Submitted:
21 July 2025
Posted:
24 July 2025
You are already at the latest version
Abstract
The gut microbiome plays a pivotal role in shaping both innate and adaptive immune responses, with growing evidence linking microbial composition to vaccine efficacy, particularly in respiratory infections. Probiotics, defined as live microorganisms that confer health benefits, have emerged as promising tools to modulate immunity through mucosal and systemic pathways. This review explores the mechanisms by which probiotics enhance host immunity, including activation of pattern recognition receptors, modulation of cytokine profiles, and promotion of regulatory T cells and IgA production. The gut-lung axis is highlighted as a key pathway in mucosal defense against respiratory pathogens. Beyond immunomodulation, probiotics are being explored as vaccine adjuvants and vectors. Clinical and preclinical studies suggest certain strains can improve vaccine responses and reduce respiratory infection severity. A growing number of trials are evaluating these approaches, although findings remain variable. Key challenges include strain specificity, dosing optimization, and inconsistent efficacy across populations. Continued research is needed to clarify their role and advance probiotic-based strategies as accessible tools for respiratory disease prevention.
Keywords:
respiratory vaccines and probiotics
; gut-lung axis immune modulation
; probiotics as vaccine adjuvants
; microbiome-vaccine interactions
; clinical trials
; respiratory vaccines
1. Introduction
Respiratory infections remain a significant global health concern with high morbidity and mortality rates that disproportionately affect children, the elderly, and immunocompromised individuals [1,2]. Despite the availability of vaccines, limitations such as production cost, cold-chain dependency, and suboptimal mucosal immunity requiring multiple booster shots contribute to reduced efficacy [3]. In recent years, the gut microbiome has emerged as a key player in shaping host immunity and vaccine responsiveness, offering new avenues to enhance protective strategies against respiratory pathogens [1].
A healthy and diverse microbiome is especially crucial during early life, where it supports immune system maturation and long-term immunologic memory [1]. This relationship, initiated at birth, persists throughout life and is particularly relevant to respiratory health, given the importance of mucosal immune responses in protecting the airway [1]. The gut microbiota also contributes to immune tolerance through the interaction of pattern recognition receptors (PRRs) with pathogen-associated molecular patterns (PAMPs), leading to the production of epigenetic modulators such as short-chain fatty acids (SCFAs), which are integral to mucosal immune homeostasis [4].
Disruptions to this microbial balance, known as dysbiosis, have been associated with impaired immunity [5]. Dysbiosis reduces protective T cell subsets like Th1, Th17, and inducible regulatory T cells (iTregs), while weakening systemic IgG responses, components essential for pathogen clearance and vaccine efficacy [5]. Excessive antibiotic use, particularly in early childhood, can interfere with immune memory formation and diminish vaccine responsiveness [1,5]. For instance, studies have shown that antibiotic exposure can cause a 10,000-fold reduction in gut microbial diversity and impair humoral responses, including H1N1-specific IgG1 and IgA production in individuals with low pre-existing antibody titers [6]. Germ-free animal models further underscore the microbiome’s role in immune development, as these mice display underdeveloped lymphoid structures, reduced antibody production, and heightened vulnerability to infections [7].
In this context, probiotics have garnered attention for their ability to modulate host immunity. Defined as live microorganisms that confer health benefits when administered in adequate amounts, probiotics are commonly found in fermented foods and dairy products [8]. Certain strains have demonstrated the ability to enhance both innate and adaptive immune responses, suggesting their utility in vaccine support [1]. Building on this concept, probiotic-based vaccines have emerged as a novel strategy to boost immunogenicity [1]. As adjuvants, probiotics can enhance host responses to co-administered vaccines, while as vectors, they can be engineered to express and deliver antigens directly to the immune system [1,9].
Probiotic-based vaccines offer several advantages over conventional formulations, including improved mucosal stimulation, thermal stability, and a favorable safety profile with minimal risk of reversion to pathogenicity [5]. These features are particularly valuable in the context of respiratory diseases, where mucosal immunity is paramount. Randomized controlled trials have demonstrated that certain probiotic strains enhance the immunogenicity of influenza vaccines by strengthening humoral responses [10]. Moreover, emerging data suggest that specific gut microbial profiles may improve the response to COVID-19 vaccines [11,12]. Given their potential to serve both as immunomodulators and antigen delivery systems, probiotics represent a promising platform for respiratory disease prevention. This review aims to elucidate the mechanisms by which probiotics influence immune function and to assess the current landscape of clinical trials evaluating probiotic-based vaccines in respiratory infections.
2. Effects of Probiotics on Immune Response
2.1. Enhancing Innate Immune Response
Probiotics demonstrate the ability to stimulate both the innate and adaptive immune systems. They interact with pattern recognition receptors, such as toll-like receptors (TLRs), expressed on immune cells, to activate important signaling pathways like nuclear factor κB (NF-κB) and mitogen-activated protein kinases [13]. This interaction leads to the modulation of innate immune responses, resulting in the production of both pro- and anti-inflammatory cytokines and chemokines [13]. In a preclinical study, four different probiotic strains, including Lactobacillus rhamnosus and Lacticaseibacillus casei were shown to enhance innate immunity by increasing phagocytosis in human monocyte-derived macrophages and elevating levels of reactive oxygen species [13]. Probiotics also stimulate antigen-presenting cells (APCs), including dendritic cells, macrophages, and natural killer cells, all of which contribute to immune surveillance by recognizing pathogens and presenting their antigens to naïve T cells to initiate adaptive immune responses. Upon activation, APCs release cytokines like TNF-α, IL-6, IL-12, and IFN-γ to enhance antigen presentation and trigger the downstream activation of effector T cells to elicit an immune response [5,14].
Additionally, probiotics exert antiviral effects through multifaceted mechanisms involving chemical, physical, and immunological defences. Key probiotic genera such as Lactobacillus, Bifidobacterium, Enterococcus, and Bacillus produce antiviral metabolites such as bacteriocins, hydrogen peroxide, lactic acid, SCFAs, and extracellular vesicles that disrupt viral envelopes, block replication, or interfere with viral attachment to host cell [15]. These metabolites also enhance mucosal defense by promoting mucin secretion and maintaining acidic microenvironments hostile to viral entry [15]. Probiotics can also directly bind viruses or block host cell receptors, thereby outcompeting viral attachment and entry [15]. Probiotics reinforce the physical barrier by upregulating tight junction proteins and stimulating epithelial renewal [15]. Immunologically, probiotics activate plasmacytoid dendritic cells (pDCs) which leads to increased production of type I interferons (IFN) [16,17]. IFN is a key driver of innate antiviral immunity leading to enhancement of NK cell activity, macrophage phagocytosis, production of antiviral cytokines (such as IFN-γ, IL-2, IL-6, IL-12, IL-18, TNF-α, IL-10), and expression of IFN stimulated genes that block various stages of viral infection [15,16].
