
Submitted:
31 July 2025
Posted:
04 August 2025
You are already at the latest version
Abstract
The human face serves as a crucial component of attractiveness and identity, playing a pivotal role in social interactions. The genetic legacy of the human face reaches back to the evolution of the vertebrate jaw, via transformation of the third gill-arch of lamprey-like ancestors. Teleost fish models, particularly zebrafish, have offered valuable insights into the developmental and molecular mechanisms governing craniofacial skeletogenesis, owing to the molecular conservation in these processes. However, knowledge regarding the morphogenic processes shaping facial soft tissues remains surprisingly sparse. The diverse family of Cichlidae constitute ~10% of teleost fish species and represent an exceptional opportunity for evolutionary biology research due to their extensive ecological diversity and rapid speciation rates. Cichlid fishes also exhibit remarkable craniofacial morphological diversity, making them excellent models for studying both craniofacial skeletal and soft tissue morphogenesis. Utilizing the wealth of natural mutants within cichlid populations, this short review pitches cichlid fish models as valuable tools for investigating the genetic regulators and interactions underlying facial soft tissue formation. By synthesising developmental and regulatory mechanisms that influence morphological variations in facial soft tissues in cichlids and other model organisms, we create a blueprint for forthcoming molecular genetic investigations into facial diversity across various species and facial deformities in humans.
Keywords:
craniofacial development
; facial soft tissues
; Gene regulation
; cichlids
; morphological diversity
Introduction
In recent years, the investigation of how facial soft tissues develop has gained significant attention due to its implications for human health and overall well-being (Chaimongkhol and Mahakkanukrauh, 2022; Cotofana and Lachman, 2019; Giri et al., 2024; Hersberger-Zurfluh et al., 2018; Hînganu et al., 2024; Qian et al., 2022; Richmond et al., 2018; Weber et al., 2021). The face holds a central position in both attractiveness and identity, crucial for shaping interpersonal interactions among humans. Any abnormalities in facial appearance, whether present from birth or acquired later, can profoundly affect an individual's quality of life, often resulting in social stigma and negative emotional consequences due to perceived deviations from societal norms (Bull and Rumsey, 1988; Little et al., 2011; Samson et al., 2010). Furthermore, society's increasing focus on facial aesthetics underscores the importance of focusing on the complex molecular processes involved in the formation of facial soft tissues.
Facial soft tissues make significant contributions to a complex array of structures, including the lips, nose, ears, cheeks, chin, and forehead, each with distinct structural and functional characteristics (Nanda et al., 1990). The formation of these tissues involves complex molecular pathways that regulate cellular differentiation, proliferation, and patterning (Landi et al., 2022; Quinto-Sánchez et al., 2018; Szabo-Rogers et al., 2008; Weber et al., 2021). Despite the unique importance of facial soft tissues, our understanding of the molecular processes governing their morphogenesis, and how these contribute to overall facial geometry, remains limited. For example, genetic association studies in humans have implicated various genetic factors in shaping facial soft tissues structures (Naqvi et al., 2022). Other studies have shown that the morphology of specific parts of the human face, such as the forehead, upper lips, and nose, exhibit high heritability, offering these anatomical regions as prime candidates for exploring the genetic contributions to facial soft tissue variation (Giri et al., 2024; Hersberger-Zurfluh et al., 2018; Jelenkovic et al., 2010; Qian et al., 2022; Richmond et al., 2018; Song et al., 2018). However, further functional and developmental investigations are needed to dissect the molecular mechanisms through which they operate. Traditional mammalian models for genetic studies have provided valuable insights into various biological processes, including craniofacial development. But ethical considerations and practical constraints limit the feasibility of conducting extensive genetic manipulations and experimental studies in mammalian models (Higashiyama et al., 2025; Xie et al., 2024). Furthermore, the long gestation period and relatively low reproductive rates of mammalian models can impede the pace of research and limit the scope of studies investigating the molecular mechanisms underlying facial soft tissue morphogenesis. On the other hand, teleost fish models, such as zebrafish, although instrumental in elucidating the developmental and molecular mechanisms governing craniofacial skeletogenesis, lack pronounced facial soft tissue structures comparable to mammals (Ibarra and Atit, 2020).
In contrast to zebrafish, the naturally diverse family of Cichlidae emerges as model for investigating morphological variation in facial soft tissues. Cichlid fishes exhibit remarkable craniofacial morphological diversity (Conith and Albertson, 2021; Cooper et al., 2010; Fryer and Iles, 1972; Liem, 1973; Powder et al., 2015, 2014; Powder and Albertson, 2016), with exaggerated facial soft tissues and complex facial structures (Colombo et al., 2013; Concannon and Albertson, 2015; Conith et al., 2019, 2018; Duenser et al., 2023; Henning et al., 2017; Lecaudey et al., 2021, 2019; Machado-Schiaffino et al., 2017; Machii et al., 2025; Rometsch et al., 2021). Notably, the extensive diversity among cichlids has evolved in the context of gene flow and overall low levels of genetic divergence, making this system ideal for genetic/association mapping (Albertson et al., 2003; Brawand et al., 2014; Carmona Baez et al., 2025; Hulsey et al., 2017; Kautt et al., 2020; Parnell et al., 2012; Santos et al., 2023; Torres-Dowdall and Meyer, 2021). Furthermore, the availability of high quality annotated genomic data from diverse but phylogenetically closely related species of cichlids, coupled with established genetic manipulation tools like CRISPR/Cas9 (Clark et al., 2023; Marconi et al., 2024; Santos et al., 2014), positions cichlid fishes as powerful models for studying the genetic regulators and interactions underlying facial soft tissue formation. The wealth of natural mutants among cichlids and their experimental tractability make them amenable to elucidate the molecular pathways shaping facial soft tissues and their role in morphological variation.
This review provides a concise overview of recent molecular findings in cichlids related to facial soft tissue morphogenesis (excluding muscles and tendons). By connecting these findings to previous research in humans and other species, we seek to establish cichlid fish models as valuable tools for investigating facial diversity across various species and for decoding its underlying genetic regulatory networks. Overall, this review highlights the timeliness and significance of utilizing cichlid fish models in advancing our understanding of facial soft tissue morphogenesis and its implications for human health.
Facial Soft Tissues
In recent decades, craniofacial genetic research has transitioned from primarily focusing on anomalies to understanding the biological basis of normal facial variation. This expanded focus is aided by advancements in high-resolution three-dimensional systems for precise facial feature capture and sequencing technology for exploring genetic impacts on facial characteristics (Richmond et al., 2018). Investigating the genetics of normal-range facial morphology is essential for several reasons. Firstly, it elucidates the interplay between environmental factors and parental biological contributions, revealing the origins of individual appearance. Secondly, in certain instances specific but subclinical craniofacial shape in parents can indicate an elevated risk of dysmorphology in children (El Sergani et al., 2021). Furthermore, genetic data can determine ancestry, sex, and specific facial features, with applications in healthcare and forensics. Finally, studying historical selection and adaptation informs craniofacial evolution research, while exploring shared facial traits, medical conditions, and genes offers insights into commonalities and differences among individuals (Richmond et al., 2018).
Most craniofacial tissues originate from cranial neural crest cells (CNCCs), which undergo epithelial-to-mesenchymal transition and migrate ventrally to form most of the craniofacial skeleton and connective tissue (Som and Naidich, 2013). During early embryonic development, CNCCs divide into frontonasal and pharyngeal arches, with the frontonasal prominence giving rise to the forehead and nasal bones (Cordero et al., 2011). Subsequently, nasal placodes develop into lateral and medial nasal prominences, while mandibular prominences merge to form the mandible. Fusion of nasal and maxillary prominences leads to the formation of central nose structures (Cordero et al., 2011; Som and Naidich, 2013). Finally, fusion of remaining facial structures occurs, with subsequent growth and maturation throughout the embryogenesis. The development facial soft tissues also involve complex embryonic processes closely linked to CNCCs (Richmond et al., 2018). In mammals, facial soft tissue development comprises distinct components, including frontonasal (forehead and upper eyelid), medial nasal (nose and upper lip/philtrum), lateral nasal (sidewalls of the nostrils and base of the nose), maxillary (lower eyelids, cheeks and lateral parts of the upper lip), and mandibular (the entire lower lip) regions, which fuse at different stages of embryonic development (Richmond et al., 2018). Molecular studies highlight the complex interactions of various factors and pathways, such as fibroblast growth factors, hedgehog proteins, bone morphogenetic proteins, Wnt signaling, retinoic acid signaling, homeobox genes, etc., in regulating facial primordia growth and patterning (Ahi, 2016; Richmond et al., 2018). Disruptions in facial process fusion can lead to facial, lip, or palate clefts. In human, the postnatal facial growth follows general somatic growth patterns, with distinct periods of steady and rapid growth, peaking at puberty (Som and Naidich, 2013). The timing, direction, and duration of growth surges vary between genders and populations, contributing to overall facial variation (Matthews et al., 2018).
Facial variation is a consequence of genetic and environmental factors (Naqvi et al., 2022). Genetic influences are evident through familial resemblance and similarities between monozygotic twins compared to dizygotic twins (Giri et al., 2024; Hersberger-Zurfluh et al., 2018). Shared facial characteristics within ancestries and sexes, as well as distinctive features associated with genetic conditions, further illustrate genetic influence on facial appearance. Studies estimating facial heritability traditionally involve comparing facial similarity among relatives like twins or parents and offspring. With advancements in technology, large-scale analyses using genome-wide association study (GWAS) have become feasible (Finucane et al., 2015), even in cichlid fishes (Singh et al., 2025). Strong genetic influences, particularly on upper and midfacial regions (e.g. forehead, upper lip and nose) (Figure 1A), have been consistently reported (Hoskens et al., 2018; Tsagkrasoulis et al., 2017), whereas stronger environmental contributions observed in the lower parts of the face (e.g. lower lip, mandibular regions and chin) (Naqvi et al., 2022). Environmental influences such as nutrition, aging, oral function, prenatal exposure to teratogens, and climatic factors also contribute to facial shape variation.
Cichlid Diversity in Facial Morphology Beyond Skeletal Tissues
Cichlid fishes, particularly those from East African lakes, are renowned for their exceptional craniofacial diversity (Santos et al., 2023), making them a prime model for studying the genetic and developmental mechanisms underlying craniofacial variation (Powder and Albertson, 2016). This diversity encompasses a wide array of jaw structures, skull shapes, and facial soft tissues, each adapted to specific ecological niches and feeding strategies. While much research has focused on craniofacial skeletogenesis (Albertson and Kocher, 2006; Roberts et al., 2011, Singh et al 2017, WGCNA paper 2021, Ahi et al 3-lake qPCR paper), more recent studies have begun to explore the diversity in facial soft tissues, providing new insights into the evolutionary biology of these species (Concannon and Albertson, 2015; Conith et al., 2019, 2018; Duenser et al., 2023; Henning et al., 2017; Lecaudey et al., 2021, 2019). Historically, research has predominantly focused on the bony components of cichlid craniofacial anatomy. Studies have elucidated how variations in jaw morphology and skull structure contribute to the exploitation of diverse trophic resources (Fraser et al., 2009; Powder and Albertson, 2016). For instance, differences in jaw shape and kinematics and dental arrangements have been linked to specific feeding habits, such as algae scraping or mollusk crushing (Ahi et al., 2019; Fraser et al., 2009; Hu and Albertson, 2014; Singh et al., 2021). These skeletal adaptations are often governed by conserved molecular pathways, including Wnt and Hedgehog signaling, which modulate bone development and morphogenesis (Navon et al., 2020; Parsons et al., 2014; Roberts et al., 2011); however, novel genes have also been implicated (Powder et al., 2014; Gilbert et al., 2021), which underscores the potential of the cichlid system to reveal new molecular mechanisms into the regulation of craniofacial shape.
