
Submitted:
05 August 2025
Posted:
06 August 2025
You are already at the latest version
Abstract
Amphistegina lobifera, a symbiont-bearing benthic foraminifera native to the Indo-Pacific, is ex-panding its distribution range in the Mediterranean Sea due to Lessepsian migration through the Suez Canal. This study investigates the colonization dynamics and ecological impact of A. lobifera on Pantelleria’ island, located in the central Mediterranean. Nine samples were collected from various substrates at different depths (-4 to –20 m) and analysed for composition, abundance, and ecological diversity. Results reveal a significant dominance of A. lobifera, especially in shallow un-vegetated substrates, with relative abundances exceeding 50%. Conversely, more vegetated sub-strates host more diverse and balanced assemblages. Diversity indexes are consistently lower where A. lobifera is dominant, indicating reduced biodiversity. Comparison with similar studies in Malta, Linosa, and Corfu confirms a 20% relative abundance threshold for A. lobifera as indicative of potential ecological imbalance. The results suggest that it represents a growing threat to native microbenthic biodiversity and reinforce its usefulness as a bioindicator species. Pantelleria represents a key transition zone for monitoring the dynamics of invasive species in the Mediterranean, and this study highlights the urgent need for long-term surveillance of benthic biodiversity.
Keywords:
benthic foraminifera
; non-indigenous species
; Amphistegina lobifera
; marine biodiversity
; Mediterranean Sea
1. Introduction
In the Mediterranean Sea the phenomenon of Lessepsian migration, through the Suez Canal, is well documented in many multicellular organisms such as fishes, jellyfishes, crustaceans, molluscs, soft corals, and algae [1,2,3,4,5,6,7]. This migration has also been favoured in recent years by the progressive warming of the sea surface waters of the eastern Mediterranean (Levantine Basin), which has made the environment more favourable for subtropical-tropical organisms of Indo-Pacific origin [8,9]. In this context, numerous species of Indo-Pacific origin have successfully established in the eastern Mediterranean and are gradually expanding towards the central Mediterranean, while they are poorly represented in the western Mediterranean. The term Non-Indigenous Species (NIS) has been introduced to indicate organism’s native to a specific geographic area that, through accidental or intentional causes, are introduced into ecosystems outside their natural range. According to the definition provided by the International Union for Conservation of Nature 2000 [10], these are species that successfully establish in natural or semi-natural habitats, acting as agents of change and posing a threat to endemic biodiversity. Such species can compete with native species, becoming invasive and causing serious ecological imbalances, and are referred to as Invasive Alien Species (IAS).
Although the presence of NIS in the Mediterranean Sea is well documented with over 1,000 species [5], only in recent years has there been growing interest from the EU and local governments, with laws and implementing measures to regulate the various aspects related to their presence and to assess any potential damage to the marine environment. At the European level, the European Union's Regulation on IAS no. 1143/2014 (EU 2014, EC 2019) represents the main regulatory framework [11] and references therein] This regulation establishes rules for the prevention and management of the introduction and spread of IAS, including marine ones. Among the NIS, some exotic species of benthic foraminifera have also been described [12,13,14,15] and references therein]. Recently, [16] published an updated list consisting of 44 species of alien benthic foraminifera introduced via the Suez Canal pathway.
Foraminifera are unicellular marine microorganisms (planktonic or benthic) that may have a carbonate or arenaceous shell. Benthic forms can exist as infaunal or epifaunal forms, the latter also including epiphytic forms. Foraminifera are essential organisms for the marine ecosystem as they constitute a fundamental link in the food chain and are widely studied as climatic, ecological, and environmental bioindicators [17,18,19]. The abundance of different species is closely linked to various environmental factors, better known as limiting factors. Among these, the most important are temperature, dissolved oxygen, pH, nutrients, turbidity, bathymetry, the presence of bottom vegetation, water transparency, etc. Finally, some polluting factors can influence assemblages by modifying specific diversity and association types [20,21,22,23,24,25].
Among benthic foraminifera, Amphistegina lobifera [26] is the most important exotic species considered as NIS as well as IAS. In particular, A. lobifera is a symbiont-bearing species, with a thick and robust carbonatic shell with dimensions up to 1-2 mm. [27] demonstrated in laboratory experiments that A. lobifera cease all movements when exposed to temperatures below 12°C. [26] reported that the occurrences of amphisteginids in modern oceans are delimited by the 14°C winter isotherm [28,29]. Today it is widely present in tropical and subtropical areas of the Indo-Pacific and the Red Sea [29,30]. It lives both as an epiphytic and epifaunal form and consumes an increasing concentration of oxygen as the temperature increases from 16 to 32°C [31]. Furthermore, in tropical areas is well known its ability to significantly contribute to the formation of biogenic carbonatic sands, which from a sedimentological point of view together with the building corals contributes to the formation of sands in the atolls [32,33]. Recently, [34] affirmed that A. lessonii, due to its high reproductive capacity to carbonate production shows an optimum at slightly elevated pCO2 levels (700 ppm).
Amphistegina lobifera was first recognized in the eastern Mediterranean in the early 2000s off the coasts of Israel [35], Malta [36], Turkey [37], Greece [38], Corfù [39] and Cyprus [40], in the islands of the Sicily Channel [13]. In recent years, it has been reaching increasingly higher percentages along the Mediterranean coasts, progressively migrating towards west. [14] described three stages of colonization for this species: i) early stage of colonization with abundance values < 20%; ii) medium stage of colonization with values between 20 and 50%; iii) advanced stage of colonization with values > 50%. A recent study [41], carried out by our team at Malta island, allowed us to predate the first appearance of A. lobifera during the Second World War, in 1943, with a strong increase starting from the 90s. A. lobifera is very abundant along the coasts of Greece [38,42], and it is spreading along the coasts of Albania [43] but to date, the low winter temperatures of the Adriatic Sea limit its spread towards the north. The progressive colonization of south-eastern Sicily has developed from an early to a medium stage of colonization, while it is well established with a medium-advanced stage of colonization in the islands located in the Sicilian Channel i.e. Pelagian and Pantelleria islands [13,14,15]. Instead, along the coasts of the Egadi Islands (the westernmost part of Sicily) it is still in the early phase of colonization, with percentages well below 5% [14]. In the southern Tyrrhenian Sea and the western Mediterranean, it has not yet been reported [13 and unpublished data]. The aim of this work is to verify the colonization process of A. lobifera at Pantelleria island in comparison with the data reported by [14], and secondly to evaluate the impact on foraminiferal biodiversity. Pantelleria’ island is located in the center of the Sicilian Channel and its geographical position between the western and eastern Mediterranean makes it an ideal site for marine biology studies, also useful for monitoring the progressive colonization of organisms of Indo-Pacific affinity from east to west. The presence of different NIS reflects the environmental fluctuations to which the water masses of the Sicilian Channel are subject and the interactions between species of Atlantic affinity from the western Mediterranean and those of Indo-Pacific affinity.
2. Study Area
2.1. Geological and Environmental Setting
The island of Pantelleria is located in the central sector of the Sicilian Channel (Central Mediterranean, Figure 1A), approximately 110 km southwest off the Sicilian coast (Cape Granitola) and 70 km northeast off the coast of north Africa (Cape Kelibia, Tunisia). Pantelleria is elliptical in shape, extending in a NW-SE direction, and is the largest of the islands in the Sicilian Channel, approximately 80 km2 and 51 km of coastline.
From a geographical and geological point of view, it represents a unique environment characterized by a complex geology, with predominantly volcanic rocky coasts and a narrow, jagged continental shelf. This island has been called the "Black Pearl" of the Mediterranean because it is entirely of volcanic origin, and therefore its landscape is typically dark in colour. Pantelleria represents the emerging tip of a complex volcanic edifice, mostly submerged, which reaches a depth of approximately 1,200 meters and is located in the axial part of the Sicilian Channel Rift. The first submarine volcanic activity that gave rise to the island dates back to approximately 1 million years ago, then the island emerged stably around 300,000 years ago with the onset of the first explosive subaerial volcanic activity. The oldest volcanic rocks that outcrop have an age of approximately 324 ka [44] and vary in composition from weakly alkaline basalts to trachytes and rhyolites with peralkaline affinity [45]. Pantelleria is dominated by Montagna Grande, a relief 836 meters above sea level which is a volcanic caldera formed approximately 45 ka [44,46]. From a lithological point of view, the most common rocks are pantellerites, basalts, green tuff and pyroclastic deposits. One of the most recent episodes of eruptive activity occurred in 1831 in underwater environment, north-west of the island, and created a small island known as Isola Ferdinandea [47]. This remained above sea level for only eight months and then disappeared completely in 1832, forming a vast rocky platform (Graham Bank).
Today, it lies eight meters below sea level and is a rich fishing area, recognized as a biodiversity hotspot thanks to the presence of many protected pelagic and benthic marine species. Pantelleria’ island is protected as "Pantelleria Island National Park," established in 2016 and covering 80% of its territory. The park covers only the land area, not the marine area. It constitutes a highly complex ecological system, characterized by significant biodiversity, both terrestrial and marine. The marine-coastal environment, characterized by volcanic rocky substrates, submerged reefs, Posidonia oceanica meadows, marine algae and, more rarely, small sandy areas, provides an ideal habitat for a wide variety of marine organisms. Here, the marine environment is characterized by an excellent degree of transparency of the waters, as reported by [15] at Linosa Island. Transparency permits light to reach deeper depths, allowing symbiont-bearing organisms to extend their habitat.
Furthermore, the waters surrounding the island are influenced by the interaction between Atlantic and Levantine water masses, giving the area particularly dynamic oceanographic characteristics. The local marine climate is typically warm-temperate, but in recent decades it has shown signs of tropicalization, consistent with trends observed in other areas of the southern Mediterranean. Generally, in recent years, the Sea Surface Temperature (SST) of the central Mediterranean, between Sicily and North Africa, has fluctuated between 15°C and 29°C [48].
In particular, SST data show that the coasts of south-western Sicily are characterized by more or less stable low temperatures during the winter (15-16°C), while they are characterized by wide oscillations during the summer (17-26°C). However, in the south-eastern part of Sicily and in the Sicilian Channel (around Pantelleria, Pelagian Islands, and Malta), average summer temperatures are higher and more stable with smaller oscillations.
2.2. Exotic Species in the Pantelleria’ Island
From the point of view of terrestrial biodiversity and the presence of exotic species, Pantelleria has been the object of numerous studies [49,50,51,52,53,54,55]. The marine algal flora shows strong North African affinity [56,57,58,59,60]. As regards exotic marine organisms, several species have been described, such as Rhopilema nomadica [61], Portunus segnis [61], the algae Caulerpa cylindracea and Caulerpa taxifolia [63], Parupeneus forsskali [64]. On the contrary, as regards marine microorganisms (i.e. benthic foraminifera), the work of [14], is the only study focused on benthic foraminiferal assemblages in this island. In particular, these authors reported the presence of NIS and several cryptogenic species. They recorded A. lobifera at all 4 study sites (sampled in 2017) with abundances ranging from 2% to 82%, even if in that study the authors reported only the total assemblages not discriminating dead and living specimens. Thus, probably the percentages were overestimated hypothesizing a stage of colonization from early to advanced. In this study we fix better this evaluation.
3. Materials and Methods
In October 2024, a scuba diving campaign was conducted on the seabed of Pantelleria. Starting from the northeast and proceeding clockwise, three sampling sites were selected (Gadir, Cala Tramontana, and Balata dei Turchi, respectively; Figure 1, Figure 2, Figure 3 and Figure 4).
Two to four algal and/or sediment samples were collected at each site at different depths. A total of nine samples were collected: eight algal samples, and one marine sediment sample. Table 1 shows the geographic coordinates of the sampling stations, their depths, and the type of sample collected. Sea Surface Temperature and salinity were also measured at the three sampling sites (Table 1). Underwater photographs were taken with GOPRO 8 camera.
The samples were collected at depths between 4 and 20 meters, divided into four aliquots, and stored in cylindrical polyethylene containers (Falcon). One aliquot was preserved as it is for possible sedimentological and/or geochemical analyses. Another aliquot was treated with a buffered Bengal Rose solution (2 g of Bengal Rose in one liter of ethanol) to differentiate living (coloured) benthic foraminifera from those already dead (uncoloured)at the time of sampling [65]. Two aliquots of each sample were also treated with two different chemical products (Qiagen PowerProtect DNA/RNA reagent and RNAlaterTM Solution Invitrogen by Thermo Fischer Scientific) to preserve both DNA and RNA and subsequently carry out genetic studies on living foraminifera.