2.2. Enhancing Adaptive Immune Response
In addition to activating T cells via APCs, probiotics also encourage the development of anti-inflammatory regulatory T cells (Tregs) and plasma B cells to produce more IgA [14]. Clinical trials have further substantiated probiotics’ immune-boosting effects in adaptive immune response. Researchers administered seven probiotic strains to healthy adult volunteers before and after oral vaccination against cholera toxin and enterotoxigenic Escherichia coli [18]. Two strains, Bifidobacterium lactis and Lactobacillus acidophilus significantly increased serum IgG levels compared to controls, indicating the activation of the adaptive immunity and memory systemically [18]. The serum IgG elevation was evident in the early response period (days 0–21) and persisted into the late response period (days 21–28) [18]. IgG is produced primarily by memory B cells and long-lived plasma cells and the presence of serum IgG indicates that B cells have undergone class switching from IgM for long-term immune memory against a specific pathogen [18]. These findings suggest that specific probiotics could serve as adjuvants to enhance humoral immune responses, potentially leading to improved and sustained vaccine efficacy.
2.3. Modulating Inflammatory Cytokines and Overactive Immune Responses
Probiotics play a crucial role in modulating inflammatory responses by influencing the balance between pro-inflammatory and anti-inflammatory cytokine production. A comprehensive meta-analysis of 42 randomized clinical trials involving 2,258 participants revealed that probiotic supplementation significantly reduced serum concentrations of pro-inflammatory markers, including hs-CRP, TNF-α, IL-6, IL-12, and IL-4 [19]. Concurrently, the analysis showed a significant increase in IL-10, an anti-inflammatory cytokine following probiotic supplementation [19]. These findings align with another study that found probiotics can promote the development of Tregs. Tregs produce Il-10 which is crucial for controlling overactive immune cells and reducing inflammation [19,20]. For instance, Bifidobacterium infantis has been observed to increase the proportion of Foxp3+ Treg lymphocytes in peripheral blood, leading to decreased levels of pro-inflammatory cytokines in various inflammatory conditions [13]. Kwon et al. found that another probiotic mixture (L. acidophilus, L. casei, L. reuteri, B. bifidum, and S. thermophilus) also increased numbers of CD4+ Foxp3+ r Tregs and decreased numbers of T helper (Th) 1, Th2, and Th17 cytokines in animal models [21]. These studies strongly suggest that certain probiotic strains can reduce overactive immune cells and promote anti-inflammatory and tissue-healing T cells. Altogether, these findings underscore the potential of probiotics to manage inflammatory conditions and control overactive immune responses.
2.4. Mucosal Immunity Modulation
A key mechanism by which probiotics exert their effects on respiratory immunity is mediated by the gut-lung axis. Probiotics administered orally not only alter gut microbiota composition but also impact distal mucosal sites, including the respiratory tract, through immune crosstalk mediated by dendritic cells (DCs) [22,23]. DCs play a crucial role in lung immunity by modulating both innate and adaptive responses. Probiotics can stimulate DCs and lung stromal cells to produce type I and II interferons (IFNs), activating natural killer (NK) cells and priming antigen-presenting cells such as DCs, macrophages, and B cells [24]. This leads to the activation of CD4+ and CD8+ T cells, which release cytokines that promote B cell production of secretory IgA in mucosal immunity but also secretory IgG and IgM to neutralize viral infectivity [23,24]. Additionally, the production of SCFAs has been found to prime hematopoiesis of DC precursors in the bone marrow and boost CD8+ T cell activity for antiviral functions while simultaneously promoting Treg cell differentiation and IL-10 to mitigate inflammation-induced lung damage [23,25]. Probiotics’ ability to reduce inflammatory responses may be very useful in reducing lung damage and progression to severe symptoms caused by respiratory infections such as COVID-19, seeing that overzealous inflammation and cytokine storm marked by increased levels of proinflammatory cytokines is what causes extensive systemic and tissue damage [23].
In experimental settings, the administration of probiotics has shown various protective effects against respiratory infections. First, various clinical and preclinical studies have elucidated that probiotics help maintain epithelial barrier integrity, reduce pathogen adhesion, and increase both IgA and IgG levels in serum and the respiratory tract while simultaneously decreasing systemic inflammation [24]. Probiotics have also demonstrated the ability to reduce viral loads in lung tissue [24]. Furthermore, feeding mice milk containing Lactobacillus rhamnosus prior to lethal influenza challenges increased IFN-γ and IL-12 levels in the lungs showed a shift toward antiviral Th1 response, and the increased levels of sIgA further support humoral involvement against viral invasion [26]. In clinical studies, various probiotic supplementations in senior and vulnerable populations have been shown to decrease the incidence, duration and severity of upper respiratory tract infections [26]. Altogether, these results demonstrate that specific probiotic strains serve as an effective strategy for controlling respiratory virus infections. The key actions and benefits of probiotics have been outlined in Figure 1.
3. Probiotic Use in Vaccine Research
3.1. Probiotics as Vaccine Adjuvants
Due to their natural protective effects against respiratory infections via mucosal immunity, probiotics have been evaluated as adjuvants in mucosal vaccine strategies for respiratory pathogens. Their ability to deliver immunogenic molecules via mucosal routes has been associated with an enhanced humoral and cellular immune response at both the primary (gut) and secondary (respiratory) mucosal sites [25]. The integration of probiotic supplementation with vaccination strategies has been shown to enhance immunogenicity, particularly against respiratory viruses such as influenza and SARS-CoV-2 [25]. The mechanisms underlying these effects involve stimulation of mucosal immunity and regulation of cytokine profiles [25].
Evidence from human studies supports these mechanistic insights. A randomized controlled trial conducted by Fonollá et al. demonstrated that supplementation with Lactobacillus coryniformis K8 enhanced the immunogenicity of influenza vaccination in elderly subjects, a population known for diminished vaccine responsiveness [27]. Participants in the trial exhibited higher post-vaccination antibody titers and fewer respiratory symptoms, suggesting probiotics can serve as effective adjuvants in vulnerable populations [27]. Several additional clinical trials have reported that probiotics reduce respiratory symptoms, improve vaccine efficacy, and modulate gut microbiota by increasing beneficial microbes following flu vaccinations in elderly individuals [28]. Similarly, a meta-analysis confirmed that individuals receiving probiotics exhibited a notable increase in vaccine-induced antibody titers and a reduction in the incidence and severity of respiratory tract infections, supporting the translational potential of probiotic adjuvants in clinical settings [29].
