Submitted:
18 July 2025
Posted:
21 July 2025
You are already at the latest version
Abstract
In the face of recent setbacks to coral reefs and coral reef restoration due to intensifying marine heat waves, climate smart strategies have been developed to accelerate natural processes of coral reef adaptation and recovery. In 2024 "Reefs of Hope" became the first coral-focused restoration and adaptation program to be endorsed by UNESCO as an Ocean Decade action. https://oceandecade.org/actions/reefs-of-hope/ This paper shares progress made, methods used, and translates the new paradigm into a national coral-focused adaptation plan for Fiji. The primary goal of a national coral-focused adaptation plan is to keep coral species alive despite increasingly lethal temperatures due to ocean warming, and in doing so to retain as much genetic diversity as possible within secure gene bank nurseries located in ideal reef locations, with a focus on heat-adapted bleaching resistant corals which are vital to adaptation. Secondary goals are to restore sexual reproduction to declining and rare coral species, and to support natural larval-based recovery and adaptation processes via creation of “regeneration patches” which enhance and restore natural recovery processes while facilitating the spread of heat adapted genetic diversity of both host and symbionts.
Keywords:
Facilitated recovery
; Coral reef restoration
; Assisted evolution
; Coral reef adaptation
; Reefs of Hope
; Bleaching resistance
; Regeneration patch
; Coral gene banking
Introduction
The Immense Value of Coral Reefs and Their Ongoing Loss
Coral reefs have grown over the eons, building expansive geographical features in tropical oceans, growing into extensive barrier reef, fringing reef, and atoll systems [1], often rising hundreds of meters from the depths. Coral reefs have created atoll nations [2] and have been uplifted to form raised limestone islands and coasts, building entire nations and parts of nations, with rocky cliffs, caves, and karst formations. Over millions of years, immense stores of carbon have been sequestered into coral skeletons, removing it from the atmosphere as essentially permanent storage and thus moderating the earth’s climate. Modern coral reefs, protect shorelines by absorbing waves [3] and generating coralline sand [4], helping to mitigate against changing sea levels, while providing food, livelihoods, and well-being to communities [5] throughout multiple countries in three oceans. Coral reefs have provided for island and coastal populations for thousands of years and have nurtured the development of diverse ethnic groups and nations which to this day continue to rely heavily on coral reefs for food security and sustenance.
The white sandy beaches, rich biodiversity, and stunning beauty of coral reefs have served as the backbone of the tourism industry [6] in multiple nations for over half a century, lifting the prosperity of these nations to formerly undreamt-of levels. However, the ocean is now warming rapidly due to climate change [7,8] and coral reefs globally have become the first marine ecosystem actively collapsing in the face of climate change, predicted to be mostly gone by 2050 [9]. The collapse of coral reefs and the extinction of coral species has immense and dire implications for the future of not only cultures and nations [10], but also for Planet Earth. While governments and NGOs have expended considerable efforts over the past decades to put coral reef conservation measures in place, setting aside no-take marine reserves and cleaning up coastal waters, these measures have proven ineffective in protecting coral reefs from collapse in the face of severe marine heat waves resulting from climate change [11]. Corals continue to bleach white in the heat and are dying in increasing numbers as temperatures become increasingly lethal [12]. Recurring marine heat waves threaten to wipe away decades of progress, killing off even the corals located on pristine reefs isolated from other man-made influences [13].
Buying Time for Coral Reefs
Even though an impending collapse of coral reefs in the face of rapid climate change is universally recognized as being imminent by the coral reef scientific community [14,15,16], there is no scientific consensus on what to do as far as the corals themselves are concerned [17,18]. The reality we face is that by the time society transitions to non-carbon energy sources, it is unlikely that much of anything will be left due to the rapid warming of the oceans. At this late point, the best we can hope for is to buy time for coral reefs by focusing on preventing the extinction of coral species. As heat-adapted individuals of most coral species have been found to survive unbleached at temperatures of 33–35°C and sometimes higher [19,20,21,22,23] these resilient corals have a good chance of surviving into the future, especially if relocated to cooler waters [24,25].
In discussions with colleagues in recent years, key points of agreement on active strategies to keep coral species alive into the future have emerged:
1. Further loss of coral genetic diversity must be prevented by securing what remains.
2. Sexual reproduction of rare and declining coral species must be restored and maintained.
3. Restoration efforts must focus on heat-adapted, bleaching resistant corals.
4. Coral focused work must support natural coral reef regeneration and adaptation.
These four points of agreement have been expanded to address the threats posed by ocean warming and to delay coral reef collapse, becoming part of a new coral-focused paradigm [13]. This “Reefs of Hope” paradigm was endorsed by UNESCO as an Ocean Decade Action in February 2024. https://oceandecade.org/actions/reefs-of-hope/ The present paper attempts to translate this Reefs of Hope model into a national coral-focused climate change adaptation plan, using available information on coral reefs to provide new lines of action designed to prevent coral death in the face of severe marine heat waves. The new active adaptation strategies have potential to reinforce and reinvigorate existing coral reef conservation efforts and MPAs, and as such represent new tools in the fight to keep coral reefs alive in the face of climate change, should conservationists and governments choose to move in this direction.
Ocean warming intensified suddenly in 2023 and is holding at >1.5°C above pre-industrial levels. https://climatereanalyzer.org/clim/sst_daily/ This has caused a new urgency to emerge: the need to rescue and secure heat-adapted corals from areas of the hottest waters before lethal temperatures arrive. This is based on data from the most impacted coral reefs of the Caribbean and South Pacific, where entire populations of corals of shallow lagoons and reef flats have died in the face of bleaching of an intensity that Fiji has not yet experienced [34], but which is certain to arrive at come point in the future. Many of these shallow stressed coral populations are situated in a “hot tub” thermal layer which forms at the surface during low tide on cloudless, windless days. These corals commonly withstand temperatures of 34-37°C without bleaching. However, at the present rate of ocean warming, these areas can be expected to reach lethal temperatures of >38-40°C in the coming years.
While some may insist that more information is needed before developing plans involving coral rescue and precautionary gene banking, the situation with coral reefs has become so dire with the recent spike in ocean heat that actions to save corals from approaching marine heat waves are needed urgently. Such precautionary actions were carried out in Florida in 2023 during a severe record-breaking marine heat wave, moving thousands of corals of many species and genotypes into land-based facilities successfully, saving them from heat stress and mass bleaching, while most of what was left in-situ on Florida’s coral reefs bleached white and died [26,27,28,29]. However just to the south in the Caribbean, in country after country such rescue measures were not possible and the intense heat waves of 2023 and 2024 caused mass mortality events with local-scale extinction of coral species, particularly devastated were the Acropora species [30,31,32,33]. The ocean temperature spike of 2023 then spilled into the Pacific at the end of the year, causing extreme extinction-level bleaching and massive coral die-offs in Kiribati and Tuvalu in 2024, and in PNG in 2025 (predicted by NOAA data and confirmed by observations), with major bleaching and coral loss in most Pacific Island nations. Fiji’s first back-to-back bleaching occurring in 2023 and 2024 as well, extending into 2025 in some sites. The record-breaking global bleaching event resulted in 84% of reefs impacted by heat stress since January 2023 in the fourth and largest coral bleaching event on record, impacting reefs across at least 83 countries and territories. The event is ongoing, with bleaching alerts continuing to develop across the world (ICRI and NOAA press releases, 2025).
Despite the endgame for coral reefs approaching, with some reefs already experiencing local extinction of formerly abundant coral species [34], no coral focused adaptation strategies have yet been approved and set in place for any nation. This paper will attempt to do this for the first time in history, with Fiji as the model, translating the strategies described in the UNESCO-endorsed Reefs of Hope Ocean Decade Action into a national coral-focused climate change adaptation plan to prevent coral species extinction in the face of rapid climate change.
The Need for National Coral-Focused Adaptation Plans
Existing strategies and government programs do a good job in many countries at addressing pollution, controlling negative tourism impacts, and preventing overfishing on coral reefs; however, none of these programs actively intervene to keep declining coral species alive and ecologically intact in the face of increasing ocean temperatures.
Although most coral restoration efforts failed in the recent mass bleaching events, much progress has been made in developing effective coral propagation and out-planting methods, which now can be re-purposed to create intensive coral-focused adaptation strategies along the lines of Reefs of Hope. Coral-focused actions have the added advantage of being nested within and supporting existing national coral reef conservation strategies, serving as mutual reinforcement, as well as insuring against loss due to future marine heat waves and associated severe mass bleaching and mortality.
The Reefs of Hope Ocean Decade Action strategies are being facilitated by the NGO Corals for Conservation and serve as the foundation for the present document. The diverse strategies were developed based on lessons learned at the leading edge of coral reef collapse, where extinction-level extreme marine heat waves had previously hit areas of the South Pacific. These UNESCO-endorsed nature-based solutions are based on forecasting future impacts and intervening to buy time for coral reefs. As global temperatures have reached >1.5°C above pre-industrial levels and continue to rise quickly [35,36], it is vital that we plan for what is coming and begin acting to secure what remains, rather than simply hoping for the best.
Promoting Coral-Focused Adaptation
The development and implementation of national-level coral-focused adaptation plans along the lines of what is being proposed by the Reefs of Hope Ocean Decade Action can best be implemented as a series of small trials, each monitored and evaluated before upscaling. In this way action plans and strategies are precautionary and can be adjusted as new information and results emerge. Only by adopting such a learn-as-you-go adaptive management approach can successful directions be established and rapid progress made. In this way, if a working hypothesis is wrong, it soon becomes apparent, and adjustments can be made. Reefs of Hope strategies can in this manner be refined and adjusted based on the realities of each reef system and country, likely differing in emphasis between severely impacted coral reefs versus coral reefs that remain largely intact.
Continuing to wait for more information before acting carries significant risk to coral survival; just as a boat can’t be steered while stopped and idling, only by engaging the engine and moving forward (or backward) can the boat be steered away from peril. Through movement the location of the vessel becomes known so that its direction can be adjusted. Likewise, only by acting on new strategies and working hypotheses of coral-focused adaptation can they be verified and modified for diverse situations, helping steer a course that prevents the loss of heat adapted coral and symbiont genetic diversity. The goal is to move forward with new strategies designed to reboot natural recovery processes and facilitate adaptation, while at the same time strengthening the functioning of existing MPAs and coral reef conservation programs. With few exceptions, corals are presently being left to die both during and after severe marine heat waves. Heat-adapted corals which are so important to future coral reef adaptation and survival are subsequently being eliminated by coral predators (e.g., Drupella and COTS) in the face of vastly skewed predator to prey ratios, undermining adaptation potential [37,38].
Mainstreaming ROH Strategies into Existing Programs and Policies
For coral-focused adaptation plans to be mainstreamed, they should strive to contribute to existing government policies and plans, becoming nested under existing National Adaptation Plans, adding coral-focused interventions and strategies designed to actively secure coral species, to promote bleaching resistance, and to restore active reproduction and recruitment in declining coral species to halt further coral species decline. The most basic coral-focused adaptation measure that is already in place in some conservation areas is COTS removal and control. If predator impacts are not part of existing management strategies, this is where to start for many reef systems.
SPREP, the South Pacific Regional Environmental Programme, an intergovernmental organization with the mandate of guiding governments of the South Pacific region, has developed the Pacific Coral Reef Action Plan which includes addressing climate change and coral-focused adaptation. The plan has been endorsed as a UNESCO Ocean Decade project along with Reefs of Hope and both are nested under the South Pacific Community’s Ocean Decade Programme: Pacific Solutions to Save Our Ocean. https://oceandecade.org/actions/pacific-solutions-to-save-our-ocean/
Ministries of Environment and other government agencies of most nations have developed action plans to reduce biodiversity loss on land and sea due to various and diverse stressors via National Biodiversity Strategy and Action Plans, which emphasize the integration of biodiversity values into national accounts, encouraging participation of government, businesses, and stakeholders at all levels. Among the accomplishments are hundreds of MPAs established to safeguard coral reef habitats, as well as reef-safe regulations for the tourism industry, with restrictions on fishing, anchoring, and other potentially harmful activities. National Adaptation Plans (NAP) have also been developed in partnership with the UN Environment Programme (UNEP) and are designed to enhance climate resilience.
In Fiji, the focus of coral reef management is within the context of Locally Managed Marine Areas (LMMAs). The restoration of the customary practice of setting aside "tabu areas" (MPAs) for the rehabilitation of overfished resources, traditionally widespread but discontinued for several generations, was first reestablished in Cuvu District in 2001, with the creation of four tabu areas [39]. From this initial work and other community-led efforts, a national movement emerged, resulting in over 300 tabu areas (FLMMA, pers. comm.) within Fiji's LMMA network, most functioning as rotational no-take zones [40,41]. While this represents a major accomplishment in community-based resource management, climate change now threatens to erase much of these gains, as coral mortality from increasingly severe marine heatwaves degrades essential fisheries habitat [14,42]. Reinforcing LMMAs with heat-adapted corals offers a promising emerging strategy to mitigate these impacts and support ecological and social resilience [13,18,23].
Communities and resorts in Fiji have shown great interest in planting corals as a conservation measure [43], but unfortunately most coral work has failed due to the random selection of corals for the work. However, coral reef adaptation plans for Fiji can take advantage of this receptivity and widespread grassroots energy by focusing on capacity building to reinforce existing community-based and resort-based coral reef conservation efforts. The goal must be to empower reef-dependent communities and tourism businesses on the front line of coral reef collapse and whose livelihoods are severely impacted by coral loss, to build resilience within their MPAs and areas of operation.
Background Important to Developing an Adaptation Plan for Fiji’s Coral Reefs
Fiji has the most extensive coral reefs of any island group in the Pacific Ocean, with over 10,000 km2 of coral reefs (Figure 1). Fiji’s reefs contain some 300 species of coral and are home to over 475 mollusc and 2,000 fish species (WWF 2021). Connectivity between the reefs of Fiji appears is very high [44], as indicated by fish genetics studies and by the rapid recovery rates after disturbance, which is dependent on intact breeding populations of corals at up current reefs. Fiji’s shallow, leeward reefs have shown notable resilience and recovery following mass bleaching disturbances [45], and cyclones [46]. Thus far the coral reefs of Fiji retain good species diversity, functionality, and coral cover despite multiple challenges which include land-based runoff and associated siltation, ecological imbalances due to overfishing, COTS plagues, and mass coral bleaching [47,48].
These findings underscore the importance of connectivity and larval supply in sustaining reef resilience. The prevailing southeast trade winds and corresponding westward-flowing surface currents play a key role in transporting coral larvae from the more pristine reefs of the Lau Group toward the more environmentally challenged reefs of Viti Levu and Vanua Levu [49,50]. However, during La Niña years, the reversal or weakening of trade winds and changes in regional oceanography can alter current directions [51], potentially sending coral larvae from the Great Sea Reef and western reef systems back toward Lau and the eastern islands. This latter pattern, while speculative, is based on long-term field observations and experience.
With global warming, mass coral bleaching has now become a problem for the coral reefs of Fiji, with the first mass coral bleaching resulting from a severe marine heat wave (>12 DHW, Condition 3), which struck the nation in 2000. Mass coral die-offs occurred, especially on the southern reefs of Fiji. Additional equally severe coral bleaching events leading to major mortality followed in 2014, 2023, and 2024, indicating an increasing frequency as the oceans warm. Five moderate, condition 2, (>8 DHW) bleaching events have also hit Fiji since records began in 1985, all after 2000 and with patchy impacts and moderate to low levels of mortality (Figure 2).
The maximum heat stress that Fiji has experienced has resulted in four major mass bleaching events, but this stress has been comparatively moderate thus far, condition 3 bleaching at 12-13 DHW. In comparison, most of the Caribbean has recently experienced stressful condition 5 bleaching of 21-26 DHW, while Christmas Atoll, Kiribati has experienced 27 DHW, over twice what Fiji has experienced (Figure 3). Figure 4 is a previously unpublished graph that compares bleaching stress levels and frequencies of the Pacific Islands nations against each other and to selected nations of the Caribbean. Clear trends are visible along the north-south transect from PNG through the various latitudinal regions of the Great Barrier Reef, and these same trends of greater heat stress on reefs closest to the equator are repeated in the Pacific Islands nations as well, with the exceptions being Tuvalu and the Solomon Islands, which have suffered less than expected for their latitudes, as well the most southerly reefs of the Austral Islands. The Caribbean region had similar trends up until 2023, when the entire region had a >20 DHW mass bleaching event.
Fiji’s Resilient Reefs in Peril
Because breeding coral populations continue to survive intact on many reefs of Fiji, the recovery of coral reefs has continued to proceed rapidly via larval-based processes. However, this resiliency depends on healthy breeding populations of corals in up current areas. Sexual reproduction often fails after a mass coral die-off event when the remaining colonies of a particular coral species become too few and far apart for effective fertilization to occur (generally >10M) termed the “alee effect” [52]. Once breeding populations of corals are no longer located up-current on Fiji’s reefs, larval-based recovery processes will fail at reef scale. The increased bleaching frequency now evident in Fiji will eventually begin to harm coral reproduction and thus slow coral recovery- if it hasn’t already. Bleaching is predicted to become an annual event in the coming years [53], threatening the larval-based recovery process, and with the recent back-to-back bleaching years, we appear to be drifting closer to that reality. However, while bleached and partially bleached corals may not produce viable coral larvae due to the great loss of energy resources due to the stress, bleaching resistant corals can continue to produce offspring effectively, assuming that we can secure these corals within reproductively viable, close-proximity patches [13].
An Ideal Crucible for the Evolution of Thermal Tolerance in Corals
While climate resilience is particularly high on Fiji’s coral reefs due to strong larval based connectivity, and the simple fact that the coral reefs cover such a broad extent: roughly 500km x 500km, provides added resilience because even major cyclones and bleaching events will spare many reefs, allowing breeding populations to persist.
Another possible factor in Fiji’s impressive resilience is the presence of extensive shallow heat-stressed reefs throughout the islands. Fiji has by far the greatest expanse of such shallow-water reefs in the Pacific Islands region, as indicated by surveying the region with Google Earth (Figure 5). The extensive shallow leeward reefs of the two main islands provide excellent potential for the evolutionary process of adaptation to heat stress.
A major driver of evolutionary selection within these wide reef flats and shallow nearshore waters is thought to be the hot surface layer, a "hot tub” thermal layer, which forms in the summer months during midday low tides, becoming especially hot on cloudless, windless days. These thermally stressed areas have undoubtedly been in existence for hundreds if not thousands of years, providing selection pressure for heat adaptation and bleaching resistance. Thermally resistant coral populations have been confirmed to exist in such chronically stressed shallow reef areas in many places globally [54,55,56]. Even the corals of adjacent deeper and cooler waters in these evolutionary hot spots are of heightened interest as they have a high chance of receiving the larvae of heat adapted corals, as genetically based heat resilience would naturally spill out of the hot pocket reefs and into the general coral population via spawning and subsequent larval transport and recruitment processes. Despite the potential importance of Fiji’s shallow hot pocket reefs to continued thermal resiliency and climate change adaptation, biological surveys of these shallow reefs have not yet been carried out.
Other smaller reef areas of known heat-adapted coral populations are scattered throughout Fiji, of particular note are the 500-1000 Meter wide fringing reefs of Fiji’s Coral Coast, where reef flats abut the land, forming elevated tidepools where an algal rim retains water well above MLW during low tide, creating ideal conditions for thermal heating at low tide, and with high temperatures >33C during January through April and >35C during marine heat waves (personal observations). These same reefs experience thermal stress from cold waters from June through September.
