable tumor-type specificity: ALDH+, CD44+, and CD133+ are predominantly expressed in head and neck cancer CSCs [25]; CD200+ and CD166+ are highly expressed in colorectal CSCs [26,27]; while pancreatic CSCs specifically overexpress markers including CD44/CD24 and epithelial-specific antigens [28]. Although CSCs typically constitute only 0.01-2% of the total tumor cell population, they play a pivotal role in tumor initiation, progression, metastasis, and recurrence [29]. Furthermore, CSCs frequently reside in a quiescent state within the cell cycle while overexpressing various drug resistance-associated molecules (such as ABC transporters and DNA damage repair enzymes), thereby serving as crucial mediators of therapeutic resistance. Mare et al. [30] demonstrated that paclitaxel-pretreated MCF7 breast cancer cells exhibited significantly enhanced stem cell properties, as evidenced by increased tumor sphere formation capacity, providing direct evidence for the central role of CSCs in chemoresistance. Recent studies have further revealed that beyond their remarkable self-renewal capacity, CSCs display unique "phenotypic plasticity" -the ability to undergo epithelial-mesenchymal transition (EMT) or dedifferentiation in response to microenvironmental cues (such as hypoxia and inflammatory cytokines), thereby generating adaptive tumor cell subpopulations. This plasticity not only substantially contributes to tumor heterogeneity but also plays a critical role in metastasis and treatment resistance [31]. At the molecular level, CSCs maintain their stemness and regulate vital biological processes (including survival, proliferation, and differentiation) through aberrant activation of key signaling pathways (such as Wnt/β-catenin, Notch, and PI3K/AKT/mTOR) accompanied by specific epigenetic modifications (including DNA methylation and histone modifications) [32].
4. Role of Exosomal miRNAs in Cancer
Exosomal miRNAs serve as pivotal mediators of intercellular communication in cancer, orchestrating long-distance regulatory effects through their stable circulatory transport system to play essential roles in tumor progression (Figure 2 and Table 1). These molecular regulators exert multifaceted effects by: (1) remodeling the tumor microenvironment through angiogenesis promotion and immunosuppression induction to facilitate immune evasion; (2) activating epithelial-mesenchymal transition (EMT) to enhance metastatic potential; and (3) transmitting drug-resistance genes and reprogramming cellular metabolism to promote tumor cell survival and proliferation.
4.1. Promotion of Tumor Growth and Metastasis
Exosomal miRNAs can modulate the expression of oncogenes or tumor suppressor genes in recipient cells, thereby influencing cellular growth processes and promoting tumor metastasis [33]. Zhang et al. uncovered that miR-200b derived from exosomes of colorectal cancer cells can specifically bind to the 3' untranslated region of the tumor suppressor p27, leading to a reduction in p27 expression at the post-transcriptional level and consequently facilitating the growth of tumor cells [34]. TGF-β activated kinase-1 (TAK1) can suppress the onset of hepatocellular carcinoma by regulating the MAPK-JNK signaling pathway. However, miR-584 within exosomes can target TAK1 and its associated signaling cascades, resulting in the downregulation of TAK1 expression and subsequently promoting the growth and metastasis of hepatocellular carcinoma [35]. In glioblastoma, exosomal miR-148a relieves the inhibitory effect of cell adhesion molecule 1 (CADM1) on the STAT3 pathway by targeting CADM1, significantly enhancing the proliferative and invasive metastatic capabilities of tumor cells[36]. In addition to regulating proliferation-related signaling pathways, exosomal miRNAs can also drive tumor progression by interfering with cell cycle checkpoints and apoptotic pathways. Research has demonstrated that miR-1246 disrupts the cell cycle regulatory network by specifically downregulating the expression of cyclin-G2 (CCNG2), thereby augmenting the proliferative capacity of breast cancer cells and inducing chemotherapy resistance [37]. Concurrently, miR-128 in exosomes derived from breast cancer cells can inhibit the initiation of apoptosis in tumor cells by regulating the expression balance of Bcl-2 family proteins (such as the pro-apoptotic factor Bax), thereby enhancing the tumor's metastatic potential [38]. These findings systematically unveil that exosomal miRNAs can promote tumor growth and metastasis through a coordinated, multi-target, and multi-pathway mechanism.
Epithelial-mesenchymal transition (EMT) represents a fundamental biological process driving cancer invasion and metastasis, characterized by distinct molecular alterations including: (1) downregulation of epithelial markers (E-cadherin, β-catenin), (2) upregulation of mesenchymal markers (vimentin, N-cadherin), and (3) loss of cellular polarity with enhanced motility [39]. Exosomes serve as critical EMT mediators by delivering specific miRNAs that modulate tumor invasive potential. Li et al. demonstrated that tumor-derived exosomal miR-21 post-transcriptionally regulates Snail (a master EMT transcription factor), concurrently upregulating vimentin while downregulating E-cadherin expression, thereby markedly enhancing cancer cell migration and invasion [40]. Wang's group revealed that CCRCC stem cell-secreted exosomal miR-19b-3p suppresses PTEN (a tumor suppressor) expression in recipient cells, effectively inducing EMT and promoting metastatic dissemination [41]. Under hypoxic conditions, BMSC-derived exosomal miR-193a-3p, miR-210-3p, and miR-5100 promote the invasion of lung cancer cells by inducing EMT through STAT3 signaling activation [42]. Sun et al. demonstrated that exosome-delivered miR-335-5p promotes colorectal cancer (CRC) cell invasion and metastasis by inducing epithelial-mesenchymal transition (EMT) through targeted inhibition of RAS p21 protein activator 1 (RASA1) [43].
4.2. Promotion of Tumor Angiogenesis
Angiogenesis supplies tumor cells with nutrients and oxygen while providing escape routes for metastasis, and is therefore recognized as a critical event in cancer progression [44]. Exosomal miRNAs play pivotal roles in angiogenesis by transmitting pro-angiogenic signals, regulating target gene expression, and activating key signaling pathways. For example, hepatocellular carcinoma (HCC)-derived exosomal miR-103 targets multiple endothelial junction proteins, including VE-cadherin, p120-catenin, and zonula occludens-1, thereby enhancing vascular permeability and promoting tumor metastasis [45]. Exosomal miR-23a secreted by nasopharyngeal carcinoma (NPC) cells not only promotes endothelial cell proliferation, migration, and tube formation but also enhances angiogenesis by targeting and suppressing TSGA10 [46]. Hypoxia, a major driving force of tumor angiogenesis, can modulate the activity of various cytokines, including the expression of exosomal miRNAs. Under hypoxic conditions, leukemia cell–derived exosomal miR-210 is highly expressed and promotes endothelial angiogenic activity by targeting Ephrin-A3 and activating the VEGF/VEGFR2 signaling pathway [47]. Further studies have shown that hypoxia upregulates the expression of tissue inhibitor of metalloproteinases-1 (TIMP-1), which in turn activates the PI3K/AKT/HIF-1 signaling pathway to promote miR-210 transcription. By suppressing its target proteins, miR-210 enhances the pro-angiogenic capacity of exosomes derived from TIMP-1–overexpressing cells [48]. In addition, exosomal miR-21 can induce STAT3 pathway activation, leading to upregulation of vascular endothelial growth factor (VEGF) expression in recipient cells, ultimately mediating angiogenesis and malignant transformation in human bronchial epithelial (HBE) cells [49]. Exosomal miR-23a from hypoxic lung cancer cells suppresses prolyl hydroxylase (PHD) and the tight junction protein zonula occludens-1 (ZO-1), thereby increasing vascular permeability and enhancing tumor transendothelial migration [50]. Exosomal miRNAs can also directly promote neovascularization by activating angiogenesis-related signaling pathways. In ovarian cancer, small extracellular vesicles (sEVs) secreted by the SKOV-3 cell line are enriched in miR-141-3p, which activates the JAK-STAT3 pathway in endothelial cells and promotes angiogenesis [51]. Mesenchymal stem cell (MSC)–derived exosomal miR-100 regulates the mTOR/HIF-1α/VEGF axis to stimulate angiogenesis in breast cancer [52]. In NPC, exosomal miR-9 modulates the phosphoinositide-dependent kinase (PDK)/Akt pathway to inhibit NPC cell migration and angiogenesis [53].
4.3. Promotion of Tumor Chemoresistance
Chemoresistance is a major cause of therapeutic failure and poor prognosis in cancer patients, primarily driven by tumor cell heterogeneity and remodeling of the tumor microenvironment. Exosomal miRNAs play critical roles in the initiation and progression of chemoresistance. Tumor cells can transfer drug resistance-associated miRNAs to drug-sensitive cells via exosomes, thereby directly regulating target gene expression and inducing resistance. For example, in colorectal cancer, exosomal miR-21 derived from resistant cells induces resistance to 5-fluorouracil (5-FU) by downregulating PTEN and hMSH2 expression [54], whereas in breast cancer, exosomal miR-221/222 from tamoxifen-resistant cells suppresses p27 and ERα expression, thereby enhancing tamoxifen resistance in recipient cells [55]. In addition, exosomal miRNAs can promote drug efflux by regulating the expression of drug transporters, further exacerbating resistance. Tao et al. [56] reported that, in colorectal cancer, exosomal miR-208b targets PDCD4 to promote the activation of regulatory T cells (Tregs), indirectly enhancing tumor resistance to oxaliplatin. Exosomal miRNAs can also inhibit chemotherapy-induced apoptosis by degrading or repressing the mRNAs of pro-apoptotic genes, thereby reducing drug sensitivity. For instance, miR-155 is upregulated in multiple cancers and suppresses PTEN and Fas ligand expression, thus blocking apoptotic signaling and decreasing chemosensitivity[57,58]. In head and neck cancer, cancer-associated fibroblast (CAF)–derived exosomal miR-196a targets CDKN1B and ING5, promoting cell proliferation and inhibiting apoptosis, which confers cisplatin resistance to recipient cells [59]. Moreover, exosomal miRNAs can enhance drug resistance by promoting EMT. Studies have shown that miR-21 and miR-27a are upregulated in pancreatic cancer[60] and breast cancer [61,62],where they suppress E-cadherin and induce the expression of N-cadherin and vimentin, thereby promoting the EMT process and consequently enhancing chemoresistance and metastatic potential.
