
Submitted:
14 July 2025
Posted:
15 July 2025
You are already at the latest version
Abstract
Background/Objectives: This review explores whether astrological character typologies—rooted in symbolic archetypes—correlate with measurable patterns in personality psychology, psychiatric epidemiology, and developmental chronobiology. Methods: Drawing on over 100 peer-reviewed studies across psychology, psychiatry, neuroendocrinology, and chronobiology, we examined data on birth-month effects, seasonal neurodevelopment, personality inventories (e.g., Big Five), and psychiatric incidence. These were systematically cross-compared with astrological archetypes to evaluate symbolic alignment with biological traits. Results: Statistically significant associations were observed between season-of-birth and neuropsychiatric outcomes—for example, higher schizophrenia rates in winter births and increased ADHD diagnoses in spring—echoing traditional astrological depictions of psychological imbalance and stimulation. Fire-sign analogues (e.g., Aries, Leo, Sagittarius) showed correlations with extraversion and novelty-seeking, while water-sign types aligned with emotional sensitivity and affective vulnerability. Perinatal light exposure and circadian hormone cycles (e.g. melatonin, dopamine) appear to mediate these patterns. Conclusions: Astrological archetypes may symbolically reflect underlying biopsychological rhythms—capturing trends in temperament, personality structure, and psychiatric risk shaped by seasonal and chronobiological factors.
Keywords:
astrology
; personality traits
; typologies
; birth month
; seasonality of birth
; chronobiology
; psychiatric disorders
; mental health
; neurodevelopment
; epigenetics
; zodiac signs
; MBTI
; psychometrics
; integrative medicine
1. Introduction
1.1. Background
Astrology, often dismissed as metaphysical lore,
may in fact encode a symbolic system of characterological archetypes with deep
psychological, biological, and even neurocognitive relevance[1–3]. Rooted in ancient traditions yet
psychologically formalized by Carl Jung, astrological typology aligns with the
collective unconscious through elemental theory—Fire, Earth, Air, and
Water—representing motivational and affective constellations of human behavior [4]. These four elements parallel modern constructs
in temperament theory (e.g., choleric, melancholic) and are echoed in
neuroscience via dominant affective circuits, such as dopaminergic
novelty-seeking (Fire), serotonergic regulation (Water), or prefrontal-executive
grounding (Earth) [5,6].
Carl Jung viewed astrology not as superstition, but
as a pre-modern psychology encoded in the language of the stars—a symbolic
system reflecting projections of the unconscious onto cosmic order. In his
letters to Freud and others [4,7], Jung
emphasized astrology’s capacity to mirror personality dynamics and
synchronicity patterns, forming a bridge between inner psychodynamics and outer
temporal patterns. Freud himself, while less enthusiastic, acknowledged the
complex role of archetypal symbolism in the psyche, laying the groundwork for
modern typological approaches [8,9].
1.2. Modern Relevance and Research Gap
Astrology’s persistent influence among
decision-makers in fields such as finance, fashion, and politics underscores
its cultural relevance. Far from obsolete, it continues to function as a
symbolic system for interpreting timing, temperament, and strategic choice.
Historically, astrology was central to medicine [21–24]. The Hippocratic-Galenic tradition viewed
celestial patterns as influencing health through the balance of humors, lunar
cycles, and organ vitality [25,26]. In this
ancient system, each zodiac sign governed specific body parts and physiological
functions—a notion not far removed from today’s exploration of chronobiology,
endocrine rhythms [27,28], and seasonal gene
expression [29–32]. Recent
chronobiological studies (see Section 4.4) demonstrate that melatonin-serotonin cycles influenced
by perinatal light exposure shape long-term emotional and cognitive profiles
[33–37].
Despite its cultural marginalization, astrology’s
intuitive typology may fill a critical gap in contemporary psychiatry and
psychogenetics. Modern personality science has made significant strides in
identifying heritable patterns of behavior (e.g., through GWAS [38–40] and polygenic scores [41–43]), but struggles to explain the symbolic,
qualitative complexity of the psyche. Likewise, psychiatry’s diagnostic systems
(e.g., DSM-5) remain symptom-based, often neglecting the developmental,
archetypal, and seasonal roots of character and cognition [44,45].
Astrology may also represent a forgotten limb of
cosmophysical science—an observational model for understanding how large-scale
celestial systems (e.g., solar activity, planetary orbits, lunar cycles)
modulate biological rhythms on Earth [46–51].
Rather than invoking mysticism, this view locates astrology within the spectrum
of biophysical resonance, phase-locked entrainment, and temporal
patterning—principles foundational to chronobiology and systems neuroscience.
This review explores the hypothesis that astrology
encodes biopsychological patterns that can be statistically and biologically
validated. By synthesizing data from peer-reviewed studies on birth timing,
personality traits, psychiatric vulnerability, and chrononeuroendocrinology, we
examine whether astrological archetypes reflect reproducible trends in
neurodevelopment and behavior. In particular, we investigate correlations
between season-of-birth and psychiatric disorders (e.g., schizophrenia, ADHD), zodiac
analogues and personality traits, and neurocognitive outcomes modulated by
circadian and hormonal cycles.
We propose that astrology functions as a symbolic
interface—a culturally derived system encoding biopsychological patterns
through archetypes, timing, and temperament. Rather than causality, its
structure mirrors developmental and neuroendocrine rhythms, warranting renewed
transdisciplinary attention.
2. Theoretical Background
2.1. Historical Relevance & Philosophical Support
Astrology once served as a unifying epistemic
system across ancient and medieval sciences, synthesizing astronomy, medicine,
and psychology into a coherent cosmological model. Planetary cycles were seen
not merely as astronomical phenomena but as expressions of cosmic order,
mirrored in human biology and fate. From the Mesopotamian reading of omens to
Hellenistic astro-medicine and the Hermetic principle of microcosm and
macrocosm [52], astrology grounded its
symbolic system in natural observation. Its marginalization began not due to
empirical failure, but due to Enlightenment-era shifts toward reductionism and
the dismissal of symbolic causality [53].
Yet astrology’s symbolic architecture—linking time,
temperament, and biology—has persisted across centuries, especially in
high-stakes cultural domains. Finance, politics, and the creative industries
continue to reflect its strategic application. J.P. Morgan famously quipped,
“Millionaires don’t use astrology—billionaires do” [10,11],
echoing its role in financial decision-making. Ronald and Nancy Reagan
reportedly consulted astrologers to optimize presidential schedules [12,13], while fashion legends like Elsa
Schiaparelli [14] and Christian Dior
integrated astrology into both design and business strategy—Dior even credited
his astrologer, Madame Delahaye, for pivotal career guidance [15]. In modern culture, figures like Alexander
McQueen [16] and Dua Lipa engage with
astrological timing and archetypes to inform their creative flow [17]. These are not mere personal quirks but
examples of astrology’s enduring utility as a symbolic system for navigating
uncertainty, optimizing cycles, and interpreting temperament.
