
Submitted:
14 July 2025
Posted:
15 July 2025
You are already at the latest version
Abstract
Free-living amoebae (FLA) are widely present in the environment and may be pathogenic for animals and humans. Studies on the prevalence of FLA in European soils are few. This study aimed to molecularly identify the species and genotype of FLA occurring in soil from Southern Poland. Forty soil samples were collected in June 2024 in the Babiogórski National Park. Amoebae cultures and a thermal-tolerance test were conducted. Five PCR protocols were applied for amplification of FLA SSU rRNA fragments. The following FLA, including potentially pathogenic ones, were detected: Acanthamoeba T4 and T13 genotypes in 79.1% of positive samples, Naegleria gruberi and Naegleria galeacystis in 25%, Vermamoeba vermiformis in 12.5%, and Paravahlkampfia sp. and Ptolemeba bulliensis in 8.3%. Species and genotype identification were determined by sequence comparison and phylogenetic analysis. This study reports the first time isolation of N. galeacystis from soil and N. gruberi and V. vermiformis from soil collected in Europe. The used primer sets have different usefulness for Naegleria species identification and their phylogenetic analysis. The primers applied in this study may not reveal the full diversity of amoebae in soil; therefore, it is necessary to design new primers for this purpose.
Keywords:
free-living amoebae
; soil
; SSU rRNA
; genotyping
1. Introduction
Free-living amoebae (FLA) are a polyphyletic group of protists that belong to three supergroups within the kingdom Protozoa: Amoebozoa, e.g., Acanthamoeba spp., Vermamoeba vermiformis (formerly Hartmannella vermiformis), Balamuthia spp., and Sappinia spp.; Excavata, e.g., Naegleria spp.; and Rhizaria, e.g., Paulinella [1─3]. FLA can survive and reproduce in the environment, and depending on external conditions, they may occur as active, feeding trophozoites or dormant, resistant cysts. They play critical ecological roles and interact with other microorganisms, forming a predator-prey, symbiotic, or host-parasite relationship with bacteria. FLA may serve as hosts for many animal and human pathogens, contributing to their survival, spread, and transmission [1,4,5]. Some free-living amoebae are pathogenic and may cause infections of the central nervous system (Acanthamoeba spp., Balamuthia mandrillaris, Sappinia pedata, and Naegleria fowlerii) or keratitis (Acanthamoeba spp. and Vermamoeba vermiformis) [2,3,5,6,7]. A few authors suggest the possibility of FLA presence in the gut microbiome of mammals and other animals [8].
Some FLA infecting humans have been well genetically analyzed and divided into genotypes, linked to their pathogenicity. Based on 18S rRNA gene sequence analysis, 23 genotypes (T1 to T23) of Acanthamoeba species have been distinguished. Some genotypes cause granulomatous amoebic encephalitis (GAE), Acanthamoeba keratitis, or both. In the case of N. fowlerii, five of the eight genotypes, established based on the sequence of ITS1 and 5.8S rRNA gene, are pathogenic and cause primary amoebic meningoencephalitis (PAM). There is no genotype classification for B. mandrillaris due to a lack of or low variation in its 18S rRNA and mitochondrial 16S rRNA genes, respectively [2]. The pathogenicity of V. vermiformis is questionable, and limited molecular analyses of this species demonstrated a low diversity of its 18S rRNA gene. However, the latest research, based on multi-locus analysis, revealed a high degree of genetic diversity and the presence of multiple cryptic species within V. vermiformis [2,9]. S. pedata is linked with only one case of encephalitis. The sequences of the 18S rRNA gene and ITS1, 5.8S, and ITS2 regions of the four analyzed isolates demonstrated a size variation, but little is known about the genetic diversity of this species [2]. In the case of Acanthamoeba, thermotolerance and osmotolerance are also considered to evaluate the strain's pathogenicity. However, further studies are required to clarify this question [10,11].
FLA are widely present in various natural and artificial environments and have been isolated from different water sources, solid matrices, and air [5,12]. Many studies have been conducted on the prevalence and genetic diversity of FLA in various solid matrices, including soil, mud, sand, sediments, compost, and dust [12]. However, there are few European studies on this topic, especially regarding non-Acanthamoeba FLA. Almost all previous studies were based on amoebae cultures before their identification. Using environmental DNA (eDNA), total DNA isolated from environmental samples (e.g., soil) originating from various organisms, helps reduce the time needed for examinations. It also allows for the detection of more species from the same sample. The challenge of the eDNA approach is the presence of PCR inhibitors or the abundance of different organisms in the same sample [13,14], which can lead to false-negative or false-positive results, respectively.
This study aims to identify FLA species occurring in soil at recreational mountain areas in Southern Poland, using PCR and sequencing, their genotyping, and verification of thermal tolerance. The next aim is to compare the usefulness of eDNA and DNA isolated from amoebae cultures for PCR amplification of FLA 18S rRNA gene fragments. The purpose is also to compare the sensitivity and specificity of five PCR protocols for FLA detection. The next aim was to evaluate the usefulness of the obtained SSU rRNA gene fragments for species identification, genotyping, and phylogenetic analysis. The knowledge regarding the prevalence, genetic diversity, and phylogenetic relationship of the detected amoebae will shed light on their biogeography and ecology and help evaluate their pathogenic potential and the health threat to visitors in the analyzed area.
2. Results
2.1. Results of Amoebae Cultures, Thermal Tolerance Test, PCRs, and Sequencing
The PCR with JDP1/JDP2 primers and eDNA isolated directly from soil yielded a negative result. Using other primers, only non-specific products were obtained with eDNA.
Among all forty soil samples cultured at 28°C, FLA were observed under a microscope in 24 (60%). After incubation of the transferred trophozoites at 37°C and 42°C, FLA were observed respectively on 24 plates (60% of all samples, 100% of all observed FLA) and 12 plates (30% of all samples, 50% of all observed FLA). All the FLA that grew at 37°C were washed off the plates, and DNA was extracted from the rinsed material. Next, PCR reactions were performed to detect FLA. The Acanthamoeba genus was detected using the JDP1/JDP2 primer set in 19 of 24 (79.1%) DNA samples. The Naegleria genus was detected using the FLA-F/FLA-R, Ami6F1/Ami9R, and AmeF977/AmeR1534 primer sets in 6/24 (25%) DNA samples. V. vermiformis was detected using the FLA-F/FLA-R, AmeF977/AmeR1534, and HARTfor/HARTrev primer sets in 3/24 (12.5%) DNA samples. Other amoebae (Paravahlkampfia sp. and Ptolemeba bulliensis) were detected using the FLA-F/FLA-R and AmeF977/AmeR1534 primer sets, respectively, in 2/24 (8.3%) DNA samples. Co-occurrence of two different amoebae was detected in 7/24 (29.16%) of the DNA samples. Among all Acanthamoeba strains, 63.2% (12/19) represent the T4 genotype, and 36.8% (7/19) the T13 genotype. The detailed results of amoebae cultures at 42°C, PCRs with individual primer pairs, and sequencing are presented in Table 1.
2.2. Results of Sequence Comparison and Phylogenetic Analysis
The Acanthamoeba genotype and the species of the remaining amoebae were identified by aligning their sequences with those in the GenBank database using the BLAST tool, as well as by analyzing the topology of the phylogenetic trees (Figure 1, Figure 2, Figure 3 and Figure 4). The detailed alignment results for Acanthamoeba, Naegleria, and Vermamoeba vermiformis are presented in Table 2, Table 3 and Table 4. The sequence obtained from sample 4 using the FLA-F/FLA-R primer set shows 99.25% identity to the sequence DQ388521 from the GenBank database, representing Paravahlkampfia sp. The sequence obtained from sample 19 using the AmeF977/AmeR1534 primer set shows 98.94% identity to the sequence PP732398 from the GenBank database, representing Ptolemeba bulliensis isolated from water (Philippines).