4.4. Promotion of Tumor Immunosuppression
Tumor cells can suppress the maturation and differentiation of immune cells, remodel the immunosuppressive microenvironment, and induce immune tolerance, thereby establishing a synergistic regulatory network that facilitates malignant progression. As critical carriers of intercellular communication, exosomal miRNAs can reprogram immune regulators and effector cells, including tumor-associated macrophages (TAMs), dendritic cells (DCs), natural killer (NK) cells, and T lymphocytes [63]. TAMs are central components of the tumor microenvironment, with the M1 phenotype exerting antitumor activity, whereas the M2 phenotype promotes tumor growth and metastasis by creating an immunosuppressive niche [64]. Ying et al. [65] reported that exosomal miR-222-3p derived from epithelial ovarian cancer (EOC) cells targets and suppresses SOCS3, activates the STAT3 signaling pathway, and drives macrophage polarization toward a tumor-promoting M2 phenotype, thereby enhancing EOC proliferation and metastasis. Under hypoxic conditions, pancreatic cancer (PC) cell–derived exosomal miR-301a-3p suppresses PTEN and activates the PI3Kγ signaling pathway, similarly inducing M2 macrophage polarization [66]. DCs initiate T-cell–mediated immune responses through antigen processing and presentation. Their pattern recognition receptors (PRRs), such as Toll-like receptors (TLRs), are activated upon recognition of pathogen-associated molecular patterns (PAMPs) or damage-associated molecular patterns (DAMPs), triggering innate immune signaling cascades [67]. Exosomal miR-21 and miR-29a from non-small cell lung cancer (NSCLC) can activate NF-κB in a TLR-dependent manner, inducing a tumor-promoting inflammatory microenvironment that drives tumor growth and metastasis [68]. Ding et al. [69] demonstrated that pancreatic cancer cell–derived exosomal miR-212-3p downregulates the expression of regulatory factor X-associated protein (RFXAP), leading to reduced levels of the antigen-presenting molecule MHC class II and the induction of immune tolerance.Exosomal miRNAs can also facilitate immune evasion by impairing immune cell cytotoxicity. NK cells typically recognize and eliminate tumor cells via the NKG2D receptor–ligand interaction. However, high levels of exosomal miR-20a can downregulate NKG2D ligands (MICA and MICB), thereby weakening NK cell recognition and cytotoxicity, ultimately promoting immune escape and enhancing the metastatic capacity of breast cancer cells [70]. In nasopharyngeal carcinoma (NPC), exosomal miRNAs can concurrently inhibit T-cell proliferation and differentiation by targeting the MAPK1 and JAK/STAT signaling pathways, thereby facilitating tumor progression [71]. T helper 1 (Th1) cells, a differentiated subset of naïve CD4⁺ T cells, secrete IFN-γ, a key effector molecule in antitumor immunity. Exosomal miR-let-7d suppresses Th1 cell proliferation and IFN-γ production, inducing tumor immune tolerance [72]. Furthermore, exosomal miR-24-3p can target PTEN, activate the PI3K/AKT signaling pathway, and upregulate PD-L1 expression, thereby inhibiting T-cell function and promoting NPC malignant progression [73].
4.5. Promotion of Tumor Metabolism
Exosomal miRNAs can modulate the expression or activity of metabolic enzymes and reprogram tumor cell metabolic pathways, thereby influencing energy supply, biosynthetic processes, and ultimately regulating tumor growth and metastasis. In glycolysis, exosomal miRNAs can target key rate-limiting enzymes. For instance, Jiang et al. [74] reported that miR-143 suppresses glycolysis by targeting hexokinase 2 (HK2), whereas miR-155 indirectly upregulates HK2 expression by inhibiting miR-143, thereby enhancing glycolysis and promoting tumor cell proliferation. Glucose transporters (GLUTs) are essential for glucose uptake, and miR-144 has been shown to upregulate GLUT1 expression in lung cancer, increasing glucose uptake and lactate production, which in turn supports cancer cell growth [75]. Beyond glucose metabolism, exosomal miRNAs also participate in the regulation of fatty acid metabolism. Upregulation of miR-21 in NSCLC cells promotes cell growth and migration, while increasing intracellular lipid accumulation and elevating key lipid metabolic enzymes, including fatty acid synthase (FASN), acetyl-CoA carboxylase 1 (ACC1), and fatty acid-binding protein 5 (FABP5) [76]. Moreover, exosomal miRNAs can regulate the expression of metabolic signaling molecules such as PI3K, AKT, or mTOR, thereby affecting glucose metabolism, lipid biosynthesis, and protein translation. Notably, miR-21 has been reported to activate the PI3K/AKT pathway in colorectal and lung cancers, promoting glycolysis and tumor cell proliferation [77,78]. AMP-activated protein kinase (AMPK) acts as a central energy sensor, activated under low-energy conditions to promote catabolic pathways and maintain energy homeostasis. Evidence suggests that miR-451 may modulate tumor metabolic reprogramming by targeting the AMPK pathway, thereby facilitating tumor growth [79].
4.6. Regulation of Autophagy
Autophagy is a highly conserved cellular self-degradation process that maintains intracellular homeostasis by degrading damaged organelles and misfolded proteins via lysosomes. In cancer, autophagy plays a dual role: on one hand, it suppresses tumor initiation by clearing harmful components; on the other hand, in established tumors, autophagy provides metabolic substrates and energy to cancer cells, promoting their survival, drug resistance, and metastasis [80]. Exosomes can deliver miRNAs to target cells, regulating autophagy-related genes or signaling pathways, thereby influencing the formation, elongation, and degradation of autophagosomes. This process plays a critical role in disease treatment, intercellular communication, and pathophysiological processes. Exosomes secreted by ovarian cancer cells carry miR-328-3p, which targets Raf1 to inhibit the activation of the mTOR signaling pathway, thereby promoting autophagy and enhancing tumor cell invasiveness [81]. miR-224-5p in exosomes derived from human umbilical cord mesenchymal stem cells (hUCMSCs-exo) regulates autophagy by targeting and suppressing HOXA5 expression, affecting the proliferation and apoptosis of breast cancer (BC) cells [82]. In NSCLC, exosomal miR-425-3p activates autophagy in recipient cells by targeting AKT1, ultimately leading to chemoresistance [83]. Wang et al. [84] reported that exosomal miR-1910-3p promotes breast cancer cell proliferation, metastasis, and autophagy by targeting MTMR3 and activating the NF-κB signaling pathway. Osteoblast-derived exosomal miR-140-3p targets ACER2 and promotes prostate cancer progression by inhibiting autophagy via the AKT/mTOR pathway [85].
5. Roles of Exosomal miRNAs in Cancer Stem Cells
Exosomal miRNAs can precisely regulate the activity of oncogene promoters and the expression of tumor suppressor genes, thereby significantly influencing multiple functional mechanisms, biological behaviors, and phenotypic characteristics of cancer stem cells (CSCs), as shown in Figure 3. Their core regulatory roles are mainly manifested in: modulating the self-renewal capacity, differentiation direction, invasive and metastatic potential of CSCs, as well as the development of therapeutic resistance.
5.1. Maintenance of CSC Stemness Characteristics and Self-Renewal
The Wnt, Notch, and Hedgehog signaling pathways are key regulators of tumor stemness [86]. Exosomal miRNAs modulate these pathways to regulate the sphere-forming ability and tumorigenicity of CSCs, thereby maintaining and promoting their stem-like phenotype. Wu et al. found that miR-483-5p promotes proliferation, invasion, and self-renewal of gastric cancer stem cells by activating the Wnt/β-catenin signaling pathway [87]. Under the regulation of the transcription factor HIF-1α, miR-1275 activates both the Wnt/β-catenin and Notch pathways, enhancing stemness in lung adenocarcinoma cells and promoting tumorigenicity, recurrence, and metastasis [88]. Yang et al. [89] reported that chemotherapy activates the EZH2/STAT3 axis in breast cancer cells, leading to secretion of exosomes containing miR-378a-3p, which upon uptake by surviving cells activate Wnt/Notch pathways to enhance stemness and confer drug resistance. Exosomal miR-454 from breast cancer cells maintains CSC stemness in ovarian cancer by activating the PRRT2/Wnt axis [90]. M2 macrophage-derived exosomal miR-27a-3p promotes liver cancer CSC characteristics, proliferation, and tumorigenicity in vivo via activation of the TXNIP pathway [91]. Additionally, exosomal miRNAs target stemness-related gene expression to promote CSC traits. Chemotherapy induces breast cancer cells to secrete exosomes carrying multiple miRNAs such as miR-9-5p, miR-195-5p, and miR-203a-3p, which target ONECUT2, induce CSC phenotypes, and upregulate stemness-associated gene expression [92]. miR-328-3p is significantly upregulated in ovarian CSCs, maintaining stemness by targeting and suppressing DNA damage-binding protein 2. Concurrently, low reactive oxygen species levels reduce ERK signaling activity, favoring enhanced miR-328 expression and CSC maintenance [93].
5.2. Mediation of CSC Therapy Resistance
CSCs are considered key contributors to tumor drug resistance. Increasing evidence shows that CSC-derived exosomes deliver miRNAs to non-CSC cells, promoting chemoresistance through multiple mechanisms including apoptosis regulation, enhanced drug efflux, and induction of drug resistance in chemo-sensitive cells [94,95,96]. CSC-derived exosomes reprogram recipient cells’ cell cycle and apoptosis-related genes to facilitate resistance. Yin et al. [97] demonstrated that under hypoxia, miR-30b-3p in glioma CSC-derived exosomes targets RHOB, reducing apoptosis and promoting resistance to temozolomide. Zhuang et al. [98] reported that exosomal miR-146a-5p from cancer-associated fibroblasts regulates cell cycle and apoptosis pathways, maintaining bladder CSC stemness and enhancing chemoresistance, thus establishing a pro-resistant tumor microenvironment niche. Yang et al. [99] showed that exosomes from gemcitabine-resistant pancreatic CSCs transfer miR-210 to drug-sensitive cells, conferring resistance accompanied by upregulation of MDR1, YB-1, BCRP, and activation of the mTOR pathway, suggesting miR-210 mediates resistance by promoting drug efflux in non-CSC subpopulations. miR-221 suppresses the tumor suppressor QKI by targeting its mRNA 3′-UTR, leading to aberrant activation of downstream stemness pathways, which promotes tumorigenicity and chemoresistance of colorectal CSCs [100]. CSC-derived exosomes transmit miRNAs and other genetic materials that enable horizontal transfer of resistance to sensitive cells. Santos et al. experimentally confirmed that miR-155 secreted by breast CSC exosomes downregulates C/EBP-β and inhibits TGF-β, C/EBP-β, and FOXO3a expression, inducing EMT and chemoresistance in sensitive cells [101]. miR-485-5p specifically suppresses keratin 17 (KRT17) expression, thereby regulating integrin-mediated FAK/Src/ERK signaling and β-catenin nuclear translocation, ultimately affecting oral CSC stemness maintenance and chemoresistance phenotypes [102].
6. Translational Applications of Exosomal miRNAs in Cancer Diagnosis and Therapy
6.1. Potential Diagnostic and Prognostic Biomarkers
Exosomal miRNAs, owing to their remarkable stability, high tissue and disease specificity, and widespread presence in various body fluids, are regarded as promising diagnostic and prognostic biomarkers with significant clinical potential [103]. They have demonstrated important value in the detection and evaluation of malignancies, neurological disorders, and cardiovascular diseases. Wu et al. [104] examined the expression profiles of eight serum and serum-derived exosomal miRNAs (including miR-21-5p and miR-141-3p) in healthy individuals, patients with benign lung lesions, and patients with early-stage (NSCLC, stage I/II). Their findings revealed that serum exosomal miRNAs outperformed serum miRNAs as biomarkers for early NSCLC diagnosis, and the combination of both further enhanced diagnostic accuracy. Wang et al. [84] reported that the combination of CA15-3 in exosomes and serum miR-1910-3p served as an effective biomarker, improving the reliability of breast cancer diagnosis. In prostate cancer patients, plasma exosomal miR-141-5p was upregulated, while miR-125a-5p was downregulated, suggesting their potential as diagnostic indicators [105]. In oral squamous cell carcinoma (OSCC), elevated plasma exosomal miR-130a levels were identified as independent predictors of overall survival and recurrence-free survival [106]. Liu et al. [107] demonstrated that serum exosomal miR-106b-3p was significantly increased in metastatic CRC compared to non-metastatic cases, indicating its potential as a prognostic biomarker and therapeutic target. Additionally, urinary exosomal miR-155-5p, miR-15a-5p, and miR-21-5p were markedly elevated in bladder cancer patients compared to healthy controls, supporting their utility as non-invasive diagnostic indicators [108].
