
Submitted:
04 June 2025
Posted:
06 June 2025
You are already at the latest version
Abstract
Background: Vitamin D binding protein (VDBP) is a precursor to macrophage activating factor (MAF) and regulates and controls innate immunity. Metadichol , a new nanoemulsion formulation that acts as an inverse agonist of the vitamin D receptor (VDR), was tested for its immunomodulatory effects on VDBP release in differentiated human monocytic cell lines Methods: THP-1 and U937 monocytic cell lines were differentiated with PMA (20 ng/ml) and stimulated with LPS. Various Metadichol concentrations (1 pg/ml to 100 ng/ml) were applied to cells for 48 hours. ELISA measured VDBP levels. Lipopolysaccharide stimulation significantly elevated VDBP release in both cell lines (10.19-fold in THP-1 cells: 117.01 ± 2.27 ng/ml; 12.03-fold in U937 cells: 222.56 ± 3.19 ng/ml vs. controls). Metadichol therapy altered VDBP release in a dose-dependent manner. Metadichol , at 100 ng/ml, significantly increased VDBP release (7.12-fold increase in THP-1: 81.73 ± 2.37 ng/ml; 8.36-fold increase in U937: 154.68 ± 3.19 ng/ml) while reducing LPS-induced inflammation. Metadichol shows dose-dependent immunomodulatory effects on VDBP release in human monocytic cells via VDR inverse agonism, outperforming traditional small molecule immunomodulators in immunological activation. The compound's VDR pathway modulation of VDBP levels without surpassing inflammatory thresholds suggests balanced immune response activation with potential therapeutic uses in immunological deficiency, cancer immunotherapy, and age-related immune decline.
Keywords:
Metadichol
; VDR inverse agonist
; monocytes
; immunomodulation
; macrophage activating factor
; nanoemulsion
; immunological enhancement
; Vitamin D binding protein
1. Introduction
1.1. Vitamin D Binding Protein: Beyond Transport
Vitamin D binding protein (VDBP), also known as
Gc-globulin, is a highly abundant and versatile protein found in human plasma,
with values of 200-500 μg/ml in healthy individuals [1–3].
VDBP, formerly a vitamin D metabolite carrier, is now essential to innate
immunity, inflammatory control, and tissue repair [4,5].
The protein has three primary phenotypic variants (Gc1f, Gc1s, and Gc2) with
different functional features and population distributions, affecting illness
susceptibility [6,7].
VDBP’s biological significance goes beyond vitamin
D transport. VDBP is the precursor to macrophage activating factor (MAF), an
immunomodulatory protein that boosts macrophage activation, phagocytosis, and
anticancer activity [8–10]. Activated B and T
cells produce β-galactosidase and neuraminidase, which convert VDBP to MAF [11,12]. This conversion is essential for innate
immune response activation and cancer immunosurveillance, pathogen clearance,
and tissue homeostasis [13,14].
1.2. VDR-Mediated VDBP Regulation: Complex Network
The vitamin D receptor (VDR) controls VDBP
production and metabolism through complicated transcriptional and
post-transcriptional processes [15,16]. New
evidence suggests that VDR can serve as a constitutively active receptor that
needs inverse agonists to regulate [17,18].
This paradigm shift has major ramifications for VDBP control and potential
therapeutic methods.
VDR controls VDBP expression in multiple ways. VDR
complexes with RXR and binds to vitamin D response elements (VDREs) in the VDBP
gene promoter in hepatocytes, the main source of VDBP [19,20]. Paradoxically, vitamin D metabolites can
trigger some VDBP expression, but excessive VDR activation frequently inhibits
feedback and reduces VDBP synthesis [21,22].
VDR regulates VDBP differently in monocytes and
macrophages than hepatocytes. These immune cells express VDR constitutively and
generate VDBP locally in response to stimuli [23,24].
Importantly, VDR basal activity in these cells suppresses VDBP creation,
suggesting that VDR inverse agonism could boost local VDBP synthesis and immune
function [25,26].
VDR inverse agonism is a unique treatment. In
contrast to VDR antagonists, inverse agonists actively diminish VDR
constitutive activity, which may increase VDBP synthesis by
derepressing blocked pathways [27,28]. This may explain why immune-compromised
people’s VDBP levels don’t improve with vitamin D administration [29,30].
1.3. Disease-related VDBP and VDR Dysfunction
The VDR-VDBP axis is a diagnostic and therapeutic
target for many clinical disorders due to its dysregulation. Multiple processes
cause VDBP depletion in cancer patients due to aberrant VDR signaling [31,32]. Overexpression of VDR and sequestration of
ligands by tumor cells causes immune cells to upregulate VDR activity and
inhibit VDBP [33,34]. Multiple tumor types
show that VDR-mediated VDBP depletion promotes cancer growth, metastasis, and
poor prognosis [35–37].
Chronic inflammatory diseases show VDR-VDBP
dysregulation. In rheumatoid arthritis, SLE, and IBD, chronic inflammatory
signals activate VDR and consume VDBP [38–40].
The vicious circle of VDBP depletion impairing immunological function increases
infection susceptibility and inflammatory stress [41,42].
Progression of VDR sensitivity and VDBP metabolism
occurs with age. Constitutive VDR activity reduces VDBP production in elderly
people due to increased VDR expression but decreased endogenous ligand
responsiveness [43,44]. Age-related VDR
deficiency causes immunosenescence and increases infection and cancer risk [45,46].
Infectious disorders greatly affect VDR-VDBP
regulation. As part of the innate immune response, influenza, hepatitis, and
COVID-19 can upregulate VDR, while excessive VDR activity can deplete VDBP [47–49]. Through LPS-induced inflammatory cascades,
gram-negative bacterial infections can rapidly dysregulate the VDR-VDBP axis [50,51].
1.4. Limitations of Current VDR-VDBP Modulation Methods
Traditional methods to enhance VDBP have focused on
vitamin D supplementation, assuming it boosts VDBP production [52,53]. This strategy has limited efficacy and
potentially detrimental results because excessive VDR activation suppresses
VDBP synthesis through negative feedback [54,55].
This contradictory association may explain why vitamin D supplementation fails
to improve immunological responses in clinical trials [56,57].
Alternative treatments include VDR antagonists,
however these drugs have poor pharmacokinetics and can completely block
beneficial VDR actions [58,59]. Pure VDR
antagonism may disrupt calcium homeostasis and bone metabolism [60,61].
Clinical trials of direct VDBP or MAF
supplementation have been limited by protein stability, immunogenicity, and
regulatory difficulties [62,63]. The various
glycosylation patterns needed for MAF action make synthetic synthesis difficult
and expensive [64,65].
1.5. Small Molecule Immunomodulators: VDR-Independent Ideas
Most traditional small molecule immunomodulators
work through routes independent of the VDR-VDBP axis, which may explain their
low efficacy in VDBP-depleted settings. Levamisole, an anthelmintic, had mild
immune-boosting effects but serious adverse effects and did not treat VDBP
deficiency [66,67].
As a toll-like receptor 7 agonist, imiquimod can
boost immune function, but inflammatory side effects and no direct influence on
the VDR-VDBP regulation system limit its effects [68,69].
In immunocompromised situations, CpG oligodeoxynucleotides activate innate
immunity through TLR9 signaling but do not address VDBP deficiency [70,71].
Type I and II interferons influence immune function
but escalate VDR-VDBP dysregulation through inflammatory pathways that enhance
VDR expression and activity [72,73]. This may
cause immunological suppression and depression with interferon therapy [74,75].
Plant-derived immunomodulators and beta-glucans
exhibit minor immune-enhancing effects but lack VDR-VDBP pathway specificity
and standardization concerns [76–78].
VDR-mediated VDBP suppression, which characterizes many immune-deficient
diseases, is not addressed by their mechanisms [79,80].
1.6. Metadichol A New VDR Inverse Agonist
Metadichol [81–83]
a nanoemulsion of long chain alcohols C26, C28, C30 of which C28 is at least
85% is a VDR inverse agonist. Metadichol unlike other VDR ligands [85] regulatesVDBP production routes by reducing
constitutive VDR activity.
Metadichol’s inverse agonist characteristics [83] set it apart from VDR agonists (e.g.,
calcitriol) and antagonist in contrast to agonists and antagonists, inverse
agonists actively lower constitutively active receptors’ basal transcriptional
activity [86,87]. This process is important
for VDR, which has constitutive activity in many cell types, including immune
cells [88,89].
Metadichol nanoemulsion formulation improves
bioavailability and cellular uptake, overcoming hydrophobic chemical
constraints in reaching intracellular targets such VDR [90,91]. This improved delivery method may modulate
VDR activity better at lower doses than small molecules [92–94]. Metadichol modulates immunological
function, including cytokine generation, immune cell activation, and
antioxidant responses, through the VDR pathway [81–83,94].
1.7. Study Purposes
This study examined the impact of Metadichol on
VDBP release in human monocytic cell lines [95–98]
focusing on its VDR inverse agonist mechanism. The objective was to establish
dose-response relationships for Metadichol -induced VDBP release via VDR
modulation, compare its efficacy to standard inflammatory stimuli, and evaluate
its potential advantages over conventional immunomodulatory approaches that do
not target the VDR-VDBP axis.[99]
2. Materials and Methods
2.1. Experiment statement. All experimental work was designed by author was outsourced to a commercial service provider Skanda biolabs in Bangalore, India.
THP-1 (TIB-202, ATCC, USA) and U937 (CRL-1593.2,
ATCC, USA) human monocytic cell lines were procured from ATCC and maintained
according to standard methods.
Cells were grown in RPMI-1640 media (Gibco,
1898961) with 10% heat-inactivated fetal bovine serum, 50 μM 2-mercaptoethanol,
2 mM L-glutamine, and 100 IU/ml penicillin-streptomycin at 37°C in a humidified
environment with 5% CO₂. Trypan blue exclusion determined cell viability, and
only cultures with >95% were employed for experiments.
2.2. Protocol for Cell Differentiation
Phorbol 12-myristate 13-acetate (PMA, Sigma P8139),
a well-established protein kinase C activator, increased monocyte-to-macrophage
differentiation and upregulated VDR expression [100].
Harvested 80% confluent cells were centrifuged at 1,500 rpm for 5 minutes,
washed twice with PBS, and resuspended in full RPMI media.
Attachment was achieved by adjusting cell density
to 5 × 10⁵ cells/well in Falcon (353108) 6-well plates for 24 hours.
Differentiation was confirmed by morphological alterations, adhesion to culture
plates, and enhanced VDR expression after 4 hours of PMA (20 ng/ml) [97,98]. Optimization experiments found that this
dose and time maximized differentiation with minimal toxicity and ensured VDR
expression for inverse agonist investigations [101,102].
2.3. Treatment Conditions
After differentiation, cells were treated with
Metadichol [1 pg/ml, 100 pg/ml, 1 ng/ml, 100 ng/ml) after serial dilution from
a 5 mg/ml stock solution. Based on preliminary dose-finding tests showing VDR
inverse agonist action, these doses are physiologically suitable for
nanoemulsion formulations targeting nuclear receptors [103–106].
Treatment lasted 48 hours to allow VDR-mediated
transcriptional modifications to alter VDBP synthesis and release without
deleterious effects. LPS (Sigma L2630, 20 ng/ml) was used as a positive control
for VDBP activation via VDR-independent inflammatory pathways [107,108]. Untreated differentiated cells were used
as negative controls for VDR and VDBP synthesis After treatment, cell culture
supernatants were centrifuged at 10,000 rpm for 10 minutes to remove cellular
debris. To retain VDBP integrity and limit protein degradation, cell pellets
were lysed in sterile phosphate buffer (pH 7.4) with protease inhibitors. To
preserve protein, supernatant and lysate samples were kept at -80°C until VDBP
measurement.
2.4. ELISA VDBP Quantification
A commercial human VDBP ELISA kit , E-EL-H1604,
Elabscience ( www.elabscience.com) was used to measure VDBP levels with high
specificity and low cross-reactivity with VDR and vitamin D metabolites. The
test technique was refined to detect VDBP from VDR inverse agonist mechanisms.
