
Submitted:
22 July 2025
Posted:
23 July 2025
You are already at the latest version
Abstract
Bacteria must constantly adapt to changing environments, and iron has served as a critical cofactor in essential biochemical processes since life first evolved. Over the years, it has become apparent that iron homeostasis is intricately tangled with RNA metabolism. Diverse proteins and RNAs are sensitive to the levels and redox states of iron, and can trigger various signaling cascades in response to iron starvation, oxidative stress, and other challenges. While it is widely known that iron-binding proteins can serve as environmental sensors, those that also bind to RNA may have far-reaching impacts beyond iron homeostasis. Here, the impact of iron on the metabolism of RNA in bacteria is reviewed, ranging from single-atom cofactors for RNA folding to the widespread post-transcriptional regulatory effects of iron-binding proteins. In this review, the term degradation is used specifically to refer to complete breakdown of an RNA, while the terms processing and maturation refer to an RNA being converted to a shorter, stable, functional form through endonucleolytic and/or exonucleolytic cleavage. We use the term stability to refer to the degradation rate of an RNA. Finally, we consider RNA metabolism an umbrella term encompassing all aspects of RNA synthesis (transcription), post-transcriptional modifications, and processing and degradation by RNases.
Keywords:
iron-sulfer clusters
; Fe-S proteins
; RNA metabolism
; RNA degradation
; RNA processing
; mycobacteria
1. In the Beginning: The Prehistoric Chemical Landscape Shaped the Evolution of Biochemical Pathways
When life first originated about 4 billion years ago, approximately 9.5 billion years after the Big Bang and 1 billion years after the formation of the Earth, we were born into an anoxic environment where ferrous iron (Fe2+) was highly abundant [1,2,3]. As such, the Iron-Sulfur World hypothesis proposes that our microbial ancestors came to life in a reduced environment that was rich in both iron and sulfur, particularly around hydrothermal vents in the seabed [4]. While the exact composition of Earth’s surface and atmosphere over time remains a topic of intense study, the Iron-Sulfur World hypothesis suggests that the first simple metabolic reactions developed on the surface of iron-sulfide minerals. Eventually, these early “energy flows” gave rise to the genetic material we know today, beginning with RNA. The RNA World hypothesis suggests that, for about half a billion years after life evolved, early organisms used RNA-based chemistries for all metabolic and genetic processes [5] (Figure 1). It is unclear exactly who these early organisms were or how they functioned, since the earliest evidence for prokaryotes is dated around 3.7 billion years ago [6,7,8]. It is also unknown when or how exactly DNA emerged, either from an organism creating a “mutated” 2’-deoxygenated ribonucleotide, or spontaneously from the primordial soup.
Approximately 2.4 billion years ago, environmental O2 levels increased drastically in a phenomenon called the Great Oxidation Event (GOE) (Figure 1) [9]. The catalysts of the GOE are believed to be the cyanobacteria, who emerged either from freshwater environments or on land, and became the first organisms to produce O2 by way of photosynthesis [10,11,12]. Before the GOE, small amounts of oxygen reacted readily with the strong electron donors in the environment [13,14]. However, with the proliferation of oxygenic bacteria and geological changes such as the composition of volcanic gases, environmental oxygen sinks became saturated [15,16,17]. This rapid accumulation of atmospheric oxygen, which heretofore had largely only been found in water [14,18], fundamentally altered the landscape of the ancient Earth. Some of the elements and organic compounds that were previously ubiquitous in the environment, namely Fe2+, sulfur, hydrogen sulfide, and methane, were depleted. The GOE thus gave rise to the substitution of magnesium, manganese, and other metals largely replacing iron as cofactors for chemical reactions [19,20,21,22,23,24,25,26]. The increase in atmospheric O2 caused Fe2+ to become oxidized to its ferric (Fe3+) form on a never-before-seen scale, a shift that exposed early organisms to iron-mediated redox stress for which they had no defensive or adaptative strategies, likely devastating the primeval microbial population [27]. It is believed that some of the surviving Archaea underwent symbiogenesis with some members of the Proteobacteria (also called Pseudomonodota) phylum, leading to the evolution of the mitochondrion and, eventually, to multicellular life forms [28,29,30,31].
Figure 1.
Timeline approximating biological innovations and the oxidation of the Earth.
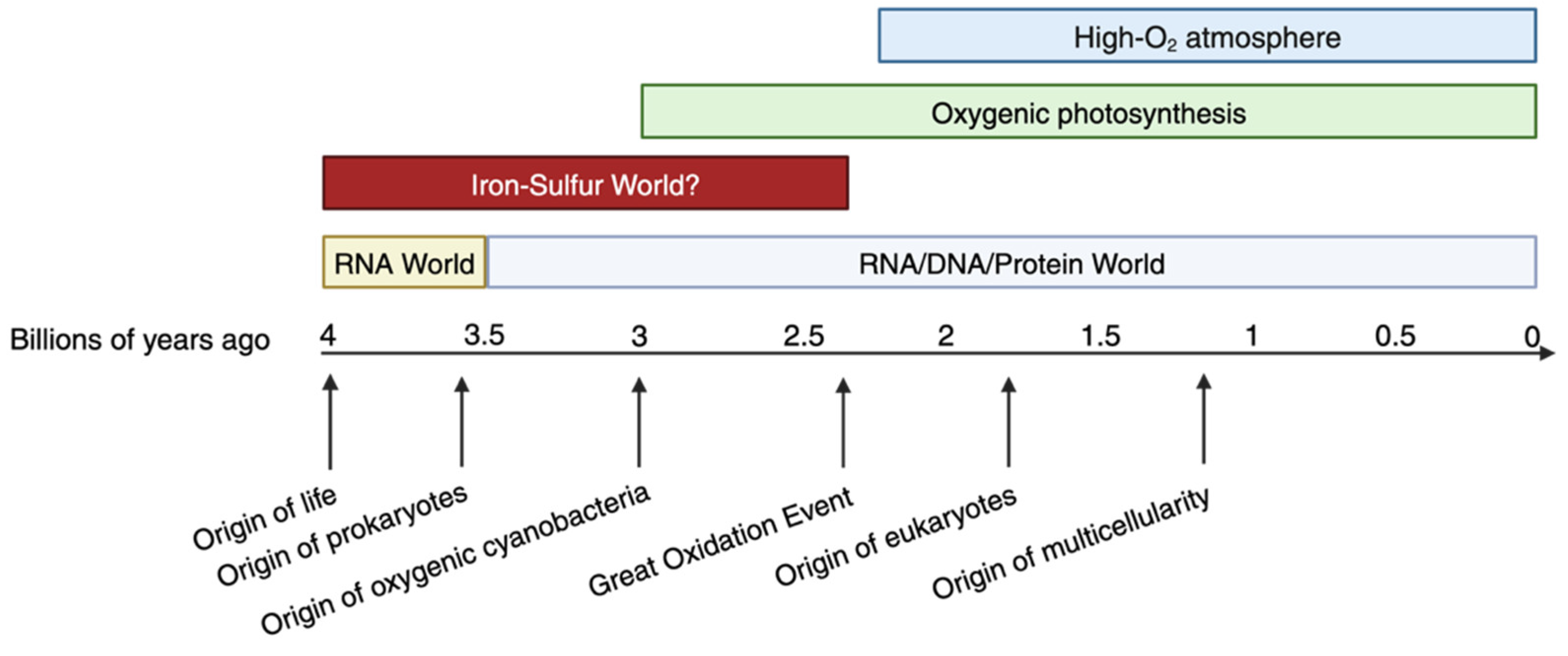
While the GOE eliminated much of the first life forms, the ghosts of the ancient Fe2+-mediated RNA folding, catalysis, and environmental sensing are still found in the descendants of those bacteria who survived the GOE. For example, iron has been shown to participate in folding the 23S ribosomal RNA (rRNA) from the thermophilic deep-sea vent-dwelling bacteria, Thermus thermophilus, in conditions that mimic the ancient Earth in vitro. Strikingly, the 23S rRNA was found to fold in the presence of Fe2+ or Mn2+ to the same extent as in the presence of Mg2+, which previously had been thought to be the primary cofactor for rRNA folding. In fact, a lower concentration of Fe2+ was required to achieve the same extent of folding compared to Mg2+ [32]. Since rRNA folding is a crucial step in ribosome assembly, this suggests that iron has played a role in mediating translation since prehistory.
Iron has also been implicated as an ancient cofactor for catalysis by ribozymes. Early ribozymes, or RNA molecules capable of catalytic activity, were likely an important step in the evolution of proteins, with Fe2+ serving as an early RNA cofactor pre-GOE and subsequently being replaced by Mg2+ during and after the GOE. The hammerhead ribozyme, which was first discovered in plant viroids and is conserved in all kingdoms [33,34], was shown to display 3x higher rate of cleavage in the presence of Fe2+ than the presence of the same concentration of Mg2+ [35]. Several other modern RNAs were shown to be able to catalyze a single-electron transfer in a horseradish peroxidase activity assay in the presence of Fe2+ in an anoxic environment [36]. Since electron transfer is the foundation of metabolic reactions, this suggests that many RNA catalytic activities were lost or altered upon the onset of the GOE, as iron became oxidized and early life forms had to transition to utilize other metals.
Aside from RNA folding and ribozymes, iron mediates catalysis by a wide range of enzymes, including some nucleic acid-processing enzymes in vitro. A DNA ligase from an acidophilic ferrous iron-oxidizing archeaon, Ferroplasma acidiphilum, coordinates two Fe3+ ions at a pH optimum between 2.0-3.0. It is neither dependent on nor stimulated by Mg2+, which is required by most other known DNA ligases [37]. While this might represent a highly specialized protein evolved in an organism according to its specific environment, there is other evidence that Fe2+ can be used by other nucleic acid processing enzymes. One study tested the activity of the Deep Vent® (exo-) DNA polymerase from New England Biolabs, T7 RNA polymerase, and T4 DNA ligase in the presence of Fe2+ instead of Mg2+. They showed that Fe2+ could replace Mg2+ as a cofactor for the DNA polymerase, and that the DNA ligase could function with Fe2+ instead of Mg2+, albeit with less efficiency [38]. The RNA polymerase synthesized RNA from a DNA template at lower concentrations of Fe2+ than was required to complete the RNA polymerization using Mg2+ [38]. The fact that these enzymes, which today are commonly thought to prefer Mg2+ or Mn2+, can seemingly revert to an ancient activity in pre-GOE conditions provides further evidence of a conserved biochemical function.
Biological metal-ligand complexes formed by divalent metal ions are ranked in order of the stability of the interaction between the metal and its binding partner in water in the Irving-Williams series: Mg2+ < Mn2+ < Fe2+ < Co2+ < Ni2+ < Cu2+ > Zn2+ [39]. The physiological “free” (unbound) concentrations of these metal ions in cells follows the reverse trend, such that the ions bound in biological complexes most weakly (Mg2+, Mn2+, and Fe2+) are the most bioavailable [40]. Proteins that selectively regulate the transport of these metals are generally able to bind a range of other metals. Consequently, metal homeostasis in cells is thought to depend more on selective preference informed by environmental conditions (such as bioavailable ion concentrations) to bind the cognate ion than on binding specificities [40,41]. Interestingly, protein-Fe3+ complexes in aqueous biological systems are typically more stable than protein-Fe2+ complexes, because Fe2+ has a 3d6 electron configuration, while Fe3+ has a 3d5 configuration. This is because the half-filled d-orbital of Fe3+ is more energetically stable than that of Fe2+; furthermore, having fewer electrons than Fe2+ makes Fe3+ a smaller ion and allows it to form stronger coordinate bonds due to having a greater electrostatic attraction to negatively charged ligands [41]. This suggests the possibility that while Fe3+ complexes may be more stable, the ability to adopt the Fe2+ state, or to convert between Fe2+ and Fe3+, is beneficial for biological processes where such reactivity drives signaling pathways.
Many of the best-characterized (and most highly conserved) iron-binding proteins are those that function in iron storage and mobilization or in electron transport reactions. For example, cytochromes contain complex heme groups that have a ferrous iron atom at the core in its oxygenated state and can be found in organisms across all domains of life. They function as crucial redox-active proteins in the electron transport chain. Ferritins, also present in nearly all organisms, are the primary iron storage proteins that sequester iron inside of cells and release it in a controlled manner according to the cell’s needs. As we try to better understand how evolution and the changing environment have shaped the bacteria that exist today, extensive work has gone into the study of the regulation of iron homeostasis, that is, the expression of transcripts coding for proteins that function in various aspects of iron metabolism. The relationship between gene expression and iron homeostasis can be thought of as cyclical, wherein cellular levels and redox states of iron directly influence the expression of genes that enact important processes to respond to the current iron status [42]. In order to learn as much as possible about the bacteria that pose health threats today, it is important to also consider the more oblique angle of the RNA-iron relationship—the impact that iron can have on post-transcriptional RNA modifications, processing and maturation by ribonucleases (RNases), and degradation (Figure 2). Below is a synopsis of disseminated reports about the functions of iron in various aspects of RNA metabolism.
Figure 2.
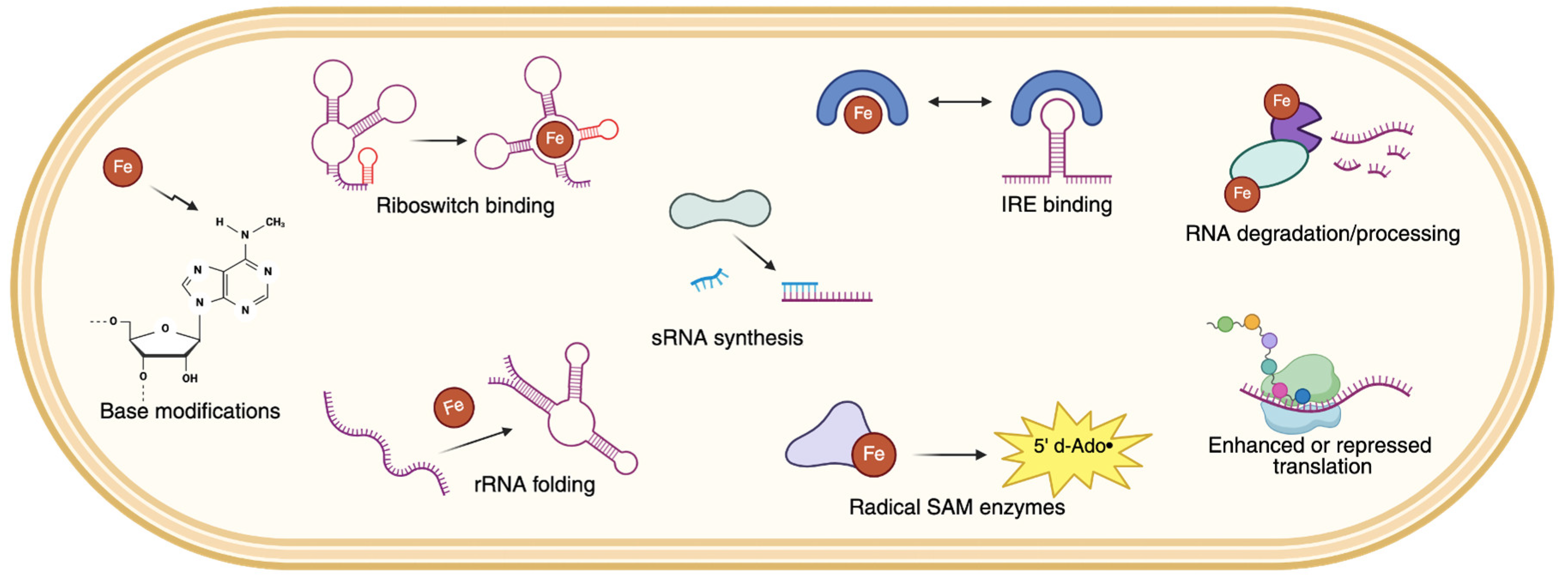
The various impacts of iron on bacterial RNA metabolism. Examples of each mechanism discussed in this work are as follows: Base modifications, such as post-transcriptional methylation. Riboswitch folding, such as czcD in some gut bacteria. rRNA folding, as seen with Thermus thermophilus 23S rRNA in vitro. IRE binding, such as E. coli aconitase. sRNA synthesis, such as E. coli RyhB. Radical SAM enzymes, such as methylthiotransferases. RNA degradation/processing, such as B. subtilis RicT and the Y-complex. Enhanced or repressed translation, as a result of RNA abundance and stability.
Figure 2.
The various impacts of iron on bacterial RNA metabolism. Examples of each mechanism discussed in this work are as follows: Base modifications, such as post-transcriptional methylation. Riboswitch folding, such as czcD in some gut bacteria. rRNA folding, as seen with Thermus thermophilus 23S rRNA in vitro. IRE binding, such as E. coli aconitase. sRNA synthesis, such as E. coli RyhB. Radical SAM enzymes, such as methylthiotransferases. RNA degradation/processing, such as B. subtilis RicT and the Y-complex. Enhanced or repressed translation, as a result of RNA abundance and stability.
2. Direct Roles of Iron in RNA Cleavage
A Brief Overview of the Machinery That Processes and Degrades Bacterial RNA
Degradation of RNA in bacteria is generally thought to be governed by multiprotein complexes called RNA degradosomes, or more transiently interacting proteins in degradosome-like networks. RNA degradation proteins function not just in the turnover of mRNA, but also in the maturation of rRNAs, as discussed in more detail later in this section. When bacteria grow rapidly, mRNA degradation generally occurs at high rates transcriptome-wide. In contrast, it has been shown that when bacteria are faced with stressful conditions, global RNA degradation is generally slowed, stabilizing the transcriptome and allowing the cells to conserve energy (reviewed in [43]). Furthermore, transcript-specific changes in degradation rates allow for upregulation of certain genes implicated in the response to that stressor [44,45]. Thus, the ability to regulate transcript abundance, which is a balance between transcription and degradation, is critical to bacterial survival and adaptation.