Recent studies also suggest that probiotics may enhance vaccine responses in the context of emerging respiratory pathogens, such as SARS-CoV-2 [30]. Clinical trials employing Limosilactobacillus reuteri have suggested that probiotic supplementation elicited a significant humoral immune response against COVID-19 antigens [30]. The treatment group that consumed L. reuteri had significantly higher serum anti-Spike IgG as well as anti-receptor binding domain IgG and IgA levels post vaccination, indicating a specific humoral response to SARS-CoV-2 [30]. Furthermore, a retrospective cohort study found that a probiotic mixture, when combined with best available treatment, reduced mortality in adults with severe COVID-19 pneumonia compared to those receiving standard treatment alone [31]. While the authors acknowledged limitations such as the retrospective design and lack of prospective data, these findings highlight the need for more rigorous clinical trials given the ongoing challenges in optimizing vaccine efficacy amid rapidly evolving respiratory pathogens.
3.2. Probiotics as Vaccine Vectors and Potential Candidates
Beyond their adjuvant capabilities, probiotics have gained attention as promising antigen delivery vehicles due to their ability to modulate mucosal innate and adaptive immune responses. Probiotic-based vaccine vectors have emerged as a promising strategy for respiratory diseases by combining the intrinsic immunomodulatory properties of probiotics with the ability to deliver heterologous antigens at mucosal surfaces [32]. This strategy typically involves engineering probiotic microorganisms, most commonly lactic acid bacteria, to express conserved antigens from respiratory pathogens [32]. When modified to express components such as influenza virus or SARS-CoV-2 proteins, probiotics can activate antigen-presenting cells (APCs) in the mucosal-associated lymphoid tissue (MALT), thereby inducing secretory IgA at the site of pathogen entry and systemic IgG for long-lasting immunity [17].
Although clinical evidence for probiotic-based vaccine vectors is still emerging, preclinical studies suggest their potential to induce mucosal and systemic immune responses. For instance, a live oral probiotic vaccine expressing H1N1 influenza antigens induced HA- and M2e-specific IgG antibodies in mice [33]. M2e-based vaccines in particular hold great potential as the M2e epitope is a conserved region across all influenza A viruses [33]. Another study using Lactococcus lactis expressing a cell wall-anchored version of the rotavirus VP6 vaccine demonstrated a 79.5% reduction in rotavirus shedding in mice [34]. An E. coli membrane vesicle vaccine outperformed two licensed vaccines against Streptococcus pneumoniae, with the CPS14+ vaccine eliciting significantly stronger IgG responses that persisted for a year in both systemic circulation and the lungs [35]. In the context of COVID-19, L. plantarum and L. lactis engineered to express the receptor-binding domain of the SARS-CoV-2 spike protein on their surfaces induced mucosal and systemic humoral responses in mice, evidenced by elevated serum IgG and fecal IgA [36,37]. Additionally, oral immunization with an L. lactis-based DNA mucosal vaccine encoding Mycobacterium tuberculosis antigens increased IFN-γ, TNF-α, IL-17 levels, and antigen-specific IgA, reflecting enhanced cellular and mucosal immunity [38].
Overall, probiotic vaccine vectors leverage natural mucosal targeting, safety, and adjuvanticity to deliver heterologous respiratory antigens. This dual function not only stimulates balanced immune responses at both mucosal and systemic levels but also presents a scalable, cost-effective platform for vaccine development. Key distinctions between probiotic use as adjuvants versus live vaccine vectors are illustrated in Figure 2. While clinical studies have demonstrated probiotic adjuvants can enhance the efficacy of influenza and COVID-19 vaccines, more preclinical and clinical research is needed to establish the proof-of-concept for live probiotic vaccines. Moving forward, efforts should focus on validating efficacy, optimizing dosing regimens, and ensuring long-term protection in diverse and at-risk populations.
4. Current Status of Human Clinical Trials
A search of ongoing and completed clinical trials investigating the use of probiotic vaccines to elicit protection against respiratory infections was conducted to provide insight into the current landscape of clinical research. The search identified 26 studies with start dates ranging from 2015 to 2025, as shown in Table 1. Of these, 8 were excluded because they did not involve the use of probiotics as vaccine adjuvants or as live probiotic vaccines targeting respiratory diseases.
Notably, the registered clinical trials to date have only examined probiotics as vaccine adjuvants in combination with influenza or COVID-19 vaccines. Two studies did not appear in the initial search but meet the criteria as live probiotic vaccine candidates are the bacTRL-Spike vaccine (Table 1, #18) and the Recombinant B. subtilis vaccine (Table 1, #19) [39,40]. The bacTRL-Spike vaccine is a completed phase I randomized, observer-blind, placebo-controlled trial that evaluated the use of live Bifidobacterium longum engineered to express SARS-CoV-2 spike protein DNA in healthy adults. Results from this study are not yet been known to be published [39]. To date, the Recombinant B. subtilis vaccine is the only other known live probiotic vector vaccine tested in human subjects however, its clinical results were retracted due to a lack of ethical approval for the research [41]. This study demonstrated a significant increase in neutralizing antibody against sRBD recombinantly expressed on B. subtilis spores following oral administration with no adverse health events in human subjects [41].
5. Discussion
As global vaccine needs continues to expand, novel immunization strategies are urgently needed to overcome persistent immunologic barriers and improve efficacy among vulnerable populations. Probiotic-based vaccines offer a biologically grounded and potentially scalable solution. Their compatibility with oral delivery, interaction with the mucosal immune system, and role in microbiome modulation position them as innovative tools for the next generation of vaccinology, especially in the context of respiratory diseases where the gut-lung axis plays a central immunologic role.
The need for accessible and immunologically innovative vaccines is pressing, especially as global vaccine equity remains elusive. Unlike traditional injectable vaccines, probiotic vectors and adjuvants are orally administered, thermally stable, and cost-effective to produce [9]. These attributes make them exceptionally well-suited for developing countries, where cold chain infrastructure and healthcare access can be limited [9]. Additionally, the concept of consuming beneficial microbes is already culturally embedded in many global diets through fermented foods, facilitating greater public acceptance [8]. Oral delivery also offers a practical advantage of allowing self-administration, which reduces the need for clinical oversight and expands accessibility during pandemics [9]. Beyond logistical benefits, the bidirectional communication between gut microbiota and respiratory immunity via the gut lung axis also positions probiotics uniquely to modulate mucosal responses and enhance defenses against respiratory pathogens [22,23].
Conventional vaccines also often show reduced efficacy in vulnerable populations such as children, older adults, and those with compromised immunity due to immunosenescence, microbiome dysbiosis, or comorbid disease states [44,45]. In developing countries, compounding factors such as malnutrition, poor sanitation, endemic parasitic infections, and high HIV/AIDS burdens further hinder vaccine performance [46]. For instance, oral rotavirus vaccine efficacy drops from over 80% in high-income nations to roughly 50% in resource-limited settings where the disparity is partially attributed to enteric pathogen load and environmental enteropathy [47,48]. A sanitation-focused intervention trial in Zimbabwe demonstrated improved rotavirus vaccine responses following enhanced water, sanitation, and hygiene conditions, underlining the critical interplay between microbiota health and vaccine outcomes [46]. Because probiotics can potentially restore gut microbial balance and improve mucosal immunity, they are promising adjuncts or platforms for vaccine delivery in such contexts.
