Submitted:
06 August 2025
Posted:
06 August 2025
You are already at the latest version
Abstract
In Chile Santiago, urban plants are highly vulnerable to drought, as they are mainly exotic from the northern hemisphere with temperate origins. We hypothesize that, although species may generally differ in their response to water restriction, we would find high growth and survival rates among native species of central Chile. The goal was to determine the effect of the year season and an irrigation gradient on the growth and survival of native plant, in order to evaluate potential plant for use in urban green areas of central Chile. Four plots of 20 m2 were located in the Santiago center. In June 2024 twelve species were planted and from November 2024 to March 2025 were irrigated with 13.3, 10.1, 1.7 and 1.4 L/m2/day in each o the lots. The GLM and Kaplan-Meier survival analyses were used. Shoot growth rate was highly variable among species, among irrigation treatments applied, and among year seasons. Eight species shown water-related growth and shoot growth during the winter was very small and higher in spring. Two species showed evidence of water-related survival; in the other 10 species, no significant differences were found between irrigation treatments. Winter was the season with the highest survival rates for eleven species. In conclusion, the evidence suggests that using native species with low water requirements is an effective and efficient way to use water in the urban green area of central Chile.
Keywords:
urban plant
; native plant
; plant survival
; plant growth
; water use
; central Chile
1. Introduction
Urban vegetation is known to improve air quality, ameliorate summer heat island temperatures, lower cooling costs [1], reduce storm water runoff [2], reducing atmospheric carbon dioxide [3], and improves various indicators of social and individual development among residents [4,5,6,7,8]. Urban populations have a very positive average opinion of urban vegetation, particularly valuing its shade, aesthetics, air quality, and noise reduction [9].
The lack of vegetation in central Chilean cities (e.g., only 5.7% of Santiago’s population has access to >9 m2 green space/inhabitant) is accentuated by the unsustainable design of urban areas [10], and characterized by high maintenance and irrigation demands and driven by the selection of plant species that are not adapted to regional urban conditions (soil, atmosphere, water, nutrients). Urban vegetation in cities in central Chile is also deteriorated and stressed by climate change [11], along with adverse growing conditions including compacted soil, extreme heat, lack of nutrients, drought, damage from cars, pruning, and vandalism [12]. Any plan for the development or replacement of urban vegetation in the region should thus be based on experimental evidence in real urban areas, at least considering the survival and growth of potential urban plants. Unfortunately, experimental studies with native species are not common in urban areas located in the Mediterranean-type climate region of central Chile.
Vegetation survival and growth in an urban setting depends on planting location, installation, and post-planting care, between others [13,14]. For example, the most common environmental conditions influencing urban tree mortality are related to water stress, nutrient deficiency, and soil compaction [15,16]. Plant species that successfully colonize and persist in public areas would likely possess traits for stress-tolerance or avoidance. In seasonally dry ecosystems, such as those of central Chile, water is the limiting factor for vegetation growth and survival [17]. It is important to know which species persist and how plant communities change over time so that urban area can be designed to support urban biodiversity for future generations.
Due to the variability in urban tree care during and after planting, it is important to choose plant species with a high chance of survival and growth, and with minimal supplemental care required after installation [18]. Published studies have primarily focused on urban trees’ survival and growth, and the majority of research about urban tree success comes from experiments conducted in relatively controlled nursery settings rather than in sites exposed to urban tree environments. However, other living forms such as urban herbs and shrubs can provide diversity and functionality to urban ecosystems, while survival and growth are conditioned by the urban habitat.
A study of this characteristic has important implications for urban species selection in a changing climate. Cities located in Mediterranean-type climates are particularly vulnerable to climate change. Mainly are predicted to face reduced annual precipitation, temperature increase, and intensified rainfall events [19]. In Santiago, Chile, urban trees are highly vulnerable to drought, as they are mainly exotic species that not adapted to water scarcity, forcing municipalities to use potable water for irrigation [20]. Assessing tolerance or resistance to climate change for potential urban plants helps architects and urban landscape designers select plants suited to a city facing water scarcity [21].
In this study, we estimated the survival and growth rates of seedlings over 10 consecutive months through seasonal monitoring of 12 potential native species of central Chile for use in urban green infrastructure in Santiago. For the analyses, the cumulative survival rate and the growth index of the shoot per season of the year were used for each species tested. These indicators are widely used in demography and population ecology.
We hypothesize that, although species may generally differ in their response to water restriction, we would find high survival rates among native species in central Chile during the first year of the trial in an urban area. The main purpose of the study was to determine the effect of the season and an irrigation gradient on the growth and survival of native species in order to evaluate potential plant species for use in green areas in Mediterranean-type climate cities in central Chile.
2. Results
2.1. Shoot Growth
Shoot growth varied among species (Figure 2A). The species with the highest average growth during all the study period were the shrubs Baccharis linearis (16.7 ± 1.8 cm), Andeimalva chilensis (14.3 ± 1.1 cm), Sphaeralcea obtusiloba (12.0 ± 1.6 cm), and the tree Vachelia caven (13.0 ± 1.8 cm). At the opposite end of the spectrum, the woody Cistanthe laxiflora (0.6 ± 1.3) and the succulent herbs Puya alpestri (2.9 ± 1.8 cm) and P. coerulea (2.0 ± 1.9 cm) showed the lowest average growth of the species studied during the study period. The average growth of woody species (9.5 ± 0.6 cm) during the study period was higher than that of non-woody species (3.3 ± 0.8 cm), with a significant difference between both (F = 37.98; p < 0.001).
Shoot growth rate varied among the irrigation treatments applied in the 66.6% of the evaluated species (Table 1). In particular, in all species which showed evidence of water-sensitive growth, individuals subjected to a higher irrigation volume tended to show greater shoot average growth. Overall, the combined water-sensitive growth species in the treatment with higher irrigation (average 9.9 cm throughout the test) was > 50% higher than the combined water-sensitive growth species in the treatment with lower irrigation (average 3.3 cm throughout the test).
The woody Cistanthe laxiflora (-3.2 ± 1.4 cm), the herb Nasella laevissima (0.9 ± 1.2 cm), the succulents P. alpestri (1.3 ± 1.1 cm) and P. coerulea (0,02 ± 1.1 cm) showed very low average growth in the lowest irrigation volume treatment (Table 1). In fact, C. laxiflora and S. cumingii stand out because they were the species that showed negative growth in response to the lowest water treatments of 1.4 L/m2/day and 1.7 L/m2/day, respectively (Table 1).
The remaining four species were Baccharis linearis, Balbisia peduncularis, Encelia canescens, and Sphaeralcea obtusiloba. They did not exhibit water-sensitive growth, and there was no statistical difference in shoot growth between the applied irrigation treatments (Table 1).
Shoot growth was highly variable among year seasons in all the species tested (Table 2). In the 12 species, shoot growth during the winter season was very small, it was even negative in E. canescens (F = 12.25; p ≤ 0.001), and in S. obtusiloba (F = 19.65; p ≤ 0.001) although without differences among treatments in both species (Table 2). However, spring was the most productive season, except for the tree V. caven (Figure 1A), which grew faster in summer (F = 20.31; p < 0.001). In contrast, growth in C. laxiflora decreased significantly and was negative during the summer (F = 10.09; p < 0.01). The combined growth of the 12 species was also significantly higher in spring than in winter or summer (F = 52.53; p < 0.001). A similar pattern, although more pronounced, was found when comparing woody species with non-woody species combined (Figure 1B, Table 2).
2.2. Species Survival
The final cumulative survival rate for the 12 species combined averaged at 85.9%. At one extreme, B. linearis, P. alpestri, and V. caven had a final cumulative survival rate of 100% (Figure 2A). At the other extreme, the species A. chilensis and E. canescens achieved a final cumulative survival of 75.0% and 59.6%, respectively (Figure 2A). Even E. canescens achieved a statistical survival rate lower than A. chilensis (χ2 = 4.56; p = 0.033) and, consequently, lower than all the species tested. The rest of the 10 species showed a cumulative final survival rate of at least 85.4%.
The 12 species showed survival rates above 80% in each season monitored (Figure 2A). Winter was the season with the highest survival rates for all species except E. canescens. Ten species showed 100% survival during winter (Figure 2A). The species C. odorífera, P. coerulea, and S. obtusiloba achieved a final survival rate of 100% in at least two seasons (Figure 2A). Four species had a final survival rate of 100% during the summer: B. linearis, P. alpestri, P. coerulea, and V. caven.