The samples were stored in a cool bag, then transported to the laboratory and refrigerated until ready for analysis.
The algal species were identified in situ and subsequently rechecked in the laboratory for taxonomic identification.
Following the FOBIMO protocol [66], after 14 days each Bengal Rose treated sample was removed from the polyethylene container, gently washed on a 63-µm mesh sieve, and placed in an oven to dry for 24 hours at 40°C. The algal samples were prepared by carefully removing any benthic foraminifera attached to the algae or P. oceanica rhizomes. The samples were then placed in small plastic containers, labelled, and stored until microscopic analysis.
In the first phase, a qualitative analysis was performed on the samples using a binocular microscope, and the benthic foraminiferal species present were recognized and classified following [30,67,68,69]. Regarding the genus Amphistegina, the presence of a particular morphotype was observed, similar to A. lessonii but characterized by a very small size, a very flattened spiral side, and a pronounced carina, which was identified, according to [15], as Amphistegina morphotype alfa. This morphotype also corresponds to Amphistegina cf. lessonii, first reported in [13]. The genus Amphistegina have been photographed both by using a Leica optical system and a SEM PHENOM PROX (ATeN Center, Università di Palermo).
Subsequently, quantitative analysis was performed on benthic foraminifera. Each sample was split into smaller aliquots using an Otto microsplitter. The splitted fraction was weighed and all the individuals present were counted, distinguishing those alive at the time of sampling (coloured) from those dead (uncoloured).
To evaluate the structure of the benthic foraminiferal community, four diversity indexes were calculated using the Paleontological Statistics Data Analysis (PAST) software [70]: 1) Species richness (S), i.e. the number of species in each sample; 2) Dominance (D), an index ranging from 0 to 1 that indicates how single species are distributed within the assemblages; 3) Shannon index (H), which measures both the richness (number of species) and the evenness (distribution of abundances) of a biological community; 4) Fisher-α index, i.e. the relationship between the number of species and the number of individuals in an assemblage [71,72].
4. Results
In the studied samples, the assemblages are dominated by living foraminifera and the dead specimens constitutes only a small percentage of the total assemblage (Table 2). In some cases, the assemblages were only characterized by living specimens (PANT 24-3-1A, PANT 24-3-1 and PANT 24-3-2). Moreover, in the other samples dead specimens constitute less than 6% of the total assemblage except for the samples PANT 24-3-3 and PANT 24-5-2, 20.81% and 27.93%, respectively. For this reason, we decided to focus our attention on the living assemblages.
A total of 30 species belonging to 23 genera were recognized. In Table 3 we have reported only the living benthic foraminiferal species (29), not including Elphidium sp. as only one dead individual was found in the studied samples.
Relative abundances (percentages) of all the living species recorded are reported in Table 3 as numerical values, while the most relevant species are reported as histograms in Figure 5. The analysis of the association highlights an overall community dominated by a small number of species, with strong variability between samples in relation to depth and type of substrate. Among the most abundant species, Amphistegina lobifera stands out, with percentages ranging between 7.3% (PANT 24-4-3, -20m) and 69.9% (PANT 24-5-2, -5m). Its dominance is particularly marked in samples with shallower bathymetries. Amphistegina lessonii is present in all samples, with percentages ranging from 4.5% to 31.4% (PANT 24-3-1A, -5 m) and is abundant at deeper depths. Amphistegina morphotype alfa is present in only five samples, with very low percentages, ranging from 0.4% (PANT 24-4-2, -9 m) to 1.69% (PANT 24-3-3, -17.5 m).
Among the native and common species in the Mediterranean Sea, Miliolids are always present. Among these, Miliolinella subrotunda shows significant percentages in almost all samples (minimum value 2.4% at PANT 24-5-2 and maximum value 18.9% in PANT 24-4-3). Peneroplis pertusus is present in eight samples with values ranging from 3.2% at PANT 24-3-2 to 16.2% in PANT 24-5-1. Laevipeneroplis sp. is present in only three samples with the highest percentage at PANT 24-3-1A (8.1%). Individuals belonging to the genus Quinqueloculina are characterized by variable abundances that, however, never exceed 7.5%.
Textularia pala, a foraminifer with an agglutinating shell, is present in all samples, with percentages ranging from 1.4% at PANT 24-3-2 to 16.1% at PANT 24-3-3.
Rosalina obtusa and Asterigerinata mamilla, typical epiphytic species, are almost always present, but with percentages below 6.3% and 11.7%, respectively. Other species, such as Adelosina sp., Lobatula lobatula, Planorbulina acervalis and Vertebralina striata, are present only at particular sites with highly variable abundances.
The samples have also been analysed to assess the diversity and structure of the foraminiferal community. The results obtained using PAST software highlight some variability among samples in terms of Species Richness (S), Dominance (D), Shannon Index (H'), and Fisher's α (Figure 6).
The number of taxa (Species Richness, S) ranges from 11 to 21 species, with the highest values found in samples PANT 24-4-1 (S = 21) and PANT 24-4-2 (S = 19), suggesting a more diverse community. In contrast, samples PANT 24-5-1 and PANT 24-5-2 show the lowest species richness (S = 11 and 12, respectively). The dominance index (D), which expresses the degree of dominance of a single species, ranges from 0.109 to 0.502. The lowest values (D ≈ 0.11–0.14) correspond to samples PANT 24-4-2, PANT 24-4-1, and PANT 24-3-1A and indicate more evenly distributed communities, while the PANT 24-5-2 sample (D = 0.502). Shannon index (H′) values range between 1.237 and 2.512, with the most diverse samples always corresponding to those with low dominance (i.e., PANT 24-4-2, H′ = 2.512). Samples with H′ < 1.6 (e.g., PANT 24-5-2 and PANT 24-3-2) instead show low diversity, with an association dominated by a few taxa.
Finally, the Fisher's α index shows values between 2.38 and 5.75, confirming the greater specific heterogeneity of the PANT 24-4-1 (α = 5.75) and PANT 24-3-1A (α = 5.60) samples and a simpler structure in the high-dominance samples.
Overall, the results highlight a consistent correlation between the four indexes analysed: the samples on algal substrates show greater diversity and more even distribution, while the sediment sample PANT 24-5-2 is characterized by lower diversity and is dominated by a few high-dominance species (D = 0.502).
5. Discussions
5.1. Benthic Foraminiferal Assemblages and Diversity Indexes
Benthic foraminiferal analysis highlights an assemblages dominated by a limited number of species, with a distribution strongly influenced by substrate type, depth, and the presence of vegetation, dominated by the genus Amphistegina. In particular, A. lobifera is the dominant species, which is widely represented in almost shallow sites, with values exceeding 50%, essentially attached as epiphyte to the Posidonia oceanica rhizomes and Halopteris scoparia, and in the sandy substrate as an epifaunal species. A. lobifera is associated with A. lessonii and Amphistegina morphotype alfa, albeit in lower percentages. The highest percentages of A. lobifera occur in samples with bathymetries lower than 11 meters and in substrates where P. oceanica is strongly degraded, due to the high number of summer anchorages that damage the sites where there is a high summer tourist influx. This type of damage also facilitates predation by the fish species Sarpa salpa and the echinoderm Paracentrotus lividus, which feeds on P. oceanica. In these cases, the predominance of A. lobifera corresponds to a drastic reduction in species diversity, highlighted by ecological indexes (H′ and Fisher's α), suggesting a significant ecological impact on the local foraminiferal assemblage. Conversely, in samples with deeper bathymetries, the species' abundance is lower, and the foraminiferal assemblage is more balanced, presumably also favoured by greater variability of plant and algal species. In these environments, A. lobifera coexists with other species without significantly altering the overall composition of the assemblage. Amphistegina lessonii is present in all samples with variable percentages, suggesting a possible competitive coexistence with both A. lobifera and the morphotype alfa with partially overlapping microenvironmental preferences.
In particular, A. lessonii is one of the most abundant species in the deeper algal samples (31.4% in PANT 24-3-3 and 19.6% in PANT 24 4-3, at -17.5 and -20 m depth, respectively; Figure 7, photo A).
This pattern suggests that A. lessonii tends to be dominant in similar, but deeper environments, and tends to occupy complementary niches. The presence of Amphistegina morphotype alfa is marginal, with percentages ≤1.7%, indicating a presence with no significant impact on the structure of the assemblages. From a taxonomic point of view, this morphotype cannot be clearly classified and therefore it requires greater attention through genetic analysis. This will be further developed in the future, also thanks to the 18SrDNA analyses currently being conducted at the STEBICEF laboratories under our supervision.
Analysis of diversity indexes reveals patterns consistent with literature regarding foraminiferal community structure. Diversity indexes highlighted considerable variability in foraminiferal community structure, likely reflecting the different environmental characteristics of the substrates. The living benthic foraminiferal assemblage shows high variability in terms of dominance and species richness, with a clear distinction between samples collected on vegetated substrates and those on incoherent sediments or sandy substrates.
The dominance index (D) highlighted a greater concentration of abundance of a few species at some sites, indicating less structured communities dominated by opportunistic taxa, including A. lobifera. Conversely, high values of the Shannon index (H') and Fisher's α in other samples reflect more diverse and uniformly distributed communities, suggesting more stable and/or less disturbed environments. Samples taken from vegetated substrates show high values of Species Richness (S), Shannon index (H'), and Fisher's α, associated with low dominance (D). This suggests that the presence of vegetation contributes for creating a more stable and complex microhabitat, capable to support a more diverse and balanced community. Overall, samples collected from sites with abundant vegetation show a more heterogeneous benthic foraminiferal community, with abundances distributed among different species, while samples on sandy substrates without vegetation are characterized by a high dominance of A. lobifera (Figure 7, photos B, C, D, E), accompanied by an overall reduction in diversity indexes. In contrast, the sediment sample PANT 24-5-2 (-5m) is characterized by a strong dominance (D > 0.5), low diversity (H′ < 1.3), lower species richness (S = 12), and reduced Fisher's α values, indicating an ecologically simplified community. In particular, the abundant presence of A. lobifera in this latter sample (69.86%) is indicative of the competitive and colonizing capacity of this non-native species, whose expansion in the eastern and central Mediterranean Sea is now well documented [14,15,73].
This pattern confirms the structuring role of the substrate type and the invasiveness of the Lessepsian species in the organization of the benthic foraminiferal communities on the island. Less stable sedimentary conditions, devoid of vegetation, subject to wave energy, and highly transparent waters that favour the penetration of sunlight facilitate the dominance of A. lobifera, a symbiont-bearing, opportunistic, and highly adaptable species, which tends to reduce community complexity and modify microbenthic trophic chain. Conversely, vegetated substrates offer shade, stability, and greater roughness, elements that favour the coexistence of multiple taxa and prevent, at least partially, the expansion of invasive taxa. In some sites, summer anchoring causes a high level of deterioration of P. oceanica meadow, this triggers an increase of leaf predation leaving the rhizomes exposed, which are promptly colonized by Amphistegina.
The preference of A. lobifera in well-lit waters and poor vegetated sand substrates and is consistent with data recorded in the Pelagian and Malta Islands, where they dominate surface sediments, actively contributing to biogenic carbonate [14,15]. During scuba divers and sampling, we have observed that the presence of A. lobifera is lower where P. oceanica meadow is well developed without deterioration. In particular, in samples where there was a good vegetation higher species equity has been observed, with an enrichment of native taxa such as Miliolinella subrotunda, Peneroplis pertusus, Quinqueloculina spp., and Rosalina obtusa. This distribution suggests that the presence of benthic macrophytes acts as a stabilizing factor, capable for limiting the dominance of invasive species and promoting more diverse communities. Conversely, in unstable or more degraded environments, interspecific competition is unbalanced in favour of A. lobifera, whose high abundance is also linked to a reduction in diversity indexes and the alteration of the benthic ecological balance [15,74]. Furthermore, it is important to highlight the presence in all samples of Textularia pala, a foraminifer with an agglutinating shell. The genus Textularia is generally reported in the literature, in the Mediterranean Sea, as a typical form found in sandy and detrital bottoms in the infralittoral and circalittoral zones [75] but also as epiphytic species in P. oceanica rhizomes [76]. In our samples it is always present and abundant, probably due to the high energy of the marine environment which bring into suspension minerals (quartz, feldspars) and clasts deriving from the volcanic rocks erosion which are used to build its agglutinant test.
