Submitted:
05 August 2025
Posted:
07 August 2025
Read the latest preprint version here
Abstract
Food allergies are a rising global health concern, influenced by complex interac-tions among genetic predisposition, environmental exposures, dietary habits, and gut microbiota. Sensitization typically occurs through dermal, respiratory, or gas-trointestinal routes, with epithelial barrier dysfunction and Th2-skewed immune responses playing key roles. Understanding the mechanisms and pathways of sensitization is critical to developing effective mitigation strategies. Both thermal and non-thermal food processing technologies are being explored for their poten-tial to reduce the allergenicity of common foods such as milk, egg, peanut, tree nuts, and seafood. Thermal methods can denature allergenic proteins but may al-so compromise nutritional and sensory quality. In contrast, non-thermal technol-ogies—including high-pressure processing, pulsed light, cold plasma, and ultra-sound—offer milder alternatives that modify protein structures, mask or degrade IgE-binding epitopes, and reduce immunoreactivity while preserving food quality. However, outcomes are highly context-dependent, influenced by food matrix, processing parameters, and assessment methods. Limitations in current in vitro and in vivo models, lack of standardized allergenicity testing protocols, and in-sufficient clinical validation remain key barriers. This review synthesizes current knowledge of allergy mechanisms with technological approaches for allergen re-duction, highlighting promising interventions, existing constraints, and the need for integrated research to develop safe, hypoallergenic foods.
Keywords:
food allergy non-thermal processing
; allergenicity reduction
; pulsed electric fields
; high hydrostatic pressure
; ultraviolet
; ultrasonication
; cold plasma
1. Food Allergy and Sensitization: Mechanisms, Pathways, and Risk Factors
Food allergy is defined as an adverse immune response to specific food proteins and affects approximately 6% of children and 3%–4% of adults. Clinical manifestations vary widely but most commonly involve the skin, gastrointestinal tract, and respiratory system. Immunologically, food allergies are classified into IgE-mediated and non–IgE-mediated (cellular) responses [1,2].
Under normal circumstances, the immune system maintains tolerance to dietary antigens—a process known as oral tolerance. Food allergy represents a failure of this immunological tolerance. Although a wide range of foods can elicit allergic reactions, a limited number of foods account for most clinically significant cases. These include milk, eggs, peanuts, tree nuts, fish, and shellfish [2,3,4].
Diagnosis of food allergy requires a structured, evidence-based approach. The process typically begins with a thorough clinical history and is followed by diagnostic tools such as serologic testing, skin prick tests, elimination diets, and, when indicated, oral food challenges. Advances in molecular allergology have led to the characterization of many food allergens, enhancing the understanding of underlying immune mechanisms and paving the way for novel diagnostic and therapeutic strategies. Nevertheless, the current mainstay of management remains strict avoidance of known allergens, comprehensive patient education, and readiness to manage accidental exposures—primarily through the use of emergency epinephrine [2].
Food allergies can develop through various routes of sensitization. Class 1 food allergies result from oral exposure to food proteins and are commonly associated with proteins that are resistant to heat, acid, and enzymatic degradation [5,6,7]. These allergens are often introduced during infancy or early childhood, a period when the immune system is particularly susceptible to dysregulation. In contrast, class 2 food allergies result from primary sensitization to inhaled allergens, such as pollens, that share structural similarity with proteins in raw fruits or vegetables. This can lead to cross-reactivity and symptoms consistent with pollen–food allergy syndrome [5,6].
Research suggests that sensitization to certain class 1 allergens—such as peanuts and eggs—may also occur through the skin, especially in individuals with impaired skin barrier function [8,9]. Many class 1 food allergens are water-soluble glycoproteins ranging from 10 to 70 kDa in size and demonstrate resistance to digestion and thermal processing. Examples include caseins in milk, ovomucoid in eggs, vicilins in peanuts, and nonspecific lipid transfer proteins found in apples (Mal d 3) and corn (Zea m 14) [5,6]. Bet v 1, the major birch pollen allergen, exemplifies a class 2 allergen. It can elicit respiratory sensitization and cause oral symptoms when cross-reactive proteins such as Mal d 1 (apple) or Dau c 1 (carrot) are ingested. Most food allergens fall into a limited number of protein families, including the Cupin and Prolamin superfamilies, and pathogenesis-related proteins [5,6].
The clinical relevance of specific sensitizations varies. For instance, peanut allergens Ara h 1, Ara h 2, and Ara h 3 are often implicated in severe IgE-mediated allergic reactions, whereas Ara h 8—a Bet v 1 homolog—is usually associated with milder symptoms and oral allergy syndrome [10]. Despite structural similarities between proteins, true clinical cross-reactivity is often less common than laboratory tests suggest [1,6].
Food allergies can result in a range of clinical disorders affecting one or more organ systems. Many gastrointestinal disorders attributed to food allergies exhibit overlapping symptoms, but they can often be distinguished through careful clinical evaluation and diagnostic testing [11,12,13]. Additionally, some pediatric gastrointestinal conditions—such as colic, constipation, and reflux—are occasionally suspected to have allergic etiologies (Figure 1).
Anaphylaxis represents one of the most severe outcomes of food allergy, with foods being the leading cause of anaphylactic episodes in outpatient settings [14]. Fatal cases often involve peanut or tree nut exposure in adolescents or young adults with pre-existing food allergies and asthma, and typically occur in the absence of prompt epinephrine administration [12,15].
Adverse food reactions more broadly, termed food intolerances, arise from a variety of mechanisms. These reactions are categorized as toxic or non-toxic. Non–immune-mediated intolerances—such as those due to enzymatic deficiencies (e.g., lactose intolerance) or sensitivity to biogenic amines—are more common than immune-mediated food allergies. Nonetheless, immune-mediated food allergies, though less prevalent, affect millions globally and pose significant medical and economic burdens. In their most severe form, these allergies can cause life-threatening anaphylaxis [2,16,17]. The National Institute of Allergy and Infectious Diseases (NIAID) defines food allergy as “a specific immune response to a food that results in reproducible adverse health effects” [18,19], encompassing both adaptive and innate immune pathways.
The concept of allergy was introduced by Clemens von Pirquet in 1906, who described hypersensitivity reactions such as serum sickness in children receiving antiserum therapies [20,21]. Later, hypersensitivity reactions were classified into four immunological types [22]. The most common food allergy mechanism is Type I hypersensitivity, mediated by IgE antibodies against food allergens. These reactions involve complex immune cascades, including T cell activation and eosinophil recruitment [16]. Diagnosis often includes detection of specific IgE antibodies or IgE-related cellular responses [2].
Although there has been speculation regarding the involvement of food-specific IgG antibodies in Type II or Type III hypersensitivity reactions, current scientific evidence does not support a pathogenic role. Accordingly, professional societies discourage the use of IgG-based food allergy testing [23,24,25]. Type IV hypersensitivity reactions, mediated by T cells, contribute to conditions such as celiac disease, in which gluten proteins trigger autoimmune inflammation in the gut [26]. Another T cell–mediated disorder is food protein-induced enterocolitis syndrome (FPIES), which typically affects infants and is not associated with IgE [27].
Recent studies also implicate the innate immune system in food-induced inflammation. Molecules such as wheat amylase-trypsin inhibitors and certain milk oligosaccharides activate Toll-like receptor 4 (TLR4), initiating intestinal inflammation [4,28]. Such mechanisms may underlie disorders like non-celiac gluten sensitivity [4,29,30].
In industrialized nations, IgE-mediated food allergies affect 3%–8% of children and 1%–3% of adults [2,17,31]. These conditions impose not only physical health risks but also emotional and social burdens due to dietary limitations and lifestyle adjustments. The most common allergenic foods include milk, eggs, wheat, peanuts, tree nuts, sesame, fish, fruits, and vegetables [24,31]. Allergies to milk, eggs, and wheat often resolve in childhood, while allergies to peanuts, tree nuts, and seafood tend to persist [2].
Estimating the true prevalence of food allergies remains challenging due to variability in diagnostic methodologies and reliance on self-reported versus clinically confirmed cases [32]. Nevertheless, evidence suggests that both the incidence and severity of food allergies are increasing, driven by a complex interplay of genetic and environmental factors, including diet and lifestyle [9,33,34,35].
Epigenetic research has revealed that children with IgE-mediated food allergies exhibit distinct DNA methylation patterns in CD4+ T cells, suggesting a role for gene–environment interactions in allergy pathogenesis [36]. The “hygiene hypothesis,” first proposed by Strachan [37], posits that reduced microbial exposure—due to smaller family sizes and improved sanitation—impairs immune system development, increasing susceptibility to allergic disease. Supporting this, children raised in anthroposophic environments—with minimal antibiotic or vaccine exposure and adherence to organic diets—show lower rates of allergies [38,39].
Limited dietary and microbial diversity in early life may impede immune tolerance, contributing to the growing prevalence of allergic and inflammatory conditions in developed societies [40]. Allergic sensitization refers to the immune system’s initial response upon first exposure to an allergen [4,41]. Two primary routes of sensitization have been described. Class 1 allergens—such as milk, egg, and peanut—induce sensitization through the gastrointestinal tract [42]. Class 2 allergens, such as Bet v 1 from birch pollen, sensitize via the respiratory tract and can cross-react with homologous food proteins (e.g., Mal d 1 in apples), resulting in oral allergy syndrome [4,43,44,45,46].
Epicutaneous sensitization—where allergens enter through inflamed or damaged skin—has also been proposed as a sensitization route. In murine models, such exposure leads to intestinal mast cell expansion and systemic anaphylaxis [47,48,49].
Multiple factors influence the likelihood of sensitization:
- Adjuvant effects of food matrices: Nonallergenic components and microbial products in foods may act as immunological adjuvants, promoting sensitization [6].
Conversely, secretory immunoglobulins—particularly SIgA and SIgM—strengthen mucosal defenses and promote oral tolerance. Mice deficient in SIgA transport exhibit increased intestinal permeability and greater vulnerability to allergen-induced anaphylaxis, which can be reversed by the induction of regulatory T cells [54,55].
2. Mitigation Strategies for Food Allergens
2.1. Allergen Elimination and Control Strategies
Elimination or reduction of allergens in the food supply chain requires a multifaceted and preventive approach. Genetic modification or selective breeding is one strategy used to remove or silence genes encoding allergenic proteins, such as the reduction or knockout of Ara h 2 and Ara h 6 in peanuts [56]. Enzymatic hydrolysis is another promising approach, in which proteolytic enzymes cleave allergenic epitopes to decrease immunoreactivity. This method has proven effective in hydrolyzed milk and wheat products [57,58]. However, proteolysis may not fully eliminate allergenic potential. For example, Sen et al. [59] observed that the peanut allergen Ara h 2 retained IgE-binding epitopes post-hydrolysis, especially when disulfide bonds were intact. This highlights the importance of linear epitopes in allergen stability, and suggests that reducing disulfide bonds alone may not sufficiently degrade allergenic regions.
In addition to ingredient-level interventions, allergen control measures begin with careful raw material management. This includes supplier verification and traceability systems to ensure allergen-free inputs [60]. In manufacturing, dedicated lines, validated sanitation, and equipment separation are necessary to avoid cross-contact [61,62,63]. Hazard Analysis and Critical Control Points (HACCP), ingredient monitoring, and accurate food labeling are central to allergen risk management. Finally, workforce training and allergen awareness programs are vital for maintaining safety throughout the supply chain [64].
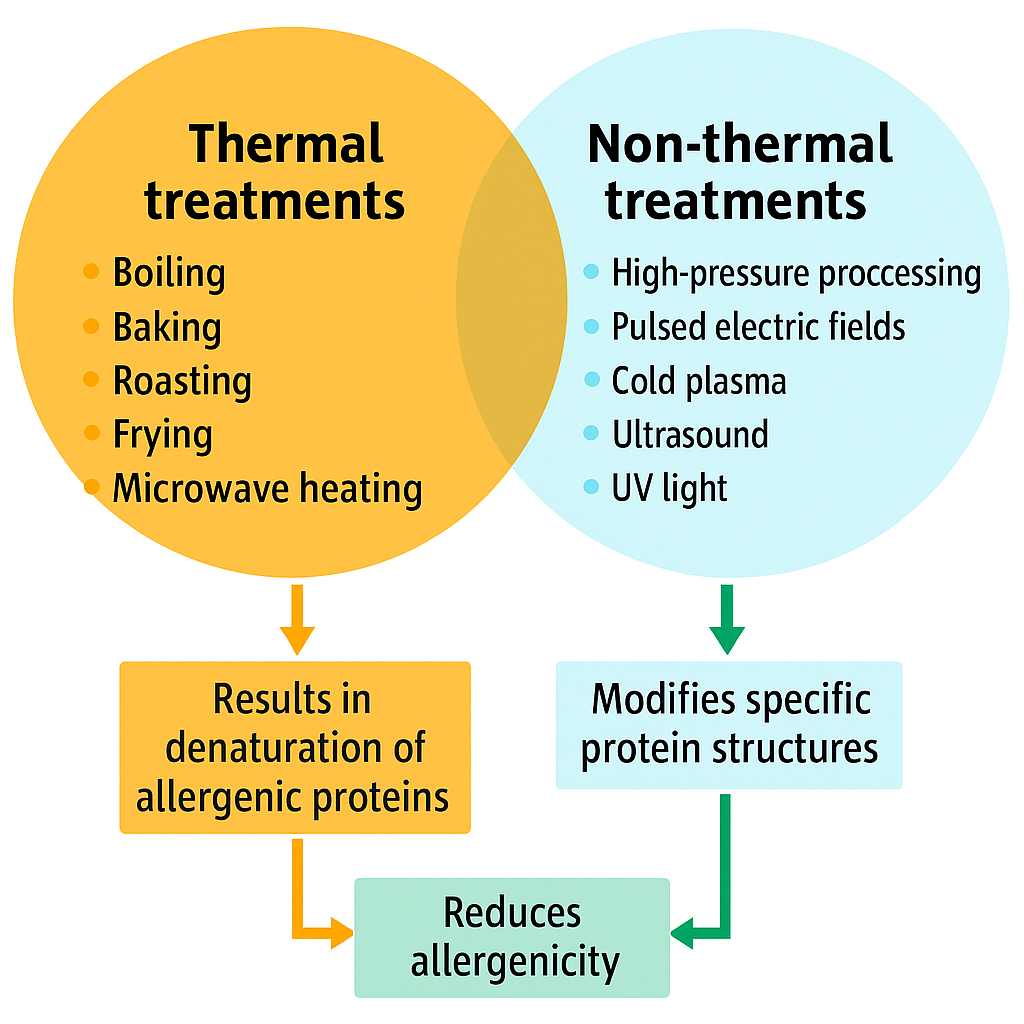
2.2. Effects of Food Processing on Allergen Control
Food processing plays a critical role in modifying allergenic properties while enhancing safety, shelf life, and nutritional value. Physical, chemical, and biochemical processing methods can influence allergenicity based on food type and processing conditions [65,66,67]. The structural integrity and allergenic potential of proteins are significantly influenced by processing. Changes in physicochemical properties and protein stability can alter immunogenicity [57,68,69]. The food matrix can modulate these effects, at times enhancing allergenic responses [70,71]. These treatments may reduce, increase, or have no effect on allergenicity. Notably, some techniques can lead to the formation of neoallergens—new antigenic compounds created during processing—which may intensify allergic reactions [72,73,74].
Thermal and non-thermal food processing can alter epitope structures, reducing allergenicity by disrupting existing immunodominant epitopes or creating neoallergens. The phenomenon of neoallergen formation has been recognized since the 1970s, when it was reported that processed foods could elicit allergic responses even when raw forms did not [75]. Subsequent findings identified neoallergens in processed pecans [76] and wheat flour [77]. Processing typically reduces allergenicity in many foods. Fermentation not only enhances nutritional quality and preservation but also affects allergenicity. Lactic acid bacteria, for example, degrade IgE-binding epitopes in milk proteins, such as β-lactoglobulin, thus reducing allergenicity [78]. However, fermentation’s effects are product-specific. Soy sauce, a fermented product, retains allergenicity despite microbial activity [79], while fermented dairy products such as yogurt often show reduced immunogenicity due to acid denaturation and proteolysis [80,81].
2.3. Thermal Processing and Its Effects on Allergenicity
Thermal treatment is one of the most commonly used technologies in food processing and can significantly alter the structure and immunoreactivity of food allergens. Heat-induced denaturation leads to unfolding of proteins, disruption of disulfide bonds, and potential destruction of IgE-binding epitopes. For example, heating egg proteins such as ovomucoid or ovalbumin at 100°C for extended periods reduces their IgE-binding capacity [81]. Similarly, roasting or boiling peanuts can modify allergenic proteins (e.g., Ara h 1, Ara h 2) and reduce or sometimes enhance allergenicity depending on the conditions (Table 1) [82,83].
Baking and extrusion may reduce the allergenicity of wheat and soy by Maillard reactions and protein aggregation, although new allergenic determinants can sometimes be formed [84]. However, in some cases, thermal processing increases the resistance of certain allergens to digestion, potentially enhancing their allergenic potential. Thus, heat treatments must be carefully optimized based on food type and target allergen.
