
Submitted:
17 July 2025
Posted:
18 July 2025
You are already at the latest version
Abstract
The response of the hypothalamic-pituitary-thyroid (HPT) to energy demands is perturbed by previous chronic stress perceived during the neonatal or the adult periods, We examined sex-specific effects of chronic variable stress (CVS) during adolescence on the responses of these axes and target peripheral tissues to volun-tary wheel-running (Ex) in adult rats. Two weeks of Ex stimulated mediobasal hypothalamus (MBH)-Pomc expression in both sexes, but this response was blunted by CVS. CVS altered basal HPA activity only of adult males increasing paraventricular nucleus (PVN) Gr and arcuate-Npy expression, and serum Cort levels. Ex reduced serum Cort concentration in controls. CVS decreased MBH-Dio2 and PVN-Trh expression, raising total and free T3 levels in females. In males, CVS reduced MBH-Dio2, -Trhde, and PVN-Trh expression, impairing exercise-induced expression and reducing serum T3/T4 ratio. The exer-cise-induced gene-expression of Dio2 and Pgc1a in skeletal muscle, of Adrb3, Dio2, Pparg, Hsl in inguinal and gonadal white-adipose tissues, and of Adrb3 and Ucp1 brown adipose tissue was blunted by CVS in both sexes except Hsl in in-guinal of both and BAT-Dio2 in brown adipose tissue of females that were not affected by stress. These results support that adolescent stress impacts adult met-abolic and neuroendocrine responses to exercise in a sex-specific manner.
Keywords:
HPT axis
; HPA axis
; chronic variable stress
; exercise
; adolescence
; sex differences
; skeletal muscle
; white adipose tissue
; brown adipose tissue
1. Introduction
Modern societies bear the burden of a wide range of metabolic, cardiovascular, and psychological pathologies, many of which stem from a sedentary lifestyle and/or chronic stress, even in the early stages of development [1,2,3]. Exercise not only activates metabolism but also reduces stress and its detrimental effects, such as depression, preventing or ameliorating many of these diseases [4,5]. Exercise performance requires an immediate and adequate supply of energy, which depends on the concerted and efficient responses of the somatic, sympathetic, and neuroendocrine systems, which communicate with participating organs through neurotransmitters, hormones, and exerkines [5,6,7,8]. The hypothalamus is the center of energy homeostasis, as its diverse nuclei decode environmental, nutritional, and psychological signals, integrate them, and send neuronal signals as appropriate commands to the behavioral, sympathetic, and endocrine systems [8]. Neurons of the arcuate nucleus (ARC) expressing POMC or AgRP/NPY sense the energetic status of the organism due to a “loose” brain barrier in this area [8,9]; many of these neurons send afferents to the paraventricular nucleus (PVN) that, through multisynaptic contacts, project to the brainstem, to sympathetic noradrenergic neurons that innervate peripheral organs; in turn, sensory neurons send information back to the brain, establishing bidirectional communication [9,10,11,12,13].
At the PVN level, neuroendocrine control relies on hypophysiotropic neurons that synthesize thyrotropin-releasing hormone (TRH) or corticotropin-releasing hormone (CRH) and release them from nerve terminals at the median eminence, triggering the activity of the hypothalamus-pituitary-thyroid (HPT: TRH: thyrotropin (TSH): T4,T3) [14,15] or hypothalamus-pituitary-adrenal (HPA: corticotrophin-releasing hormone (CRH): corticotrophin (ACTH): cortisol/corticosterone (Cort) axes [16]. Peripheral hormones, such as T3/T4 and glucocorticoids, exert feedback control at the hypothalamic and pituitary levels, maintaining homeostasis [14,15,16,17] (Supplementary Figure 1). T3/T4 and glucocorticoids are pleiotropic hormones and crucial endocrine regulators of energy homeostasis, as they regulate carbohydrate and lipid metabolism in multiple tissues and mobilize energy substrates to oxidizing tissues [17,18]. T3, the transcriptionally active hormone, is formed from T4 released from the thyroid and transformed to T3 by deiodinase 1 (DIO1) in the liver or DIO2 in the median eminence, thyroid, pituitary, and many target tissues, while T3 may be inactivated by DIO3; DIO2 is exquisitely regulated in a tissue-specific manner, setting the concentration of active T3 in situ [19,20]. T3 involved in the feedback of Trh transcription is probably formed mainly by DIO2 present in tanycytes, which are special glial cells lining the third ventricle whose prolongations are near TRH nerve terminals and portal vessels in the median eminence, and express the TRH-degrading ectoenzyme (TRH-DE) that inactivates released TRH into the median eminence before entering the portal vessels, thus regulating the concentration of TRH that reaches the thyrotrophs [21,22,23]. Thus, in “basal conditions” rigorous homeostatic mechanisms keep tissue levels of thyroid hormones with minor alterations mainly due to circadian cycle variations, to feedback inhibition by T3 and to the regulated activity of tissue deiodinases [14,15,19,23].
The elements involved in HPT axis activity are stimulated in a fast and transient manner by energy demands such as acute cold exposure or increased physical activity, which increase Trh expression in hypophysiotropic neurons of the PVN and TSH serum concentration in male rats within the first hour, while serum T4 or T3 concentration increases may be detected later [24,25,26,27,28]. In contrast, situations of negative energy balance such as fasting, food restriction, exhaustive exercise, disease as well as various forms of either acute or chronic stress inhibit HPT axis activity [17,20,21,24,26,29,30]. However, the exercise-induced activation of the HPT axis has been questioned due to controversial reports on circulating levels of TSH or thyroid hormones in either humans or animals, which could be due to the type of exercise or energy status at the time of sampling [31]. Involvement of thyroid hormones in exercise is, however, supported by the diminished exercise performance of hypothyroid animals or humans [32].
After cortical motor command generation, exercise initiates with the contraction of skeletal muscles (SKM) that utilize carbohydrates and lipids as their main energy sources in a proportion that depends on intensity and duration of exercise, nutritional status and sex [8,33]. Muscle contraction provokes the release of myokines and metabolites (such as lactate) that signal the brain and various peripheral tissues to activate key metabolic pathways, such as lipolysis in adipose tissue, followed by adaptive changes in the participating organs [8,34]. Fatty acid oxidation and tissue adaptations continue during early recovery after exercise (up to 4h) [35]. Thyroid hormones (TH) are essential in the maintenance and regeneration of muscle cells, as well as for mitochondrial biogenesis by activating peroxisome proliferator-activated receptor γ coactivator 1α (PGC1-α) [36,37]. TH interact with the adrenergic signaling in a coordinated or synergistic fashion in white and brown adipose tissues (WAT, BAT) where they activate lipolysis, de novo lipogenesis, or thermogenesis [38,39,40]. Furthermore, TH participate in the adequate functioning of the cardiorespiratory system [41].
A diminished activity of the HPT axis may cause effects of hypothyroidism, as demonstrated in adult male rats exposed to a Cort injection or, to two weeks of chronic stress prior to one hour of cold exposure; stressed rats fail to show increased PVN Trh expression or serum TSH concentration, their body temperature decreases, and cold-induced BAT thermogenesis is absent [42,43]. Chronic stress also inhibits the HPT response to voluntary exercise: neither the expected increase in PVN Trh or Pomc expression nor the loss of fat mass is detected [44]. Long-term effects caused by stress perceived during critical developmental periods like that imposed by maternal separation, which causes altered HPA axis reactivity in adults [45], blunt the cold-induced activation of PVN TRH neurons and the expression of BAT thermogenic proteins, as well as the response to exercise in adult rats [46,47]. The effects of stress on HPT responses to food deprivation or exercise are sex dimorphic [44,46,47].
Results obtained when combining stress exposure before cold or voluntary exercise support the interference of stress with the fast and efficient responses of the HPT axis required for adequate energy availability, which led us to propose that it may be one of the causes of subclinical hypothyroidism (SCH) [47,48]. Patients with SCH have high serum TSH concentrations but T4 or T3 within the normal range, hypothermia, exercise intolerance, and complaints of fatigue [32,49]. Women have higher incidence of SCH than men, obesity, thyroid diseases and many of stress-related disorders as anxiety and depression; they are also very susceptible to the long-term effects caused by stress during adolescence [17,48,49,50,51,52,53]. It is during adolescence that circuits between the prefrontal cortex and limbic regions are formed and consolidated, and the HPA axis matures. Stress-disrupting effects may last until adulthood, leading to increased stress reactivity and impulsivity, altered metabolic and neuroendocrine functions, and a high risk of developing psychiatric pathologies [54,55,56,57,58].
To demonstrate that stress-induced HPT axis dysfunction alters the expression of T3 targets in tissues involved in exercise, such as SKM, WAT, and BAT, we studied the long-term effects of chronic variable stress (CVS) during adolescence, a developmental period in which females show higher susceptibility to stress than males. Hence, male and female rats (control or CVS) were exposed to voluntary exercise for two weeks in adulthood; a pair-fed group was included to account for exercise-induced diminished food intake [27,44]. Elements involved in HPT and HPA axes activities (Supplementary Figure 1) were measured to evaluate responses to stress, exercise and food intake; Pomc and Npy expression in the ARC and serum concentration of leptin as sensors of energy status [9,59], and serum triglyceride concentration as an indirect measure of lipid mobilization [60]. We also measured in SKM and adipose tissues the expression profiles of Dio2 that is regulated in some tissues by either stress, glucocorticoids or exercise [37,61,62], of Pgc-1α that mediates biogenesis of mitochondria and oxidative capacity, fatty acid utilization and fiber-type switching [39,40], of adrenergic receptor β3 (Adrb3) responsible of sympathetic-induced activation in all adipose tissues [63], of peroxisome proliferator-activated receptor γ (Pparg), a transcription factor that acts as lipid sensor and is involved in lipolytic and lipogenic actions as well as in exercise-induced browning of inguinal WAT (iWAT), and in glucose homeostasis in perigonadal WAT (pgWAT) [64], of hormone sensitive lipase (HSL, Lipe) that is the rate limiting step of lipolysis [65], and finally of uncoupling protein-1 (Ucp1) responsible for thermogenesis in BAT [66].
2. Results
2.1. Behavioral and Ponderal Variables During Adolescence
Behavioral tests were conducted throughout the CVS procedure (Figure 1A) to evaluate the state of animals during treatment, compared to rats from the control group. At day 7 of CVS (at PND 37), spontaneous physical activity was measured in PAS, the least invasive procedure that quantifies spontaneous locomotor activity; compared to controls (C), only CVS females had reduced ambulatory movements in the center and in the periphery (Figure 1B), whereas the total number of movements was similar to that of males (not shown). At PND 44, rats were subjected to the open field test (OFT), in which CVS females spent less time and covered less distance in the center than C females, while males showed no difference, except that total distance values were lower than those of C females (Figure 1C).
The EPM shows an anxious state more clearly, as the animal fears exploring the open arms of the maze; at PND 51, CVS females spent less time and traveled less distance in the open arms than C females (Figure 1D), whilst C and CVS males had similar values (not shown). OFT was repeated at PND58 and again, only CVS females differed from controls with diminished time and distance in the center (Figure 1E). Males were tested in the EPM at PND 65, and at this time, CVS males showed more anxious behavior than unstressed controls (Figure 1F). Thus, when behavior was evaluated within the time frame of the CVS procedure, the results agreed with various reports on the higher susceptibility of females to stress during adolescence, as they showed increased emotional reactivity in the OFT and EPM, compared to males [76,77,78,79].
Food intake was higher in males than in females during adolescence (Figure 1G), but if calculated by body weight (relative food intake, RFI), it was higher in females [44] (Supplementary Table 3). Stressed females increased their food intake during the last two weeks of the CVS procedure (Figure 1G) with no difference in body weight (BW) compared to unstressed rats (Figure 1H). CVS males ate as controls (Figure 1G) but had a lower BW by PND 70 (Figure 1H), resulting thus in a higher RFI and a lower food efficiency (FE) than controls as reported [78] (Supplementary Table 3).
2.2. Long-Term Effects of CVS on Voluntary Exercise
WR is a less stressful paradigm compared to other types of aerobic exercise, in which rats do not run continuously but in bouts, and more intensely during the first hours of dark period [69,80]. Rats were sacrificed three hours after lights were turned on, cage-changed and fresh food provided, at an unknown time after the last running bout; biochemical measurements correspond therefore to a recovery period from the last exercise bout and an unknown postprandial time.
Plots of individual distance covered per day revealed various peaks in C-Ex females, many separated by 4, some by 5 days, coinciding with the estrous cycle stage in rodents [81] (Figure 2A), which contrasts with the flat trend observed in C-Ex males (Figure 2C). Similar patterns were observed in the CVS-Ex groups (Figure 2B,D). Females gradually increased their daily running distance per day (Figure 2A,B). Females had a higher level of locomotion than males in the running wheel (Figure 2A,C), almost five times more (total distance run: females: C-Ex 23.27 km, CVS-Ex 25.74 km; males C-Ex 5.892 km, CVS-Ex 4.992 km) as reported [44,47,80] and observed with these rats in the OFT and EPM performed during adolescence (Figure 1C, E).
Vaginal smears were not taken before exercise (to avoid additional stress) but immediately after decapitation to verify the effect of experimental conditions on the cycle, as stress diminishes serum estradiol concentration or affects cyclicity [82]. At the end of the experiments, the four phases of the cycle were distributed similarly in control and stressed groups, with slight variations in the % of rats in diestrous or metaestrous (Figure 2E). Mean values of the serum concentrations of estradiol and prolactin were not different between experimental groups except for progesterone whose concentration increased in the CVS pair-fed rats (Figure 2F). These results support previous reports suggesting that female rats randomly cycling do not limit overall exercise performance [83].
2.3. Voluntary Exercise Decreases Food Intake
Exercise modifies food intake in a manner that depends on its type and intensity, as well as the animal’s sex and nutritional status [9,59]. As previously published, two weeks of WR diminished food intake in both sexes compared to sedentary rats [27,44,47], in both C and CVS rats, especially during the first 5 days, and this gradually normalized later in females (Figure 3A,B). Total food intake was lower in females than in males, either for C-Sed or C-Ex rats (Supplementary Table 4), even though females ran much more than males (Figure 2A-D); C-Ex females diminished food intake by 34% compared to that consumed by C-Sed, while in C-Ex males it was 20% lower (Supplementary Table 4) (females: C-Sed 100±2%, C-Ex 66±3%, CVS-Ex 72±3%; Males: C-Sed 100±13%, C-Ex 76±4%, CVS-Ex 82±5%) [44,47]; this level of reduced calorie intake (less than 40%) is considered mild and beneficial [84]. Sedentary rats gained (7-10%) BW from day 1 to 14th of the exercise trial; males more than females, and only CVS males gained less than controls (Supplementary Table 4). Food restriction (PF-rats) reduced body weight gain (BWg) to 2% of the control values in both sexes. Exercise also decreased BWg, with notable reductions observed in CVS-Ex females and C-Ex males, while no significant loss was detected in CVS-Ex males (Figure 3C,E, Supplementary Table 4).
Lipolysis of WAT provides energy for muscle activity and other oxidizing tissues in situations of negative energy balance, such as food restriction or exercise; the utilization of different fat depots differs according to the task and sex [85]. Compared to C-Sed rats (= 100%), food restriction (C-PF) reduced only pgWAT mass in females (Figure 3D) probably due to their lower food intake, whereas exercise (C-Ex) decreased both visceral (gonadal and retroperitoneal) and subcutaneous (interscapular) WAT mass (Figure 3D). In contrast, CVS in females led to a similar reduction in WAT mass, including rWAT in CVS-PF and CVS-Ex (Figure 3D). Males of both C and CVS groups lost WAT weight only after exercise (Figure 3F). Variations in WAT mass relative to body weight showed similar trends (Supplementary Table 4). Females diminished food intake more than males, results not consistent with the reported higher susceptibility of female than male-pgWAT to mild calorie restriction, but reports are controversial [84,85]. The lack of correspondence between the reduction in BWg and WAT loss supports stress-induced loss of lean mass in males [86].
WAT secretes various hormones, such as leptin, which signals the energy status of the organism. Serum leptin levels correlated positively with iscWAT mass (Figure 3G,J), which agrees with the proportionality of fat mass to leptin blood concentration [87]. Leptin concentration was decreased by food restriction and by exercise in sera of females (Figure 3H), being lower in CVS-PF than C-PF due to the additional inhibitory effect of stress [88]; however, this decrease was counteracted in CVS-Ex (Figure 3H), confirming the effect of exercise in reducing stress [89]. Leptin serum concentration decreased to a similar extent in C-PF, CVS-PF or -Ex in males (Figure 3K) albeit WAT weight did not decrease in PF (Figure 3F) as BWg did (Figure 3E), in agreement with the determining role of general energy status on leptin release and not only of fat mass [87].
Serum triglycerides (Tg) and free fatty acids represent a major energy supply to active tissues; serum Tg concentration is proportional to WAT mass in basal situations, increases in the postprandial state and decreases by removal by active tissues in situations of chronic stress, cold or exercise [27,35,90,91]. There was a clear sex difference in the variations produced by food restriction or exercise. Female Tg serum concentration was lower than that of C-Sed in C-PF, CVS-PF and in C-Ex but not in CVS-Ex (Figure 3I), even though visceral WAT mass decreased in this group. In males, circulating levels of Tg were slightly reduced in C-PF while strongly in both exercised groups (Figure 3L) that did not show as high fat loss as females; these results suggest different dynamics in removal of Tg from blood and probably variations in their source (whether postprandial or from WAT lipolysis). Similar sexual dimorphisms in variation of serum Tg concentrations after exercise have been reported in humans and mice [90,91].
2.4. Long Term Effects of CVS on ARC and Serum Markers of Energy Homeostasis in Mild Food Restriction and Exercise
POMC and NPY expressing neurons of the ARC are key players in energy homeostasis as they sense the energy and nutritional status of the organism, inhibiting or stimulating their activity (respectively), as in situations of negative energy balance like food restriction that decreased ARC Pomc expression and increased that of Npy in PF groups of both females (Figure 4A,B) and males (Figure 4D,E) [87]. As reported for males, exercise increased Pomc expression considerably in C-Ex rats of both sexes (Figure 4A,D) [44,92] despite the decrease in serum leptin concentration (Figure 3H,K), but consistent with the role of ARC POMC neurons in physical activity control, and the existence of different subpopulations of POMC neurons containing particular receptors that respond specifically to different stimuli [9,59]. ARC Pomc expression was reduced in CVS-PF more than in C-PF, particularly in males (Figure 4A,D). Moreover, the exercise-induced increase in Pomc expression observed in controls was abolished in the CVS group of both sexes (Figure 4A,D).
Although leptin decreased similarly after food restriction or exercise suggesting negative energy status in both cases, Npy expression increased in PF rats as published [92,93] but not in exercised rats (Figure 4B,E); both groups ate equal amounts but while in PF reduced food intake was forced, in exercise, it was voluntary. A plausible explanation is that NPY is expressed in distinct populations of neurons within the ARC, such that some of these neuronal populations are activated by food restriction, whereas others are inhibited by exercise training. In the present study, ARC Npy mRNA levels were elevated in CVS-Sed males, consistent with previous reports in chronically stressed animals. Furthermore, Npy expression was further increased in CVS-Ex males (Figure 4E). Females in contrast responded as controls, increasing Npy levels in CVS-PF with no change in CVS-Ex (Figure 4B), with levels correlating negatively with ARC Pomc values in these latter groups (Figure 4C). Tg correlated negatively with MBH-Pomc expression in C-Ex and CVS-PF male rats (Figure 4F). The stress-induced increased expression of ARC Npy in males but not in females may be attributable to Cort and testosterone stimulatory transcriptional effects, absent in females probably due to estrogen inhibitory effects [96,97].
2.5. CVS Alters HPA Responses to Mild Food Restriction and Exercise
Long-term effects of CVS during adolescence on HPA responsivity to acute psychological stressors show sexual dimorphism [76,77,78,79]. We thus evaluated the long-term effects in basal HPA axis activity and in the stress response to food restriction or exercise at adulthood to evaluate changes in HPA axis responses. Basal HPA axis activity was not affected by CVS in adult females, PVN Crh, Avp or Gr expression as well as Cort serum concentration were similar to those of C-Sed (Figure 4G,H). Crh or Avp expression did not change either in CVS-Sed males, but Gr expression and serum Cort concentration increased (Figure 4I,J). The response to calorie restriction differed; in females, PVN Crh and Avp expression diminished in C-PF and CVS-PF and, Gr expression increased in CVS-PF compared to C-Sed; albeit due to the high dispersion of results Crh data did not achieve significance in the CVS-PF group (Figure 4G). In males, expression of Crh and Avp decreased only CVS-PF rats (Figure 4I), as reported in calorie-restricted male rats previously stressed by individual housing [92] or in animals under higher levels of calorie restriction or longer periods [98]. However, all pair-fed groups increased serum Cort concentration as reported [99] (Figure 4H,J); even though females had higher basal Cort concentration than males (C-Sed females: 275±26 ng/mL vs males: 49±3 ng/mL) as reported [47,51], the extent of increase was higher in males (Figure 4H,J).
Exercise prevented the inhibition of PVN-Crh and -Avp expression caused by food restriction in C-PF as levels in C-Ex or CVS-Ex normalized to those of female C-Sed or male CVS-Sed, except for Avp expression in CVS-Ex females which values remained as low as those of CVS-PF (Figure 4G,I). Gr expression increased only in CVS-ex males. In both sexes, serum Cort concentration diminished in C-Ex to lower levels than in C-Sed, consistent with the stress-reducing effect of exercise [80]; even though increased serum Cort concentration by WR has been reported [44,100], this did not occur after CVS as values in CVS-Ex remained as high as in CVS-PF (Figure 4H,J).
2.6. Long Term Effects of CVS on HPT Axis Activity in Mild Food Restriction and Exercise
As we have demonstrated that the small food restriction provoked by exercise inhibits the HPT axis in males [27] we included the pair-fed group to evaluate to distinguish the effects of exercise from those caused by the negative energy balance in the neuroendocrine responses of males and females and, to determine if changes could relate to effects on target organs. At the central level, TRH hypophysiotropic activity is regulated by two important enzymes synthesized in tanycytes, present in the MBH fragment dissected: DIO2, which provides T3 for feedback effects on Trh transcription [21] and TRH-DE, which inactivates released TRH at the median eminence [22,23]; both enzymes are modulated by fasting [21,101,102].
In females, MBH Dio2 expression decreased by stress (CVS-Sed) and as reported, increased by exercise (Figure 5A) [27,44]; expression was lower than in C-Sed in all CVS-groups, slightly less in CVS-PF and CVS-Ex groups (Figure 5A). MBH Trhde expression was not modified in either group. PVN-Trh expression diminished by chronic stress (CVS-Sed) and food restriction (C-PF, CVS-PF) compared to C-Sed (Figure 5A). The inhibitory effect of NPY on PVN Trh expression [21] supports the negative relationship between the expression of both genes that extends to other elements of the axis such as TSH and T4 serum concentrations (Figure 5B).
