
Submitted:
17 July 2025
Posted:
18 July 2025
You are already at the latest version
Abstract
Nuclear receptors (NRs) are ligand-activated transcription factors that mediate diverse cellular processes, including signalling, survival, proliferation, immune response and metabolism, through both genomic and non-genomic mechanisms in response to hor-mones and metabolic ligands. Given their central role in inter-organ, tissue, and cellular communication, NRs are critical for maintaining homeostasis and have become a major focus in biomedical research and drug discovery due to their association with numerous diseases.
Among NRs, the NR4A subfamily (NR4A1/Nur77, NR4A2/Nurr1, and NR4A3/Nor1) re-sponds to various stimuli—such as insulin, growth factors, inflammatory cytokines, and β-adrenergic signals—though their endogenous ligands remain unidentified. Their ex-pression is tissue-dependent, particularly in energy-demanding tissues, where they mod-ulate leukocyte function and promote an anti-inflammatory profile. Like other NRs, NR4As regulate acute and chronic inflammation by suppressing pro-inflammatory tran-scription factors (e.g., NF-κB) or enhancing their inhibitors, thereby polarising macro-phages toward an anti-inflammatory phenotype.
This review summarises current knowledge on the role of NR4A receptors in immune re-sponses. Given their well-documented involvement in autoimmune diseases, inflammato-ry conditions, and cancer, elucidating their contributions to neuro-immune-endocrine crosstalk may uncover their therapeutic potential for immunopathological disorders.
Keywords:
nuclear receptors
; NR4A subfamily
; NR4A1/Nur77/NBFIB
; NR4A2/Nurr1
; NR4A3/Nor1
; immune response
1. Introduction
Nuclear receptors (NRs) are ligand-activated transcription factors that mediate diverse cellular processes, including signalling, survival, proliferation, immune response and metabolism, through both genomic and non-genomic mechanisms in response to hormones and metabolic ligands [1]. Given their pivotal role in inter-organ, tissue, and cellular communication, for homeostasis, NRs are essential for maintaining homeostasis and have emerged as key targets in biomedical research and drug discovery due to their dysregulation in numerous diseases [2].
The NRs superfamily is classified into six subfamilies based on evolutionary sequence conservation. Structurally, NRs comprise four functional domains:
- The variable N-terminal A/B domain, which contains the activation function-1 (AF-1) region and mediates interactions with coactivators and corepressors to modulate transcriptional activity.
- The highly conserved central C domain, housing the DNA-binding domain (DBD) that enables NRs to regulate gene expression by binding promoter sequences as homo-/heterodimers or monomers.
- The flexible hinge D domain, which connects the DBD to the ligand-binding domain (LBD) and facilitates LBD rotation for transcriptional responses; it also contains a nuclear localisation signal (NLS) for nuclear import.
NRs orchestrate developmental and homeostatic processes by modulating gene networks across cell types, thereby maintaining organ-specific functions. Under pathological conditions, their transcriptional activity, is often amplified as part of compensatory responses to restore homeostasis, underscoring their dual roles in physiology and disease [1], Stromal and immune cells fine-tune immune function through surface receptors and NRs, with the latter primarily regulated by lipophilic ligands (e.g., steroids, retinoids, phospholipids, oxysterols, and vitamins). Ligand binding triggers NRs to bind genomic DNA [5], recruit co-regulators, and remodel chromatin to activate or repress target genes. Consequently, NR dysregulation is implicated in cancer, autoimmunity, and chronic inflammation [6].
While some NRs—such as the glucocorticoid receptor (GR), retinoic acid receptor (RAR), and vitamin D receptor (VDR)—are well-characterised and therapeutically exploited, others have only recently attracted attention. Among these are the NR4A subfamily receptors (NR4A1/Nur77, NR4A2/Nurr1, and NR4A3/Nor1), discovered in the late 1980s–early 1990s [7,8,9]. Initially thought to function predominantly in the nervous system, emerging evidence has revealed their roles in diverse tissues, particularly in immune cells, where they modulate homeostasis and inflammation in a context-dependent manner [10].
This review synthesises current knowledge on NR4A receptors, emphasising their regulation of immune responses and potential as therapeutic targets.
2. General Characteristics of NR4A Subfamily
As previously mentioned, the NR4A subfamily consists of three transcription factors - NR4A1 (Nur77), NR4A2 (Nurr1), and NR4A3 (Nor1) - which belong to the nuclear receptor superfamily of 48 members These receptors share the characteristic modular structure of nuclear receptors, featuring an N-terminal domain containing the ligand-independent activation function-1 (AF-1) region, a central DBD, and a C-terminal LBD with the activation function-2 (AF-2) region. While structurally similar, the N-terminal domain shows only 26-28% sequence homology among NR4A members, yet it plays crucial roles in transcriptional regulation, post-translational modifications, and functional diversification [10,11,12].
NR4A receptors regulate gene expression by binding to DNA response elements either as monomers or dimers. As monomers, they recognise the NGFI-B response element (NBRE; AAAGGTCA) [10], while as homodimers or heterodimers with retinoid X receptors (RXRs), they bind to Nur response elements (NuRE; AAT(G/A)(C/T)CA) or DR5 elements [13,14]. Despite their structural similarities, the three NR4A members exhibit distinct preferences for specific cofactors and response elements, contributing to their functional diversity [15]. Originally identified for their roles in the nervous system, NR4A receptors are now known to be widely expressed and to regulate critical cellular processes including proliferation, apoptosis, and metabolism in a tissue-specific manner [12].
These receptors are classified as immediate early genes, showing rapid induction within minutes of cellular stimulation. They respond to diverse signals, including T-cell receptor activation, cellular stress, GPCR signalling, and cAMP pathways [16,17]. Unlike classical NRs, NR4As are considered “orphan receptors” as no endogenous ligands have been conclusively identified. Structural studies reveal that their LBDs contain a collapsed orthosteric pocket obstructed by bulky hydrophobic residues, making conventional ligand binding unlikely [18]. Current models suggest that NR4A activity is primarily regulated through post-translational modifications, protein-protein interactions, and changes in cellular localisation [19,20]. However, emerging evidence indicates that the LBD may adopt alternative conformations capable of binding unsaturated fatty acids, prostaglandins, and dopamine metabolites, though the physiological significance of these interactions requires further investigation [21].
In the immune system, NR4A receptors play well-established roles in modulating Nuclear Factor κB (NF-κB) activity, particularly in myeloid cells. Under resting conditions, NF-κB is sequestered in the cytoplasm by inhibitory IκB proteins. Following cellular activation, IκB phosphorylation and degradation allows NF-κB nuclear translocation and subsequent inflammatory gene expression. NR4A receptors, like other nuclear receptors, can suppress this pathway at multiple levels, and their loss can lead to excessive NF-κB activation and hyperinflammatory responses [10].
In lymphoid cells, NR4A receptors are rapidly induced by T-cell and B-cell receptor signaling [22,23,24], as well as by inflammatory cytokines and stress stimuli [13,25,26,27]. In During thymocyte development, they mediate apoptosis of self-reactive cells, contributing to central tolerance [25,28]. In mature T cells, they influence effector differentiation, affecting Th1, Th17, and regulatory T cell functions [29,30,31,32]. Notably, NR4A1 has been shown to prevent exhaustion in chronically stimulated CD8+ T cells [33], enhancing the suppressive activity of regulatory T cells [34]. In B cells, these receptors regulate survival, proliferation, and antibody production; their dysregulation is implicated in autoimmune and lymphoproliferative disorders [35]. The diverse, context-dependent functions of NR4A receptors - ranging from pro-apoptotic to pro-survival and anti-inflammatory to pro-inflammatory - highlight their potential as therapeutic targets for immune-related diseases. The following sections will explore these roles in greater detail, focusing on their contributions to myeloid and lymphoid cell function.
3. NR4A Receptors in Myeloid Cells
Myeloid cells—including monocytes, macrophages, dendritic cells (DCs), and granulocytes, play central roles in innate immunity and inflammation. These cells patrol tissues for pathogens and damage, responding through phagocytosis, cytokine/chemokine secretion, and antigen presentation to bridge innate and adaptive immunity. Myeloid cells exhibit remarkable plasticity, adopting either pro-inflammatory (e.g., M1 macrophages) or anti-inflammatory (e.g., M2 macrophages, tolerogenic DCs) phenotypes depending on microenvironmental cues. Dysregulation of myeloid function contributes to various diseases, from atherosclerosis to cancer, where aberrant cytokine production or immunosuppressive behaviour can either exacerbate pathology or hinder therapy. Nuclear receptors NR4A1-3 are emerging as key regulators of myeloid cell differentiation, metabolic reprogramming and effector functions, making them attractive therapeutic targets. While their roles in neutrophils and mast cells remain less explored, current research has significantly advanced our understanding of NR4A functions in macrophages and DCs [36]. The main functions described for NR4A receptors in myeloid cells are summarized in Figure 1.
3.1. Macrophages
Macrophages serve as crucial sentinels of the innate immune system, responding to infection or injury by producing effector molecules like nitric oxide (NO) and pro-inflammatory cytokines. The robust inflammatory response of classically activated (M1) macrophages is supported by profound metabolic reprogramming that fuels their antimicrobial functions [37]. NR4A receptors are potently induced by various inflammatory stimuli, including cytokines, TLR ligands, and oxidised lipids, suggesting their involvement in macrophage inflammatory responses. The NF-κB signalling pathway appears to be a principal regulator of inducible NR4A expression in these cells [17].
- NR4A1 plays complex roles in macrophage biology, upregulating genes involved in inflammation, apoptosis, and cell cycle regulation. In the context of immune response, while it generally suppresses NF-κB activity and limits inflammatory cytokine production, its effects on the high inflammatory cytokine Tumoral Necrotic Factor α (TNF-α), appear distinct - neither expression nor secretion of this cytokine is altered in NR4A1-deficient macrophages, This exception may relate to succinate dehydrogenase (SDH) activity, which exerts independent anti-inflammatory effects [37]. NR4A1 deficiency leads to enhanced NF-κB activation, evidenced by increased p65 phosphorylation, and promotes a pro-inflammatory macrophage phenotype [10].
Beyond inflammation regulation, NR4A1 influences macrophage metabolism. Growing evidence highlights the intricate link between immune response and cellular metabolism, demonstrating that metabolic reprogramming—including rewiring of the TCA cycle and activation of alternative shunt pathways—is essential for activated macrophages to efficiently produce inflammatory effector molecules [38]. Koenis et al. demonstrated that NR4A1 modulates mitochondrial metabolism to restrain excessive inflammatory responses [37]. It is worth mentioning that, in a sepsis model, LPS stimulation of TLR4 triggers NR4A1 phosphorylation via p38α, leading to NF-κB p65 release and pro-inflammatory gene transcription [39].
NR4A1 also contributes to macrophage differentiation, with its expression increasing during the transition from classical M1 to non-classical M2 monocytes in murine models [17]. NR4A1-deficient macrophages from atherosclerotic mice show elevated pro-inflammatory markers (e.g., MHC class II) and reduced anti-inflammatory markers (e.g., Arginase-1), supporting its role in polarisation [40,41].
- NR4A2 was first linked to NF-κB regulation through studies in TLR4-stimulated microglia, where its depletion exacerbated pro-inflammatory responses. Upon TLR4 activation, NR4A2 undergoes SUMOylation and phosphorylation, enabling it to bind phosphorylated NF-κB/p65 at target gene promoters. This interaction recruits the Co-REST repressor complex, displacing NF-κB/p65 and suppressing pro-inflammatory gene expression [10,42].