Emerging clinical evidence strongly supports the role of probiotics in enhancing vaccine efficacy, preventing respiratory symptoms, and reducing the incidence of acute upper respiratory tract infections [28,29]. Studies involving Lactobacillus coryniformis K8 and Limosilactobacillus reuteri have demonstrated significant improvements in vaccine-induced immune responses in older adults, a vulnerable population often overlooked despite poor vaccine responsiveness [27,30]. These trials reported elevated post-vaccination antibody titers, reduced respiratory symptoms, and enhanced mucosal and systemic immunity, including increased SARS-CoV-2-specific IgG and IgA levels [27,30]. Meta-analyses reinforce these findings across various respiratory pathogens, highlighting probiotics’ consistent ability to modulate immune responses and reduce infection severity [29]. Early-stage clinical efforts are also beginning to explore probiotics as live vaccine vectors. For example, the bacTRL-Spike platform uses genetically engineered Bifidobacterium longum to deliver SARS-CoV-2 spike DNA to the gut [39]. Though results are pending, it marks a pivotal step in translating probiotic vector vaccines into human use and provides a foundation for future innovation in probiotic vaccine development.
To fully realize the potential of probiotic vaccines, future research must:
- Identify and characterize immunologically active strains, with a focus on their antigen-presenting and immune-modulatory mechanisms.
- Optimize delivery platforms and dosing strategies across different age groups and health conditions.
- Evaluate long-term immune memory and durability, including booster responses and cross-protection.
- Establish standardized global safety benchmarks, accounting for geographic and ethnic variations in microbiota composition.
- Integrate probiotic-based vaccine approaches into existing immunization programs, particularly in resource-limited settings and for diseases with mucosal entry routes.
If successful, probiotic vaccines could fill critical gaps in current immunization systems, particularly in settings where injectable vaccines face logistical and immunological hurdles. They hold promise not only in bolstering pandemic preparedness but also as scalable tools for vaccine equity globally. As a low-cost, orally delivered, culturally accepted, and biologically rational platform, probiotic vaccines may soon redefine what global vaccine accessibility and innovation look like.
References
- Kazemifard, N.; Dehkohneh, A.; Baradaran Ghavami, S. Probiotics and Probiotic-Based Vaccines: A Novel Approach for Improving Vaccine Efficacy. Front. Med. 2022, 9, 940454, . [CrossRef]
- Muthukutty, P.; MacDonald, J.; Yoo, S.Y. Combating Emerging Respiratory Viruses: Lessons and Future Antiviral Strategies. Vaccines 2024, 12, 1220, . [CrossRef]
- Md Khairi, L.N.H.; Fahrni, M.L.; Lazzarino, A.I. The Race for Global Equitable Access to COVID-19 Vaccines. Vaccines 2022, 10, 1306, . [CrossRef]
- Jiao, Y.; Wu, L.; Huntington, N.D.; Zhang, X. Crosstalk Between Gut Microbiota and Innate Immunity and Its Implication in Autoimmune Diseases. Front. Immunol. 2020, 11, . [CrossRef]
- Singh, S.; Singh, M.; Gaur, S. Probiotics as Multifaceted Oral Vaccines against Colon Cancer: A Review. Front. Immunol. 2022, 13, . [CrossRef]
- Hagan, T.; Cortese, M.; Rouphael, N.; Boudreau, C.; Linde, C.; Maddur, M.S.; Das, J.; Wang, H.; Guthmiller, J.; Zheng, N.-Y.; et al. Antibiotics-Driven Gut Microbiome Perturbation Alters Immunity to Vaccines in Humans. Cell 2019, 178, 1313-1328.e13, . [CrossRef]
- Wu, H.-J.; Wu, E. The Role of Gut Microbiota in Immune Homeostasis and Autoimmunity. Gut Microbes 2012, 3, 4–14, . [CrossRef]
- Xu, C.; Aqib, A.I.; Fatima, M.; Muneer, S.; Zaheer, T.; Peng, S.; Ibrahim, E.H.; Li, K. Deciphering the Potential of Probiotics in Vaccines. Vaccines 2024, 12, 711, . [CrossRef]
- Taghinezhad-S, S.; Mohseni, A.H.; Bermúdez-Humarán, L.G.; Casolaro, V.; Cortes-Perez, N.G.; Keyvani, H.; Simal-Gandara, J. Probiotic-Based Vaccines May Provide Effective Protection against COVID-19 Acute Respiratory Disease. Vaccines 2021, 9, 466, . [CrossRef]
- Lei, W.-T.; Shih, P.-C.; Liu, S.-J.; Lin, C.-Y.; Yeh, T.-L. Effect of Probiotics and Prebiotics on Immune Response to Influenza Vaccination in Adults: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Nutrients 2017, 9, 1175, . [CrossRef]
- Ng, S.C.; Peng, Y.; Zhang, L.; Mok, C.K.; Zhao, S.; Li, A.; Ching, J.Y.; Liu, Y.; Yan, S.; Chan, D.L.S.; et al. Gut Microbiota Composition Is Associated with SARS-CoV-2 Vaccine Immunogenicity and Adverse Events. 2022, . [CrossRef]
- Leung, J.S.M. Interaction between Gut Microbiota and COVID-19 and Its Vaccines. World J. Gastroenterol. 2022, 28, 5801–5806, . [CrossRef]
- Liu, Y.; Wang, J.; Wu, C. Modulation of Gut Microbiota and Immune System by Probiotics, Pre-Biotics, and Post-Biotics. Front. Nutr. 2022, 8, . [CrossRef]