There are controversial reports on the effects of calorie restriction on male and female rodents, but conditions vary considerably between groups [84]. In our conditions, reduction of circulating TSH concentration by calorie restriction was detected only in CVS-PF, as reported in young rats kept in individual cages and under 33% food restriction for 3 weeks [102]. Serum TSH concentration also diminished in C-Ex but not in CVS-Ex, unlike the pattern detected in PVN Trh expression, supporting other factors and dynamics involved at hypothalamic and pituitary levels. No significant effects were observed in the serum concentration of T4 (Figure 5C) while that of T3 increased in C-PF and CVS groups; the serum T3/T4 ratio increased in C-PF, C-Ex and CVS-Sed females but not in CVS-PF or CVS-Ex due to the increased T4 concentration (Figure 5C).
Free thyroid hormones were quantified only in experiment 2 (n=5-6/group); serum fT3 concentration of all CVS groups was higher than in sedentary controls supporting a main effect of chronic stress; the serum fT3/fT4 ratio, an index of metabolic conversion of T4 to T3 [103], followed the trend of fT3 changes but, was significantly different only in CVS-Ex (Figure 5D). Changes in serum fT4 concentration correlated positively with progesterone serum concentration in C-Sed whereas PVN Trh expression and serum fT3/fT4 concentrations did in the CVS-Sed and CVS-EX group, respectively (Figure 5E).
In males, unlike the reported upregulation of hypothalamic Dio2 after 25 days of 40% food restriction in male rats [104], it was not affected after of 14 days of 25% restriction (C-PF, Figure 6A). Similar to females, and as reported [27,44], the expression of Dio2 increased in C-Ex but diminished in all CVS groups (Figure 6A). Expression of Trhde decreased by calorie restriction in control animals. CVS-PF and CVS-Ex Trhde expressions were higher than CVS-Sed but similar to C-Sed (Figure 6A). MBH Trhde expression correlated with that of Dio2, explained by the stimulatory effect of T3 on Trhde expression and activity (Figure 6B) [22,23]. Trh mRNA expression increased in C-Ex while as in females, decreased in C-PF [47] and in all CVS groups (Figure 6A). Circulating TSH concentration decreased by calorie restriction and exercise in controls (C-PF, C-EX) and to a similar extent in all stressed rats (CVS-Sed, -PF, -Ex) (Figure 6C). Serum T4 concentration tended to increase but only C-PF values were significantly higher than C-Sed, while those of C-Ex (p=0.053) or CVS- achieved no significance (Figure 6C). No differences were detected on serum T3 concentration (Figure 6C) even though levels in C-Ex correlated positively with distance ran (Figure 6E). However, the ratio of T3/T4 tended to diminish by stress, significantly only in the CVS-Ex group (Figure 6C).
Serum concentration of fT4 decreased by stress, in CVS-Sed and CVS-PF but not in CVS-Ex whose concentration tended to increase (Figure 6D). fT3 concentration was reduced in C-PF and in CVS-PF or CVS-Ex; thus, in consequence, fT3/fT4 was similar and modified only in stressed groups albeit with high variability; statistical differences were detected between CVS-Sed and CVS-PF and CVS-Ex (Figure 6D). Serum concentrations of C-Ex T4 and T3 correlated positively with distance [27,47]; while fT4/fT3 negatively (Figure 6E).
2.7. Long Term Effects of CVS on Gene Expression in SKM, WAT and BAT in Mild Food Restriction and Exercise
Although changes in the expression of the elements of HPT axis modified by either calorie restriction, exercise or stress are small, the importance of thyroid hormones is evident in cases of thyroid dysfunction that diminish exercise performance [32]. The expression of genes known to be regulated by either exercise and/or thyroid hormones was thus evaluated in tissues involved in exercise such as muscle, white and brown adipose tissues.
T3 modulates multiple processes involved in muscle function and metabolism, including activation of PGC-1α, a key regulator in mitochondrial biogenesis [105,106]. Considering that glucocorticoids decrease Dio2 expression in muscle [107] and exercise stimulates the expression of Dio2 and Pgc1a in muscle of mice and rats after treadmill running, and that Dio2 inhibition attenuates the exercise-induced stimulation of Pgc1a in soleus and gastrocnemius SKM [37], we evaluated if chronic stress would alter their expression or the effect of voluntary exercise. Dio2 and Pgc1a expression levels indeed increased after WR in C-Ex, the latter more in females than males, while pair-feeding caused no effect in either sex, except for a small increase in Dio2 expression of female rats (Figure 7A,B). Stress decreased Dio2 expression in CVS-Sed males, and of Pgc1a in CVS-PF females, while preventing the exercise-induced increase of both genes detected in C-Ex, though only partially in Pgc1a of CVS-Ex males (Figure 7A,B). TSH serum concentration correlated with Dio2 expression in C-Sed female and C-Ex male rats coincident with the TSH stimulatory effect reported in BAT of cold exposed male rats and in cultured brown adipocytes [108,109].
2.7.1. Inguinal and Perigonadal WAT (iWAT, pgWAT)
Lipolysis is activated in adipose tissues by the stimulated sympathetic nervous system through the adrenergic receptor β3 (Adrb3) [60] which stimulates PKA-induced phosphorylation of deiodinase 2 and of hormone-sensitive lipase, as well as the synthesis of Dio2 and of PPARg, a transcription factor involved in many aspects of fatty acid metabolism and glucose homeostasis [110]. We thus evaluated the expression of these genes in subcutaneous inguinal WAT that undergoes several adaptations to exercise, and in visceral WAT that provides fuel in negative energy situations whose utilization differs according to the task and sex [85,111].
Expression of the adrenergic receptor β3 (Adrb3) decreased by chronic stress in both WAT depots of female and male CVS-Sed rats (Figure 8A-D) which could relate to the inhibitory effect of dexamethasone on cultured adipocytes [112,113]. This receptor synthesis was also diminished by food restriction in either C- or CVS-PF rats of both sexes (Figure 8A-D) as reported in male rats submitted to conditions of higher calorie restriction [114,115]. A similar expression profile was detected for Dio2 after food restriction, yet inhibited by stress only in CVS-Sed males (Figure 8B,D). Pparg expression differed among WAT depots, it was slightly decreased in iWAT of C-PF females but more in pgWAT, whose levels were lower than C-Sed in C-PF of both sexes and lowest in CVS-PF of males (Figure 8A,D); the higher reduction in CVS-PF is consistent with the inhibitory effect of Cort [116] and of stress-induced inhibition of this transcription factor in visceral WAT proposed as one of the mechanisms responsible in stress-induced diminished glucose tolerance [64].
Exercise stimulated the expression of Adbr3 as expected [111,117,118] so as that of Dio2 and Pparg [119] in both WAT depots (Figure 8A-D), increasing similarly in pgWAT and iWAT of females (Figure 8A,C) and pgWAT of males (Figure 8D) while only barely in males-iWAT (Figure 8B). Stress prevented the exercise-induced stimulation of this receptor in iWAT, levels of CVS-Ex remained as low as CVS-PF compared to a lesser inhibition in pgWAT since levels were higher than CVS-Sed though still lower than those of C-Ex (Figure 8A-D). Dio2 expression followed that of Adrb3 in all groups (Figure 8D) except in CVS-Ex of female iWAT whose levels were higher than C-Sed or CVS-Sed but lower than C-Ex (Figure 8A) consistent with the sex differences in WAT response to activation [63]. Exercise induced stimulation of Pparg similarly in all groups and this was curtailed by CVS.
In contrast to the general inhibition caused by food restriction on the expression of the analyzed genes, (Figure 8, Supplementary Figure 2), that of hormone sensitive lipase differed. HSL is the rate limiting enzyme in triglyceride hydrolysis [65], whose activity is regulated by β-adrenergic receptor-stimulated cAMP-PKA induced phosphorylation [65] and its expression is increased by glucocorticoids, exercise and calorie restriction [65,119,120,121]. Hsl (Lipe) expression increased by exercise and also by food restriction in both fat depots of C-Ex and C-PF, more in males than females even though females ran much more than males (Figure 8A-D). CVS increased Hsl expression in iWAT of CVS-Sed males, and more in CVS-PF and CVS-Ex than CVS-Sed in both sexes but higher stimulation in males than females (Figure 8 A,B). In contrast, Hsl expression of pgWAT was inhibited in CVS-Sed more in females than males, values remained low in CVS-PF or CVS-Ex except for a small increase in CVS-PF compared to CVS-Sed of males (Figure 8C,D).
Various parameters of the HPT axis correlated positively with the expression of Adrb3 in pgWAT of both sexes (Figure 8E,G) similarly with iWAT (Supplementary Figure 5,6) and with Hsl in iWAT (Figure 8F,H). A positive effect of T4 on the expression of Adrb3 has been reported [38]. Positive correlation between HPT and Hsl coincides with lipolytic actions [39].
2.7.2. Brown Adipose Tissue
BAT is a highly metabolically active adipose tissue activated by cold exposure where norepinephrine and T3 act in synergy, enhancing thermogenesis by the energy released from mitochondrial membranes due to the uncoupling protein [66]. Involvement of BAT in exercise has been questioned due to conflicting results, but its participation in lipid and glucose homeostasis, as well as its activation during exercise, is now recognized [84,119,123]. BAT response to either stress, food restriction or exercise varied in a sex dependent manner; mRNA levels of Adrb3 and of Ucp1 decreased by stress in CVS-Sed males, and by calorie restriction in C-PF males (C-Sed) (Figure 9C). In contrast, Adrb3 expression increased in stressed males and females under calorie restriction (CVS-PF) (Figure 9A,C). Exercise enhanced mRNA levels of Adrb3, Dio2 and Ucp1 in C-Ex males and females (Figure 9A,C) whereas stress prevented the exercise-induced stimulation of Adrb3 and Ucp1 in CVS-Ex females and of Dio2 and Ucp1 in CVS-Ex males (Figure 9C) [62]. Adrb3 levels of CVS-Ex males were as low as those of CVS-PF (Figure 9C) while Dio2 expression in female increased to the same level as in C-Ex (Figure 9C). Adrb3 expression varied proportionally to Cort concentrations only in females where it was stimulated in CVS-Sed, while decreased in males, consistent with the dual effect of Cort on the expression of this receptor [124].
2.8. Correlation Analyses
Linear correlation analyses revealed several changes in either gene expression or serum hormone concentrations that were proportional to variations in several of the variables measured (some depicted in corresponding figures) and also among different tissues; although they do not represent a causal relationship, various correlations reflected known physiological functions (mentioned in various figures) and thus supported the reliability of results. Representing the correlations among the different variables in Hive plots allowed us to recognize many associations in the exercised rats between components of the HPT axis and target molecules in peripheral tissues (Figure 10; note: abbreviations of tissues and genes in this section correspond to those used in the hive plots).
Exercise induced associations in with various elements of the HPT axis (Figure 10). For example, in females Trh related negatively with scW, FI consistent with the inhibitory action of TRH on food intake [125]; TSH correlation with gW Pprg, BAT D2, BAT Ucp1, scW mass, and FI, coincides with known effects of TSH [109,122]. Stress hindered most of C-Ex correlations but generated others, more negative than in C-Ex (Figure 10 and Supplementary Figure 5,6). Many more were related in C-Ex males than in C-Ex females as for example, in males, variations of Trh and TSH values associated positively with M D2, gW Ab3 and BAT D2 and negatively with gW D2; Trh correlated positively with BAT Ab3, negatively with T4, T3 and gW Hsl, while TSH positively with PRL, gW Hsl, negative with iW Hsl and Crh.
Compared to the exercised controls, less associations were found in the pair-fed groups and even less in sedentaries, many of them different from those registered in exercised groups (Supplementary Figure 3,4). Hive plots illustrated various “hubs” where one variable associated with several others, for example between the concentrations of Npy and with several variables in C-Pf CVS-Sed and C-Pf of both sexes and CVS-Ex of females supported by the NPY effects on lipid and carbohydrate metabolism, energy expenditure and body temperature regulation [126]. TSH showed over 3 different associations in CVS-PF of females as well as the C-PF, CVS-Pf, C-Ex and CVS-Ex of males (Supplementary Figure 3,4); T4 in CVS-Sed and CVS-Ex females whereas T3 in CVS-PF and CVS-Ex females; multiple associations are justified by the effects of TH and TSH including the presence of TSH-receptor in several tissues [108,109,122]. Correlation of Trh with elements of SkM and BAT is supported by the effect of TRHergic neurons of the PVN on sympathetic neurons that contact these tissues [127].
Importantly, Hive plots revealed several correlations among tissues that support the concerted responses of various tissues in response to energy demands. These include muscle Dio2 in CVS-Sed females and of males to a lesser degree, so as in C-PF. Males displayed various hubs between tissues: gWPparg in C-PF and CVS-Ex; iW Pparg in C-PF, iW Hsl in C-Ex, UCP1 in C- and CVS-Sed (Figure 10, Supplementary Figure 4).
3. Discussion
Chronic stress has been implicated in various metabolic and psychiatric disorders, especially when experienced during critical developmental periods, such as the perinatal stage or adolescence, leading to long-lasting effects [3,54,128]. The ameliorative effects of exercise on stress-induced disruptions are actively investigated, with most research focusing on exercise performed before or concurrently with stress exposure [129]. Fewer studies address the immediate or long-term effects of chronic stress prior to exercise on performance and plasticity [130]. We previously reported that rats exposed to stress during the neonatal period or adulthood exhibit inhibited HPT axis responses to energy demands, such as cold exposure or exercise. Here, we examined how chronic stress during adolescence affects the neuroendocrine and metabolic responses to exercise in adult male and female rats. Adolescence was chosen to ensure detectable long-term effects in females, who are more resilient to stress at other stages of life [76,78,131].
Behavioral tests during adolescent chronic variable stress (CVS) confirmed that females exhibited stress effects from early puberty [77,131,132], whereas males showed effects lasting until late adolescence, possibly reflecting a near adult stage [57,133]. The results summarized in Table 1 reveal the differential stress effects on adult responses to negative energy challenges, such as mild calorie restriction (CR) and voluntary exercise. Mild CR inhibited several studied parameters, while exercise stimulated them; prior chronic stress prevented these exercise-induced effects. Validation of the paradigm through the evaluation of HPA axis components in adulthood revealed persistent effects of adolescent CVS in both sexes. Although behavioral tests confirmed increased anxiety in CVS females, basal HPA axis alterations were only observed in males. CVS sedentary males showed activation characterized by elevated serum corticosterone, PVN glucocorticoid receptor, and ARC Npy expression [76,77]. Previous studies on adult male CVS effects have been inconsistent, reporting either no changes in PVN Trh or circulating TSH/thyroid hormones [43,134] or basal hormonal suppression [29,30,135,136].
In our study, adolescent CVS reduced PVN-Trh expression and serum-free T4 levels in males, as well as MBH Dio2 and Trhde expression. Such reductions could favor HPT axis activation by diminishing inhibitory T3 feedback and enhancing Trh transcription and TRH availability to the thyrotrope. However, clarification requires kinetic studies, as these changes may be transient. In females, PVN-Trh and MBH-Dio2 also decreased slightly, whereas total and free T3 serum concentrations increased (Table 1; detailed T3 discussion follows).
Consistent with previous reports [27,44], exercised rats consumed 24–28% less food than controls, paralleling CR levels known to extend lifespan. Responses to CR differ by sex: females lose less white adipose tissue (WAT) and lower energy expenditure more than males at equivalent intake (30 or 40% CR) [84,114]. Here, females ran more and consumed less than males, explaining findings such as gonadal WAT loss, which was evident only in pair-fed females. Most expected metabolic changes appeared in both sexes at varying degrees. Exceptions included similar leptin reductions and ARC Pomc decreases with ARC Npy increases in control pair-fed rats [87,88,98]. CR inhibited central HPA activity in pair-fed females (reduced PVN Crh and Avp expression), paralleling findings in CVS females post-stress [78]; both CVS pair-fed females and males showed diminished PVN Crh and Avp mRNA levels [92,98]. The CR-induced PVN Crh and Avp suppression in pair-fed females does not seem to be mediated by feedback since serum corticosterone was similarly elevated across groups, although PVN glucocorticoid receptor upregulation was observed only in CVS males, including sedentary animals. Despite elevated corticosterone, CR in pair-fed males did not alter HPA activity or PVN Crh expression in CVS males, potentially attributable to MBH AGRP neurons stimulating CRH release at the median eminence without affecting PVN Crh [137], supported by increased MBH Npy and its correlation with serum corticosterone in CVS pair-fed males, indicating the involvement of NPY/AGRP neurons.
HPT axis activity decreases at multiple levels during negative energy balance [138]. Even mild CR inhibited PVN Trh expression more in females than in males among pair-fed controls; levels in CVS pair-fed groups matched those in CVS sedentary rats, underscoring the stress effects. Fasting-induced tertiary hypothyroidism is attributed to increased tanycyte Dio2 expression, which produces T3 that suppresses Trh transcription via leptin reduction and corticosterone elevation [21,139]. However, both pair-fed groups showed low leptin and high corticosterone levels, and Dio2 expression was unaffected in pair-fed controls and decreased in CVS pair-fed rats. Moreover, Dio2 was reduced in CVS sedentary animals despite elevated corticosterone levels but normal leptin levels, in contrast to increased Dio2 levels in corticosterone-treated male mice [140]. MBH Trhde expression, T3-responsive, and paralleling Dio2 during fasting [72] were also decreased in CVS sedentary and pair-fed controls. Trhde levels normalized in CVS pair-fed and CVS exercise groups, implying additional factors beyond Dio2-generated T3; POMC neurons may contribute given the positive correlations between Pomc, Dio2, and Trhde in CVS sedentary rats. Circulating TSH levels decreased in pair-fed groups, except in pair-fed females, possibly reflecting the stimulatory effects of estrogen [141]. Thyroid hormone concentrations differed by sex: females exhibited slight T3 increases in control and CVS pair-fed groups and elevated free T3 in the CVS exercise group; males showed decreased T4 (control pair-fed), free T4 (CVS pair-fed), and free T3 (control and CVS pair-fed) levels. The reduction in male T4 levels aligns with CR-induced axis suppression, whereas the elevation in female T3 levels suggests additional regulatory mechanisms.
Exercise imposes a negative energy balance, further enhanced by voluntary food restriction, necessitating careful differentiation between exercise- and intake-related effects. The functional heterogeneity of ARC POMC and NPY neurons is crucial [9,94]. Exercise elevates the firing of ARC POMC neurons and suppresses NPY neurons independently of leptin, despite leptin’s serum decline during exercise [94]. Conversely, chronic restraint stress reduces excitatory input to ARC POMC neurons, thereby diminishing their responsiveness [142]. Exercise stimulated MBH-Pomc expression similarly in unstressed males and females, with females running more than males. Prior CVS exposure attenuated this response, although MBH Pomc expression did not correlate with the distance run, likely due to neuronal heterogeneity in receptors, co-transmitters, and projection targets [9,93,94].
Voluntary exercise does not induce exhaustion or stress; instead, it promotes neuroplasticity, increases Bdnf, and reduces glucocorticoid receptor expression in the hippocampus and frontal cortex, thereby enhancing cognition and reducing stress [100,129,143,144]. Correspondingly, exercise normalized PVN Crh and Avp expression suppressed by food restriction and lowered serum corticosterone below sedentary controls in both sexes, although these effects were absent in the CVS exercise groups. Whether prolonged exercise can counteract CVS effects and sex differences therein remains to be elucidated [129].
Exercise also triggers mechanisms that restore energy balance, including transient HPT axis activation, as shown by the correlations between exercise distance and circulating thyroid hormones [27,47]. Previously, we used pair-fed controls to isolate exercise effects [27,44,47], finding numerous changes similar to those observed in CR or stress (Figure 11). Only PVN Trh levels were significantly elevated in control exercised males. These subtle and transient HPT axis changes depend on timing, with kinetic studies showing peak PVN Trh expression and serum TSH within one hour post-acute cold exposure [25] or physical activity [26], normalizing thereafter; similar transient serum TSH increases occur after swimming [145] or treadmill running [24,28]. The precise regulation of deiodinases, transporters, and clearance enzymes ensures rapid homeostatic recovery [146]. Except in pathological or extreme conditions, such as fasting [21], HPT axis dysfunction is more apparent during energetic challenges, such as cold or exercise.
Skeletal muscle, a primary thyroid hormone target that is critical for exercise performance, exhibits sex-dependent responses. Thyroid hormones are essential for muscle function and maintenance [17,36,37,105,106,107]. Exercise stimulates Dio2, increasing local T3 and Pgc1a expression [37], observed in control exercised but not in CVS rats. Females had similar exercise-induced Dio2 expression but higher Pgc1a expression than males, potentially linked to elevated circulating T4. Among the analyzed tissues, skeletal muscle Dio2 showed the greatest exercise response (Supplementary Figure 2). A recent study demonstrated increased gastrocnemius T3 and Dio2 levels following repeated mild treadmill running [147]. CVS blunted the Dio2 response, explaining the diminished Pgc1a upregulation. The lack of exercise response in stressed animals suggests impaired energy substrate utilization and muscle fiber maintenance, and elevated glucocorticoids contribute mainly to muscle atrophy [148].
White adipose tissue (WAT) depots supply energy substrates with sex-specific roles [63,85,149]. Exercise induces subcutaneous WAT adaptations, including sympathetic innervation, mitochondrial biogenesis, and metabolic gene expression [150]; visceral WAT regulates glucose homeostasis and inflammatory tone via cytokines [151]. We measured parameters reported as exercise-stimulated or CR-responsive in WAT and reproduced these findings: exercise enhanced, whereas CR reduced adrenergic receptor, Dio2, and Pparg expression. Tissue comparisons (Supplementary Figure 2) revealed that exercise-induced Adrb3 expression was similar in subcutaneous and visceral WAT and BAT in females, while males exhibited higher Adrb3 expression in visceral WAT. Inguinal WAT showed greater susceptibility to CVS inhibition than gonadal WAT or BAT in both sexes, whereas CR effects were uniform in WAT but milder in BAT, which remained at or above the control levels. Exercise increased Dio2 more in perigonadal than inguinal WAT, more so in males, and CR and CVS effects were similarly inhibitory across tissues. Pparg responses in controls did not parallel Dio2, except for lower stimulation in female inguinal WAT compared to perigonadal WAT, blunted by CVS but not inhibited by CR, likely reflecting PPARG’s multifaceted regulatory roles and correlation with TSH and TH in exercised groups, consistent with direct TSH effects and potential myokine-mediated WAT interactions [123,152]. In contrast, Hsl expression responses to exercise or CR differed markedly by adipose depot and sex; exercise stimulated Hsl more in males, particularly in inguinal WAT, but also in pgWAT. Reports on HSL distribution and CR effects are inconsistent [65,121], reflecting regulation via multiple factors: adrenergic agonists rapidly increase HSL enzymatic activity through PKA-induced phosphorylation without altering mRNA levels, while dexamethasone elevates Hsl mRNA over threefold in vitro [120]. CVS caused higher CR- or Ex-induced Hsl expression in iWAT while decreasing it in pgWAT in all groups. Pathological conditions such as metabolic syndrome show altered local GC metabolism [153]; in mice, reduced local GC production due to adipose-specific H6PDH inhibition leads to decreased expression of Hsl and Atgl in epididymal WAT [154], which may contribute to stress-induced obesity in humans.