Studies have demonstrated that moist heat can significantly reduce the allergenic potential of certain foods. For instance, sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) revealed a marked loss of protein bands in canned tuna and salmon extracts compared to raw or conventionally cooked samples. Immunoblotting further showed minimal IgE-specific binding in the canned extracts. ELISA inhibition assays and oral challenges with canned salmon in allergic individuals confirmed reduced allergenicity, suggesting that some fish allergens are more heat-labile than previously thought [60,85].
Similarly, kiwi fruit allergy provides a compelling case of heat-mediated allergenicity reduction. Variability in patient responses has been attributed to protein heterogeneity and cross-reactivity with other allergens, such as pollens. However, steam cooking kiwi at 100°C for 5 minutes followed by homogenization effectively eliminated sensitivity in certain allergic children [86,87,88].
In the case of milk allergens, the IgE-binding capacity of β-lactoglobulin, whether as an isolated protein or within whole milk, was assessed using FEIA-CAP inhibition. While mild heating at 74°C caused only slight reductions, significant decreases in IgE binding were observed at 90°C. Despite reduced binding, a sufficiently high concentration of β-lactoglobulin could still inhibit a similar total amount of IgE antibodies across all heat treatments [80].
Allergic reactions are triggered by IgE antibody binding to specific protein epitopes. Cross-linking of IgE on mast cells or basophils leads to the release of histamine and other mediators [89]. Thermal sensitivity varies by epitope type: conformational epitopes are more easily denatured, while linear epitopes are more heat-resistant [57,90]. As a result, heating may reduce allergenicity in some foods like lentils, enhance it in others like peanuts, or have no effect, as observed in boiled peas [91,92].
Pollen-related allergens in fruits and vegetables often become less immunogenic after heating. However, allergens in foods like shrimp can remain heat-stable and retain immunogenicity post-cooking. The type of epitope also affects stability: conformational epitopes, reliant on three-dimensional protein folding, are more susceptible to denaturation during processing, whereas linear epitopes may resist heat but are subject to enzymatic or chemical alteration. Genetic engineering allows precise modification of such epitopes to reduce allergenicity.
Protein structures respond differently to thermal conditions: mild heat (70–80°C) disrupts secondary structures, moderate heat (80–90°C) affects disulfide bonding, and intense heat (90–100°C) induces aggregation [90,93]. Heat may also initiate Maillard reactions, where lysine residues form covalent bonds with sugars, generating advanced glycation end products that could increase allergenicity [90].
Heating can enhance digestibility and reduce allergenicity in legumes such as chickpeas, lentils, lupins, and black gram [94]. Conversely, it may reduce digestibility in peanut allergens like Ara h 1 and Ara h 2 [83,95]. Thermal processing may also produce neoantigens, which elicit stronger immune responses [69,83]. The Maillard reaction is a key mechanism behind this neoantigen formation, where heat-induced protein-sugar interactions yield glycation products with heightened immunogenic potential [96]. In a study by [97], sera from 57 patients with suspected meat allergy showed stronger IgE binding to raw meat extracts than to those heated at 140°C for 20 minutes, suggesting thermal degradation of most allergens. However, chicken was an exception: six of 24 sera reacted more strongly to heated chicken meat. Common allergenic proteins in raw and cooked chicken were identified at 17–66 kDa, while heat-labile proteins (45 and 150 kDa) disappeared and neoallergens emerged in the 14–90 kDa range.
The IgE-binding capacity of wheat proteins significantly decreases with increasing temperature and heating time. When wheat flour and dough (flour mixed with 50% water) were subjected to heat treatments at 80°C, 100°C, and 120°C for 10, 20, and 60 minutes, both neutral and acidic protein extracts showed reduced IgE reactivity in skin prick tests (SPT) and radioallergosorbent tests (RAST) using sera from cereal-sensitive adults. No further reduction in allergenicity was observed beyond 10 minutes at 120°C, indicating a thermal stability threshold. Pooled sera RAST scores dropped from class 4 (untreated flour) to class 2 following heat treatment. Notably, heated dough demonstrated lower allergenic potential than heated flour at 80°C and 100°C, though this difference was no longer evident at 120°C [98].
Heat treatment by microwave heating of kiwi fruit to 40, 60, 80, and 90°C resulted in a progressive reduction in allergenicity, as assessed by skin prick testing (SPT) in three kiwi-allergic individuals. At 90°C, SPT responses were negative in two patients and markedly reduced in the third, indicating significant thermal degradation of allergenic proteins [99].
Protein solubility in minced beef decreases with extended heat treatment. Following heating at 85°C for up to 2 hours, SDS–PAGE revealed faint bands at 17.8, 19, 14, 20, 45, and >60 kDa, which were also detectable in well-cooked samples (80°C, 20 min) via immunoblotting. Among these, the 17.8 kDa protein showed the highest IgE-binding activity in patients with confirmed allergy to cooked beef (positive DBPCFC). Bovine serum albumin and gamma-globulin were undetectable in minced beef heated at 80°C for 10 and 3 minutes, respectively. However, isolated serum albumin remained stable at 95°C for 15 minutes, whereas isolated gamma-globulin was fully denatured at 65°C within 15 min [100].
Dry heat processing can influence the allergenic potential of foods through complex chemical reactions such as the Maillard reaction and enzymatic browning. These processes can irreversibly alter or destroy conformational epitopes. For example, Gruber et al. [101] demonstrated that cherry allergen Pru av 1 loses its IgE-binding capacity due to the Maillard reaction and polyphenol oxidase–catalyzed oxidation. Similarly, Hansen et al. [102] reported that roasting hazelnuts reduced their allergenicity, particularly among individuals sensitized to birch pollen–related allergens such as Cor a 1.04 and Cor a 2. However, five out of seventeen birch pollen–allergic individuals still reacted to roasted hazelnuts in double-blind placebo-controlled food challenge tests, indicating that roasting does not eliminate allergenicity for all patients. These findings suggest that individual sensitivity to conformational versus linear epitopes is a critical consideration in evaluating the allergenic potential of thermally processed foods. Conversely, Maleki et sl. [103] found that thermal processing actually increased the IgE-binding capacity of major peanut allergens Ara h 1 and Ara h 2, indicating that heating may enhance peanut allergenicity in some cases.
Because processing alone may not eliminate all allergens, integrating multiple strategies is more effective [104]. Thermal techniques such as blanching, pasteurization, canning, and roasting offer accessible and cost-effective allergen mitigation and microbial control [105]. Freezing inhibits microbial growth below −9.5°C (15°F), contributing to food stability [60,79]. Drying enhances shelf life and lowers storage costs, though its allergen-modifying effects vary. Edible crickets with three major allergens identified: tropomyosin, actin, and hexamerin-like protein 2, particularly Gryllus bimaculatus (two-spotted cricket), are gaining attention as a high-protein food source; however, knowledge about how processing affects their allergenicity is limited. It was proven that tray drying led to a more pronounced reduction in the allergenicity of heat-labile proteins [106].
Fiocchi et al. [107] investigated the allergenicity of freeze-dried and homogenized beef in ten children with confirmed beef allergy, all of whom had positive skin prick tests (SPT) to both raw and heated beef and positive DBPCFC responses to 180 g of heated beef (5 min, 100°C). All children showed positive SPT reactions to untreated bovine serum albumin, and five reacted positively to it in DBPCFC. In contrast, only one child had a positive SPT to freeze-dried beef, and none reacted in DBPCFC. Homogenized beef also elicited no positive responses in either test. In a related study, the same group [106] reported weak allergenic potential of freeze-dried and homogenized beef, as well as freeze-dried sheep meat, in just two of 12 children with clinically confirmed beef allergy [88].
2.4. Non-Thermal Processing for Allergen Reduction
Non-thermal food processing technologies have gained considerable attention as promising strategies to reduce the allergenicity of food proteins without compromising nutritional value, sensory quality, or functional properties. Unlike traditional thermal treatments, non-thermal methods such as high-pressure processing (HPP), pulsed electric fields (PEF), cold plasma, ultrasound, and ultraviolet (UV) treatments offer more targeted approaches for modifying allergenic proteins. These techniques can disrupt protein structures, mask or cleave IgE-binding epitopes, and trigger biochemical changes that reduce immunoreactivity. As the prevalence of food allergies continues to rise globally, non-thermal interventions provide a compelling alternative for developing hypoallergenic food products while maintaining consumer acceptability and regulatory compliance (Table 2).
2.4.1. High Hydrostatic Pressure and Allergenicity in Foods
High hydrostatic pfressure (HHP) processing is a non-thermal food preservation method that typically subjects food products to pressures between 100 and 600 MPa, often at ambient or refrigerated temperatures. This treatment is particularly impactful on protein molecules, as it disrupts non-covalent interactions such as hydrogen bonds, ionic bonds, hydrophobic forces, and van der Waals interactions [108,109]. These interactions are fundamental to maintaining the secondary, tertiary, and quaternary structures of proteins. However, unlike thermal processing, HHP generally does not cleave covalent bonds such as peptide bonds or disulfide bridges unless extremely high pressures or extended treatment durations are applied [110]. As a result, while the primary amino acid sequence typically remains intact, HHP can lead to significant unfolding and denaturation of proteins, altering their structural and functional properties [111].
One of the most critical implications of HHP-induced protein denaturation is its effect on allergenicity. Food allergens are proteins or glycoproteins that possess immunologically active regions—known as epitopes—that are recognized by Immunoglobulin E (IgE) antibodies in sensitized individuals [112]. These epitopes can be either conformational, depending on the three-dimensional folding of the protein, or linear, comprising a continuous stretch of amino acids. HHP has the potential to disrupt these epitopes, especially conformational ones, by unfolding the protein structure and eliminating the spatial arrangements necessary for epitope recognition. Consequently, the IgE-binding capacity of many food allergens is reduced after HHP treatment [84].
In addition to epitope disruption, HHP can also cause protein aggregation. During unfolding, reactive groups become exposed, leading to intermolecular interactions and the formation of protein aggregates. This aggregation can result in the masking of linear epitopes or the creation of neoepitopes—new epitopic structures that may be more or less allergenic depending on the protein and processing conditions [113]. Furthermore, HHP-treated proteins often become more susceptible to proteolysis. The unfolding of the protein enhances accessibility for digestive enzymes such as pepsin and trypsin, promoting more effective breakdown during gastrointestinal digestion. This reduced stability and increased enzymatic degradation decrease the likelihood of allergenic epitopes surviving digestion and triggering immune responses [112].
The efficacy of HHP in reducing allergenicity is not uniform across all food proteins and is influenced by several factors, including pressure intensity, exposure time, temperature, pH, and the composition of the surrounding food matrix. For instance, pressures exceeding 400 MPa are generally required to significantly reduce allergenicity in robust proteins like β-lactoglobulin (from milk), tropomyosin (from shellfish), or ovomucoid (from egg) [114]. Additionally, the presence of other food components—such as lipids, carbohydrates, or salts—can stabilize protein structures or shield epitopes, thereby diminishing the effectiveness of HHP [115,116]. These matrix effects must be considered when designing HHP-based allergen control strategies.
HHP can lead to structural alterations in allergenic proteins, potentially reducing their immunoreactivity by destroying conformational epitopes. However, in some cases, HHP can expose hidden epitopes or modify existing ones, resulting in unchanged or even increased allergenicity. The outcome is dependent on the specific protein, the pressure level, and whether other treatments (e.g., heat) are combined with HHP. In the case of β-lactoglobulin (BLG), a major milk allergen, treatment at 600 MPa caused irreversible changes in secondary and tertiary structures, including the release of free thiol groups and surface hydrophobicity [117]. These structural changes enhanced the protein’s IgE-binding capacity, indicating increased allergenicity post-treatment.
Kato et al. [118] demonstrated that HHP, when combined with 8 M urea, significantly reduced rice allergen levels. The proposed mechanism involved structural damage to allergens caused by pressure, followed by enhanced extraction with urea to remove these proteins. Egg allergenicity was shown to decrease when pressure was applied in combination with heat. A study reported that the synergistic application of pressure and heat significantly reduced the allergenic response to hen’s egg, compared to either treatment alone [119]. HHP applied to soybean sprouts at 400 MPa resulted in significant reductions in the levels of Gly m 1, a key soybean allergen [120]. The treatment also reduced antigenicity of soybean protein isolates, with modifications in protein structure likely responsible for the lowered immunoreactivity. Overall, HHP is a promising tool for allergen mitigation in food systems. However, the effects are protein-specific and process-dependent. When combined with thermal treatments or other chemical agents, HHP may significantly reduce allergenicity, making it an important strategy in the development of hypoallergenic foods (Table 3).
2.4.2. Pulsed Electric Fields (PEF) Technology and Its Impact on Food Allergenicity
Pulsed electric field (PEF) technology represents a cutting-edge, non-thermal processing method that applies short bursts of high-voltage pulses—typically between 1 and 50 kV/cm—to biological materials such as tissues or fluids. This approach results in electroporation, a temporary increase in cell membrane permeability caused by dielectric breakdown, facilitating improved mass transfer and promoting the release of internal components such as proteins, lipids, and bioactive molecules [121,122,123,124]. PEF has gained broad acceptance across multiple domains within the food industry. Its applications range from microbial inactivation and juice extraction to drying pre-treatment, seed enhancement, detoxification of mycotoxins, and starch transformation [125,126,127,128]. Fundamentally, PEF’s mechanism involves inducing electroporation in cell membranes through high-intensity, short-duration pulses. These pulses lead to reversible or irreversible structural modifications in the membrane, thereby enhancing the transfer of substances and aiding in the recovery of intracellular compounds [123,124].
The performance of PEF systems is closely tied to the electrical behavior of cell membranes, which are largely composed of lipid bilayers acting as insulators. These membranes isolate the conductive environments inside and outside the cell, making each cell resemble a microscopic capacitor [122]. When placed in a conductive solution and exposed to an electric field, the cell disrupts the uniformity of the field, concentrating the intensity at the poles of the membrane aligned with the field [124].
A typical PEF setup includes a high-voltage pulse generator (ranging from 10 to 80 kV/cm), a treatment chamber with insulated electrodes, a cooling system, and a flow system for uniform exposure [122,125]. The system design is tailored through parameters like pulse duration, waveform, frequency, and electrode configuration. Pulse modes can be exponential decay, sinusoidal, monopolar, or bipolar. For instance, monopolar pulses have a constant polarity, while bipolar ones alternate it, minimizing polarization effects on electrodes. Different waveforms—such as square, decaying exponential, logarithmic, or oscillatory—offer varying benefits. Square waves, for example, ensure consistent membrane disruption, whereas decaying pulses can reduce energy use and thermal damage. The choice of pulse configuration is critical for achieving the desired effects—whether microbial inactivation, compound extraction, or textural modification—while preserving the nutritional and sensory quality of food products. Advanced control and safety systems are incorporated into industrial PEF equipment to ensure continuous, hygienic, and reliable processing [122,125,129,130]. Upon the application of the electric field, the cell—functioning as a dielectric sphere in a conductive environment—undergoes enhanced localized electric field intensity at its poles. These intensified regions are key sites for electroporation, often experiencing a field strength far exceeding the average applied to the bulk medium [124].
Although research on the influence of PEF on protein structure remains relatively limited, several studies have reported notable structural and functional changes depending on the protein type [131] and evidences suggest that PEF can disrupt the secondary and tertiary structures of proteins. These alterations are likely due to ionization of specific chemical groups and the disruption of electrostatic interactions [132,133]. Initially, protein molecules become polarized, leading to the exposure of hydrophobic amino acid residues to the surrounding solvent. This process subsequently results in protein unfolding and aggregation under the influence of high-intensity electric fields [134]. PEF processing has gained considerable interest in the food industry because of its energy efficiency, ability to preserve nutritional quality, and its suitability for liquid food matrices [135]. For instance, treatment of soybean protein isolates with PEF has demonstrated alterations in physicochemical properties, including denaturation and aggregation behaviors [136]. Similarly, investigations involving purified enzymes such as horseradish peroxidase and pectin esterase revealed a decline in enzymatic activity post-treatment, which was primarily attributed to conformational changes in protein structure [137]. However, it is important to consider that these changes may not result solely from the electric field itself; thermal effects due to associated Ohmic heating could also contribute to the observed outcomes [137]. The study conducted by Johnson et al. [138] explored the effects of PEF processing on the structural integrity of selected food allergens, including peanut allergens Ara h 2 and Ara h 6, and apple allergens Mal d 3 and Mal d 1b, all expressed heterologously. Structural changes were assessed using circular dichroism spectroscopy and gel-filtration chromatography. The findings showed that PEF treatment did not lead to any significant alterations in the secondary structure or aggregation state of the allergens. These results suggest that PEF is a structurally non-invasive food processing method with minimal impact on the conformation of purified food allergens.
A study investigating the effect of pulsed electric field (PEF) treatment at 25 kV/cm and 50 °C on Pru p 3, the major allergenic protein in peach, revealed that PEF induced structural denaturation of the protein, as determined by ELISA using rabbit IgG. However, the PEF treatment did not affect the IgE-binding capacity of Pru p 3, as shown by a competitive fluorescent immunoassay with sera from peach-allergic patients. Additionally, skin prick test results varied among individuals; more than 50% of participants exhibited increased skin reactivity following PEF treatment, indicating that patient-specific sensitization patterns may persist despite structural alterations in the allergen [139].
