
Submitted:
29 July 2025
Posted:
30 July 2025
You are already at the latest version
Abstract
The human microbiome is increasingly recognized as a critical modulator of host physiology, offering a dynamic and targetable axis in the landscape of precision medicine. This review critically examines the mechanistic pathways through which the microbiome influences immunity, metabolism, pharmacological response, and gene regulation, with emphasis on disease-specific interactions spanning oncology, immunology, metabolic disorders, and neuropsychiatry. Multi-omics technologies, including metagenomics, metatranscriptomics, metaproteomics, and metabolomics, are explored as integrative tools that generate a systems-level understanding of host–microbe interactions. Computational frameworks for data integration and interpretation are discussed, alongside clinical applications in microbiome-informed diagnostics, patient stratification, and therapeutic interventions such as fecal microbiota transplantation and engineered probiotics. Persistent challenges, ranging from inter-individual variability and data standardization to ethical and regulatory complexities, are evaluated with reference to emerging solutions, including synthetic biology, personalized microbial interventions, and decision-support algorithms. The review underscores the need for collaborative, mechanistically anchored, and longitudinal approaches to fully translate microbiome science into actionable precision health strategies.
Keywords:
microbiome-guided precision medicine
; multi-omics integration
; host–microbe interactions
; microbiome-based diagnostics
; personalized therapeutics
Introduction
The human microbiome, comprising trillions of bacteria, viruses, archaea, and fungi, has emerged as a powerful modulator of health and disease. Once considered a passive passenger, it is now understood to actively shape host immunity, metabolism, neurological function, and drug response [1,2]. This shift in understanding coincides with the rise of precision medicine, which aims to tailor healthcare based on an individual's genetic, environmental, and lifestyle factors. Yet, one of the most dynamic and modifiable of these factors, the microbiome, has yet to be fully integrated into precision medicine frameworks.
Microbiome variation has been linked to a wide range of conditions, including inflammatory bowel disease [3], type 2 diabetes [4], cancer [5], and neuropsychiatric disorders [1]. In many of these contexts, microbial biomarkers show potential not just for disease detection but also for predicting outcomes and guiding therapy. Interventions such as dietary modulation, prebiotics, probiotics, and fecal microbiota transplantation (FMT) are being explored to shift disease trajectories by reshaping the microbiome [6,7]. Still, despite an explosion of microbiome association studies, their translation into robust clinical tools remains limited. Most efforts remain descriptive or correlative, lacking mechanistic depth or reproducibility.
The expanding landscape of multi-omics, metagenomics, metabolomics, metatranscriptomics, and host-microbiome interaction studies, offers unprecedented opportunities to resolve these gaps. However, integrative frameworks that link microbiome data with host phenotypes in a clinically meaningful way are still emerging and often fragmented [8]. This review aims to consolidate what we know, what remains unclear, and what is needed to move microbiome science into the heart of precision medicine. While disease-specific or technology-focused reviews exist, few provide a cross-cutting synthesis of mechanistic insights, multi-omic strategies, and translational potential across disease domains. As the field matures, this broader lens is urgently needed. We argue that microbiome-informed precision medicine is not a speculative goal but an achievable frontier, one that demands deeper integration of mechanistic understanding with clinically actionable outcomes.
Mechanistic Foundations of Microbiome-Driven Precision Medicine
The integration of the microbiome into precision medicine requires a mechanistic understanding of how microbial communities influence host physiology. These mechanisms operate at multiple levels, ranging from molecular interactions and immune modulation to drug metabolism and gene regulation, and are context-dependent across tissues and disease states. Rather than being passive indicators, microbes are increasingly recognized as active participants in shaping host phenotypes, with the potential to mediate health trajectories.
One of the most well-characterized domains of host–microbiome interaction is the immune system. Commensal microbes are critical for immune maturation, tolerance, and homeostasis, influencing everything from gut barrier integrity to the balance between pro-inflammatory and regulatory responses [9]. For instance, segmented filamentous bacteria promote Th17 cell differentiation, while Bacteroides fragilis secretes polysaccharide A to induce regulatory T cells [10]. Dysbiosis can disrupt this balance, predisposing individuals to autoimmune diseases and inflammatory disorders, a foundational mechanism now being explored for immunotherapy stratification.
The microbiome also exerts profound metabolic influence. Microbial fermentation of dietary fiber produces short-chain fatty acids (SCFAs) such as acetate, propionate, and butyrate, which regulate epithelial integrity, energy metabolism, and even appetite signaling via G-protein-coupled receptors [11]. Specific microbial species can synthesize or degrade metabolites that affect systemic lipid and glucose homeostasis, offering mechanistic links between microbiome composition and metabolic diseases like type 2 diabetes and obesity.
Beyond intrinsic host processes, the microbiome can modulate drug efficacy and toxicity, a domain now termed pharmacomicrobiomics. Microbial enzymes can activate, inactivate, or toxify drugs; a classic example is the reactivation of the chemotherapeutic irinotecan by microbial β-glucuronidases, leading to severe gastrointestinal toxicity [12]. Similarly, microbial composition has been shown to influence the efficacy of immune checkpoint inhibitors in cancer therapy, suggesting that baseline microbiome profiling could predict therapeutic response [13]. These findings have sparked interest in microbiome-aware drug design and adjunct therapies that target microbial pathways.
The specificity of dysbiosis across disease contexts also points to distinct mechanistic pathways. In colorectal cancer, for instance, certain Fusobacterium nucleatum strains promote tumorigenesis by activating β-catenin signaling and recruiting immunosuppressive cells [14]. In contrast, in neurological disorders such as Parkinson’s disease, microbial dysbiosis may influence disease onset and progression through altered bile acid metabolism, systemic inflammation, and gut-brain signaling [15]. These disease-specific mechanisms underscore the need for tailored microbiome profiling and interventions based on underlying pathophysiology.
Finally, emerging evidence highlights a triangular crosstalk between the host genome, microbiome, and environmental exposures. Host genetic variants influence microbial colonization patterns, such as the link between LCT genotype and Bifidobacterium abundance [16], while microbial metabolites can, in turn, regulate host gene expression via epigenetic modification [17]. This bidirectional interaction suggests that microbiome-informed precision medicine must move beyond profiling microbes in isolation, toward integrated models that consider host-microbe co-evolution and systems-level dynamics. A modular framework outlining how the microbiome modulates host systems and informs precision medicine strategies is shown in Figure 3.
