
Submitted:
21 July 2025
Posted:
22 July 2025
You are already at the latest version
Abstract
We investigated the impact of liver damage on systemic inter-organ communication in an extensive observational case-control study of 923 patients with severe obesity and biopsy-confirmed metabolic dysfunction-associated steatotic liver disease (MASLD) or metabolic dysfunction-associated steatohepatitis (MASH) undergoing bariatric surgery. Using a comprehensive panel of circulating organokines, including fibroblast growth factor (FGF) 19, FGF21, adiponectin, galectin-3, irisin, and leptin, along with choline metabolites, we characterized metabolic signaling patterns associated with liver disease severity. Compared to controls, patients with MASLD/MASH exhibited significantly lower levels of FGF19, choline, and trimethylamine, while FGF21, galectin-3, irisin, and leptin were elevated. Sex-specific alterations in leptin and adiponectin were observed in patients with severe obesity but not in controls. Network analysis revealed a complex and individualized interplay among organokines, shaped by age, sex, and anthropometric factors. Despite this complexity, a reproducible dysregulation of the FGF21–adiponectin axis was associated with more advanced liver involvement. The large cohort and comprehensive organokine profiling studied provide valuable insights into the role of the FGF21–adiponectin axis on systemic metabolic alterations in severe obesity and their potential clinical implications.
Keywords:
adipose tissue-liver axis
; choline metabolism
; interorgan crosstalk
; MASLD
; MASH
; microbiota
; organokines
1. Introduction
Different organisms have evolved sophisticated communication systems among their organs to coordinate metabolic responses efficiently in changing environmental and nutritional conditions. However, chronic nutrient excess and impaired nutrient processing can disrupt these inter-organ communication pathways, compromising metabolic homeostasis. This disruption has led to a growing public health challenge characterized by obesity and associated metabolic diseases, including liver dysfunction [1,2]. These metabolic abnormalities often coexist and may further interfere with organ crosstalk, especially in severe or morbid obesity, complicating personalized management strategies. Clinically, severe or morbid obesity is defined by a body mass index (BMI) of 40 kg/m² or greater, or a BMI of 35 kg/m² or greater with associated comorbidities. This condition frequently involves liver damage and is linked to an increased risk of serious metabolic complications [3,4].
Understanding the relationship between severe obesity and the progression of metabolic dysfunction-associated steatotic liver disease (MASLD) is critical. Patients advancing from MASLD to metabolic dysfunction-associated steatohepatitis (MASH) and fibrosis face a substantially heightened risk of cirrhosis and hepatocellular carcinoma [5]. Importantly, adipose tissue expansion beyond a certain threshold leads to metabolic dysregulation, highlighting the necessity for proactive interventions such as aggressive weight loss strategies and emerging pharmacological therapies [6,7].
Recent advances have reframed adipose tissue as an active endocrine organ coordinating systemic metabolism rather than merely an energy reservoir [8,9,10]. Communication among metabolic organs, including adipose tissue, liver, gut, and skeletal muscle, is primarily mediated by secreted factors known as organokines [11,12]. This “organokine crosstalk” is a complex network where hormones and metabolites function as signaling molecules that modulate energy homeostasis and tissue function [13,14]. For example, these signals can regulate liver fat accumulation and impact gut microbiota composition, bile acid metabolism, and lipid handling [15,16,17]. Yet, the detailed molecular mechanisms governing these interactions remain incompletely understood, particularly in the context of severe obesity, due to a lack of studies combining large patient cohorts, biopsy-confirmed liver disease, and comprehensive investigation of circulating organokines and metabolic markers [18].
Treatment options for severe obesity are limited, with bariatric surgery being the only consistently effective intervention for sustained weight loss and metabolic improvement. However, not all patients are candidates, and long-term outcomes remain variable [19]. Understanding the metabolic alterations and disrupted inter-organ communication that accompany this disease is essential to identifying new biomarkers and therapeutic targets. Unfortunately, few studies have addressed these mechanisms in depth, especially in large cohorts with biopsy-proven liver disease and comprehensive organokine and metabolite profiling.
Building on previous work implicating mitochondrial dysfunction and organelle interplay in liver metabolic impairment [20,21], the present study aims to deepen the understanding of how organokine networks are altered across the spectrum of liver damage and metabolic dysfunction in a extensive cohort of patients with severe obesity. By systematically profiling key circulating organokines and related metabolites, we seek to elucidate the complex inter-organ communication patterns that characterize metabolic complications in severe obesity and explore their potential clinical implications.
2. Results
2.1. Participants
Anthropometric measurements and laboratory tests confirmed the link between excess body fat, metabolic dysfunction, and liver damage (Table 1). The patients with severe obesity shared a similar ethnic background and originated from the same geographical area. Sex distribution was noticeably skewed, with a significantly higher proportion of women than men. Patients showed a significantly higher prevalence of type 2 diabetes mellitus, hypertension, and dyslipidemia, which was reflected in elevated levels of glucose, insulin, triglycerides, and aminotransferases. In terms of organokines and choline metabolites, they showed lower concentrations of fibroblast growth factor (FGF) 19, choline, and trimethylamine (TMA), and higher concentrations of FGF21, irisin, and leptin. Analysis of liver histological features showed no significant differences based on sex. Therefore, we present data for the entire cohort (Supplementary Tables S1 and S2). The prevalence of MASLD was high, characterized by a complex distribution of liver lesions across a broad spectrum. Only 43 patients had a nonalcoholic fatty liver disease activity score (NAS) of 0, while 217 (24.2%) exhibited definite MASH (Supplementary Table S1 and Supplementary Figure S1). Metabolic dysfunction correlated with the severity of liver damage. In patients with MASH, older age, higher anthropometric measurements, comorbidities, elevated plasma levels of FGF21 and lower adiponectin levels were evident (Supplementary Table S2).
No sex-related differences were observed in organokines and metabolites within the control group. However, among patients with severe obesity, plasma levels of leptin and adiponectin were higher in women than in men (Supplementary Figure S2).
2.2. Fibroblast Growth Factor 19 and Choline Metabolism
Patients with severe obesity had significantly lower circulating FGF19 levels than controls, which inversely correlated with body size measurements (Figure 1a). These differences were independent of dyslipidemia, hypertension, or diabetes (Figure 1b). Plasma FGF19 levels remained unaffected by varying degrees of liver damage (Supplementary Table S2). Specifically, there were no significant differences between patients with and without MASH, or associations with liver histologic features (Supplementary Table S3).
As shown in Supplementary Figure S3, the small intestine is the primary source of FGF19, and circulating metabolites related to choline metabolism can influence communication between the gut and the liver. Our study found that plasma levels of choline and TMA were significantly lower in individuals with severe obesity compared to the control group, and these levels were not associated with body measurements. The differences in betaine and trimethilamine N-oxide (TMAO) levels did not reach statistical significance (Figure 1c), nor did we observe any relationships with metabolic comorbidities. However, ordinal logistic regression analyses revealed significant associations with hepatic histological features. Specifically, plasma choline levels increased with the NAS and were associated with a higher fibrosis score. Plasma levels of choline and TMAO were higher in patients with the highest ballooning scores (Figure 1c and Supplementary Table S3).
2.3. Fibroblast Growth Factor 21
Patients with severe obesity had significantly higher levels of circulating FGF21 than the control group. These elevated levels positively correlated with various measures of body size (Figure 2a). Plasma FGF21 levels were also associated with common markers of liver injury and glucose metabolism. Although we did not find any significant associations with dyslipidemia or hypertension, it is noteworthy that patients with severe obesity and diabetes had higher FGF21 concentrations than those without diabetes (Figure 2b).
The liver is the primary source of FGF21, and circulating FGF21 levels closely correlate with liver histopathology. Patients with MASH exhibited significantly higher plasma FGF21 concentrations than those without MASH. Furthermore, there were robust correlations between FGF21 levels and the NAS, with positive relationships for lobular inflammation, steatosis, and fibrosis. Receiver operating characteristic curve analyses indicated that FGF21 has limited accuracy as a non-invasive diagnostic marker. Nonetheless, its significant associations with metabolic comorbidities and liver pathology suggest potential utility in guiding therapeutic strategies (Figure 2c and Supplementary Tables S2 and S3).
2.4. Galectin-3
Galectin-3, previously known as macrophage antigen 2 (Mac-2), is a multifunctional protein produced and secreted by various types of cells. In our study cohort, we found that plasma levels of galectin-3 were positively correlated with BMI and were significantly higher in patients with severe obesity compared to controls (Supplementary Figure S4a). Additionally, patients with diabetes had markedly elevated levels of galectin-3 in their circulation, with no associated changes related to hypertension or dyslipidemia (Supplementary Figure S4b). MASH did not affect plasma galectin-3 levels. However, we did find that plasma galectin-3 decreased in patients with greater liver fibrosis. Furthermore, galectin-3 was consistently associated with liver macrophages (Supplementary Figure S4c and Supplementary Tables S2 and S3).
2.5. Irisin
The muscle generates the highest amounts of irisin. We observed that patients with severe obesity had higher circulating irisin levels than controls. However, plasma irisin levels showed a poor correlation with anthropometric measurements (Supplementary Figure S5a). In contrast to controls, the presence of dyslipidemia and hypertension was significantly associated with lower circulating levels of irisin in patients with severe obesity. There was no significant relationship between irisin levels and diabetes (Supplementary Figure S5b). Although we found a positive correlation with the ballooning score, our results do not indicate strong associations with MASH in these patients (Supplementary Figure S5c and Supplementary Tables S2 and S3).
2.6. Leptin and Adiponectin: The Relevance of Sex Dimorphism
When analyzing the entire cohort, we found that circulating leptin levels were higher in patients with severe obesity than in controls, and there was a significant positive correlation between anthropometric measures and leptin levels (Figure 3a). However, plasma leptin levels were lower in patients with metabolic comorbidities, with a statistically significant trend noted only among those with diabetes (Figure 3b). Liver damage did not appear to influence the levels of this adipokine, and we found no appreciable differences between patients with and without MASH (Supplementary Table S2). Interestingly, patients with more ballooned liver cells had significantly higher plasma leptin concentrations than those without ballooning (Figure 3c, Supplementary Table S3).
We also noted significant sex differences in our findings. Women with severe obesity had higher circulating leptin levels than men (Supplementary Figure S2), and these levels strongly correlated with body size measurements for both sexes (Supplementary Figure S6a). Notably, the impact of diabetes varied by sex; it was not significant in men, while women with diabetes had lower plasma leptin levels compared to those without diabetes (Supplementary Figure S6b). The pattern was not observed in the control group. Furthermore, the relationship between circulating leptin and ballooning score was significant only in women (Supplementary Figure S6c, Supplementary Table S4).
In contrast to the findings related to leptin, when analyzing the entire cohort, plasma adiponectin levels did not show significant differences between patients with severe obesity and the control group. Additionally, the correlation between adiponectin levels and body size measurements was weak (Figure 4a). However, circulating adiponectin levels were lower in men compared to women (Supplementary Figures S2 and S7a). When the data was separated by sex, there were no notable changes in the association with morphometric values. On the other side, metabolic comorbidities significantly affected patients with obesity; we observed that lower plasma adiponectin levels were associated with dyslipidemia, hypertension, and diabetes, regardless of sex (Figure 4b and Supplementary Figure S7b). Furthermore, liver damage had a significant impact, and lower plasma adiponectin levels could modestly predict the presence of MASH in patients (Supplementary Table S2). We also found significant associations between plasma adiponectin levels and steatosis, inflammation, and NAS (Fig. 4c and Supplementary Table S3) with only minor differences observed between the sexes (Supplementary Figure S7c, Supplementary Table S4).