Coordinated expression patterns across adipose tissues support their integrative functions. BAT, once questioned in exercise roles, is strongly activated by exercise with increases in Adrb3, Dio2, and Ucp1, the latter of which mediates thermogenesis [66,118,119,123]. Recently, Ucp1 induction in male mice has been linked to exercise-induced muscle-derived factors such as IL-16 [155]. Unlike other tissues where CR and stress blunted marker expression and suppressed exercise responses, female BAT was resistant to CVS effects; Dio2 expression in CVS exercised females matched controls, and CVS pair-fed females even exceeded the controls. Beyond energy expenditure, increased BAT DIO2 may contribute to elevated circulating T3 levels [156,157].
T3 concentrations in the CVS pair-fed, CVS exercised, and control pair-fed groups were higher than those in the control sedentary or exercised rats, suggesting additional mechanisms, such as Dio3 inhibition or altered thyroid hormone clearance. DIO3 seems unlikely to explain these results since chronic stress does not affect Dio3 activity in the cerebral brain areas of male rats [61], and fasting induces WAT Dio3 mRNA [158], reducing serum and liver T3 and T4 levels. Furthermore, glucocorticoids suppress Dio3 in vitro and upregulate the expression of liver enzymes involved in thyroid hormone clearance in vivo [158,159,160]. Other players, such as transporters or binding proteins, warrant further investigation. In particular, because CVS-induced stimulation was more pronounced for free T3, this suggests the regulation of additional factors, such as thyroid hormone-binding proteins. Although stress generally inhibits HPT axis activity, elevated T3 levels have been reported in postwar PTSD veterans and other psychiatric disorders, including bipolar I disorder [161,162,163].
Chronic stress blunted nearly all exercise-induced increases in gene expression. Many effects may arise from corticosterone-mediated transcriptional interference via cross-talk mechanisms, including the inhibition of PKA pathways regulating Trh and Crh synthesis in the PVN [164,165] and Dio2 expression in BAT [62]. However, these interactions occur within narrow temporal windows dictated by glucocorticoid receptor dynamics. It is increasingly recognized that chronic stress effects stem less from sustained corticosterone elevations and more from altered brain circuitry modulating neuronal activity and stress perception dynamically [166]. Similarly, exercise-induced hippocampal plasticity reduces stress reactivity and promotes resilience, which requires time—approximately three weeks in females and] in males [167]. Thus, the two-week exercise used here revealed early perturbations from prior stress that may influence subsequent processes.
The effects of chronic stress on HPT axis responses to exercise vary by stress paradigm and sex and are not fully dependent on HPA axis alterations. Unlike stress models previously studied, this study focuses on adolescent-restricted CVS, distinct from adult intermittent restraint or post-weaning social isolation [44] and neonatal maternal separation [47]. Adult restraint stress abolishes exercise-induced WAT loss in both sexes but not PVN- Trh induction in males [44], whereas adolescent CVS does not impair exercise-induced WAT loss but inhibits PVN Trh upregulation. Adolescent isolation induces stronger stress responses in males and reduces exercise-induced MBH Pomc and Dio2 expression in both sexes, similar to our findings [44]. In contrast, exercise rescues fat loss in maternally separated males but not females and suppresses exercise-induced increases in MBH Pomc levels in females [47]. Our study confirms that adolescent CVS causes persistent HPA activation in males, elevated anxiety-like behavior in females, and disrupts exercise-induced gene expression regulating HPT axis activity and peripheral metabolic tissues. The effect of adolescent stress on increased circulating total and free T3 may involve effects on some of the brain circuits involved, as it is at this stage that adolescent circuits mature and females are more susceptible to stress effects; however, this awaits resolution. Future studies should explore whether prolonged exercise can overcome the effects of chronic stress and consider psychological history when evaluating exercise performance clinically.
Caveats: Most exercise studies have addressed dynamic and transient neuronal and molecular responses. Here, voluntary wheel running led to individual variability in the timing and duration of activity. Our analyses represent a snapshot taken three hours post-possible last running session, during the animals’ inactive phase, recovery period, and fluctuating sleep and feeding bouts. Further studies exploring different times of sacrifice and different days of exercise are required.
4. Materials and Methods
4.1. Animals and Experimental Groups
Wistar rats were bred and raised at the Institute’s animal pathogen-free installation, with a 12h light/dark cycle (lights at 7:00h), independent filtered air and positive pressure in each room. The rat colony is renewed every five years with 10 non-related male and 15 female Wistar rats from Charles River Laboratories (MA, USA). Only two people took care of the animal husbandry keeping strict registry of crossbreeding to avoid family inter-crossing [43]. Rats were kept with food (Teklad 2018SX, Envigo, USA) and purified water (in glass bottles to limit endocrine disruptors) ad libitum. All procedures followed the Guide for the care and use of laboratory animals [67], the Mexican norm NOM-062-ZOO-1999, and Institute Bioethics Committee authorizations (No. 273, 318), taking care to keep stress at a minimum [42,43].
Two independent experiments were performed for either males or females. For each experiment, 12 female virgin rats (3 months old) were mated with unrelated males, the day of birth registered, and the litter culled to 10 pups/mother; at weaning (post-natal day [PND] 21) pups of each mother were placed by sex in a cage. Each experiment required 6 groups of 5-6 rats each; 3 groups were submitted to CVS and the other were controls; groups were formed at PND 30 placing one pup of each mother in a different cage, ending with 2 pups/cage using 3 cages/group with no siblings; 9 cages were used as control [C] and the other nine for CVS. CVS rats were housed in a different room from C rats to avoid the stress of noise produced by taking cages out of the room every day for the CVS procedure, and of the noise of stressed rats. Eighteen rats were submitted, during the adolescent period, every day to the CVS paradigm applying one different stressor per day during the light period according to [43,68] modified as described in Supplementary Table 1.
Because the period of adolescence extends over puberty that is restricted to hormonal and physical changes, the period of adolescence was chosen according to [56], from PND 31-60 for females and PND 31-70 for males, to guarantee the inclusion of the sensible periods considering the existing controversies regarding the end of adolescence in rats [54,55,56,57,58] (Figure 1A). Behavioral tests were conducted throughout the CVS procedure to evaluate the state of animals during treatment.
4.2. Behavioral Tests
Rats were submitted to behavioral tests during the light period, starting at 10:00 h. Spontaneous locomotor activity At PND 37 locomotor activity was evaluated by placing rats individually for 5 minutes in a standard cage with a photobeam activity system (PAS-Home Cage, San Diego Instruments Co.) connected to a computer that registers: central and peripheral activities, ambulatory and fine movements, and rearing. Photobeams are placed around the cage and the program can distinguish fine movements from locomotion in the cage.
Open Field Test (OFT)
At PND 44 and 58, rats were placed for 5 minutes and left to explore freely in an enclosed squared arena of 100×100×60 cm made of opaque black acrylic (San Diego Instruments Co.), under dim instead of bright light to distinguish small changes [26]. Total distance traveled (measure of total locomotion), distance and time spent in the center of the area, and the number of entries to the center were registered and analyzed with the SMART 2.5 software (Panlab). Reduced time and distance spent in the center indicate anxiety behavior (inferred by the avoidance of crossing the center of the field).
Elevated Plus Maze (EPM)
At PND 51 male and female rats and only on PND 65 male rats were submitted to the EPM [26] (San Diego Instruments Co.). Rats were placed for 5 min at the center of the EPM, time spent in the open or closed arms, and distance traveled was registered, and analyzed with the software SMART 2.5 (Panlab).
4.3. Voluntary Exercise
Two weeks after the end of adolescence and of CVS or C treatment, rats were exposed to voluntary wheel running (WR), at PND 74 for females, PND 84 for males [55]. Control and CVS animals were divided into 3 groups (5-6 rats/group), avoiding siblings within groups: sedentary (Sed) fed ad libitum, pair-fed (PF) to food intake of exercised rats, and exercised (Ex) [27,44]. C-Sed and CVS-Sed rats were left undisturbed in a different room from Ex and PF groups. As rodents are nocturnal and run most of the time during the dark period [69], C-Ex and CVS-Ex rats were placed individually, only during the dark period (at light change, from 19:00 to 7:00h) for 14 days, in a cage with a running wheel (AccuScan Instruments INC) with fresh food and water; at 7:00h animals were returned with their cage mate with food and water. Food intake was monitored after each cage change (at 7:00h and 19:00h), and distance registered at 7:00h. The average amount of food consumed by the Ex rats at each period was provided to the PF rats; during the dark period control PF rats were placed in individual cages to account for the 12h-stress of isolation [70) that exercised rats had in the cage with the running wheel; during the light period they were returned with their home cage mate.
4.4. Tissue Collection
After 2 weeks of voluntary WR, on day 15th rats were euthanized; each rat was introduced, starting at 9:00h, to the nearby laboratory to be decapitated with a sharp guillotine by an experienced technician. To avoid time differences between groups that may involve circadian variations in hormone or mRNA levels, rats were alternated between the different groups. Trunk blood was collected, serum was separated by centrifugation and stored in aliquots at -20 °C. Vaginal smears were taken immediately after decapitation to avoid stress and analyze by cytology the stage of estrous cycle. The brain was carefully excised from the skull, ensuring the optic nerve was severed beforehand to preserve the median eminence and placed in aluminum foil over dry ice. Peripheral tissues such as gastrocnemius muscle, WAT and BAT were rapidly removed; perigonadal (pg), retroperitoneal (r) and interscapular (isc) WAT depots were weighed fresh before freezing, and inguinal WAT (iWAT) were dissected and frozen without weighing (to avoid RNA degradation due to the dissection time). All tissues were wrapped in aluminum foil and kept at -70 °C. At least five people were involved in the dissection procedure to minimize time between sacrifices.
4.5. Brain Dissections
Frozen brains were placed in a cryostat (Thermo Scientific HM525) for coronal sectioning; after verifying alignment and recognizing the end of anterior PVN in 20 μm slices (toluidine blue staining), three consecutive 200 μm coronal slices were cut, which included the median-caudal PVN (Bregma -1.3 to -1.9 mm), and 5 slices for the medial-basal hypothalamus (MBH, Bregma -1.92 to -2.92 mm) [71]. Each section was placed horizontally on a glass cover of a petri dish over dry ice. For the 3 anterior slices, the PVN from each side of the 3rd ventricle was cut with a 0.5 mm internal diameter sample corer (Fine Science Tools), and stored in tubes at -70 °C. The following 5 slices included the ARC and the median eminence with the lower third of the 3rd ventricle containing the β2 tanycytes that express Dio2 and thyrotropin-releasing hormone-degrading ectoenzyme (Trhde) [22,72]; from each slice, a 0.5 mm internal diameter sample corer was placed at the center and most ventral end of hypothalamus.
4.6. Biochemical Measurements
Hormone quantification in serum
TSH (reagents and protocol from NIDDK, antibody ID AB_2891204) and Cort (Merck-Millipore, antibody ID AB_90543) were quantified by radioimmunoassay and other hormones with ELISA kits following manufacturer’s instructions, except that for total T3 (cat. IIDE 2021, Diagnóstica International, Zapopan, México) and T4 (cat. IIDE 2022, Diagnóstica International), an aliquot of 25 µL of hypothyroid rat serum was added to the standard curves as reported [44]. Free T4 (fT4) (cat. E-EL-0122) and free T3 (fT3) (cat. E-EL-0079) were measured using kits from Elabscience only in sera of rats from experiment 2. Estradiol (cat. 80548), progesterone (cat. 80558), prolactin (cat. 80560) and leptin (cat. 90040) were quantified using kits from Crystal Chem. Samples were measured in duplicate, the mean was taken as n = 1; intra-assay and inter-assay coefficients of variation were <10%.
4.7. RNA Extraction and mRNA Quantification
Total RNA was isolated from PVN, MBH, and gastrocnemius muscle by the acid guanidinium thiocyanate-phenol-chloroform method [73]. For adipose tissues, modifications included a first centrifugation at 870g for 10 min and, an additional chloroform wash to remove fat. Purity and integrity of RNA were verified by electrophoresis and S18 and 28 bands quantified; samples with a 28/18 ratio of less than 1.5 were discarded. One µg RNA was retro-transcribed (M-MLV, 200 U/µL, Invitrogen), cDNA amplified by end-point PCR for MBH, gastrocnemius muscle, BAT and WATs, using recombinant Taq DNA polymerase (Biotecnologías Moleculares, México) and linearity verified against internal markers cyclophilin A (Ppia) or hypoxanthine phosphoribosyl transferase 1 (Hprt) [44,46]. Genes studied in the PVN were quantified by qPCR (Sygreen Mix, PCR Biosystems) on the RotorGene Q (Qiagen) with an initial denaturing stage at 95 °C for 10 min, followed by 40 cycles at 95 °C for 10 s, annealing temperature of 60 °C for 20 s, and a 72 °C extension period for 20 s. Melting curve analysis was performed to verify that only one PCR product was detected in each sample. The relative expression of the target genes with respect to the reference genes was calculated using the Pfaffl method [74]. The expression level of target genes was normalized against Hprt or Ppia, as indicated in figures. End-point PCR and qPCR primer sequences used are described in Supplementary Table 2.
4.8. Statistical Analyses
Two independent experiments were performed/sex, n=5-6/group in each experiment; mean of C-Sed values of each experiment was taken as 100% and the value for each animal calculated as % of C-Sed mean; % values of the two experiments were pooled, calculated as the mean ± S.E.M and analyzed with GraphPad Prism 10. Two-way ANOVA was used to determine the significance of treatment and/or sex for variables measured before exercise; three-way ANOVA to determine the significance of stress, exercise, and sex; food intake and body weight gain were analyzed by two-way ANOVA of repeated measures. In some cases, a two-tailed t-test was used, as stated in figure legends. Statistical significance was set at p ≤0.05. ANOVA data are presented in Supplementary Tables 5A-C. All comparisons were made against C-Sed unless specified otherwise in the text.
We evaluated the Pearson correlation coefficient and the significance of the interaction, between all variables of all tissues measured, using GraphPad Prism 10; for visual analysis of correlation networks we developed hive plots (Figure 10) [75] [created with software: https://codeberg.org/rgarcia-herrera/neuro_endocrine_correlations]. Only significant correlations showing linearity were included, discarding links between two separate sets of data.
Supplementary Materials
The following supporting information can be downloaded at website of this paper posted on Preprints.org, S1: Supplementary tables; S2: Supplementary figures.
Author Contributions
Conceptualization, PJB; methodology, MPMO and KG; hive plots, RGH; statistical analysis, LJH and MPMO; writing—original draft preparation, PJB; writing—review and editing, PJB, JLC and LJH; funding acquisition, PJB. All authors have read and agreed to the published version of the manuscript.”
Funding
This research was funded by DGAPA-UNAM IN213459 and IN217422.
Institutional Review Board Statement
The animal study protocol was approved by the Institutes Bioethics Committee, protocol No. 273, 318.
Data Availability Statement
The original data presented in the study are openly available in FigShare at DOI 10.6084/m9.figshare.29579270.
Acknowledgments
The authors thank the technical assistance of Dr. A. Cote Vélez, B.S. F. Romero, B.S. M. Cisneros, B.S. E. Mata, and S. Gonzalez (UNAM). Supported in part by grants from DGAPA-UNAM IN213459 andIN217422. Dr. M. Parra-Montes de Oca, fellow of the Postgraduate Program in Biochemical Sciences (UNAM), was supported by CONACYT and DGAPA-UNAM fellowships.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| ACTH: | corticotrophin |
| Adrb3: | adrenergic receptor β3 |
| AgRP: | Agouti-related peptide |
| ARC: | arcuate nucleus |
| Avp: | Arginine vasopressin |
| BAT: | brown adipose tissue |
| BW: | body weight |
| BWg: | body weight gain |
| C: | control |
| Cort: | corticosterone |
| CR: | calorie restriction |
| CRH: | corticotropin-releasing hormone |
| CVS: | chronic variable stress |
| Dio1: | deiodinase 1 |
| Dio2: | deiodinase 2 |
| Dio3: | deiodinase 3 |
| EPM: | Elevated Plus Maze |
| Ex: | exercised (voluntary wheel-running) |
| FE: | food efficiency |
| FI: | Food Intake |
| fT3: | free T3 |
| fT4: | free T4 |
| Gr: | Glucocorticoid receptor |
| HPA: | hypothalamus-pituitary-adrenal |
| Hprt: | hypoxanthine phosphoribosyl transferase 1 |
| HPT: | hypothalamic-pituitary-thyroid |
| HSL (Lipe): | hormone sensitive lipase |
| IL-16: | Interleukin-16 |
| iWAT: | inguinal WAT |
| MBH: | mediobasal hypothalamus |
| NPY: | Neuropeptide Y |
| OFT: | Open Field Test |
| P4: | Progesterone |
| PAS: | photobeam activity system |
| PF: | pair-fed |
| Pgc-1α: | peroxisome proliferator-activated receptor γ coactivator 1α |
| pgWAT: | perigonadal WAT |
| PND: | post-natal day |
| POMC: | Pro-opiomelanocortin |
| Ppia: | cyclophilin A |
| Pparg: | peroxisome proliferator-activated receptor γ |
| PRL: | Prolactin |
| PVN: | paraventricular nucleus |
| RFI: | relative food intake |
| SCH: | subclinical hypothyroidism |
| Sed: | sedentary |
| SKM: | skeletal muscles |
| T3: | Triiodothyronine |
| T4: | Thyroxine |
| Tg: | triglycerides |
| TH: | Thyroid hormones |
| TRH: | thyrotropin-releasing hormone |
| TRH-DE (Trhde): | TRH-degrading ectoenzyme |
| TSH: | thyrotropin |
| Ucp1: | uncoupling protein-1 |
| WAT: | white adipose tissues |
| WR: | wheel running |
References
- Kerr,N.R.; Booth,F.W. Contributions of physical inactivity and sedentary behavior to metabolic and endocrine diseases. Trends Endocrinol Metab. 2022;33(12):817-827. [CrossRef]
- McEwen,B.S.; Akil,H. Revisiting the Stress Concept:Implications for Affective Disorders. J Neurosci. 2020;40(1):12-21. [CrossRef]
- Agorastos A.; Chrousos G.P. The neuroendocrinology of stress:the stress-related continuum of chronic disease development. Mol Psychiatry. 2022;27(1):502-513. [CrossRef]
- Hird EJ; Slanina-Davies A; Lewis G; Hamer M; Roiser JP. From movement to motivation:a proposed framework to understand the antidepressant effect of exercise. Transl Psychiatry. 2024;14(1):273. [CrossRef]
- Esteves JV; Stanford KI. Exercise as a tool to mitigate metabolic disease. Am J Physiol Cell Physiol. 2024;327(3):C587-C598. [CrossRef]
- Chow LS; Gerszten RE; Taylor JM; Pedersen BK; van Praag H; Trappe S; Febbraio MA; Galis ZS; Gao Y; Haus JM; Lanza IR; Lavie CJ; Lee CH; Lucia A; Moro C; Pandey A; Robbins JM; Stanford KI; Thackray AE; Villeda S; Watt MJ; Xia A; Zierath JR; Goodpaster BH; Snyder MP. Exerkines in health; resilience; disease. Nat Rev Endocrinol. 2022;18(5):273-289. [CrossRef]
- Murphy,R.M.; Watt,M.J.; Febbraio,M.A. Metabolic communication during exercise. Nat Metab. 2020;2(9):805-816. [CrossRef]
- Hwang,E.; Portillo,B.; Grose,K.; Fujikawa,T.; Williams,K.W. Exercise-induced hypothalamic neuroplasticity:Implications for energy and glucose metabolism. Mol Metab. 2023;73:101745. [CrossRef]
- Quarta C; Claret M; Zeltser LM; Williams KW; Yeo GSH; Tschöp MH; Diano S; Brüning JC; Cota D. POMC neuronal heterogeneity in energy balance; beyond:an integrated view. Nat Metab. 2021;3(3):299-308. [CrossRef]
- Lorsignol A; Rabiller L; Labit E; Casteilla L; Pénicaud L. The nervous system; adipose tissues:a tale of dialogues. Am J Physiol Endocrinol Metab. 2023;325(5):E480-E490. [CrossRef]
- Zhang Z; Su J; Tang J; Chung L; Page JC; Winter CC; Liu Y; Kegeles E; Conti S; Zhang Y; Biundo J; Chalif JI; Hua CY; Yang Z; Yao X; Yang Y; Chen S; Schwab JM; Wang KH; Chen C; Prerau MJ; He Z. -Spinal projecting neurons in rostral ventromedial medulla co-regulate motor; sympathetic tone. Cell. 2024;187(13):3427-3444. e21. [CrossRef]
- Carpentier AC; Blondin DP; Haman F; Richard D. Brown Adipose Tissue-A Translational Perspective. Endocr Rev. 2023;44(2):143-192d. oi:10.1210/endrev/bnac015.13. Mishra G; Townsend KL. The metabolic; functional roles of sensory nerves in adipose tissues. Nat Metab. 5(9):1461-1474. [CrossRef]