6.2. Potential Therapeutic Targets
6.2.1. Targeted Delivery of Exosomal miRNAs to Tumor Cells
Exosomes can be engineered to encapsulate specific tumor-suppressive miRNAs for precise delivery to target cells, offering novel therapeutic strategies. For instance, miR-122 loaded into MSC-derived exosomes effectively targeted hepatocellular carcinoma cells, exerting anti-tumor effects [109]. miR-375, a tumor suppressor inversely correlated with EMT, was delivered via tumor-derived exosomes in the form of mimics by Rezaei et al. [110], thereby suppressing migration and invasion in colon cancer cells. In gastric cancer, CAF-derived exosomes loaded with miR-139 or miR-34 mimics significantly inhibited tumor cell proliferation and metastasis [111,112]. Similarly, miR-205 mimics transfected into bone marrow mesenchymal stem cell-derived exosomes suppressed RHPN2 expression, impeding prostate cancer progression [113]. In a recent study, Cui et al. [114] employed electroporation to load miR-486-5p into bone marrow mesenchymal stem cell-derived exosomes and deliver them specifically to tumor sites. The results demonstrated that this strategy effectively inhibited glycolysis and stemness in colorectal cancer cells by targeting NEK2, thereby significantly suppressing tumor growth and progression. In addition to delivering tumor-suppressive miRNAs, exosomes can serve as carriers for miRNA inhibitors or antagonists to block oncogenic miRNA activity. This can be achieved using anti-miRNA oligonucleotides or inhibitors that interfere with transcription, processing, or stability. For example, Wang et al. [115] encapsulated a miR-21 inhibitor into engineered exosomes and delivered it to gastric cancer cells, achieving greater inhibitory effects and lower cytotoxicity compared to conventional transfection methods. Liang et al. [54] co-loaded the chemotherapeutic drug 5-FU and a miR-21 inhibitor (miR-21i) into exosomes for targeted delivery to Her2-positive colorectal cancer and 5-FU-resistant cell lines. This approach restored the function of tumor suppressor PTEN and DNA repair protein hMSH2, inducing cell cycle arrest and apoptosis. Furthermore, transfection of an miRNA-BART1-5p antagonist into exosomes significantly inhibited angiogenesis in tumor tissues and induced apoptosis [116]. Collectively, these studies highlight the substantial potential of exosomes in miRNA-based targeted cancer therapy, offering novel strategies and insights for precision oncology.
6.2.2. Targeted Delivery of Exosomal miRNAs to Cancer Stem Cells
The maintenance of CSC stemness is highly dependent on the regulation of multiple signaling pathways within the tumor microenvironment. As essential intercellular communication vehicles, exosomes can deliver specific miRNAs to CSCs, modulating various stemness-related signaling pathways and thereby altering their biological properties to suppress tumor growth and progression. In recent years, the advent of engineered exosomes has provided a feasible and efficient strategy for targeted miRNA delivery to CSCs. Epithelial cell adhesion molecule (EpCAM) is one of the classic CSC markers. Studies have shown that engineering exosomes to specifically recognize and target EpCAM-expressing liver CSCs, while loading them with β-catenin–specific siRNA, can effectively block activation of the Wnt/β-catenin signaling pathway and significantly inhibit CSC proliferation [117]. Similarly, tumor-suppressive miRNAs can be loaded into exosomes for CSC targeting. For example, Alessia et al. [118] demonstrated that exosomes derived from human liver stem cells (HLSCs) can deliver miR-145 and miR-200 to renal CSCs, inducing apoptosis and markedly suppressing proliferation, sphere-forming capacity, and invasiveness.SOX9, a critical regulator of CSC stemness maintenance, self-renewal, and tumor progression, has emerged as an important therapeutic target for CSC-directed interventions. Wu et al. [119] reported that bone marrow mesenchymal stem cell–derived exosomes carrying miR-145-5p specifically target SOX9 in non-small cell lung cancer, thereby significantly impairing CSC stemness maintenance and inhibiting tumor cell proliferation, migration, and invasion. In addition, Lang et al. [120] demonstrated that transducing miR-124a into MSCs enables the generation of miR-124a-enriched exosomes (Exo-miR-124a), which can be effectively delivered to glioblastoma stem cells (GSCs). In vitro, Exo-miR-124a markedly reduced GSC viability and clonogenic potential, while in an orthotopic GSC xenograft mouse model, systemic administration of Exo-miR-124a resulted in 50% of tumor-bearing mice achieving long-term tumor-free survival, indicating complete tumor eradication. Beyond tumor-suppressive miRNAs, engineered exosomes can also be utilized to deliver inhibitors of pro-stemness miRNAs (anti-miRs) to attenuate CSC tumorigenicity. For instance, Naseri et al. [121] employed electroporation to load anti-miR-142-3p into exosomes for targeted delivery to tumor sites, effectively suppressing endogenous miR-142-3p expression and function. This intervention disrupted signaling pathways associated with breast CSC tumorigenicity, significantly reducing CSC tumorigenic potential and impeding malignant progression of breast cancer.
7. Conclusions and Future Perspectives
Exosome-derived miRNAs, as key mediators of intercellular communication, play crucial roles in cancer and CSC biology by reshaping the tumor microenvironment, regulating CSC stemness maintenance, and modulating therapeutic resistance. They are deeply involved in tumor initiation and progression, metastasis, and treatment resistance. Moreover, the detectability and specificity of exosomal miRNAs in liquid biopsies provide a solid foundation for their use as biomarkers for early cancer diagnosis, therapeutic response assessment, and prognostic prediction. In recent years, strategies employing engineered exosomes as miRNA delivery vehicles have achieved promising results in preclinical studies, offering new therapeutic avenues for targeting CSCs and overcoming drug resistance. However, several challenges remain in this field. First, the mechanisms underlying exosomal miRNA biogenesis, selective packaging, and secretion are not yet fully elucidated, limiting their precise application in early disease diagnosis. Second, the techniques for exosome isolation and purification, miRNA loading efficiency, and targeted in vivo delivery still require optimization to ensure safety and controllability. Furthermore, the functions of exosomal miRNAs within CSCs and the tumor microenvironment are highly context-dependent and exhibit spatiotemporal specificity, adding complexity to clinical translation. Future research should focus on the following directions: Elucidating the dynamic regulatory networks of exosomal miRNAs in CSC stemness maintenance and tumor progression; Developing efficient, standardized techniques for exosome isolation and quantitative miRNA detection to improve reproducibility and accuracy in clinical testing; Advancing the application of engineered exosomes in miRNA-targeted delivery by optimizing loading efficiency and targeting specificity to achieve CSC-specific eradication; Integrating single-cell omics, multi-omics, and spatial transcriptomics to uncover the key mechanisms by which exosomal miRNAs regulate tumor heterogeneity and resistance evolution; Accelerating the clinical translation of exosomal miRNA-based diagnostic and therapeutic strategies, grounded in rigorous safety evaluations. In summary, research on exosome-derived miRNAs in cancer and CSCs is evolving from fundamental mechanistic studies toward clinical application. With deeper insights into their molecular mechanisms and the refinement of engineering-based delivery technologies, exosomal miRNAs are poised to become integral components of future precision oncology, offering innovative solutions to the challenges of tumor recurrence, metastasis, and therapeutic resistance.
Author Contributions
SM Wang: Literature review and original manuscript writing; SK Jin: Article review and editing; XY Wang: Conceptualization, supervision, and funding. All authors contributed to the article. All authors have read and agreed to the published version of the manuscript.
Funding
We are grateful for the financial support received from various sources, including the National Natural Science Foundation of China (82360044), the “12345 Future Talent Training Plan” of Zunyi Medical University (Future Science and Technology Elite Talent Project, ZYSE-2024-04), and the Guizhou Provincial Natural Science Foundation (Qian Ke He Ji Chu-ZK[2024]-269).
Conflicts of Interest
The authors declare no competing interests.