The general model of the degradosome includes a scaffolding endoribonuclease (typically RNase E or RNase Y), an RNA helicase, and a 3’ to 5’ exoribonuclease (typically PolyNucleotide Phosphorylase [PNPase]) [46,47,48,49,50,51,52]. Auxiliary proteins, including additional RNases, helicases, and glycolytic enzymes such as enolase, have been associated with RNA degradation as well [49,53,54]. While the leading knowledge of degradosome characterization has historically come from Escherichia coli, species-specific diversity of RNases or other proteins have been described [55,56,57,58,59]. For example, in mycobacteria, the GC-rich family of bacteria containing deadly pathogens including Mycobacterium tuberculosis, RNase E plays a crucial role in bulk mRNA degradation, while the dual endo/5-to-3’ exonuclease RNase J has a specialized role in degradation of highly structured transcript fragments [60,61,62,63,64,65]. In Bacillus subtilis, a degradosome-like network has been proposed instead of a fixed multiprotein complex, due to the transitory nature of protein-protein interactions that have only been captured after crosslinking [62,66,67,68]. RNase Y is the central endonuclease in Firmicutes, which canonically encode an RNase J1 and RNase J2 [69,70,71].
RNases require metal ions for enzymatic activity, typically divalent manganese or magnesium (Figure 3). RNase E requires either divalent magnesium or manganese for catalysis, and requires zinc for structural stabilization [72,73,74]. RNase Y typically requires magnesium, but it has been suggested that it can also use manganese or zinc [75]. There is less evidence for metal cofactors bound by RNase G, but it is believed to require magnesium or manganese [76,77,78], which RNase J typically requires for activity as well [70,79]. The single-stranded 3’ to 5’ exoribonucleases RNase II and RNase R, the latter of which can also cleave some double-stranded RNAs, typically use a two-metal mechanism involving one monovalent and one divalent cation [80,81,82,83,84]. Lastly, there is evidence of PNPase using multiple metal ions, which will be addressed in more detail in the next section.
Figure 3.
Venn Diagram of the major RNases in well-characterized bacterial phyla and their relevant metal ions. Note that this is not an exhaustive list of all RNases, RNA-binding proteins, or RNA degradation-associated proteins, nor the diversity of functional analogs.
Figure 3.
Venn Diagram of the major RNases in well-characterized bacterial phyla and their relevant metal ions. Note that this is not an exhaustive list of all RNases, RNA-binding proteins, or RNA degradation-associated proteins, nor the diversity of functional analogs.
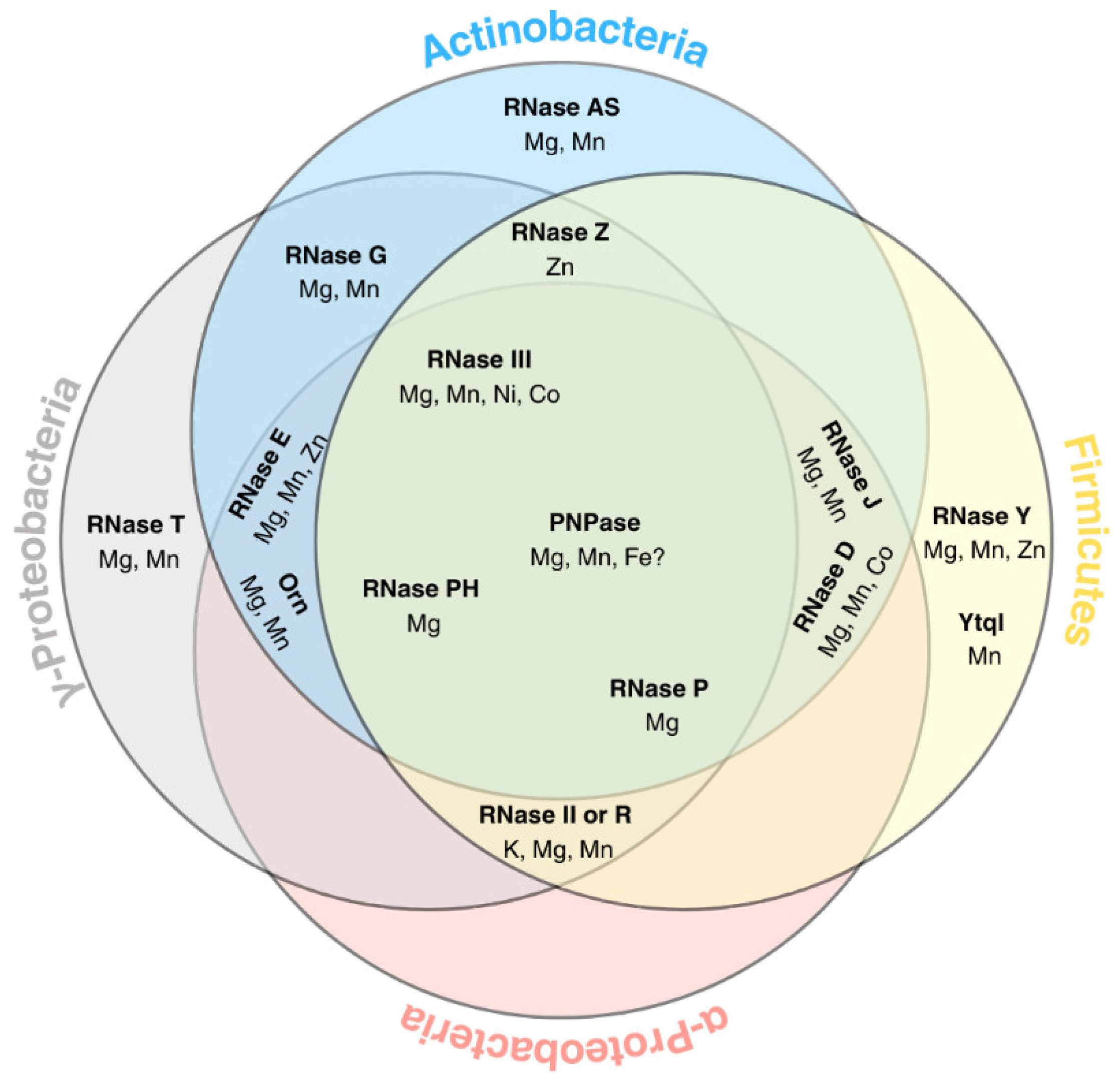
Aside from bulk turnover of mRNA, RNases also process stable RNAs by cleaving and/or trimming longer precursors RNAs into mature RNAs that perform important functions in the cell. A major example is rRNA. In bacteria, rRNAs are encoded in a single, highly-structured polycistron that is processed by RNases to produce the three mature rRNAs: 23S, 16S, and 5S [60,85,86]. Precursors of the mature rRNAs are produced by endoribonucleolytic cleavages in the leader, spacer, and trailer regions, and further trimmed by exoribonucleases [60,87,88]. The enzymes thought to participate in rRNA maturation in bacteria are PNPase, RNase E, RNase G, or RNase Y, the double-stranded endonuclease RNase III, RNase J, and the 3’ to 5’ exoribonuclease RNase T [60,89,90,91,92,93,94]. RNase III is typically most efficient with magnesium, but has also been shown to use manganese, nickel, or cobalt, and can also bind calcium but is not functionally active [95,96]. Actinobacteria canonically do not encode RNase T, but an enzyme with a similar structure has been identified in mycobacteria and was named RNase AS for its strong specificity for adenylate-rich RNAs [97]. Oligoribonuclease (Orn) degrades small fragments five nucleotides or shorter, and is broadly conserved among Proteobacteria and Actinobacteria. While Orn is absent from the Firmicutes, a functional analog, Ytql, has been identified in B. subtilis. Orn, as well as RNase T, require divalent cations for activity such as magnesium or manganese [98,99,100].
Aside from rRNAs, tRNAs—some of which are encoded within rRNA operons or within mRNAs—are processed by several RNases across species, including RNase D, RNase P, RNase T, RNase Z, PNPase, and RNase PH [91,93,101,102,103]. The phosphorolytic RNase PH has been shower to require magnesium for its enzymatic function, using inorganic phosphate as a substrate for nucleophilic attack on the phosphodiester backbone [104]. The ribozyme RNase P also requires magnesium [105,106]. The 3’ to 5’ exoribonuclease RNase D, thought to be involved in processing of tRNAs, 5S rRNA, and other small stable RNAs, has been shown to require divalent magnesium, manganese, or cobalt for activity [107]. The endonuclease RNase Z coordinates zinc and participates in the 3’ maturation of tRNAs [108].
PNPase Is a Multifunctional Metal-Binding Enzyme
Among the proteins that carry out mRNA degradation and stable RNA processing, PNPase is a unique enzyme with multiple catalytic capabilities. It has phosphorolytic 3’-to-5’ exoribonucleolytic activity, converting the 3’ phosphodiesters of RNAs to nucleoside diphosphates (NDPs), and broadly participates in RNA degradation and processing [109]. PNPase can also polymerize RNA by using NDPs as substrates to catalyze addition of NMPs to the 3’ hydroxyl termini of RNAs, releasing inorganic phosphate [109,110,111]. Lastly, PNPase can function as a 3’ DNA polymerase, using dADP as a substrate [112]. PNPase requires the coordination of metal ions for catalysis, using divalent manganese for DNA substrates [113,114] and divalent magnesium for RNA substrates [112,115,116]. This one of many examples across the tree of life of multifunctional enzymes that can perform different reactions having vastly different downstream effects depending on the identity of the metal cofactor. One such enzyme is acireductone dioxygenase, an enzyme first isolated from Klebsiella pneumoniae that when bound to Fe2+ catalyzes the penultimate step in the methionine salvage pathway, but when oxidized to Fe3+ is enzymatically inactive, and when bound to Mn2+, Ni2+, and Co2+ can catalyze off-pathway reactions that have been linked to carcinogenesis in humans [117,118,119,120,121,122,123,124]. Thus, both the identity and oxidation state of a metal cofactor can profoundly influence enzymatic activity.
There is some evidence in the literature, albeit scant, that PNPase can use iron as a cofactor. The PNPase isolated from Micrococcus luteus was able to polymerize DNA in vitro when incubated with dNDPs and FeCl3) [125], but not with MgCl2.Additionally, the PNPase purified from E. coli was able to polymerize dNDPs in vitro using oligo-A as a primer in the presence of Fe3+, but not Fe2+, at concentrations less than 0.5 mM, which are low enough to occur physiologically in cells [126]. The widespread conservation of PNPase suggests it originated early in microbial evolution and may preserve traces of ancient activity from the Iron-Sulfur World to the post-GOE era [58,127,128].
Some Iron-Sulfur Proteins Participate in RNA Processing
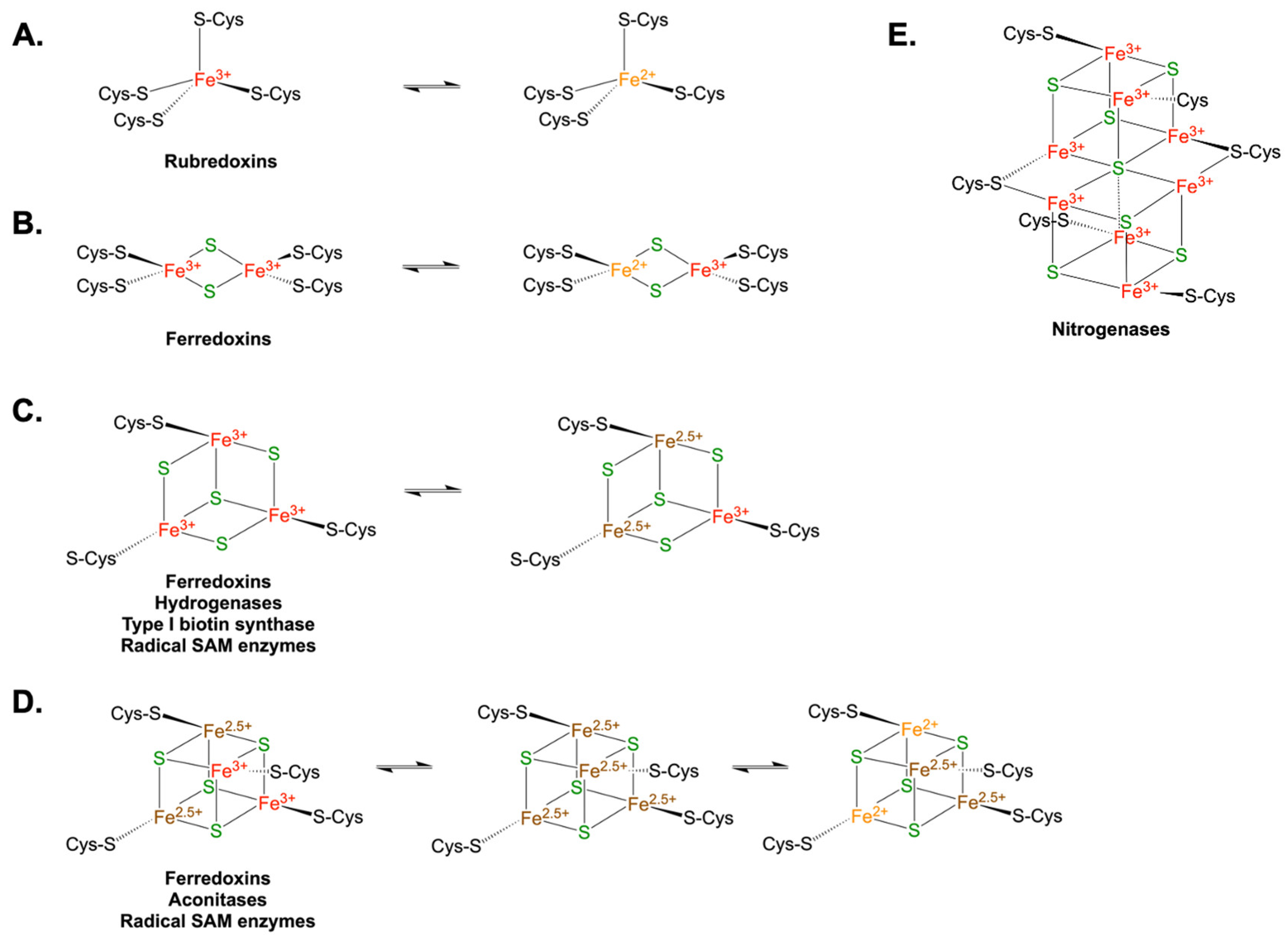
A wide breadth of proteins participates in the synthesis, cleavage, and modification of RNA in cells. One remarkable family of proteins whose roles in various RNA metabolic processes continue to emerge are iron-sulfur (Fe-S) proteins, so named because they coordinate Fe-S clusters. FeS clusters are highly diverse, varying widely in the number of iron and sulfur atoms that comprise the clusters, their typical physiological oxidation state(s), and the amino acid residues of the protein that participate in cluster coordination [129,130,131,132]. In rubredoxins, small sulfur-metabolizing proteins found in bacteria and archaea, one iron atom is bound by four cysteine residues (1Fe-0S) (Figure 4A) [133]. A Rieske cluster is formed when two iron atoms and two sulfurs (2Fe-2S) are coordinated by a combination of two cysteine and two histidine residues [134]. Other rhombic 2Fe-2S clusters coordinated by four cysteines are commonly found in ferredoxins, as well as ferrochelatases (Figure 4B) [135,136]. Ferredoxins, hydrogenases, type I biotin synthetases, and some radical SAM enzymes possess Fe-S clusters with three iron atoms and four sulfur atoms (3Fe-4S) (Figure 4C) [137,138,139]. Other radical SAM enzymes, which will be discussed in more detail in a later section, as well as other ferredoxins and aconitases, have clusters containing four iron atoms and four sulfur atoms (4Fe-4S) (Figure 4D) [140,141]. These are typically referred to as cubane clusters. Larger, more complex Fe-S clusters include the P-cluster found in nitrogenases of nitrogen-fixing bacteria, as well as clusters with other moieties such as molybdenum, homocitrate, or heme groups (Figure 4E) [142,143,144,145,146,147].
Figure 4.
General schematic of common iron-sulfur clusters with examples of possible redox states and proteins in which they are typically found. A. 1Fe-0S clusters. B. 2Fe-2S or rhombic clusters. C 3Fe-4S clusters. A charge of 2.5+ indicates a shared electron valence shell. D 4Fe-4S or cubane clusters. A charge of 2.5+ indicates a shared valence shell. E. An all-ferric 8Fe-7S P-cluster, which can also adopt partially oxidized or fully reduced forms.
Figure 4.
General schematic of common iron-sulfur clusters with examples of possible redox states and proteins in which they are typically found. A. 1Fe-0S clusters. B. 2Fe-2S or rhombic clusters. C 3Fe-4S clusters. A charge of 2.5+ indicates a shared electron valence shell. D 4Fe-4S or cubane clusters. A charge of 2.5+ indicates a shared valence shell. E. An all-ferric 8Fe-7S P-cluster, which can also adopt partially oxidized or fully reduced forms.
The crux of Fe-S cluster functionality is a property that also makes them difficult to study; Fe-S clusters are inherently highly prone to oxidation. As such, they are extremely sensitive to the levels of not just iron and sulfur but also O2, nitric oxide, and other redox-active molecules in their environments. This property underlies the utility of iron as a cofactor for electron-shuttling enzymes as well as a component of environmental sensors. Many Fe-S proteins thus regularly cycle between different oxidation states and/or atomic stoichiometries in cells, with each having a unique function or activity [130]. One can imagine how easily the structures and functions of such proteins can be mischaracterized, overlooked, or otherwise confounded in the lab if great care is not taken to stabilize them. Such sensitivity to the elemental milieu is thought to be an evolutionary adaptation that conferred upon our ancestors the ability to detect changing conditions and readily signal for homeostatic regulation.
While Fe-S clusters can assemble spontaneously in certain conditions, provided the optimal concentrations of iron, sulfur, O2 (or lack thereof), and polypeptides, all domains of life encode a compendium of proteins dedicated to the biogenesis of Fe-S clusters, underscoring the complexity of the regulation of these proteins and their downstream effects [130]. Generally speaking, Fe-S cluster biosynthesis takes place in three steps: donation of a sulfur moiety typically mediated by a cysteine desulfurase, assembly of a cluster on a scaffold protein, and transfer of the cluster to a target protein [148,149]. Three distinct Fe-S biogenesis operons have been identified in bacteria: nif (named for nitrogen fixation), suf (named for sulfur mobilization), and isc (named for iron-sulfur clusters). Unsurprisingly, there is variation between species in which operons are encoded or used. For example, the E. coli suf operon is only expressed upon stress, and the isc operon is typically used to assemble the clusters under normal conditions [150,151]. However, in M. tuberculosis, the suf machinery is the major Fe-S biogenesis pathway [151].