- Joseph-Bravo P; Jaimes-Hoy L; Uribe RM; Charli JL. 2015;60 YEARS OF NEUROENDOCRINOLOGY:TRH; the first hypophysiotropic releasing hormone isolated:control of the pituitary-thyroid axis. J Endocrinol. 226(2):T85-T100. [CrossRef]
- Costa-E-Sousa,R.H.; Rorato,R.; Hollenberg,A.N.; Vella,K.R. Regulation of Thyroid Hormone Levels by Hypothalamic Thyrotropin-Releasing Hormone Neurons. Thyroid. 2023;33(7):867-876. [CrossRef]
- Herman JP. The neuroendocrinology of stress:Glucocorticoid signaling mechanisms. Psychoneuroendocrinology. 137:105641. 2021. [CrossRef]
- Sinha RA; Yen PM. Metabolic Messengers:Thyroid Hormones. Nat Metab. 2024;6(4):639-650. [CrossRef]
- Jimeno B; Verhulst S. Meta-analysis reveals glucocorticoid levels reflect variation in metabolic rate; not ‘stress’. Elife. 2023;12:RP88205. [CrossRef]
- Russo SC; Salas-Lucia F; Bianco AC. Deiodinases; the Metabolic Code for Thyroid Hormone Action. Endocrinology. 2021;162(8):bqab059. [CrossRef]
- Köhrle,J.; Frädrich,C. Deiodinases control local cellular and systemic thyroid hormone availability. Free Radic Biol Med. 2022;193(Pt 1):59-79. [CrossRef]
- Fekete,C.; Lechan; R. M. Central regulation of hypothalamic-pituitary-thyroid axis under physiological; pathophysiological conditions. Endocr Rev. 2014;35(2), 159-194; [CrossRef]
- Sánchez E; Vargas MA; Singru PS; Pascual I; Romero F; Fekete C; Charli JL; Lechan RM. Tanycyte pyroglutamyl peptidase II contributes to regulation of the hypothalamic-pituitary-thyroid axis through glial-axonal associations in the median eminence. Endocrinology. 2009;150(5):2283-91. [CrossRef]
- Rodríguez-Rodríguez A; Lazcano I; Sánchez-Jaramillo E; Uribe RM; Jaimes-Hoy L; Joseph-Bravo P; Charli JL. Tanycytes; the Control of Thyrotropin-Releasing Hormone Flux Into Portal Capillaries. Front Endocrinol (Lausanne). 2019;10:401. [CrossRef]
- Ruisseau PD; Taché Y; Brazeau P; Collu R. Pattern of adenohypophyseal hormone changes induced by various stressors in female; male rats. Neuroendocrinology. 1978;27(5-6):257-71. [CrossRef]
- Uribe RM; Redondo JL; Charli JL; Joseph-Bravo P. Suckling; cold stress rapidly; transiently increase TRH mRNA in the paraventricular nucleus. Neuroendocrinology. 1993;58(1):140-5. [CrossRef]
- Gutiérrez-Mariscal M; Sánchez E; García-Vázquez A; Rebolledo-Solleiro D; Charli JL; Joseph-Bravo P. Acute response of hypophysiotropic thyrotropin releasing hormone neurons; thyrotropin release to behavioral paradigms producing varying intensities of stress; physical activity. Regul Pept. 2012;179(1-3):61-70. [CrossRef]
- Uribe RM; Jaimes-Hoy L; Ramírez-Martínez C; García-Vázquez A; Romero F; Cisneros M; Cote-Vélez A; Charli JL; Joseph-Bravo P. Voluntary exercise adapts the hypothalamus-pituitary-thyroid axis in male rats. Endocrinology. 2014;155(5):2020-30. [CrossRef]
- Fortunato RS; Ignácio DL; Padron AS; Peçanha R; Marassi MP; Rosenthal D; Werneck-de-Castro JPS; Carvalho DP. The effect of acute exercise session on thyroid hormone economy in rats. J Endocrinol. 2008;198(2):347-53. [CrossRef]
- Joseph-Bravo,P.; Jaimes-Hoy,L.; Charli,J.L. Regulation of TRH neurons and energy homeostasis-related signals under stress. J Endocrinol. 2015;224(3):R139-59. [CrossRef]
- Kyriacou,A.; Tziaferi,V.; Toumba,M. Stress, Thyroid Dysregulation, and Thyroid Cancer in Children and Adolescents:Proposed Impending Mechanisms. Horm Res Paediatr. 4477;2023;96(1):44-53. [CrossRef]
- Hackney,A.C.; Saeidi,A. The thyroid axis, prolactin, and exercise in humans. Curr Opin Endocr Metab Res. 9, 45–50. 2019. [CrossRef]
- Lankhaar,J.A.; deVries,W.R.; Jansen,J.A.; Zelissen,P.M.; Backx,F.J. Impact of overt and subclinical hypothyroidism on exercise tolerance:a systematic review. Research quarterly for exercise and sport, 85(3), 365–389. 2014. [CrossRef]
- Athanasiou,N.; Bogdanis,G.C.; Mastorakos,G. Endocrine responses of the stress system to different types of exercise. Rev Endocr Metab Disor. 24(2), 251–266. 2023. [CrossRef]
- Murphy,R.M.; Watt,M.J.; Febbraio,M.A. Metabolic communication during exercise. Nature metabolism, 2(9), 805–816. 2020. [CrossRef]
- Lundsgaard,A.M.; Fritzen,A.M.; Kiens,B. The Importance of Fatty Acids as Nutrients during Post-Exercise Recovery. Nutrients, 12(2), 280. 2020. [CrossRef]
- Bloise FF; Cordeiro A; Ortiga-Carvalho TM. Role of thyroid hormone in skeletal muscle physiology. J Endocrinol. 2018;236(1):R57-R68. [CrossRef]
- Bocco BM; Louzada RA; Silvestre DH; Santos MC; Anne-Palmer E; Rangel IF; Abdalla S; Ferreira AC; Ribeiro MO; Gereben B; Carvalho DP; Bianco AC; Werneck-de-Castro JP. Thyroid hormone activation by type 2 deiodinase mediates exercise-induced peroxisome proliferator-activated receptor-γ coactivator-1α expression in skeletal muscle. J Physiol. 2016;594(18):5255-69. [CrossRef]
- Silva JE; Bianco SD. Thyroid-adrenergic interactions:physiological; clinical implications. Thyroid. 2008;18(2):157-65. [CrossRef]
- Mullur R; Liu YY; Brent GA. Thyroid hormone regulation of metabolism. Physiol Rev. 2014;94(2):355-82. [CrossRef]
- Guilherme A; Yenilmez B; Bedard AH; Henriques F; Liu D; Lee A; Goldstein L; Kelly M; Nicoloro SM; Chen M; Weinstein L; Collins S; Czech MP. Control of Adipocyte Thermogenesis; Lipogenesis through β3-Adrenergic; Thyroid Hormone Signal Integration. Cell Rep. 2020;31(5):107598. [CrossRef]
- Müller P; Leow MK-S; Dietrich JW. Minor perturbations of thyroid homeostasis and major cardiovascular endpoints—Physiological mechanisms and clinical evidence. Front. Cardiovasc. Med. 2022;9:942971. [CrossRef]
- Sotelo-Rivera I; Jaimes-Hoy L; Cote-Vélez A; Espinoza-Ayala C; Charli JL; Joseph-Bravo P. An acute injection of corticosterone increases thyrotrophin-releasing hormone expression in the paraventricular nucleus of the hypothalamus but interferes with the rapid hypothalamus pituitary thyroid axis response to cold in male rats. J Neuroendocrinol. 2014;26(12):861-9. [CrossRef]
- Castillo-Campos A; Gutiérrez-Mata A; Charli JL; Joseph-Bravo P. Chronic stress inhibits hypothalamus-pituitary-thyroid axis; brown adipose tissue responses to acute cold exposure in male rats. J Endocrinol Invest. 2021;44(4):713-723. [CrossRef]
- Parra-Montes de Oca MA; Gutiérrez-Mariscal M; Salmerón-Jiménez MF; Jaimes-Hoy L; Charli JL; Joseph-Bravo P. Voluntary Exercise-Induced Activation of Thyroid Axis; Reduction of White Fat Depots Is Attenuated by Chronic Stress in a Sex Dimorphic Pattern in Adult Rats. Front Endocrinol (Lausanne). 2019;10:418. [CrossRef]
- Weaver IC; Diorio J; Seckl JR; Szyf M; Meaney MJ. Early environmental regulation of hippocampal glucocorticoid receptor gene expression:characterization of intracellular mediators; potential genomic target sites. Ann N Y Acad Sci. 2004;1024:182-212. [CrossRef]
- Jaimes-Hoy L; Romero F; Charli JL; Joseph-Bravo P. Sex Dimorphic Responses of the Hypothalamus-Pituitary-Thyroid Axis to Maternal Separation; Palatable Diet. Front Endocrinol (Lausanne). 2019;10:445. [CrossRef]
- Jaimes-Hoy L; Pérez-Maldonado A; Narváez Bahena E; de la Cruz Guarneros N; Rodríguez-Rodríguez A; Charli JL; Soberón X; Joseph-Bravo P. Sex Dimorphic Changes in Trh Gene Methylation; Thyroid-Axis Response to Energy Demands in Maternally Separated Rats. Endocrinology. 2021;162(8):bqab110. [CrossRef]
- Vella KR; Hollenberg AN. Early Life Stress Affects the HPT Axis Response in a Sexually Dimorphic Manner. Endocrinology. 2021;162(9):bqab137. [CrossRef]
- Biondi B; Cooper DS. Thyroid hormone therapy for hypothyroidism. Endocrine. 2019;66(1):18-26. [CrossRef]
- 49. Baksi,S.; Pradhan,A. Thyroid hormone:sex-dependent role in nervous system regulation and disease. Biology of sex differences, 12(1), 25. 2021. [CrossRef]
- Heck AL; Handa RJ. Sex differences in the hypothalamic-pituitary-adrenal axis’ response to stress:an important role for gonadal hormones. Neuropsychopharmacology. 2019;44(1):45-58. [CrossRef]
- Parra-Montes de Oca MA; Sotelo-Rivera I; Gutiérrez-Mata A; Charli JL; Joseph-Bravo P. Sex Dimorphic Responses of the Hypothalamus-Pituitary-Thyroid Axis to Energy Demands; Stress Front Endocrinol (Lausanne). 2021;12:746924. [CrossRef]
- Chen J; Zhang L; Zhang X. Overall, sex-and race/ethnicity-specific prevalence of thyroid dysfunction in US adolescents aged 12–18 years. Front Public Health 12:1366485. [CrossRef]
- Romeo RD. The metamorphosis of adolescent hormonal stress reactivity:A focus on animal models. Front Neuroendocrinol. 2018;49:43-51. [CrossRef]
- Drzewiecki,C.M.; Juraska,J.M. The structural reorganization of the prefrontal cortex during adolescence as a framework for vulnerability to the environment. Pharmacol Biochem Behav. 2020;199, 173044. [CrossRef]
- Schneider M. Adolescence as a vulnerable period to alter rodent behavior. Cell Tissue Res. 2013;354(1):99-106. [CrossRef]
- McCormick CM. Methods; Challenges in Investigating Sex-Specific Consequences of Social Stressors in Adolescence in Rats:Is It the Stress or the Social or the Stage of Development? Curr Top Behav Neurosci. 2022;54:23-58. [CrossRef]
- Holder MK; Blaustein JD. Puberty; adolescence as a time of vulnerability to stressors that alter neurobehavioral processes. Front Neuroendocrinol. 2014;35(1):89-110. [CrossRef]
- Cavalcanti-de-Albuquerque,J.P.; Donato,J.; Jr. Rolling out physical exercise and energy homeostasis:Focus on hypothalamic circuitries. Front Neuroendocrinol. 2021;63:100944. [CrossRef]
- nt. Neuroendocrinol. 2009;63, 100944. 2021. Lafontan M; Langin D. Lipolysis; lipid mobilization in human adipose tissue. Prog Lipid Res. 48(5):275-97. [CrossRef]
- Baumgartner,A.; Hiedra,L.; Pinna,G.; Eravci,M.; Prengel,H.; Meinhold,H. Rat brain type II 5’-iodothyronine deiodinase activity is extremely sensitive to stress. J Neurochem. 1998;71(2), 817–826. [CrossRef]
- Martinez-deMena,R.; Calvo,R.M.; Garcia,L.; Obregon,M.J. Effect of glucocorticoids on the activity, expression and proximal promoter of type II deiodinase in rat brown adipocytes. Mol Cell Endocrinol. 2016;428, 58–67. [CrossRef]
- Queathem ED; Welly RJ; Clart LM; Rowles CC; Timmons H; Fitzgerald M; Eichen PA; Lubahn DB; Vieira-Potter VJ. White Adipose Tissue Depots Respond to Chronic Beta-3 Adrenergic Receptor Activation in a Sexually Dimorphic; Depot Divergent Manner Cells. 2021;10(12):3453. [CrossRef]
- Pereira,V.H.; Marques,F.; Lages,V.; Pereira,F.G.; Patchev,A.; Almeida,O.F.; Almeida-Palha,J.; Sousa,N.; Cerqueira,J.J. Glucose intolerance after chronic stress is related with downregulated PPAR-γ in adipose tissue. Cardiovascular diabetology, 15(1), 114. 2016. [CrossRef]
- Holm C; Osterlund T; Laurell H; Contreras JA. Molecular mechanisms regulating hormone-sensitive lipase; lipolysis. Annu Rev Nutr. 2000;20:365-93. [CrossRef]
- Ikeda K; Yamada T. UCP1 Dependent; Independent Thermogenesis in Brown; Beige Adipocytes. Front Endocrinol (Lausanne). 2020;11:498. [CrossRef]
- National Research Council (US) Committee for the Update of the Guide for the Care; Use of Laboratory Animals. Guide for the Care; Use of Laboratory Animals. 2011;8th ed. Washington (DC):National Academies Press (US);. PMID:21595115.
- Toth E; Gersner R; Wilf-Yarkoni A; Raizel H; Dar DE; Richter-Levin G; Levit O; Zangen A. Age-dependent effects of chronic stress on brain plasticity; depressive behavior. J Neurochem. 2008;107(2):522-32. [CrossRef]
- Basso JC; Morrell JI. Using wheel availability to shape running behavior of the rat towards improved behavioral; neurobiological outcomes. J Neurosci Methods. 2017;290:13-23. 10.1016/j.jneumeth.2017.07.009.
- Perelló M; Chacon F; Cardinali DP; Esquifino AI; Spinedi E. Effect of social isolation on 24-h pattern of stress hormones; leptin in rats. Life Sci. 2006;78(16):1857-62. [CrossRef]
- Paxinos; George; Watson; Charles. The Rat Brain in Stereotaxic Coordinates–The New Coronal Set, 5th Edn.
- Goodman T.; Hajihosseini M.K. Hypothalamic tanycytes-masters and servants of metabolic, neuroendocrine, and neurogenic functions. Front Neurosci. 2015;9:387. [CrossRef]
- Chomczynski P; Sacchi N. The single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction:twenty-something years on. Nat Protoc. 2006;1(2):581-5. [CrossRef]
- Pfaffl MW. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001;29(9):e45. [CrossRef]
- Krzywinski M; Birol I; Jones S; Marra M. Hive Plots — Rational Approach to Visualizing Networks. Brief Bioinform. 2012;13(5):627-44. [CrossRef]
- Jankord,R.; Solomon,M.B.; Albertz,J.; Flak,J.N.; Zhang,R.; Herman,J.P. Stress vulnerability during adolescent development in rats. Endocrinology, 152, 629–638. [CrossRef]
- Cotella EM; Gómez AS; Lemen P; Chen C; Fernández G; Hansen C; Herman JP; Paglini MG. Long-term impact of chronic variable stress in adolescence versus adulthood. Prog Neuropsychopharmacol Biol Psychiatry. 2019;10; 88:303-310. [CrossRef]
- Wulsin,A.C.; Wick-Carlson,D.; Packard,B.A.; Morano,R.; Herman,J.P. Adolescent chronic stress causes hypothalamo-pituitary-adrenocortical hypo-responsiveness and depression-like behavior in adult female rats. Psychoneuroendocrinol. 65, 109–117. 2016. [CrossRef]
- Smith,B.L.; Morano,R.L.; Ulrich-Lai,Y.M.; Myers,B.; Solomon,M.B.; Herman,J.P. Adolescent environmental enrichment prevents behavioral and physiological sequelae of adolescent chronic stress in female (but not male) rats. Stress (Amsterdam, Netherlands), 21(5), 464–473. 2018. [CrossRef]
- Greenwood BN; Fleshner M. Voluntary Wheel Running:A Useful Rodent Model for Investigating the Mechanisms of Stress Robustness; Neural Circuits of Exercise Motivation. Curr Opin Behav Sci. 2019;28:78-84. [CrossRef]
- Mathis V; Wegman-Points L; Pope B; Lee CJ; Mohamed M; Rhodes JS; Clark PJ; Clayton S; Yuan LL. Estrogen-mediated individual differences in female rat voluntary running behavior. J Appl Physiol. 1985;2024 136(3):592-605. [CrossRef]
- Casillas F; Flores-González A; Juárez-Rojas L; López A; Betancourt M; Casas E; Bahena I; Bonilla E; Retana-Márquez S. Chronic stress decreases fertility parameters in female rats. Syst Biol Reprod Med. 2023;69(3):234-244. [CrossRef]
- Aguiar AS Jr; Speck AE; Amaral IM; Canas PM; Cunha RA. The exercise sex gap; the impact of the estrous cycle on exercise performance in mice. Sci Rep. 2018;8(1):10742. [CrossRef]
- Suchacki KJ; Thomas BJ; Ikushima YM; Chen KC; Fyfe C; Tavares AAS; Sulston RJ; Lovdel A; Woodward HJ; Han X; Mattiucci D; Brain EJ; Alcaide-Corral CJ; Kobayashi H; Gray GA; Whitfield PD; Stimson RH; Morton NM; Johnstone AM; Cawthorn WP. The effects of caloric restriction on adipose tissue; metabolic health are sex-; age-dependent. Elife. 2023;12:e88080. [CrossRef]
- Mauvais-Jarvis F. Sex differences in energy metabolism:natural selection; mechanisms; consequences. Nat Rev Nephrol. 2024;20(1):56-69. [CrossRef]
- Harris RB. Chronic; acute effects of stress on energy balance:are there appropriate animal models? Am J Physiol Regul Integr Comp Physiol. 2015;308(4):R250-65. [CrossRef]
- Münzberg,H.; Singh,P.; Heymsfield,S.B.; Yu,S.; Morrison,C.D. Recent advances in understanding the role of leptin in energy homeostasis. F1000Res. 2020 ; 9:F1000 Faculty Rev-451. [CrossRef]
- López López AL; Escobar Villanueva MC; Brianza Padilla M; Bonilla Jaime H; Alarcón Aguilar FJ. Chronic unpredictable mild stress progressively disturbs glucose metabolism; appetite hormones in rats. Acta Endocrinol (Buchar). 2018;14(1):16-23. [CrossRef]
- Sasse SK; Nyhuis TJ; Masini CV; Day HE; Campeau S. Central gene expression changes associated with enhanced neuroendocrine; autonomic response habituation to repeated noise stress after voluntary wheel running in rats. Front Physiol. 2013;4:341. [CrossRef]
- Mikkelsen M; Wilsgaard T; Grimsgaard S; Hopstock LA; Hansson P. Associations between postprandial triglyceride concentrations; sex; age; body mass index:cross-sectional analyses from the Tromsø study 2015-2016. Front Nutr. 2023. 10:1158383. [CrossRef]
- Holcomb LE; Rowe P; O’Neill CC; DeWitt EA; Kolwicz SC Jr. Sex differences in endurance exercise capacity; skeletal muscle lipid metabolism in mice. Physiol Rep. 2022;10(3):e15174. [CrossRef]
- Brady LS; Smith MA; Gold PW; Herkenham M. Altered expression of hypothalamic neuropeptide mRNAs in food-restricted; food-deprived rats. Neuroendocrinology. 1990;52(5):441-7. [CrossRef]
- Loh K; Herzog H; Shi YC. Regulation of energy homeostasis by the NPY system. Trends Endocrinol Metab. 2015;26(3):125-35. [CrossRef]
- He,Z.; Gao,Y.; Alhadeff,A.L.; Castorena,C.M.; Huang,Y.; Lieu,L.; Afrin,S.; Sun,J.; Betley,J.N.; Guo,H.; Williams,K.W. Cellular and synaptic reorganization of arcuate NPY/AgRP and POMC neurons after exercise. Mol Metab. 18, 107–119. 2018. [CrossRef]
- Sergeyev V; Fetissov S; Mathé AA; Jimenez PA; Bartfai T; Mortas P; Gaudet L; Moreau JL; Hökfelt T. Neuropeptide expression in rats exposed to chronic mild stresses. Psychopharmacology (Berl). 2005;178(2-3):115-24. [CrossRef]
- J H Urban; A C Bauer-Dantoin; J E Levine; Neuropeptide Y gene expression in the arcuate nucleus:sexual dimorphism; modulation by testosterone; Endocrinology; 132; Issue 1:139–145. 1993. [CrossRef]
- Rebouças EC; Leal S; Sá SI. Regulation of NPY; α-MSH expression by estradiol in the arcuate nucleus of Wistar female rats:a stereological study. Neurol Res. 2016;38(8):740-7. [CrossRef]
- Lindblom J; Haitina T; Fredriksson R; Schiöth HB. Differential regulation of nuclear receptors; neuropeptides; peptide hormones in the hypothalamus; pituitary of food restricted rats. Brain Res Mol Brain Res. 2005;133(1):37-46. [CrossRef]
- Levay EA; Tammer AH; Penman J; Kent S; Paolini AG. Nutr Res. 2010;30(5):366-73. [CrossRef]
- Chen C; Nakagawa S; An Y; Ito K; Kitaichi Y; Kusumi I. The exercise-glucocorticoid paradox:How exercise is beneficial to cognition; mood; the brain while increasing glucocorticoid levels. Front Neuroendocrinol. 2017;44:83-102. [CrossRef]
- Joseph-Bravo P; Lazcano I; Jaimes-Hoy L; Gutierrez-Mariscal M; Sanchez-Jaramillo E; Uribe RM; Charli JL. Sexually dimorphic dynamics of thyroid axis activity during fasting in rats. Front Biosci (Landmark Ed). 2020;25(7):1305-1323. [CrossRef]
- van Haasteren GA; Linkels E; van Toor H; Klootwijk W; Kaptein E; de Jong FH; Reymond MJ; Visser TJ; de Greef WJ. Effects of long-term food reduction on the hypothalamus-pituitary-thyroid axis in male; female rats. J Endocrinol. 1996;150(2):169-78. [CrossRef]
- Zhang S; Su Z; Wen X. Associationof T3/T4 ratio with inflammatory indicatorsand all-cause mortality in stroke survivors. Front Endocrinol. 2025;15:1509501. [CrossRef]
- Araujo RL; Andrade BM; da Silva ML; Ferreira AC; Carvalho DP. Tissue-specific deiodinase regulation during food restriction; low replacement dose of leptin in rats. Am J Physiol Endocrinol Metab 296:E1157–E1163. [CrossRef]
- Weitzel JM; Iwen KA. Coordination of mitochondrial biogenesis by thyroid hormone. Mol Cell Endocrinol. 2011;342(1-2):1-7. [CrossRef]
- Nappi A; Moriello C; Morgante M; Fusco F; Crocetto F; Miro C. Effects of thyroid hormones in skeletal muscle protein turnover. J Basic Clin Physiol Pharmacol. 2024;35(4-5):253-264. [CrossRef]
- Toyoda N; Yasuzawa-Amano S; Nomura E; Yamauchi A; Nishimura K; Ukita C; Morimoto S; Kosaki A; Iwasaka T; Harney JW; Larsen PR; Nishikawa M. Thyroid hormone activation in vascular smooth muscle cells is negatively regulated by glucocorticoid. Thyroid. 2009;19(7):755-63. [CrossRef]
- Martinez-deMena R; Anedda A; Cadenas S; Obregon MJ. TSH effects on thermogenesis in rat brown adipocytes. Mol Cell Endocrinol. 2015;404:151-8. [CrossRef]
- Masami Murakami; Yuji Kamiya; Tadashi Morimura; Osamu Araki; Makoto Imamura; Takayuki Ogiwara; Haruo Mizuma; Masatomo Mori; Thyrotropin Receptors in Brown Adipose Tissue:Thyrotropin Stimulates Type II Iodothyronine Deiodinase; Uncoupling Protein-1 in Brown Adipocytes; Endocrinology; 142(3):1195–1201. 2001. [CrossRef]
- Sun,C.; Mao; S.; Chen,S.; Zhang; W.; Liu,C.PPARs-Orchestrated Metabolic Homeostasis in the Adipose Tissue. Int J Mol Sci. 8974;22 2021. [CrossRef]
- Tsiloulis T; Watt MJ. Exercise; the Regulation of Adipose Tissue Metabolism. Prog Mol Biol Transl Sci. 2015;135:175-201. [CrossRef]
- Granneman JG; Lahners KN. Regulation of mouse beta 3-adrenergic receptor gene expression; mRNA splice variants in adipocytes. Am J Physiol. 1995;268(4 Pt 1):C1040-4. [CrossRef]
- Fève B; Baude B; Krief S; Strosberg AD; Pairault J; Emorine LJ. dexamethasone down-regulates β3-adrenergic receptors in 3T3-F442A adipocytes. J Biol Chem 267(22):15909–15915. 1992. [CrossRef]
- Valle A; García-Palmer FJ; Oliver J; Roca P. Sex differences in brown adipose tissue thermogenic features during caloric restriction. Cell Physiol Biochem. 2007;19(1-4):195-204. [CrossRef]
- Jeremic N, Chaturvedi P, Tyagi SC. Browning of White Fat:Novel Insight Into Factors, Mechanisms, and Therapeutics. J Cell Physiol. 2017;232(1):61-8. [CrossRef]
- Ayala-Sumuano JT; Velez-delValle C; Beltrán-Langarica A; Marsch-Moreno M; Hernandez-Mosqueira C; Kuri-Harcuch W. Glucocorticoid paradoxically recruits adipose progenitors; impairs lipid homeostasis; glucose transport in mature adipocytes. Sci Rep. 2013;3:2573. [CrossRef]
- Laurens C; de Glisezinski I; Larrouy D; Harant I; Moro C. Influence of Acute; Chronic Exercise on Abdominal Fat Lipolysis:An Update. Front Physiol. 2020;11:575363. [CrossRef]
- Balagova,L.; Graban,J.; Puhova,A.; Jezova,D. Opposite Effects of Voluntary Physical Exercise on β3-Adrenergic Receptors in the White and Brown Adipose Tissue. Horm Metab Res. 2019;51(9):608-617. [CrossRef]
- Lehnig AC; Stanford KI. Exercise-induced adaptations to white; brown adipose tissue. J Exp Biol. 2018;221(Pt Suppl 1):jeb161570. [CrossRef]
- Slavin BG; Ong JM; Kern PA. Hormonal regulation of hormone-sensitive lipase activity; mRNA levels in isolated rat adipocytes. J Lipid Res. 1994;35(9):1535-41.