- In macrophages, NR4A2 expression is induced through the PI3K-Akt-mTOR pathway, which attenuates innate inflammatory responses. NR4A2 promotes M2 polarisation and protects against endotoxin-induced sepsis, suggesting it functions as an inflammatory brake [43]. Interestingly, in autoimmune conditions like bullous pemphigoid, pro-inflammatory macrophages show elevated NR4A2 levels, possibly representing a compensatory mechanism to restrain inflammation [44].
- NR4A3 demonstrates distinct functions in macrophage biology. Silencing NOR1 in human IL-4-polarized macrophages downregulates alternative activation markers (Mannose Receptor, IL-1Ra, CD200R, F13A1, IL-10, PPARγ) while increasing MMP9 expression and activity - typically associated with M1 phenotypes [45]. In atherosclerosis, NR4A3 promotes early disease events by enhancing monocyte adhesion to endothelium. Inflammatory signals like NF-κB activate NR4A3, which induces expression of adhesion molecules (either directly on monocytes or indirectly via endothelial VCAM-1/ICAM-1 upregulation), facilitating immune cell recruitment to vascular walls [46].
3.2. Dendritic Cells
DCs are professional antigen-presenting cells that bridge innate and adaptive immunity by activating pathogen-specific T cells, playing essential roles in both initiating adaptive immunity and maintaining immune tolerance [47]. Their development from bone marrow progenitors involves upregulated expression of all three NR4A receptors during the transition from pre-DCs to mature DCs [17].
- NR4A1 serves as a critical immunoregulator in DCs. Expressed across human and murine DC subsets, its expression rapidly increases upon TLR stimulation. NR4A1-deficient DCs display hyperinflammatory responses with enhanced NF-κB-dependent cytokine production (IL-6, TNFα, IL-12) and increased T-cell stimulatory capacity [41,48]. Conversely, pharmacological activation of NR4A1 suppresses cytokine and attenuates DC-driven allogeneic T-cell proliferation. positioning it as a key checkpoint against excessive immune activation [48].
- NR4A2, while less abundantly expressed in DCs than other family members [17], plays a specialised role in immune tolerance. Saini et al. demonstrated that NR4A2 drives a regulatory phenotype in bone marrow-derived DCs (BMDCs), suppressing autoimmune neuroinflammation through Treg expansion. These findings suggest NR4A2 can reprogram immunogenic DCs toward tolerogenic states [49].
- NR4A3 shows preferential expression in migratory DCs and in essential for their lymph node homing. NR4A3-deficient DCs exhibit reduced CCR7 expression - the key chemokine receptor for DC migration - both at steady-state and upon activation [47,50]. While NR4A3 doesn’t directly bind the CCR7 promoter, it may regulate migration indirectly through transcription factors like FOXO1 or broader migratory programs [17,50].
Functional studies reveal that NR4A3 knockdown impairs DC-mediated T-cell proliferation, and NR4A3 KO mice show increased infection susceptibility due to DC migration defects [51]. Additionally, NR4A3 is required for in vitro monocyte-to-DC differentiation, with its absence causing up to 10-fold reductions in DC generation [17].
4. NR4A Receptors in Lymphoid Cells
Lymphoid cells - including T cells, B cells, and natural killer (NK) cells - form the backbone of adaptive and innate immunity. These cells coordinate antigen-specific responses (T and B cells) and provide rapid cytotoxic activity (NK cells). The NR4A nuclear receptors (NR4A1-3) critically regulate lymphoid cell function: NR4A1 modulates T cell activation and prevents autoimmunity, NR4A3 influences CD8+ T cell exhaustion in chronic infections, and NR4A2 contributes to B cell receptor signaling. Dysregulation of these pathways underlies various immune pathologies, highlighting their therapeutic potential [28].
4.1. T Cells
T cells demonstrate remarkable functional diversity, with subsets specialized for distinct immune roles. CD4+ helper T cells differentiate into lineages including Th1 (antiviral/antitumor), Th2 (anti-helminth/allergy), Th17 (mucosal defense/autoimmunity), and regulatory T cells (Tregs; immune suppression). CD8+ cytotoxic T cells directly eliminate infected or malignant cells. These subsets develop through thymic selection, where NR4A receptors play critical roles in both positive selection (promoting survival of useful clones) and negative selection (eliminating self-reactive cells) [28,52,53,54].
T cells recognize foreign peptide-MHC complexes through their T-cell receptor (TCR), a highly diverse receptor generated via somatic gene rearrangement. Thymic selection plays a critical role in shaping the T-cell repertoire, ensuring self-tolerance while maintaining the ability to detect pathogen-derived antigens. During the CD4+CD8+ double-positive (DP) stage, thymocytes that fail to engage the TCR with peptide-MHC complexes undergo programmed cell death. In contrast, those with intermediate TCR affinity are positively selected to differentiate into either CD4+ or CD8+ single-positive T cells. Thymocytes exhibiting excessive self-reactivity are eliminated through negative selection (primarily at the DP stage) or redirected into Treg lineages to promote immune tolerance. Following selection, mature thymocytes upregulate CCR7, enabling their migration to the medulla for further screening against tissue-restricted antigens [55].
4.1.1. CD4+ T Cells
- CD4+ T cells are the master regulators of adaptive immunity, coordinating immune responses against pathogens and cancer by directing the activity of B cells, macrophages, and CD8+ T cells. Upon recognizing antigen via MHC class II on dendritic cells, naïve CD4+ T cells proliferate and differentiate into specialized subsets dictated by the inflammatory environment: Th1 (IFN-γ producers that activate macrophages and CD8+ T cells), Th2 (IL-4-secreting helpers for B cell class-switching to IgE), Th17 (IL-17-dependent recruiters of neutrophils), Tfh (follicular helpers for B cell germinal center responses), and Tregs (suppressors of autoimmunity). Post-infection, most effector CD4+ T cells undergo apoptosis, but a subset persists as long-lived memory cells—mirroring the CD8+ T cell response [28]. Regarding the diverse repertoire of different populations of T cells, NR4A family is among the few factors directly implicated in thymic selection, orchestrating thymic deletion and Treg diversion, establishing a cell-intrinsic tolerance program upon self-antigen recognition. This process likely serves as a critical fail-safe against autoimmunity [56].
- NR4A1 serves as a master regulator of CD4+ T cells. While expressed at low levels In naïve cells, it rapidly induces upon antigen engagement [28]. suppresses effector T cell responses by inhibiting Th1/Th17 differentiation and cytokine production (IFN-γ, IL-17) while promoting Treg development and function [57]. Mechanistically, NR4A1 competes with AP-1 transcription factors at shared DNA binding sites, directly repressing IL-2 transcript [58]. It also modulates T cell metabolism, with NR4A1 deficiency leading to enhanced glycolysis and oxidative phosphorylation that fuels unchecked proliferation [59,60].
T cells undergo profound metabolic reprogramming to meet the bioenergetic and biosynthetic demands of immune activation. This metabolic plasticity enables the transition from quiescent naive cells to rapidly proliferating effectors and long-lived memory populations. Naive T cells primarily utilise oxidative phosphorylation (OXPHOS) fueled by fatty acid oxidation (FAO), reflecting their low energy requirements in the resting state. Upon antigen recognition, activated effector T cells shift toward aerobic glycolysis, to support rapid proliferation and effector molecule synthesis. Memory T cells, by contrast, re-establish metabolic flexibility with enhanced mitochondrial spare respiratory capacity, allowing rapid recall responses through efficient OXPHOS [60]. This metabolic hierarchy ensures the appropriate allocation of energy across T cell differentiation states while maintaining redox balance. NR4A1 has emerged as a critical regulator interfacing T cell metabolism with functional outcomes [59].
Studies demonstrate that NR4A1 constrains T cell activation by limiting both cell cycle progression and metabolic reprogramming. Genetic ablation of NR4A1 results in hyperactive CD4+ T cells exhibiting elevated oxidative respiration, glycolytic flux, and glycolytic capacity - metabolic changes that likely fuel the unchecked proliferation observed in NR4A1-deficient T cells [60]. Mechanistically, NR4A1 appears to function as a metabolic checkpoint, preventing excessive activation by modulating both mitochondrial and glycolytic pathways, though the precise molecular targets remain under investigation [59].
At the transcriptional level, NR4A1 exerts broad immunosuppressive effects through competitive interference with pro-inflammatory signalling networks. Chromatin immunoprecipitation sequencing (ChIP-seq) analyses reveal that NR4A1 co-localizes with both canonical NBRE motifs and AP-1 consensus sequences, suggesting functional antagonism with bZIP transcription factors [58]. This competition was experimentally validated through multiple approaches: NR4A1 overexpression reduces c-Jun occupancy at shared genomic targets, while electrophoretic mobility shift assays (EMSAs) and luciferase reporter systems demonstrate direct repression of AP-1-driven transcription [28]. Notably, this molecular interplay inhibits IL-2 production - a critical growth factor for T cell expansion - providing a mechanism for NR4A1-mediated suppression of effector responses. The ability of NR4A1 to simultaneously modulate metabolic programs and compete with activation-associated transcription factors positions it as a multifaceted regulator of T cell homeostasis.
The NR4A family, particularly NR4A1, plays indispensable roles in thymocyte development and selection. During thymic education, NR4A1 expression dynamically responds to TCR signal strength, with highest induction in thymocytes undergoing negative selection [26,61]. This expression pattern correlates with its dual mechanisms of clonal deletion: (1) transcriptional upregulation of pro-apoptotic BIM (Bcl2l11) through canonical nuclear receptor activity, and (2) direct mitochondrial targeting where it binds BCL-2, exposing its BH3 domain to trigger caspase-independent apoptosis [26,62]. While these pathways ensure elimination of strongly self-reactive clones, the contextual signals determining whether NR4A1 promotes deletion or Treg differentiation remain incompletely understood. Recent work by Moran et al. [61] highlights NR4A1’s quantitative response to TCR signal intensity, with graded expression levels potentially serving as a rheostat for selection outcomes.
NR4A1 plays a central role in Tregs. These cells represent a specialized CD4+ lineage critical for maintaining immune tolerance. Their suppressive arsenal includes cell-contact-dependent mechanisms (CTLA-4, CD25), anti-inflammatory cytokines (IL-10, TGF-β), and active suppression of pro-inflammatory signals (IFN-γ, IL-2) [63]. Tregs further modulate immune responses by restraining dendritic cell-mediated T cell priming and inhibiting follicular helper T (Tfh) cell differentiation, with specialized follicular regulatory T (Tfr) cells directly regulating germinal centre reactions [63].
NR4A1 serves as a master regulator of Treg biology through multiple non-redundant functions. It directly activates Foxp3 transcription - the lineage-defining transcription factor - and maintains Treg stability by sustaining expression of effector molecules (Ikzf4, CD25) while repressing inflammatory cytokines (IL-4, IL-21) [23,25,63,64]. In NR4A1-deficient models, Tregs lose Foxp3 expression and aberrantly differentiate into Th2 and Tfh-like cells, culminating in fatal autoimmunity [26,64]. The preferential enrichment of NR4A1 in peripheral Tregs versus conventional T cells [26] underscores its specialised role in preserving immune tolerance beyond thymic development.