Both volatile and versatile, Fe-S proteins participate in an extensive variety of cellular processes and can trigger diverse signaling cascades [130]. Perhaps the most commonly-reported role for Fe-S clusters is to serve as electron carriers in redox reactions, including but not limited to aerobic respiration and photosynthesis [130]. Fe-S clusters can also serve as cofactors for enzyme catalysis; for example, the nitrogenases in nitrogen-fixing bacteria are Fe-S cluster proteins that convert nitrogen gas into biologically available ammonia [152]. While their roles in electron carrying and oxidative sensing have been well-documented, over the past decade or so, more reports have been published of Fe-S clusters participating in binding to and processing various nucleic acids across the domains of life [153]. This was originally not expected, as iron (especially ferrous iron), can readily generate harmful reactive oxygen species (ROS) when reacting with hydrogen peroxide, which can cause nucleic acid damage. Nonetheless, the RNA polymerase domains of eukaryotic DNA primases contain an iron-sulfur cluster predicted to be in the 4Fe-4S structure [154]. While conserved in archaea, it appears that bacterial DNA primases lack this cluster. That being said, there is a growing body of evidence that Fe-S proteins have roles in RNA metabolism in bacteria.
To the best of our knowledge, there is only one report to date of an Fe-S protein participating directly in RNA cleavage. The aforementioned degradosome-like network of B. subtilis is comprised of RNase Y, RNase J1, RNase J2, PNPase, the DEAD-box helicase CshA, enolase, phosphofructokinase, and sometimes glyceraldehyde 3-phosphate dehydrogenase A and/or smaller scaffolding proteins [66,70,75,155,156,157]. Distinct from the degradosome-like network, the B. subtilis Y-complex is a 1:1:1 heterotrimer formed by RicA (also called YmcA), RicF (also called YblF), and RicT (also called YaaT). RicT contains a cubane 4Fe-4S cluster, and it recently was shown that a second 4Fe-4S cluster forms in the Y-complex [158]. The Y-complex is thought to present RicT to RNase Y, forming a RicT-RNase Y complex. Both of the Fe-S clusters are required for the formation of a stable RicT-RNase Y complex [158,159]. While it has been shown that these two clusters are not necessary for the Y-complex to stay together, they are essential for the maturation of the cggR-gapA operon [159], which encodes a transcriptional repressor of genes involved in glycolysis (CggR) and glyceraldehyde-3-phosphate dehydrogenase (GapA). RNase Y cleavage near the 3’ end of the cggR coding sequence results in that portion of the operon being degraded while the downstream gapA coding sequence remains stable [160]. In strains lacking each component of the Y-complex, there was a loss of shorter gapA-encoding transcript isoforms processed from longer RNA precursors via RNase Y. The RicT-RNase Y heterodimer may thus act as the functional entity for maturation of that operon. The Fe-S clusters may participate in coordinating the interaction between RNase Y and its target RNA, perhaps by stabilizing a partially-unwound loop region of the transcript. At least 20 other operons were found to have Y-complex-dependent processing, indicating that it is needed to direct cleavage by RNase Y at a subset of sites across the transcriptome [160].
The identification of Fe-S clusters participating in RNA degradation could represent an emerging role for Fe-S clusters that has not previously been identified in bacteria. While RNase Y and the Ric proteins are generally conserved in Firmicutes, more work will be necessary to identify functional analogs of the Y-complex or other Fe-S proteins involved in RNA degradation in other species. To date, transient Fe-S clusters that form only when specific proteins are in complex have been reported mainly in the context of Fe-S biosynthesis or repair [161]. It may be possible that the Y-complex has an undescribed role in Fe-S biosynthesis or repair. Additionally, the three Ric proteins were originally identified as participating in B. subtilis sporulation, competence, and biofilm formation [162,163,164], underscoring the possibility that these clusters participate in diverse biochemical processes, some of which may have yet to be discovered.
Iron-Mediated Degradation of Ribosomal RNAs
There are some reports implicating iron in the mediation of rRNA degradation in vitro. Internal transesterification, also called in-line cleavage or strand scission, refers to a non-enzymatic, non-oxidative RNA self-cleavage, where the 2′-hydroxyl attacks the adjacent phosphate group, and is typically facilitated by Mg2+. However, E. coli rRNA was recently shown to be cleaved by redox-active Fe2+ in an in-line manner, with a rate constant 200x greater than the same experiment using Mg2+ [165].
Aside from in vitro work, in the budding yeast Saccharomyces cerevisiae, ribosome-bound iron has been shown to induce rRNA in-line attacks in oxidative stress, suggesting that some rRNAs may be primed for degradation as part of a redox-sensing stress-response mechanism [166]. Furthermore, it was found that Mn2+ competes with Fe2+ for rRNA-binding sites, and that protection of rRNA from Fe2+-mediated in-line cleavage by supplementation of excess Mn2+ restores cell viability [167]. This may represent physiological processes in cells. Additionally, a protective effect of Mn2+ being substituted for Fe2+ has been observed in some enzymes in bacteria, with oxidative stress inducing transcriptional changes causing Mn2+ uptake and Fe2+ sequestration, resulting in improved cell fitness [25,168,169]. It is possible that a similar stress response mechanism exists in bacteria to protect rRNA, perhaps another relic from the early struggle to survive in a changing landscape.
3. Iron-Mediated RNA Modifications
A Brief Introduction to Post-Transcriptional RNA Modifications
The function and lifespan of a given RNA is determined not only by its sequence, but also by post-transcriptional modifications. While prokaryotic RNAs lack the canonical 7-methylgyanosine 5’ cap found in eukaryotes, there is recent evidence showing that 5’ chemistry may affect vulnerability of RNA to degradation by RNases. RNAs are generally synthesized with 5’ triphosphates, but these can be converted to 5’ di- or monophosphates by RNA pyrophosphohydolases. The 5’ phosphorylation status, as well as other 5’ end chemistries originating from the use of non-canonical initiating nucleotides during transcription, have been implicated in regulation of transcript stability in E. coli by triggering degradation or stabilization of subpopulations of RNAs [170,171]. Additionally, and in contrast to eukaryotes, 3’ polyadenylation of bacterial transcripts typically promotes degradation [172]. There are several polyadenylate polymerases in bacteria, including PNPase [173]. The 3’ poly(A) extensions can recruit RNases, among them PNPase, to degrade the RNA [174].
Aside from modifications to transcript termini, many RNAs are modified by the addition of a chemical group or moiety to a nitrogenous base, such as methylation, hydroxylation, and acetylation. Other modifications include deamination, isomerization, and conjugation with other amino acids or sugars. Such modifications are an important tool for binding with protein partners and regulating transcript abundance and occur in all kingdoms. In bacteria, these modifications occur in mRNAs, tRNAs, and rRNAs. Certain base modifications, including methylations, thiouridylation, and pseudouridylation, are thought to have arisen early in evolution. Indeed, about 50% of the protein domains that are known to have some role in RNA metabolism—including base modifications, degradation, and polyadenylation—are highly conserved and can be traced back to our last universal common ancestor. Additionally, it has been suggested that nucleotide-modified RNAs provided a key stepping stone in the transition from the RNA World to the present DNA-Protein World; the earliest rRNAs and tRNAs were derived from chemical edits to pre-existing RNAs, giving rise to new RNA functions that eventually led to translation of proteins [175].
Radical SAM Enzymes Use an Fe-S Cluster to Generate a Radical Intermediate with Widespread Effects
There are myriad mechanisms by which RNA bases can be chemically modified [176,177], many of which do not involve iron as a cofactor. That said, one of the major players responsible for the chemical modification of various RNAs, as well as a multitude of other biochemical processes, does characteristically contain at least one 4Fe-4S cluster. The cluster participates in cleaving S-adenosyl-L-methionine (SAM), producing a radical intermediate, 5-deoxyadenosyl (5’-dAdo•) that is involved in the catalysis of diverse chemical reactions (Figure 5). Aptly named radical SAM enzymes, this enzyme superfamily is a diverse group of enzymes present in all domains of life. The 5’-dAdo• species can be used in many “difficult” chemical reactions, such as forming certain covalent carbon-carbon and carbon-sulfur bonds, sulfur transfers, isomerizations, ring insertions, and more [178]. Some biochemical processes that radical SAM enzymes participate in include the production of antibiotics, metalloprotein cluster biogenesis, post-translational modifications of proteins, and lipid metabolism, among others [179].
Radical SAM enzymes in bacteria have been reported to chemically modify various RNAs by addition of different moieties. One subgroup of radical SAM enzymes that act post-transcriptionally are methyltransferases, which donate methyl groups to specific bases on tRNA or rRNAs [180,181]. Interestingly, the general mechanism of methyltransferase reactions is a nucleophilic attack that does not involve redox chemistry. Thus, it seems unlikely that the redox reactivity of the Fe-S cluster participates directly in the catalytic activity of these enzymes, instead functioning primarily in the formation of the 5’-dAdo• intermediate, which then serves as the catalytic center [182,183]. Similarly, methylthiotransferases contain two 4Fe-4S clusters and at least one radical SAM catalytic domain [184,185,186,187]. They catalyze the methylthiolation of both RNAs and proteins [177,188].
Figure 5.
Simplified mechanism of action of a radical SAM enzyme 4Fe-4S cluster participating in SAM cleavage, generating the 5’-dAdo• radical species. The •CH2 radical is drawn in addition to the hydrocarbon skeleton for emphasis. Reactions that 5’-dAdo• can catalyze are indicated by arrows.
Figure 5.
Simplified mechanism of action of a radical SAM enzyme 4Fe-4S cluster participating in SAM cleavage, generating the 5’-dAdo• radical species. The •CH2 radical is drawn in addition to the hydrocarbon skeleton for emphasis. Reactions that 5’-dAdo• can catalyze are indicated by arrows.
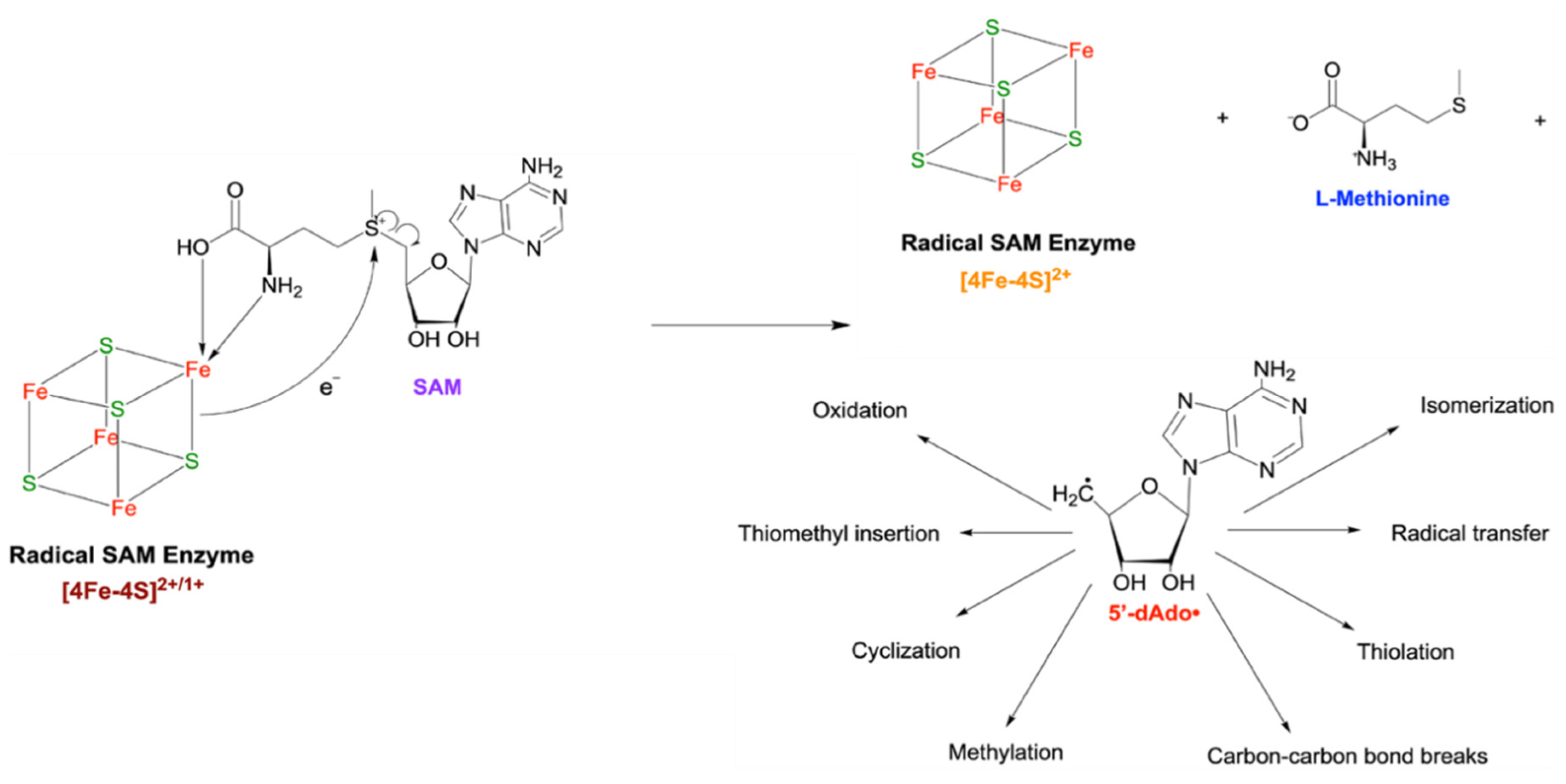
One area where radical SAM enzyme activity is critical is the chemical modification of rRNAs. Complete assembly of the mature ribosome requires base modifications such as methylation at important functional sites in the ribosome, including the large subunit-small subunit interface, the mRNA and tRNA binding sites, the peptidyl transfer center, and the nascent peptide exit channel [189,190]. Thus, rRNA chemical modifications are critical for translational fidelity and efficiency, having broader impacts on cell fitness, virulence, and host evasion [191]. There are several reports implicating Fe-S proteins in bacterial rRNA chemical modifications. Rlha (also called YdcP) is a radical SAM enzyme responsible for hydroxylating 23S rRNA at the C2501 position, producing 5-hydroxycytidine 2501 (ho5C2501) [192]. The production of ho5C2501 has been implicated in E. coli survival during oxidative stress [193]. Two other Fe-S proteins that modify rRNA in E. coli are RumA and RumB. RumA catalyzes transfer of a methyl group from SAM to the 23S U1939, producing a 5-methyluridine [194,195]. By a similar mechanism, RumB methylates U747 of the 23S [196]. Upon oxidation, the Fe-S cluster of RumA is disassembled, pointing at a possible regulatory function where the activity of RumA is regulated based on the oxidative environment or stresses within the cell [197]. Together, these studies reveal that Fe-S proteins play critical roles in rRNA modification, linking ribosome function to environmental adaptation.
tRNAs are also subject to iron-mediated chemistry in cells. Modified tRNA nucleotides have been implicated in translational fidelity [188,198,199,200]. Thiolated or methylthiolated tRNAs are required for efficient translation and stress responses in bacteria [201,202]. They are essential for proper recognition of the codons in mRNA or stabilization of tRNA structure [202]. Diverse reactions take place to accomplish these modifications [203,204]. For example, in E. coli and Salmonella enterica serovar typhimurium, the methylthiotransferase MiaB uses one radical SAM domain to synthesize the 5’-dAdo• radical that interacts with the RNA substrate and a second SAM domain to donate a sulfur atom to the RNA substrate [205,206]. YqeV similarly adds methylthiolate moieties to N6-threonylcarbamoyladenosines at position 37 of tRNAs, which are first made from adenosines by other enzymes [186]. Another modified nucleoside, queuosine, exists in the wobble position of tRNAs that have a 5’-GUN-3’ anticodon. The base is synthesized in bacteria using guanosine-5′-triphosphate and a series of eight enzymatic reactions. The last step in this pathway is catalyzed by epoxyqueuosine reductase, which is not a radical SAM enzyme but has been recently found to contain two 4Fe-4S clusters coordinated by eight cysteines and require cobalamin (vitamin B12) as a cofactor [207]. In the anaerobic hyperthermophile Thermotoga maritima, the epoxyqueuosine reductase homolog QueH has one 4Fe-4S cluster alongside an adjacent coordinated iron ion [208]. Together, these findings highlight the widespread use of Fe-S clusters in tRNA modification pathways across bacteria.
The oxidation states of Fe-S clusters and/or the abundance of iron has been shown to regulate tRNA modifications. For instance, the Fe-S protein MnmA produces 2-thiouridine (s2U) derivatives in the anticodons (position 34) of RNALys, tRNAGlu, and tRNAGln. An experiment with T. thermophilus MnmA, which has a 4Fe-4S cluster, only produced s2U34 in tRNALys and tRNAGln in vitro in anaerobic conditions, suggesting that oxidation of the cluster impairs catalytic activity [209]. In S. typhimurium, Klebsiella pneumoniae, and Pseudomonas aeruginosa, the methylthiolation of tRNATrp, yielding 2-methylthio-N6-isopentenyladenosine (ms2i6a), does not occur when the bacteria are deprived of iron [210]. The same modification has been reported to occur in E. coli, where tRNATyr, tRNAPhe, and tRNASer are similarly affected [211]. This is not necessarily ubiquitous across bacteria, however, as the same modification of tRNAtrp in Serratia marcescens was not affected by the presence or absence of iron [210]. Additionally, as opposed to the methyltransferases RumA and RumB, the E. coli tRNA m5U54 methyltransferase TrmA employs a nucleophilic attack on the SAM methyl group and does not have an Fe-S cluster itself [212]. Together, these findings illustrate that Fe-S proteins and iron availability play central and sometimes condition-dependent roles in regulating critical rRNA and tRNA modifications, with broad implications for bacterial translation, stress responses, and adaptation.
4. Iron-Mediated Regulation of Transcription and mRNA Degradation
Transcript Abundance Is Regulated in Response to Changing Conditions
All organisms must regulate gene expression in order to respond to environmental stimuli and survive, and genes involved in the sensing, storage, and metabolism of iron are essential for bacterial survival. Many known bacterial transcription factors contain Fe-S clusters, and as such are finely-tuned for sensing and responding to important signals including nitric oxide, oxygen, and reactive species to mediate adaptive responses [215]. For example, in E. coli, the transcription factor SoxR has two 2Fe-2S clusters and is activated by nitric oxide or other superoxides, responding to oxidative stress by activating genes implicated in the superoxide response [215,216,217]. When the clusters are reduced, such as under anaerobic conditions, its activity is lost; it can be reactivated by oxidation. Cluster oxidation results from a change in the redox balance of electron chain intermediates that maintain SoxR in its inactive reduced form [216]. In Actinobacteria, the WhiB-like family of transcription factors contain characteristic 4Fe-4S clusters [218]. Of these, M. tuberculosis WhiB3, which responds to changes in environmental gases and fluctuation of reducing cofactors such as NADH and NADPH, is one example of an Fe-S protein exhibiting different functions depending on the chemical composition of the cluster. Oxidized apo-WhiB3 binds DNA strongly, while reduced apo-WhiB3 does not bind DNA at all, and holo-WhiB3 with either (4Fe-4S)1+ or (4Fe-4S)2+ binds DNA weakly [219,220].