Figure 1.
Mechanistic pathways linking the human microbiome to host physiology and precision medicine outcomes. This schematic depicts how the human microbiome influences host systems through specific mechanistic pathways to support precision healthcare. a. The human microbiome, comprising diverse microbes, forms the central hub of interaction. b. It acts through molecular mechanisms including metabolite production, immune modulation, pathogen exclusion, and gut barrier regulation. c. These actions affect key host systems such as the gut, endocrine organs, brain, and immune network. d. Insights from these interactions enable precision medicine applications like personalized therapeutics, drug response prediction, patient stratification, and targeted interventions.
Figure 1.
Mechanistic pathways linking the human microbiome to host physiology and precision medicine outcomes. This schematic depicts how the human microbiome influences host systems through specific mechanistic pathways to support precision healthcare. a. The human microbiome, comprising diverse microbes, forms the central hub of interaction. b. It acts through molecular mechanisms including metabolite production, immune modulation, pathogen exclusion, and gut barrier regulation. c. These actions affect key host systems such as the gut, endocrine organs, brain, and immune network. d. Insights from these interactions enable precision medicine applications like personalized therapeutics, drug response prediction, patient stratification, and targeted interventions.
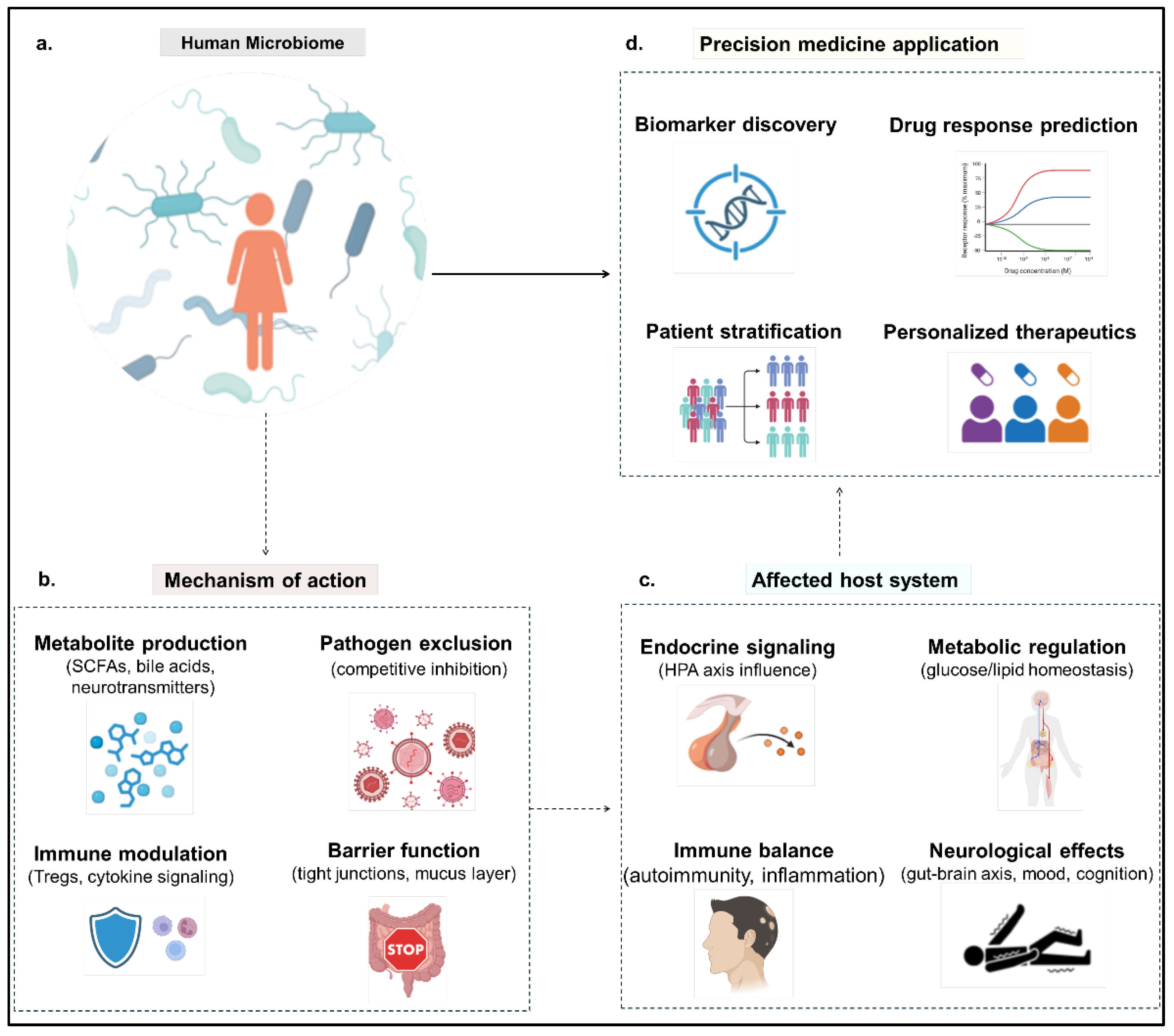
In sum, the microbiome contributes to host physiology through multifaceted and interdependent mechanisms. A clear understanding of these interactions, anchored in immune regulation, metabolic function, pharmacological response, and gene-environment interplay, is essential for translating microbiome science into clinical precision. These mechanisms also provide potential intervention points, where therapies can be tailored not only to a patient's genome but to their microbiome-informed phenotype.
Multi-Omics Approaches to Microbiome Profiling
Traditional microbiome studies relying solely on 16S rRNA gene surveys or metagenomics have illuminated broad community structures and taxonomic associations with disease. However, to decipher the functional roles of microbial communities and identify clinically actionable biomarkers, multi-omics approaches are increasingly being adopted. These strategies aim to capture different layers of microbial activity, from gene content to expression, proteins, and metabolites, providing a systems-level view of host-microbiome interactions.