NR4A1’s functional impact varies across T cell subsets and activation contexts. In Tfh cells, its effects appear context-dependent, modulating but not being essential for their development [63]. Notably, NR4A1 induction provides a sensitive biomarker of recent TCR engagement, outperforming traditional markers like CD69 in specificity for antigen receptor signalling [65] [6. This property, combined with its roles in metabolic regulation, apoptosis induction, and transcriptional modulation, makes NR4A1 an attractive target for immunotherapeutic interventions. Ongoing research continues to elucidate how NR4A1 integrates these diverse functions to maintain immune homeostasis while permitting appropriate responses to pathogens.
- NR4A2. While NR4A1 and NR4A3 serve as primary regulators of Treg biology, NR4A2 exhibits unique and context-dependent functions in immune homeostasis. Unlike its family members, NR4A2 protein remains undetectable in stimulated thymocytes despite mRNA expression, suggesting divergent regulatory mechanisms during thymic selection [66]. In mature CD4+ T cells, NR4A2 demonstrates partial functional redundancy with other NR4As, as evidenced by only modest reductions in Foxp3 and CD25 expression in NR4A2 single-knockout models [63]. However, its specific capacity to suppress IL-4 promoter activity in reporter assays [26] and maintain Treg identity through conserved Foxp3 enhancer elements (CNS1/2) [67,68,69] indicates specialised roles in peripheral immune regulation.
NR4A2 presents a paradoxical association with autoimmune inflammation. In multiple sclerosis (MS) and experimental autoimmune encephalomyelitis (EAE), T cells show marked upregulation of NR4A2 transcripts, and the receptor overexpression enhances production of pathogenic cytokines IFN-γ and IL-17 [26]. However, mechanistic studies reveal that NR4A2 is required for terminal Th17 differentiation through an IL-21-dependent autocrine loop. NR4A2-deficient Th17 precursors maintain RORγt expression but fail to upregulate IL-23R or produce IL-17/IL-21, defects rescued by exogenous IL-21 [29]. This establishes NR4A2 as a critical checkpoint in Th17 maturation, explaining its dual association with both disease pathogenesis and developmental control of this inflammatory lineage [70]
Recent work by Zhang et al. [71]. identified NR4A2 as the crucial molecular link between coactivator signalling and Treg differentiation. Their studies demonstrated that 1) SRC2 (Steroid Receptor Coactivator 2) physically interacts with NFAT1 to drive NR4A2 transcription, 2) SRC2 deficiency impairs NR4A2 expression and FoxP3 induction, causing autoimmune pathology, and 3) NR4A2 restoration rescues Treg development in SRC2-knockout cells. Mechanistically, SRC2 recruits NFAT1 to the NR4A2 promoter, creating a feed-forward loop that amplifies FoxP3 expression. This SRC2-NFAT1-NR4A2 axis represents a novel therapeutic target for modulating immune tolerance, with NR4A2 serving as the essential transcriptional bridge between upstream signaling and Treg lineage commitment [71].
- NR4A3 plays dual yet distinct roles in thymocyte development and Treg differentiation. During thymic selection, NR4A3 functions redundantly with NR4A1 to enforce central tolerance, particularly in mediating negative selection of self-reactive thymocytes [62]. Both nuclear receptors trigger apoptosis through a shared mitochondrial pathway - following nuclear export, they bind and convert anti-apoptotic BCL-2 into a pro-apoptotic form by exposing its BH3 domain [28]. This non-transcriptional mechanism complements their transcriptional regulation of apoptotic genes, providing a fail-safe for eliminating autoreactive clones.
The receptor’s role shifts markedly during Treg development, where persistent NR4A3 expression serves as a molecular signature of ongoing differentiation [72,73]. Unlike NR4A1 which responds to transient TCR signals, NR4A3 specifically integrates strong, sustained TCR stimulation through direct induction by NFAT transcription factors: NFAT, AP1, and NF-κB [73]. Notably, NFAT directly induces NR4A3 [74] inking sustained TCR signals to Treg lineage commitment. This higher activation threshold positions NR4A3 as a specialized sensor of prolonged antigen recognition - the type of signal that drives Treg lineage commitment rather than deletion. The continued expression of NR4A3 in peripheral Tregs suggests these cells maintain aspects of this signaling program even after thymic egress [73].
Notably, NR4A3 exhibits several unique regulatory features that make it particularly valuable for studying T cell activation. Along with NR4A1, it is rapidly induced by TCR engagement and ERK signaling [73], but its expression dynamics provide superior resolution of strong versus weak activation compared to conventional markers like CD69 [61,72]. These properties, combined with its specific association with Treg development, make NR4A3 both a critical functional mediator and a sensitive reporter of high-affinity TCR interactions throughout T cell maturation.
4.1.2. CD8+ T Cells
CD8+ T cells play essential roles in immune protection against infections and cancer. Upon activation, they differentiate into effector cells capable of secreting cytokines (e.g., IFN-γ, TNF-α) and cytolytic molecules (e.g., perforin, granzymes) to eliminate target cells. Chronic antigen exposure in persistent infections or cancer can induce T cell exhaustion, characterized by reduced functionality and increased expression of inhibitory receptors (e.g., PD-1, LAG-3). These cells also generate diverse memory subsets, including self-renewing central memory (TCM) cells, rapidly responsive effector memory (TEM) cells, and tissue-resident memory (TRM) cells that provide local barrier protection. Their differentiation and function are tightly regulated by transcription factors (e.g., T-bet, EOMES, NR4A family), metabolic pathways, and microenvironmental cues, ensuring balanced immune responses and self-tolerance [28,75].
- NR4A1 orchestrates CD8+ T cell responses through stage-specific functions. During initial T cell receptor (TCR) activation, NR4A1 expression peaks rapidly (1-3 hours post-stimulation), with single-cell RNA sequencing demonstrating its transcription levels directly correlate with TCR signal strength [76]. This immediate-early response supports initial T cell activation while simultaneously establishing regulatory checkpoints. Under conditions of chronic antigen exposure, NR4A1 undergoes functional switching - upregulating the pro-apoptotic factor BIM to trigger mitochondrial apoptosis of overactivated clones, thereby preventing immunopathology [28]. This dual role as both activation promoter and termination signal is evidenced in NR4A1-deficient models, which exhibit enhanced antitumor activity but also dysregulated T cell expansion [12,59].
A central mechanism of NR4A1-mediated regulation involves direct suppression of Interferon Regulatory Factor 4 (IRF4) transcription. Loss of this regulatory brake in NR4A1-deficient CD8+ T cells leads to uncontrolled IRF4 expression, driving hyperproliferation and excessive differentiation into short-lived effector cells (SLECs) [28,77]. This IRF4 dysregulation results in heightened cytokine production and accelerated exhaustion, positioning NR4A1 as a critical modulator of effector-memory balance.
NR4A1 further contributes to immune tolerance by enforcing T cell anergy. This process is regulated by a dynamic balance between co-stimulatory and co-inhibitory signals, where dominant co-inhibitory pathways (e.g., PD-1) enforce tolerance and drive T cell dysfunction. Transcriptomic analyses reveal its upregulation in anergic T cells correlates with suppressed effector function - particularly reduced IFNγ production in CD8+ T cells. Conversely, NR4A1 deletion enhances proliferative capacity and cytokine secretion (IL-2, IFNγ), establishing its role as a transcriptional mediator of exhaustion [78,79]. These findings establish NR4A1 as a central transcriptional mediator of T cell exhaustion.
Further supporting the role of NR4A1 in exhaustion, studies in tumour microenviroment demonstrated that NR4A1 promotes surface expression of exhaustion markers PD-1 and TIM-3 on CD8+ T cells, as well as reduces effector cytokine production. Furthermore, it expression correlates with exhaustion-associated gene signatures in tumor-infiltrating lymphocytes [80]. The therapeutic implications of NR4A1 modulation are substantial. Genetic deletion of NR4A1 in T cells enhances antitumor responses through Increased IL-2 and IFNγ production, Reduced PD-1 expression and Improved T cell persistence [58,81]. Strikingly, adoptive transfer of triple NR4A knockout CAR-T cells in murine solid tumour models resulted in complete tumour eradication and improved survival [79], highlighting the therapeutic potential of targeting this pathway to enhance T cell function in cancer immunotherapy.
- NR4A2. As well as NR4A1, NR4A2 participates in CD8+ T cell exhaustion. A recent work of Srirat et al. demonstrated that genetic deletion of NR4A2 in CD8+ T cells reduced tumour size, with the double-knockout in NR4A1/2 showing a strong antitumoral response [82].
Regarding its participation in memory response, studies evaluating the heterogeneity of TRM cells following LCMV infection by scRNA-Seq have revealed that the highly functional CD28+ subset of CD8+ TRM cells is particularly enriched in NR4A1-3 and that knockdown of NR4A2 specifically decreased the proportion of these CD28+ TRM cells [28,75].
- NR4A3. While NR4A1 and NR4A3 share overlapping expression patterns, accumulating evidence reveals their distinct functional roles in T cell biology. NR4A3 serves as a specific marker for thymocytes receiving strong TCR signals during negative selection, with NR4A3 deficiency impairing the deletion of autoreactive clones [55]. Their differential expression patterns further highlight this functional divergence - NR4A1 responds to both positive and negative selection signals (albeit more strongly to the latter), while NR4A3 induction occurs exclusively in response to high-affinity TCR engagement [28]. These findings fundamentally challenge the concept of complete redundancy between these nuclear receptor family members.
In CD8+ T cell differentiation, NR4A3 plays unique regulatory roles through epigenetic mechanisms. During early activation, it restricts chromatin accessibility at bZIP transcription factor binding sites (including Fos/Jun family motifs), thereby suppressing memory-promoting programs while favouring short-lived effector cell (SLEC) differentiation [28,76]. This explains why NR4A3 deficiency enhances central memory (TCM) formation, demonstrating its specialised role in balancing effector and memory fates [76]. The restriction of bZIP factor binding represents a key mechanism through which NR4A family members coordinate T cell responses.
The NR4A receptors exhibit stage-specific modulation of CD8+ T cell responses during immune challenges. While both NR4A1 and NR4A3 suppress cytokine production during the effector phase, only NR4A3 actively promotes SLEC differentiation [28]. Notably, NR4A3 shows differential regulation from NR4A1, as it remains unresponsive to tonic signalling in both T and B cells - a property confirmed by early TCR stimulation sequencing data [28,76]. This suggests NR4A3’s effects on differentiation occur independently of tonic signalling pathways.
In thymocyte development, NR4A1 and NR4A3 demonstrate both shared and specialized functions. Both receptors are sharply upregulated during negative selection and cooperate with BIM to eliminate self-reactive clones through dual mechanisms: transcriptional activation of pro-apoptotic genes and direct mitochondrial targeting of BCL-2 to expose its BH3 domain [28]. Despite this functional overlap in clonal deletion, they maintain distinct expression dynamics and activation thresholds that tailor their responses to different TCR signal strengths.
Beyond thymic selection, the NR4A family plays an indispensable role in regulatory T cell (Treg) biology. Complete deletion of NR4A1 and NR4A3 (with or without NR4A2) leads to near-total Treg loss and severe autoimmunity in mice. These receptors reinforce Foxp3 expression in thymic Treg precursors and maintain peripheral Treg stability by suppressing inflammatory cytokine production while preserving suppressive function. Their coordinated activity links strong self-antigen recognition to Treg lineage commitment while preventing aberrant Th1/Th2 differentiation in conventional CD4+ T cells, highlighting their central role in maintaining immune tolerance at multiple levels.