RNases have been shown to play roles in regulation of transcripts that respond to iron levels in the cell. In E. coli, PNPase forms a complex with the global small RNA (sRNA) chaperone Hfq, which has been well-characterized in Gram-negative bacteria but is absent from others. The PNPase-Hfq complex stabilizes some sRNAs that in turn impact mRNA levels [221]. Deletion of the pnp gene resulted in faster degradation of several sRNAs, including RyhB and CyaR, which are involved in regulation of iron-storage proteins and response to nutrient sensing, respectively [222]. Furthermore, in Staphylococcus aureus, the gene encoding ferritin is regulated by RNase III and PNPase acting at the 3’ UTR to degrade the ftnA transcript [223]. Deletion of the ftnA 3’ UTR therefore allowed ferritin to be expressed more highly. Notably, S. aureus expressing ftnA with its 3’ UTR deleted had a growth defect in iron-starved conditions. This may suggest that high levels of ferritin in iron-starved conditions result in excess iron sequestration, at a cost to the essential functions that require iron as a cofactor [223]. Thus, while more experimental work is needed to clarify any iron-binding capacities, the regulation of iron-sensing genes by PNPase points at its broader role in transcript stabilization and iron metabolism.
Aconitase and Iron-Response Elements
Aconitases are ancient, ubiquitous Fe-S enzymes [224,225]. Eukaryotes have one aconitase isozyme in the mitochondria (m-Acn) that was probably evolved from bacterial aconitase, and a second is in the cytosol (c-Acn) [226]. Aconitase catalyzes the reversible isomerization of citrate to isocitrate via a cis-aconitate intermediate in the citric acid cycle, and also plays a regulatory role by binding RNA. This bifunctionality is regulated by the assembly and disassembly of its Fe-S cluster, making the two functions mutually exclusive [224,227,228]. In eukaryotes, only c-Acn, also called iron regulatory protein 1 (IRP1) exhibits this bifunctionality. The RNA binding of aconitase occurs via the iron-response or iron-regulatory element (IRE). The IRE is a regulatory motif present in the untranslated regions of mRNAs involved in various aspects of iron homeostasis. The IRE is typically an ~30-nt long stem loop region that is bound by iron-response element-binding proteins (IRE-BP, also called IRBP or IRP). Canonical IREs are extensively characterized in eukaryotes [229]. Apo-IRP1 undergoes structural rearrangements upon Fe-S cluster disassembly that allow it to recognize and bind to IREs that regulate the abundance and/or translation of the mRNAs for such genes as ferritins and transferritin receptors. The impact of Apo-IRP1 binding to IREs depends on the location of the IRE. If the IRE is located in the 5′ UTR, binding of IRP1 will inhibit translation; if the IRE is located in the 3′ UTR, binding of IRP1 will increase transcript stability and result in enhanced translation [230,231,232,233,234].
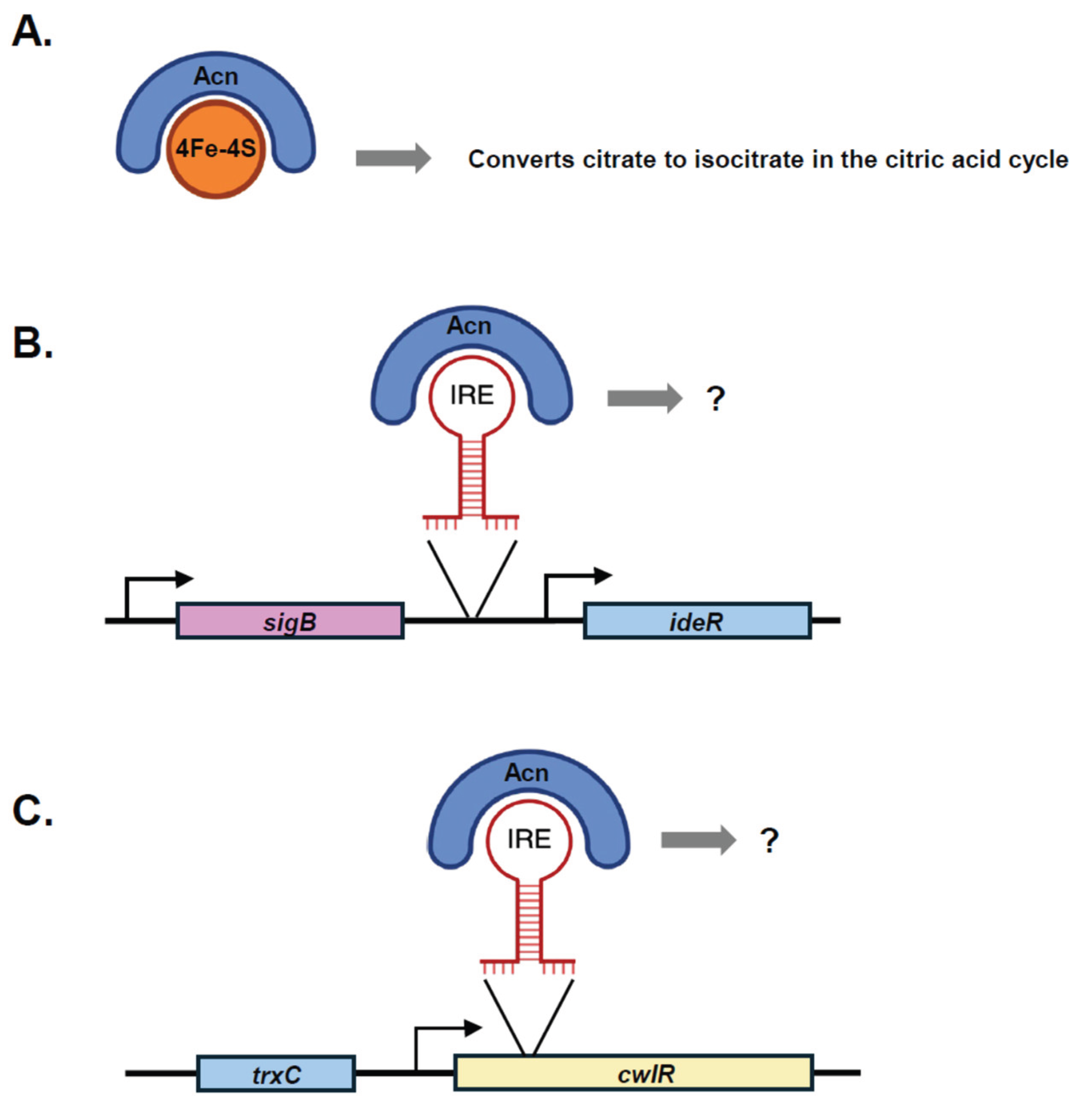
Evidence for the function and structures of prokaryotic aconitases is more recent than that of the eukaryotic homologs. The bifunctionality of aconitase has been shown in several species, including Streptomyces [235], E. coli [236], and mycobacteria [237]. Generally, aconitase with a 4Fe-4S cluster is enzymatically active, transporting electrons during the conversion of citrate to isocitrate. However, in the enzymatically inactive form that does not have the 4Fe-4S cluster, aconitase can act as an IRE-BP and modulate transcript stability or translation similar to the eukaryotic aconitase [140,238,239]. IRE-like sequences found in some bacteria are considered a departure from the traditional eukaryotic IRE model, as the loop sequence or position can vary [230]. In M. tuberculosis, IREs are also present in the UTRs of the aconitase (Acn) transcript as well as that of succinate dehydrogenase, another citric acid cycle enzyme, possibly suggesting that that iron-binding proteins may contribute to regulation of energy metabolism in response to iron [240]. Under iron-depleted conditions, M. tuberculosis Acn was reported to bind to IRE-like sequences present at the 3′ UTR of the thioredoxin (trxC) transcript and the 5′ UTR of the iron-dependent regulator ideR transcript [238]. However, inspection of the IRE locations together with data on transcription start site (TSS) locations reveals that they may not be co-transcribed with the reported genes [241] (Figure 6) . The IRE reported to be in the ideR 5’ UTR is upstream of the TSS closest to ideR, and may instead be in the 3’ UTR of sigB (Figure 6). The IRE reported to be in the 3’ UTR of trxC is located far downstream in the coding sequence of the following gene, cwlR, which has its own TSS (Figure 6). Many open questions therefore remain regarding the regulation of gene expression by Acn in M. tuberculosis. Additionally, it is not known if the form of Acn that binds RNA is fully Apo-Acn or a form with an altered Fe-S cluster. For example, an 3Fe-4S intermediate may form due to loss of the fourth labile iron in some organisms [242,243,244,245].
The two aconitases encoded by E. coli are AcnA and AcnB. Apo-AcnA enhances translation of superoxide dismutase (SodA), which converts superoxide into oxygen and hydrogen peroxide, protecting the cell from harmful ROS [246]. Apo-AcnA also binds and stabilizes its own transcript, enhancing AcnA translation [247]. E. coli AcnB, which is highly unstable and readily loses the fourth labile iron in its cluster, is thought to sense and respond to low levels of iron in the environment [248]. Apo-AcnB binds mRNA and represses translation of the sodA transcript. [246]. Curiously, mycobacterial Acn has higher sequence identity to E. coli AcnA than AcnB. Since the E. coli AcnA is induced during oxidative stress and stationary phase [236], and M. tuberculosis Acn is downregulated under nutrient starvation conditions and upregulated in iron toxicity [249,250], it is possible that the bacterial aconitase regulates additional transcripts with roles in energy metabolism aside from those involved in iron-associated regulation.
Figure 6.
Examples of M. tuberculosis iron-dependent Aconitase (Acn) functionality. A. With an intact 4Fe-4S cluster, Acn participates in the citric acid cycle. Upon iron depletion, Acn binds (B) an IRE that was reported to be in the 5’ UTR of the ideR transcript but may in fact be in the 3’ UTR of the sigB transcript, and (C) an IRE that was reported to be in the 3’ UTR of the trxC transcript but is in fact in the cwlR coding sequence and may be encoded on a separate transcript. In both cases, the impacts on stability and translation are unknown.
Figure 6.
Examples of M. tuberculosis iron-dependent Aconitase (Acn) functionality. A. With an intact 4Fe-4S cluster, Acn participates in the citric acid cycle. Upon iron depletion, Acn binds (B) an IRE that was reported to be in the 5’ UTR of the ideR transcript but may in fact be in the 3’ UTR of the sigB transcript, and (C) an IRE that was reported to be in the 3’ UTR of the trxC transcript but is in fact in the cwlR coding sequence and may be encoded on a separate transcript. In both cases, the impacts on stability and translation are unknown.
Aconitases in other bacteria have been implicated in the regulation of genes that are important for other cellular functions. The B. subtilis aconitase, CitB, binds the 3′ UTR of gerE, a transcriptional activator and repressor involved in sporulation [251], and this activity appears to be important for sporulation [252]. Similar to Acn in M. tuberculosis, while CitB has aconitase activity in the presence of iron, its RNA binding capacity is abolished by iron [252]. There is also evidence of regulatory activity for the aconitase of Xanthomonas campestris [253]. The aconitase of H. pylori, AcnB, regulates abundance of cell wall-modifying enzyme peptidoglycan deacetylase, PgdA. Apo-AcnB binds to the 3′-untranslated region of the pgdA transcript, an activity that is abolished upon the addition of iron, enhancing its stability and increasing abundance of the pgdA RNA [254]. Lastly, the aconitase of Caulobacter crescentus has been shown to bind the C-terminal domain of RNase E, suggesting some participation in RNA degradation [50].
Iron-Regulated sRNAs
sRNAs are important post-transcriptional regulators that affect a variety of cellular functions. Iron-regulated sRNAs have been reported to mediate the degradation or translation of diverse transcripts related to iron homeostasis as well as other aspects of bacterial physiology including responses to acid and oxidative stress, amino acid biosynthesis, carbon storage, virulence factors, and photosynthesis [255]. In E. coli, Fe-S biogenesis is itself mediated by sRNAs. Suf Fe-S cluster biosynthesis machinery is regulated by the iron-sensing protein Fur. Under iron-rich conditions, Fur-Fe2+ represses transcription of the suf operon. Fur-Fe2+ also represses transcription of the sRNA RyhB [256]. When iron becomes limited, repression by Fur is lifted, leading to rapid synthesis of RyhB. RyhB, with the help of the sRNA chaperone protein Hfq, represses transcripts encoding non-essential iron-binding proteins by promoting their degradation by RNase E [257,258]. RyhB also promotes siderophore production through both direct and indirect mechanisms (reviewed in [259]). RyhB also targets the isc operon, leading to the degradation of the 3’ end of the mRNA, inhibiting translation of most ISC proteins but allowing translation of IscR, which activates the suf operon [260]. This shift prioritizes Suf-dependent Fe-S cluster assembly when iron is scarce. A similar process has been observed for two sRNAs, PrrF1 and PrrF2 in P. aeruginosa [261], and for IsrR in S. aureus [262], which represses translation of MiaB under iron-deficient conditions. In mycobacteria, the sRNA MrsI is induced in iron-deprived conditions and targets the mRNA for bacterioferritin A [263,264,265,266].
Iron-Regulated Riboswitches
Riboswitches are short, non-coding regions of RNA that form secondary structures and undergo conformational changes in response to ligand binding. These ligands can include metal ions. Riboswitches are typically located in the 5’ UTR of an mRNA, and their changes in secondary structure can in some cases regulate whether that transcript is degraded or translated into protein, or in other cases regulate whether transcription is terminated within the 5’ UTR or continued through the coding sequence. Metal-responsive riboswitches have been recently proposed to regulate storage and transport of those metals that bind relatively weakly in biological complexes [41]. To date, the only known class of bacterial iron-binding riboswitches, found in numerous human gut microbiota and pathogenic bacteria, is the czcD riboswitch. Also called NiCo, this riboswitch was originally believed to respond to specifically nickel and cobalt [267]. The genes regulated by the czcD riboswitch in various bacteria include a magnesium importer, a likely Fe2+ exporter, and some cation diffusion facilitators thought to function in export of divalent metal ions including Fe2+ [268]. Over the course of studying czcD, first using an fluorescent aptamer [268] and then isothermal titration calorimetry [269], it was confirmed that czcD of Listeria monocytogenes, Erysipelotrichaceae bacterium, and Clostridium cellulolyticum responds preferentially to iron, which likely was overlooked because the initial work had been performed in aerobic conditions. A report of another alleged iron-responsive riboswitch in the sexually-transmitted pathogen Haemophilus ducreyi was later retracted [270].
Some riboswitches are influenced indirectly by Fe-S proteins [41]. For example, since radical SAM enzymes influence the availability of SAM, this can in turn influence SAM riboswitches. Some Gram-positive bacteria encode SAM riboswitches in a number of genes that code for methionine or cysteine synthesis [271,272,273]. I B. subtilis, the Y-complex, which contains two Fe-S clusters, has been shown to have a global effect on the abundance of riboswitch-containing transcripts [160]. Additionally, while RNase Y by itself does not contain an Fe-S cluster, it has been implicated in turnover of SAM riboswitches by cleaving upstream of the SAM-binding domain, producing fragments that are further cleaved by PNPase, RNase R, and RNase J1 [274]. These findings may suggest that Fe-S proteins have a broader role in regulating riboswitches, either directly or indirectly.
5. From Past to Future
It is well-documented that iron is one of the most important metals for biological processes, and has been since life first evolved. With respect to RNA metabolism, it is apparent that some iron-binding proteins may have roles regulating specific transcripts or participating in RNA degradation or processing, and ongoing work continues to elucidate these roles (Figure 2). The major challenge to studying iron-binding proteins, especially those containing Fe-S clusters, is the fact that iron is readily oxidized, often necessitating experiments to be performed in anaerobic conditions. Another challenge will be parsing through existing literature that may have drawn inaccurate conclusions due to such constraints.
From packaging the earliest nucleic acids to serving as a cofactor for the SARS-CoV-2 RNA polymerase [275], iron, sulfur, and sometimes oxygen have been critical to RNA chemistry for billions of years. It is no wonder why Fe-S clusters may have played a crucial role in the evolution of important biomolecules and metabolic reactions, as they have the remarkable capacity to integrate with peptides and serve as a center of catalytic activity as well as a structural element [153]. While Fe-S proteins have diverse functions not limited to the metabolism of RNA, the fact that most of these proteins seem to participate in essential functions may suggest that any Fe-S proteins performing auxiliary functions were selected against as the world became less accommodating to these fragile structures.
Author Contributions
Conceptualization and Writing (original draft): J.G.R.; Writing (review & editing): S.S.S.
Funding
This work was supported by the NSF grant number 1652756 (to SSS) and NIH grant numbers AI143575 and AI156415 (to SSS).
Acknowledgements
We thank members of the Shell lab for helpful discussions.
Conflict of Interest
The authors declare no conflict of interest.
References
- Holland, H.D. The Oceans; A Possible Source of Iron in Iron-Formations. Econ. Geol. 1973, 68, 1169–1172. [Google Scholar] [CrossRef]
- Klein, C. Some Precambrian banded iron-formations (BIFs) from around the world: Their age, geologic setting, mineralogy, metamorphism, geochemistry, and origins. Am. Miner. 2005, 90, 1473–1499. [Google Scholar] [CrossRef]
- Hazen, R.M.; Ferry, J.M. Mineral Evolution: Mineralogy in the Fourth Dimension. Elements 2010, 6, 9–12. [Google Scholar] [CrossRef]
- Wächtershäuser, G. Before enzymes and templates: theory of surface metabolism. Microbiol. Rev. 1988, 52, 452–484. [Google Scholar] [CrossRef] [PubMed]
- A Benner, S.; Ellington, A.D.; Tauer, A. Modern metabolism as a palimpsest of the RNA world. Proc. Natl. Acad. Sci. USA 1989, 86, 7054–7058. [Google Scholar] [CrossRef]
- Cleland, CE. 2007. Epistemological issues in the study of microbial life: alternative terran biospheres? Stud Hist Philos Biol Biomed Sci 38:847–861.