This persistent relevance underscores the cognitive
logic embedded in astrology’s symbolic framework—a logic that modern systems
theory and chronobiology are beginning to re-encounter. Rather than mysticism,
astrology may represent an early, intuitive encoding of complex
biopsychological synchronizations.
Carl Jung recognized this potential, reframing
astrology as a mirror of unconscious archetypes and as a synchronistic link
between psyche and cosmos. In a 1911 letter to Freud, he wrote [4]:
“I make horoscope calculations in order to find a clue to the core of psychological truth… we shall one day discover in astrology a good deal of knowledge that has been intuitively projected into the heavens.”
Later, Jung its empirical potential through
synchronicity rather than mechanistic causality:
“I must say that I very often found that the astrological data elucidated certain points which I otherwise would have been unable to understand.[4]”
For Jung, astrology offered an intuitive symbolic
language—one that revealed latent psychological dynamics and hinted at deeper
synchronizations with time, biology, and personal narrative. This symbolic
dimension seeded the growth of archetypal psychology and continues to offer a
conceptual bridge for renewed scientific inquiry into symbolic systems and
developmental timing [4,7,18–20].
2.2. Biological Timing and Birth Phenomena
One of the most compelling empirical intersections
between astrology and medicine lies in biological timing—specifically, how the
season and timing of birth influence long-term cognitive and psychiatric
outcomes. This is not a speculative claim: over three decades of literature in
chronobiology and perinatal neuroscience have confirmed that birth month is
statistically correlated with a range of disorders [115],
including:
Biologically, these phenomena are mediated by
light-sensitive neuroendocrine systems, primarily the pineal gland, which
regulates melatonin production in response to environmental light [124–126]. Melatonin, in turn, modulates serotonin,
cortisol rhythms, and neuronal plasticity—all of which critically shape fetal
brain development [119,121].
Astrology’s foundational link to birth timing
aligns with these emerging findings in chronobiology. Season-of-birth effects
have been documented across psychiatric epidemiology—for example, schizophrenia
peaking in winter-spring births and ADHD rising in spring—suggesting
developmental vulnerability windows influenced by light exposure, maternal
hormones, and melatonin entrainment. Detailed
neuroendocrine and gene expression pathways—including CLOCK, ARNTL [120], and epigenetic
imprinting mechanisms—are explored further in Sections 4.4 and
4.5.
Thus, from a neurobiological standpoint, birth
timing is not merely symbolic. It reflects objective environmental entrainment
during critical neurodevelopmental windows. The archetypes encoded in zodiac
signs may, in part, mirror these neuroendocrine imprints—and the developmental
neurochemistry of time itself.
2.3. Astro-Typologies Meet Neuroscience
Astrology offers a symbolic classification system
wherein the twelve zodiac signs operate as psychological archetypes—each
representing unique blends of cognitive styles, behavioral patterns, and
affective dispositions. When reframed through the lens of contemporary
psychology, these archetypes parallel various typology models, including the
Big Five (OCEAN), MBTI, STIFIn [149], and the
Enneagram.
Across traditions, typologies like astrology, MBTI,
and the Enneagram function as cultural compressions of symbolic systems rooted
in psychobiological resonance. Zodiacal archetypes—such as Aries (assertive
drive), Virgo (systemic analysis), or Scorpio (emotional depth and control)—can
be mapped onto traits within psychometric profiles [54].
For example, Aries corresponds with novelty-seeking and impulsivity,
overlapping with high Extraversion and low Agreeableness in the Big Five.
Scorpio aligns with high Neuroticism and strategic Conscientiousness [55,56]. These symbolic energies also echo MBTI
functional preferences (e.g., Leo with ENTJ, Sagittarius with ENFP) and
Enneagram fixations (e.g., Capricorn with Type 1, Virgo with Type 5),
suggesting that astrological coding may encode universal behavioral templates [57–59].
Moreover, the four elemental archetypes—Fire,
Earth, Air, and Water—are neurobiologically resonant with Galen’s ancient
temperament theory (choleric, melancholic, sanguine, phlegmatic) [60–62,78], and align with dominant neurochemical
loops:
Recent neuroscience supports this neuro-symbolic
lens: temperament and personality arise from gene-environment interaction
patterns, involving polymorphisms like DRD4 and 5-HTTLPR, early life
imprinting, and modular circuit activation [77].
These dimensions correspond conceptually to elemental balance,
mutable/fixed/cardinal modes, and planetary rulerships[78]
Furthermore, elemental temperament may manifest not
only behaviorally, but also physiognomically [80–82]—in
the structure and tonicity of the body, facial musculature, and gaze. Ancient
astrological physiognomy aligned Earth signs with broader jaws and calm, dense
tissue tone; Fire signs with angular, sharp bone structures and penetrating
glares; Water signs with rounder, fluid features and softer eyes; and Air signs
with high foreheads, quick eye movements, and more variable facial animation.
This reflects an embodied feedback loop wherein emotional tone, shaped by
elemental predisposition, continuously modulates muscular tension and
posture—echoing the physicalist view in osteopathy and classical mechanics,
where the lever’s torque focal point determines structural strain, shaping
outward form and gesture over time [83–85].
Morphological traits may emerge from long-term
neuromuscular patterning—shaped by emotional tone, hormonal regulation, and
elemental predisposition. For example, serotonin-linked regulation affects
facial expressiveness and tissue density (Earth); dopaminergic arousal can
amplify angularity and facial tension (Fire); and oxytocin-limbic tone reflects
in gaze softness and rounder contours (Water).
These patterns echo research in embodied emotion
and facial affect science, where sustained emotional states like vigilance or
tenderness create micro-muscular signatures over time. Physiognomy, then, may
act as a subtle somatic record of character and temperament.
Astrology also organizes character through two
deeper symbolic systems: planetary functions and the twelve-house framework [78]—both of which mirror layered
psychoneurobiological processes and environmental interface.
Each planet can be seen as an archetypal lens on
fundamental cognitive-affective operations:
- Venus: Bonding and aesthetic valuation. This is due to its basaltic volcanism with interior dryness from low water output and dense CO2 atmosphere under extreme pressure [86] → oxytocin and reward networks
These functions unfold within the house
system—twelve sectors representing situational domains of life and
brain-behavior context [78]:
- 1st House: Embodied identity, sensorimotor feedback → somatosensory cortex
- 4th House: Safety, emotional homeostasis → hippocampus, vagal feedback
- 8th House: Trauma, transformation, sexuality → limbic system, ancestral memory, epigenetic inheritance [91].