The phylogenetic trees based on the sequences obtained in this study and derived from the GenBank database are presented in Figure 1, Figure 2, Figure 3 and Figure 4. The twelve Acanthamoeba sequences (samples 4, 6, 12, 15, 19, 21, 27, 29, 31, 37, 39, and 40) obtained in this study belong to a clade consisting of T4 genotype sequences from the GenBank database. The seven sequences (samples 2, 10, 23, 28, 32, 33, and 35) belong to a clade that contains T13 genotype sequences from GenBank (Figure 1). These results are consistent with the results of sequence comparison (Table 3). The trees based on the products of the FLA-F/FLA-R and Ami6F1/Ami9R primer sets for the Naegleria genus (Figure 2 and Figure 3) have a similar topology. The three sequences obtained in this study (samples 8, 11, 24) make a group with the N. gruberi sequences from GenBank, while the two sequences (samples 7 and 10) are grouped with the N. galeacystis sequence from GenBank. These results are consistent with the results of sequence comparison (Table 3). The tree based on the products of the AmeF977/AmeR1534 primer set (Figure 4) has a different topology than the other two trees for the Naegleria genus. The sequences obtained from the two samples (7 and 10) belong to the same clade as the N. galeacystis sequence from GenBank, which is consistent with the sequence comparison (Table 3) and the trees’ topology presented in Figure 2 and Figure 3. The remaining sequences, obtained from samples 8, 11, and 24, form a clade with different Naegleria species within this tree. Using the AmeF977/AmeR1534 primer set, it is not possible to determine the species of the Naegleria strains detected in samples 8, 11, and 24 based on either the tree topology or sequence comparison.
2.3. Results of Statistical Analysis
Statistical analysis shows significant differences in the prevalence of genera Acanthamoeba and Naegleria; Acanthamoeba genus and Vermamoeba vermiformis; and Acanthamoeba genus and other amoebae (Paravahlkampfia sp. and Ptolemeba bulliensis). The p-values were 0.0262, 0.314, and 0.0338, respectively. The differences between Naegleria genus, V. vermiformis, and other amoebae, as well as between T4 and T13 genotypes of Acanthamoeba, are not statistically significant. The analysis does not show statistically significant differences in the prevalence of different amoebae between the samples collected in the particular zones.
3. Discussion
Studies on FLA prevalence in the environment include cultures of amoebae, which increase their number and reduce the amount of inhibitors. However, this approach is time-consuming and allows the detection of only those amoebae that can be grown under the used conditions. In this study, an attempt was made to molecularly detect FLA in eDNA isolates obtained directly from the soil. The use of a kit intended for DNA isolation from soil enabled the effective removal of inhibitors, as PCR products were obtained from eDNA using all primer pairs except the JDP1/JDP2 pair. However, all obtained products were nonspecific and derived from non-amoebae organisms. Using DNA isolates obtained from amoebae cultures, specific products were obtained with all primer pairs. The Acanthamoeba genus-specific primer set, JDP1/JDP2, was the most specific of all used, but it was not sensitive enough to amplify amoebae DNA without prior culture. Thus, amoebae cultures are necessary to detect these organisms in soil samples using the primers applied in this study.
The primer pairs used in this study have been applied in other papers for FLA detection in cultures from soil samples [15─19]. However, the authors did not compare their sensitivity and specificity. In this study, FLA-F/FLA-R and AmeF977/AmeR1534 sets appeared to be the most sensitive among the primers intended for different FLA DNA amplification. They both amplified the amoebae DNA in seven samples. However, in one sample, Paravahlkampfia sp. was detected only with the FLA-F/FLA-R set, and in another two samples, Ptolemeba bulliensis and Naegleria sp. were detected only with the AmeF977/AmeR1534 set. The Ami6F1/Ami9R set was the least sensitive and allowed for the detection of N. gruberi in the three samples in which this species was detected using the FLA-F/FLA-R set. The sequences amplified using FLA-F/FLA-R and Ami6F1/Ami9R pairs overlap to a large extent, and their use in phylogenetic analysis resulted in the creation of phylogenetic trees with very similar topologies. Topology of these trees, combined with sequence comparison, enabled the identification of species in Naegleria strains isolated in this study. The disadvantage of the AmeF977/AmeR1534 set was the inability to identify the species of Naegleria in 3 out of 5 samples based on both sequence comparison and phylogenetic tree topology. The FLA-F/FLA-R set seems to be the best choice, but it was not sensitive enough to amplify amoebae DNA in the two samples positive with the AmeF977/AmeR1534 set. None of the primer pairs intended for different FLA allowed for the detection of Acanthamoeba. Reyes-Batlle et al. [20] used the FLA-F/FLA-R primer set, and they also failed to detect Acanthamoeba in any of the 24 soil samples from El Hierro Island (Canary Islands), despite this species being one of the most common in soil and occurring in the Canary Islands [21]. They identified only V. vermiformis in 20.8% of the samples. In this study, V. vermiformis was detected in only one sample using FLA-F/FLA-R and AmeF977/AmeR1534 pairs, in contrast to three samples with the use of the HARTfor/HARTrev set, which is intended for the detection of the former genus Hartmannella. The application of the primers for a wide range of FLA used in this study does not reveal the full diversity of soil amoebae. It is necessary to design new, sensitive primers enabling the detection and species identification of FLA in soil and other environmental samples. A major challenge would be to create these primers so that their sensitivity and specificity are sufficient to detect FLA in eDNA isolated directly from soil. The use of metagenomics techniques would provide complete knowledge of the amoebae diversity in soil.
Studies on the prevalence of a wide range of FLA in soil and their genotyping are few in Europe. In Poland, Hendiger-Rizo et al. [16] detected Acanthamoeba spp. in most of the soil samples (78.2%) from Warsaw parks and squares, followed by Platyamoeba placida, Stenamoeba berchidia, and Allovahlkampfia sp. Denet et al. [22] examined the diversity of culturable FLA at the genus level in French alpine soils, based on PCR and morphological analysis. They revealed that Acanthamoeba was the dominant genus (77%) among all detected FLA, and the remaining detected amoebae belonged to Tetramitus and Stachyamoeba genera, or were unidentified. In this study, Acanthamoeba sp. was also detected in the majority of positive cultures (79.1%) and occurred significantly more frequently than other identified amoebae: Naegleria spp., H. vermiformis, P. bulliensis, and Paravahlkampfia sp. The differences between this study and the others may result from distinct climate or soil properties, or, in the case of the French study, may be caused by methodological differences. Other studies on the presence of different amoebae in soil used samples from outside Europe. In samples from Turkey, Iran, Tenerife, Santiago Island of Cape Verde, and the Canary Islands, Acanthamoeba sp. was the most prevalent FLA [17─19,21,23,24]. V. vermiformis was the most prevalent FLA in Northern Iran, Bolivia, and Guadeloupe [25─27]. Naegleria and Tetramitus genera dominated in Vietnam and Burkina Faso, respectively [22]. Acanthamoeba sp. seems to be a dominant genus in European soils, and there are considerable differences in FLA composition between sampling areas worldwide. Additional studies on the prevalence and distribution of FLA in soil are necessary, and this research provides valuable insights into the subject. Acanthamoeba spp., Naegleria spp., and V. vermiformis have frequently been found in soil [9,12,15,17─20,23─25,28]. However, the Hartmannellid genus Ptolemeba and the Valkhampfiidae amoebae Paravahlkampfia sp. and Naegleria other than N. fowleri have been rarely detected in environmental samples. Ptolemeba bulliensis and Ptolemeba noxubium have been isolated from soil and water in Mississippi, respectively [29]. P. bulliensis has also been found in the gills of rainbow trout (Oncorhynchus mykiss) in Russia [30] and in water from the Philippines (PP732398, GenBank, unpublished). Paravahlkampfia lenta has been isolated from soil samples collected from a Scottish farm [31], and Paravahlkampfia ustiana from soil samples collected in Thailand [28]. N. gruberi has been isolated from soil samples from the UK and California [32]. This study is the first to reveal the presence of N. galeacystis in soil, as well as N. gruberi and V. vermiformis in soil from Europe.