Metagenomics and Beyond
Shotgun metagenomics remains foundational, offering species- and strain-level resolution of microbial composition along with insights into genetic potential. Yet, DNA-based profiling provides only a static snapshot of potential functionality. To understand which genes are actively expressed, metatranscriptomics , the sequencing of microbial mRNA, is essential. This layer has uncovered context-specific microbial responses in disease states, such as microbial stress gene upregulation in inflammatory bowel disease [18] or carbohydrate metabolism shifts in type 2 diabetes [3].
Metaproteomics complements transcriptomics by measuring actual protein abundance, capturing post-transcriptional regulation and microbial-host protein interactions [19]. Though still technically challenging due to sample complexity and dynamic range, it offers invaluable insight into microbial functional phenotypes, particularly in mucosal and inflammatory niches.
Metabolomics, profiling small molecules produced by microbiota or the host, often provides the most direct readout of microbe-host crosstalk. For instance, altered levels of short-chain fatty acids (SCFAs), bile acids, and tryptophan metabolites have been implicated in diseases ranging from colorectal cancer to depression [20,21]. These molecules often function as signaling mediators and are more proximate to disease-relevant physiological changes, making them attractive as biomarkers or therapeutic targets.
To consolidate these complementary omics strategies, Table 1 provides a comparative overview of metagenomics, metatranscriptomics, metaproteomics, and metabolomics, including their major outputs, strengths, limitations, and translational relevance.
Integration Strategies: Temporal and Spatial Resolution
Despite the richness of individual omics layers, their integration poses conceptual and analytical challenges. Longitudinal multi-omics sampling is crucial for understanding microbial dynamics, such as responses to antibiotics, diet, or immune perturbations. For example, multi-timepoint metagenomic and metabolomic profiling has revealed personalized but stable microbiome-metabolome trajectories in healthy individuals, forming the basis for precision nutrition algorithms [22,23].
Emerging single-cell technologies offer yet another dimension, enabling deconvolution of microbial heterogeneity that is masked in bulk analyses. Single-cell genomics and transcriptomics, though still in early stages for microbiome research, have begun to uncover niche-specific functional roles within polymicrobial communities [24].
An integrated multi-omics pipeline (Figure 2) facilitates the transition from raw microbiome data to clinical decisions by harmonizing metagenomic, transcriptomic, proteomic, and metabolomic datasets into actionable outputs.
Computational Frameworks and Machine Learning
Given the high dimensionality and sparsity of multi-omics microbiome data, integrative computational tools are indispensable. Methods such as similarity network fusion [25], multi-omics factor analysis (MOFA+) [26], and Bayesian latent variable models have been deployed to identify cross-omic patterns and infer causal relationships.
Machine learning (ML) approaches, particularly random forests, support vector machines, and neural networks, have been applied to predict clinical phenotypes from integrated omics features. Recent developments in interpretable ML, such as SHAP (SHapley Additive exPlanations), have improved the transparency of these models, allowing identification of specific microbial taxa or metabolites driving predictions [27].
Case Studies and Translational Examples
Integrated multi-omics has already demonstrated translational value. In a landmark study on inflammatory bowel disease, Lloyd-Price et al. (2019) combined metagenomic, transcriptomic, metabolomic, and proteomic data to stratify patients by immune state and microbial function, revealing mechanistic subtypes beyond clinical diagnosis [2].
In cancer immunotherapy, multi-omics analyses have identified not only taxonomic biomarkers (e.g., Akkermansia muciniphila) but also associated bile acid signatures that modulate T cell activity and therapy response [28]. Similarly, microbiome-informed predictions of glycemic response have outperformed standard clinical predictors, as seen in the personalized nutrition studies [23,29].
Clinical Applications and Current Landscape
The shift from association-based microbiome studies to translational applications has begun to take shape, particularly in the domains of diagnostics, therapeutics, and patient stratification. Despite challenges in reproducibility and standardization, the clinical potential of microbiome-guided precision medicine is increasingly being realized through carefully designed trials and early-stage interventions.
Microbiome-Based Diagnostics and Prognostics
Numerous studies have shown that microbial profiles can act as reliable biomarkers for disease detection and risk assessment. For instance, microbial signatures have been linked with early-stage colorectal cancer, allowing for non-invasive detection approaches with performance comparable to or exceeding fecal occult blood tests [30]. Similarly, dysbiosis scores based on the presence or absence of key microbial taxa have been proposed for IBD, metabolic disorders, and even psychiatric conditions such as depression [21]. These biomarkers, especially when integrated with host genomic or metabolomic data, offer enhanced predictive accuracy, yet remain constrained by variability across cohorts and platforms.
Fecal Microbiota Transplantation (FMT) and Engineered Probiotics
FMT is among the most clinically mature microbiome-based interventions, having shown high efficacy in treating recurrent Clostridioides difficile infection [31]. Beyond this indication, trials are ongoing to assess FMT's efficacy in ulcerative colitis, metabolic syndrome, and even neurodevelopmental disorders such as autism [32]. However, clinical outcomes remain heterogeneous, in part due to donor variability, undefined microbial compositions, and patient-specific host-microbiome interactions.
This has led to a growing interest in rationally designed microbial therapeutics. Next-generation probiotics, engineered strains or defined microbial consortia, are being developed to target metabolic pathways, immune modulation, or colonization resistance [33]. For instance, Bacteroides thetaiotaomicron engineered to produce immunomodulatory molecules has shown preclinical promise in ameliorating inflammation in IBD models [34]. These approaches offer more predictable outcomes than FMT but face challenges in regulatory approval, scalability, and ecological stability within diverse host environments (see Table 2 for a comparative summary of current and emerging microbiome-based therapeutic strategies).
Role in Immunotherapy Response
The interplay between the gut microbiome and cancer immunotherapy has emerged as a critical frontier. Studies have demonstrated that the presence of specific bacterial taxa, such as Akkermansia muciniphila and Bifidobacterium longum, correlates with enhanced response to immune checkpoint inhibitors in melanoma and non-small cell lung cancer [13,40]. Mechanistically, these bacteria may enhance dendritic cell maturation and T-cell activation via microbial metabolites and pattern recognition receptor signaling.