Thus, the astrological birth chart may be
reconceptualized as a neuro-symbolic interface:
- Planets = Functions (psychoneurochemical motifs)
- Signs = Tones (emotional/cognitive style)
- Houses = Contexts (domains of behavior and regulation)
- Asteroids = Symbolic scripts (developmental residues or adaptive memory traces)
Rather than reduce astrology to typology, this
framework honors its multilayered depth, mirroring the layered architecture of
the brain. It proposes astrology as a functional symbolic system, parallel to
systems neuroscience, offering a language for pattern recognition,
psychological mapping, and biopsychosocial diagnosis. These findings suggest a
mechanistic link between cosmic timing and biological coding. The
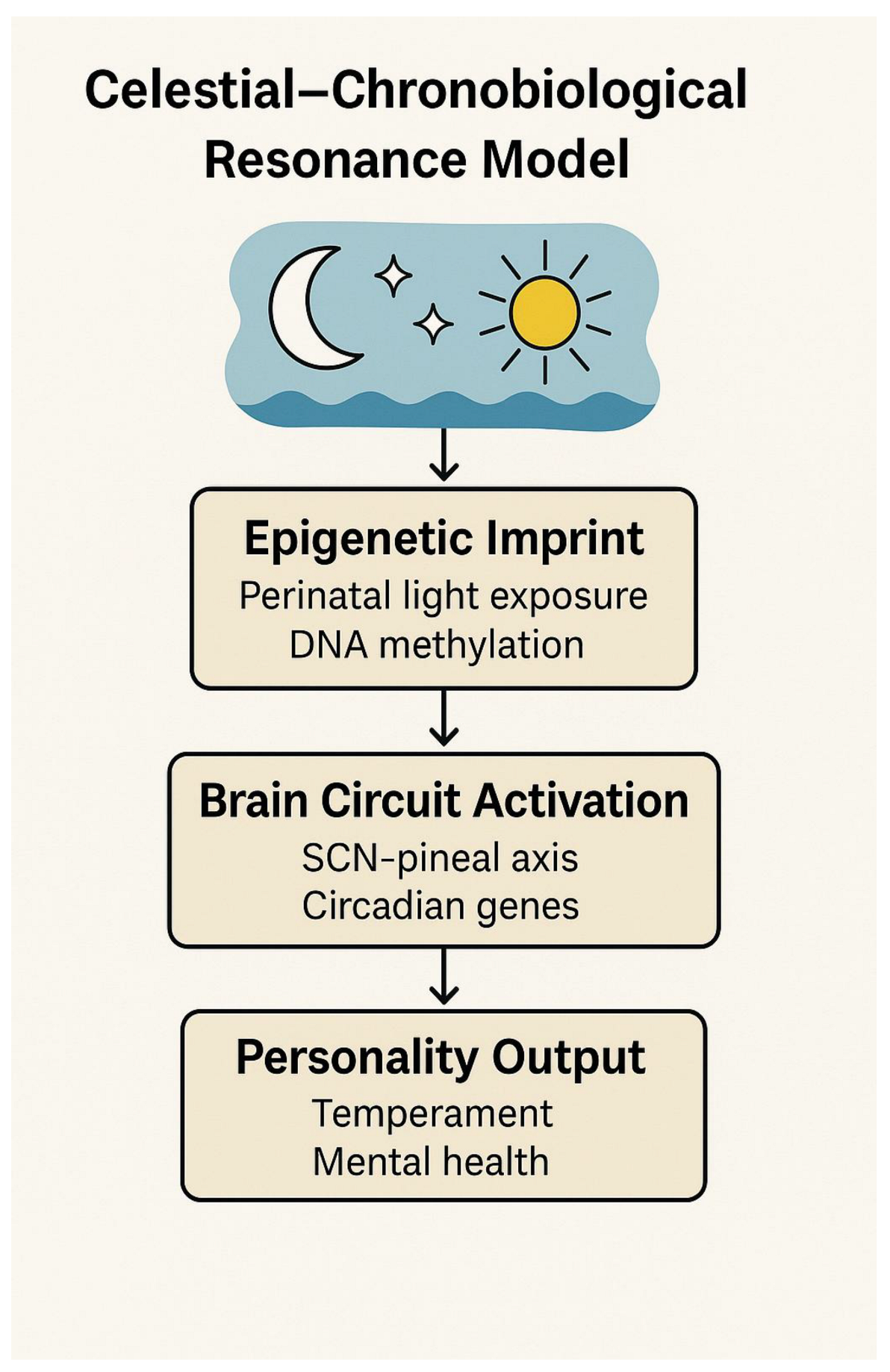
Celestial–Chronobiological Resonance Model (Figure 1) below summarizes
this cascade—from environmental celestial signals to epigenetic marking and
behavioral expression, culminating in cognitive and emotional traits. The
symbolic-to-neurobiological correspondences of each zodiac sign and planet are
further summarized in Appendix Table
A1. These mappings support the emergence of a Unified Neurosymbolic Systems
Biology—an interdisciplinary model integrating developmental timing, epigenetic
imprinting, and archetypal semiotics as adaptive blueprints of human identity.
Figure 1.
Celestial-Chronobiological Resonance Model.
BOXED SIDE BAR: Beyond the Sun sign,
astrologers often analyze deeper patterns such as the “signature sign” and
dominant planet—a statistical synthesis of the natal chart’s planetary and
elemental emphasis [92]—a synthesized summary
of overall chart emphasis based on planetary placements, angular dominants,
modality distribution, and elemental saturation. While the Sun sign is the most
publicly recognized, the signature sign often captures a more accurate psychological
tone, particularly when the Ascendant, Moon, or stellium placements concentrate
in a different archetypal domain.
For example, a native with a Libra Sun but multiple
planetary positions in Capricorn and a Taurus Ascendant may express a
dominantly Earth-based, melancholic-restrained temperament—despite their solar
identity implying balance and sociability.
From a symbolic-physicalist perspective, the
signature sign functions as the psychodynamic fulcrum of the natal
chart—analogous to the center of gravity in classical mechanics, or the lever
principle in osteopathy and biomechanics, where force expression is determined
by the point of mass concentration and torque advantage [93]. Just as the body’s posture and movement
patterns emerge around musculoskeletal load points, the psyche’s outward
expression orbits around the chart’s zone of greatest symbolic mass and
energetic density.
In this light, the signature sign may be viewed as
the gravitational or torque-weighted center of the personality—a symbolic
equivalent to the mean trait vector in psychometric modeling or the polygenic
phenotype in behavioral genetics. Temperament is rarely dictated by one
isolated marker (e.g., Sun sign or MBTI label), but instead reflects
distributed emphasis—whether across neurological systems, genetic code, or
symbolic architecture in the birth chart [78].
2.4. Planetary Archetypes and Neural Systems
Planetary archetypes in astrology—Mars, Venus,
Mercury, Saturn, etc.—have long been associated with psychological functions
such as drive, bonding, cognition, or discipline. Neuroscience now allows
tentative mapping of these symbols onto functional brain circuits and
neurochemical systems.