Studies on the prevalence of Acanthamoeba genotypes in European soils were conducted in Poland, Austria, the Netherlands, Hungary, and Sardinia. The composition of Acanthamoeba genotypes was different depending on the examined area. The T4 genotype was the only one detected in the soil [16] and sandboxes [33] in Poland, as well as in rhizosphere samples from Hungary [34]. This genotype dominated in soils from Austria [35] and Sardinia [36], and accounted for half of all the detected Acanthamoeba strains in soil from the Netherlands [36]. Additionally, the T2 genotype has been found in soil from Austria and the Netherlands [35,36], T16 in soil from the Netherlands, and T13 in soil from Sardinia [36]. In this study, the T4 and T13 genotypes were identified. The first one dominated; however, the difference was not statistically significant. The T4 genotype was the most common in the European soil samples examined so far, and the share of the remaining genotypes was different in each examined area. Additional studies are necessary to investigate whether T4 predominance is typical in European soils and whether other genotypes, not detected so far, occur. The share of FLA species and Acanthamoeba genotypes differed even between relatively close locations, such as central [16,33] and southern Poland (this study). The composition of FLA in soil, therefore, appears to be influenced by local conditions, such as soil properties. However, in this study, there were no significant differences in the occurrence of individual FLAs in different zones of the Babiogórski National Park.
Both Acanthamoeba genotypes identified in this study and V. vermiformis can cause keratitis, and the T4 Acanthamoeba genotype is an etiological agent of granulomatous amoebic encephalitis [6,37]. The Paravahlkampfia genus was previously considered a human pathogen [38─40]. The presence of these amoebae in the examined samples may indicate a potential health threat to individuals visiting the studied area. Tourists frequently visit the Babiogórski National Park, and maintaining hygiene, such as hand washing, is difficult in this location, which increases the risk of infection. Thermal-tolerant Acanthamoeba strains are considered potentially pathogenic [10,11]. All Acanthamoeba strains detected in this study were grown at a temperature of 28°C and 37°C. Half of the T4 strains (6/12) and 42,8% of the T13 strains (3/7) were grown at a temperature of 42°C. However, the study of Kahraman et al. [10] suggests that thermotolerance does not indicate the pathogenicity of Acanthamoeba.
4. Materials and Methods
4.1. Study Area and Soil Sampling
Babia Góra National Park is situated in the Western Carpathians and is a popular destination for tourists. Babia Góra is the highest peak in the park (1,725 meters above sea level), and mixed forests cover most of the park area. The main tree species growing in the park are common beech (Fagus sylvatica), Norway spruce (Picea abies), and silver fir (Abies alba). Fir and beech forests, including Norway spruce and sycamore (Acer pseudoplatanus), dominate the lower montane zone (up to 1,150 m). Norway spruce grows in the upper montane zone (up to 1,390 m) with an admixture of rowan (Sorbus aucuparia). The next zone (up to 1,650 m) is dominated by dwarf mountain pine (Pinus mugo). However, other species, such as dwarf forms of spruce, rowan, Silesian willow (Salix silesiaca), rock currant (Ribes petraeum), and mountain juniper (Juniperus communis subsp. alpina) are also present. Only low grasslands, mosses, and lichens grow in the Alpine zone (up to 1,725 m).
Forty soil samples (approximately 5.0 mL each) were collected in June 2024 from Babia Góra National Park and its surrounding areas in southern Poland (Figure 5). Thirty-two samples (1-15, 21-28, 31-37, 39, and 40) were taken from the lower montane zone, four samples (16, 29, 30, 38) from the upper montane zone, three samples (17-19) from the zone of dwarf mountain pine, and one sample (20) from the Alpine zone. The samples were taken directly from the ground surface along marked tourist trails. During the sampling, the air temperature oscillated between 15 and 20°C. The obtained material was placed in sterile Eppendorf tubes and stored at 4°C until further processing and analysis in the laboratory.
4.2. Amoebae Cultures and Thermal Tolerance Test
1g of each soil sample was seeded onto a Petri dish with NN-agar coated with heat-killed Escherichia coli. The agar plates were incubated at 28°C and examined daily under an optical microscope until amoebae were observed (up to 72h). Then, two sets of new Petri dishes were prepared as described above. Small pieces of agar (approximately 5 x 5 mm) with trophozoites were transferred onto new Petri dishes and placed top side down on the agar surface. One set was incubated at 37°C and the second at 42°C until amoebae were observed under the microscope (up to 72 hours). The amoebae obtained from cultures incubated at 37°C and 42°C were washed with sterile PBS buffer (1 ml per plate).
4.3. DNA Extraction from Soil and Cultures, and PCR Protocols
Two hundred microliters of PBS with washed amoebae from each agar plate were used for DNA extraction with the QIAamp DNA Mini Kit (Qiagen, Hilden, Germany). The Soil DNA Mini Kit (Syngen, Wrocław, Poland) was used for direct DNA extraction from soil samples (0.5 g of soil per isolation). DNA extractions were performed according to the manufacturer’s protocols. Five PCR protocols, as previously described by the authors, were used to amplify the SSU rRNA gene fragments and detect the presence of FLA in the examined samples (Table 5). Except for three primer pairs specific for a broad spectrum of FLA, genus-specific primers were used for Acanthamoeba and the former genus Hartmannella, as they are common in the environment [12] and have been isolated earlier from environmental samples in Poland [41]. PCR products were separated by electrophoresis in a 1.5% agarose gel, stained with ethidium bromide, and visualized under UV light.
4.4. Sequencing, Genotyping, and Phylogenetic Analysis
Both strands of all obtained PCR products were sequenced using the amplification primers (Table 1). The sequencing was performed at Macrogen Europe (Amsterdam, the Netherlands). The obtained sequences were compared with other homologous sequences deposited in the GenBank database using the Basic Local Alignment Search Tool (BLAST) at the National Center for Biotechnology Information. Separate alignments were performed for sequences of different genera of FLA, using the MUSCLE algorithm (the MEGA 12 software), which allowed for comparison to each other and to other homologous sequences from GenBank. The ends of the sequences in the alignments were trimmed to form blunt ends. The multiple alignment for the Acanthamoeba genus covered nucleotides corresponding to positions 720 to 1,300 of the Acanthamoeba sp. genotype T4 sequence with GenBank accession number AY702983 (JDP1/JDP2 primer set). For the Naegleria genus, the multiple alignment covered nucleotides corresponding to positions 680-1343 (Ami6F1/Ami9R primer set), 756-1476 (FLA-F/FLA-R primer set), and 1277-1662 (AmeF977/AmeR1534 primer set) of the N. gruberi sequence with GenBank accession number OR769034. The phylogenetic trees were constructed using Mega 12 software, based on a multiple alignment and the neighbor-joining statistical method with the Kimura 2-parameter model, with 1,000 bootstrap samples [47]. Different fragments of the same sequences from GenBank were used to construct the trees based on the 18S rRNA sequence of Naegleria spp. (Figure 2, Figure 3 and Figure 4). The results of the sequence comparison using BLAST and analysis of the phylogenetic trees' topology were used to determine the genotype of Acanthamoeba and the species of Naegleria strains examined in this study. The genus or species of the remaining amoebae was determined based on the sequence comparison using BLAST. Phylogenetic trees for V. vermiformis, P. bulliensis, and Paravahlkampfia sp. were not constructed, as all obtained V. vermiformis sequences represent the same species and are all identical to those in the GenBank database. Sequences of the Ptolemeba and Paravahlkampfia genera deposited in GenBank are too few to use for phylogenetic analysis. The sequences analyzed in this study have been deposited in the GenBank database under accession numbers: PV867401-PV867419 (Acanthamoeba spp.), PV867462-PV867474 (Naegleria spp.), PV867476-PV867480 (V. vermiformis), PV867809 (P. bulliensis), and PV873343 (Paravahlkampfia sp.).