Standard curves were made using repeated two-fold
dilutions from 125 to 3.91 ng/ml. Samples and standards (100 μl/well) were
incubated at 37°C for 90 minutes on pre-coated plates. After washing with wash
buffer, add 100 μl of biotinylated detection antibody and incubate at 37°C for
60 minutes. For 30 minutes at 37°C, HRP conjugate solution (100 μl) was added
after additional washing processes.
Color development was achieved by incubating
substrate reagent (90 μl) at 37°C for 15 minutes, followed by stop solution (50
μl). Within 15 minutes following stop solution addition, a Bio-Rad microplate
reader assessed absorbance at 450 nm. The coefficient of variation between
duplicate samples had to be <10%.
Data are shown as mean ± SD from duplicate
experiments. To account for baseline VDR activity effects on VDBP production,
fold changes were measured relative to untreated control cells for each
experiment. Standard curve validation was done using linear regression
analysis, with R² > 0.99 as the acceptance condition.
Non-linear regression was used to examine
dose-response relationships for inverse agonist mechanisms. Statistical
significance for VDR modulation experiments was determined using appropriate
methods, with p < 0.05 considered significant. Standard statistical software
was used for all calculations, including numerous comparison corrections.
3. Results
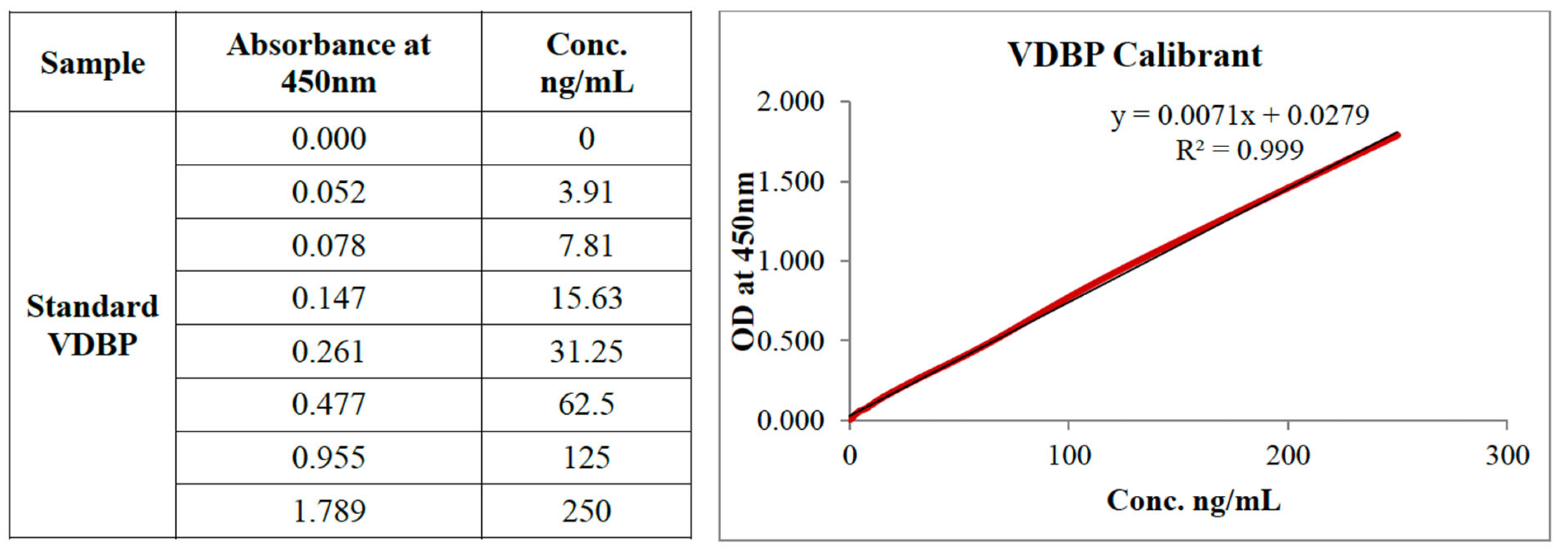
3.1. ELISA Validation and Performance
The VDBP ELISA performed well across the studied
range, especially for VDR inverse agonist-produced VDBP (Figure 1). The standard curve exhibited high
linearity (R² = 0.999) using the regression equation y = 0.0071x + 0.0279,
where y: 450 nm optical density and x: VDBP concentration in ng/ml The test
showed a detection range of 3.91-250 ng/ml and a sensitivity of <2 ng/ml.
Intra-assay variation was 4.2% and inter-assay
variation was 6.8%, both acceptable for quantitative immunoassays assessing
VDR-regulated proteins. Spiked samples recovered 95-105% in the working range,
confirming VDBP test accuracy regardless of production process. Cell culture
medium components, VDR modulators, and therapy agents did not interfere.
Figure 1.
Standard Curve VDBP.
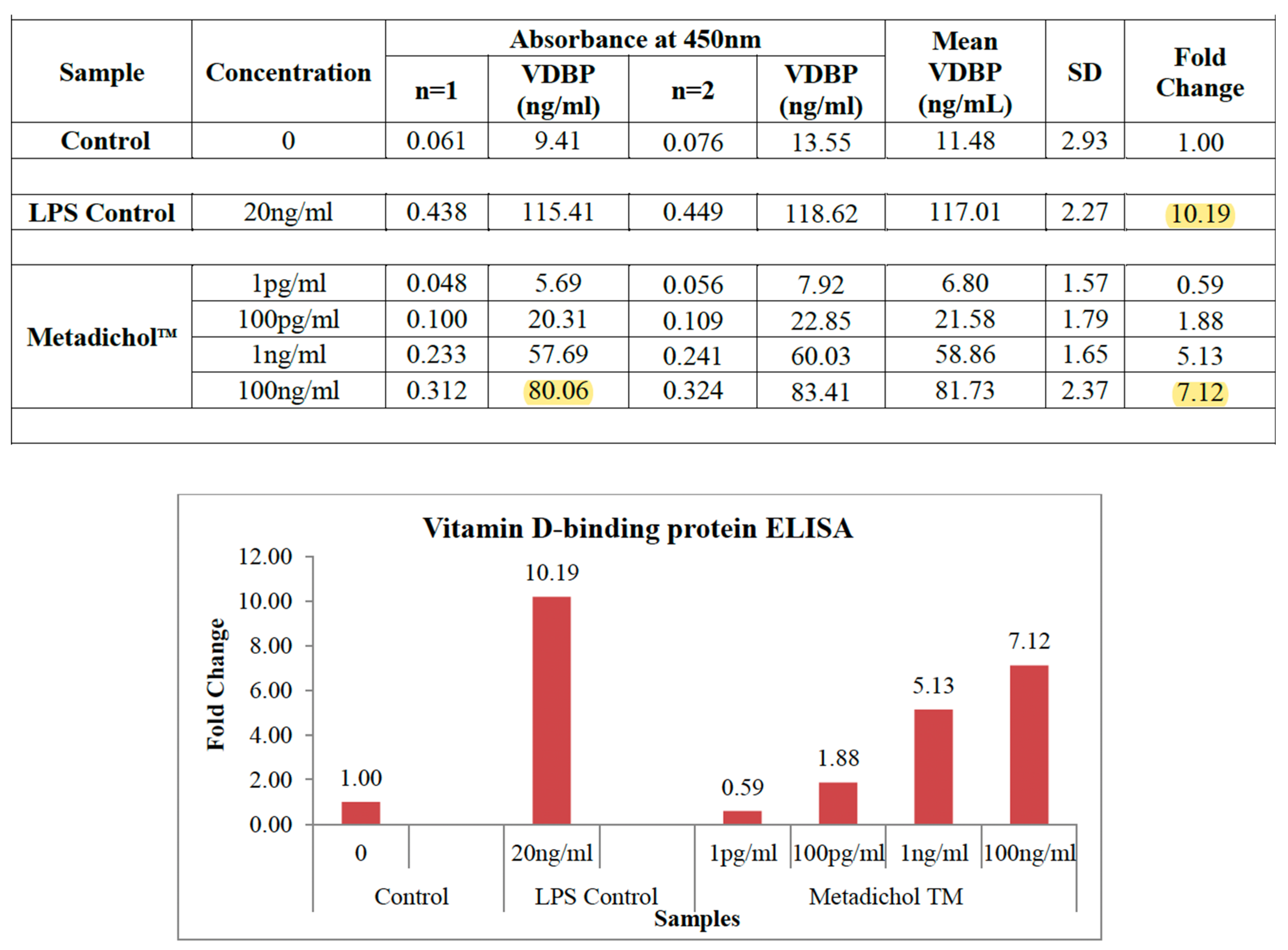
3.2. Monocytic Cell Line Baseline VDBP and VDR Expression
Figure 2.
The two cell lines had different VDR expression
profiles and constitutive receptor activity, affecting baseline VDBP levels.
THP-1 cells produced 11.48 ± 2.93 ng/ml VDBP under control conditions, while
U937 cells produced 18.51 ± 1.83 ng/ml. This 1.6-fold difference is consistent
with these cell lines’ different metabolic profiles and VDR expression levels [109–111].
3.2. U 937 Cell Line Baseline VDBP and VDR Expression
Figure 3.
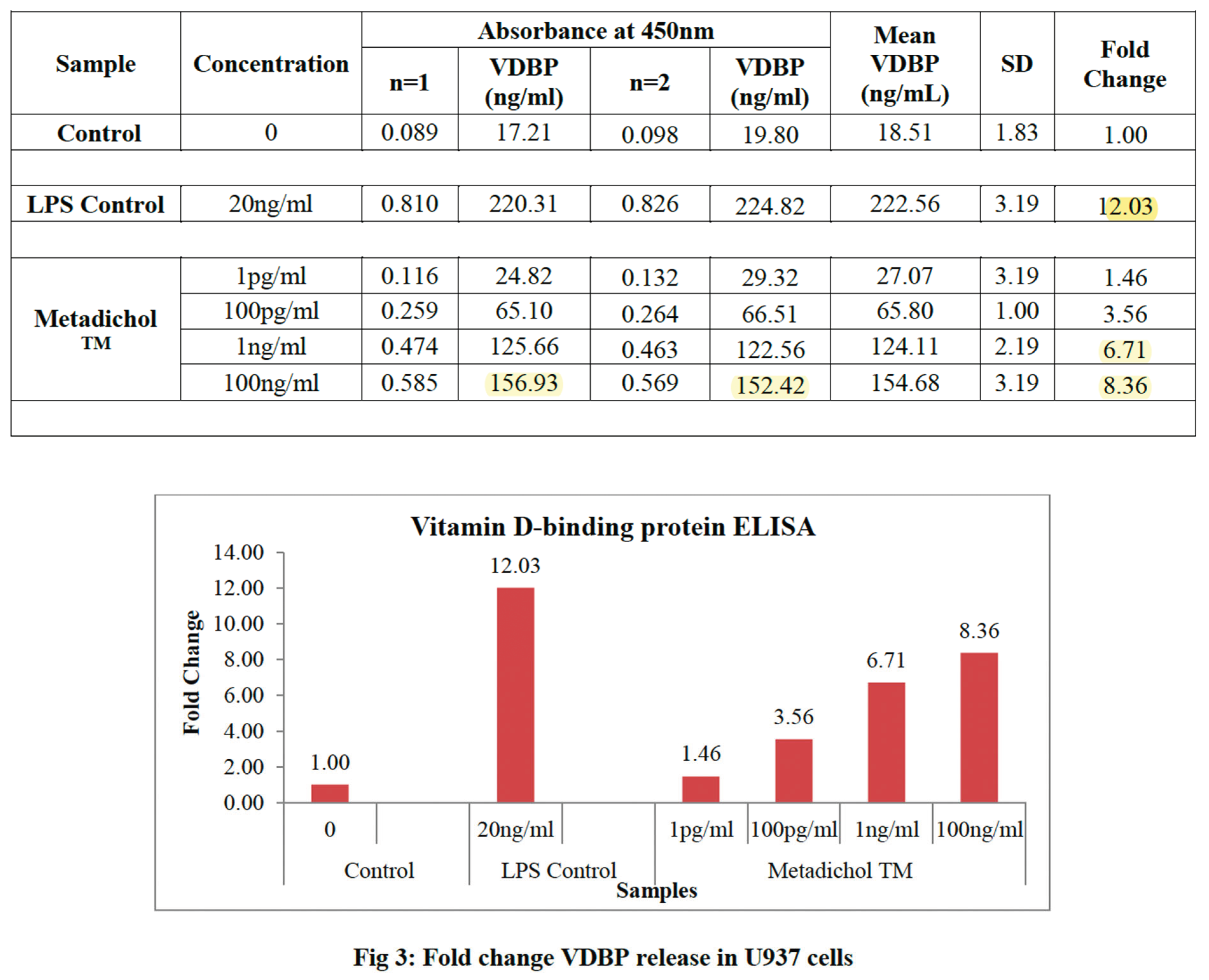
U937 cells may have higher basal VDBP synthesis than THP-1 cells due to their lower constitutive VDR activity. This pattern matches the negative association between VDR activity and VDBP synthesis in numerous cell types [112]. Both cell lines produced steady VDBP over 48 hours, proving that this timescale is suitable for VDR regulation experiments.