- Zhou, P.; Chen, C.; Patil, S.; Dong, S. Unveiling the Therapeutic Symphony of Probiotics, Prebiotics, and Postbiotics in Gut-Immune Harmony. Front. Nutr. 2024, 11, . [CrossRef]
- Wang, Y.; Moon, A.; Huang, J.; Sun, Y.; Qiu, H.-J. Antiviral Effects and Underlying Mechanisms of Probiotics as Promising Antivirals. Front. Cell. Infect. Microbiol. 2022, 12, 928050, . [CrossRef]
- Jounai, K.; Ikado, K.; Sugimura, T.; Ano, Y.; Braun, J.; Fujiwara, D. Spherical Lactic Acid Bacteria Activate Plasmacytoid Dendritic Cells Immunomodulatory Function via TLR9-Dependent Crosstalk with Myeloid Dendritic Cells. PLOS ONE 2012, 7, e32588, . [CrossRef]
- Kanauchi, O.; Low, Z.X.; Jounai, K.; Tsuji, R.; AbuBakar, S. Overview of Anti-Viral Effects of Probiotics via Immune Cells in Pre-, Mid- and Post-SARS-CoV2 Era. Front. Immunol. 2023, 14, 1280680, . [CrossRef]
- Paineau, D.; Carcano, D.; Leyer, G.; Darquy, S.; Alyanakian, M.-A.; Simoneau, G.; Bergmann, J.-F.; Brassart, D.; Bornet, F.; Ouwehand, A.C. Effects of Seven Potential Probiotic Strains on Specific Immune Responses in Healthy Adults: A Double-Blind, Randomized, Controlled Trial. FEMS Immunol. Med. Microbiol. 2008, 53, 107–113, . [CrossRef]
- Milajerdi, A.; Mousavi, S.M.; Sadeghi, A.; Salari-Moghaddam, A.; Parohan, M.; Larijani, B.; Esmaillzadeh, A. The Effect of Probiotics on Inflammatory Biomarkers: A Meta-Analysis of Randomized Clinical Trials. Eur. J. Nutr. 2020, 59, 633–649, . [CrossRef]
- Mazziotta, C.; Tognon, M.; Martini, F.; Torreggiani, E.; Rotondo, J.C. Probiotics Mechanism of Action on Immune Cells and Beneficial Effects on Human Health. Cells 2023, 12, 184, . [CrossRef]
- Kwon, H.-K.; Lee, C.-G.; So, J.-S.; Chae, C.-S.; Hwang, J.-S.; Sahoo, A.; Nam, J.H.; Rhee, J.H.; Hwang, K.-C.; Im, S.-H. Generation of Regulatory Dendritic Cells and CD4+Foxp3+ T Cells by Probiotics Administration Suppresses Immune Disorders. Proc. Natl. Acad. Sci. U. S. A. 2010, 107, 2159–2164, . [CrossRef]
- Dang, A.T.; Marsland, B.J. Microbes, Metabolites, and the Gut–Lung Axis. Mucosal Immunol. 2019, 12, 843–850, . [CrossRef]
- Cruz, C.S.; Ricci, M.F.; Vieira, A.T. Gut Microbiota Modulation as a Potential Target for the Treatment of Lung Infections. Front. Pharmacol. 2021, 12, . [CrossRef]
- Wang, Y.-H.; Limaye, A.; Liu, J.-R.; Wu, T.-N. Potential Probiotics for Regulation of the Gut-Lung Axis to Prevent or Alleviate Influenza in Vulnerable Populations. J. Tradit. Complement. Med. 2023, 13, 161–169, . [CrossRef]
- Amrouche, T.; Chikindas, M.L.; Amrouche, T.; Chikindas, M.L. Probiotics for Immunomodulation in Prevention against Respiratory Viral Infections with Special Emphasis on COVID-19. AIMS Microbiol. 2022, 8, 338–356, . [CrossRef]
- Song, J.A.; Kim, H.J.; Hong, S.K.; Lee, D.H.; Lee, S.W.; Song, C.S.; Kim, K.T.; Choi, I.S.; Lee, J.B.; Park, S.Y. Oral Intake of Lactobacillus Rhamnosus M21 Enhances the Survival Rate of Mice Lethally Infected with Influenza Virus. J. Microbiol. Immunol. Infect. 2016, 49, 16–23, . [CrossRef]
- Fonollá, J.; Gracián, C.; Maldonado-Lobón, J.A.; Romero, C.; Bédmar, A.; Carrillo, J.C.; Martín-Castro, C.; Cabrera, A.L.; García-Curiel, J.M.; Rodríguez, C.; et al. Effects of Lactobacillus Coryniformis K8 CECT5711 on the Immune Response to Influenza Vaccination and the Assessment of Common Respiratory Symptoms in Elderly Subjects: A Randomized Controlled Trial. Eur. J. Nutr. 2019, 58, 83–90, . [CrossRef]
- Sandionigi, A.; De Giani, A.; Tursi, F.; Michelotti, A.; Cestone, E.; Giardina, S.; Zampolli, J.; Di Gennaro, P. Effectiveness of Multistrain Probiotic Formulation on Common Infectious Disease Symptoms and Gut Microbiota Modulation in Flu-Vaccinated Healthy Elderly Subjects. BioMed Res. Int. 2022, 2022, 3860896, . [CrossRef]
- Zhao, Y.; Dong, B.R.; Hao, Q. Probiotics for Preventing Acute Upper Respiratory Tract Infections - Zhao, Y - 2022 | Cochrane Library.
- Forsgård, R.A.; Rode ,Julia; Lobenius-Palmér ,Karin; Kamm ,Annalena; Patil ,Snehal; Tacken ,Mirriam G. J.; Lentjes ,Marleen A. H.; Axelsson ,Jakob; Grompone ,Gianfranco; Montgomery ,Scott; et al. Limosilactobacillus Reuteri DSM 17938 Supplementation and SARS-CoV-2 Specific Antibody Response in Healthy Adults: A Randomized, Triple-Blinded, Placebo-Controlled Trial. Gut Microbes 2023, 15, 2229938, . [CrossRef]
- Ceccarelli, G.; Borrazzo, C.; Pinacchio, C.; Santinelli, L.; Innocenti, G.P.; Cavallari, E.N.; Celani, L.; Marazzato, M.; Alessandri, F.; Ruberto, F.; et al. Oral Bacteriotherapy in Patients With COVID-19: A Retrospective Cohort Study. Front. Nutr. 2021, 7, . [CrossRef]
- Levit, R.; Cortes-Perez, N.G.; Leblanc, A. de M. de; Loiseau, J.; Aucouturier, A.; Langella, P.; LeBlanc, J.G.; Bermúdez-Humarán, L.G. Use of Genetically Modified Lactic Acid Bacteria and Bifidobacteria as Live Delivery Vectors for Human and Animal Health. Gut Microbes 2022.