In summary, NR4A receptors emerge as central regulators of T cell function and biology, with their diverse actions schematized in Figure 2.
4.2. B Cells
B cells constitute a fundamental component of adaptive immunity, responsible for antibody production, pathogen clearance, and immunological memory establishment. Developing in the bone marrow, they undergo activation in secondary lymphoid organs following antigen encounter, ultimately differentiating into antibody-secreting plasma cells or memory B cells. Their activation and function depend critically on B cell receptor (BCR) signaling and productive interactions with helper T cells. Similar to T cells, B cells rapidly upregulate NR4A family members upon BCR engagement, with expression detectable at transcriptional, translational, and reporter levels. Among these nuclear receptors, NR4A1 shows the highest basal and activation-induced expression, followed by NR4A3, while NR4A2 remains minimally expressed in B cells [60].
- NR4A1. During the GC response, B cell clones compete for entry and dominance, with selection typically favouring those expressing high-affinity B cell receptors (BCRs). However, GCs are not exclusively dominated by high-affinity clones; they can sustain a heterogeneous population, including low-affinity B cells, over extended periods. This suggests the presence of regulatory mechanisms that prevent early monopolization by dominant clones. Recent studies highlight the orphan nuclear receptor NR4A1 as a key negative regulator in this process. Upon BCR-antigen engagement, NR4A1 is rapidly induced, forming a negative feedback loop that curbs B cell proliferation and restricts the early expansion of high-affinity clones [83].
NR4A1 and NR4A3 jointly restrain B cell survival and proliferation when signal 1 (antigen) is received in the absence of signal 2 (costimulation via T-cell help or presence of Pathogen-Associated Molecular Patterns). However, strong signal 2 overrides this suppression. Notably, NR4As also inhibit BCR-induced upregulation of key interaction molecules CD86 (critical for CD28 co-stimulation), CCL3/CCL4 (which recruit T cells via CCR5), and ICAM1/2 (essential for T-B cell conjugation)—all of which are required for efficient B cell expansion in T-dependent responses [84]. This suppression confers a competitive disadvantage on NR4A-deficient B cells in competing for scarce T-cell help [66].
Functionally, NR4A receptors temper the early dominance of high-affinity clones in polyclonal immune responses, enabling the participation of lower-affinity or subdominant clones [83]. This mechanism may safeguard lower-affinity but potentially neutralising antibody responses elicited during infection. Acting as a negative feedback loop in B cells, NR4As help maintain clonal diversity in the GC reaction, which could ultimately optimise long-term affinity maturation [66].
In summary, these findings reveal a molecular pathway that restrains immunodominance and preserves clonal diversity during humoral responses. This regulation may prevent gaps in the post-immune repertoire that pathogens could exploit while enhancing the affinity maturation of long-lived plasma cells [83].
While NR4A2 shows minimal expression and no apparent role in B cell signalling, NR4A3 functions redundantly with NR4A1 despite its lower expression levels [84].
4.3. NKT Cell
Natural Killer T (NKT) cells are a unique subset of T lymphocytes that bridge innate and adaptive immunity by recognizing lipid antigens presented by CD1d molecules rather than peptide-MHC complexes. These cells express both TCRs) and natural killer cell markers, enabling rapid cytokine production upon activation. NKT cells are classified into two main subsets: type I (or invariant) NKT cells expressing a semi-invariant TCRα chain, and type II NKT cells with more diverse TCRs. Upon activation, NKT cells can quickly secrete large amounts of Th1, Th2, or Th17 cytokines, thereby modulating various immune responses including tumor surveillance, antimicrobial defense, and autoimmune regulation. Their unique antigen recognition properties and rapid response capabilities make NKT cells crucial players in immune homeostasis [85].
iNKT cells arise in the thymus from precursors that share a developmental trajectory with conventional T cells until the CD4+CD8+ double-positive (DP) stage. Beyond this point, iNKT lineage commitment is driven by the recognition of self-lipid antigens presented by CD1d molecules, which engage their semi-invariant TCR. This interaction, along with cytokine signals such as IL-7 and IL-15, initiates a unique transcriptional program involving key regulators like NR4A1 [61,66,85]. Recent studies have highlighted NR4A1 as a critical checkpoint in iNKT cell biology, influencing not only development but also immune tolerance and effector function. Nur77 enforces self-tolerance by two major mechanisms: 1) inducing caspase-3–mediated apoptosis during thymic selection (negative selection) and 2) promoting a hyporesponsive state in peripheral iNKT cells. These regulatory roles are distinct from their functions in conventional T cells, emphasising the unique pathways governing iNKT cell homeostasis. Given their potent immunomodulatory capacity, tight control of iNKT cell activation is essential to prevent autoimmunity or inflammatory pathology. One proposed mechanism involves Nur77-dependent feedback inhibition, where excessive TCR or cytokine signalling upregulates NR4A1 to restrain iNKT cell activity [85].
5. NR4A Receptors in Pathology
NR4A receptors have key physiological roles (besides immunological ones), which include proliferation, apoptosis, DNA repair, cellular stress, memory, endocrinology, neuronal signaling, and hematopoietic, immune and metabolic processes, that have been demonstrated in many cell types. Moreover, these receptors are involved in the onset and progression of numerous diseases, such as obesity, atherosclerosis, inflammation and cancer (Table 1) [86]. In autoimmune diseases, NR4A1–3 modulate immune cell function and tolerance, particularly by influencing Th17 differentiation and NF-κB signaling. These receptors act as early regulators of inflammation, with aberrant expression linked to immune-mediated rheumatic diseases (IMRDs), where they are found in inflamed synovial tissue (e.g., in arthritis) and psoriatic lesions [10,11,41,87]. NR4A2, for instance, promotes TNF-α production in synovial cells [87,88], while in multiple sclerosis (MS), it governs IL-17 expression. Silencing NR4A2 in experimental autoimmune encephalomyelitis (EAE) mitigates disease severity [88], whereas NR4A1 deficiency exacerbates EAE via enhanced CNS macrophage infiltration [89].
In hematologic malignancies, the expression and function of NR4A receptors have been studied in lymphomas, where decreased nuclear levels of NR4A1 and NR4A3 were observed in follicular lymphoma and diffuse large B-cell lymphoma (DLBCL) compared to their normal cell counterparts. Reduced NR4A1 expression was associated with more aggressive disease and poorer patient survival, whereas NR4A2 was inconsistently detected and showed no significant difference between tumour and non-tumour tissues. Notably, NR4A3 overexpression was linked to a favourable response to chemotherapy in DLBCL patients, suggesting a potential tumour-suppressive role in these malignancies. Evidence also indicates that the loss of NR4A1 and NR4A3 may contribute to the development and progression of certain leukaemias and lymphomas [16,90,91,92].
In contrast, NR4A receptors, particularly NR4A1, have been extensively investigated in solid tumours, where they often exhibit oncogenic properties. While most in vivo studies suggest that NR4A1 promotes tumour growth in cancers such as breast, lung, pancreatic, ovarian, colon, and others, high expression of this receptor is associated with poor patient survival. Interestingly, this contrasts with its tumour-suppressive role in hematologic malignancies, highlighting a tissue-specific duality in NR4A1 function. Although NR4A2 and NR4A3 have been less studied in solid tumours, existing data suggest that NR4A2 may also contribute to oncogenic processes. These differences underscore the complex and context-dependent roles of NR4A receptors in cancer biology [16,81,93].
In another hand, NR4A receptors are crucial regulators of energy metabolism, influencing obesity and diabetes through their roles in glucose and lipid homeostasis. They modulate insulin sensitivity, gluconeogenesis, and mitochondrial function, with tissue-specific effects: promoting glucose utilisation in muscle and fat while stimulating hepatic glucose production and reducing lipid accumulation in muscle. However, their impacts on liver and adipose tissue metabolism remain incompletely understood, and their effects on insulin secretion and resistance vary across experimental models. While NR4As participate in organ-specific metabolic control, their systemic coordination of whole-body energy balance through inter-organ communication remains unresolved [94].
Although NR4As do not rely on endogenous ligands for their transcriptional activity and regulation, several modulators have been identified. These include potential endogenous ligands such as prostaglandins and tryptophan/indolic metabolites, as well as exogenous ligands like cytosporone B, celastrol, and bis-indole-derived compounds [21,95,96,97,98]. Both agonists and antagonists—particularly those selective for specific NR4A family members—could have broad applications in autoimmunity, inflammatory diseases, cancer, transplantation, and vaccine development. Given their role in immune regulation, these nuclear receptors present a unique opportunity to modulate pathways in both myeloid and lymphoid cells. Such strategies may prove especially valuable in chronic diseases, where persistent immune dysregulation disrupts homeostasis. Further research into NR4A functions in immunity and their ligand interactions will be crucial for developing targeted therapies.
Author Contributions
writing—original draft preparation SLF; Conceptualization, Funding acquisition, Writing– original draft, Writing– review & editing N.S. All authors have read and agreed to the published version of the manuscript.