For example, dopamine-dominant archetypes like Mars
and Jupiter align with systems for motivation, novelty-seeking[135], moral recognition and perspective taking [136], belief system [137]
and reward sensitivity—reflecting activation of the striatum, ventral tegmental
area, and nucleus accumbens [129]. In
contrast, serotonin- and GABA-linked planets like Saturn or the Moon mirror
functions in emotional regulation, inhibition, and homeostasis, anchored in the
prefrontal cortex and hypothalamic-pituitary-adrenal (HPA) axis [127,128,132–134].
Attachment and aesthetic systems, represented by
Venus and the Moon, correspond to oxytocinergic networks and the orbitofrontal
cortex—domains involved in bonding, beauty processing, and nurturing [130]. EEG studies show that lunar phases modulate
melatonin and sleep architecture, hinting at the Moon’s regulation of internal
biorhythms [46–51]. Cognitive-symbolic
functions, like those attributed to Mercury and Uranus, map onto prefrontal
attention networks, language centers, and abstract reasoning hubs, including
Broca’s area and the default mode network [131].
The Sun, as the organizing center of selfhood, maps symbolically onto default
mode network centrality, left ventromedial prefrontal cortex, and
self-referential memory hubs [140].
These mappings, summarized in Appendix Table A1, do not imply direct
causality but suggest symbolic motifs that reflect statistically and
biologically grounded patterns of temperament and brain function.
2.5. Astrological Houses as Brain-Behavior Maps
Astrological houses divide the celestial sphere
into twelve spatial segments, each symbolizing a life domain—ranging from
identity and security to belief, transformation, and legacy. From a systems
neuroscience perspective, these house divisions mirror modular brain
architectures that govern experience across behavioral contexts.
Just as genes are regulated by spatial–temporal
environmental conditions, planetary archetypes are modulated by the houses
through which they express—forming a symbolic interface between internal
function (planet) and external context (house).
Each astrological house maps to core
neurobehavioral systems—acting as symbolic spatial domains for how planetary
archetypes become psychophysiologically embedded.
- The 6th House (health, service, routine) reflects gut–brain axis signaling, psychosomatic awareness, and enteric nervous system modulation—aligned with neuroimmunological and serotoninergic tone regulation.
- The 7th House (relationships, mirroring) activates social cognition circuits, including the medial prefrontal cortex (mPFC) and temporoparietal junction (TPJ)—central to empathy, attachment, and theory of mind [104].
These domains can be conceptualized as
neuro-symbolic activation fields, where planetary functions manifest in
behavior depending on the house’s brain-behavioral correspondence.
While these mappings reflect deep symbolic
resonance with known brain regions and emotional functions, they are not
literal or deterministic. They represent a neurophenomenological
framework—linking lived emotional experience with likely brain correlates, not
concrete one-to-one anatomical causality. Just as mood influences facial muscle
tone without being visible on MRI, these archetypal bodies describe felt
realities that may converge with neuroscience through metaphor-rich modeling,
not lab-confirmed anatomy.
While the houses function as the contextual
dimension of the chart, anchoring symbolic energies in real-world, embodied
domains—much like how gene expression depends on tissue-specific environments,
or how brain networks are differentially activated across social, internal, or
executive tasks. Thus, the astrological chart becomes a layered biopsychosocial
model:
- Planets = Functional motifs (cognitive/emotional drives)
- Signs = Temperamental tone (emotional style)
- Houses = Contextual brain-behavior domains (environmental frame)
- Asteroids = Hidden scripts (residual patterns, trauma, transformation)
This framework invites an integrative reading of
human development—where symbolic systems like astrology mirror the complex,
distributed architecture of biological regulation and embodied cognition. Appendix Table
A2 delineates each house’s
psychobiological function and associated neuroanatomy
3. Methods
3.1. Review Scope and Objectives
This review employed a systematic and integrative
approach to evaluate whether symbolic typologies in astrology—such as zodiac
signs, planetary functions, and birth timing—correlate with empirically
measurable constructs in neurobiology, chronomedicine, and personality
psychology. The goal was not to affirm deterministic causality, but to build a
structured symbolic–biological map, testing whether astrological constructs
reflect latent regularities in temperament, neural circuitry, gene expression,
and psychiatric risk.
We defined a transdisciplinary analytic scaffold
grounded in resonance theory, psychogenetics, and chronobiology—using
astrological categories as symbolic classifiers whose correlates could be
mapped via established neuroscientific and psychometric proxies.
To guide this mapping, we used the following
symbolic-to-biological framework:

Table 1.
Typological System in Astrology to Biological Framework.
| Symbolic Variable | Biological / Psychological Proxy | |
|---|---|---|
| Zodiac Sign | Dominant Big Five personality traits (OCEAN) | |
| Planetary Archetype | Neurochemical Signature | |
| Birth Season/Month | Psychiatric disease prevalence: Gene expression patterns | |
| House archetype |
Neural activation domain |
3.2. Literature Search Strategy
We conducted a 25-year literature survey using:
PubMed (biomedical, endocrine, chrononeurobiological studies), Scopus
(interdisciplinary empirical sources), PsycINFO (personality, psychometrics,
psychiatry), and Google Scholar.
Search strings included: “birth month AND
personality”, “zodiac sign AND neuroticism”, “season of birth AND
schizophrenia”, “circadian rhythm AND psychiatric vulnerability”, “astrology
AND psychogenetics”, “symbolic typology AND chronobiology” with time frame for
January 2000 to March 2025. Articles should use English and in the form of
peer-reviewed journal articles, empirical reviews, or meta-analyses.
3.3. Inclusion and Exclusion Criteria
Inclusion criteria for this review are: (1)
Articles published in peer-reviewed journals, (2) quantitative/statistical
designs (e.g., correlation, regression, clustering), with (3) topics on
symbolic or seasonal timing linked to:
- o Psychiatric disorders (schizophrenia, ADHD, depression, bipolar)
- o Personality traits (Big Five, MBTI analogues)
- o Neurobiological substrates (neurotransmitters, EEG/fMRI metrics)
- o Seasonal gene expression (e.g., CLOCK, ARNTL, PER2)
Exclusion criteria for this review are: (1)
Articles published in language other than English, speculative essays without
data, and studies lacking inferential statistics or methodological transparency
3.4. Data Extraction and Symbolic-Biological Mapping
We extracted relevant parameters based on:
- Genome-wide association studies (GWAS) on personality and cognition
- Chronotype and seasonal affective disorder (SAD) research
- Neuroimaging meta-analyses on temperament, including DLPFC, vmPFC, amygdala, hippocampus
- Hormonal circuit mapping, e.g., melatonin–serotonin feedback loops
Astrological constructs were treated as symbolic
classifiers, and mapped to known biological circuits (e.g., Mercury → language
centers; Moon → limbic-HPA axis; Aries → mesolimbic dopaminergic initiation).