References
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin 2024, 74, 229–263. [Google Scholar] [CrossRef]
- Huang, T.; Song, X.; Xu, D.; Tiek, D.; Goenka, A.; Wu, B.; Sastry, N.; Hu, B.; Cheng, S.Y. Stem cell programs in cancer initiation, progression, and therapy resistance. Theranostics 2020, 10, 8721–8743. [Google Scholar] [CrossRef] [PubMed]
- Bhat, G.R.; Sethi, I.; Sadida, H.Q.; Rah, B.; Mir, R.; Algehainy, N.; Albalawi, I.A.; Masoodi, T.; Subbaraj, G.K.; Jamal, F.; et al. Cancer cell plasticity: from cellular, molecular, and genetic mechanisms to tumor heterogeneity and drug resistance. Cancer Metastasis Rev 2024, 43, 197–228. [Google Scholar] [CrossRef] [PubMed]
- Seo, N.; Akiyoshi, K.; Shiku, H. Exosome-mediated regulation of tumor immunology. Cancer Sci 2018, 109, 2998–3004. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhou, T.; Chen, J.; Li, R.; Chen, H.; Luo, S.; Chen, D.; Cai, C.; Li, W. The role of Exosomal miRNAs in cancer. J Transl Med 2022, 20, 6. [Google Scholar] [CrossRef]
- Chong, Z.X. Roles of miRNAs in regulating ovarian cancer stemness. Biochim Biophys Acta Rev Cancer 2024, 1879, 189191. [Google Scholar] [CrossRef]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367. [Google Scholar] [CrossRef]
- Rajput, A.; Varshney, A.; Bajaj, R.; Pokharkar, V. Exosomes as New Generation Vehicles for Drug Delivery: Biomedical Applications and Future Perspectives. Molecules 2022, 27. [Google Scholar] [CrossRef]
- Lee, Y.; El Andaloussi, S.; Wood, M.J. Exosomes and microvesicles: extracellular vesicles for genetic information transfer and gene therapy. Hum Mol Genet 2012, 21, R125–134. [Google Scholar] [CrossRef]
- Hurley, J.H. ESCRTs are everywhere. Embo j 2015, 34, 2398–2407. [Google Scholar] [CrossRef]
- Adell, M.A.; Vogel, G.F.; Pakdel, M.; Müller, M.; Lindner, H.; Hess, M.W.; Teis, D. Coordinated binding of Vps4 to ESCRT-III drives membrane neck constriction during MVB vesicle formation. J Cell Biol 2014, 205, 33–49. [Google Scholar] [CrossRef]
- Tang, S.; Buchkovich, N.J.; Henne, W.M.; Banjade, S.; Kim, Y.J.; Emr, S.D. ESCRT-III activation by parallel action of ESCRT-I/II and ESCRT-0/Bro1 during MVB biogenesis. Elife 2016, 5. [Google Scholar] [CrossRef]
- Chen, J.; Li, C.; Li, R.; Chen, H.; Chen, D.; Li, W. Exosomes in HIV infection. Curr Opin HIV AIDS 2021, 16, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Krylova, S.V.; Feng, D. The Machinery of Exosomes: Biogenesis, Release, and Uptake. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef] [PubMed]
- Kosaka, N.; Iguchi, H.; Yoshioka, Y.; Takeshita, F.; Matsuki, Y.; Ochiya, T. Secretory mechanisms and intercellular transfer of microRNAs in living cells. J Biol Chem 2010, 285, 17442–17452. [Google Scholar] [CrossRef] [PubMed]
- Kosaka, N.; Iguchi, H.; Hagiwara, K.; Yoshioka, Y.; Takeshita, F.; Ochiya, T. Neutral sphingomyelinase 2 (nSMase2)-dependent exosomal transfer of angiogenic microRNAs regulate cancer cell metastasis. J Biol Chem 2013, 288, 10849–10859. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, L.; Zou, W.; Chen, X.; Roizman, B.; Zhou, G.G. hnRNPA2B1 Associated with Recruitment of RNA into Exosomes Plays a Key Role in Herpes Simplex Virus 1 Release from Infected Cells. J Virol 2020, 94. [Google Scholar] [CrossRef]
- Lin, F.; Zeng, Z.; Song, Y.; Li, L.; Wu, Z.; Zhang, X.; Li, Z.; Ke, X.; Hu, X. YBX-1 mediated sorting of miR-133 into hypoxia/reoxygenation-induced EPC-derived exosomes to increase fibroblast angiogenesis and MEndoT. Stem Cell Res Ther 2019, 10, 263. [Google Scholar] [CrossRef]
- Shurtleff, M.J.; Temoche-Diaz, M.M.; Karfilis, K.V.; Ri, S.; Schekman, R. Y-box protein 1 is required to sort microRNAs into exosomes in cells and in a cell-free reaction. Elife 2016, 5. [Google Scholar] [CrossRef]
- McKenzie, A.J.; Hoshino, D.; Hong, N.H.; Cha, D.J.; Franklin, J.L.; Coffey, R.J.; Patton, J.G.; Weaver, A.M. KRAS-MEK Signaling Controls Ago2 Sorting into Exosomes. Cell Rep 2016, 15, 978–987. [Google Scholar] [CrossRef]
- Yokoi, A.; Villar-Prados, A.; Oliphint, P.A.; Zhang, J.; Song, X.; De Hoff, P.; Morey, R.; Liu, J.; Roszik, J.; Clise-Dwyer, K.; et al. Mechanisms of nuclear content loading to exosomes. Sci Adv 2019, 5, eaax8849. [Google Scholar] [CrossRef]
- Ayob, A.Z.; Ramasamy, T.S. Cancer stem cells as key drivers of tumour progression. J Biomed Sci 2018, 25, 20. [Google Scholar] [CrossRef]
- Lapidot, T.; Sirard, C.; Vormoor, J.; Murdoch, B.; Hoang, T.; Caceres-Cortes, J.; Minden, M.; Paterson, B.; Caligiuri, M.A.; Dick, J.E. A cell initiating human acute myeloid leukaemia after transplantation into SCID mice. Nature 1994, 367, 645–648. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Guo, Z.; Li, G.; Zhang, Y.; Liu, X.; Li, B.; Wang, J.; Li, X. Cancer stem cells and their niche in cancer progression and therapy. Cancer Cell Int 2023, 23, 305. [Google Scholar] [CrossRef] [PubMed]
- Gomez, K.E.; Wu, F.; Keysar, S.B.; Morton, J.J.; Miller, B.; Chimed, T.S.; Le, P.N.; Nieto, C.; Chowdhury, F.N.; Tyagi, A.; et al. Cancer Cell CD44 Mediates Macrophage/Monocyte-Driven Regulation of Head and Neck Cancer Stem Cells. Cancer Res 2020, 80, 4185–4198. [Google Scholar] [CrossRef] [PubMed]
- Eini, L.; Naseri, M.; Karimi-Busheri, F.; Bozorgmehr, M.; Ghods, R.; Madjd, Z. Preventive cancer stem cell-based vaccination modulates tumor development in syngeneic colon adenocarcinoma murine model. J Cancer Res Clin Oncol 2023, 149, 4101–4116. [Google Scholar] [CrossRef]
- Zhang, S.S.; Huang, Z.W.; Li, L.X.; Fu, J.J.; Xiao, B. Identification of CD200+ colorectal cancer stem cells and their gene expression profile. Oncol Rep 2016, 36, 2252–2260. [Google Scholar] [CrossRef]
- Li, C.; Heidt, D.G.; Dalerba, P.; Burant, C.F.; Zhang, L.; Adsay, V.; Wicha, M.; Clarke, M.F.; Simeone, D.M. Identification of pancreatic cancer stem cells. Cancer Res 2007, 67, 1030–1037. [Google Scholar] [CrossRef]
- Walcher, L.; Kistenmacher, A.K.; Suo, H.; Kitte, R.; Dluczek, S.; Strauß, A.; Blaudszun, A.R.; Yevsa, T.; Fricke, S.; Kossatz-Boehlert, U. Cancer Stem Cells-Origins and Biomarkers: Perspectives for Targeted Personalized Therapies. Front Immunol 2020, 11, 1280. [Google Scholar] [CrossRef]
- de la Mare, J.A.; Sterrenberg, J.N.; Sukhthankar, M.G.; Chiwakata, M.T.; Beukes, D.R.; Blatch, G.L.; Edkins, A.L. Assessment of potential anti-cancer stem cell activity of marine algal compounds using an in vitro mammosphere assay. Cancer Cell Int 2013, 13, 39. [Google Scholar] [CrossRef]
- Loh, J.J.; Ma, S. Hallmarks of cancer stemness. Cell Stem Cell 2024, 31, 617–639. [Google Scholar] [CrossRef]
- Zeng, Z.; Fu, M.; Hu, Y.; Wei, Y.; Wei, X.; Luo, M. Regulation and signaling pathways in cancer stem cells: implications for targeted therapy for cancer. Mol Cancer 2023, 22, 172. [Google Scholar] [CrossRef]
- Sun, Z.; Shi, K.; Yang, S.; Liu, J.; Zhou, Q.; Wang, G.; Song, J.; Li, Z.; Zhang, Z.; Yuan, W. Effect of exosomal miRNA on cancer biology and clinical applications. Mol Cancer 2018, 17, 147. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xing, T.; Chen, Y.; Xiao, J. Exosome-mediated miR-200b promotes colorectal cancer proliferation upon TGF-β1 exposure. Biomed Pharmacother 2018, 106, 1135–1143. [Google Scholar] [CrossRef] [PubMed]
- Kogure, T.; Lin, W.L.; Yan, I.K.; Braconi, C.; Patel, T. Intercellular nanovesicle-mediated microRNA transfer: a mechanism of environmental modulation of hepatocellular cancer cell growth. Hepatology 2011, 54, 1237–1248. [Google Scholar] [CrossRef] [PubMed]
- Cai, Q.; Zhu, A.; Gong, L. Exosomes of glioma cells deliver miR-148a to promote proliferation and metastasis of glioblastoma via targeting CADM1. Bull Cancer 2018, 105, 643–651. [Google Scholar] [CrossRef]
- Li, X.J.; Ren, Z.J.; Tang, J.H.; Yu, Q. Exosomal MicroRNA MiR-1246 Promotes Cell Proliferation, Invasion and Drug Resistance by Targeting CCNG2 in Breast Cancer. Cell Physiol Biochem 2017, 44, 1741–1748. [Google Scholar] [CrossRef]
- Wei, Y.; Li, M.; Cui, S.; Wang, D.; Zhang, C.Y.; Zen, K.; Li, L. Shikonin Inhibits the Proliferation of Human Breast Cancer Cells by Reducing Tumor-Derived Exosomes. Molecules 2016, 21. [Google Scholar] [CrossRef]
- Brabletz, T.; Kalluri, R.; Nieto, M.A.; Weinberg, R.A. EMT in cancer. Nat Rev Cancer 2018, 18, 128–134. [Google Scholar] [CrossRef]
- Li, L.; Li, C.; Wang, S.; Wang, Z.; Jiang, J.; Wang, W.; Li, X.; Chen, J.; Liu, K.; Li, C.; et al. Exosomes Derived from Hypoxic Oral Squamous Cell Carcinoma Cells Deliver miR-21 to Normoxic Cells to Elicit a Prometastatic Phenotype. Cancer Res 2016, 76, 1770–1780. [Google Scholar] [CrossRef]
- Wang, L.; Yang, G.; Zhao, D.; Wang, J.; Bai, Y.; Peng, Q.; Wang, H.; Fang, R.; Chen, G.; Wang, Z.; et al. CD103-positive CSC exosome promotes EMT of clear cell renal cell carcinoma: role of remote MiR-19b-3p. Mol Cancer 2019, 18, 86. [Google Scholar] [CrossRef]
- Zhang, X.; Sai, B.; Wang, F.; Wang, L.; Wang, Y.; Zheng, L.; Li, G.; Tang, J.; Xiang, J. Hypoxic BMSC-derived exosomal miRNAs promote metastasis of lung cancer cells via STAT3-induced EMT. Mol Cancer 2019, 18, 40. [Google Scholar] [CrossRef]
- Sun, X.; Lin, F.; Sun, W.; Zhu, W.; Fang, D.; Luo, L.; Li, S.; Zhang, W.; Jiang, L. Exosome-transmitted miRNA-335-5p promotes colorectal cancer invasion and metastasis by facilitating EMT via targeting RASA1. Mol Ther Nucleic Acids 2021, 24, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Lugano, R.; Ramachandran, M.; Dimberg, A. Tumor angiogenesis: causes, consequences, challenges and opportunities. Cell Mol Life Sci 2020, 77, 1745–1770. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.H.; Zhang, Z.J.; Shang, L.R.; Luo, Y.W.; Lin, Y.F.; Yuan, Y.; Zhuang, S.M. Hepatoma cell-secreted exosomal microRNA-103 increases vascular permeability and promotes metastasis by targeting junction proteins. Hepatology 2018, 68, 1459–1475. [Google Scholar] [CrossRef] [PubMed]