Experimental studies have demonstrated that high-intensity PEF treatments, particularly at 25–35 kV/cm for durations ranging from 60 to 180 µs, significantly impact the structural conformation of ovalbumin, including the α-helix content, thereby altering its immunogenic properties. The most substantial reduction in immunoreactivity was observed at 35 kV/cm for 180 µs [140], indicating PEF’s potential to lower the allergenicity of egg proteins.
In a molecular dynamics simulation study, Vanga et al. [141] explored the effects of oscillating and static electric fields (2450 MHz, 0.05 V/nm) at various temperatures (300, 380, and 425 K) on the structure of Ara h 6, a major peanut allergen. The simulations revealed significant conformational modifications under all tested conditions, implying potential impacts on the allergen’s functional properties. Subsequently, Vanga et al. [142] experimentally treated peanut flour with electric field intensities of 10, 15, and 20 kV for 60–180 minutes. Results indicated time-dependent increases in α-helix conformational changes, likely attributed to the emergence of new random coil structures and protein aggregates during processing.
However, not all findings confirm a consistent reduction in allergenicity. For instance, Johnson et al. [138] reported no significant structural changes in Ara h 2 and Ara h 6 (peanut 2S albumins), Mal d 1, and Mal d 3 (apple allergens) following PEF treatment at electric field strengths ranging from 0 to 35 kV/cm, with 2 Hz frequency and up to 130 kJ/kg energy input. Similarly, Paschke [143] found no notable reduction in celery allergen content when treated with a 10 kV PEF at 50 Hz.
A recent study investigated the modulation of ovalbumin (OVA) allergenicity using PEF treatment, highlighting structural changes linked to reduced immunoreactivity. PEF treatment at 6 kHz inhibited IgE and IgG1 binding by 30.41%, accompanied by visible microstructural surface cracks and unfolding of the protein’s secondary structure. Spectroscopic analyses revealed a blue shift in the amide I band, reductions in α-helix and β-sheet content, and conformational changes in disulfide bonds. Increased fluorescence intensity suggested exposure of hydrophobic residues such as tryptophan and tyrosine. Molecular dynamics simulations further confirmed reduced structural stability and hydrogen bonding. These findings suggest that PEF disrupts allergenic epitopes by altering protein conformation, supporting its potential application in developing hypoallergenic egg products [144]. In comparison to thermal and other non-thermal techniques, the impact of PEF on the structural modification and immunoreactivity of food allergens appears to be relatively limited. These inconsistent outcomes highlight the need for further optimization of PEF parameters to achieve reliable and reproducible allergen mitigation.
2.4.3. Pulsed Ultraviolet (PUV) Light and Its Impact on Food Allergenicity
Pulsed light (PL) technology utilizes a series of extremely brief, high-intensity bursts of broad-spectrum white light, composed primarily of ultraviolet (UV) light (54%), followed by visible light (26%) and infrared radiation (20%) [145]. The PUV light spans wavelengths from 200 nm to 1000 nm, allowing it to deliver intense light energy within a very short duration [146]. As a result, PUV light can achieve energy intensities up to a thousand times greater than those of traditional continuous UV systems (Shriver and Yang 2011). The technology utilizes xenon flash lamps to generate short-duration (typically <1 ms), high-energy light bursts at a frequency ranging from 1 to 20 Hz. These pulses result in rapid microbial inactivation via photochemical, photothermal, and photophysical mechanisms, primarily targeting nucleic acids and protein structures. In addition to microbial control, PUV has been investigated for its potential to alter food allergens, degrade toxins, and induce structural and functional changes in biomolecules such as proteins and lipids, without significantly elevating product temperature or compromising sensory and nutritional qualities [147,148,149,150].
In food applications, PUV light has been explored for its ability to modify the structural configuration of allergenic proteins. This includes altering conformational epitopes and promoting protein aggregation, which may reduce allergenicity [146,150]. However, post-treatment re-association of peptide fragments can lead to the development of neo-epitopes—new structures that could increase the allergenic potential of the food [151].
The functional mechanisms of PUV light inactivation can be grouped into photochemical, photothermal, and photophysical processes. These effects contribute to chemical transformations and modifications in protein structure, influenced by rapid heating and the intermittent delivery of high-energy pulses [146,152].
When applied to peanut samples—including raw, roasted, and peanut butter extracts—PUV light at distances of 10.8, 14.6, and 18.2 cm for 1–6 minutes caused a marked reduction in the intensity of allergenic proteins Ara h 1, Ara h 2, and Ara h 3, as evidenced by SDS-PAGE analysis. Increased energy doses (111.6 to 223.2 J/cm2) and longer exposure times further enhanced this reduction, while greater distance from the light source diminished the effect. ELISA results revealed that IgE binding decreased approximately 3-fold in raw peanut extracts and 7-fold in peanut butter slurry compared to untreated controls. These effects are attributed to changes in protein solubility and the formation of insoluble aggregates due to PUV treatment [153].
PUV light treatment of soy extracts for 2, 4, and 6 minutes led to a time-dependent decline in major allergens such as glycinin and β-conglycinin. Indirect ELISA using sera from soy-allergic individuals showed that IgE binding was reduced by 20%, 44%, and 50%, respectively. The observed reduction is likely associated with aggregation of allergenic proteins under PUV exposure [146]. PUV light also reduced the immunoreactivity of Gly m5 and Gly m6 allergens in soy when applied at a distance of 8–10 cm for up to 6 minutes. Gel bands corresponding to Gly m5 disappeared after just 2 minutes of treatment, while Gly m6 bands remained visible even after 6 minutes, indicating differential susceptibility to degradation or precipitation [154].
Similarly, ultraviolet-C (UV-C) treatment has shown promising effects in reducing the allergenic potential of milk proteins. Significant reductions in IgE binding were also observed in milk allergens including α-casein, α-lactalbumin, and β-lactoglobulin after 15 minutes of UV-C exposure. The decrease in immunoreactivity is likely due to alterations in discontinuous epitope structures. The enhanced effect in whey solutions has been linked to the greater pulse intensity and energy associated with PUV treatment [155]. The enhanced allergen reduction observed in whey proteins subjected to PUV treatment could be linked to the higher energy and pulse rate involved in the process [156].
For egg white proteins, ultraviolet exposure caused both aggregation and backbone cleavage; however, no substantial changes in immunoreactivity were observed compared to untreated controls. This suggests that the structural changes induced by UV light may not significantly affect allergenic epitopes in egg proteins [157].
Exposure of raw and boiled shrimp extracts (5 mg/mL) to pulsed ultraviolet (PUV) light at a pulse width of 360 µs, frequency of 3 pulses per second, and a distance of 10 cm led to a notable and irreversible reduction in allergenic reactivity. This reduction has been attributed to the formation of high molecular weight protein complexes, likely due to cross-linking between tropomyosin and other heat-sensitive proteins [145,146]. This effect was attributed to the formation of high molecular weight compounds via cross-linking between tropomyosin and other heat-sensitive proteins during the treatment [145].
Overall, PUV light has shown the capacity to lower allergenic potential in soy, peanut, milk, and shrimp products, though it appears ineffective for modifying egg allergens. Despite these promising results, especially for producing hypoallergenic food items, further investigations—including clinical and in vivo studies—are necessary to confirm its broader applicability in food processing [142].
However, not all studies report a reduction in allergenicity with UV processing. For instance, Manzocco et al. [157] found that although ultraviolet exposure led to aggregation and backbone cleavage in egg white proteins, these structural changes did not significantly affect their immunoreactivity compared to untreated samples, suggesting that the epitope structures remained largely unchanged.
Overall, the evidence suggests that PUV light may be effective in reducing allergenicity in foods such as soy, milk, shrimp, and peanuts, while it appears to have limited impact on egg allergens (Table 4). Although the technology shows potential for producing hypoallergenic food products, further investigation—particularly clinical and in vivo studies—is essential before broader adoption in the food industry [104].
2.4.4. Gamma Irradiation and Its Impact on Food Allergenicity
Radiation refers to the transfer or emission of energy through space or a material medium in the form of waves or particles. It is broadly classified into two types: nonionizing and ionizing radiation. Nonionizing radiation lacks the energy required to remove electrons from atoms or molecules and is typically considered non-harmful, causing little to no chemical changes. This category includes ultraviolet light, visible light, infrared radiation, microwaves, and radio waves—all of which are lower in energy and generally not used for food processing. In contrast, ionizing radiation carries sufficient energy to dislodge electrons, resulting in the formation of ions. This form includes energetic electromagnetic waves and subatomic particles capable of altering molecular structures. Among ionizing methods, gamma irradiation is considered one of the most straightforward techniques [164,165]. The reduction in immunoreactivity was linked to protein denaturation, assessed through parameters such as turbidity, surface hydrophobicity, and chromogenic reactivity. These findings suggest that coagulation of allergenic proteins may play a crucial role in minimizing allergenicity post-irradiation [166]. Irradiation of wheat germ agglutinin (WGA) was found to initially cause polypeptide chain fragmentation, followed by the formation of large, insoluble amorphous aggregates, ultimately leading to a decrease in allergenicity [167].
Radiation has proven to be an effective method for preserving food while maintaining its nutritional value and sensory qualities. It induces structural modifications in food proteins—including fragmentation, aggregation, cross-linking, and alterations to amino acids—that can influence their immunogenic properties [168]. These changes are primarily driven by reactive oxygen species formed during the radiolysis of water when proteins are irradiated in aqueous environments.
Exposure to ionizing radiation at doses of 10 and 50 kGy (applied at 10 °C with a rate of 10 kGy/h) altered the conformational epitopes in peanuts, leading to a significant reduction in IL-4 cytokine production by splenocytes from sensitized mice [169]. Similarly, Luo et al. [168] demonstrated that irradiation of purified peanut allergen Ara h 6 and whole peanut extract (at 1, 3, 5, and 10 kGy, 10 °C) resulted in a notable decline in IgG binding, as detected by ELISA, with higher radiation doses enhancing the effect. Interestingly, up to 5 kGy, the IgG response to whole peanut extract was greater than that of Ara h 6 alone, potentially due to the presence of other components in the extract that shielded Ara h 6 epitopes.
A comparable pattern was reported by Zhenxing et al. [170] in shrimp, where protein extracts irradiated at doses between 3 and 15 kGy (10 °C, 1 kGy/h) showed reduced IgE binding. However, intact shrimp muscle initially exhibited increased IgE reactivity up to 5 kGy, which then declined at higher doses.
A dose-dependent response to irradiation was observed in a study on cow milk allergy by Lee et al. [156], where IgE binding to isolated β-lactoglobulin (β-lg) increased up to a dose of 5 kGy. Beyond this level, protein agglomeration occurred, likely masking specific epitopes and resulting in reduced allergenic potential. In contrast, Kaddouri et al. [171] reported enhanced recognition of anti-β-lg IgG antibodies following irradiation (3–10 kGy at 13 Gy/min) of both liquid and freeze-dried cow milk and whey. These differing results may be attributed to the sample form—whether β-lg was isolated or present within a milk matrix—and the type of antibodies used in detection. These findings suggest that the allergenic response of purified proteins and whole food extracts to irradiation can vary, which holds relevance for food industry applications. Furthermore, differences in dose rate among studies indicate that the impact of varying dose rates at a constant radiation dose on protein allergenicity remains unclear.
In other research, irradiation of whole almonds, cashews, and walnuts did not result in structural changes to allergenic proteins or alter their allergenic potential [172]. Conversely, studies have shown that irradiation may increase the immune reactivity of gliadin and wheat flour [173]. These opposing results may stem from the physical state of the samples—solid versus solution—which can influence how effectively irradiation reduces allergenicity. Overall, applying irradiation to food allergens in liquid form appears to be the most promising approach for minimizing their immunoreactivity.
Seo et al. [174] observed a decrease in the presence of the egg allergen ovalbumin when exposed to gamma radiation using cobalt-60 at a dose of 100 kGy, while treatment at 10 kGy showed no significant impact. This reduction was believed to result from alterations in the protein’s molecular weight. Gamma irradiation is known to promote protein crosslinking—such as disulfide bond formation—and enhance hydrophobic interactions, both of which can lead to protein aggregation [175]. However, a 100 kGy dose exceeds levels typically considered safe for food processing, as doses up to around 10 kGy are generally recognized as safe for consumption [176]. In a separate study, Seo et al. [177] also reported reduced ovalbumin immunoreactivity in white cake samples irradiated at 10–20 kGy.
In studies involving crustacean allergens, such as shrimp tropomyosin, gamma irradiation at doses of 7 kGy and higher resulted in the disappearance of the characteristic 36 kDa tropomyosin band on SDS–PAGE, indicating structural degradation. Additionally, IgE binding was significantly diminished at the highest doses tested [178]. Multiple studies have demonstrated the effectiveness of gamma irradiation in modifying allergenic proteins. For instance, Seo et al. [174] reported that ovalbumin, a principal allergen. Gamma irradiation can enhance protein crosslinking through the formation of disulfide bonds and intensified hydrophobic interactions, which promote aggregation and possible loss of immunoreactive epitopes (Table 5) [166].
2.4.5. High-Intensity Ultrasound and Its Impact on Food Allergenicity
Ultrasound technology has gained considerable attention in food processing due to its non-thermal and energy-efficient nature. It involves the application of sound waves with frequencies above the range of human hearing, typically between 20 kHz and 100 kHz in food systems [180]. When ultrasound is applied at high intensity, it induces a phenomenon known as acoustic cavitation—the formation, growth, and implosive collapse of microbubbles within a liquid medium [181]. The core mechanism involves cavitation, wherein alternating compression and rarefaction cycles form microbubbles that collapse violently, generating localized temperatures up to 5000 K and pressures reaching 1000 atm [182]. These extreme conditions facilitate protein denaturation, potentially altering allergenic structures and epitopes. This collapse generates localized hot spots with extremely high temperatures (up to 5000 K) and pressures (up to 1000 atm), although these effects are confined to the microscopic scale and last for only a few microseconds [183]. Alongside thermal effects, cavitation leads to intense shear forces, microjetting, turbulence, and the production of free radicals (e.g., hydroxyl radicals from water sonolysis), all of which can contribute to physical and chemical transformations in food matrices [184].
High-intensity ultrasound (HIU) has been explored for various food processing operations, including peeling of fruits and vegetables [185], reduction of oil content and shelf-life extension of fried products [186], and shortening of parboiling time in rice processing [187]. More recently, its potential to alter protein structure and reduce food allergenicity has attracted scientific interest. The primary mechanism by which HIU affects allergens lies in its capacity to alter the three-dimensional structure of allergenic proteins. During sonication, proteins can undergo unfolding, aggregation, or fragmentation due to mechanical stress and localized heating, leading to changes in epitope exposure and antigenicity. Additionally, free radicals generated during cavitation may oxidize specific amino acid residues, contributing further to conformational changes [58,166].
In a study by Li et al. [187], shrimp samples were subjected to high-intensity ultrasound at a frequency of 30 kHz for durations ranging from 130 to 180 minutes. This treatment significantly reduced IgE-binding capacity in both isolated tropomyosin (the major shrimp allergen) and in crude shrimp protein extracts. Specifically, IgE-binding to the isolated tropomyosin was reduced by 81.3–88.5%, whereas binding in the shrimp extract decreased by approximately 68.9%, based on ELISA assays. Immunoblotting also revealed that prolonged ultrasound exposure led to the formation of new, lower-molecular-weight protein bands, suggesting fragmentation of the allergenic protein. This structural disruption likely contributed to the observed decrease in allergenic reactivity. Interestingly, the treatment duration was relatively long, yet no significant temperature rise or quality deterioration of the shrimp product was reported. This highlights one of the advantages of HIU as a non-thermal method for allergen mitigation. Zhenxing et al. [170] further confirmed that shrimp allergenicity decreased more significantly at elevated temperatures (50 °C) during ultrasound treatment. In another study, Zhang et al. [188] subjected shrimp tropomyosin (TM) to 15 minutes of ultrasound (100–800 W), reporting a substantial degradation of TM and associated reduction in allergenicity, as evidenced by ELISA and immunoblotting using sera from allergic patients. However, some shellfish allergens appear more resistant to ultrasound. Chen et al. [189] found minimal degradation and unchanged IgE-binding activity of arginine kinase (AK) in crayfish even after ultrasound treatment at 200 W and 30 °C for up to 180 minutes. Dairy proteins also showed limited response; Tammineedi et al. [155] observed no significant changes in SDS-PAGE profiles of α-casein, β-lactoglobulin, and α-lactalbumin after treatment with 500 W ultrasound at 20 kHz for up to 30 minutes. Similar findings were reported in other milk allergen studies [155,190].