2.7. Network Modeling and Insights into Patterns: The Functional Overlap Between FGF21 and Adiponectin
In patients with severe obesity, we often encounter varied data modalities. We used mixed graphical models to investigate the relationships between different organokines, liver damage, and other metabolic comorbidities. This approach provides a flexible framework for representing conditional dependencies (Figure 5a,b). Network analyses revealed that all organokines are interconnected, and clinical factors and histological features can complicate the interpretation of their interactions. Multivariate statistical techniques and regression models did not identify specific clusters of co-regulated organokines, and adjusted models did not clarify their potential use in diagnostics.
The disruption in communication is most noticeable in the context of liver damage. For example, the collective evaluation of organokines was unable to differentiate between MASH and non-MASH patients. The degree of liver damage has varying effects on circulating signals, indicating distinct metabolic profiles (Figure 5c-f). The principal component analysis did not effectively separate the groups, and the heatmap illustrates the complex clustering of patients based on circulating organokine levels. Similarly, the receiver operating characteristic curves show only slight variations in the already modest performance of models that include clinical data and organokine levels. However, the plot depicting variable importance in projection scores highlights the significant roles of FGF-21 and adiponectin in distinguishing the effects of liver lesions (Figure 5d).
This finding is significant because FGF21 and adiponectin exhibit functional similarities despite differences in their plasma concentrations. Our data suggest that an impaired FGF21–adiponectin axis may be crucial to developing MASH. In patients with MASH, circulating FGF21 levels were elevated, while plasma adiponectin concentrations were decreased compared to patients with a less severe form of MASLD. Additionally, we observed sex-based differences in these levels. The results regarding circulating FGF21 levels were consistent for both men and women; however, while there was a similar trend for circulating adiponectin levels, the differences did not reach statistical significance in either males or females (Fig. 6a). To clarify this relationship, we calculated the FGF21/adiponectin ratio and observed that patients with MASH had elevated values compared to those without it. These results were consistent across both males and females and demonstrated a modest predictive value for liver damage (Fig. 6b). (Figure 6b).
3. Discussion
Understanding the complex interplay between severe obesity, metabolic dysfunction, and liver disease is essential for developing effective diagnostic and therapeutic strategies. In this study, we examined an extensive series of patients with severe obesity and biopsy-proven liver disease, integrating clinical, histological, and biochemical data. By analyzing a comprehensive panel of circulating organokines and choline-related metabolites, we aimed to identify key signaling alterations that reflect or potentially drive hepatic and metabolic changes.
Severe obesity disrupts the homeostatic balance between major metabolic organs, including the liver, adipose tissue, skeletal muscle, and gut, partly through impaired organokine communication [6,10,22,23]. Our findings reinforce this concept, showing significant alterations in circulating levels of FGF19, FGF21, leptin, irisin, and adiponectin. Together, these data provide robust evidence that severe obesity is associated with a dysregulated organokine network and metabolic dysfunction, contributing to the development and progression of liver disease.
The liver serves as the primary line of defense against gut-derived substances, positioning it at the center of the interplay between intestinal signals and systemic metabolism. In severe obesity, excessive adiposity alters this gut-liver axis, potentially disrupting hepatocyte-mediated regulation of FGF19, an enterokine produced in the ileum in response to bile acid signaling. This relationship provides a strong rationale for continued research into how FGF19 influences bile acid metabolism and gut microbiota function in the context of obesity-related liver disease [15,16,17,18,24,25]. Our findings show that plasma levels of FGF19 were decreased in patients with severe obesity. This reduction is consistent with a downregulation of the farnesoid X receptor (FXR) in the gut, a key regulator of FGF19 expression. Given the role of FXR in bile acid homeostasis and metabolic signaling, these observations strengthen the rationale for exploring therapeutic strategies that target FXR pathways, including peroxisome proliferator-activated receptors and FGF19-based analogues [26,27,28].
Notably, the observed decrease in FGF19 levels was not directly associated with comorbidities. However, metabolic signals from the gut encompass more than protein hormones alone. In this context, our findings on choline and TMAO metabolism offer additional insight into gut-liver crosstalk in severe obesity. Despite the known variability and inconsistency in the literature [29], our data reveal a significant reduction in circulating choline and TMA levels compared to controls, with both correlating with liver fibrosis scores. Interestingly, plasma TMAO levels remained stable, which may reflect a compensatory hepatic response involving FGF19-regulated pathways, such as bile acid signaling and one-carbon metabolism [30]. Although FGF19 plays a well-established role in bile acid homeostasis and metabolic regulation, we did not observe a significant correlation between its plasma levels and the severity of liver injury in our cohort. This may appear counterintuitive, but it is possible that circulating FGF19 reflects intestinal FXR activity and gut-derived metabolic signals more than the degree of hepatic damage [26,27,28]. Moreover, in severe obesity, alterations in enterohepatic signaling or fibroblast growth factor receptor 4 responsiveness may occur independently of histological liver injury [31]. Our findings further support this interpretation: despite lower FGF19 levels, circulating TMAO concentrations remained unchanged, suggesting a compensatory hepatic adaptation in bile acid and choline metabolism [32]. Taken together, these data support the idea that low FGF19 levels are a feature of severe obesity, but not necessarily a marker of fibrosis severity.
Most circulating FGF21 is produced in the liver and affects other tissues through specific receptors [33,34]. In contrast to FGF19, plasma FGF21 concentrations were elevated in patients with severe obesity, especially in those with diabetes, and were strongly associated with liver damage. This finding complicates our understanding of the potential role of the gut-liver axis [11]. The elevated FGF21 levels observed in patients with MASH likely reflect a compensatory response to metabolic stress, underscoring its role as a key regulator in conditions of hepatic and systemic dysfunction. Recent reviews have examined the therapeutic effect of FGF21 analogues in clinical trials, highlighting novel variants such as aldafermin and pegozafermin. The benefits of these therapies for treating MASH by reducing hepatic fat can vary among patients [35,36]. Non-responders to treatment may include individuals with FGF21 resistance or with differences in hepatic FGF21 expression [37]. Our data indicate that such difference in expression is closely related to liver damage in severe obesity characterized by a high amount of fat deposited in adipose tissue and may contribute to the modest diagnostic accuracy for MASH observed in our cohort [18,38,39,40]. Moreover, the FGF21 to adiponectin ratio improves the interpretation.
Galectin-3, a β-galactose-binding lectin, is widely recognized as a key mediator of inflammation and fibrogenesis [41,42]. Circulating galectin-3 has also been proposed as a prognostic biomarker for hepatocellular carcinoma [43]. In our study, plasma galectin-3 levels were elevated in patients with severe obesity compared to those in the control group. However, an unexpected inverse correlation emerged between galectin-3 levels and liver fibrosis scores, which contrasts with its established pro-fibrotic role. One possible explanation is that galectin-3 levels are stage-dependent: while they contribute to fibrogenesis in early disease, they may plateau or decline in more advanced stages, when tissue remodeling slows. The concomitant increase in FGF21 and decrease in FGF19 further support a shift in hepatic stress and bile acid signaling. Indeed, FGF21 may exert anti-inflammatory effects, suppressing galectin-3 expression [33,34]. Additionally, chronic obesity may alter the function of fibroblasts or macrophages, thereby limiting galectin-3 production. These mechanisms may collectively explain the lower galectin-3 levels observed despite the progression of fibrosis. This interpretation, however, remains speculative. Notably, we confirmed galectin-3 expression in hepatic macrophages in our cohort, supporting its involvement in obesity-related metabolic inflammation. Ongoing clinical trials with Selvigaltin, a galectin-3 inhibitor, may provide further insight into its role in the pathogenesis and progression of MASH [44,45].
Circulating irisin, a myokine associated with muscle size and activity, was elevated in patients with severe obesity compared to controls. This finding is consistent with earlier research on individuals who are overweight or obese [46]; however, it is somewhat surprising given that these patients generally engage in low levels of physical activity. Like other organokines, irisin promotes energy expenditure by facilitating non-shivering thermogenesis [47]. Our results indicate a complex regulatory network in which organ systems respond differently to various organokines. We observed in patients with low plasma irisin levels a significant metabolic dysregulation, particularly dyslipidemia and hypertension, suggesting a regulatory role in severe obesity. Its contribution to liver pathology is unclear [38,48]. While circulating irisin levels did not correlate with the NAS score, we did find a positive correlation between irisin levels and ballooning in the liver.
Sex differences significantly influence the course of diseases. We have consistently observed variations in the prevalence of severe obesity and their responses to treatment between men and women. Fat distribution differs between the sexes, which is linked to metabolic health. Generally, men are more susceptible to metabolic comorbidities, resulting in a more pronounced clinical response to weight loss [49,50]. In our cohort, we specifically noted sexual dimorphism in the circulating adipokines leptin and adiponectin. Plasma leptin levels were significantly higher in patients with severe obesity and were closely correlated with BMI and waist circumference. These elevated circulating leptin levels support the concept of leptin resistance, which contributes to the chronic positive energy balance [51,52]. While leptin is a reliable marker of body fat, its role in liver pathology is not appreciated. Despite the importance of communication between adipose tissue and the liver [6], leptin did not correlate with MASLD or MASH in our cohort.
Our study found no significant differences in circulating adiponectin levels between patients with severe obesity and the control group. However, we did observe that plasma adiponectin concentrations were lower in men than in women, and the correlation between adiponectin levels and body size was weak. Additionally, low levels of adiponectin were significantly associated with metabolic dysfunction. In patients with MASH, circulating adiponectin levels were also significantly lower, which aligns with the role of adiponectin as an adipokine that protects against liver steatosis and fibrosis [14,53,54,55]. Our research revealed an inverse relationship between adiponectin levels and the severity of liver disease, emphasizing its potential as a protective factor. Therefore, lower levels of adiponectin may contribute to the progression of MASH. The therapeutic implications are significant, as interventions to increase adiponectin levels could potentially slow or prevent liver disease progression in obese individuals [56,57,58]. Interestingly, we identified a contrast between FGF21 and adiponectin concerning metabolic comorbidities. While circulating adiponectin levels were reduced in patients with MASH, plasma FGF21 levels were increased. This suggests that the imbalance in the FGF21-adiponectin axis is crucial for liver health. Conversely, it is also possible that liver disease itself contributes to the distortion of the FGF21-adiponectin axis.
Network analysis visualizes the complexity of organokine communication and the conditional dependencies associated with metabolic abnormalities and liver damage. Unfortunately, the multivariate analysis and regression models did not reveal any potential clusters of co-regulated organokines or predictive models that could distinguish between patients with and without MASH and could be proposed as useful biomarkers. However, addressing adipose tissue and liver interactions is important. Some data indicate the importance of the adipose tissue-liver axis in restoring liver homeostasis and the critical role of weight loss [38,39,58,59,60]. Recent treatments for MASH that depend on weight loss include peptide-based incretin therapies, which focus on glucagon-like peptide-1 and glucose-dependent insulinotropic polypeptides [61]. There are also promising drugs for MASH treatment that function independently of weight loss. These include FGF21 analogues, which increase adiponectin levels, and thyroid hormone receptor beta agonists like resmetirom [62,63]. However, clinical trials have not yet clarified the role of inter-organ communication, and drugs effective in alleviating liver disease often inadequately address concurrent metabolic morbidities, and vice versa [64]. We demonstrate relevant factors that can significantly impact the expression and function of organokines. While there are indications of interconnectedness among circulating organokines, this relationship is unclear in patients with severe obesity, a situation that could explain why most medications do not produce a consistent effect.
4. Strenghts and Limitations of the Study
This study provides a comprehensive analysis of circulating organokines and choline metabolites in a well-characterized cohort of patients with severe obesity undergoing bariatric surgery, with liver histology available for most cases. The simultaneous assessment of multiple signaling pathways allowed us to explore their associations with both metabolic traits and liver histological features. However, the cross-sectional design limits causal interpretation, and circulating levels may not fully reflect tissue-specific activity or receptor responsiveness. Additionally, our study cohort included predominantly female participants, reflecting the typical profile of patients undergoing bariatric surgery. This sex imbalance limits the generalizability of our findings and highlights the need to further explore sex-specific differences in organokine signaling and disease progression in future studies. Moreover, findings may not be generalizable to other populations, as lifestyle, genetic background, and microbiome composition can influence these molecular patterns. Consequently, some secondary findings should be interpreted as exploratory and warrant confirmation in independent cohorts.