While skeletal structures have been extensively studied, there has been a burgeoning interest in the diversity of craniofacial soft tissues among cichlids. Soft tissue features, such as hypertrophied lips, forehead/nuchal humps, and nasal protrusions, play significant roles in feeding strategies, mate attraction, and species recognition (Baumgarten et al., 2015; Concannon and Albertson, 2015; Conith et al., 2019; Darrin Hulsey et al., 2018; Duenser et al., 2023; Henning et al., 2017; Lecaudey et al., 2021, 2019). These traits often evolve rapidly and can exhibit remarkable variability even among closely related species. Recent transcriptomic analyses have begun to uncover the molecular underpinnings of these soft tissue adaptations (Duenser et al., 2023; Lecaudey et al., 2021, 2019). For example, studies on species with pronounced lip hypertrophy have identified differential expression of genes involved in cell proliferation, extracellular matrix formation, and lipid metabolism (Henning et al., 2017; Lecaudey et al., 2021). Similarly, research into the development of nuchal humps, a prominent forehead swelling observed in certain cichlid species, has revealed the involvement of genes regulating adipogenesis, cell growth, and craniofacial morphogenesis (Lecaudey et al., 2019). These findings suggest that modifications in gene expression and signaling pathways contribute to the evolution of soft tissue diversity in cichlid fishes. Examples of soft tissue diversification in cichlids that are studied at molecular level are:
Hypertrophied Lips: In species such as Gnathochromis permaxillaris (so-called vacuum cleaner cichlid), exaggerated lip structures have evolved, potentially enhancing their ability to forage by creating a more effective seal against substrates. Transcriptomic studies have identified upregulation of genes associated with tissue remodeling and structural integrity in these species (Lecaudey et al., 2021). A recent comprehensive study assessed the convergent transcriptional changes underlying hypertrophied lips across the three Great Lakes of Africa, using Haplochromis chilotes, Placidochromis milomo, and Lobochilotes labiatus as models. Interestingly, they found a conserved set of overlapping ECM remodeling genes at the core of these repeated phenotypes (Machii et al., 2025).
Nuchal Humps: The dolphin cichlid (Cyrtocara moorii) from Lake Malawi in Eastern Africa exhibits a prominent nuchal hump (in forehead region), which may serve as a visual signal in social interactions or play a role in fat storage. Gene expression profiling of the nuchal hump tissue has highlighted the activation of pathways related to cell proliferation and differentiation (Lecaudey et al., 2019), indicating a complex regulatory network governing this trait.
Nasal Protrusions: Certain cichlid species from Lakes Tanganyika and Malawi, so called cat cichlids, display hypertrophied nasal structures, which are thought to be involved in either enhanced foraging efficiency or sexual selection. Investigations into these features have revealed the involvement of conserved cell, tissue and molecular players also implicated in human facial development, underscoring the relevance of cichlid models in understanding vertebrate facial diversity (Concannon and Albertson, 2015; Conith et al., 2019, 2018; Duenser et al., 2023).
In short, the rapid diversification of craniofacial traits in cichlids, driven by ecological pressures and sexual selection, offers insights into how genetic and environmental factors can interplay to produce phenotypic variation. Moreover, the molecular players identified during the morphogenesis of cichlid facial soft appear to be conserved across vertebrates (see examples in Figure 1), making these fishes pertinent models for understanding human craniofacial development, dysmorphology and diseases.
Examples of Conserved Genes Underlying Facial Soft Tissue Morphogenesis
Facial soft tissue morphogenesis is shaped by conserved genetic pathways that regulate mesenchymal proliferation, ECM remodeling, and epithelial-mesenchymal interactions. Many key genes involved in human and mouse craniofacial development also play roles in cichlid facial soft tissue formation. This section highlights a handful of example genes with known functions in mammalian facial development that have also been identified in cichlids with distinct nasal, forehead and lip hypertrophy. While not exhaustive, these examples illustrate the molecular parallels between cichlids and mammals, reinforcing their value as a model for studying vertebrate facial soft tissue evolution and morphogenesis (Figure 2).
Growth regulators: Growth factor signals play a central role in facial soft tissue morphogenesis, directing mesenchymal proliferation, differentiation, and extracellular matrix (ECM) remodeling. In cichlid fish, these pathways contribute to three key facial modifications, nasal, forehead (nuchal hump), and lip hypertrophy, through a conserved set of genetic regulators. The genes amad12, tgf-β1, fgf22, foxf1, foxa2, bmp2, and tfap4 drive nasal protrusion development (Conith et al., 2018; Duenser et al., 2023), while pdgfrb, btg3, and tcf12 regulate forehead hypertrophy (Lecaudey et al., 2019). Meanwhile, foxp1, tcf12, and foxf1 contribute to lip hypertrophy (Lecaudey et al., 2021). The shared features among these three morphogenetic processes likely reflect a conserved role in driving similar tissue-level outcomes, particularly the hypertrophy of facial soft tissues.
In human, TGF-β1, BMP2, FGF22, and TFAP4 play critical roles in epithelial-mesenchymal interactions and mesenchymal proliferation, facilitating expansion of nasal soft tissues (Gupta et al., 2020). TGF-β1, a key regulator of tissue remodeling and fibroblast activation, has been linked to nasal tissue expansion in cichlids, similar to its role in human craniofacial development (Gupta et al., 2020). BMP2, a fundamental driver of soft tissue morphogenesis, influences cell fate and ECM production, reinforcing nasal mesenchymal structure (Tan et al., 2017). FGF22, involved in epidermal homeostasis and tissue repair, contributes to the regulation of epithelial integrity during nasal expansion (Quigley et al., 2004). FOXA2 and FOXF1, transcription factors crucial for mesodermal and endodermal tissue specification, have been identified in nasal mesenchyme, where they modulate mesenchymal-epithelial signaling, further supporting their role in soft tissue patterning across vertebrates (Dines et al., 2019; Kucharczyk et al., 2014). TFAP4, a transcription factor regulating cell proliferation and differentiation, has been implicated in as potential upstream regulator of gene networks involved in epithelial thickening in nasal protrusions (Gervasini et al., 2007), which is also a feature of nasal expansion in cichlid species (Conith et al., 2019).
For forehead hypertrophy (nuchal hump formation), pdgfrb, btg3 and tcf12 regulate fibroblast proliferation, mesenchymal expansion, and connective tissue remodeling (Lecaudey et al., 2019). Pdgfrb (Platelet-Derived Growth Factor Receptor Beta) is a major player in connective tissue expansion and fibroblast proliferation, and its expression is significantly upregulated in cichlids with pronounced forehead structures (Lecaudey et al., 2019), mirroring its role in mammalian craniofacial soft tissue development (Bredrup et al., 2018; Fantauzzo and Soriano, 2016; McCarthy et al., 2016; Saultz et al., 2016; Takenouchi et al., 2015). In human, BTG3, a cell cycle regulator, modulates mesenchymal proliferation, ensuring controlled facial soft tissue thickening (Errichiello et al., 2016). In mammals, TCF12, a transcription factor essential for tissue differentiation and patterning, contributes to species-specific variation in forehead structure, demonstrating its conserved function in soft tissue morphogenesis (Le Tanno et al., 2014; Piard et al., 2015; Sharma et al., 2013).
In the upper lip specific hypertrophy, foxp1, tcf12 and foxf1 are implicated play role in epithelial thickening, mesenchymal expansion, and fibroblast differentiation (Lecaudey et al., 2021). FOXP1 and FOXF1, both involved in mesenchymal-epithelial interactions, are suggested to play roles in fibroblast differentiation and ECM organization in cichlid lips, most likely mirroring their function in human craniofacial soft tissue development (Meerschaut et al., 2017; Shaw-Smith, 2010; Xu et al., 2016). TCF12, also found in forehead hypertrophy, is suggested to contribute to tissue remodeling and connective tissue specialization in the upper lip region, reinforcing its widespread role in facial soft tissue variation (Piard et al., 2015). The recurrent use of TCF12 and FOXF1 in multiple regions suggests that growth factor signaling is modular and adaptable, allowing for diverse morphological outcomes across species. This highlights cichlids as an effective model system for investigating the molecular basis of facial soft tissue development and variation in vertebrates, including humans.
Regulators of cell differentiation: Cell differentiation is a fundamental process in facial soft tissue morphogenesis, directing the specialization of fibroblasts, mesenchymal cells, and epithelial cells across different facial regions. In cichlid fish, conserved regulatory genes have been identified in nasal protrusions, forehead hypertrophy, and lip hypertrophy, playing key roles in tissue-specific differentiation. Among these, scx (Scleraxis) and six2 are implicated in nasal protrusion development, particularly in modulating ligamentous tissues, connective tissue and mesenchymal cell specification (Conith et al., 2018; Duenser et al., 2023). Scx, a well-known transcription factor regulating tendon and ligament differentiation, has been implicated in the structural integrity of nasal mesenchyme (Sugimoto et al., 2013), ensuring proper tissue remodeling and flexibility in species with elongated nasal protrusions (Conith et al., 2018). Similarly, six2, which plays a major role in craniofacial mesenchymal stem cell regulation, is crucial for proliferation and fate determination in nasal soft tissues (Hufnagel et al., 2016; Okello et al., 2017), supporting its role in vertebrate facial morphogenesis.
In forehead hypertrophy (nuchal hump formation), genes such as dlk1, papp-a, dlx5a, fosl1a, and mycn regulate mesenchymal differentiation, connective tissue expansion, and adipogenesis (Lecaudey et al., 2019). Dlk1 (Delta-Like 1 Homolog), a well-known Notch signaling modulator, functions as an inhibitor of adipocyte differentiation, potentially influencing fat accumulation patterns in the nuchal hump, and in mammals is also involved in facial soft tissue morphogenesis (Abdallah et al., 2011; Begemann et al., 2012). In contrast, Papp-a (Pregnancy-Associated Plasma Protein-A) enhances IGF signaling, promoting mesenchymal growth and expansion in the forehead region, as shown in mice (Conover et al., 2004). Dlx5, a transcription factor with conserved role in morphogenesis of various craniofacial soft tissue (Chung et al., 2010; Holleville et al., 2003; Talbot et al., 2010; Vera-Carbonell et al., 2012), is also upregulated in cichlids with prominent forehead structures. In human, FOSL1A, part of the Ap-1 transcription factor complex, is linked to connective tissue patterning (Mirzamohammadi et al., 2018; Van Mater et al., 2013; Yu et al., 2018), while MYCN, a key cell cycle regulator, drives mesenchymal cell proliferation (Mirzamohammadi et al., 2018; Van Mater et al., 2013; Yu et al., 2018), supporting its proposed role in formation of the nuchal hump morphology.
For the upper lip specific hypertrophy, rasal3, fhl2 and igf2 regulate fibroblast differentiation, epithelial remodeling, and mesenchymal expansion (Lecaudey et al., 2021). RASAL3, a Ras-GTPase activator, modulates fibroblast signaling, ensuring balanced lip tissue differentiation and proliferation (Draaken et al., 2013; Kosaki et al., 2011). FHL2, a LIM-domain protein, integrates mechanical and biochemical signals to regulate muscle and connective tissue adaptation, potentially influencing lip thickness and elasticity in hypertrophied cichlid lips (Labalette et al., 2008; Manousaki et al., 2013; Ng et al., 2011). IGF2 (Insulin-like Growth Factor 2), a key driver of tissue growth and differentiation, is upregulated in cichlids with exaggerated lips, where it likely promotes mesenchymal proliferation and ECM remodeling (Peñaherrera et al., 2010). Together, these differentiation regulators illustrate the molecular conservation of facial soft tissue morphogenesis, emphasizing the functional parallels between cichlid and mammalian craniofacial development.
Modulators of tissue remodeling: The morphogenesis of facial soft tissues relies not only on structural ECM components but also critically on modulators that guide tissue remodeling, including matrix turnover, cellular behavior, and microenvironmental signaling. These modulators orchestrate dynamic ECM remodeling processes that shape the face during development and evolution. In cichlid fishes, several genes with conserved roles in human craniofacial morphogenesis have been identified as key players in region-specific hypertrophic phenotypes, particularly in the nasal protrusion, forehead, and lip regions. Their functional convergence across these domains highlights the modular yet integrated nature of facial soft tissue evolution.