The combined non-woody species had better survival rates during winter and summer as well, while the combined woody species had higher survival rates only during winter (Figure 2B).
On the other hand, only 2 species showed evidence of water-sensitive survival (Table 1). The species C. laxiflora and N. laevisima showed statistically lower survival rates in the lower irrigation treatment (1.4 L/m2/day). In the other 10 species tested, no significant differences were found between irrigation treatments.
No evidence was found that shoot growth rate was associated with the final survival rate or the survival rate achieved according to the irrigation treatment applied to the species tested.
3. Discussion
Vegetation in public areas of cities has emerged as a key component of nature-based solutions in socio-ecologically stressed urban environments, although the biogeographical origin of its components has not been sufficiently emphasized in central Chile [36]. Several studies of Mediterranean-type climate regions around the globe suggest that native species should be preferred because they contribute more to ecosystem functioning and the ecological integrity of urban environments, and provide a greater number of ecosystem services [37,38,39,40,41]. Additionally, it could be crucial to select native species not only for their ecological features, but also having evolved in particular habitats characterized by summer drought stress similar to those present in the urban area of central Chile [42]. This further inclusion could greatly expand the number of plants potentially suitable for growing in cities with Mediterranean-type climates. The present study fits within this conceptual framework.
In regions with a Mediterranean-type climate, methodologies have been designed to identify potential native species that are not used in various urban vegetation contexts. These methodologies use bibliographic information on the species ecology, distribution, and performance in their natural conditions [43]. At first approach, this is an appropriate strategy for defining the potential components of urban vegetation that should be selected. However, it should not be forgotten that experimental evidence should also be gathered in the contexts of diverse urban habitats [44].
One set of evidence comes from assessing the condition and health of historic vegetation in different areas of the city [45]. A second set comes from managing and monitoring vegetation from the moment an urban green area is implemented, even if it was not considered an experimental design from the outset [46]. This study is closer to the second group, as it sought potential native species to be used in public areas in experimental urban contexts in a central Chile city. Although published information on native species from central Chile was used for an initial selection of potential species (see Table 3), the plants were then subjected to an experimental procedure to evaluate their performance in real urban conditions.
The results of this study showed that the irrigation gradient tested did not affect survival in 83% of the potential native species selected for use in urban areas of central Chile. The result is consistent with studies of Mediterranean species in natural conditions, where survival differences due to irrigation are not always observed, probably because these species have low water requirements [47,48,49]. In fact, the results show that overall final survival is greater than 80%, with the exception of the E. canescens shrub. It should be noted that the first year is a bottleneck for urban plant survival, due to the stress suffered by individuals during their transfer and transplantation, including the effects of environmental pollution [13,50]. Our results even showed that survival was high after summer water stress.
On the other hand, the results showed that shoot growth in the tested species was highly variable between species (Figure 1A). They also showed that approximately 66% of the species studied were significantly affected by the irrigation gradient. However, all species studied had shoot growth practically stop during the winter season regardless of the treatment applied (Figure 1A). The highest shoot growth in woody and non-woody species was in the spring season (Figure 1B).
Consequently, the research shows that the species studied exhibit temporal variation in their water behavior patterns consistent with species from Mediterranean-type climates, characterized in central Chile by a winter influenced by the polar jet stream, numerous frosts with snow at higher elevations, an extended period of summer drought, and high interannual variability and temperatures [51]. Results show that these plants stop or slow down shoot growth during the winter and summer seasons, respectively. However, survival was not significantly affected in both seasons. According to the literature, cold ocean currents on the west coast of Mediterranean-type climate regions moderate temperatures, allowing survival and plant growth in the late winter and early spring [22].
Although this topic needs to be studied in greater detail, growth during winter 2024 was probably affected by low temperatures, since the year was particularly cold in central Chile [52]. According to the experimental design, during winter the plots did not receive additional irrigation. The plots were not watered until late spring (November and December) and maintained during the winter with the water that initially saturated the soil and with the natural rainfall and atmospheric humidity regime. Water availability and the water potential of the soil and atmosphere were thus very similar across all experimental plots and sufficient for plant survival at the beginning of spring (Figure 3). Slightly above-zero growth during winter in this study is therefore likely due to low temperatures.
The results of this study showed that only the herb N. laevissima and the shrub C. laxiflora presented highly water-sensitive responses, as both growth and survival were significantly affected (Table 1). In fact, growth was negative in C. laxiflora and zero in N. laevissima, and in both species, survival was lower in the treatment with less water availability (Table 1, Table 2). Both species concentrated their shoot growth during the spring, and the results even show that C. laxiflora suffered significant self-thinning during the summer. ROS (Reactive Oxygen Species) analyses, which were carried out in parallel with this study, indicate potential oxidative stress in the herb N. laevissima in midsummer for low irrigation treatments (unpublished data). The environmental stress suffered by this species could explain its low abundance in natural conditions in the Metropolitan Region of central Chile [53] and its displacement by exotic annual plants that use water more efficiently [54]. On the contrary, the same ROS analysis shows the absence of oxidative stress in the C. laxiflora shrub in any of the treatments applied during the same period of the year, probably due to the decrease in both the transpiration surface area and water loss suffered by the species during this period of greater water stress [55]. However, C. laxiflora survival was negatively affected (Figure 2A), probably due to a decrease in the photosynthetic area caused by self-thinning during the summer.
The results of this study also showed that 50% of the species studied were water-sensitive, as only growth was significantly affected by the treatments applied (Table 1). These species are characterized by high survival rates, which are not affected by the irrigation gradient applied. However, growth was practically stopped in low irrigation conditions, especially during winter.
The succulent herbs P. alpestri and P. coerulea stand out in this group, as they have adapted their photosynthesis to avoid water stress in the summer [56]. Although the results showed that the species were water-sensitive growth, practically stopped in both species when the plants were subjected to 1.7 and 1.4 L/m2/day, however survival was 100% in all treatments (Table 1). This pattern demonstrates the ability of both succulent species to colonize sites exposed to solar radiation (Table 3) and to avoid water stress in the summer [56]. Even during this season, H2O2 levels associated with water stress are normal in both succulents of this study (unpublished data). However, during the winter, growth was zero in both species, probably indicating stress levels due to cold weather events and good resistance to the same phenomenon, as the average survival rate was close to 100%. A pattern very similar to that found in P. alpestri and P. coerulea can be seen in the shrubs S. cumingii, B. linearis, and in the tree V. caven. The plants of S. cumingii have deep roots that perform hydraulic lifting, allowing them to reach more humid soil layers and move water to dry surface layers, thus avoiding water stress during the summer [57,58]. However, we can also see that summer is the season with the highest mortality rate and the season with the highest growth rate in urban areas as well. With regard to V. caven, a dominant species of thorny shrubland [59] and represented in the native urban flora of central Chile [20], research shows high survival rates throughout the year, although growth has been limited under severe water restrictions. On the other hand, unlike S. cumingii, in the tree V. caven there is no differentiated use of water within the root zone, but rather constant activity to capture the water available at a depth of 100 cm [60]. The last representative of the species water-sensitive is B. linearis, which develops shallow roots. This shrub is suitable for colonizing barren soils and remediating soils contaminated by mining [61], making it an attractive candidate for rehabilitating urban soils with abundant fill.
Analysis of the results also detected a group of four potential species that were not water-sensitive to the treatments applied and showed high survival rates, with the exception of E. canescens (Table 1). This shrub from arid regions showed a very low survival rate during winter and spring (Figure 2A), which could not even be reverted with the increased water availability in the irrigation gradient applied (Table 1). Interestingly, the literature shows that E. canescens is a common species that is resistant to low water availability [62]. It is suggested that plants from more arid areas with similar behavior could benefit from the establishment of seedlings in shaded environments, although this hypothesis would need to be evaluated more precisely [18,63]. The ROS analysis for E. canescens was consistent with the results for survival and shoot growth in the water gradient (Table 1) because it showed potential oxidative stress during the summer in all treatments, and was even reinforced in the plot with low water availability. The low survival rate of E. canescens during winter is probably due to frost stress, which cannot be reversed and recovered from before the onset of summer water stress in plots that are completely exposed to solar radiation. In contrast, although the S. obtusiloba shrub also showed oxidative stress during the summer, which was intensified in the low irrigation treatment (data not shown), its survival rate was high with growth not stopping under any water conditions. In this species, there is probably a mechanism associated with secondary metabolites such as proline, sugars, or antioxidants to prevent water stress, which should be evaluated in further studies (Claudia Ortiz-Calderón, pers. comm.).