Funding
The authors declare that financial support was received for the research, authorship, and/or publication of this article. This work was supported by grants from Fond for Scientific and Technological Research-FONCyT-(PICT-2019-00044) and Facultad de Ciencias Medicas, Universidad Nacional de Rosario, Rosario, Argentina.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Volle, D.H. Nuclear receptors in physiology and pathophysiology. Mol. Asp. Med. 2021, 78, 100956. [Google Scholar] [CrossRef]
- Xu, P. Nuclear Receptors in Health and Diseases. Int. J. Mol. Sci. 2023, 24, 9153. [Google Scholar] [CrossRef]
- Sever, R.; Glass, C.K. Signaling by Nuclear Receptors. Cold Spring Harb. Perspect. Biol. 2013, 5, a016709–a016709. [Google Scholar] [CrossRef]
- Volle, D.H. Nuclear receptors as pharmacological targets, where are we now? Cell. Mol. Life Sci. 2016, 73, 3777–3780. [Google Scholar] [CrossRef]
- Kiss, M.; Czimmerer, Z.; Nagy, L. The role of lipid-activated nuclear receptors in shaping macrophage and dendritic cell function: From physiology to pathology. J. Allergy Clin. Immunol. 2013, 132, 264–286. [Google Scholar] [CrossRef] [PubMed]
- Fan, R.; Pineda-Torra, I.; Venteclef, N. Editorial: Nuclear Receptors and Coregulators in Metabolism and Immunity. Front. Endocrinol. 2021, 12, 828635. [Google Scholar] [CrossRef] [PubMed]
- Milbrandt, J. Nerve growth factor induces a gene homologous to the glucocorticoid receptor gene. Neuron 1988, 1, 183–188. [Google Scholar] [CrossRef]
- Law SW, Conneely OM, DeMayo FJ, O’Malley BW (1992) Identification of a new brain-specific transcription factor, NURR1. Molecular Endocrinology 6:2129–2135. [CrossRef]
- Ohkura, N.; Hijikuro, M.; Yamamoto, A.; Miki, K. Molecular Cloning of a Novel Thyroid/Steroid Receptor Superfamily Gene from Cultured Rat Neuronal Cells. Biochem. Biophys. Res. Commun. 1994, 205, 1959–1965. [Google Scholar] [CrossRef]
- Murphy, E.P.; Crean, D. Molecular Interactions between NR4A Orphan Nuclear Receptors and NF-κB Are Required for Appropriate Inflammatory Responses and Immune Cell Homeostasis. Biomolecules 2015, 5, 1302–1318. [Google Scholar] [CrossRef]
- Ranhotra, H.S. The NR4A orphan nuclear receptors: mediators in metabolism and diseases. J. Recept. Signal Transduct. 2014, 35, 184–188. [Google Scholar] [CrossRef] [PubMed]
- Herring JA, Elison WS, Tessem JS (2019) Function of nr4a orphan nuclear receptors in proliferation, apoptosis and fuel utilization across tissues. Cells 8. [CrossRef]
- Crean D, Murphy EP (2021) Targeting NR4A Nuclear Receptors to Control Stromal Cell Inflammation, Metabolism, Angiogenesis, and Tumorigenesis. Front Cell Dev Biol 9:. [CrossRef]
- Yu X, He Y, Kamenecka TM, Kojetin DJ (2025) Towards a unified molecular mechanism for liganddependent activation of NR4A-RXR heterodimers. bioRxiv. [CrossRef]
- de Vera, I.M.S.; Munoz-Tello, P.; Zheng, J.; Dharmarajan, V.; Marciano, D.P.; Matta-Camacho, E.; Giri, P.K.; Shang, J.; Hughes, T.S.; Rance, M.; et al. Defining a Canonical Ligand-Binding Pocket in the Orphan Nuclear Receptor Nurr1. Structure 2019, 27, 66–77.e5. [Google Scholar] [CrossRef]
- Safe, S.; Karki, K. The Paradoxical Roles of Orphan Nuclear Receptor 4A (NR4A) in Cancer. Mol. Cancer Res. 2020, 19, 180–191. [Google Scholar] [CrossRef]
- Boulet, S.; Le Corre, L.; Odagiu, L.; Labrecque, N. Role of NR4A family members in myeloid cells and leukemia. Curr. Res. Immunol. 2022, 3, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Kurakula, K.; Koenis, D.S.; van Tiel, C.M.; de Vries, C.J. NR4A nuclear receptors are orphans but not lonesome. Biochim. et Biophys. Acta (BBA) - Mol. Cell Res. 2014, 1843, 2543–2555. [Google Scholar] [CrossRef] [PubMed]
- Zárraga-Granados, G.; Muciño-Hernández, G.; Sánchez-Carbente, M.R.; Villamizar-Gálvez, W.; Peñas-Rincón, A.; Arredondo, C.; Andrés, M.E.; Wood, C.; Covarrubias, L.; Castro-Obregón, S.; et al. The nuclear receptor NR4A1 is regulated by SUMO modification to induce autophagic cell death. PLOS ONE 2020, 15, e0222072. [Google Scholar] [CrossRef]
- McMorrow JP, Murphy EP (2011) Inflammation: A role for NR4A orphan nuclear receptors? Biochem Soc Trans 39:688–693.
- Munoz-Tello, P.; Lin, H.; Khan, P.; de Vera, I.M.S.; Kamenecka, T.M.; Kojetin, D.J. Assessment of NR4A Ligands That Directly Bind and Modulate the Orphan Nuclear Receptor Nurr1. J. Med. Chem. 2020, 63, 15639–15654. [Google Scholar] [CrossRef]
- Liu, Z.-G.; Smith, S.W.; McLaughlin, K.A.; Schwartz, L.M.; Osborne, B.A. Apoptotic signals delivered through the T-cell receptor of a T-cell hybrid require the immediate–early gene nur77. Nature 1994, 367, 281–284. [Google Scholar] [CrossRef]
- Woronicz, J.D.; Calnan, B.; Ngo, V.; Winoto, A. Requirement for the orphan steroid receptor Nur77 in apoptosis of T-cell hybridomas. Nature 1994, 367, 277–281. [Google Scholar] [CrossRef]
- Hiwa, R.; Brooks, J.F.; Mueller, J.L.; Nielsen, H.V.; Zikherman, J. NR4A nuclear receptors in T and B lymphocytes: Gatekeepers of immune tolerance*. Immunol. Rev. 2022, 307, 116–133. [Google Scholar] [CrossRef]
- Sekiya, T.; Kashiwagi, I.; Yoshida, R.; Fukaya, T.; Morita, R.; Kimura, A.; Ichinose, H.; Metzger, D.; Chambon, P.; Yoshimura, A. Nr4a receptors are essential for thymic regulatory T cell development and immune homeostasis. Nat. Immunol. 2013, 14, 230–237. [Google Scholar] [CrossRef]
- Bonta, P.I.; van Tiel, C.M.; Vos, M.; Pols, T.W.; van Thienen, J.V.; Ferreira, V.; Arkenbout, E.K.; Seppen, J.; Spek, C.A.; van der Poll, T.; et al. Nuclear Receptors Nur77, Nurr1, and NOR-1 Expressed in Atherosclerotic Lesion Macrophages Reduce Lipid Loading and Inflammatory Responses. Arter. Thromb. Vasc. Biol. 2006, 26, 2288–2288. [Google Scholar] [CrossRef] [PubMed]
- Jeanneteau, F.; Barrère, C.; Vos, M.; De Vries, C.J.; Rouillard, C.; Levesque, D.; Dromard, Y.; Moisan, M.-P.; Duric, V.; Franklin, T.C.; et al. The Stress-Induced Transcription Factor NR4A1 Adjusts Mitochondrial Function and Synapse Number in Prefrontal Cortex. J. Neurosci. 2018, 38, 1335–1350. [Google Scholar] [CrossRef] [PubMed]
- Odagiu, L.; May, J.; Boulet, S.; Baldwin, T.A.; Labrecque, N. Role of the Orphan Nuclear Receptor NR4A Family in T-Cell Biology. Front. Endocrinol. 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Raveney, B.J.E.; Oki, S.; Yamamura, T.; Stangel, M. Nuclear Receptor NR4A2 Orchestrates Th17 Cell-Mediated Autoimmune Inflammation via IL-21 Signalling. PLOS ONE 2013, 8, e56595. [Google Scholar] [CrossRef]
- Hibino, S.; Chikuma, S.; Kondo, T.; Ito, M.; Nakatsukasa, H.; Omata-Mise, S.; Yoshimura, A. Inhibition of Nr4a Receptors Enhances Antitumor Immunity by Breaking Treg-Mediated Immune Tolerance. Cancer Res. 2018, 78, 3027–3040. [Google Scholar] [CrossRef]
- Bandukwala, H.S.; Rao, A. 'Nurr'ishing Treg cells: Nr4a transcription factors control Foxp3 expression. Nat. Immunol. 2013, 14, 201–203. [Google Scholar] [CrossRef]
- Wang, L.-M.; Zhang, Y.; Li, X.; Zhang, M.-L.; Zhu, L.; Zhang, G.-X.; Xu, Y.-M. Nr4a1 plays a crucial modulatory role in Th1/Th17 cell responses and CNS autoimmunity. Brain, Behav. Immun. 2018, 68, 44–55. [Google Scholar] [CrossRef]
- Seo, H.; Chen, J.; González-Avalos, E.; Samaniego-Castruita, D.; Das, A.; Wang, Y.H.; López-Moyado, I.F.; Georges, R.O.; Zhang, W.; Onodera, A.; et al. TOX and TOX2 transcription factors cooperate with NR4A transcription factors to impose CD8+ T cell exhaustion. Proc. Natl. Acad. Sci. USA 2019, 116, 12410–12415. [Google Scholar] [CrossRef]
- Sekiya, T.; Kashiwagi, I.; Yoshida, R.; Fukaya, T.; Morita, R.; Kimura, A.; Ichinose, H.; Metzger, D.; Chambon, P.; Yoshimura, A. Nr4a receptors are essential for thymic regulatory T cell development and immune homeostasis. Nat. Immunol. 2013, 14, 230–237. [Google Scholar] [CrossRef]
- Tan, C.; Mueller, J.L.; Noviski, M.; Huizar, J.; Lau, D.; Dubinin, A.; Molofsky, A.; Wilson, P.C.; Zikherman, J. Nur77 Links Chronic Antigen Stimulation to B Cell Tolerance by Restricting the Survival of Self-Reactive B Cells in the Periphery. J. Immunol. 2019, 202, 2907–2923. [Google Scholar] [CrossRef]
- Boulet, S.; Le Corre, L.; Odagiu, L.; Labrecque, N. Role of NR4A family members in myeloid cells and leukemia. Curr. Res. Immunol. 2022, 3, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Koenis, D.S.; Medzikovic, L.; van Loenen, P.B.; van Weeghel, M.; Huveneers, S.; Vos, M.; Gogh, I.J.E.-V.; Bossche, J.V.D.; Speijer, D.; Kim, Y.; et al. Nuclear Receptor Nur77 Limits the Macrophage Inflammatory Response through Transcriptional Reprogramming of Mitochondrial Metabolism. Cell Rep. 2018, 24, 2127–2140.e7. [Google Scholar] [CrossRef] [PubMed]
- Ryan DG, O’Neill LAJ (2017) Krebs cycle rewired for macrophage and dendritic cell effector functions. FEBS Lett 591:2992–3006. [CrossRef]
- Li, L.; Liu, Y.; Chen, H.-Z.; Li, F.-W.; Wu, J.-F.; Zhang, H.-K.; He, J.-P.; Xing, Y.-Z.; Chen, Y.; Wang, W.-J.; et al. Impeding the interaction between Nur77 and p38 reduces LPS-induced inflammation. Nat. Chem. Biol. 2015, 11, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Hanna, R.N.; Shaked, I.; Hubbeling, H.G.; Punt, J.A.; Wu, R.; Herrley, E.; Zaugg, C.; Pei, H.; Geissmann, F.; Ley, K.; et al. NR4A1 (Nur77) Deletion Polarizes Macrophages Toward an Inflammatory Phenotype and Increases Atherosclerosis. Circ. Res. 2012, 110, 416–427. [Google Scholar] [CrossRef]
- Murphy, E.P.; Crean, D. NR4A1-3 nuclear receptor activity and immune cell dysregulation in rheumatic diseases. Front. Med. 2022, 9, 874182. [Google Scholar] [CrossRef]
- Saijo, K.; Winner, B.; Carson, C.T.; Collier, J.G.; Boyer, L.; Rosenfeld, M.G.; Gage, F.H.; Glass, C.K. A Nurr1/CoREST Pathway in Microglia and Astrocytes Protects Dopaminergic Neurons from Inflammation-Induced Death. Cell 2009, 137, 47–59. [Google Scholar] [CrossRef]
- Mahajan, S.; Saini, A.; Chandra, V.; Nanduri, R.; Kalra, R.; Bhagyaraj, E.; Khatri, N.; Gupta, P. Nuclear Receptor Nr4a2 Promotes Alternative Polarization of Macrophages and Confers Protection in Sepsis. J. Biol. Chem. 2015, 290, 18304–18314. [Google Scholar] [CrossRef]
- A Solís-Barbosa, M.; Santana, E.; Muñoz-Torres, J.R.; Segovia-Gamboa, N.C.; Patiño-Martínez, E.; A Meraz-Ríos, M.; Samaniego, R.; Sánchez-Mateos, P.; Sánchez-Torres, C. The nuclear receptor Nurr1 is preferentially expressed in human pro-inflammatory macrophages and limits their inflammatory profile. Int. Immunol. 2023, 36, 111–128. [Google Scholar] [CrossRef]
- De Paoli, F.; Eeckhoute, J.; Copin, C.; Vanhoutte, J.; Duhem, C.; Derudas, B.; Dubois-Chevalier, J.; Colin, S.; Zawadzki, C.; Jude, B.; et al. The neuron-derived orphan receptor 1 (NOR1) is induced upon human alternative macrophage polarization and stimulates the expression of markers of the M2 phenotype. Atherosclerosis 2015, 241, 18–26. [Google Scholar] [CrossRef]
- Zhao, Y.; Howatt, D.A.; Gizard, F.; Nomiyama, T.; Findeisen, H.M.; Heywood, E.B.; Jones, K.L.; Conneely, O.M.; Daugherty, A.; Bruemmer, D. Deficiency of the NR4A Orphan Nuclear Receptor NOR1 Decreases Monocyte Adhesion and Atherosclerosis. Circ. Res. 2010, 107, 501–511. [Google Scholar] [CrossRef]
- Boulet, S.; Daudelin, J.-F.; Odagiu, L.; Pelletier, A.-N.; Yun, T.J.; Lesage, S.; Cheong, C.; Labrecque, N. The orphan nuclear receptor NR4A3 controls the differentiation of monocyte-derived dendritic cells following microbial stimulation. Proc. Natl. Acad. Sci. 2019, 116, 15150–15159. [Google Scholar] [CrossRef] [PubMed]
- Tel-Karthaus, N.; Kers-Rebel, E.D.; Looman, M.W.; Ichinose, H.; de Vries, C.J.; Ansems, M. Nuclear Receptor Nur77 Deficiency Alters Dendritic Cell Function. Front. Immunol. 2018, 9, 1797. [Google Scholar] [CrossRef] [PubMed]
- Saini, A.; Mahajan, S.; Gupta, P. Nuclear receptor expression atlas in BMDCs: Nr4a2 restricts immunogenicity of BMDCs and impedes EAE. Eur. J. Immunol. 2016, 46, 1842–1853. [Google Scholar] [CrossRef]
- Park, K.; Mikulski, Z.; Seo, G.-Y.; Andreyev, A.Y.; Marcovecchio, P.; Blatchley, A.; Kronenberg, M.; Hedrick, C.C. The transcription factor NR4A3 controls CD103+ dendritic cell migration. J. Clin. Investig. 2016, 126, 4603–4615. [Google Scholar] [CrossRef]
- Nagaoka, M.; Yashiro, T.; Uchida, Y.; Ando, T.; Hara, M.; Arai, H.; Ogawa, H.; Okumura, K.; Kasakura, K.; Nishiyama, C. The Orphan Nuclear Receptor NR4A3 Is Involved in the Function of Dendritic Cells. J. Immunol. 2017, 199, 2958–2967. [Google Scholar] [CrossRef]
- Dejean, A.S.; Joulia, E.; Walzer, T. The role of Eomes in human CD4 T cell differentiation: A question of context. Eur. J. Immunol. 2018, 49, 38–41. [Google Scholar] [CrossRef]
- Savino W, Mendes-Da-Cruz DA, Lepletier A, Dardenne M (2016) Hormonal control of T-cell development in health and disease. Nat Rev Endocrinol 12:77–89.