5. Materials and Methods
5.1. Study Design
We conducted an observational case-control study involving 923 patients with severe obesity and biopsy-proven MASLD from the EOM cohort (ClinicalTrials.org reference number NCT05554224). All eligible patients met the inclusion criteria for bariatric surgery at the Hospital Universitari de Sant Joan in Reus, Spain. These criteria required participants to be over 18 years old and to have a BMI ≥35 kg/m². All patients underwent laparoscopic sleeve gastrectomy, during which liver biopsy samples were systematically obtained for histological assessment. We excluded individuals with clinical or analytical evidence of severe illness, chronic or acute inflammation, cancer or infectious diseases. Patients showing histologic patterns of hepatocellular damage suggestive of alcoholic disease were also excluded. The reference group comprised 258 non-obese individuals without liver disease and matched for age and ethnic background. These participants were drawn from a population study and follow-up conducted by the Institut d'Investigació Sanitària Pere Virgili in our area by the procedures outlined in the Declaration of Helsinki [65]. The study received approval from our institution's ethics committee (EPIMET PI21/00510_083, PL4NASH112/2021 and EOM 244/2024), and all participants provided written informed consent.
5.2. Sampling and Data Collection
We collected blood samples from all participants between 8:00 and 9:00 A.M., ensuring a 10-hour fasting period beforehand. The samples were processed within two hours and plasma and serum aliquots were stored at -80ºC until analysis. We followed conventional protocols and laboratory methods to obtain anthropometric measurements and to diagnose type 2 diabetes mellitus, hypertension, and dyslipidemia in all participants [10]. In patients, we examined liver biopsies obtained during surgery. We conducted laboratory analyses and ultrasonography for the control group to confirm the likely absence of significant liver disease.
5.3. Biochemical and Histologic Assessments
We obtained four-micron sections from liver tissues fixed in formalin and embedded in paraffin. We assessed steatosis, ballooning, and lobular inflammation on hematoxylin and eosin-stained sections following the validated NAS [38,66]. A re-review protocol determined that 27 biopsies lacked sufficient quality and were excluded from the analysis. As a result, histological analysis was conducted on 896 biopsies. Using a final score ranging between 0 and 8, we identified 340 patients with NAS <3 (non-MASH), 339 with NAS 3-4 (uncertain MASH), and 217 with NAS of 5 or greater (definite MASH). Fibrosis staging was performed on Masson's trichrome-stained sections, and in some cases, we used the SAF algorithm (steatosis, activity and fibrosis) for reassessment [10]. This categorization provides insights into the different manifestations of liver conditions. When necessary, we used immunochemistry as described [25]. We analyzed serum levels of commonly measured analytes, including glucose, insulin, triglycerides, total cholesterol, high-density lipoprotein cholesterol, low-density lipoprotein cholesterol, and aminotransferases, using a COBAS® 8000 automatic analyzer (Roche Diagnostics, Basel, Switzerland). Additionally, we measured the plasma concentrations of a selected panel of organokines that previously showed responses to nutritional states in an exploratory study [25]. This panel included FGF19, FGF21, galectin-3, leptin, adiponectin, and irisin, which were analyzed using enzyme-linked immunosorbent assay (ELISA) kits from R&D Systems (Minneapolis, MN, USA), following the manufacturer's instructions.
5.4. Mass Spectrometry for Selected Metabolites
We included the measurement of betaine, choline, TMA, and TMAO concentrations using liquid chromatography coupled to triple quadrupole mass spectrometry. Their association with organokines and their role in the metabolic crosstalk between the liver, gut, and adipose tissue has been described [29]. Briefly, 100 μL of plasma were mixed with 900 μL of methanol: water (8:2) and ten μL of internal standard (betaine-D9, choline-D9, TMAO-D9 at ten ppm each). Afterwards, the mix was incubated at -20ºC for 2 hours to precipitate proteins. Samples were centrifuged at 14,000 rpm for 10 minutes at 4ºC, and the supernatants were mixed with 75 μL of 50 mM tert-butyl bromoacetate in acetonitrile and ten μL of 70% ammonium hydroxide. Samples were vortexed for 1 min and incubated for 30 min at room temperature. We added 50 μL of 1% formic acid in acetonitrile, and samples were vortexed and centrifuged again at 14,000 rpm for 5 minutes at 4ºC. Supernatants were collected into chromatographic vials for subsequent analysis. Samples (two μL) were injected into an Ultra-High-Performance Liquid Chromatography (UHPLC) 1290 Infinity II Series instrument (Agilent, Santa Clara, CA, USA), coupled to a triple quadrupole equipped with an electrospray ionization source QqQ/MS 6470 Series (Agilent) operating in positive-ion mode. The chromatographic column for the UHPLC was an ACQUITY UPLC BEH HILIC 1.7 μm, 2.1 x 100 mm (Waters, Milford, MA, USA). The mobile phase consisted of A: acetonitrile: water (1:9) and B: acetonitrile: water (9:1) in 10 mM ammonium formate and 0.125% formic acid. The gradient consisted of 0% A for 1 minute, 10% A at 4 minutes, and 55% for 5 minutes at a constant flow rate of 0.5 mL/minute. Instrument conditions included a nebulizer gas pressure of 50 psi, a gas temperature of 300°C, a gas flow rate of 11 L/min, a sheath gas temperature of 400°C, a sheath gas flow rate of 12 L/min, a capillary voltage of 2,500 V, and a nozzle voltage of 500 V. Each metabolite was quantified by correcting the peak areas with their corresponding internal standard and calibration curve using the MassHunter Workstation Quantitative Analysis B.10.0 software (Agilent).
5.5. Statistical Analyses
Statistical analyses were performed using RStudio (R version 4.0.2). The Shapiro–Wilk normality test was used to assess the normality of the variables. Since some variables did not present normal distributions, non-parametric methods were used to ensure consistency. Two- and multi-group comparisons were performed using the Mann–Whitney U and Kruskal–Wallis tests. For categorical comparisons, we used the Fisher Exact test. The “Readxl” and “dplyr” packages were used for data management in R. Descriptive statistics, including participants’ characteristics, were generated with the “Tableone” package, with continuous variables summarized as medians and interquartile ranges and categorical variables as counts and percentages. Graphical resources were created using the “ggplot2”, “ggpur”, and “pROC” packages to obtain box and bar plots, correlation plots, and receiver operating characteristic curves. For ordinal logistic regression, we utilized the “ordinal” package. We used the R “mgm package” to implement and visualize mixed graphical models over continuous and discrete variables. Finally, the MetaboAnalyst 6R was used to obtain hierarchically clustered heatmaps and to run Partial Least Square Discriminant Analysis.
5. Conclusions
Providing personalized care for patients with severe obesity remains a challenge due to the heterogeneous response to treatment, although weight loss remains the most effective intervention. Our findings highlight the role of inter-organ crosstalk in the development of MASLD and other metabolic complications in these patients. We identified disruptions in organokine signaling, particularly involving adipose tissue and the liver. FGF21 and adiponectin emerged as key mediators, showing interconnected patterns and similar associations with metabolic health and liver disease. These results suggest that therapeutic strategies aimed at restoring the FGF21–adiponectin axis, together with weight reduction, may enhance the management of MASLD. Longitudinal studies could help clarify the temporal dynamics of this axis, though the need for sequential liver biopsies remains a significant limitation.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Histological features of liver biopsies of patients with severe obesity; Table S2: Clinical and laboratory characteristics according to liver damage distribution; Figure S1: Histopathologic diagnosis of metabolic dysfunction-associated steatohepatitis (MASH) in patients with severe obesity; Figure S2: Box-and-whisker plots illustrate the variability outside the upper and lower quartiles for the measured variables; Table S3: Ordinal logistic regression analysis summarizing the associations between organokines and metabolites with histopathology; Figure S3: Schematic representation of the gut-liver axis, illustrating the potential relationships among the fibroblast growth factor (FGF19), bile acids, microbiota composition, and choline metabolism; Figure S4: The relationship between circulating galectin-3 and comorbidities; Figure S5: The relationship between circulating irisin and comorbidities; Figure S6: Leptin levels in men and women. Table S4: The effect of sex on the associations between liver histologic features and plasma leptin and adiponectin levels; Figure S7: Adiponectin levels in men and women show notable differences.
Author Contributions
Conceptualization, H.C., J.C. and J.J.; methodology, H.C. and A.J.F.; software, H.C. and A.J.F.; validation, A.J.F. and J.C.; formal analysis, H.C. and A.J.F.; investigation, H.C., A.J.F., A.I.O., V.C.C., A.H.A., D.P., F.R. and A.Z.; resources, A.Z. and J.J.; data curation, H.C. and A.J.F. X.X.; writing—original draft preparation, H.C. and J.J.; writing—review and editing, J.C. and J.J.; visualization, J.C.; supervision, J.J.; project administration, J.C. and J.J.; funding acquisition, A.Z. and J.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by LA CAIXA FOUNDATION, Barcelona, Spain, under the project code HR21-00430, and by the INSTITUTO DE SALUD CARLOS III, Madrid, Spain, through the consecutive projects co-funded by the European Union, codes PI18/00921, PI21/00510, and PI24/01146.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Institutional Review Board (or Ethics Committee) of INSTITUT D’INVESTIGACIÓ PERE VIRGILI (protocol codes EPIMET PI21/00510_083, PL4NASH112/2021 and EOM 244/2024).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The datasets generated during and/or analyzed during the current study are available from the corresponding authors on reasonable request.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Abbreviations
The following abbreviations are used in this manuscript:
| BMI | Body mass index |
| ELISA | Enzyme-linked immunosorbent assay |
| FGF | Fibroblast growth factor |
| FXR | Farnesoid X receptor |
| MASLD | Metabolic dysfunction-associated steatotic liver disease |
| MASH | Metabolic dysfunction-associated steatohepatitis |
| NAS | Nonalcoholic fatty liver disease activity score |
| TMA | Trimethylamine |
| TMAO | Trimethylamine N-oxide |
| UHPLC | Ultra-High-Performance Liquid Chromatography |
References
- Blüher, M. Obesity: Global epidemiology and pathogenesis. Nat. Rev. Endocrinol. 2019, 15, 288–298. https://doi.org/10.1038/s41574-019-0176-8.
- Ellison-Barnes, A.; Johnson, S.; Gudzune, K. Trends in obesity prevalence among adults aged 18 through 25 years, 1976-2018. JAMA 2021, 326, 2073–2074. [CrossRef]
- Fryar, C.D.; Carroll, M.D.; Afful, J. Prevalence of overweight, obesity, and severe obesity among adults aged 20 and over: United States, 1960–1962 Through 2017–2018. https://www.cdc.gov/nchs/data/hestat/obesity-adult-17-18/obesity-adult.htm (Accessed 18 Jul 2025).