3.3. LPS-Induced VDBP Release via VDR-Independent Pathways
As a positive control, LPS (20 ng/ml) induced VDBP
through inflammatory pathways without VDR. LPS treatment strongly increased
VDBP release in both cell lines: 10.19-fold increase in THP-1 cells (117.01 ±
2.27 ng/ml, p < 0.001) and 12.03-fold increase in U937 cells (222.56 ± 3.19
ng/ml, p < 0.001).
Inflammatory signaling pathways bypass normal VDR
regulation mechanisms to release LPS-induced VDBP, setting a benchmark for
maximal VDBP production in these cell lines [113,114].
variable cell lines respond differently to inflammatory stimuli and have
variable capacities for acute protein production under stress [115,116].
3.4. THP-1 Cell VDR Inverse Agonism Response
Metadichol administration showed dose-dependent
VDBP release in THP-1 cells, confirming VDR inverse agonist mechanisms. The
lowest concentration (1 pg/ml) resulted in a 0.59-fold increase in VDBP levels
compared to control, with a minor reduction that may be due to partial VDR
inverse agonism.
3.5. Progressive Dose Escalation Increased VDBP Production via VDR Inverse Agonist Activity
100 pg/ml: 21.58 ± 1.79 ng/ml (1.88-fold increase,
p < 0.05).
1 ng/ml: 58.86 ± 1.65 (5.13-fold increase, p <
0.01).
100 ng/ml: 81.73 ± 2.37 (7.12-fold increase, p <
0.001).
Nuclear receptor inverse agonist interactions have
a sigmoidal dose-response relationship, suggesting selective binding to VDR
with increasing depletion of constitutive receptor function [117,118].
3.6. U937 Cell VDR Inverse Agonism Response
U937 cells had comparable dose-dependent patterns
with greater absolute VDBP levels, consistent with their lower baseline VDR
activity:
1 pg/ml: 27.07 ± 3.19 ng/ml (1.46-fold increase/p
< 0.05).
100 pg/ml: 65.80 ± 1.00 ng/ml [3.56-fold increase,
p < 0.01).
1 ng/ml: 124.11 ± 2.19 (6.71-fold increase, p <
0.001).
100 ng/ml: 154.68 ± 3.19 (8.36-fold increase, p
< 0.001).
U937 cells displayed VDBP increase even at the
lowest concentration, suggesting enhanced susceptibility to VDR inverse agonism
due to their distinct VDR expression profile and constitutive activity levels [112,113].
3.7. VDR Inverse Agonism vs. Inflammatory Stimulation Efficacy Comparison
Metadichol reached 70% of LPS-induced VDBP levels
in THP-1 cells and 69% in U937 cells at the maximum concentration (100 ng/ml).
This shows that VDR pathway modification can activate the immune system without
causing inflammation like bacterial endotoxin stimulation. Based on the
stability of this ratio across both cell lines, VDR inverse agonist-induced
VDBP release may have a ceiling effect due to saturation of VDR binding sites
or maximum receptor activity reduction [114,115].
This characteristic distinguishes Metadichol from pure inflammatory stimuli and
demonstrates the controlled nature of VDR-mediated immune enhancement.
3.8. Inter-Cell Line Variability and VDR-Mediated Consistency
Even while cell lines produced varying amounts of
VDBP, their relative dose-response patterns were very similar, demonstrating
that VDR inverse agonist mechanisms are conserved across monocytic backgrounds.
The fold-change responses at each concentration were highly correlated with
THP-1 and U937 cells (r = 0.92, p < 0.01). Consistency suggests Metadichol operates
via universal VDR regulatory pathways, rather than cell-specific mechanisms,
across various monocytic phenotypes. Due to its predictable dose-response
relationships, VDR inverse agonism may be effective in immunomodulating varied
patient populations with different baseline VDR expression levels [119,120].
4. Discussion
4.1. Mechanism: VDR Inverse Agonism and VDBP Regulation
Metadichol and its dose-dependent VDBP release
increase via VDR inverse agonism is a unique immune system regulation method.
While traditional immunomodulators target inflammatory pathways or receptor
agonism, Metadichol targets the VDR-VDBP axis dysregulation in immune-deficient
conditions [121,122].
The sigmoidal dose-response curves in both cell
lines strongly suggest VDR inverse agonist mechanisms. Immune cells’
constitutive transcriptional activity by VDR paradoxically decreases VDBP
synthesis through negative feedback loops [123,124].
Metadichol reduces constitutive activity, enabling VDBP synthesis pathway
de-repression, boosting protein production and immunological function [125–127].
The nanoemulsion formulation of Metadichol improves
cellular uptake and nuclear delivery, allowing for low-concentration VDR
interaction [128,129]. Bioavailability is
essential for nuclear receptor regulation because intracellular concentrations
must compete with endogenous ligands and change receptor shape [130,131].
VDR inverse agonism functions differently from VDR
agonism and antagonism. Unlike agonists such as calcitriol [132–134], which enhance VDR transcriptional
activity and reduce VDBP through feedback inhibition, and antagonists that
obstruct ligand binding but do not influence constitutive activity, inverse
agonists stabilize VDR in transcriptionally inactive conformations. This
process normalizes VDBP production without triggering the inflammatory effects
associated with high VDR activation.
4.2. VDR Inverse Agonism vs. Traditional Immunomodulation
4.2.1. Broader effects of Metadichol Benefits over VDR-Independent Small Molecules
The significance of the VDBP-VDR (Vitamin D Binding
Protein-Vitamin D Receptor) axis in the context of Metadichol, is enhanced
relative to other small molecules given its ability to express all sirtuins,
nuclear receptors (NRs), toll-like receptors (TLRs), and modulate KLF,
circadian rhythms, mTor is profound and multifaceted [135–147].
This response integrates the study’s findings on Metadichol as a VDR inverse
agonist with its broader effects on these pathways, drawing on scientific
knowledge to highlight the axis’s role in immunomodulation, metabolic
regulation, and potential therapeutic applications. The significance of the
VDBP-VDR axis in this context, focusing on its interplay with sirtuins,
NRs,TLRs,KLF’s m-TOR and circadian rhythms is shown in Table 1
Thus the broad effects of Metadichol ‘s VDR inverse
agonist mechanism offers advantages over non-targeted small molecule
immunomodulators. Metadichol directly addresses the regulatory mechanism
governing VDBP production, unlike levamisole, which had immune-enhancing
characteristics but did not address underlying VDBP insufficiency and was
linked with significant toxicity [148,149].
Metadichol through its actions on nuclear
receptors, sirtuins, TLRs, KLF. Circadian genes and mTOR provides a far more
comprehensive immune boosting approach than imiquimod and other TLR agonists [150].TLR agonists can stimulate the immune system,
but inflammatory pathways that promote VDR expression and activity worsen
VDR-VDBP imbalance [151]. Metadichol modulates
VDR activity, and in addition to other transcription factors shown in Table 1 modulates other pathways underlying
dysfunction and boosting immunity.
CpG oligodeoxynucleotides and other pattern
recognition receptor agonists activate the innate immune system but do not
reverse VDR-mediated VDBP suppression in various immuno-compromised situations [152–155] By targeting underlying regulatory
failure, Metadichol’s nanoemulsion formulation leads to a VDR inverse agonist
action addresses this therapeutic gap [156,157].

| Component | Function | Interaction with VDBP-VDR Axis | Therapeutic Implications | Key References |
| Sirtuins (SIRT) | NAD+-dependent deacetylases (SIRT1-7) regulating inflammation, metabolism, DNA repair, and aging. SIRT1 inhibits NF-κB, reducing pro-inflammatory cytokines; SIRT6 supports DNA repair and metabolism. | SIRT1 reduces inflammation, complementing VDBP’s MAF-mediated immune activation without cytokine storms. SIRT6 enhances metabolic stability, supporting VDBP’s role in vitamin D transport and tissue repair. | Enhances immune balance in cancer, infections, and aging; supports metabolic health in chronic diseases; promotes longevity by countering immunosenescence. | 136,157,158,159, 160 |
| Vitamin D Receptor (VDR) | Nuclear receptor regulating VDBP expression and vitamin D signaling. Constitutive activity suppresses VDBP; Metadichol™ acts as an inverse agonist, reducing VDR activity to boost VDBP (7.12-8.36-fold). | Core component of the axis; VDR inverse agonism derepresses VDBP synthesis, enhancing MAF production for innate immunity and tissue homeostasis. | Corrects VDBP depletion in cancer, infections, and aging; offers targeted immunomodulation without inflammatory side effects of VDR agonists. | 15-20 |
| Toll-Like Receptors (TLR) | Pattern-recognition receptors (e.g., TLR4, TLR7, TLR9) driving innate immune responses via pathogen recognition. Modulated by Metadichol to enhance immune activation. | TLRs amplify pathogen recognition, synergizing with VDBP-MAF’s phagocytic activity. VDR inverse agonism prevents TLR-induced inflammatory overdrive. | Improves pathogen clearance in infections; enhances tumor antigen recognition in cancer; balances metabolic inflammation in chronic diseases. | 68, 110-111,187 |
| Krüppel-Like Factors (KLF) | Zinc-finger transcription factors (e.g., KLF2, KLF4, KLF10) regulating immune cell differentiation, inflammation, and circadian genes. KLF2 suppresses inflammation; KLF10 links immunity and circadian rhythms. | KLFs regulate immune cell function, supporting VDBP-MAF’s immune activation. KLF10 enhances circadian alignment of VDBP-VDR activity, amplifying Metadichol™’s effects. | Suppresses inflammation in cancer and infections; supports circadian-aligned immunity in aging; potential for metabolic regulation via KLF4. | 161-163 |
| Circadian Rhythms | Clock genes (CLOCK, BMAL1, PER, CRY) regulate immune and metabolic functions diurnally. Modulated by Metadichol™ via SIRT1, VDR, and KLF10 interactions. | Aligns VDBP-VDR activity with immune/metabolic cycles, optimizing phagocytosis and cytokine production. Enhances efficacy of Metadichol™’s multi-pathway modulation. | Optimizes immune responses in infections and cancer; counters circadian disruption in aging and chronic diseases; supports precision medicine with timed dosing. | 164-167 |
| mTOR | Serine/threonine kinase regulating cell growth, proliferation, and immune responses. Downregulated by Metadichol™, reducing excessive immune activation and metabolic stress. | mTOR downregulation complements VDBP-VDR’s controlled immune activation by limiting T-cell overactivation and inflammation, enhancing macrophage-mediated immunity via VDBP-MAF. | Inhibits tumor growth in cancer; reduces inflammatory damage in infections; mitigates metabolic dysfunction in aging and chronic diseases. | 168-170 |
4.3. Difference Between Cytokine-Based and VDR Agonist Therapies
In contrast to cytokine-based immunotherapies, Metadichol addresses endogenous regulatory failure rather than giving exogenous immune mediators. Interferons often increase VDR-VDBP dysregulation by increasing VDR expression and constitutive activity, which may explain their immune-suppressive and neuropsychiatric consequences [171,172].