FMT from responders into germ-free mice or non-responding patients has recapitulated enhanced immunotherapeutic efficacy, suggesting a causal role for the microbiome [41]. Despite these promising results, reproducibility across studies is limited, and contextual host factors likely play a key role in determining outcome. Nonetheless, modulation of the microbiome, via diet, probiotics, or even FMT, is increasingly being explored as a co-therapy to improve immunotherapy efficacy.
Stratification of Patients Using Microbial Biomarkers
Personalized medicine requires stratifying patients into subgroups with differential risk or treatment response. The microbiome offers a novel axis of stratification, as demonstrated in studies where microbial diversity or taxonomic composition predicted relapse in IBD [42] or treatment efficacy in colorectal cancer [43]. Integrating microbiome data with host transcriptomic or metabolomic profiles enables more granular subtyping, potentially guiding therapeutic decisions.
For example, in metabolic syndrome, patients with low microbial gene richness are less likely to benefit from high-fiber dietary interventions [44]. Similarly, stratifying patients based on enterotypes has been proposed for tailoring dietary interventions and probiotic formulations. However, standardizing microbial biomarkers for clinical use remains an unsolved challenge, especially given population-specific microbiome signatures and inter-individual variability.
Challenges and Limitations
Despite a robust body of research linking the microbiome to various disease states, the field continues to grapple with significant translational hurdles. A major challenge lies in the high interindividual variability of microbiome composition, which complicates the development of universal biomarkers or therapies. Even within disease cohorts, microbial signatures often vary across geographic regions, ethnicities, diets, and environmental exposures, limiting reproducibility and generalizability [45,46]. While efforts like the Human Microbiome Project (HMP) and MetaHIT have provided foundational data, they remain disproportionately focused on Western populations [47].
Another obstacle is the context-dependent nature of host–microbe interactions. The same bacterial species can exert protective or pathogenic effects depending on host genetics, immune status, or metabolic context [48]. This nuance undermines simple causative models and raises the bar for designing interventions with predictable outcomes.
Moreover, a lack of methodological standardization, in sample collection, sequencing platforms, data processing pipelines, and analytical frameworks, further hampers clinical utility. Comparative analyses across studies often suffer from batch effects or inconsistent definitions of “dysbiosis” [49]. The absence of standardized reference materials or diagnostic thresholds makes it difficult to translate microbiome data into regulatory-grade diagnostics or treatment decision tools. International initiatives for microbiome biobanking and metadata harmonization, such as MBQC and MIxS, are emerging but remain underutilized [50]. A global consensus on reference datasets, quality controls, and longitudinal repositories is urgently needed.
Ethical, legal, and social implications (ELSI) also pose critical challenges, particularly in light of microbiome data’s potential for identifiability, population-level variation, and incidental findings [51]. Issues such as informed consent, data sharing across borders, and benefit-sharing from commercial microbiome products raise unresolved concerns. Additionally, the patentability of microbial strains and the ownership of microbiota-derived insights continue to stir debate [52].
Finally, many studies are underpowered, cross-sectional, and correlative in nature. Longitudinal, mechanistic investigations, particularly those incorporating multi-omics layers (e.g., metatranscriptomics, metabolomics), remain scarce but are essential for delineating causal relationships and actionable targets [53]. Without these, clinical translation remains speculative, even when associations appear robust. Furthermore, integrative computational frameworks that can robustly link microbiome features with host phenotypes are still in development. Issues of overfitting, confounding, and false discovery remain prevalent.
Taken together, the translational bottleneck is not due to a lack of associations but to the complexity of disentangling microbiome–host–environment interactions and converting them into reproducible, clinically meaningful interventions. Addressing these foundational issues is critical for realizing the full potential of microbiome-informed precision medicine.
Future Directions and Research Gaps
As microbiome science transitions from correlation to causation, the path ahead demands strategic integration of emerging technologies, personalized interventions, and robust regulatory support.
Personalized Pre/Probiotics and Diet–Microbiome Interfaces
A major frontier lies in customized microbiome modulation, especially through precision-designed prebiotics, probiotics, and synbiotics tailored to individual microbial and metabolic profiles [54,55]. Current interventions are largely empirical, with mixed outcomes due to host-specific microbiome variability [56]. Advances in metagenomic profiling and functional inference are enabling the design of next-generation probiotics with targeted mechanisms, including competitive exclusion of pathogens, SCFA production, and immune modulation.
Simultaneously, nutritional microbiomics aims to decode how specific dietary components interact with the gut microbiota to influence host physiology, offering personalized dietary recommendations to modulate health outcomes [56]. Despite promise, translating these interventions requires long-term, randomized clinical trials that stratify participants by baseline microbiome signatures and track longitudinal responses. Moreover, diet–microbiome relationships are often nonlinear and context-dependent, demanding sophisticated computational models that integrate host genetics, lifestyle factors, and microbial function [54].
Synthetic Biology Tools for Precision Microbiome Editing
The application of synthetic biology offers exciting tools for targeted manipulation of the microbiome. Engineered microbes can be designed to sense disease states, secrete therapeutic molecules, or compete with pathogenic strains [33,57]. Technologies such as CRISPR-based antimicrobials and designer bacteriophages provide organism-specific targeting without disrupting the broader microbial ecology [58].
However, delivery, stability, and containment remain critical challenges. Ensuring that engineered microbes function predictably in diverse gut environments while avoiding unintended ecological consequences requires refined safety switches and containment systems [33]. Regulatory frameworks must also evolve to accommodate these live biotherapeutics under appropriate risk-benefit assessments [57].
Integrating Microbiome Data into EHRS and Clinical Decision Support
As microbial diagnostics inch closer to clinical reality, a key research gap lies in the integration of microbiome data into Electronic Health Records (EHRs) and clinical decision support systems (CDSS) [59]. Currently, microbial data exists largely in silos, disconnected from other patient data streams. Integration would allow clinicians to interpret microbiome results in the context of comorbidities, medication use, lifestyle, and genomics [60].
To achieve this, interoperability standards, user-friendly visualization tools, and interpretation frameworks must be developed. Moreover, AI-driven models capable of synthesizing multi-modal data (e.g., metagenomics, transcriptomics, lab tests) into actionable outputs will be critical to support decision-making in real time [60]. But issues around data standardization, quality control, and clinical validation remain unresolved [61].