These results indicate that algal vegetation acts as an ecological buffer, supporting more balanced communities in the presence of P. oceanica meadows or macroalgae (PANT 24-3-3, PANT 24-4-3), while less structured substrates (sediments) favour the dominance of A. lobifera. This is consistent with observations in other areas of the Mediterranean, where species prevalence above 20% is associated with a decline in the specific diversity of benthic foraminifera [15].
These findings confirm that foraminiferal diversity can be used as a sensitive bioindicator to assess the ecological status of benthic seabeds and to monitor the presence and impact of alien species, such as A. lobifera, in Mediterranean coastal environments, including islands like Pantelleria.
In Pantelleria, A. lobifera percentages exceed 20% in 8 out of 9 samples, a threshold considered in the literature to be a potential indicator of ecological imbalance in coastal environments [15,74].
This suggests that the island of Pantelleria, although characterized by a still complex and diverse benthic habitat, is already affected by active bioinvasion by A. lobifera.
5.2. Benthic Foraminiferal Assemblages and Diversity Indexes
The expansion of A. lobifera in the Mediterranean is a well-documented phenomenon, with differential impacts on the structure of benthic communities depending on the relative presence of the species, the nature of the substrate and bathymetry. The results obtained fit coherently with the colonization of A. lobifera in other site of the eastern Mediterranean Sea, a Lessepsian species now widely documented in shallow marine environments in the eastern and central sectors of the basin. The previous studies carried out in the Pelagian Islands and the eastern Aegean Sea have highlighted similar ecological patterns. In the Pelagian islands, A. lobifera was reported by [15] as one of the most abundant species in sandy surface sediments, where it tends to progressively replace the native component, particularly in sites with low vegetation coverture. Sediments collected off the coast of Linosa island show a marked sedimentary "switch," from a dark volcanic matrix to almost entirely white biogenic sediments, due to the accumulation of calcareous shells of A. lobifera. This process has led to the formation of microenvironments similar to those observed in tropical regions, where this species is known for its ability to produce large-scale carbonate sediments. This phenomenon has also been observed at underwater sites off of Pantelleria, although this island does not favour the formation of sand beaches along the coastline due to the particular geological conformation, where biogenic sand accumulations can occur few meters below the sea level. In fact, in areas where the P. ocenica meadow is severely degraded, significant accumulations of Amphistegina shells are forming above the rhizomes, creating biogenic sand. In the Aegean Sea, [38,74] found that the massive presence of A. lobifera is strongly correlated with sandy substrates and well-lit environments, with critical dominance values observed starting from thresholds >20%. In particular, [74] proposed an ecological threshold of 20% in samples off the coasts of Corfù (Greece), that is the relative abundance beyond which a statistically significant effect on the diversity and balance of microbenthic communities is observed. In detail, this study demonstrated that A. lobifera values above 20% are associated with a significant reduction in the Shannon index (H′), an increase in the dominance index (Berger–Parker), a partial reduction in Fisher’s α and in Species Richness, although less marked.
The data obtained from Pantelleria are fully consistent with these observations. In eight of the nine analysed samples, A. lobifera exceeds the threshold of 20%, with a peak of 69.86% in sample PANT 24-5-2 (sedimentary substrate). In these samples, H′ values were lower than 1.6, consistent with strong taxonomic dominance, dominance (D) values up to 0.50, and low Fisher's α, confirming the ecological pressure exerted by the species, especially in the absence of vegetated substrates. In samples with a presence lower than 20% (i.e., PANT 24-4-3, 7.3%), diversity indexes were more balanced, and the benthic community maintained a higher level of diversity, confirming the role played by algal and vegetated substrates.
This convergence between the data from Corfù and Pantelleria supports the hypothesis that the 20% threshold value represents a reliable indicator of potential ecological imbalance linked to the invasiveness of A. lobifera and suggests the usefulness of this parameter as an early warning criterion in benthic biomonitoring programs in Mediterranean areas subject to colonization by Lessepsian species.
Comparison of data from Pantelleria with those from Pelagian Islands and the Aegean Sea suggests that A. lobifera responds consistently to the same favourable environmental conditions, displaying synergistic invasive behaviour throughout the central-eastern Mediterranean basin. Its progressive expansion along the south-western coasts of Sicily and in the Sicilian Channel clearly demonstrates the process of "tropicalization" of the Mediterranean, triggered by the opening and widening of the Suez Canal and the increase in Mediterranean SSTs over the past 30 years, especially in the eastern Mediterranean.
Since 1993, ocean SSTs have increased globally by an average of more than 0.4 (± 0.02)°C, while in the Mediterranean they have increased by about 1.5°C (± 0.02) [77]. In this context, Pantelleria represents an ecological transition area of particular interest, where the coexistence of indigenous and Lessepsian species reflects ongoing dynamics of competition and adaptation. Long-term monitoring of benthic communities and assessment of the impact of the A. lobifera invasion are therefore essential to understanding the evolutionary trajectory of these ecosystems. The comparison with Pantelleria highlights how, despite the island's geological and environmental peculiarities (volcanic substrates, variable depths, presence of photophilous macroalgae), the patterns of microhabitat dominance and preference of A. lobifera are replicable and comparable with other areas of the central and eastern Mediterranean. The presence of favourable habitats (shallow depth, incoherent sediments, water transparency) makes the area vulnerable to the persistence and potential expansion of this species. At the same time, the presence of benthic macrophytes and the structural complexity of some microhabitats appear to exert a stabilizing effect, favouring more heterogeneous communities and reducing the absolute dominance of A. lobifera, as also observed in Linosa and Corfù islands.
The distribution and abundance of the genus Amphistegina closely matches the surface temperatures recorded on the website [77].
The areas around southern Sicily, characterized by strong summer temperature variations linked to current circulation and atmospheric conditions (mistral storms causing upwelling of colder deep water), likely limit their distribution during the summer and, to date, prevent their spread and colonization in the Tyrrhenian area. Although they can tolerate temperatures of 14-15°C, they are unable to proliferate at these low temperatures. Throughout the year, temperatures around Pantelleria’ island range from 15°C to 28-29°C, with higher and more stable average values during the summer compared to the coasts of southern Sicily. These high temperatures (Figure 8) favour the development of the genus Amphistegina which reaches an advanced stage of colonization (Figure 9).
Biological invasions are currently one of the main drivers of habitat degradation and a major cause of biodiversity loss in both marine and terrestrial ecosystems. In particular, in the Mediterranean Sea the spread of invasive species of Lessepsian origin poses a particularly serious environmental threat to islands, which are biodiversity hotspots and host peculiar habitats and endemic species, especially small islands that are highly vulnerable to biodiversity loss. For this reason, several European directives have been enacted to address this issue. The Marine Strategy Framework Directive (MSFD) 2008/56/EC [78] considers the presence of non-native species as one of the 11 descriptors of environmental quality (Descriptor 2: "Introduced non-native species"). The Directive requires Member States to ensure that "the introduction of non-native species is minimized and that these species do not cause negative impacts on marine ecosystems." Furthermore, Regulation (EU) No. 1143/2014 [79] states that "After the introduction of an invasive alien species, early detection and rapid eradication measures are crucial to prevent their establishment and spread". In Italy, the implementation of these regulations has led to the development of monitoring plans made by the Regional Agency for Environmental Protection (ARPA), and the Higher Institute for Environmental Protection and Research (ISPRA) and the collection of data through networks such as the International Commission for Scientific Exploration of the Mediterranean (CIESM) or the European Alien Species Information Network (EASIN).
In addition, to complement traditional monitoring efforts, citizen science is increasingly being adopted. In this context, the use of Local Ecological Knowledge (LEK) has proven effective for tracking non-native species [80].
The crucial point is that many species are also commercially exploited, and it is difficult to initiate an eradication process, while on the other hand, it is even more difficult, if not impossible, to eradicate microorganisms such as foraminifera i.e. the genus Amphistegina. Their spread, coupled with rising temperatures, may pose a serious threat to the future of typical Mediterranean species in the coming decades. On the other hand, the increase of the genus Amphistegina in the Mediterranean could increase the production of carbonate in sea surface waters, counteracting the acidification processes due to the increase of CO2 in the atmosphere.
What we are witnessing is essentially a true biological revolution, with ecosystems undergoing the fastest changes after the last glacial maximum.
6. Conclusions
This study confirms the active colonization of the non-native benthic foraminifera Amphistegina lobifera in the coastal environments of Pantelleria’ island, providing new data on its environmental preferences and its impact on native benthic communities. Analysis of living benthic foraminiferal assemblages highlighted the increasing incidence of A. lobifera, whose ecological success appears to be favoured by local environmental conditions such as incoherent sediments, well-lit waters and substrates few vegetated. The consistent association between high abundances of A. lobifera and low biodiversity indexes (Shannon index, Fisher's α, species richness) highlights its potential role as a factor in ecological imbalance in microbenthic communities. In particular, sedimentary substrates devoid of vegetation are more vulnerable to invasion, while habitats with algae and Posidonia oceanica, where the associations exhibit greater equality and a significant representation of native species, appear to mitigate the dominance of this invasive species, promoting more diverse and stable associations. The presence of A. lobifera in all analysed samples, with abundances exceeding the ecological threshold of 20% in 8 of 9 samples, with peaks higher that 68%, indicates that on Pantelleria it is no longer in the initial phase of colonization, but rather in an intermediate or advanced stage. Comparison with other Mediterranean contexts (i.e., the Pelagian Islands, the Aegean Sea) confirms that the dynamics here observed are part of a broader process of tropicalization of the Mediterranean Sea, triggered by the construction of the artificial Suez Canal and favoured by maritime traffic the and the progressive warming of eastern Mediterranean surface waters that facilitated the migration of Indo-Pacific organisms.
In this context, A. lobifera is confirmed as an effective bioindicator of ongoing changes, both from ecological and sedimentological point of view, as it causes significant changes in both local endemic biodiversity and coastal sedimentation in areas where it is proliferating due to the massive accumulation of its carbonate shells.
Thus, the island of Pantelleria represents a biogeographical frontier area of particular importance for the Mediterranean Sea for studying the interactions between native and invasive species. Continuous monitoring of these communities and the integration of ecological and sedimentological approaches will be crucial for understanding the future evolution of Mediterranean benthic ecosystems and for guiding potential management and conservation strategies, however challenging. Given the limited possibilities for controlling or eradicating invasive microbial species, long-term monitoring and conservation of vegetated substrates become essential tools for mitigating their ecological impact. The results obtained highlight the urgent need to integrate benthic foraminifera data into broader marine monitoring programs and to use diversity indexes as early warning indicators of bioinvasion in Mediterranean coastal ecosystems.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, C.C. and A.C.; methodology, C.C. and A.C..; software, C.C..; validation, C.C. and A.C.; formal analysis, C.C. and A.C.; investigation, C.C. and A.C; resources, C.C. and A.C.; data curation, C.C. and A.C.; writing—original draft preparation, C.C..; writing—review and editing, A.C.; visualization, C.C. and A.C.; supervision, C.C. and A.C.; project administration, C.C. and A.C.; funding acquisition, C.C. and A.C. All authors have read and agreed to the published version of the manuscript.”
Funding
This research was funded by Claudia Cosentino FFR 2024 (Fondo Finalizzato Ricerca di Ateneo, UNIPA) and PRJ 0686 (Antonio Caruso).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data presented in this study have been inserted in the manuscript. Further inquiries can be directed to the Corresponding Author (C.C.).