In the context of nasal protrusion development, three modulator genes; adam12, ugdh, and hmx2, have been shown to exhibit coordinated expression and function (Duenser et al., 2023). ADAM12, a metalloprotease-disintegrin, is known to influence ECM remodeling by modulating cell-matrix interactions and activating latent growth factors such as TGF-β (Ruff et al., 2015), which are crucial for mesenchymal proliferation and migration. Its involvement in nasal protrusion formation in cichlids mirrors its known role in midfacial growth in mammals (Conith et al., 2018; Feng et al., 2009). UGDH (UDP-glucose 6-dehydrogenase), by catalyzing the production of glycosaminoglycan precursors such as hyaluronic acid, plays a central role in regulating ECM hydration and compressibility, factors essential for soft tissue expansion and morphogenesis (Alhamoudi et al., 2020). HMX2, a transcription factor, modulates gene networks involved in facial development and has been associated with nasal and midfacial prominence shaping in vertebrates (Miller et al., 2009). Together, these three genes likely form a regulatory module that facilitates nasal protrusion via localized ECM remodeling and cellular proliferation.
A similar tissue remodeling theme underlies the exaggerated soft tissue phenotype observed in the forehead region, particularly in species exhibiting nuchal humps (Lecaudey et al., 2019). Dermatopontin (dpt), a small ECM protein with known roles in collagen fibrillogenesis and tissue organization, is markedly upregulated in species with this trait. In mammals, DPT is involved in skin matrix integrity and dermal fibroblast function (Krishnaswamy et al., 2017; Krishnaswamy and Korrapati, 2015; Liu et al., 2013), and its upregulation in cichlid forehead tissues suggests a role in maintaining the tensile and structural properties of the expanded connective tissue mass. Unlike purely structural ECM components, DPT interacts dynamically with other matrix molecules, modifying the extracellular environment to support prolonged soft tissue hypertrophy.
The upper lip region provides another example of how tissue remodeling modulators contribute to localized facial outgrowths. Genes such as lama5, apoda, ggt5a, and col21a1 act in synergy to drive lip hypertrophy (Lecaudey et al., 2021; Masonick et al., 2023). LAMA5, a laminin subunit, contributes to the basement membrane and mediates epithelial–mesenchymal interactions crucial for lip tissue adhesion and outgrowth (Peixoto da-Silva et al., 2012). Apoda, a member of the apolipoprotein family, has putative roles in lipid transport and may contribute to the metabolic support of growing soft tissues (Dassati et al., 2014; Manousaki et al., 2013). GGT5A (gamma-glutamyltransferase 5) participates in redox balance and amino acid metabolism, potentially influencing cellular proliferation within remodeling regions (Pinchefsky et al., 2017). Meanwhile, COL21A1, a FACIT collagen, modulates the interaction of fibrillar collagens with surrounding ECM components and plays a more regulatory than structural role in maintaining ECM architecture (Mohamad Shah et al., 2019). These genes collectively modulate the microenvironment of the lip region, promoting region-specific hypertrophy via ECM remodeling and enhanced tissue plasticity.
Most strikingly, a conserved module of ECM regulator genes; decorin (dcn), lumican (lum), and asporin (aspn), has been recently implicated in the hypertrophy of the entire lips (both upper and lower), a trait observed repeatedly and independently across the three Great African Lakes (Victoria, Malawi, and Tanganyika) (Machii et al., 2025). These genes encode small leucine-rich proteoglycans (SLRPs) that modulate collagen fibrillogenesis, matrix organization, and tissue elasticity. In mammals, these SLRPs are essential for soft connective tissue integrity and remodeling, particularly in skin and cartilage (Pang et al., 2020). Their shared upregulation in cichlids with hypertrophied lips suggests that these genes orchestrate a conserved remodeling program capable of generating robust, convergent phenotypes (Machii et al., 2025). Their repeated recruitment across evolutionary lineages supports a model where modular ECM remodeling pathways are co-opted to drive parallel facial adaptations in distinct ecological and phylogenetic contexts. Thus, dcn, lum, and aspn exemplify how conserved genetic pathways can underpin repeated morphogenetic outcomes across species.
Structural factors: Structural genes encoding core ECM components and scaffolding proteins support facial morphology by providing biomechanical structure and regulating signaling pathways. In cichlid fishes, which show diverse facial soft tissue forms, regionally expressed structural genes are linked to distinct hypertrophic traits. These patterns are consistent with their roles in vertebrate craniofacial development and suggest relevance to human facial morphogenesis.
In the context of nasal protrusion, three structural genes, adprhl1, dusp22, and lyve1, have emerged as potential important players (Duenser et al., 2023). ADPRHL1 encodes a muscle-specific, actin-associated protein related to the ADP-ribosylation family. While it has been most studied in cardiac and skeletal muscle contexts, its association with tissue scaffolding and actin cytoskeleton remodeling suggests a structural role in shaping the soft nasal tissue matrix during protrusion (De Pater et al., 2005). DUSP22, though primarily known as a dual-specificity phosphatase, has emerging links to cytoskeletal dynamics and tissue architecture, potentially acting through regulation of cell-matrix interactions (Hosono et al., 2020). LYVE1, a lymphatic vessel endothelial hyaluronan receptor, is an ECM-interacting protein implicated in hyaluronan transport and matrix hydration (Mitteldorf et al., 2018). Its expression in nasal protrusion zones of cichlids suggests it contributes to the viscoelastic properties and expansion potential of this soft tissue region, echoing its role in mucosal tissues in mammals.
The most extensive array of structural ECM genes is associated with the whole-lip hypertrophy phenotype observed in multiple cichlid lineages across the African Great Lakes. This striking convergence has been underpinned by upregulation of collagen type I alpha chains (col1a1a, col1a1b, col1a2), fibrillar collagens that provide tensile strength and form the backbone of connective tissues (Machii et al., 2025). These genes are fundamental in maintaining dermal structure and are conserved components in vertebrate skin and oral tissue (Arseni et al., 2018; Lim et al., 2020). Their repeated involvement in lip thickening across lakes highlights their evolutionary utility in reinforcing expanded soft tissues. Accompanying the collagen framework are elastin microfibril interface-located proteins (emilin1b, emilin2a, emilin2b, emilin3b), which regulate elastogenesis and microfibril formation (Machii et al., 2025). These proteins not only maintain elasticity and structural resilience in soft tissues like lips but also influence TGF-β signaling, linking them to morphogenetic regulation. In humans, EMILIN family members are expressed in elastic connective tissues such as the dermis and oral mucosa, suggesting deep conservation of their function in lip flexibility and expansion (Fitoussi et al., 2019; Schiavinato et al., 2024).
Further supporting this ECM complex are hyaluronan and proteoglycan link proteins (hapln1a/b, hapln3) and versican (vcanb), which stabilize the hyaluronan-based matrix and confer compressibility and hydration to soft tissues. These genes contribute to matrix swelling and pliability, crucial properties in the hypertrophied lips of many cichlid species (Machii et al., 2025). Similarly, periostin (postna/b), an ECM glycoprotein known for its role in collagen cross-linking and tissue remodeling, is enriched in cichlid lip tissues and is recognized in mammals for its involvement in periodontal ligament and skin ECM maintenance (Machii et al., 2025). Collectively, these genes form a robust structural gene network that underlies the convergent evolution of full-lip hypertrophy in geographically and phylogenetically distinct cichlid lineages.
Finally, some other structural contributors like cldn7 and actb involved in ECM remodeling may also play supportive role (Colombo et al., 2013). CLDN7 (claudin 7), a tight junction protein, helps maintain epithelial integrity and may influence localized adhesion and barrier properties at the lip margin. ACTB (beta-actin), while broadly expressed, contributes to cytoskeletal tension and interacts with ECM structures during tissue expansion. Though not ECM in the classical sense, these proteins support the structural integrity of soft tissue outgrowths and interact closely with matrix components.
Other potential molecular players: Beyond structural and remodeling genes, other molecular players, such as ion transporters, and immune regulators, support facial soft tissue morphogenesis by coordinating signaling and tissue homeostasis. In cichlids, their region-specific expression suggests roles in traits like nasal protrusion, nuchal humps, and lip hypertrophy, offering insights into conserved regulatory mechanisms across vertebrates.
Among mediators of cell communication and ion transporters, several genes have been associated with distinct facial regions. In nasal protrusion, prodh and rac1 stand out (Duenser et al., 2023). PRODH (proline dehydrogenase) is involved in proline metabolism and influences redox signaling, which can affect cell proliferation and tissue remodeling during craniofacial development (Guilmatre et al., 2010). RAC1, a small GTPase, regulates actin cytoskeletal dynamics, cell adhesion, and migration, functions that are critical for shaping the protruding nasal soft tissues (Reijnders et al., 2017; Thomas et al., 2010). These genes likely work together to modulate the cellular environment during tissue outgrowth, contributing to the distinctive nasal elongation seen in certain cichlid species.
In the forehead region, characterized in some species by a hypertrophic nuchal hump, ago3 (argonaute 3) has been identified (Lecaudey et al., 2019). This gene belongs to the Argonaute family involved in RNA interference and gene silencing. Through its regulatory influence on post-transcriptional gene expression, AGO3 may modulate the activity of other morphogenetic genes during forehead development, particularly those involved in ECM production or cell proliferation. In human, the function of AGO3 has been linked to in facial morphogenesis including forehead, nasal and palpebral tissues (Tokita et al., 2015). Thus, its expression in the forehead region suggests a role in finely tuning gene expression programs during soft tissue thickening or expansion, although functional studies are still needed to elucidate the exact mechanisms.
In the upper lip region, a different set of genes, including kcnj16, kcnj2a, and cyp1a, appear to mediate region-specific soft tissue hypertrophy (Lecaudey et al., 2021). Kcnj16 and kcnj2a encode inward-rectifier potassium channels, which regulate cellular membrane potential and ionic balance, potentially influencing cell volume and tissue tension in lip outgrowth areas (Lecaudey et al., 2021). These ion transporters may indirectly affect signaling pathways or mechanical properties of cells during hypertrophy. CYP1A, a member of the cytochrome P450 family, is involved in xenobiotic metabolism and may modulate local oxidative states, contributing to localized tissue remodeling or differentiation (Linnenkamp et al., 2020; Stuppia et al., 2011). Together, these genes suggest a complex interplay between bioelectric signaling, metabolism, and tissue growth in shaping the unique upper lip profiles of certain cichlid species.
In parallel, inflammatory and immune-related genes have also been implicated in region-specific facial traits, particularly in the forehead and upper lip. In the forehead, CD163, a scavenger receptor expressed by macrophages, and SOCS2 (Suppressor of Cytokine Signaling 2), which modulates cytokine responses (Lecaudey et al., 2019), point to a role for immune regulation in soft tissue hypertrophy. These genes may contribute to the regulation of inflammation during active tissue growth or maintenance of tissue homeostasis in hypertrophic zones, especially in contexts of rapid remodeling or expansion (Farquharson and Ahmed, 2013; Greenhalgh et al., 2005; Greenhalgh and Alexander, 2004; MacRae et al., 2009; Metcalf et al., 2000; Sandell et al., 2015; Vos et al., 2005). The upregulation of immune genes in this region underscores the emerging view that developmental and immune pathways are deeply intertwined.
In the upper lip specific hypertrophy, GIMAP8 (GTPase of the immunity-associated protein family) has been identified (Lecaudey et al., 2021). Known for roles in lymphocyte survival and immune modulation (Zhang et al., 2024), GIMAP8 may influence lip tissue morphogenesis by shaping the local immune environment, possibly affecting cell turnover or response to mechanical stress during lip growth (Manousaki et al., 2013). The presence of inflammatory regulators in facial tissues further suggests that localized immune activity may serve as a permissive or modulatory factor during morphogenesis, rather than being restricted to host defense.
Conclusions
Facial soft tissue morphogenesis represents a complex and largely understudied frontier in developmental biology; one shaped by the interplay of genetic regulation, cellular plasticity, and evolutionary innovation. Cichlid fishes offer a uniquely powerful model for unraveling the mechanisms driving facial soft tissue diversity, thanks to their rich natural variation and experimental accessibility. Importantly, the genes involved in shaping facial soft tissues often serve multiple roles across different biological processes. For instance, regulators like adam12, known for roles in ECM remodeling, may also influence signaling environments and cell behavior, demonstrating how a single gene can contribute to distinct outcomes depending on context. This genetic pleiotropy begs the need to move beyond linear gene-function models and toward integrative frameworks that consider how genetic networks interact within dynamic multidimendional developmental landscapes. Cichlid models now position us to explore these complexities at both molecular and evolutionary scales. As we begin to map the genetic architecture underlying soft tissue variation, we not only deepen our understanding of vertebrate facial development but also lay the groundwork for translational insights into human facial diversity and congenital disorders.