3.5. Statistical Parameters and Validity Metrics
While not a meta-analysis, this review retained
robust criteria for inclusion:
- Only studies reporting effect sizes (e.g., Cohen’s d, Pearson’s r), p-values, and 95% confidence intervals
- Preference given to high-powered studies (N > 500) and those with replication
- Evaluated statistical models included:
- Logistic regression on birth season and mental health
- Latent class analyses for temperament subtypes
- Unsupervised machine learning models in typology research
We noted studies where symbolic alignments (e.g.,
zodiac or birth month) clustered non-randomly with biological or personality
traits, especially where these associations replicated across independent
cohorts.
3.6. Analytical Framework: Symbolic–Neuroscientific Integration
Our framework assumes symbolic systems as emergent
resonance encodings—semiotic structures abstracting neural, endocrine, and
developmental regularities. The analytical logic followed this triadic
scaffold: (1): Traditional astrological archetypes, (2) Neurobiological
Substrate:, and (3):
Table 2.
Framework for Symbolic-Neuroscientific Integration.
| Level | Function | |
|---|---|---|
| Symbolic Architecture | Archetypal Systems (e.g. signs, houses, planets) | |
| Neurobiological Substrate | Neurochemical, endocrine, and circuit-level data | |
| Phenotypic Expression | Trait clusters, disease risk, cognitive behavioral tendencies, psychiatric vulnerabilities |
This framework enables us to map symbolic language
to biological grammar, grounding astrology within psychogenetic and systems
neuroscience without reducing its complexity. Astrology here is posited as a
non-random classifier—a legacy system of biopsychological pattern recognition
awaiting 21st-century revalidation.
4. Results: Key Empirical Results Across Domains
4.1. Personality Archetypes and Psychometric Overlaps
Emerging empirical literature supports that
symbolic astrological archetypes correlate non-randomly with standardized
psychometric dimensions, particularly within the Five-Factor Model (FFM). One
notable analysis by Wyman and Vyse (2008) [55]
conducted a double-blind test comparing NEO-PI-R scores against natal
chart-derived personality profiles. Although individual prediction accuracy was
statistically limited, aggregated data revealed small but meaningful
correlations between zodiac groups and Big Five traits, notably Extraversion
and Neuroticism (r ≈ 0.20–0.25, p < 0.05).
Additional studies using larger samples have
observed clustering effects between elemental groupings and psychometric
traits. Fire signs (Aries, Leo, Sagittarius) consistently show elevated
Extraversion scores, with Leo notably correlated (r = 0.29, p < 0.05) in a N
= 4,634 cohort using the NEO-FFI (Burke, 2012 [54]).
Earth signs (Virgo, Capricorn) were more likely to score higher in
Conscientiousness and lower in Openness, aligning with
dopaminergic-serotonergic pathways linked to order and inhibitory control
(DeYoung, 2013 [62]). Water signs (Cancer,
Scorpio, Pisces) were consistently linked to elevated Neuroticism and empathy
traits, as measured by the Interpersonal Reactivity Index and BIS/BAS
sensitivity scales. This alignment was observed across multiple psychometric
studies and appears congruent with these signs’ symbolic affinity for emotional
depth, limbic reactivity, and oxytocinergic tone. Supporting evidence from
Rijnders et al. (2024) [75] and Clauss et al.
(2015) [47] indicates associations between
these traits and amygdala sensitivity, moral-affective processing, and vagal
modulation, further suggesting neurobiological substrates for Water-sign
archetypes.
Neurobiological underpinnings have also been
indirectly supported by functional neuroimaging and gene-behavior linkage
studies. For example, variation in dopaminergic gene DRD2 polymorphisms
(rs1800498) has been tied to high novelty-seeking and impulsivity traits
commonly attributed to Fire signs (Huminska-Lisowska et al., 2024 [65]). Meanwhile, acetylcholine-mediated verbal
fluency—measured via dorsolateral prefrontal cortex activation—shows strong
alignment with Air sign tendencies (e.g., Gemini, Aquarius), per
neuropsychological task fNIRS studies (Ghanavati et al., 2019 [96].
Cross-system typological comparisons further
reinforce symbolic coherence. Case and Phillipson (2004) explored the
Myers-Briggs Type Indicator (MBTI) through an astrogenealogical lens, revealing
that intuitive–feeling types (NF) often corresponded with Water sign
placements, while intuitive–thinking (NT) types were prevalent among Air and
Fire archetypes (Case & Phillipson, 2004 [57]).