- Nutman, A.P.; Bennett, V.C.; Friend, C.R.L.; Van Kranendonk, M.J.; Chivas, A.R. Rapid emergence of life shown by discovery of 3,700-million-year-old microbial structures. Nature 2016, 537, 535–538. [Google Scholar] [CrossRef]
- Davies, P.C.; Benner, S.A.; Cleland, C.E.; Lineweaver, C.H.; McKay, C.P.; Wolfe-Simon, F. Signatures of a Shadow Biosphere. Astrobiology 2009, 9, 241–249. [Google Scholar] [CrossRef]
- Atkins JF, Gesteland RF, Cech T. 2011. RNA worlds: from life’s origins to diversity in gene regulation. Cold Spring Harbor Laboratory Press.
- Schirrmeister, B.E.; Gugger, M.; Donoghue, P.C.J.; Smith, A. Cyanobacteria and the Great Oxidation Event: evidence from genes and fossils. Palaeontology 2015, 58, 769–785. [Google Scholar] [CrossRef]
- Fournier, G.P.; Moore, K.R.; Rangel, L.T.; Payette, J.G.; Momper, L.; Bosak, T. The Archean origin of oxygenic photosynthesis and extant cyanobacterial lineages. Proc. R. Soc. B: Biol. Sci. 2021, 288, 20210675. [Google Scholar] [CrossRef]
- Garcia-Pichel, F.; Lombard, J.; Soule, T.; Dunaj, S.; Wu, S.H.; Wojciechowski, M.F.; Giovannoni, S.J. Timing the Evolutionary Advent of Cyanobacteria and the Later Great Oxidation Event Using Gene Phylogenies of a Sunscreen. mBio 2019, 10, e00561–19. [Google Scholar] [CrossRef]
- Rasmussen B, Buick R. 1999. Redox state of the Archean atmosphere: Evidence from detrital heavy minerals in ca. 3250–2750 Ma sandstones from the Pilbara Craton, Australia. Geology 27:115–118.
- Anbar AD, Duan Y, Lyons TW, Arnold GL, Kendall B, Creaser RA, Kaufman AJ, Gordon GW, Scott C, Garvin J, Buick R. 2007. A Whiff of Oxygen Before the Great Oxidation Event? Science 317:1903–1906.
- Bachan, A.; Kump, L.R. The rise of oxygen and siderite oxidation during the Lomagundi Event. Proc. Natl. Acad. Sci. USA 2015, 112, 6562–6567. [Google Scholar] [CrossRef] [PubMed]
- Holland, HD. 2002. Volcanic gases, black smokers, and the great oxidation event. Geochimica et Cosmochimica Acta 66:3811–3826.
- Catling, D.C.; Zahnle, K.J.; McKay, C.P. Biogenic Methane, Hydrogen Escape, and the Irreversible Oxidation of Early Earth. Science 2001, 293, 839–843. [Google Scholar] [CrossRef] [PubMed]
- Olson, S.L.; Kump, L.R.; Kasting, J.F. Quantifying the areal extent and dissolved oxygen concentrations of Archean oxygen oases. Chem. Geol. 2013, 362, 35–43. [Google Scholar] [CrossRef]
- Harel A, Bromberg Y, Falkowski PG, Bhattacharya D. 2014. Evolutionary history of redox metal-binding domains across the tree of life. Proceedings of the National Academy of Sciences of the United States of America 111:7042–7047.
- Aguirre, J.D.; Culotta, V.C. Battles with Iron: Manganese in Oxidative Stress Protection. J. Biol. Chem. 2012, 287, 13541–13548. [Google Scholar] [CrossRef]
- Ushizaka, S.; Kuma, K.; Suzuki, K. Effects of Mn and Fe on growth of a coastal marine diatom Talassiosira weissflogii in the presence of precipitated Fe(III) hydroxide and EDTA-Fe(III) complex. Fish. Sci. 2011, 77, 411–424. [Google Scholar] [CrossRef]
- Torrents, E.; Aloy, P.; Gibert, I.; Rodríguez-Trelles, F. Ribonucleotide Reductases: Divergent Evolution of an Ancient Enzyme. J. Mol. Evol. 2002, 55, 138–152. [Google Scholar] [CrossRef]
- Martin JE, Imlay JA. 2011. The alternative aerobic ribonucleotide reductase of Escherichia coli, NrdEF, is a manganese-dependent enzyme that enables cell replication during periods of iron starvation. Molecular Microbiology 80:319–334.
- Cotruvo, J.A.; Stubbe, J. Class I Ribonucleotide Reductases: Metallocofactor Assembly and Repair In Vitro and In Vivo. Annu. Rev. Biochem. 2011, 80, 733–767. [Google Scholar] [CrossRef]
- Anjem, A.; Varghese, S.; Imlay, J.A. Manganese import is a key element of the OxyR response to hydrogen peroxide in Escherichia coli. Mol. Microbiol. 2009, 72, 844–858. [Google Scholar] [CrossRef]
- Wolfe-Simon F, Starovoytov V, Reinfelder JR, Schofield O, Falkowski PG. 2006. Localization and role of manganese superoxide dismutase in a marine diatom. Plant Physiology 142:1701–1709.
- Athavale, S.S.; Petrov, A.S.; Hsiao, C.; Watkins, D.; Prickett, C.D.; Gossett, J.J.; Lie, L.; Bowman, J.C.; O'NEill, E.; Bernier, C.R.; et al. RNA Folding and Catalysis Mediated by Iron (II). PLOS ONE 2012, 7, e38024. [Google Scholar] [CrossRef]
- Schirrmeister, B.E.; de Vos, J.M.; Antonelli, A.; Bagheri, H.C. Evolution of multicellularity coincided with increased diversification of cyanobacteria and the Great Oxidation Event. Proc. Natl. Acad. Sci. USA 2013, 110, 1791–1796. [Google Scholar] [CrossRef]
- Crockford, P.W.; Kunzmann, M.; Bekker, A.; Hayles, J.; Bao, H.; Halverson, G.P.; Peng, Y.; Bui, T.H.; Cox, G.M.; Gibson, T.M.; et al. Claypool continued: Extending the isotopic record of sedimentary sulfate. Chem. Geol. 2019, 513, 200–225. [Google Scholar] [CrossRef]
- Crockford, P.W.; Bar On, Y.M.; Ward, L.M.; Milo, R.; Halevy, I. The geologic history of primary productivity. Curr. Biol. 2023, 33, 4741–4750.e5. [Google Scholar] [CrossRef] [PubMed]
- Knoll AH, Nowak MA. 2017. The timetable of evolution. Science Advances 3:e1603076.
- Bray MS, Lenz TK, Haynes JW, Bowman JC, Petrov AS, Reddi AR, Hud NV, Williams LD, Glass JB. 2018. Multiple prebiotic metals mediate translation. Proceedings of the National Academy of Sciences of the United States of America 115:12164–12169.
- Prody, G.A.; Bakos, J.T.; Buzayan, J.M.; Schneider, I.R.; Bruening, G. Autolytic Processing of Dimeric Plant Virus Satellite RNA. Science 1986, 231, 1577–1580. [Google Scholar] [CrossRef]
- Hammann, C.; Luptak, A.; Perreault, J.; de la Peña, M. The ubiquitous hammerhead ribozyme. RNA 2012, 18, 871–885. [Google Scholar] [CrossRef] [PubMed]
- Athavale, S.S.; Petrov, A.S.; Hsiao, C.; Watkins, D.; Prickett, C.D.; Gossett, J.J.; Lie, L.; Bowman, J.C.; O'NEill, E.; Bernier, C.R.; et al. RNA Folding and Catalysis Mediated by Iron (II). PLOS ONE 2012, 7, e38024. [Google Scholar] [CrossRef]
- Hsiao, C.; Chou, I.-C.; Okafor, C.D.; Bowman, J.C.; O'Neill, E.B.; Athavale, S.S.; Petrov, A.S.; Hud, N.V.; Wartell, R.M.; Harvey, S.C.; et al. RNA with iron(II) as a cofactor catalyses electron transfer. Nat. Chem. 2013, 5, 525–528. [Google Scholar] [CrossRef]
- Ferrer, M.; Golyshina, O.V.; Beloqui, A.; Böttger, L.H.; Andreu, J.M.; Polaina, J.; De Lacey, A.L.; Trautwein, A.X.; Timmis, K.N.; Golyshin, P.N. A purple acidophilic di-ferric DNA ligase from Ferroplasma. Proc. Natl. Acad. Sci. USA 2008, 105, 8878–8883. [Google Scholar] [CrossRef]
- Okafor, C.D.; Lanier, K.A.; Petrov, A.S.; Athavale, S.S.; Bowman, J.C.; Hud, N.V.; Williams, L.D. Iron mediates catalysis of nucleic acid processing enzymes: support for Fe(II) as a cofactor before the great oxidation event. Nucleic Acids Res. 2017, 45, 3634–3642. [Google Scholar] [CrossRef]
- Irving, H.; Williams, R.J.P. Order of Stability of Metal Complexes. Nature 1948, 162, 746–747. [Google Scholar] [CrossRef]
- Foster AW, Young TR, Chivers PT, Robinson NJ. 2022. Protein metalation in biology. Current Opinion in Chemical Biology 66:102095.
- Xu J, Cotruvo JA. 2022. Iron-responsive riboswitches. Current opinion in chemical biology 68:102135.
- Andrews, S.C.; Robinson, A.K.; Rodríguez-Quiñones, F. Bacterial iron homeostasis. FEMS Microbiol. Rev. 2003, 27, 215–237. [Google Scholar] [CrossRef]
- Vargas-Blanco, D.A.; Shell, S.S. Regulation of mRNA Stability During Bacterial Stress Responses. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Vargas-Blanco, D.A.; Zhou, Y.; Zamalloa, L.G.; Antonelli, T.; Shell, S.S.; Barkan, D. mRNA Degradation Rates Are Coupled to Metabolic Status in Mycobacterium smegmatis. mBio 2019, 10. [Google Scholar] [CrossRef]
- Vargas-Blanco, D.A.; Shell, S.S. Regulation of mRNA Stability During Bacterial Stress Responses. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Carpousis, A.J.; Van Houwe, G.; Ehretsmann, C.; Krisch, H.M. Copurification of E. coli RNAase E and PNPase: Evidence for a specific association between two enzymes important in RNA processing and degradation. Cell 1994, 76, 889–900. [Google Scholar] [CrossRef] [PubMed]
- Carpousis, A.J. The RNA Degradosome of Escherichia coli: An mRNA-Degrading Machine Assembled on RNase E. Annu. Rev. Microbiol. 2007, 61, 71–87. [Google Scholar] [CrossRef] [PubMed]
- Marcaida, M.J.; DePristo, M.A.; Chandran, V.; Carpousis, A.J.; Luisi, B.F. The RNA degradosome: life in the fast lane of adaptive molecular evolution. Trends Biochem. Sci. 2006, 31, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Morita, T.; Kawamoto, H.; Mizota, T.; Inada, T.; Aiba, H. Enolase in the RNA degradosome plays a crucial role in the rapid decay of glucose transporter mRNA in the response to phosphosugar stress in Escherichia coli. Mol. Microbiol. 2004, 54, 1063–1075. [Google Scholar] [CrossRef]
- Hardwick, S.W.; Chan, V.S.Y.; Broadhurst, R.W.; Luisi, B.F. An RNA degradosome assembly in Caulobacter crescentus. Nucleic Acids Res. 2010, 39, 1449–1459. [Google Scholar] [CrossRef]
- Tejada-Arranz, A.; Galtier, E.; El Mortaji, L.; Turlin, E.; Ershov, D.; De Reuse, H.; Charpentier, E. The RNase J-Based RNA Degradosome Is Compartmentalized in the Gastric Pathogen Helicobacter pylori. mBio 2020, 11. [Google Scholar] [CrossRef]
- Bernardini, A.; Martínez, J.L. Genome-wide analysis shows that RNase G plays a global role in the stability of mRNAs in Stenotrophomonas maltophilia. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Li, Z.; Pandit, S.; Deutscher, M.P. RNase G (CafA protein) and RNase E are both required for the 5′ maturation of 16S ribosomal RNA. EMBO J. 1999, 18, 2878–2885. [Google Scholar] [CrossRef]
- Durand, S.; Condon, C.; Storz, G.; Papenfort, K. RNases and Helicases in Gram-Positive Bacteria. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef]
- Aït-Bara, S.; Carpousis, A.J.; Quentin, Y. RNase E in the γ-Proteobacteria: conservation of intrinsically disordered noncatalytic region and molecular evolution of microdomains. Mol. Genet. Genom. 2014, 290, 847–862. [Google Scholar] [CrossRef] [PubMed]
- Tejada-Arranz, A.; Galtier, E.; El Mortaji, L.; Turlin, E.; Ershov, D.; De Reuse, H.; Charpentier, E. The RNase J-Based RNA Degradosome Is Compartmentalized in the Gastric Pathogen Helicobacter pylori. mBio 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Liu T-Y, Tsai S-H, Chen J-W, Wang Y-C, Hu S-T, Chen Y-Y. 2021. Mab_3083c Is a Homologue of RNase J and Plays a Role in Colony Morphotype, Aggregation, and Sliding Motility of Mycobacterium abscessus. Microorganisms 9:676.
- Aït-Bara, S.; Carpousis, A.J. RNA degradosomes in bacteria and chloroplasts: classification, distribution and evolution of RNase E homologs. Mol. Microbiol. 2015, 97, 1021–1135. [Google Scholar] [CrossRef] [PubMed]
- Maeda, T.; Wachi, M. Corynebacterium glutamicum RNase E/G-type endoribonuclease encoded by NCgl2281 is involved in the 5′ maturation of 5S rRNA. Arch. Microbiol. 2011, 194, 65–73. [Google Scholar] [CrossRef]
- Taverniti V, Forti F, Ghisotti D, Putzer H. 2011. Mycobacterium smegmatis RNase J is a 5’-3’ exo-/endoribonuclease and both RNase J and RNase E are involved in ribosomal RNA maturation. Molecular microbiology 82:1260–1276.
- Płociński, P.; Macios, M.; Houghton, J.; Niemiec, E.; Płocińska, R.; Brzostek, A.; Słomka, M.; Dziadek, J.; Young, D.; Dziembowski, A. Proteomic and transcriptomic experiments reveal an essential role of RNA degradosome complexes in shaping the transcriptome of Mycobacterium tuberculosis. Nucleic Acids Res. 2019, 47, 5892–5905. [Google Scholar] [CrossRef]
- Zhou Y, Sun H, Rapiejko AR, Vargas-Blanco DA, Martini MC, Chase MR, Joubran SR, Davis AB, Dainis JP, Kelly JM, Ioerger TR, Roberts LA, Fortune SM, Shell SS. 2023. Mycobacterial RNase E cleaves with a distinct sequence preference and controls the degradation rates of most Mycolicibacterium smegmatis mRNAs. Journal of Biological Chemistry 299:105312.
- Jaiswal, L.K.; Singh, R.K.; Nayak, T.; Kakkar, A.; Kandwal, G.; Singh, V.S.; Gupta, A. A comparative analysis of mycobacterial ribonucleases: Towards a therapeutic novel drug target. Infect. Genet. Evol. 2024, 123, 105645. [Google Scholar] [CrossRef]
- TEJADA-ARRANZ A, de CRECY-LAGARD V, DE REUSE H. 2020. Bacterial RNA degradosomes. Trends Biochem Sci 45:42–57.
- Martini, B.A.; Grigorov, A.S.; Skvortsova, Y.V.; Bychenko, O.S.; Salina, E.G.; Azhikina, T.L. Small RNA MTS1338 Configures a Stress Resistance Signature in Mycobacterium tuberculosis. Int. J. Mol. Sci. 2023, 24, 7928. [Google Scholar] [CrossRef]
- Lehnik-Habrink, M.; Pförtner, H.; Rempeters, L.; Pietack, N.; Herzberg, C.; Stülke, J. The RNA degradosome in Bacillus subtilis: identification of CshA as the major RNA helicase in the multiprotein complex. Mol. Microbiol. 2010, 77, 958–971. [Google Scholar] [CrossRef]
- Haq IU, Müller P, Brantl S. 2024. A comprehensive study of the interactions in the B. subtilis degradosome with special emphasis on the role of the small proteins SR1P and SR7P. Molecular microbiology 121:40–52.
- Commichau FM, Rothe FM, Herzberg C, Wagner E, Hellwig D, Lehnik-Habrink M, Hammer E, Völker U, Stülke J. 2009. Novel activities of glycolytic enzymes in Bacillus subtilis: interactions with essential proteins involved in mRNA processing. Molecular & cellular proteomics : MCP 8:1350–1360.
- Roux, C.M.; DeMuth, J.P.; Dunman, P.M. Characterization of Components of the Staphylococcus aureus mRNA Degradosome Holoenzyme-Like Complex. J. Bacteriol. 2011, 193, 5520–5526. [Google Scholar] [CrossRef]
- Even, S.; Pellegrini, O.; Zig, L.; Labas, V.; Vinh, J.; Bréchemmier-Baey, D.; Putzer, H. Ribonucleases J1 and J2: two novel endoribonucleases in B.subtilis with functional homology to E.coli RNase E. Nucleic Acids Res. 2005, 33, 2141–2152. [Google Scholar] [CrossRef] [PubMed]
- Bugrysheva, J.V.; Scott, J.R. The ribonucleases J1 and J2 are essential for growth and have independent roles in mRNA decay in Streptococcus pyogenes. Mol. Microbiol. 2010, 75, 731–743. [Google Scholar] [CrossRef] [PubMed]
- Callaghan AJ, Redko Y, Murphy LM, Grossmann JG, Yates D, Garman E, Ilag LL, Robinson CV, Symmons MartynF, McDowall KJ, Luisi BF. 2005. “Zn-Link”: A Metal-Sharing Interface that Organizes the Quaternary Structure and Catalytic Site of the Endoribonuclease, RNase E. Biochemistry 44:4667–4675.