The last water-insensitive species, the shrub B. peduncularis, reduces its water deficit after the application of more irrigation in the summer season. The reviewed literature estimates that B. peduncularis responds efficiently to increases in surface water availability and is therefore dependent on rainfall and surface soil moisture [64].
These types of studies must consider the importance of both current environmental conditions and future climate scenarios in areas undergoing urbanization in central Chile. We selected a group of species that are not only found in the central Chilean Mediterranean-type climate region, but also in more arid areas in northern Chile. Biodiversity in Mediterranean climate areas is particularly susceptible to global change [65]. Urban vegetation proposals should not only improve the adaptability of urban green infrastructure in a changing environment, but also contribute to the broader goals of biodiversity conservation in a particularly vulnerable and endangered Mediterranean-type area [65,66,67].
4. Materials and Methods
4.1. Study Site
Santiago (33° S; 70° W; 550 m.a.s.l.), the Chilean capital, has a Mediterranean climate type, characterized by mild wet winters and warm dry summers (Figure 3) [22,23]. The Santiago metropolitan area currently has about 6.3 million people, with a population density of roughly 8,497 inhabitants per Km2 [24]. Since the late 20th and early 21th centuries, urban growth has also spread to surrounding areas, mostly consisting of agricultural lands and smaller remnants of semi-natural vegetation [25,26,27]. Urban vegetation of Santiago is dominated by exotic flora, which represents more than 80% of the total urban flora [20]. We chose Santiago as a study site in central Chile due to the Bio-urban Shelter project designed for public area in Santiago [28].
4.2. Species Selection
We used the landscape value method to select 12 potential native plant species from central Chile for use in urban vegetation in the same region (Fernández et al. 2025). The method consists of four selection criteria: a) environmental criteria, which are native species adapted to the environment where they will be used; b) aesthetic criteria, morphological features that have aesthetic value for the landscape use of the species; c) cultural criteria, which refer to the meanings and values of the use of the flora; and d) management criteria, which is the ability of the species to respond to management and conditions in public area. Consideration was given to ensuring that the selected species were available in nurseries in the region as well, due to potential demand from local governments for use in the Bio-urban Shelter project designed [28].
4.3. Experimental Design
The experimental plots are located in the historic center of the city of Santiago, Chile. Two experimental plots were set up on the campus of the Universidad Tecnológica Metropolitana (UTEM, 33.451071°S, 70.656622°W) and two similar plots at the Universidad Central de Chile (UCEN; 33.451360°S, 70.653433°W).
Each plot had an area of 20 m2. The soil in the plots was homogenized with a structure suitable for plant growth. Consequently, 60% of the existing soil in the top 150 cm of the plots was cleaned and loosened for recovery. The remaining 50% of the planting substrate consisted of 20% coarse sand, 15% gravel, and 5% compost, which increased the organic matter in the soil. A 5 cm layer of organic mulch was also added to the surface to prevent weed growth and excessive water loss from the soil. The chemical and biological composition of the soil on the plots was suitable for plant growth, on average: nitrogen = 32 mg/kg, phosphorus = 67 mg/kg, potassium = 374 mg/kg, % organic matter = 2.7; % water retention = 47.86; pH = 7.8 and electrical conductivity = 1.25 mS/cm; alkaline phosphatase = 24.88 ug/g soil h; DAF activity = 39.95 ug Fluorescein/g soil; urease = 17.23 ug N-NH4 /g soil; biomass carbon = 484.96 mg C/kg soil.
The 12 native species selected from central Chile, including woody and herbaceous perennials, were planted in each plot (Figure 4A). Three species of geophytes were also planted in the plots as part of a complementary study. The spatial arrangement of the species was the same in the four experimental plots and responded to landscape and monitoring criteria, as the experiment is part of a project on bio-urban shelters in public area in Santiago.
Juvenile plants under two years old were obtained from certified nurseries and, after being purchased, were immediately planted in the plots. The plants were planted in May 2024 in water-saturated soil to avoid stress and mortality. The irrigation system installed was a long-lasting, low-yield automated underground drip system with a pipe with a built-in emitter and Copper Shield technology (Figure 4B), which protects the dripper from root intrusion [34]. The hose has a diameter of 16 mm and a flow rate of 2.3 L/h. The automatic irrigation was controlled by a Rainbird ESP LXM programmer. The system was fed by the drinking water system. In each plot, the irrigation lines were buried at a depth of 30 cm and spaced 30.5 cm apart. Each plot was subject to a different water regime. The 2 plots located at UTEM were irrigated daily during the irrigation season with an average of 10.1 L/m2/day and 1.7 L/m2/day. Meanwhile, the 2 plots located at UCEN had an average daily irrigation regime of 13.3 L/m2/day and 1.4 L/m2/day. Irrigation began on 22 November 2024 and continued when the soil moisture content reached 100 cb in the two plots with the higher water availability and 200 cb (2 December 2024) in the other two plots. The water gradient applied represents the recommended irrigation for public green areas in the Santiago Metropolitan Region at the upper end, and conditions considered to be water stress for urban plants at the lower end [35]. The experiment lasted from 29 July 2024 to 03 March 2025.
4.4. Statistical Design
To evaluate survival in woody and herbaceous perennial species, plant shoots were visually monitored at the end of the 2024 winter season, the end of spring 2024, and the end of summer 2025. If the plant shoot was completely dry, the individual was considered dead, considering that it could subsequently regrow at the base of the shoot and its condition could change to alive. To determine the effect of water treatment on species survival, Kaplan-Meier survival analyses were performed. This analysis is based on estimating the conditional probabilities of survival at each monitoring event or season (winter, spring, and summer) and considering the limit of the product of those probabilities to estimate the survival rate at each event. Kaplan-Meier analyses were also done to determine the effect of water treatment on the survival of woody versus non-woody species in combination.
The growth of each individual will be estimated using the following growth index:
GI = (A + B + C)/3
A = plant height, B = canopy length, C = canopy width
The growth per individual for each season of the year was estimated by:
GIt1 − GIt0
t1 = end of the current year season,
t0 = immediately preceding year season.
The growth index of each individual was monitored at the end of three seasons of the year (winter on 13 September 2024, spring on 17 December 2024, and summer on 03 March 2024).
The growth index was analyzed using a generalized linear model of repeated measures. This analysis determines the effect of the four irrigation treatments on the growth index by performing inter-subject tests. Bonferroni was used for comparisons between pairs of treatments. The data were transformed to log10 if they did not meet normality and homoscedasticity. The effect of the season and its interaction with the treatments was determined using intra-subject effect tests. When the sphericity condition was not met according to Mauchly’s test, the Greenhouse-Geisser correction or, failing that, the Huynh-Feldt correction was performed in the intra-subject effect tests. A significance level of p < 0.05 was considered in the analyses. Statistical analyses were performed using the IBM SPSS Statistics for PC, version 30.0 (IBM SPSS, Armonk, NY, USA: IBM Corp.2024).
For the purposes of this study: we defined a species as being highly water-sensitive if significant differences in plant growth and survival were observed between the different water treatments applied during the trial. A species is water-sensitive if significant differences in plant growth or survival is observed between the different water treatments applied during the trial. Finally, a species is considered non-sensitive if there are no significant differences in plant growth and survival between water treatments.
5. Conclusions
Survival results show that almost 80% of the native central Chileans species selected for this study were not affected by the water treatments applied, and even reached high percentages in the first year of establishment, which is considered a survival bottleneck for urban plants. However, when water resources were limited to ≤ 1.7 L/m2/day, shoot growth can be slowed significantly, because they are water-sensitive species, consequently survival rates may remain high. The study also showed that the main growing season for both woody and non-woody species is spring. Growth is then slowed during the summer, with even biomass being lost.
Finally, growth is stopped during the winter season. In summary, the evidence obtained in this study suggests that using native species with low water requirements in urban projects is an effective and efficient way to use water in Mediterranean-type climate regions such as central Chile.