Acknowledgments
The authors wish to thank the Ente Parco Nazionale “Isola di Pantelleria” for authorizing the sampling and, in particular, Andrea Biddittu for helpful discussions on the marine environment of Pantelleria Island. The authors are also grateful to the ATEN Center - University of Palermo for support during the acquisition of SEM photographs of foraminifera.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Azzurro, E.; Carnevali, O.; Bariche, M.; Andaloro, F. Reproductive features of the nonnative Siganus luridus (Teleostei, Siganidae) during early colonization at Linosa Island (Sicily Strait, Mediterranean Sea). J. Appl. Ichthyol. 2006, 23, 640–645. [Google Scholar] [CrossRef]
- Lodola, A.; Savini, D.; Occhipinti-Ambrogi, A. Alien species in the Central Mediterranean Sea: the case study of Linosa Island (Pelagian Islands, Italy). Biol. Mar. Mediterr. 2012, 19, 257–258. [Google Scholar]
- Bariche, M.; Torres, M.; Azzurro, E. The presence of the invasive Lionfish Pterois miles in the Mediterranean Sea. Mediterr. Mar. Sci. 2013, 14, 292–294. [Google Scholar] [CrossRef]
- Ahnelt, H. Translocations of tropical and subtropical marine fish species into the Mediterranean. A case study based on Siganus virgatus (Teleostei: Siganidae). Biology 2016, 71, 952–959. [Google Scholar] [CrossRef]
- Zenetos, A.; Albano, P.G.; López Garcia, E.; Stern, N.; Tsiamis, K.; Galanidi, M. Established non-indigenous species increased by 40% in 11 years in the Mediterranean Sea. Mediterr. Mar. Sci. 2022, 23. [Google Scholar] [CrossRef]
- Nativ, H.; Galili, O.; Almuly, R.; Einbinder, S.; Tchernov, D.; Mass, T. New record of Dendronephthya hemprichi (Family: Nephtheidae) from Mediterranean, Israel - an evidence for tropicalization? Biology 2023, 12, 1220. [Google Scholar] [CrossRef]
- Tsirintanis, K.; Sini, M.; Ragkousis, M.; Zenetos, A.; Katsanevakis, S. Cumulative Negative Impacts of Invasive Alien Species on Marine Ecosystems of the Aegean Sea. Biology 2023, 12, 933. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Morri, C. Global sea warming and “tropicalization” of the Mediterranean Sea: biogeographic and ecological aspects. Biogeographia 2003, 24, 319–327. [Google Scholar] [CrossRef]
- Raitsos, D.E.; Beaugrand, G.; Georgopoulos, D.; Zenetos, A.; Pancucci-Papadopoulou, M.A.; Theocharis, A.; Papathanassiou, E. Global climate change amplifies the entry of tropical species into the Eastern Mediterranean Sea. Limnol. Oceanogr. 2010, 55, 1478–1484. [Google Scholar] [CrossRef]
- IUCN. Guidelines for the Prevention of Biodiversity Loss caused by Alien Invasive Species. In Proceedings of the Fifth Meeting of the Conference of the Parties to the Convention on Biological Diversity, Nairobi, Kenya, 15–26 May 2000. [Google Scholar]
- Mannino, A.M.; Balistreri, P. Invasive alien species in Mediterranean Marine Protected Areas: the Egadi Islands (Italy) case study. Biodiversity 2021. [Google Scholar] [CrossRef]
- Yokeş, M.B.; Meriç, E.; Avşar, N. On the presence of alien foraminifera Amphistegina lobifera Larsen on the coasts of the Maltese Islands. Aquat. Invasions 2007, 2, 439–441. [Google Scholar] [CrossRef]
- Caruso, A.; Cosentino, C. The first colonization of the Genus Amphistegina and other exotic benthic foraminifera of the Pelagian Islands and South-Eastern Sicily (Central Mediterranean Sea). Mar. Micropaleontol. 2014a, 111, 38–52. [Google Scholar] [CrossRef]
- Guastella, R.; Marchini, A.; Caruso, A.; Cosentino, C.; Evans, J.; Weinmann, A.; Langer, M.; Mancin, N. “Hidden invaders” conquer the Sicily Channel and knock on the door of the Western Mediterranean Sea. Estuar. Coast. Shelf Sci. 2019, 225, 106234. [Google Scholar] [CrossRef]
- Cosentino, C.; Guastella, R.; Mancin, N.; Caruso, A. Spatial and vertical distribution of the genus Amphistegina and its relationship with the indigenous benthic foraminiferal assemblages in the Pelagian Archipelago (Central Mediterranean Sea). Mar. Micropaleontol. 2024, 188, 102344. [Google Scholar] [CrossRef]
- Stulpinaite, R.; Hyams-Kaphzan, O.; Langer, M. Alien and cryptogenic Foraminifera in the Mediterranean Sea: A revision of taxa as part of the EU 2020 Marine Strategy Framework Directive. Mediterr. Mar. Sci. 2020, 21, 719–758. [Google Scholar] [CrossRef]
- Sen Gupta, B.K. Foraminifera in marginal marine environments. In Modern Foraminifera; Sen Gupta, B.K., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1999; pp. 141–159. [Google Scholar]
- Scott, D.B.; Medioli, F.S.; Schafer, C.T. Monitoring in Coastal Environments Using Foraminifera and Thecamoebian Indicators; Cambridge University Press: Cambridge, UK, 2001; p. 177. [Google Scholar]
- Murray, J.W. Ecology and Applications of Benthic Foraminifera; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Yanko, V.; Kronfeld, J.; Flexer, A. Response of benthic foraminifera to various pollution sources: implications for pollution monitoring. J. Foraminifer. Res. 1994, 24, 1–17. [Google Scholar] [CrossRef]
- Frontalini, F.; Buosi, C.; Da Pelo, S.; Coccioni, R.; Cherchi, A.; Bucci, C. Benthic foraminifera as bio-indicators of trace element pollution in the heavily contaminated Santa Gilla lagoon (Cagliari, Italy). Mar. Pollut. Bull. 2009, 58, 858–877. [Google Scholar] [CrossRef]
- Caruso, A.; Cosentino, C.; Tranchina, L.; Brai, M. Response of benthic foraminifera to heavy metal contamination in marine sediments (Sicilian coasts, Mediterranean Sea). Chem. Ecol. 2011, 27, 9–30. [Google Scholar] [CrossRef]
- Cosentino, C.; Pepe, F.; Scopelliti, G.; Calabrò, M.; Caruso, A. Benthic foraminiferal response to trace element pollution. The case study of the Gulf of Milazzo, NE Sicily (Central Mediterranean Sea). Environ. Monit. Assess. 2013, 185, 8777–8802. [Google Scholar] [CrossRef]
- Machain-Castillo, M.L.; Ruiz-Fernández, A.C.; Alonso-Rodríguez, R.; Sanchez-Cabeza, J.A.; Gío-Argáez, F.R.; Rodríguez-Ramírez, A.; Villegas-Hernández, R.; Mora-Garcíad, A.I.; Gómez-Ponce, M.A.; Pérez-Bernal, L.H. Anthropogenic and natural impacts in the marine area of influence of the Grijalva - Usumacinta River (Southern Gulf of Mexico) during the last 45 years. Mar. Pollut. Bull. 2020, 156, 111245. [Google Scholar] [CrossRef]
- Bergamin, L.; Pierfranceschi, G.; Romano, E. Anthropogenic impact due to mining from a sedimentary record of a marine coastal zone (SW Sardinia, Italy). Mar. Micropaleontol. 2021, 169, 102036. [Google Scholar] [CrossRef]
- Larsen, A.R. Studies of recent Amphistegina, taxonomy and some ecological aspects. Isr. J. Earth Sci. 1976, 25, 1–26. [Google Scholar]
- Zmiri, A.; Kahan, D.; Hochstein, S.; Reiss, Z. Phototaxis and thermotaxis in some species of Amphistegina (Foraminifera). J. Protozool. 1974, 21, 133–138. [Google Scholar] [CrossRef]
- Langer, M.R.; Hottinger, L. Biogeography of selected “larger” foraminifera. Micropaleontology 2000, 46, 105–126. [Google Scholar]
- Langer, M.R. Foraminifera from the Mediterranean and the Red Sea. In Aqaba-Eilat, the Improbable Gulf; Por, F.D., Ed.; Magnes Press: Jerusalem, 2008; pp. 397–415. [Google Scholar]
- Hottinger, L.; Halicz, E.; Reiss, Z. Recent Foraminiferida from the Gulf of Aqaba, Red Sea. Dela SAZU, Classis IV, Historia Naturalis, Vol. 33; Slovenska akademija znanosti in umetnosti: Ljubljana, 1993. [Google Scholar]
- Palme, T.; Nagy, M.; Heinz, P. Quantifying rates of oxygen production and consumption in the benthic foraminifer Amphistegina lobifera at different temperatures. Mar. Biol. 2025, 172, 101. [Google Scholar] [CrossRef]
- Langer, M.R.; Silk, M.T.; Lipps, J.H. Global ocean carbonate and carbon dioxide production; the role of reef Foraminifera. J. Foraminifer. Res. 1997, 27, 271–277. [Google Scholar] [CrossRef]
- Resig, J.M. Age and preservation of Amphistegina (foraminifera) in Hawaiian beach sand: implications for sand turnover rate and resource renewal. Mar. Micropaleontol. 2004, 50, 225–236. [Google Scholar] [CrossRef]
- Dämmer, L.K.; Ivkić, A.; de Nooijer, L.; Renema, W.; Webb, A.E.; Reichart, G.-J. Impact of dissolved CO2 on calcification in two large, benthic foraminiferal species. PLoS ONE 2023, 18, e0289122. [Google Scholar] [CrossRef]
- Hyams, O.; Almogi-Labin, A.; Benjamini, C. Larger foraminifera of the south-eastern Mediterranean shallow continental shelf off Israel. Isr. J. Earth Sci. 2002, 51, 169–179. [Google Scholar] [CrossRef]
- Yokeş, M.B.; Meriç, E.; Avşar, N. On the presence of alien foraminifera Amphistegina lobifera Larsen on the coasts of the Maltese Islands. Aquat. Invasions 2007, 2, 439–441. [Google Scholar] [CrossRef]
- Meriç, E.; Avşar, N.; Nazik, A.; Yokeş, M.B.; Dinçer, F. A review of benthic foraminifers and ostracodes of the Antalya coast. Micropaleontology 2008, 54, 199–240. [Google Scholar] [CrossRef]
- Triantaphyllou, M.V.; Koukousioura, O.; Dimiza, M.D. The presence of the Indo-Pacific symbiont-bearing foraminifer Amphistegina lobifera in Greek coastal ecosystems (Aegean Sea, Eastern Mediterranean). Mediterr. Mar. Sci. 2009, 10, 73–85. [Google Scholar] [CrossRef]
- Zenetos, A.; Gofas, S.; Verlaque, M.; Çinar, M.E.; García Raso, J.G.; Bianchi, C.N.; Morri, C.; Azzurro, E.; Bilecenoglu, M.; Froglia, C.; et al. Alien species in the Mediterranean Sea by 2010. A contribution to the application of European Union's Marine Strategy Framework Directive (MSFD). Part I. Spatial distribution. Mediterr. Mar. Sci. 2010, 11, 381–493. [Google Scholar] [CrossRef]
- Abu Tair, N.K.; Langer, M.R. Foraminiferal Invasions: The Effect of Lessepsian Migration on the Diversity and Composition of Benthic Foraminiferal Assemblage Around Cyprus (Mediterranean Sea). Forams 2010-International Symposium on Foraminifera, Bonn, 2010, p. 42 (Abstract).
- Guastella, R.; Marchini, A.; Caruso, A.; Evans, J.; Cobianchi, M.; Cosentino, C.; Langone, L.; Lecci, R.; Mancin, N. Reconstructing Bioinvasion Dynamics through Micropaleontologic Analysis highlights the Role of Temperature Change as a driver of Alien Foraminifera Invasion. Front. Mar. Sci. 2021, 8, 675807. [Google Scholar] [CrossRef]
- Koukousioura, O.; Dimiza, M.D.; Triantaphyllou, M.V. Alien foraminifers from Greek coastal areas (Aegean Sea, Eastern Mediterranean). Mediterr. Mar. Sci. 2010, 11, 155–172. [Google Scholar] [CrossRef]
- Langer, M.R.; Mouanga, G.H. Invasion of amphisteginid foraminifera in the Adriatic Sea. Biol. Invasions 2016, 18, 1335–1349. [Google Scholar] [CrossRef]
- Mahmood, K.; Hildreth, W. Geology of Pantelleria, a peralkaline volcano in the Strait of Sicily. Bull. Volcanol. 1986, 48, 143–172. [Google Scholar] [CrossRef]
- Neave, D.A.; Fabbro, G.; Herd, R.A.; Petrone, C.M.; Edmonds, M. Melting, Differentiation and Degassing at the Pantelleria Volcano, Italy. J. Petrology 2012, 53, 637–663. [Google Scholar] [CrossRef]
- Rotolo, S.G.; Scaillet, B.; La Felice, S.; Vita-Scaillet, L. Paroxysmal eruption of Pantelleria volcano (Italy) at 45 ka: insight into caldera formation, magma recharge and compositional evolution. J. Petrology 2013, 54, 767–800. [Google Scholar] [CrossRef]
- Caruso, A.; Censi, P.; Aricò, P.; Meli, C.; Sprovieri, M. Astronomical dating of two Pliocene alkaline volcanic ash layers in the Capo Rossello area (southern Sicily, Italy): implications for the beginning of the rifting in the Sicily Channel. Bull. Soc. Géol. France 2009, 180, 95–104. [Google Scholar] [CrossRef]
- Copernicus Marine Service. Available online: https://data.marine.copernicus.eu/.