Regulatory Frameworks and Commercialization Prospects
The current regulatory landscape for microbiome-based products is fragmented. While probiotics and dietary supplements are often regulated as foods, microbiome-derived therapeutics and diagnostics fall under drug or medical device categories depending on jurisdiction [62,63]. A coherent international regulatory framework is urgently needed to facilitate innovation while ensuring patient safety.
Furthermore, commercialization pathways remain unclear for many microbiome-based products. Questions around IP protection, biomarker qualification, and reimbursement models must be addressed [64]. Public-private partnerships, like those seen in cancer immunotherapy, could accelerate development and implementation by aligning academic discovery with industrial scaling and regulatory insight [63]. A strategic comparison of existing challenges versus envisioned solutions across infrastructure, clinical integration, and governance is summarized in Table 3, highlighting key leverage points for transitioning to microbiome-guided precision medicine.
Conclusions
Microbiome-informed precision medicine stands at the frontier of biomedical innovation, poised to redefine how we diagnose, treat, and prevent disease. Across a spectrum of conditions, from inflammatory bowel disease and metabolic disorders to cancer immunotherapy responsiveness, the human microbiome is no longer viewed as a passive background entity, but as an active, modulatable interface between host biology and environmental exposures [65,66]. The mounting evidence linking microbial signatures with clinical phenotypes, treatment outcomes, and disease trajectories signals a paradigm shift from one-size-fits-all medicine toward more nuanced, individualized strategies [13,67].
However, realizing the full potential of microbiome-guided precision health demands more than just descriptive profiling. It requires systemic integration of multi-omics data, AI-enabled analytics, and context-aware clinical frameworks [66,68]. The complexity and plasticity of the microbiome, influenced by host genetics, diet, lifestyle, geography, and drug regimens, highlight the limitations of simplistic correlative models. Rather, mechanistic elucidation, bridging functional metagenomics with causal inference and host-microbe interactions, will be key to identifying actionable microbial targets [69,70]. Moreover, precision interventions such as personalized probiotics, microbiota-directed foods, and engineered microbial consortia must be tailored not just to taxa, but to the metabolic and immunological landscapes of individual patients [71,72].
Our analysis underscores the urgent need for longitudinal cohort studies, harmonized data repositories, and standardized clinical endpoints to strengthen translational pipelines [45,50]. Short-term, geographically isolated studies, while insightful, often fail to capture the temporal and inter-individual variability necessary for clinical generalization. Integrating microbiome data into Electronic Health Records (EHRs) and real-time decision-support tools would allow dynamic tracking and optimization of interventions, but such integration remains in its infancy and fraught with technical and regulatory hurdles [73].
Equally pressing are the ethical, legal, and societal challenges. Issues around identifiability, consent, data ownership, and equitable benefit-sharing are not peripheral, they are central to the responsible clinical adoption of microbiome-based tools [51,74]. Establishing global governance models that are both robust and inclusive will be essential for building public trust and ensuring that the benefits of this technology are shared broadly.
Ultimately, unlocking the promise of microbiome-informed precision health will require a concerted, interdisciplinary effort. Collaboration among microbiologists, clinicians, computational scientists, ethicists, and policymakers is not optional but foundational. Mechanistic understanding must be married with systems-level modeling; innovation must be guided by reproducibility and equity [75,76].
We are at a pivotal juncture. With careful design, ethical foresight, and translational ambition, microbiome science can move from fragmented discovery to a cohesive, clinically actionable discipline. It is time to shift the lens from "what microbes are present" to "what microbes are doing", and more importantly, how we can harness them to promote health, resilience, and individualized care at scale.
Author Declarations
The author declares no conflict of interest and received no external funding for this work. No new data were created or analysed in this study. The author is solely responsible for the conceptualization, literature review, writing, and editing of the manuscript.
References
- C. Jf et al., “The Microbiota-Gut-Brain Axis,” Physiological reviews, vol. 99, no. 4, Oct. 2019. [CrossRef]
- J. Lloyd-Price et al., “Multi-omics of the gut microbial ecosystem in inflammatory bowel diseases,” Nature, vol. 569, no. 7758, pp. 655–662. May 2019. [CrossRef]
- E. A. Franzosa et al., “Gut microbiome structure and metabolic activity in inflammatory bowel disease,” Nat Microbiol, vol. 4, no. 2, pp. 293–305, Feb. 2019. [CrossRef]
- Q. J et al., “A metagenome-wide association study of gut microbiota in type 2 diabetes,” Nature, vol. 490, no. 7418, Oct. 2012. [CrossRef]
- G. Ws, “Cancer and the microbiota,” Science (New York, N.Y.), vol. 348, no. 6230, Apr. 2015. [CrossRef]
- L. Hj et al., “Therapeutic Modulation of Gut Microbiota in Functional Bowel Disorders,” Journal of neurogastroenterology and motility, vol. 23, no. 1, Jan. 2017. [CrossRef]
- Y. B. Mutalub et al., “Gut Microbiota Modulation as a Novel Therapeutic Strategy in Cardiometabolic Diseases,” Foods, vol. 11, no. 17, Art. no. 17, Jan. 2022. [CrossRef]