Author Contributions
Conceptualization, J.F. and F.F.C.; methodology, J.F. and F.F.C.; software, J.F.; validation, J.F., R.C.-J., A.C.-C. and F.F.C.; formal analysis, J.F.; investigation, J.F., R.C.-J., A.C.-C. and F.F.C.; data curation, J.F. and F.F.C.; writing—original draft preparation, J.F.; writing—review and editing, J.F.; visualization, J.F. and F.F.C.; supervision, J.F.; project administration, R.C.-J., A.C.-C.; funding acquisition, R.C.-J., A.C.-C., F.F.C. and J.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by ANID/IDEA ID23I10043.
Data Availability Statement
The data presented in this study are available on request from the corresponding author J.F.
Acknowledgments
We would like to thank Carlos Bustamante-Oleart, Carlos Bustamante Espina, M. Villagrán-Escobar, Javier Ríos Vilche, José Cerón Córdova, Consuelo Chaparro Gómez and Margarita Reyes for maintaining the databases and providing technical support at various stages of the project. We would like to express our sincere gratitude to Claudia Ortiz-Calderón for her valuable contributions, which have enabled us to improve our analysis of the results.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- McPherson, E.G. Cooling urban heat islands with sustainable landscapes. In The ecological city: preserving and restoring urban biodiversity; Platt, R.H., Rowntree, R.A., Muick, P.C, Eds.; University of Massachusetts Press: Massachusetts, USA, 1994; pp. 151-171.
- Richter, M.; Heinemann, K.; Meiser, N.; Dickhaut, W. Trees in sponge cities—a systematic review of trees as a component of blue-green infrastructure, vegetation engineering principles, and Stormwater management. Water 2024, 16, 655. [Google Scholar] [CrossRef]
- Rasoolzadeh, R.; Mobarghaee, N.; Esmaeilzadeh, H.; Rashidi, Y.; Marcu, M.V.; Sadeghi, S.M.M. Carbon sequestration and storage of urban trees in a polluted semiarid city. Forests 2024, 15, 1488. [Google Scholar] [CrossRef]
- Burch Jr, W.R., Grove, J.M. People, trees and participation on the urban frontier. Unasylva 1993, 44, 19-27.
- Dwyer, J.F.; McPherson, E.G.; Schroeder, H.W. Rowntree, R.A. Assessing the benefits and costs of the urban forest. J. Arboric. 1992, 18, 227–234. [Google Scholar] [CrossRef]
- Simpson, J. R.; McPherson, E.G. San Francisco bay area state of the urban forest final report. Center for Urban Forest Research USDA Forest Service, Pacific Southwest Research Station: California, USA, 2007; p. 81.
- Faber Taylor, A.; Kuo, F.E. Could exposure to everyday green spaces help treat ADHD? Evidence from children’s play settings. Appl. Psychol Health Well Being. 2011, 3, 281–303. [Google Scholar] [CrossRef]
- Bele, A.; Chakradeo, U. Public perception of biodiversity: a literature review of its role in urban green spaces. J. Land. Ecol. 2021, 14, 1–28. [Google Scholar] [CrossRef]
- Lohr, V.I.; Pearson-Mims, C.H.; Tarnai, J.; Dillman, D.A. How urban resident rate and rank the benefits and problems associated with trees in cities. Arboric. Urban For. 2004, 30, 28–35. [Google Scholar] [CrossRef]
- Irarrázaval, F. El imaginario” verde” y el verde urbano como instrumento de consumo inmobiliario: configurando las condiciones ambientales del área Metropolitana de Santiago. Revista INVI 2012, 27, 73–103. [Google Scholar] [CrossRef]
- Uribe, S.V.; Villaseñor, N.R. Inequities in urban tree care based on socioeconomic status. Urban For. Urban Green. 2024, 96, 128363. [Google Scholar] [CrossRef]
- Kornienko, V.; Reuckaya, V.; Shkirenko, A.; Meskhi, B.; Olshevskaya, A.; Odabashyan, M.; Shevchenko, V.; Teplyakova, S. Silvicultural and Ecological Characteristics of Populus bolleana Lauche as a Key Introduced Species in the Urban Dendroflora of Industrial Cities. Plants 2025, 14, 2052. [Google Scholar] [CrossRef]
- de la Fuente, L.M.; Ovalle, J.F.; Arellano, E.C.; Ginocchio, R. Does woody species with contrasting root architecture require different container size in nursery? Madera y Bosques 2018, 24, e2421419. [Google Scholar] [CrossRef]
- Allen, K.S.; Harper, R.W.; Bayer, A.; Brazee, N.J. A review of nursery production systems and their influence on urban tree survival. Urban For. Urban Green. 2017, 21, 183–191. [Google Scholar] [CrossRef]
- Esperon-Rodriguez, M.; Rymer, P.D.; Power, S.A.; Barton, D.N.; Cariñanos, P.; Dobbs, C., et al., Tjoelker, M.G. Assessing climate risk to support urban forests in a changing climate. Plants People Planet. 2022, 4, 201-213. [CrossRef]
- Kim, Y.J.; Yoo, G. Suggested key variables for assessment of soil quality in urban roadside tree systems. J. Soils Sediments 2021, 21, 2130–2140. [Google Scholar] [CrossRef]
- Vico, G.; Thompson, S.E.; Manzoni, S.; Molini, A.; Albertson, J.D.; Almeida-Cortez, J.S.; Fay, P.A.; Feng, X.; Guswa, A.J.; Liu, H.; Wilson, T.G.; Porporato, A. Climatic, ecophysiological, and phenological control son plant ecohydrological strategies in seasonally dry ecosystems. Ecohydrol. 2015, 8, 660–681. [Google Scholar] [CrossRef]
- Yáñez, M.A.; Espinoza, S.E.; Magni, C.R.; Martínez-Herrera, E. Early Growth and physiological acclimation to shade and water restriction of seven sclerophyllous species of the Mediterranean forests of central Chile. Plants 2024, 13, 2410. [Google Scholar] [CrossRef]
- Fuentes, I.; Fuster, R.; Avilés, D.; Vervoort, W. Water scarcity in central Chile: the effect of climate and land cover changes on hydrologic resources. Hydrolog. Sci. J. 2021, 66, 1028–1044. [Google Scholar] [CrossRef]
- Figueroa, J.A.; Teillier, S.; Guerrero-Leiva, N.; Ray-Bobadilla, C.; Rivano, S.; Saavedra, D.; Castro, S.A. Flora vascular en el espacio público de Santiago, Chile. Gayana Bot. 2016, 73, 85–103. [Google Scholar] [CrossRef]
- Ekren, E.; Çorbacı, Ö.L.; Kordon, S. Evaluation of plants based on ecological tolerance criteria: A case study of urban open green spaces in Rize, Türkiye. Turk J For Sci. 2024, 8, 107–131. [Google Scholar] [CrossRef]
- Rundel, P.W.; Arroyo, M.T.; Cowling, R.M.; Keeley, J.E.; Lamont, B.B.; Vargas, P. Mediterranean biomes: evolution of their vegetation, floras, and climate. Annu. Rev. Ecol. Evol. Syst. 47, 2016, 383-407. [CrossRef]
- McPhee, J.; Cortés, G.; Rojas, M.; García, L.; Descalzi, A.; Vargas, L. Downscaling climate changes for Santiago: What effects can be expected? In Climate adaptation Santiago; Krellenberg, K., Hansjürgens, B. Eds.; Springer-Verlag: Berlin Heidelberg, 2014; pp. 19-41.
- Densidad de Población y Vivienda. Censo 2017. Available online: https://storymaps.arcgis.com/stories/fc03ad1481f44b6299b81c22c91497fe (accessed on 28 July 2025).
- De Mattos, C. Globalización y transformación metropolitana en el caso de Santiago. In Los nuevos modos de gestión de la metropolización; Hidalgo, R., Arenas, F., Coll, J.L., Eds.; Pontificia Universidad Católica de Chile: Santiago, Chile, 2003; pp. 27–55.