- Cabré, N.; Luciano-Mateo, F.; Baiges-Gayà, G.; Fernández-Arroyo, S.; Rodríguez-Tomàs, E.; Hernández-Aguilera, A.; París, M.; Sabench, F.; Del Castillo, D.; López-Miranda, J.; Menéndez, J.A.; Camps, J.; Joven, J. Plasma metabolic alterations in patients with severe obesity and non-alcoholic steatohepatitis. Aliment. Pharmacol. Ther. 2020, 51, 374–387. [CrossRef]
- Eslam, M.; Sanyal, A.J.; George, J.; International Consensus Panel. MAFLD: A consensus-driven proposed nomenclature for metabolic associated fatty liver disease. Gastroenterology 2020, 158, 1999–2014. [CrossRef]
- Azzu, V.; Vacca, M.; Virtue, S.; Allison, M.; Vidal-Puig, A. Adipose tissue-liver cross talk in the control of whole-body metabolism: Implications in nonalcoholic fatty liver disease. Gastroenterology 2020, 158, 1899–1912. [CrossRef]
- Lin, X.; Li, H. Obesity: Epidemiology, pathophysiology, and therapeutics. Front. Endocrinol. (Lausanne) 2021, 12, 706978. [CrossRef]
- Cypess, A.M. Reassessing human adipose tissue. New Engl. J. Med. 2022, 386, 768–779. [CrossRef]
- Chouchani, E.T.; Kajimura, S. Metabolic adaptation and maladaptation in adipose tissue. Nat. Metab. 2019, 1, 189–200. [CrossRef]
- Jiménez-Franco, A.; Castañé, H.; Martínez-Navidad, C., Placed-Gallego, C.; Hernández-Aguilera, A.; Fernández-Arroyo, S.; Samarra, I.; Canela-Capdevila, M.; Arenas, M.; Zorzano, A.; Hernández-Alvarez, M.I.; Castillo, D.D.; Paris, M.; Menendez, J.A.; Camps, J.; Joven, J. Metabolic adaptations in severe obesity: Insights from circulating oxylipins before and after weight loss. Clin. Nutr. 2024, 43, 246–258. [CrossRef]
- Tilg, H.; Adolph, T.E.; Trauner, M. Gut-liver axis: Pathophysiological concepts and clinical implications. Cell. Metab. 2022, 34, 1700–1718. https://. [CrossRef]
- Gérard, P. Gut microbiota and obesity. Cell. Mol. Life Sci. 2016, 73, 147–162. [CrossRef]
- Stern, J.H.; Rutkowski, J.M.; Scherer, P.E. Adiponectin, leptin, and fatty acids in the maintenance of metabolic homeostasis through adipose tissue crosstalk. Cell. Metab. 2016, 23, 770–784. [CrossRef]
- Jangam, T.C.; Desai, S.A.; Patel, V.P.; Pagare, N.B.; Raut, N.D. Exosomes as therapeutic and diagnostic tools: Advances, challenges, and future directions. Cell. Biochem. Biophys. 2025, Online ahead of print. [CrossRef]
- Jadhav, K.; Cohen, T.S. Can you trust your gut? Implicating a disrupted intestinal microbiome in the progression of NAFLD/NASH. Front. Endocrinol. 2020, 11, 592157. [CrossRef]
- Ferrell, J.M.; Dilts, M.; Pokhrel, S.; Stahl, Z.; Boehme, S.; Wang, X.; Chiang, J.Y.L. Fibroblast growth factor 19 alters bile acids to induce dysbiosis in mice with alcohol-induced liver disease. Cell. Mol. Gastroenterol. Hepatol. 2024, 18, 71–87. [CrossRef]
- Canyelles, M.; Tondo, M.; Cedó, L.; Farràs, M.;, Escolà-Gil, J.C.; Blanco-Vaca, F. Trimethylamine N-oxide: A link among diet, gut microbiota, gene regulation of liver and intestine cholesterol homeostasis and HDL function. Int. J. Mol. Sci. 2018, 19, 3228. [CrossRef]
- Camps, J.; Castañé, H.; Rodríguez-Tomàs, E.; Baiges-Gaya, G.; Hernández-Aguilera, A.; Arenas, M.; Iftimie, S.; Joven, J. On the role of paraoxonase-1 and chemokine ligand 2 (C-C motif) in metabolic alterations linked to inflammation and disease. A 2021 update. Biomolecules 2021, 11, 971. [CrossRef]
- Aderinto, N.; Olatunji, G.; Kokori, E.; Olaniyi, P.; Isarinade, T.; Yusuf, I.A. Recent advances in bariatric surgery: a narrative review of weight loss procedures. Ann. Med. Surg. (Lond). 2023, 85, 6091–6104. [CrossRef]
- Hernández-Alvarez, M.I.; Sebastián, D.; Vives, S.; Ivanova, S.; Bartoccioni, P.; Kakimoto, P.; Plana, N.; Veiga, S.R.; Hernández, V.; Vasconcelos, N.; Peddinti, G.; Adrover, A.; Jové, M.; Pamplona, R.; Gordaliza-Alaguero, I.; Calvo, E.; Cabré, N.; Castro, R.; Kuzmanic, A.; Boutant, M.; Sala, D.; Hyotylainen, T.; Orešič, M.; Fort, J.; Errasti-Murugarren, E.; Rodrígues, C.M.P.; Orozco, M.; Joven, J.; Cantó, C.; Palacin, M.; Fernández-Veledo, S.; Vendrell, J.; Zorzano, A. Deficient endoplasmic reticulum-mitochondrial phosphatidylserine transfer causes liver disease. Cell 2019, 177, 881–895.e17. [CrossRef]
- Naón, D.; Hernández-Alvarez, M.I.; Shinjo, S.; Wieczor, M.; Ivanova, S.; Martins de Brito, O.; Quintana, A.; Hidalgo, J.; Palacín, M.; Aparicio, P.; Castellanos, J.; Lores, L.; Sebastián, D.; Fernández-Veledo, S.; Vendrell, J.; Joven, J.; Orozco, M.; Zorzano, A.; Scorrano, L. Splice variants of mitofusin 2 shape the endoplasmic reticulum and tether it to mitochondria. Science 2023, 380, eadh9351. [CrossRef]
- Priest, C.; Tontonoz, P. Inter-organ cross-talk in metabolic syndrome. Nat. Metab. 2019, 1, 1177–1188. [CrossRef]
- Santos, J.P.M.D.; Maio, M.C.; Lemes, M.A.; Laurindo, L.F.; Haber, J.F.D.S.; Bechara, M.D.; Prado, P.S.D. Jr.; Rauen, E.C.; Costa, F.; Pereira, B.C.A.; Flato, U.A.P.; Goulart, R.A.; Chagas, E.F.B.; Barbalho, S.M. Non-alcoholic steatohepatitis (NASH) and organokines: What is now and what will be in the future. Int. J. Mol. Sci. 2022, 23, 498. https:/doi: 10.3390/ijms23010498.
- Ma, F.; Longo, M.; Meroni, M.; Bhattacharya, D.; Paolini, E.; Mughal, S.; Hussain, S.; Anand, S.K.; Gupta, N.; Zhu, Y.; Navarro-Corcuera, A.; Li, K.; Prakash, S.; Cogliati, B.; Wang, S.; Huang, X.; Wang, X.; Yurdagul, A. Jr.; Rom, O.; Wang, L.; Fried, S.K.; Dongiovanni, P.; Friedman, S.L.; Cai, B. EHBP1 suppresses liver fibrosis in metabolic dysfunction-associated steatohepatitis. Cell. Metab. 2025, 37, 1152–1170.e7. [CrossRef]
- Sharpton, S.R.; Schnabl, B.; Knight, R.; Loomba, R. Current concepts, opportunities, and challenges of gut microbiome-based personalized medicine in nonalcoholic fatty liver disease. Cell. Metab. 2021, 33, 21–32. [CrossRef]
- van de Wiel, S.M.W.; Bijsmans, I.T.G.W.; van Mil, S.W.C.; van de Graaf, S.F.J. Identification of FDA-approved drugs targeting the farnesoid X receptor. Sci. Rep. 2019, 9, 2193. [CrossRef]
- Wahlström, A.; Aydin, Ö.; Olsson, L.M.; Sjöland, W.; Henricsson, M.; Lundqvist, A.; Marschall, H.U.; Franken, R.; van de Laar, A.; Gerdes, V.; Meijnikman, A.S.; Hofsø, D.; Groen, A.K.; Hjelmesæth, J.; Nieuwdorp, M.; Bäckhed, F. Alterations in bile acid kinetics after bariatric surgery in patients with obesity with or without type 2 diabetes. EBioMedicine 2024, 106, 105265. [CrossRef]
- Barb, D.; Kalavalapalli, S.; Godinez Leiva, E.; Bril, F.; Huot-Marchand, P.; Dzen, L.; Rosenberg, J.T.; Junien, J.L.; Broqua, P.; Rocha, A.O.; Lomonaco, R.; Abitbol, J.L.; Cooreman, M.P.; Cusi, K. Pan-PPAR agonist lanifibranor improves insulin resistance and hepatic steatosis in patients with T2D and MASLD. J. Hepatol. 2025, 82, 979-991. [CrossRef]
- Chen, X.; Qiu, W.; Ma, X.; Ren, L.; Feng, M.; Hu, S.; Xue, C.; Chen, R. Roles and mechanisms of choline metabolism in nonalcoholic fatty liver disease and cancers. Front. Biosci. (Landmark Ed). 2024, 29, 182. [CrossRef]
- Schugar, R.C.; Gliniak, C.M.; Osborn, L.J.; Massey, W.; Sangwan, N.; Horak, A.; Banerjee, R.; Orabi, D.; Helsley, R.N.; Brown, A.L.; Burrows, A.; Finney, C.; Fung, K.K.; Allen, F.M.; Ferguson, D.; Gromovsky, A.D.; Neumann, C.; Cook, K.; McMillan, A.; Buffa, J.A.; Anderson, J.T.; Mehrabian, M.; Goudarzi, M.; Willard, B.; Mak, T.D.; Armstrong, A.R.; Swanson, G.; Keshavarzian, A.; Garcia-Garcia, J.C.; Wang, Z.; Lusis, A.J.; Hazen, S.L.; Brown, J.M. Gut microbe-targeted choline trimethylamine lyase inhibition improves obesity via rewiring of host circadian rhythms. Elife 2022, 11, e63998. [CrossRef]
- Ge, H.; Zhang, J.; Gong, Y.; Gupte, J.; Ye, J.; Weiszmann, J.; Samayoa, K.; Coberly, S.; Gardner, J.; Wang, H.; Corbin, T.; Chui, D.; Baribault, H.; Li, Y. Fibroblast growth factor receptor 4 (FGFR4) deficiency improves insulin resistance and glucose metabolism under diet-induced obesity conditions. J. Biol. Chem. 2014, 289, 30470–30480. [CrossRef]
- Zhou, D.; Zhang, J.; Xiao, C.; Mo, C.; Ding, B.S. Trimethylamine-N-oxide (TMAO) mediates the crosstalk between the gut microbiota and hepatic vascular niche to alleviate liver fibrosis in nonalcoholic steatohepatitis. Front. Immunol. 2022, 13, 964477. [CrossRef]
- Kharitonenkov, A.; Shiyanova, T.L.; Koester, A.; Ford, A.M.; Micanovic, R.; Galbreath, E.J.; Sandusky, G.E.; Hammond, L.J.; Moyers, J.S.; Owens, R.A.; Gromada, J.; Brozinick, J.T.; Hawkins, E.D.; Wroblewski, V.J.; Li, D.S.; Mehrbod, F.; Jaskunas, S.R.; Shanafelt, A.B. FGF-21 as a novel metabolic regulator. J. Clin. Invest. 2005, 115, 1627–1635. [CrossRef]
- Kliewer, S.A.; Mangelsdorf, D.J. A dozen years of discovery: Insights into the physiology and pharmacology of FGF21. Cell. Metab. 2019, 29, 246–253. [CrossRef]
- Koh, B.; Xiao, J.; Ng, C.H.; Law, M.; Gunalan, S.Z.; Danpanichkul, P.; Ramadoss, V.; Sim, B.K.L.; Tan, E.Y.; Teo, C.B.; Nah, B.; Teng, M.; Wijarnpreecha, K.; Seko, Y.; Lim, M.C.; Takahashi, H.; Nakajima, A.; Noureddin, M.; Muthiah, M.; Huang, D.Q.; Loomba, R. Comparative efficacy of pharmacologic therapies for MASH in reducing liver fat content: Systematic review and network meta-analysis. Hepatology 2024, Online ahead of print. [CrossRef]