References
- Abdallah, B.M.; Ditzel, N.; Mahmood, A.; Isa, A.; Traustadottir, G.A.; Schilling, A.F.; Ruiz-Hidalgo, M.-J.; Laborda, J.; Amling, M.; Kassem, M. DLK1 is a novel regulator of bone mass that mediates estrogen deficiency-induced bone loss in mice. J. Bone Miner. Res. 2011, 26, 1457–71. [Google Scholar] [CrossRef] [PubMed]
- Ahi, E.P. Signalling pathways in trophic skeletal development and morphogenesis: Insights from studies on teleost fish. Dev. Biol. 2016, 420, 11–31. [Google Scholar] [CrossRef]
- Ahi, E.P.; Singh, P.; Duenser, A.; Gessl, W.; Sturmbauer, C. Divergence in larval jaw gene expression reflects differential trophic adaptation in haplochromine cichlids prior to foraging. BMC Evol. Biol. 2019, 19, 150. [Google Scholar] [CrossRef]
- Albertson, R.C.; Kocher, T.D. Genetic and developmental basis of cichlid trophic diversity. Heredity (Edinb) 2006, 97, 211–221. [Google Scholar] [CrossRef]
- Albertson, R.C.; Streelman, J.T.; Kocher, T.D. Directional selection has shaped the oral jaws of Lake Malawi cichlid fishes. PNAS 2003, 100, 5252–5257. [Google Scholar] [CrossRef]
- Alhamoudi, K.M.; Bhat, J.; Nashabat, M.; Alharbi, M.; Alyafee, Y.; Asiri, A.; Umair, M.; Alfadhel, M. A Missense Mutation in the UGDH Gene Is Associated With Developmental Delay and Axial Hypotonia. Front. Pediatr. 2020, 8. [Google Scholar] [CrossRef]
- Arseni, L.; Lombardi, A.; Orioli, D. From Structure to Phenotype: Impact of Collagen Alterations on Human Health; Int. J. Mol. Sci., 2018; Vol. 19, Page 1407 19. [Google Scholar] [CrossRef]
- Baumgarten, L.; Machado-Schiaffino, G.; Henning, F.; Meyer, A. What big lips are good for: On the adaptive function of repeatedly evolved hypertrophied lips of cichlid fishes. Biol. J. Linn. Soc. 2015, 115, 448–455. [Google Scholar] [CrossRef]
- Begemann, M.; Spengler, S.; Kordaß, U.; Schröder, C.; Eggermann, T. Segmental maternal uniparental disomy 7q associated with DLK1/GTL2 (14q32) hypomethylation. Am. J. Med. Genet. Part A 2012, 158A, 423–428. [Google Scholar] [CrossRef]
- Brawand, D., Wagner, C.E., Li, Y.I., Malinsky, M., Keller, I., Fan, S., Simakov, O., Ng, A.Y., Lim, Z.W., Bezault, E., Turner-Maier, J., Johnson, J., Alcazar, R., Noh, H.J., Russell, P., Aken, B., Alföldi, J., Amemiya, C., Azzouzi, N., Baroiller, J.-F., Barloy-Hubler, F., Berlin, A., Bloomquist, R., Carleton, K.L., Conte, M. a., D’Cotta, H., Eshel, O., Gaffney, L., Galibert, F., Gante, H.F., Gnerre, S., Greuter, L., Guyon, R., Haddad, N.S., Haerty, W., Harris, R.M., Hofmann, H. a., Hourlier, T., Hulata, G., Jaffe, D.B., Lara, M., Lee, A.P., MacCallum, I., Mwaiko, S., Nikaido, M., Nishihara, H., Ozouf-Costaz, C., Penman, D.J., Przybylski, D., Rakotomanga, M., Renn, S.C.P., Ribeiro, F.J., Ron, M., Salzburger, W., Sanchez-Pulido, L., Santos, M.E., Searle, S., Sharpe, T., Swofford, R., Tan, F.J., Williams, L., Young, S., Yin, S., Okada, N., Kocher, T.D., Miska, E. a., Lander, E.S., Venkatesh, B., Fernald, R.D., Meyer, A., Ponting, C.P., Streelman, J.T., Lindblad-Toh, K., Seehausen, O., Di Palma, F., 2014. The genomic substrate for adaptive radiation in African cichlid fish. Nature 513, 375–381. [CrossRef]
- Bredrup, C.; Stokowy, T.; McGaughran, J.; Lee, S.; Sapkota, D.; Cristea, I.; Xu, L.; Tveit, K.S.; Høvding, G.; Steen, V.M.; Rødahl, E.; Bruland, O.; Houge, G. A tyrosine kinase-activating variant Asn666Ser in PDGFRB causes a progeria-like condition in the severe end of Penttinen syndrome. Eur. J. Hum. Genet. 2018, 1. [Google Scholar] [CrossRef] [PubMed]
- Bull, R.; Rumsey, N. The Social Psychology of Facial Appearance, The Social Psychology of Facial Appearance; Springer New York, 1988. [Google Scholar] [CrossRef]
- Carmona Baez, A.; Ciccotto, P.J.; Moore, E.C.; Peterson, E.N.; Lamm, M.S.; Roberts, N.B.; Coyle, K.P.; Barker, M.K.; Dickson, E.; Cass, A.N.; Pereira, G.S.; Zeng, Z.-B.; Guerrero, R.F.; Roberts, R.B. Gut length evolved under sexual conflict in Lake Malawi cichlids. Genetics 2025, 230. [Google Scholar] [CrossRef] [PubMed]
- Chaimongkhol, T.; Mahakkanukrauh, P. The facial soft tissue thickness related facial reconstruction by ultrasonographic imaging: A review. Forensic Sci. Int. 2022, 337, 111365. [Google Scholar] [CrossRef]
- Chung, I.-H.; Han, J.; Iwata, J.; Chai, Y. Msx1 and Dlx5 function synergistically to regulate frontal bone development. genesis 2010, 48, 645–655. [Google Scholar] [CrossRef]
- Clark, B.; Kuwalekar, M.; Fischer, B.; Woltering, J.; Biran, J.; Juntti, S.; Kratochwil, C.F.; Santos, M.E.; Almeida, M.V. Genome editing in East African cichlids and tilapias: state-of-the-art and future directions. Open Biol 2023, 13. [Google Scholar] [CrossRef]
- Colombo, M.; Diepeveen, E.T.; Muschick, M.; Santos, M.E.; Indermaur, A.; Boileau, N.; Barluenga, M.; Salzburger, W. The ecological and genetic basis of convergent thick-lipped phenotypes in cichlid fishes. Mol. Ecol. 2013, 22, 670–684. [Google Scholar] [CrossRef] [PubMed]
- Concannon, M.R.; Albertson, R.C. The genetic and developmental basis of an exaggerated craniofacial trait in East African cichlids. J. Exp. Zool. B. Mol. Dev. Evol. 2015, 324, 662–70. [Google Scholar] [CrossRef] [PubMed]
- Conith, A.J.; Albertson, R.C. The cichlid oral and pharyngeal jaws are evolutionarily and genetically coupled. Nat. Commun. 2021, 2021 121 12, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Conith, M.R.; Conith, A.J.; Albertson, R.C. Evolution of a soft-tissue foraging adaptation in African cichlids: Roles for novelty, convergence, and constraint. Evolution (N. Y). 2019, 73, 2072–2084. [Google Scholar] [CrossRef]
- Conith, M.R.; Hu, Y.; Conith, A.J.; Maginnis, M.A.; Webb, J.F.; Craig Albertson, R. Genetic and developmental origins of a unique foraging adaptation in a Lake Malawi cichlid genus. Proc. Natl. Acad. Sci. U. S. A. 2018, 115, 7063–7068. [Google Scholar] [CrossRef]
- Conover, C.A.; Bale, L.K.; Overgaard, M.T.; Johnstone, E.W.; Laursen, U.H.; Füchtbauer, E.-M.; Oxvig, C.; van Deursen, J. Metalloproteinase pregnancy-associated plasma protein A is a critical growth regulatory factor during fetal development. Development 2004, 131, 1187–94. [Google Scholar] [CrossRef]
- Cooper, W.J.; Parsons, K.; McIntyre, A.; Kern, B.; McGee-Moore, A.; Albertson, R.C. Bentho-Pelagic Divergence of Cichlid Feeding Architecture Was Prodigious and Consistent during Multiple Adaptive Radiations within African Rift-Lakes. PLoS One 2010, 5, e9551. [Google Scholar] [CrossRef]
- Cordero, D.R.; Brugmann, S.; Chu, Y.; Bajpai, R.; Jame, M.; Helms, J.A. Cranial neural crest cells on the move: their roles in craniofacial development. Am. J. Med. Genet. A 2011, 155A, 270–279. [Google Scholar] [CrossRef]
- Cotofana, S.; Lachman, N. Anatomy of the Facial Fat Compartments and their Relevance in Aesthetic Surgery. JDDG J. der Dtsch. Dermatologischen Gesellschaft 2019, 17, 399–413. [Google Scholar] [CrossRef]
- Darrin Hulsey, C.; Zheng, J.; Holzman, R.; Alfaro, M.E.; Olave, M.; Meyer, A. Phylogenomics of a putatively convergent novelty: Did hypertrophied lips evolve once or repeatedly in Lake Malawi cichlid fishes? BMC Evol. Biol. 2018, 18, 179. [Google Scholar] [CrossRef] [PubMed]
- Dassati, S.; Waldner, A.; Schweigreiter, R. Apolipoprotein D takes center stage in the stress response of the aging and degenerative brain. Neurobiol. Aging. 2014. [Google Scholar] [CrossRef] [PubMed]
- De Pater, J.M.; Nikkels, P.G.J.; Poot, M.; Eleveld, M.J.; Stigter, R.H.; Van Der Sijs-Bos, C.J.M.; Loneus, W.H.; Engelen, J.J.M. Striking Facial Dysmorphisms and Restricted Thymic Development in a Fetus with a 6-Megabase Deletion of Chromosome 14q. Pediatr. Dev. Pathol. 2005, 8, 497–503. [Google Scholar] [CrossRef]
- Dines, J.N.; Liu, Y.J.; Neufeld-Kaiser, W.; Sawyer, T.; Ishak, G.E.; Tully, H.M.; Racobaldo, M.; Sanchez-Valle, A.; Disteche, C.M.; Juusola, J.; Torti, E.; McWalter, K.; Doherty, D.; Dipple, K.M. Expanding phenotype with severe midline brain anomalies and missense variant supports a causal role for FOXA2 in 20p11.2 deletion syndrome. Am. J. Med. Genet. Part A 2019, 179, ajmg.a.61281. [Google Scholar] [CrossRef]
- Draaken, M.; Mughal, S.S.; Pennimpede, T.; Wolter, S.; Wittler, L.; Ebert, A.-K.; Rösch, W.; Stein, R.; Bartels, E.; Schmidt, D.; Boemers, T.M.; Schmiedeke, E.; Hoffmann, P.; Moebus, S.; Herrmann, B.G.; Nöthen, M.M.; Reutter, H.; Ludwig, M. Isolated bladder exstrophy associated with a de novo 0.9 Mb microduplication on chromosome 19p13.12. Birth Defects Res. Part A Clin. Mol. Teratol. 2013, 97, 133–139. [Google Scholar] [CrossRef]
- Duenser, A.; Singh, P.; Lecaudey, L.A.; Sturmbauer, C.; Albertson, R.C.; Gessl, W.; Ahi, E.P. Conserved Molecular Players Involved in Human Nose Morphogenesis Underlie Evolution of the Exaggerated Snout Phenotype in Cichlids. Genome Biol. Evol. 2023, 15. [Google Scholar] [CrossRef]
- El Sergani, A.M.; Brandebura, S.; Padilla, C.; Butali, A.; Adeyemo, W.L.; Valencia-Ramírez, C.; Muñeton, C.P.R.; Moreno, L.M.; Buxó, C.J.; Long, R.E.; Neiswanger, K.; Shaffer, J.R.; Marazita, M.L.; Weinberg, S.M. Parents of Children With Nonsyndromic Orofacial Clefting Show Altered Palate Shape. Cleft Palate-Craniofacial J 2021, 58, 847–853. [Google Scholar] [CrossRef]
- Errichiello, E.; Novara, F.; Cremante, A.; Verri, A.; Galli, J.; Fazzi, E.; Bellotti, D.; Losa, L.; Cisternino, M.; Zuffardi, O. Dissection of partial 21q monosomy in different phenotypes: clinical and molecular characterization of five cases and review of the literature. Mol. Cytogenet. 2016, 9, 21. [Google Scholar] [CrossRef] [PubMed]
- Fantauzzo, K.A.; Soriano, P. PDGFRβ regulates craniofacial development through homodimers and functional heterodimers with PDGFRα. Genes Dev 2016, 30, 2443–2458. [Google Scholar] [CrossRef] [PubMed]