- Z. M et al., “Multiomics approach reveals the comprehensive interactions between nutrition and children’s gut microbiota, and microbial and host metabolomes,” Nutrition journal, vol. 24, no. 1, Mar. 2025. [CrossRef]
- B. Y and H. Oj, “Homeostatic Immunity and the Microbiota,” Immunity, vol. 46, no. 4, Apr. 2017. [CrossRef]
- R. Jl and M. Sk, “Inducible Foxp3+ regulatory T-cell development by a commensal bacterium of the intestinal microbiota,” Proceedings of the National Academy of Sciences of the United States of America, vol. 107, no. 27, Jul. 2010. [CrossRef]
- Koh, F. De Vadder, P. Kovatcheva-Datchary, and F. Bäckhed, “From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites,” Cell, vol. 165, no. 6, pp. 1332–1345, Jun. 2016. [CrossRef]
- W. Bd et al., “Alleviating cancer drug toxicity by inhibiting a bacterial enzyme,” Science (New York, N.Y.), vol. 330, no. 6005, Nov. 2010. [CrossRef]
- V. Gopalakrishnan et al., “Gut microbiome modulates response to anti–PD-1 immunotherapy in melanoma patients,” Science, vol. 359, no. 6371, pp. 97–103, Jan. 2018. [CrossRef]
- K. Ad et al., “Fusobacterium nucleatum potentiates intestinal tumorigenesis and modulates the tumor-immune microenvironment,” Cell host & microbe, vol. 14, no. 2, Aug. 2013. [CrossRef]
- S. Tr et al., “Gut Microbiota Regulate Motor Deficits and Neuroinflammation in a Model of Parkinson’s Disease,” Cell, vol. 167, no. 6, Dec. 2016. [CrossRef]
- G. Jk et al., “Human genetics shape the gut microbiome,” Cell, vol. 159, no. 4, Nov. 2014. [CrossRef]
- K. Ka, F. K. Ka, F. J, and B. F, “Gut microbial metabolites as multi-kingdom intermediates,” Nature reviews. Microbiology, vol. 19, no. 2, Feb. 2021. [CrossRef]
- S. M et al., “Dynamics of metatranscription in the inflammatory bowel disease gut microbiome,” Nature microbiology, vol. 3, no. 3, Mar. 2018. [CrossRef]
- Z. X et al., “The oral and gut microbiomes are perturbed in rheumatoid arthritis and partly normalized after treatment,” Nature medicine, vol. 21, no. 8, Aug. 2015. [CrossRef]
- A, P. J, and S. H, “Gut Microbiota Regulation of Tryptophan Metabolism in Health and Disease,” Cell host & microbe, vol. 23, no. 6, Jun. 2018. [CrossRef]
- M. Valles-Colomer et al., “The neuroactive potential of the human gut microbiota in quality of life and depression,” Nat Microbiol, vol. 4, no. 4, pp. 623–632, Apr. 2019. [CrossRef]
- J. Aj et al., “Daily Sampling Reveals Personalized Diet-Microbiome Associations in Humans,” Cell host & microbe, vol. 25, no. 6, Jun. 2019. [CrossRef]
- Z. D et al., “Personalized Nutrition by Prediction of Glycemic Responses,” Cell, vol. 163, no. 5, Nov. 2015. [CrossRef]
- X. Wu et al., “Single-cell sequencing to multi-omics: technologies and applications,” Biomarker Research, vol. 12, no. 1, p. 110, Sep. 2024. [CrossRef]
- B. Wang et al., “Similarity network fusion for aggregating data types on a genomic scale,” Nat Methods, vol. 11, no. 3, pp. 333–337, Mar. 2014. [CrossRef]
- A. R et al., “MOFA+: a statistical framework for comprehensive integration of multi-modal single-cell data,” Genome biology, vol. 21, no. 1. May 2020. [CrossRef]
- T. Bd, L. Na, R. Mt, W. J, and S. Pd, “A Framework for Effective Application of Machine Learning to Microbiome-Based Classification Problems,” mBio, vol. 11, no. 3, Jun. 2020. [CrossRef]
- M. Lf et al., “Microbiome-derived inosine modulates response to checkpoint inhibitor immunotherapy,” Science (New York, N.Y.), vol. 369, no. 6510, Sep. 2020. [CrossRef]
- M.-S. H et al., “Model of personalized postprandial glycemic response to food developed for an Israeli cohort predicts responses in Midwestern American individuals,” The American journal of clinical nutrition, vol. 110, no. 1, Jul. 2019. [CrossRef]
- T. Am et al., “Metagenomic analysis of colorectal cancer datasets identifies cross-cohort microbial diagnostic signatures and a link with choline degradation,” Nature medicine, vol. 25, no. 4, Apr. 2019. [CrossRef]
- van, N. E et al., “Duodenal infusion of donor feces for recurrent Clostridium difficile,” The New England journal of medicine, vol. 368, no. 5, Jan. 2013. [CrossRef]
- K. Dw et al., “Microbiota Transfer Therapy alters gut ecosystem and improves gastrointestinal and autism symptoms: an open-label study,” Microbiome, vol. 5, no. 1, Jan. 2017. [CrossRef]
- M. R. Charbonneau, V. M. Isabella, N. Li, and C. B. Kurtz, “Developing a new class of engineered live bacterial therapeutics to treat human diseases,” Nat Commun, vol. 11, no. 1, p. 1738, Apr. 2020. [CrossRef]
- H. Iy et al., “Engineered probiotic Escherichia coli can eliminate and prevent Pseudomonas aeruginosa gut infection in animal models,” Nature communications, vol. 8, Apr. 2017. [CrossRef]
- S. Reddi et al., “Fecal microbiota transplantation to prevent acute graft-versus-host disease: pre-planned interim analysis of donor effect,” Nat Commun, vol. 16, no. 1, p. 1034, Jan. 2025. [CrossRef]
- S. Khanna et al., “SER-109: An Oral Investigational Microbiome Therapeutic for Patients with Recurrent Clostridioides difficile Infection (rCDI),” Antibiotics, vol. 11, no. 9, Art. no. 9, Sep. 2022. [CrossRef]
- C. Dimopoulou et al., “Potential of using an engineered indole lactic acid producing Escherichia coli Nissle 1917 in a murine model of colitis,” Sci Rep, vol. 14, no. 1, p. 17542, Jul. 2024. [CrossRef]
- K. Mok et al., “Synbiotic-driven modulation of the gut microbiota and metabolic functions related to obesity: insights from a human gastrointestinal model,” BMC Microbiology, vol. 25, no. 1, p. 250, Apr. 2025. [CrossRef]
- R. M. Dedrick et al., “Phage Therapy of Mycobacterium Infections: Compassionate Use of Phages in 20 Patients With Drug-Resistant Mycobacterial Disease,” Clinical Infectious Diseases, vol. 76, no. 1, pp. 103–112, Jan. 2023. [CrossRef]
- R. B et al., “Gut microbiome influences efficacy of PD-1-based immunotherapy against epithelial tumors,” Science (New York, N.Y.), vol. 359, no. 6371, Jan. 2018. [CrossRef]