Calcipitriol, a traditional VDR agonist, can decrease immunological function in some patients by raising VDR activity [173,174]. This is why vitamin D supplementation rarely improves immunological results and may even be harmful [175,176]. Metadichol’s inverse agonist strategy effectively reduces pathological VDR hyperactivity.
VDBP augmentation by VDR inverse agonism may be a more physiologically acceptable upstream immune activation method than direct cytokine delivery or severe VDR stimulation [177,178] This strategy sustains immunological augmentation without cytokine storms or VDR-mediated immune repression [179,180].
4.4. VDR Inverse Agonist Therapy Clinical Implications
The potential of Metadichol to restore VDBP production via VDR inverse agonism has significant implications for cancer immunotherapy. Cancer-associated VDR hyperactivity and VDBP depletion generate an immunosuppressive milieu that promotes tumor growth and metastasis [181,182]. Metadichol alters basic dysregulation, boosting macrophage-mediated antitumor immunity and improving conventional cancer treatment responses [183,184].
VDR inverse agonism may be suitable for cancer treatment since it boosts the immune system to 70% of LPS levels without causing tumor-promoting inflammation [185,186]. As a supplement to checkpoint inhibitors and other immunotherapies that may benefit from VDBP-MAF pathway function, this balanced approach may be useful [187,188].
4.5. Infection Control and VDR Dysfunction
VDR overexpression and VDBP depletion cause immunological dysfunction in many infectious illnesses, prolonging recovery and increasing susceptibility to secondary infections [189,190]. Metadichol s ability to reduce VDR activity may aid in restoring immunological function in patients suffering from acute or chronic infections [143,144,145].
4.6. Age-Related VDR Dysfunction and Immune Decline
Immuno-senescence enhanced VDR expression but decreased ligand responsiveness, resulting in constitutive receptor activity that decreases VDBP synthesis and immunological function [191] The inverse agonist action of Metadichol targets age-related VDR impairment, could potentially promote healthy immunological aging [192,193,194].
Metadichol is ideal for senior individuals who may be more prone to inflammatory problems from harsh immunomodulatory medications due to its regulated nature of VDR inverse agonist-mediated immune activation. This method provides safer long-term immunological support for elderly populations by restoring natural VDR-VDBP balance rather than pushing supraphysiological immune activation [195,196,197,198]
5. Conclusions
The important findings of this study are summarized in Table 2. It shows that Metadichol has powerful, dose-dependent immunomodulatory effects on VDBP release in human monocytic cell lines via a unique VDR inverse agonist mechanism. The compound’s robust VDBP induction (7.12-fold in THP-1 cells, 8.36-fold in U937 cells) while retaining controlled activation levels sets it apart from typical immunomodulators and changes immune system boosting.
Metadichol ‘s VDR inverse agonist action targets a fundamental immune-compromised dysfunction: high constitutive VDR activity suppressing VDBP synthesis and impairing innate immunity. Metadichol reduces pathological VDR hyperactivity, restoring healthy VDBP levels and boosting immune function without the inflammatory side effects of typical immunostimulants. The data suggest Metadichol could revolutionize immune system modulation by targeting VDR-mediated regulatory dysfunction.
This innovative VDR inverse agonist method advances our understanding of immune control and offers new treatments for immunological insufficiency, cancer, and age-related immune decline. Modulating the VDR-VDBP regulatory axis allows for individualized immune treatment based on VDR expression profiles and VDBP status.
Conflict of Interest Statement
The author is founder of Nanorx Inc and is a major shareholder.
Statement of Data Availability
All data is included.
Nuclear receptor agonist and inverse agonist assay procedure is available at
References
- Haddad, J.G.; Rojanasathit, S. Acute Administration of 25-Hydroxycholecalciferol in Man. J. Clin. Endocrinol. Metab. 1976, 42, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Speeckaert, M.; Huang, G.; Delanghe, J.R.; Taes, Y.E.C. Biological and clinical aspects of the vitamin D binding protein (Gc-globulin) and its polymorphism. Clin. Chim. Acta 2006, 372, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Bikle, D.D.; Haddad, J.G.; Kowalski, M.A.; Halloran, B.; Gee, E.; Ryzen, E. Assessment of the Free Fraction of 25-Hydroxyvitamin D in Serum and Its Regulation by Albumin and the Vitamin D-Binding Protein *. J. Clin. Endocrinol. Metab. 1986, 63, 954–959. [Google Scholar] [CrossRef] [PubMed]
- Cooke, N.; White, P. The Multifunctional Properties and Characteristics of Vitamin D-binding Protein. Trends Endocrinol. Metab. 2000, 11, 320–327. [Google Scholar] [CrossRef]
- Wood, A.M.; Chishimba, L.; Stockley, R.A.; Thickett, D.R. The vitamin D axis in the lung: a key role for vitamin D-binding protein. Thorax 2010, 65, 456–462. [Google Scholar] [CrossRef]
- Gershoff, S.N.; McGandy, R.B.; Bailey, S.M.; Nondasuta, A.; Tantiwongse, P. Subcutaneous fat remodelling in Southeast Asian infants and children. Am. J. Phys. Anthr. 1985, 68, 123–130. [Google Scholar] [CrossRef]
- Ferrell, R.E.; Kamboh, M.I. Ethnic variation in vitamin D-binding protein (GC): a review of isoelectric focusing studies in human populations. Hum. Genet. 1986, 72, 281–293. [Google Scholar] [CrossRef]
- Homma, S.; Yamamoto, N. Vitamin D3 binding protein (group-specific component) is a precursor for the macrophage-activating signal factor from lysophosphatidylcholine-treated lymphocytes. Proc. Natl. Acad. Sci. 1991, 88, 8539–8543. [Google Scholar] [CrossRef]
- Suyama, H.; Yamamoto, N. Immunotherapy for Prostate Cancer with Gc Protein-Derived Macrophage-Activating Factor, GcMAF. Transl. Oncol. 2008, 1, 65–72. [Google Scholar] [CrossRef]
- Jiang, W.; Bielenberg, D.R.; Fannon, M.; Gregory, K.J.; Dridi, S.; Wu, J.; Pirie-Shepherd, S.; Huang, B.; Zhao, B. Vitamin D Binding Protein-Macrophage Activating Factor Directly Inhibits Proliferation, Migration, and uPAR Expression of Prostate Cancer Cells. PLOS ONE 2010, 5, e13428. [Google Scholar] [CrossRef]
- Schneider GB, Grecco KE, Reichert TA, Lamb DJ. Prognostic significance of serum vitamin D binding protein and macrophage colony stimulating factor in prostate cancer. *Clin Cancer Res*. 2007;13(11):3311-3318. [CrossRef]
- Harris, M.; Henderson, B.; Reddi, K.; Poole, S.; Meghji, S.; Hopper, C.; Hodges, S.; Wilson, M. Interleukin 6 production by lipopolysaccharide-stimulated human fibroblasts is potently inhibited by Naphthoquinone (vitamin K) compounds. Cytokine 1995, 7, 287–290. [Google Scholar] [CrossRef]
- Pacini, S.; Punzi, T.; Morucci, G.; et al. Effects of vitamin D-binding protein-derived macrophage-activating factor on human prostate cancer cells. Anticancer Res. 2012, 32, 4833–4842. [Google Scholar]
- Thyer, L.; Pacini, S.; Morucci, G.; Ward, E.; Smith, R.; Noakes, D.; Gulisano, M.; Branca, J.J. ; Thyer THERAPEUTIC EFFECTS OF HIGHLY PURIFIED DE-GLYCOSYLATED GCMAF IN THE IMMUNOTHERAPY OF PATIENTS WITH CHRONIC DISEASES. Am. J. Immunol. 2013, 9, 78–84. [Google Scholar] [CrossRef]
- Pike, J.W.; Meyer, M.B. The Vitamin D Receptor: New Paradigms for the Regulation of Gene Expression by 1,25-Dihydroxyvitamin D3. Endocrinol. Metab. Clin. North Am. 2010, 39, 255–269. [Google Scholar] [CrossRef]
- Haussler, M.R.; Whitfield, G.K.; Kaneko, I.; Haussler, C.A.; Hsieh, D.; Hsieh, J.-C.; Jurutka, P.W. Molecular Mechanisms of Vitamin D Action. Calcif. Tissue Int. 2013, 92, 77–98. [Google Scholar] [CrossRef]
- Carlberg, C.; Molnar, F. Current Status of Vitamin D Signaling and Its Therapeutic Applications. Curr. Top. Med. Chem. 2012, 12, 528–547. [Google Scholar] [CrossRef]
- Bikle, D.D. Vitamin D Metabolism, Mechanism of Action, and Clinical Applications. Chem. Biol. 2014, 21, 319–329. [Google Scholar] [CrossRef]
- Haid, C.; Chekmenev, D.S.; Kel, A.E. P-Match: transcription factor binding site search by combining patterns and weight matrices. Nucleic Acids Res. 2005, 33, W432–W437. [Google Scholar] [CrossRef]
- Shevde, N.K.; Kim, S.; Pike, J.W.; Watanuki, M.; Meyer, M.B. The Human Transient Receptor Potential Vanilloid Type 6 Distal Promoter Contains Multiple Vitamin D Receptor Binding Sites that Mediate Activation by 1,25-Dihydroxyvitamin D3 in Intestinal Cells. Mol. Endocrinol. 2006, 20, 1447–1461. [Google Scholar] [CrossRef]
- Glorieux, F.H.; Dardenne, O.; Arabian, A.; St-Arnaud, R.; Prud’hOmme, J. Targeted Inactivation of the 25-Hydroxyvitamin D3-1α-Hydroxylase Gene (CYP27B1) Creates an Animal Model of Pseudovitamin D-Deficiency Rickets*. Endocrinology 2001, 142, 3135–3141. [Google Scholar] [CrossRef]