In soy protein, Tammineedi et al. [155] demonstrated a 24% decrease in immunoreactivity following treatment with high-intensity ultrasound at 37 kHz for 10 minutes. The observed reduction is attributed to alterations in secondary and tertiary structures. Similarly, significant allergen reduction has been observed in roasted peanuts; Li et al. [187] reported decreases of 84.8% in Ara h 1 and 4.88% in Ara h 2 after exposure to 50 Hz ultrasound for two hours. Structural modifications in the α-helical IgE-binding regions of these proteins are believed to account for this change [191,192]. Zhenxing et al. [170] further confirmed that shrimp allergenicity decreased more significantly at elevated temperatures (50 °C) during ultrasound treatment. In another study, Zhang, He et al. (2018b) subjected shrimp tropomyosin (TM) to 15 minutes of ultrasound (100–800 W), reporting a substantial degradation of TM and associated reduction in allergenicity, as evidenced by ELISA and immunoblotting using sera from allergic patients.
The efficiency of ultrasound appears to increase when combined with thermal treatment, suggesting a synergistic effect. Ultrasound processing offers several benefits, including reduced energy consumption, minimal chemical usage, enhanced mass transfer, and better retention of sensory qualities [193]. Given these advantages and its proven potential in reducing immunoreactivity in soy, peanuts, and shellfish, ultrasound is a compelling alternative to conventional processing methods. Nevertheless, optimization of key parameters—such as frequency, treatment duration, and temperature—is essential to maximize its efficacy in allergen reduction across various food types. Overall, high-intensity ultrasound offers a promising approach to reduce the allergenic potential of food proteins through a combination of physical and chemical effects (Table 6). While more data is needed to understand optimal parameters and food matrix-specific responses, current evidence supports its utility as part of an integrated strategy for allergen control in food systems [165].
2.4.6. Cold Plasma and Its Impact on Food Allergenicity
Cold plasma is primarily used for microbial inactivation and decontamination of food due to its lower energy consumption and reduced temperature requirements compared to traditional methods [199]. Ionized gas, often referred to as the fourth state of matter, consists of a mixture of reactive species such as UV photons, ions, electrons, free radicals, molecules, and excited atoms. These components are formed through the excitation and ionization of gases and are capable of interacting with proteins, resulting in conformational changes. Since most food allergens are proteins, it is hypothesized that cold plasma treatment could similarly alter their structures through these reactive agents [165,199]. Multiple studies have shown the potential of plasma treatment in reducing food allergenicity. For example, Venkataratnam et al. [200] applied cold atmospheric plasma (80 kV) for different durations (0 to 60 minutes) to dry, defatted peanut flour and whole peanuts. Competitive ELISA using bovine serum albumin showed up to a 43% reduction in antigenicity, accompanied by secondary structure changes detected through circular dichroism. In another investigation, direct dielectric discharge cold plasma treatment for 5 minutes at 30 kV and 60 Hz led to a 76% decrease in shrimp allergen tropomyosin’s immunoreactivity [166].
In one study, the impact of both direct and remote cold atmospheric pressure plasma (CAPP) on the immunoreactivity of soy protein isolate was examined [165]. Researchers measured factors such as sample weight, surface temperature, pH, and hydrogen peroxide content. SDS-PAGE analysis revealed a noticeable reduction in the intensity of protein bands corresponding to the primary soy allergens β-conglycinin (Gly m5) and glycinin (Gly m6). The most substantial decrease in immunoreactivity—ranging from 91% to 100%—was observed in the soluble protein fraction following direct CAPP treatment. Remote CAPP exposure also reduced immunoreactivity, though to a slightly lesser extent, with reductions of up to 89%. These reductions in immunoreactivity are attributed to the disruption or masking of conformational epitopes by free radicals produced during plasma exposure, which hinders IgE antibody recognition [201]. Meinlschmidt et al. [154] reported that CAPP treatment of soy protein extracts for up to 10 minutes at 9, 10, and 11 kVpp (3.0 kHz) nearly eliminated Gly m5 allergen reactivity. This was likely due to alterations in conformational and linear epitopes or destruction of antibody binding sites, as shown by sandwich ELISA using human sera and monoclonal antibodies.
On the other hand, Tammineedi et al. [155] treated milk a-casein with cold atmospheric plasma at 13.56 MHz radiofrequency and 30.7 L/min argon gas flow for 5, 10, and 15 minutes. They found no significant reduction in immunoreactivity, possibly due to limited plasma exposure or insufficient treatment intensity. These varied findings across food types suggest that cold plasma’s effectiveness in allergen reduction is not uniform and depends on factors such as sample composition and processing conditions. To date, reductions in allergen levels have been observed in soy, peanuts, shrimp, and wheat after cold plasma treatment, while no notable changes have been reported in milk proteins. Furthermore, concerns remain regarding the quality degradation associated with this method. Several studies have linked cold plasma processing to undesirable effects, including accelerated lipid oxidation, nutrient loss (such as vitamins), and compromised sensory qualities [202,203,204]. Therefore, while cold plasma holds potential, its limitations highlight the need for continued research and the development of alternative food processing technologies.
2.4.7. Genetic Modification and Its Role in Reducing Allergenicity
Genetic modification (GM) refers to the deliberate alteration of an organism’s genetic material using biotechnology, allowing for the insertion, deletion, or silencing of specific genes to achieve desired traits. In the context of food production, GM is employed to enhance crop yield, resistance to pests or diseases, and improve nutritional or functional characteristics. Over recent years, its potential to reduce food allergenicity has become a growing area of research, although concerns regarding safety, stability, and consumer perception remain significant. One strategy to reduce allergenicity in foods through GM involves silencing the expression of specific allergenic proteins by employing post-transcriptional gene silencing or co-suppression mechanisms. This approach aims to prevent the production of allergenic proteins during gene expression. However, uncertainties persist regarding the long-term stability of these modifications. Incomplete suppression or loss of gene silencing could reintroduce allergens, posing potential health risks to sensitized individuals. Additionally, removing key proteins may affect the structural and functional integrity of food products, as many allergenic proteins play vital roles in the metabolism and development of the source organism [79].
Public skepticism also surrounds the possibility that modifying protein structures may inadvertently result in the emergence of new allergenic determinants or epitopes that the immune system may fail to recognize as harmless. Therefore, a more targeted strategy—such as altering specific IgE-binding epitopes while maintaining the overall structure and functionality of the protein—may offer a safer and more effective route [205].
Experimental studies provide insights into these strategies. For instance, Rupa et al. [206] modified the IgE-binding sites of ovomucoid, a major egg allergen, by introducing glycosylation at known epitope locations. This post-translational modification significantly reduced IgE-mediated immune responses in a mouse model, demonstrating the feasibility of epitope-focused interventions. However, since the modified proteins were expressed in yeast cells rather than in the natural egg matrix, it remains unclear how such changes would impact the final food product. In a study targeting peanut allergens, Chu et al. [207] applied RNA interference to suppress the expression of Ara h 2 and Ara h 6, two major peanut allergens. Although this intervention significantly decreased IgE binding to these allergens, the whole peanut extract still retained immunoreactivity, likely due to the presence of other allergenic proteins. Interestingly, the silencing of these genes did not lead to any noticeable morphological changes in the peanut plants. Similarly, Dodo et al. [56] reported reduced IgE binding following genetic silencing of Ara h 2. In another example, Herman et al. [208] demonstrated that silencing Gly m Bd 30, a key soybean allergen, nearly eliminated IgE binding. Detailed protein analyses and microscopic evaluations confirmed that the modified soybeans were structurally and compositionally similar to their non-GM counterparts and did not produce any new allergens. Although these findings highlight the potential of GM approaches to reduce allergenicity (Table 7), broader questions concerning consumer acceptance, regulatory approval, and long-term safety must still be addressed.
3. Conclusions
The rising prevalence of food allergies necessitates the development of effective mitigation strategies that ensure both consumer safety and food quality. A growing body of evidence highlights the complex interplay between the molecular characteristics of food allergens, processing conditions, and the surrounding food matrix in determining allergenic potential. Different processing technologies—thermal and non-thermal—induce distinct physicochemical modifications in food proteins, thereby influencing their digestibility, bioavailability, and IgE-binding capacity.
Thermal treatments such as moist heat generally reduce allergenicity by disrupting protein conformation and enhancing proteolytic susceptibility, while dry heat methods (e.g., roasting and baking) may increase allergenicity through the formation of neo-epitopes via Maillard reactions. Non-thermal technologies, including HPP, PEF, cold plasma, PUV, US, and gamma irradiation, offer promising alternatives for allergen mitigation by inducing conformational changes without compromising nutritional and sensory attributes. Additionally, microbial fermentation and enzymatic hydrolysis can effectively target linear epitopes, further reducing allergenic potential.
Despite these advances, translating reductions in IgE reactivity observed in vitro to clinically meaningful outcomes remain a significant challenge. The variability in digestive stability, structural resilience of allergenic proteins, and individual immune responses necessitates the use of complementary in vivo assessments such as skin prick tests, oral food challenges, and mediator release assays to validate hypoallergenic claims. Furthermore, a comprehensive understanding of both conformational and linear epitope modifications is essential for the design of processing strategies that minimize elicitation thresholds without compromising safety.
Future research should focus on standardizing allergenicity assessment protocols, integrating multi-hurdle approaches for simultaneous allergen and microbial risk reduction, and expanding studies to encompass emerging alternative proteins. Ultimately, a multidisciplinary approach that bridges food processing technology, immunology, and clinical validation is crucial for developing safe, effective, and industry-relevant solutions to mitigate food allergenicity and improve public health outcomes.
Author Contributions
Gulsun Akdemir Evrendilek: Literature review, writing original manuscript; Alper Güven: writing manuscript, formatting.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Sicherer, S.H. Clinical Implications of Cross-Reactive Food Allergens. Journal of Allergy and Clinical Immunology 2001, 108, 881–890. [Google Scholar] [CrossRef]
- Sicherer, S.H.; Sampson, H.A. Food Allergy: Epidemiology, Pathogenesis, Diagnosis, and Treatment. Journal of Allergy and Clinical Immunology 2014, 133, 291–307.e5. [Google Scholar] [CrossRef]
- Renz, H.; Allen, K.J.; Sicherer, S.H.; Sampson, H.A.; Lack, G.; Beyer, K.; Oettgen, H.C. Food Allergy. Nat Rev Dis Primers 2018, 4, 17098. [Google Scholar] [CrossRef]
- Valenta, R.; Hochwallner, H.; Linhart, B.; Pahr, S. Food Allergies: The Basics. Gastroenterology 2015, 148, 1120–1131.e4. [Google Scholar] [CrossRef] [PubMed]
- Breiteneder, H.; Ebner, C. Molecular and Biochemical Classification of Plant-Derived Food Allergens. Journal of Allergy and Clinical Immunology 2000, 106, 27–36. [Google Scholar] [CrossRef]
- Breiteneder, H.; Mills, E.N.C. Molecular Properties of Food Allergens. Journal of Allergy and Clinical Immunology 2005, 115, 14–23. [Google Scholar] [CrossRef]
- Chehade, M.; Mayer, L. Oral Tolerance and Its Relation to Food Hypersensitivities. Journal of Allergy and Clinical Immunology 2005, 115, 3–12. [Google Scholar] [CrossRef]
- Hsieh, K.-Y.; Tsai, C.-C.; Herbert Wu, C.H.; Lin, R.-H. Epicutaneous Exposure to Protein Antigen and Food Allergy. Clinical & Experimental Allergy 2003, 33, 1067–1075. [Google Scholar] [CrossRef]
- Lack, G. Update on Risk Factors for Food Allergy. Journal of Allergy and Clinical Immunology 2012, 129, 1187–1197. [Google Scholar] [CrossRef] [PubMed]
- Mittag, D.; Akkerdaas, J.; Ballmer-Weber, B.K.; Vogel, L.; Wensing, M.; Becker, W.-M.; Koppelman, S.J.; Knulst, A.C.; Helbling, A.; Hefle, S.L.; et al. Ara h 8, a Bet v 1–Homologous Allergen from Peanut, Is a Major Allergen in Patients with Combined Birch Pollen and Peanut Allergy. Journal of Allergy and Clinical Immunology 2004, 114, 1410–1417. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, S.; Crowe, S.E. Gastrointestinal Food Allergy: New Insights into Pathophysiology and Clinical Perspectives. Gastroenterology 2005, 128, 1089–1113. [Google Scholar] [CrossRef]
- Sampson, H.A.; Sicherer, S.H.; Birnbaum, A.H. AGA Technical Review on the Evaluation of Food Allergy in Gastrointestinal Disorders. Gastroenterology 2001, 120, 1026–1040. [Google Scholar] [CrossRef] [PubMed]
- Sampson, H.A.; Anderson, J.A. Summary and Recommendations: Classification of Gastrointestinal Manifestations Due to Immunologic Reactions to Foods in Infants and Young Children. J Pediatr Gastroenterol Nutr 2000, 30 Suppl, S87–94. [Google Scholar] [CrossRef]
- Yocum, M.W.; Butterfield, J.H.; Klein, J.S.; Volcheck, G.W.; Schroeder, D.R.; Silverstein, M.D. Epidemiology of Anaphylaxis in Olmsted County: A Population-Based Study. Journal of Allergy and Clinical Immunology 1999, 104, 452–456. [Google Scholar] [CrossRef]
- Bock, S.A.; Muñoz-Furlong, A.; Sampson, H.A. Fatalities Due to Anaphylactic Reactions to Foods. Journal of Allergy and Clinical Immunology 2001, 107, 191–193. [Google Scholar] [CrossRef]
- Badina, L.; Barbi, E.; Berti, I.; Radillo, O.; Matarazzo, L.; Ventura, A.; Longo, G. The Dietary Paradox in Food Allergy: Yesterday’s Mistakes, Today’s Evidence and Lessons for Tomorrow. Current Pharmaceutical Design 2012, 18, 5782–5787. [Google Scholar] [CrossRef] [PubMed]
- de Silva, R.; Dasanayake, W.M.D.K.; Wickramasinhe, G.D.; Karunatilake, C.; Weerasinghe, N.; Gunasekera, P.; Malavige, G.N. Sensitization to Bovine Serum Albumin as a Possible Cause of Allergic Reactions to Vaccines. Vaccine 2017, 35, 1494–1500. [Google Scholar] [CrossRef] [PubMed]
- Boyce, J.A.; Assa’ad, A.; Burks, A.W.; Jones, S.M.; Sampson, H.A.; Wood, R.A.; Plaut, M.; Cooper, S.F.; Fenton, M.J.; Arshad, S.H.; et al. Guidelines for the Diagnosis and Management of Food Allergy in the United States: Summary of the NIAID-Sponsored Expert Panel Report. Journal of Pediatric Nursing 2011, 26, e2–e17. [Google Scholar] [CrossRef]
- Finkelman, F.D.; Boyce, J.A.; Vercelli, D.; Rothenberg, M.E. Key Advances in Mechanisms of Asthma, Allergy, and Immunology in 2009. Journal of Allergy and Clinical Immunology 2010, 125, 312–318. [Google Scholar] [CrossRef]
- Silverstein, A.M. Clemens Freiherr von Pirquet: Explaining Immune Complex Disease in 1906. Nat Immunol 2000, 1, 453–455. [Google Scholar] [CrossRef]
- Turk, J.L. Von Pirquet, Allergy and Infectious Diseases: A Review. J R Soc Med 1987, 80, 31–33. [Google Scholar] [CrossRef]
- Palmiere, C.; Comment, L.; Mangin, P. Allergic Reactions Following Contrast Material Administration: Nomenclature, Classification, and Mechanisms. Int J Legal Med 2014, 128, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Kagan, R.S. Food Allergy: An Overview. Environmental Health Perspectives 2003, 111, 223–225. [Google Scholar] [CrossRef] [PubMed]
- Molina, M.A.F.; Kram, Y.E.; Lanser, B.J. The Global Burden of Food Allergy. Immunology and Allergy Clinics 2025, 45, 325–337. [Google Scholar] [CrossRef]
- Stapel, S.O.; Kleine-Tebbe, J. Allergy Testing in the Laboratory. In Allergy and Allergic Diseases; John Wiley & Sons, Ltd., 2008; pp. 1346–1367 ISBN 978-1-4443-0091-8.