- Farquharson, C.; Ahmed, S.F. Inflammation and linear bone growth: the inhibitory role of SOCS2 on GH/IGF-1 signaling. Pediatr. Nephrol. 2013, 28, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.; Leach, S.M.; Tipney, H.; Phang, T.; Geraci, M.; Spritz, R.A.; Hunter, L.E.; Williams, T. Spatial and Temporal Analysis of Gene Expression during Growth and Fusion of the Mouse Facial Prominences. PLoS One 2009, 4, e8066. [Google Scholar] [CrossRef]
- Finucane, H.K.; Bulik-Sullivan, B.; Gusev, A.; Trynka, G.; Reshef, Y.; Loh, P.R.; Anttila, V.; Xu, H.; Zang, C.; Farh, K.; Ripke, S.; Day, F.R.; Purcell, S.; Stahl, E.; Lindstrom, S.; Perry, J.R.B.; Okada, Y.; Raychaudhuri, S.; Daly, M.J.; Patterson, N.; Neale, B.M.; Price, A.L. Partitioning heritability by functional annotation using genome-wide association summary statistics. Nat. Genet. 2015, 2015 4711 47, 1228–1235. [Google Scholar] [CrossRef]
- Fitoussi, R.; Beauchef, G.; Guéré, C.; André, N.; Vié, K. Localization, fate and interactions of Emilin-1 in human skin. Int. J. Cosmet. Sci. 2019, 41, 183–193. [Google Scholar] [CrossRef]
- Fraser, G.J.; Hulsey, C.D.; Bloomquist, R.F.; Uyesugi, K.; Manley, N.R.; Streelman, J.T. An ancient gene network is co-opted for teeth on old and new jaws. PLoS Biol 2009, 7, e31. [Google Scholar] [CrossRef]
- Fryer, G.; Iles, T.D. The Cichlid Fishes of the Great lakes of Africa: their Biology and Evolution. In Oliver and Boyd; Edinburgh, 1972. [Google Scholar]
- Gervasini, C.; Castronovo, P.; Bentivegna, A.; Mottadelli, F.; Faravelli, F.; Giovannucci-Uzielli, M.L.; Pessagno, A.; Lucci-Cordisco, E.; Pinto, A.M.; Salviati, L.; Selicorni, A.; Tenconi, R.; Neri, G.; Larizza, L. High frequency of mosaic CREBBP deletions in Rubinstein-Taybi syndrome patients and mapping of somatic and germ-line breakpoints. Genomics 2007, 90, 567–573. [Google Scholar] [CrossRef]
- Giri, J.; Bockmann, M.; Brook, A.; Gurr, A.; Hughes, T. Genetic and environmental contributions to the development of soft tissue facial profile: a twin study. Eur. J. Orthod. 2024, 46. [Google Scholar] [CrossRef]
- Greenhalgh, C.J.; Alexander, W.S. Suppressors of cytokine signalling and regulation of growth hormone action. Growth Horm. IGF Res. 2004, 14, 200–6. [Google Scholar] [CrossRef]
- Greenhalgh, C.J.; Rico-Bautista, E.; Lorentzon, M.; Thaus, A.L.; Morgan, P.O.; Willson, T.A.; Zervoudakis, P.; Metcalf, D.; Street, I.; Nicola, N.A.; Nash, A.D.; Fabri, L.J.; Norstedt, G.; Ohlsson, C.; Flores-Morales, A.; Alexander, W.S.; Hilton, D.J. SOCS2 negatively regulates growth hormone action in vitro and in vivo. J. Clin. Invest. 2005, 115, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Guilmatre, A.; Legallic, S.; Steel, G.; Willis, A.; Di Rosa, G.; Goldenberg, A.; Drouin-Garraud, V.; Guet, A.; Mignot, C.; Des Portes, V.; Valayannopoulos, V.; Van Maldergem, L.; Hoffman, J.D.; Izzi, C.; Espil-Taris, C.; Orcesi, S.; Bonafé, L.; Le Galloudec, E.; Maurey, H.; Ioos, C.; Afenjar, A.; Blanchet, P.; Echenne, B.; Roubertie, A.; Frebourg, T.; Valle, D.; Campion, D. Type I hyperprolinemia: genotype/phenotype correlations. Hum. Mutat. 2010, 31, 961–965. [Google Scholar] [CrossRef]
- Gupta, P.; Tripathi, T.; Singh, N.; Bhutiani, N.; Rai, P.; Gopal, R. A review of genetics of nasal development and morphological variation. J. Fam. Med. Prim. Care 2020, 9, 1825. [Google Scholar] [CrossRef]
- Henning, F.; Machado-Schiaffino, G.; Baumgarten, L.; Meyer, A. Genetic dissection of adaptive form and function in rapidly speciating cichlid fishes. Evolution (N. Y). 2017, 71, 1297–1312. [Google Scholar] [CrossRef]
- Hersberger-Zurfluh, M.A.; Papageorgiou, S.N.; Motro, M.; Kantarci, A.; Will, L.A.; Eliades, T. Facial soft tissue growth in identical twins. Am. J. Orthod. Dentofac. Orthop. 2018, 154, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Higashiyama, H.; Kuroda, S.; Iwase, A.; Irie, N.; Kurihara, H. On the Maxillofacial Development of Mice, Mus musculus. J. Morphol. 2025, 286, e70032. [Google Scholar] [CrossRef] [PubMed]
- Hînganu, M.V.; Cucu, R.P.; Hînganu, D. Personalized Research on the Aging Face—A Narrative History; J. Pers. Med., 2024; Vol. 14, Page 343 14. [Google Scholar] [CrossRef]
- Holleville, N.; Quilhac, A.; Bontoux, M.; Monsoro-Burq, A.-H.; élèn. BMP signals regulate Dlx5 during early avian skull development. Dev. Biol. 2003, 257, 177–189. [Google Scholar] [CrossRef]
- Hoskens, H.; Li, J.; Indencleef, K.; Gors, D.; Larmuseau, M.H.D.; Richmond, S.; Zhurov, A.I.; Hens, G.; Peeters, H.; Claes, P. Spatially Dense 3D Facial Heritability and Modules of Co-heritability in a Father-Offspring Design. Front. Genet. 2018, 9, 554. [Google Scholar] [CrossRef] [PubMed]
- Hosono, K.; Kawase, K.; Kurata, K.; Niimi, Y.; Saitsu, H.; Minoshima, S.; Ohnishi, H.; Yamamoto, Takahiro; Hikoya, A.; Tachibana, N.; Fukao, T.; Yamamoto, Tetsuya; Hotta, Y. A case of childhood glaucoma with a combined partial monosomy 6p25 and partial trisomy 18p11 due to an unbalanced translocation. Ophthalmic Genet 2020, 41, 175–182. [Google Scholar] [CrossRef]
- Hu, Y.; Albertson, R.C. Hedgehog signaling mediates adaptive variation in a dynamic functional system in the cichlid feeding apparatus. Proc. Natl. Acad. Sci. 2014, 1323154111. [Google Scholar] [CrossRef]
- Hufnagel, R.B.; Zimmerman, S.L.; Krueger, L.A.; Bender, P.L.; Ahmed, Z.M.; Saal, H.M. A new frontonasal dysplasia syndrome associated with deletion of the SIX2 gene. Am. J. Med. Genet. Part A 2016, 170, 487–491. [Google Scholar] [CrossRef]
- Hulsey, C.D.; Machado-Schiaffino, G.; Keicher, L.; Ellis-Soto, D.; Henning, F.; Meyer, A. The Integrated Genomic Architecture and Evolution of Dental Divergence in East African Cichlid Fishes (Haplochromis chilotes x H. nyererei). G3 GenesGenomesGenetics 2017, 7, 3195–3202. [Google Scholar] [CrossRef]
- Ibarra, B.A.; Atit, R. What Do Animal Models Teach Us About Congenital Craniofacial Defects? Adv. Exp. Med. Biol. 2020, 1236, 137–155. [Google Scholar] [CrossRef] [PubMed]
- Jelenkovic, A.; Poveda, A.; Susanne, C.; Rebato, E. Common genetic and environmental factors among craniofacial traits in Belgian nuclear families: Comparing skeletal and soft-tissue related phenotypes. HOMO 2010, 61, 191–203. [Google Scholar] [CrossRef]
- Kautt, A.F.; Kratochwil, C.F.; Nater, A.; Machado-Schiaffino, G.; Olave, M.; Henning, F.; Torres-Dowdall, J.; Härer, A.; Hulsey, C.D.; Franchini, P.; Pippel, M.; Myers, E.W.; Meyer, A. Contrasting signatures of genomic divergence during sympatric speciation. Nat 2020, 2020 5887836 588, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Kosaki, K.; Saito, H.; Kosaki, R.; Torii, C.; Kishi, K.; Takahashi, T. Branchial arch defects and 19p13.12 microdeletion: Defining the critical region into a 0.8 M base interval. Am. J. Med. Genet. Part A 2011, 155, 2212–2214. [Google Scholar] [CrossRef] [PubMed]
- Krishnaswamy, V.R.; Balaguru, U.M.; Chatterjee, S.; Korrapati, P.S. Dermatopontin augments angiogenesis and modulates the expression of transforming growth factor beta 1 and integrin alpha 3 beta 1 in endothelial cells. Eur. J. Cell Biol. 2017, 96, 266–275. [Google Scholar] [CrossRef]
- Krishnaswamy, V.R.; Korrapati, P.S. Role of Dermatopontin in re-epithelialization: Implications on keratinocyte migration and proliferation. Sci. Rep. 2015, 4, 7385. [Google Scholar] [CrossRef]
- Kucharczyk, M.; Kochański, A.; Jezela-Stanek, A.; Kugaudo, M.; Sielska-Rotblum, D.; Gutkowska, A.; Krajewska-Walasek, M. The first case of a patient with de novo partial distal 16q tetrasomy and a data’s review. Am. J. Med. Genet. Part A 2014, 164, 2541–2550. [Google Scholar] [CrossRef]
- Labalette, C.; Nouët, Y.; Sobczak-Thepot, J.; Armengol, C.; Levillayer, F.; Gendron, M.C.; Renard, C.A.; Regnault, B.; Chen, J.; Buendia, M.A.; Wei, Y. The LIM-only protein FHL2 regulates cyclin D1 expression and cell proliferation. J. Biol. Chem. 2008, 283, 15201–15208. [Google Scholar] [CrossRef]
- Landi, F.; Barraclough, J.; Evteev, A.; Anikin, A.; Satanin, L.; O’Higgins, P. The role of the nasal region in craniofacial growth: An investigation using path analysis. Anat. Rec. 2022, 305, 1892–1909. [Google Scholar] [CrossRef]
- Le Tanno, P.; Poreau, B.; Devillard, F.; Vieville, G.; Amblard, F.; Jouk, P.-S.; Satre, V.; Coutton, C. Maternal complex chromosomal rearrangement leads to TCF12 microdeletion in a patient presenting with coronal craniosynostosis and intellectual disability. Am. J. Med. Genet. Part A 2014, 164, 1530–1536. [Google Scholar] [CrossRef]
- Lecaudey, L.A.; Singh, P.; Sturmbauer, C.; Duenser, A.; Gessl, W.; Ahi, E.P. Transcriptomics unravels molecular players shaping dorsal lip hypertrophy in the vacuum cleaner cichlid, Gnathochromis permaxillaris. BMC Genomics 2021, 22, 506. [Google Scholar] [CrossRef]
- Lecaudey, L.A.; Sturmbauer, C.; Singh, P.; Ahi, E.P. Molecular mechanisms underlying nuchal hump formation in dolphin cichlid, Cyrtocara moorii. Sci. Rep. 2019. [Google Scholar] [CrossRef]
- Liem, K.F. Evolutionary strategies and morphological innovations: cichlid pharyngeal jaws. Syst. Zool. 1973, 22, 425–441. [Google Scholar] [CrossRef]
- Lim, M.H.; Jeun, J.H.; Kim, D.H.; Park, S.H.; Kim, S.J.; Lee, W.S.; Hwang, S.H.; Lim, J.Y.; Kim, S.W. Evaluation of Collagen Gel-Associated Human Nasal Septum-Derived Chondrocytes As a Clinically Applicable Injectable Therapeutic Agent for Cartilage Repair. Tissue Eng. Regen. Med. 2020, 17, 387–399. [Google Scholar] [CrossRef] [PubMed]
- Linnenkamp, B.D.W.; Raskin, S.; Esposito, S.E.; Herai, R.H. A comprehensive analysis of AHRR gene as a candidate for cleft lip with or without cleft palate. Mutat. Res.-Rev. Mutat. Res. 2020. [Google Scholar] [CrossRef] [PubMed]
- Little, A.C.; Jones, B.C.; Debruine, L.M. Facial attractiveness: evolutionary based research. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 1638–1659. [Google Scholar] [CrossRef]