Bleaching resistance is the ability of a coral to maintain photosynthetic processes despite extreme temperatures. Photosynthetic integrity is more complex in corals than in macroalgae or flowering plants, as corals have the added complication of being a combination of species, and bleaching resilience can be based on either the host animal, the algal symbionts, symbiotic bacteria, or in combination [57,58,59]. Evolutionary selection acts on each component of the holobiont, which then impacts the whole [60,61]. However, the primary driver of resilience in corals is the algal component and is based on resistant algal species or clades (e.g., Durusdinium trenchii) [62,63,64]. However, certain coral hosts have been shown to be capable of maintaining even sensitive algae within their tissues, presumably by neutralizing the impact of free radicals which would otherwise burn the coral tissues [65,66].
While acclimation or stress hardening may be involved to a certain extent in the increase of bleaching resistance in corals [19,54,67]. This acquired resilience is limited and is not passed on to the next generation of the coral host animal via genetics. Evolutionary theory dictates that inheritable traits which give a survival advantage to a species are the result of natural selection among individuals over time. Natural selection in corals requires environmental stress, which inhibits the survival or reproduction of individuals with less desirable traits and selects for those individuals best able to survive, thrive, and pass on these traits to the next generation [68,69]. For corals to evolve a high degree of bleaching resistance, thermal stress levels beyond simple acclimation potential are required [65,70], resulting in lethal levels of stress to coral holobionts without genetic traits for thermal tolerance among the component members, weeding out poorly adapted individuals.
Heat-adapted corals from shallow heat-stressed reefs have been shown to retain their resilience [22,71] when translocated to cooler waters. However, these same corals have been shown to already be living close to their upper temperature threshold, and thus unable to adapt further to increasing temperatures due to rapid global warming [23,72]. This provides a rationale for human-assisted migration to rescue and secure these heat-adapted corals while restoring cooler degraded reefs with thermally resistant corals from these stressed reefs. Global studies have shown that the range in coral thermal tolerance across a single reef can be as large as differences observed across vast latitudinal gradients [19,21,55]. This indicates that long-distance translocation is not required to keep corals alive into the future and provides the rationale for the implementation of local-scale translocation of corals as a strategy to keep heat-adapted corals alive into the future as the ocean warms, assuming strong thermal gradients exist within a coral reef system [20,73].
Our operating assumption for the Reefs of Hope paradigm is that areas less than one meter deep at low tide have been the driving force behind the evolution of thermal resistance over the millennia, as this is where a “hot tub” thermal layer forms at low tide during summer months, reversing and becoming the coolest waters in the winter months [13,22,73]. However, with increasingly severe marine heat waves, these stressed reefs have begun to experience lethal levels of heat stress, putting entire coral communities in peril [34].
Coral breeding work, which has now become commonplace [24,74], focuses on the coral animal rather than the symbionts, and therefore to be effective must first select corals with host-derived bleaching resilience as broodstock [70,75]. Recent studies have found that heat-adapted corals are widespread in some coral reef systems [21,23], even in deeper less heat-stressed environments, only showing their resilience during mass bleaching events. It thus appears that while heat resilience evolves in the hot-stressed shallows, heat-adapted traits can spread throughout coral reefs via larval dispersal resulting from sexual reproduction within shallow heat-adapted coral populations [19,76].
As the larvae of spawning corals such as Acropora do not carry with them algal symbionts [77], nor have they been found to carry bacterial symbionts, it is the host genetics of these species that spreads with the larvae, not the symbiont genetics. Brooding coral species such as Pocillopora, on the other hand, carry algal [78] and bacterial components [79,80] best adapted to the conditions of the source reefs (hot pocket reefs); however, these symbionts may be mal adapted to their new environment [81,82]. Therefore, coral bleaching resilience in corals found in cooler or deeper waters is more strongly associated with host-based factors than symbiont-based factors [19,83,84], while in shallow-water environments, corals are more likely to be resilient based on heat-tolerant symbionts like Durusdinium, which play a greater role in thermal tolerance [20,63]. Understanding this tendency could help revolutionize the collection of corals for breeding purposes [13], with researchers targeting coral populations of cooler or deeper waters during mass bleaching events as a more effective way to seek out corals exhibiting host-derived resilience, while targeting larger/older coral colonies of hot pocket coral populations, even during non-bleaching seasons, to acquire heat-adapted algal and bacterial symbionts. During extreme heat stress these same shallow areas could be targeted to seek out bleaching resilient colonies which might contain multiple sources of resilience, both host-derived and symbiont-derived.
With rising temperatures already near the upper limit of adaptation potential, lethal temperatures are becoming problematic within the heat-stressed reefs [72,86]. Unfortunately, it has been difficult to convince the coral reef management community of this time-sensitive urgency; that the very coral populations where all sources of thermal resilience have evolved may soon be gone, initially transitioning away from Acropora dominance into lower diversity steady-state reefs dominated by only the most resilient genera.
The Reefs of Hope Ocean Decade Action solution seeks to systematically move jeopardized corals of the more sensitive genera as well as uncommon coral species from shallow areas of extreme thermal stress to cooler deeper waters. A depth of 1.5 meters or so deeper at low tide is generally enough to get the corals out of the hot thermal layer and into slightly cooler waters nearby, where maximum temperatures remain below 33 °C even during major stress events, so that the coral holobionts can escape increasingly lethal temperatures while remaining within the thermal regime where they evolved. As climate change unfolds over the coming decades, the expectation is that rising sea levels will cool off shallow hot pocket reefs via thermal buffering; increased circulation and depth, enabling conditions where these same corals could potentially be moved back to the reefs where they were originally collected.
Breakthroughs in Facilitating Natural Processes of Adaptation and Recovery
To facilitate natural coral reef adaptation and recovery processes we must understand the ecological basis of coral reef recovery and adaptation. However, coral reef restoration has rarely focused on facilitating natural processes, instead the focus has mainly been on the development of nursery and out-planting methods, and “up-scaling” has usually been based on the numbers of corals planted, using few coral species of few genetic strains, and with considerable ecological risks, such as creating genetic bottlenecks and highly altered species compositions [87]. Significant opposition and criticism have been levied against the field of coral reef restoration for these and other reasons. Following the recent death of most of the corals planted in the multi-million-dollar restoration efforts by the off-scale marine heat wave that hit Florida and the Caribbean in 2023–24, a re-think of restoration strategies is now occurring [13,88,89]. The emerging realization is that restoration as formerly practiced is no longer possible in the presence of recurring severe marine heat waves, just like planting trees becomes futile during drought conditions and in the face of an approaching firestorm. Efforts must now turn to securing coral genetic diversity and preventing its further loss [13,90,91].
The fundamental aspect of a coral-focused adaptation strategy is to prevent further loss of coral species and coral genetic diversity by mobilizing to collect and secure rare and declining coral species within gene bank nurseries located on coral reefs, with an emphasis on heat-adapted coral genotypes. In the process of securing corals from further decline, diverse genotypes are brought closely together, which in turn restores reproduction to declining and rare coral species via increased fertilization rates [13]. As an emphasis is placed on pre-adapted heat-resistant corals, inheritable traits conducive to bleaching resistance will likely be passed to some of the resulting larvae.
Saving heat adapted corals is an important first step in preventing coral species extinction; securing coral broodstock so that the natural processes of coral reproduction, recovery, and adaptation can be maintained and assisted [13]. Ensuring that larger numbers of heat adapted corals and individuals of increasingly rare species are relatively secure and restored to breeding condition will require moving significant numbers of jeopardized corals from the hottest coral reef areas to cooler reef areas [13]. And when bleaching hits, special efforts should be made to collect unbleached heat-adapted corals from both shallow and deeper environments to add to the nurseries, further increasing both symbiont and host derived bleaching resistance within the gene banks.
To maintain gene bank nurseries over the years as the corals grow, considerable trimming of corals must occur to prevent overgrowth and competitive interactions between the corals. These trimmed coral branches are then used to create “regeneration patches” in degraded reef areas, designed to maximize fish and invertebrate habitat for improved ecological functioning and to reboot natural larval-based recovery processes via creation of strong settlement signals which enhance coral larval settlement to the reef. The fastest growing corals will produce the most biomass for out planting, and thus might be considered over-replicated in the restoration efforts, however, they are the best corals for the job of patch creation due to their rapid growth and ideal habitat forming characteristics, and the initial low diversity patch (in theory) will subsequently attract coral larvae of many species, resulting in a highly diverse naturally structured reef patch over time [13]. Where possible, regeneration patches should be established within MPAs and linked to strategies addressing problems of overfishing, pollution, COTS over abundance, and other negative impacts on coral reefs. In this way, previous advances in coral reef conservation can build a strong foundation on which a coral-focused adaptation strategy can be built.
Materials and Methods
The primary goal of a national coral-focused adaptation plan is to keep coral species alive despite increasingly lethal temperatures due to ocean warming, and in doing so to retain as much genetic diversity as possible, with a focus on heat-adapted bleaching resistant corals which are vital to adaptation. Secondary goals are to restore sexual reproduction to declining and rare coral species, and to support natural recovery and adaptation processes while facilitating the retention and sharing of heat-resistant algal symbionts.
Reefs Hope Operational Strategy
- Remote sensing and field searches to locate potential shallow water heat-adapted coral populations as well as to locate ideal cooler-water nursery sites as nearby as possible.
-
A national coral-focused strategy must first seek out and collect heat-adapted corals of declining species on which to build a programme.
- -
- Collect heat-adapted coral stock of all declining species; 10 genotypes can retain approximately 50% of the original genetic diversity, while 35 genotypes can retain roughly 90% of coral genetic diversity within the species [92].
- -
- Two strategies are employed towards this aim, the first uses mass coral bleaching events as an opportunity to collect unbleached and thus heat-resistant corals, collected as the temperature begins to drop but before partially bleached corals regain their color.
- -
- A second strategy involves identifying heat adapted coral populations of the extreme shallows where corals are chronically exposed to high levels of thermal stress. Corals located in these “hot pocket” areas at the upper threshold of coral survival continually risk death from exposure during extreme low tides and during hot summer temperatures and high UV levels, especially during midday low tides on windless and cloudless days, worsening now due to ocean warming. Once these hot pocket reefs are identified, coral colonies of selected for relocating to cooler-water nurseries nearby, 1-2 meters deeper, to prevent lethal levels of heat stress as the oceans warm, as well as to protect the corals from COTS and other predators.
- -
- The goal is to keep the corals in the gene bank within a thermal stress regime similar to that which they have adapted to before the present climate crisis, to prevent the possibility of symbionts losing their heat resilience over time via local-scale adaptation, which could potentially happen over the decades if a nursery were in much cooler conditions.
- Carry out long-term monitoring by installing temperature loggers in collecting sites and nursery sites, as well as coral monitoring plots in the collecting sites and areas adjacent to nurseries to assess changes to the coral populations over time as the ocean warms, or conversely as hot nearshore waters potentially cool, should rapid sea-level rise commence. Bleaching is also monitored within the gene bank nurseries as well as in the collecting sites during marine heat wave events as climate change and ocean warming develops. Corals within the nurseries that exceed their bleaching thresholds are tagged and translocated to sites where temperatures have remained 1-2C cooler, as indicated by logger data. Corals proven bleaching resistant to extreme temperatures on the reefs can be added to the nurseries as space opens, focusing on collecting species of exceptional resilience represented by fewer genotypes, or representing unusual coloration or other characteristics.
-
Regeneration patches are created in subsequent years using fragments trimmed from colonies within the nurseries as the corals grow larger. These patches start with a single or a few rapidly growing open-branched coral species and are intended to jump-start natural recovery processes which in turn facilitate the transition of the patch into a highly diverse patch representing a balanced natural ecosystem. Corals of rapidly growing branching Acropora species within the nurseries are trimmed annually to prevent their overgrowth, and fragments are used to create regeneration patches for the accelerated regeneration of degraded reef areas (synonymous with nucleation patches of Bowden-Kerby 2023). Planted to various structures designed to enhance survival and ecological impact, the focus is on creating strong settlement signals, enhancing fish habitat, and restoring sexual reproduction to corals.
- -
- As best practices for the creation of regeneration patches are not yet known, there is an important need for experimental comparisons between methods and between coral species used. Optimal patch size and comparisons among the various planting structures designed to receive coral fragments and to enhance fish habitat are needed. Single-species patches of various species, growth forms, and genera as well as mixed-species patches must be set out as experimental comparisons of relative impacts on larval-based coral recovery as well as their ability to provide habitat for various fish species. In this way great strides can be made in better understanding natural larval-based recovery processes and interactions between corals and fish, with best practices for enhancing or accelerating the recovery process established.
The primary method presently used for locating heat-adapted coral populations to collect for relocation to secure nurseries is via the study of satellite photographs available on Google Earth. Figure 6, Figure 7 and Figure 8 show representative collecting areas and potential gene bank nursery sites. The shallow heat-stressed reef areas can readily be identified by the light blue and white colors of sand reflecting the light, against the contrast of the brown colors of corals and algae. Coral colonies are readily distinguished by color and texture on zooming in, although it is not possible to determine if the corals are living or dead, so further field scoping is then required. While the present work uses boats and snorkelling, drones offer a potential to greatly speed the process, as living corals can be readily identified at that finer scale.
Once the collecting sites are identified, ideal nursery sites must be found nearby to create long-term gene bank nurseries, also using satellite photos followed by ground truthing.
Ideal nursery sites provide good water flow yet protection from storm generated waves and currents. While deeper areas might be considered for counties where resources allow, for developing countries like Fiji with less access to technology and funding, we only consider shallow areas ranging from 2-4 meters deep, which allows nursery maintenance using snorkelling and without the use of SCUBA. Nurseries are ideally situated immediately behind a shallow reef break, where waves consistently break during high tide or rough weather. Situating the nursery a meter or so lower than the protective reef allows waves breaking on the nearby reef to roll over the top of the nursery, leaving the nursery unscathed even during stormy weather. A possible advantage to shallow water nurseries is that water motion and proximity to breaking waves may help increase coral growth rates, nutrient flows, and could potentially accelerate the diffusion of toxic free radicals during heat stress events. However shallow nurseries will have greater temperature fluxes and higher UV levels than deeper ones.
In a major shift in approach to collecting corals, the Reefs of Hope strategies recognize the imminent threat that heat adapted and declining coral species face in shallow water locations, so rather than taking only a small portion of a coral colony as is common practice, entire coral colonies are removed from situations of extreme heat stress and air exposure during extreme low tides, moving and planting them within nurseries located 1-2 meters deeper and 2-3⁰C cooler on elevated nursery structures. After being secured, rescued heat-adapted corals are grown within the nurseries over the years and serve as “mother colonies”, trimmed as they grow, with seed fragments used to accelerate nature-based adaptation and recovery processes [13].
Where heat-adapted coral colonies continue to exist in abundance in jeopardized conditions where long-term survival is impossible, such as colonies exposed at low tide or located within extremely heat stressed tide pools, colonies can be removed for direct planting into “regeneration patches” on cooler reefs, designed to facilitate the recovery and adaptation of damaged reef areas via creation of fish habitat and settlement signals to reboot natural larval-based recovery processes. In this manner, copious coral genetic diversity can be saved in some locations by direct translocation of corals from hotter to cooler waters nearby, averting losing at least a portion of corals during local mass die-off events. However, in such cases these corals will not be as secure as those located within elevated nurseries, as they will remain vulnerable to coral predators such as COTS, as well as perhaps being even more vulnerable to cyclones.
In both nurseries and regeneration patches, coral colonies of unique genotypes, are clustered by species to restore sexual reproduction to declining and increasingly rare coral species, overcoming the alee effect so that copious numbers of coral larvae are released into the wider coral reef system. While the long-term outcome of this strategy may be difficult to monitor, the present situation of declining and rare coral species releasing gametes during mass spawning events with little or no chance for fertilization is a tragedy, and simply by restoring sexual reproduction, nature is enabled to do the rest. Reefs of hope is about restoring natural recovery and adaptation processes, rather that forcing nature into less natural configurations such as with the planting of thousands of less diverse coral fragments, as is the common paradigm.
The coral rescue strategies proposed in this document are time-sensitive and therefore of great urgency, as marine heat waves of extinction-level intensity are now being experienced on many reefs and threaten to hit all coral reefs in the coming years [52]. A systematic rescue of jeopardized corals can potentially save many thousands of corals located in extreme environments where they will otherwise perish. These same corals, once secured, would provide an immense resource for accelerating coral breeding and recovery aspects of the program, helping keep pace with the accelerating rate of ocean warming.
Nursery Design and Construction
Genebank nurseries are intended to serve as long-term refuges for corals and therefore must be built to last for decades. They should be designed to become self-maintaining within a year or so, cleaned by herbivorous and planktivorous fish populations that increase within the corals over time. Nurseries should be located close to the reef to encourage larger herbivorous fish to cross over and help clean the structure. If several meters of sand separate the nursery from the reef, small grazing fish will be prevented from crossing, and so in such cases “fish bridges” are then installed, consisting of corals planted to mesh A-frames or welded reef stars, providing fish with shelter so that they can safely cross open ground to access the nursery.
The preferred gene bank nursery structure is built of 16mm deformed steel bars. Each bar is six meters long and is typically cut in half for ease of transport and use. A basic small nursery is therefore slightly smaller than 3m x 3m (2.8 x 2.8m). Supporting legs are made by bending bars into U-or L-shapes to form supporting legs, hammered into the sand or rubble sediments. The unbent bars are laid out into a square on the sand or rubble substratum to outline the nursery, and legs are then hammered into place before lifting the straight bars and attaching them with heavy, UV-resistant (black) cable ties. To create the nursery top, heavy 3mm gauge, 200x200mm welded reinforcing floor mesh is used. Available widely as 3x12 meter sheets, the mesh is cut into four 3x3m sections, each coated with two coats of epoxy resin to form four nursery tabletops. One of these sheets is attached to the top of the completed 2.8 x 2.8m metal bar frame substructure to complete the “table nursery”. Nurseries can be easily expanded by linking them together, and sections of rope nurseries can also be included for easy cultivation of large branching species. Braces are also added to the completed nurseries for increased cyclone resistance, and the design has proven to be highly resistant.
Designs or locations that depend on regular human maintenance for existence are risky, as far too much precious coral diversity is at stake. Personal experience during the COVID pandemic showed that disruptions can prevent access for months or even years, with suspended rope nurseries growing so heavy that the ropes broke and some nurseries collapsed under the weight of the corals, with many corals falling to the sand below and perishing. While design can certainly vary, methods such as tree nurseries and other sorts of nurseries suspended from floats are never used, as they are neither self-cleaned by fish, nor are they structurally sound over many years. Rope nurseries suspended from metal bar frames, if they are placed adjacent to reefs and fish populations, can be used, as long as they have additional braces installed to reinforce the structure and additional support bars for the ropes, allowing the corals to grow for many years without falling, even if abandoned. However, as fish do not typically live within rope nurseries, table nurseries are integrated into rope nursery design to provide fish shelter habitat (Figure 9 and Figure 10). Another factor in gene bank nursery design is ease of shading, should an off-scale marine heat wave impact the nursery. The standard design of rope nurseries lends itself to ease of shading during bleaching emergencies, and shading can also be installed during high UV summer months, enabling the coral work to continue without delay (Figure 11).