- Narita T; Kobayashi M; Itakura K; Itagawa R; Kabaya R; Sudo Y; Okita N; Higami Y. Differential response to caloric restriction of retroperitoneal; epididymal; subcutaneous adipose tissue depots in rats. Exp Gerontol. 2018;104:127-137. [CrossRef]
- Walczak,K.; Sieminska; LObesity; Thyroid Axis. Int J Environ Res Public Health. 2021;18(18):9434. [CrossRef]
- Dewal RS; Stanford KI. Effects of exercise on brown; beige adipocytes. Biochim Biophys Acta Mol Cell Biol Lipids. 2019;1864(1):71-78. [CrossRef]
- Bakopanos E; Silva JE. Opposing effects of glucocorticoids on beta(3)-adrenergic receptor expression in HIB-1B brown adipocytes. Mol Cell Endocrinol. 2002;190(1-2):29-37. [CrossRef]
- Vargas Y; Castro Tron AE; Rodríguez Rodríguez A; Uribe RM; Joseph-Bravo P; Charli JL. Thyrotropin-Releasing Hormone; Food Intake in Mammals:An Update. Metabolites. 2024;4(6):302. [CrossRef]
- Yan Su; Ewout Foppen; Eric Fliers; Andries Kalsbeek; Effects of Intracerebroventricular Administration of Neuropeptide Y on Metabolic Gene Expression; Energy Metabolism in Male Rats; Endocrinology; Volume 157(8):3070–3085. 2016. [CrossRef]
- Martelli,D.; Brooks; V.L.Leptin Increases:Physiological Roles in the Control of Sympathetic Nerve Activity; Energy Balance; the Hypothalamic–Pituitary–Thyroid Axis. Int J Mol Sci. 2023;24, 2684. [CrossRef]
- Dearing,C.; Handa,R.J.; Myers,B. Sex differences in autonomic responses to stress:implications for cardiometabolic physiology. Am J Physiol Endocrinol Metab. 2022;323(3):E281-E289. [CrossRef]
- Tanner MK; Mellert SM; Fallon IP; Baratta MV; Greenwood BN. Multiple Sex-; Circuit-Specific Mechanisms Underlie Exercise-Induced Stress Resistance. Curr Top Behav Neurosci. 2024;67:37-60. [CrossRef]
- Russell VA; Zigmond MJ; Dimatelis JJ; Daniels WM; Mabandla MV. The interaction between stress; exercise; its impact on brain function. Metab Brain Dis. 2014;29(2):255-60. [CrossRef]
- Bourke CH; Neigh GN. Behavioral effects of chronic adolescent stress are sustained; sexually dimorphic. Horm Behav. 2011;60(1):112-20. [CrossRef]
- Vieira IH; Rodrigues D; Paiva I. The Mysterious Universe of the TSH Receptor. Front. Endocrinol. 2022;13:944715. [CrossRef]
- Sequeira-Cordero A; Salas-Bastos A; Fornaguera J; Brenes JC. Behavioural characterisation of chronic unpredictable stress based on ethologically relevant paradigms in rats. Sci Rep. 2019;9(1):17403. [CrossRef]
- Nair BB; Khant Aung Z; Porteous R; Prescott M; Glendining KA; Jenkins DE; Augustine RA; Silva MSB; Yip SH; Bouwer GT; Brown CH; Jasoni CL; Campbell RE; Bunn SJ; Anderson GM; Grattan DR; Herbison AE; Iremonger KJ. Impact of chronic variable stress on neuroendocrine hypothalamus; pituitary in male; female C57BL/6J mice. J Neuroendocrinol. 2021;33(5):e12972. [CrossRef]
- Guo TY; Liu LJ; Xu LZ; Zhang JC; Li SX; Chen C; He LG; Chen YM; Yang HD; Lu L; Hashimoto K. Alterations of the daily rhythms of HPT axis induced by chronic unpredicted mild stress in rats. Endocrine. 2015;48(2):637-43. [CrossRef]
- Zhang J; Huang J; Aximujiang K; Xu C; Ahemaiti A; Wu G; Zhong L; Yunusi K. Thyroid Dysfunction; Neurological Disorder; Immunosuppression as the Consequences of Long-term Combined Stress. Sci Rep. 2018;8(1):4552. [CrossRef]
- Yesmin R; Watanabe M; Sinha AS; Ishibashi M; Wang T; Fukuda A. A subpopulation of agouti-related peptide neurons exciting corticotropin-releasing hormone axon terminals in median eminence led to hypothalamic-pituitary-adrenal axis activation in response to food restriction. Front Mol Neurosci. 2022;15:990803. [CrossRef]
- Joseph-Bravo,P.; Gutiérrez-Mariscal,M.; Jaimes-Hoy,L.; Charli; JL. Thyroid Axis and Energy Balance:Focus on Animals and Implications for Humankind. In:Preedy, V., Patel, V. 2017;(eds) Handbook of Famine, Starvation, and Nutrient Deprivation. Springer, Cham.
- Coppola A; Meli R; Diano S. Inverse shift in circulating corticosterone; leptin levels elevates hypothalamic deiodinase type 2 in fasted rats. Endocrinology. 146(6):2827-33. [CrossRef]
- Wray JR; Davies A; Sefton C; Allen TJ; Adamson A; Chapman P; Lam BYH; Yeo GSH; Coll AP; Harno E; White A. Global transcriptomic analysis of the arcuate nucleus following chronic glucocorticoid treatment. Mol Metab. 2019;26:5-17. [CrossRef]
- Benvenga S; Di Bari F; Granese R; Borrielli I; Giorgianni G; Grasso L; Le Donne M; Vita R; Antonelli A. Circulating thyrotropin is upregulated by estradiol. J Clin Transl Endocrinol. 2018;11:11-17. [CrossRef]
- Ha GE; Cheong E. Chronic Restraint Stress Decreases the Excitability of Hypothalamic POMC Neuron; Increases Food Intake. Exp Neurobiol. 30(6):375-386. [CrossRef]
- Stranahan,A.M.; Lee,K.; Mattson,M.P. Central mechanisms of HPA axis regulation by voluntary exercise. Neuromolecular Med. 2008;10, 118–227. [CrossRef]
- Kim, Y.J; Kim H.J.; Lee W.J.; Seong J.K. A comparison of the metabolic effects of treadmill and wheel running exercise in mouse model. Lab Anim Res. 2020;36:3. [CrossRef]
- Aguilar-Valles A; Sánchez E; de Gortari P; García-Vazquez AI; Ramírez-Amaya V; Bermúdez-Rattoni F; Joseph-Bravo P. The expression of TRH; its receptors; degrading enzyme is differentially modulated in the rat limbic system during training in the Morris water maze. Neurochem Int. 2007;50(2):404-17. [CrossRef]
- Bianco AC, Dumitrescu A, Gereben B, Ribeiro MO, Fonseca TL, Fernandes GW, Bocco BMLC. Paradigms of Dynamic Control of Thyroid Hormone Signaling. Endocr Rev. 2019;40(4):1000-1047. [CrossRef]
- Giacco A; Cioffi F; Cuomo A; Simiele R; Senese R; Silvestri E; Amoresano A; Fontanarosa C; Petito G; Moreno M; Lanni A; Lombardi A; de Lange P. Mild Endurance Exercise during Fasting Increases Gastrocnemius Muscle; Prefrontal Cortex Thyroid Hormone Levels through Differential BHB; BCAA-Mediated BDNF-mTOR Signaling in Rats. Nutrients. 2022;14(6):1166. [CrossRef]
- Shen,S.; Liao,Q.; Liu,J.; Pan,R.; Lee,S.M.; Lin,L. Myricanol rescues dexamethasone-induced muscle dysfunction via a sirtuin 1-dependent mechanism. J Cachexia Sarcopenia Muscle 10, 429–444. [CrossRef]
- Haque,N.; Tischkau; S.A.Sexual Dimorphism inAdipose-Hypothalamic Crosstalkand the Contribution of ArylHydrocarbon Receptor to RegulateEnergy Homeostasis. Int J Mol Sci. 2022;23, 7679. [CrossRef]
- Vidal P; Stanford KI. Exercise-Induced Adaptations to Adipose Tissue Thermogenesis. Front Endocrinol (Lausanne). 2020;11:270. [CrossRef]
- Münzberg H; Singh P; Heymsfield SB; Yu S; Morrison CD. Recent advances in understanding the role of leptin in energy homeostasis. F1000Res. 9:F1000 Faculty Rev-451. [CrossRef]
- Scheel AK, Espelage L, Chadt A. Many Ways to Rome:Exercise, Cold Exposure and Diet-Do They All Affect BAT Activation and WAT Browning in the Same Manner? Int J Mol Sci. 2022;23(9):4759. [CrossRef]
- Recazens E, Mouisel E, Langin D. Hormone-sensitive lipase:sixty years later. Prog Lipid Res. 2021;82:101084. [CrossRef]
- Wang J, Wang Y, Liu L, Lutfy K, Friedman TC, Liu Y, Jiang M, Liu Y. Lack of adipose-specific hexose-6-phosphate dehydrogenase causes inactivation of adipose glucocorticoids and improves metabolic phenotype in mice. Clin Sci (Lond). 2019;133(21):2189-2202. [CrossRef]
- Kim HJ, Kim YJ, Seong JK. AMP-activated protein kinase activation in skeletal muscle modulates exercise-induced uncoupled protein 1 expression in brown adipocyte in mouse model. J Physiol. 2022;600(10):2359-2376. [CrossRef]
- Hellstrom IC, Dhir SK, Diorio JC, Meaney MJ. Maternal licking regulates hippocampal glucocorticoid receptor transcription through a thyroid hormone-serotonin-NGFI-A signalling cascade. Philos Trans R Soc Lond B Biol Sci. 2012;367(1601):2495-510. [CrossRef]
- Louzada RA, Santos MC, Cavalcanti-de-Albuquerque JP, Rangel IF, Ferreira AC, Galina A, Werneck-de-Castro JP, Carvalho DP. Type 2 iodothyronine deiodinase is upregulated in rat slow- and fast-twitch skeletal muscle during cold exposure. Am J Physiol Endocrinol Metab. 2014;307(11):E1020-9. [CrossRef]
- Hernandez A, St Germain DL. Dexamethasone inhibits growth factor-induced type 3 deiodinase activity and mRNA expression in a cultured cell line derived from rat neonatal brown fat vascular-stromal cells. Endocrinology. 2002;143(7):2652-8. [CrossRef]
- de Vries EM, van Beeren HC, van Wijk ACWA, Kalsbeek A, Romijn JA, Fliers E, Boelen A. Regulation of type 3 deiodinase in rodent liver and adipose tissue during fasting. Endocr Connect. 2020;9(6):552-562. [CrossRef]
- Ayyar VS, Almon RR, DuBois DC, Sukumaran S, Qu J, Jusko WJ. Functional proteomic analysis of corticosteroid pharmacodynamics in rat liver:Relationship to hepatic stress, signaling, energy regulation, and drug metabolism. J Proteomics. 2017;160:84-105. [CrossRef]
- Mason J, Southwick S, Yehuda R, Wang S, Riney S, Bremner D, et al. Elevation of serum free triiodothyronine, total triiodothyronine, thyroxine-binding globulin, and total thyroxine levels in combat-related posttraumatic stress disorder. Arch Gen Psychiatry (1994) 51(8):629–41. [CrossRef]
- Raise-Abdullahi P, Meamar M, Vafaei AA, Alizadeh M, Dadkhah M, Shafia S, Ghalandari-Shamami M, Naderian R, Afshin Samaei S, Rashidy-Pour A. Hypothalamus and Post-Traumatic Stress Disorder:A Review. Brain Sci.; 13(7):1010. [CrossRef]
- Dietrich JW, Hoermann R, Midgley JEM, Bergen F, Müller P. The Two Faces of Janus:Why Thyrotropin as a Cardiovascular Risk Factor May Be an Ambiguous Target. Front Endocrinol (Lausanne). 2020;11:542710. [CrossRef]
- Sotelo-Rivera I, Cote-Vélez A, Uribe RM, Charli JL, Joseph-Bravo P. Glucocorticoids curtail stimuli-induced CREB phosphorylation in TRH neurons through interaction of the glucocorticoid receptor with the catalytic subunit of protein kinase A. Endocrine. 2017;55(3):861-871. [CrossRef]
- Osterlund C, Spencer RL. Corticosterone pretreatment suppresses stress-induced hypothalamic-pituitary-adrenal axis activity via multiple actions that vary with time, site of action, and de novo protein synthesis. J Endocrinol. 2011 ; 208(3):311-22. [CrossRef]
- Jaszczyk A, Juszczak GR. Glucocorticoids, metabolism and brain activity. Neurosci Biobehav Rev. 2021;126:113-145. [CrossRef]
- Fallon IP, Tanner MK, Greenwood BN, Baratta MV. Sex differences in resilience:Experiential factors and their mechanisms. Eur J Neurosci. 2020;52(1):2530-2547. [CrossRef]
Figure 1.
Chronic variable stress (CVS) during adolescence affects anxiety-like behavior, food intake, and body weight more in female rats than in males. (A) Male and female rats underwent the CVS paradigm during adolescence (PND30–70 for males and PND30–60 for females). Behavioral tests were conducted throughout the CVS procedure to assess changes during this period. Exercise was initiated 39 days after the onset of puberty (PND45 for males and PND35 for females). Figure created using BioRender. (B) Spontaneous locomotor activity was assessed using a photobeam activity system (PAS) on PND37 in female rats. The measurements included fine and ambulatory movements in the center of the cage (Cf and Ca) and ambulatory activity in the periphery (Pa). Time spent in the center and distance traveled in the open field test (OFT) were recorded at PND44 (OFT1; (C)) and PND51 (OFT2; (E)) for both sexes. Time and distance traveled in the open arms of the elevated plus maze test (EPM) were evaluated at PND58 in females (D) and PND65 in males (EPM2; (F)) Food intake (G) and body weight changes (H) were monitored throughout the CVS procedure during adolescence in male and female rats. Results are expressed as mean ± S.E.M. and analyzed by Two-way ANOVA for each PND, followed by Tukey’s post-hoc test. * P<0.05 vs. C group; σ P<0.05 vs. males of the same group. C: control group; CVS: chronic variable stress group.
Figure 1.
Chronic variable stress (CVS) during adolescence affects anxiety-like behavior, food intake, and body weight more in female rats than in males. (A) Male and female rats underwent the CVS paradigm during adolescence (PND30–70 for males and PND30–60 for females). Behavioral tests were conducted throughout the CVS procedure to assess changes during this period. Exercise was initiated 39 days after the onset of puberty (PND45 for males and PND35 for females). Figure created using BioRender. (B) Spontaneous locomotor activity was assessed using a photobeam activity system (PAS) on PND37 in female rats. The measurements included fine and ambulatory movements in the center of the cage (Cf and Ca) and ambulatory activity in the periphery (Pa). Time spent in the center and distance traveled in the open field test (OFT) were recorded at PND44 (OFT1; (C)) and PND51 (OFT2; (E)) for both sexes. Time and distance traveled in the open arms of the elevated plus maze test (EPM) were evaluated at PND58 in females (D) and PND65 in males (EPM2; (F)) Food intake (G) and body weight changes (H) were monitored throughout the CVS procedure during adolescence in male and female rats. Results are expressed as mean ± S.E.M. and analyzed by Two-way ANOVA for each PND, followed by Tukey’s post-hoc test. * P<0.05 vs. C group; σ P<0.05 vs. males of the same group. C: control group; CVS: chronic variable stress group.
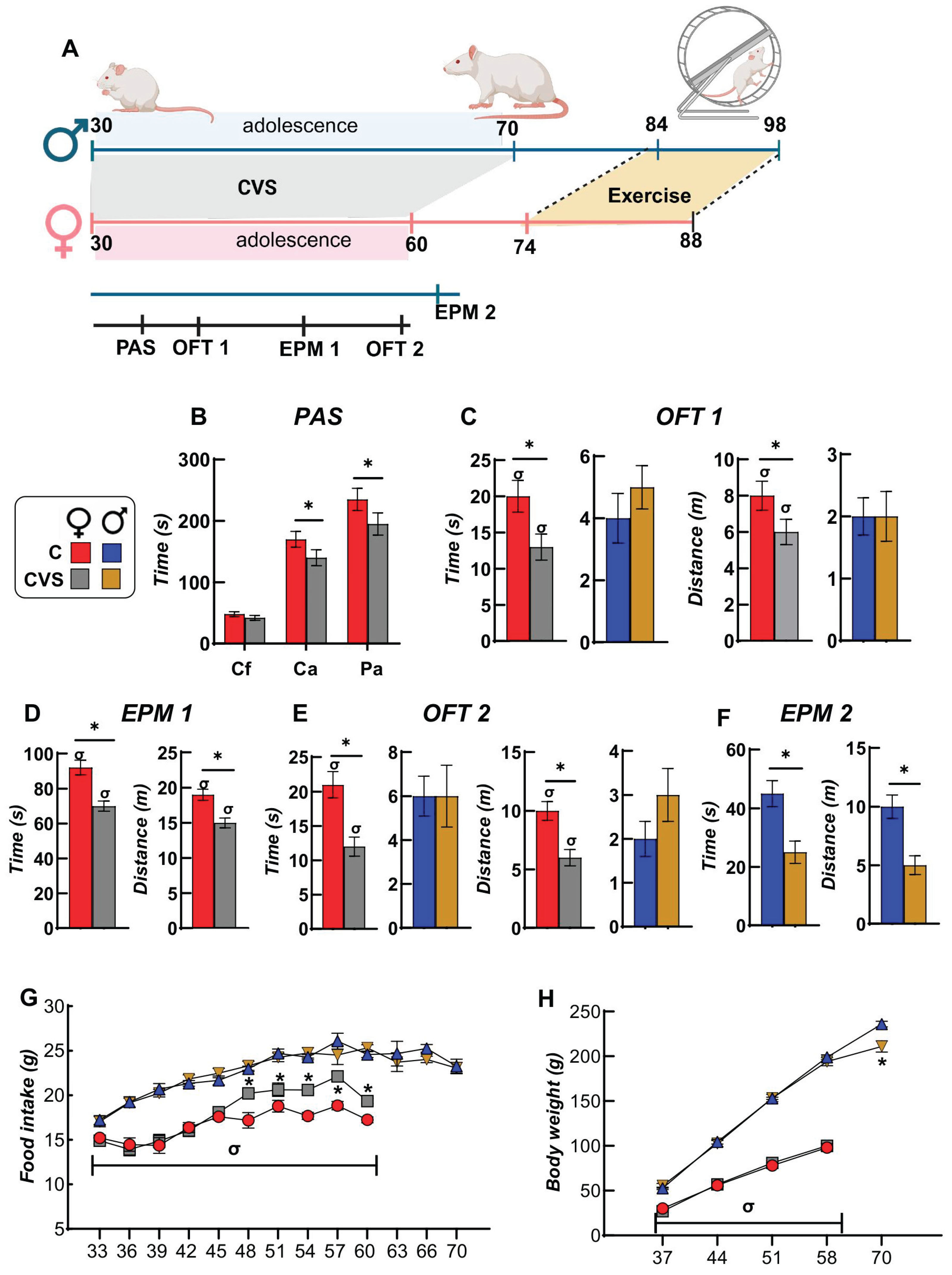
Figure 2.
CVS does not affect the activity patterns of cycling female rats, or their hormonal profiles. Daily exercise patterns of female control (A) and rats submitted to CVS (B) reveal periodic peaks aligned with the 4–5-day estrous cycle (C-Ex, indicated by arrows), contrasting with the flat activity trend observed in control (C) and CVS (D) male rats. (E) Distribution of estrous cycle phases determined by cytology according to (Cora et al. Toxicol Pathol. 2015 Aug;43(6):776-93), of control and stressed groups were similar (figure shows all female rats). (F) Serum concentrations of estradiol (E2, pg/mL), prolactin (PRL, ng/mL), and progesterone (P4, ng/mL) in sedentary (Sed), pair-fed (PF), or exercised (Ex) female rats under control or CVS conditions. Stress had a significant effect on serum progesterone levels (P=0.001). Results are expressed as mean ± S.E.M. and analyzed by Two-way ANOVA followed by Tukey’s post-hoc test. *: P<0.05 vs. C-SED; #: P < 0.05 vs. CVS-Ex. P, proestrus; E, estrus; M, metestrus; Di, diestrus.
Figure 2.