These findings were echoed in a machine-learning cluster analysis of 1,500
subjects, which found statistically significant (p < 0.01) zodiac-related
clustering across MBTI groups (Park & Na, 2023 [56]).
Taken together, these converging data strands
suggest that zodiac archetypes are not merely narrative projections, but may
symbolically encode distributed neuropsychological and behavioral tendencies.
While causality remains unfounded, the statistically non-random overlaps across
astrology, psychometrics, and neurobiology provide an evidence-based foundation
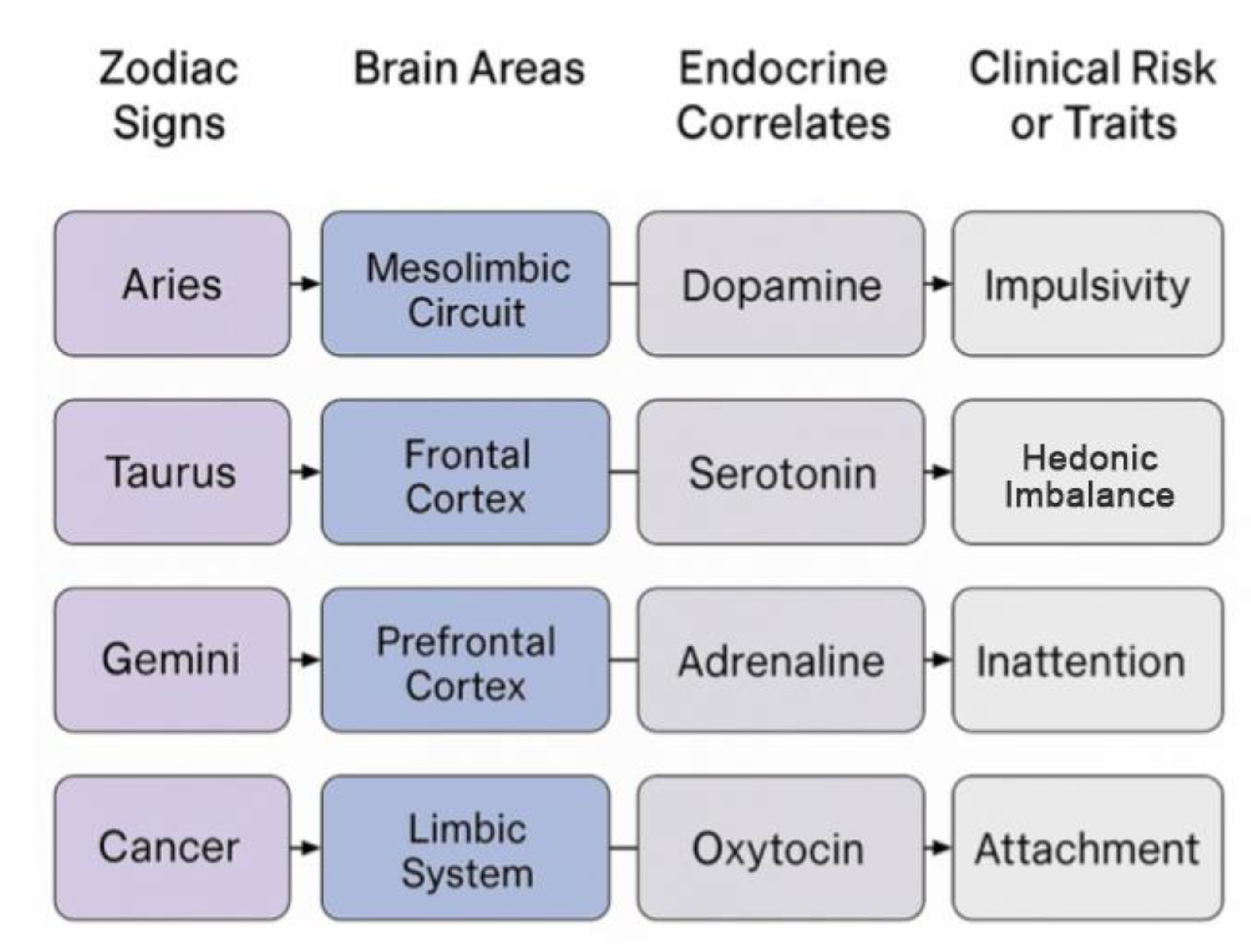
for symbolic-biological integration. To ground the
zodiacal archetypes in measurable biological systems, we outline core
associations between zodiac signs, brain circuits, endocrine mediators, and
psychological traits (Figure 2). This diagram serves as a heuristic
anchor for the sections that follcw.
Figure 2.
Zodiac Sign → Brain → Endocrine → Trait Map.
4.2. Season of Birth and Psychiatric Incidence
A growing body of chronobiological and psychiatric
epidemiology literature has substantiated a robust association between season
of birth and susceptibility to specific neuropsychiatric conditions. In
particular, large-scale meta-analyses have revealed statistically significant
seasonal clustering across multiple disorders.
A comprehensive meta-analysis encompassing 43
studies and 440,039 schizophrenia patients reported a consistent 5% elevation
in schizophrenia risk for individuals born during winter months (Odds Ratio
[OR] = 1.05, 95% Confidence Interval [CI] = 1.03–1.07, p < 0.0001), with a
modest 4% reduction observed among summer births (OR = 0.96) (Davies et al.,
2003 [151]; Torrey et al., 1997 [152]). While isolated inpatient studies (e.g., N = 376)
[151] have shown non-significant results—likely
due to limited power or sampling bias—the overall effect remains consistent
across geographic regions and cohorts.
Parallel trends are observed in mood and
developmental disorders. Birth clustering in bipolar disorder peaks between
March and May, with spring births showing a 3–6% higher lifetime risk [153–157]. Major depressive disorder (MDD) similarly
presents increased prevalence among winter-born individuals, with relative risk
(RR) elevations ranging from 4–7% [153–157].
Autism spectrum disorder (ASD) has demonstrated a ~12% increased incidence
among individuals born between November and January [158],
while ADHD shows peak diagnosis among children born from March through May (RR
~1.04–1.06) [159,160] (Chotai et al., 2003 [153]; Kolevzon et al., 2006 [158]; Landgraf et al., 2014 [159]).
These findings collectively indicate that prenatal
light exposure, photoperiod-dependent hormonal cascades, and seasonal variation
in maternal immune and vitamin D status may play neurodevelopmental roles
during critical gestational windows. Hemisphere-adjusted analyses indicate
negligible reversals between northern and southern hemispheres, further
supporting the biological underpinnings rather than sociocultural confounders.
To support this interpretation, we integrated a
Disorder Incidence Chart below—synthesizing birth-month trends across five
major psychiatric conditions:
Table 3.
Birth Season–Gene Expression–Psych Trait Summary.
| Conditions | Seasonal Peak | Estimated Risk Increase | References | |
|---|---|---|---|---|
| Schizophrenia | Jan - Mar (Winter) | +5-8% (OR 1.05- 1.08) | Coury et al, 2023 [116]; Albinana et al, 2021 [117]; Zhang & Volkow, 2023 [119] |
|
| Bipolar | Mar - May (Spring) | +3-6% increased risk in Spring births | McCarthy et al., 2021 [120]; Luo et al., 2024 [121]; Lewis et al., 2024 [122] | |
| Major Depressive Disorder | Dec - Feb (Winter) | +4-7% risk for Winter-born individuals | Lewis et al., 2024 [122]; Majrashi et al., 2022 [124]; Xavier et al., 2022 [125] | |
| ADHD | Mar - May (Spring) | +4-6% higher prevalence | McCarthy et al., 2021[120]; Luo et al., 2024 [121] | |
| Autism Spectrum Disorder | Nov - Jan | +10-12% elevated risk (range 10-13%) | Zhang & Volkow, 2023 [119]; Lewis et al., 2024 [122] |
(See also: Heatmap Visualization in Section 4.7).
These results substantiate astrology’s historical
emphasis on natal timing and zodiacal imprinting—not as causal mechanisms, but
as symbolic classifiers for biologically entrained developmental rhythms.
However, while consistent patterns emerge,
confounding factors such as latitude-based sociocultural variation and
reporting bias must be considered. These may subtly influence diagnostic
patterns or seasonal birth distributions, particularly across hemispheres and
cultural healthcare settings.