- Romero, H.; Vásquez, A. Evaluación ambiental del proceso de urbanización de las cuencas del piedemonte andino de Santiago de Chile. Eur Secur. 2005, 94, 97–118. [Google Scholar] [CrossRef]
- Romero, H.; Molina, M.; Moscoso, C.; Sarricolea, P.; Smith, P.; Vásquez, A. Caracterización de los cambios de usos y coberturas de suelo causados por la expansión urbana de Santiago, análisis de sus factores explicativos e inferencias ambientales. In Movilidad espacial y Reconfiguración Metropolitana; de Mattos, C., Hidalgo, R., Eds.; Pontificia Universidad Católica de Chile: Santiago, Chile, 2007; pp. 251-270.
- Chandia-Jaure, R.; Cataldo-Cunich, A.; Bustamante-Oleart, C.; Fernandez Cano, F.; Figueroa, J.A.; Villagrán-Escobar, M. Bio-Urban Shelters. Neighborhood-scale intervention model for water sensitive urban design. In 14° Encuentro Diseño Urbano, Readu, Punta Arenas, Chile, 6 November 2024.
- Agrupación de especies nativas según condiciones agroecológicas aptas para su cultivo. Available online: https://www.pumahuida.cl/wp-content/uploads/2022/03/AGRUPACI%C3%93N-DE-ESPECIES-NATIVAS-SEG%C3%9AN-CONDICIONES-AGROECOL%C3%93GICAS-APTAS-PARA-SU-CULTIVO-M.-MUSALEM_VI-CONGRESO-DE-FLORA-NATIVA-2019_opt.pdf, accessed on 28 July 2025.
- Listas de Especies Nativas para Distintas Situaciones de Paisaje. Available online: https://www.pumahuida.cl/informacion-tecnica/ (accessed on 28 July 2025).
- Rodríguez, R.; Marticorena, C.; Alarcón, D.; Baeza, C.; Cavieres, L.; Finot, V.L.; Fuentes, N.; Kiessling, A.; Mihoc, M.; Pauchard, A.; Ruiz, E.; Sanchez, P.; Marticorena, A. Catálogo de las plantas vasculares de Chile. Gayana Bot. 2018, 75, 1–430. [Google Scholar] [CrossRef]
- Hoffmann, J.A. Flora Silvestre de Chile. Zona central. Una guía para la identificación de las especies vegetales más frecuentes. 5ta ed.; Fundación Claudio Gay: Santiago, Chile, 2012; p. 250.
- Luebert, F.; Pliscoff, P. Sinopsis bioclimática y vegetacional de Chile. Editorial Universitaria: Santiago, Chile. 2017; p. 316.
- Ivelic-Sáez, J.; Reckmann, O.; López, R.; Uribe, H.; Valenzuela, J.; Ibarra, D. Bases para el riego en Magallanes. Boletín INIA (Chile). 2021, 459, 92. [Google Scholar]
- Fernández, F.; Chiang, L.; Figueroa, J.A. Guía de recomendaciones para jardines eficientes en el espacio público en la Región Metropolitana. Universidad Central de Chile: Santiago, Chile. 2025; p. 63.
- Figueroa, J.A.; Castro, S.A.; Reyes, M.; Teillier, S. Urban park area and age determine the richness of native and exotic plants in parks of a Latin American city: Santiago as a case study. Urban Ecosyst. 2018, 21, 645–655. [Google Scholar] [CrossRef]
- Blasi, C.; Biondi, E.; Izco, J. 100 years of plant sociology: A celebration. Plant Biosyst.-J. Deal. Asp. Plant Biol. 2011, 145, 1–3. [Google Scholar] [CrossRef]
- Säumel, I.; Weber, F.; Kowarik, I. Toward livable and healthy urban streets: Roadside vegetation provides ecosystem services where people live and move. Environ. Sci. Policy 2016, 62, 24–33. [Google Scholar] [CrossRef]
- Conway, T.M.; Almas, A.D.; Coore, D. Ecosystem services, ecological integrity, and native species planting: How to balance these ideas in urban forest management? Urban For. Urban Green. 2019, 41, 1–5. [Google Scholar] [CrossRef]
- Arcos-LeBert, G.; Aravena-Hidalgo, T.; Figueroa, J.A.; Jaksic, F.M.; Castro, S.A. Native trees provide more benefits than exotic trees when ecosystem services are weighted in Santiago, Chile. Trees 2021, 35, 1663–1672. [Google Scholar] [CrossRef]
- D’Amato, L.; Bartoli, F.; Savo, V.; Caneva, G. Promoting native biodiversity: An evaluation of multifactorial and bioclimatic selection criteria for street trees in Italian cities. Urban For. Urban Green. 2025, 107, 128784. [Google Scholar] [CrossRef]
- Rundel, P.W. Landscape disturbance in Mediterranean-type ecosystems: an overview. In Landscape disturbance and biodiversity in Mediterranean-type ecosystems; Rundel, P.W., Montenegro, G., Jaksic, F.M., Eds.; Springer-Verleg: Berlin Heidelberg, Germany, 1998; pp. 3-22.
- Fernández, F.; Delaunoy, J.; Chiang, L.; Reyes, M. Metodología de valor paisajístico: Selección de plantas nativas para la infraestructura verde pública de la Región Metropolitana. In Biodiversidad Urbana en Chile: Estado del arte y los desafíos futuros. Figueroa, J.A., Lazzoni, I. Eds.; 2da. ed.; Ediciones Universidad Central de Chile: Santiago, Chile; in press.
- Grimm, N.B.; Pickett, S.T.; Hale, R.L.; Cadenasso, M.L. Does the ecological concept of disturbance have utility in urban social–ecological–technological systems? Ecosyst. Health Sustain. 2017, 3, e01255. [Google Scholar] [CrossRef]
- Degerickx, J.; Roberts, D.A.; McFadden, J.P., Hermy, M.; Somers, B. Urban tree health assessment using airborne hyperspectral and LiDAR imagery. Int. J. Appl. Earth. Geoinf. 2018, 73, 26-38. [CrossRef]
- Ward, E.B.; Doroski, D.A.; Felson, A.J.; Hallett, R.A.; Oldfield, E.E.; Kuebbing, S.E.; Bradford, M.A. Positive long-term impacts of restoration on soils in an experimental urban forest. Ecol. Appl. 2021, 31, e02336. [Google Scholar] [CrossRef] [PubMed]
- Armesto, J.J.; Pickett, S.T.A. Experiments on disturbance in old-field plant communities: impact on species richness and abundance. Ecology 1985, 66, 230–240. [Google Scholar] [CrossRef]
- Becerra, P.I.; Cruz, G.; Ríos, S.; Castelli, G. Importance of irrigation and plant size in the establishment success of different native species in a degraded ecosystem of central Chile. Bosque 2013, 34, 23–24. [Google Scholar] [CrossRef]
- Becerra, P.I.; Arellano, E.C.; Vilagrosa, A.; Hernández, G.; Figueroa, C. The provision of water and shade but not soil amendments in degraded habitats increases the seedling survival of woody species in restoration processes of the Chilean sclerophyllous forest. Trees 2024, 38, 523–535. [Google Scholar] [CrossRef]
- Razzaghmanesh, M.; Beecham, S.; Kazemi, F. The growth and survival of plants in urban green roofs in a dry climate. Sci. Total Environ. 2014, 476, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Armesto, J.J.; Arroyo, M.T.K.; Hinojosa, F.L. The Mediterranean environment of central PM Chile. In The Physical Geography of South America; Veblen, T.T., Young, K.R., Orme, A.R., Eds.; Oxford Univ. Press: Oxford, UK, 2007; pp. 184–199. [Google Scholar]
- Dirección General de Aeronáutica Civil. Dirección Meteorológica de Chile—Servicios climáticos. Available online: https://climatologia.meteochile.gob.cl, accessed on 28 July 2025.
- Teillier, S.; Macaya-Berti, J.; García, N.; Marticorena, A.; Rojas, G.; Niemeyer, H.M. Flora de la Región Metropolitana de Santiago. Guía para la identificación de las especies. Asia Pacific Offset: China; 2022. 671 p.