Planting Corals to Table Nurseries
Coral colonies are usually planted directly to the table nurseries, attached with UV-resistant cable ties in areas of active current flow, or for calmer areas nested unattached onto the mesh, recessed within the spaces of the 200mm x 200mm mesh where they will self-attach within several months. Larger and heavy massive coral colonies are generally left unattached and remain stable during cyclones due to their weight and the sheltered nature of the nurseries. Another method is to cement the corals to concrete “cookies”, large 30-40cm disks with 4 holes in them for ease of attachment to the structure, allowing ease of movement later, while corals placed directly onto the nursery will adhere strongly and become hard to move over time. Tabulate and short-branched species of digitate and corymbose Acropora corals are well-suited to attachment to concrete disks (Figure 12), while large branched staghorn growth forms are not. For staghorn corals collected from Stegastes farmer damselfish territories and with dead lower branches, the dead portions are trimmed off and 5-6 branches are then tied into a sort of bouquet using long thin cable ties or fishing line, with the completed bundle nested within the mesh spaces of the nursery, or these large branching corals can simply be grown on ropes.
Experience indicates that arrangement of corals within the nursery according to growth form is important. Fast growing open branching coral species are given more room and are separated from slower growing tightly branched and massive species. However, including the larger branched corals in the nursery greatly helps ensure functionality and low maintenance by attracting abundant resident fish populations, as long as overgrowth is kept in check. Planktivorous fish such as blue Chromis and black and white banded Dascyllus, help keep drifting cyanobacteria out of their host colonies, and herbivorous fish such as juvenile parrotfish and surgeonfish, work to keep the dead parts of corals and the nursery table clean. All of these fish species prefer large branching corals therefore it is important to locate clusters of colonies of these species throughout the nursery.
The goal of coral arrangements within the nursery is to pair each coral species with 2-3 other genotypes of its kind within the nursery, to encourage successful spawning and larval generation. Another goal is to produce corals for establishing regeneration patches on degraded reefs. Annual cool-season trimming of especially fast-growing corals needs to occur to prevent competition and overgrowth between corals. Slower growing massive corals and tightly branched corals can serve as donor colonies for restoration work, but trimming is rarely required as a maintenance measure. Where particular coral species have become rare, it is important to create a duplicate nursery elsewhere, as insurance against loss due to unforeseen disasters. However, all corals trimmed for inclusion within regeneration patches are already backed up, should the patches fail. The chances of successful recruitment of larvae originating from within the gene bank and regeneration patches and settling to areas of the wider reef system will increase with each passing year, however it may not be possible to verify or quantify these impacts, so we trust that nature will put the larvae to good use.
Regeneration Patch Design and Construction
Corals trimmed from the nurseries as part of routine annual maintenance are put to good use in creating “regeneration patches” as the primary out-planting strategy for degraded areas of the wider reef system. Using this approach, trimmed fragments, generally between 10-30cm in length, are attached via cable ties to mesh A-frames, created from the same 200x200mm heavy floor mesh as used for the nursery table nursery tops. Four frames, eight squares wide each can be cut from a single 12-meter sheet, bent into an A-shape, and then coated with epoxy resin. Coral fragments trimmed from a single mother colony are ideally kept within separate containers so that they can all be planted together onto a single section of an A-frame, with the goal of the branches merging into a single reproductive coral colony within 1-2 years. At least two genotypes of each coral species, preferably 4-6 genotypes of a species should be planted in this manner onto a single A-frame, creating an effective reproductive unit for deployment to the field. Fragments are usually attached at each welded cross-junction to prevent movement, and a planted A-frame contains 104 coral fragments, avoiding the bottom row of junctions to avoid direct contact with the substratum by the corals and some elevation as partial protection against benthic predators (Figure 13). The A-frames are generally planted in a boat or on shore, regularly sprinkling the corals with seawater during production, and the planted A-frames are then transported to and deployed into the desired site, undergoing the same sprinkling. Once deployed, the A-frames are secured either metal stakes, with concrete block weights tied to the A-frame bottom via short metal rods.
There are many unknowns as far as regeneration patch design, and there are so many possible combinations, so an experimental approach should be taken. Single or multiple A-frames (or other structures) can be used to create a regeneration patch or various sizes, and various coral genera and growth forms can be used individually or in combination. Beyond restoring sexual reproduction and dispersing bleaching resistant symbionts more widely throughout the coral reef system, the other purposes of the regeneration patches should not be forgotten: to create ideal fish habitat for positive impacts to the corals and substratum around the patch, and to create strong olfactory, auditory, and visual settlement signals for incoming coral larvae. Experience hints at trends, such as Acropora being the smelliest of the genera, while certain Pocillopora species are the noisiest due to infaunal crustaceans.
Results
Proof of Concept for Proactive Coral Rescue in the Face of a Marine Heat Wave
In Fiji, the 2023 Caribbean mass die-off was taken as a warning, and when the NOAA Coral Reef Watch predicted an intense marine heat wave due to hit Fiji in February-April 2024, plans were made to rescue heat adapted corals from reef areas of extreme heat stress in Malolo District and receiving nurseries were begun at the end of the cool season, in October 2023. local-scale translocation of corals was then carried out, focusing on securing corals found within a shallow “hot pocket” reef. Over 1,300 coral colonies were moved from a shallow <0.5 M deep hot pocket reef flat area at West Nuku Reef over the coming months (Figure 14), relocated only a few hundred meters away and about a meter deeper, within waters about 2C cooler. The nursery was built of 16mm metal rebars hammered into the sand, and topped with 200x200mm heavy welded mesh, constructed in the shape of letters to spell out “BULA”, the largest (16M x 45M) word ever written under the sea. Bula is a greeting that means life, and the rescue nursery, in addition to its function as a coral gene bank as a shout out for action on addressing climate change (Figure 15, Figure 16 and Figure 17). The hot pocket reached >35C during the bleaching event, while the nursery reached 33C. Tens of thousands of corals left behind bleached severely and died, while none of the translocated corals bleached and all survived the marine heat wave event. https://icriforum.org/bula-reef/ 80% of the corals on the shallow source reef died due to bleaching and of the 20% of corals that remained, most were subsequently devoured by over-abundant COTS, and coral cover of the source reef went down to about 5% within a year. Most surviving corals were either unpalatable species or were located within protective Stegastes farmer damselfish territories. The nurseries were proven totally immune to COTS predation. This intervention represents the first major local-scale translocation of corals from hotter to cooler conditions as a proactive measure to rescue corals from an approaching marine heat wave plus a subsequent COTS plague. https://www.youtube.com/watch?v=C9OmNWZEt94
Challenges of Establishing Gene Bank Nurseries
Although all the 1300+ corals on Bula Nursery survived the marine heat wave unbleached, several challenges were encountered later in the season; small butterflyfish of several species became abundant and targeted the tabulate and tightly branched corymbose coral species, spreading a white band type of disease. Several staghorn type species of branched Acropora also caught the disease, but not colonies with abundant planktivorous fish. When multiple corals began to perish, we began removing all diseased colonies into a rubble quarantine area to try to stop the spread, but the disease ultimately led to the retirement of about a quarter of all the corals in the nursery over a one-year period. On the positive side, the Acropora corals remaining in the nursery have proven disease resistant as well as heat resistant. The butterflyfish are now less abundant, perhaps as large predatory fish visit the nursery and the coral-dwelling fish have increased greatly. As additional space in the nursery opened, we replaced the corals with large-branched corals from the nearby rope nursery known to be bleaching resistant. Knowing which coral species do poorly in this particular low-energy site, an additional nursey was created further out in the reef system with better flow, for housing the tabulate and corymbose growth forms as a possible solution.
Towards the end of the cool season in September/October, we faced another crisis at Bula Nursery, with an outbreak of the black hairy/ filamentous cyanobacteria, Lyngbya. We were fortunately familiar with dealing with this problem, which is seasonal [93]. The algae had become a major problem in established nurseries twice in years past, in 2019 and in 2021. According to the literature, this alga is iron-limited [94]. We have noticed that once the iron mesh has cured in the ocean for many months, or if it is double coated with epoxy, that these algae growing on the nursery structures become less problematic. At the peak of Lyngbya blooms, for over a month, clumps of algae lifted off reef flats via small oxygen bubbles which form in the afternoon, and the algae then drifted into and snagged on the branching corals, with rope nurseries especially affected. (Figure 18). This can result in the algae covering the corals to the point whereby the corals become invisible, and if not quickly removed, results in high mortality [93]. During the height of the first outbreak, we noticed that Lyngbya cover continued to remain low on the natural reef as Stegastes farmer damselfish work to weed the algae from corals and dispose of it, keeping the main reef clean and alive. A solution was found for treating badly impacted nursery ropes; untying and placing Lyngbya-covered ropes within Stegastes territories resulted in 100% of the algae being removed within 24 hours [93]. However, for table nurseries, this procedure was not so feasible, so extensive hand cleaning was required. In more recent years, and with Bula Nursery, we have observed that the more abundant the planktivorous blue/green Chromis and black and white banded Dascyllus fish are, the less impacted by algae the coral colonies become. Although quantitative data is needed to better quantify this, we appear to have found some very helpful coral gardeners specializing in weeding noxious filamentous algae! Using large coral colonies to establish gene bank nurseries greatly speeds the recruitment of planktivorous and juvenile grazing fish (parrotfish and surgeonfish), which are important to the ecological functioning of coral nurseries and lower the need for maintenance.
Mass Coral Bleaching as a Major Selection Event for Bleaching Resistant Corals
In two other Fiji sites, Uluibau Village on Moturiki Island in 2023, and Naidiri Village on the Coral Coast in 2025, mass coral bleaching events were used to identify and collect bleaching resistant coral colonies as soon as the heat stress began to abate, but before any surviving bleached corals could regain their color. These corals were incorporated into table nurseries to secure the corals and to create broodstock of heat adapted corals for future coral work by the communities, while supporting climate resilience, coral reef regeneration, reproduction, and ecological functionality.
While the gene bank nurseries will facilitate successful spawning, some species are represented by only one genetic individual, and so more collections must occur during future bleaching events or from hot pocket reefs to restore reproductive functionality. These three sites can now serve as model training sites on which a Fiji national coral reef adaptation plan can be built.
While multiple regeneration patches have been created in all three of C4C’s Fiji sites, time has proven inadequate to establish results-based verification. However, as strong larval settlement signals for incoming coral larvae are now undoubtedly being emitted night and day by these restored patch communities in the form of chemical cues (smell), visual cues (sight), and acoustic cues (sound) [95,96,97,98,99], these areas may serve as important larval attractors and sources of reef recovery, and the results should become quantifiable within 3-4 years, indicated by a significant increase in the numbers of juvenile corals around and down current from the regeneration patches, as compared to reef areas without regeneration patches. From a community standpoint, the increased fish populations already evident in the patches which support increased MPA functionality are proof of success.
Developing a Fiji National Coral Reef Adaptation Plan
Fiji has tremendous advantages over other independent Pacific Island nations; not only are Fiji’s coral reefs more intact in comparison to the other nations due the factors discussed in the introduction, but the numbers of NGOs operating coral reef conservation initiatives is impressive. This includes Fiji offices of the big international NGOs: WWF, WCS, and CI, as well as LMMA, the Locally Managed Marine Area Network, Blue Prosperity Fiji, and several smaller NGOS, including Reef Explorer Fiji, Pacific Blue Foundation, Vatuvara Foundation, Coral Gardeners, Global Vision International, and Corals for Conservation, as well as others. All of these organizations work in partnership and under the scope of the Fiji Ministry of Environment and Climate Change, and the Fiji Ministry of Fisheries and Forests.
Fiji also has the additional advantage of having a foundation of well-established academic institutions; the University of the South Pacific with its marine science program, and Fiji National University with its fisheries science program, with hundreds of Pacific Islanders graduating with qualifications to fill roles as marine field officers at government and NGO level. A credentials-level capacity building curriculum has been developed specifically for the Reefs of Hope Ocean Decade Action and awaits approval by SPC for implementation through a certified academic institution.
Another source of support for a national coral-focused adaptation program is the tourism industry of Fiji, which is the largest of the independent Pacific region. As coral reefs are an essential foundation of Fiji’s tourism industry, resorts have been major supporting partners for coral-focused work, providing access to field sites and logistics support, hiring trained university graduates as marine officers, and funding actions such as the Bula rescue nursery in the Mamanucas. The tourism industry with their keen interest could potentially absorb dozens of trained coral reef adaptation professionals to implement the coral-focused objectives of a Fiji national coral reef adaptation plan as a means for securing and enhancing the tourism product. Tourism Fiji as well as the Fiji Ministry of Tourism and individual resorts should thus be regarded as important partners in a national coral reef adaptation plan and efforts to secure coral reefs into the future.
And last by not least, the indigenous communities and locally managed marine areas offer an immense source of support, youthful energy, and stewardship that is unequaled in most parts of the world and must be nurtured, guided, and utilized to enable coral-focused adaptation work to expand into hundreds of community-based MPAs around the country. In some areas of the country, tourism-community partnerships also offer a potential source of multi-generational conservation and adaptation support that can stretch far into the future, independent of the need for external funding and management.
This proactive plan to save the coral diversity of Fiji from demise in the coming decades in the face of worsening marine heat waves due to climate change must build on the existing site work of Government Ministries, Academic Institutions, NGOs, communities, and resorts, weaving the various coral elements into existing conservation and capacity building programs as insurance against the inevitable impacts of extinction-level bleaching impacts that may be unavoidable now due to increasing levels of carbon in the atmosphere and oceans.
What is required now is for these coral reef focused institutions to study the plans and discuss the importance and potentials of working together towards unified coral-focused outcomes. While UNESCO has already endorsed these strategies, an endorsement of the general concepts is needed at government and NGO levels. The specific implementation plans suggested below are flexible and are given as an example of what implementation might look like (Figure 16), as a beginning at formulating a national coral-focused adaptation action plan. The sites were selected based on ideal adaptation potential for thermal resistance, ease of access (roads, dependable boats, or airports), and existing support for marine conservation.
Phases of a Fiji-Wide National Coral Reef Adaptation Plan
Phase One: Coral Species Rescue and Stabilization
In this initial phase focus is placed on actions to stop further coral loss; to keep each coral species alive and with enough genetic variability to remain in a viable condition into the future. The goal is to secure each coral species within gene bank nurseries located within ‘sweet spots’ in the ocean: suitable temperature regime, adequate current flow, shelter from storms, and with easy access, to maintain the corals in healthy condition over time, increase their biomass, and trim excess material for use in subsequent reef regeneration efforts.
The work of securing declining coral species begins with environmental scoping and assessment to determine threats to the most vulnerable genera and seeking out the most heat-resilient variants of the species that remain- the bleaching resistant corals located in the hottest waters- the extreme shallows and restricted lagoons, where lethal temperatures are becoming inevitable as the oceans warm.
Based on the findings of scoping exercises, locally managed marine areas and tourism areas are assessed for inclusion and support for the work in the various stages of implementation. Where special reef areas are under attack by COTS, removal activities will be implemented. Efforts are made for supervised involvement of communities and youth in the work of seeking out and collecting the corals for relocation to coral gene bank nursery sites situated in areas with good potential for future survival of the corals. At this stage in the work, the gene bank nurseries are created using table nurseries and rope nurseries as both are elevated above sandy substratum as protection from coral predators.
This simple act of saving heat-adapted corals from imminent death due to extreme temperatures and predators, by moving them into cooler water nurseries has been shown to be highly effective, providing a source of sustainably obtained corals for community and resort-based restoration work. This provides an effective point of entry by those most impacted by the loss of coral reefs due to climate change and warming seas, helping to spread thermal resilience so that coral reefs adapt and survive into the future.
Once gathered into centrally located and cooler water genebank nurseries, the corals are propagated to increase coral biomass, with surplus heat adapted corals then trimmed for use in a community and resort-focused program. The out-planting stage plants corals strategically into regeneration patches within facilitated coral reef recovery sites. Each coral genotype is maintained within the nursery to keep the genotype under protection and monitored over time; the goal is to secure the shallow water corals of Fiji to mid century and beyond. The work will take several years before all areas of the country have gene bank nurseries and most coral species are secured and reproduction restored, with reference to the rare species. The suggested goal is for a minimum of 50 genotypes of each of the most vulnerable coral species secured around the country, securing 99% of the genetic diversity of the nation’s corals [92], with a focus on the more vulnerable bleaching impacted of Fiji’s 300+ coral species. If represented by 50 genotypes per 200 coral species total, that would come to 10,000 coral genotypes secured throughout the combined nurseries of the country.
Based on ongoing work, a 15m x 12m nursery can secure about 600 corals, and so 20 nurseries could secure over 12 thousand corals of the nation. Doubling the size of each nursery or adding a second nursery would increase that number to 24,000 corals. The out-planting phase of the work 2-3 years after nursery establishment will spread the risk of loss from natural disasters even more, sending corals of as many coral genotypes as practical out to multiple sites locally.
The most vulnerable species group is Acropora, which is also the fastest growing, most genetically and morphologically diverse, and is the most vital habitat forming coral species group [13]. Acropora is also the coral that produces a strong pheromone smell which attracts the larvae of corals and potentially many other species, indicating that a reef area is “prime real estate” so that the larvae can settle out in the best possible habitat. Pristine reefs of the past were most often dominated by Acropora but are now becoming dominated by more resilient species [13]. This is why Acropora species are a major target when securing corals from localized extinction due to severe marine heat waves. The more resilient coral species such as Porites, Montipora, and Pocillopora can also be planted where Acropora is in short supply, even though these corals grow more slowly and may not provide the same chemical settlement signals nor do they provide the same sorts of fish habitat, however their presence might still help reboot recovery as the small crabs and shrimp which live among Pocillopora in particular, are very noisy and sound attracts the larvae of both fish and corals [13].
The work of selecting heat-adapted broodstock corals for the gene banks and finding the ideal nursery sites proceeds based on local knowledge and experience, with temperature logger data and remote sensing data, where possible used to identify the best areas for collecting heat adapted corals as well as the areas of less thermal stress for developing nursery and regeneration sites. The fate (bleaching and mortality) of the corals left behind in heat stressed sites plus environmental data of these more stressed reef areas as compared to nursery and out-planting sites, will serve as controls over the years, recording the advance of global warming. As the program develops, areas where corals are dying out as well as areas where corals are doing better in the face of mass bleaching will provide an opportunity to tease out the underlying factors so that the adaptation models and strategies might be improved, contributing to the wider objectives of the program.
A collaborative and coordinated approach to carry out the plan is required, with the major stakeholders being the Fiji Government Ministries of Environment and Climate Change, Fisheries, Tourism, Indigenous Affairs, the indigenous reef owning communities, tourism industry partners, conservation NGOs, and scientists. Each site requires a lead organization or management committee of all those involved in the site. A multi-partner national coordination body should give scientific oversight and oversee reporting standards and assist with problem solving, and implementation requires reporting and feedback mechanisms and standards. All implementing staff must be trained via a coordinated training program (Figure17).
Phase Two: Restoring Sexual Reproduction
When corals become rare, reproduction and larval recruitment fails. A problem arises when coral colony abundance becomes low. The “allele effect” results when the surviving adult coral colonies are separated by too great a distance for effective fertilization. When coral colonies are 10-15 meters apart, virtually no fertilization and thus no larval formation occurs, greatly impacting coral reef recovery. A simple strategy of relocation of isolated corals to restore sexual reproduction is integral to the development of an adaptation plan.
A central strategy of a recovery plan focuses on searching the reefs to find and secure rare and declining coral species and to gather these heat-adapted survivors of bleaching together to restore sexual reproduction within nurseries and regeneration patches. This enables the corals to recover through natural larval recruitment processes, with the intended long-term result being less vulnerable heat-adapted coral reefs.