- Schuppan, D.; Junker, Y.; Barisani, D. Celiac Disease: From Pathogenesis to Novel Therapies. Gastroenterology 2009, 137, 1912–1933. [Google Scholar] [CrossRef]
- Caubet, J.-C.; Ponvert, C. Vaccine Allergy. Immunology and Allergy Clinics 2014, 34, 597–613. [Google Scholar] [CrossRef]
- Junker, Y.; Zeissig, S.; Kim, S.-J.; Barisani, D.; Wieser, H.; Leffler, D.A.; Zevallos, V.; Libermann, T.A.; Dillon, S.; Freitag, T.L.; et al. Wheat Amylase Trypsin Inhibitors Drive Intestinal Inflammation via Activation of Toll-like Receptor 4. J Exp Med 2012, 209, 2395–2408. [Google Scholar] [CrossRef]
- Catassi, C.; Bai, J.C.; Bonaz, B.; Bouma, G.; Calabrò, A.; Carroccio, A.; Castillejo, G.; Ciacci, C.; Cristofori, F.; Dolinsek, J.; et al. Non-Celiac Gluten Sensitivity: The New Frontier of Gluten Related Disorders. Nutrients 2013, 5, 3839–3853. [Google Scholar] [CrossRef] [PubMed]
- Ruiter, B.; Shreffler, W.G. The Role of Dendritic Cells in Food Allergy. Journal of Allergy and Clinical Immunology 2012, 129, 921–928. [Google Scholar] [CrossRef]
- Badina, L.; Barbi, E.; Berti, I.; Radillo, O.; Matarazzo, L.; Ventura, A.; Longo, G. The Dietary Paradox in Food Allergy: Yesterday’s Mistakes, Today’s Evidence and Lessons for Tomorrow. Current Pharmaceutical Design 2012, 18, 5782–5787. [Google Scholar] [CrossRef]
- Burney, P.; Jarvis, D.; Perez-Padilla, R. The Global Burden of Chronic Respiratory Disease in Adults. The International Journal of Tuberculosis and Lung Disease 2015, 19, 10–20. [Google Scholar] [CrossRef]
- Baumann, S.; Lorentz, A. Obesity - A Promoter of Allergy? Int Arch Allergy Immunol 2013, 162, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Gray, C.L.; Levin, M.E. Epidemiology of Food Allergy: Review Article. Current Allergy & Clinical Immunology 2014, 27, 170–176. [Google Scholar]
- Marrs, T.; Bruce, K.D.; Logan, K.; Rivett, D.W.; Perkin, M.R.; Lack, G.; Flohr, C. Is There an Association between Microbial Exposure and Food Allergy? A Systematic Review. Pediatric Allergy and Immunology 2013, 24, 311–320.e8. [Google Scholar] [CrossRef]
- Martino, D.; Neeland, M.; Dang, T.; Cobb, J.; Ellis, J.; Barnett, A.; Tang, M.; Vuillermin, P.; Allen, K.; Saffery, R. Epigenetic Dysregulation of Naive CD4+ T-Cell Activation Genes in Childhood Food Allergy. Nat Commun 2018, 9, 3308. [Google Scholar] [CrossRef]
- Strachan, D.P. Hay Fever, Hygiene, and Household Size. BMJ 1989, 299, 1259–1260. [Google Scholar] [CrossRef]
- Alm, B.; Åberg, N.; Erdes, L.; Möllborg, P.; Pettersson, R.; Norvenius, S.G.; Goksör, E.; Wennergren, G. Early Introduction of Fish Decreases the Risk of Eczema in Infants. Archives of Disease in Childhood 2009, 94, 11–15. [Google Scholar] [CrossRef] [PubMed]
- McGeady, S.J. Immunocompetence and Allergy. Pediatrics 2004, 113, 1107–1113. [Google Scholar] [CrossRef]
- Thorburn, A.N.; Macia, L.; Mackay, C.R. Diet, Metabolites, and “Western-Lifestyle” Inflammatory Diseases. Immunity 2014, 40, 833–842. [Google Scholar] [CrossRef]
- Valenta, R. The Future of Antigen-Specific Immunotherapy of Allergy. Nat Rev Immunol 2002, 2, 446–453. [Google Scholar] [CrossRef]
- Han, Y.; Kim, J.; Ahn, K. Food Allergy. Korean J Pediatr 2012, 55, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Ebner, C. Allergene. In Allergologie; Heppt, W., Renz, H., Röcken, M., Eds.; Springer: Berlin, Heidelberg, 1998; ISBN 978-3-662-05660-8. [Google Scholar]
- Ebner, C.; Hoffmann-Sommergruber, K.; Breiteneder, H. Plant Food Allergens Homologous to Pathogenesis-Related Proteins. Allergy 2001, 56, 43–44. [Google Scholar] [CrossRef]
- Ortolani, C.; Ispano, M.; Ansaloni, R.; Rotondo, F.; Incorvaia, C.; Pastorello, E.A. Diagnostic Problems Due to Cross-reactions in Food Allergy. Allergy 1998, 53, 58–61. [Google Scholar] [CrossRef]
- Ortolani, C.; Pastorello, E.A. Food Allergies and Food Intolerances. Best Practice & Research Clinical Gastroenterology 2006, 20, 467–483. [Google Scholar] [CrossRef]
- Asero, R.; Antonicelli, L. Does Sensitization to Foods in Adults Occur Always in the Gut? Int Arch Allergy Immunol 2011, 154, 6–14. [Google Scholar] [CrossRef]
- Bartnikas, L.M.; Phipatanakul, W. Turning Up the Heat on Skin Testing for Baked Egg Allergy. Clin Exp Allergy 2013, 43, 1095–1096. [Google Scholar] [CrossRef]
- Noti, M.; Kim, B.S.; Siracusa, M.C.; Rak, G.D.; Kubo, M.; Moghaddam, A.E.; Sattentau, Q.A.; Comeau, M.R.; Spergel, J.M.; Artis, D. Exposure to Food Allergens through Inflamed Skin Promotes Intestinal Food Allergy through the Thymic Stromal Lymphopoietin–Basophil Axis. Journal of Allergy and Clinical Immunology 2014, 133, 1390–1399.e6. [Google Scholar] [CrossRef] [PubMed]
- Astwood, J.D.; Leach, J.N.; Fuchs, R.L. Stability of Food Allergens to Digestion in Vitro. Nat Biotechnol 1996, 14, 1269–1273. [Google Scholar] [CrossRef]
- Groschwitz, K.R.; Hogan, S.P. Intestinal Barrier Function: Molecular Regulation and Disease Pathogenesis. Journal of Allergy and Clinical Immunology 2009, 124, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Irvine, A.D.; McLean, W.H.I.; Leung, D.Y.M. Filaggrin Mutations Associated with Skin and Allergic Diseases. New England Journal of Medicine 2011, 365, 1315–1327. [Google Scholar] [CrossRef]
- Perrier, C.; Corthésy, B. Gut Permeability and Food Allergies. Clinical & Experimental Allergy 2011, 41, 20–28. [Google Scholar] [CrossRef]
- Karlsson, M.R.; Johansen, F.-E.; Kahu, H.; Macpherson, A.; Brandtzaeg, P. Hypersensitivity and Oral Tolerance in the Absence of a Secretory Immune System. Allergy 2010, 65, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Thyssen, J.P.; Linneberg, A.; Menné, T.; Johansen, J.D. The Epidemiology of Contact Allergy in the General Population – Prevalence and Main Findings. Contact Dermatitis 2007, 57, 287–299. [Google Scholar] [CrossRef] [PubMed]
- Dodo, H.W.; Konan, K.N.; Chen, F.C.; Egnin, M.; Viquez, O.M. Alleviating Peanut Allergy Using Genetic Engineering: The Silencing of the Immunodominant Allergen Ara h 2 Leads to Its Significant Reduction and a Decrease in Peanut Allergenicity. Plant Biotechnology Journal 2008, 6, 135–145. [Google Scholar] [CrossRef]
- Mills, E.N.C.; Sancho, A.I.; Rigby, N.M.; Jenkins, J.A.; Mackie, A.R. Impact of Food Processing on the Structural and Allergenic Properties of Food Allergens. Molecular Nutrition & Food Research 2009, 53, 963–969. [Google Scholar] [CrossRef]
- Rahaman, T.; Vasiljevic, T.; Ramchandran, L. Effect of Processing on Conformational Changes of Food Proteins Related to Allergenicity. Trends in Food Science & Technology 2016, 49, 24–34. [Google Scholar] [CrossRef]
- Sen, M.; Kopper, R.; Pons, L.; Abraham, E.C.; Burks, A.W.; Bannon, G.A. Protein Structure Plays a Critical Role in Peanut Allergen Stability and May Determine Immunodominant IgE-Binding Epitopes. J Immunol 2002, 169, 882–887. [Google Scholar] [CrossRef]
- Taylor, S.L. Food Allergies and Other Food Sensitivities. 2001, 55. 55.
- Crevel, R.W.R. 3 - Food Allergen Risk Assessment and Management. In Handbook of Food Allergen Detection and Control; Flanagan, S., Ed.; Woodhead Publishing Series in Food Science, Technology and Nutrition; Woodhead Publishing, 2015; pp. 41–66 ISBN 978-1-78242-012-5.
- Crevel, R.W.R.; Baumert, J.L.; Baka, A.; Houben, G.F.; Knulst, A.C.; Kruizinga, A.G.; Luccioli, S.; Taylor, S.L.; Madsen, C.B. Development and Evolution of Risk Assessment for Food Allergens. Food and Chemical Toxicology 2014, 67, 262–276. [Google Scholar] [CrossRef]
- Crevel, R.W.R.; Baumert, J.L.; Luccioli, S.; Baka, A.; Hattersley, S.; Hourihane, J.O.; Ronsmans, S.; Timmermans, F.; Ward, R.; Chung, Y. Translating Reference Doses into Allergen Management Practice: Challenges for Stakeholders. Food and Chemical Toxicology 2014, 67, 277–287. [Google Scholar] [CrossRef]
- Petersen, K.D.; Kronborg, C.; Larsen, J.N.; Dahl, R.; Gyrd-Hansen, D. Patient Related Outcomes in a Real Life Prospective Follow up Study: Allergen Immunotherapy Increase Quality of Life and Reduce Sick Days. World Allergy Organ J 2013, 6, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Arora, R.; Kumar, A.; Singh, I.K.; Singh, A. Pathogenesis Related Proteins: A Defensin for Plants but an Allergen for Humans. International Journal of Biological Macromolecules 2020, 157, 659–672. [Google Scholar] [CrossRef]
- Basant, N.; Jain, K.; Mittal, N.; Pandey, S.; Srivastava, V.; Pandey, K. ; Ayushi; Sharma, S.; Tiwari, N. Food Allergies and Their Management. In Traditional Foods; CRC Press, 2025 ISBN 978-1-00-351424-4.
- Cabanillas, B.; Novak, N. Effects of Daily Food Processing on Allergenicity. Critical Reviews in Food Science and Nutrition 2019, 59, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Sathe, S.K.; Teuber, S.S.; Roux, K.H. Effects of Food Processing on the Stability of Food Allergens. Biotechnology Advances 2005, 23, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Sathe, S.K.; Sharma, G.M. Effects of Food Processing on Food Allergens. Molecular Nutrition & Food Research 2009, 53, 970–978. [Google Scholar] [CrossRef]
- Kroghsbo, S.; Bøgh, K.L.; Rigby, N.M.; Mills, E.N.C.; Rogers, A.; Madsen, C.B. Sensitization with 7S Globulins from Peanut, Hazelnut, Soy or Pea Induces IgE with Different Biological Activities Which Are Modified by Soy Tolerance. Int Arch Allergy Immunol 2011, 155, 212–224. [Google Scholar] [CrossRef]
- Madsen, J.L.; Kroghsbo, S.; Madsen, C.B.; Pozdnyakova, I.; Barkholt, V.; Bøgh, K.L. The Impact of Structural Integrity and Route of Administration on the Antibody Specificity against Three Cow’s Milk Allergens - a Study in Brown Norway Rats. Clin Transl Allergy 2014, 4, 25. [Google Scholar] [CrossRef]
- An, S.B.; Yang, B.-G.; Jang, G.; Kim, D.-Y.; Kim, J.; Oh, S.-M.; Oh, N.; Lee, S.; Moon, J.-Y.; Kim, J.-A.; et al. Combined IgE Neutralization and Bifidobacterium Longum Supplementation Reduces the Allergic Response in Models of Food Allergy. Nat Commun 2022, 13, 5669. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.N.; Dupre, R.A.; Ebmeier, C.C.; Patil, S.; Smith, B.; Mattison, C.P. Heating Differentiates Pecan Allergen Stability: Car i 4 Is More Heat Labile Than Car i 1 and Car i 2. Food Science & Nutrition 2025, 13, e4747. [Google Scholar] [CrossRef]
- Garcia-Calvo, E.; García-García, A.; Madrid, R.; Martin, R.; García, T. From Polyclonal Sera to Recombinant Antibodies: A Review of Immunological Detection of Gluten in Foodstuff. Foods 2021, 10, 66. [Google Scholar] [CrossRef]
- Spies, J.R. Allergens. Available online: https://pubs.acs.org/doi/pdf/10.1021/jf60191a005 (accessed on 1 August 2025).
- Malanin, K.; Lundberg, M.; Johansson, S.G.O. Anaphylactic Reaction Caused by Neoallergens in Heated Pecan Nut. Allergy 1995, 50, 988–991. [Google Scholar] [CrossRef]
- Gayraud, J.; Mairesse, M.; Fontaine, J.F.; Thillay, A.; Leduc, V.; Rancé, F.; Parisot, L.; Moneret-Vautrin, D.A. The Prevalence of Sensitization to Lupin Flour in France and Belgium: A Prospective Study in 5,366 Patients, by the Allergy Vigilance Network.
- Abbring, S.; Xiong, L.; Diks, M.A.P.; Baars, T.; Garssen, J.; Hettinga, K.; Esch, B.C.A.M. van Loss of Allergy-Protective Capacity of Raw Cow’s Milk after Heat Treatment Coincides with Loss of Immunologically Active Whey Proteins. Food & Function 2020, 11, 4982–4993. [Google Scholar] [CrossRef]
- Hefle, S.L.; Taylor, S.L. Revealing and Diagnosing Food Allergies and Intolerances. Clinical Nutrition Insight 2005, 31, 1. [Google Scholar]
- Ehn, B.-M.; Ekstrand, B.; Bengtsson, U.; Ahlstedt, S. Modification of IgE Binding during Heat Processing of the Cow’s Milk Allergen β-Lactoglobulin. J. Agric. Food Chem. 2004, 52, 1398–1403. [Google Scholar] [CrossRef]
- Mine, Y.; Yang, M. Recent Advances in the Understanding of Egg Allergens: Basic, Industrial, and Clinical Perspectives. J. Agric. Food Chem. 2008, 56, 4874–4900. [Google Scholar] [CrossRef]
- Maleki, S.J.; Schmitt, D.A.; Galeano, M.; Hurlburt, B.K. Comparison of the Digestibility of the Major Peanut Allergens in Thermally Processed Peanuts and in Pure Form. Foods 2014, 3, 290–303. [Google Scholar] [CrossRef] [PubMed]
- Maleki, S.J.; Sathe, S.K. The Effects of Processing Methods on Allergenic Properties of Food Proteins. In Food Allergy; John Wiley & Sons, Ltd., 2006; pp. 309–322 ISBN 978-1-68367-175-6.