- Everard, K.; Seabloom, E.W.; Harpole, W.S.; De Mazancourt, C. Plant water use affects competition for nitrogen: why drought favors invasive species in California. Amer. Naturalist 2010, 175, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Burlett, R.; Trueba, S.; Bouteiller, X.P.; Forget, G.; Torres-Ruiz, J.M.; Martin-StPaul, N.K., et al. Delzon, S. Minimum leaf conductance during drought: unravelling its variability and impact on plant survival. New Phytol. 2025, 246, 1001-1014. [CrossRef]
- Ortuño, M.A.; Machtig, A.E.; Chacón, M.A.; Cuzmar, J.; Fontúrbel, F.E. Spatial distribution of Puya coerulea Lindl. in response to abiotic factors and accompanying species in the Río Clarillo National Reserve. Gayana Bot. 2019, 76, 115–118. [Google Scholar] [CrossRef]
- Prieto, I.; Martínez-Tillería, K.; Martínez-Manchego, L.; Montecinos, S.; Pugnaire, F.I.; Squeo, F.A. Hydraulic lift through transpiration suppression in shrubs from two arid ecosystems: patterns and control mechanisms. Oecologia 2010, 163, 855–865. [Google Scholar] [CrossRef]
- León, M.F.; Squeo, F.A.; Gutiérrez, J.R.; Holmgren, M. Rapid root extension during water pulses enhances establishment of shrub seedlings in the Atacama Desert. J. Veg. Sci. 2011, 22, 120–129. [Google Scholar] [CrossRef]
- Gerstmann, C.; Miranda, M.; Condal, A. Description of space-time variability of the potential productivity of Acacia caven espinales based on MODIS images and the Enhanced Vegetation Index (EVI). Int. J. Agric. Nat. Res. 2010, 37, 63–73. [Google Scholar] [CrossRef]
- Sepúlveda, M.; Bown, H.E.; Fernandez, B. Stomatal conductance responses of Acacia caven to seasonal patterns of water availability at different soil depths in a Mediterranean savanna. Water 2018, 10, 1534. [Google Scholar] [CrossRef]
- Ginocchio, R.; de la Fuente, L.M.; Orrego, F.; Díaz, M.J.; Báez, J.; Ovalle, J.F. A novel fast-vegetative propagation technique of the pioneer shrub Baccharis linearis on mine tailings by adding compost. Int. J. Phytoremediation. 2021, 23, 1169–1174. [Google Scholar] [CrossRef]
- Martínez-Tillería, K.; Loayza, A.P.; Sandquist, D.R.; Squeo, F.A.; Ward, D. No evidence of a trade-off between drought and shade tolerance in seedlings of six coastal desert shrub species in north-central Chile. J. Veg. Sci. 2012, 23, 1051–1061. [Google Scholar] [CrossRef]
- Carvajal, D.E.; Loayza, A.P.; López, R.P.; Toro, P.J.; Squeo, F.A. Growth and early seedling survival of four Atacama Desert shrub species under experimental light and water availability regimes. Rev. Chil. Hist. Nat. 2014, 87, 1–9. [Google Scholar] [CrossRef]
- Torres, R.; Squeo, F.A.; Jorquera, C.; Aguirre, E.; Ehleringer, J.R. Evaluación de la capacidad estacional de utilizar eventos de precipitación en tres especies de arbustos nativos de Chile con distintos sistemas radiculares. Rev. Chil. Hist. Nat. 2002, 75, 737–749. [Google Scholar] [CrossRef]
- Newbold, T.; Oppenheimer, P.; Etard, A.; Williams, J.J. Tropical and Mediterranean biodiversity is disproportionately sensitive to land-use and climate change. Nat. Ecol. Evol. 2020, 4, 1630–1638. [Google Scholar] [CrossRef] [PubMed]
- Caneva, G.; Kumbaric, A.; Savo, V.; Casalini, R. Ecological approach in selecting extensive green roof plants: A data-set of Mediterranean plants. Plant Biosyst. 2015, 149, 374–383. [Google Scholar] [CrossRef]
- Bambach, N.; Meza, F.J.; Gilabert, H.; Miranda, M. Impacts of climate change on the distribution of species and communities in the Chilean Mediterranean ecosystem. Reg. Environ. Change 2013, 13, 1245–1257. [Google Scholar] [CrossRef]
Figure 1.
(a) Shoot yield index for winter, spring, and summer seasons in 12 native plant species from central Chile. Bars represent ±1 E.D. * indicate statistically significant differences among seasonal for each species assessed according to Bonferroni test (p ≤ 0.05); (b) Shoot yield index for winter, spring, and summer seasons for woody and non-woody species combined. Bars represent ±1 E.D. Different letters (a-c) above the bars indicate statistically significant differences among seasonal according to Bonferroni test (p ≤ 0.05).
Figure 1.
(a) Shoot yield index for winter, spring, and summer seasons in 12 native plant species from central Chile. Bars represent ±1 E.D. * indicate statistically significant differences among seasonal for each species assessed according to Bonferroni test (p ≤ 0.05); (b) Shoot yield index for winter, spring, and summer seasons for woody and non-woody species combined. Bars represent ±1 E.D. Different letters (a-c) above the bars indicate statistically significant differences among seasonal according to Bonferroni test (p ≤ 0.05).
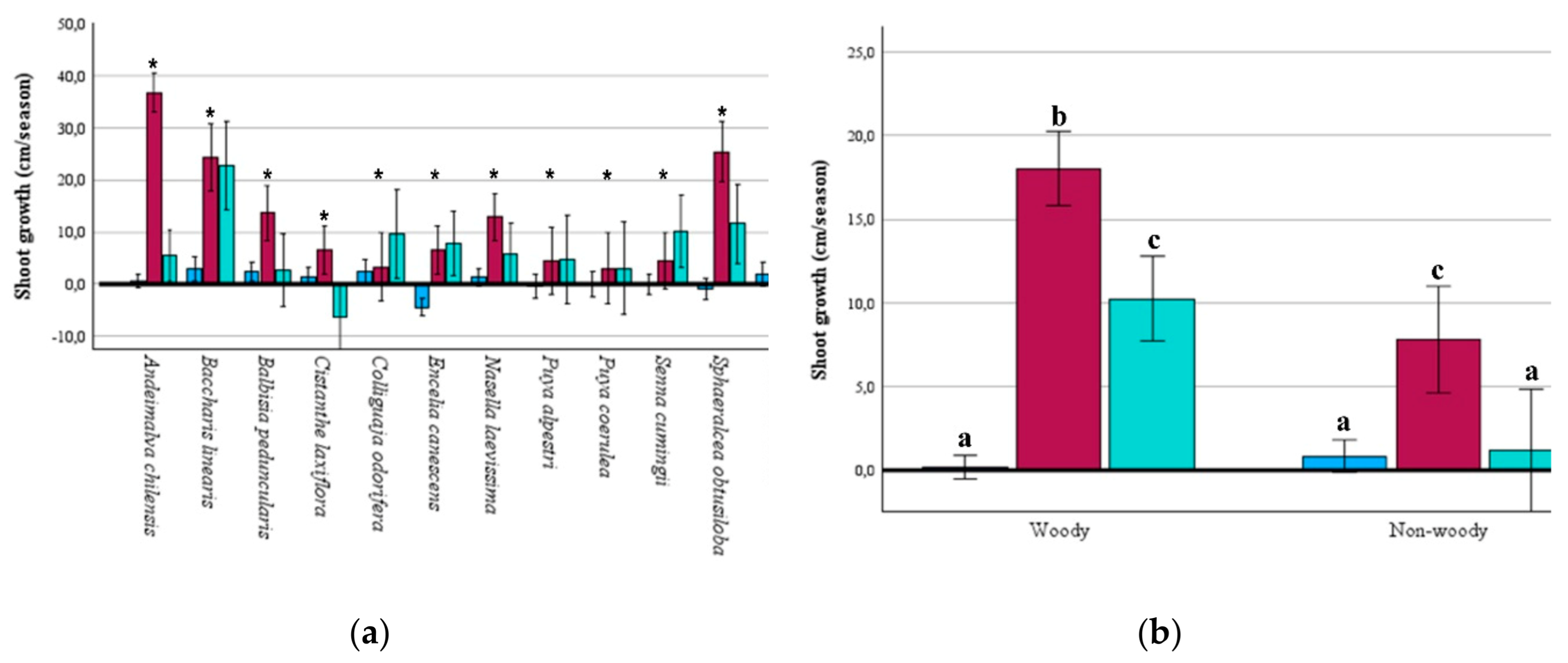
Figure 2.
(a) Survival percentage during for winter, spring, and summer seasons in 12 native plant species from central Chile; (b) Survival percentage for winter, spring, and summer seasons for woody and non-woody species combined.