Natural coral reef recovery is reliant on a good supply of coral larvae, which requires reproductively viable populations of corals in the vicinity and up-current of impacted reefs. Coral reefs with good connectivity to up-current populations of diverse corals and thus abundant coral larvae are able to recover from disturbance rapidly, within 5-10 years, while reefs with poor connectivity and thus cut off from abundant larval sources take much longer to recover. However, reefs with poor connectivity to good larval sources can sometimes recover as an alternative coral community- a phase shift dominated by brooding coral species that produce larvae able to settle immediately (Porites rus, Pocillopora), and thus more likely to settle locally near the parent corals. For broadcast spawners such as Acropora, the coral larvae require development while drifting in the larval state, and so the source reefs are best located 4-10 days up current from the recovering reefs, so that the larvae can settle within that time period. The relative distance this represents can be as high as several hundred km, or as low as just a few km, should the larvae get caught within lagoons, or caught within the eddies that can form down current from islands.
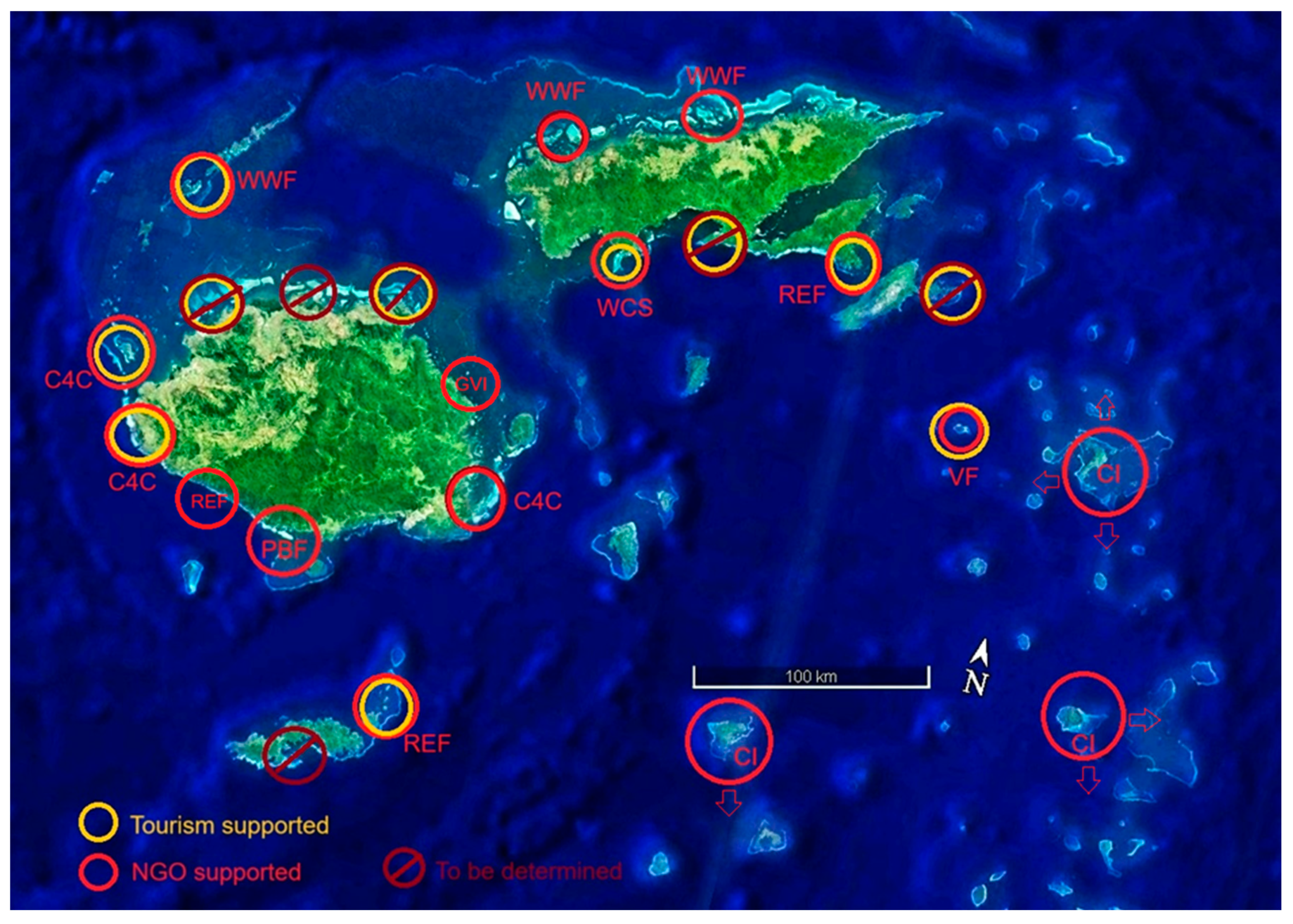
Figure 18.
Existing NGO supported sites (red) and sites with potential tourism industry support (yellow). Double circles indicate both sources of support for coral reef conservation. Conservation International (CI), Wildlife Conservation Society (WCS), World Wildlife Fund (WWF), Vatuvara Foundation (VF), Reef Explorer Fiji (REF), Pacific Blue Foundation (PBF), Global Vision International (GVI), and Corals for Conservation (C4C). Government support and oversight covers all sites.
Figure 18.
Existing NGO supported sites (red) and sites with potential tourism industry support (yellow). Double circles indicate both sources of support for coral reef conservation. Conservation International (CI), Wildlife Conservation Society (WCS), World Wildlife Fund (WWF), Vatuvara Foundation (VF), Reef Explorer Fiji (REF), Pacific Blue Foundation (PBF), Global Vision International (GVI), and Corals for Conservation (C4C). Government support and oversight covers all sites.
Phase Three: Facilitating Natural Recovery Processes
A largely unrecognized problem when Acropora corals become rare, is that the strong smell they emit is missing from the reef. This “reef” smell is due to the presence of a pheromone attractant that strongly induces larval settlement. Once Acropora corals are missing from the reef, coral larvae have a more difficult time finding the proper place to settle. This is why a special focus is placed on large branching staghorn corals, as they are particularly smelly, grow the fastest of any corals, and serve as important fish habitat. As most Acropora species have declined throughout the islands, it is important that they be secured within gene bank nurseries and propagated for later use in the creation of reef regeneration patches designed to attract coral and fish larvae to degraded reefs, which in turn accelerates natural adaptation and recovery.
Out-planting proceeds using second generation corals trimmed from nurseries and planted to reefs with good ecological balance and lower stress and thus with good prospects for long-term survival of heat adapted corals. The goal is to create multi-genetic patches of each coral species close enough to each other to restore high rates of fertilization as well as to emit a strong smell to help the larvae find their way home. Established as discrete and concentrated patches, the corals can be more easily monitored and cared for, as opposed to haphazard planting of smaller fragments. Regular gardening activities can be conducted to remove predators and excessive seaweeds and to control disease. This work should be focused in reef areas 2-3°C cooler than where the corals originated, and if possible, where some corals have escaped death due to bleaching in past marine heat waves.
Assuming that there are initially 10-20 gene bank nurseries of heat adapted corals established, each of these nurseries should be producing surplus corals to create recovery patches in multiple sites of the surrounding reefs as the corals grow and are trimmed. With at least 100 species of branching corals in each nursery, by the 3rd year, at least a hundred densely planted recovery patches would be created per year per nursery, 1,000 to 2,000 coral patches spread throughout the islands, facilitating both sexual reproduction and recruitment-based recovery, as well as heat adaptation, vastly increasing the stability and resilience of coral reefs in the nation, regardless of warming seas.
Phase Four: Additional Measures to Support Coral Reef Health
This phase might include Diadema sea urchin cultivation, giant clam hatcheries and cultivation, lobster habitats, and a professional level educational program to train larger numbers of people, to fill coral-focused jobs created and supported at government facilities and at resorts. At this point in the strategy and following biosecurity protocols, corals could be sent between sites within the nation where a species is either locally extinct or of low genetic diversity.
Captive Breeding of Corals
While less community appropriate, more costly strategies such as land-based flowing seawater facilities for the spawning and rearing of large numbers of juvenile bleaching-resistant corals, can also be justified in an upscaled national strategy. However, these and all strategic efforts are dependent on securing an abundant collection of diverse heat-adapted coral broodstock, secured within gene bank nurseries.
As resources allow captive breeding programs between resilient coral hosts to begin, the coral genotypes used should be more fully investigated as far as the source of resilience: host, algal symbionts, or probiotic symbionts. Captive breeding carried out within flowing seawater systems only makes sense between two or more genotypes of host-based heat resilient corals. The goal should be to inoculate the resulting juvenile corals with heat resilient symbionts and probiotics, giving two resistant parents, resistant algal symbionts, plus resilient bacteria.
A more low-tech version of gamete harvesting and captive larval rearing strategy is also possible once the source of resilience of the coral broodstock in a gene bank nursery is known. Resilient host-based coral strains can then be used to create single-species regeneration patches of multiple coral genotypes. The resulting gametes released from these patches, will all be from heat resistant parents, which can be captured via nets placed directly above patches during spawning. The resulting larvae can then be reared to the point of settlement within temporary tanks and allowed to settle onto specially designed (movable) substrates in an environment dominated by heat resistant symbionts. An alternate strategy would be to be release ready to settle larvae directly onto coral reefs with well-established regeneration patches planted with heat-resilient holobionts, giving the juvenile corals a high probability of accessing heat-resilient symbionts as they settle around the patches.
Including Tridacnid Clams in the Coral Reef Adaptation Work
In addition to corals, giant clams of the genus Tridacna are another photosynthetic coral reef animal that is vulnerable to bleaching due to hot waters. Just as with corals, heat adapted clams can be moved from hot to cooler and from extreme shallows to deeper waters as an adaptation strategy. This would help prevent giant clam death during marine heat waves in extremely hot and shallow environments.
Tridacnid clams contain the same species and strains of symbiotic algae within their tissues as do corals, and the clams shed live symbionts in copious amounts in their feces, which have been shown to be picked up by newly settled corals [100]. One potential strategy therefore could focus on using heat adapted clams to facilitate adaptation. The strategy would involve moving heat adapted clams from hot pocket reefs to cooler waters to give the clams a better chance at survival, and so the clams can potentially share their algae throughout the reef to become “teachers of the reef” shedding resistant algae into the environment, which are then picked up by the newly settled juvenile corals.
The present clam hatchery at Makogai Island also has the potential to become focused on supporting the national coral reef adaptation plan; intentionally introducing heat-resistant symbiotic algae to the juvenile clams, thereby generating large numbers of heat adapted clams for sending out to LMMAs, which would in turn help spread heat resistant symbionts throughout the country, increasing the adaptation potential of coral reefs.
Measures to Combat Ocean Acidification
While not an important factor presently, measures to combat against the impacts of ocean acidification may in the future need to be incorporated into gene bank nurseries. Such measures might involve farming seaweeds on ropes alternating with ropes of corals or perhaps locating nurseries above healthy seagrass beds to combat declining pH levels. The present practice of locating nurseries above lagoon sand may serve to buffer the impacts of acidification as well. Corals adapted to acidified waters may also be sought for incorporation into the program, such as corals potentially found associated with mangrove areas far from freshwater influences. Numerous potential sites for the development of low pH adapted corals are found on leeward low-rainfall reefs (Figure 18).
Detailed Components of the Proposed Coral Reef Adaptation Strategy for Fiji
- Build vision and support for the national coral reef adaptation plan as a collaborative partnership between government ministries, NGOs, academic institutions, and scientists. Create a multi-stakeholder management structure and mechanisms for moving forward, including obtaining financial support for a ten-year program.
- Conduct training of managers and field staff in the strategies and methods to build unified vision and upskill local capacity.
- Work within existing NGO implementation sites using existing data, remote sensing, aerial photos, and local expertise to create coral reef adaptation site plans.
- Identify jeopardized heat-adapted coral populations located within shallow reef areas.
- Identify ideal coral refuge sites where waters are expected to remain 2-4°C cooler and make plans for relocation and securing heat adapted coral colonies.
- Focus on rare and less common coral species within shallow heat stressed reefs, collect and translocate at least 5-10 genetically distinct colonies of each coral species per site to reefs where heat stress is less intense, rescuing them from near certain demise in the short term as the ocean warms, and securing them within gene bank nurseries.
- Use mass bleaching events as selection events for collecting bleaching resistant corals from the wider reef area, incorporating them into the gene bank nurseries to secure them from COTS and to ensure that they are cloned and used to reboot reproduction and natural recovery within regeneration patches.
- As the corals in the nurseries grow, branching species are trimmed and fragments used for rapid growth within suspended rope nurseries located in shallow, moderately stressed waters, where they are cultivated for out-planting work. Slow-growing massive coral species can also be propagated via micro-fragmentation techniques and planted onto concrete domes to create adult-sized colonies within 2-3 years.
- The corals in the rope nurseries grow rapidly and are harvested annually during low heat and low UV stress, with the ropes replanted. These second-generation corals are used for out-planting to create multi-genetic and reproductive recovery patches of each coral species on the reef. Either whole colonies are planted directly to degraded reefs, or branches can be planted onto A-frame or reef star structures, segregated by genotype into quadrants on the frame, so that the branches merge into larger reproductive colonies as they grow.
- Duplicate gene bank nurseries by sending samples to secondary nurseries on each atoll as insurance against loss due to unforeseen conditions.
- Establish temperature loggers in donor and recipient sites to monitor marine heat waves over time, monitoring both the corals left behind and those translocated.
- Encourage community and resort COTS removal activities on high-value reefs and within LMMAs, especially around regeneration patches to help ensure their success.
- Include the translocation of giant clams and rare or uncommon species of sea cucumbers and sea urchins in the work and within no-take MPAs, as these important coral reef species also face demise in extreme temperatures. Monitor these species closely to prevent predators from killing them in the new sites and as needed, locate clams within structures designed to shelter them from octopus, eagle rays and triggerfish.
- Continue to assess which coral species remain in each area of the country to determine which species are common, which are uncommon, which have become rare, and which are now locally extinct, with plans for species recovery via restoration of sexual reproduction. Missing or rare species may require cross-site exchange of samples.
- Implement a formal professional-level credentials training programme, adapted from the Reefs of Hope curriculum approved by the South Pacific Community.
- Through trained and experienced facilitators, involve the community and youth in the coral work, building capacity and hope for the future.
- Include the tourism industry in the work, while maintaining high standards by requiring the oversight and employment of professionally certified coral reef regeneration technicians.
- NGO-supported gene bank nurseries will provide community-based efforts with multiple genotypes of heat adapted corals for the work within LMMAs.
- Monitor the work and continue with the steps above until most of the coral reef LMMAs of the nation have a coral reef regeneration and recovery programme, focused on active support for the adaptation and recovery of coral reefs.
Recommendations Based on Decades of Lessons Learned
Strategies for Collecting and Moving Corals
- For coral recovery programmes, whole coral colonies should be targeted for inclusion within gene bank nurseries, so that the programme will not have delays waiting for corals to grow to the appropriate reprodctive adult size. We must remember that this is a coral rescue, and what is left behind will be monitored until it likely dies in a severe marine heat wave at some point in the future.
- To move as quickly as possible to save more coral biomass, where patches of branching corals are extensive, duplicate fragments of each coral can be collected at the same time as the gene bank nursery is created, with the intention of creating recovery patches simultaneously. This strategy would therefore also collect 15-20cm fragments for planting to frames and stars.
- A systematic approach to collections on the various hot pocket reefs is important to avoid re-collecting the same genotypes.
- Collecting the few unbleached corals at the end of a mass bleaching event as the waters begin to cool is an effective way to ensure that the corals are truly bleaching resistant. However, if this is done, whole colonies do better than fragments, as fragments often bleach severely in the high UV due to increased exposure, unless the fragments are planted under shade.
- When transporting corals, care must be taken to avoid oxidative stress, which can result when using non-aerated containers, or when fragments bunch up together in a container. In Fiji we get 100% survival from transporting coral colonies for hours on deck out of water, if the corals are shaded and constantly sprayed with cool seawater.
Guidance for Selecting Cooler Water Gene Bank Nursery Sites
- Desktop scoping for hot-pocket coral populations and genebank nursery sites should be conducted using Google Earth satellite photos. Coral populations can be identified among the extensive shallow waters and reef flats as potential hot water collection sites. Prospective cooler water nursery sites can also be located. Ground-truthing visits to the promising sites is the second step, to see what corals remain and to verify assumptions. This desktop and field scoping process is required before any plan of action to rescue heat adapted corals can be devised and carried out. The use of drones might be important to fine-tune strategies and save time, once collection areas are identified.
- Follow the principle that outer reefs are generally cooler than inner reefs and smaller reefs with less shallow water area will be cooler than larger reefs with extensive shallow areas. If shallow water temperature data exists, that would also be useful.
- With Reefs of Hope strategies, ideally corals are not translocated into highly different thermal or light regimes, to prevent acclimation pressures away from their original highly resistant state. The goal is to keep heat-adapted corals within the natural stress regimes where they evolved, which now requires local scale translocation, as thermal conditions are shifting due to warming oceans. As global warming progresses, the location of the gene bank nurseries may thus need to shift into cooler waters in the years to come.
- Even short distance translocation a few hundred meters, if taken from <1m deep and moved to ~2m deep, can make a huge difference in reducing temperatures, as it moves corals from the hot thermal layer that forms at the surface and into the cooler layer below.
- Gene bank nurseries ideally should be secure from wave damage, being located behind a shallow reef facing the incoming wave direction.
- As winds and wave directions can be expected to change during shifting monsoons and storms, the nursery should be located where reefs protect it on all sides, not just the prevailing seasonal or fair-weather wave direction.
- Nurseries are situated in waters 2-3 meters deep, to optimize growth and to allow for more community involvement. Situated at least 1-2 meters deeper than the nearby reef flats or coral heads and situated close-up against the reef, so that breaking waves will roll right over and above the nursery during storms. In this way the nursery will have both good flow and excellent protection, and reef fish will visit the nursery to clean the structures.
- Nurseries located near populations of surgeonfish and juvenile parrotfish are cleaned naturally by the fish. However, if the fish are too afraid to cross barren sand or rubble to the nursery, habitat bridges can be created. Tree nurseries are not recommended due to the high cost of hand cleaning, as well as vulnerability during big wave events.
- Elevated table and rope nurseries help prevent coral predation and are less impacted by resuspended silt and associated disease.
- A-frames and reef stars can be used as easily deployed mini gene bank nurseries in shallow water and hardground sites, with up to six genotypes of a single species per frame and one per star, in close proximity so that effective sexual reproduction is restored.
- If a primary goal is to produce fragments for out-planting, rope nurseries can be established on 3m x 3m iron frame tables linked together over sand or rubble substrates, with each rope representing a single genotype of a particular coral species. The corals are trimmed annually, or one colony is fragmented to replant a replacement rope, and the other colonies are used for out-planting.
- 12. Out-planting for restoration is best focused on cooler water reef areas, and is done as mixed genetic patches, to restore reproductive potential to declining species. If reef stars or A-frames are used, 2-3 distinct genotypes should be planted to each star, with each genotype planted to a specific section, to encourage fusion of branches, resulting in merging into adult colonies over a shorter time frame.