4.5. Statistical Analysis
Statistical analyses were performed using a chi-squared test to investigate the differences in the prevalence of various amoebae and the prevalence of amoebae in samples collected from specific zones in the Babiogórski National Park. Statistical significance was defined as p < 0.05. The Statistica 13.3 software (StatSoft Inc., USA) was used for the analysis.
5. Conclusions
The knowledge of FLA diversity in European soils is insufficient, and more studies regarding this topic are needed. This study revealed that the Acanthamoeba genus (T4 and T13 genotypes) dominates among FLA occurring in soil from the Babiogórski National Park and its surroundings, followed by Naegleria spp. and V. vermiformis. It also reports the isolation of Paravahlkampfia sp., P. bulliensis, and N. gruberi, which are rarely detected in soil, as well as the first-time isolation of N. galeacystis from soil and N. gruberi and V. vermiformis from soil collected in Europe. Some of the detected FLAs are potentially pathogenic and may pose a health threat to humans. Among the used primer sets, genus-specific primers (JDP1/JDP2 and HARTfor/HARTrev) are more sensitive than the three pairs specific for a wide range of FLA: FLA-F/FLA-R, Ami6F1/Ami9R, and AmeF977/AmeR1534. It is necessary to design new, sensitive, and specific primers enabling the detection, species identification, and genotyping of FLA in soil.
Funding
Co-financed by the Minister of Science under the "Regional Excellence Initiative" Program for 2024-2027 (RID/SP/045/2024/01).
Data Availability Statement
All data generated or analyzed in this study are included in this published article. The accession numbers of the DNA sequences obtained in this study are provided in the Materials and Methods section and are available in the GenBank database.
Conflicts of Interest
The author declares no conflicts of interest.
References
- Ali, M.; Rice, C.A.; Byrne, A.W.; Paré, P.E.; Beauvais, W. Modelling dynamics between free-living amoebae and bacteria. Environ. Microbiol. 2024, 26, e16623. [CrossRef]
- Otero-Ruiz, A.; Gonzalez-Zuñiga, L.D.; Rodriguez-Anaya, L.Z.; Lares-Jiménez, L.F.; Gonzalez-Galaviz, J.R.; Lares-Villa, F. Distribution and current state of molecular genetic characterization in pathogenic free-living amoebae. Pathogens 2022, 11, 1199. [CrossRef]
- Siddiqui, R.; Makhlouf, Z.; Khan, N.A. The increasing importance of Vermamoeba vermiformis. J. Eukaryot. Microbiol. 2021, 68, e12857. [CrossRef]
- Dinda, S.K.; Hazra, S.; De, A.; Datta, A.; Das, L.; Pattanayak, S.; Kumar, K.; Dey, M.D.; Basu, A.; Manna, D. Amoebae: beyond pathogens – exploring their benefits and future potential. Front. Cell. Infect. Microbiol. 2024, 14, 1518925. [CrossRef]
- Fan, S.; Shen, Y.; Qian, L. Social life of free-living amoebae in aquatic environment – comprehensive insights into interactions of free-living amoebae with neighboring microorganisms. Front. Microbiol. 2024, 15, 1382075. [CrossRef]
- Aykur, M.; Selver, O.B.; Dagci, H.; Palamar, M. Vermamoeba vermiformis as the ethiological agent in a patient with suspected non-Acanthamoeba keratitis. Parasitol. Res. 2024, 123, 323. [CrossRef]
- Salazar-Ardiles, C.; Valencia, K.P.; Andrade, D.C. Amoebas: the omnipotent organism and silent assassin. Mol Biol Rep. 2025, 52, 160. [CrossRef]
- Cardoso, I.R.; de Lima, C.S.; dos Reis, R.B.; Pinto, A.C.A.; Pissinatti, T.; Kugelmeier, T.; da Costa Neto, S.F.; da Silva, F.A.; Santos, H.L.C. Occurrence of free-living amoebae in non-human primate gut. Trop. Med. Infect. Dis. 2024, 9, 108. [CrossRef]
- Zurita-Artaloitia, J.M.; Riviera, J; Vinuesa, P. Extensive cryptic diversity and ecological associations uncovered among Mexican and global collections of Naegleria and Vermamoeba species by 18S ribosomal DNA, internal transcribed spacer, and cytochrome oxidase subunit I sequence analysis. Microbiol. Spectr. 2023, 11(2), 03795–22. [CrossRef]
- Kahraman, M.; Polat, Z.A. Are thermotolerant and osmotolerant characteristics of Acanthamoeba species an indicator of pathogenicity? Türkiye Parazitol Derg. 2024, 48(1), 15–20. [CrossRef]
- Lorenzo-Morales, J.; Khan, N.A.; Walochnik, J. An update on Acanthamoeba keratitis: diagnosis, pathogenesis and treatment. Parasite 2015, 22, 10. [CrossRef]
- Chaúque, B.J.M.; da Silva, T.C.B.; dos Santos, D.L.; Benitez, G.B.; Chaúque, L.G.H.; Benetti, A.D; Zanette, R.A.; Rott, M.B. Global prevalence of free-living amoebae in solid matrices – a systematic review with meta-analysis. Acta Trop. 2023, 247, 107006. [CrossRef]
- Bass, D.; Stentiford, G.D.; Littlewood, D.T.J.; Hartikainen, H. Diverse applications of environmental DNA methods in parasitology. Trends Parasitol. 2015, 31(10), 499–513. https://dx.doi.org/10.1016/j.pt.2015.06.013.