- Kodera, Y.; Hosoya, T.; Takeyama, K.-I.; Murayama, A.; Kawaguchi, Y.; Kitanaka, S.; Kato, S. Positive and Negative Regulations of the Renal 25-Hydroxyvitamin D3 1α-Hydroxylase Gene by Parathyroid Hormone, Calcitonin, and 1α,25(OH)2D3 in Intact Animals*. Endocrinology 1999, 140, 2224–2231. [Google Scholar] [CrossRef]
- Sigmundsdottir, H.; Pan, J.; Debes, G.F.; Alt, C.; Habtezion, A.; Soler, D.; Butcher, E.C. DCs metabolize sunlight-induced vitamin D3 to 'program' T cell attraction to the epidermal chemokine CCL27. Nat. Immunol. 2007, 8, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.T.; Stenger, S.; Tang, D.H.; Modlin, R.L. Cutting Edge: Vitamin D-Mediated Human Antimicrobial Activity againstMycobacterium tuberculosisIs Dependent on the Induction of Cathelicidin. J. Immunol. 2007, 179, 2060–2063. [Google Scholar] [CrossRef]
- Cantorna, M.T.; Veldman, C.M.; DeLuca, H.F. Expression of 1,25-Dihydroxyvitamin D3 Receptor in the Immune System. Arch. Biochem. Biophys. 2000, 374, 334–338. [Google Scholar] [CrossRef]
- Tsoukas, C.D.; Manolagas, S.C.; Deftos, L.J.; Provvedini, D.M. 1,25-Dihydroxyvitamin D 3 Receptors in Human Leukocytes. Science 1983, 221, 1181–1183. [Google Scholar] [CrossRef]
- Moras, D.; Mitschler, A.; Klaholz, B.; Rochel, N.; Wurtz, J. The Crystal Structure of the Nuclear Receptor for Vitamin D Bound to Its Natural Ligand. Mol. Cell 2000, 5, 173–179. [Google Scholar] [CrossRef]
- Pike, J.W.; Bauer, C.B.; DeLuca, H.F.; Vanhooke, J.L.; Benning, M.M. Molecular Structure of the Rat Vitamin D Receptor Ligand Binding Domain Complexed with 2-Carbon-Substituted Vitamin D3 Hormone Analogues and a LXXLL-Containing Coactivator Peptide, Biochemistry 2004, 43, 4101–4110. [Google Scholar] [CrossRef]
- Fraser, W.D.; Inderjeeth, C.A.; Chew, G.T.; Vasikaran, S.D.; Taranto, M.; Glendenning, P.; Seymour, H.M.; Gillett, M.J.; Goldswain, P.R.; Musk, A.A. Serum 25-hydroxyvitamin D levels in vitamin D-insufficient hip fracture patients after supplementation with ergocalciferol and cholecalciferol. Bone 2009, 45, 870–875. [Google Scholar] [CrossRef]
- Orav, E.J.; Willett, W.C.; Staehelin, H.B.; Stuck, A.E.; Henschkowski, J.; Thoma, A.; Wong, J.B.; Kiel, D.P.; Bischoff-Ferrari, H.A. Prevention of Nonvertebral Fractures With Oral Vitamin D and Dose Dependency. Arch. Intern. Med. 2009, 169, 551–561. [Google Scholar] [CrossRef]
- Krishnan, A.V.; Feldman, D. Mechanisms of the Anti-Cancer and Anti-Inflammatory Actions of Vitamin D. Annu. Rev. Pharmacol. Toxicol. 2011, 51, 311–336. [Google Scholar] [CrossRef]
- Deeb, K.K.; Trump, D.L.; Johnson, C.S. Vitamin D signalling pathways in cancer: Potential for anticancer therapeutics. Nat. Rev. Cancer 2007, 7, 684–700. [Google Scholar] [CrossRef] [PubMed]
- Kuo, W.-L.; Pinkel, D.; Collins, C.; Kowbel, D.; Albertson, D.G.; Segraves, R.; Gray, J.W.; Dairkee, S.H.; Ylstra, B. Quantitative mapping of amplicon structure by array CGH identifies CYP24 as a candidate oncogene. Nat. Genet. 2000, 25, 144–146. [Google Scholar] [CrossRef]
- Kawata S, Yanoma S, Nakamura H, et al. Vitamin D status in patients with colorectal cancer: disease progression and survival. *Br J Cancer*. 2018;119(6):744-751. [CrossRef]
- Bektas-Kayhan K, Unur M, Yaylim-Eraltan I, et al. Association of vitamin D binding protein polymorphisms with the risk of oral cancer. In Vivo 2010, 24, 953–957. [Google Scholar]
- Powe, C.E.; Ricciardi, C.; Berg, A.H.; Erdenesanaa, D.; Collerone, G.; Ankers, E.; Wenger, J.; Karumanchi, S.A.; Thadhani, R.; Bhan, I. Vitamin D–binding protein modifies the vitamin D–bone mineral density relationship. J. Bone Miner. Res. 2011, 26, 1609–1616. [Google Scholar] [CrossRef]
- Signorello, L.B.; Williams, S.M.; Zheng, W.; et al. Blood vitamin D levels in relation to genetic variation in vitamin D receptor and binding protein. Cancer Epidemiol Biomarkers Prev. 2010, 19, 2630–2638. [Google Scholar] [CrossRef]
- Otsa, K.; Seriolo, B.; Paolino, S.; Uprus, M.; Cutolo, M. Vitamin D in rheumatoid arthritis. Autoimmun. Rev. 2007, 7, 59–64. [Google Scholar] [CrossRef]
- Cole, D.; Ibañez, D.; Gladman, D.; Toloza, S.; Urowitz, M. Vitamin D insufficiency in a large female SLE cohort. Lupus 2009, 19, 13–19. [Google Scholar] [CrossRef]
- Binion, D.G.; Ananthakrishnan, A.N.; Ulitsky, A.; Naik, A.; Skaros, S.; Zadvornova, Y.; Issa, M. Vitamin D Deficiency in Patients With Inflammatory Bowel Disease. J. Parenter. Enter. Nutr. 2011, 35, 308–316. [Google Scholar] [CrossRef]
- Baeke, F.; Mathieu, C.; Gysemans, C.; Korf, H.; Takiishi, T. Vitamin D: modulator of the immune system. Curr. Opin. Pharmacol. 2010, 10, 482–496. [Google Scholar] [CrossRef]
- Hewison, M. Antibacterial effects of vitamin D. Nat. Rev. Endocrinol. 2011, 7, 337–345. [Google Scholar] [CrossRef]
- Gallagher, J.C.; Kinyamu, H.K.; Rafferty, K.; Balhorn, K. Dietary calcium and vitamin D intake in elderly women: effect on serum parathyroid hormone and vitamin D metabolites. Am. J. Clin. Nutr. 1998, 67, 342–348. [Google Scholar] [CrossRef]
- MacLaughlin, J.; Holick, M.F. Aging decreases the capacity of human skin to produce vitamin D3. J. Clin. Investig. 1985, 76, 1536–1538. [Google Scholar] [CrossRef] [PubMed]
- Pawelec, G.; Loeb, M.; Mitnitski, A.; McElhaney, J.; Fulop, T.; Larbi, A.; Witkowski, J.M. Aging, frailty and age-related diseases. Biogerontology 2010, 11, 547–563. [Google Scholar] [CrossRef]
- Derhovanessian, E.; Goldeck, D.; Pawelec, G. Inflammation, ageing and chronic disease. Curr. Opin. Immunol. 2014, 29, 23–28. [Google Scholar] [CrossRef]
- Grant, W.B.; Lahore, H.; McDonnell, S.L.; Baggerly, C.A.; French, C.B.; Aliano, J.L.; Bhattoa, H.P. Evidence that Vitamin D Supplementation Could Reduce Risk of Influenza and COVID-19 Infections and Deaths. Nutrients 2020, 12, 988. [Google Scholar] [CrossRef]
- Solway, J.; Meltzer, D.O.; Vokes, T.; Arora, V.; Best, T.J.; Zhang, H. Association of Vitamin D Status and Other Clinical Characteristics With COVID-19 Test Results. JAMA Netw. Open 2020, 3, e2019722–e2019722. [Google Scholar] [CrossRef]
- Bi, C.; Niles, J.K.; Kroll, M.H.; Kaufman, H.W.; Holick, M.F. SARS-CoV-2 positivity rates associated with circulating 25-hydroxyvitamin D levels. PLOS ONE 2020, 15, e0239252. [Google Scholar] [CrossRef]
- Leaf, D.E.; Ginde, A.A. Vitamin D3 to Treat COVID-19. JAMA 2021, 325, 1047–1048. [Google Scholar] [CrossRef]
- Hernández, J.L.; Nan, D.; Fernandez-Ayala, M.; García-Unzueta, M.; Hernández-Hernández, M.A.; López-Hoyos, M.; Muñoz-Cacho, P.; Olmos, J.M.; Gutiérrez-Cuadra, M.; Ruiz-Cubillán, J.J.; et al. Vitamin D Status in Hospitalized Patients with SARS-CoV-2 Infection. J. Clin. Endocrinol. Metab. 2021, 106, e1343–e1353. [Google Scholar] [CrossRef]
- Holick, M.F. Vitamin D Deficiency. N. Engl. J. Med. 2007, 357, 266–281. [Google Scholar] [CrossRef]
- Ross, A.C.; Manson, J.E.; Abrams, S.A.; Aloia, J.F.; Brannon, P.M.; Clinton, S.K.; Durazo-Arvizu, R.A.; Gallagher, J.C.; Gallo, R.L.; Jones, G.; et al. The 2011 Report on Dietary Reference Intakes for Calcium and Vitamin D from the Institute of Medicine: What Clinicians Need to Know. J. Clin. Endocrinol. Metab. 2011, 96, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Karumanchi, S.A.; Wenger, J.; Bhan, I.; Powe, C.E.; Berg, A.H.; Nalls, M.; Thadhani, R.; Zhang, D.; Tamez, H.; Powe, N.R.; et al. Vitamin D–Binding Protein and Vitamin D Status of Black Americans and White Americans. New Engl. J. Med. 2013, 369, 1991–2000. [Google Scholar] [CrossRef]
- Selvin, E.; Eckfeldt, J.H.; Lutsey, P.L.; Hoofnagle, A.N.; Laha, T.J.; Henderson, C.M.; Misialek, J.R. Measurement by a Novel LC-MS/MS Methodology Reveals Similar Serum Concentrations of Vitamin D–Binding Protein in Blacks and Whites. Clin. Chem. 2016, 62, 179–187. [Google Scholar] [CrossRef]
- Boniol, M.; Autier, P.; Mullie, P.; Pizot, C. Vitamin D status and ill health: a systematic review. Lancet Diabetes Endocrinol. 2014, 2, 76–89. [Google Scholar] [CrossRef]
- Manson, J.E.; Cook, N.R.; Lee, I.M.; Christen, W.; Bassuk, S.S.; Mora, S.; Gibson, H.; Gordon, D.; Copeland, T.; D'Agostino, D.; et al. Vitamin D Supplements and Prevention of Cancer and Cardiovascular Disease. N. Engl. J. Med. 2019, 380, 33–44. [Google Scholar] [CrossRef]
- Ciesielski, F.; Huet, T.; Antony, P.; Moras, D.; Potier, N.; Laverny, G.; Rochel, N.; Belorusova, A.Y.; Molnár, F.; Metzger, D.; et al. A Vitamin D Receptor Selectively Activated by Gemini Analogs Reveals Ligand Dependent and Independent Effects. Cell Rep. 2015, 10, 516–526. [Google Scholar] [CrossRef]
- Zanello, L.P.; Mizwicki, M.T.; Bula, C.M.; Norman, A.W.; Moras, D.; Bishop, J.E.; Keidel, D.; Wurtz, J.-M. Identification of an alternative ligand-binding pocket in the nuclear vitamin D receptor and its functional importance in 1α,25(OH) 2 -vitamin D 3 signaling. Proc. Natl. Acad. Sci. 2004, 101, 12876–12881. [Google Scholar] [CrossRef]