Discussion
Government Incentives and Policies Are Needed to Support Coral Reef Adaptation
The mainstreaming of coral-focused adaptation at the tourism industry level could be incentivised by establishing government enabling mechanisms through creation of new policies and tax incentives. Enabling mechanisms For communities would be through the creation of new laws supporting the establishment and enforcement of no-take marine areas, support for economic incentives such as alternative livelihoods to fishing, activities such as a bounty on COTS removal, continued support for giant clam restocking into permanent MPAs, support for sea cucumber restoration, and support for youth involvement in coral-focused work. Increased coordination and capacity building at village, district, and provincial levels should focus on maintaining local initiative to continue carrying out the lion’s share of the work at community level.
With a more proactive approach being taken by government, the various local and international NGOs can better align behind and support government initiatives. In the past NGOs had to develop their own priorities and took on the primary leadership role, which was appropriate during Fiji’s several coups and in post-coup realities. However, a stronger leadership role is now being taken by government as times become stable and government capacity increases. Government capacity is an essential aspect of funding for a national climate change adaptation program, as only governments can tap into the billions of UN dollars set aside to fund climate change adaptation.
A promising development in proactive management and leadership by the Ministry of Environment & Climate Change was launched in August 2024- the Fiji National Hub for Coral Reef Conservation, with the goal of becoming the central coordination body to work with NGOs, Ministries, and UN agencies to develop Fiji’s Coral Reef Action Plan. The ongoing work aims for completion by mid 2025, and is being facilitates in partnership with WCS, the Wildlife Conservation Society and UNDP/UNEP, and with input from all stakeholders.
Representative Estimated Budget
Budget estimated for a comprehensive coral reef adaptation plan. The true costs of national implementation will require more fine-scale analysis.
| ITEM | Per year site cost | 20 Sites/ year | Ten years |
| Personnel | €100,000. | € 2,000,000. | €20,000,000. |
| Boats, Fuel, Travel | 50,000. | 1,000,000. | 10,000,000. |
| Nursery Materials | 10,000. | 200,000. | 2,000,000. |
| Out-planting Materials | 10,000. | 200,000. | 2,000,000. |
| Community Workshops | 10,000. | 200,000. | 2,000,000. |
| SUBTOTALS | € 180,000. | € 3,600,000. | € 36,000,000. |
| Additional support for coral spawning systems to double as giant clam hatcheries |
100,000. |
Five sites/ yr 500,000. |
Ten Years € 5,000,000. |
| TOTALS | € 275,000. | € 4,100,000. | € 41,000,000. |
Reinterpreting of the Precautionary Principle of Science
The coral reef conservation community uses the precautionary "do no harm" approach as far as applying coral-focused interventions at scale, waiting for substantial scientific proofs before acting, assuming corals will survive and adapt without intervention until the relevant science is accepted and published in peer-reviewed literature. However, it is now clear that science has moved too slowly to save the corals, as a series of intense marine heat waves and mass coral bleaching events has begun to hit, killing off most of the genetic diversity in multiple countries, with little possibility for recovering the genetic diversity that has been lost.
While basic proof-of-concept for Reefs of Hope solutions has been established and shared widely at conferences and on social media, there has been little change in strategies. Most scientists continue to adhere to older models and a strictly conventional interpretation of the "precautionary principle of science", not understanding that the precautionary principle should be turned on its head whenever there is an extinction risk. "Innocent until proven guilty" turns into "guilty until proven innocent" whenever a factor potentially threatens to kill an endangered species or ecosystem. There is very low risk in using this approach, as even if we end up securing corals whose source populations survive the next mass bleaching event, nothing is lost, especially considering that the existential threats to heat adapted coral populations will only increase with each passing year as the planet warms.
Conclusion
There are no quick-fix solutions, all we can do is buy more time for coral reefs and coral species. Reefs of Hope solutions will NOT solve the overarching problem of coral reef collapse due to climate change. Rather, our goal is to help those involved with coral planting to do it in such a way that it keeps coral species alive and coral reefs recovering in the face of severe marine heat waves, buying more time. What better solution than to prevent the die-off of our most heat adapted corals and to work with these genetic treasures to reboot natural adaptation and recovery processes?
Global warming is causing the nearshore and hot reef flats to become lethal environments during marine heat waves, and the heat-adapted corals living there can not move themselves, so we must urgently move these corals to maintain them within the shifting thermal environment as the waters warm. Perhaps one day, when sea level rises 30-50cm, the thermal environment of these warmer waters will cool again, and we can put these corals back where they came from? But for now, we must act swiftly to keep them alive!
Ultimately, the long-term success of any interventions requires a transformation of energy production away from fossil fuels. However, with carbon emissions and temperatures increasing unhindered, nature may impose her own solutions, such as massive ice loss resulting in sea level rise and the collapse of ocean overturning circulation [101,102,103]. While these events pose major risks globally, some models suggest that they could temporarily ameliorate reef heat stress by slowing global warming rates or cooling surface waters regionally [104,105]. Even a moderate amount of sea level rise could help cool shallow reef areas and lagoons due to increased circulation and increased depth of reef flats, lowering heat stress on the corals that remain [106,107,108]. These “silver lining” types of changes are unpredictable, but they would come with enormous costs to humanity which might finally galvanize effective action, while buying coral reefs more time?
References
- Drake JL, Mass T, Stolarski J, Von Euw S, van de Schootbrugge B, Falkowski PG. How corals made rocks through the ages. Glob. Chang. Biol. 2020, 26, 31–53. [Google Scholar] [CrossRef] [PubMed]
- Barnett J, Jarillo S, Swearer SE, et al. Nature-based solutions for atoll habitability. Philos. Trans. R. Soc. B Biol. Sci. 2022, 377.
- Ferrario F, Beck MW, Storlazzi CD, Micheli F, Shepard CC, Airoldi L. The effectiveness of coral reefs for coastal hazard risk reduction and adaptation. Nat. Commun. 2014, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hasan MM, Kabir MB, Prodhan MSR, et al. The Physical and Mechanical Properties of Coral Sand. Eur. J. Theor. Appl. Sci. 2024, 2, 313–337. [Google Scholar] [CrossRef] [PubMed]
- Hoegh-Guldberg O, Poloczanska ES, Skirving W, Dove S. Coral reef ecosystems under climate change and ocean acidification. Front. Mar. Sci. 2017, 4, 252954. [Google Scholar] [CrossRef]
- Spalding, M, Burke L, Wood SA, Ashpole J, Hutchison J, zu Ermgassen P. Mapping the global value and distribution of coral reef tourism. Mar. Policy 2017, 82, 104–113. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O. Climate change, coral bleaching and the future of the world’s coral reefs. Mar. Freshw. Res. 1999, 50, 839–866. [Google Scholar] [CrossRef]
- Voolstra CR, Peixoto RS, Ferrier-Pagès C. Mitigating the ecological collapse of coral reef ecosystems. EMBO Rep. 2023, 2023, 24. [Google Scholar]
- United Nations Environment Program. (2021). Why are coral reefs dying? 2021, November 12. https://www.unep.org/news-and-stories/story/why-are-coral-reefs-dying.
- Najeeb S, Khan RAA, Deng X, Wu C. Drivers and consequences of degradation in tropical reef island ecosystems: strategies for restoration and conservation. Front. Mar. Sci. 2025, 12, 1518701. [Google Scholar] [CrossRef]
- Mulà C, Bradshaw CJA, Cabeza M, Manca F, Montano S, Strona G. Restoration cannot be scaled up globally to save reefs from loss and degradation. Nat Ecol Evol. 2025, 9, 822–832. [Google Scholar] [CrossRef] [PubMed]
- Leggat WP, Camp EF, Suggett DJ, et al. Rapid Coral Decay Is Associated with Marine Heatwave Mortality Events on Reefs. Current Biology. 2019, 29, 2723–2730.e4. [Google Scholar] [CrossRef] [PubMed]
- Bowden-Kerby, A. Coral-focused climate change adaptation and restoration based on accelerating natural processes: Launching the “Reefs of Hope” paradigm. Oceans 2023, 4, 13–26. [Google Scholar] [CrossRef]
- Hughes TP, Kerry JT, Álvarez-Noriega M, Álvarez-Romero JG, Anderson KD, Baird AH, et al. Global warming and recurrent mass bleaching of corals. Nature 2017, 543, 373–377. [Google Scholar] [CrossRef] [PubMed]
- IPCC. (2019). IPCC Special Report on the Ocean and Cryosphere in a Changing Climate. Intergovernmental Panel on Climate Change. https://www.ipcc.ch/srocc/.
- Hoegh-Guldberg O, Mumby PJ, Hooten AJ, Steneck RS, Greenfield P, Gomez E, et al. Coral reefs under rapid climate change and ocean acidification. Science 2007, 318, 1737–1742. [Google Scholar] [CrossRef] [PubMed]
- National Academies of Sciences, Engineering, and Medicine. (2019). A Research Review of Interventions to Increase the Persistence and Resilience of Coral Reefs. Washington, DC: The National Academies Press. [CrossRef]
- Bay RA, Palumbi SR, Davies SW, et al. Coral conservation in the age of genome editing. Proceedings of the National Academy of Sciences 2019, 116, 12134–12141. [Google Scholar]
- Palumbi SR, Barshis DJ, Traylor-Knowles N,; Bay RA. Mechanisms of reef coral resistance to future climate change. Science 2014, 344, 895–898. [Google Scholar] [CrossRef] [PubMed]
- Oliver TA & Palumbi, SR. Do fluctuating temperature environments elevate coral thermal tolerance. Coral Reefs 2011, 30, 429–440. [Google Scholar] [CrossRef]
- Safaie A, Silbiger NJ, McClanahan TR, et al. High frequency temperature variability reduces the risk of coral bleaching. Nat Commun 2018, 9, 1671. [Google Scholar] [CrossRef] [PubMed]
- Thomas L, Rose NH, Bay RA, Lopez EH, Morikawa MK, Ruiz-Jones L, Palumbi SR. Mechanisms of thermal tolerance in reef-building corals across a fine-grained environmental mosaic: Lessons from Ofu, American Samoa. Frontiers in Marine Science 2018, 5, 434. [Google Scholar] [CrossRef]
- Morikawa MK,; Palumbi SR. Using naturally occurring climate resilient corals to construct bleaching-resistant nurseries. Proceedings of the National Academy of Sciences 2019, 116, 10586–10591. [Google Scholar] [CrossRef] [PubMed]
- van Oppen MJH, Oliver JK, Putnam HM, Gates RD. Building coral reef resilience through assisted evolution. Proceedings of the National Academy of Sciences 2015, 112, 2307–2313. [Google Scholar] [CrossRef] [PubMed]
- Wood S, Stephen J, Pandolfi JM, Palumbi SR. Potential for conservation via relocation: Spatial genetic structure of thermally tolerant corals and implications for assisted migration. Global Change Biology 2023, 29, 999–1013. [Google Scholar]
- NOAA. (2023). Unprecedented marine heatwave impacts Florida coral reefs. National Oceanic and Atmospheric Administration Coral Reef Watch. https://coralreefwatch.noaa.gov.
- Lindsey, R. (2023). NOAA and partners race to rescue remaining Florida corals from historic ocean heat wave. https://www.climate.gov/news-features/event-tracker/noaa-and-partners-race-rescue-remainig-florida-corals-historic-ocean.
- Muller EM, Bartels E, Baums IB, et al. Widespread coral bleaching in Florida linked to anomalously high sea temperatures. Frontiers in Marine Science 2023, 10, 1227745. [Google Scholar]
- Florida Keys National Marine Sanctuary. (2023). 2023 Coral Rescue Operation Summary. NOAA Office of National Marine Sanctuaries.
- Eakin CM, Heron SF, Liu G, et al. (2023). Global coral bleaching continues into 2023: record heat stress in the Atlantic and Caribbean. NOAA Coral Reef Watch Special Report, September 2023.
- AGRRA (Atlantic and Gulf Rapid Reef Assessment). (2023). Preliminary Report on Coral Mortality in the Caribbean following 2023 Heatwaves. https://www.agrra.
- 32 Alemu J, Laydoo R, et al. (2024). Coral bleaching and mortality in Trinidad and Tobago during the 2023–2024 marine heatwave. Marine Pollution Bulletin (early view).
- Hernández-Delgado EA, Rodríguez-González YM. Runaway Climate Across the Wider Caribbean and Eastern Tropical Pacific in the Anthropocene: Threats to Coral Reef Conservation, Restoration, and Social–Ecological Resilience. Atmosphere. 2025, 16, 575. [Google Scholar] [CrossRef]
- Bowden-Kerby A, Romero L, and Kirata T. (2021). Chapter 17: Line Islands Case Study. In: Active Coral Restoration: Techniques for a changing planet, David Vaughn, Editor. 610pp. https://www.jrosspub.com/science/environmental-science/active-coral-restoration.html.
- IPCC. 2023. AR6 Synthesis Report: Climate Change (2023). Intergovernmental Panel on Climate Change.
- Copernicus Climate Change Service. (2024). Global Climate Highlights 2023: Hottest Year on Record. European Union, ECMWF. https://climate.copernicus.
- Wooldridge, S. A. , & Pratchett, M. S. Coral predators exert selection pressure on corals with heat tolerance traits: implications for reef recovery under climate change. Frontiers in Ecology and Evolution 2019, 7, 221. [Google Scholar]
- Kayal, M. , Vercelloni, J., Lison de Loma, T., Bosserelle, P., Chancerelle, Y., Geoffroy, S., Stievenart, C., Michonneau, F., Penin, L., Planes, S., & Adjeroud, M. Predator crown-of-thorns starfish (Acanthaster planci) outbreak, mass mortality of corals, and cascading effects on reef fish and benthic communities. PLoS ONE 2012, 7, e47363. [Google Scholar]
- Bowden-Kerby, A. (2003). Community-based Management of Coral Reefs: An Essential Requisite for Certification of Marine Aquarium Products Harvested from Reefs Under Customary Marine Tenure. Chapter 11: 141-166 In: Marine Ornamental Species: Collection, Culture and Conservation. Cato, J and C. Brown Editors. Iowa State Press/ Blackwell Scientific Publications, NY, 395 pp.
- Govan H, Tawake A,; Tabunakawai K. Community-based marine resource management in the South Pacific. SPC Traditional Marine Resource Management and Knowledge Information Bulletin 2009, 25, 11–23. [Google Scholar]
- Jupiter SD, Cohen PJ, Weeks R, Tawake A,; Govan H. Locally-managed marine areas: multiple objectives and diverse strategies. Pacific Conservation Biology 2014, 20, 165–179. [Google Scholar] [CrossRef]
- McClanahan TR, Darling ES, Maina J, Muthiga NA, et al. Climate change and coral reef fisheries: shared challenges, converging solutions. Proceedings of the Royal Society B 2021, 288, 20210927. [Google Scholar]
- Bowden-Kerby, A. (2020). Coral restoration for climate change adaptation in the South Pacific. Pages 54-57 In: Hein et al. Coral Reef Restoration as a strategy to improve ecosystem services – A guide to coral restoration methods. UNEP Report, 60pp.
- Eastwood EK, López EH, Drew JA. Population Connectivity Measures of Fishery-Targeted Coral Reef Species to Inform Marine Reserve Network Design in Fiji. Sci Rep. 2016, 6, 19318. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Harding SP, Solandt JL, Walker RC, Walker D, Taylor J, Haycock S, Davis MT, and Raines P. Reef Check Data Reveal Rapid Recovery from Coral Bleaching In the Mamanucas, Fiji. Silliman Journal 2022, 44. [Google Scholar]
- WCS (2020). https://fiji.wcs.org/News-Room/ID/15026/ Research Expedition to Assess Coral Reef Health and Recovery from Tropical Cyclone Winston.
- Rowlands G, Comley J, and Raines P, (2005). The Coral Coast, Viti Levu, Fiji, A Marine Resource Assessment. Published: by Coral Cay Foundation and SPREP. 124pp.
- Bruckner, AW. (2014). Global Reef Expedition: Lau Province, Fiji. Field Report. Khaled bin Sultan Living Oceans Foundation, Landover MD. 33 pp.
- Lequeux C, Dumas P,; Andréfouët S. Connectivity of coral reefs in the Fiji Islands: insights from oceanographic modeling and implications for conservation. Marine Pollution Bulletin 2018, 135, 684–697. [Google Scholar]
- Thompson A, Mellegers M, & Souter D. (2018). Fiji Marine Environment Monitoring Report 2005–2015. Global Coral Reef Monitoring Network.