- Sengupta, M.E.; Lynggaard, C.; Mukaratriwa, S.; Vennervald, B.J.; Stensgaard, A.S. Environmental DNA in human and veterinary parasitology – current applications and future prospects for monitoring and control. Food and Waterborne Parasitol. 2022, 29, e00183. [CrossRef]
- Albuquerque, P.; Nicola, A.M.; Almeida, D.; Magnabosco, G.; da Silveira Derengowski, L.; Crisóstomo, L.S.; Xavier, L.C.G.; de Oliveira Frazão, S.; Guilhelmelli, F.; de Oliveira, M.A.; do Nascimento Dias, J.; Hurtado, F.A.; de Melo Teixeira, M.; Guimarães, A.J.; Paes, H.C.; Bagagli, E.; Felipe, M.S.S.; Casadevall, A.; Silva-Pereira, I. A hidden battle in dirt: Soil amoebae interactions with Paracoccidioides spp. NTDs 2019, 13(10), e0007742. [CrossRef]
- Hendiger-Rizo, E.B.; Chmielewska-Jeznach, M.; Poreda, K.; Liendo, A.R.; Koryszewska-Bagińska, A.; Olędzka, G.; Padzik, M. Potentially pathogenic free-living amoebae isolated from soil samples from Warsaw parks and squares. Pathogens 2024, 13, 895. [CrossRef]
- Pérez-Pérez, P.; Reyes-Batlle, M.; Morchón, R.; Piñero, J.E.; Lorenzo-Morales, J. Isolation and molecular identification of pathogenic free-living amoebae from environmental samples in Tenerife, Canary Islands. ACS EST Water 2025, 5, 2861–2869. [CrossRef]
- Sousa-Ramos, D.; Reyes-Batlle, M.; Bellini, N.K.; Rodriguez-Expósito, R.; Piñero.; J.E.; Lorenzo-Morales, J. Free-living amoebae in soil samples from Santiago Island, Cape Verde. Parasitol. Res. 2021, 121, 2399–2404. [CrossRef]
- Sousa-Ramos, D.; Reyes-Batlle, M.; Bellini, N.K.; Rodriguez-Expósito, R.; Martin-Real, C.; Piñero, J.E.; Lorenzo-Morales, J. Pathogenic free-living amoebae from water sources in Cape Verde. Microorganisms 2022, 9, 1460. [CrossRef]
- Reyes-Batlle, M.; Wagner, C.; Zamora-Herrera, J.; Vargas-Mesa, A.; Sifaoui, I.; González, A.C.; López-Arencibia, A.; Valladares, B.; Martinez-Carretero, E.; Piñero, J.E., Lorenzo-Morales, J. Isolation and molecular identification of Vermamoeba vermiformis strains from soil sources in El Hierro Island, Canary Islands, Spain. Curr. Microbiol. 2016, 73, 104–107. [CrossRef]
- Reyes-Batlle, M.; Diaz, F.J.; Sifaoui, I.; Rodriguez-Expósito, R.L.; Rizo-Liendo, A.; Piñero, J.E.; Lorenzo-Morales, J. Free living amoebae isolation in irrigation waters and soils of an insular arid agroecosystem. Sci. Total Environ. 2021, 753, 141833. [CrossRef]
- Denet, E.; Coupat-Goutaland, B.; Nazaret, S.; Pélandakis, M.; Favre-Bonté, S. Diversity of free-living amoebae in soils and their associated human opportunistic bacteria. Parasitol. Res. 2017,116, 3151–3162. [CrossRef]
- Aykur, M.; Dagci, H. Molecular identification of Acanthamoeba spp., Balamuthia mandrillaris and Naegleria fowleri in soil samples using quantitative real-time PCR assay in Turkey; Hidden danger in the soil! Acta Trop. 2023, 244, 106956. [CrossRef]
- Pazoki, H.; Niyyati, M.; Javanmard, E.; Iasjerdi, Z.; Spotin, A.; Mirjalali, H.; Behravan, M.R. Isolation and phylogenetic analysis of free-living amoebae (Acanthamoeba, Naegleria, and Vermamoeba) in the farmland soils and recreational places in Iran. Acta Parasitol. 2020, 65, 36–43. [CrossRef]
- Pérez-Pérez, P.; Artigas, P.; Reyes-Batlle, M.; Córdoba-Lanús, E.; Rodriguez-Expósito, R.L.; Cuervo, P.F.; Dominguez-de Barros, A.; Garcia-Pérez, O.; Valero, M.A.; De Elias, A.; Anglés, R.; Mas-Coma, S.; Piñero, J.E.; Bargues, M.D.; Lorenzo-Morales, J. Potentially pathogenic free-living amoebae at very high altitude: Detection by multiplex qPCR in the Northern Altiplano fascioliasis hyperendemic area in Bolivia. OH 2025, 20, 100985. [CrossRef]
- Reynaud, Y.; Ducat, C.; Talarmin, A.; Marcelino, I. Cartography of free-living amoebae in soil in Guadeloupe (French West Indies) using DNA metabarcoding. Pathogens 2020, 9, 440. [CrossRef]
- Tanzifi, A.; Moghaddam, Y.; Dodangeh, S.; Daryani, A.; Shahabeddin, S.; Gholami, S.; Hosseini, S.A.; Chegeni, T.N.; Hosseininejad, Z. Detection and molecular characterization of potentially pathogenic free-living amoebae from recreational and public soils in Mazandaran, Northern Iran. Iran J Parasitol. 2021, 16(2), 295–304.
- Noinarin, P.; Chareonsudjai, P.; Wangsomnuk, P.; Wongratanacheewin, S.; Chareonsudjai, S. Environmental free-living amoebae isolated from soil in Khon Kaen, Thailand, antagonize Burkholderia pseudomallei. PLoS ONE 2016, 11(11), e0167355. [CrossRef]
- Watson, P.M.; Sorrell, S.C.; Brown, M.W. Ptolemeba n. gen., a novel genus of Hartmannellid amoebae (Tubulinea, Amoebozoa); with an emphasis on the taxonomy of Saccamoeba. J. Eukaryot. Microbiol. 2014, 61, 611–619. [CrossRef]
- Kudryavtsev, A.; Volkova, E.; Parshukov, A. Ptolemeba bulliensis Watson et al. 2014 (Amoebozoa, Tubulinea) from freshwater NGD-affected rainbow trout (Oncorhynchus mykiss Walbaum, 1792) gills tolerates brackish water conditions. J. Fish Dis. 2025, 0, e14132. [CrossRef]
- Brown, S.; De Jonckheere, J.F. Isolation of a new valkhampfiid amoeba from soil: Paravahlkampfia lenta n. sp. Eur. J. Protistol. 2004, 40, 289–294. [CrossRef]
- McLaughlin, G.L.; Brandt, F.H.; Visvesvara, G.S. Restriction fragment length polymorphisms of the DNA of selected Naegleria and Acanthamoeba amebae. J. Clin. Microbiol. 1988, 26(9), 1655–1658.
- Cholewiński, M.; Solarczyk, P.; Derda, M.; Wojtkowiak-Giera, A.; Hadaś, E. Presence of potential pathogenic genotypes of free-living amoebae isolated from sandboxes in children’s playgrounds. Folia Parasitol. 2015, 62: 064. [CrossRef]
- Orosz, E.; Posta, K. Genotyping of Acanthamoeba spp. from rizosphere in Hungary. Acta Microbiol Immunol Hung. 2020, 67(3), 171–175. [CrossRef]
- Schmitz-Esser, S.; Toenschoff, E.R.; Haider, S.; Heinz, E.; Hoenninger, V.M.; Wagner, M.; Horn, M. Diversity of bacterial endosymbionts of environmental Acanthamoeba isolates. Appl. Environ. Microbiol. 2008, 74(18), 5822–5831. [CrossRef]
- Geisen, S.; Fiore-Donno, A.M.; Walochnik, J.; Bonkowski, M. Acanthamoeba everywhere: high diversity of Acanthamoeba in soils. Parasitol. Res. 2014, 113, 3151–3158. [CrossRef]
- Wang, Y.; Jiang, L.; Zhao, Y.; Ju, X.; Wang, L.; Jin, L.; Fine, R.D.; Li, M. Biological characteristics and pathogenicity of Acanthamoeba. Front. Microbiol. 2023, 14, 11470077. [CrossRef]
- Garcia, A.; Goñi, P.; Clavel, A.; Lobez, S.; Fernandez, M.T.; Ormad, M.P. Potentially pathogenic free-living amoebae (FLA) isolated in Spanish wastewater treatment plants. Environ. Biol. Rep. 2011, 3(5): 622–626. [CrossRef]
- Ozkoc, S.; Tuncay, S.; Delibas, S.B.; Akisu, C.; Ozbek, Z.; Durak, I.; Walochnik, J. Identification of Acanthamoeba genotype T4 and Paravahlkampfia sp. from two clinical isolates. J. Med. Microbiol. 2008, 57. [CrossRef]
- Visvesvara, G.S.; Sriram, R.; Qvarnstrom, Y.; Bandyopadhyay, K.; da Silva, A.J.; Pieniążek, N.J.; Cabral, G.A. Paravahlkampfia francinae n. sp. masquerading as an agent of primary amoebic meningocephalitis. J. Eukaryot. Microbiol. 2009, 56(4), 357–366 https://doi.org/10.1111/j.1550-7408.2009.00410.x.