Figure 2.
(a) Survival percentage during for winter, spring, and summer seasons in 12 native plant species from central Chile; (b) Survival percentage for winter, spring, and summer seasons for woody and non-woody species combined.
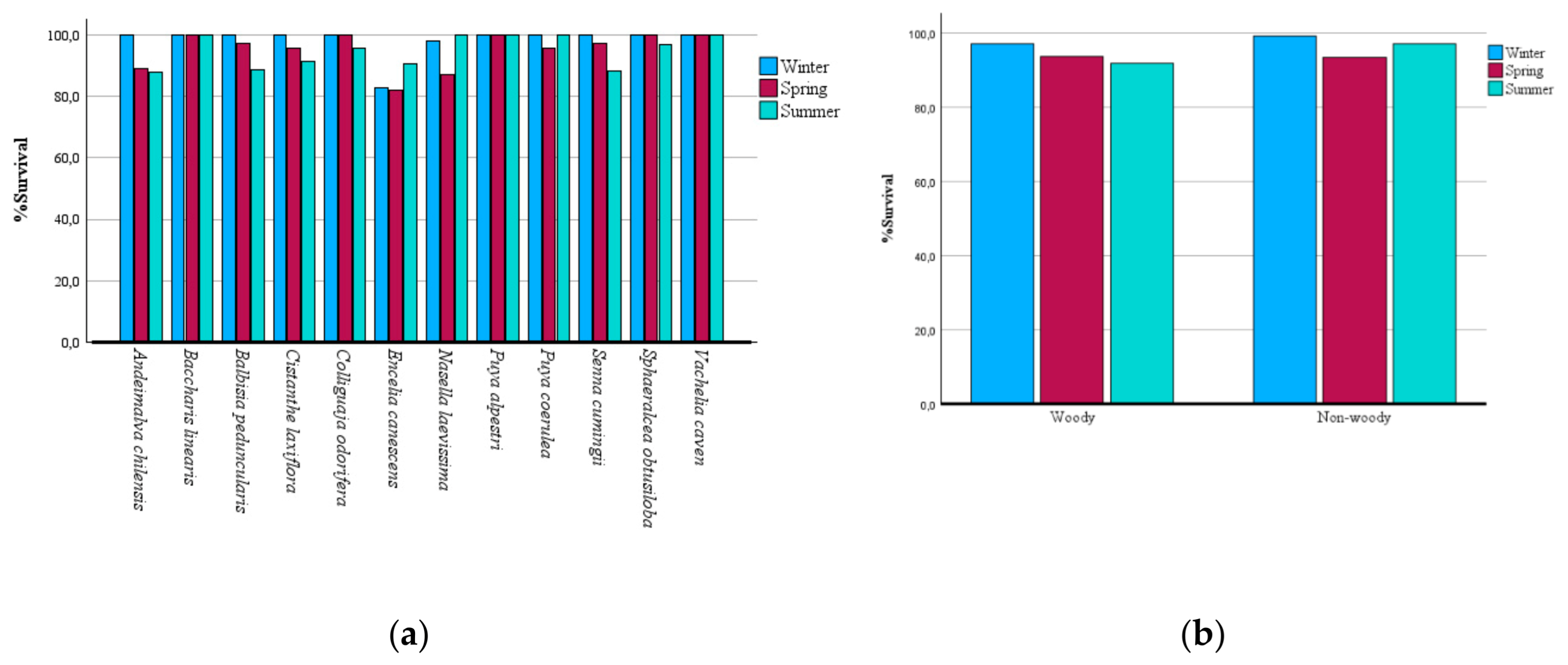
Figure 3.
Mean monthly maximum and minimum temperature and precipitation for Universidad Central de Chile, Santiago, for April 2024 to March 2025.
Figure 3.
Mean monthly maximum and minimum temperature and precipitation for Universidad Central de Chile, Santiago, for April 2024 to March 2025.
Figure 4.
(a) Planting plan for the four experimental plots. Two plots were located Universidad Tecnológica Metropolitana and another two Universidad Central de Chile, in the historic center of the city of Santiago, central Chile. Vc = Vachelia caven, cl = Cistanthe laxiflora, nl = Nasella laevissima, Ac = Andeimalva chilensis, Co = Colliguaja odorífera, Pa = Puya alpestri, Pco = Puya coerulea, Senna cumingii, Bl = Baccharis linearis, Bp = Balbisia peduncularis, Ec = Encelia canescens, So = Sphaeralcea obtusiloba; (b) Long-lasting, low-yield automated underground drip system with a pipe with a built-in emitter and Copper Shield technology.
Figure 4.
(a) Planting plan for the four experimental plots. Two plots were located Universidad Tecnológica Metropolitana and another two Universidad Central de Chile, in the historic center of the city of Santiago, central Chile. Vc = Vachelia caven, cl = Cistanthe laxiflora, nl = Nasella laevissima, Ac = Andeimalva chilensis, Co = Colliguaja odorífera, Pa = Puya alpestri, Pco = Puya coerulea, Senna cumingii, Bl = Baccharis linearis, Bp = Balbisia peduncularis, Ec = Encelia canescens, So = Sphaeralcea obtusiloba; (b) Long-lasting, low-yield automated underground drip system with a pipe with a built-in emitter and Copper Shield technology.


Table 1.
Shoot growth index (±1 E.D.) and survival rate (final %) for each irrigation treatment applied to the 12 species of native plants from central Chile tested in the experimental plots. N = total number of individuals tested per species. Note: * p ≤ 0.05; ** p ≤ 0.01; *** p ≤ 0.001; n.s. = p > 0.05.
Table 1.
Shoot growth index (±1 E.D.) and survival rate (final %) for each irrigation treatment applied to the 12 species of native plants from central Chile tested in the experimental plots. N = total number of individuals tested per species. Note: * p ≤ 0.05; ** p ≤ 0.01; *** p ≤ 0.001; n.s. = p > 0.05.
| Species | N | Dependent variable | 13.3 L/m2/day | 10.1 L/m2/day | 1.7 L/m2/day | 1.4 L/m2/day | p-Value | Responses |
|---|---|---|---|---|---|---|---|---|
| Cistanthe laxiflora | 45 | Growth | 2.0 (1.4) a,b | 2.8 (1.4) a | 0.4 (1.4) a,b | -3.2 (1.4) b | * | Highly sensitive |
| Survival | 83.3 a,b | 100 a | 91.7 a,b | 75.0 b | * | |||
| Nasella laevissima | 48 | Growth | 7.4 (1.2) a | 10.5 (1.2) a | 7.9 (1.2) a | 0.9 (1.2) b | *** | Highly sensitive |
| Survival | 83.3 a,b | 91.7 a,b | 100 a | 66.7 b | * | |||
| Andeimalva chilensis | 72 | Growth | 22.3 (3.3) a | 17.7 (3.3) a | 5.6 (3.3) b | 11.6 (3.3) a,b | ** | Sensitive |
| Survival | 83.3 | 72.2 | 61.1 | 83.3 | n.s. | |||
| Colliguaja odorifera | 24 | Growth | 10.7 (2.1) a | 5.0 (2.1) a,b | 1.4 (2.1) b | 3.5 (2.1) a,b | * | Sensitive |
| Survival | 100 | 100 | 83,3 | 100 | n.s. | |||
| Puya alpestri | 24 | Growth | 5.6 (1.1) a | 3.6 (1.1) a,b | 1.2 (1.1) b | 1.3 (1.1) b | * | Sensitive |
| Survival | 100 | 100 | 100 | 100 | n.s. | |||
| Puya coerulea | 23 | Growth | 3.8 (0.8) a | 3.6 (0.8) a | 0.02 (0.9) b | 0.02 (0.9) b | ** | Sensitive |
| Survival | 100 | 100 | 100 | 80 | n.s. | |||
| Senna cumingii | 35 | Growth | 11.0 (2.2) a | 4.1 (2.3) a,b | -0.4 (2.2) b | 4.7 (2.2) a,b | ** | Sensitive |
| Survival | 88,9 | 75,0 | 77,8 | 100 | n.s. | |||
| Vachelia caven | 24 | Growth | 16.7 (2,7) a,b | 21.6 (2.7) a | 6.2 (2.7) b | 7.3 (2.7) b | *** | Sensitive |
| Survival | 100 | 100 | 100 | 100 | n.s. | |||
| Baccharis linearis | 24 | Growth | 15.3 (2.1) | 19.5 (2.1) | 19.5 (2.1) | 12.4 (2.1) | n.s. | Non-sensitive |
| Survival | 100 | 100 | 100 | 100 | n.s. | |||
| Balbisia peduncularis | 36 | Growth | 8.1 (2.7) | 3.1 (2.7) | 7,6 (2.7) | 6.2 (2.7) | n.s. | Non-sensitive |
| Survival | 77.8 | 66.7 | 100 | 100 | n.s. | |||
| Encelia canescens | 47 | Growth | 0.9 (2.9) | 4.1 (2.9) | 5.4 (2.9) | 2.9 (3.1) | n.s. | Non-sensitive |
| Survival | 41.7 | 58.3 | 75.0 | 63.3 | n.s. | |||
| Sphaeralcea obtusiloba | 32 | Growth | 13.9 (3.0) | 16.0 (3.5) | 7.9 (3.0) | 11.2 (3.0) | n.s. | Non-sensitive |
| Survival | 100 | 100 | 87,5 | 100 | n.s. |
Table 2.