- Ganachaud A, Sen Gupta A, Brown JN, et al. Projecting changes in the South Pacific Ocean circulation using IPCC models. Climate Dynamics 2011, 36, 1231–1246. [Google Scholar]
- Mumby PJ, Sartori G, Buccheri E, Alessi C, Allan H, Doropoulos C, Rengiil G, Ricardo G. Allee effects limit coral fertilization success. Proc Natl Acad Sci U S A 2024, 121, e2418314121. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Mellin C, Brown S, Cantin N, Klein-Salas E, Mouillot D, Heron SF, Fordham DA. Cumulative risk of future bleaching for the world's coral reefs. Sci Adv. 2024, 10, eadn9660. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Schoepf V, Stat M, Falter JL, McCulloch MT. Thermally tolerant corals have limited capacity to acclimatize to future warming. Global Change Biology 2019, 25, 968–982. [Google Scholar]
- Brown KT, Martynek MP,; Barott KL. Local habitat heterogeneity rivals regional differences in coral thermal tolerance. Coral Reefs 2024, 43, 571–585. [Google Scholar] [CrossRef]
- Klepac CN, and Barshis DJ. Reduced thermal tolerance of massive coral species in a highly variable environment. Proc. R. Soc. B. 2020, 28720201379. [Google Scholar] [CrossRef]
- Rosenberg E, Koren O, Reshef L. et al. The role of microorganisms in coral health, disease and evolution. Nat Rev Microbiol 2007, 5, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Bourne DG, Morrow KM,; Webster NS. Insights into the coral microbiome: Underpinning the health and resilience of reef ecosystems. Annual Review of Microbiology 2016, 70, 317–340. [Google Scholar] [CrossRef] [PubMed]
- Suggett DJ,; Smith DJ. Coral bleaching patterns are the outcome of complex biological and environmental networking. Global Change Biology 2020, 26, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Apprill, A. The role of symbioses in the adaptation and resilience of coral reefs to environmental change. Annual Review of Marine Science 2020, 12, 489–516. [Google Scholar]
- Ziegler M, Arif C, Burt JA, Dobretsov S, Roder C, LaJeunesse TC, Voolstra CR. Biogeography and molecular diversity of coral symbionts in the genus Symbiodinium around the Arabian Peninsula. Journal of Biogeography 2017, 44, 674–686. [Google Scholar] [CrossRef] [PubMed]
- Rowan, R. Thermal adaptation in reef coral symbionts. Nature 2004, 430, 742. [Google Scholar] [CrossRef] [PubMed]
- Berkelmans R,; van Oppen MJH. The role of zooxanthellae in the thermal tolerance of corals: A ‘nugget of hope’ for coral reefs in an era of climate change. Proceedings of the Royal Society B 2006, 273, 2305–2312. [Google Scholar] [CrossRef] [PubMed]
- Cunning R, Silverstein RN, & Baker, AC. Investigating the causes and consequences of symbiont shuffling in a multi-partner coral symbiosis under environmental stress. Proceedings of the Royal Society B, 0141. [CrossRef]
- Barshis DJ, Ladner JT, Oliver TA, Seneca FO, Traylor-Knowles N, Palumbi SR. Genomic basis for coral resilience to climate change. Proceedings of the National Academy of Sciences 2013, 110, 1387–1392. [Google Scholar] [CrossRef] [PubMed]
- Downs CA, Fauth JE, Halas JC, Dustan P, Bemiss J, Woodley CM. Oxidative stress and seasonal coral bleaching. Free Radical Biology and Medicine 2002, 33, 533–543. [Google Scholar] [CrossRef] [PubMed]
- Middlebrook R, Anthony KRN,; Hoegh-Guldberg O. Thermal priming affects symbiont photosynthesis but does not alter bleaching susceptibility in Acropora millepora. Journal of Experimental Marine Biology and Ecology 2008, 357, 40–45. [Google Scholar] [CrossRef]
- Bay RA,; Palumbi SR. Multilocus adaptation associated with heat resistance in reef-building corals. Current Biology 2014, 24, 2952–2956. [Google Scholar] [CrossRef] [PubMed]
- van Oppen MJH, Oliver JK, Putnam HM,; Gates RD. Building coral reef resilience through assisted evolution. Proceedings of the National Academy of Sciences 2015, 112, 2307–2313. [Google Scholar] [CrossRef] [PubMed]
- Dixon GB, Davies SW, Aglyamova GA, Meyer E, Bay LK,; Matz MV. Genomic determinants of coral heat tolerance across latitudes. Science 2015, 348, 1460–1462. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth TD, Heron SF, Ortiz JC, Mumby PJ, Grech A, Ogawa D, Eakin CM, Leggat W. Climate change disables coral bleaching protection on the Great Barrier Reef. Science 2016, 352, 338–342. [Google Scholar] [CrossRef] [PubMed]
- Schoepf V, Carrion SA, Pfeifer SM, Naugle M, Dugal L, Bruyn J, McCulloch MT. Stress-resistant corals may not acclimatize to ocean warming but maintain heat tolerance under cooler temperatures. Nat Commun. 2019, 10, 4031. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kenkel CD, Meyer E,; Matz MV. Gene expression under chronic heat stress in populations of the mustard hill coral (Porites astreoides) from different thermal environments. Molecular Ecology 2013, 22, 4322–4334. [Google Scholar] [CrossRef] [PubMed]
- Quigley KM, Bay LK, van Oppen MJH. Selective breeding of corals provides insight into the genetic basis of thermal tolerance. Frontiers in Marine Science 2020, 7, 796. [Google Scholar]
- Bay RA, Palumbi SR. Multilocus adaptation associated with heat resistance in reef-building corals. Current Biology 2014, 24, 2952–2956. [Google Scholar] [CrossRef] [PubMed]
- Quigley KM, Warner PA, Bay LK. The genetic architecture of inherited variation in thermal tolerance in the reef-building coral Acropora millepora. Molecular Ecology 2019, 28, 3864–3880. [Google Scholar]
- Schwarz JA, Brokstein PB, Voolstra C, Coffroth MA. Late larval development and onset of symbiosis in the coral Acropora palmata. Biological Bulletin 1999, 196, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Hirose M, Kinzie RA, Hidaka M. Vertical transmission of symbiotic dinoflagellates in the planulae of the coral Pocillopora damicornis. Zoological Science 2001, 18, 515–518. [Google Scholar]
- Apprill A, Marlow HQ, Martindale MQ, Rappé MS. The onset of microbial associations in the coral Pocillopora meandrina. ISME Journal 2009, 3, 685–699. [Google Scholar] [CrossRef] [PubMed]
- Leite DCA, Salles JF, Calderon EN, Castro CB, Bianchini A, Marques JA, Parente TE, Tsai SM, Peixoto RS. Broadcast spawning coral species associate with specific bacterial communities during early developmental stages. Ecological Indicators 2017, 81, 48–57. [Google Scholar]
- Putnam HM, Mayfield AB, Fan TY, Chen CS, Gates RD. The physiological and molecular responses of larvae from the reef-building coral Pocillopora damicornis exposed to near-future increases in temperature and pCO2. Marine Biology 2012, 160, 2157–2173. [Google Scholar]
- Voolstra CR, Buitrago-López C, Perna G, Cárdenas A, Hume BCC, Rädecker N, Barshis DJ. Standardized short-term acute heat stress assays resolve historical differences in coral thermotolerance across microhabitat reef sites. Global Change Biology 2021, 27, 4307–4319. [Google Scholar]
- Stat M,; Gates RD. Clade D Symbiodinium in Scleractinian Corals: A “Nugget” of Hope, a Selfish Opportunist, an Ominous Sign, or All of the Above? Journal of Marine Biology 2011, 2011, 730715. [Google Scholar]
- Barott KL, et al. Coral bleaching response is unaltered by community composition in long-term warmed reefs. PNA 2021, 118, e2025435118. [Google Scholar]
- Hughes TP, Kerry JT, Baird AH, Connolly SR, Dietzel A, Eakin CM, Heron SF, Hoey AS, Hoogenboom MO, Liu G, McWilliam MJ, Pears RJ. Global warming transforms coral reef assemblages. Nature 2018, 556, 492–496. [Google Scholar] [CrossRef] [PubMed]
- Pears RJ, Devlin M, Hammerman N. et al. (2024). CORDAP R&D Technology Roadmap on Managing the Ecological Risks of Coral Reef Interventions. CORDAP. cordap.org.
- NOAA. (2023). Record-breaking marine heatwave devastates Florida coral reefs. Coral Reef Watch, National Oceanic and Atmospheric Administration.
- Morais RA, Smith TB, Williams DE, & Edmunds PJ. (2024). Caribbean corals under extreme heat stress: insights from the 2023 bleaching event. Coral Reefs.
- Vardi T, Williams DE, Sandin SA,; Zgliczynski BJ. Shifting paradigms in coral restoration: building resilience in the age of climate change. Ecological Applications 2021, 31, e02315. [Google Scholar]
- Bay RA, Rose NH, Logan, CA,; Palumbi, SR. Genomic models predict successful coral adaptation if future ocean warming rates are reduced. Science Advances 2019, 5. [Google Scholar]
- Shearer TL, Porto I, Zubillaga AL. Restoration of coral populations in light of genetic diversity estimates. Coral Reefs. 2009, 28, 727–733. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- McDougall, A. (2020). Territorial Damselfishes as Coral Gardening Partners during a Bloom of the Cyanobacterium Lyngbya majuscula. Master’s Thesis, University of Miami, Coral Gables, FL, USA, 2020. [Google Scholar]
- Paerl HW,; Paul VJ. Climate change: Links to global expansion of harmful cyanobacteria. Environmental Microbiology Reports 2012, 4, 27–37. [Google Scholar]
- Vermeij MJA, Marhaver KL, Huijbers CM, Nagelkerken I,; Simpson SD. Coral larvae move toward reef sounds. PLoS ONE 2010, 5, e10660. [Google Scholar]
- Lillis A, Eggleston DB,; Bohnenstiehl DR. Soundscape variation from a larval perspective: the role of habitat identity and temporal dynamics in a coral reef. Marine Ecology Progress Series 2013, 472, 1–16. [Google Scholar]
- Mason B, Beard M,; Miller MW. Coral larvae settle at a higher frequency on red surfaces. Coral Reefs 2011, 30, 667–676. [Google Scholar] [CrossRef]
- Erwin PM, Szmant AM. Settlement induction of Acropora palmata planulae by a GLW-amide neuropeptide. Coral Reefs 2010 2010, 29, 929–939. [Google Scholar]
- Takahashi T, Hatta M. The Importance of GLWamide Neuropeptides in Cnidarian Development and Physiology. The Importance of GLWamide Neuropeptides in Cnidarian Development and Physiology. J. Amino Acids 2011 2011, 424501. [Google Scholar]
- Morishima SY, Yamashita H, Ohara S, Nakamura Y, Quek VZ, Yamauchi M, et al. Study on expelled but viable zooxanthellae from giant clams, with an emphasis on their potential as subsequent symbiont sources. PLoS ONE 2019, 14, e0220141. [Google Scholar] [CrossRef]
- Kleypas JA, Anthony KRN, Castruccio FS, et al. Designing a blueprint for coral reef survival under rapid climate change. Global Change Biology 2021, 27, 1186–1200. [Google Scholar]
- Hoegh-Guldberg O, Poloczanska ES, Skirving W, Dove S. Coral reef ecosystems under climate change and ocean acidification. Frontiers in Marine Science 2018, 4, 158. [Google Scholar]
- Tittensor DP, Novaglio C, Harrison CS, et al. A global agenda for advancing climate resilience of marine ecosystems. Nature Climate Change 2023, 13, 395–404. [Google Scholar]
- Morrison AK, Griffies SM, Winton M, et al. Mechanisms and impacts of a partial AMOC collapse in the GFDL CM4 climate model. Journal of Climate 2020, 33, 8043–8064. [Google Scholar]
- Heinze C, Meyer S, Goris N, et al. The quiet crossing of ocean tipping points. Proceedings of the National Academy of Sciences 2021, 118, e2008478118. [Google Scholar] [CrossRef] [PubMed]
- Lowe RJ, Falter JL, Monismith SG, Atkinson MJ. Wave-driven circulation of a coastal reef–lagoon system. Journal of Physical Oceanography 2009, 39, 873–893. [Google Scholar] [CrossRef]
- van Woesik R, Houk P, Isechal AL, et al. Climate-change refugia in the sheltered bays of Palau: analogs of future reefs. Ecology and Evolution 2012, 2, 2474–2484. [Google Scholar] [CrossRef] [PubMed]
- Kennedy EV, Perry CT, Halloran PR, Iglesias-Prieto R, Schönberg CHL, Wisshak M, et al. Avoiding coral reef functional collapse requires local and global action. Current Biology 2020, 30, R995–R1000. [Google Scholar]
Figure 1.
Overview of Fiji’s coral reefs except for Southern Lau, which extends several hundred kms to the south.
Figure 1.
Overview of Fiji’s coral reefs except for Southern Lau, which extends several hundred kms to the south.
Figure 2.
History of marine heat stress and ocean surface temperatures on Fiji’s coral reefs, with bottom graph showing summertime bleaching stress levels in “degree heating weeks”, DHW, with the top graph showing mean ocean temperature. Notice the rainbow- like appearance of the top graph, with the oldest data in blue and the newest in red, grey, and black, clearly showing a warming ocean since the 1980s and 90s.
Figure 2.
History of marine heat stress and ocean surface temperatures on Fiji’s coral reefs, with bottom graph showing summertime bleaching stress levels in “degree heating weeks”, DHW, with the top graph showing mean ocean temperature. Notice the rainbow- like appearance of the top graph, with the oldest data in blue and the newest in red, grey, and black, clearly showing a warming ocean since the 1980s and 90s.
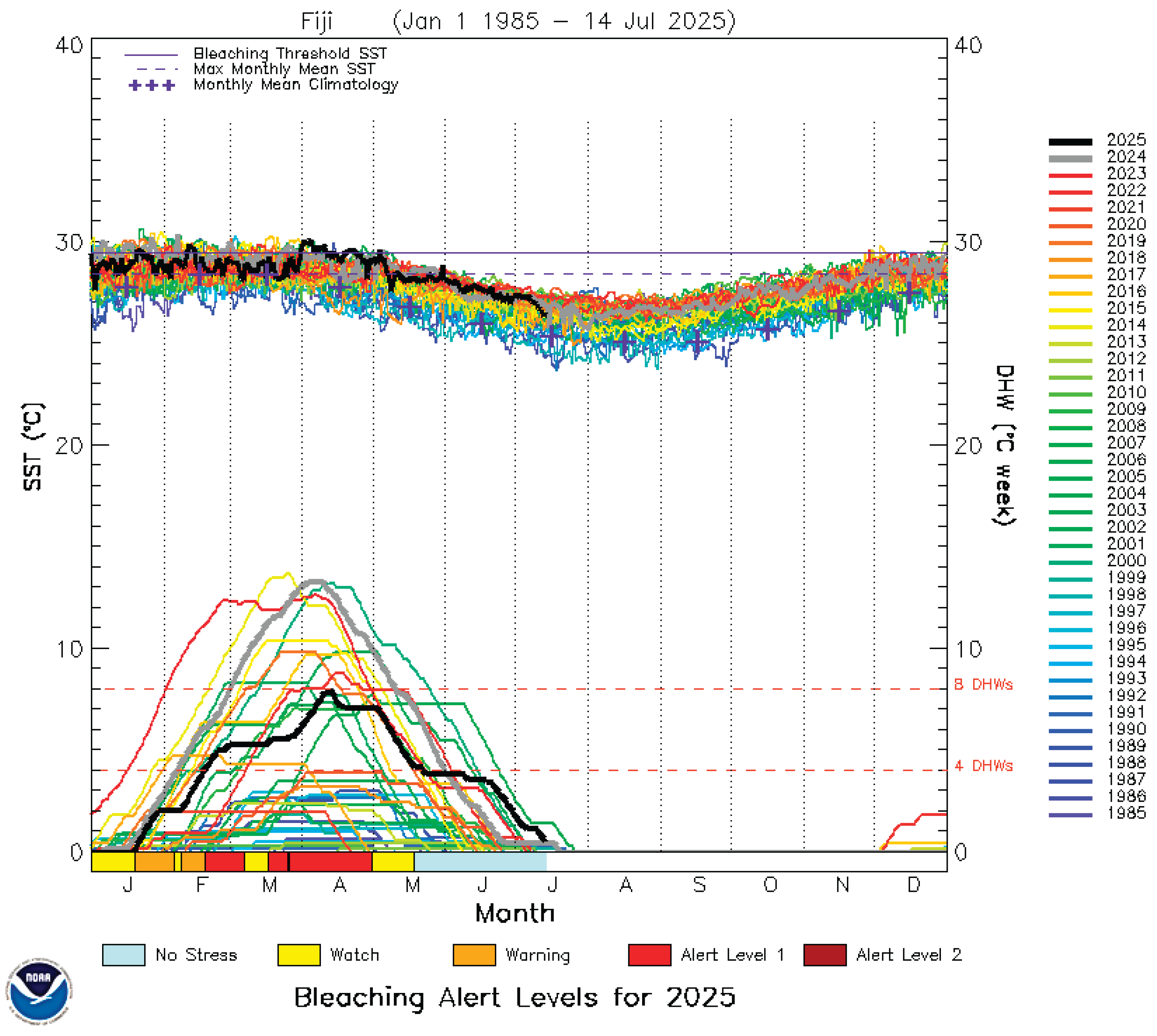
Figure 3.
Heat stress levels in Jamaica (left) and equatorial Kiribati (right) since 1985. Jamaica, is at a similar latitude to Fiji, and represents what Fiji can expect in the future. Fiji has had four 13-14 DHW events while Jamaica had previously only experienced 10 DHWs once, but Jamaica has now experienced consecutive years of 19 and 23 DHW, extinction level coral bleaching. The Line Islands of Kiribati have experienced 27 DHW as well as six years >15 DHW, with multiple coral species becoming locally extinct. Fiji still has time to prepare for this level of heat stress via hot to cooler translocation of corals beforehand, which sparing Fiji some of the long-lasting impacts and saving a wide diversity of corals.
Figure 3.
Heat stress levels in Jamaica (left) and equatorial Kiribati (right) since 1985. Jamaica, is at a similar latitude to Fiji, and represents what Fiji can expect in the future. Fiji has had four 13-14 DHW events while Jamaica had previously only experienced 10 DHWs once, but Jamaica has now experienced consecutive years of 19 and 23 DHW, extinction level coral bleaching. The Line Islands of Kiribati have experienced 27 DHW as well as six years >15 DHW, with multiple coral species becoming locally extinct. Fiji still has time to prepare for this level of heat stress via hot to cooler translocation of corals beforehand, which sparing Fiji some of the long-lasting impacts and saving a wide diversity of corals.
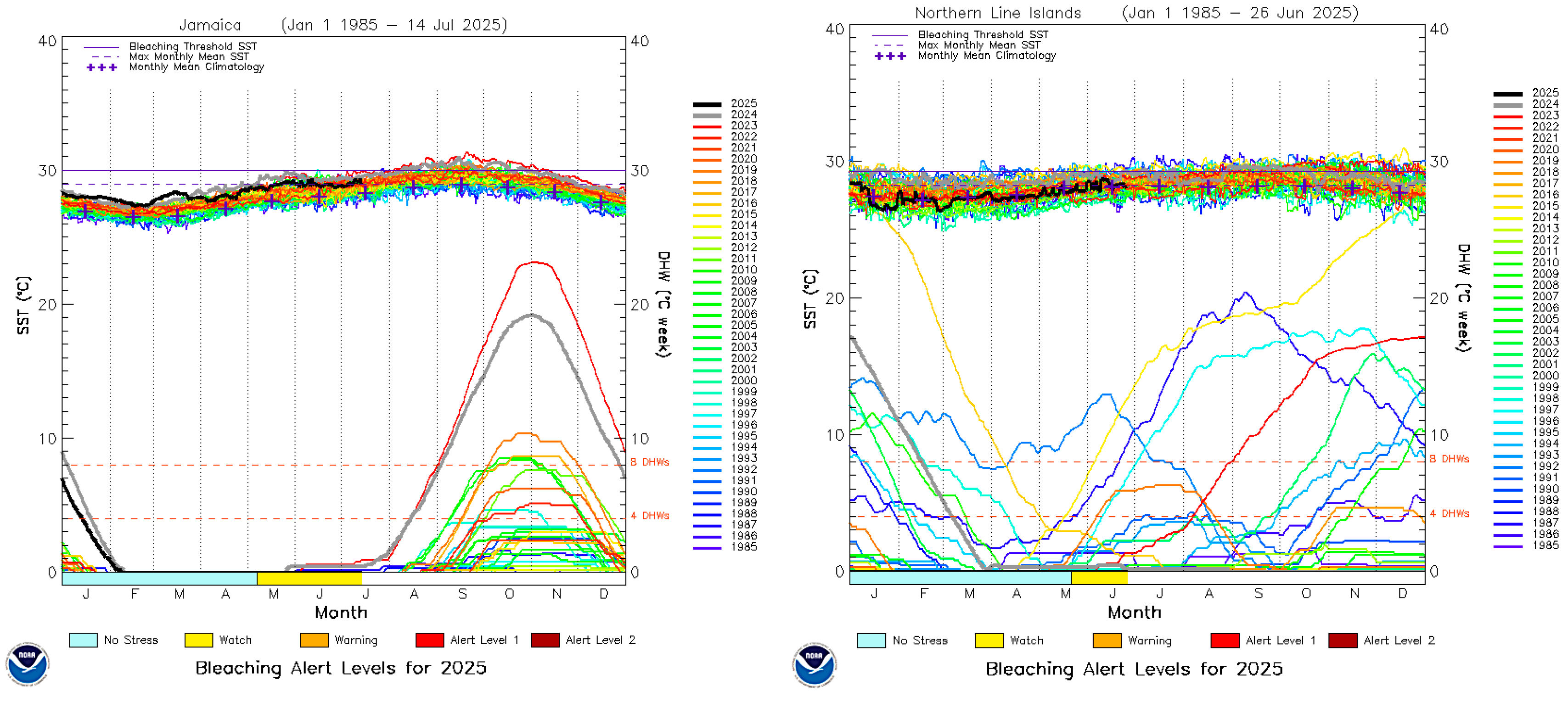
Figure 4.
Previously unpublished analysis using data obtained from NOAA Coral Reef Watch website and using the newly expanded thermal stress scale based on “degree heating weeks”. Comparing South Pacific nations for bleaching frequency and severity, arranged by geographic area and latitude. Arrows indicate latitudinal anomalies: the Solomons and Tuvalu are on the same latitude as PNG, and likewise Society and Tuamotu with Fiji.