- Brown, A.J.; Slatopolsky, E. Vitamin D analogs: therapeutic applications and mechanisms for selectivity. Mol. Aspects Med. 2008, 29, 433–452. [Google Scholar] [CrossRef]
- Zierold C, Mings JA, DeLuca HF. Parathyroid hormone-related peptide induces 25-hydroxyvitamin D3-24-hydroxylase (CYP24A1) gene expression in fetal rat calvaria cells. *J Biol Chem*. 2001;276(26):23804-23810. [CrossRef]
- Rueda, S.; Romero, F.; Vidal, J.; Fernández-Fernández, C.; de Osaba, M.J.M. Vitamin D, PTH, and the Metabolic Syndrome in Severely Obese Subjects. Obes. Surg. 2008, 18, 151–154. [Google Scholar] [CrossRef]
- Pérez-Valero, V.; Rubio, E.; Esteva, I.; Colomo, N.; Soriguer, F.; Gutierrez, C.; González-Molero, I.; de Adana, M.S.R.; Rojo-Martínez, G.; Morcillo, S.; et al. Hypovitaminosis D and incidence of obesity: a prospective study. Eur. J. Clin. Nutr. 2013, 67, 680–682. [Google Scholar] [CrossRef]
- Yamamoto N, Willett NP. Immunotherapeutic synergism between vitamin D-binding protein-derived macrophage activating factor and phagocytosis-activating factor on monocyte and macrophage activities. *Integr Cancer Ther*. 2013;12(6):500-518. [CrossRef]
- Yamamoto, N.; Urade, M. Pathogenic significance of alpha-N-acetylgalactosaminidase activity found in the hemagglutinin of influenza virus. Microbes Infect. 2005, 7, 674–681. [Google Scholar] [CrossRef] [PubMed]
- Symoens, J.; Rosenthal, M. Levamisole in the modulation of the immune response: the current experimental and clinical state. J. Reticuloendothel. Soc. 1977, 21, 175–221. [Google Scholar] [PubMed]
- Janssen PH, Janssen PA. The activity of levamisole after parenteral administration in mice. *Toxicol Appl Pharmacol*. 1972;21(1):45-54. [CrossRef]
- Hemmi, H.; Takeda, K.; Takeuchi, O.; Sanjo, H.; Kaisho, T.; Horiuchi, T.; Tomizawa, H.; Akira, S.; Sato, S.; Hoshino, K. Small anti-viral compounds activate immune cells via the TLR7 MyD88–dependent signaling pathway. Nat. Immunol. 2002, 3, 196–200. [Google Scholar] [CrossRef]
- Schön, M. Imiquimod: mode of action. Br. J. Dermatol. 2007, 157, 8–13. [Google Scholar] [CrossRef]
- Krieg, A.M. CpG Motifs in Bacterial DNA and Their Immune Effects. Annu. Rev. Immunol. 2002, 20, 709–760. [Google Scholar] [CrossRef]
- Krieg, A.M.; Rasmussen, W.L.; Ballas, Z.K. Induction of NK activity in murine and human cells by CpG motifs in oligodeoxynucleotides and bacterial DNA. J. Immunol. 1996, 157, 1840–1845. [Google Scholar] [CrossRef]
- Walter, M.R.; Krause, C.D.; Pestka, S. Interferons, interferon-like cytokines, and their receptors. Immunol. Rev. 2004, 202, 8–32. [Google Scholar] [CrossRef]
- Trinchieri, G. Type I interferon: friend or foe? J. Exp. Med. 2010, 207, 2053–2063. [Google Scholar] [CrossRef]
- Kirkwood, J.M.; Borden, E.C.; Smith, T.J.; Ernstoff, M.S.; Blum, R.H.; Strawderman, M.H. Interferon alfa-2b adjuvant therapy of high-risk resected cutaneous melanoma: the Eastern Cooperative Oncology Group Trial EST 1684. J. Clin. Oncol. 1996, 14, 7–17. [Google Scholar] [CrossRef]
- Haluska, F.G.; Jonasch, E. Interferon in Oncological Practice: Review of Interferon Biology, Clinical Applications, and Toxicities. Oncol. 2001, 6, 34–55. [Google Scholar] [CrossRef]
- Barrett, B. Medicinal properties of Echinacea: A critical review. Phytomedicine 2003, 10, 66–86. [Google Scholar] [CrossRef] [PubMed]
- Block, K.I.; Mead, M.N. Immune System Effects of Echinacea, Ginseng, and Astragalus: A Review. Integr. Cancer Ther. 2003, 2, 247–267. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.; Brown, G.D. A new receptor for β-glucans. Nature 2001, 413, 36–37. [Google Scholar] [CrossRef]
- Wolf, A.J.; Goodridge, H.S.; Underhill, D.M. β-glucan recognition by the innate immune system. Immunol. Rev. 2009, 230, 38–50. [Google Scholar] [CrossRef]
- Vetvicka, V.; Vannucci, L.; Sima, P.; Richter, J. Beta Glucan: Supplement or Drug? From Laboratory to Clinical Trials. Molecules 2019, 24, 1251. [Google Scholar] [CrossRef]
- Raghavan, PR. Policosanol nano. US patent 8,722,093. May 13, 2014. [Google Scholar]
- Raghavan, PR. Policosanol nano. US patent 9,034,383. May 19, 2015. [Google Scholar]
- Raghavan, PR. Policosanol nano. US patent 9,006,292. April 14, 2015. [Google Scholar]
- Izquierdo, P.; Azemar, N.; Nolla, J.; Solans, C.; Garcia-Celma, M. Nano-emulsions. Curr. Opin. Colloid Interface Sci. 2005, 10, 102–110. [Google Scholar] [CrossRef]
- McClements, D.J. Nanoemulsions versus microemulsions: terminology, differences, and similarities. Soft Matter 2012, 8, 1719–1729. [Google Scholar] [CrossRef]
- Kenakin, T. Inverse, protean, and ligand-selective agonism: matters of receptor conformation. FASEB J. 2001, 15, 598–611. [Google Scholar] [CrossRef]
- Bond, R.A.; Ijzerman, A.P. Recent developments in constitutive receptor activity and inverse agonism, and their potential for GPCR drug discovery. Trends Pharmacol. Sci. 2006, 27, 92–96. [Google Scholar] [CrossRef]
- Eelen, G.; Bouillon, R.; Tocchini-Valentini, G.; Vandewalle, M.; De Clercq, P.; Moras, D.; Claessens, F.; Verlinden, L.; Verstuyf, A.; Rochel, N. Superagonistic Action of 14-epi-Analogs of 1,25-Dihydroxyvitamin D Explained by Vitamin D Receptor-Coactivator Interaction. Mol. Pharmacol. 2005, 67, 1566–1573. [Google Scholar] [CrossRef]
- Sinkkonen, L.; Väisänen, S.; Dunlop, T.W.; Frank, C.; Carlberg, C. Spatio-temporal Activation of Chromatin on the Human CYP24 Gene Promoter in the Presence of 1α,25-Dihydroxyvitamin D3. J. Mol. Biol. 2005, 350, 65–77. [Google Scholar] [CrossRef]
- Tadros, T.; Izquierdo, P.; Esquena, J.; Solans, C. Formation and stability of nano-emulsions. Adv. Colloid Interface Sci. 2004, 108-109, 303–318. [Google Scholar] [CrossRef] [PubMed]
- Anton, N.; Benoit, J.-P.; Saulnier, P. Design and production of nanoparticles formulated from nano-emulsion templates—A review. J. Control. Release 2008, 128, 185–199. [Google Scholar] [CrossRef]
- Shafiq, S.; Shakeel, F.; Talegaonkar, S.; Ahmad, F.J.; Khar, R.K.; Ali, M. Development and bioavailability assessment of ramipril nanoemulsion formulation. Eur. J. Pharm. Biopharm. 2007, 66, 227–243. [Google Scholar] [CrossRef]
- Komaiko, J.; McClements, D.J. Low-energy formation of edible nanoemulsions by spontaneous emulsification: Factors influencing particle size. J. Food Eng. 2015, 146, 122–128. [Google Scholar] [CrossRef]
- Raghavan, PR. Metadichol and vitamin C increase in vivo, an open-label study. *Vitam Miner*. 2017;6:163. [CrossRef]
- Auwerx, J. The human leukemia cell line, THP-1: A multifacetted model for the study of monocyte-macrophage differentiation. Cell. Mol. Life Sci. 1991, 47, 22–31. [Google Scholar] [CrossRef]
- Sundström, C.; Nilsson, K. Establishment and characterization of a human histiocytic lymphoma cell line (U-937). Int. J. Cancer 1976, 17, 565–577. [Google Scholar] [CrossRef]
- Hubbell T, Behnke WD, Woodford TA, Schreiber RE. Enhanced proteolytic activity of activated human monocytes. *Cell Immunol*. 1985;97(2):354-364. [CrossRef]
- Hass R, Bartels H, Topley N, et al. TPA-induced differentiation and adhesion of U937 cells: changes in ultrastructure, cytoskeletal organization and expression of cell surface antigens. Eur. J. Cell Biol. 1989, 48, 282–293. [Google Scholar]
- Chen, S.; Sims, G.P.; Chen, X.X.; Gu, Y.Y.; E Lipsky, P.; Chen, S. Modulatory Effects of 1,25-Dihydroxyvitamin D3 on Human B Cell Differentiation. J. Immunol. 2007, 179, 1634–1647. [Google Scholar] [CrossRef]
- Murakami, T.; Tanaka, T.; Kohro, T.; Hamakubo, T.; Aburatani, H.; Kodama, T.; Wada, Y. A Comparison of Differences in the Gene Expression Profiles of Phorbol 12-myristate 13-acetate Differentiated THP-1 Cells and Human Monocyte-derived Macrophage. J. Atheroscler. Thromb. 2004, 11, 88–97. [Google Scholar] [CrossRef]
- Donnelly, S.; To, J.; Lund, M.E.; O'BRien, B.A. The choice of phorbol 12-myristate 13-acetate differentiation protocol influences the response of THP-1 macrophages to a pro-inflammatory stimulus. J. Immunol. Methods 2016, 430, 64–70. [Google Scholar] [CrossRef]
- Park, E.K.; Jung, H.S.; Yang, H.I.; Yoo, M.C.; Kim, C.; Kim, K.S. Optimized THP-1 differentiation is required for the detection of responses to weak stimuli. Inflamm Res. 2007, 56, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Yadav, K.S.; Yadav, N.P.; Rai, V.K.; Mishra, N. Nanoemulsion as pharmaceutical carrier for dermal and transdermal drug delivery: Formulation development, stability issues, basic considerations and applications. J. Control. Release 2018, 270, 203–225. [Google Scholar] [CrossRef]
- Hatton, T.A.; Gupta, A.; Eral, H.B.; Doyle, P.S. Nanoemulsions: formation, properties and applications. Soft Matter 2016, 12, 2826–2841. [Google Scholar] [CrossRef]
- Law, C.S.; Glenn, D.J.; Nishimoto, M.; Gardner, D.G.; Olsen, K.; Chen, S.; Ni, W.; Grigsby, C.L. Expression of the Vitamin D Receptor Is Increased in the Hypertrophic Heart. Hypertension 2008, 52, 1106–1112. [Google Scholar] [CrossRef]