- Mezhenskaya, D.; Isakova-Sivak, I.; Gupalova, T.; Bormotova, E.; Kuleshevich, E.; Kramskaya, T.; Leontieva, G.; Rudenko, L.; Suvorov, A. A Live Probiotic Vaccine Prototype Based on Conserved Influenza a Virus Antigens Protect Mice against Lethal Influenza Virus Infection. Biomedicines 2021, 9, 1515, . [CrossRef]
- Temprana, C.F.; Argüelles, M.H.; Gutierrez, N.M.; Barril, P.A.; Esteban, L.E.; Silvestre, D.; Mandile, M.G.; Glikmann, G.; Castello, A.A. Rotavirus VP6 Protein Mucosally Delivered by Cell Wall-Derived Particles from Lactococcus Lactis Induces Protection against Infection in a Murine Model. PLOS ONE 2018, 13, e0203700, . [CrossRef]
- Nakao, R.; Kobayashi, H.; Iwabuchi, Y.; Kawahara, K.; Hirayama, S.; Ramstedt, M.; Sasaki, Y.; Kataoka, M.; Akeda, Y.; Ohnishi, M. A Highly Immunogenic Vaccine Platform against Encapsulated Pathogens Using Chimeric Probiotic Escherichia Coli Membrane Vesicles. Npj Vaccines 2022, 7, 1–17, . [CrossRef]
- Li, L.; Wang, M.; Hao, J.; Han, J.; Fu, T.; Bai, J.; Tian, M.; Jin, N.; Zhu, G.; Li, C. Mucosal IgA Response Elicited by Intranasal Immunization of Lactobacillus Plantarum Expressing Surface-Displayed RBD Protein of SARS-CoV-2. Int. J. Biol. Macromol. 2021, 190, 409–416, . [CrossRef]
- Xuan, B.; Park, J.; Yoo, J.H.; Kim, E.B. Oral Immunization of Mice with Cell Extracts from Recombinant Lactococcus Lactis Expressing SARS-CoV-2 Spike Protein. Curr. Microbiol. 2022, 79, 167, . [CrossRef]
- de Castro, C.P.; Souza, B.M.; Mancha-Agresti, P.; Pereira, V.B.; Zurita-Turk, M.; Preisser, T.M.; da Cunha, V.P.; dos Santos, J.S.C.; Leclercq, S.Y.; Azevedo, V.; et al. Lactococcus Lactis FNBPA+ (pValac:E6ag85a) Induces Cellular and Humoral Immune Responses After Oral Immunization of Mice. Front. Microbiol. 2021, 12, 676172, . [CrossRef]
- Symvivo Corporation A Phase 1, Randomized, Observer-Blind, Placebo-Controlled Trial to Evaluate the Safety, Tolerability and Immunogenicity of the bacTRL-Spike Oral Candidate Vaccine for the Prevention of COVID-19 in Healthy Adults; clinicaltrials.gov, 2022;
- Sung, J.C.-C.; Liu, Y.; Wu, K.-C.; Choi, M.-C.; Ma, C.H.-Y.; Lin, J.; He, E.I.C.; Leung, D.Y.-M.; Sze, E.T.-P.; Hamied, Y.K.; et al. Expression of SARS-CoV-2 Spike Protein Receptor Binding Domain on Recombinant B. Subtilis on Spore Surface: A Potential COVID-19 Oral Vaccine Candidate. Vaccines 2021, 10, 2, . [CrossRef]
- Sung, J.C.-C.; Liu, Y.; Wu, K.-C.; Choi, M.-C.; Ma, C.H.-Y.; Lin, J.; He, E.I.C.; Leung, D.Y.-M.; Sze, E.T.-P.; Hamied, Y.K.; et al. Retraction: Sung et al. Expression of SARS-CoV-2 Spike Protein Receptor Binding Domain on Recombinant B. Subtilis on Spore Surface: A Potential COVID-19 Oral Vaccine Candidate. Vaccines 2022, 10, 2. Vaccines 2022, 10, 1852, . [CrossRef]
- ClinicalTrials.Gov Glossary Terms | ClinicalTrials.Gov Available online: https://clinicaltrials.gov/study-basics/glossary (accessed on 15 April 2025).
- Search for: Respiratory Disease, Other Terms: Probiotic Vaccine | List Results | ClinicalTrials.Gov Available online: https://clinicaltrials.gov/search?cond=Respiratory%20Disease&term=Probiotic%20vaccine (accessed on 11 July 2025).
- Peroni, D.G.; Morelli, L. Probiotics as Adjuvants in Vaccine Strategy: Is There More Room for Improvement? Vaccines 2021, 9, 811, . [CrossRef]
- Akatsu, H. Exploring the Effect of Probiotics, Prebiotics, and Postbiotics in Strengthening Immune Activity in the Elderly. Vaccines 2021, 9, 136, . [CrossRef]
- Church, J.A.; Rukobo, S.; Govha, M.; Lee, B.; Carmolli, M.P.; Chasekwa, B.; Ntozini, R.; Mutasa, K.; McNeal, M.M.; Majo, F.D.; et al. The Impact of Improved Water, Sanitation, and Hygiene on Oral Rotavirus Vaccine Immunogenicity in Zimbabwean Infants: Substudy of a Cluster-Randomized Trial. Clin. Infect. Dis. 2019, 69, 2074–2081, . [CrossRef]
- Jiang, V.; Jiang, B.; Tate, J.; Parashar, U.D.; Patel, M.M. Performance of Rotavirus Vaccines in Developed and Developing Countries. Hum. Vaccin. 2010, 6, 532–542, . [CrossRef]
- Levine, M.M. Immunogenicity and Efficacy of Oral Vaccines in Developing Countries: Lessons from a Live Cholera Vaccine. BMC Biol. 2010, 8, 129, . [CrossRef]
Figure 1.
Immunomodulatory Effects of Probiotics on Innate, Adaptive, and Mucosal Immunity [13,14,18,19,20,21,22,23,24]. Overview of the diverse immunological effects of probiotics on immune function across innate, adaptive, and mucosal pathways. Innate Immunity: Probiotics activate dendritic cells (DCs), macrophages, and NK cells to produce TNF-α, IL-6, IL-12, and IFN-γ, enhancing antigen presentation and effector T cell activation. Adaptive Immunity: Probiotics promote regulatory T cell (Treg) development and stimulate plasma B cells to produce IgA and IgG, supporting immune memory and long-term protection. Immune Modulation: Probiotics reduce pro-inflammatory markers (e.g., TNF-α, IL-6, IL-12), increase CD4⁺Foxp3⁺ Tregs, and suppress Th1, Th2, and Th17 responses to limit inflammation. Mucosal Immunity: Probiotics stimulate IFN production via DCs, activate NK cells and APCs, and enhance T and B cell responses. SCFAs promote hematopoiesis and CD8⁺ T cell and Treg differentiation. Figure created with BioRender.
Figure 1.
Immunomodulatory Effects of Probiotics on Innate, Adaptive, and Mucosal Immunity [13,14,18,19,20,21,22,23,24]. Overview of the diverse immunological effects of probiotics on immune function across innate, adaptive, and mucosal pathways. Innate Immunity: Probiotics activate dendritic cells (DCs), macrophages, and NK cells to produce TNF-α, IL-6, IL-12, and IFN-γ, enhancing antigen presentation and effector T cell activation. Adaptive Immunity: Probiotics promote regulatory T cell (Treg) development and stimulate plasma B cells to produce IgA and IgG, supporting immune memory and long-term protection. Immune Modulation: Probiotics reduce pro-inflammatory markers (e.g., TNF-α, IL-6, IL-12), increase CD4⁺Foxp3⁺ Tregs, and suppress Th1, Th2, and Th17 responses to limit inflammation. Mucosal Immunity: Probiotics stimulate IFN production via DCs, activate NK cells and APCs, and enhance T and B cell responses. SCFAs promote hematopoiesis and CD8⁺ T cell and Treg differentiation. Figure created with BioRender.