- Bao, L.; You, B.; Shi, S.; Shan, Y.; Zhang, Q.; Yue, H.; Zhang, J.; Zhang, W.; Shi, Y.; Liu, Y.; et al. Metastasis-associated miR-23a from nasopharyngeal carcinoma-derived exosomes mediates angiogenesis by repressing a novel target gene TSGA10. Oncogene 2018, 37, 2873–2889. [Google Scholar] [CrossRef]
- Tadokoro, H.; Umezu, T.; Ohyashiki, K.; Hirano, T.; Ohyashiki, J.H. Exosomes derived from hypoxic leukemia cells enhance tube formation in endothelial cells. J Biol Chem 2013, 288, 34343–34351. [Google Scholar] [CrossRef]
- Cui, H.; Seubert, B.; Stahl, E.; Dietz, H.; Reuning, U.; Moreno-Leon, L.; Ilie, M.; Hofman, P.; Nagase, H.; Mari, B.; et al. Tissue inhibitor of metalloproteinases-1 induces a pro-tumourigenic increase of miR-210 in lung adenocarcinoma cells and their exosomes. Oncogene 2015, 34, 3640–3650. [Google Scholar] [CrossRef]
- Liu, Y.; Luo, F.; Wang, B.; Li, H.; Xu, Y.; Liu, X.; Shi, L.; Lu, X.; Xu, W.; Lu, L.; et al. STAT3-regulated exosomal miR-21 promotes angiogenesis and is involved in neoplastic processes of transformed human bronchial epithelial cells. Cancer Lett 2016, 370, 125–135. [Google Scholar] [CrossRef]
- Hsu, Y.L.; Hung, J.Y.; Chang, W.A.; Lin, Y.S.; Pan, Y.C.; Tsai, P.H.; Wu, C.Y.; Kuo, P.L. Hypoxic lung cancer-secreted exosomal miR-23a increased angiogenesis and vascular permeability by targeting prolyl hydroxylase and tight junction protein ZO-1. Oncogene 2017, 36, 4929–4942. [Google Scholar] [CrossRef]
- Masoumi-Dehghi, S.; Babashah, S.; Sadeghizadeh, M. microRNA-141-3p-containing small extracellular vesicles derived from epithelial ovarian cancer cells promote endothelial cell angiogenesis through activating the JAK/STAT3 and NF-κB signaling pathways. J Cell Commun Signal 2020, 14, 233–244. [Google Scholar] [CrossRef]
- Pakravan, K.; Babashah, S.; Sadeghizadeh, M.; Mowla, S.J.; Mossahebi-Mohammadi, M.; Ataei, F.; Dana, N.; Javan, M. MicroRNA-100 shuttled by mesenchymal stem cell-derived exosomes suppresses in vitro angiogenesis through modulating the mTOR/HIF-1α/VEGF signaling axis in breast cancer cells. Cell Oncol (Dordr) 2017, 40, 457–470. [Google Scholar] [CrossRef]
- Lu, J.; Liu, Q.H.; Wang, F.; Tan, J.J.; Deng, Y.Q.; Peng, X.H.; Liu, X.; Zhang, B.; Xu, X.; Li, X.P. Exosomal miR-9 inhibits angiogenesis by targeting MDK and regulating PDK/AKT pathway in nasopharyngeal carcinoma. J Exp Clin Cancer Res 2018, 37, 147. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.; Zhu, Y.; Ali, D.J.; Tian, T.; Xu, H.; Si, K.; Sun, B.; Chen, B.; Xiao, Z. Engineered exosomes for targeted co-delivery of miR-21 inhibitor and chemotherapeutics to reverse drug resistance in colon cancer. J Nanobiotechnology 2020, 18, 10. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Lai, X.; Yu, S.; Chen, S.; Ma, Y.; Zhang, Y.; Li, H.; Zhu, X.; Yao, L.; Zhang, J. Exosomal miR-221/222 enhances tamoxifen resistance in recipient ER-positive breast cancer cells. Breast Cancer Res Treat 2014, 147, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Ning, T.; Li, J.; He, Y.; Zhang, H.; Wang, X.; Deng, T.; Liu, R.; Li, H.; Bai, M.; Fan, Q.; et al. Exosomal miR-208b related with oxaliplatin resistance promotes Treg expansion in colorectal cancer. Mol Ther 2021, 29, 2723–2736. [Google Scholar] [CrossRef]
- Bayraktar, R.; Van Roosbroeck, K. miR-155 in cancer drug resistance and as target for miRNA-based therapeutics. Cancer Metastasis Rev 2018, 37, 33–44. [Google Scholar] [CrossRef]
- Sayyed, A.A.; Gondaliya, P.; Mali, M.; Pawar, A.; Bhat, P.; Khairnar, A.; Arya, N.; Kalia, K. MiR-155 Inhibitor-Laden Exosomes Reverse Resistance to Cisplatin in a 3D Tumor Spheroid and Xenograft Model of Oral Cancer. Mol Pharm 2021, 18, 3010–3025. [Google Scholar] [CrossRef]
- Qin, X.; Guo, H.; Wang, X.; Zhu, X.; Yan, M.; Wang, X.; Xu, Q.; Shi, J.; Lu, E.; Chen, W.; et al. Exosomal miR-196a derived from cancer-associated fibroblasts confers cisplatin resistance in head and neck cancer through targeting CDKN1B and ING5. Genome Biol 2019, 20, 12. [Google Scholar] [CrossRef]
- Chen, C.; Demirkhanyan, L.; Gondi, C.S. The Multifaceted Role of miR-21 in Pancreatic Cancers. Cells 2024, 13. [Google Scholar] [CrossRef]
- Arisan, E.D.; Rencuzogullari, O.; Cieza-Borrella, C.; Miralles Arenas, F.; Dwek, M.; Lange, S.; Uysal-Onganer, P. MiR-21 Is Required for the Epithelial-Mesenchymal Transition in MDA-MB-231 Breast Cancer Cells. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Gong, C.; Yao, Y.; Wang, Y.; Liu, B.; Wu, W.; Chen, J.; Su, F.; Yao, H.; Song, E. Up-regulation of miR-21 mediates resistance to trastuzumab therapy for breast cancer. J Biol Chem 2011, 286, 19127–19137. [Google Scholar] [CrossRef]
- Nail, H.M.; Chiu, C.C.; Leung, C.H.; Ahmed, M.M.M.; Wang, H.D. Exosomal miRNA-mediated intercellular communications and immunomodulatory effects in tumor microenvironments. J Biomed Sci 2023, 30, 69. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Li, F.; Jin, S.; Ho, P.C.; Liu, P.S.; Xie, X. Functional polarization of tumor-associated macrophages dictated by metabolic reprogramming. J Exp Clin Cancer Res 2023, 42, 245. [Google Scholar] [CrossRef] [PubMed]
- Ying, X.; Wu, Q.; Wu, X.; Zhu, Q.; Wang, X.; Jiang, L.; Chen, X.; Wang, X. Epithelial ovarian cancer-secreted exosomal miR-222-3p induces polarization of tumor-associated macrophages. Oncotarget 2016, 7, 43076–43087. [Google Scholar] [CrossRef]
- Wang, X.; Luo, G.; Zhang, K.; Cao, J.; Huang, C.; Jiang, T.; Liu, B.; Su, L.; Qiu, Z. Hypoxic Tumor-Derived Exosomal miR-301a Mediates M2 Macrophage Polarization via PTEN/PI3Kγ to Promote Pancreatic Cancer Metastasis. Cancer Res 2018, 78, 4586–4598. [Google Scholar] [CrossRef]
- Thwe, P.M.; Amiel, E. The role of nitric oxide in metabolic regulation of Dendritic cell immune function. Cancer Lett 2018, 412, 236–242. [Google Scholar] [CrossRef]
- Fabbri, M.; Paone, A.; Calore, F.; Galli, R.; Gaudio, E.; Santhanam, R.; Lovat, F.; Fadda, P.; Mao, C.; Nuovo, G.J.; et al. MicroRNAs bind to Toll-like receptors to induce prometastatic inflammatory response. Proc Natl Acad Sci U S A 2012, 109, E2110–2116. [Google Scholar] [CrossRef]
- Ding, G.; Zhou, L.; Shen, T.; Cao, L. IFN-γ induces the upregulation of RFXAP via inhibition of miR-212-3p in pancreatic cancer cells: A novel mechanism for IFN-γ response. Oncol Lett 2018, 15, 3760–3765. [Google Scholar] [CrossRef]
- Wang, B.; Wang, Q.; Wang, Z.; Jiang, J.; Yu, S.C.; Ping, Y.F.; Yang, J.; Xu, S.L.; Ye, X.Z.; Xu, C.; et al. Metastatic consequences of immune escape from NK cell cytotoxicity by human breast cancer stem cells. Cancer Res 2014, 74, 5746–5757. [Google Scholar] [CrossRef]
- Ye, S.B.; Li, Z.L.; Luo, D.H.; Huang, B.J.; Chen, Y.S.; Zhang, X.S.; Cui, J.; Zeng, Y.X.; Li, J. Tumor-derived exosomes promote tumor progression and T-cell dysfunction through the regulation of enriched exosomal microRNAs in human nasopharyngeal carcinoma. Oncotarget 2014, 5, 5439–5452. [Google Scholar] [CrossRef]
- Okoye, I.S.; Coomes, S.M.; Pelly, V.S.; Czieso, S.; Papayannopoulos, V.; Tolmachova, T.; Seabra, M.C.; Wilson, M.S. MicroRNA-Containing T-Regulatory-Cell-Derived Exosomes Suppress Pathogenic T Helper 1 Cells. Immunity 2014, 41, 503. [Google Scholar] [CrossRef] [PubMed]
- Ye, S.B.; Zhang, H.; Cai, T.T.; Liu, Y.N.; Ni, J.J.; He, J.; Peng, J.Y.; Chen, Q.Y.; Mo, H.Y.; Jun, C.; et al. Exosomal miR-24-3p impedes T-cell function by targeting FGF11 and serves as a potential prognostic biomarker for nasopharyngeal carcinoma. J Pathol 2016, 240, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Zhang, L.F.; Zhang, H.W.; Hu, S.; Lu, M.H.; Liang, S.; Li, B.; Li, Y.; Li, D.; Wang, E.D.; et al. A novel miR-155/miR-143 cascade controls glycolysis by regulating hexokinase 2 in breast cancer cells. Embo j 2012, 31, 1985–1998. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Gao, J.; Huang, Q.; Jin, Y.; Wei, Z. Downregulating microRNA-144 mediates a metabolic shift in lung cancer cells by regulating GLUT1 expression. Oncol Lett 2016, 11, 3772–3776. [Google Scholar] [CrossRef]
- Ni, K.; Wang, D.; Xu, H.; Mei, F.; Wu, C.; Liu, Z.; Zhou, B. miR-21 promotes non-small cell lung cancer cells growth by regulating fatty acid metabolism. Cancer Cell Int 2019, 19, 219. [Google Scholar] [CrossRef]
- Lai, C.Y.; Yeh, K.Y.; Liu, B.F.; Chang, T.M.; Chang, C.H.; Liao, Y.F.; Liu, Y.W.; Her, G.M. MicroRNA-21 Plays Multiple Oncometabolic Roles in Colitis-Associated Carcinoma and Colorectal Cancer via the PI3K/AKT, STAT3, and PDCD4/TNF-α Signaling Pathways in Zebrafish. Cancers (Basel) 2021, 13. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, W.; Zhao, Q.; Zhang, R.; Wang, J.; Pan, P.; Shang, H.; Liu, C.; Wang, C. Down-Regulating the Expression of miRNA-21 Inhibits the Glucose Metabolism of A549/DDP Cells and Promotes Cell Death Through the PI3K/AKT/mTOR/HIF-1α Pathway. Front Oncol 2021, 11, 653596. [Google Scholar] [CrossRef]
- Godlewski, J.; Nowicki, M.O.; Bronisz, A.; Nuovo, G.; Palatini, J.; De Lay, M.; Van Brocklyn, J.; Ostrowski, M.C.; Chiocca, E.A.; Lawler, S.E. MicroRNA-451 regulates LKB1/AMPK signaling and allows adaptation to metabolic stress in glioma cells. Mol Cell 2010, 37, 620–632. [Google Scholar] [CrossRef]
- Wei, J.; Wang, X.; Yu, D.; Tu, Y.; Yu, Y. MicroRNA-mediated autophagy and drug resistance in cancer: mechanisms and therapeutic strategies. Discov Oncol 2024, 15, 662. [Google Scholar] [CrossRef]
- Sun, H.; Huo, X.; Bi, X.; Cao, D.; Yang, J.; Shen, K.; Peng, P. Exosome transmit the ability of migration and invasion in heterogeneous ovarian cancer cells by regulating autophagy via targeting hsa-miR-328. Gynecol Oncol 2025, 194, 60–70. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, P.; Zhao, L.; Chen, X.; Lin, Z.; Zhang, L.; Li, Z. miR-224-5p Carried by Human Umbilical Cord Mesenchymal Stem Cells-Derived Exosomes Regulates Autophagy in Breast Cancer Cells via HOXA5. Front Cell Dev Biol 2021, 9, 679185. [Google Scholar] [CrossRef]