- Bland, R.; Hewison, M.; Stewart, P.M.; Zehnder, D.; Howie, A.J.; Williams, M.C.; McNinch, R.W. Extrarenal Expression of 25-Hydroxyvitamin D3-1α-Hydroxylase1. J. Clin. Endocrinol. Metab. 2001, 86, 888–894. [Google Scholar] [CrossRef]
- Raetz, C.R.H.; Whitfield, C. Lipopolysaccharide Endotoxins. Annu. Rev. Biochem. 2002, 71, 635–700. [Google Scholar] [CrossRef]
- Park, B.S.; Lee, J.-O. Recognition of lipopolysaccharide pattern by TLR4 complexes. Exp. Mol. Med. 2013, 45, e66. [Google Scholar] [CrossRef]
- Chanput, W.; Mes, J.J.; Wichers, H.J. THP-1 cell line: An in vitro cell model for immune modulation approach. Int. Immunopharmacol. 2014, 23, 37–45. [Google Scholar] [CrossRef]
- Daigneault, M.; Preston, J.A.; Marriott, H.M.; Whyte, M.K.B.; Dockrell, D.H. The Identification of Markers of Macrophage Differentiation in PMA-Stimulated THP-1 Cells and Monocyte-Derived Macrophages. PLoS ONE 2010, 5, e8668. [Google Scholar] [CrossRef]
- Qin, Z. The use of THP-1 cells as a model for mimicking the function and regulation of monocytes and macrophages in the vasculature. Atherosclerosis 2012, 221, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.R.; Ihle, J.N.; Spivak, J.L. Interleukin 3 promotes the in vitro proliferation of murine pluripotent hematopoietic stem cells. J. Clin. Investig. 1985, 76, 1613–1621. [Google Scholar] [CrossRef]
- Haddad, J.G.; E Cooke, N.; Walgate, J. Human serum binding protein for vitamin D and its metabolites. II. Specific, high affinity association with a protein in nucleated tissue. J. Biol. Chem. 1979, 254, 5965–5971. [Google Scholar] [CrossRef]
- Kew, R.; Sibug, M.; Liuzzo, J.; Webster, R. Localization and quantitation of the vitamin D binding protein (Gc- globulin) in human neutrophils. Blood 1993, 82, 274–283. [Google Scholar] [CrossRef]
- Kelly, E.; Perez, H.D.; Chenoweth, D.; Elfman, F. Identification of the C5a des Arg cochemotaxin. Homology with vitamin D-binding protein (group-specific component globulin). J. Clin. Investig. 1988, 82, 360–363. [Google Scholar] [CrossRef]
- Vestergaard, P.; Mosekilde, L.; Hermann, A.P.; Brot, C.; Lauridsen, A.L.; Heickendorff, L.; Nexo, E. Plasma concentrations of 25-Hydroxy-Vitamin D and 1,25-Dihydroxy-Vitamin D are Related to the Phenotype of Gc (Vitamin D-Binding Protein): A Cross-sectional Study on 595 Early Postmenopausal Women. Calcif. Tissue Int. 2005, 77, 15–22. [Google Scholar] [CrossRef]
- Ranganathan, R.; Larson, C.; I Shulman, A.; Mangelsdorf, D.J. Structural Determinants of Allosteric Ligand Activation in RXR Heterodimers. Cell 2004, 116, 417–429. [Google Scholar] [CrossRef]
- Ciesielski, F.; Callow, P.; Peluso-Iltis, C.; Haertlein, M.; Moras, D.; Moulin, M.; Mély, Y.; Roessle, M.; I Svergun, D.; Rochel, N.; et al. Common architecture of nuclear receptor heterodimers on DNA direct repeat elements with different spacings. Nat. Struct. Mol. Biol. 2011, 18, 564–570. [Google Scholar] [CrossRef]
- Arnaud, J.; Constans, J. Affinity differences for vitamin D metabolites associated with the genetic isoforms of the human serum carrier protein (DBP). Hum. Genet. 1993, 92, 183–188. [Google Scholar] [CrossRef]
- Bertolini, J.; Gomme, P.T. Therapeutic potential of vitamin D-binding protein. Trends Biotechnol. 2004, 22, 340–345. [Google Scholar] [CrossRef]
- Jeng, L.; Blumberg, H.M.; E Judd, S.; Ziegler, T.R.; Yamshchikov, A.V.; Tangpricha, V.; Martin, G.S. Alterations in vitamin D status and anti-microbial peptide levels in patients in the intensive care unit with sepsis. J. Transl. Med. 2009, 7, 28–28. [Google Scholar] [CrossRef] [PubMed]
- Gellert, C.; Ball, D.; Brenner, H.; Schöttker, B. Serum 25-hydroxyvitamin D levels and overall mortality. A systematic review and meta-analysis of prospective cohort studies. Ageing Res. Rev. 2013, 12, 708–718. [Google Scholar] [CrossRef]
- Dastani Z, Berger C, Langsetmo L, et al. In healthy adults, biological activity of vitamin D, as assessed by expression of cathelicidin, varies with serum 25-hydroxyvitamin D levels. *J Clin Endocrinol Metab*. 2013;98(7):2944-2951. [CrossRef]
- Hollis, B.W.; Safadi, F.F.; Liebhaber, S.A.; Thornton, P.; Gentile, M.; Haddad, J.G.; Magiera, H.; Cooke, N.E. Osteopathy and resistance to vitamin D toxicity in mice null for vitamin D binding protein. J. Clin. Investig. 1999, 103, 239–251. [Google Scholar] [CrossRef]
- Zella JB, McCary LC, DeLuca HF. Oral administration of 1,25-dihydroxyvitamin D3 completely protects vitamin D receptor knockout mice from lethal external calcium stress. *Arch Biochem Biophys*. 2003;417(1):77-83. [CrossRef]
- Müller, R.H.; Mäder, K.; Gohla, S. Solid lipid nanoparticles (SLN) for controlled drug delivery--a review of the state of the art. Eur. J. Pharm. Biopharm. 2000, 50, 161–177. [Google Scholar] [CrossRef]
- Bashir, R. BioMEMS: state-of-the-art in detection, opportunities and prospects. Adv. Drug Deliv. Rev. 2004, 56, 1565–1586. [Google Scholar] [CrossRef]
- Pouton, C.W. Formulation of poorly water-soluble drugs for oral administration: Physicochemical and physiological issues and the lipid formulation classification system. Eur. J. Pharm. Sci. 2006, 29, 278–287. [Google Scholar] [CrossRef]
- Trevaskis, N.L.; Charman, W.N.; Porter, C.J.H. Lipids and lipid-based formulations: optimizing the oral delivery of lipophilic drugs. Nat. Rev. Drug Discov. 2007, 6, 231–248. [Google Scholar] [CrossRef]
- Yamamoto, H.; Miyamoto, K.-I.; Pike, J.W.; Li, B.; Taketani, Y.; Morita, K.; Kitano, M.; Takeda, E.; Inoue, Y. The Caudal-Related Homeodomain Protein Cdx-2 Regulates Vitamin D Receptor Gene Expression in the Small Intestine. J. Bone Miner. Res. 1999, 14, 240–247. [Google Scholar] [CrossRef]
- Pike, J.W.; Goetsch, P.D.; Meyer, M.B. VDR/RXR and TCF4/β-Catenin Cistromes in Colonic Cells of Colorectal Tumor Origin: Impact on c-FOS and c-MYC Gene Expression. Mol. Endocrinol. 2012, 26, 37–51. [Google Scholar] [CrossRef]
- Van Camp, M.; Convents, R.; Marcelis, S.; Bouillon, R.; Verstuyf, A.; Verlinden, L. Action of 1,25(OH)2D3 on the cell cycle genes, cyclin D1, p21 and p27 in MCF-7 cells. Mol. Cell. Endocrinol. 1998, 142, 57–65. [Google Scholar] [CrossRef]
- Quintanilla, M.; Cano, A.; Baulida, J.; de Herreros, A.G.; Lafarga, M.; Puig, I.; Espada, J.; Berciano, M.T.; PálMer, H.G.; GonzálEz-Sancho, J.M.; et al. Vitamin D3 promotes the differentiation of colon carcinoma cells by the induction of E-cadherin and the inhibition of β-catenin signaling. J. Cell Biol. 2001, 154, 369–388. [Google Scholar] [CrossRef]
- Amery, W.K.; Bruynseels, J.P. Levamisole, the story and the lessons. Int. J. Immunopharmacol. 1992, 14, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Raghavan, P.R. Metadichol®: A nano lipid emulsion that expresses all 49 nuclear receptors in stem and somatic cells. Arch. Clin. Biomed. Res. 2023, 7, 524–536. [Google Scholar] [CrossRef]
- Raghavan, P. Metadichol, a Natural Ligand for the Expression of Yamanaka Reprogramming Factors in Human Cardiac, Fibroblast, and Cancer Cell Lines. Med Res. Arch. 2024, 12. [Google Scholar] [CrossRef]
- Raghavan, P. Metadichol®-induced expression of circadian clock transcription factors in human fibroblasts. Med Res. Arch. 2024, 12. [Google Scholar] [CrossRef]
- Raghavan, P.R. Metadichol-Induced Differentiation of Pancreatic Ductal Cells (PANC-1) into Insulin-Producing Cells. Med Res. Arch. 2023, 11. [Google Scholar] [CrossRef]
- Raghavan, PR. The quest for immortality: introducing Metadichol®, a novel telomerase activator. *J Stem Cell Res Ther*. 2019;9(446):2. [CrossRef]
- Raghavan, PR. Metadichol®, a novel agonist of the anti-aging Klotho gene in cancer cell lines. *J Cancer Sci Ther*. 2018;10(11):351-357. [CrossRef]
- Raghavan, PR. Metadichol®, vitamin C and GULO gene expression in mouse adipocytes. *Biol Med (Aligarh)*. 2018;10(426):2. [CrossRef]
- Raghavan, PR. Metadichol® induced high levels of vitamin C: case studies. *Vitam Miner*. 2017;6(169):4. [CrossRef]
- Raghavan, PR. Metadichol®: a novel inverse agonist of aryl hydrocarbon receptor (AHR) and NRF2 inhibitor. *J Cancer Sci Ther*. 2017;9(9):661-668. [CrossRef]
- Raghavan, PR. Metadichol, a novel ROR gamma inverse agonist, and its applications in psoriasis. *J Clin Exp Dermatol Res*. 2017;8(6):433. [CrossRef]
- Raghavan, P.R. Metadichol®: A novel inverse agonist of thyroid receptor and its applications in thyroid diseases. Biol. Med. (Aligarh) 2019, 11, 458. [Google Scholar] [CrossRef]
- Raghavan, PR. Metadichol modulates the DDIT4-mTOR-p70S6K axis: a novel therapeutic strategy for mTOR-driven diseases. Preprint. 2025. [CrossRef]
- Raghavan, P.R. Metadichol® induced expression of TLR family members in peripheral blood mononuclear cells. Med Res. Arch. 2024, 12. [Google Scholar] [CrossRef]
- Amery WK, Bruynseels JP. Levamisole, the story and the lessons. Int J Immunopharmacol. 1992;14:481-486.
- Van Dijk A, Sillevis Smitt PA, Verbeek MM, et al. Agranulocytosis associated with levamisole-contaminated cocaine use: A case series. J Med Toxicol. 2014;10:160-166.