- van Hamburg, J.P.; Tas, S.W. Molecular mechanisms underpinning T helper 17 cell heterogeneity and functions in rheumatoid arthritis. J. Autoimmun. 2018, 87, 69–81. [Google Scholar] [CrossRef]
- Boulet, S.; Odagiu, L.; Dong, M.; Lebel, M.; Daudelin, J.-F.; Melichar, H.J.; Labrecque, N. NR4A3 Mediates Thymic Negative Selection. J. Immunol. 2021, 207, 1055–1064. [Google Scholar] [CrossRef]
- Nielsen, H.V.; Yang, L.; Mueller, J.L.; Ritter, A.J.; Hiwa, R.; Proekt, I.; Rackaityte, E.; Aylard, D.; Gupta, M.; Scharer, C.D.; et al. Nr4a1 and Nr4a3 redundantly control clonal deletion and contribute to an anergy-like transcriptome in auto-reactive thymocytes to impose tolerance in mice. Nat. Commun. 2025, 16, 1–22. [Google Scholar] [CrossRef]
- Wang, L.-M.; Zhang, Y.; Li, X.; Zhang, M.-L.; Zhu, L.; Zhang, G.-X.; Xu, Y.-M. Nr4a1 plays a crucial modulatory role in Th1/Th17 cell responses and CNS autoimmunity. Brain, Behav. Immun. 2018, 68, 44–55. [Google Scholar] [CrossRef]
- Liu, X.; Wang, Y.; Lu, H.; Li, J.; Yan, X.; Xiao, M.; Hao, J.; Alekseev, A.; Khong, H.; Chen, T.; et al. Genome-wide analysis identifies NR4A1 as a key mediator of T cell dysfunction. Nature 2019, 567, 525–529. [Google Scholar] [CrossRef]
- Fujii Y, Matsuda S, Takayama G, Koyasu S (2008) ERK5 is involved in TCR-induced apoptosis through the modification of Nur77. Genes to Cells 13:411–419. [CrossRef]
- Liebmann, M.; Hucke, S.; Koch, K.; Eschborn, M.; Ghelman, J.; Chasan, A.I.; Glander, S.; Schädlich, M.; Kuhlencord, M.; Daber, N.M.; et al. Nur77 serves as a molecular brake of the metabolic switch during T cell activation to restrict autoimmunity. Proc. Natl. Acad. Sci. 2018, 115, 201721049–E8026. [Google Scholar] [CrossRef]
- Moran, A.E.; Holzapfel, K.L.; Xing, Y.; Cunningham, N.R.; Maltzman, J.S.; Punt, J.; Hogquist, K.A. T cell receptor signal strength in Treg and iNKT cell development demonstrated by a novel fluorescent reporter mouse. J. Exp. Med. 2011, 208, 1279–1289. [Google Scholar] [CrossRef]
- Hiwa, R.; Nielsen, H.V.; Mueller, J.L.; Mandla, R.; Zikherman, J. NR4A family members regulate T cell tolerance to preserve immune homeostasis and suppress autoimmunity. J. Clin. Investig. 2021, 6. [Google Scholar] [CrossRef]
- Sekiya, T.; Kondo, T.; Shichita, T.; Morita, R.; Ichinose, H.; Yoshimura, A. Suppression of Th2 and Tfh immune reactions by Nr4a receptors in mature T reg cells. J. Exp. Med. 2015, 212, 1623–1640. [Google Scholar] [CrossRef]
- Yoshimura, A.; Ito, M.; Mise-Omata, S.; Ando, M. SOCS: negative regulators of cytokine signaling for immune tolerance. Int. Immunol. 2021, 33, 711–716. [Google Scholar] [CrossRef]
- Ashouri, J.F.; Weiss, A. Endogenous Nur77 Is a Specific Indicator of Antigen Receptor Signaling in Human T and B Cells. J. Immunol. 2017, 198, 657–668. [Google Scholar] [CrossRef]
- Bending, D.; Zikherman, J. Nr4a nuclear receptors: markers and modulators of antigen receptor signaling. Curr. Opin. Immunol. 2023, 81, 102285. [Google Scholar] [CrossRef]
- Park, T.-Y.; Jang, Y.; Kim, W.; Shin, J.; Toh, H.T.; Kim, C.-H.; Yoon, H.S.; Leblanc, P.; Kim, K.-S. Chloroquine modulates inflammatory autoimmune responses through Nurr1 in autoimmune diseases. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Won, H.Y.; Shin, J.H.; Oh, S.; Jeong, H.; Hwang, E.S. Enhanced CD25+Foxp3+ regulatory T cell development by amodiaquine through activation of nuclear receptor 4A. Sci. Rep. 2017, 7, 16946–16946. [Google Scholar] [CrossRef]
- Ogawa C, Tone Y, Tsuda M, et al. (2014) TGF-β–Mediated Foxp3 Gene Expression Is Cooperatively Regulated by Stat5, Creb, and AP-1 through CNS2. The Journal of Immunology 192:475–483. [CrossRef]
- Takahashi, H.; Tsuboi, H.; Asashima, H.; Hirota, T.; Kondo, Y.; Moriyama, M.; Matsumoto, I.; Nakamura, S.; Sumida, T. cDNA microarray analysis identifies NR4A2 as a novel molecule involved in the pathogenesis of Sjögren's syndrome. Clin. Exp. Immunol. 2017, 190, 96–109. [Google Scholar] [CrossRef]
- Zhang, W.; Cao, X.; Zhong, X.; Wu, H.; Feng, M.; Gwack, Y.; Isakov, N.; Sun, Z. Steroid nuclear receptor coactivator 2 controls immune tolerance by promoting induced T reg differentiation via up-regulating Nr4a2. Sci. Adv. 2022, 8, eabn7662. [Google Scholar] [CrossRef]
- Bending, D.; Martín, P.P.; Paduraru, A.; Ducker, C.; Marzaganov, E.; Laviron, M.; Kitano, S.; Miyachi, H.; Crompton, T.; Ono, M. A timer for analyzing temporally dynamic changes in transcription during differentiation in vivo. J. Cell Biol. 2018, 217, 2931–2950. [Google Scholar] [CrossRef]
- Jennings, E.; Elliot, T.A.; Thawait, N.; Kanabar, S.; Yam-Puc, J.C.; Ono, M.; Toellner, K.-M.; Wraith, D.C.; Anderson, G.; Bending, D. Nr4a1 and Nr4a3 Reporter Mice Are Differentially Sensitive to T Cell Receptor Signal Strength and Duration. Cell Rep. 2020, 33, 108328. [Google Scholar] [CrossRef]
- Scott-Browne, J.P.; López-Moyado, I.F.; Trifari, S.; Wong, V.; Chavez, L.; Rao, A.; Pereira, R.M. Dynamic Changes in Chromatin Accessibility Occur in CD8 + T Cells Responding to Viral Infection. Immunity 2016, 45, 1327–1340. [Google Scholar] [CrossRef]
- Kurd, N.S.; He, Z.; Louis, T.L.; Milner, J.J.; Omilusik, K.D.; Jin, W.; Tsai, M.S.; Widjaja, C.E.; Kanbar, J.N.; Olvera, J.G.; et al. Early precursors and molecular determinants of tissue-resident memory CD8 + T lymphocytes revealed by single-cell RNA sequencing. Sci. Immunol. 2020, 5. [Google Scholar] [CrossRef]
- Odagiu L, Boulet S, de Sousa DM, et al. (2020) Early programming of CD8+ T cell response by the orphan nuclear receptor NR4A3. Proc Natl Acad Sci U S A 117:24392–24402. [CrossRef]
- Nowyhed, H.N.; Huynh, T.R.; Thomas, G.D.; Blatchley, A.; Hedrick, C.C. Cutting Edge: The Orphan Nuclear Receptor Nr4a1 Regulates CD8+ T Cell Expansion and Effector Function through Direct Repression of Irf4. J. Immunol. 2015, 195, 3515–3519. [Google Scholar] [CrossRef]
- Liu, X.; Wang, Y.; Lu, H.; Li, J.; Yan, X.; Xiao, M.; Hao, J.; Alekseev, A.; Khong, H.; Chen, T.; et al. Genome-wide analysis identifies NR4A1 as a key mediator of T cell dysfunction. Nature 2019, 567, 525–529. [Google Scholar] [CrossRef]
- Chen, J.; López-Moyado, I.F.; Seo, H.; Lio, C.-W.J.; Hempleman, L.J.; Sekiya, T.; Yoshimura, A.; Scott-Browne, J.P.; Rao, A. NR4A transcription factors limit CAR T cell function in solid tumours. Nature 2019, 567, 530–534. [Google Scholar] [CrossRef]
- Lith SC, van Os BW, Seijkens TTP, de Vries CJM (2020) ‘Nur’turing tumor T cell tolerance and exhaustion: novel function for Nuclear Receptor Nur77 in immunity. Eur J Immunol 50:1643–1652.