- Ma, Y.; Yuwen, D.; Chen, J.; Zheng, B.; Gao, J.; Fan, M.; Xue, W.; Wang, Y.; Li, W.; Shu, Y.; et al. Exosomal Transfer Of Cisplatin-Induced miR-425-3p Confers Cisplatin Resistance In NSCLC Through Activating Autophagy. Int J Nanomedicine 2019, 14, 8121–8132. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Mao, J.H.; Wang, B.Y.; Wang, L.X.; Wen, H.Y.; Xu, L.J.; Fu, J.X.; Yang, H. Exosomal miR-1910-3p promotes proliferation, metastasis, and autophagy of breast cancer cells by targeting MTMR3 and activating the NF-κB signaling pathway. Cancer Lett 2020, 489, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, S.; Guo, K.; Ma, S.; Wang, X.; Liu, Q.; Yan, R.; Huang, Y.; Li, T.; He, S.; et al. Osteoblast-derived exosomal miR-140-3p targets ACER2 and increases the progression of prostate cancer via the AKT/mTOR pathway-mediated inhibition of autophagy. Faseb j 2024, 38, e70206. [Google Scholar] [CrossRef] [PubMed]
- Clara, J.A.; Monge, C.; Yang, Y.; Takebe, N. Targeting signalling pathways and the immune microenvironment of cancer stem cells - a clinical update. Nat Rev Clin Oncol 2020, 17, 204–232. [Google Scholar] [CrossRef]
- Wu, K.; Ma, L.; Zhu, J. miR-483-5p promotes growth, invasion and self-renewal of gastric cancer stem cells by Wnt/β-catenin signaling. Mol Med Rep 2016, 14, 3421–3428. [Google Scholar] [CrossRef]
- Jiang, N.; Zou, C.; Zhu, Y.; Luo, Y.; Chen, L.; Lei, Y.; Tang, K.; Sun, Y.; Zhang, W.; Li, S.; et al. HIF-1ɑ-regulated miR-1275 maintains stem cell-like phenotypes and promotes the progression of LUAD by simultaneously activating Wnt/β-catenin and Notch signaling. Theranostics 2020, 10, 2553–2570. [Google Scholar] [CrossRef]
- Yang, Q.; Zhao, S.; Shi, Z.; Cao, L.; Liu, J.; Pan, T.; Zhou, D.; Zhang, J. Chemotherapy-elicited exosomal miR-378a-3p and miR-378d promote breast cancer stemness and chemoresistance via the activation of EZH2/STAT3 signaling. J Exp Clin Cancer Res 2021, 40, 120. [Google Scholar] [CrossRef]
- Wang, L.; He, M.; Fu, L.; Jin, Y. Exosomal release of microRNA-454 by breast cancer cells sustains biological properties of cancer stem cells via the PRRT2/Wnt axis in ovarian cancer. Life Sci 2020, 257, 118024. [Google Scholar] [CrossRef]
- Li, W.; Xin, X.; Li, X.; Geng, J.; Sun, Y. Exosomes secreted by M2 macrophages promote cancer stemness of hepatocellular carcinoma via the miR-27a-3p/TXNIP pathways. Int Immunopharmacol 2021, 101, 107585. [Google Scholar] [CrossRef]
- Shen, M.; Dong, C.; Ruan, X.; Yan, W.; Cao, M.; Pizzo, D.; Wu, X.; Yang, L.; Liu, L.; Ren, X.; et al. Chemotherapy-Induced Extracellular Vesicle miRNAs Promote Breast Cancer Stemness by Targeting ONECUT2. Cancer Res 2019, 79, 3608–3621. [Google Scholar] [CrossRef]
- Srivastava, A.K.; Banerjee, A.; Cui, T.; Han, C.; Cai, S.; Liu, L.; Wu, D.; Cui, R.; Li, Z.; Zhang, X.; et al. Inhibition of miR-328-3p Impairs Cancer Stem Cell Function and Prevents Metastasis in Ovarian Cancer. Cancer Res 2019, 79, 2314–2326. [Google Scholar] [CrossRef]
- Mashouri, L.; Yousefi, H.; Aref, A.R.; Ahadi, A.M.; Molaei, F.; Alahari, S.K. Exosomes: composition, biogenesis, and mechanisms in cancer metastasis and drug resistance. Mol Cancer 2019, 18, 75. [Google Scholar] [CrossRef] [PubMed]
- Su, C.; Zhang, J.; Yarden, Y.; Fu, L. The key roles of cancer stem cell-derived extracellular vesicles. Signal Transduct Target Ther 2021, 6, 109. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Teng, Y. Harnessing cancer stem cell-derived exosomes to improve cancer therapy. J Exp Clin Cancer Res 2023, 42, 131. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Ge, X.; Shi, Z.; Yu, C.; Lu, C.; Wei, Y.; Zeng, A.; Wang, X.; Yan, W.; Zhang, J.; et al. Extracellular vesicles derived from hypoxic glioma stem-like cells confer temozolomide resistance on glioblastoma by delivering miR-30b-3p. Theranostics 2021, 11, 1763–1779. [Google Scholar] [CrossRef]
- Zhuang, J.; Shen, L.; Li, M.; Sun, J.; Hao, J.; Li, J.; Zhu, Z.; Ge, S.; Zhang, D.; Guo, H.; et al. Cancer-Associated Fibroblast-Derived miR-146a-5p Generates a Niche That Promotes Bladder Cancer Stemness and Chemoresistance. Cancer Res 2023, 83, 1611–1627. [Google Scholar] [CrossRef]
- Yang, Z.; Zhao, N.; Cui, J.; Wu, H.; Xiong, J.; Peng, T. Exosomes derived from cancer stem cells of gemcitabine-resistant pancreatic cancer cells enhance drug resistance by delivering miR-210. Cell Oncol (Dordr) 2020, 43, 123–136. [Google Scholar] [CrossRef]
- Mukohyama, J.; Isobe, T.; Hu, Q.; Hayashi, T.; Watanabe, T.; Maeda, M.; Yanagi, H.; Qian, X.; Yamashita, K.; Minami, H.; et al. miR-221 Targets QKI to Enhance the Tumorigenic Capacity of Human Colorectal Cancer Stem Cells. Cancer Res 2019, 79, 5151–5158. [Google Scholar] [CrossRef]
- Santos, J.C.; Lima, N.D.S.; Sarian, L.O.; Matheu, A.; Ribeiro, M.L.; Derchain, S.F.M. Exosome-mediated breast cancer chemoresistance via miR-155 transfer. Sci Rep 2018, 8, 829. [Google Scholar] [CrossRef]
- Jang, T.H.; Huang, W.C.; Tung, S.L.; Lin, S.C.; Chen, P.M.; Cho, C.Y.; Yang, Y.Y.; Yen, T.C.; Lo, G.H.; Chuang, S.E.; et al. MicroRNA-485-5p targets keratin 17 to regulate oral cancer stemness and chemoresistance via the integrin/FAK/Src/ERK/β-catenin pathway. J Biomed Sci 2022, 29, 42. [Google Scholar] [CrossRef] [PubMed]
- Preethi, K.A.; Selvakumar, S.C.; Ross, K.; Jayaraman, S.; Tusubira, D.; Sekar, D. Liquid biopsy: Exosomal microRNAs as novel diagnostic and prognostic biomarkers in cancer. Mol Cancer 2022, 21, 54. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Yu, L.; Lin, X.; Zheng, Q.; Zhang, S.; Chen, D.; Pan, X.; Huang, Y. Combination of Serum miRNAs with Serum Exosomal miRNAs in Early Diagnosis for Non-Small-Cell Lung Cancer. Cancer Manag Res 2020, 12, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Dong, Y.; Wang, K.J.; Deng, Z.; Zhang, W.; Shen, H.F. Plasma exosomal miR-125a-5p and miR-141-5p as non-invasive biomarkers for prostate cancer. Neoplasma 2020, 67, 1314–1318. [Google Scholar] [CrossRef]
- He, T.; Guo, X.; Li, X.; Liao, C.; Wang, X.; He, K. Plasma-Derived Exosomal microRNA-130a Serves as a Noninvasive Biomarker for Diagnosis and Prognosis of Oral Squamous Cell Carcinoma. J Oncol 2021, 2021, 5547911. [Google Scholar] [CrossRef]
- Liu, H.; Liu, Y.; Sun, P.; Leng, K.; Xu, Y.; Mei, L.; Han, P.; Zhang, B.; Yao, K.; Li, C.; et al. Colorectal cancer-derived exosomal miR-106b-3p promotes metastasis by down-regulating DLC-1 expression. Clin Sci (Lond) 2020, 134, 419–434. [Google Scholar] [CrossRef]
- Piao, X.M.; Cha, E.J.; Yun, S.J.; Kim, W.J. Role of Exosomal miRNA in Bladder Cancer: A Promising Liquid Biopsy Biomarker. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Lou, G.; Song, X.; Yang, F.; Wu, S.; Wang, J.; Chen, Z.; Liu, Y. Exosomes derived from miR-122-modified adipose tissue-derived MSCs increase chemosensitivity of hepatocellular carcinoma. J Hematol Oncol 2015, 8, 122. [Google Scholar] [CrossRef]
- Rezaei, R.; Baghaei, K.; Amani, D.; Piccin, A.; Hashemi, S.M.; Asadzadeh Aghdaei, H.; Zali, M.R. Exosome-mediated delivery of functionally active miRNA-375-3p mimic regulate epithelial mesenchymal transition (EMT) of colon cancer cells. Life Sci 2021, 269, 119035. [Google Scholar] [CrossRef]
- Xu, G.; Zhang, B.; Ye, J.; Cao, S.; Shi, J.; Zhao, Y.; Wang, Y.; Sang, J.; Yao, Y.; Guan, W.; et al. Exosomal miRNA-139 in cancer-associated fibroblasts inhibits gastric cancer progression by repressing MMP11 expression. Int J Biol Sci 2019, 15, 2320–2329. [Google Scholar] [CrossRef]
- Shi, L.; Wang, Z.; Geng, X.; Zhang, Y.; Xue, Z. Exosomal miRNA-34 from cancer-associated fibroblasts inhibits growth and invasion of gastric cancer cells in vitro and in vivo. Aging (Albany NY) 2020, 12, 8549–8564. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Mo, C.; Guo, S.; Zhuang, J.; Huang, B.; Mao, X. Human bone marrow mesenchymal stem cells-derived microRNA-205-containing exosomes impede the progression of prostate cancer through suppression of RHPN2. J Exp Clin Cancer Res 2019, 38, 495. [Google Scholar] [CrossRef] [PubMed]
- Cui, F.; Chen, Y.; Wu, X.; Zhao, W. Mesenchymal stem cell-derived exosomes carrying miR-486-5p inhibit glycolysis and cell stemness in colorectal cancer by targeting NEK2. BMC Cancer 2024, 24, 1356. [Google Scholar] [CrossRef]
- Wang, J.J.; Wang, Z.Y.; Chen, R.; Xiong, J.; Yao, Y.L.; Wu, J.H.; Li, G.X. Macrophage-secreted Exosomes Delivering miRNA-21 Inhibitor can Regulate BGC-823 Cell Proliferation. Asian Pac J Cancer Prev 2015, 16, 4203–4209. [Google Scholar] [CrossRef]
- Wang, J.; Liu, Y.; Zhang, Y.; Li, X.; Fang, M.; Qian, D. Targeting exosomes enveloped EBV-miR-BART1-5p-antagomiRs for NPC therapy through both anti-vasculogenic mimicry and anti-angiogenesis. Cancer Med 2023, 12, 12608–12621. [Google Scholar] [CrossRef]
- Ishiguro, K.; Yan, I.K.; Lewis-Tuffin, L.; Patel, T. Targeting Liver Cancer Stem Cells Using Engineered Biological Nanoparticles for the Treatment of Hepatocellular Cancer. Hepatol Commun 2020, 4, 298–313. [Google Scholar] [CrossRef]
- Brossa, A.; Fonsato, V.; Grange, C.; Tritta, S.; Tapparo, M.; Calvetti, R.; Cedrino, M.; Fallo, S.; Gontero, P.; Camussi, G.; et al. Extracellular vesicles from human liver stem cells inhibit renal cancer stem cell-derived tumor growth in vitro and in vivo. Int J Cancer 2020, 147, 1694–1706. [Google Scholar] [CrossRef]
- Yan, W.; Yang, H.; Duan, D.; Wu, Y.; Liu, Y.; Mao, J.; Zhao, Y.; Ye, J. Bone marrow mesenchymal stem cells-derived exosomal miR-145-5p reduced non-small cell lung cancer cell progression by targeting SOX9. BMC Cancer 2024, 24, 883. [Google Scholar] [CrossRef]
- Lang, F.M.; Hossain, A.; Gumin, J.; Momin, E.N.; Shimizu, Y.; Ledbetter, D.; Shahar, T.; Yamashita, S.; Parker Kerrigan, B.; Fueyo, J.; et al. Mesenchymal stem cells as natural biofactories for exosomes carrying miR-124a in the treatment of gliomas. Neuro Oncol 2018, 20, 380–390. [Google Scholar] [CrossRef]
- Naseri, Z.; Oskuee, R.K.; Forouzandeh-Moghadam, M.; Jaafari, M.R. Delivery of LNA-antimiR-142-3p by Mesenchymal Stem Cells-Derived Exosomes to Breast Cancer Stem Cells Reduces Tumorigenicity. Stem Cell Rev Rep 2020, 16, 541–556. [Google Scholar] [CrossRef]
Figure 1.