- Adamska, M.; Leońska-Duniec, A.; Łanocha, N.; Skotarczak, B. Thermophilic, potentially pathogenic amoebae isolated from natural water bodies in Poland and their molecular characterization. Acta Parasitol. 2014, 59(3), 433–441. [CrossRef]
- Schroeder, J.M.; Booton, G.C.; Hay, J.; Niszl, I.A.; Seal, D.V.; Markus, M.B.; Fuerst, P.A.; Byers, T.J. Use of subgenic 18S ribosomal DNA PCR and sequencing for genus and genotype identification and Acanthamoebae from humans with keratitis and from sewage sludge. J. Clin. Microbiol. 2001, 39(5), 1903–1911. [CrossRef]
- Scheikl, U.; Sommer, R.; Kirschner, A.; Rameder, A.; Schrammel, B.; Zweimüller, I.; Wesner, W.; Hinker, M.; Walochnik, J. Free-living amoebae (FLA) co-occurring with legionellae in industrial waters. Eur. J. Protistol. 2014, 50, 422–429. https://dx.doi.org/10.1016/j.ejop.2014.04.002.
- Tsvetkova, N.; Schild, M.; Panaiotov, S.; Kurdova-Mintcheva, R.; Gottstein, B.; Walochnik, J.; Aspӧck, H.; Lucas, M.S.; Müller, N. The identification of free-living environmental isolates of amoebae from Bulgaria. Parasitol. Res. 2004, 92, 405–413. [CrossRef]
- Thomas, V.; Herrera-Rimann, K.; Blanc, D.S.; Greub, G. Biodiversity of amoebae and amoeba-resisting bacteria in a hospital water network. Appl. Environ. Microbiol. 2006, 72(4), 2428–2438. [CrossRef]
- Liang, S.-Y.; Ji, D.-R.; Hsia, K.-T.; Hung, C.-C.; Sheng, W.-H.; Hsu, B.-M.; Chen, J.-S.; Wu, M.-H.; Lai, C.-H.; Ji, D.-D. Isolation and identification of Acanthamoeba species related to amoebic encephalitis and nonpathogenic free-living amoeba species from the rice field. J. Appl. Microbiol. 2010, 109, 1422–1429. [CrossRef]
- Kumar, S.; Stecher, G.; Suleski, M.; Sanderford, M.; Sharma, S.; Tamura, K. MEGA12: Molecular Evolutionary Genetic Analysis Version 12 for Adaptive and Green Computing. Mol. Biol. Evol. 2024, 41, 1–9. [CrossRef]
Figure 1.
Phylogenetic tree constructed using the neighbor-joining statistical method and the Kimura 2-parameter model with 1000 bootstrap sampling, based on forty-five SSU rRNA sequences of Acanthamoeba. The forty sequences with accession numbers are from GenBank, and the remaining five sequences were obtained in this study using the JDP1/JDP2 primer set. Identical sequences are counted as a single sequence. Only bootstrap values ≥50 are shown.
Figure 1.
Phylogenetic tree constructed using the neighbor-joining statistical method and the Kimura 2-parameter model with 1000 bootstrap sampling, based on forty-five SSU rRNA sequences of Acanthamoeba. The forty sequences with accession numbers are from GenBank, and the remaining five sequences were obtained in this study using the JDP1/JDP2 primer set. Identical sequences are counted as a single sequence. Only bootstrap values ≥50 are shown.

Figure 2.
Phylogenetic tree constructed using the neighbor-joining statistical method and the Kimura 2-parameter model with 1000 bootstrap sampling, based on twenty-four SSU rRNA sequences of the Naegleria genus. Twenty-one sequences with accession numbers are from GenBank, and the remaining three sequences were obtained in this study using the FLA-F/FLA-R primer set. Identical sequences are counted as a single sequence. Only bootstrap values ≥50 are shown.
Figure 2.
Phylogenetic tree constructed using the neighbor-joining statistical method and the Kimura 2-parameter model with 1000 bootstrap sampling, based on twenty-four SSU rRNA sequences of the Naegleria genus. Twenty-one sequences with accession numbers are from GenBank, and the remaining three sequences were obtained in this study using the FLA-F/FLA-R primer set. Identical sequences are counted as a single sequence. Only bootstrap values ≥50 are shown.

Figure 3.
Phylogenetic tree constructed using the neighbor-joining statistical method and the Kimura 2-parameter model with 1000 bootstrap sampling, based on twenty-three SSU rRNA sequences of the Naegleria genus. Twenty-one sequences with accession numbers are from GenBank, and the remaining two sequences were obtained in this study using the Ami6F1/Ami9R primer set. Identical sequences are counted as a single sequence. Only bootstrap values ≥50 are shown.
Figure 3.
Phylogenetic tree constructed using the neighbor-joining statistical method and the Kimura 2-parameter model with 1000 bootstrap sampling, based on twenty-three SSU rRNA sequences of the Naegleria genus. Twenty-one sequences with accession numbers are from GenBank, and the remaining two sequences were obtained in this study using the Ami6F1/Ami9R primer set. Identical sequences are counted as a single sequence. Only bootstrap values ≥50 are shown.

Figure 4.
Phylogenetic tree constructed using the neighbor-joining statistical method and the Kimura 2-parameter model with 1000 bootstrap sampling, based on twenty-three SSU rRNA sequences of the Naegleria genus. Twenty-one sequences with accession numbers are from GenBank, and the remaining two sequences were obtained in this study using the AmeF977/AmeR1534 primer set. Identical sequences are counted as a single sequence. Only bootstrap values ≥50 are shown.
Figure 4.
Phylogenetic tree constructed using the neighbor-joining statistical method and the Kimura 2-parameter model with 1000 bootstrap sampling, based on twenty-three SSU rRNA sequences of the Naegleria genus. Twenty-one sequences with accession numbers are from GenBank, and the remaining two sequences were obtained in this study using the AmeF977/AmeR1534 primer set. Identical sequences are counted as a single sequence. Only bootstrap values ≥50 are shown.