- B. En et al., “Fecal microbiota transplant promotes response in immunotherapy-refractory melanoma patients,” Science (New York, N.Y.), vol. 371, no. 6529, Feb. 2021. [CrossRef]
- Z. Y et al., “Gut Microbiota Offers Universal Biomarkers across Ethnicity in Inflammatory Bowel Disease Diagnosis and Infliximab Response Prediction,” mSystems, vol. 3, no. 1, Jan. 2018. [CrossRef]
- F. B et al., “The oral microbiota in colorectal cancer is distinctive and predictive,” Gut, vol. 67, no. 8, Aug. 2018. [CrossRef]
- E. Le Chatelier et al., “Richness of human gut microbiome correlates with metabolic markers,” Nature, vol. 500, no. 7464, pp. 541–546, Aug. 2013. [CrossRef]
- G. Falony et al., “Population-level analysis of gut microbiome variation,” Science, vol. 352, no. 6285, pp. 560–564, Apr. 2016. [CrossRef]
- G. Vk et al., “A predictive index for health status using species-level gut microbiome profiling,” Nature communications, vol. 11, no. 1, Sep. 2020. [CrossRef]
- J. Lloyd-Price et al., “Strains, functions and dynamics in the expanded Human Microbiome Project,” Nature, vol. 550, no. 7674, pp. 61–66, Oct. 2017. [CrossRef]
- Z. N, Z. D, K. T, S. E, and E. E, “Taking it Personally: Personalized Utilization of the Human Microbiome in Health and Disease,” Cell host & microbe, vol. 19, no. 1, Jan. 2016. [CrossRef]
- S. R, A. Cc, W. O, K. R, and H. C, “The microbiome quality control project: baseline study design and future directions,” Genome biology, vol. 16, Dec. 2015. [CrossRef]
- R. Knight et al., “Best practices for analysing microbiomes,” Nature Reviews Microbiology, vol. 16, no. 7, pp. 410–422, Jul. 2018. [CrossRef]
- E. Hs, P. M, and L. B, “Ethical challenges in conducting and the clinical application of human microbiome research,” Journal of medical ethics and history of medicine, vol. 16, Jul. 2023. [CrossRef]
- X. Sun, J. L. A. Fiala, and D. Lowery, “Modulating the human microbiome with live biotherapeutic products: intellectual property landscape,” Nature Reviews Drug Discovery, vol. 15, no. 4, pp. 224–225, Apr. 2016. [CrossRef]
- R. D et al., “Environment dominates over host genetics in shaping human gut microbiota,” Nature, vol. 555, no. 7695, Mar. 2018. [CrossRef]
- S. Kp, G. Sw, S. Po, F. Hj, and D. Sh, “The influence of diet on the gut microbiota,” Pharmacological research, vol. 69, no. 1, Mar. 2013. [CrossRef]
- D. M and van H. V. Je, “Fate, activity, and impact of ingested bacteria within the human gut microbiota,” Trends in microbiology, vol. 23, no. 6, Jun. 2015. [CrossRef]
- Z. N, S. J, and E. E, “You are what you eat: diet, health and the gut microbiota,” Nature reviews. Gastroenterology & hepatology, vol. 16, no. 1, Jan. 2019. [CrossRef]
- M. M, C. Rj, and L. Tk, “Microbiome therapeutics - Advances and challenges,” Advanced drug delivery reviews, vol. 105, no. Pt A, Oct. 2016. [CrossRef]
- R. C, C. Sp, C. V, Y. Sj, and W. Hh, “Metagenomic engineering of the mammalian gut microbiome in situ,” Nature methods, vol. 16, no. 2, Feb. 2019. [CrossRef]
- S.-F. R. Sm et al., “A longitudinal big data approach for precision health,” Nature medicine, vol. 25, no. 5. May 2019. [CrossRef]
- T. Ej, “A decade of digital medicine innovation,” Science translational medicine, vol. 11, no. 498, Jun. 2019. [CrossRef]
- B. Al, B. Y, and S. Ja, “The human skin microbiome,” Nature reviews. Microbiology, vol. 16, no. 3, Mar. 2018. [CrossRef]
- V. N. Ga, “Drugs and Devices: Comparison of European and U.S. Approval Processes,” JACC. Basic to translational science, vol. 1, no. 5, Aug. 2016. [CrossRef]
- P. C. Kashyap, N. Chia, H. Nelson, E. Segal, and E. Elinav, “Microbiome at the Frontier of Personalized Medicine,” Mayo Clinic proceedings, vol. 92, no. 12, p. 1855, Dec. 2017. [CrossRef]
- E.-J. Song and J.-H. Shin, “Personalized Diets based on the Gut Microbiome as a Target for Health Maintenance: from Current Evidence to Future Possibilities,” Journal of Microbiology and Biotechnology, vol. 32, no. 12, p. 1497, Oct. 2022. [CrossRef]
- J. A. Gilbert, M. J. Blaser, J. G. Caporaso, J. K. Jansson, S. V. Lynch, and R. Knight, “Current understanding of the human microbiome,” Nat Med, vol. 24, no. 4, pp. 392–400, Apr. 2018. [CrossRef]
- L. M. Proctor et al., “The Integrative Human Microbiome Project,” Nature, vol. 569, no. 7758, pp. 641–648. May 2019. [CrossRef]
- K. Zy and L. Sk, “The Human Gut Microbiome - A Potential Controller of Wellness and Disease,” Frontiers in microbiology, vol. 9, Aug. 2018. [CrossRef]
- H. D, “Microbiome Research Is Becoming the Key to Better Understanding Health and Nutrition,” Frontiers in genetics, vol. 9, Jun. 2018. [CrossRef]
- Z. A et al., “Population-based metagenomics analysis reveals markers for gut microbiome composition and diversity,” Science (New York, N.Y.), vol. 352, no. 6285, Apr. 2016. [CrossRef]
- M. Levy, A. A. Kolodziejczyk, C. A. Thaiss, and E. Elinav, “Dysbiosis and the immune system,” Nat Rev Immunol, vol. 17, no. 4, pp. 219–232, Apr. 2017. [CrossRef]
- Z. N et al., “Personalized Gut Mucosal Colonization Resistance to Empiric Probiotics Is Associated with Unique Host and Microbiome Features,” Cell, vol. 174, no. 6, Sep. 2018. [CrossRef]
- S. Sa, H. F, L. L, S. H, and de V. Wm, “Intestinal microbiome landscaping: insight in community assemblage and implications for microbial modulation strategies,” FEMS microbiology reviews, vol. 41, no. 2, Mar. 2017. [CrossRef]
- W. Md et al., “The FAIR Guiding Principles for scientific data management and stewardship,” Scientific data, vol. 3, Mar. 2016. [CrossRef]
- M. van de Guchte, H. M. Blottière, and J. Doré, “Humans as holobionts: implications for prevention and therapy,” Microbiome, vol. 6, no. 1, p. 81. May 2018. [CrossRef]
- K. Hou et al., “Microbiota in health and diseases,” Sig Transduct Target Ther, vol. 7, no. 1, p. 135, Apr. 2022. [CrossRef]
- D. A. Relman, “Thinking about the microbiome as a causal factor in human health and disease: philosophical and experimental considerations,” Curr Opin Microbiol, vol. 54, pp. 119–126, Apr. 2020. [CrossRef]
Figure 2.