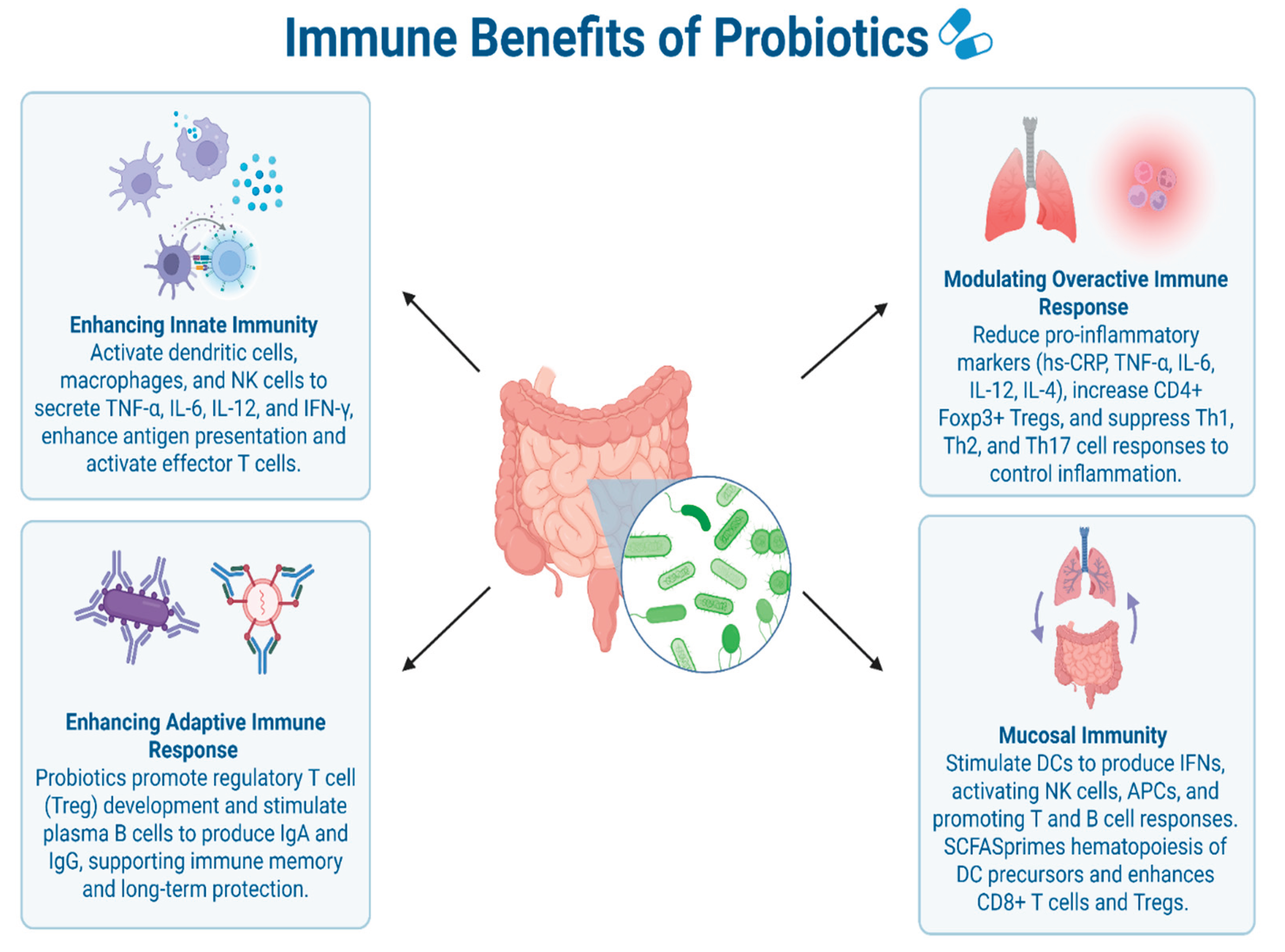
Figure 2.
Summary of Probiotic-Based Vaccine Strategies: Adjuvants vs. Vectors [17,25,26,27,28,29,30,31,32,33,34,35,36,37,38]. Comparison of the two primary approaches to probiotic vaccine development: (A) probiotic-based adjuvants, which enhance immune responses to conventional vaccines, and (B) genetically engineered probiotic vectors that deliver heterologous respiratory pathogen antigens directly at mucosal sites. Both strategies are non-invasive and promote robust mucosal and systemic immunity, though they differ in mechanism, development stage, and translational maturity. Vaccine Adjuvant (Top Panel): Enhances immune response via cytokine modulation, mucosal antigen presentation, and activation of humoral and cellular immunity. Benefits include oral delivery, improved vaccine efficacy, and reduced incidence/severity of infections. Vaccine Vector (Bottom Panel): Uses engineered bacteria to deliver antigens, triggering secretory and systemic immunity. Offers dual-functionality, stability, and adaptability to emerging pathogens or immune-compromised individuals. Figure created with BioRender.
Figure 2.
Summary of Probiotic-Based Vaccine Strategies: Adjuvants vs. Vectors [17,25,26,27,28,29,30,31,32,33,34,35,36,37,38]. Comparison of the two primary approaches to probiotic vaccine development: (A) probiotic-based adjuvants, which enhance immune responses to conventional vaccines, and (B) genetically engineered probiotic vectors that deliver heterologous respiratory pathogen antigens directly at mucosal sites. Both strategies are non-invasive and promote robust mucosal and systemic immunity, though they differ in mechanism, development stage, and translational maturity. Vaccine Adjuvant (Top Panel): Enhances immune response via cytokine modulation, mucosal antigen presentation, and activation of humoral and cellular immunity. Benefits include oral delivery, improved vaccine efficacy, and reduced incidence/severity of infections. Vaccine Vector (Bottom Panel): Uses engineered bacteria to deliver antigens, triggering secretory and systemic immunity. Offers dual-functionality, stability, and adaptability to emerging pathogens or immune-compromised individuals. Figure created with BioRender.