- Liu, X.; Meng, L.; Shi, Q.; Liu, S.; Cui, C.; Hu, S.; Wei, Y. Dermatopontin promotes adhesion, spreading and migration of cardiac fibroblasts in vitro. Matrix Biol 2013, 32, 23–31. [Google Scholar] [CrossRef]
- Machado-Schiaffino, G.; Kautt, A.F.; Torres-Dowdall, J.; Baumgarten, L.; Henning, F.; Meyer, A. Incipient speciation driven by hypertrophied lips in Midas cichlid fishes? Mol. Ecol. 2017, 26, 2348–2362. [Google Scholar] [CrossRef]
- Machii, N.; Hatashima, R.; Niwa, T.; Taguchi, H.; Kimirei, I.A.; Mrosso, H.D.; Aibara, M.; Nagasawa, T.; Nikaido, M. Pronounced expression of extracellular matrix proteoglycans regulated by Wnt pathway underlies the parallel evolution of lip hypertrophy in East African cichlids. Elife 2025, 13. [Google Scholar] [CrossRef]
- MacRae, V.E.; Horvat, S.; Pells, S.C.; Dale, H.; Collinson, R.S.; Pitsillides, A.A.; Ahmed, S.F.; Farquharson, C. Increased bone mass, altered trabecular architecture and modified growth plate organization in the growing skeleton of SOCS2 deficient mice. J. Cell. Physiol. 2009, 218, 276–284. [Google Scholar] [CrossRef]
- Manousaki, T.; Hull, P.M.; Kusche, H.; Machado-Schiaffino, G.; Franchini, P.; Harrod, C.; Elmer, K.R.; Meyer, A. Parsing parallel evolution: ecological divergence and differential gene expression in the adaptive radiations of thick-lipped Midas cichlid fishes from Nicaragua. Mol. Ecol. 2013, 22, 650–69. [Google Scholar] [CrossRef]
- Marconi, A.; Vernaz, G.; Karunaratna, A.; Ngochera, M.J.; Durbin, R.; Santos, M.E. Genetic and Developmental Divergence in the Neural Crest Program between Cichlid Fish Species. Mol. Biol. Evol. 2024, 41. [Google Scholar] [CrossRef]
- Masonick, P.; Meyer, A.; Hulsey, C.D. A Kiss of Deep Homology: Partial Convergence in the Genomic Basis of Hypertrophied Lips in Cichlid Fish and Human Cleft Lip. Genome Biol. Evol. 2023, 15. [Google Scholar] [CrossRef]
- Matthews, H.S.; Penington, A.J.; Hardiman, R.; Fan, Y.; Clement, J.G.; Kilpatrick, N.M.; Claes, P.D. Modelling 3D craniofacial growth trajectories for population comparison and classification illustrated using sex-differences. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef]
- McCarthy, N.; Liu, J.S.; Richarte, A.M.; Eskiocak, B.; Lovely, C. Ben; Tallquist, M.D.; Eberhart, J.K. Pdgfra and Pdgfrb genetically interact during craniofacial development. Dev. Dyn. 2016, 245, 641–652. [Google Scholar] [CrossRef] [PubMed]
- Meerschaut, I.; Rochefort, D.; Revençu, N.; Pètre, J.; Corsello, C.; Rouleau, G.A.; Hamdan, F.F.; Michaud, J.L.; Morton, J.; Radley, J.; Ragge, N.; García-Miñaúr, S.; Lapunzina, P.; Bralo, M.P.; Mori, M.A.; Moortgat, S.; Benoit, V.; Mary, S.; Bockaert, N.; Oostra, A.; Vanakker, O.; Velinov, M.; De Ravel, T.J.L.; Mekahli, D.; Sebat, J.; Vaux, K.K.; DiDonato, N.; Hanson-Kahn, A.K.; Hudgins, L.; Dallapiccola, B.; Novelli, A.; Tarani, L.; Andrieux, J.; Parker, M.J.; Neas, K.; Ceulemans, B.; Schoonjans, A.S.; Prchalova, D.; Havlovicova, M.; Hancarova, M.; Budisteanu, M.; Dheedene, A.; Menten, B.; Dion, P.A.; Lederer, D.; Callewaert, B. FOXP1-related intellectual disability syndrome: A recognisable entity. J. Med. Genet. 2017, 54, 613–623. [Google Scholar] [CrossRef] [PubMed]
- Metcalf, D.; Greenhalgh, C.J.; Viney, E.; Willson, T.A.; Starr, R.; Nicola, N.A.; Hilton, D.J.; Alexander, W.S. Gigantism in mice lacking suppressor of cytokine signalling-2. Nature 2000, 405, 1069–1073. [Google Scholar] [CrossRef] [PubMed]
- Miller, N.D.; Nance, M.A.; Wohler, E.S.; Hoover-Fong, J.E.; Lisi, E.; Thomas, G.H.; Pevsner, J. Molecular (SNP) analyses of overlapping hemizygous deletions of 10q25.3 to 10qter in four patients: Evidence for HMX2 and HMX3 as candidate genes in hearing and vestibular function. Am. J. Med. Genet. Part A 2009, 149A, 669–680. [Google Scholar] [CrossRef]
- Mirzamohammadi, F.; Kozlova, A.; Papaioannou, G.; Paltrinieri, E.; Ayturk, U.M.; Kobayashi, T. Distinct molecular pathways mediate Mycn and Myc-regulated miR-17-92 microRNA action in Feingold syndrome mouse models. Nat. Commun. 2018, 9, 1352. [Google Scholar] [CrossRef]
- Mitteldorf, C.; Llamas-Velasco, M.; Schulze, H.-J.; Thoms, K.-M.; Mentzel, T.; Tronnier, M.; Kutzner, H. Deceptively bland cutaneous angiosarcoma on the nose mimicking hemangioma-A clinicopathologic and immunohistochemical analysis. J. Cutan. Pathol. 2018, 45, 652–658. [Google Scholar] [CrossRef]
- Mohamad Shah, N.S.; Sulong, S.; Wan Sulaiman, W.A.; Halim, A.S. Two novel genes TOX3 and COL21A1 in large extended Malay families with nonsyndromic cleft lip and/or palate. Mol. Genet. Genomic Med. 2019, 7, e635. [Google Scholar] [CrossRef]
- Nanda, R.S.; Meng, H.; Kapila, S.; Goorhuis, J. Growth changes in the soft tissue facial profile. Angle Orthod 1990, 60, 177–190. [Google Scholar] [CrossRef]
- Naqvi, S.; Hoskens, H.; Wilke, F.; Weinberg, S.M.; Shaffer, J.R.; Walsh, S.; Shriver, M.D.; Wysocka, J.; Claes, P. Decoding the Human Face: Progress and Challenges in Understanding the Genetics of Craniofacial Morphology. Annu. Rev. Genomics Hum. Genet. 2022, 23, 383–412. [Google Scholar] [CrossRef]
- Navon, D.; Male, I.; Tetrault, E.R.; Aaronson, B.; Karlstrom, R.O.; Craig Albertson, R. Hedgehog signaling is necessary and sufficient to mediate craniofacial plasticity in teleosts. Proc. Natl. Acad. Sci. U. S. A. 2020, 117, 19321–19327. [Google Scholar] [CrossRef]
- Ng, C.F.; Ng, P.K.S.; Lui, V.W.Y.; Li, J.; Chan, J.Y.W.; Fung, K.P.; Ng, Y.K.; Lai, P.B.S.; Tsui, S.K.W. FHL2 exhibits anti-proliferative and anti-apoptotic activities in liver cancer cells. Cancer Lett 2011, 304, 97–106. [Google Scholar] [CrossRef]
- Okello, D.O.; Iyyanar, P.P.R.; Kulyk, W.M.; Smith, T.M.; Lozanoff, S.; Ji, S.; Nazarali, A.J. Six2 Plays an Intrinsic Role in Regulating Proliferation of Mesenchymal Cells in the Developing Palate. Front. Physiol. 2017, 8, 955. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.; Dong, N.; Zheng, Z. Small leucine-rich proteoglycans in skin wound healing. Front. Pharmacol. 2020, 10, 501915. [Google Scholar] [CrossRef]
- Parnell, N.F.; Hulsey, C.D.; Streelman, J.T. THE GENETIC BASIS OF A COMPLEX FUNCTIONAL SYSTEM. Evolution (N. Y). 2012, 66, 3352–3366. [Google Scholar] [CrossRef] [PubMed]
- Parsons, K.J.; Trent Taylor, A.; Powder, K.E.; Albertson, R.C. Wnt signalling underlies the evolution of new phenotypes and craniofacial variability in Lake Malawi cichlids. Nat. Commun. 2014, 5, 3629. [Google Scholar] [CrossRef] [PubMed]
- Peixoto da-Silva, J.; Lourenço, S.; Nico, M.; Silva, F.H.; Martins, M.T.; Costa-Neves, A. Expression of laminin-5 and integrins in actinic cheilitis and superficially invasive squamous cell carcinomas of the lip. Pathol. Res. Pract. 2012, 208, 598–603. [Google Scholar] [CrossRef] [PubMed]
- Peñaherrera, M.S.; Weindler, S.; Van Allen, M.I.; Yong, S.-L.; Metzger, D.L.; McGillivray, B.; Boerkoel, C.; Langlois, S.; Robinson, W.P. Methylation profiling in individuals with Russell-Silver syndrome. Am. J. Med. Genet. Part A 2010, 152A, 347–355. [Google Scholar] [CrossRef]
- Piard, J.; Rozé, V.; Czorny, A.; Lenoir, M.; Valduga, M.; Fenwick, A.L.; Wilkie, A.O.M.; Van Maldergem, L. TCF12 microdeletion in a 72-year-old woman with intellectual disability. Am. J. Med. Genet. Part A 2015, 167, 1897–1901. [Google Scholar] [CrossRef]
- Pinchefsky, E.; Laneuville, L.; Srour, M. Distal 22q11.2 Microduplication. Child Neurol. Open 2017, 4, 2329048X1773765. [Google Scholar] [CrossRef] [PubMed]
- Powder, K.E.; Albertson, R.C. Cichlid fishes as a model to understand normal and clinical craniofacial variation. Dev. Biol. 2016, 415, 338–346. [Google Scholar] [CrossRef] [PubMed]
- Powder, K.E.; Cousin, H.; McLinden, G.P.; Craig Albertson, R. A nonsynonymous mutation in the transcriptional regulator lbh is associated with cichlid craniofacial adaptation and neural crest cell development. Mol. Biol. Evol. 2014, 31, 3113–24. [Google Scholar] [CrossRef] [PubMed]
- Powder, K.E.; Milch, K.; Asselin, G.; Albertson, R.C. Constraint and diversification of developmental trajectories in cichlid facial morphologies. Evodevo 2015, 6, 25. [Google Scholar] [CrossRef]
- Qian, W.; Zhang, M.; Wan, K.; Xie, Y.; Du, S.; Li, J.; Mu, X.; Qiu, J.; Xue, X.; Zhuang, X.; Wu, Y.; Liu, F.; Wang, S. Genetic evidence for facial variation being a composite phenotype of cranial variation and facial soft tissue thickness. J. Genet. Genomics 2022, 49, 934–942. [Google Scholar] [CrossRef]
- Quigley, D.I.; Kaiser-Rogers, K.; Aylsworth, A.S.; Rao, K.W. Submicroscopic deletion 9(q34.4) and duplication 19(p13.3): Identified by subtelomere specific FISH probes. Am. J. Med. Genet. 2004, 125A, 67–72. [Google Scholar] [CrossRef]
- Quinto-Sánchez, M.; Muñoz-Muñoz, F.; Gomez-Valdes, J.; Cintas, C.; Navarro, P.; De Cerqueira, C.C.S.; Paschetta, C.; De Azevedo, S.; Ramallo, V.; Acuña-Alonzo, V.; Adhikari, K.; Fuentes-Guajardo, M.; Hünemeier, T.; Everardo, P.; De Avila, F.; Jaramillo, C.; Arias, W.; Gallo, C.; Poletti, G.; Bedoya, G.; Bortolini, M.C.; Canizales-Quinteros, S.; Rothhammer, F.; Rosique, J.; Ruiz-Linares, A.; Gonzalez-Jose, R. Developmental pathways inferred from modularity, morphological integration and fluctuating asymmetry patterns in the human face. Sci. Reports 2018, 2018 81 8, 1–15. [Google Scholar] [CrossRef]
- Reijnders, M.R.F.; Ansor, N.M.; Kousi, M.; Yue, W.W.; Tan, P.L.; Clarkson, K.; Clayton-Smith, J.; Corning, K.; Jones, J.R.; Lam, W.W.K.; Mancini, G.M.S.; Marcelis, C.; Mohammed, S.; Pfundt, R.; Roifman, M.; Cohn, R.; Chitayat, D.; Millard, T.H.; Katsanis, N.; Brunner, H.G.; Banka, S. RAC1 Missense Mutations in Developmental Disorders with Diverse Phenotypes. Am. J. Hum. Genet. 2017, 101, 466–477. [Google Scholar] [CrossRef]
- Richmond, S.; Howe, L.J.; Lewis, S.; Stergiakouli, E.; Zhurov, A. Facial Genetics: A Brief Overview. Front. Genet. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Roberts, R.B.; Hu, Y.; Albertson, R.C.; Kocher, T.D. Craniofacial divergence and ongoing adaptation via the hedgehog pathway. Proc. Natl. Acad. Sci. U. S. A. 2011, 108, 13194–9. [Google Scholar] [CrossRef] [PubMed]