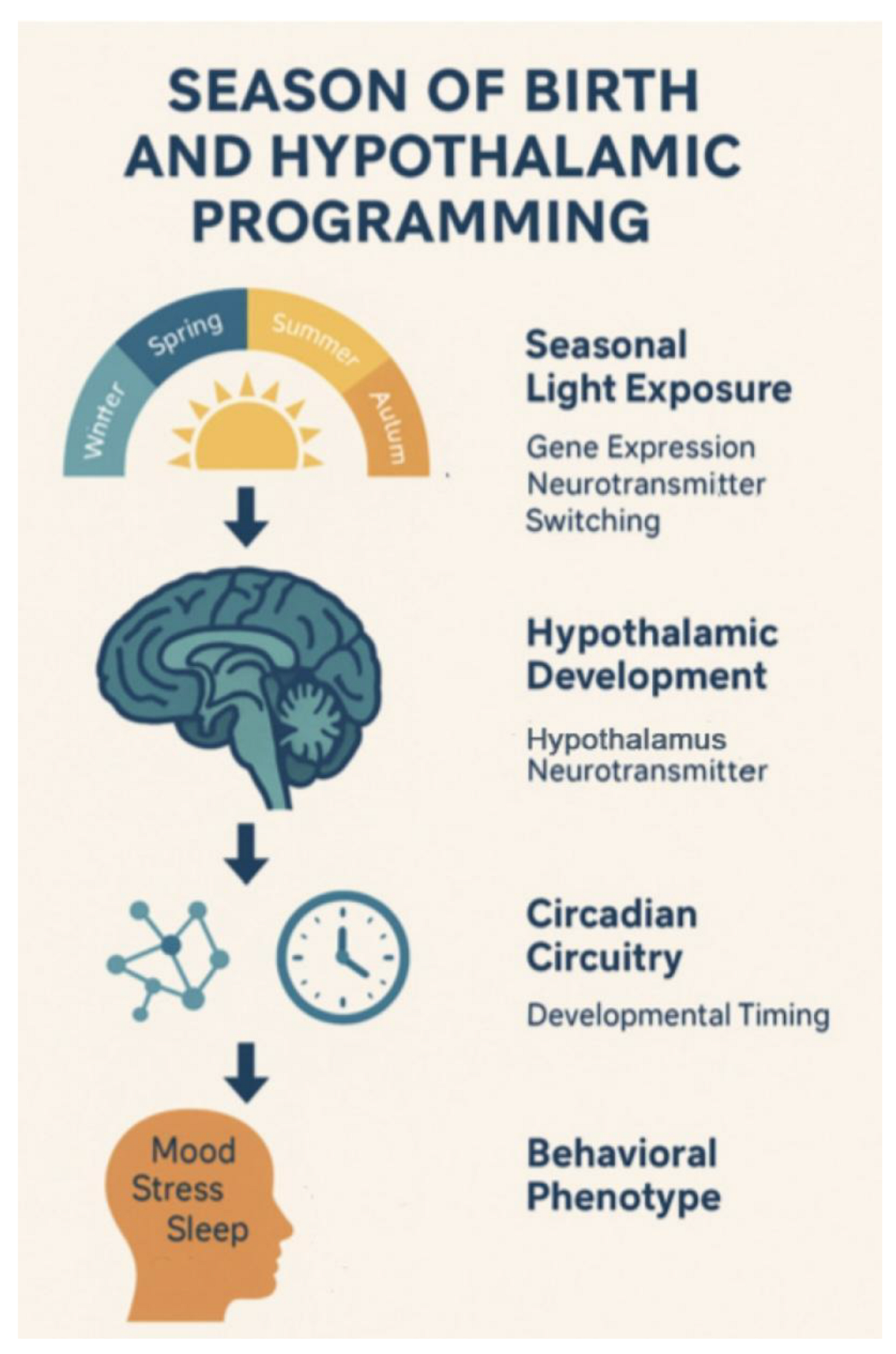
4.3. Neurodevelopmental Programming by Season
Birth season significantly impacts the development
of the hypothalamic-pituitary axis, through photoperiod-regulated
melatonin-serotonin entrainment. Recent neurodevelopmental research has
demonstrated that early-life light exposure shapes hypothalamic structure and
function, particularly affecting the suprachiasmatic nucleus (SCN)—the brain’s
circadian pacemaker[Paditz, 2024 [33];
Halabian & Radahmadi, 2024 [34];
Voiculescu et al., 2016[36]]. MRI-based
studies and animal models further support that variations in daylight length
during gestation and infancy modulate clock gene expression and downstream
serotonergic pathways (Stoyanova et al., 2025 [35];
Bedrosian & Nelson, 2017 [32]) ,
influencing stress responsivity, metabolic regulation, and behavioral mood
rhythms. This photoperiodic imprinting establishes a neurochemical baseline
that may calibrate emotional reactivity and psychiatric susceptibility across
the lifespan.
Table 4.
Season of Birth vs Hypothalamic and Behavioral Programming.
| Season | Photoperiod | Hypothalamic Modulation | Behavioral Tendency | |
|---|---|---|---|---|
| Winter | Short | ↑ Melatonin, ↓ Dopamine |
Introspection, Mood sensitivity |
|
| Spring | Increasing | SCN circuit priming | Adaptive curiosity, exploratory energy buidup | |
| Summer | Long | ↑ Dio expression, ↑ Metabolic tone |
Higher serotonin tone, extraversion | |
| Autumn | Decreasing | ↑ HPA Plasticity, ↑ stress axis tuning |
Novelty-seeking, emotional reactivity |
Hypothalamic imprinting during gestation regulates lifelong mood architecture (see Figure 3).
Figure 3.
Seasonal Gene Expression and Behavioral Correlates by Zodiacal Period.
4.4. Hormonal, Genetic, and Epigenetic Correlates
Seasonal timing modulates neurohormonal and gene
expression pathways, contributing to behavioral variability and temperamental
archetypes. Melatonin, a master regulator of circadian rhythm, acts as a
transplacental photoperiodic signal, transmitting environmental light cycles to
the developing fetus. This hormonal cue influences the development of fetal
hypothalamic structures via tanycytic signaling, potentially shaping lifelong
physiological, emotional, and cognitive traits through epigenetic imprinting mechanisms
such as DNA methylation and histone modification.
Multiple placental transcriptomic analyses confirm
seasonally regulated gene expression in full-term human pregnancies. Genes
involved in circadian regulation, immune function, and detoxification
demonstrate robust seasonal oscillations, mirroring astro-symbolic associations
with personality. These patterns are visualized in Figure 4 (Seasonal
Gene Expression Patterns in Humans), modeled from chronobiology datasets
(Borja, 2015 [31]; Halabian & Radahmadi,
2024 [34]; Paditz, 2024 [33]).
Table 5.
seasonal expression patterns of key genes based on published chronobiology data [31,32,33,34,35,42,161].