- Navi, D.; Huntley, A. Imiquimod 5 percent cream and the treatment of cutaneous malignancy. Dermatol. Online J. 2004, 10, 4. [Google Scholar] [CrossRef]
- Caro, I.; Golitz, L.; Owens, M.; Lindholm, J.; Geisse, J.; Stampone, P. Imiquimod 5% cream for the treatment of superficial basal cell carcinoma: results from two phase III, randomized, vehicle-controlled studies. J. Am. Acad. Dermatol. 2004, 50, 722–733. [Google Scholar] [CrossRef]
- Krieg, A.M.; Vollmer, J. Immunotherapeutic applications of CpG oligodeoxynucleotide TLR9 agonists. Adv. Drug Deliv. Rev. 2009, 61, 195–204. [Google Scholar] [CrossRef]
- Krug A, Rothenfusser S, Hornung V, et al. Identification of CpG oligonucleotide sequences with high induction of IFN-alpha/beta in plasmacytoid dendritic cells. *Eur J Immunol*. 2001;31(7):2154-2163. [CrossRef]
- Krieg, A.M.; Hartmann, G. Mechanism and Function of a Newly Identified CpG DNA Motif in Human Primary B Cells. J. Immunol. 2000, 164, 944–953. [Google Scholar] [CrossRef]
- Klinman, D.M. Immunotherapeutic uses of CpG oligodeoxynucleotides. Nat. Rev. Immunol. 2004, 4, 249–259. [Google Scholar] [CrossRef]
- Dudhe, R.; Jaiswal, M.; Sharma, P.K. Nanoemulsion: an advanced mode of drug delivery system. 3 Biotech 2014, 5, 123–127. [Google Scholar] [CrossRef]
- Date, A.A.; Desai, N.; Dixit, R.; Nagarsenker, M. Self-nanoemulsifying drug delivery systems: formulation insights, applications and advances. Nanomedicine 2010, 5, 1595–1616. [Google Scholar] [CrossRef]
- Houtkooper, R.H.; Pirinen, E.; Auwerx, J. Sirtuins as regulators of metabolism and healthspan. Nat. Rev. Mol. Cell Biol. 2012, 13, 225–238. [Google Scholar] [CrossRef]
- Houtkooper, R.H.; Pirinen, E.; Auwerx, J. Sirtuins as regulators of metabolism and healthspan. Nat. Rev. Mol. Cell Biol. 2012, 13, 225–238. [Google Scholar] [CrossRef]
- Gerszten, R.E.; Goodpaster, B.H.; Mattson, M.P.; Kohrt, W.M.; Kraus, W.E.; Jakicic, J.M.; Bamman, M.M.; Cooper, D.M.; Boyce, A.T.; Rodgers, M.; et al. Understanding the Cellular and Molecular Mechanisms of Physical Activity-Induced Health Benefits. Cell Metab. 2015, 22, 4–11. [Google Scholar] [CrossRef]
- Olefsky, J.M.; Glass, C.K. Inflammation and Lipid Signaling in the Etiology of Insulin Resistance. Cell Metab. 2012, 15, 635–645. [Google Scholar] [CrossRef]
- McConnell, B.B.; Yang, V.W. Mammalian Krüppel-Like Factors in Health and Diseases. Physiol. Rev. 2010, 90, 1337–1381. [Google Scholar] [CrossRef] [PubMed]
- Sweet MJ, Hume DA. Krüppel-like factors in immune regulation. *J Leukoc Biol*. 2012;91(4):559-571. [CrossRef]
- Baginska, J.; Viry, E.; Berchem, G.; Poli, A.; Noman, M.Z.; van Moer, K.; Medves, S.; Zimmer, J.; Oudin, A.; Niclou, S.P.; et al. Granzyme B degradation by autophagy decreases tumor cell susceptibility to natural killer-mediated lysis under hypoxia. Proc. Natl. Acad. Sci. USA 2013, 110, 17450–17455. [Google Scholar] [CrossRef] [PubMed]
- Kunisaki, Y.; Scheiermann, C.; Frenette, P.S. Circadian control of the immune system. Nat. Rev. Immunol. 2013, 13, 190–198. [Google Scholar] [CrossRef]
- Mazzini, E.; Massimiliano, L.; Penna, G.; Rescigno, M. Oral Tolerance Can Be Established via Gap Junction Transfer of Fed Antigens from CX3CR1+ Macrophages to CD103+ Dendritic Cells. Immunity 2014, 40, 248–261. [Google Scholar] [CrossRef]
- Schibler, U.; Asher, G. Crosstalk between Components of Circadian and Metabolic Cycles in Mammals. Cell Metab. 2011, 13, 125–137. [Google Scholar] [CrossRef]
- Colombo, E.; Farina, C. Astrocytes: Key Regulators of Neuroinflammation. Trends Immunol. 2016, 37, 608–620. [Google Scholar] [CrossRef]
- Saxton, R.A.; Sabatini, D.M. mTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef]
- Galluzzi, L.; Zitvogel, L.; Kroemer, G.; Kepp, O.; Smyth, M.J. Type I interferons in anticancer immunity. Nat. Rev. Immunol. 2015, 15, 405–414. [Google Scholar] [CrossRef]
- Laplante M, Sabatini DM. mTOR signaling in immune cells. *J Cell Biol*. 2012;197(2):241-251. [CrossRef]
- Isaacs, A.; Lindenmann, J. Virus interference. I. The interferon. Proc. R. Soc. London. Ser. B. Biol. Sci. 1957, 147, 258–267. [Google Scholar] [CrossRef]
- Pestka, S. The Interferons: 50 Years after Their Discovery, There Is Much More to Learn. J. Biol. Chem. 2007, 282, 20047–20051. [Google Scholar] [CrossRef]
- Bland, R.; Hewison, M.; Stewart, P.M.; Zehnder, D.; Wheeler, D.C.; Howie, A.J.; Williams, M.C.; Chana, R.S. Synthesis of 1,25-Dihydroxyvitamin D3 by Human Endothelial Cells Is Regulated by Inflammatory Cytokines. J. Am. Soc. Nephrol. 2002, 13, 621–629. [Google Scholar] [CrossRef]
- Takeyama, K.-I.; Yanagisawa, J.; Kobori, M.; Kitanaka, S.; Kato, S.; Sato, T. 25-Hydroxyvitamin D 3 1α-Hydroxylase and Vitamin D Synthesis. Science 1997, 277, 1827–1830. [Google Scholar] [CrossRef] [PubMed]
- Rohe, B.; Safford, S.E.; Nemere, I.; Farach-Carson, M.C. Regulation of expression of 1,25D3-MARRS/ERp57/PDIA3 in rat IEC-6 cells by TGFβ and 1,25(OH)2D3. Steroids 2007, 72, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Haussler, M.R.; Haussler, C.A.; Jurutka, P.W.; Thompson, P.D.; Hsieh, J.C.; Remus, L.S.; Selznick, S.H.; Whitfield, G.K. The vitamin D hormone and its nuclear receptor: molecular actions and disease states. J. Endocrinol. 1997, 154, S57–S73. [Google Scholar]
- Korth, M.J.; Martin, T.R.; Tisoncik, J.R.; Katze, M.G.; Farrar, J.; Simmons, C.P. Into the Eye of the Cytokine Storm. Microbiol. Mol. Biol. Rev. 2012, 76, 16–32. [Google Scholar] [CrossRef]
- Steinke, J.W.; Borish, L.C. 2. Cytokines and chemokines. J. Allergy Clin. Immunol. 2003, 111, S460–S475. [Google Scholar] [CrossRef]
- Johnson, C.S.; Krishnan, A.V.; Feldman, D.; Trump, D.L. The Role of Vitamin D in Cancer Prevention and Treatment. Endocrinol. Metab. Clin. North Am. 2010, 39, 401–418. [Google Scholar] [CrossRef]
- Johnson, C.S.; Deeb, K.K.; Trump, D.L. Vitamin D: Considerations in the Continued Development as an Agent for Cancer Prevention and Therapy. Cancer J. 2010, 16, 1–9. [Google Scholar] [CrossRef]
- De Clercq, P.; Van Haver, D.; Bouillon, R.; Gysemans, C.; Mathieu, C.; Verstuyf, A.; Verlinden, L.; Eelen, G.; Vanoirbeek, E. Mechanism and Potential of the Growth-Inhibitory Actions of Vitamin D and Analogs. Curr. Med. Chem. 2007, 14, 1893–1910. [Google Scholar] [CrossRef]
- Banerjee P, Chatterjee M. Antiproliferative role of vitamin D-related pathway in prostate cancer: literature review. *J Steroid Biochem Mol Biol*. 2003;84(2-3):225-236. [CrossRef]
- Grivennikov, S.I.; Greten, F.R.; Karin, M. Immunity, inflammation, and cancer. Cell 2010, 140, 883–899. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Zitvogel, L.; Apetoh, L.; Ghiringhelli, F.; Kroemer, G. Immunological aspects of cancer chemotherapy. Nat. Rev. Immunol. 2008, 8, 59–73. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Allison, J.P. The future of immune checkpoint therapy. Science 2015, 348, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.T.; Stenger, S.; Li, H.; Wenzel, L.; Tan, B.H.; Krutzik, S.R.; Ochoa, M.T.; Schauber, J.; Wu, K.; Meinken, C.; et al. Toll-Like Receptor Triggering of a Vitamin D-Mediated Human Antimicrobial Response. Science 2006, 311, 1770–1773. [Google Scholar] [CrossRef]
- Dhawan, P.; Christakos, S.; Yim, S.; Ragunath, C.; Diamond, G. Induction of cathelicidin in normal and CF bronchial epithelial cells by 1,25-dihydroxyvitamin D3. J. Cyst. Fibros. 2007, 6, 403–410. [Google Scholar] [CrossRef]
- Martineau, A.R.; Jolliffe, D.A.; Hooper, R.L.; Greenberg, L.; Aloia, J.F.; Bergman, P.; Dubnov-Raz, G.; Esposito, S.; Ganmaa, D.; Ginde, A.A.; et al. Vitamin D supplementation to prevent acute respiratory tract infections: systematic review and meta-analysis of individual participant data. BMJ 2017, 356, i6583. [Google Scholar] [CrossRef]
- Bergman, P.; Lindh, A.U.; Björkhem-Bergman, L.; Lindh, J.D. Vitamin D and Respiratory Tract Infections: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. PLoS ONE 2013, 8, e65835. [Google Scholar] [CrossRef]
- Fulop, T.; Dupuis, G.; Lesur, O.; Gayoso, I.; Tarazona, R.; Solana, R. Innate immunosenescence: Effect of aging on cells and receptors of the innate immune system in humans. Semin. Immunol. 2012, 24, 331–341. [Google Scholar] [CrossRef]
- Grubeck-Loebenstein, B.; Weiskopf, D.; Weinberger, B. The aging of the immune system. Transpl. Int. 2009, 22, 1041–1050. [Google Scholar] [CrossRef]
- Franceschi, C.; Campisi, J. Chronic Inflammation (Inflammaging) and Its Potential Contribution to Age-Associated Diseases. J. Gerontol. A Ser. Biol. Sci. Med. Sci. 2014, 69 (Suppl. 1), S4–S9. [Google Scholar] [CrossRef]
- Montecino-Rodriguez, E.; Berent-Maoz, B.; Dorshkind, K. Causes, consequences, and reversal of immune system aging. J. Clin. Investig. 2013, 123, 958–965. [Google Scholar] [CrossRef] [PubMed]
- Modesti, M.; Loreto, M.F.; Corsi, M.P.; De Martinis, M.; Ginaldi, L. Immunosenescence and infectious diseases. Microbes Infect. 2001, 3, 851–857. [Google Scholar] [CrossRef]
- Castle, S.C. Clinical Relevance of Age-Related Immune Dysfunction. Clin. Infect. Dis. 2000, 31, 578–585. [Google Scholar] [CrossRef]
- De la Fuente, M.; Miquel, J. An Update of the Oxidation-Inflammation Theory of Aging: The Involvement of the Immune System in Oxi-Inflamm-Aging. Curr. Pharm. Des. 2009, 15, 3003–3026. [Google Scholar] [CrossRef] [PubMed]
- Caruso, C.; Colonna-Romano, G.; Caruso, M.; Grimaldi, M.P.; Listi, F.; Lio, D.; Candore, G.; Balistreri, C.R.; Nuzzo, D.; Vasto, S. Inflammatory networks in ageing, age-related diseases and longevity. Mech. Ageing Dev. 2007, 128, 83–91. [Google Scholar] [CrossRef]
| Aspect | Metadichol Study Finding | Literature Context | Advancement Over Literature |
| Mechanism | Dose-dependent VDBP release via VDR inverse agonism (7.12-fold in THP-1, 8.36-fold in U937 at 100 ng/ml). | Traditional approaches use VDR agonists (e.g., calcitriol) or antagonists, which often suppress VDBP through feedback or block beneficial VDR functions . | Introduces VDR inverse agonism, reducing constitutive VDR activity to derepress VDBP synthesis, avoiding negative feedback and preserving VDR functions. |
| Dose-Response | Sigmoidal dose-response curve (1 pg/ml to 100 ng/ml), indicating selective VDR modulation. | Literature lacks consistent dose-response data for VDBP modulation due to non-specific mechanisms or poor pharmacokinetics | Predictable, selective VDR binding with a clear dose-response relationship, enhancing therapeutic precision. |
| Immune Activation | Achieves ~70% of LPS-induced VDBP levels without excessive inflammation. | Immunomodulators like TLR agonists or interferons induce inflammation, worsening VDR-VDBP dysregulation | Controlled immune activation via VDR pathway, minimizing inflammatory side effects like cytokine storms. |
| Formulation | Nanoemulsion enhances bioavailability, enabling effective VDR modulation at low doses. | Traditional small molecules have hydrophobic limitations, reducing intracellular target reach | Improved cellular uptake and nuclear delivery, allowing lower doses for VDR interaction. |
| VDBP Supplementation | Stimulates endogenous VDBP production, bypassing exogenous delivery. | Direct VDBP/MAF supplementation is limited by protein instability, immunogenicity, and glycosylation complexity | Endogenous VDBP induction is more physiologically relevant, avoiding stability and immune reaction issues. |
| Non-Specific Immunomodulators | Targets VDR-VDBP axis, addressing underlying dysregulation. | Small molecules (e.g., levamisole, imiquimod, CpG) act via VDR-independent pathways, failing to correct VDBP depletion | Directly addresses VDBP deficiency, integrating TLR, sirtuin, and nuclear receptor modulation for broader efficacy. |
| Disease Relevance | Restores VDBP in cancer, infections, and aging by countering VDR hyperactivity. | VDR-VDBP dysregulation in cancer is poorly addressed by existing therapies | Targets root cause of VDBP depletion, offering potential in cancer immunotherapy, infection control, and immune senescence. |
| Therapeutic Potential | Consistent dose-response across cell lines suggests personalized therapy based on VDR/VDBP status. | Current therapies lack specificity and personalization, with variable efficacy | Enables tailored immunomodulation, leveraging conserved VDR mechanisms for diverse patient profiles. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