- Rometsch, S.J.; Torres-Dowdall, J.; Machado-Schiaffino, G.; Karagic, N.; Meyer, A. Dual function and associated costs of a highly exaggerated trait in a cichlid fish. Ecol. Evol. 2021, 11, 17496–17508. [Google Scholar] [CrossRef] [PubMed]
- Ruff, M.; Leyme, A.; Le Cann, F.; Bonnier, D.; Le Seyec, J.; Chesnel, F.; Fattet, L.; Rimokh, R.; Baffet, G.; Théret, N. The Disintegrin and Metalloprotease ADAM12 Is Associated with TGF-β-Induced Epithelial to Mesenchymal Transition. PLoS One 2015, 10, e0139179. [Google Scholar] [CrossRef]
- Samson, N.; Fink, B.; Matts, P.J. Visible skin condition and perception of human facial appearance. Int. J. Cosmet. Sci. 2010, 32, 167–184. [Google Scholar] [CrossRef]
- Sandell, R.F.; Carter, J.M.; Folpe, A.L. Solitary (juvenile) xanthogranuloma: a comprehensive immunohistochemical study emphasizing recently developed markers of histiocytic lineage. Hum. Pathol. 2015, 46, 1390–1397. [Google Scholar] [CrossRef]
- Santos, M.E.; Braasch, I.; Boileau, N.; Meyer, B.S.; Sauteur, L.; Böhne, A.; Belting, H.G.; Affolter, M.; Salzburger, W. The evolution of cichlid fish egg-spots is linked with a cis-regulatory change. Nat. Commun. 2014, 2014 51 5, 1–11. [Google Scholar] [CrossRef]
- Santos, M.E.; Lopes, J.F.; Kratochwil, C.F. East African cichlid fishes. EvoDevo 2023, 14, 1–21. [Google Scholar] [CrossRef]
- Saultz, J.N.; Kaffenberger, B.H.; Taylor, M.; Heerema, N.A.; Klisovic, R. Novel Chromosome 5 Inversion Associated With PDGFRB Rearrangement in Hypereosinophilic Syndrome. JAMA Dermatology 2016, 152, 1391. [Google Scholar] [CrossRef]
- Schiavinato, A.; Marcous, F.; Zuk, A. V.; Keene, D.R.; Tufa, S.F.; Mosquera, L.M.; Zigrino, P.; Mauch, C.; Eckes, B.; Francois, K.; De Backer, J.; Hunzelmann, N.; Moinzadeh, P.; Krieg, T.; Callewaert, B.; Sengle, G. New insights into the structural role of EMILINs within the human skin microenvironment. Sci. Reports 2024, 2024 141 14, 1–15. [Google Scholar] [CrossRef]
- Sharma, V.P.; Fenwick, A.L.; Brockop, M.S.; McGowan, S.J.; Goos, J.A.C.; Hoogeboom, A.J.M.; Brady, A.F.; Jeelani, N.O.; Lynch, S.A.; Mulliken, J.B.; Murray, D.J.; Phipps, J.M.; Sweeney, E.; Tomkins, S.E.; Wilson, L.C.; Bennett, S.; Cornall, R.J.; Broxholme, J.; Kanapin, A.; Consortium; 500 Whole-Genome Sequences (WGS500); Johnson, D.; Wall, S.A.; Spek; van der, P.J.; Mathijssen, I.M.J.; Maxson, R.E.; Twigg, S.R.F.; Wilkie, A.O.M. Mutations in TCF12, encoding a basic helix-loop-helix partner of TWIST1, are a frequent cause of coronal craniosynostosis. Nat. Genet. 2013, 453 45, 304. [Google Scholar] [CrossRef] [PubMed]
- Shaw-Smith, C. Genetic factors in esophageal atresia, tracheo-esophageal fistula and the VACTERL association: Roles for FOXF1 and the 16q24.1 FOX transcription factor gene cluster, and review of the literature. Eur. J. Med. Genet. 2010. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Ahi, E.P.; Sturmbauer, C. Gene coexpression networks reveal molecular interactions underlying cichlid jaw modularity. BMC Ecol. Evol. 2021, 21, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Tschanz-Lischer, H.; Ford, K.; Ahi, E.P.; Haesler, M.P.; Mwaiko, S.; Meier, J.I.; Marques, D.; Bruggman, R.; Kishe, M.; Seehausen, O. Highly modular genomic architecture underlies combinatorial mechanism of speciation and adaptive radiation. bioRxiv 2025, 2025.07.07.663194. [Google Scholar] [CrossRef]
- Som, P.M.; Naidich, T.P. Illustrated review of the embryology and development of the facial region, part 1: Early face and lateral nasal cavities. AJNR. Am. J. Neuroradiol. 2013, 34, 2233–2240. [Google Scholar] [CrossRef]
- Song, J.; Chae, H.S.; Shin, J.W.; Sung, J.; Song, Y.M.; Baek, S.H.; Kim, Y.H. Influence of heritability on craniofacial soft tissue characteristics of monozygotic twins, dizygotic twins, and their siblings using Falconer’s method and principal components analysis. Korean J. Orthod. 2018, 49, 3–11. [Google Scholar] [CrossRef]
- Stuppia, L.; Capogreco, M.; Marzo, G.; La Rovere, D.; Antonucci, I.; Gatta, V.; Palka, G.; Mortellaro, C.; Tetè, S. Genetics of Syndromic and Nonsyndromic Cleft Lip and Palate. J. Craniofac. Surg. 2011, 22, 1722–1726. [Google Scholar] [CrossRef]
- Sugimoto, Y.; Takimoto, A.; Akiyama, H.; Kist, R.; Scherer, G.; Nakamura, T.; Hiraki, Y.; Shukunami, C. Scx+/Sox9+ progenitors contribute to the establishment of the junction between cartilage and tendon/ligament. Development 2013, 140, 2280–2288. [Google Scholar] [CrossRef]
- Szabo-Rogers, H.L.; Geetha-Loganathan, P.; Nimmagadda, S.; Fu, K.K.; Richman, J.M. FGF signals from the nasal pit are necessary for normal facial morphogenesis. Dev. Biol. 2008, 318, 289–302. [Google Scholar] [CrossRef]
- Takenouchi, T.; Yamaguchi, Y.; Tanikawa, A.; Kosaki, R.; Okano, H.; Kosaki, K. Novel Overgrowth Syndrome Phenotype Due to Recurrent De Novo PDGFRB Mutation. J. Pediatr. 2015, 166, 483–486. [Google Scholar] [CrossRef] [PubMed]
- Talbot, J.C.; Johnson, S.L.; Kimmel, C.B. hand2 and Dlx genes specify dorsal, intermediate and ventral domains within zebrafish pharyngeal arches. Development 2010, 137, 2507–17. [Google Scholar] [CrossRef] [PubMed]
- Tan, T.Y.; Gonzaga-Jauregui, C.; Bhoj, E.J.; Strauss, K.A.; Brigatti, K.; Puffenberger, E.; Li, D.; Xie, L.Q.; Das, N.; Skubas, I.; Deckelbaum, R.A.; Hughes, V.; Brydges, S.; Hatsell, S.; Siao, C.J.; Dominguez, M.G.; Economides, A.; Overton, J.D.; Mayne, V.; Simm, P.J.; Jones, B.O.; Eggers, S.; Le Guyader, G.; Pelluard, F.; Haack, T.B.; Sturm, M.; Riess, A.; Waldmueller, S.; Hofbeck, M.; Steindl, K.; Joset, P.; Rauch, A.; Hakonarson, H.; Baker, N.L.; Farlie, P.G. Monoallelic BMP2 Variants Predicted to Result in Haploinsufficiency Cause Craniofacial, Skeletal, and Cardiac Features Overlapping Those of 20p12 Deletions. Am. J. Hum. Genet. 2017, 101, 985–994. [Google Scholar] [CrossRef]
- Thomas, P.S.; Kim, J.; Nunez, S.; Glogauer, M.; Kaartinen, V. Neural crest cell-specific deletion of Rac1 results in defective cell-matrix interactions and severe craniofacial and cardiovascular malformations. Dev. Biol. 2010, 340, 613–625. [Google Scholar] [CrossRef]
- Tokita, M.J.; Chow, P.M.; Mirzaa, G.; Dikow, N.; Maas, B.; Isidor, B.; Le Caignec, C.; Penney, L.S.; Mazzotta, G.; Bernardini, L.; Filippi, T.; Battaglia, A.; Donti, E.; Earl, D.; Prontera, P. Five children with deletions of 1p34.3 encompassing AGO1 and AGO3. Eur. J. Hum. Genet. 2015, 23, 761–765. [Google Scholar] [CrossRef]
- Torres-Dowdall, J.; Meyer, A. Sympatric and Allopatric Diversification in the Adaptive Radiations of Midas Cichlids in Nicaraguan Lakes. In The Behavior, Ecology and Evolution of Cichlid Fishes; Springer; Dordrecht, 2021; pp. 175–216. [Google Scholar] [CrossRef]
- Tsagkrasoulis, D.; Hysi, P.; Spector, T.; Montana, G. Heritability maps of human face morphology through large-scale automated three-dimensional phenotyping. Sci. Reports 2017, 2017 71 7, 1–18. [Google Scholar] [CrossRef]
- Van Mater, D.; Knelson, E.H.; Kaiser-Rogers, K.A.; Armstrong, M.B. Neuroblastoma in a pediatric patient with a microduplication of 2p involving the MYCN locus. Am. J. Med. Genet. Part A 2013, 161, 605–610. [Google Scholar] [CrossRef] [PubMed]
- Vera-Carbonell, A.; Moya-Quiles, M.R.; Ballesta-Martínez, M.; López-González, V.; Bafallíu, J.A.; Guillén-Navarro, E.; López-Expósito, I. Rapp–Hodgkin syndrome and SHFM1 patients: Delineating the p63–Dlx5/Dlx6 pathway. Gene 2012, 497, 292–297. [Google Scholar] [CrossRef]
- Vos, J.A.; Abbondanzo, S.L.; Barekman, C.L.; Andriko, J.W.; Miettinen, M.; Aguilera, N.S. Histiocytic sarcoma: a study of five cases including the histiocyte marker CD163. Mod. Pathol. 2005, 18, 693–704. [Google Scholar] [CrossRef]
- Weber, M.; Wehrhan, F.; Deschner, J.; Sander, J.; Ries, J.; Möst, T.; Bozec, A.; Gölz, L.; Kesting, M.; Lutz, R. The Special Developmental Biology of Craniofacial Tissues Enables the Understanding of Oral and Maxillofacial Physiology and Diseases; Int. J. Mol. Sci., 2021; Vol. 22, Page 1315 22. [Google Scholar] [CrossRef]
- Xie, M.; Kaiser, M.; Gershtein, Y.; Schnyder, D.; Deviatiiarov, R.; Gazizova, G.; Shagimardanova, E.; Zikmund, T.; Kerckhofs, G.; Ivashkin, E.; Batkovskyte, D.; Newton, P.T.; Andersson, O.; Fried, K.; Gusev, O.; Zeberg, H.; Kaiser, J.; Adameyko, I.; Chagin, A.S. The level of protein in the maternal murine diet modulates the facial appearance of the offspring via mTORC1 signaling. Nat. Commun. 2024, 15, 1–15. [Google Scholar] [CrossRef]
- Xu, J.; Liu, H.; Lan, Y.; Aronow, B.J.; Kalinichenko, V. V.; Jiang, R. A Shh-Foxf-Fgf18-Shh Molecular Circuit Regulating Palate Development. PLOS Genet 2016, 12, e1005769. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Hu, L.; Liu, X.; Zhan, G.; Mei, M.; Wang, H.; Zhang, X.; Qiu, Z.; Zhou, W.; Yang, L. A Novel MYCN Variant Associated with Intellectual Disability Regulates Neuronal Development. Neurosci. Bull. 2018, 34, 854–858. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, S.; Liu, D.; Zhao, Z.; Song, H.; Peng, K. Identification and validation of GIMAP family genes as immune-related prognostic biomarkers in lung adenocarcinoma. Heliyon 2024, 10, 33111. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Examples of cichlid fish models for studying the heritable morphologies of human face. (A) Heritability map of human face. Red and blue areas indicate high and low levels of heritability, respectively (www.heritabilitymaps.info). (B) African cichlids as models to study facial areas with high of levels of heritability in human (e.g. forehead, nose and upper lip). (C) Examples of predicted co-expression of facial morphogenic genes in human soft tissues (www.gtexportal.org) with potentially conserved role in cichlids.