Biogenesis, secretion, and cellular uptake of exosomes. Exosome formation initiates with plasma membrane invagination to generate early endosomes. These compartments mature into late endosomes through additional membrane remodeling, during which they selectively incorporate diverse molecular cargo (including mRNAs, proteins, and lipids) through sophisticated sorting mechanisms. The resulting multivesicular bodies (MVBs) face two distinct fates: (1) lysosomal fusion leading to content degradation, or (2) plasma membrane fusion resulting in exosome secretion.
Figure 1.
Biogenesis, secretion, and cellular uptake of exosomes. Exosome formation initiates with plasma membrane invagination to generate early endosomes. These compartments mature into late endosomes through additional membrane remodeling, during which they selectively incorporate diverse molecular cargo (including mRNAs, proteins, and lipids) through sophisticated sorting mechanisms. The resulting multivesicular bodies (MVBs) face two distinct fates: (1) lysosomal fusion leading to content degradation, or (2) plasma membrane fusion resulting in exosome secretion.
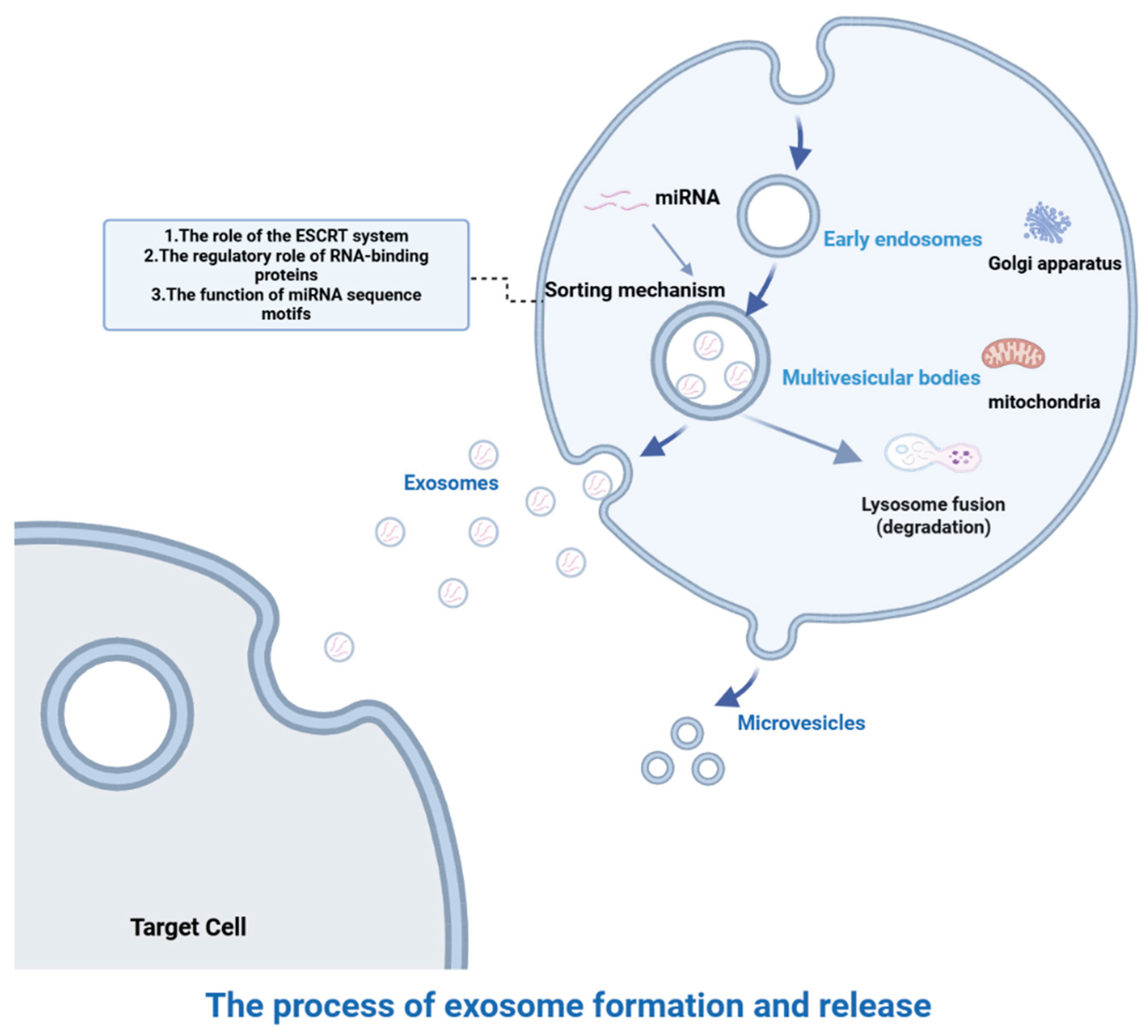
Figure 2.
Roles of exosomal miRNAs in tumor progression. Exosomal miRNAs can accelerate tumor metastasis, induce angiogenesis, enhance drug resistance and tumor metabolic reprogramming, and contribute to the formation of a tumor-promoting and immunosuppressive microenvironment.
Figure 2.
Roles of exosomal miRNAs in tumor progression. Exosomal miRNAs can accelerate tumor metastasis, induce angiogenesis, enhance drug resistance and tumor metabolic reprogramming, and contribute to the formation of a tumor-promoting and immunosuppressive microenvironment.
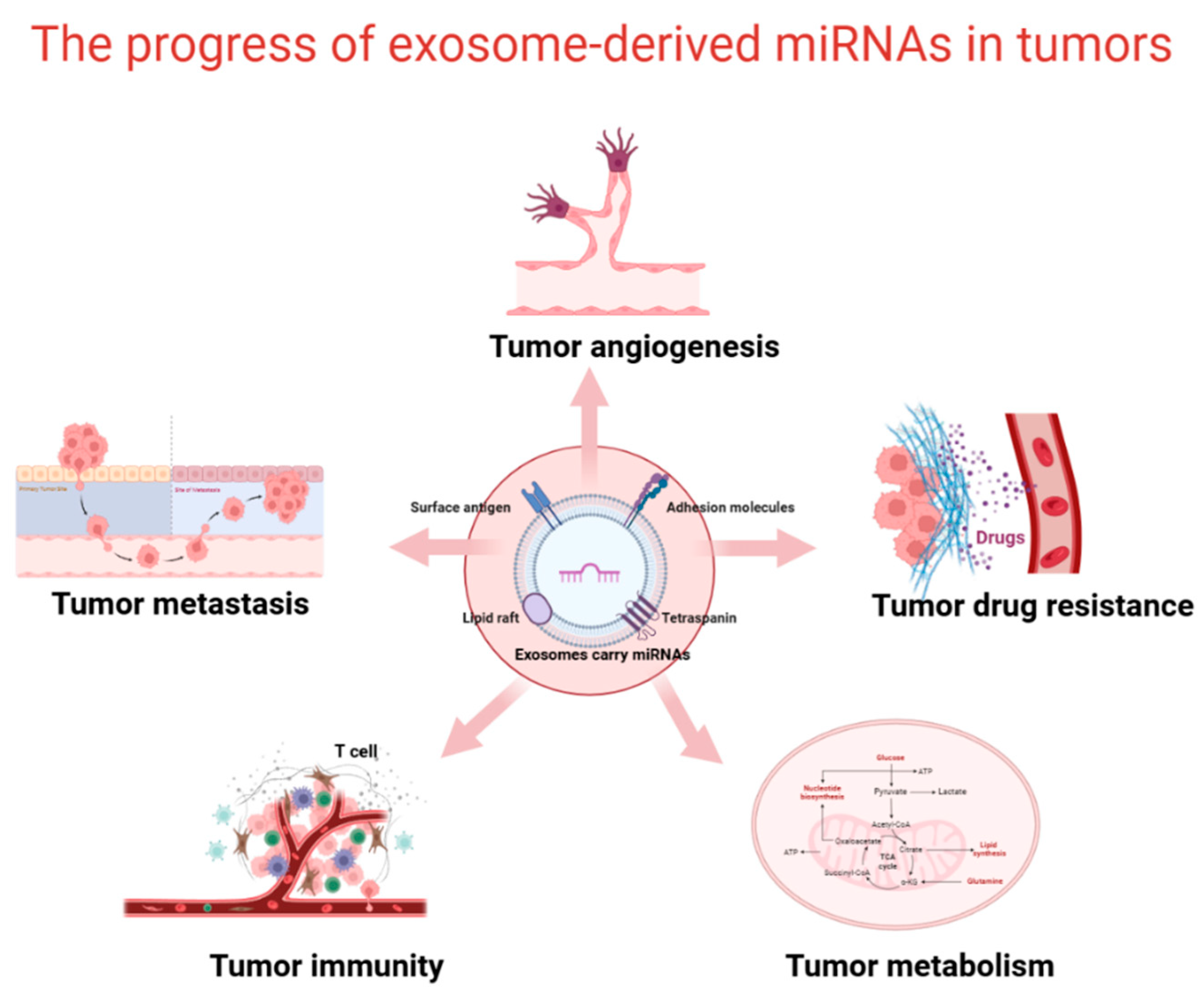
Figure 3.