CVS does not affect the activity patterns of cycling female rats, or their hormonal profiles. Daily exercise patterns of female control (A) and rats submitted to CVS (B) reveal periodic peaks aligned with the 4–5-day estrous cycle (C-Ex, indicated by arrows), contrasting with the flat activity trend observed in control (C) and CVS (D) male rats. (E) Distribution of estrous cycle phases determined by cytology according to (Cora et al. Toxicol Pathol. 2015 Aug;43(6):776-93), of control and stressed groups were similar (figure shows all female rats). (F) Serum concentrations of estradiol (E2, pg/mL), prolactin (PRL, ng/mL), and progesterone (P4, ng/mL) in sedentary (Sed), pair-fed (PF), or exercised (Ex) female rats under control or CVS conditions. Stress had a significant effect on serum progesterone levels (P=0.001). Results are expressed as mean ± S.E.M. and analyzed by Two-way ANOVA followed by Tukey’s post-hoc test. *: P<0.05 vs. C-SED; #: P < 0.05 vs. CVS-Ex. P, proestrus; E, estrus; M, metestrus; Di, diestrus.
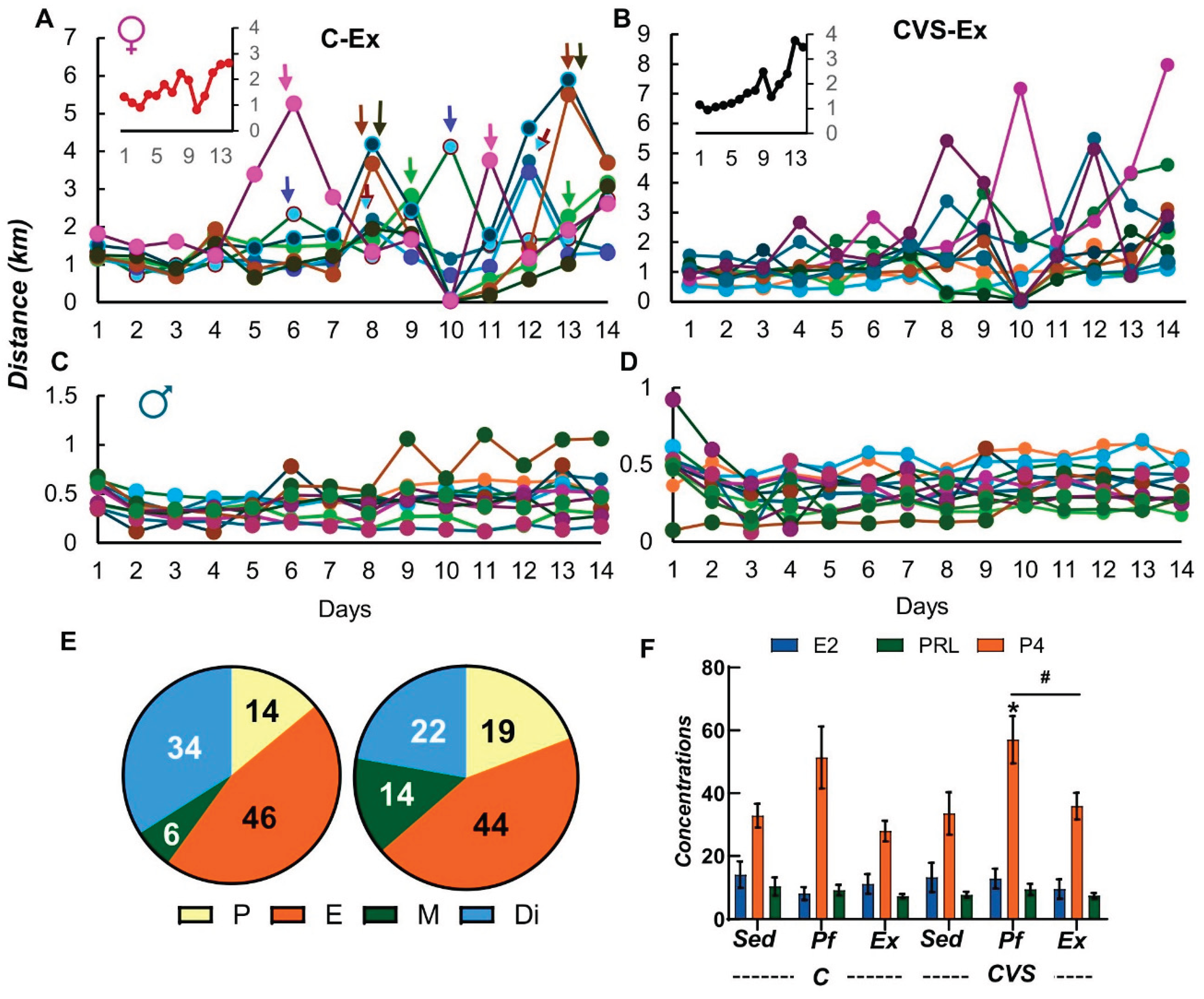
Figure 3.
CVS affects the voluntary exercise response of ponderal variables and metabolic hormones in a sex-dependent manner. Female rats began voluntary exercise at PND74, and males at PND84 (considering the adulthood of each sex). Average food consumption over the 14-day exercise protocol in females (A) and males (B). Body weight gain (BWg) and weights of perigonadal (pg), retroperitoneal (r), and interscapular (isc) white adipose tissue (WAT) are shown for females (C, D) and males (E, F). Correlation between iscWAT mass and serum leptin concentration in female control (G) and male PF groups (J). Serum leptin and.triglyceride (Tg) concentrations in female (H, I) and male (K, L) rats. Results are expressed as mean ± SEM and analyzed using three-way ANOVA, followed by Tukey’s post-hoc test. A main effect of exercise was found for adipose tissue weights (P < 0.0001) and for leptin and triglyceride concentrations (P < 0.0001). A significant interaction between exercise and sex was observed for triglyceride levels (P < 0.0001). $: P < 0.05 vs. different day periods within the same group; *: P < 0.05 vs. C-Sed; x: P < 0.05 vs. CVS-Sed; #: P < 0.05 vs. CVS-Ex; &: P < 0.05 vs. CVS-PF or CVS-Ex; σ: P < 0.05 vs. females of the same group; t: P < 0.05 by t-test. C-: control; CVS-: chronic variable stress; Sed: sedentary; PF: pair-fed; Ex: exercise.
Figure 3.
CVS affects the voluntary exercise response of ponderal variables and metabolic hormones in a sex-dependent manner. Female rats began voluntary exercise at PND74, and males at PND84 (considering the adulthood of each sex). Average food consumption over the 14-day exercise protocol in females (A) and males (B). Body weight gain (BWg) and weights of perigonadal (pg), retroperitoneal (r), and interscapular (isc) white adipose tissue (WAT) are shown for females (C, D) and males (E, F). Correlation between iscWAT mass and serum leptin concentration in female control (G) and male PF groups (J). Serum leptin and.triglyceride (Tg) concentrations in female (H, I) and male (K, L) rats. Results are expressed as mean ± SEM and analyzed using three-way ANOVA, followed by Tukey’s post-hoc test. A main effect of exercise was found for adipose tissue weights (P < 0.0001) and for leptin and triglyceride concentrations (P < 0.0001). A significant interaction between exercise and sex was observed for triglyceride levels (P < 0.0001). $: P < 0.05 vs. different day periods within the same group; *: P < 0.05 vs. C-Sed; x: P < 0.05 vs. CVS-Sed; #: P < 0.05 vs. CVS-Ex; &: P < 0.05 vs. CVS-PF or CVS-Ex; σ: P < 0.05 vs. females of the same group; t: P < 0.05 by t-test. C-: control; CVS-: chronic variable stress; Sed: sedentary; PF: pair-fed; Ex: exercise.
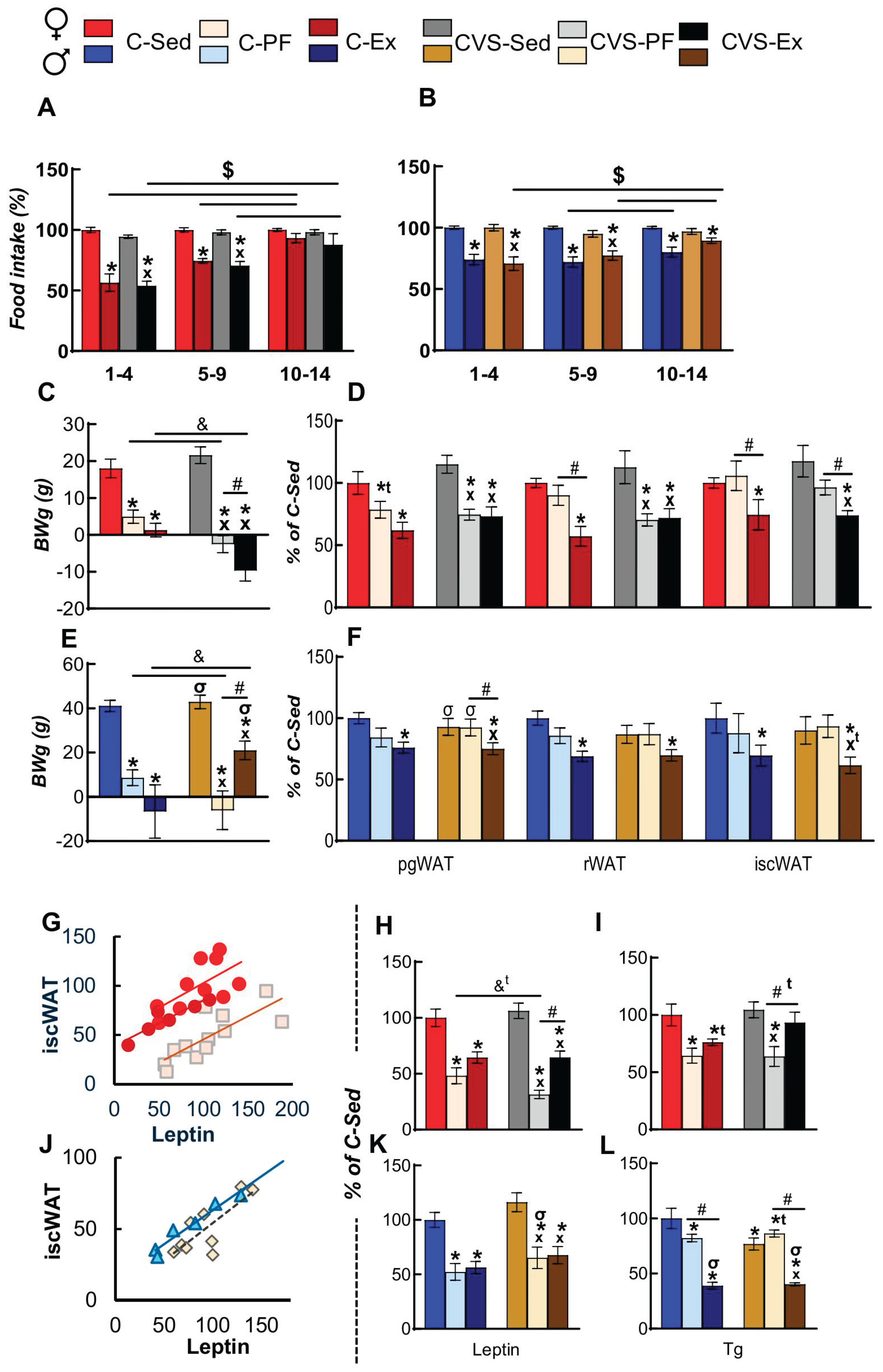
Figure 4.
Impact of CVS and voluntary exercise on mRNA levels of neuropeptides in the medial-basal hypothalamus (MBH) and hypothalamic-pituitary-adrenal (HPA) axis activity. mRNA levels of pro-opiomelanocortin (Pomc) (A) and neuropeptide Y (Npy) (B) in the medial-basal hypothalamus (MBH) of female rats. (C) MBH Npy expression was negatively correlated with Pomc expression in the CVS-PF and CVS-Ex female groups. Expression levels of Pomc (D) and Npy (E) in the MBH of male rats. (F) Pomc mRNA levels were negatively correlated with serum Tg concentrations in the C-Ex and CVS-Sed male groups. Expression of mRNA levels of Crh, Avp, and Gr in the medial caudal paraventricular nucleus (PVN) of female (G) and male (I) rats. Serum corticosterone (Cort) concentrations in females (H) and males (J). Results are expressed as mean ± SEM and analyzed using three-way ANOVA, followed by Tukey’s post-hoc test. Pomc and Npy were significantly affected by activity and stress (P < 0.000). Furthermore, Npy expression was influenced by the main effect of sex (P < 0.0001). Exercise was a significant factor for Crh (P = 0.004) and Avp (P < 0.0001) expression levels. Stress also affected Avp (P < 0.0001) and Gr expression (P = 0.0002), whereas sex had a significant effect on Gr alone (P < 0.0001). Analysis of corticosterone (Cort) levels revealed a significant interaction between activity, stress, and sex (P = 0.0001), and each factor independently showed a significant main effect (P < 0.0001). *: P < 0.05 vs. C-Sed; x: P < 0.05 vs. CVS-Sed; #: P < 0.05 vs. -PF; &: P < 0.05 vs. CVS-PF or CVS-Ex; σ: P < 0.05 vs. females of the same group; t: P < 0.05 by t-test. C-: control; CVS-: chronic variable stress; Sed: sedentary; PF: pair-fed; Ex: exercise.
Figure 4.
Impact of CVS and voluntary exercise on mRNA levels of neuropeptides in the medial-basal hypothalamus (MBH) and hypothalamic-pituitary-adrenal (HPA) axis activity. mRNA levels of pro-opiomelanocortin (Pomc) (A) and neuropeptide Y (Npy) (B) in the medial-basal hypothalamus (MBH) of female rats. (C) MBH Npy expression was negatively correlated with Pomc expression in the CVS-PF and CVS-Ex female groups. Expression levels of Pomc (D) and Npy (E) in the MBH of male rats. (F) Pomc mRNA levels were negatively correlated with serum Tg concentrations in the C-Ex and CVS-Sed male groups. Expression of mRNA levels of Crh, Avp, and Gr in the medial caudal paraventricular nucleus (PVN) of female (G) and male (I) rats. Serum corticosterone (Cort) concentrations in females (H) and males (J). Results are expressed as mean ± SEM and analyzed using three-way ANOVA, followed by Tukey’s post-hoc test. Pomc and Npy were significantly affected by activity and stress (P < 0.000). Furthermore, Npy expression was influenced by the main effect of sex (P < 0.0001). Exercise was a significant factor for Crh (P = 0.004) and Avp (P < 0.0001) expression levels. Stress also affected Avp (P < 0.0001) and Gr expression (P = 0.0002), whereas sex had a significant effect on Gr alone (P < 0.0001). Analysis of corticosterone (Cort) levels revealed a significant interaction between activity, stress, and sex (P = 0.0001), and each factor independently showed a significant main effect (P < 0.0001). *: P < 0.05 vs. C-Sed; x: P < 0.05 vs. CVS-Sed; #: P < 0.05 vs. -PF; &: P < 0.05 vs. CVS-PF or CVS-Ex; σ: P < 0.05 vs. females of the same group; t: P < 0.05 by t-test. C-: control; CVS-: chronic variable stress; Sed: sedentary; PF: pair-fed; Ex: exercise.
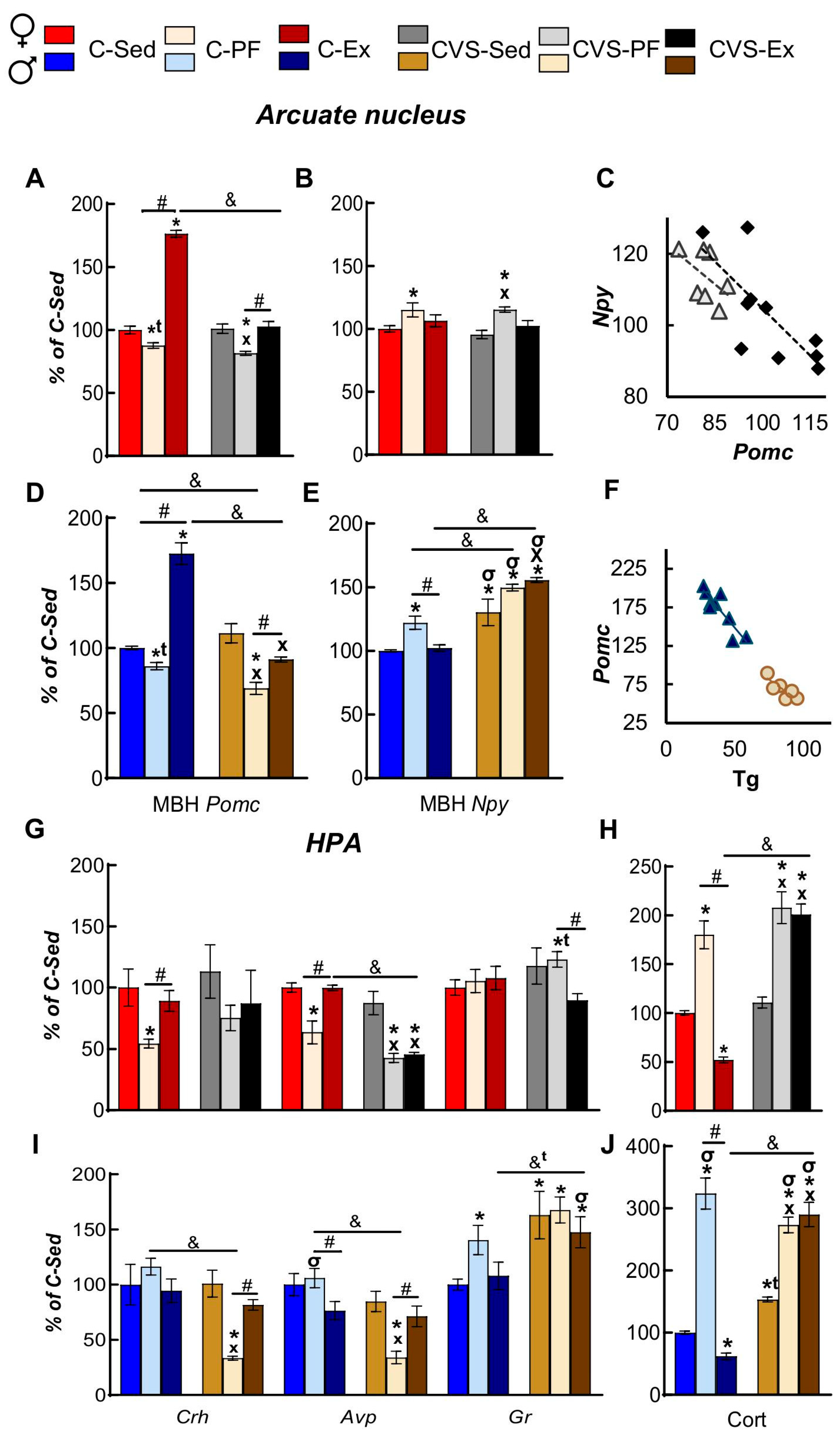
Figure 5.
CVS affects the basal activity of the hypothalamus-pituitary-thyroid (HPT) axis and its response to voluntary exercise in female rats. (A) mRNA expression levels of deiodinase type 2 (Dio2) and TRH-degrading enzyme (Trhde) in the medio-basal hypothalamus (MBH) and Trh in the medial-caudal paraventricular nucleus (PVN). (B) Correlation between MBH Npy expression levels and PVN Trh expression (C-PF), TSH (CVS-Ex), and T4 (CVS-Sed) serum concentrations. (C) Serum concentrations of TSH, total T4, and T3 and T3/T4 ratios. (D) Serum concentrations of free T4 and T3 (fT4 and fT3) and the fT3/fT4 ratio. (E) Correlation between serum progesterone and PVN Trh expression levels (CVS-Sed), serum fT4 concentration (C-Ex), and fT3/fT4 ratio (CVS-Ex). Results are expressed as mean ± SEM and analyzed using three-way ANOVA, followed by Tukey’s post-hoc test. Exercise and stress impacted MBH Dio2 and Trh in the MBH and PVN, respectively (P < 0.0001). Sex was also a significant factor in MBH Trhde and PVN Trh (P= 0.001). This pattern extended to circulating hormones, where activity was the main effect for TSH (P < 0.0001) and T4 (P = 0.03) levels. The effect of stress was significant for T4 (P= 0.03), T3 (P= 0.0006), and fT3 (P < 0.0001), whereas sex influenced the concentrations of T3, fT4, and fT3 (P < 0.001).*: P < 0.05 vs. C-Sed; x: P < 0.05 vs. CVS-Sed; #: P < 0.05 vs. -PF; &: P < 0.05 vs. CVS-PF or CVS-Ex; t: P < 0.05 by t-test. C-: control; CVS-: chronic variable stress; Sed: sedentary; PF: pair-fed; Ex: exercise.
Figure 5.
CVS affects the basal activity of the hypothalamus-pituitary-thyroid (HPT) axis and its response to voluntary exercise in female rats. (A) mRNA expression levels of deiodinase type 2 (Dio2) and TRH-degrading enzyme (Trhde) in the medio-basal hypothalamus (MBH) and Trh in the medial-caudal paraventricular nucleus (PVN). (B) Correlation between MBH Npy expression levels and PVN Trh expression (C-PF), TSH (CVS-Ex), and T4 (CVS-Sed) serum concentrations. (C) Serum concentrations of TSH, total T4, and T3 and T3/T4 ratios. (D) Serum concentrations of free T4 and T3 (fT4 and fT3) and the fT3/fT4 ratio. (E) Correlation between serum progesterone and PVN Trh expression levels (CVS-Sed), serum fT4 concentration (C-Ex), and fT3/fT4 ratio (CVS-Ex). Results are expressed as mean ± SEM and analyzed using three-way ANOVA, followed by Tukey’s post-hoc test. Exercise and stress impacted MBH Dio2 and Trh in the MBH and PVN, respectively (P < 0.0001). Sex was also a significant factor in MBH Trhde and PVN Trh (P= 0.001). This pattern extended to circulating hormones, where activity was the main effect for TSH (P < 0.0001) and T4 (P = 0.03) levels. The effect of stress was significant for T4 (P= 0.03), T3 (P= 0.0006), and fT3 (P < 0.0001), whereas sex influenced the concentrations of T3, fT4, and fT3 (P < 0.001).*: P < 0.05 vs. C-Sed; x: P < 0.05 vs. CVS-Sed; #: P < 0.05 vs. -PF; &: P < 0.05 vs. CVS-PF or CVS-Ex; t: P < 0.05 by t-test. C-: control; CVS-: chronic variable stress; Sed: sedentary; PF: pair-fed; Ex: exercise.