- Kleberg, J.; Nataraj, A.; Xiao, Y.; Podder, B.R.; Jin, Z.; Tithi, T.I.; Zheng, G.; Smalley, K.S.M.; Moser, E.K.; Safe, S.; et al. Targeting Lineage-Specific Functions of NR4A1 for Cancer Immunotherapy. Int. J. Mol. Sci. 2025, 26, 5266. [Google Scholar] [CrossRef]
- Srirat, T.; Hayakawa, T.; Mise-Omata, S.; Nakagawara, K.; Ando, M.; Shichino, S.; Ito, M.; Yoshimura, A. NR4a1/2 deletion promotes accumulation of TCF1+ stem-like precursors of exhausted CD8+ T cells in the tumor microenvironment. Cell Rep. 2024, 43, 113898. [Google Scholar] [CrossRef]
- Brooks, J.F.; Tan, C.; Mueller, J.L.; Hibiya, K.; Hiwa, R.; Vykunta, V.; Zikherman, J. Negative feedback by NUR77/Nr4a1 restrains B cell clonal dominance during early T-dependent immune responses. Cell Rep. 2021, 36, 109645–109645. [Google Scholar] [CrossRef]
- Tan, C.; Hiwa, R.; Mueller, J.L.; Vykunta, V.; Hibiya, K.; Noviski, M.; Huizar, J.; Brooks, J.F.; Garcia, J.; Heyn, C.; et al. NR4A nuclear receptors restrain B cell responses to antigen when second signals are absent or limiting. Nat. Immunol. 2020, 21, 1267–1279. [Google Scholar] [CrossRef]
- Kumar, A.; Hill, T.M.; Gordy, L.E.; Suryadevara, N.; Wu, L.; Flyak, A.I.; Bezbradica, J.S.; Van Kaer, L.; Joyce, S. Nur77 controls tolerance induction, terminal differentiation, and effector functions in semi-invariant natural killer T cells. Proc. Natl. Acad. Sci. 2020, 117, 17156–17165. [Google Scholar] [CrossRef]
- Chen, L.; Fan, F.; Wu, L.; Zhao, Y. The nuclear receptor 4A family members: mediators in human disease and autophagy. Cell. Mol. Biol. Lett. 2020, 25, 1–15. [Google Scholar] [CrossRef]
- Safe, S.; Jin, U.-H.; Morpurgo, B.; Abudayyeh, A.; Singh, M.; Tjalkens, R.B. Nuclear receptor 4A (NR4A) family – orphans no more. J. Steroid Biochem. Mol. Biol. 2016, 157, 48–60. [Google Scholar] [CrossRef]
- Doi, Y.; Oki, S.; Ozawa, T.; Hohjoh, H.; Miyake, S.; Yamamura, T. Orphan nuclear receptor NR4A2 expressed in T cells from multiple sclerosis mediates production of inflammatory cytokines. Proc. Natl. Acad. Sci. 2008, 105, 8381–8386. [Google Scholar] [CrossRef]
- Wang, L.-M.; Zhang, Y.; Li, X.; Zhang, M.-L.; Zhu, L.; Zhang, G.-X.; Xu, Y.-M. Nr4a1 plays a crucial modulatory role in Th1/Th17 cell responses and CNS autoimmunity. Brain, Behav. Immun. 2018, 68, 44–55. [Google Scholar] [CrossRef]
- Deutsch, A.J.; Rinner, B.; Pichler, M.; Prochazka, K.; Pansy, K.; Bischof, M.; Fechter, K.; Hatzl, S.; Feichtinger, J.; Wenzl, K.; et al. NR4A3 Suppresses Lymphomagenesis through Induction of Proapoptotic Genes. Cancer Res. 2017, 77, 2375–2386. [Google Scholar] [CrossRef]
- Fechter K, Feichtinger J, Prochazka K, et al. (2018) Cytoplasmic location of NR4A1 in aggressive lymphomas is associated with a favourable cancer specific survival. Sci Rep 8. [CrossRef]
- Li, Q.-X.; Ke, N.; Sundaram, R.; Wong-Staal, F. NR4A1, 2, 3 – an orphan nuclear hormone receptor family involved in cell apoptosis and carcinogenesis. 2006, 21, 533–540. [Google Scholar] [CrossRef]
- Wang JR, Gan WJ, Li XM, et al. (2014) Orphan nuclear receptor Nur77 promotes colorectal cancer invasion and metastasis by regulating MMP-9 and E-cadherin. Carcinogenesis 35:2474–2484. [CrossRef]
- Zhang, C.; Zhang, B.; Zhang, X.; Sun, G.; Sun, X. Targeting Orphan Nuclear Receptors NR4As for Energy Homeostasis and Diabetes. Front. Pharmacol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Safe, S.; Shrestha, R.; Mohankumar, K. Orphan nuclear receptor 4A1 (NR4A1) and novel ligands. Essays Biochem. 2021, 65, 877–886. [Google Scholar] [CrossRef]
- Lee, M.; Upadhyay, S.; Mariyam, F.; Martin, G.; Hailemariam, A.; Lee, K.; Jayaraman, A.; Chapkin, R.S.; Lee, S.-O.; Safe, S. Flavone and Hydroxyflavones Are Ligands That Bind the Orphan Nuclear Receptor 4A1 (NR4A1). Int. J. Mol. Sci. 2023, 24, 8152. [Google Scholar] [CrossRef]
- Hu, M.; Luo, Q.; Alitongbieke, G.; Chong, S.; Xu, C.; Xie, L.; Chen, X.; Zhang, D.; Zhou, Y.; Wang, Z.; et al. Celastrol-Induced Nur77 Interaction with TRAF2 Alleviates Inflammation by Promoting Mitochondrial Ubiquitination and Autophagy. Mol. Cell 2017, 66, 141–153.e6. [Google Scholar] [CrossRef] [PubMed]
- Safe, S. Natural products and synthetic analogs as selective orphan nuclear receptor 4A (NR4A) modulators. Histol Histopathol. 2024, 39, 543–556. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Regulatory roles of NR4A receptors in macrophages and DCs. Macrophages and DCs utilize NR4A receptors (NR4A1, NR4A2, NR4A3) to fine-tune immune responses. In macrophages, NR4A1 suppresses NF-κB activity (except for TNF-α) and reprograms metabolism (e.g., via SDH and TCA cycle) to restrain inflammation, while promoting M2-like polarization. Furthermore, NR4A2 inhibits TLR4-induced inflammation via SUMOylation-mediated NF-κB/p65 displacement and enhances M2 polarization, while NR4A3 promotes monocyte adhesion in atherosclerosis and modulates M1/M2 marker expression (e.g., suppresses IL-10 in M2 macrophages). In DCs, NR4A1 limits hyperinflammatory cytokine production (IL-6, TNFα) and T-cell stimulation, with NR4A2 driving a tolerogenic DC differentiation and Treg expansion. By its part, NR4A3 facilitates CCR7-dependent DC migration to lymph nodes and monocyte-to-DC differentiation. Figure created with BioRender.com and Servier Medical Art.
Figure 1.
Regulatory roles of NR4A receptors in macrophages and DCs. Macrophages and DCs utilize NR4A receptors (NR4A1, NR4A2, NR4A3) to fine-tune immune responses. In macrophages, NR4A1 suppresses NF-κB activity (except for TNF-α) and reprograms metabolism (e.g., via SDH and TCA cycle) to restrain inflammation, while promoting M2-like polarization. Furthermore, NR4A2 inhibits TLR4-induced inflammation via SUMOylation-mediated NF-κB/p65 displacement and enhances M2 polarization, while NR4A3 promotes monocyte adhesion in atherosclerosis and modulates M1/M2 marker expression (e.g., suppresses IL-10 in M2 macrophages). In DCs, NR4A1 limits hyperinflammatory cytokine production (IL-6, TNFα) and T-cell stimulation, with NR4A2 driving a tolerogenic DC differentiation and Treg expansion. By its part, NR4A3 facilitates CCR7-dependent DC migration to lymph nodes and monocyte-to-DC differentiation. Figure created with BioRender.com and Servier Medical Art.
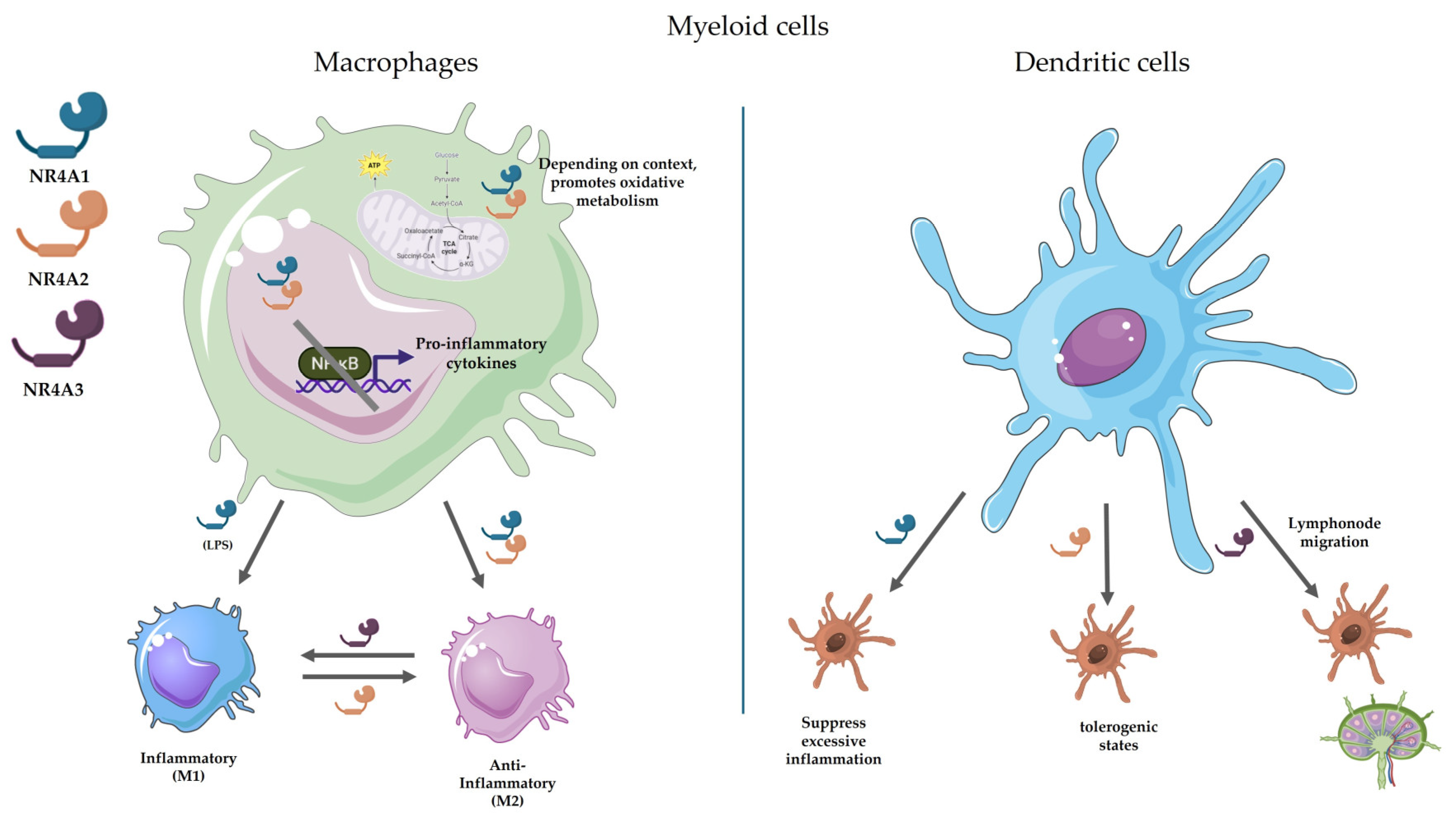
Figure 2.