- Carapezza, A. Gli Eterotteri Dell’Isola Di Pantelleria (Insecta, Heteroptera). Il Nat. Sicil. 1981, 4, 73–91. [Google Scholar]
- Galasso, G.; Montoleone, E.; Federico, C. Persicaria senegalensis (Polygonaceae), entità nuova per la flora italiana, e chiave di identificazione delle specie del genere Persicaria in Italia. NHS Nat. Hist. Sci. Atti Soc. Ital. Sci. Nat. Museo Civ. Storia Nat. Milano 2014, 1, 13–18. [Google Scholar] [CrossRef]
- Di Luca, M.; Toma, L.; Severini, F.; Boccolini, D.; D’Avola, S.; Todaro, D.; Stancanelli, A.; Antoci, F.; La Russa, F.; Casano, S.; Sotera, S.D.; Carraffa, E.; Versteirt, V.; Schaffner, F.; Romi, R.; Torina, A. First record of the invasive mosquito species Aedes (Stegomyia) albopictus (Diptera: Culicidae) on the southernmost Mediterranean islands of Italy and Europe. Parasites Vectors 2017, 10, 543. [Google Scholar] [CrossRef] [PubMed]
- Cristofaro, M.; Sforza, R.F.H.; Roselli, G.; Paolini, A.; Cemmi, A.; Musmeci, S.; Anfora, G.; Mazzoni, V.; Grodowitz, M. Effects of Gamma Irradiation on the Fecundity, Fertility, and Longevity of the Invasive Stink Bug Pest Bagrada hilaris (Burmeister) (Hemiptera: Pentatomidae). Insects 2022, 13, 787. [Google Scholar] [CrossRef]
- Mori, E.; Magoga, G.; Mazza, G. New records based on citizen-science report alien land planarians in the three remaining Italian regions and Pantelleria Island, and first record of Dolichoplana striata (Platyhelminthes Tricladida Contineticola Geoplanidae) in Italy. Redia 2022, 105, 91–95. [Google Scholar] [CrossRef]
- Minissale, P.; Cambria, S.; Montoleone, E.; Tavilla, G.; Giusso del Galdo, G.; Sciandrello, S.; Badalamenti, E.; La Mantia, T. The alien vascular flora of the Pantelleria Island National Park (Sicily Channel, Italy): new insights into the distribution of some potentially invasive species. BioInvasions Rec. 2023, 12, 861–885. [Google Scholar] [CrossRef]
- Paolini, A.; Musmeci, S.; Mainardi, C.E.; Peccerillo, C.; Cemmi, A.; Di Sarcina, I.; Marini, F.; Sforza, R.F.H.; Cristofaro, M. Age-dependent variation in longevity, fecundity and fertility of gamma-irradiated Bagrada hilaris (Hemiptera: Pentatomidae): insights for a sustainable SIT program. Insects 2025, 16, 408. [Google Scholar] [CrossRef]
- Giaccone, G.; Scammacca, B.; Cinelli, F.; Sartoni, G.; Furnari, G. Studio preliminare sulla tipologia della vegetazione sommersa del Canale di Sicilia e isole vicine. Giorn. Bot. Ital. 1972, 106, 211–229. [Google Scholar] [CrossRef]
- Giaccone, G.; Sortino, M.; Solazzi, A.; Tolomio, C. Tipologia e distribuzione estiva della vegetazione sommersa dell’isola di Pantelleria. Lav. Reale Ist. Bot. Reale Giard. Colon. Palermo 1973, 25, 103–109. [Google Scholar]
- Barone, R.; Calvo, S.; Sortino, M. Contributo alla conoscenza della flora sommersa del porto di Pantelleria (Canale di Sicilia). Giorn. Bot. Ital. 1978, 112, 239–248. [Google Scholar] [CrossRef]
- Calvo, S.; Sortino, M. Tipologia e distribuzione della vegetazione sommersa del porto di Pantelleria (Canale di Sicilia). Inform. Bot. Ital. 1979, 11, 189–195. [Google Scholar]
- Marletta, G.; Lombardo, A. The Fucales (Ochrophyta, Phaeophyceae) of the Island of Pantelleria (Sicily Channel, Mediterranean Sea): a new contribution. Ital. Bot. 2023, 15, 137–163. [Google Scholar] [CrossRef]
- Crocetta, F.; Agius, D.; Balistreri, P.; Bariche, M.; Bayhan, Y.K.; Çakir, M.; Ciriaco, S.; Corsini-Foka, M.; Deidun, A.; El Zrelli, R.; Ergüden, D.; Evans, J.; Ghelia, M.; Giavasi, M.; Kleitou, P.; Kondylatos, G.; Lipej, L.; Mifsud, C.; Özvarol, Y.; Pagano, A.; Portelli, P.; Poursanidis, D.; Rabaoui, L.; Schembri, P.J.; Taşkin, E.; Tiralongo, F.; Zenetos, A. New Mediterranean Biodiversity Records (October 2015). Mediterr. Mar. Sci. 2015, 16, 682–702. [Google Scholar] [CrossRef]
- Castriota, L.; Falautano, M.; Maggio, T.; Perzia, P. The Blue Swimming Crab Portunus segnis in the Mediterranean Sea: Invasion Paths, Impacts and Management Measures. Biology 2022, 11, 1473. [Google Scholar] [CrossRef]
- Agenzia Regionale per la Protezione dell’Ambiente, Arpa Sicilia. Progetto Caulerpa – Indagini ambientali e rilievi sulla diffusione della Caulerpa nel Canale di Sicilia, 2014, 79 pp.
- Title of Site. Available online: https://www.regionieambiente.it/specie-aliene-ispra/ (accessed on 28 July 2025).
- Walton, W.R. Techniques for recognition of living foraminifera. Contrib. Cushman Found. Foram. Res. 1952, 3, 56–60. [Google Scholar]
- Schönfeld, J.; Alve, E.; Geslin, E.; Jorissen, F.; Korsun, S.; Spezzaferri, S.; Members of The, Fobimo. The Fobimo (FOraminiferal BIo-MOnitoring) initiative—towards a standardized protocol for soft-bottom benthic foraminiferal monitoring studies. Mar. Micropaleontol. 2012, 94–95, 1–13. [Google Scholar] [CrossRef]
- Loeblich, A.R.; Tappan, J.H. Foraminiferal Genera and their Classification, 4th ed.; Van Nostrand Reinhold: New York, USA, 1987. [Google Scholar]
- Cimerman, F.; Langer, M.R. Mediterranean Foraminifera. Slovenska Akademija Znanosti in Umetnosti, 33, Ljubljana, 1991.
- Caruso, A.; Cosentino, C. Classification and Taxonomy of Modern Benthic Shelf Foraminifera of the Central Mediterranean Sea. In Foraminifera: Aspects of Classification; Georgescu, M.D., Ed.; Nova Publishers: New York, USA, 2014b; pp. 249–313. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Fisher, R.A.; Corbet, A.S.; Williams, C.B. The relationship between the number of species and the number of individuals in a random sample of an animal population. J. Anim. Ecol. 1943, 12, 42–58. [Google Scholar] [CrossRef]
- Murray, J.W. Distribution and Ecology of Living Benthic Foraminiferids; Heinemann Educational Books: London, UK, 1973. [Google Scholar]
- Guastella, R.; Evans, J.; Mancin, N.; Caruso, A.; Marchini, A. Assessing the effect of Amphistegina lobifera invasion on infralittoral benthic foraminiferal assemblages in the Sicily Channel (Central Mediterranean). Mar. Environ. Res. 2023, 192, 106247. [Google Scholar] [CrossRef]
- Weinmann, A.E.; Koukousioura, O.; Triantaphyllou, M.V.; Langer, M.R. Invasive shallow water foraminifera impacts local biodiversity mostly at densities above 20 %: the case of Corfu Island. Web Ecol. 2023, 23, 71–86. [Google Scholar] [CrossRef]
- Sgarrella, F.; Moncharmont Zei, M. Benthic foraminifera of the Gulf of Naples (Italy): systematics and autoecology. Boll. Soc. Paleontol. Ital. 1993, 32, 145–264. [Google Scholar]
- Langer, M.R. Epiphytic foraminifera. Mar. Micropaleontol. 1993, 20, 235–265. [Google Scholar] [CrossRef]
- Title of Site. Available online: https://marine.copernicus.eu/it/ocean-climate-portal/sea-surface-temperature (accessed on 28 July 2025).
- DIRECTIVE 2008/56/EC OF THE EUROPEAN PARLIAMENT AND OF THE COUNCIL of 17 June 2008 establishing a framework for community action in the field of marine environmental policy (Marine Strategy Framework Directive). Official Journal of the European Union.
- EU. Regulation (EU) No 1143/2014 of the European Parliament and of the Council of 22 October 2014 on the prevention and management of the introduction and spread of invasive alien species. Official Journal of the European Union L 317, 35–55.
- Perzia, P.; Cillari, T.; Crociata, G.; Deidun, A.; Falautano, M.; Franzitta, G.; Galdies, J.; Maggio, T.; Vivona, P.; Castriota, L. Using Local Ecological Knowledge to Search for Non-Native Species in Natura 2000 Sites in the Central Mediterranean Sea: An Approach to Identify New Arrivals and Hotspot Areas. Biology 2023, 12, 1158. [Google Scholar] [CrossRef]
Figure 1.
Location map of the studied area in the Sicily Channel (Mediterranean Sea, A) and location of the sampling stations around the island of Pantelleria (B).
Figure 1.
Location map of the studied area in the Sicily Channel (Mediterranean Sea, A) and location of the sampling stations around the island of Pantelleria (B).