- Verhoeckx, K.C.M.; Vissers, Y.M.; Baumert, J.L.; Faludi, R.; Feys, M.; Flanagan, S.; Herouet-Guicheney, C.; Holzhauser, T.; Shimojo, R.; van der Bolt, N.; et al. Food Processing and Allergenicity. Food and Chemical Toxicology 2015, 80, 223–240. [Google Scholar] [CrossRef]
- Bernhisel-Broadbent, J.; Scanlon, S.M.; Sampson, H.A. Fish Hypersensitivity: I. In Vitro and Oral Challenge Results in Fish-Allergic Patients. Journal of Allergy and Clinical Immunology 1992, 89, 730–737. [Google Scholar] [CrossRef]
- Alemán, A.; Sastre, J.; Quirce, S.; de las Heras, M.; Carnés, J.; Fernández-Caldas, E.; Pastor, C.; Blázquez, A.B.; Vivanco, F.; Cuesta-Herranz, J. Allergy to Kiwi: A Double-Blind, Placebo-Controlled Food Challenge Study in Patients from a Birch-Free Area. Journal of Allergy and Clinical Immunology 2004, 113, 543–550. [Google Scholar] [CrossRef]
- Fiocchi, A.; Bouygue, G.R.; Albarini, M.; Restani, P. Molecular Diagnosis of Cow’s Milk Allergy. Current Opinion in Allergy and Clinical Immunology 2011, 11, 216. [Google Scholar] [CrossRef]
- Fiocchi, A.; Pecora, V.; Petersson, C.J.; Dahdah, L.; Borres, M.P.; Amengual, M.J.; Huss-Marp, J.; Mazzina, O.; Di Girolamo, F. Sensitization Pattern to Inhalant and Food Allergens in Symptomatic Children at First Evaluation. Ital J Pediatr 2015, 41, 96. [Google Scholar] [CrossRef]
- Aalberse, R.C. Structural Biology of Allergens. Journal of Allergy and Clinical Immunology 2000, 106, 228–238. [Google Scholar] [CrossRef] [PubMed]
- Thomas, K.; Herouet-Guicheney, C.; Ladics, G.; Bannon, G.; Cockburn, A.; Crevel, R.; Fitzpatrick, J.; Mills, C.; Privalle, L.; Vieths, S. Evaluating the Effect of Food Processing on the Potential Human Allergenicity of Novel Proteins: International Workshop Report. Food and Chemical Toxicology 2007, 45, 1116–1122. [Google Scholar] [CrossRef]
- Iwan, M.; Vissers, Y.M.; Fiedorowicz, E.; Kostyra, H.; Kostyra, E.; Savelkoul, H.F.J.; Wichers, H.J. Impact of Maillard Reaction on Immunoreactivity and Allergenicity of the Hazelnut Allergen Cor a 11. J. Agric. Food Chem. 2011, 59, 7163–7171. [Google Scholar] [CrossRef]
- Vissers, Y.M.; Blanc, F.; Skov, P.S.; Johnson, P.E.; Rigby, N.M.; Przybylski-Nicaise, L.; Bernard, H.; Wal, J.-M.; Ballmer-Weber, B.; Zuidmeer-Jongejan, L.; et al. Effect of Heating and Glycation on the Allergenicity of 2S Albumins (Ara h 2/6) from Peanut. PLOS ONE 2011, 6, e23998. [Google Scholar] [CrossRef] [PubMed]
- Prabakaran, S.; Damodaran, S. Thermal Unfolding of β-Lactoglobulin: Characterization of Initial Unfolding Events Responsible for Heat-Induced Aggregation. J. Agric. Food Chem. 1997, 45, 4303–4308. [Google Scholar] [CrossRef]
- Bindslev-Jensen, C.; Skov, P.S.; Roggen, E.L.; Hvass, P.; Brinch, D.S. Investigation on Possible Allergenicity of 19 Different Commercial Enzymes Used in the Food Industry. Food and Chemical Toxicology 2006, 44, 1909–1915. [Google Scholar] [CrossRef]
- Kopper, R.A.; Odum, N.J.; Sen, M.; Helm, R.M.; Stanley, J.S.; Burks, A.W. Peanut Protein Allergens: The Effect of Roasting on Solubility and Allergenicity. Int Arch Allergy Immunol 2005, 136, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, K.; Hamajima, N.; Hirai, T.; Kato, T.; Koike, K.; Inoue, M.; Takezaki, T.; Tajima, K. Aldehyde Dehydrogenase 2(ALDH2)Genotype Affects Rectal Cancer Susceptibility Due to Alcohol Consumption. Journal of Epidemiology 2002, 12, 70–76. [Google Scholar] [CrossRef]
- Ayuso, R.; Lehrer, S.B.; Tanaka, L.; Ibanez, M.D.; Pascual, C.; Burks, A.W.; Sussman, G.L.; Goldberg, B.; Lopez, M.; Reese, G. IgE Antibody Response to Vertebrate Meat Proteins Including Tropomyosin. Annals of Allergy, Asthma & Immunology 1999, 83, 399–405. [Google Scholar] [CrossRef]
- Varjonen, E.; Björkstén, F.; Savolainen, J. Stability of Cereal Allergens. Clinical & Experimental Allergy 1996, 26, 436–443. [Google Scholar] [CrossRef]
- Posch, A.; Chen, Z.; Dunn, M.J.; Wheeler, C.H.; Petersen, A.; Leubner-Metzger, G.; Baur, X. Latex Allergen Database. Electrophoresis 1997, 18, 2803–2810. [Google Scholar] [CrossRef]
- Werfel, S.J.; Cooke, S.K.; Sampson, H.A. Clinical Reactivity to Beef in Children Allergic to Cow’s Milk. J Allergy Clin Immunol 1997, 99, 293–300. [Google Scholar] [CrossRef]
- Gruber, P.; Vieths, S.; Wangorsch, A.; Nerkamp, J.; Hofmann, T. Maillard Reaction and Enzymatic Browning Affect the Allergenicity of Pru Av 1, the Major Allergen from Cherry (Prunus Avium). J. Agric. Food Chem. 2004, 52, 4002–4007. [Google Scholar] [CrossRef]
- Hansen, K.S.; Ballmer-Weber, B.K.; Lüttkopf, D.; Skov, P.S.; Wüthrich, B.; Bindslev-Jensen, C.; Vieths, S.; Poulsen, L.K. Roasted Hazelnuts – Allergenic Activity Evaluated by Double-Blind, Placebo-Controlled Food Challenge. Allergy 2003, 58, 132–138. [Google Scholar] [CrossRef]
- Maleki, S.J.; Viquez, O.; Jacks, T.; Dodo, H.; Champagne, E.T.; Chung, S.-Y.; Landry, S.J. The Major Peanut Allergen, Ara h 2, Functions as a Trypsin Inhibitor, and Roasting Enhances This Function. Journal of Allergy and Clinical Immunology 2003, 112, 190–195. [Google Scholar] [CrossRef]
- Vanga, S.K.; Singh, A.; Raghavan, V. Review of Conventional and Novel Food Processing Methods on Food Allergens. Critical Reviews in Food Science and Nutrition 2017, 57, 2077–2094. [Google Scholar] [CrossRef]
- Pi, X.; Zhu, L.; Liu, J.; Zhang, B. Effect of Thermal Processing on Food Allergenicity: Mechanisms, Application, Influence Factor, and Future Perspective. J. Agric. Food Chem. 2024, 72, 20225–20240. [Google Scholar] [CrossRef] [PubMed]
- Khammeethong, T.; Phiriyangkul, P.; Inson, C.; Sinthuvanich, C. Effect of Microwave Vacuum Drying and Tray Drying on the Allergenicity of Protein Allergens in Edible Cricket, Gryllus Bimaculatus. Food Control 2024, 160, 110328. [Google Scholar] [CrossRef]
- Fiocchi, A.; Restani, P.; Riva, E.; Mirri, G.P.; Santini, I.; Bernardo, L.; Galli, C.L. Heat Treatment Modifies the Allergenicity of Beef and Bovine Serum Albumin. Allergy 1998, 53, 798–802. [Google Scholar] [CrossRef] [PubMed]
- Campus, M. High Pressure Processing of Meat, Meat Products and Seafood. Food Eng. Rev. 2010, 2, 256–273. [Google Scholar] [CrossRef]
- Messens, W.; Van Camp, J.; Huyghebaert, A. The Use of High Pressure to Modify the Functionality of Food Proteins. Trends in Food Science & Technology 1997, 8, 107–112. [Google Scholar] [CrossRef]
- Tauscher, B. Pasteurization of Food by Hydrostatic High Pressure: Chemical Aspects. Z Lebensm Unters Forch 1995, 200, 3–13. [Google Scholar] [CrossRef]
- Considine, K.M.; Kelly, A.L.; Fitzgerald, G.F.; Hill, C.; Sleator, R.D. High-Pressure Processing – Effects on Microbial Food Safety and Food Quality. FEMS Microbiol Lett 2008, 281, 1–9. [Google Scholar] [CrossRef]
- Dhakal, S.; Liu, C.; Zhang, Y.; Roux, K.H.; Sathe, S.K.; Balasubramaniam, V.M. Effect of High Pressure Processing on the Immunoreactivity of Almond Milk. Food Research International 2014, 62, 215–222. [Google Scholar] [CrossRef]
- Barba, F.J.; Esteve, M.J.; Frígola, A. High Pressure Treatment Effect on Physicochemical and Nutritional Properties of Fluid Foods During Storage: A Review. Comprehensive Reviews in Food Science and Food Safety 2012, 11, 307–322. [Google Scholar] [CrossRef]
- Chicón, R.; Belloque, J.; Alonso, E.; López-Fandiño, R. Antibody Binding and Functional Properties of Whey Protein Hydrolysates Obtained under High Pressure. Food Hydrocolloids 2009, 23, 593–599. [Google Scholar] [CrossRef]
- Heremans, K. The Effects of High Pressure on Biomaterials. In Ultra High Pressure Treatments of Foods; Hendrickx, M.E.G., Knorr, D., Ludikhuyze, L., Van Loey, A., Heinz, V., Eds.; Springer US: Boston, MA, 2001; ISBN 978-1-4615-0723-9. [Google Scholar]
- Peñas, E.; Préstamo, G.; Luisa Baeza, M.; Martínez-Molero, M.I.; Gomez, R. Effects of Combined High Pressure and Enzymatic Treatments on the Hydrolysis and Immunoreactivity of Dairy Whey Proteins. International Dairy Journal 2006, 16, 831–839. [Google Scholar] [CrossRef]
- Kleber, N.; Maier, S.; Hinrichs, J. Antigenic Response of Bovine β-Lactoglobulin Influenced by Ultra-High Pressure Treatment and Temperature. Innovative Food Science & Emerging Technologies 2007, 8, 39–45. [Google Scholar] [CrossRef]
- Kato, T.; Katayama, E.; Matsubara, S.; Omi, Y.; Matsuda, T. Release of Allergenic Proteins from Rice Grains Induced by High Hydrostatic Pressure. J. Agric. Food Chem. 2000, 48, 3124–3129. [Google Scholar] [CrossRef] [PubMed]
- Odani, S.; Kanda, Y.; Hara, T.; Matuno, M.; Suzuki, A. Effects of High Hydrostatic Pressure Treatment on the Allergenicity and Structure of Chicken Egg White Ovomucoid. High Pressure Bioscience and Biotechnology, 2007, 1, 252–258. [Google Scholar] [CrossRef]
- Wang, X.-S.; Tang, C.-H.; Li, B.-S.; Yang, X.-Q.; Li, L.; Ma, C.-Y. Effects of High-Pressure Treatment on Some Physicochemical and Functional Properties of Soy Protein Isolates. Food Hydrocolloids 2008, 22, 560–567. [Google Scholar] [CrossRef]
- Akdemir Evrendilek, G. Chapter 14 - Pulsed Electric Field Processing: Food Pasteurization, Tissue Treatment, and Seed Disinfection. In Food Packaging and Preservation; Jaiswal, A.K., Shankar, S., Eds.; Academic Press, 2024; pp. 259–273 ISBN 978-0-323-90044-7.
- Barba, F.J.; Parniakov, O.; Pereira, S.A.; Wiktor, A.; Grimi, N.; Boussetta, N.; Saraiva, J.A.; Raso, J.; Martin-Belloso, O.; Witrowa-Rajchert, D.; et al. Current Applications and New Opportunities for the Use of Pulsed Electric Fields in Food Science and Industry. Food Research International 2015, 77, 773–798. [Google Scholar] [CrossRef]
- Chatzimitakos, T.; Athanasiadis, V.; Kalompatsios, D.; Mantiniotou, M.; Bozinou, E.; Lalas, S.I. Pulsed Electric Field Applications for the Extraction of Bioactive Compounds from Food Waste and By-Products: A Critical Review. Biomass 2023, 3, 367–401. [Google Scholar] [CrossRef]
- Yarmush, M.L.; Golberg, A.; Serša, G.; Kotnik, T.; Miklavčič, D. Electroporation-Based Technologies for Medicine: Principles, Applications, and Challenges. Annual Review of Biomedical Engineering 2014, 16, 295–320. [Google Scholar] [CrossRef]
- Evrendilek, G.A. Pulsed Electric Field Treatment for Beverage Production and Preservation; Springer International Publishing, 2017; ISBN 978-3-319-32886-7.
- Evrendilek, G.A. Influence of High Pressure Processing on Food Bioactives. In Retention of Bioactives. In Retention of Bioactives in Food Processing; Jafari, S.M., Capanoglu, E., Eds.; Food Bioactive Ingredients; Springer International Publishing: Cham, 2022; ISBN 978-3-030-96885-4. [Google Scholar]
- Lebovka, N.; Vorobiev, E.; Chemat, F. Enhancing Extraction Processes in the Food Industry; CRC Press, 2016; ISBN 978-1-4398-4595-0.
- Toepfl, S.; Heinz, V.; Knorr, D. High Intensity Pulsed Electric Fields Applied for Food Preservation. Chemical Engineering and Processing: Process Intensification 2007, 46, 537–546. [Google Scholar] [CrossRef]
- Akdemir Evrendilek, G.; Tanriverdi, H.; Demir, I.; Uzuner, S. Shelf-Life Extension of Traditional Licorice Root “Sherbet” with a Novel Pulsed Electric Field Processing. Frontiers in Food Science and Technology 2023, 3. [Google Scholar] [CrossRef]
- Akdemir Evrendilek, G.; Hitit Özkan, B. Pulsed Electric Field Processing of Fruit Juices with Inactivation of Enzymes with New Inactivation Kinetic Model and Determination of Changes in Quality Parameters. Innovative Food Science & Emerging Technologies 2024, 94, 103678. [Google Scholar] [CrossRef]
- Van Loey, A.; Verachtert, B.; Hendrickx, M. Effects of High Electric Field Pulses on Enzymes. Trends in Food Science & Technology 2001, 12, 94–102. [Google Scholar] [CrossRef]
- Wei, J.-N.; Zeng, X.-A.; Tang, T.; Jiang, Z.; Liu, Y.-Y. Unfolding and Nanotube Formation of Ovalbumin Induced by Pulsed Electric Field. Innovative Food Science & Emerging Technologies 2018, 45, 249–254. [Google Scholar] [CrossRef]
- Zhao, W.; Yang, R. Comparative Study of Inactivation and Conformational Change of Lysozyme Induced by Pulsed Electric Fields and Heat. Eur Food Res Technol 2008, 228, 47–54. [Google Scholar] [CrossRef]
- Zhao, W.; Yang, R. Effect of High-Intensity Pulsed Electric Fields on the Activity, Conformation and Self-Aggregation of Pepsin. Food Chemistry 2009, 114, 777–781. [Google Scholar] [CrossRef]
- Han, Z.; Cai, M.; Cheng, J.-H.; Sun, D.-W. Effects of Electric Fields and Electromagnetic Wave on Food Protein Structure and Functionality: A Review. Trends in Food Science & Technology 2018, 75, 1–9. [Google Scholar] [CrossRef]
- Li, Y.; Chen, Z.; Mo, H. Effects of Pulsed Electric Fields on Physicochemical Properties of Soybean Protein Isolates. LWT - Food Science and Technology 2007, 40, 1167–1175. [Google Scholar] [CrossRef]
- Zhang, R.; Cheng, L.; Wang, L.; Guan, Z. Inactivation Effects of PEF on Horseradish Peroxidase (HRP) and Pectinesterase (PE). IEEE Transactions on Plasma Science 2006, 34, 2630–2636. [Google Scholar] [CrossRef]
- Johnson, P.E.; Van der Plancken, I.; Balasa, A.; Husband, F.A.; Grauwet, T.; Hendrickx, M.; Knorr, D.; Mills, E.N.C.; Mackie, A.R. High Pressure, Thermal and Pulsed Electric-Field-Induced Structural Changes in Selected Food Allergens. Molecular Nutrition & Food Research 2010, 54, 1701–1710. [Google Scholar] [CrossRef]
- Tobajas, A.P.; Agulló-García, A.; Cubero, J.L.; Colás, C.; Segura-Gil, I.; Sánchez, L.; Calvo, M.; Pérez, M.D. Effect of High Pressure and Pulsed Electric Field on Denaturation and Allergenicity of Pru p 3 Protein from Peach. Food Chemistry 2020, 321, 126745. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Tu, Z.; Wang, H.; Zhang, L.; Gao, Y.; Li, X.; Tian, M. Immunogenic and Structural Properties of Ovalbumin Treated by Pulsed Electric Fields. International Journal of Food Properties 2017, 20, S3164–S3176. [Google Scholar] [CrossRef]
- Vanga, S.K.; Singh, A.; Raghavan, V. Effect of Thermal and Electric Field Treatment on the Conformation of Ara h 6 Peanut Protein Allergen. Innovative Food Science & Emerging Technologies 2015, 30, 79–88. [Google Scholar] [CrossRef]
- Vanga, S.K.; Singh, A.; Kalkan, F.; Gariepy, Y.; Orsat, V.; Raghavan, V. Effect of Thermal and High Electric Fields on Secondary Structure of Peanut Protein. International Journal of Food Properties 2016, 19, 1259–1271. [Google Scholar] [CrossRef]
- Paschke, A. Aspects of Food Processing and Its Effect on Allergen Structure. Molecular Nutrition & Food Research 2009, 53, 959–962. [Google Scholar] [CrossRef]
- Dong, L.; Lu, X.; Zeng, X.; Lin, S. Regulation of Ovalbumin Allergenicity and Structure-Activity Relationship Analysis Based on Pulsed Electric Field Technology. International Journal of Biological Macromolecules 2024, 261, 129695. [Google Scholar] [CrossRef]
- Shriver, S.K.; Yang, W.W. Thermal and Nonthermal Methods for Food Allergen Control. Food Eng. Rev. 2011, 3, 26–43. [Google Scholar] [CrossRef]
- Chung, S.-Y.; Yang, W.; Krishnamurthy, K. Effects of Pulsed UV-Light on Peanut Allergens in Extracts and Liquid Peanut Butter. Journal of Food Science 2008, 73, C400–C404. [Google Scholar] [CrossRef]
- Garvey, M.; Rowan, N. Pulsed UV as a Potential Surface Sanitizer in Food Production Processes to Ensure Consumer Safety. Current Opinion in Food Science 2019, 26, 65–70. [Google Scholar] [CrossRef]
- Gómez-lópez, V.M. Continuous UV-C Light. In Decontamination of Fresh and Minimally Processed Produce; John Wiley & Sons, Ltd., 2012; pp. 365–378 ISBN 978-1-118-22918-7.