Generalized linear model of repeated measures results showing the effect Intra-subject, contrast Intra-subject and of the four irrigation treatments on the growth index by performing Inter-subject tests. N = total number of individuals tested per species. Note: * p ≤ 0.05; ** p ≤ 0.01; *** p ≤ 0.001; n.s. = p > 0.05.
Table 2.
Generalized linear model of repeated measures results showing the effect Intra-subject, contrast Intra-subject and of the four irrigation treatments on the growth index by performing Inter-subject tests. N = total number of individuals tested per species. Note: * p ≤ 0.05; ** p ≤ 0.01; *** p ≤ 0.001; n.s. = p > 0.05.
| Especie | N | Effect Intra-subject | Contrast Intra-subject | Effect Inter-subject | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| TIME | TIME × TREAT | TIME | TIME × TREAT | TREAT | |||||||
| F-Value | p-Value | F-Value | p-Value | F-Value | p-Value | F-Value | p-Value | F-Value | p-Value | ||
| Andeimalva chilensis | 72 | 41.43 | *** | 4.00 | ** | 79.58 | *** | 5.37 | ** | 4.78 | ** |
| Baccharis linearis | 24 | 36.96 | *** | 2.61 | * | 63.80 | *** | 2.67 | n.s. | 2.68 | n.s. |
| Balbisia peduncularis | 36 | 7.71 | ** | 1.62 | n.s. | 17.03 | *** | 1.71 | n.s. | 0.70 | n.s. |
| Cistanthe laxiflora | 45 | 10.09 | ** | 0.29 | n.s. | 10.24 | ** | 0.11 | n.s. | 3.61 | * |
| Colliguaja odorifera | 24 | 4.34 | * | 3.21 | * | 6.04 | * | 4.88 | * | 3.80 | * |
| Encelia canescens | 47 | 12.25 | *** | 0.97 | n.s. | 20.26 | *** | 0.57 | n.s. | 0.74 | n.s. |
| Nasella laevissima | 48 | 30.19 | *** | 3.53 | ** | 33.20 | *** | 0.34 | n.s. | 11.33 | *** |
| Puya alpestri | 24 | 13.82 | *** | 5.74 | *** | 18.74 | *** | 5.48 | ** | 3.55 | * |
| Puya coerulea | 23 | 7.71 | ** | 2.60 | n.s. | 18.19 | *** | 6.30 | ** | 6.10 | ** |
| Senna cumingii | 35 | 7.59 | ** | 2.33 | n.s. | 10.35 | ** | 2.37 | n.s. | 4.71 | ** |
| Sphaeralcea obtusiloba | 32 | 19.65 | *** | 3.38 | ** | 27.26 | *** | 5.06 | ** | 1.2 | n.s. |
| Vachelia caven | 24 | 20.31 | *** | 4.59 | ** | 47.77 | *** | 6.92 | ** | 7.59 | ** |
| Non-woody | 140 | 21.03 | *** | 1.45 | n.s. | 34.11 | *** | 0.09 | n.s. | 8.63 | *** |
| Woody | 294 | 63.15 | *** | 8.27 | *** | 46.97 | *** | 7.44 | *** | 5.73 | *** |
Table 3.
Plant species from central Chile selected for the study, indicating family affiliation, life form, and environmental, aesthetic, cultural, and management criteria considered in the potential landscape value of the species for use in urban vegetation in central Chile. Information obtained in [29,30,31,32].
Table 3.
Plant species from central Chile selected for the study, indicating family affiliation, life form, and environmental, aesthetic, cultural, and management criteria considered in the potential landscape value of the species for use in urban vegetation in central Chile. Information obtained in [29,30,31,32].
| Species | Family | Life form | Criteria | |||
|---|---|---|---|---|---|---|
| Environmental | Aesthetic | Cultural | Management | |||
|
Cistanthe laxiflora |
Montiaceae | Perennial herb | Endemic. Tolerant of saline soils, poor in nutrients, and low moisture. Flowers attract native pollinators | Pink fuchsia flowers in spring and summer | No information | Plant in full sun in association with many species |
|
Nasella laevissima |
Poaceae | Perennial herb | Endemic. Tolerant in low-moisture soils and poor in nutrients and eroded. | Species with diverse shapes, colors, and subject to the wind | No information | Plant in full sun. Fast growth rate |
|
Andeimalva chilensis |
Malvaceae | Shrub | Endemic. Tolerant of saline soils, poor in nutrients, and low moisture. Attracts a diversity of floral fauna. | Extensive pink coloration and attractive appearance of flowering | No information | Plant in full sun, tolerates pruning, and resistant to pests and diseases |
|
Colliguaja odorifera |
Euphorbiaceae | Shrub | Endemic. Tolerant in low-moisture soils and poor in nutrients. | Edges of leaves and flowers are reddish in color. Ballistic seed dispersal | Medicinal. Insecticide. Dyeing. Handcrafted | Plant in full sun. Fast growth rate, resists pruning |
| Puya alpestri | Bromeliaceae | Perennial herb | Endemic. Tolerant in low-moisture soils and poor in nutrients. Attracts hummingbirds and butterflies | Attractive colors, textures, structure, and shapes of the plant. Flowers in spring and fruits in summer | Edible | Plant in full sun and resists pruning |
| Puya coerulea | Bromeliaceae | Perennial herb | Endemic. Tolerant in low-moisture soils and poor in nutrients. Attracts hummingbirds and butterflies | Attractive colors, textures, structure, and shapes of the plant. Flowers in spring and fruits in summer | No information | Plant in full sun and resists pruning |
| Senna cumingii | Fabaceae | Shrub | Endemic. Tolerant of saline soils, poor in nutrients, low moisture and fixes nitrogen | Golden yellow flowers in spring and summer | No information | Plant in full sun |
| Vachelia caven | Fabaceae | Tree | Endemic. Tolerant of saline soils, poor in nutrients, low moisture and fixes nitrogen. Attracts birds | A variety of colors throughout the year | Edible. Medicinal. Fuel. Handcrafted. Dye |
Plant in full sun. Fast growth rate |
|
Baccharis linearis |
Asteraceae | Shrub | Native. Tolerant in low-moisture soils and eroded. Attracts pollinators | Attractive colors, textures, structure, and shapes of the plant | Medicinal | Plant in full sun. Fast growth rate, tolerates pruning, and resistant to pests and diseases |
|
Balbisia peduncularis |
Vivianiaceae | Shrub | Native. Tolerant of saline soils, poor in nutrients, and low moisture. | Attractive flowers in spring and summer. | No information. | Plant in full sun. |
|
Encelia canescens |
Asteraceae | Shrub | Native. Tolerant in low-moisture soils and poor in nutrients. Attracts native pollinating bees | Yellow flowers in spring and fall | No information | Plant in full sun. Fast growth rate |
|
Sphaeralcea obtusiloba |
Malvaceae | Shrub | Endemic. Tolerant in low-moisture soils and poor in nutrients. Attracts native pollinating bees | Lilac flowers in spring and fall. Attractive colors, textures, and shapes of the foliage | Medicinal | Plant in full sun. Fast growth rate and resistant to pests |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