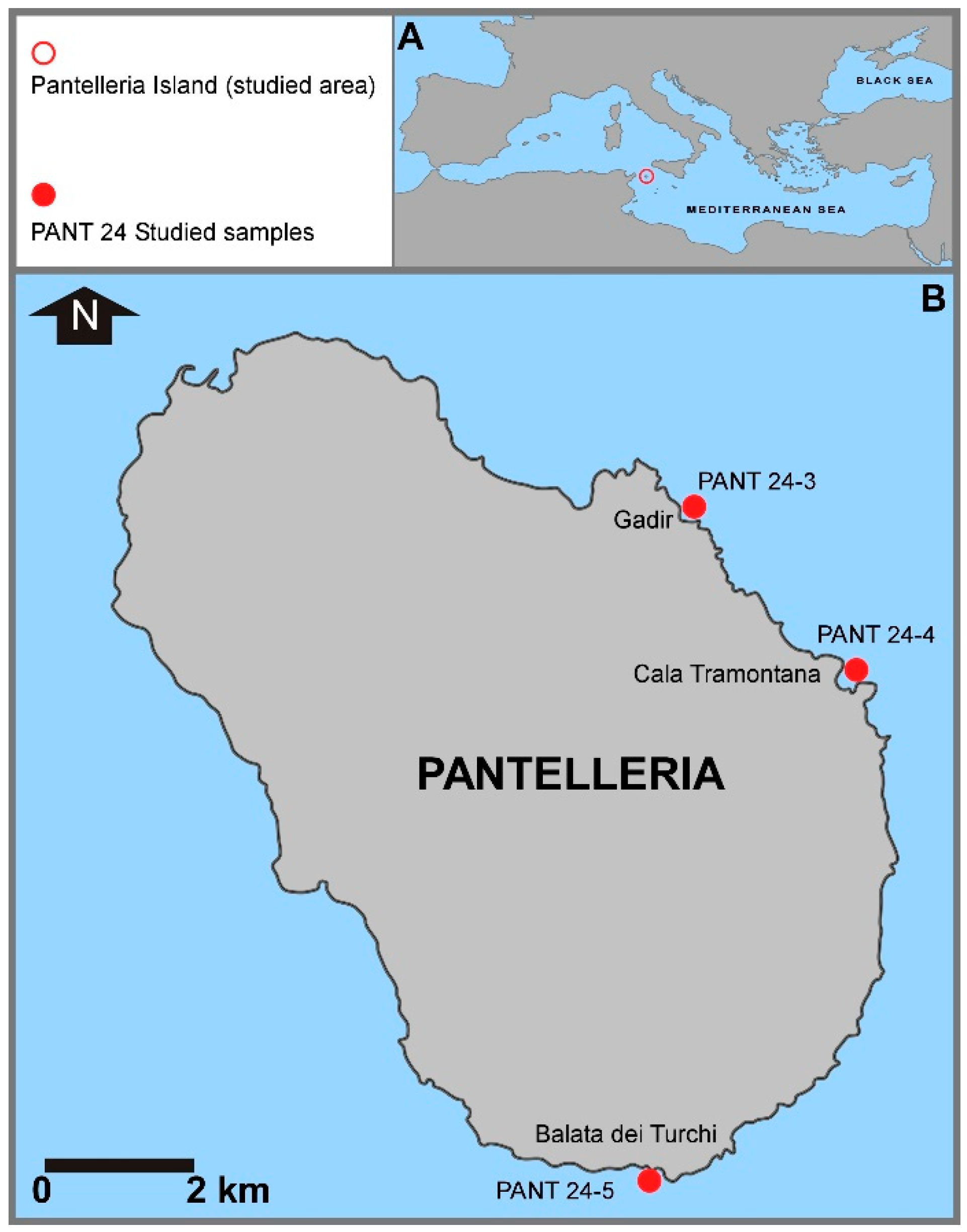
Figure 2.
A) Location of samples collected at the Gadir site (imagine from Google Earth); (B) particular of the PANT 24-3-2 sample stored in a cylindrical polyethylene container (Falcon), all the white dots visible even to the naked eye are shells of living Amphistegina attached as epiphytes on algae.
Figure 2.
A) Location of samples collected at the Gadir site (imagine from Google Earth); (B) particular of the PANT 24-3-2 sample stored in a cylindrical polyethylene container (Falcon), all the white dots visible even to the naked eye are shells of living Amphistegina attached as epiphytes on algae.

Figure 3.
A) Location of samples collected at Cala Tramontana (imagine from Google Earth); (B) PANT 24-4-1 (-7 m), Posidonia oceanica and the brown algae Halopteris scoparia are clearly visible on the rocky substrate; (C) PANT 24-4-2 (-9 m), particular of P. oceanica, on the right only rhizomes were found, since the meadow was destroyed by touristic boats during summer anchorages. The rest of rhizomes are rich in Amphistegina shells (white dots).
Figure 3.
A) Location of samples collected at Cala Tramontana (imagine from Google Earth); (B) PANT 24-4-1 (-7 m), Posidonia oceanica and the brown algae Halopteris scoparia are clearly visible on the rocky substrate; (C) PANT 24-4-2 (-9 m), particular of P. oceanica, on the right only rhizomes were found, since the meadow was destroyed by touristic boats during summer anchorages. The rest of rhizomes are rich in Amphistegina shells (white dots).
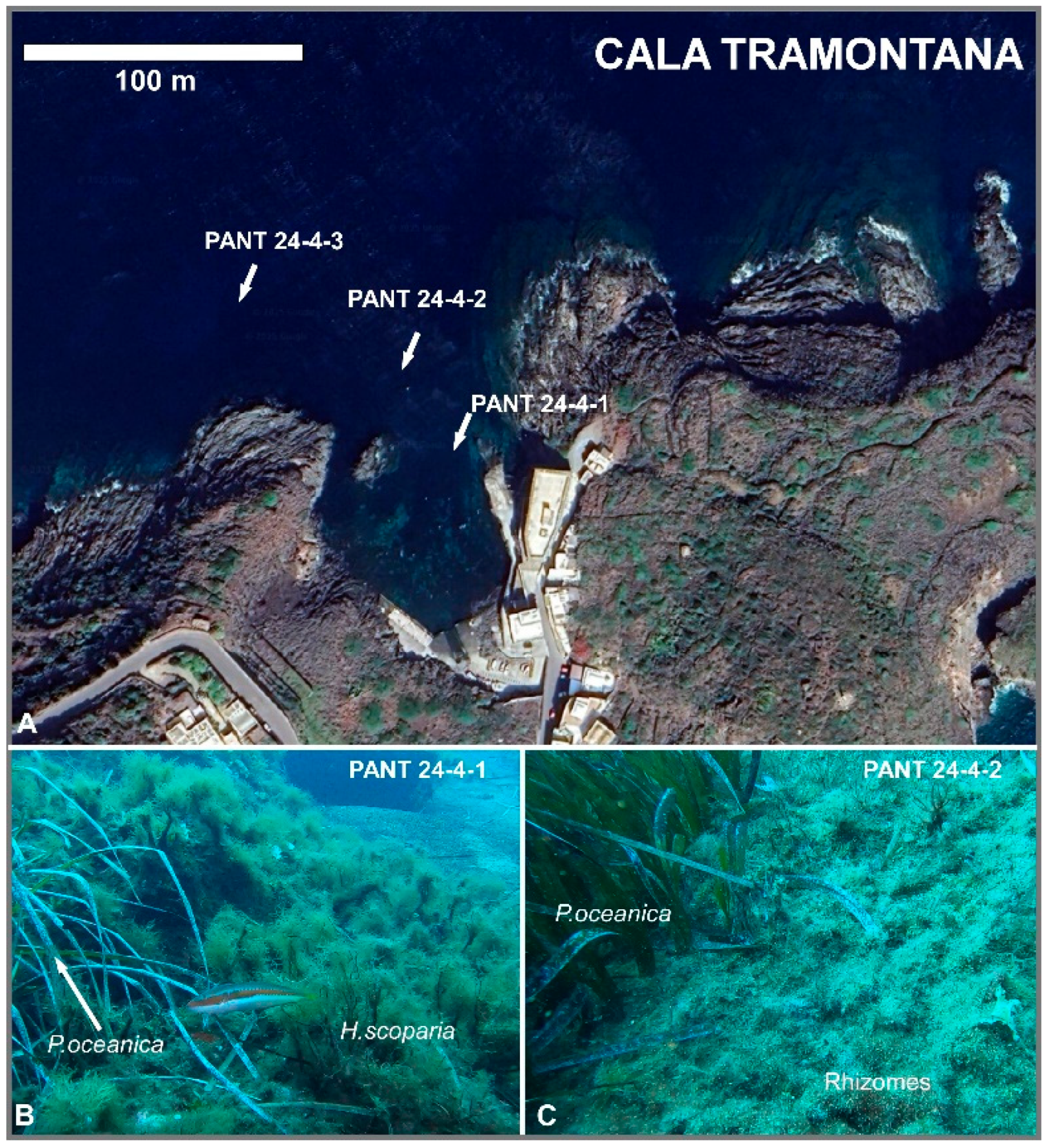
Figure 4.
Location of samples collected at Balata dei Turchi (imagine from Google Earth).

Figure 5.
Relative abundances of living benthic foraminifera in samples from Pantelleria’ island. A. lobifera reaches percentages >20% except in the sample at -20 m depth. In the deeper samples A. lessonii is more abundant with respect to A. lobifera.
Figure 5.
Relative abundances of living benthic foraminifera in samples from Pantelleria’ island. A. lobifera reaches percentages >20% except in the sample at -20 m depth. In the deeper samples A. lessonii is more abundant with respect to A. lobifera.
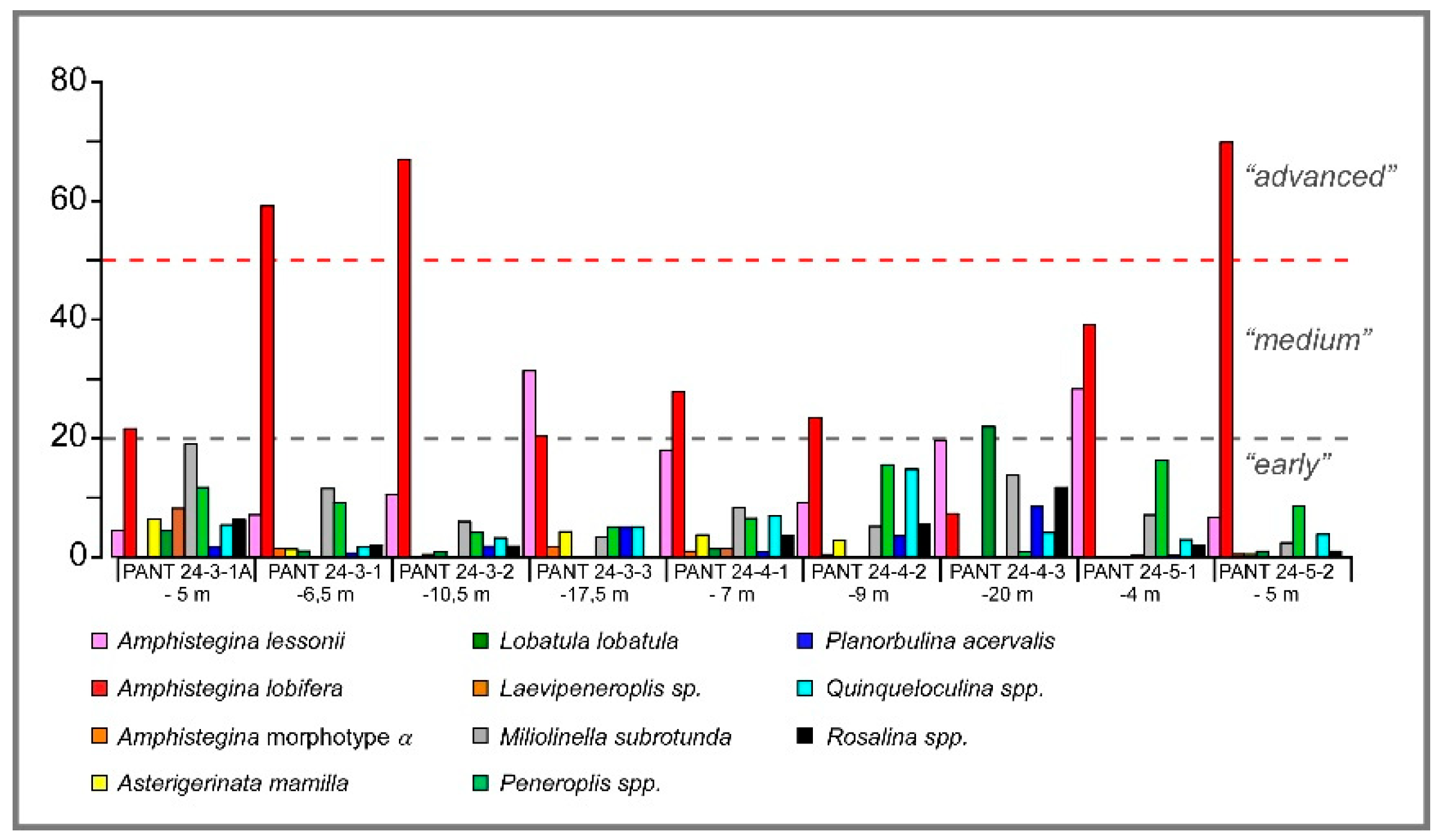
Figure 6.
Diversity indexes calculated for living benthic foraminiferal assemblages around Pantelleria’ island.
Figure 6.
Diversity indexes calculated for living benthic foraminiferal assemblages around Pantelleria’ island.