- Gómez-López, V.M.; Koutchma, T.; Linden, K. Chapter 8 - Ultraviolet and Pulsed Light Processing of Fluid Foods. In Novel Thermal and Non-Thermal Technologies for Fluid Foods. In Novel Thermal and Non-Thermal Technologies for Fluid Foods; Cullen, P.J., Tiwari, B.K., Valdramidis, V.P., Eds.; Academic Press: San Diego, 2012; ISBN 978-0-12-381470-8. [Google Scholar]
- Krishnamurthy, K.; Tewari, J.C.; Irudayaraj, J.; Demirci, A. Microscopic and Spectroscopic Evaluation of Inactivation of Staphylococcus Aureus by Pulsed UV Light and Infrared Heating. Food Bioprocess Technol 2010, 3, 93–104. [Google Scholar] [CrossRef]
- Gorman, S.; McGlade, J.P.; Lambert, M.J.M.; Strickland, D.H.; Thomas, J.A.; Hart, P.H. UV Exposure and Protection against Allergic Airways Disease. Photochem Photobiol Sci 2010, 9, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Keklik, N.M.; Krishnamurthy, K.; Demirci, A. 12 - Microbial Decontamination of Food by Ultraviolet (UV) and Pulsed UV Light. In Microbial Decontamination in the Food Industry; Demirci, A., Ngadi, M.O., Eds.; Woodhead Publishing Series in Food Science, Technology and Nutrition; Woodhead Publishing, 2012; pp. 344–369 ISBN 978-0-85709-085-0.
- Yang, W.W.; Mwakatage, N.R.; Goodrich-Schneider, R.; Krishnamurthy, K.; Rababah, T.M. Mitigation of Major Peanut Allergens by Pulsed Ultraviolet Light. Food Bioprocess Technol 2012, 5, 2728–2738. [Google Scholar] [CrossRef]
- Meinlschmidt, P.; Ueberham, E.; Lehmann, J.; Reineke, K.; Schlüter, O.; Schweiggert-Weisz, U.; Eisner, P. The Effects of Pulsed Ultraviolet Light, Cold Atmospheric Pressure Plasma, and Gamma-Irradiation on the Immunoreactivity of Soy Protein Isolate. Innovative Food Science & Emerging Technologies 2016, 38, 374–383. [Google Scholar] [CrossRef]
- Tammineedi, C.V.R.K.; Choudhary, R.; Perez-Alvarado, G.C.; Watson, D.G. Determining the Effect of UV-C, High Intensity Ultrasound and Nonthermal Atmospheric Plasma Treatments on Reducing the Allergenicity of α-Casein and Whey Proteins. LWT - Food Science and Technology 2013, 54, 35–41. [Google Scholar] [CrossRef]
- Lee, J.-W.; Kim, J.-H.; Yook, H.-S.; Kang, K.-O.; Lee, S.-Y.; Hwang, H.-J.; Byun, M.-W. Effects of Gamma Radiation on the Allergenic and Antigenic Properties of Milk Proteins. Journal of Food Protection 2001, 64, 272–276. [Google Scholar] [CrossRef] [PubMed]
- Manzocco, L.; Panozzo, A.; Nicoli, M.C. Effect of Ultraviolet Processing on Selected Properties of Egg White. Food Chemistry 2012, 135, 522–527. [Google Scholar] [CrossRef]
- Surowsky, B.; Schlüter, O.; Knorr, D. Interactions of Non-Thermal Atmospheric Pressure Plasma with Solid and Liquid Food Systems: A Review. Food Eng Rev 2015, 7, 82–108. [Google Scholar] [CrossRef]
- Meza-Morales, W.; Estévez-Carmona, M.M.; Alvarez-Ricardo, Y.; Obregón-Mendoza, M.A.; Cassani, J.; Ramírez-Apan, M.T.; Escobedo-Martínez, C.; Soriano-García, M.; Reynolds, W.F.; Enríquez, R.G. Full Structural Characterization of Homoleptic Complexes of Diacetylcurcumin with Mg, Zn, Cu, and Mn: Cisplatin-Level Cytotoxicity in Vitro with Minimal Acute Toxicity in Vivo. Molecules 2019, 24, 1598. [Google Scholar] [CrossRef]
- Shriver, S.; Yang, W.; Chung, S.-Y.; Percival, S. Pulsed Ultraviolet Light Reduces Immunoglobulin E Binding to Atlantic White Shrimp (Litopenaeus Setiferus) Extract. International Journal of Environmental Research and Public Health 2011, 8, 2569–2583. [Google Scholar] [CrossRef]
- Jiménez-Saiz, R.; Benedé, S.; Molina, E.; López-Expósito, I. Effect of Processing Technologies on the Allergenicity of Food Products. Critical Reviews in Food Science and Nutrition 2015, 55, 1902–1917. [Google Scholar] [CrossRef]
- Uesugi, A.R.; Danyluk, M.D.; Harris, L.J. Survival of Salmonella Enteritidis Phage Type 30 on Inoculated Almonds Stored at −20, 4, 23, and 35°C. Journal of Food Protection 2006, 69, 1851–1857. [Google Scholar] [CrossRef]
- Guerrero-Beltrán, J.; Welti-Chanes, J.; Barbosa-Cánovas, G.V. Ultraviolet-C light processing of grape, cranberry and grapefruit juices to inactivate Saccharomyces cerevisiae. Journal of Food Process Engineering 2009, 32, 916–932. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1745-4530.2008.00253.x (accessed on 4 August 2025). [CrossRef]
- Pathak, B.; Omre, P.K.; Bisht, B.; Saini, D. Effect of thermal and non-thermal processing methods on food allergens.
- Tolouie, H.; Mohammadifar, M.A.; Ghomi, H.; Hashemi, M. Cold Atmospheric Plasma Manipulation of Proteins in Food Systems. Critical Reviews in Food Science and Nutrition 2018, 58, 2583–2597. [Google Scholar] [CrossRef]
- Shriver, S.K.; Yang, W.W. Thermal and Nonthermal Methods for Food Allergen Control. Food Eng. Rev. 2011, 3, 26–43. [Google Scholar] [CrossRef]
- Vaz, A.F.M.; Souza, M.P.; Carneiro-da-Cunha, M.G.; Medeiros, P.L.; Melo, A.M.M.A.; Aguiar, J.S.; Silva, T.G.; Silva-Lucca, R.A.; Oliva, M.L.V.; Correia, M.T.S. Molecular Fragmentation of Wheat-Germ Agglutinin Induced by Food Irradiation Reduces Its Allergenicity in Sensitised Mice. Food Chemistry 2012, 132, 1033–1039. [Google Scholar] [CrossRef]
- Luo, C.; Hu, C.; Gao, J.; Li, X.; Wu, Z.; Yang, A.; Chen, H. A Potential Practical Approach to Reduce Ara h 6 Allergenicity by Gamma Irradiation. Food Chemistry 2013, 136, 1141–1147. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.; Jang, D.-I.; Lee, J.-W.; Kim, J.-H.; Byun, M.-W.; Lee, S.-Y. Evaluation of Reduced Allergenicity of Irradiated Peanut Extract Using Splenocytes from Peanut-Sensitized Mice. Radiation Physics and Chemistry 2009, 78, 615–617. [Google Scholar] [CrossRef]
- Zhenxing, L.; Hong, L.; Limin, C.; Jamil, K. The Influence of Gamma Irradiation on the Allergenicity of Shrimp (Penaeus Vannamei). Journal of Food Engineering 2007, 79, 945–949. [Google Scholar] [CrossRef]
- Kaddouri, H.; Mimoun, S.; El-Mecherfi, K.E.; Chekroun, A.; Kheroua, O.; Saidi, D. Impact of γ-Radiation on Antigenic Properties of Cow’s Milk β-Lactoglobulin. Journal of Food Protection 2008, 71, 1270–1272. [Google Scholar] [CrossRef]
- Su, M.; Venkatachalam, M.; Teuber, S.S.; Roux, K.H.; Sathe, S.K. Impact of γ-Irradiation and Thermal Processing on the Antigenicity of Almond, Cashew Nut and Walnut Proteins. Journal of the Science of Food and Agriculture 2004, 84, 1119–1125. [Google Scholar] [CrossRef]
- Leszczynska, J.; Łącka, A.; Szemraj, J.; Lukamowicz, J.; Zegota, H. The Influence of Gamma Irradiation on the Immunoreactivity of Gliadin and Wheat Flour. Eur Food Res Technol 2003, 217, 143–147. [Google Scholar] [CrossRef]
- Seo, J.-H.; Kim, J.-H.; Lee, J.-W.; Yoo, Y.-C.; Kim, M.R.; Park, K.-S.; Byun, M.-W. Ovalbumin Modified by Gamma Irradiation Alters Its Immunological Functions and Allergic Responses. International Immunopharmacology 2007, 7, 464–472. [Google Scholar] [CrossRef]
- Davies, K.J.; Delsignore, M.E. Protein Damage and Degradation by Oxygen Radicals. III. Modification of Secondary and Tertiary Structure. Journal of Biological Chemistry 1987, 262, 9908–9913. [Google Scholar] [CrossRef]
- Byun, M.-W.; Kim, J.-H.; Lee, J.-W.; Park, J.-W.; Hong, C.-S.; Kang, I.-J. Effects of Gamma Radiation on the Conformational and Antigenic Properties of a Heat-Stable Major Allergen in Brown Shrimp. Journal of Food Protection 2000, 63, 940–944. [Google Scholar] [CrossRef]
- Seo, J.-H.; Lee, J.-W.; Lee, Y.-S.; Lee, S.-Y.; Kim, M.-R.; Yook, H.-S.; Byun, M.-W. Change of an Egg Allergen in a White Layer Cake Containing Gamma-Irradiated Egg White. Journal of Food Protection 2004, 67, 1725–1730. [Google Scholar] [CrossRef]
- Zhao, J.; Zhu, W.; Zeng, J.; Liu, Y.; Li, H.; Wang, H.; Zhang, Z.; Lin, H.; Li, Z. Insight into the Mechanism of Allergenicity Decreasing in Recombinant Sarcoplasmic Calcium-Binding Protein from Shrimp (Litopenaeus Vannamei) with Thermal Processing via Spectroscopy and Molecular Dynamics Simulation Techniques. Food Research International 2022, 157, 111427. [Google Scholar] [CrossRef] [PubMed]
- Kume, T.; Furuta, M.; Todoriki, S.; Uenoyama, N.; Kobayashi, Y. Status of Food Irradiation in the World. Radiation Physics and Chemistry 2009, 78, 222–226. [Google Scholar] [CrossRef]
- Piyasena, P.; Mohareb, E.; McKellar, R.C. Inactivation of Microbes Using Ultrasound: A Review. International Journal of Food Microbiology 2003, 87, 207–216. [Google Scholar] [CrossRef]
- Chandrapala, J.; Oliver, C.; Kentish, S.; Ashokkumar, M. Ultrasonics in Food Processing. Ultrasonics Sonochemistry 2012, 19, 975–983. [Google Scholar] [CrossRef]
- Soria, A.C.; Villamiel, M. Effect of Ultrasound on the Technological Properties and Bioactivity of Food: A Review. Trends in Food Science & Technology 2010, 21, 323–331. [Google Scholar] [CrossRef]
- Chemat, F.; Rombaut, N.; Sicaire, A.-G.; Meullemiestre, A.; Fabiano-Tixier, A.-S.; Abert-Vian, M. Ultrasound Assisted Extraction of Food and Natural Products. Mechanisms, Techniques, Combinations, Protocols and Applications. A Review. Ultrasonics Sonochemistry 2017, 34, 540–560. [Google Scholar] [CrossRef]
- Knorr, D.; Froehling, A.; Jaeger, H.; Reineke, K.; Schlueter, O.; Schoessler, K. Emerging Technologies in Food Processing. Annual Review of Food Science and Technology 2011, 2, 203–235. [Google Scholar] [CrossRef]
- Mason, T.J. Ultrasonic Cleaning: An Historical Perspective. Ultrasonics Sonochemistry 2016, 29, 519–523. [Google Scholar] [CrossRef]
- Zhou, C.; Sun, D.; Sun, X.; Zhu, C.; Wang, Q. Combining Ultrasound and Microwave to Improve the Yield and Quality of Single-Cell Oil from Mortierella Isabellina NTG1−121. Journal of the American Oil Chemists’ Society 2018, 95, 1535–1547. [Google Scholar] [CrossRef]
- Liu, X.; Guo, Y.; Wang, S.; Wang, X.; Wang, Z.; Wang, Z. Effects of Ultrasound Combined with Hydrothermal Treatment on the Quality of Brown Rice. LWT 2024, 196, 115874. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, X.; Chen, W.; Zhou, P. Conformation Stability, in Vitro Digestibility and Allergenicity of Tropomyosin from Shrimp (Exopalaemon Modestus) as Affected by High Intensity Ultrasound. Food Chemistry 2018, 245, 997–1009. [Google Scholar] [CrossRef]
- Chen, H.-L.; Mao, H.-Y.; Cao, M.-J.; Cai, Q.-F.; Su, W.-J.; Zhang, Y.-X.; Liu, G.-M. Purification, Physicochemical and Immunological Characterization of Arginine Kinase, an Allergen of Crayfish (Procambarus Clarkii). Food and Chemical Toxicology 2013, 62, 475–484. [Google Scholar] [CrossRef]
- Stanic-Vucinic, D.; Stojadinovic, M.; Atanaskovic-Markovic, M.; Ognjenovic, J.; Grönlund, H.; van Hage, M.; Lantto, R.; Sancho, A.I.; Velickovic, T.C. Structural Changes and Allergenic Properties of β-Lactoglobulin upon Exposure to High-Intensity Ultrasound. Molecular Nutrition & Food Research 2012, 56, 1894–1905. [Google Scholar] [CrossRef]
- Barre, A.; Jacquet, G.; Sordet, C.; Culerrier, R.; Rougé, P. Homology Modelling and Conformational Analysis of IgE-Binding Epitopes of Ara h 3 and Other Legumin Allergens with a Cupin Fold from Tree Nuts. Molecular Immunology 2007, 44, 3243–3255. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Lin, L.; Chau, F. Ultrasound-Assisted Extraction of Ginseng Saponins from Ginseng Roots and Cultured Ginseng Cells. Ultrasonics Sonochemistry 2001, 8, 347–352. [Google Scholar] [CrossRef]
- Wang, J.; Vanga, S.K.; McCusker, C.; Raghavan, V. A Comprehensive Review on Kiwifruit Allergy: Pathogenesis, Diagnosis, Management, and Potential Modification of Allergens Through Processing. Compr Rev Food Sci Food Saf 2019, 18, 500–513. [Google Scholar] [CrossRef]
- Dong, X.; Wang, J.; Raghavan, V. Critical Reviews and Recent Advances of Novel Non-Thermal Processing Techniques on the Modification of Food Allergens. Critical Reviews in Food Science and Nutrition 2021, 61, 196–210. [Google Scholar] [CrossRef] [PubMed]
- Jadhav, H.B.; Annapure, U.S.; Deshmukh, R.R. Non-Thermal Technologies for Food Processing. Front. Nutr. 2021, 8. [Google Scholar] [CrossRef]
- Li, Y.-Q.; Chen, Q.; Liu, X.-H.; Chen, Z.-X. Inactivation of Soybean Lipoxygenase in Soymilk by Pulsed Electric Fields. Food Chemistry 2008, 109, 408–414. [Google Scholar] [CrossRef]
- Li, Y.; Wang, J.; Zeng, Q.-H.; Wang, L.; Wang, J.J.; Li, S.; Zhu, J.; Zeng, X.-A. Novel Thawing Method of Ultrasound-Assisted Slightly Basic Electrolyzed Water Improves the Processing Quality of Frozen Shrimp Compared with Traditional Thawing Approaches. Ultrasonics Sonochemistry 2024, 107, 106931. [Google Scholar] [CrossRef]
- Sun, Y.; Zhang, M.; Bhandari, B.; Yang, C. Ultrasound Treatment of Frozen Crayfish with Chitosan Nano-Composite Water-Retaining Agent: Influence on Cryopreservation and Storage Qualities. Food Research International 2019, 126, 108670. [Google Scholar] [CrossRef]
- Ekezie, F.-G.C.; Cheng, J.-H.; Sun, D.-W. Effects of Atmospheric Pressure Plasma Jet on the Conformation and Physicochemical Properties of Myofibrillar Proteins from King Prawn (Litopenaeus Vannamei). Food Chemistry 2019, 276, 147–156. [Google Scholar] [CrossRef]
- Venkataratnam, H.; Cahill, O.; Sarangapani, C.; Cullen, P.J.; Barry-Ryan, C. Impact of Cold Plasma Processing on Major Peanut Allergens. Sci Rep 2020, 10, 17038. [Google Scholar] [CrossRef] [PubMed]
- Nooji, J.K. 2011. Reduction of wheat allergen potency by pulsed ultraviolet light, high hydrostatic pressure, and non-thermal plasma (Doctoral dissertation, University of Florida).