Multi-omics integration pipeline for microbiome-guided precision medicine. This infographic outlines a structured pipeline for integrating multi-omics data to support microbiome-guided precision healthcare. a. The process begins with the collection of biological specimens, such as stool, saliva, or tissue biopsies, using standardized protocols to ensure sample integrity and comparability across individuals. b. From these samples, multi-layered data are generated: metagenomics to profile microbial communities and gene content; metatranscriptomics to capture actively expressed genes; metaproteomics to identify functional proteins; and metabolomics to quantify small-molecule metabolites influenced by host–microbiome interactions. c. These diverse omics layers are then integrated using advanced computational tools including machine learning, network analysis, and multivariate modeling to uncover functionally relevant and biologically meaningful relationships. d. The resulting insights are translated into actionable outcomes with real-world applications in precision nutrition, predictive diagnostics, and microbiome-informed therapeutic strategies.
Figure 2.
Multi-omics integration pipeline for microbiome-guided precision medicine. This infographic outlines a structured pipeline for integrating multi-omics data to support microbiome-guided precision healthcare. a. The process begins with the collection of biological specimens, such as stool, saliva, or tissue biopsies, using standardized protocols to ensure sample integrity and comparability across individuals. b. From these samples, multi-layered data are generated: metagenomics to profile microbial communities and gene content; metatranscriptomics to capture actively expressed genes; metaproteomics to identify functional proteins; and metabolomics to quantify small-molecule metabolites influenced by host–microbiome interactions. c. These diverse omics layers are then integrated using advanced computational tools including machine learning, network analysis, and multivariate modeling to uncover functionally relevant and biologically meaningful relationships. d. The resulting insights are translated into actionable outcomes with real-world applications in precision nutrition, predictive diagnostics, and microbiome-informed therapeutic strategies.


Table 1.
Characterizing key microbiome features through multi-omics approaches.
| Microbiome Feature | Relevance to Precision Medicine | Omics Approaches Used | Specific Insights Enabled |
|---|---|---|---|
| Taxonomic composition | Determines disease-linked dysbiosis; informs microbial biomarkers | Metagenomics (16S rRNA, WGS) | Detection of microbial signatures across diseases |
| Functional potential | Reveals biosynthetic capacities, AMR genes, virulence traits | Metagenomics, Metaproteomics | Prediction of functional shifts before phenotypic onset |
| Gene expression activity | Identifies active microbial pathways | Metatranscriptomics | Differentiation between latent and active microbial functions |
| Metabolite production | Directly influences host metabolism and immune signaling | Metabolomics (LC-MS, GC-MS, NMR) | Discovery of disease-linked metabolites (e.g., SCFAs, bile acids) |
| Microbe–host crosstalk | Underpins immunomodulation, gut-brain axis, inflammation | Host transcriptomics + microbial omics integration | Host responses to microbial fluctuations; immune-metabolic links |
| Temporal dynamics | Captures microbiome fluctuations linked to diet, therapy, disease | Longitudinal multi-omics + time-series modeling | Personalized monitoring; real-time tracking of interventions |
| Ecological interactions | Community stability, competition, and resilience | Systems biology, co-occurrence networks, integrated multi-omics | Network-level vulnerabilities and keystone species identification |
Table 2.
Microbiome-targeted interventions in clinical use and development.
| Therapy | Mechanism | Disease Context | Clinical Status | Key Limitations | Study |
|---|---|---|---|---|---|
| Fecal Microbiota Transplant (FMT) | Full microbiome ecosystem transfer | rCDI, UC, graft-vs-host |
Approved for rCDI, trials ongoing for others | Donor variability, infection risk | FDA-2022-176 [35] |
| Live biotherapeutic product (SER-109) | Enriched Firmicutes spores | rCDI | Phase III completed (Seres) | Targeted efficacy, not broad spectrum | [36] |
| Engineered E. coli nissle | SCFA biosynthesis, barrier repair | IBD, inflammation | Preclinical | Safety, horizontal gene transfer | [37] |
| Precision synbiotics | Selective prebiotic + probiotic strains | T2D, obesity | Phase I/II | Response variability, diet dependency | [38] |
| Phage therapy | Targeted depletion of pathogenic species | Crohn’s, MDR infections |
Experimental use | Resistance evolution, narrow targeting | [39] |
Table 3.
Current Gaps vs Ideal Future Frameworks in Microbiome-Informed Medicine.
| Category | Current State | Ideal Future State | Actions Needed |
|---|---|---|---|
| Data standardization | Non-uniform metadata, poor reproducibility | Harmonized pipelines, shared ontologies | Adoption of MIxS, MBQC; global repositories |
| Clinical integration | Minimal use in EHRs or decision support | Embedded microbiome metrics in diagnostics | Interoperable data standards, pilot deployments |
| Personalization of therapies | Broad-spectrum approaches | Microbiome-informed individualized treatment | Multi-omics modeling, n=1 trial design |
| Regulatory guidance | Patchy, product-specific approvals | Clear frameworks for diagnostics, probiotics, live biotherapeutics | International regulatory harmonization |
| Ethical & legal oversight | Limited, fragmented by jurisdiction | Global ELSI framework respecting identifiability & consent | Policy dialogue, equitable benefit-sharing models |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