- Mardle, C.E.; Shakespeare, T.J.; Butt, L.E.; Goddard, L.R.; Gowers, D.M.; Atkins, H.S.; Vincent, H.A.; Callaghan, A.J. A structural and biochemical comparison of Ribonuclease E homologues from pathogenic bacteria highlights species-specific properties. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Thompson, K.J.; Zong, J.; Mackie, G.A.; Gourse, R.L. Altering the Divalent Metal Ion Preference of RNase E. J. Bacteriol. 2014, 197, 477–482. [Google Scholar] [CrossRef]
- Shahbabian, K.; Jamalli, A.; Zig, L.; Putzer, H. RNase Y, a novel endoribonuclease, initiates riboswitch turnover in Bacillus subtilis. EMBO J. 2009, 28, 3523–3533. [Google Scholar] [CrossRef]
- Deana, A.; Belasco, J.G. The function of RNase G in Escherichia coli is constrained by its amino and carboxyl termini. Mol. Microbiol. 2004, 51, 1205–1217. [Google Scholar] [CrossRef]
- Garrey, S.M.; Blech, M.; Riffell, J.L.; Hankins, J.S.; Stickney, L.M.; Diver, M.; Hsu, Y.-H.R.; Kunanithy, V.; Mackie, G.A. Substrate Binding and Active Site Residues in RNases E and G. J. Biol. Chem. 2009, 284, 31843–31850. [Google Scholar] [CrossRef]
- Schein, A.; Sheffy-Levin, S.; Glaser, F.; Schuster, G. The RNase E/G-type endoribonuclease of higher plants is located in the chloroplast and cleaves RNA similarly to the E. coli enzyme. RNA 2008, 14, 1057–1068. [Google Scholar] [CrossRef]
- Raj, R.; Nadig, S.; Patel, T.; Gopal, B. Structural and biochemical characteristics of two Staphylococcus epidermidis RNase J paralogs RNase J1 and RNase J2. J. Biol. Chem. 2020, 295, 16863–16876. [Google Scholar] [CrossRef]
- Amblar, M.; Arraiano, C.M. A single mutation in Escherichia coli ribonuclease II inactivates the enzyme without affecting RNA binding. FEBS J. 2004, 272, 363–374. [Google Scholar] [CrossRef]
- A Steitz, T.; A Steitz, J. A general two-metal-ion mechanism for catalytic RNA. Proc. Natl. Acad. Sci. USA 1993, 90, 6498–6502. [Google Scholar] [CrossRef]
- Zuo, Y.; Vincent, H.A.; Zhang, J.; Wang, Y.; Deutscher, M.P.; Malhotra, A. Structural Basis for Processivity and Single-Strand Specificity of RNase II. Mol. Cell 2006, 24, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Vincent, H.A.; Deutscher, M.P. The Roles of Individual Domains of RNase R in Substrate Binding and Exoribonuclease Activity. J. Biol. Chem. 2009, 284, 486–494. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.-F.; Deutscher, M.P. An Important Role for RNase R in mRNA Decay. Mol. Cell 2005, 17, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Giuliano MG, Engl C. 2021. The lifecycle of ribosomal RNA in bacteria. RNA Damage and Repair 27–51.
- Apirion, D.; Gegenheimer, P. Processing of bacterial RNA. FEBS Lett. 1981, 125, 1–9. [Google Scholar] [CrossRef]
- Deutscher, MP. 2009. Maturation and degradation of ribosomal RNA in bacteria. Progress in molecular biology and translational science 85:369–391.
- Loughney, K.; Lund, E.; Dahlberg, J.E. Ribosomal RNA precursors ofBacillus subtilis. Nucleic Acids Res. 1983, 11, 6709–6721. [Google Scholar] [CrossRef]
- Li, Z.; Pandit, S.; Deutscher, M.P. RNase G (CafA protein) and RNase E are both required for the 5′ maturation of 16S ribosomal RNA. EMBO J. 1999, 18, 2878–2885. [Google Scholar] [CrossRef]
- Wachi M, Umitsuki G, Shimizu M, Takada A, Nagai K. 1999. Escherichia coli cafA Gene Encodes a Novel RNase, Designated as RNase G, Involved in Processing of the 5′ End of 16S rRNA. Biochemical and Biophysical Research Communications 259:483–488.
- Li, Z.; Deutscher, M.P. The tRNA processing enzyme RNase T is essential for maturation of 5S RNA. Proc. Natl. Acad. Sci. USA 1995, 92, 6883–6886. [Google Scholar] [CrossRef]
- Li, Z.; Pandit, S.; Deutscher, M.P. Maturation of 23S ribosomal RNA requires the exoribonuclease RNase T. RNA 1999, 5, 139–146. [Google Scholar] [CrossRef]
- Li, Z.; Deutscher, M.P.; Lovett, S.T. Exoribonucleases and Endoribonucleases. EcoSal Plus 2004, 1. [Google Scholar] [CrossRef]
- Martini, M.C.; Zhou, Y.; Sun, H.; Shell, S.S. Defining the Transcriptional and Post-transcriptional Landscapes of Mycobacterium smegmatis in Aerobic Growth and Hypoxia. Front. Microbiol. 2019, 10, 591. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, A.W. Ribonuclease III mechanisms of double-stranded RNA cleavage. Wiley Interdiscip. Rev. RNA 2013, 5, 31–48. [Google Scholar] [CrossRef] [PubMed]
- E Campbell, F.; Cassano, A.G.; E Anderson, V.; E Harris, M. Pre-steady-state and stopped-flow fluorescence analysis of Escherichia coli ribonuclease III: insights into mechanism and conformational changes associated with binding and catalysis. J. Mol. Biol. 2002, 317, 21–40. [Google Scholar] [CrossRef] [PubMed]
- Romano, M.; van de Weerd, R.; Brouwer, F.C.; Roviello, G.N.; Lacroix, R.; Sparrius, M.; Stempvoort, G.v.D.B.-V.; Maaskant, J.J.; van der Sar, A.M.; Appelmelk, B.J.; et al. Structure and Function of RNase AS, a Polyadenylate-Specific Exoribonuclease Affecting Mycobacterial Virulence In Vivo. Structure 2014, 22, 719–730. [Google Scholar] [CrossRef]
- Zuo Y, Zheng H, Wang Y, Chruszcz M, Cymborowski M, Skarina T, Savchenko A, Malhotra A, Minor W. 2007. Crystal Structure of RNase T, an exoribonuclease involved in tRNA maturation and end-turnover. Structure 15:417–428.
- Badhwar, P.; Khan, S.H.; Taneja, B. Three-dimensional structure of a mycobacterial oligoribonuclease reveals a unique C-terminal tail that stabilizes the homodimer. J. Biol. Chem. 2022, 298, 102595. [Google Scholar] [CrossRef]
- Mechold, U.; Fang, G.; Ngo, S.; Ogryzko, V.; Danchin, A. YtqI from Bacillus subtilis has both oligoribonuclease and pAp-phosphatase activity. Nucleic Acids Res. 2007, 35, 4552–4561. [Google Scholar] [CrossRef]
- Li, Z.; Deutscher, M.P. RNase E plays an essential role in the maturation of Escherichia coli tRNA precursors. RNA 2002, 8, 97–109. [Google Scholar] [CrossRef]
- Li, Z.; Deutscher, M.P. Maturation Pathways for E. coli tRNA Precursors: A Random Multienzyme Process In Vivo. Cell 1996, 86, 503–512. [Google Scholar] [CrossRef]
- Wen, T.; Oussenko, I.A.; Pellegrini, O.; Bechhofer, D.H.; Condon, C. Ribonuclease PH plays a major role in the exonucleolytic maturation of CCA-containing tRNA precursors in Bacillus subtilis. Nucleic Acids Res. 2005, 33, 3636–3643. [Google Scholar] [CrossRef]
- Harlow, L.S.; Kadziola, A.; Jensen, K.F.; Larsen, S. Crystal structure of the phosphorolytic exoribonuclease RNase PH from Bacillus subtilis and implications for its quaternary structure and tRNA binding. Protein Sci. 2004, 13, 668–677. [Google Scholar] [CrossRef]
- Beebe, J.A.; Kurz, J.C.; Fierke, C.A. Magnesium Ions Are Required by Bacillus subtilis Ribonuclease P RNA for both Binding and Cleaving Precursor tRNAAsp. Biochemistry 1996, 35, 10493–10505. [Google Scholar] [CrossRef]
- Kazantsev, A.V.; Krivenko, A.A.; Pace, N.R. Mapping metal-binding sites in the catalytic domain of bacterial RNase P RNA. RNA 2008, 15, 266–276. [Google Scholar] [CrossRef] [PubMed]
- Cudny, H.; Zaniewski, R.; Deutscher, M. Escherichia coli RNase D. Catalytic properties and substrate specificity. J. Biol. Chem. 1981, 256, 5633–5637. [Google Scholar] [CrossRef] [PubMed]
- Redko, Y.; de la Sierra-Gallay, I.L.; Condon, C. When all's zed and done: the structure and function of RNase Z in prokaryotes. Nat. Rev. Microbiol. 2007, 5, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Mohanty BK, Kushner SR. 2000. Polynucleotide phosphorylase functions both as a 3’ right-arrow 5’ exonuclease and a poly(A) polymerase in Escherichia coli. Proceedings of the National Academy of Sciences of the United States of America 97:11966–11971.
- Mohanty BK, Kushner SR. 2000. Polynucleotide phosphorylase, RNase II and RNase E play different roles in the in vivo modulation of polyadenylation in Escherichia coli. Mol Microbiol 36:982–994.
- Jarrige, A.-C.; Bréchemier-Baey, D.; Mathy, N.; Duché, O.; Portier, C. Mutational Analysis of Polynucleotide Phosphorylase from Escherichia coli. J. Mol. Biol. 2002, 321, 397–409. [Google Scholar] [CrossRef]
- Unciuleac, M.-C.; Shuman, S. Discrimination of RNA from DNA by Polynucleotide Phosphorylase. Biochemistry 2013, 52, 6702–6711. [Google Scholar] [CrossRef]
- Unciuleac, M.-C.; Shuman, S. Distinctive Effects of Domain Deletions on the Manganese-Dependent DNA Polymerase and DNA Phosphorylase Activities of Mycobacterium smegmatis Polynucleotide Phosphorylase. Biochemistry 2013, 52, 2967–2981. [Google Scholar] [CrossRef]
- Nurmohamed, S.; Vaidialingam, B.; Callaghan, A.J.; Luisi, B.F. Crystal Structure of Escherichia coli Polynucleotide Phosphorylase Core Bound to RNase E, RNA and Manganese: Implications for Catalytic Mechanism and RNA Degradosome Assembly. J. Mol. Biol. 2009, 389, 17–33. [Google Scholar] [CrossRef]
- Yang, Z.; Liu, W.; Zhao, L.; Yin, D.; Feng, J.; Li, L.; Guo, X. Single-exonuclease nanocircuits reveal the RNA degradation dynamics of PNPase and demonstrate potential for RNA sequencing. Nat. Commun. 2023, 14, 1–14. [Google Scholar] [CrossRef]
- Syson K, Tomlinson C, Chapados BR, Sayers JR, Tainer JA, Williams NH, Grasby JA. 2008. Three metal ions participate in the reaction catalyzed by T5 flap endonuclease. The Journal of biological chemistry 283:28741–28746.
- Ju, T.; Goldsmith, R.B.; Chai, S.C.; Maroney, M.J.; Pochapsky, S.S.; Pochapsky, T.C. One Protein, Two Enzymes Revisited: A Structural Entropy Switch Interconverts the Two Isoforms of Acireductone Dioxygenase. J. Mol. Biol. 2006, 363, 823–834. [Google Scholar] [CrossRef]
- Chai, S.C.; Ju, T.; Dang, M.; Goldsmith, R.B.; Maroney, M.J.; Pochapsky, T.C. Characterization of Metal Binding in the Active Sites of Acireductone Dioxygenase Isoforms from Klebsiella ATCC 8724. Biochemistry 2008, 47, 2428–2438. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, A.R.; Pochapsky, T.C.; Petsko, G.A.; Ringe, D. Dual chemistry catalyzed by human acireductone dioxygenase. Protein Eng. Des. Sel. 2017, 30, 109–206. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, A.R.; Pochapsky, T.C.; Ringe, D. The Metal Drives the Chemistry: Dual Functions of Acireductone Dioxygenase. Chem. Rev. 2017, 117, 10474–10501. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, A.R.; Wagenpfeil, K.; Pochapsky, T.C.; Petsko, G.A.; Ringe, D. Metal-Dependent Function of a Mammalian Acireductone Dioxygenase. Biochemistry 2016, 55, 1398–1407. [Google Scholar] [CrossRef]
- Liu, X.; Garber, A.; Ryan, J.; Deshpande, A.; Ringe, D.; Pochapsky, T.C. A Model for the Solution Structure of Human Fe(II)-Bound Acireductone Dioxygenase and Interactions with the Regulatory Domain of Matrix Metalloproteinase I (MMP-I). Biochemistry 2020, 59, 4238–4249. [Google Scholar] [CrossRef]
- Liu, X.; Pochapsky, T.C. Human Acireductone Dioxygenase (HsARD), Cancer and Human Health: Black Hat, White Hat or Gray? Inorganics 2019, 7, 101. [Google Scholar] [CrossRef]
- Miłaczewska, A.; Kot, E.; Amaya, J.A.; Makris, T.M.; Zając, M.; Korecki, J.; Chumakov, A.; Trzewik, B.; Kędracka-Krok, S.; Minor, W.; et al. On the Structure and Reaction Mechanism of Human Acireductone Dioxygenase. Chem. – A Eur. J. 2017, 24, 5225–5237. [Google Scholar] [CrossRef]
- Beljanksi, M. 1995. De novo synthesis of DNA-like molecules by polynucleotide phosphorylase in vitro. Journal of Molecular Evolution 493–499.
- Cardenas, P.P.; Carzaniga, T.; Zangrossi, S.; Briani, F.; Garcia-Tirado, E.; Dehò, G.; Alonso, J.C. Polynucleotide phosphorylase exonuclease and polymerase activities on single-stranded DNA ends are modulated by RecN, SsbA and RecA proteins. Nucleic Acids Res. 2011, 39, 9250–9261. [Google Scholar] [CrossRef]
- Arraiano, C.M.; Andrade, J.M.; Domingues, S.; Guinote, I.B.; Malecki, M.; Matos, R.G.; Moreira, R.N.; Pobre, V.; Reis, F.P.; Saramago, M.; et al. The critical role of RNA processing and degradation in the control of gene expression. FEMS Microbiol. Rev. 2010, 34, 883–923. [Google Scholar] [CrossRef]
- Sarkar, D.; Fisher, P.B. Polynucleotide phosphorylase: An evolutionary conserved gene with an expanding repertoire of functions. Pharmacol. Ther. 2006, 112, 243–263. [Google Scholar] [CrossRef] [PubMed]
- Venkateswara Rao P, Holm RH. 2004. Synthetic Analogues of the Active Sites of Iron−Sulfur Proteins. Chem Rev 104:527–560.
- Vallières, C.; Benoit, O.; Guittet, O.; Huang, M.-E.; Lepoivre, M.; Golinelli-Cohen, M.-P.; Vernis, L. Iron-sulfur protein odyssey: exploring their cluster functional versatility and challenging identification. Metallomics 2024, 16. [Google Scholar] [CrossRef] [PubMed]
- Yao W, Gurubasavaraj PM, Holland PL. 2014. All-Ferrous Iron–Sulfur Clusters, p. 1–37. In Rabinovich, D (ed.), Molecular Design in Inorganic Biochemistry. Springer, Berlin, Heidelberg.
- Beinert H, Holm RH, Münck E. 1997. Iron-sulfur clusters: nature’s modular, multipurpose structures. Science 277:653–659.
- Lee, W.; Brune, D.; Lobrutto, R.; Blankenship, R. Isolation, Characterization, and Primary Structure of Rubredoxin from the Photosynthetic Bacterium, Heliobacillus mobilis. Arch. Biochem. Biophys. 1995, 318, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, C.L.; Shaw, L. A Comprehensive Phylogenetic Analysis of Rieske and Rieske-Type Iron-Sulfur Proteins. J. Bioenerg. Biomembr. 2001, 33, 9–26. [Google Scholar] [CrossRef]
- Dailey, T.A.; Dailey, H.A. Identification of [2Fe-2S] Clusters in Microbial Ferrochelatases. J. Bacteriol. 2002, 184, 2460–2464. [Google Scholar] [CrossRef]
- Kakuta Y, Horio T, Takahashi Y, Fukuyama K. 2001. Crystal Structure of Escherichia coli Fdx, an Adrenodoxin-Type Ferredoxin Involved in the Assembly of Iron−Sulfur Clusters. Biochemistry 40:11007–11012.
- Escalettes, F.; Florentin, D.; Bui, B.T.S.; Lesage, D.; Marquet, A. Biotin Synthase Mechanism: Evidence for Hydrogen Transfer from the Substrate into Deoxyadenosine. J. Am. Chem. Soc. 1999, 121, 3571–3578. [Google Scholar] [CrossRef]
- Lachowicz, J.C.; Lennox-Hvenekilde, D.; Myling-Petersen, N.; Salomonsen, B.; Verkleij, G.; Acevedo-Rocha, C.G.; Caddell, B.; Gronenberg, L.S.; Almo, S.C.; Sommer, M.O.A.; et al. Discovery of a Biotin Synthase That Utilizes an Auxiliary 4Fe–5S Cluster for Sulfur Insertion. J. Am. Chem. Soc. 2024, 146, 1860–1873. [Google Scholar] [CrossRef]
- Adams, M.W.; Eccleston, E.; Howard, J.B. Iron-sulfur clusters of hydrogenase I and hydrogenase II of Clostridium pasteurianum. Proc. Natl. Acad. Sci. USA 1989, 86, 4932–4936. [Google Scholar] [CrossRef]
- Beinert H, Kennedy MC, Stout CD. 1996. Aconitase as Iron−Sulfur Protein, Enzyme, and Iron-Regulatory Protein. Chem Rev 96:2335–2374.
- Smith, A.T.; Linkous, R.O.; Max, N.J.; Sestok, A.E.; Szalai, V.A.; Chacón, K.N. The FeoC [4Fe–4S] Cluster Is Redox-Active and Rapidly Oxygen-Sensitive. Biochemistry 2019, 58, 4935–4949. [Google Scholar] [CrossRef]
- Keable, S.M.; Zadvornyy, O.A.; Johnson, L.E.; Ginovska, B.; Rasmussen, A.J.; Danyal, K.; Eilers, B.J.; Prussia, G.A.; LeVan, A.X.; Raugei, S.; et al. Structural characterization of the P1+ intermediate state of the P-cluster of nitrogenase. J. Biol. Chem. 2018, 293, 9629–9635. [Google Scholar] [CrossRef]
- Rupnik K, Hu Y, Lee CC, Wiig JA, Ribbe MW, Hales BJ. 2012. P+ State of Nitrogenase P-Cluster Exhibits Electronic Structure of a [Fe4S4]+ Cluster. J Am Chem Soc 134:13749–13754.