- Harrison, S.A.; Rolph, T.; Knott, M.; Dubourg, J. FGF21 agonists: An emerging therapeutic for metabolic dysfunction-associated steatohepatitis and beyond. J. Hepatol. 2024, 81, 562–576. [CrossRef]
- Tang, S.; Borlak, J. A comparative genomic study across 396 liver biopsies provides deep insight into FGF21 mode of action as a therapeutic agent in metabolic dysfunction-associated steatotic liver disease. Clin. Transl. Med. 2025, 15, e70218. [CrossRef]
- Brunt, E.M.; Kleiner, D.E.; Wilson, L.A.; Belt, P.; Neuschwander-Tetri, B.A.; NASH Clinical Research Network (CRN). Nonalcoholic fatty liver disease (NAFLD) activity score and the histopathologic diagnosis in NAFLD: distinct clinicopathologic meanings. Hepatology 2011, 53, 810–820. [CrossRef]
- Cabré, N.; Luciano-Mateo, F.; Fernández-Arroyo, S.; Baiges-Gayà, G.; Hernández-Aguilera, A.; Fibla, M.; Fernández-Julià, R.; París, M.; Sabench, F.; Castillo, D.D.; Menéndez, J.A.; Camps, J.; Joven, J. Laparoscopic sleeve gastrectomy reverses non-alcoholic fatty liver disease modulating oxidative stress and inflammation. Metabolism 2019, 99, 81–89. [CrossRef]
- Chow, W.S.; Xu, A.; Woo, Y.C.; Tso, A.W.; Cheung, S.C.; Fong, C.H.; Tse, H.F.; Chau, M.T.; Cheung, B.M.; Lam, K.S. Serum fibroblast growth factor-21 levels are associated with carotid atherosclerosis independent of established cardiovascular risk factors. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 2454–2459. [CrossRef]
- Sciacchitano, S.; Lavra, L.; Morgante, A.; Ulivieri, A.; Magi, F.; De Francesco, G.P.; Bellotti, C.; Salehi, L.B.; Ricci, A. Galectin-3: One molecule for an alphabet of diseases, from A to Z. Int. J. Mol. Sci. 2018, 19, 379. [CrossRef]
- Jiang, Q.; Zhao, Q.; Li, P. Galectin-3 in metabolic disorders: mechanisms and therapeutic potential. Trends Mol. Med. 2015, 31, 424–437. [CrossRef]
- Chamseddine, S.; Yavuz, B.G.; Mohamed, Y.I.; Lee, S.S.; Yao, J.C.; Hu, Z.I.; LaPelusa, M.; Xiao, L.; Sun, R.; Morris, J.S.; Hatia, R.I.; Hassan, M.; Duda, D.G.; Diab, M.; Mohamed, A.; Nassar, A.; Amin, H.M.; Kaseb, A.O. Circulating galectin-3: A prognostic biomarker in hepatocellular carcinoma. J. Immunother. Precis. Oncol. 2024, 7, 255–262. [CrossRef]
- MacKinnon, A.C.; Humphries, D.C.; Herman, K.; Roper, J.A.; Holyer, I.; Mabbitt, J.; Mills, R.; Nilsson, U.J.; Leffler, H.; Pedersen, A.; Schambye, H.; Zetterberg, F.; Slack, R.J. Effect of GB1107, a novel galectin-3 inhibitor on pro-fibrotic signalling in the liver. Eur. J. Pharmacol. 2024, 985, 177077. [CrossRef]
- Comeglio, P.; Guarnieri, G.; Filippi, S.; Cellai, I.; Acciai, G.; Holyer, I.; Zetterberg, F.; Leffler, H.; Kahl-Knutson, B.; Sarchielli, E.; Morelli, A.; Maggi, M.; Slack, R.J.; Vignozzi, L. The galectin-3 inhibitor selvigaltin reduces liver inflammation and fibrosis in a high fat diet rabbit model of metabolic-associated steatohepatitis. Front. Pharmacol. 2024, 15, 1430109. [CrossRef]
- Jia, J.; Yu, F.; Wei, W.P.; Yang, P.; Zhang, R.; Sheng, Y.; Shi, YQ. Relationship between circulating irisin levels and overweight/obesity: A meta-analysis. World J. Clin. Cases 2019, 7, 1444–1455. [CrossRef]
- Takahashi, K.; Yamada, T.; Katagiri, H. Inter-organ communication involved in brown adipose tissue thermogenesis. Adv. Exp. Med. Biol. 2024, 1461, 161–175. [CrossRef]
- Shen, C.; Wu, K.; Ke, Y.; Zhang, Q.; Chen, S.; Li, Q.; Ruan, Y.; Yang, X.; Liu, S.; Hu, J. Circulating irisin levels in patients with MAFLD: an updated systematic review and meta-analysis. Front. Endocrinol. (Lausanne) 2024, 15, 1464951. [CrossRef]
- Mauvais-Jarvis, F.; Bairey Merz, N.; Barnes, P.J.; Brinton, R.D.; Carrero, J.J.; DeMeo, D.L.; De Vries, G.J.; Epperson, C.N.; Govindan, R.; Klein, S.L.; Lonardo, A.; Maki, P.M.; McCullough, L.D.; Regitz-Zagrosek, V.; Regensteiner, J.G.; Rubin, J.B.; Sandberg, K.; Suzuki, A. Sex and gender: modifiers of health, disease, and medicine. Lancet 2020, 396, 565–582. https.//doi: 10.1016/S0140-6736(20)31561-0.
- Friedman, J.; Halaas, J. Leptin and the regulation of body weight in Mammals. Nature 1998, 395, 763–770. [CrossRef]
- Obradovic, M.; Sudar-Milovanovic, E.; Soskic, S.; Essack, M.; Arya, S.; Stewart, A.J.; Gojobori, T.; Isenovic, E.R. Leptin and obesity: Role and clinical implication. Front. Endocrinol. (Lausanne) 2021, 12, 585887. [CrossRef]
- Maxim, M.; Soroceanu, R.P.; Vlăsceanu, V.I.; Platon, R.L.; Toader, M.; Miler, A.A.; Onofriescu, A.; Abdulan, I.M.; Ciuntu, B.M.; Balan, G.; Trofin, F.; Timofte, D.V. Dietary habits, obesity, and bariatric surgery: A review of impact and interventions. Nutrients 2025, 17, 474. [CrossRef]
- Wang, Z.; Scherer, P. Adiponectin, the past two decades. J. Mol. Cell. Biol. 2016, 8, 93–100. https://doi:10.1093/jmcb/mjw011.
- Buechler, C.; Wanninger, J.; Neumeier, M. Adiponectin, a key adipokine in obesity related liver diseases. World J. Gastroenterol. 2011, 17, 2801–2811. [CrossRef]
- Wang, H.; Zhang, H.; Zhang, Z.; Huang, B.; Cheng, X.; Wang, D.; la Gahu, Z.; Xue, Z.; Da, Y.; Li, D.; Yao, Z.; Gao, F.; Xu, A.; Zhang, R. Adiponectin-derived active peptide ADP355 exerts anti-inflammatory and anti-fibrotic activities in thioacetamide-induced liver injury. Sci. Rep. 2016, 6, 19445. [CrossRef]
- Hui, X.; Feng, T.: Liu, Q.; Gao, Y.; Xu, A. The FGF21-adiponectin axis in controlling energy and vascular homeostasis. J. Mol. Cell. Biol. 2016, 8, 110–119. [CrossRef]
- Zheng, Y.T.; Xiao, T.M.; Wu, C.X.; Cheng, J.Y.; Li, L.Y. Correlation of adiponectin gene polymorphisms rs266729 and rs3774261 with risk of nonalcoholic fatty liver disease: A systematic review and meta-analysis. Front. Endocrinol. (Lausanne) 2022, 13, 798417. [CrossRef]
- Patt, M.; Karkossa, I.; Krieg, L.; Massier, L.; Makki, K.; Tabei, S.; Karlas, T.; Dietrich, A.; Gericke, M.; Stumvoll, M.; Blüher, M.; von Bergen, M.; Schubert, K.; Kovacs, P.; Chakaroun, R.M. FGF21 and its underlying adipose tissue-liver axis inform cardiometabolic burden and improvement in obesity after metabolic surgery. EBioMedicine 2024, 110, 105458. [CrossRef]
- Jirapinyo, P.; McCarty, T.R.; Dolan, R.D.; Shah, R.; Thompson, C.C. Effect of endoscopic bariatric and metabolic therapies on nonalcoholic fatty liver disease: A systematic review and meta-analysis. Clin. Gastroenterol. Hepatol. 2022, 20, 511–524.e1. [CrossRef]
- Pais, R.; Aron-Wisnewsky, J.; Bedossa, P.; Ponnaiah, M.; Oppert, J.M.; Siksik, J.M.; Genser, L.; Charlotte, F.; Thabut, D.; Clement, K.; Ratziu, V. Persistence of severe liver fibrosis despite substantial weight loss with bariatric surgery. Hepatology 2022, 76, 456–468. [CrossRef]
- Loomba, R.; Hartman, M.L.; Lawitz, E.J.; Vuppalanchi, R.; Boursier, J.; Bugianesi, E.; Yoneda, M.; Behling, C.; Cummings, O.W.; Tang, Y.; Brouwers, B.; Robins, D.A.; Nikooie, A.; Bunck, M.C.; Haupt, A.; Sanyal, A.J.; SYNERGY-NASH Investigators. Tirzepatide for metabolic dysfunction-associated steatohepatitis with liver fibrosis. New Engl. J. Med. 2024, 391, 299–310. [CrossRef]
- Loomba, R.; Sanyal, A.J.; Kowdley, K.V.; Bhatt, D.L.; Alkhouri, N.; Frias, J.P.; Bedossa, P.; Harrison, S.A.; Lazas, D.; Barish, R.; Gottwald, M.D.; Feng, S.; Agollah, G.D.; Hartsfield, C.L.; Mansbach, H.; Margalit, M.; Abdelmalek, M.F. Randomized, controlled trial of the FGF21 analogue pegozafermin in NASH. New Engl. J. Med. 2023, 389, 998–1008. [CrossRef]
- Huang, D.Q.; Wong, V.W.S.; Rinella, M.E.; Boursier, J.; Lazarus, J.V.; Yki-Järvinen, H.; Loomba, R. Metabolic dysfunction-associated steatotic liver disease in adults. Nat. Rev. Dis. Primers 2025, 11, 14. [CrossRef]
- Huttasch, M.; Roden, M.; Kahl, S. Obesity and MASLD: Is weight loss the (only) key to treat metabolic liver disease? Metabolism 2024, 157, 155937. [CrossRef]
- Bertran, N.; Camps, J.; Fernandez-Ballart, J.; Arija, V.; Ferre, N.; Tous, M.; Simo, D.; Murphy, M.M.; Vilella, E.; Joven, J. Diet and lifestyle are associated with serum C-reactive protein concentrations in a population-based study. J. Lab. Clin. Med. 2005, 145, 41–46. [CrossRef]
- Kleiner, D.E.; Brunt, E.M.; Van Natta, M.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.C.; Torbenson, M.S.; Unalp-Arida, A.; Yeh, M.; McCullough, A.J.; Sanyal, A.J.; Nonalcoholic Steatohepatitis Clinical Research Network. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 2005, 41, 1313–1321. [CrossRef]
- Cabré, N.; Luciano-Mateo, F.; Chapski, D.J.; Baiges-Gaya, G., Fernández-Arroyo, S.; Hernández-Aguilera, A.; Castañé, H.; Rodríguez-Tomàs, E.; París, M.; Sabench, F.; Castillo, D.D.; Del Bas, J.M.; Tomé, M.; Bodineau, C.; Sola-García, A.; López-Miranda, J.; Martín-Montalvo, A.; Durán, R.V.; Vondriska, T.M.; Rosa-Garrido, M.; Camps, J.; Menéndez, J.A.; Joven,.
- J. Laparoscopic sleeve gastrectomy in patients with severe obesity restores adaptive responses leading to nonalcoholic steatohepatitis. Int. J. Mol. Sci. 2022, 23, 7830. [CrossRef]
Figure 1.