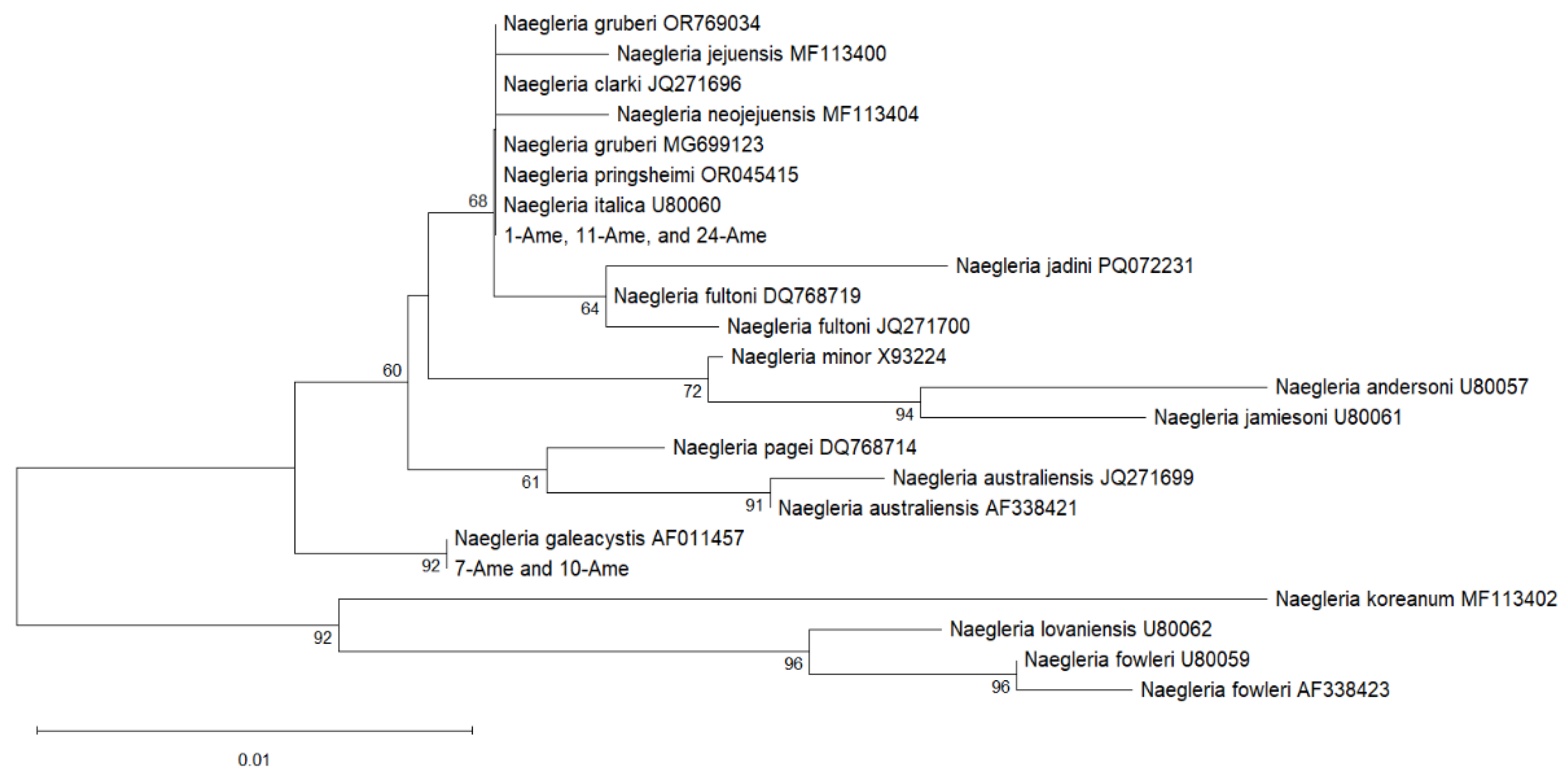
Figure 5.
Collection sites of soil samples in the Babiogórski National Park and its surroundings.
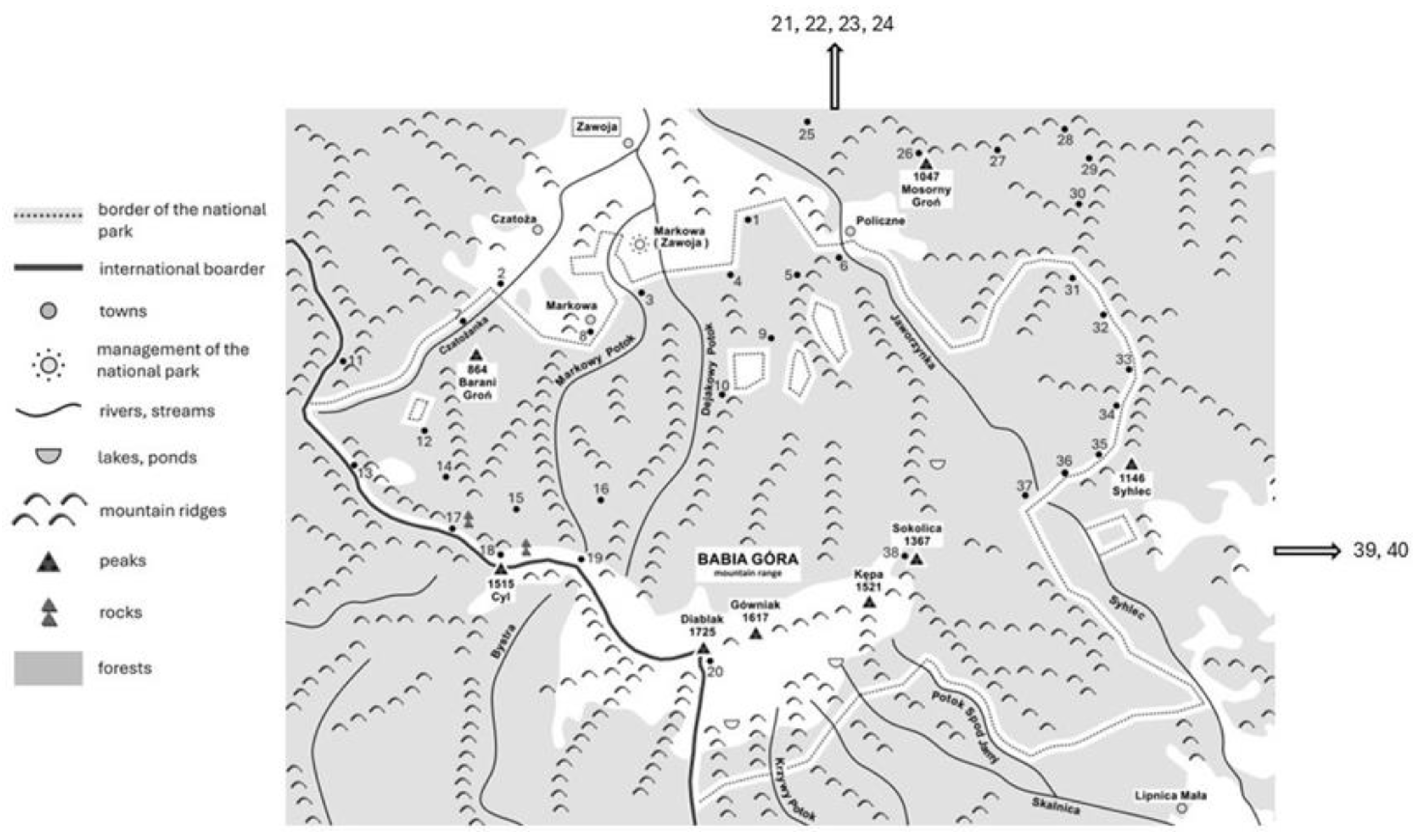

Table 1.
The results of amoebae cultures at 42°C, PCRs, and sequencing for the 26 strains that were grown at 30°C and 37°C.
Table 1.