- Osterloh, F.; Sanakis, Y.; Staples, R.J.; Münck, E.; Holm, R.H. A Molybdenum-Iron-Sulfur Cluster Containing Structural Elements Relevant to the P-Cluster of Nitrogenase. Angew. Chem. Int. Ed. Engl. 1999, 38, 2066–2070. [Google Scholar] [CrossRef]
- Lu, Y. Assembly and Transfer of Iron–Sulfur Clusters in the Plastid. Front. Plant Sci. 2018, 9, 336. [Google Scholar] [CrossRef] [PubMed]
- Frazzon, J.; Dean, D.R. Formation of iron–sulfur clusters in bacteria: an emerging field in bioinorganic chemistry. Curr. Opin. Chem. Biol. 2003, 7, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Ohki Y, Tanifuji K, Yamada N, Cramer RE, Tatsumi K. 2012. Formation of a nitrogenase P-cluster [Fe8S7] core via reductive fusion of two all-ferric [Fe4S4] clusters. Chem Asian J 7:2222–2224.
- Fontecave, M.; Ollagnier-De-Choudens, S. Iron–sulfur cluster biosynthesis in bacteria: Mechanisms of cluster assembly and transfer. Arch. Biochem. Biophys. 2008, 474, 226–237. [Google Scholar] [CrossRef]
- Roche B, Aussel L, Ezraty B, Mandin P, Py B, Barras F. 2013. Iron/sulfur proteins biogenesis in prokaryotes: Formation, regulation and diversity. Biochimica et Biophysica Acta (BBA) - Bioenergetics 1827:455–469.
- Tripathi, A.; Anand, K.; Das, M.; O’nIel, R.A.; S, S.P.; Thakur, C.; L. , R.R.R.; Rajmani, R.S.; Chandra, N.; Laxman, S.; et al. Mycobacterium tuberculosis requires SufT for Fe-S cluster maturation, metabolism, and survival in vivo. PLOS Pathog. 2022, 18, e1010475. [Google Scholar] [CrossRef]
- Hidese, R.; Mihara, H.; Kurihara, T.; Esaki, N. Global Identification of Genes Affecting Iron-Sulfur Cluster Biogenesis and Iron Homeostasis. J. Bacteriol. 2014, 196, 1238–1249. [Google Scholar] [CrossRef]
- Kim J, Rees DC. 1992. Crystallographic structure and functional implications of the nitrogenase molybdenum-iron protein from azotobacter vinelandii. Nature 360:553–560.
- White, M.F.; Dillingham, M.S. Iron–sulphur clusters in nucleic acid processing enzymes. Curr. Opin. Struct. Biol. 2012, 22, 94–100. [Google Scholar] [CrossRef]
- Klinge, S.; Hirst, J.; Maman, J.D.; Krude, T.; Pellegrini, L. An iron-sulfur domain of the eukaryotic primase is essential for RNA primer synthesis. Nat. Struct. Mol. Biol. 2007, 14, 875–877. [Google Scholar] [CrossRef]
- Oussenko IA, Abe T, Ujiie H, Muto A, Bechhofer DH. 2005. Participation of 3’-to-5’ exoribonucleases in the turnover of Bacillus subtilis mRNA. J Bacteriol 187:2758–2767.
- Gimpel M, Brantl S. 2016. Dual-function sRNA encoded peptide SR1P modulates moonlighting activity of B. subtilis GapA. RNA Biol 13:916–926.
- Ul Haq I, Brantl S. 2021. Moonlighting in Bacillus subtilis: The Small Proteins SR1P and SR7P Regulate the Moonlighting Activity of Glyceraldehyde 3-Phosphate Dehydrogenase A (GapA) and Enolase in RNA Degradation. Microorganisms 9:1046.
- Dubnau, E.; DeSantis, M.; Dubnau, D.; Babitzke, P. Formation of a stable RNase Y-RicT (YaaT) complex requires RicA (YmcA) and RicF (YlbF). mBio 2023, 14, e0126923. [Google Scholar] [CrossRef]
- Tanner AW, Carabetta VJ, Martinie RJ, Mashruwala AA, Boyd JM, Krebs C, Dubnau D. 2017. The RicAFT (YmcA-YlbF-YaaT) complex carries two [4Fe-4S]2+ clusters and may respond to redox changes. Molecular Microbiology 104:837–850.
- DeLoughery, A.; Lalanne, J.-B.; Losick, R.; Li, G.-W. Maturation of polycistronic mRNAs by the endoribonuclease RNase Y and its associated Y-complex in Bacillus subtilis. Proc. Natl. Acad. Sci. USA 2018, 115, 201803283–E5594. [Google Scholar] [CrossRef]
- Adusei-Danso F, Khaja FT, Desantis M, Jeffrey PD, Dubnau E, Demeler B, Neiditch MB, Dubnau D. 2019. [CrossRef]
- Tortosa P, Albano M, Dubnau D. 2000. Characterization of ylbF, a new gene involved in competence development and sporulation in Bacillus subtilis. Molecular Microbiology 35:1110–1119.
- Hosoya S, Asai K, Ogasawara N, Takeuchi M, Sato T. 2002. Mutation in yaaT leads to significant inhibition of phosphorelay during sporulation in Bacillus subtilis. Journal of Bacteriology 184:5545–5553.
- Branda, S.S.; González-Pastor, J.E.; Dervyn, E.; Ehrlich, S.D.; Losick, R.; Kolter, R. Genes Involved in Formation of Structured Multicellular Communities by Bacillus subtilis. J. Bacteriol. 2004, 186, 3970–3979. [Google Scholar] [CrossRef] [PubMed]
- Guth-Metzler, R.; Bray, M.S.; Frenkel-Pinter, M.; Suttapitugsakul, S.; Montllor-Albalate, C.; Bowman, J.C.; Wu, R.; Reddi, A.R.; Okafor, C.D.; Glass, J.B.; et al. Cutting in-line with iron: ribosomal function and non-oxidative RNA cleavage. Nucleic Acids Res. 2020, 48, 8663–8674. [Google Scholar] [CrossRef] [PubMed]
- Zinskie, J.A.; Ghosh, A.; Trainor, B.M.; Shedlovskiy, D.; Pestov, D.G.; Shcherbik, N. Iron-dependent cleavage of ribosomal RNA during oxidative stress in the yeast Saccharomyces cerevisiae. J. Biol. Chem. 2018, 293, 14237–14248. [Google Scholar] [CrossRef]
- Smethurst, D.G.; Kovalev, N.; McKenzie, E.R.; Pestov, D.G.; Shcherbik, N. Iron-mediated degradation of ribosomes under oxidative stress is attenuated by manganese. J. Biol. Chem. 2020, 295, 17200–17214. [Google Scholar] [CrossRef] [PubMed]
- Anjem, A.; Imlay, J.A. Mononuclear Iron Enzymes Are Primary Targets of Hydrogen Peroxide Stress. J. Biol. Chem. 2012, 287, 15544–15556. [Google Scholar] [CrossRef]
- Sobota, J.M.; Imlay, J.A. Iron enzyme ribulose-5-phosphate 3-epimerase in Escherichia coli is rapidly damaged by hydrogen peroxide but can be protected by manganese. Proc. Natl. Acad. Sci. USA 2011, 108, 5402–5407. [Google Scholar] [CrossRef]
- Vasilyev N, Gao A, Serganov A. 2019. Noncanonical features and modifications on the 5′-end of bacterial sRNAs and mRNAs. WIREs RNA 10:e1509.
- Hudeček, O.; Benoni, R.; Reyes-Gutierrez, P.E.; Culka, M.; Šanderová, H.; Hubálek, M.; Rulíšek, L.; Cvačka, J.; Krásný, L.; Cahová, H. Dinucleoside polyphosphates act as 5′-RNA caps in bacteria. Nat. Commun. 2020, 11, 1–11. [Google Scholar] [CrossRef]
- Régnier P, Marujo PE. 2013. Polyadenylation and Degradation of RNA in ProkaryotesMadame Curie Bioscience Database [Internet]. Landes Bioscience.
- Briani, F.; Carzaniga, T.; Dehò, G. Regulation and functions of bacterial PNPase. Wiley Interdiscip. Rev. RNA 2016, 7, 241–258. [Google Scholar] [CrossRef]
- Deutscher, M.P. Degradation of RNA in bacteria: comparison of mRNA and stable RNA. Nucleic Acids Res. 2006, 34, 659–666. [Google Scholar] [CrossRef]
- Giménez, J.A.M.; Sáez, G.T.; Seisdedos, R.T. On the Function of Modified Nucleosides in the RNA World. J. Theor. Biol. 1998, 194, 485–490. [Google Scholar] [CrossRef]
- Kimura S, Suzuki T. 2015. Iron–sulfur proteins responsible for RNA modifications. Biochimica et Biophysica Acta (BBA) - Molecular Cell Research 1853:1272–1283.
- Khodour Y, Kaguni LS, Stiban J. 2019. Iron–sulfur clusters in nucleic acid metabolism: Varying roles of ancient cofactors, p. 225–256. In Enzymes. Academic Press.
- Sofia HJ, Chen G, Hetzler BG, Reyes-Spindola JF, Miller NE. 2001. Radical SAM, a novel protein superfamily linking unresolved steps in familiar biosynthetic pathways with radical mechanisms: functional characterization using new analysis and information visualization methods. Nucleic acids research 29:1097–1106.
- Broderick JB, Duffus BR, Duschene KS, Shepard EM. 2014. Radical S-adenosylmethionine enzymes. Chemical Reviews 114:4229–4317.
- Fyfe, C.D.; Bernardo-García, N.; Fradale, L.; Grimaldi, S.; Guillot, A.; Brewee, C.; Chavas, L.M.G.; Legrand, P.; Benjdia, A.; Berteau, O. Crystallographic snapshots of a B12-dependent radical SAM methyltransferase. Nature 2022, 602, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Bauerle, M.R.; Schwalm, E.L.; Booker, S.J. Mechanistic Diversity of Radical S-Adenosylmethionine (SAM)-dependent Methylation. J. Biol. Chem. 2015, 290, 3995–4002. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.C.; Santi, D.V. On the mechanism and inhibition of DNA cytosine methyltransferases. Prog. Clin. Biol. Research. 1985, 198, 119–29. [Google Scholar]
- Kealey, J.; Gu, X.; Santi, D. Enzymatic mechanism of tRNA (m5U54)methyltransferase. Biochimie 1994, 76, 1133–1142. [Google Scholar] [CrossRef]
- Lee, K.-H.; Saleh, L.; Anton, B.P.; Madinger, C.L.; Benner, J.S.; Iwig, D.F.; Roberts, R.J.; Krebs, C.; Booker, S.J. Characterization of RimO, a New Member of the Methylthiotransferase Subclass of the Radical SAM Superfamily. Biochemistry 2009, 48, 10162–10174. [Google Scholar] [CrossRef]
- Hernández HL, Pierrel F, Elleingand E, García-Serres R, Huynh BH, Johnson MK, Fontecave M, Atta M. 2007. MiaB, a bifunctional radical-S-adenosylmethionine enzyme involved in the thiolation and methylation of tRNA, contains two essential [4Fe-4S] clusters. Biochemistry 46:5140–5147.
- Arragain, S.; Handelman, S.K.; Forouhar, F.; Wei, F.-Y.; Tomizawa, K.; Hunt, J.F.; Douki, T.; Fontecave, M.; Mulliez, E.; Atta, M. Identification of Eukaryotic and Prokaryotic Methylthiotransferase for Biosynthesis of 2-Methylthio-N6-threonylcarbamoyladenosine in tRNA. J. Biol. Chem. 2010, 285, 28425–28433. [Google Scholar] [CrossRef]
- Arragain, S.; Garcia-Serres, R.; Blondin, G.; Douki, T.; Clemancey, M.; Latour, J.-M.; Forouhar, F.; Neely, H.; Montelione, G.T.; Hunt, J.F.; et al. Post-translational Modification of Ribosomal Proteins: Structural and Functional Characterization of Rimo from Thermotoga Maritima, A Radical S-Adenosylmethionine Methylthiotransferase. J. Biol. Chem. 2010, 285, 5792–5801. [Google Scholar] [CrossRef]
- Wang, J.; Woldring, R.P.; Román-Meléndez, G.D.; McClain, A.M.; Alzua, B.R.; Marsh, E.N.G. Recent Advances in Radical SAM Enzymology: New Structures and Mechanisms. ACS Chem. Biol. 2014, 9, 1929–1938. [Google Scholar] [CrossRef]
- Sergiev PV, Golovina AY, Prokhorova IV, Sergeeva OV, Osterman IA, Nesterchuk MV, Burakovsky DE, Bogdanov AA, Dontsova OA. 2011. Modifications of ribosomal RNA: From enzymes to function, p. 97–110. In Ribosomes. Springer, Vienna.
- Polikanov, Y.S.; Melnikov, S.V.; Söll, D.; Steitz, T.A. Structural insights into the role of rRNA modifications in protein synthesis and ribosome assembly. Nat. Struct. Mol. Biol. 2015, 22, 342–344. [Google Scholar] [CrossRef]
- Oldenburg, M.; Krüger, A.; Ferstl, R.; Kaufmann, A.; Nees, G.; Sigmund, A.; Bathke, B.; Lauterbach, H.; Suter, M.; Dreher, S.; et al. TLR13 Recognizes Bacterial 23 S rRNA Devoid of Erythromycin Resistance–Forming Modification. Science 2012, 337, 1111–1115. [Google Scholar] [CrossRef]
- Kimura, S.; Sakai, Y.; Ishiguro, K.; Suzuki, T. Biogenesis and iron-dependency of ribosomal RNA hydroxylation. Nucleic Acids Res. 2017, 45, 12974–12986. [Google Scholar] [CrossRef]
- Fasnacht, M.; Gallo, S.; Sharma, P.; Himmelstoß, M.; A Limbach, P.; Willi, J.; Polacek, N. Dynamic 23S rRNA modification ho5C2501 benefits Escherichia coli under oxidative stress. Nucleic Acids Res. 2021, 50, 473–489. [Google Scholar] [CrossRef]
- Agarwalla, S.; Kealey, J.T.; Santi, D.V.; Stroud, R.M. Characterization of the 23 S Ribosomal RNA m5U1939 Methyltransferase from Escherichia coli. J. Biol. Chem. 2002, 277, 8835–8840. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.T.; Agarwalla, S.; Stroud, R.M. Crystal Structure of RumA, an Iron-Sulfur Cluster Containing E. coli Ribosomal RNA 5-Methyluridine Methyltransferase. Structure 2004, 12, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Pritts, J.; Zilinskas, E. 2015. Study of methyletransferase RumB in E. coli. FASEB J. 2015, 29, 711–712. [Google Scholar] [CrossRef]
- Agarwalla, S.; Stroud, R.M.; Gaffney, B.J. Redox Reactions of the Iron-Sulfur Cluster in a Ribosomal RNA Methyltransferase, RumA. J. Biol. Chem. 2004, 279, 34123–34129. [Google Scholar] [CrossRef]
- Agris, PF. The importance of being modified: roles of modified nucleosides and Mg2+ in RNA structure and function. Prog. Nucleic Acid Res. Mol. Biol. 1996, 53, 79–129. [Google Scholar]
- Urbonavičius, J.; Qian, Q.; Durand, J.M.; Hagervall, T.G.; Björk, G.R. Improvement of reading frame maintenance is a common function for several tRNA modifications. EMBO J. 2001, 20, 4863–4873. [Google Scholar] [CrossRef]
- Leipuviene, R.; Qian, Q.; BjörK, G.R. Formation of Thiolated Nucleosides Present in tRNA from Salmonella enterica serovar Typhimurium Occurs in Two Principally Distinct Pathways. J. Bacteriol. 2004, 186, 758–766. [Google Scholar] [CrossRef]
- Shigi, N. Biosynthesis and functions of sulfur modifications in tRNA. Front. Genet. 2014, 5, 67. [Google Scholar] [CrossRef]
- avužić M, Liu Y. 2017. Biosynthesis of sulfur-containing tRNA modifications: a comparison of bacterial, archaeal, and eukaryotic Pathways. Biomolecules 7.
- Bandarian, V. Radical SAM enzymes involved in the biosynthesis of purine-based natural products. Biochim. et Biophys. Acta (BBA) - Proteins Proteom. 2012, 1824, 1245–1253. [Google Scholar] [CrossRef]
- Shigi, N.; Horitani, M.; Miyauchi, K.; Suzuki, T.; Kuroki, M. An ancient type of MnmA protein is an iron–sulfur cluster-dependent sulfurtransferase for tRNA anticodons. RNA 2019, 26, 240–250. [Google Scholar] [CrossRef]
- Pierrel, F.; Douki, T.; Fontecave, M.; Atta, M. MiaB Protein Is a Bifunctional Radical-S-Adenosylmethionine Enzyme Involved in Thiolation and Methylation of tRNA. J. Biol. Chem. 2004, 279, 47555–47563. [Google Scholar] [CrossRef]
- Esberg B, Leung HCE, Tsui HCT, Bjork GR, Winkler ME. 1999. Identification of the miaB gene, involved in methylthiolation of isopentenylated A37 derivatives in the tRNA of Salmonella typhimurium and Escherichia coli. Journal of bacteriology 181:7256–7265.
- Li, Q.; Zallot, R.; MacTavish, B.S.; Montoya, A.; Payan, D.J.; Hu, Y.; Gerlt, J.A.; Angerhofer, A.; de Crécy-Lagard, V.; Bruner, S.D. Epoxyqueuosine Reductase QueH in the Biosynthetic Pathway to tRNA Queuosine Is a Unique Metalloenzyme. Biochemistry 2021, 60, 3152–3161. [Google Scholar] [CrossRef]
- Liu Y, Vinyard DJ, Reesbeck ME, Suzuki T, Manakongtreecheep K, Holland PL, Brudvig GW, Söll D. 2016. A [3Fe-4S] cluster is required for tRNA thiolation in archaea and eukaryotes. Proceedings of the National Academy of Sciences of the United States of America 113:12703–12708.