Evaluation of the gut-liver axis in severe obesity through fibroblast growth factor (FGF) 19 levels and choline metabolism markers. (a) Circulating levels of FGF19 were lower in individuals with severe obesity compared to controls and inversely correlated with body size. (b) There was no association between plasma FGF19 levels and metabolic comorbidities. (c) Choline and trimethylamine (TMA) concentrations were significantly lower in patients with obesity than in the control group, but progressively increased with fibrosis stages within the obese cohort. Statistical differences between groups were assessed using the Mann–Whitney U test. Correlations between variables were evaluated using Spearman’s rank correlation coefficient (Spearman’s ρ). ALT: Alanine aminotransferase; BMI: Body mass index; T2DM: Type 2 diabetes mellitus; TMAO: Trimethylamine N-oxide.
Figure 1.
Evaluation of the gut-liver axis in severe obesity through fibroblast growth factor (FGF) 19 levels and choline metabolism markers. (a) Circulating levels of FGF19 were lower in individuals with severe obesity compared to controls and inversely correlated with body size. (b) There was no association between plasma FGF19 levels and metabolic comorbidities. (c) Choline and trimethylamine (TMA) concentrations were significantly lower in patients with obesity than in the control group, but progressively increased with fibrosis stages within the obese cohort. Statistical differences between groups were assessed using the Mann–Whitney U test. Correlations between variables were evaluated using Spearman’s rank correlation coefficient (Spearman’s ρ). ALT: Alanine aminotransferase; BMI: Body mass index; T2DM: Type 2 diabetes mellitus; TMAO: Trimethylamine N-oxide.
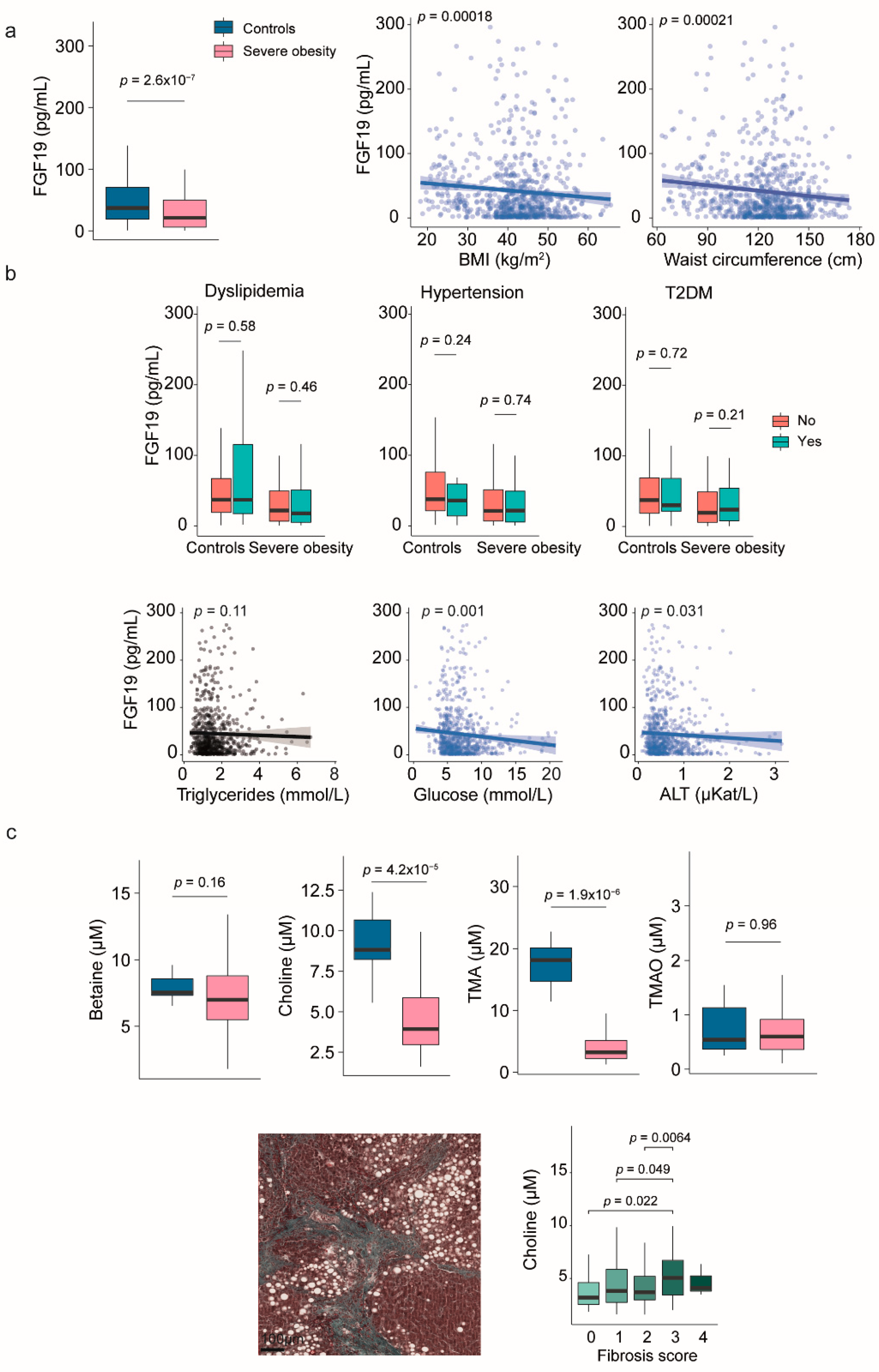
Figure 2.
The association of circulating fibroblast growth factor (FGF) 21 with hepatic histological features in severe obesity. (a) Circulating levels of FGF21 were higher in individuals with severe obesity than in the control group and showed a positive correlation with body size. (b) Box plots illustrate the association between FGF21 levels and diabetes. Scatter plots show the relationship with laboratory markers. (c) Box plots demonstrate the relationship between FGF21 levels, hepatic histological features, and predictive indicators of metabolic dysfunction-associated steatohepatitits (MASH). The receiver operating characteristic plot indicates that FGF21 is not an effective marker for distinguishing between patients with and without MASH. Statistical differences between groups were assessed using the Mann–Whitney U or the Kruskal-Wallis tests. Correlations between variables were evaluated using Spearman’s rank correlation coefficient (Spearman’s ρ). ALT: Alanine aminotransferase; AUC: Area under the curve; T2DM: Type 2 diabetes mellitus.
Figure 2.
The association of circulating fibroblast growth factor (FGF) 21 with hepatic histological features in severe obesity. (a) Circulating levels of FGF21 were higher in individuals with severe obesity than in the control group and showed a positive correlation with body size. (b) Box plots illustrate the association between FGF21 levels and diabetes. Scatter plots show the relationship with laboratory markers. (c) Box plots demonstrate the relationship between FGF21 levels, hepatic histological features, and predictive indicators of metabolic dysfunction-associated steatohepatitits (MASH). The receiver operating characteristic plot indicates that FGF21 is not an effective marker for distinguishing between patients with and without MASH. Statistical differences between groups were assessed using the Mann–Whitney U or the Kruskal-Wallis tests. Correlations between variables were evaluated using Spearman’s rank correlation coefficient (Spearman’s ρ). ALT: Alanine aminotransferase; AUC: Area under the curve; T2DM: Type 2 diabetes mellitus.
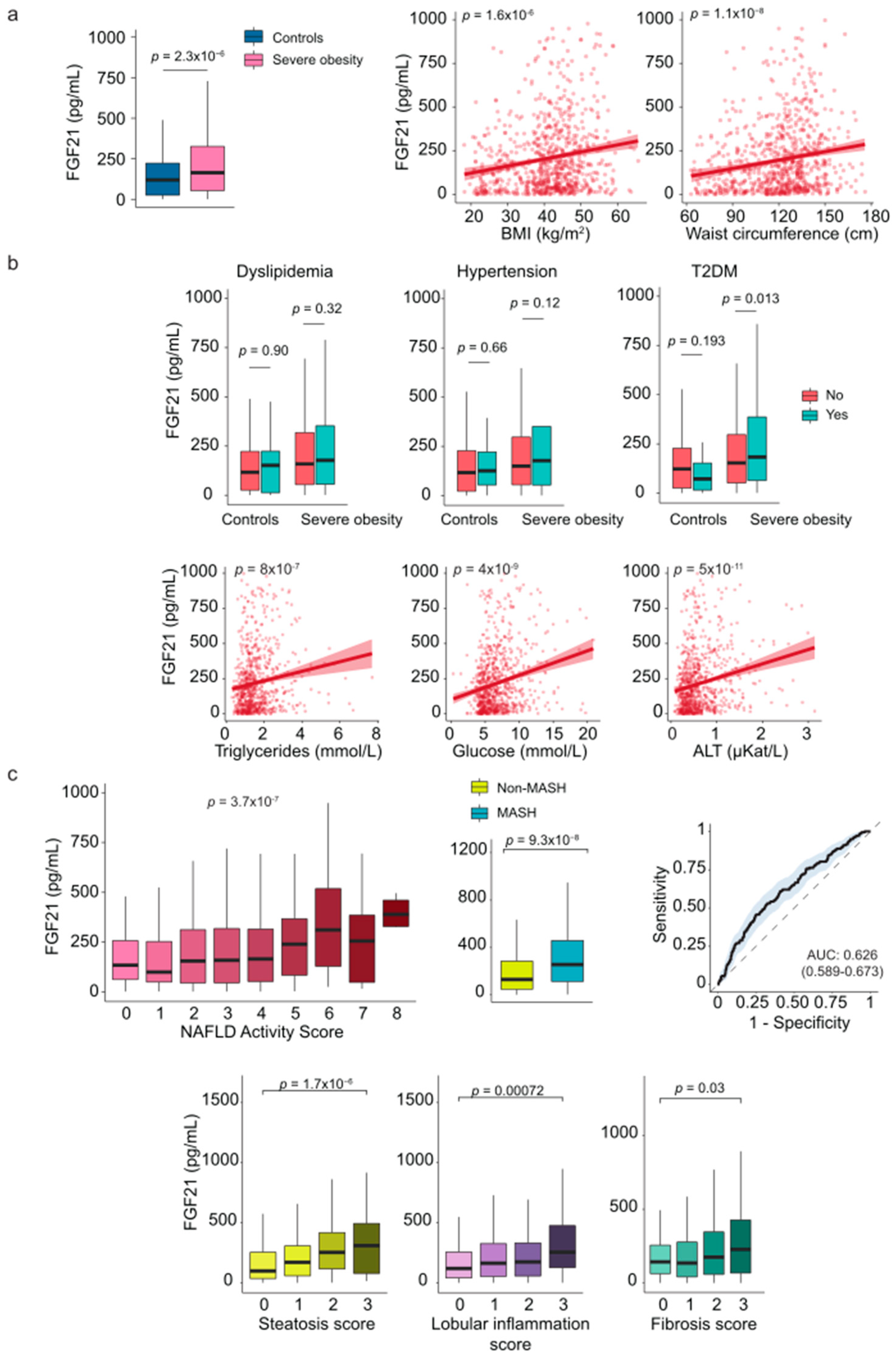
Figure 3.
The relationship between circulating leptin and comorbidities. (a) Patients with severe obesity exhibited higher circulating levels of leptin compared to controls, which positively correlated with anthropometric measurements. (b) Boxplots and scatter plots show the association between leptin levels and laboratory markers. (c) Leptin levels were associated with ballooning score. Statistical differences between groups were assessed using the Mann–Whitney U or the Kruskal-Wallis tests. Correlations between variables were evaluated using Spearman’s rank correlation coefficient (Spearman’s ρ). ALT: Alanine aminotransferase; BMI: Body mass index; MASH: Metabolic dysfunction-associated steatohepatitis; T2DM: Type 2 diabetes mellitus. .
Figure 3.