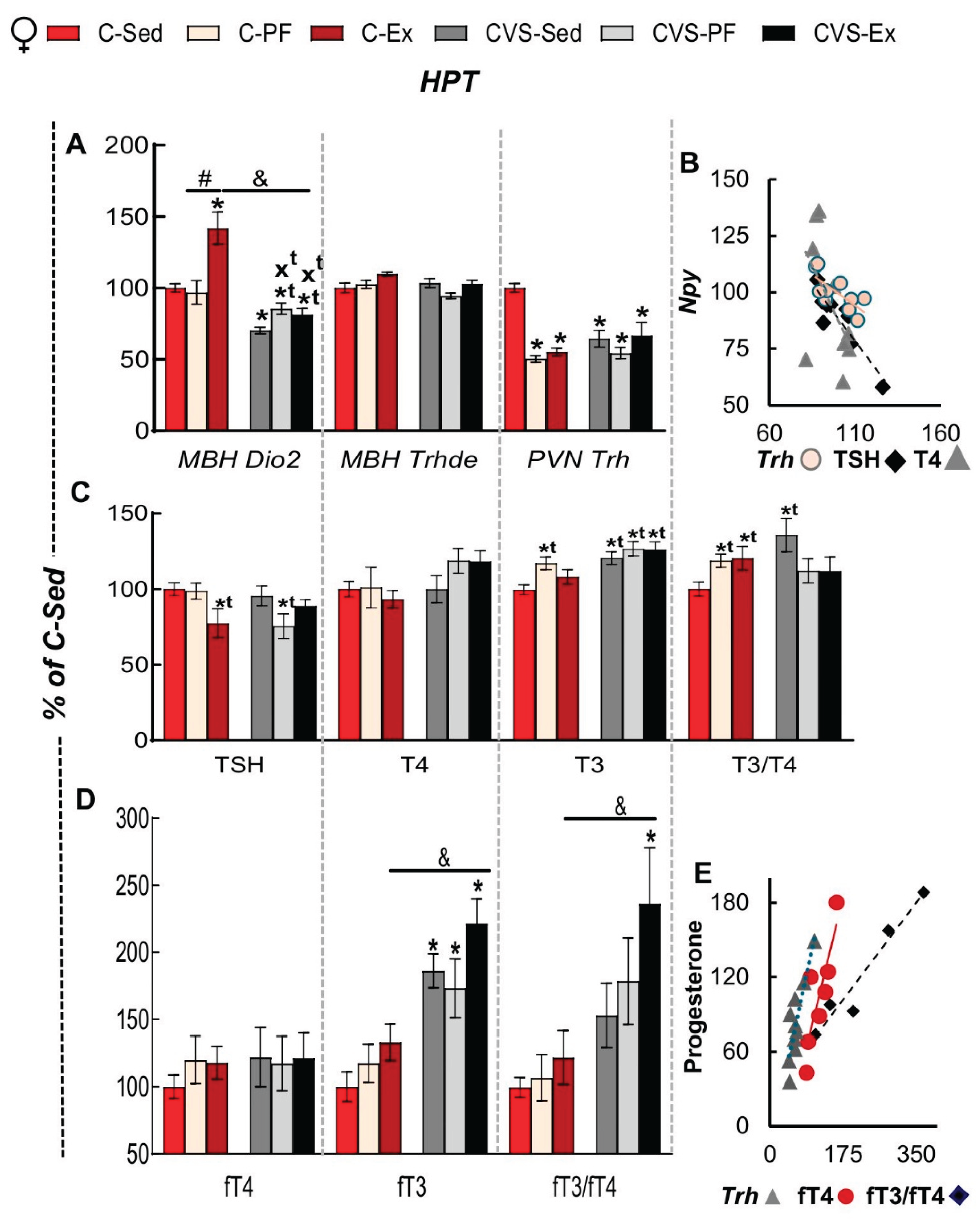
Figure 6.
Impact of CVS on the basal activity of the hypothalamus-pituitary-thyroid axis (HPT) and its response to voluntary exercise in male rats. (A) mRNA expression levels of deiodinase type 2 (Dio2) and TRH-degrading enzyme (Trhde) in the medio-basal hypothalamus (MBH) and Trh in the medial-caudal paraventricular nucleus (PVN). (B) Correlation between MBH Trhde and MBH Dio2 expression levels in the CVS groups. (C) Serum concentrations of TSH, total T4, and T3 and T3/T4 ratios. (D) Serum concentrations of free T4 and T3 (fT4 and fT3) and the fT3/fT4 ratio. (E) Correlation between distance ran and serum concentrations of T4 (C-Ex, P= 0.08), T3 (C-Ex), and fT3/fT4 ratio (CVS-Ex). Results are expressed as mean ± SEM and analyzed using three-way ANOVA, followed by Tukey’s post-hoc test. *: P < 0.05 vs. C-Sed; x: P < 0.05 vs. CVS-Sed; #: P < 0.05 vs. -PF; &: P < 0.05 vs. CVS-PF or CVS-Ex; σ: P < 0.05 vs. females of the same group; t: P < 0.05 by t-test. C-: control; CVS-: chronic variable stress; Sed: sedentary; PF: pair-fed; Ex: exercise.
Figure 6.
Impact of CVS on the basal activity of the hypothalamus-pituitary-thyroid axis (HPT) and its response to voluntary exercise in male rats. (A) mRNA expression levels of deiodinase type 2 (Dio2) and TRH-degrading enzyme (Trhde) in the medio-basal hypothalamus (MBH) and Trh in the medial-caudal paraventricular nucleus (PVN). (B) Correlation between MBH Trhde and MBH Dio2 expression levels in the CVS groups. (C) Serum concentrations of TSH, total T4, and T3 and T3/T4 ratios. (D) Serum concentrations of free T4 and T3 (fT4 and fT3) and the fT3/fT4 ratio. (E) Correlation between distance ran and serum concentrations of T4 (C-Ex, P= 0.08), T3 (C-Ex), and fT3/fT4 ratio (CVS-Ex). Results are expressed as mean ± SEM and analyzed using three-way ANOVA, followed by Tukey’s post-hoc test. *: P < 0.05 vs. C-Sed; x: P < 0.05 vs. CVS-Sed; #: P < 0.05 vs. -PF; &: P < 0.05 vs. CVS-PF or CVS-Ex; σ: P < 0.05 vs. females of the same group; t: P < 0.05 by t-test. C-: control; CVS-: chronic variable stress; Sed: sedentary; PF: pair-fed; Ex: exercise.
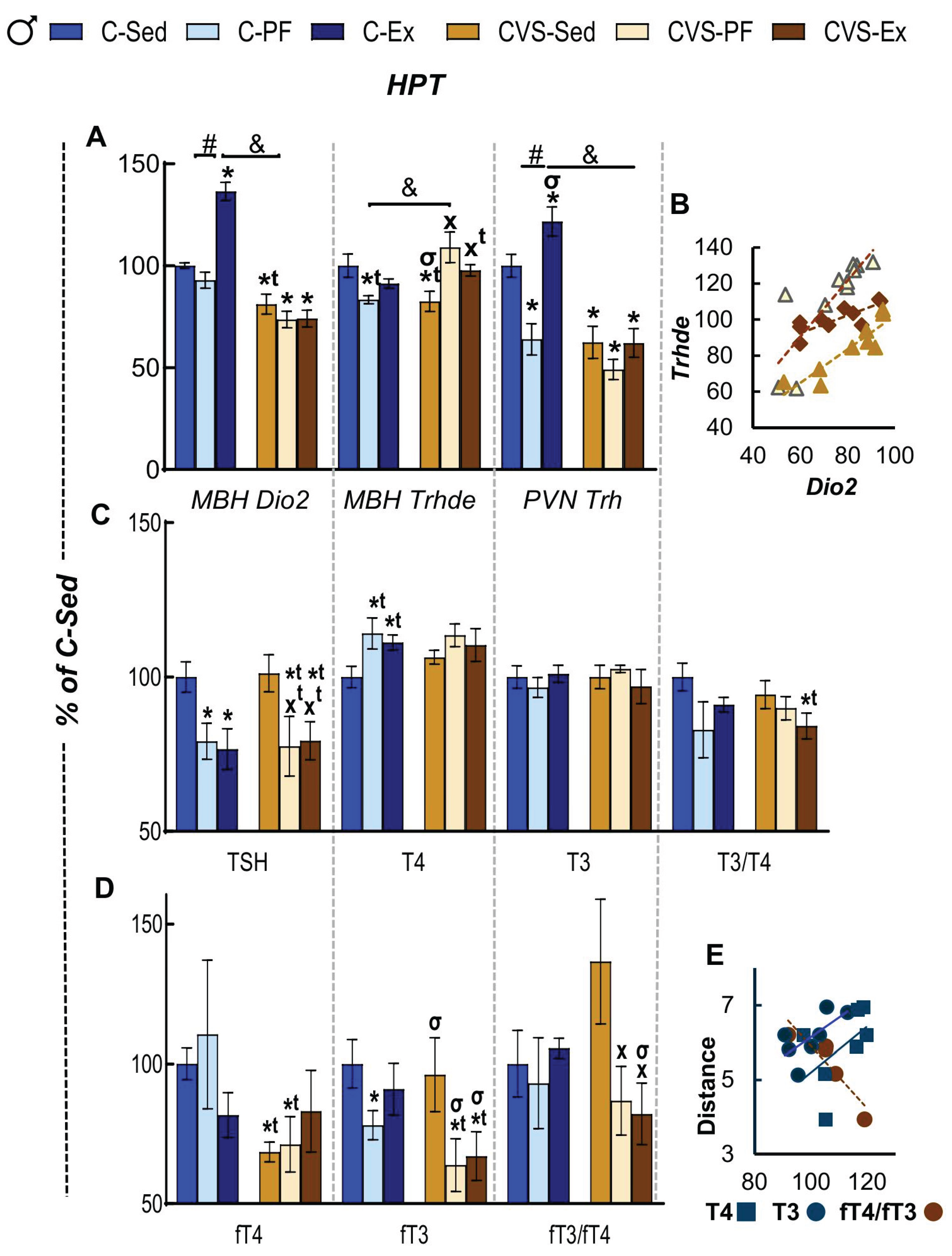
Figure 7.
CVS alters the response to voluntary exercise on mRNA expression of genes that regulate the response of thyroid hormones in skeletal muscles in a sex-dependent manner. mRNA levels of deiodinase type 2 (Dio2) and peroxisome proliferator-activated receptor γ coactivator 1α (Pgc1a) in the skeletal muscle of female (A) and male (B) rats. (C) Correlation between muscle Dio2 expression levels and serum concentrations of TSH and triglycerides (Tg) in C-Sed and CVS-PF female rats. (D) Correlation between serum corticosterone (Cort) concentration and expression levels of muscle Dio2 and Pgc1a in CVS male groups. (E) Correlation between muscle Dio2 expression and PVN Trh expression (C-Ex), serum TSH (C-Ex), and T3 (CVS-SED) concentrations. Results are expressed as mean ± SEM and analyzed using three-way ANOVA, followed by Tukey’s post-hoc test. The expression of Dio2 and Pgc1a was significantly affected by exercise (P < 0.0001) and stress (P < 0.0001). Furthermore, a main effect of sex (P = 0.0003) was.observed on Dio2 expression.*: P < 0.05 vs. C-Sed; x: P < 0.05 vs. CVS-Sed; #: P < 0.05 vs. -PF; &: P < 0.05 vs. CVS-PF or CVS-Ex; σ: P < 0.05 vs. females of the same group; t: P < 0.05 by t-test. C-: control; CVS-: chronic variable stress; Sed: sedentary; PF: pair-fed; Ex: exercise.
Figure 7.
CVS alters the response to voluntary exercise on mRNA expression of genes that regulate the response of thyroid hormones in skeletal muscles in a sex-dependent manner. mRNA levels of deiodinase type 2 (Dio2) and peroxisome proliferator-activated receptor γ coactivator 1α (Pgc1a) in the skeletal muscle of female (A) and male (B) rats. (C) Correlation between muscle Dio2 expression levels and serum concentrations of TSH and triglycerides (Tg) in C-Sed and CVS-PF female rats. (D) Correlation between serum corticosterone (Cort) concentration and expression levels of muscle Dio2 and Pgc1a in CVS male groups. (E) Correlation between muscle Dio2 expression and PVN Trh expression (C-Ex), serum TSH (C-Ex), and T3 (CVS-SED) concentrations. Results are expressed as mean ± SEM and analyzed using three-way ANOVA, followed by Tukey’s post-hoc test. The expression of Dio2 and Pgc1a was significantly affected by exercise (P < 0.0001) and stress (P < 0.0001). Furthermore, a main effect of sex (P = 0.0003) was.observed on Dio2 expression.*: P < 0.05 vs. C-Sed; x: P < 0.05 vs. CVS-Sed; #: P < 0.05 vs. -PF; &: P < 0.05 vs. CVS-PF or CVS-Ex; σ: P < 0.05 vs. females of the same group; t: P < 0.05 by t-test. C-: control; CVS-: chronic variable stress; Sed: sedentary; PF: pair-fed; Ex: exercise.
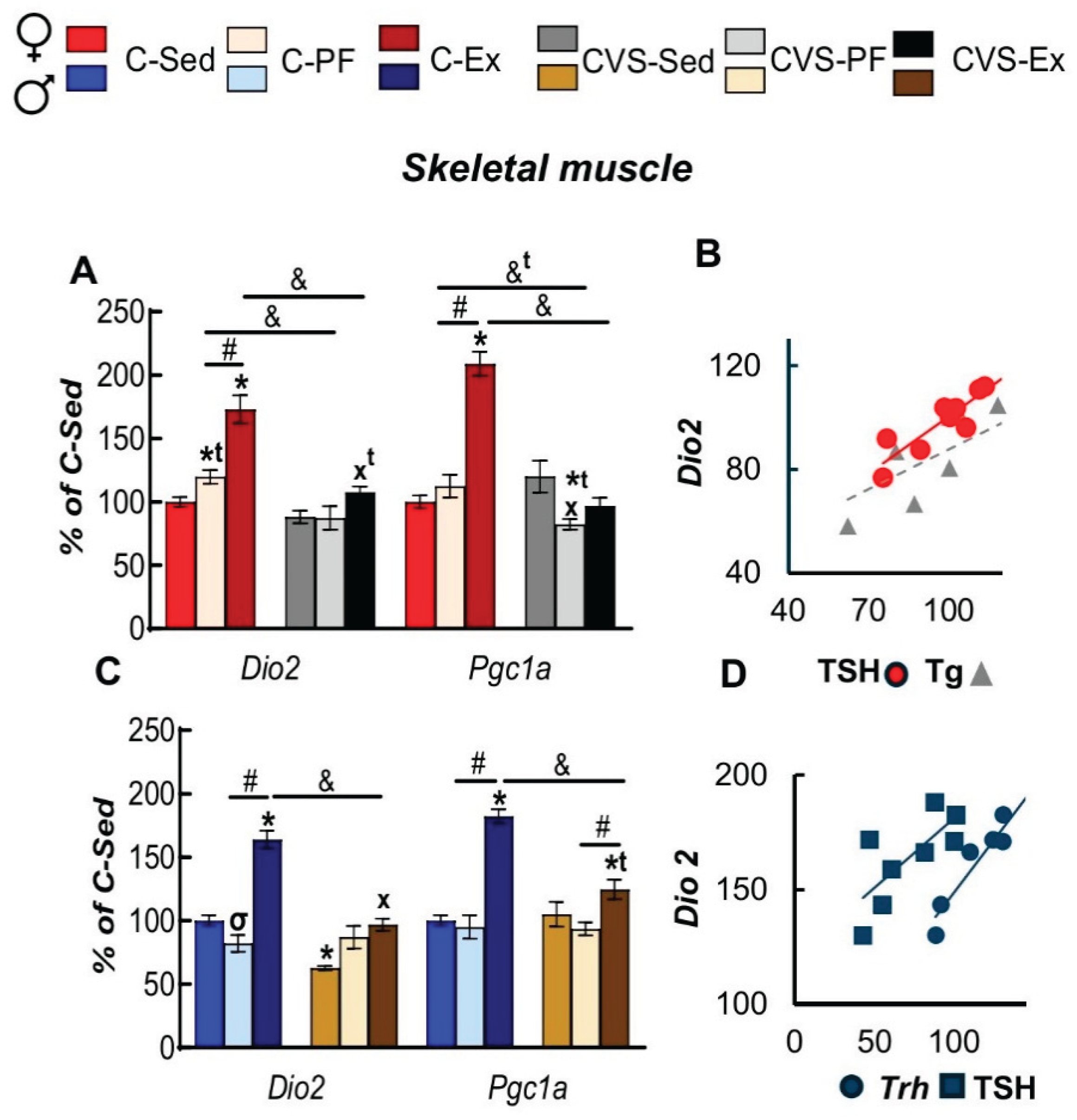
Figure 8.
Sex differences in mRNA expression of genes that regulate lipid metabolism in inguinal (iWAT) and perigonadal (pgWAT) white adipose tissues. mRNA levels of adrenergic beta 3 receptor (Adrb3), deiodinase type 2 (Dio2), peroxisome proliferator-activated receptor gamma (Pparg), and hormone-sensitive lipase (Hsl) in iWAT of female (A) and male (B) rats. mRNA levels of Adrb3, Dio2, Pparg, and Hsl in pgWAT of female (C) and male (D) rats. (E) Correlation between pgWAT Adrb3 expression levels and PVN-Trh expression (C- and CVS-Pf) and serum T4 (CVS-Sed) concentrations. (F) Correlation between iWAT Hsl expression and PVN Trh expression (CVS-Sed), serum TSH (C-Ex, CVS-Sed), and T4 (CVS-Ex) concentrations. (G) Correlation between pgWAT Hsl expression and serum corticosterone (Cort) levels in females (CVS-Ex) and males (CVS-Ex). (H) Correlation between pgWAT Adrb3 expression and PVN Trh expression (C-EX) and serum TSH (CVS-Sed) levels in male rats. (I) Correlation between iWAT Hsl expression and serum TSH levels in male CVS- groups. Results are expressed as mean ± SEM and analyzed using three-way ANOVA, followed by Tukey’s post-hoc test. Exercise (P < 0.0001) and stress (P < 0.0001) had significant effects on all genes measured (Adrb3, Dio2, Pparg, and Hsl) across both fat depots. The main effect of sex was also significant for nearly all genes, including Adrb3, Dio2, and Hsl, in both tissues (P < 0.001). The exception was Pparg, which was significantly affected by sex only in the iWAT (P = 0.001) but not in the pgWAT.*: P < 0.05 vs. C-Sed; x: P < 0.05 vs. CVS-Sed; #: P < 0.05 vs. -PF; &: P < 0.05 vs. CVS-PF or CVS-Ex; σ: P < 0.05 vs. females of the same group; t: P < 0.05 by t-test. C-: control; CVS-: chronic variable stress; Sed: sedentary; PF: pair-fed; Ex: exercise.
Figure 8.
Sex differences in mRNA expression of genes that regulate lipid metabolism in inguinal (iWAT) and perigonadal (pgWAT) white adipose tissues. mRNA levels of adrenergic beta 3 receptor (Adrb3), deiodinase type 2 (Dio2), peroxisome proliferator-activated receptor gamma (Pparg), and hormone-sensitive lipase (Hsl) in iWAT of female (A) and male (B) rats. mRNA levels of Adrb3, Dio2, Pparg, and Hsl in pgWAT of female (C) and male (D) rats. (E) Correlation between pgWAT Adrb3 expression levels and PVN-Trh expression (C- and CVS-Pf) and serum T4 (CVS-Sed) concentrations. (F) Correlation between iWAT Hsl expression and PVN Trh expression (CVS-Sed), serum TSH (C-Ex, CVS-Sed), and T4 (CVS-Ex) concentrations. (G) Correlation between pgWAT Hsl expression and serum corticosterone (Cort) levels in females (CVS-Ex) and males (CVS-Ex). (H) Correlation between pgWAT Adrb3 expression and PVN Trh expression (C-EX) and serum TSH (CVS-Sed) levels in male rats. (I) Correlation between iWAT Hsl expression and serum TSH levels in male CVS- groups. Results are expressed as mean ± SEM and analyzed using three-way ANOVA, followed by Tukey’s post-hoc test. Exercise (P < 0.0001) and stress (P < 0.0001) had significant effects on all genes measured (Adrb3, Dio2, Pparg, and Hsl) across both fat depots. The main effect of sex was also significant for nearly all genes, including Adrb3, Dio2, and Hsl, in both tissues (P < 0.001). The exception was Pparg, which was significantly affected by sex only in the iWAT (P = 0.001) but not in the pgWAT.*: P < 0.05 vs. C-Sed; x: P < 0.05 vs. CVS-Sed; #: P < 0.05 vs. -PF; &: P < 0.05 vs. CVS-PF or CVS-Ex; σ: P < 0.05 vs. females of the same group; t: P < 0.05 by t-test. C-: control; CVS-: chronic variable stress; Sed: sedentary; PF: pair-fed; Ex: exercise.
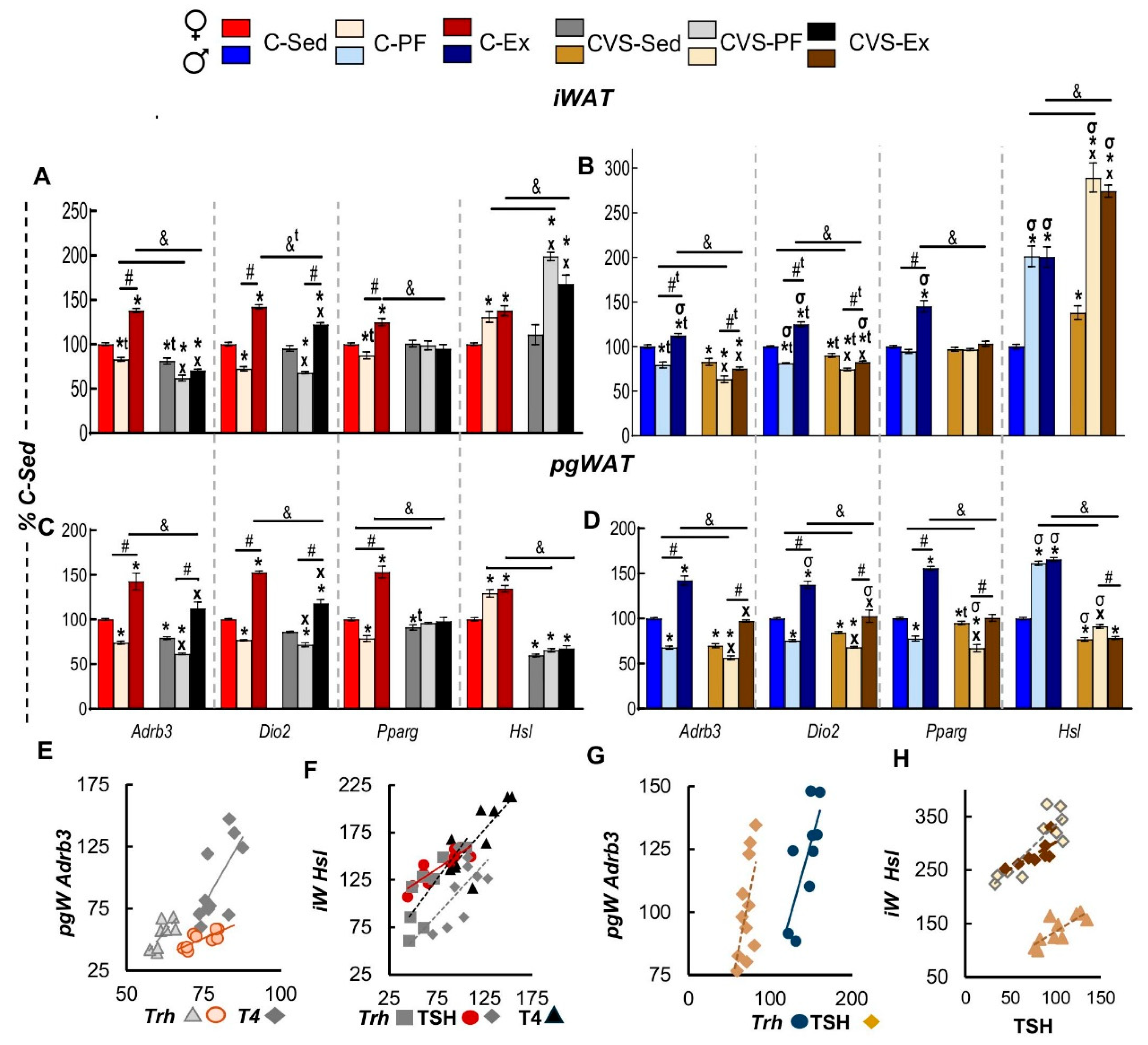
Figure 9.