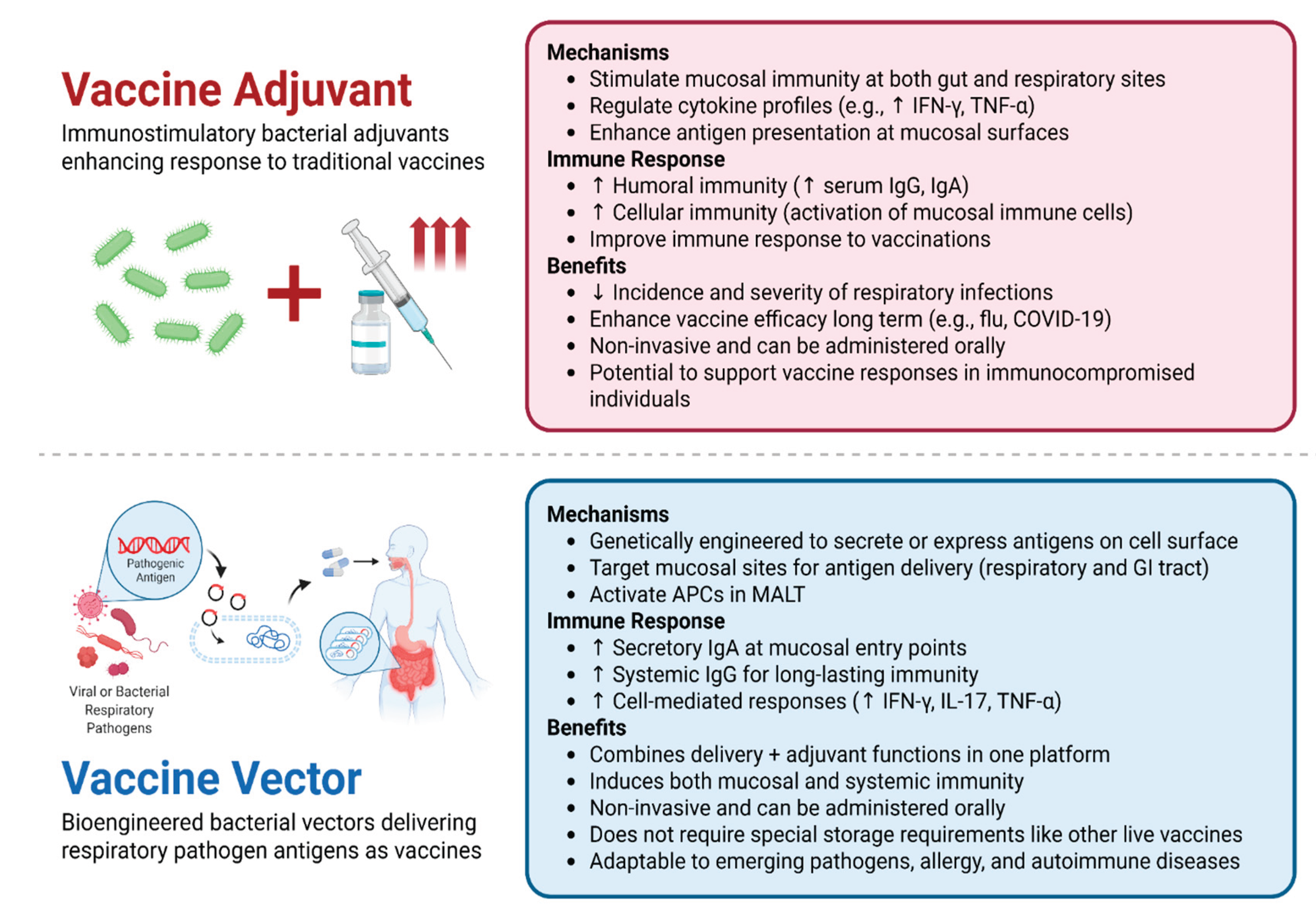

Table 1.
Ongoing and Completed Clinical Trials of Probiotic-Based Vaccines for Respiratory Infections. Summary of registered clinical trials investigating probiotics used as vaccine adjuvants or live vectors against respiratory infectious diseases, including influenza and COVID-19. Search terms included “respiratory disease” OR “respiratory infection” AND “probiotic vaccine”. Most trials were categorized under the "Not Applicable" trial phase designation, which refers to studies without FDA-defined phases including those involving medical devices or behavioral interventions [42]. As many of these studies are conducted under different regulatory authorities worldwide, the lack of standardized phase reporting may account for this designation. All trials are referenced from ClinicalTrials.gov on July 11, 2022 [43]. See the Supplemental Table S1 for more comprehensive results.
Table 1.
Ongoing and Completed Clinical Trials of Probiotic-Based Vaccines for Respiratory Infections. Summary of registered clinical trials investigating probiotics used as vaccine adjuvants or live vectors against respiratory infectious diseases, including influenza and COVID-19. Search terms included “respiratory disease” OR “respiratory infection” AND “probiotic vaccine”. Most trials were categorized under the "Not Applicable" trial phase designation, which refers to studies without FDA-defined phases including those involving medical devices or behavioral interventions [42]. As many of these studies are conducted under different regulatory authorities worldwide, the lack of standardized phase reporting may account for this designation. All trials are referenced from ClinicalTrials.gov on July 11, 2022 [43]. See the Supplemental Table S1 for more comprehensive results.
| # | Respiratory Disease | Probiotic Use (vaccine adjuvant/ live vaccine) | Bacterial Strain | Phase | NCT # |
| 1 | Influenza | vaccine adjuvant | Multistrain Probiotic (unspecified) | N/A | NCT06103994 |
| 2 | Influenza | vaccine adjuvant | Probiotic supplement (unspecified) | N/A | NCT05690373 |
| 3 | COVID-19 | vaccine adjuvant | 2-strain probiotic blend (unspecified) | N/A | NCT05195151 |
| 4 | Influenza | vaccine adjuvant | Lactiplantibacillus plantarum + P. acidilactici | N/A | NCT05157425 |
| 5 | COVID-19 | vaccine adjuvant | Bifidobacteria blend 3 strains (unpecified) | N/A | NCT04884776 |
| 6 | COVID-19 | vaccine adjuvant | Lactobacillus | N/A | NCT04756466 |
| 7 | Influenza | vaccine adjuvant | Lactobacillus acidophilus + Lactobacillus rhamnosus | Phase 4 | NCT03695432 |
| 8 | Influenza | vaccine adjuvant |
Lactobacillus coryniformis |
Phase 2 | NCT03167593 |
| 9 | Influenza | vaccine adjuvant | Lactobacillus paracasei | N/A | NCT02909842 |
| 10 | Influenza | vaccine adjuvant | Lactobacillus Gasseri + Bifidobacterium Longum + Bifidobacterium Bifidum | N/A | NCT01652066 |
| 11 | Influenza | vaccine adjuvant | Lactobacillus rhamnosus GG | Phase 1 | NCT01545349 |
| 12 | Influenza | vaccine adjuvant | Lactobacillus rhamnosus GG | Phase 1 | NCT01368029 |
| 13 | Influenza | vaccine adjuvant | Enterococcus faecium + Saccharomyces cerevisiae | Phase 1 | NCT01304771 |
| 14 | Influenza | vaccine adjuvant | Bifidobacterium longum | N/A | NCT01066377 |
| 15 | Influenza | vaccine adjuvant | Lactobacillus casei Shirota | Phase 4 | NCT00849277 |
| 16 | Influenza | vaccine adjuvant | Lactobacillus rhamnosus | Phase 1 | NCT00620412 |
| 17 | Influenza and COVID-19 | vaccine adjuvant | Saccharomyces cerevisiae blend | N/A | NCT04798677 |
| 18 | COVID-19 | Live vector vaccine | Bifidobacterium longum | Phase I | NCT04334980 |
| 19* | COVID-19 | Live vector vaccine | Bacillus subtilis | N/A | NCT05057923 |
*Results retracted [41].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