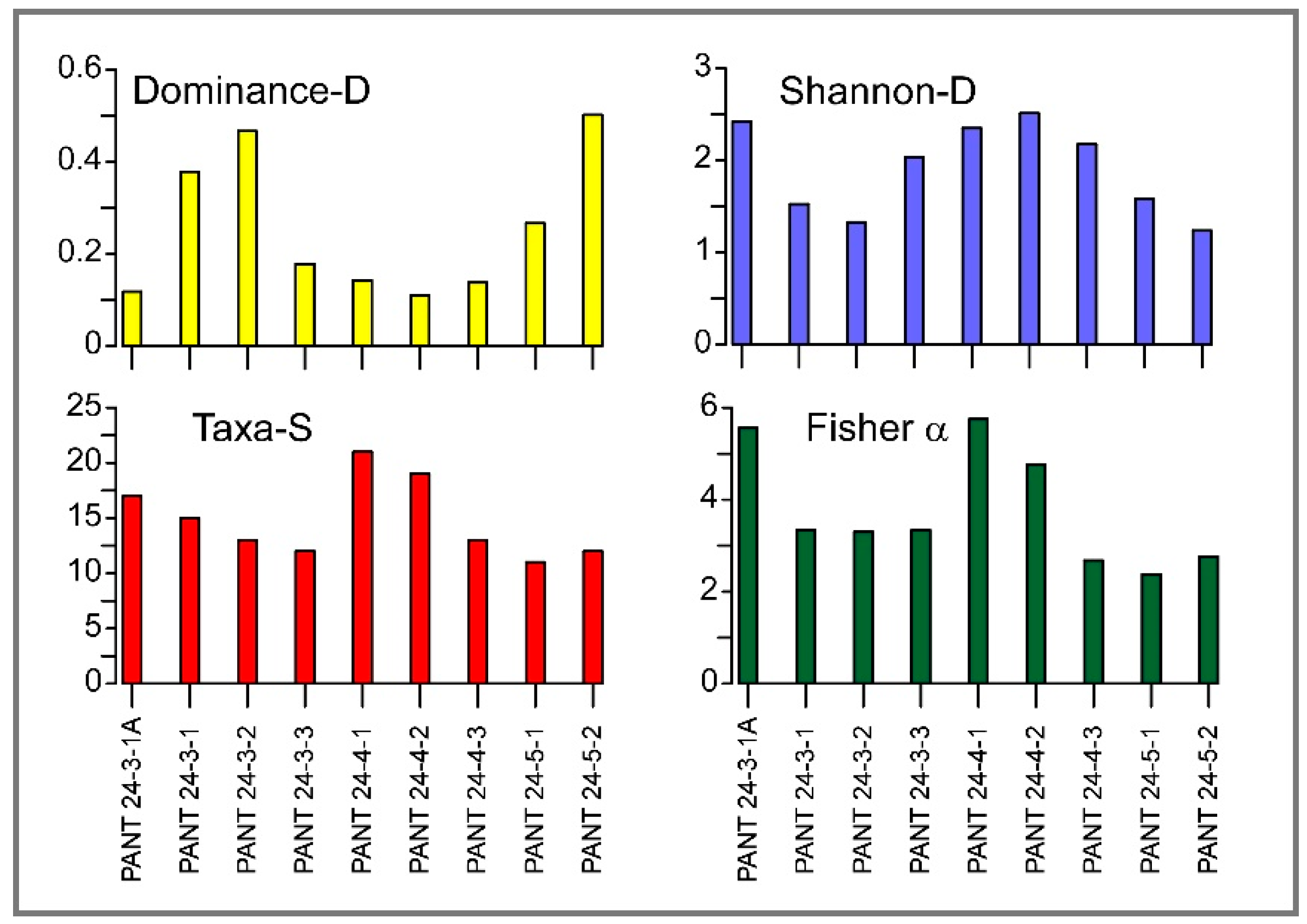
Figure 7.
A) A. lessonii, ventral side (PANT 24-3-3); B) A. lobifera, spiral side (PANT 24-3-2); C) A. lobifera, lateral view (PANT 24-3-2); D) A. lobifera, detail of the aperture (PANT 24-3-2); A. lobifera, spiral side (PANT 24-3-3). A, B, C, D: SEM photographs, D: optical microscope photograph.
Figure 7.
A) A. lessonii, ventral side (PANT 24-3-3); B) A. lobifera, spiral side (PANT 24-3-2); C) A. lobifera, lateral view (PANT 24-3-2); D) A. lobifera, detail of the aperture (PANT 24-3-2); A. lobifera, spiral side (PANT 24-3-3). A, B, C, D: SEM photographs, D: optical microscope photograph.
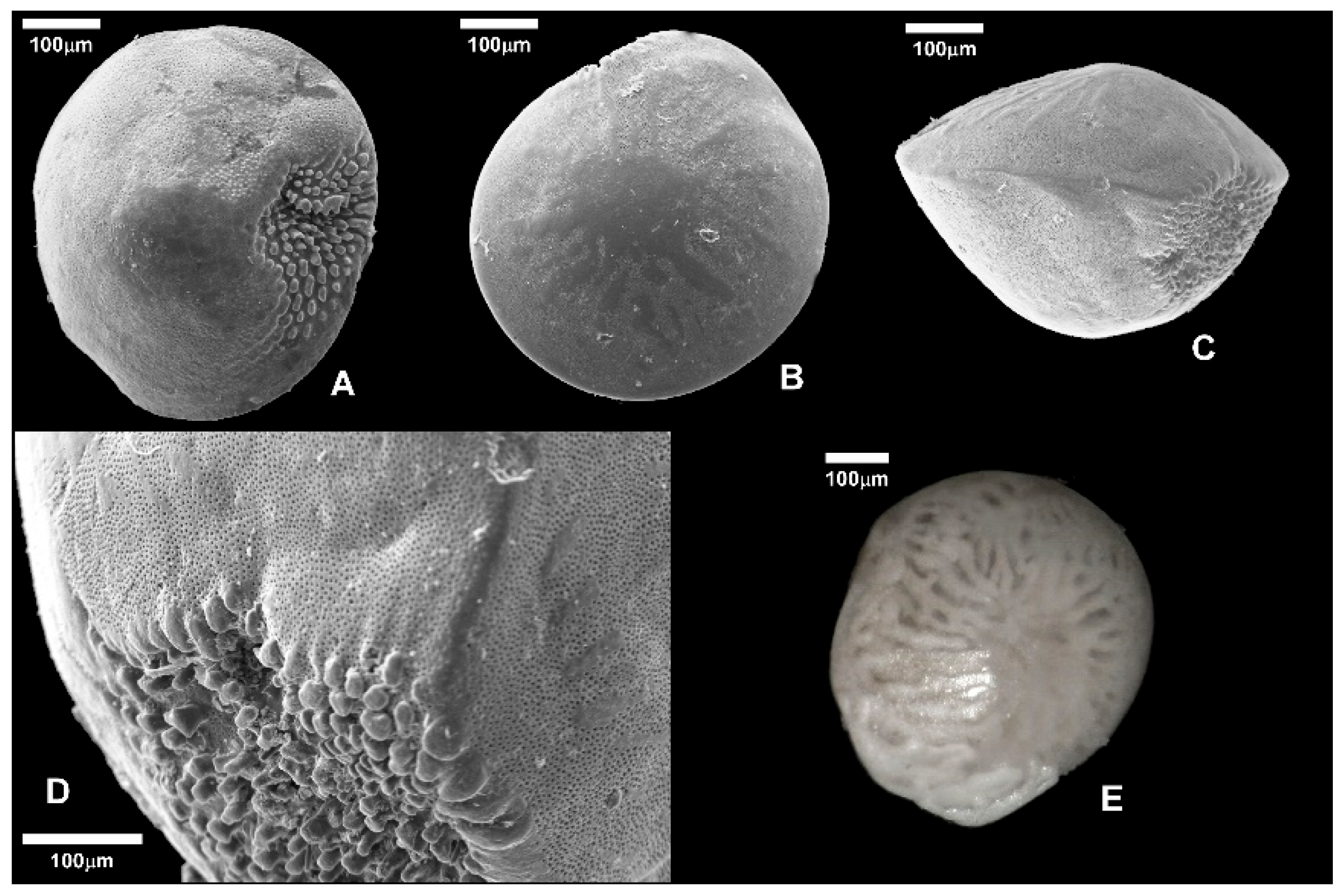
Figure 8.
An example of the Sea Surface Temperatures in the Mediterranean Sea recorded on 1th July, 2024 (data from https://data.marine.copernicus.eu/). Off the coast of the southern Sicily the temperatures are lower respect to Sicily Channel.
Figure 8.
An example of the Sea Surface Temperatures in the Mediterranean Sea recorded on 1th July, 2024 (data from https://data.marine.copernicus.eu/). Off the coast of the southern Sicily the temperatures are lower respect to Sicily Channel.
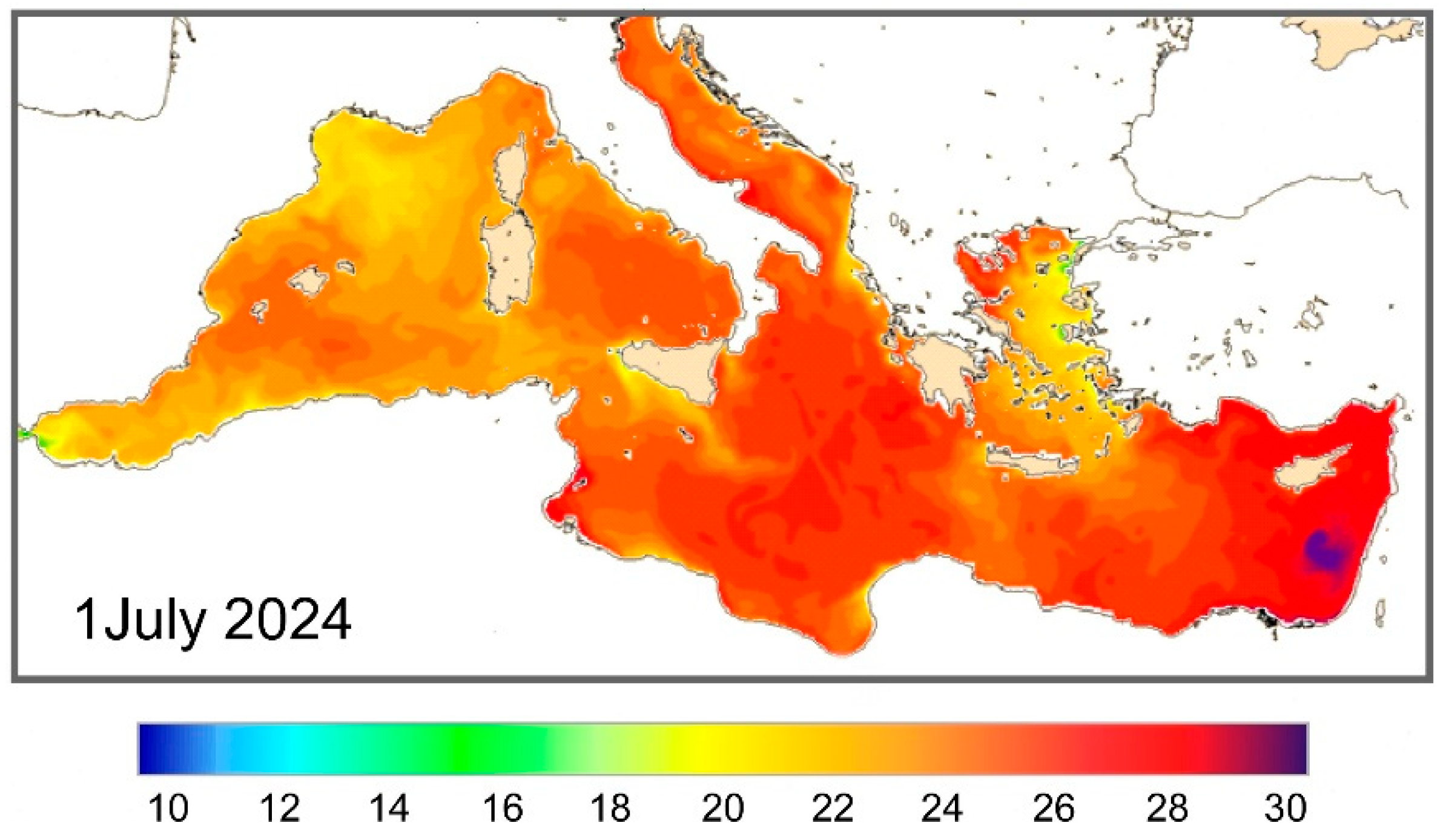
Figure 9.