The relationship between circulating leptin and comorbidities. (a) Patients with severe obesity exhibited higher circulating levels of leptin compared to controls, which positively correlated with anthropometric measurements. (b) Boxplots and scatter plots show the association between leptin levels and laboratory markers. (c) Leptin levels were associated with ballooning score. Statistical differences between groups were assessed using the Mann–Whitney U or the Kruskal-Wallis tests. Correlations between variables were evaluated using Spearman’s rank correlation coefficient (Spearman’s ρ). ALT: Alanine aminotransferase; BMI: Body mass index; MASH: Metabolic dysfunction-associated steatohepatitis; T2DM: Type 2 diabetes mellitus. .
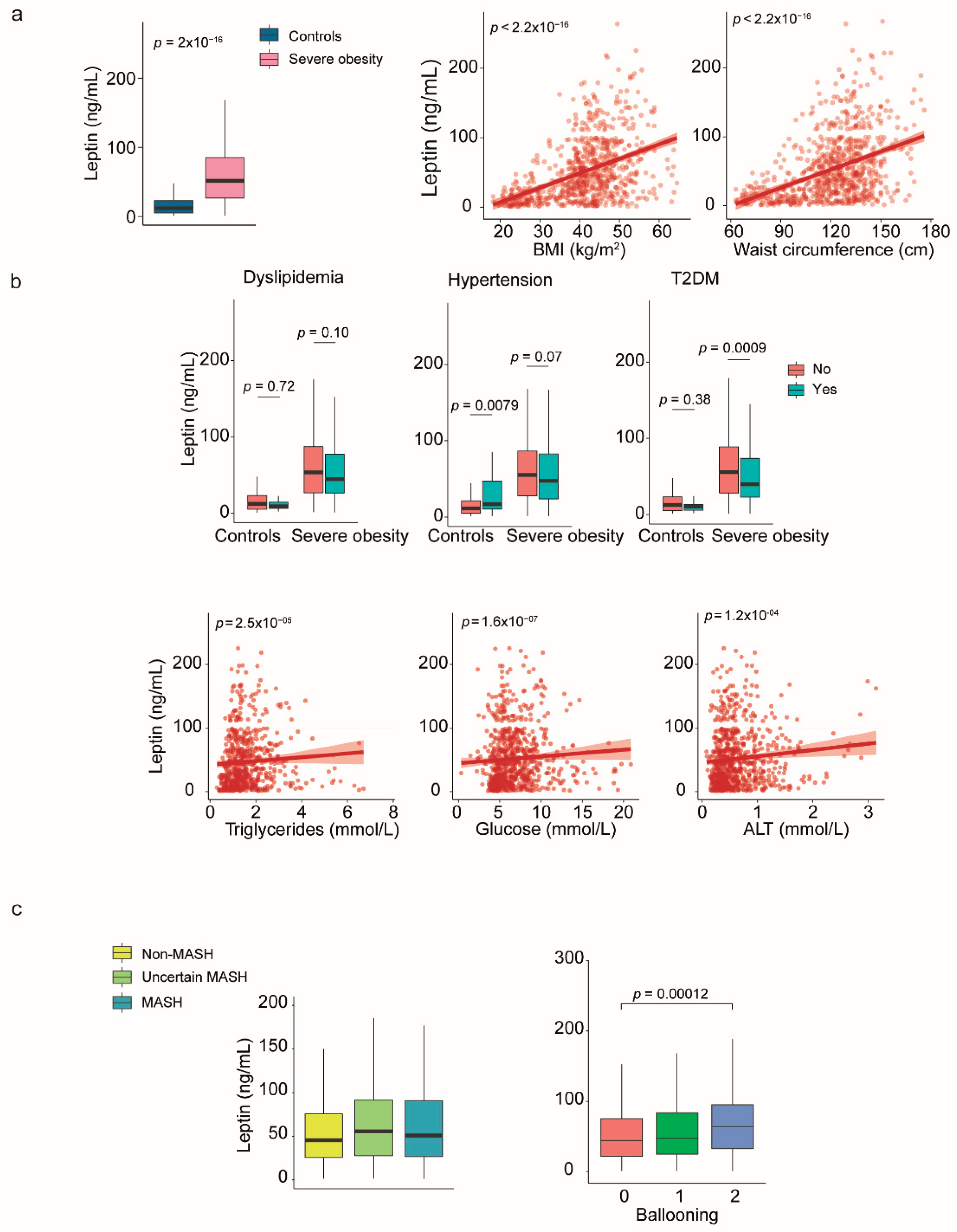
Figure 4.
The relationship between circulating adiponectin and comorbidities. (a) Plasma adiponectin levels were generally lower in patients with more severe conditions and showed little correlation with body size. (b) In contrast to healthy controls, low adiponectin levels were associated with dyslipidemia, hypertension, and diabetes. (c) Reduced adiponectin levels may serve as a marker to differentiate patients with MASH from those without. Statistical differences between groups were assessed using the Mann–Whitney U or the Kruskal-Wallis tests. Correlations between variables were evaluated using Spearman’s rank correlation coefficient (Spearman’s ρ). BMI: Body mass index; MASH: Metabolic dysfunction-associated steatohepatitis; NAFLD: Nonalcoholic fatty liver disease; T2DM: Type 2 diabetes mellitus. .
Figure 4.
The relationship between circulating adiponectin and comorbidities. (a) Plasma adiponectin levels were generally lower in patients with more severe conditions and showed little correlation with body size. (b) In contrast to healthy controls, low adiponectin levels were associated with dyslipidemia, hypertension, and diabetes. (c) Reduced adiponectin levels may serve as a marker to differentiate patients with MASH from those without. Statistical differences between groups were assessed using the Mann–Whitney U or the Kruskal-Wallis tests. Correlations between variables were evaluated using Spearman’s rank correlation coefficient (Spearman’s ρ). BMI: Body mass index; MASH: Metabolic dysfunction-associated steatohepatitis; NAFLD: Nonalcoholic fatty liver disease; T2DM: Type 2 diabetes mellitus. .
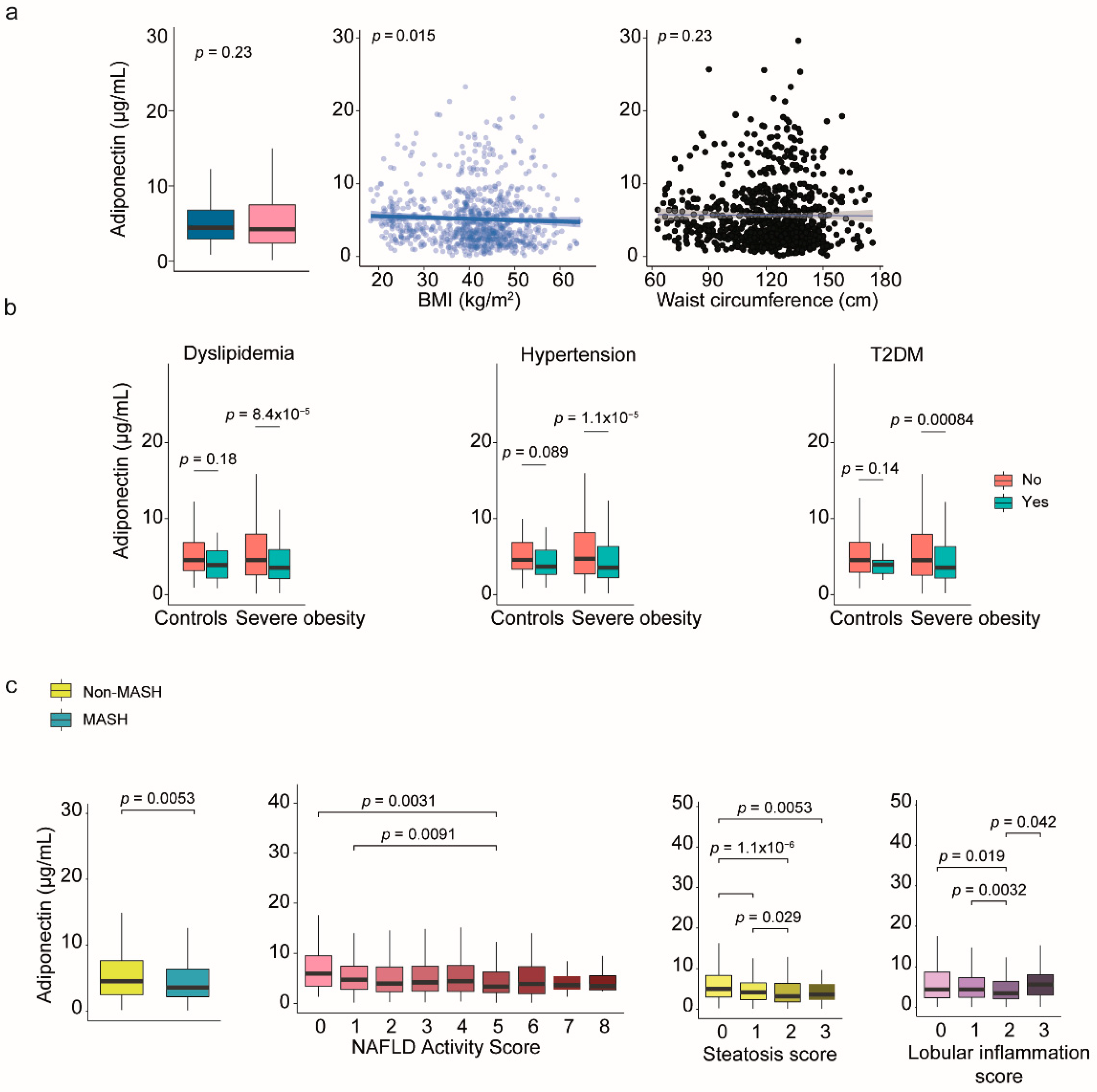
Figure 5.
Modeling the complexity of communication among organokines and clinical variables. (a) Mixed graphical models consist of variables represented as nodes, with edges indicating significant positive or negative correlations or interactions. (b) Probabilistic inferences highlight the conditional dependencies among these variables and highlight direction in relationships. (c) Principal component analysis did not reveal different patterns between patients with and without metabolic dysfunction-associated steatohepatitits (MASH). (d) The variable importance plot shows that the fibroblast growth factor (FGF) 21 and adiponectin were relevant factors in assessing MASH. (e) All classifier models demonstrated similar performance in receiver operating characteristic curves, with low areas under the curve (AUC). (f) However, analysis does not identify any potential clusters of co-regulated organokines.
Figure 5.
Modeling the complexity of communication among organokines and clinical variables. (a) Mixed graphical models consist of variables represented as nodes, with edges indicating significant positive or negative correlations or interactions. (b) Probabilistic inferences highlight the conditional dependencies among these variables and highlight direction in relationships. (c) Principal component analysis did not reveal different patterns between patients with and without metabolic dysfunction-associated steatohepatitits (MASH). (d) The variable importance plot shows that the fibroblast growth factor (FGF) 21 and adiponectin were relevant factors in assessing MASH. (e) All classifier models demonstrated similar performance in receiver operating characteristic curves, with low areas under the curve (AUC). (f) However, analysis does not identify any potential clusters of co-regulated organokines.