Exosomal miRNAs can enhance the stemness of cancer stem cells (CSCs), which is primarily manifested by increased expression of stemness markers, enhanced tumorsphere-forming ability, and strengthened self-renewal capacity.
Figure 3.
Exosomal miRNAs can enhance the stemness of cancer stem cells (CSCs), which is primarily manifested by increased expression of stemness markers, enhanced tumorsphere-forming ability, and strengthened self-renewal capacity.
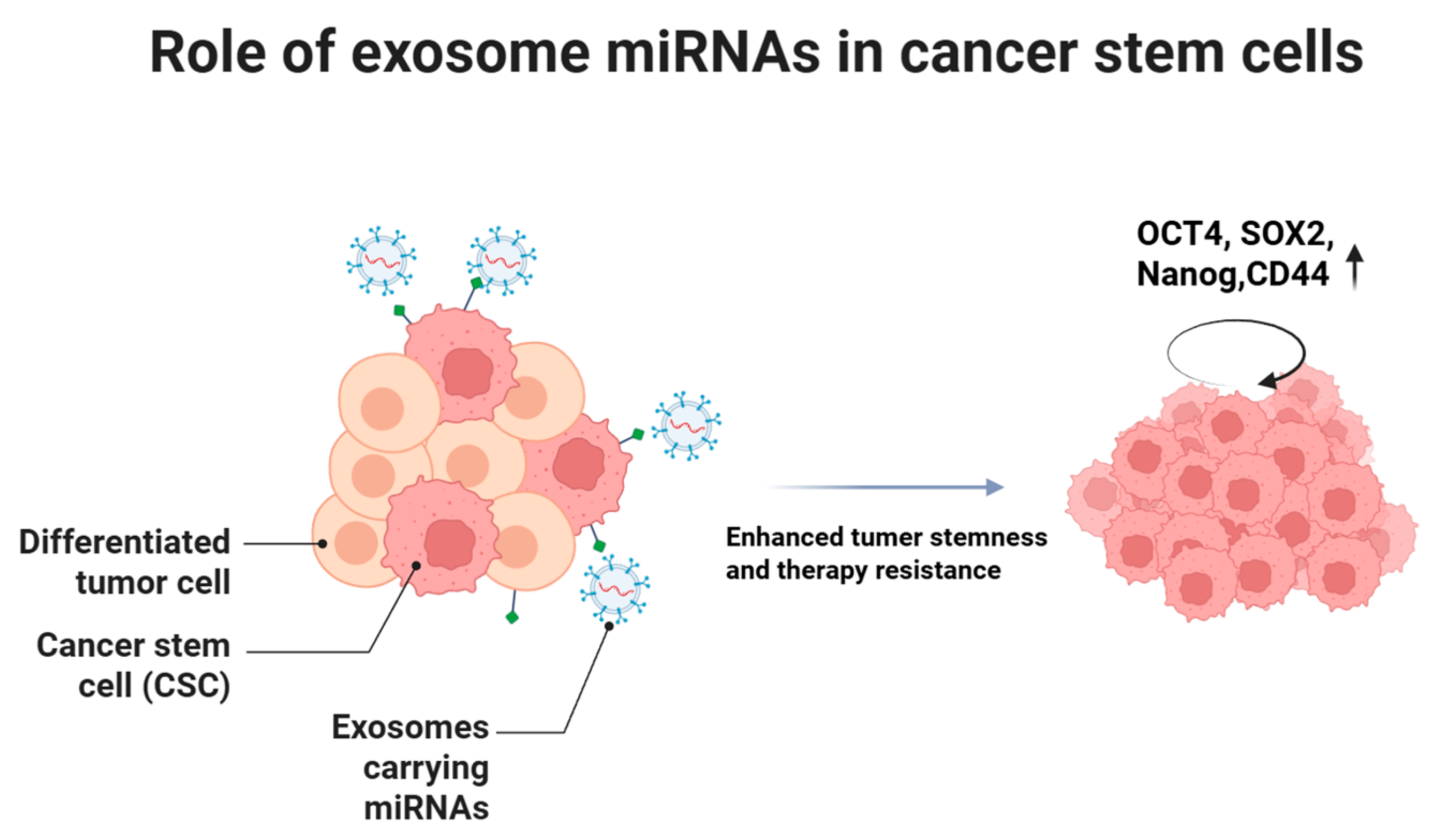

Table 1.
Roles of exosomal miRNAs in cancer progression and cancer stem cell stemness maintenance.
| Exosomal miRNA | Source | Function | Mechanism | Reference | |
|---|---|---|---|---|---|
| miR-200b | Colorectal cancer | Promotes tumor growth | Specifically binds to the 3′-UTR of tumor suppressor p27, reducing its expression | [34] | |
| miR-584 | Hepatocellular carcinoma | Promotes tumor growth | Targets TAK1 and related signaling pathways, leading to downregulation of TAK1 expression | [35] | |
| miR-148a | Glioblastoma | Promotes tumor growth | Targets CADM1 to relieve its inhibitory effect on the STAT3 pathway | [36] | |
| miR-1246 | Breast cancer | Promotes tumor growth | Specifically downregulates cyclin G2 (CCNG2), disrupting the cell cycle regulatory network | [37] | |
| miR-128 | Breast cancer | Promotes tumor metastasis | Regulates the balance of Bcl-2 family proteins (e.g., pro-apoptotic factor Bax), inhibiting initiation of apoptosis | [38] | |
| miR-21 | Oral squamous cell carcinoma | Promotes tumor metastasis | Regulates EMT core transcription factor Snail; upregulates vimentin and downregulates E-cadherin expression | [40] | |
| miR-19b-3p | Renal cell carcinoma | Promotes tumor metastasis | Reduces PTEN expression, inducing EMT | [41] | |
| miR-193a-3p, miR-210-3p, miR-5100 | Mesenchymal stem cells | Promotes tumor metastasis | Activates STAT3 signaling and induces EMT, promoting lung cancer metastasis | [42] | |
| miR-335-5p | Colorectal cancer | Promotes tumor metastasis | Targets RASA1 to promote EMT | [43] | |
| miR-103 | Hepatocellular carcinoma | Promotes tumor angiogenesis | Targets multiple endothelial junction proteins (VE-cadherin, p120-catenin, ZO-1), enhancing vascular permeability | [45] | |
| miR-23a | Nasopharyngeal carcinoma | Promotes tumor angiogenesis | Promotes endothelial cell proliferation, migration, and tube formation; targets and inhibits TSGA10 | [46] | |
| miR-210 | Leukemia | Promotes tumor angiogenesis | Targets and inhibits Ephrin-A3, activating VEGF/VEGFR2 signaling | [47] | |
| miR-21 | Lung cancer | Promotes tumor angiogenesis | Activates STAT3 signaling and upregulates VEGF expression | [49] | |
| miR-23a | Lung cancer | Promotes tumor angiogenesis | Inhibits PHD and tight junction protein ZO-1, increasing vascular permeability | [50] | |
| miR-141-3p | Ovarian cancer | Promotes tumor angiogenesis | Activates JAK-STAT3 signaling in endothelial cells | [51] | |
| miR-9 | Nasopharyngeal carcinoma | Promotes tumor angiogenesis | Regulates PDK/Akt signaling to inhibit NPC cell migration | [53] | |
| miR-21 | Colorectal cancer | Promotes chemoresistance | Downregulates PTEN and hMSH2, inducing resistance to 5-fluorouracil | [54] | |
| miR-221/222 | Breast cancer | Promotes chemoresistance | Inhibits p27 and ERα expression, enhancing tamoxifen resistance | [55] | |
| miR-208b | Colorectal cancer | Promotes chemoresistance | Targets PDCD4 to promote Treg activation, increasing oxaliplatin resistance | [56] | |
| miR-196a | Head and neck cancer | Promotes chemoresistance | Targets CDKN1B and ING5, promoting proliferation, inhibiting apoptosis, and enhancing cisplatin resistance | [59] | |
| miR-222-3p | Ovarian cancer | Promotes immune suppression | Targets SOCS3 and activates STAT3 signaling, inducing M2 macrophage polarization | [65] | |
| miR-301a-3p | Pancreatic cancer | Promotes immune suppression | Inhibits PTEN and activates PI3Kγ signaling, driving M2 macrophage polarization | [66] | |
| miR-21,miR-29 | Non-small cell lung cancer | Promotes immune suppression | Activates TLR-mediated NF-κB signaling, inducing a pro-tumor inflammatory microenvironment | [68] | |
| miR-212-3p | Pancreatic cancer | Promotes immune suppression | Inhibits RFXAP expression, reducing MHC II levels and inducing immune tolerance | [69] | |
| miR-20a | Breast cancer | Promotes immune suppression | Downregulates NKG2D ligands (MICA and MICB), reducing NK cell recognition and killing ability | [70] | |
| miR-24-3p | Nasopharyngeal carcinoma | Promotes immune suppression | Targets PTEN, activates PI3K/Akt pathway, and upregulates PD-L1, inhibiting T-cell function | [73] | |
| miR-144 | Lung cancer | Promotes tumor metabolism | Upregulates GLUT1, increasing glucose uptake and lactate production | [75] | |
| miR-21 | Non-small cell lung cancer | Promotes tumor metabolism | Increases intracellular lipid content and upregulates key lipid metabolism enzymes (FASN, ACC1, FABP5) | [76] | |
| miR-328-3p | Ovarian cancer | Promotes autophagy | Targets Raf1 and inhibits mTOR pathway activation | [81] | |
| miR-425 | Non-small cell lung cancer | Promotes autophagy | Targets AKT1 to activate autophagy | [83] | |
| miR-1910-3p | Breast cancer | Promotes autophagy | Targets MTMR3 and activates NF-κB signaling | [84] | |
| miR-378a-3p | Breast cancer | Maintains CSC stemness | Activates Wnt/Notch pathway | [89] | |
| miR-454 | Breast cancer | Maintains CSC stemness | Activates PRRT2/Wnt axis | [90] | |
| miR-328-3p | Ovarian CSCs | Maintains CSC stemness | Targets and inhibits DNA damage-binding protein 2 | [93] | |
| miR-30b-3p | Glioma CSCs | Mediates CSC therapy resistance | Targets RHOB, reducing apoptosis and enhancing TMZ resistance | [97] | |
| miR-210 | Pancreatic CSCs | Mediates CSC therapy resistance | Upregulates MDR1, YB-1, BCRP, and activates mTOR signaling | [99] | |
| miR-155 | Breast CSCs | Mediates CSC therapy resistance | Regulates C/EBP-β and suppresses TGF-β, C/EBP-β, FOXO3a, inducing EMT and chemoresistance | [101] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