- Miles, Z.D.; Myers, W.K.; Kincannon, W.M.; Britt, R.D.; Bandarian, V. Biochemical and Spectroscopic Studies of Epoxyqueuosine Reductase: A Novel Iron–Sulfur Cluster- and Cobalamin-Containing Protein Involved in the Biosynthesis of Queuosine. Biochemistry 2015, 54, 4927–4935. [Google Scholar] [CrossRef]
- McLennan, B.; Buck, M.; Humphreys, J.; Griffiths, E. Iron-related modification of bacterial transfer RNA. Nucleic Acids Res. 1981, 9, 2629–2640. [Google Scholar] [CrossRef]
- Griffiths E, Humphreys J. 1978. Alterations in tRNAs containing 2-methylthio-N6-(Δ2-isopentenyl)-adenosine during growth of enteropathogenic Escherichia coli in the presence of iron-binding proteins. European Journal of Biochemistry 82:503–513.
- Urbonavičius, J.; Jäger, G.; Björk, G.R. Amino acid residues of the Escherichia coli tRNA(m5U54)methyltransferase (TrmA) critical for stability, covalent binding of tRNA and enzymatic activity. Nucleic Acids Res. 2007, 35, 3297–3305. [Google Scholar] [CrossRef] [PubMed]
- Mettert EL, Kiley PJ. 2015. Fe–S proteins that regulate gene expression. Biochimica et Biophysica Acta (BBA) - Molecular Cell Research 1853:1284–1293.
- Crack, J.C.; Green, J.; Thomson, A.J.; Le Brun, N.E. Iron–Sulfur Clusters as Biological Sensors: The Chemistry of Reactions with Molecular Oxygen and Nitric Oxide. Accounts Chem. Res. 2014, 47, 3196–3205. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, E.; Bollinger, J.M.; Bradley, T.M.; Walsh, C.T.; Demple, B. Binuclear [2Fe-2S] Clusters in the Escherichia coli SoxR Protein and Role of the Metal Centers in Transcription. J. Biol. Chem. 1995, 270, 20908–20914. [Google Scholar] [CrossRef] [PubMed]
- Gaudu P, Weiss B. 1996. SoxR, a [2Fe-2S] transcription factor, is active only in its oxidized form. Proceedings of the National Academy of Sciences 93:10094–10098.
- Kobayashi, K.; Fujikawa, M.; Kozawa, T. Oxidative stress sensing by the iron–sulfur cluster in the transcription factor, SoxR. J. Inorg. Biochem. 2014, 133, 87–91. [Google Scholar] [CrossRef]
- Guiza Beltran D, Wan T, Zhang L. 2024. WhiB-like proteins: Diversity of structure, function and mechanism. Biochimica et Biophysica Acta (BBA) - Molecular Cell Research 1871:119787.
- Saini, V.; Farhana, A.; Steyn, A.J. Mycobacterium tuberculosisWhiB3: A Novel Iron–Sulfur Cluster Protein That Regulates Redox Homeostasis and Virulence. Antioxidants Redox Signal. 2012, 16, 687–697. [Google Scholar] [CrossRef]
- Singh, A.; Crossman, D.K.; Mai, D.; Guidry, L.; Voskuil, M.I.; Renfrow, M.B.; Steyn, A.J.C.; Bishai, W. Mycobacterium tuberculosis WhiB3 Maintains Redox Homeostasis by Regulating Virulence Lipid Anabolism to Modulate Macrophage Response. PLOS Pathog. 2009, 5, e1000545. [Google Scholar] [CrossRef]
- Luisi, B.; Dendooven, T.; Sinha, D.; Roeselova, A.; Cameron, T.; De Lay, N.; Bandyra, K. A cooperative PNPase-Hfq-RNA carrier complex facilitates bacterial riboregulation. Mol. Cell 2021, 81, 2901–2913. [Google Scholar] [CrossRef]
- Lay ND, Gottesman S. 2011. Role of polynucleotide phosphorylase in sRNA function in Escherichia coli. RNA (New York, NY) 17:1172–1189.
- Menendez-Gil, P.; Catalan-Moreno, A.; Caballero, C.J.; Toledo-Arana, A. Staphylococcus aureus ftnA 3’-Untranslated Region Modulates Ferritin Production Facilitating Growth Under Iron Starvation Conditions. Front. Microbiol. 2022, 13, 838042. [Google Scholar] [CrossRef] [PubMed]
- Beinert H, Kennedy MC. 1993. Aconitase, a two-faced protein: enzyme and iron regulatory factor. FASEB journal : official publication of the Federation of American Societies for Experimental Biology 7:1442–1449.
- Walden, WE. 2002. From bacteria to mitochondria: Aconitase yields surprises. Proceedings of the National Academy of Sciences of the United States of America 99:4138.
- Beinert, H.; Kennedy, M.C.; Stout, C.D. Aconitase as iron-sulfur protein, enzyme, and iron-regulatory protein. Chem. Rev. 2010, 96, 2335–2373. [Google Scholar] [CrossRef] [PubMed]
- Kang DK, Jeong J, Drake SK, Wehr NB, Rouault TA, Levine RL. 2003. Iron regulatory protein 2 as iron sensor: Iron-dependent oxidative modification of cysteine. The Journal of biological chemistry 278:14857–14864.
- Beinert H, Holm RH, Münck E. 1997. Iron-sulfur clusters: Nature’s modular, multipurpose structures. Science (New York, NY) 277:653–659.
- Dupuy, J.; Volbeda, A.; Carpentier, P.; Darnault, C.; Moulis, J.-M.; Fontecilla-Camps, J.C. Crystal Structure of Human Iron Regulatory Protein 1 as Cytosolic Aconitase. Structure 2006, 14, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Hentze, M.W.; Kühn, L.C. Molecular control of vertebrate iron metabolism: mRNA-based regulatory circuits operated by iron, nitric oxide, and oxidative stress. Proc. Natl. Acad. Sci. USA 1996, 93, 8175–8182. [Google Scholar] [CrossRef]
- Garza, K.R.; Clarke, S.L.; Ho, Y.-H.; Bruss, M.D.; Vasanthakumar, A.; Anderson, S.A.; Eisenstein, R.S. Differential translational control of 5′ IRE-containing mRNA in response to dietary iron deficiency and acute iron overload. Metallomics 2020, 12, 2186–2198. [Google Scholar] [CrossRef]
- Shen, M.; Goforth, J.B.; Eisenstein, R.S. Iron-dependent post transcriptional control of mitochondrial aconitase expression. Metallomics 2023, 15. [Google Scholar] [CrossRef]
- Casey, J.L.; Di Jeso, B.; Rao, K.; Klausner, R.D.; Harford, J.B. Two genetic loci participate in the regulation by iron of the gene for the human transferrin receptor. Proc. Natl. Acad. Sci. USA 1988, 85, 1787–1791. [Google Scholar] [CrossRef]
- Sanchez, M.; Galy, B.; Schwanhaeusser, B.; Blake, J.; Bähr-Ivacevic, T.; Benes, V.; Selbach, M.; Muckenthaler, M.U.; Hentze, M.W. Iron regulatory protein-1 and -2: transcriptome-wide definition of binding mRNAs and shaping of the cellular proteome by iron regulatory proteins. Blood 2011, 118, e168–e179. [Google Scholar] [CrossRef] [PubMed]
- Michta E, Schad K, Blin K, Ort-Winklbauer R, Röttig M, Kohlbacher O, Wohlleben W, Schinko E, Mast Y. 2012. The bifunctional role of aconitase in Streptomyces viridochromogenes Tü494. Environmental Microbiology 14:3203–3219.
- Jordan PA, Tang Y, Bradbury AJ, Thomson AJ, Guest JR. 1999. Biochemical and spectroscopic characterization of Escherichia coli aconitases (AcnA and AcnB). Biochemical Journal 344:739.
- Banerjee, S.; Nandyala, A.K.; Raviprasad, P.; Ahmed, N.; Hasnain, S.E. Iron-Dependent RNA-Binding Activity of Mycobacterium tuberculosis Aconitase. J. Bacteriol. 2007, 189, 4046–4052. [Google Scholar] [CrossRef] [PubMed]
- Haile, D.J.; A Rouault, T.; Tang, C.K.; Chin, J.; Harford, J.B.; Klausner, R.D. Reciprocal control of RNA-binding and aconitase activity in the regulation of the iron-responsive element binding protein: role of the iron-sulfur cluster. Proc. Natl. Acad. Sci. USA 1992, 89, 7536–7540. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Guest, J.R. Direct evidence for mRNA binding and post-transcriptional regulation by Escherichia coli aconitases. Microbiology 1999, 145, 3069–3079. [Google Scholar] [CrossRef]
- Gold B, Rodriguez GM, Marras SA, Pentecost M, Smith I. 2001. The Mycobacterium tuberculosis IdeR is a dual functional regulator that controls transcription of genes involved in iron acquisition, iron storage and survival in macrophages. Mol Microbiol 42:851–865.
- Shell, S.S.; Wang, J.; Lapierre, P.; Mir, M.; Chase, M.R.; Pyle, M.M.; Gawande, R.; Ahmad, R.; Sarracino, D.A.; Ioerger, T.R.; et al. Leaderless Transcripts and Small Proteins Are Common Features of the Mycobacterial Translational Landscape. PLOS Genet. 2015, 11, e1005641. [Google Scholar] [CrossRef]
- Brazzolotto, X.; Gaillard, J.; Pantopoulos, K.; Hentze, M.W.; Moulis, J.-M. Human Cytoplasmic Aconitase (Iron Regulatory Protein 1) Is Converted into Its [3Fe-4S] Form by Hydrogen Peroxide in Vitro but Is Not Activated for Iron-responsive Element Binding. J. Biol. Chem. 1999, 274, 21625–21630. [Google Scholar] [CrossRef]
- Jordan PA, Tang Y, Bradbury AJ, Thomson AJ, Guest JR. 1999. Biochemical and spectroscopic characterization of Escherichia coli aconitases (AcnA and AcnB).
- Kennedy, M.C.; Emptage, M.H.; Dreyer, J.L.; Beinert, H. The role of iron in the activation-inactivation of aconitase. J. Biol. Chem. 1983, 258, 11098–11105. [Google Scholar] [CrossRef]
- Robbins, A.H.; Stout, C.D. Structure of activated aconitase: formation of the [4Fe-4S] cluster in the crystal. Proc. Natl. Acad. Sci. USA 1989, 86, 3639–3643. [Google Scholar] [CrossRef]
- Tang, Y.; Quail, M.A.; Artymiuk, P.J.; Guest, J.R.; Green, J. Escherichia coli aconitases and oxidative stress: post-transcriptional regulation of sodA expression. Microbiology 2002, 148, 1027–1037. [Google Scholar] [CrossRef]
- Tang, Y.; Guest, J.R. Direct evidence for mRNA binding and post-transcriptional regulation by Escherichia coli aconitases. Microbiology 1999, 145, 3069–3079. [Google Scholar] [CrossRef]
- Varghese, S.; Tang, Y.; Imlay, J.A. Contrasting Sensitivities of Escherichia coli Aconitases A and B to Oxidation and Iron Depletion. J. Bacteriol. 2003, 185, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Betts, J.C.; Lukey, P.T.; Robb, L.C.; McAdam, R.A.; Duncan, K. Evaluation of a nutrient starvation model of Mycobacterium tuberculosis persistence by gene and protein expression profiling. Mol. Microbiol. 2002, 43, 717–731. [Google Scholar] [CrossRef] [PubMed]
- Wong DK, Lee BY, Horwitz MA, Gibson BW. 1999. Identification of fur, aconitase, and other proteins expressed by Mycobacterium tuberculosis under conditions of low and high concentrations of iron by combined two-dimensional gel electrophoresis and mass spectrometry. Infection and immunity 67:327–336.
- Serio, A.W.; Pechter, K.B.; Sonenshein, A.L. Bacillus subtilis Aconitase Is Required for Efficient Late-Sporulation Gene Expression. J. Bacteriol. 2006, 188, 6396–6405. [Google Scholar] [CrossRef] [PubMed]
- Alén C, Sonenshein AL. 1999. Bacillus subtilis aconitase is an RNA-binding protein. Proceedings of the National Academy of Sciences 96:10412–10417.
- Wilson, T.J.G.; Bertrand, N.; Tang, J.; Feng, J.; Pan, M.; Barber, C.E.; Dow, J.M.; Daniels, M.J. The rpfA gene of Xanthomonas campestris pathovar campestris, which is involved in the regulation of pathogenicity factor production, encodes an aconitase. Mol. Microbiol. 1998, 28, 961–970. [Google Scholar] [CrossRef]
- Austin, C.M.; Maier, R.J. Aconitase-Mediated Posttranscriptional Regulation of Helicobacter pylori Peptidoglycan Deacetylase. J. Bacteriol. 2013, 195, 5316–5322. [Google Scholar] [CrossRef]
- Oglesby-Sherrouse, A.G.; Murphy, E.R. Iron-responsive bacterial small RNAs: variations on a theme. Metallomics 2013, 5, 276–286. [Google Scholar] [CrossRef]
- Massé, E.; Gottesman, S. A small RNA regulates the expression of genes involved in iron metabolism in Escherichia coli. Proc. Natl. Acad. Sci. USA 2002, 99, 4620–4625. [Google Scholar] [CrossRef]
- Massé, E.; Escorcia, F.E.; Gottesman, S. Coupled degradation of a small regulatory RNA and its mRNA targets in Escherichia coli. Genes Dev. 2003, 17, 2374–2383. [Google Scholar] [CrossRef]
- Massé, E.; Gottesman, S. A small RNA regulates the expression of genes involved in iron metabolism in Escherichia coli. Proc. Natl. Acad. Sci. USA 2002, 99, 4620–4625. [Google Scholar] [CrossRef]
- Salvail, H.; Massé, E. Regulating iron storage and metabolism with RNA: an overview of posttranscriptional controls of intracellular iron homeostasis. Wiley Interdiscip. Rev. RNA 2011, 3, 26–36. [Google Scholar] [CrossRef]
- Kadner, R.J. Regulation by Iron: RNA Rules the Rust. J. Bacteriol. 2005, 187, 6870–6873. [Google Scholar] [CrossRef]
- Wilderman, P.J.; Sowa, N.A.; FitzGerald, D.J.; FitzGerald, P.C.; Gottesman, S.; Ochsner, U.A.; Vasil, M.L. Identification of tandem duplicate regulatory small RNAs in Pseudomonas aeruginosa involved in iron homeostasis. Proc. Natl. Acad. Sci. USA 2004, 101, 9792–9797. [Google Scholar] [CrossRef]
- Barrault, M.; Leclair, E.; Kumeko, E.K.; Jacquet, E.; Bouloc, P.; Wang, H. Staphylococcal sRNA IsrR downregulates methylthiotransferase MiaB under iron-deficient conditions. Microbiol. Spectr. 2024, 12, e0388823. [Google Scholar] [CrossRef] [PubMed]
- Garg, R.; Manhas, I.; Chaturvedi, D. Unveiling the orchestration: mycobacterial small RNAs as key mediators in host-pathogen interactions. Front. Microbiol. 2024, 15, 1399280. [Google Scholar] [CrossRef]
- Czyz, A.; Mooney, R.A.; Iaconi, A.; Landick, R.; Adhya, S. Mycobacterial RNA Polymerase Requires a U-Tract at Intrinsic Terminators and Is Aided by NusG at Suboptimal Terminators. mBio 2014, 5, e00931. [Google Scholar] [CrossRef] [PubMed]
- Prakash, P.; Yellaboina, S.; Ranjan, A.; Hasnain, S.E. Computational prediction and experimental verification of novel IdeR binding sites in the upstream sequences of Mycobacterium tuberculosis open reading frames. Bioinformatics 2005, 21, 2161–2166. [Google Scholar] [CrossRef] [PubMed]
- Gerrick, E.R.; Barbier, T.; Chase, M.R.; Xu, R.; François, J.; Lin, V.H.; Szucs, M.J.; Rock, J.M.; Ahmad, R.; Tjaden, B.; et al. Small RNA profiling in Mycobacterium tuberculosis identifies MrsI as necessary for an anticipatory iron sparing response. Proc. Natl. Acad. Sci. USA 2018, 115, 6464–6469. [Google Scholar] [CrossRef]
- Furukawa, K.; Ramesh, A.; Zhou, Z.; Weinberg, Z.; Vallery, T.; Winkler, W.C.; Breaker, R.R. Bacterial Riboswitches Cooperatively Bind Ni 2+ or Co 2+ Ions and Control Expression of Heavy Metal Transporters. Mol. Cell 2015, 57, 1088–1098. [Google Scholar] [CrossRef]
- Xu, J.; Cotruvo, J.A. The czcD (NiCo) Riboswitch Responds to Iron(II). Biochemistry 2020, 59, 1508–1516. [Google Scholar] [CrossRef]
- Xu, J.; Cotruvo, J.A. Reconsidering the czcD (NiCo) Riboswitch as an Iron Riboswitch. ACS Bio Med Chem Au 2022, 2, 376–385. [Google Scholar] [CrossRef]
- Bandyopadhyay, S.; Chaudhury, S.; Mehta, D.; Ramesh, A. RETRACTED ARTICLE: Discovery of iron-sensing bacterial riboswitches. Nat. Chem. Biol. 2020, 17, 924–924. [Google Scholar] [CrossRef]
- Grundy, F.J.; Henkin, T.M. The S box regulon: a new global transcription termination control system for methionine and cysteine biosynthesis genes in Gram-positive bacteria. Mol. Microbiol. 1998, 30, 737–749. [Google Scholar] [CrossRef]
- Epshtein, V.; Mironov, A.S.; Nudler, E. The riboswitch-mediated control of sulfur metabolism in bacteria. Proc. Natl. Acad. Sci. USA 2003, 100, 5052–5056. [Google Scholar] [CrossRef] [PubMed]
- Winkler, W.C.; Nahvi, A.; Sudarsan, N.; E Barrick, J.; Breaker, R.R. An mRNA structure that controls gene expression by binding S-adenosylmethionine. Nat. Struct. Mol. Biol. 2003, 10, 701–707. [Google Scholar] [CrossRef]
- Shahbabian, K.; Jamalli, A.; Zig, L.; Putzer, H. RNase Y, a novel endoribonuclease, initiates riboswitch turnover in Bacillus subtilis. EMBO J. 2009, 28, 3523–3533. [Google Scholar] [CrossRef]
- Maio, N.; Raza, K.; Li, Y.; Zhang, D.-L.; Bollinger, J.M.; Krebs, C.; Rouault, T.A. An iron–sulfur cluster in the zinc-binding domain of the SARS-CoV-2 helicase modulates its RNA-binding and -unwinding activities. Proc. Natl. Acad. Sci. USA 2023, 120. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