- Dasan, B.G.; Boyaci, I.H. Effect of Cold Atmospheric Plasma on Inactivation of Escherichia Coli and Physicochemical Properties of Apple, Orange, Tomato Juices, and Sour Cherry Nectar. Food Bioprocess Technol 2018, 11, 334–343. [Google Scholar] [CrossRef]
- Pankaj, S.K.; Wan, Z.; Keener, K.M. Effects of Cold Plasma on Food Quality: A Review. Foods 2018, 7, 4. [Google Scholar] [CrossRef] [PubMed]
- Pankaj, S.K.; Keener, K.M. Cold Plasma: Background, Applications and Current Trends. Current Opinion in Food Science 2017, 16, 49–52. [Google Scholar] [CrossRef]
- Taylor, S.L.; Hefle, S.L. Will Genetically Modified Foods Be Allergenic? Journal of Allergy and Clinical Immunology 2001, 107, 765–771. [Google Scholar] [CrossRef]
- Rupa, P.; Nakamura, S.; Katayama, S.; Mine, Y. Attenuation of Allergic Immune Response Phenotype by Mannosylated Egg White in Orally Induced Allergy in Balb/c Mice. J. Agric. Food Chem. 2014, 62, 9479–9487. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.; Faustinelli, P.; Ramos, M.L.; Hajduch, M.; Stevenson, S.; Thelen, J.J.; Maleki, S.J.; Cheng, H.; Ozias-Akins, P. Reduction of IgE Binding and Nonpromotion of Aspergillus Flavus Fungal Growth by Simultaneously Silencing Ara h 2 and Ara h 6 in Peanut. J. Agric. Food Chem. 2008, 56, 11225–11233. [Google Scholar] [CrossRef]
- Herman, E.M.; Helm, R.M.; Jung, R.; Kinney, A.J. Genetic Modification Removes an Immunodominant Allergen from Soybean, Plant Physiol 2003, 132, 36–43. [Google Scholar] [CrossRef]
Figure 1.
Classification of food allergy (adapted from [169]).
Figure 1.
Classification of food allergy (adapted from [169]).
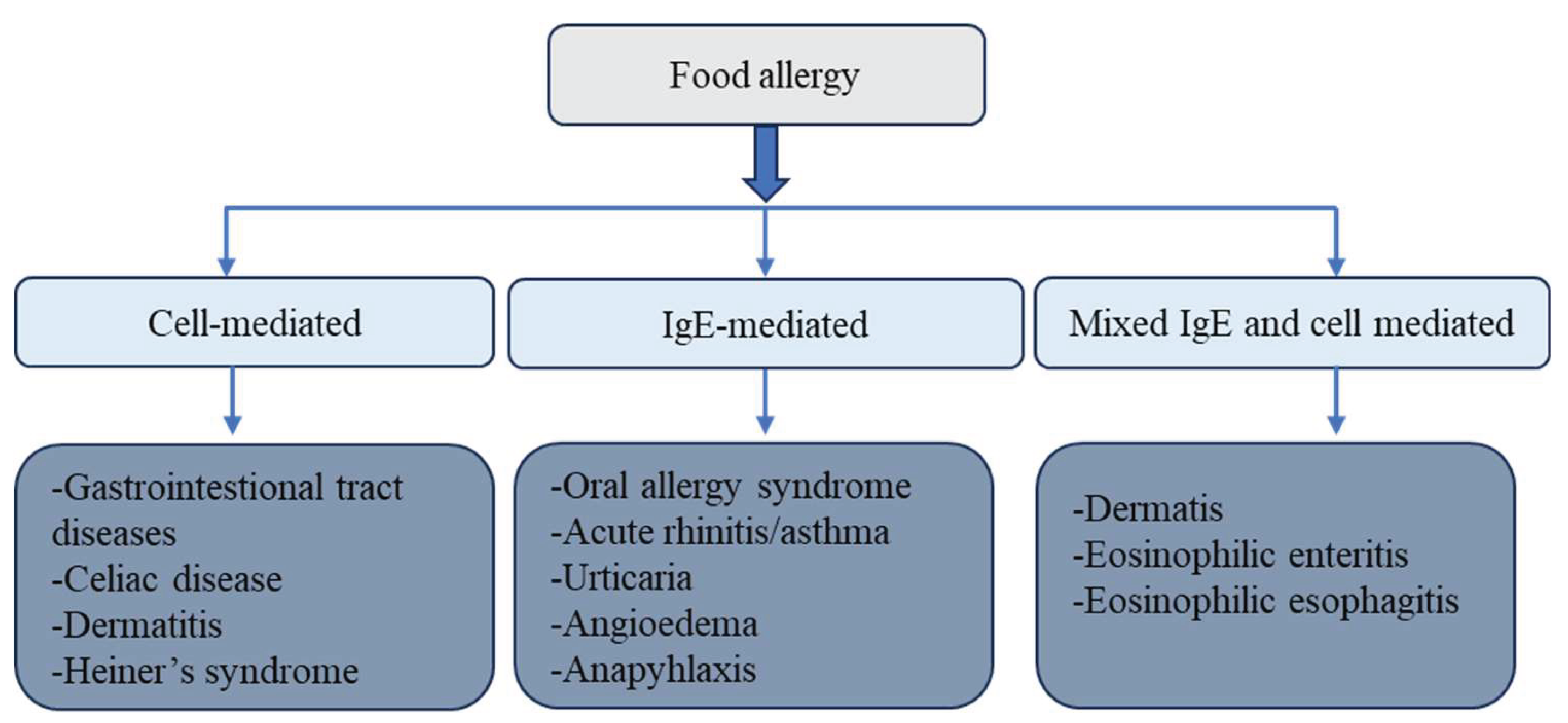

Table 1.
Influence of thermal processing on various food allergen types (adapted from [57]).
Table 1.
Influence of thermal processing on various food allergen types (adapted from [57]).
| Allergen Type | Response to Thermal Processing |
| Bet v 1-like proteins (e.g., Mal d 1 in apple, Pru av 1 in cherry) | Highly sensitive to heat; prone to unfolding. Susceptible to chemical changes such as Maillard reaction products in high-sugar foods and interactions with polyphenols, leading to decreased allergenic potential. |
| Prolamin superfamily proteins (e.g., non-specific lipid transfer proteins [nsLTPs], 2S albumins like Mal d 3; tropomyosin; parvalbumin) | Moderately heat stable; proteins unfold to a limited extent but tend to regain their structure upon cooling. Maillard reactions may still enhance allergenic potential. |
| Cupin family proteins (e.g., Ara h 1 in peanut); Lipocalins (e.g., β-lactoglobulin and α-lactalbumin in milk) | Partially resistant to denaturation; undergo partial unfolding and tend to aggregate. Can form structural networks (e.g., in emulsions or gels). Heat can also lead to Maillard reactions, which may increase allergenicity. |
| Flexible proteins (e.g., caseins in milk, gluten storage proteins in wheat, ovomucoid in egg) | Structurally dynamic and heat-resistant; maintain mobility and do not exhibit classic denaturation behavior under thermal conditions. Their allergenicity remains largely unchanged. |
Table 2.
Overview of nonthermal methods for allergen reduction (adapted from [169]).
Table 2.
Overview of nonthermal methods for allergen reduction (adapted from [169]).
| Technology | Mechanism of Action | Limitations |
| High-Pressure Processing | Disrupts non-covalent bonds within proteins; alters protein conformation through aggregation and gelation, affecting epitope exposure; enhances enzymatic hydrolysis. | Dual effects of pressure require accurate control; combining with other treatments may be necessary to optimize results. |
| Pulsed Electric Fields | Alters the secondary structure of allergenic proteins. | Often used alongside heat treatments to improve efficacy. |
| Pulsed UV Light | Delivers high-intensity UV pulses that cause photochemical modifications in proteins, including structural changes and epitope destruction | Limited penetration depth; potential degradation of sensitive nutrients; treatment uniformity can be challenging depending on food surface characteristics. |
| Cold Plasma | Reactive oxygen and nitrogen species (ROS and RNS) interact with antigens, altering protein configurations. | High equipment costs; limited understanding of how cold plasma mitigates food allergens; potential cytotoxic effects of treated liquids warrant further investigation. |
| Ultrasound | Cavitation generated by ultrasonic waves breaks peptide bonds, leading to irreversible unfolding and structural disruption of allergenic proteins. | Can negatively impact product color, flavor, and nutritional quality. |
| Gamma Irradiation | Allergenic proteins absorb radiation, altering their 3D structures; free radical formation leads to water radiolysis and modification of amino acid side chains. | Effective dose levels are not clearly established; not all protein-rich foods are currently approved for irradiation in the EU; concerns remain regarding food irradiation safety. |
Table 3.
Effects of high hydrostatic pressure on allergenity of various foods.
| Food Variety | Treatment Conditions | Key Findings | Reference |
| Soybean | 300 MPa, 40°C, 15 min | HHP notably reduced allergenicity in soybean sprouts, with only an 18% decrease in essential amino acids and overall nutritional value. HHP may offer a viable method for producing low-allergen soybean sprouts. | [2] |
| Soybean | 400 MPa | Treatment improved protein solubility and hydrophobicity, while decreasing β-sheet content. | [3] |
| Soy protein isolate (infant formula) | 300 MPa, 15 min | Allergenicity reduced by 46.8% due to structural and interaction changes in SPI, suggesting enhanced safety in allergic individuals. | [4] |
| Peanut | 150–800 MPa, 20–80°C, 10 min; 60–180 MPa | No major changes observed in allergen secondary structure under HHP. However, high-pressure microfluidization reduced Ara h 2 allergenicity by modifying its structure and increasing UV absorption and hydrophobicity. | [5,6] |
| Tofu | 300 MPa, 40°C, 15 min | HHP did not change tofu protein composition but lowered intensity of some protein bands. Allergenicity remained unchanged. | [2] |
| Rice | 100–400 MPa, 10–120 min; 300 MPa, 30 min + Protease N | Pressure facilitated allergen release into surrounding solution; with protease, allergens were nearly eliminated from rice grains. | [7] |
| Almond | 600 MPa, 4–70°C, 5–30 min | No significant change in allergen concentration or IgE-binding capacity as per SDS-PAGE, WB, and ELISA. | [8] |
| Whey protein isolate | 200–600 MPa, 30–68°C, 10–30 min | Antigenicity of β-lactoglobulin increased with pressure, temperature, and duration. | [9] |
| Skim milk | 200–600 MPa, 30–68°C, 10–30 min | β-lactoglobulin levels rose post-treatment; thermal addition mitigated allergenicity, but HHP-treated samples retained higher β-lg than controls. | [9] |
| Sweet whey | 200–600 MPa, 30–68°C, 10–30 min | β-lg antigenicity rose at moderate temperatures and pressures; higher temperatures reduced but did not eliminate antigenicity. | [9] |
| Milk | >100 MPa + chymotrypsin/trypsin, 20 min | Proteolysis of β-lg was accelerated under HHP, suggesting potential for hypoallergenic food production. | [10] |
| Apple | 400–800 MPa, 80°C, 10 min | Mal d 3 allergen lost α-helical structure, becoming a random coil; immunoreactivity declined at ≥400 MPa. | [6] |
| Apple | 700 MPa, 115°C, 10 min | Mal d 1 showed minimal changes at 20°C, but more pronounced at 80°C; Mal d 3 IgE reactivity dropped by ~30%. | [11] |
| Apple | 600 MPa, 5 min | Mal d 1 immunoreactivity decreased by >50% with combined high pressure and heat. | [11] |
| Apple | 600 MPa, 5 min; repeated consumption | Daily intake of HHP-treated apple gel for 3 weeks led to desensitization in highly allergic individuals; 90% negative skin tests post-treatment. | [12] |
| Carrot | 500 MPa, 50°C, 10 min | Slight increase in β-sheet structure of Dau c 1; no change in immunoreactivity observed. | [13] |
| Carrot juice | 400–550 MPa, 3–10 min; 500 MPa, 30–50°C, 10 min | HHP had no observable effect on the allergenic potential of carrot juice. | [13] |
| Celeriac | 700 MPa, 118°C, 10 min | Allergenicity of Api g 1 was significantly reduced through combined pressure and thermal processing. | [11] |
| Celery | 500 MPa, 50°C, 10 min | Api g 1 structural changes were pressure-dependent but did not impact allergenicity. | [14] |
Table 4.
Effect of UVC treatment on allergenity of various foods (adapted from [169]).
Table 4.
Effect of UVC treatment on allergenity of various foods (adapted from [169]).
| Food material | Target allergen | Treatment | Effect on immunoreactivity | References |
| Milk | α-casein, α-lactalbumin | UVC treatment (15 min) | 25% α-casein reduction | [166] |
| β-lactoglobulin | UVC treatment (15 min) | 27.7% whey fractions reduction | [166] | |
| Egg | Ovalbumin | UV processing (0.61 kJ/m2) | No effect | [168] |
| Ovomucoid | UV processing (63.7 kJ/m2) | No effect | [168] | |
| Shrimp | Tropomyosin | PUV sterilization (4 min) | Reduced | [170] |
| Peanut | Ara h 1, Ara h 2, Ara h 3 | PUV treatment on butter slurry (1–3 min); raw and roasted peanuts (2–6 min) | 67% reduction IgE binding of peanut butter slurry; 12.5 folds reduction, 100% reductiom total extracts | [164] |
| Soy | Gly m5 | PUV treatment (1–6 min) | 100% reduction Gly m5 reduced | [165] |
| Gly m6 | PUV treatment (1–6 min) | Gly m6 retained | [165] | |
| Soy extracts (e.g., glycinin, β-conglycinin) | PUV treatment 2 min | 20% reduction | [164] | |
| PUV treatment 4 min | 40% reduction | [164] | ||
| PUV treatment 6 min | 50% reduction | [164] |
Table 5.
Mechanisms by which PUV affects protein structure and allergenicity.
| Mechanism | Details | Impact on Allergenicity | Citations |
| Photo-oxidation | PUV emits intense UV-C light (200–280 nm) that causes oxidative modifications, particularly in amino acids like tryptophan, tyrosine, and methionine. | Oxidative damage to amino acid residues alters the structure of allergenic epitopes. | [158] |
| Disruption of Disulfide Bonds | High-energy UV pulses can break disulfide bridges that maintain protein conformation. | Destabilization of tertiary structure reduces IgE-binding ability. | [160] |
| Epitope Modification | PUV alters conformational and linear epitopes through structural denaturation. | IgE-binding is reduced due to loss of native allergenic regions. | [68,101] |
| Protein Aggregation / Fragmentation | UV treatment may lead to cross-linking or fragmentation, depending on exposure time and intensity. | Aggregation can mask epitopes; fragmentation may eliminate allergenic potential. | [101,160,161] |
| Surface Effects | PUV has limited penetration depth (~microns); primarily impacts food surface proteins. | Effective for surface allergens; limited impact on allergens embedded within the food matrix. | [162,163] |
| Dose- and Matrix-Dependent Effects | Allergen reduction is influenced by pulse energy, duration, distance, and the optical properties (color, opacity) of the food. | Matrix composition may shield proteins or influence energy absorption, thus modulating allergenicity outcomes. | [164] |
Table 6.
Effects of gamma irradiation on food allergens.
| Food Matrix | Irradiation Dose (kGy) | Effect on Allergen | Mechanism/Observation | Reference |
| Egg (Ovalbumin) | 10–100 | Reduced allergenicity | Change in molecular weight; protein aggregation and cross-linking (disulfide bonds) | [174,177] |
| White Cake (with egg) | 10–20 | Reduced ovalbumin reactivity | Reduction in IgE binding | [174] |
| Shrimp (Tropomyosin) | 7–10 | Undetectable tropomyosin band; reduced IgE binding | Protein denaturation, aggregation; turbidity and surface hydrophobicity changes | [145] |
| Shrimp (Tropomyosin) | 1–15 + heat (100°C) | 5–30-fold reduction in IgE binding | Synergistic effect with heat treatment | [170] |
| Egg (Ovomucoid) | 10 + heat | Almost undetectable levels of ovomucoid | Irradiation more effective than heat alone due to ovomucoid heat stability | [156] |
| Tree Nuts (Almonds, Cashews, Walnuts) | 1–25 ± heat | Minimal change in allergenicity | Allergens stable under irradiation and heat | [179] |
Table 7.
Overview of recent studies on the impact of ultrasound processing on food allergenicity (adapted from [194]).
Table 7.
Overview of recent studies on the impact of ultrasound processing on food allergenicity (adapted from [194]).
| Food | Target Allergen | Treatment | Immunoreactivity | References |
| Soy | Proteins | 37 kHz, 10 min | Reduced 24% | [195] |
| Milk | α-casein | 500 W and 20 kHz (10–30 min) | No effect | [155] |
| β-lactoglobulin | 500 W and 20 kHz (10–30 min) | No effect | [195] | |
| α-lactalbumin | 500 W and 20 kHz (10–30 min) | No effect | [190] | |
| Peanut | Ara h1 | 50 Hz for 5 h | Reduced 84.8% | [196] |
| Ara h2 | 50 Hz for 5 h | Reduced 4.88% | [197] | |
| Shrimp (boiled) | Proteins | 30 kHz, 800 W (0–50 °C, 0–30 min) | Reduced 40%–50% | [197] |
| Shrimp (raw) | Proteins | 30 kHz, 800 W (0–50 °C, 0–30 min) | Reduced up to 8% | [197] |
| Crayfish | Tropomyosin | 100–800 W, 15 min | Reduced | [198] |
| Shrimp and allergens | Multiple (including tropomyosin) | 30 Hz, 800 W (30–180 min) | Reduced up to 75% | [170] |
| Crayfish | Arginine kinase | 200 W, 30 °C (10–180 min) | No effect | [189] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