Figure 4.
Previously unpublished analysis using data obtained from NOAA Coral Reef Watch website and using the newly expanded thermal stress scale based on “degree heating weeks”. Comparing South Pacific nations for bleaching frequency and severity, arranged by geographic area and latitude. Arrows indicate latitudinal anomalies: the Solomons and Tuvalu are on the same latitude as PNG, and likewise Society and Tuamotu with Fiji.
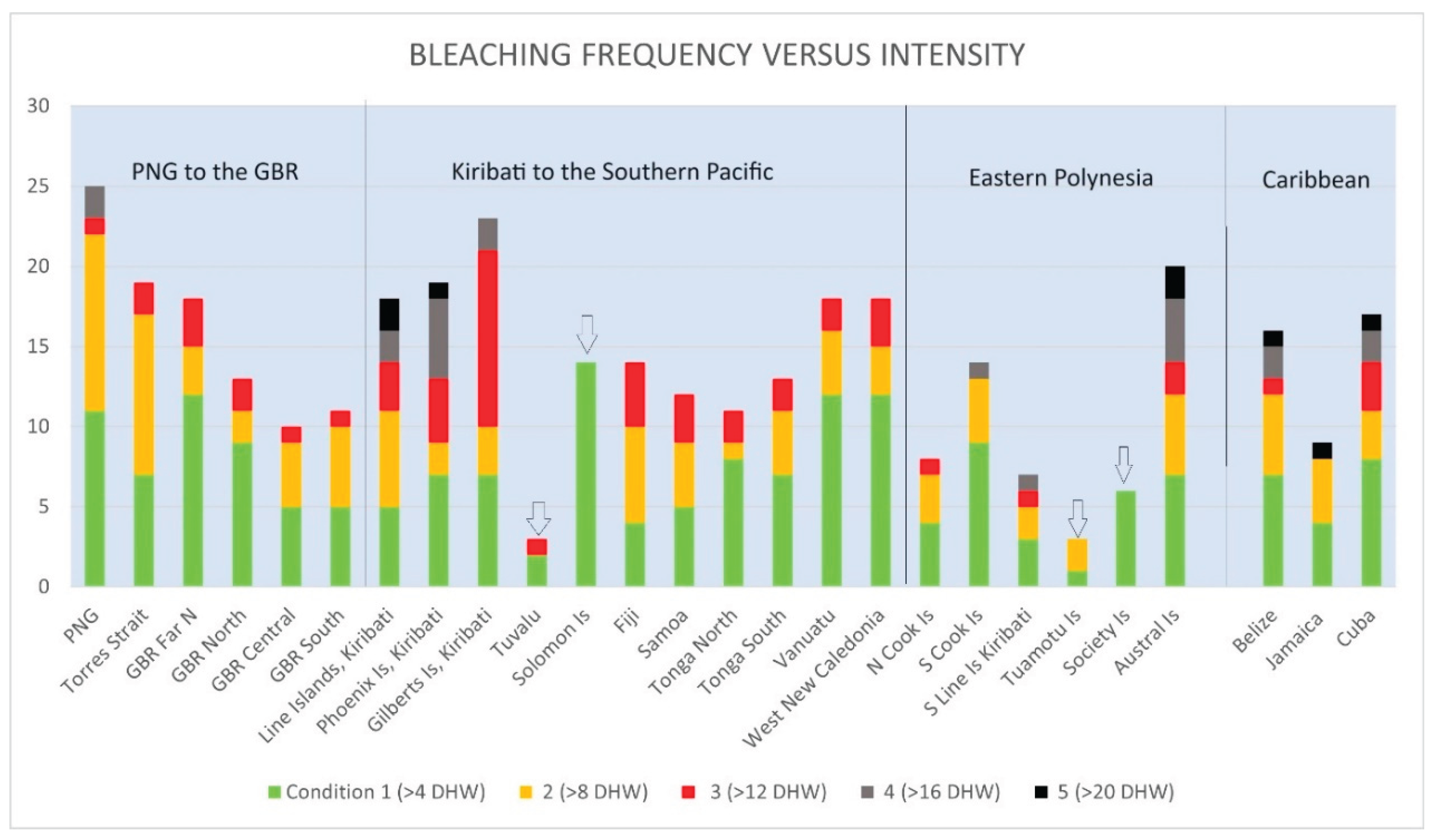
Figure 5.
Shallow coral reefs and reef flats dominate the leeward sides of the two main islands of Fiji, presenting an excellent crucible for the evolution of heat resilience. Hot Pocket Reefs retain heat-resistant coral holobionts that may be in danger of dying out in future extreme bleaching events, as they already have at more severely impacted regions.
Figure 5.
Shallow coral reefs and reef flats dominate the leeward sides of the two main islands of Fiji, presenting an excellent crucible for the evolution of heat resilience. Hot Pocket Reefs retain heat-resistant coral holobionts that may be in danger of dying out in future extreme bleaching events, as they already have at more severely impacted regions.

Figure 6.
Hot pocket reefs north of Lautoka, north coast of Viti Levu, with a cooler-water gene bank nursery site at Tivua Island where tourism activities potentially enable infrastructure, security, and ease of access. Arrows indicate local-scale translocation from hot-pocket collecting sites.
Figure 6.
Hot pocket reefs north of Lautoka, north coast of Viti Levu, with a cooler-water gene bank nursery site at Tivua Island where tourism activities potentially enable infrastructure, security, and ease of access. Arrows indicate local-scale translocation from hot-pocket collecting sites.
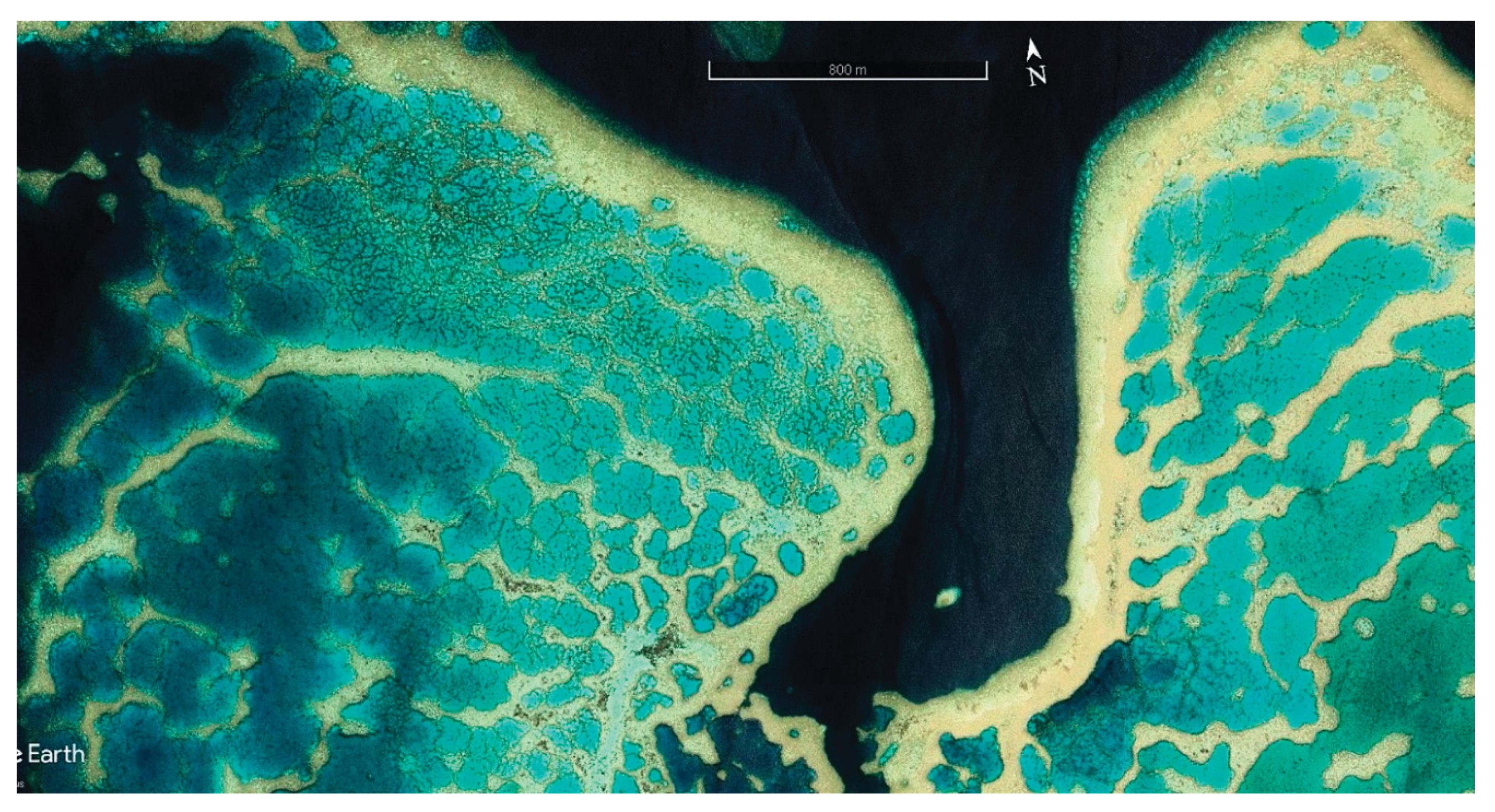
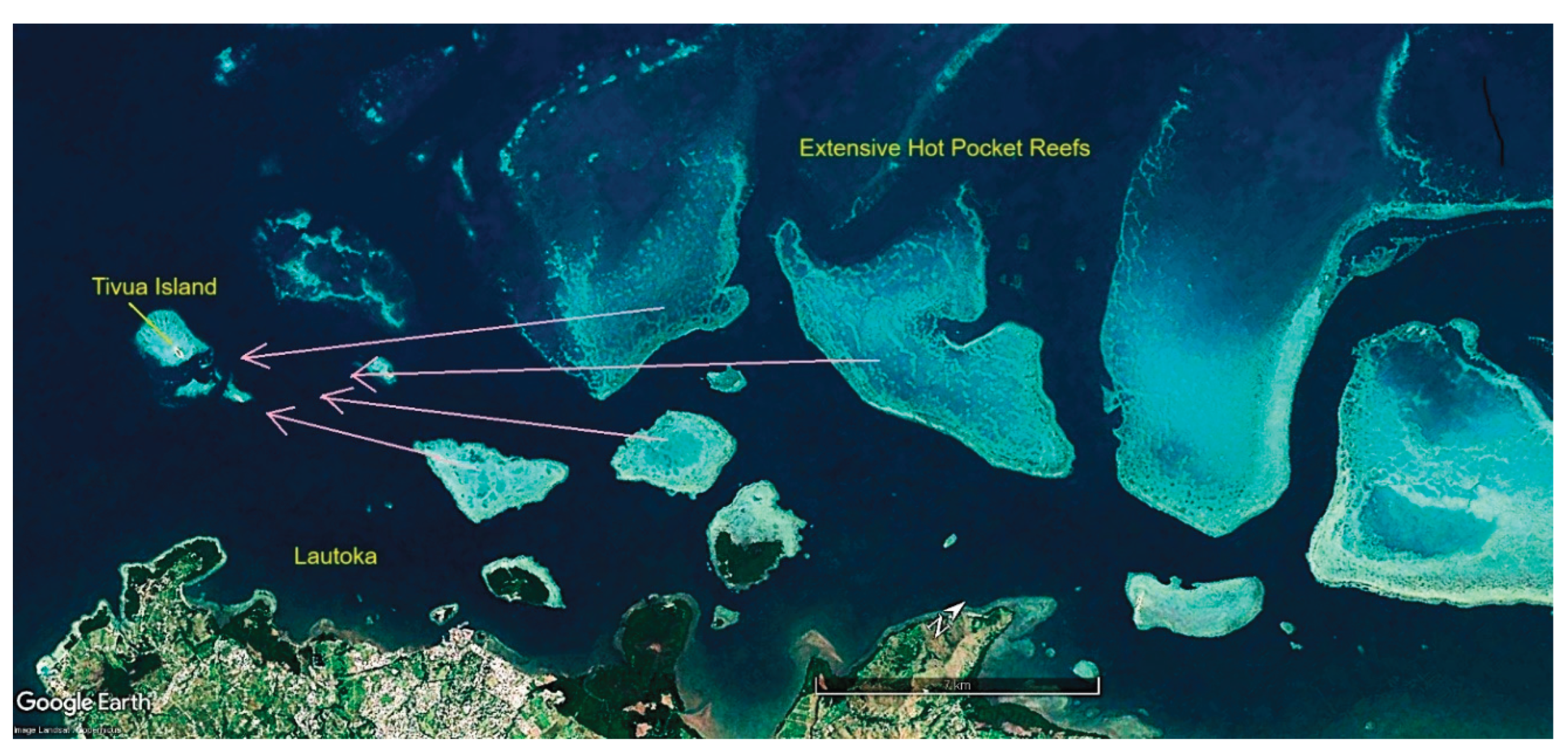
Figure 7.
Potential nursery and collecting sites of Lakeba, Lau. With regular air service for ease of access and located in a key up-current area of Fiji, Lau is important to larval based recovery mechanisms. Numerous reef systems are accessible within hours of Lakeba by boat.
Figure 7.
Potential nursery and collecting sites of Lakeba, Lau. With regular air service for ease of access and located in a key up-current area of Fiji, Lau is important to larval based recovery mechanisms. Numerous reef systems are accessible within hours of Lakeba by boat.
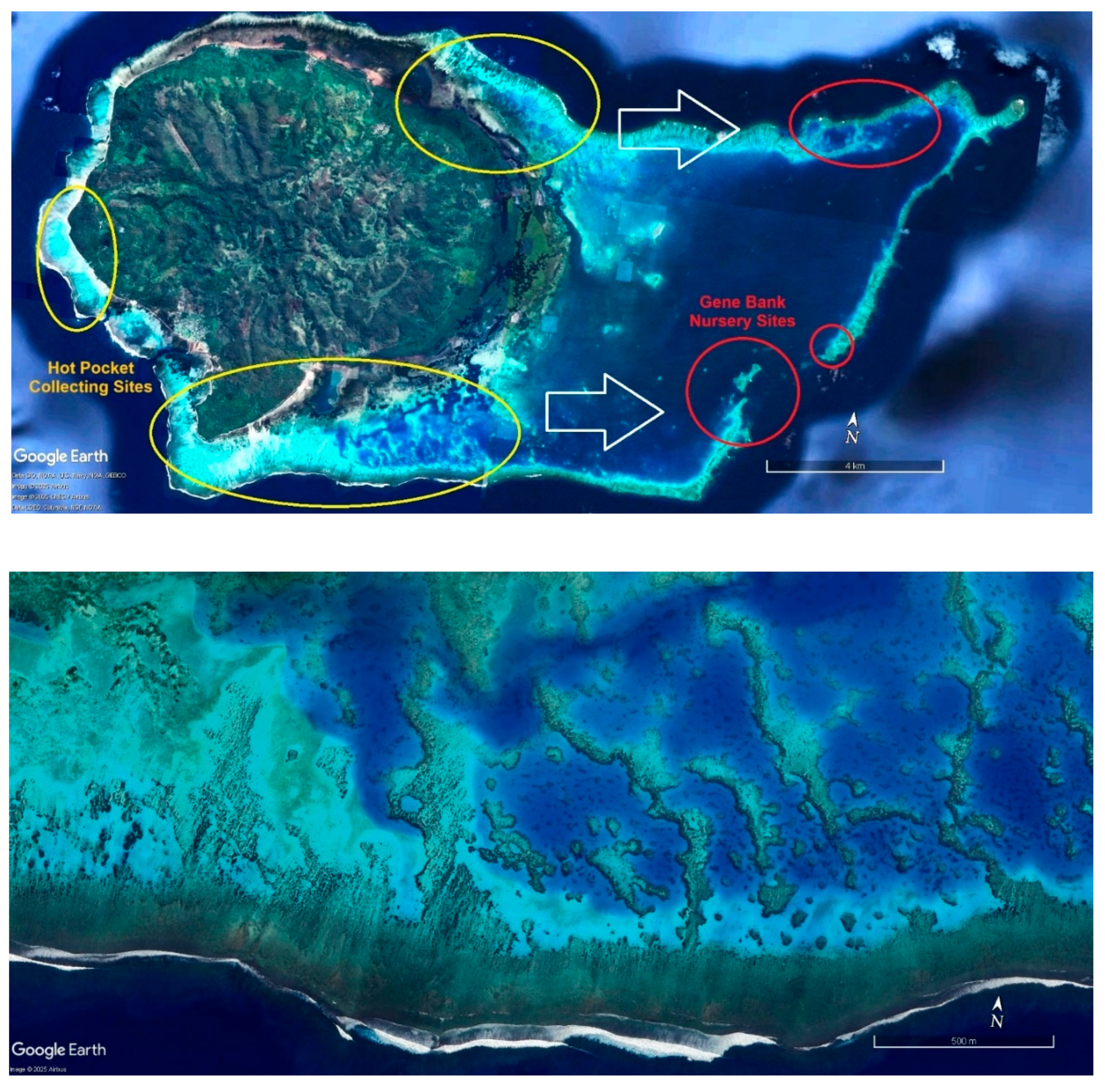
Figure 8.
Kadavu, with regular air service, offers easy access to ideal hot pocket collecting sites and sheltered nursery sites. Shallow nurseries allow for continuing moderate heat stress, faster growth rates, and cheaper scuba-free access, as well as higher community involvement.
Figure 8.
Kadavu, with regular air service, offers easy access to ideal hot pocket collecting sites and sheltered nursery sites. Shallow nurseries allow for continuing moderate heat stress, faster growth rates, and cheaper scuba-free access, as well as higher community involvement.