The results of amoebae cultures at 42°C, PCRs, and sequencing for the 26 strains that were grown at 30°C and 37°C.
| Sample number | Culture at 42°C |
PCR and sequencing results with individual primer pairs | ||||
|---|---|---|---|---|---|---|
| JDP1/JDP2 | FLA-F/FLA-R | Ami6F1/Ami9R | AmeF977/ AmeR1534 |
HARTfor/ HARTrev |
||
| 1. | Naegleria sp. | |||||
| 2. | + | Acanthamoeba T13 | V. vermiformis | |||
| 4. | Acanthamoeba T4 | Paravahlkampfia sp. | ||||
| 6. | + | Acanthamoeba T4 | ||||
| 7. | N. galeacystis | N. galeacystis | ||||
| 8. | + | N. gruberi | N. gruberi | |||
| 10. | Acanthamoeba T13 | N. galeacystis | N. galeacystis | |||
| 11. | N. gruberi | N. gruberi | Naegleria sp. | |||
| 12. | Acanthamoeba T4 | |||||
| 15. | + | Acanthamoeba T4 | ||||
| 19. | + | Acanthamoeba T4 | V. vermiformis | V. vermiformis | V. vermiformis | |
| 21. | + | Acanthamoeba T4 | ||||
| 23. | Acanthamoeba T13 | |||||
| 24. | + | N. gruberi | N. gruberi | Naegleria sp. | ||
| 27. | Acanthamoeba T4 | |||||
| 28. | Acanthamoeba T13 | P. bulliensis | ||||
| 29. | + | Acanthamoeba T4 | ||||
| 31. | + | Acanthamoeba T4 | ||||
| 32. | + | Acanthamoeba T13 | V. vermiformis | |||
| 33. | + | Acanthamoeba T13 | ||||
| 35. | + | Acanthamoeba T13 | ||||
| 37. | Acanthamoeba T4 | |||||
| 39. | Acanthamoeba T4 | |||||
| 40. | Acanthamoeba T4 | |||||
Table 2.
The results of the sequence alignment of the sequences obtained in this study and the sequences from GenBank using the BLAST tool for the Acanthamoeba genus (JDP1/JDP2 primer set).
Table 2.
The results of the sequence alignment of the sequences obtained in this study and the sequences from GenBank using the BLAST tool for the Acanthamoeba genus (JDP1/JDP2 primer set).
| The sequences obtained in this study (sample number and primer set) | The sequences from the GenBank database – accession number, isolation source, and country | Identity |
|---|---|---|
| Identical sequences: 12-JDP, 15-JDP, 19-JDP, 27-JDP, 29-JDP, 31-JDP | ON408415 Acanthamoeba T4 isolated from soil (USA) | 100% |
| 6-JDP | KT985967 Acanthamoeba T4 isolated from soil (Iran) KU894800 Acanthamoeba sp. isolated from water (Uganda) MZ404332 Acanthamoeba sp. isolated from a patient with keratitis (Brazil) |
100% |
| 39-JDP | KC164232 Acanthamoeba sp. isolated from compost (Switzerland) PQ431522, PQ431528 Acanthamoeba sp. isolated from water (France) AF019060 A. hatchetti T4 |
100% 99.48% |
| Identical sequences: 4-JDP, 21-JDP, 37-JDP, 40-JDP | JQ669659 Acanthamoeba T4 isolated from soil (USA) KT892868 Acanthamoeba T4 isolated from soil (Chile) FJ807650 Acanthamoeba sp. isolated from marsh sediment (Austria) LR813621 A. castellani isolated from coprolite (Belgium) KC164234 A. castellani isolated from compost (Switzerland) MN700280 Acanthamoeba T4 isolated from water (Malaysia) MT378239 Acanthamoeba T4 isolated from water (Iran) |
100% |
| Identical sequences: 2-JDP, 10-JDP, 23-JDP, 28-JDP, 32-JDP, 33-JDP, 35-JDP | MZ686722 Acanthamoeba T13 isolated from soil (Malaysia) | 99.48% |
Table 3.
The results of the sequence alignment of the sequences obtained in this study and the sequences from GenBank using the BLAST tool for the Naegleria genus (FLA-F/FLA-R, AmeF977/AmeR1534, and Ami6F1/Ami9R primer sets).
Table 3.
The results of the sequence alignment of the sequences obtained in this study and the sequences from GenBank using the BLAST tool for the Naegleria genus (FLA-F/FLA-R, AmeF977/AmeR1534, and Ami6F1/Ami9R primer sets).
| The sequences obtained in this study (sample number and primer set*) | The sequences from the GenBank database – accession number, isolation source, and country | Identity |
|---|---|---|
| Identical sequences: 7-FLA and 10-FLA | DQ768717 Naegleria sp. isolated from fish (Czech Republic) PP174311 Naegleria sp. |
100% |
| Identical sequences: 8-FLA and 24-FLA; Identical sequences: 8-Ami and 24-Ami |
OR769034 N. gruberi isolated from soil (USA) MG699123, AB298288 – N. gruberi |
100% |
| 11-FLA/11-Ami | 98.64%/ 99.17% |
|
| Identical sequences: 7-Ame and 10-Ame | AF011457 N. galeacystis DQ768717, DQ768718 – Naegleria sp. isolated from fishes |
100% |
| Identical sequences: 1-Ame, 11-Ame, 24-Ame |
OR769034 N. gruberi isolated from soil (USA) MG699123, AB298288 – N. gruberi AF338417, AF338419, JQ271691, JQ271692, JQ271697, JQ271705 – N. clarki isolated from fishes; JQ271704 – N. clarki isolated from sediment (Czech Republic) OR045415 – N. pringsheimi |
100% |
*FLA - FLA-F/FLA-R, Ame - AmeF977/AmeR1534, Ami - Ami6F1/Ami9R.
Table 4.
The results of the sequence alignment of the sequences obtained in this study and the sequences from GenBank using the BLAST tool for Vermamoeba vermiformis (FLA-F/FLA-R, AmeF977/AmeR1534, and HARTfor/HARTrev primer sets).
Table 4.
The results of the sequence alignment of the sequences obtained in this study and the sequences from GenBank using the BLAST tool for Vermamoeba vermiformis (FLA-F/FLA-R, AmeF977/AmeR1534, and HARTfor/HARTrev primer sets).
| The sequences obtained in this study (sample number and primer set*) |
The sequences from the GenBank database – accession number, isolation source, and country | Identity |
|---|---|---|
| 19-FLA | KU746978 V. vermiformis isolated from soil (Spain) KP792383, KP792388 V. vermiformis isolated from water (Italy) MF112024 V. vermiformis isolated from water (Pakistan) JQ271688 H. vermiformis isolated from a tapeworm (Croatia) HM363627 Hartmannella sp. isolated from a fish (Germany) |
100% |
| 19-AME | LC764480 V. vermiformis isolated from soil (Japan) LN650671 V. vermiformis isolated from water (Italy) JQ271687 V. vermiformis isolated from a fish (Czech Republic) |
100% |
| Identical sequences: 2-HART, 19-HART, 32-HART | PP732395, PP732396 V. vermiformis isolated from water (Philippines) | 100% |
*FLA - FLA-F/FLA-R, Ame - AmeF977/AmeR1534, HART - HARTfor/HARTrev.
Table 5.
Primer sets used in this study for FLA detection.
| The primer set | Detecting FLA | The amplifying fragment of the 18S rRNA gene and the product size | Hybridization temperature | References |
|---|---|---|---|---|
| JDP1/JDP2 | Acanthamoeba spp. | 897-1358 bp of A. castellani sequence (U07400); 462 bp | 55°C | [42] |
| HARTfor/HARTrev | Former genus Hartmannella | 562-1095 bp of V. vermiformis sequence (EU137741); 534 bp | 58°C | [43] |
| FLA-F/FLA-R | All FLA except Balamuthia and Sappinia | 631-1614 bp of A. castellani sequence (U07400); 984 bp | 55°C | [44] |
| Ami6F1/Ami9R | All amoebae | 641-1468 bp of A. castellani sequence (U07400); 828 bp | 55°C | [45] |
| AmeF977/AmeR1534 | All amoebae | 1179-1829 bp of A. castellani sequence (U07400); 651 bp | 60°C | [46] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