Current range and invasion success of Amphistegina lobifera in the Sicily Channel (modified from Guastella et al., 2019). The distribution and the degree of Amphistegina colonization is well correlated with Sea Surface Temperatures reported in Figure 8.
Figure 9.
Current range and invasion success of Amphistegina lobifera in the Sicily Channel (modified from Guastella et al., 2019). The distribution and the degree of Amphistegina colonization is well correlated with Sea Surface Temperatures reported in Figure 8.
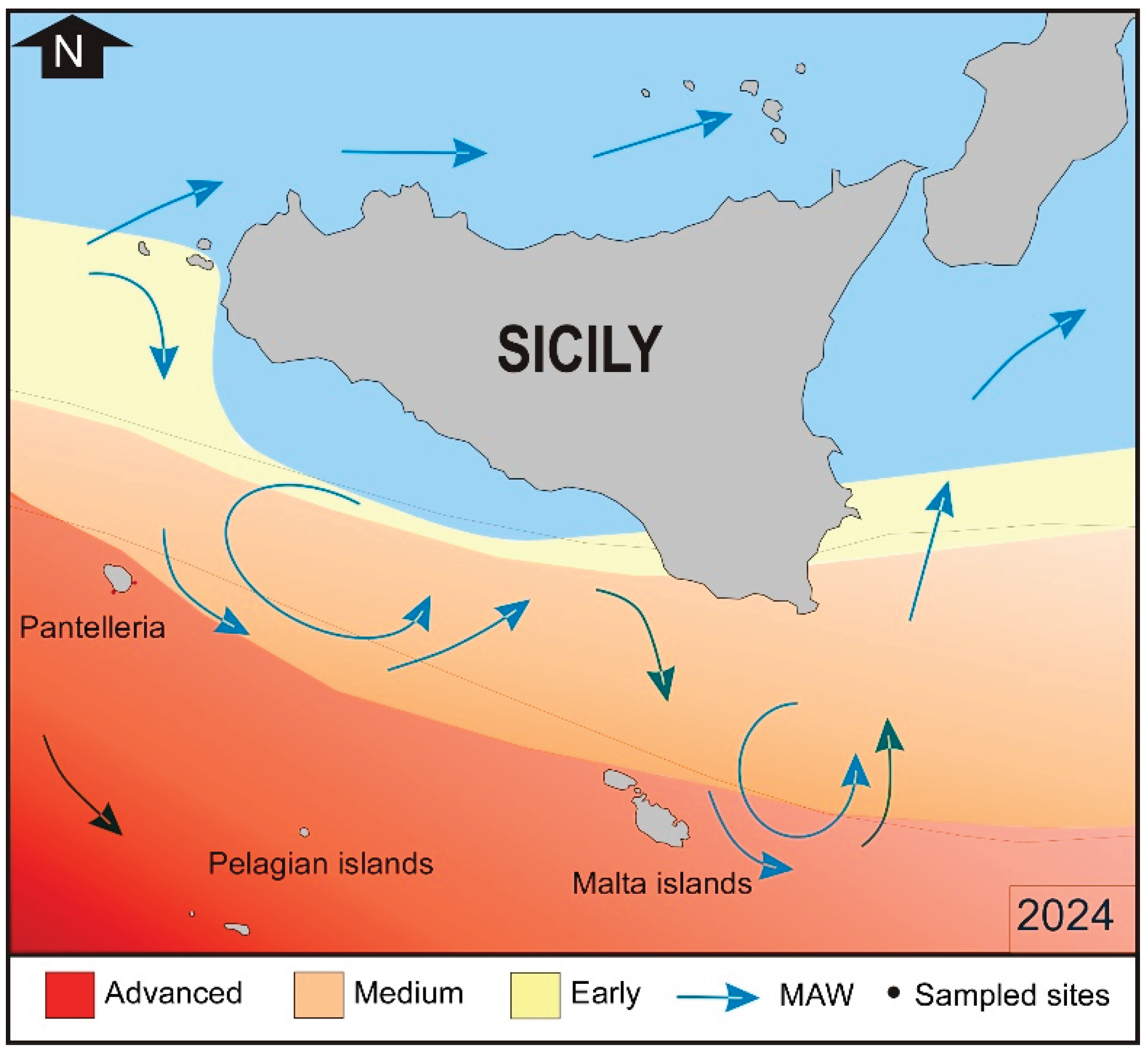

Table 1.
Geographic coordinates, bathymetry and type of sample collected at each sampling site. Two values for temperature and salinity are reported, asterisks indicate our measurements during samples’ collection while the other value is from [48].
Table 1.
Geographic coordinates, bathymetry and type of sample collected at each sampling site. Two values for temperature and salinity are reported, asterisks indicate our measurements during samples’ collection while the other value is from [48].
| Sites | Latitude | Longitude | sites | SST (°C) | SSS (‰) | Depth (m) | Sample type |
|---|---|---|---|---|---|---|---|
| Gadir | 36 48' 42'' N | 12 01' 32'' E | PANT 24-3-1A | 24* - 22.6 | 37*- 37.5 | 5 | P. oceanica's rhizomes |
| PANT 24-3-1 | 6,5 | P. oceanica's rhizomes | |||||
| PANT 24-3-2 | 10,5 | P. oceanica's rhizomes | |||||
| PANT 24-3-3 | 17,5 | P. oceanica's rhizomes | |||||
| Cala Tramontana | 36 47' 54'' N | 12 02' 52'' E | PANT 24-4-1 | 24* - 22.8 | 37* - 37.5 | 7 | brown algae Halopteris scoparia |
| PANT 24-4-2 | 9 | P. oceanica's rhizomes | |||||
| PANT 24-4-3 | 20 | brown algae Halopteris scoparia | |||||
| Balata dei Turchi | 36 44' 10'' N | 12 01' 09'' E | PANT 24-5-1 | 24* - 22.9 | 37*- 37.5 | 4 | brown algae Halopteris scoparia |
| PANT 24-5-2 | 5 | sediment |
Table 2.
Numbers and percentages of total living versus total dead foraminifera in the studied samples.
Table 2.
Numbers and percentages of total living versus total dead foraminifera in the studied samples.
| Sampling sites | N. of living specimens | N. of dead specimens | N. of total specimens (living + dead) |
Total living foraminifera (%) | Total dead foraminifera (%) |
|---|---|---|---|---|---|
| PANT 24-3-1A | 111 | 0 | 111 | 100,00 | 0,00 |
| PANT 24-3-1 | 296 | 0 | 296 | 100,00 | 0,00 |
| PANT 24-3-2 | 218 | 0 | 218 | 100,00 | 0,00 |
| PANT 24-3-3 | 118 | 31 | 149 | 79,19 | 20,81 |
| PANT 24-4-1 | 216 | 13 | 229 | 94,32 | 5,68 |
| PANT 24-4-2 | 251 | 15 | 266 | 94,36 | 5,64 |
| PANT 24-4-3 | 342 | 19 | 361 | 94,74 | 5,26 |
| PANT 24-5-1 | 240 | 3 | 243 | 98,77 | 1,23 |
| PANT 24-5-2 | 209 | 81 | 290 | 72,07 | 27,93 |
Table 3.
Percentages of living foraminiferal species.
| Benthic foraminiferal species | PANT 24-3-1A | PANT 24-3-1 | PANT 24-3-2 | PANT 24-3-3 | PANT 24-4-1 | PANT 24-4-2 | PANT 24-4-3 | PANT 24-5-1 | PANT 24-5-2 |
|---|---|---|---|---|---|---|---|---|---|
| Adelosina sp1. | 0,90 | 0,00 | 0,00 | 0,00 | 0,00 | 3,59 | 0,00 | 0,00 | 0,00 |
| Adelosina sp.2 | 0,00 | 0,00 | 0,00 | 0,00 | 0,93 | 0,00 | 0,00 | 0,00 | 0,00 |
| Amphisorus hemprichii | 0,00 | 0,34 | 0,00 | 0,00 | 0,00 | 0,00 | 0,00 | 0,00 | 0,00 |
| Amphistegina lobifera | 21,62 | 59,12 | 66,97 | 20,34 | 27,78 | 23,51 | 7,31 | 39,17 | 69,86 |
| Amphistegina lessonii | 4,50 | 7,09 | 10,55 | 31,36 | 18,06 | 9,16 | 19,59 | 28,33 | 6,70 |
| Amph. morphotype alfa | 0,00 | 1,35 | 0,00 | 1,69 | 0,93 | 0,40 | 0,00 | 0,00 | 0,48 |
| Asterigerinata mamilla | 6,31 | 1,35 | 0,46 | 4,24 | 3,70 | 2,79 | 0,00 | 0,00 | 0,48 |
| Bolivina catanensis | 0,00 | 0,00 | 0,00 | 0,00 | 0,00 | 0,40 | 0,00 | 0,00 | 0,00 |
| Cribroelphidium sp. | 0,00 | 0,00 | 0,00 | 0,00 | 0,00 | 0,80 | 0,29 | 0,00 | 0,00 |
| Cyclocibicides vermiculatus | 0,00 | 0,00 | 0,00 | 4,24 | 0,93 | 0,00 | 0,00 | 0,00 | 0,00 |
| Cymbaloporetta squammosa | 2,70 | 0,68 | 0,00 | 0,00 | 1,39 | 0,00 | 0,00 | 0,00 | 0,00 |
| Laevipeneroplis sp. | 8,11 | 0,00 | 0,00 | 0,00 | 1,39 | 0,00 | 0,00 | 0,42 | 0,00 |
| Lachlanella variolata | 2,70 | 0,00 | 0,00 | 0,00 | 0,00 | 0,00 | 0,00 | 0,00 | 0,00 |
| Lobatula lobatula | 4,50 | 1,01 | 0,92 | 0,00 | 1,39 | 0,00 | 21,93 | 0,00 | 0,96 |
| Miliolinella subrotunda | 18,92 | 11,49 | 5,96 | 3,39 | 8,33 | 5,18 | 13,74 | 7,08 | 2,39 |
| Peneroplis pertusus | 10,81 | 9,12 | 3,21 | 3,39 | 5,56 | 13,55 | 0,00 | 16,25 | 6,22 |
| Peneroplis planatus | 0,90 | 0,00 | 0,92 | 1,69 | 0,93 | 1,99 | 0,88 | 0,00 | 2,39 |
| Planorbulina acervalis | 1,80 | 0,68 | 1,83 | 5,08 | 0,93 | 3,59 | 8,48 | 0,42 | 0,00 |
| Quinqueloculina sp.1 | 0,90 | 1,35 | 0,92 | 5,08 | 3,70 | 7,17 | 2,34 | 0,83 | 1,44 |
| Quinqueloculina sp.2 | 4,50 | 0,34 | 2,29 | 0,00 | 2,78 | 7,57 | 1,75 | 2,08 | 2,39 |
| Quinqueloculina sp.3 | 0,00 | 0,00 | 0,00 | 0,00 | 0,46 | 0,00 | 0,00 | 0,00 | 0,00 |
| Quinqueloculina sp.4 | 0,00 | 0,00 | 0,00 | 0,00 | 0,00 | 0,00 | 2,34 | 0,00 | 0,00 |
| Rosalina bradyi | 0,00 | 0,00 | 0,00 | 0,00 | 0,00 | 0,80 | 0,00 | 0,00 | 0,00 |
| Rosalina obtusa | 6,31 | 2,03 | 1,83 | 0,00 | 3,70 | 4,78 | 11,70 | 2,08 | 0,96 |
| Spiroloculina excavata | 0,00 | 0,00 | 0,00 | 0,00 | 0,00 | 1,99 | 0,00 | 0,00 | 0,00 |
| Spiroloculina sp. | 0,00 | 0,00 | 0,00 | 0,00 | 0,46 | 1,99 | 0,00 | 0,00 | 0,00 |
| Textularia pala | 1,80 | 3,04 | 1,38 | 16,10 | 12,50 | 7,57 | 4,39 | 2,50 | 5,74 |
| Tretomphalus bulloides | 0,00 | 0,00 | 0,00 | 0,00 | 0,46 | 0,00 | 0,00 | 0,00 | 0,00 |
| Vertebralina striata | 2,70 | 1,01 | 2,75 | 3,39 | 3,70 | 3,19 | 5,26 | 0,83 | 0,00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