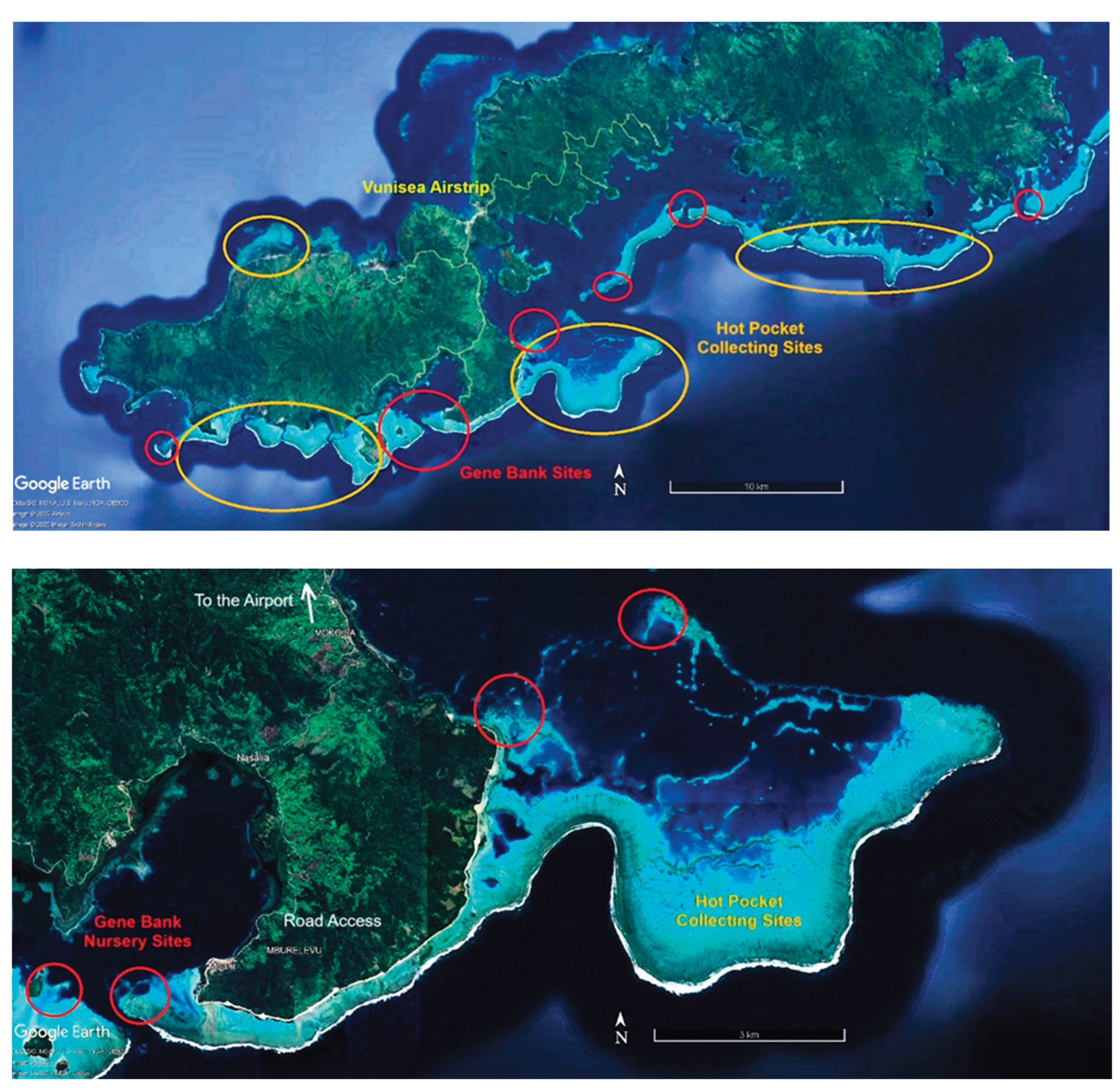
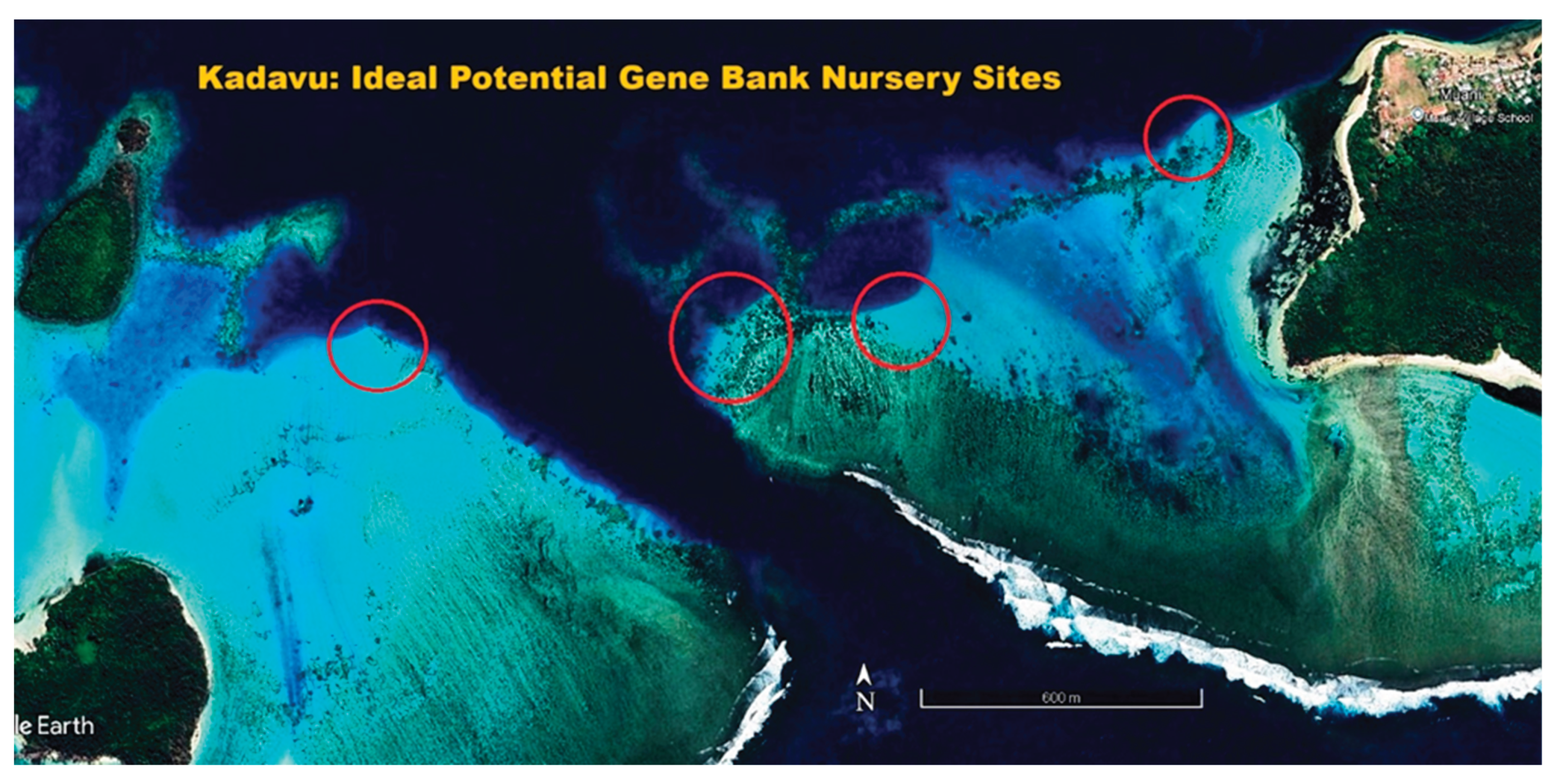
Figure 9.
Self-cleaning mixed rope and table nursery in Fiji, located in 2-3 meters of water in a moderately heat stressed site that reaches 32-33C during hot season. Note the A-frames and a small table nursery installed to create “fish bridges” to improve fish access to the nursery.
Figure 9.
Self-cleaning mixed rope and table nursery in Fiji, located in 2-3 meters of water in a moderately heat stressed site that reaches 32-33C during hot season. Note the A-frames and a small table nursery installed to create “fish bridges” to improve fish access to the nursery.

Figure 10.
Rope nursery over sand. Each rope represents a singe genotype of a species. Spaces indicate coral mortality from a severe Lyngbya infestation before the nursery was moved. Narrow table nurseries with branching corals provide fish habitat, which clean the ropes. This nursery needs support rods to prevent ropes from sagging as they become heavy.
Figure 10.
Rope nursery over sand. Each rope represents a singe genotype of a species. Spaces indicate coral mortality from a severe Lyngbya infestation before the nursery was moved. Narrow table nurseries with branching corals provide fish habitat, which clean the ropes. This nursery needs support rods to prevent ropes from sagging as they become heavy.

Figure 11.
Temporarily shaded rope nursery during the summer season, preventing reoriented coral fragments from being burned by high UV levels.
Figure 11.
Temporarily shaded rope nursery during the summer season, preventing reoriented coral fragments from being burned by high UV levels.

Figure 12.
Table nursery with corals attached to large cement disk “cookies”. Holes in the disk allow for attachment to the tabletop via cable ties for areas of higher current potential. This is a self-cleaning nursery, as the fish population help keep the corals healthy and clean.
Figure 12.
Table nursery with corals attached to large cement disk “cookies”. Holes in the disk allow for attachment to the tabletop via cable ties for areas of higher current potential. This is a self-cleaning nursery, as the fish population help keep the corals healthy and clean.

Figure 13.
Mesh A-frame out planting structures. Multiple genotypes of a single coral species are planted together to restore sexual reproduction. If located on sand, while predators may have a hard time finding the corals, but the impact on recruitment-based recovery may be limited, as sand is not a suitable settlement surface for incoming coral larvae.
Figure 13.
Mesh A-frame out planting structures. Multiple genotypes of a single coral species are planted together to restore sexual reproduction. If located on sand, while predators may have a hard time finding the corals, but the impact on recruitment-based recovery may be limited, as sand is not a suitable settlement surface for incoming coral larvae.

Figure 14.
West Nuku reef flat hot pocket, which is part of a thermal stress zone some 900 meters wide. Texture and color differentiate between corals and seagrass/macroalgae, with isolated coral patches seen beyond the sargassum and seagrass zone. The low coral cover “dead zone” of deeper waters is thought to be due to COTS predation, as aggressive coral-defending Stegastes farmer fish which on the reef flat help prevent COTS from killing corals within their territories, are absent in these deeper waters.
Figure 14.
West Nuku reef flat hot pocket, which is part of a thermal stress zone some 900 meters wide. Texture and color differentiate between corals and seagrass/macroalgae, with isolated coral patches seen beyond the sargassum and seagrass zone. The low coral cover “dead zone” of deeper waters is thought to be due to COTS predation, as aggressive coral-defending Stegastes farmer fish which on the reef flat help prevent COTS from killing corals within their territories, are absent in these deeper waters.

Figure 15.
Bula Rescue Nursery. Rebar and mesh table nurseries spelling out BULA meaning “life” in Fijian. At 45 x 16 meters, this is likely the largest rescue nursery on earth. The 24-foot boat in the photo gives perspective.
Figure 15.
Bula Rescue Nursery. Rebar and mesh table nurseries spelling out BULA meaning “life” in Fijian. At 45 x 16 meters, this is likely the largest rescue nursery on earth. The 24-foot boat in the photo gives perspective.

Figure 16.
Nuku middle-lagoon barrier reef with the Bula gene bank nursery site and other nurseries marked and he main collecting areas are indicated. The Malolo double barrier and fringing reef system is one of many examples of shallow reef systems in Fiji with extensive hot pocket areas of restricted water flow, and with a strong hot to cool thermal gradient from nearshore to offshore. Sheltered pools of the forereef areas of Nuku Reef serve as ideal nursery sites, while abundant out-planting sites are located on bleaching impacted cooler-water outer reefs, where COTS are also less abundant presently due to low coral cover.
Figure 16.
Nuku middle-lagoon barrier reef with the Bula gene bank nursery site and other nurseries marked and he main collecting areas are indicated. The Malolo double barrier and fringing reef system is one of many examples of shallow reef systems in Fiji with extensive hot pocket areas of restricted water flow, and with a strong hot to cool thermal gradient from nearshore to offshore. Sheltered pools of the forereef areas of Nuku Reef serve as ideal nursery sites, while abundant out-planting sites are located on bleaching impacted cooler-water outer reefs, where COTS are also less abundant presently due to low coral cover.

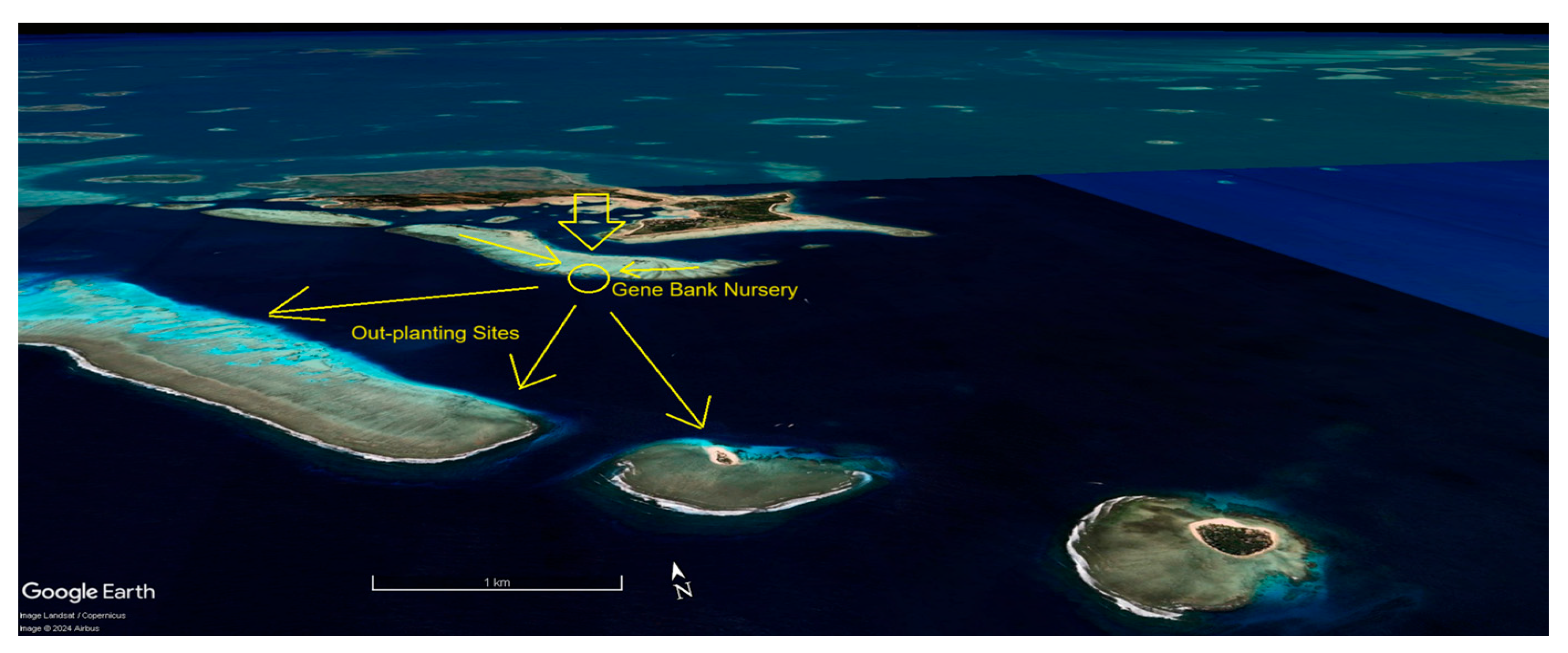
Figure 17.
Bula Nursery showing a diversity of (not yet fully sorted) corals. At one year, the nursery has for the most part become self-cleaning due to resident and visiting fish.
Figure 17.
Bula Nursery showing a diversity of (not yet fully sorted) corals. At one year, the nursery has for the most part become self-cleaning due to resident and visiting fish.

Figure 15.
Severe Lyngbya infestation of an early-stage coral nursery in 2019, before an abundant fish population could develop, compounded by being disconnected from the reef.
Figure 15.
Severe Lyngbya infestation of an early-stage coral nursery in 2019, before an abundant fish population could develop, compounded by being disconnected from the reef.

Figure 16.
Proposed key sites for full implementation of a national Coral Reef Adaptation Plan for Fiji. Sites were selected based on ideal adaptation potential for thermal resistance, ease of access (roads, dependable boats, or airports), and existing support for marine conservation.
Figure 16.
Proposed key sites for full implementation of a national Coral Reef Adaptation Plan for Fiji. Sites were selected based on ideal adaptation potential for thermal resistance, ease of access (roads, dependable boats, or airports), and existing support for marine conservation.
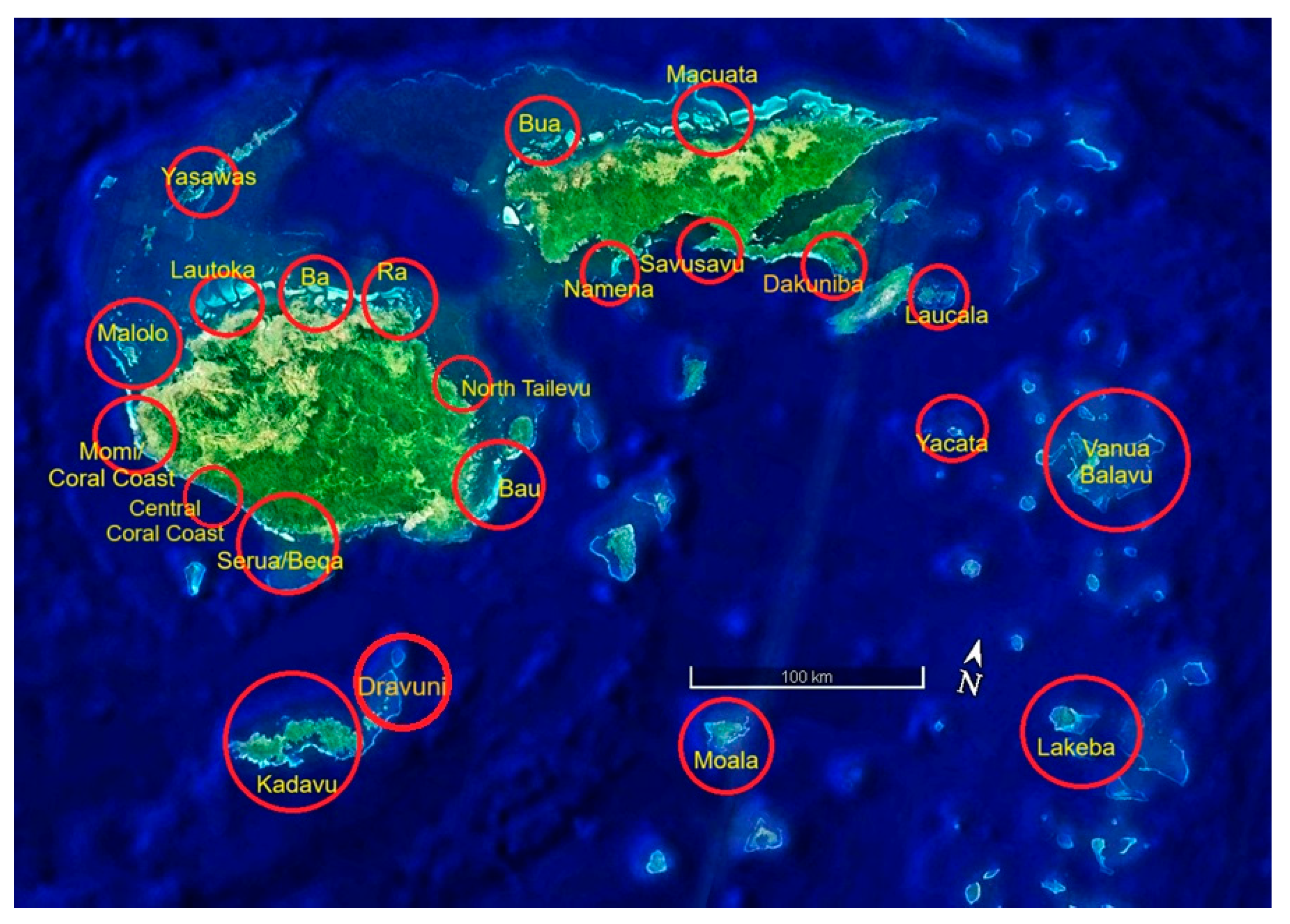
Figure 18.
Mangrove islands on restricted-flow lagoons off the northern coast of Bua, Vanua Levu. Such closed lagoon mangrove environments remote from freshwater runoff present a potential source of both low pH adapted and heat adapted corals.
Figure 18.
Mangrove islands on restricted-flow lagoons off the northern coast of Bua, Vanua Levu. Such closed lagoon mangrove environments remote from freshwater runoff present a potential source of both low pH adapted and heat adapted corals.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