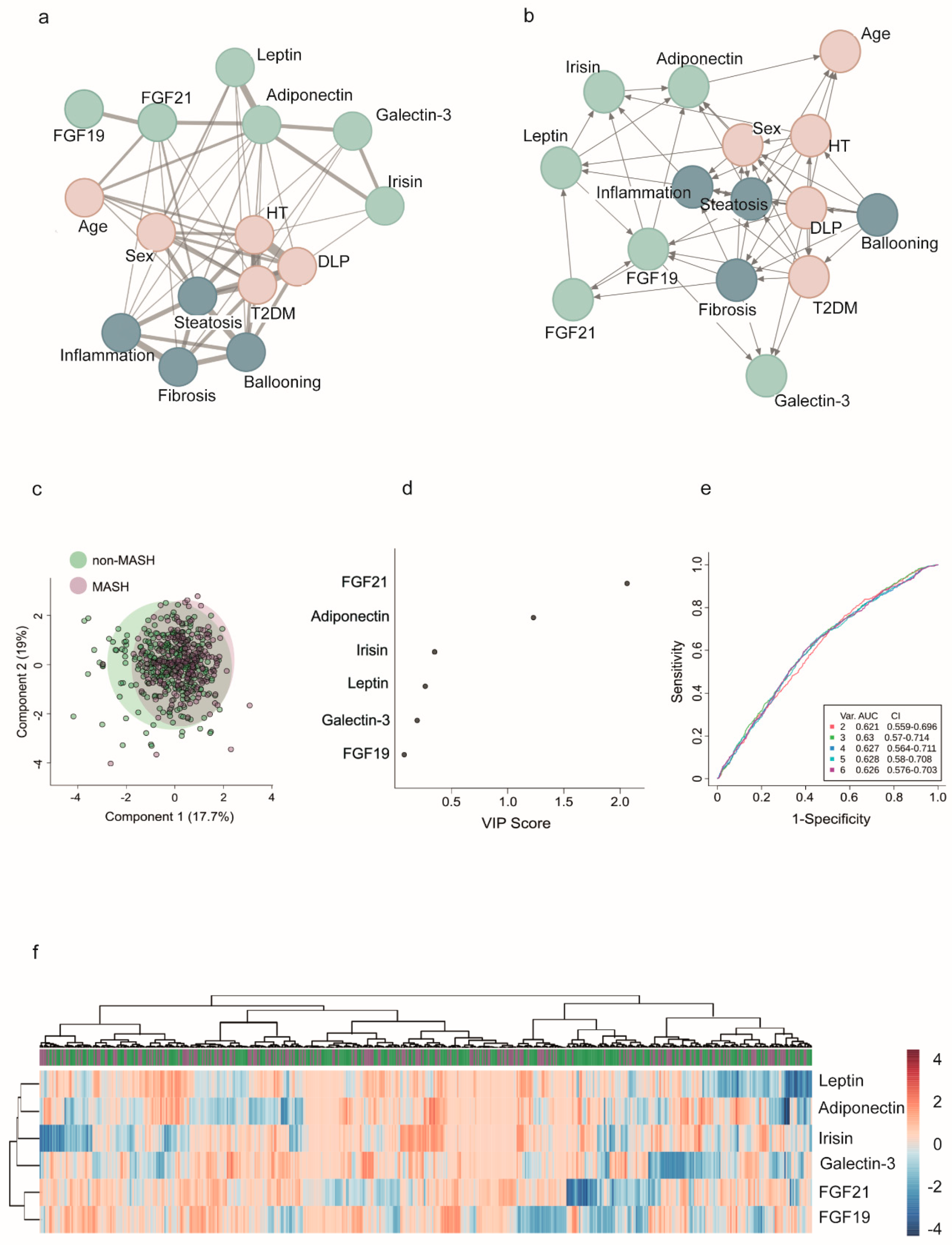
Figure 6.
Circulating fibroblast growth factor (FGF21) and adiponectin levels in patients with and without metabolic dysfunction-associated steatohepatitits (MASH). a) In patients with MASH, circulating FGF21 levels are increased, whereas plasma adiponectin concentrations are reduced. Sex influences results obtained for adiponectin. (b) This inconsistency disappeared when calculating the ratio between both hormones, which provided modest predictive value for distinguishing patients with MASH, as calculated by receiver operating characteristics curves. Statistical differences between groups were assessed using the Mann–Whitney U test. AUC: Area under the curve; NAS: Nonalcoholic fatty liver disease activity score.
Figure 6.
Circulating fibroblast growth factor (FGF21) and adiponectin levels in patients with and without metabolic dysfunction-associated steatohepatitits (MASH). a) In patients with MASH, circulating FGF21 levels are increased, whereas plasma adiponectin concentrations are reduced. Sex influences results obtained for adiponectin. (b) This inconsistency disappeared when calculating the ratio between both hormones, which provided modest predictive value for distinguishing patients with MASH, as calculated by receiver operating characteristics curves. Statistical differences between groups were assessed using the Mann–Whitney U test. AUC: Area under the curve; NAS: Nonalcoholic fatty liver disease activity score.
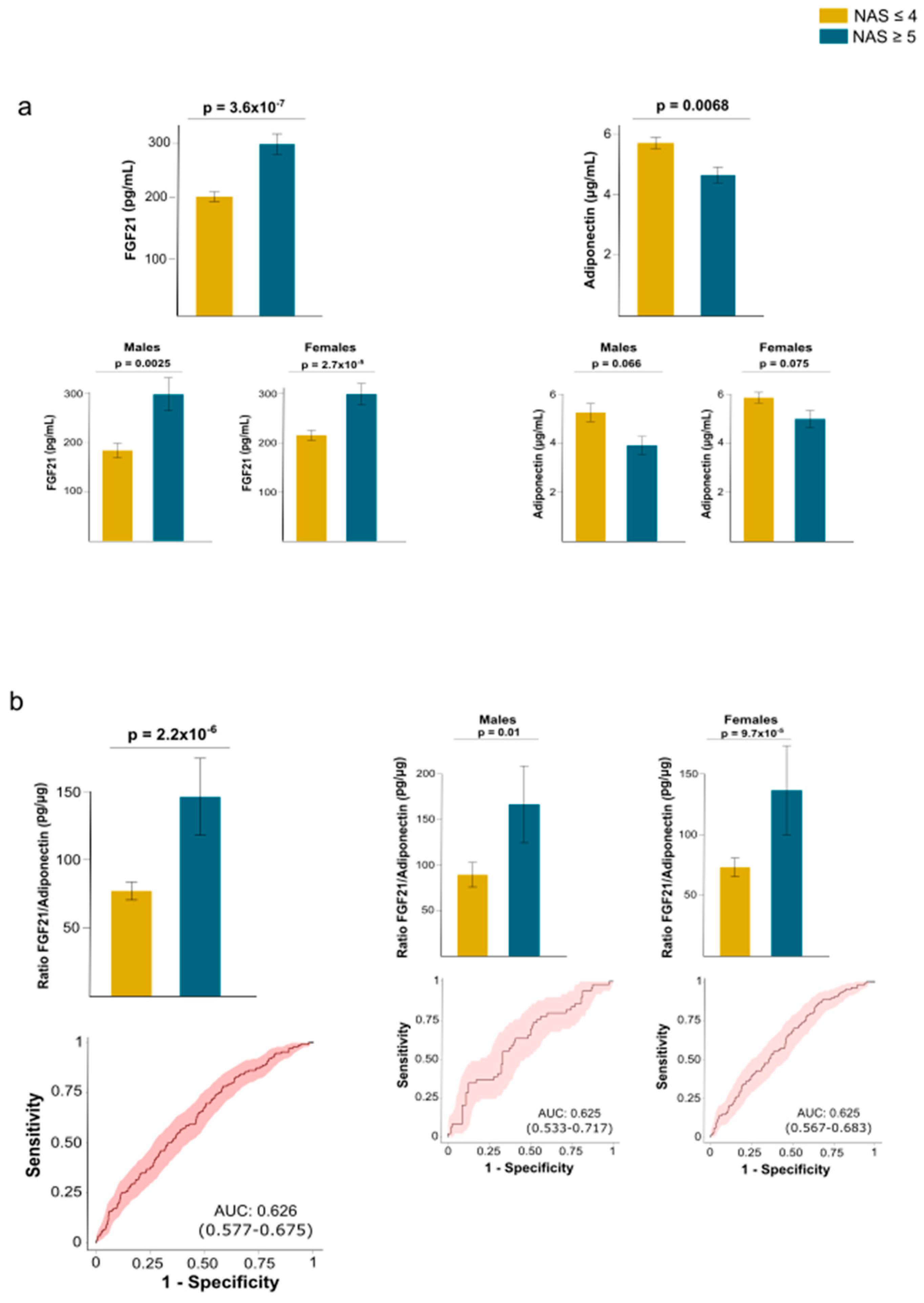

Table 1.
Demographic and clinical characteristics of participants.
| Variable |
Controls (n = 258) |
Severe obesity (n = 923) |
P-value |
| Women, n (%) | 121 (47.1) | 677 (73.4) | <0.001 |
| Age (years) | 45 (35 - 62) | 49 (41 - 56) | 0.347 |
| BMI (kg/m2) | 26.7 (23.3 - 29.8) | 43.9 (40.3 - 48.4) | <0.001 |
| Waist circumference (cm) | 89 (78 - 98) | 130 (120 - 139) | <0.001 |
| T2DM, n (%) | 18 (7.0) | 253 (27.4) | <0.001 |
| Hypertension, n (%) | 40 (15.5) | 406 (44.0) | <0.001 |
| Dyslipidemia, n (%) | 25 (9.7) | 234 (25.4) | <0.001 |
| Conventional biochemical variables | |||
| Glucose (mmol/L) | 4.7 (4.3 - 5.2) | 6.7 (5.5 - 8.5) | <0.001 |
| Insulin (pmol/L) | 47.0 (29.3 - 65.8) | 67.8 (37.7 - 109.5) | <0.001 |
| HOMA-IR | 1.4 (0.9 - 2.2) | 3.2 (1.7 - 5.7) | <0.001 |
| Triglycerides (mmol/L) | 1.0 (0.7 - 1.5) | 1.5 (1.2 - 2.0) | <0.001 |
| Cholesterol (mmol/L) | 5.2 (4.7 - 5.9) | 4.0 (3.5 - 4.7) | <0.001 |
| LDL (mmol/L) | 3.1 (2.6 - 3.8) | 2.4 (1.9 – 3.0) | <0.001 |
| HDL (mmol/L) | 1.4 (1.2 - 1.8) | 1.0 (0.8 - 1.2) | <0.001 |
| ALT (μKat/L) | 0.3 (0.2 - 0.4) | 0.6 (0.4 - 0.9) | <0.001 |
| AST (μKat/L) | 0.3 (0.3 - 0.4) | 0.6 (0.4 - 0.8) | <0.001 |
| GGT (μKat/L) | 0.2 (0.2 - 0.4) | 0.4 (0.2 - 0.6) | <0.001 |
| Organokines and metabolites | |||
| FGF19 (pg/mL) | 37.2 (19.0 – 70.7) | 21.3 (6.2 - 49.9) | <0.001 |
| Betaine (μM) | 7.5 (7.3 - 8.6) | 7.0 (5.5 - 8.8) | 0.163 |
| Choline (μM) | 8.8 (8.2 - 10.7) | 3.9 (3.0 - 5.9) | <0.001 |
| TMA (μM) | 18.1 (14.7 - 20.1) | 3.2 (2.2 - 5.2) | <0.001 |
| TMAO (μM) | 0.5 (0.4 - 1.1) | 0.6 (0.4 - 0.9) | 0.957 |
| FGF21 (pg/mL) | 119.7 (25.6 - 222.8) | 164.2 (54.5 - 326.1) | <0.001 |
| Galectin-3 (ng/mL) | 11.3 (6.2 - 17.3) | 12.7 (6.6 - 21.2) | 0.080 |
| Irisin (ng/mL) | 1.1 (0.6 - 1.6) | 1.5 (0.8 - 2.4) | <0.001 |
| Leptin (ng/mL) | 12.0 (5.4 - 23.0) | 51.6 (26.7 - 85.1) | <0.001 |
| Adiponectin (μg/mL) | 4.4 (2.9 - 6.8) | 4.2 (2.4 - 7.5) | 0.225 |
ALT: alanine aminotransferase; AST: aspartate aminotransferase; FGF: fibroblast growth factor; GGT: gamma-glutamyl transferase; HDL: high-density lipoprotein cholesterol; HOMA-IR: homeostatic model assessment for insulin resistance; LDL: low-density lipoprotein cholesterol; MASH: metabolic dysfunction-associated steatohepatitis; T2DM: type 2 diabetes mellitus; TMA: trimethylamine; TMAO: trimethylamine N-oxide.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