CVS alters the effects of voluntary exercise on mRNA expression of thermogenic genes in brown adipose tissue (BAT) in a sex-dimorphic manner. mRNA levels of adrenergic beta 3 receptor (Adrb3), deiodinase type 2 (Dio2), and uncoupling protein 1 (Ucp1) in the brown adipose tissue of female (A) and male (C) rats. (B) Correlation between BAT-Adrb3 expression and serum corticosterone (Cort) levels in C- and CVS-PF and C-Ex female rats. (D) Correlation between BAT-Adrb3 expression and PVN Trh expression (C-Ex, CVS-PF) and serum TSH levels (CVS-Ex). Results are expressed as mean ± SEM and analyzed using three-way ANOVA, followed by Tukey’s post-hoc test. Exercise influenced the expression of all measured genes: Adrb3, Dio2, and Ucp1 (P < 0.0001 for all). The main effect of sex was also significant for Adrb3 and Ucp1 expression (P < 0.0001). Finally, a significant effect of stress was observed only for Ucp1 (P < 0.0001).*: P < 0.05 vs.C-Sed; x: P < 0.05 vs. CVS-Sed; #: P < 0.05 vs. -PF; &: P < 0.05 vs. CVS-PF or CVS-Ex; σ: P < 0.05 vs. females of the same group; t: P < 0.05 by t-test. C-: control; CVS-: chronic variable stress; Sed: sedentary; PF: pair-fed; Ex: exercise.
Figure 9.
CVS alters the effects of voluntary exercise on mRNA expression of thermogenic genes in brown adipose tissue (BAT) in a sex-dimorphic manner. mRNA levels of adrenergic beta 3 receptor (Adrb3), deiodinase type 2 (Dio2), and uncoupling protein 1 (Ucp1) in the brown adipose tissue of female (A) and male (C) rats. (B) Correlation between BAT-Adrb3 expression and serum corticosterone (Cort) levels in C- and CVS-PF and C-Ex female rats. (D) Correlation between BAT-Adrb3 expression and PVN Trh expression (C-Ex, CVS-PF) and serum TSH levels (CVS-Ex). Results are expressed as mean ± SEM and analyzed using three-way ANOVA, followed by Tukey’s post-hoc test. Exercise influenced the expression of all measured genes: Adrb3, Dio2, and Ucp1 (P < 0.0001 for all). The main effect of sex was also significant for Adrb3 and Ucp1 expression (P < 0.0001). Finally, a significant effect of stress was observed only for Ucp1 (P < 0.0001).*: P < 0.05 vs.C-Sed; x: P < 0.05 vs. CVS-Sed; #: P < 0.05 vs. -PF; &: P < 0.05 vs. CVS-PF or CVS-Ex; σ: P < 0.05 vs. females of the same group; t: P < 0.05 by t-test. C-: control; CVS-: chronic variable stress; Sed: sedentary; PF: pair-fed; Ex: exercise.
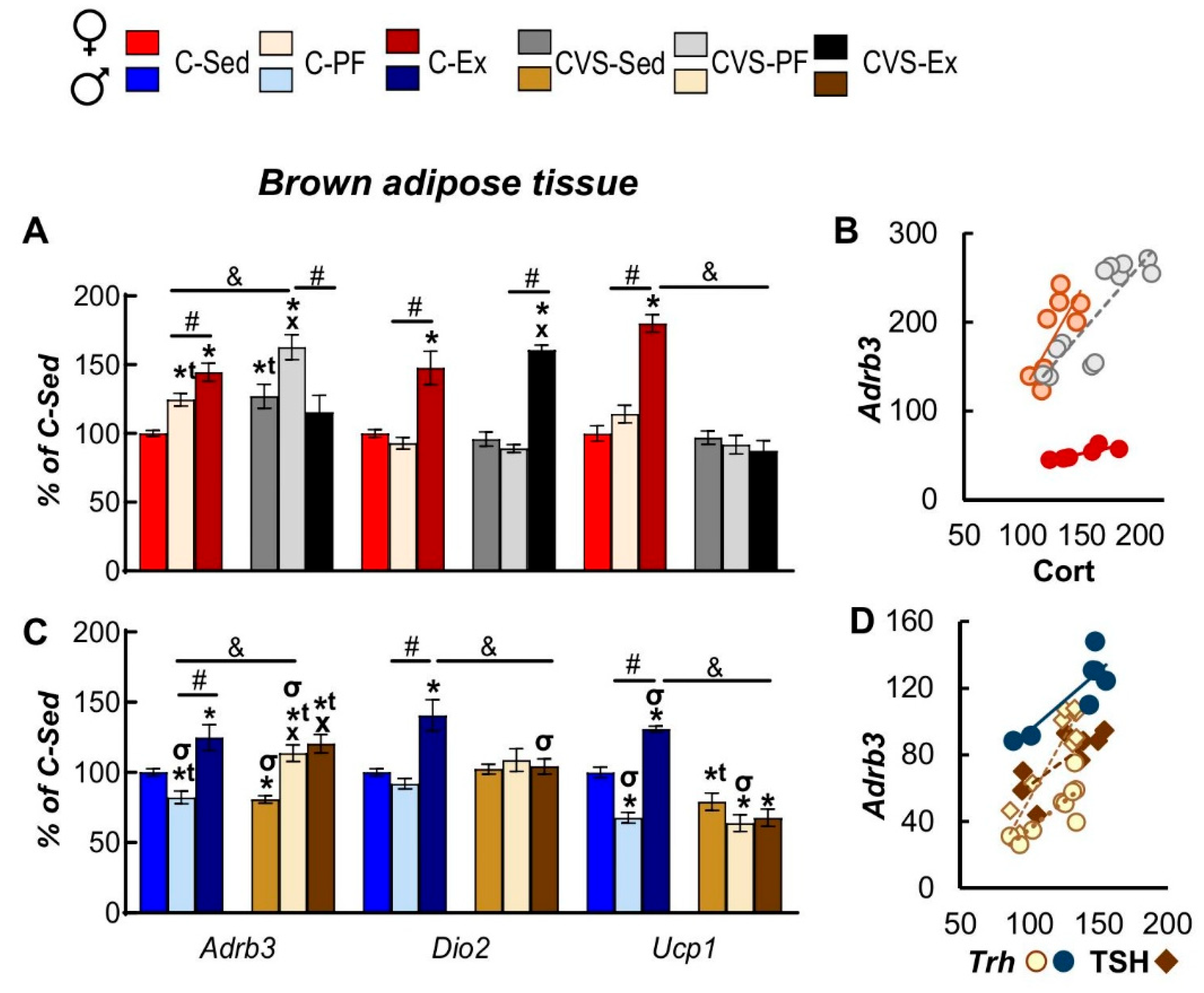
Figure 10.
Hiv eplot of correlations between gene expression in the. hypothalamus, peripheral tissues, and serum hormones of exercised rats. Hive plot diagrams illustrate significant (P < 0.05) Pearson correlations among gene expression levels in the hypothalamus and peripheral tissues and circulating hormone concentrations in exercised female and male rats. Blue edges denote positive correlations and red edges denote negative correlations. H, hypothalamus; gW, gonadal white adipose tissue (WAT); iW, inguinal WAT; M, skeletal muscle; scW, interscapular subcutaneous WAT; FI, food intake; BWc, body weight; Dist, total distance traveled (exercise performed). Hive plots created with software: https://codeberg.org/rgarcia-herrera/neuro_endocrine_correlations.
Figure 10.
Hiv eplot of correlations between gene expression in the. hypothalamus, peripheral tissues, and serum hormones of exercised rats. Hive plot diagrams illustrate significant (P < 0.05) Pearson correlations among gene expression levels in the hypothalamus and peripheral tissues and circulating hormone concentrations in exercised female and male rats. Blue edges denote positive correlations and red edges denote negative correlations. H, hypothalamus; gW, gonadal white adipose tissue (WAT); iW, inguinal WAT; M, skeletal muscle; scW, interscapular subcutaneous WAT; FI, food intake; BWc, body weight; Dist, total distance traveled (exercise performed). Hive plots created with software: https://codeberg.org/rgarcia-herrera/neuro_endocrine_correlations.
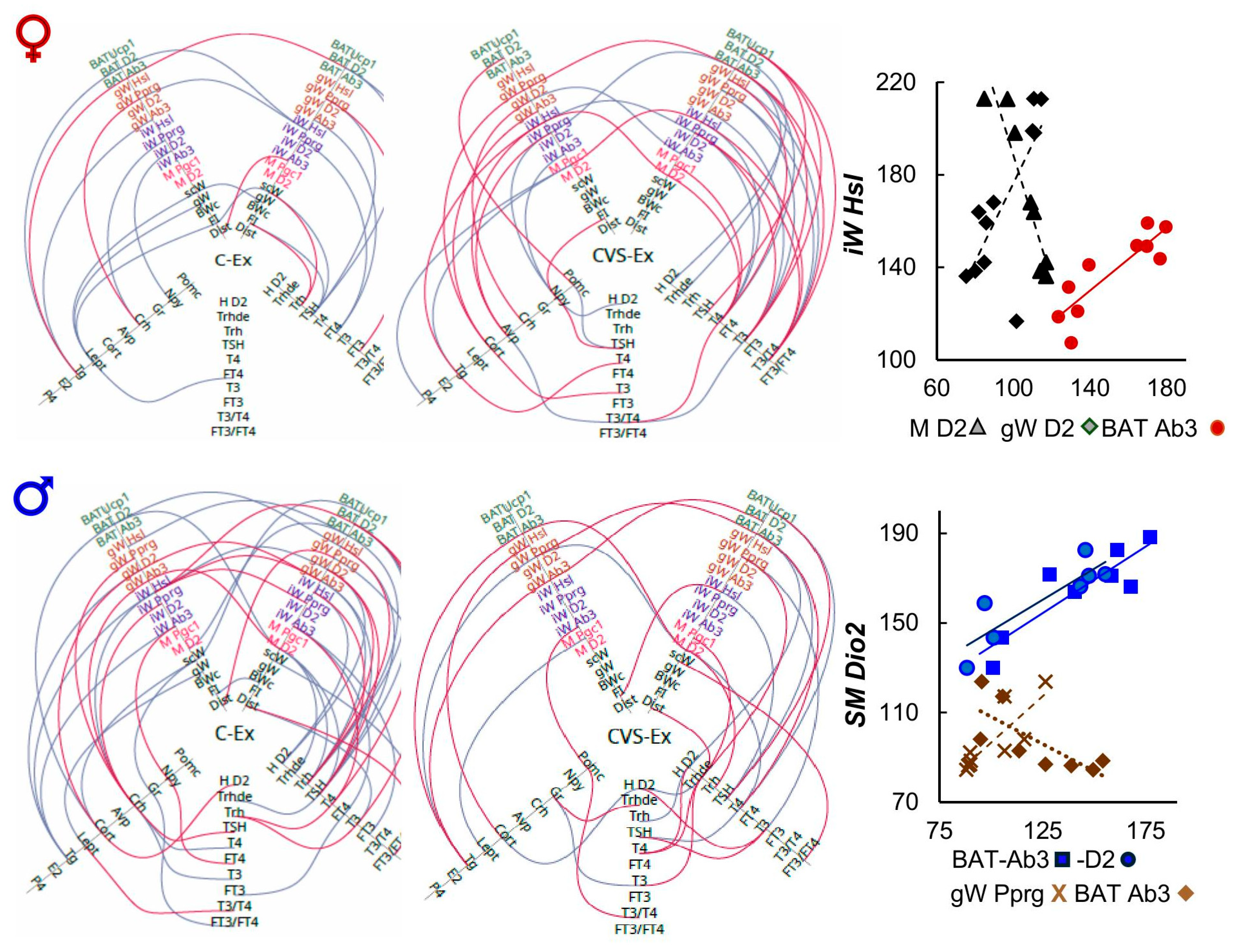
Figure 11.
Effects of exercise on the HPT, HPA axes and target tissues of. female and male controls and chronic variable stressed rats (CVS) compared to pair-fed groups. The diagram summarizes the sex-dependent long-term effects of adolescent chronic variable stress (CVS) and adult voluntary exercise (Ex) on key components of the hypothalamic-pituitary-thyroid (HPT) and adrenal (HPA) axes and thyroid hormone-responsive genes in peripheral tissues. Groups are represented by different colors for males and females; upward arrows (↑) indicate an increase, downward arrows (↓) a decrease, and dashed lines (-) indicate no significant change, relative to either the C-PF or CVS-PF group. C-: control; CVS-: chronic variable stress; PF: pair-fed; Ex: exercise. HPT axis image created using biorender.com.
Figure 11.
Effects of exercise on the HPT, HPA axes and target tissues of. female and male controls and chronic variable stressed rats (CVS) compared to pair-fed groups. The diagram summarizes the sex-dependent long-term effects of adolescent chronic variable stress (CVS) and adult voluntary exercise (Ex) on key components of the hypothalamic-pituitary-thyroid (HPT) and adrenal (HPA) axes and thyroid hormone-responsive genes in peripheral tissues. Groups are represented by different colors for males and females; upward arrows (↑) indicate an increase, downward arrows (↓) a decrease, and dashed lines (-) indicate no significant change, relative to either the C-PF or CVS-PF group. C-: control; CVS-: chronic variable stress; PF: pair-fed; Ex: exercise. HPT axis image created using biorender.com.
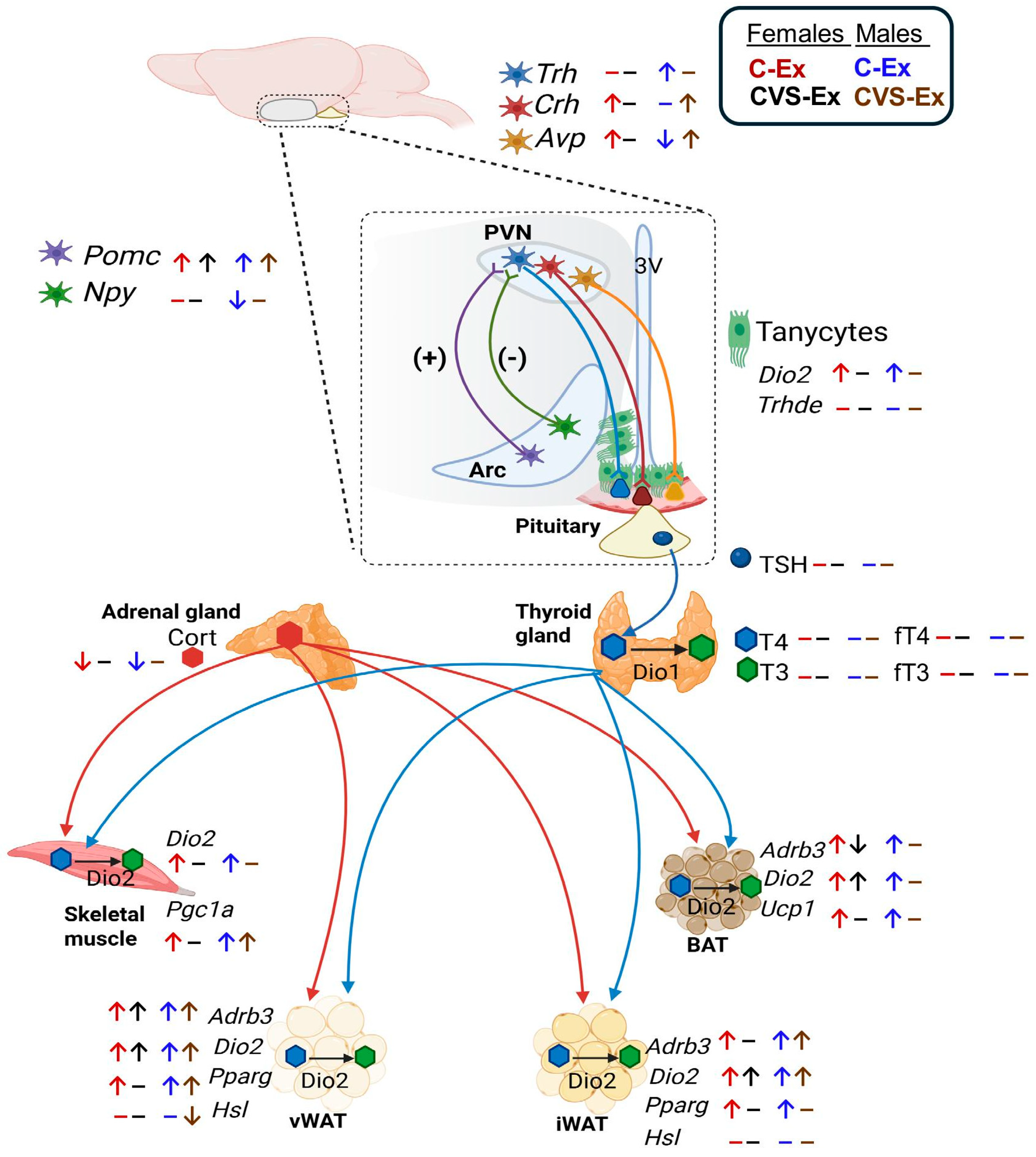

Table 1.
Summary of responses to 14 days of food restriction (pair-fed) or.exercise in female or male adult control or previously stressed rats duringadolescence (CVS). Arrows indicate significant changes compared to those in thecontrol sedentary group (C-Sed). The direction of the arrow denotes the nature ofthe change (↑ increase; ↓ decrease). The size and boldness of the arrows represent differences in the level of change. Filled cell colors highlight sex-specific effects, either a significant change observed in only one sex or opposite responses between sexes.
Table 1.
Summary of responses to 14 days of food restriction (pair-fed) or.exercise in female or male adult control or previously stressed rats duringadolescence (CVS). Arrows indicate significant changes compared to those in thecontrol sedentary group (C-Sed). The direction of the arrow denotes the nature ofthe change (↑ increase; ↓ decrease). The size and boldness of the arrows represent differences in the level of change. Filled cell colors highlight sex-specific effects, either a significant change observed in only one sex or opposite responses between sexes.
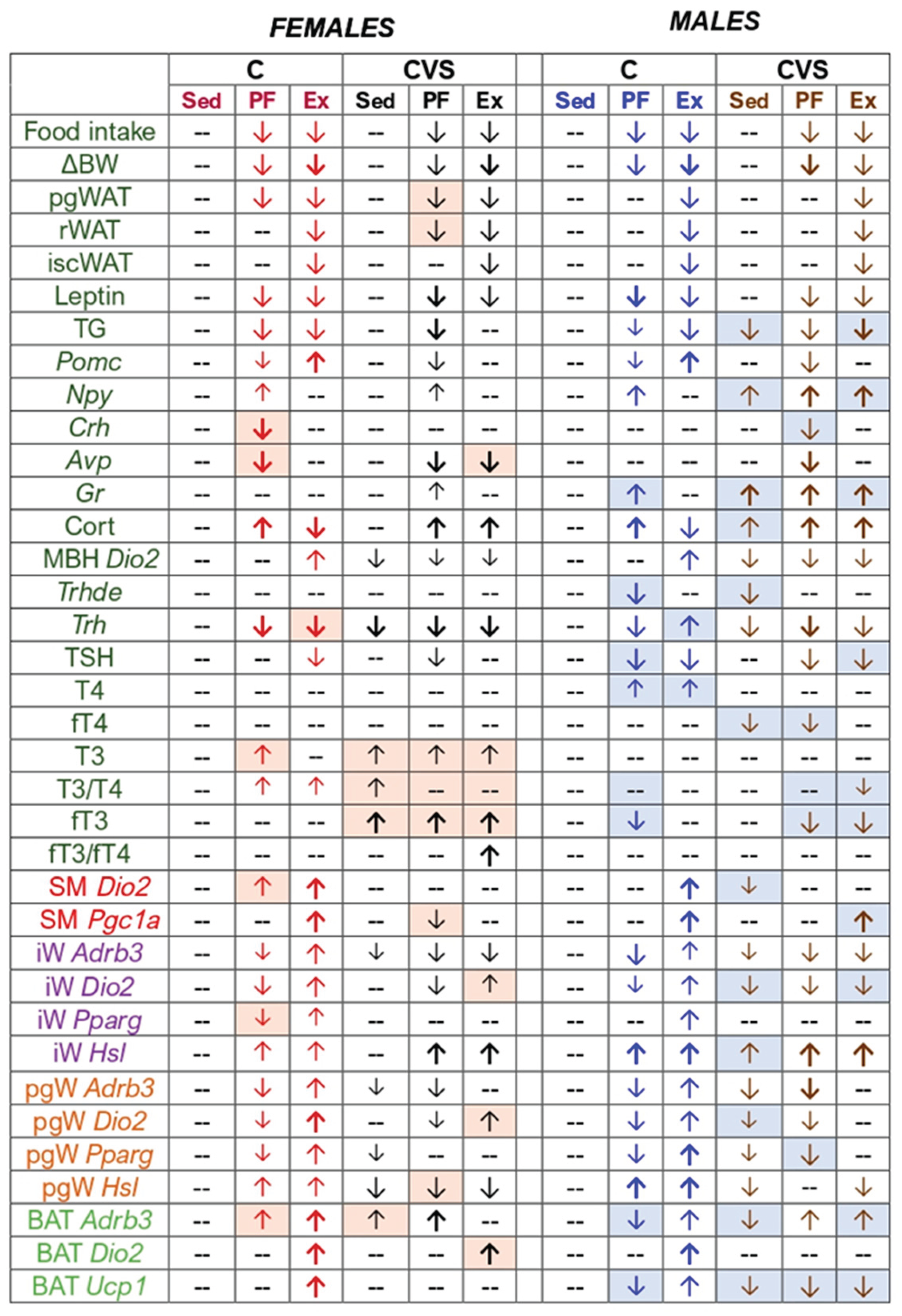 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