Roles of NR4A receptors in CD4+ and CD8+ T cell biology. The figure summarizes the involvement of NR4A family members in key T cell processes. CD4+ T cells (right): NR4A1 serves as a master regulator that ssuppresses effector T cells (inhibits Th1/Th17 differentiation and IL-2/IFN-γ production via AP-1 competition), while Promotes Treg development. Besides, constrains metabolic reprogramming (limits glycolysis/OXPHOS to prevent hyperactivation) and mediates thymic negative. Regarding NR4A2, it exbibits context-dependent roles, supporting Treg stability through Foxp3 enhancer regulation or paradoxically enabling Th17 maturation. With respect to NR4A3, it has redundant apoptotic functions with NR4A1 in thymic selection, with a strong correlation with high-affinity TCR signaling. Furthermore, it contributes to Treg lineage commitment (persistent expression in peripheral Tregs).CD8+ T cells (left). NR4A1 exhibits dual roles: (1) Promotes initial T cell activation (correlating with TCR signal strength) while establishing checkpoint control via IRF4 suppression; (2) Drives exhaustion (via PD-1/TIM-3 upregulation) and anergy under chronic stimulation. NR4A2 cooperates with NR4A1 in exhaustion and sustains CD28+ tissue-resident memory T (TRM) cells. NR4A3 regulates thymic negative selection (via strong TCR signals) and effector-memory balance by restricting bZIP transcription factor access to chromatin. Figure created with BioRender.com and Servier Medical Art.
Figure 2.
Roles of NR4A receptors in CD4+ and CD8+ T cell biology. The figure summarizes the involvement of NR4A family members in key T cell processes. CD4+ T cells (right): NR4A1 serves as a master regulator that ssuppresses effector T cells (inhibits Th1/Th17 differentiation and IL-2/IFN-γ production via AP-1 competition), while Promotes Treg development. Besides, constrains metabolic reprogramming (limits glycolysis/OXPHOS to prevent hyperactivation) and mediates thymic negative. Regarding NR4A2, it exbibits context-dependent roles, supporting Treg stability through Foxp3 enhancer regulation or paradoxically enabling Th17 maturation. With respect to NR4A3, it has redundant apoptotic functions with NR4A1 in thymic selection, with a strong correlation with high-affinity TCR signaling. Furthermore, it contributes to Treg lineage commitment (persistent expression in peripheral Tregs).CD8+ T cells (left). NR4A1 exhibits dual roles: (1) Promotes initial T cell activation (correlating with TCR signal strength) while establishing checkpoint control via IRF4 suppression; (2) Drives exhaustion (via PD-1/TIM-3 upregulation) and anergy under chronic stimulation. NR4A2 cooperates with NR4A1 in exhaustion and sustains CD28+ tissue-resident memory T (TRM) cells. NR4A3 regulates thymic negative selection (via strong TCR signals) and effector-memory balance by restricting bZIP transcription factor access to chromatin. Figure created with BioRender.com and Servier Medical Art.
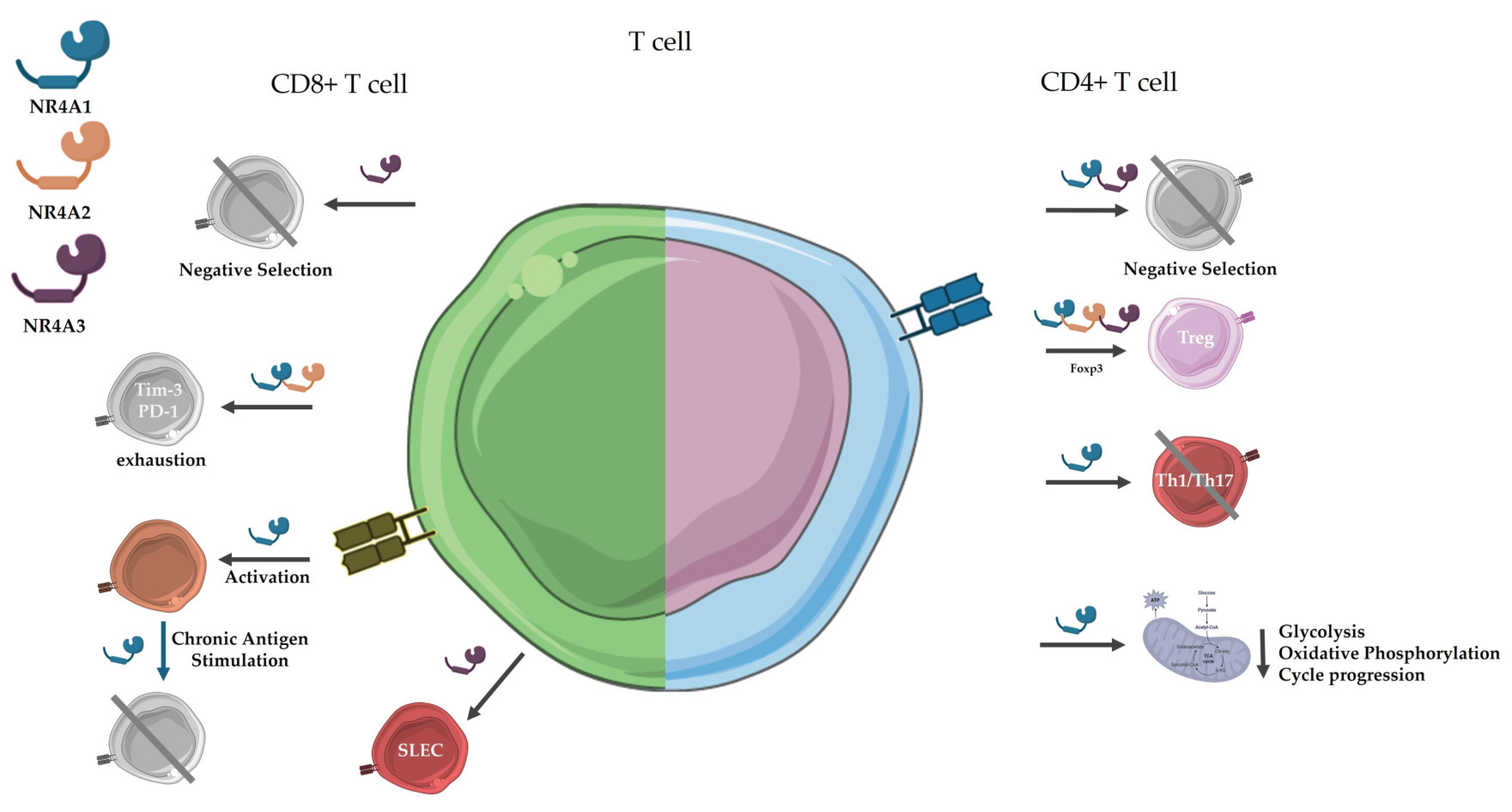

Table 1.
Overview of the NR4A Nuclear Receptor Subfamily in homeostasis and pathology.
| Feature | NR4A1 (Nur77) | NR4A2 (Nurr1) | NR4A3 (NOR1) |
|---|---|---|---|
| Gene Symbol / Synonyms | NR4A1 / Nur77, TR3, NGFI-B | NR4A2 / Nurr1, NOT, RNR1 | NR4A3 / NOR1, TEC, MINOR, CHN |
| Expression Type | Immediate early gene | Immediate early gene | Immediate early gene |
| DNA Binding | Monomer (NBRE), homodimer or heterodimer (NurRE), RXR dimerization | Monomer (NBRE), homodimer or heterodimer (NurRE), RXR dimerization | Monomer (NBRE); low affinity for NurRE; does not dimerize with RXR |
| Ligand Binding Domain (LBD) | Atypical, constitutively active; binds synthetic ligands (e.g., CsnB) | Atypical but dynamic; binds DHA, AEA, and synthetic molecules | Atypical; potential interaction with unsaturated fatty acids and prostaglandins |
| Tissue Expression | Broad (thymus, spleen, liver, brain, immune cells) | CNS (midbrain dopaminergic neurons), cartilage, immune tissues | Heart, skeletal muscle, immune cells, CNS |
| Canonical Functions | Apoptosis regulation, T cell development, inflammation modulation | Dopaminergic neuron maintenance, anti-inflammatory roles, immune regulation | Vascular remodeling, metabolic regulation, immune homeostasis |
| Role in Immune Response | Suppresses NF-κB signaling, regulates T cell activation and macrophage polarization | Restricts DC immunogenicity, promotes anti-inflammatory macrophage phenotypes | Modulates DC migration, neutrophil survival, anti-inflammatory effects in monocytes/macrophages |
| Neurological Role | Neuroprotective; expressed in cortex and hippocampus | Essential for dopaminergic neuron development; mutations linked to Parkinson’s disease | Implicated in hippocampal development, inner ear formation, depressive behavior |
| Cardiovascular Involvement | Attenuates vascular inflammation, promotes endothelial homeostasis | Limited but protective role in atherosclerosis | Regulates VSMC proliferation, modulates atherosclerosis progression, promotes cardiac hypertrophy |
| Cancer-Related Functions | Dual role: tumor suppressor or promoter depending on context; modulates immune microenvironment | Tumor suppressor; may inhibit angiogenesis and inflammatory gene expression | Tumor suppressor in AML; involved in oncogenic fusion proteins (e.g., EWS–NR4A3 in sarcomas) |
| Metabolic Regulation | Modulates glucose and lipid metabolism, mitochondrial function | Regulates insulin gene expression and β-cell function | Controls lipid/glucose homeostasis in skeletal muscle, insulin secretion |
| Modulators / Ligands | Cytosporone B, 6-mercaptopurine, PDNPA | Anandamide (AEA), DHA, CsnB, prostaglandins | Arachidonic acid, PGA1/PGA2, synthetic fatty acids |
| Therapeutic Potential | Immunotherapy, cancer, inflammation, cardiovascular disease | Parkinson’s disease, rheumatoid arthritis, sepsis, inflammatory disorders | Cardiovascular disease, AML, metabolic syndrome, neurodegeneration |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