Figure 1.
Examples of cichlid fish models for studying the heritable morphologies of human face. (A) Heritability map of human face. Red and blue areas indicate high and low levels of heritability, respectively (www.heritabilitymaps.info). (B) African cichlids as models to study facial areas with high of levels of heritability in human (e.g. forehead, nose and upper lip). (C) Examples of predicted co-expression of facial morphogenic genes in human soft tissues (www.gtexportal.org) with potentially conserved role in cichlids.
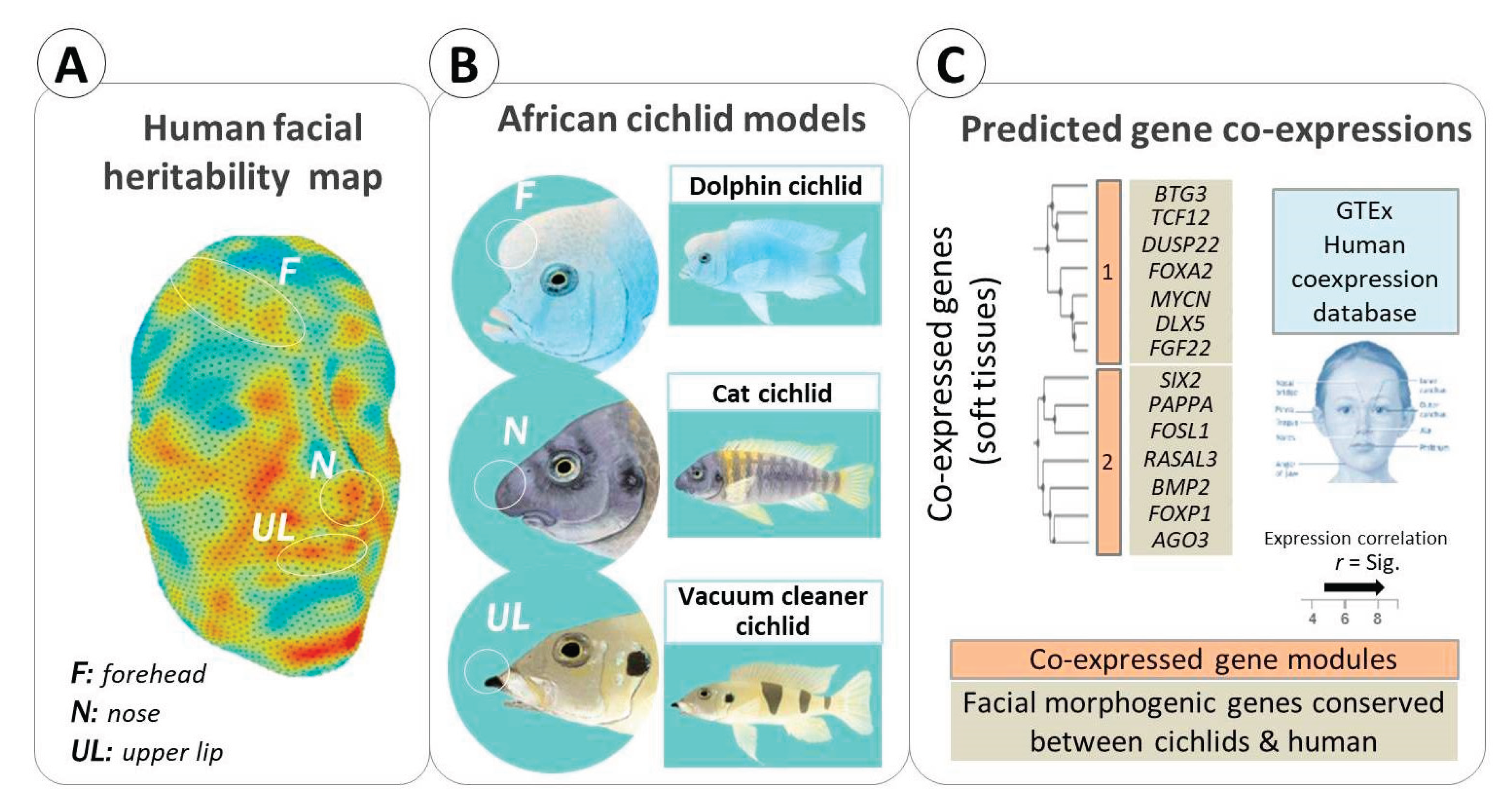
Figure 2.
Interactome maps of conserved genes involved in facial soft tissue morphogenesis in cichlid fish models and humans. Regulatory interactions among candidate genes identified as drivers of morphological changes in the forehead, nasal regions, and upper lip of selected cichlid models (with potentially similar roles in humans) are shown using the STRING database. The interactions are based on available knowledge of protein–protein associations, transcriptional regulatory connection, and biochemical interactions.
Figure 2.
Interactome maps of conserved genes involved in facial soft tissue morphogenesis in cichlid fish models and humans. Regulatory interactions among candidate genes identified as drivers of morphological changes in the forehead, nasal regions, and upper lip of selected cichlid models (with potentially similar roles in humans) are shown using the STRING database. The interactions are based on available knowledge of protein–protein associations, transcriptional regulatory connection, and biochemical interactions.
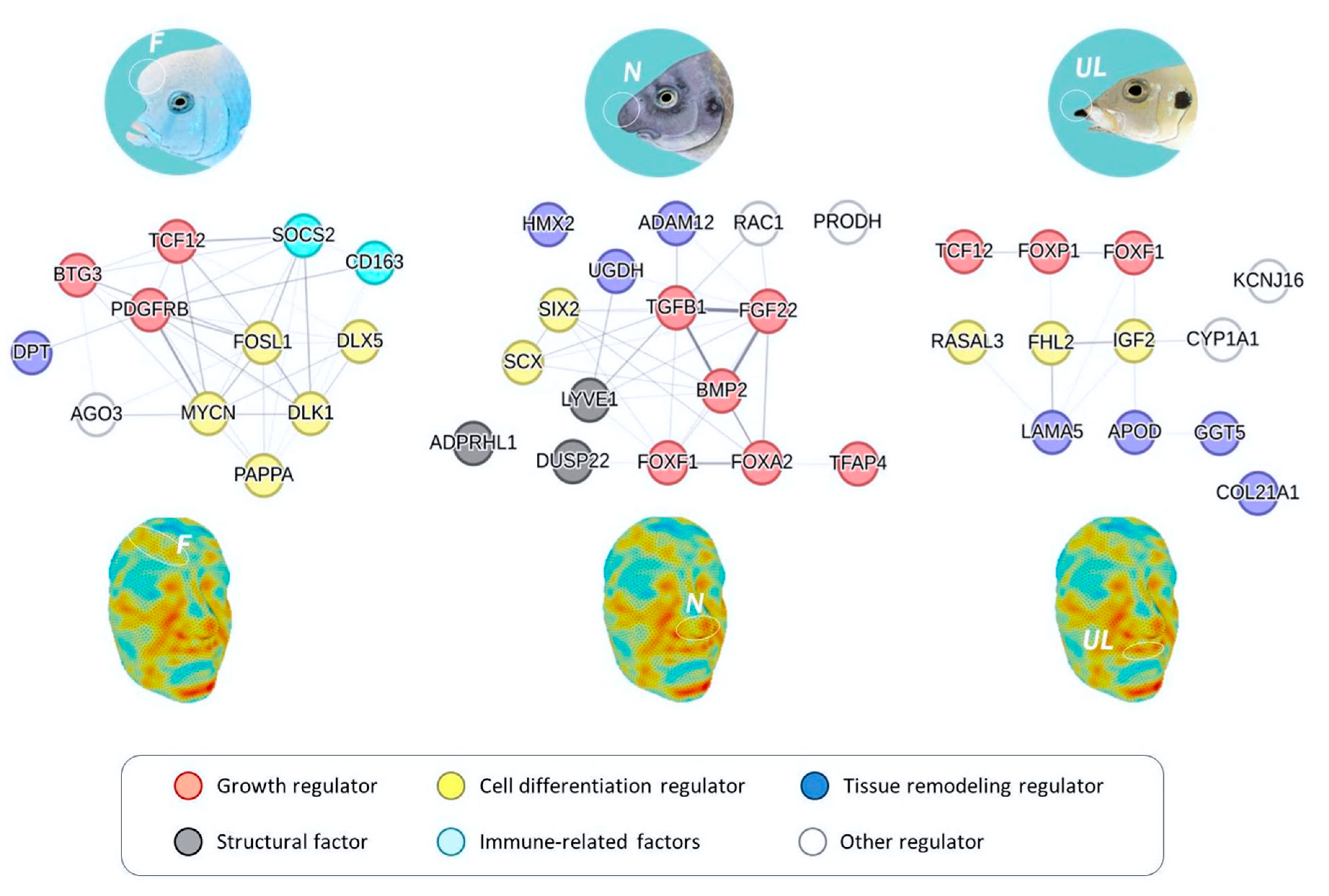
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