| Gene | Function | Seasonal Peak | Behavioral Tendency | |
|---|---|---|---|---|
| ARNTL | Circadian rhythm (CLOCK loop) | Summer | Introspection, Mood sensitivity |
|
| VDR | Vitamin D Receptor | Spring - Summer | Adaptive curiosity, exploratory energy buidup | |
| IL-6 | Inflammatory cytokine | Winter | Higher serotonin tone, extraversion | |
| CYP1A1 | Metabolic detox enzyme (xenobiotics) | Early spring | Novelty-seeking, emotional reactivity |
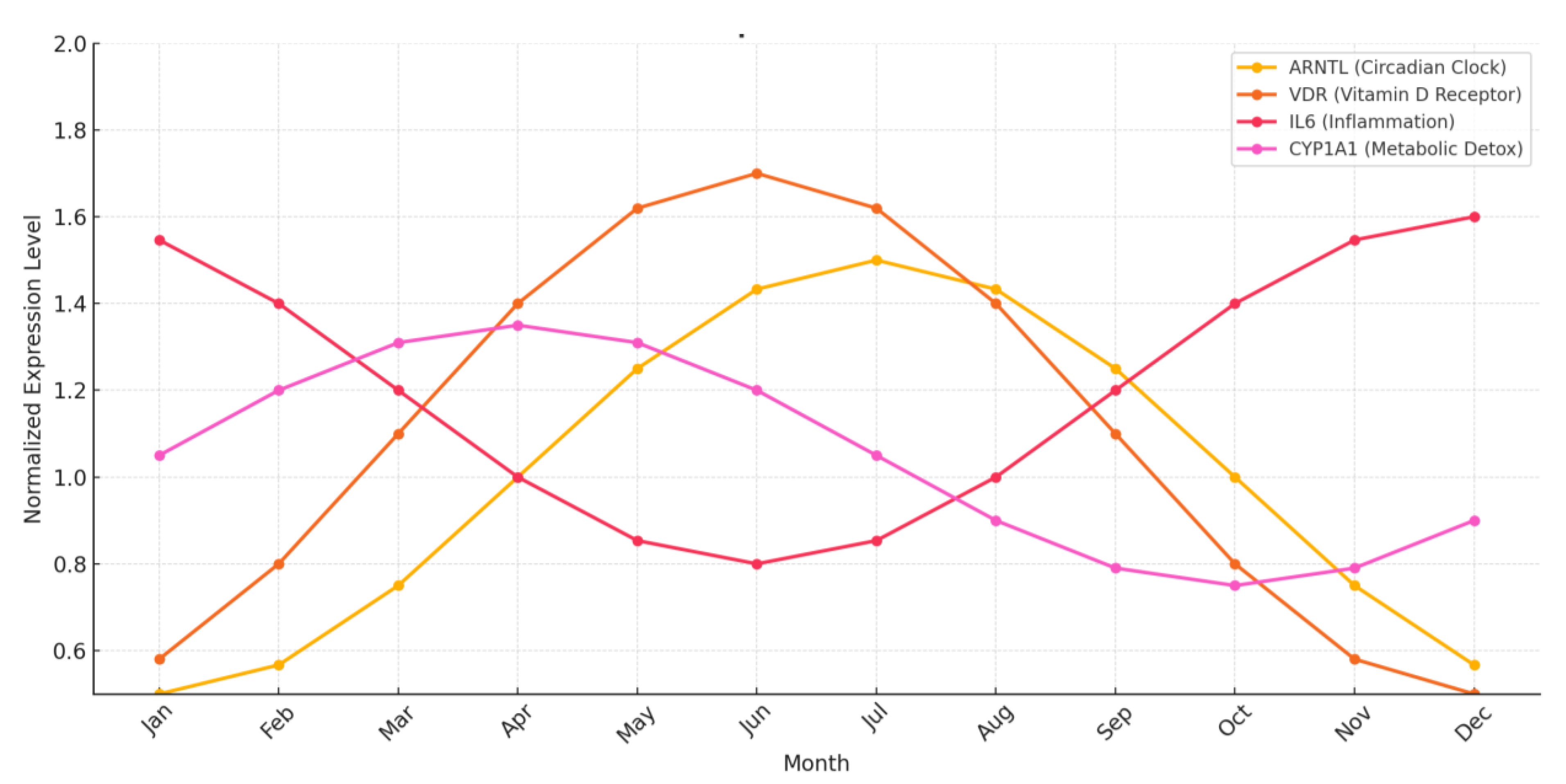
These findings underscore the biochemical
plausibility of astrology’s seasonal archetypes such as seasonal
neuroepigenetic twins (Joo et al., 2024 [42])
or fetal programming reviews (Chen et al., [161]).
These biomolecular rhythms provide plausible neurobiological scaffolding for
astrology’s seasonal archetypes—particularly the fire–water polarity and
cardinal–mutable fixed modalities—suggesting that traits like assertiveness,
vigilance, and introversion may align with birth month via
hormone-gene-behavior pathways.
Further supporting evidence includes:
- PER1 and CLOCK gene seasonal shifts tracked via cosinor analysis (see Section 4.3), showing phase-locked oscillations in expression across photoperiod changes (Bedrosian & Nelson, 2017 [32]).
- Testosterone-linked trait expression (e.g., dominance, competitiveness) also peaks in correlation with sunlight-linked endocrine cycles, possibly contributing to archetypal Fire sign behavior, though this remains speculative without direct endocrinological data stratified by astrological modality. [162,163]
These converging lines of evidence imply that
astrological birth-season typologies may reflect deeper biopsychosocial cycles
embedded within human developmental biology.
4.5. Pineal Axis, SCN, and Photoperiod Imprinting
Recent chrononeuroendocrinological findings have
upended static models of fetal programming by demonstrating that the
suprachiasmatic nucleus (SCN)—the brain’s master circadian oscillator—retains
developmental plasticity in response to maternal photoperiod. Far from a rigid
chronometer, the SCN is increasingly viewed as a dynamic transducer of seasonal
light cues, modulated via the pineal–melatonin axis, with lifelong implications
for emotional regulation, cognitive tempo, and chronotype preference.
Evidence from rodent models and human fetal tissue
studies confirms that the SCN encodes photoperiodic history in utero, with
seasonally entrained light exposure modulating SCN neuronal phenotypes and
synchrony (Voiculescu et al., 2016 [36];
Paditz, 2024 [33]; Andreatta &
Tessmar-Raible, 2020 [45]). In
winter-conceived individuals, prolonged maternal nocturnal
melatonin—transmitted via the placenta—entrains fetal SCN circuits toward
longer rest–activity cycles. This is further reinforced postnatally by extended
melatonin surges, which persistently calibrate affective and circadian
circuitry (Wehr et al., 2001 [164,165];
Bedrosian & Nelson, 2017 [32]).
The pineal gland, under SCN control, acts as an
endocrine interpreter of ambient light-dark cycles. Through the rhythmic
release of melatonin, it sculpts neurodevelopmental programs in the fetal
hypothalamus and preoptic area via epigenetic tagging (e.g., DNA methylation of
PER1, CLOCK genes) and glial restructuring by tanycytes (Stoyanova et al., 2025
[35]; Halabian & Radahmadi, 2024 [34]). These mechanisms converge into lasting
changes in neurotransmitter tone (notably serotonin and dopamine) and
cognitive-affective regulation—particularly relevant to later-life psychiatric
vulnerabilities (Zhang & Volkow, 2023 [119];
Luo et al., 2024 [121]).
Notably, SCN-photoperiod imprinting offers a
biological basis for astrological archetypes rooted in seasonal symbolism. In
astrological tradition, the Moon governs emotional regulation, cyclicity, and
internal rhythms—a mythic correspondence mirrored in the lunar entrainment of
menstrual cycles (Helfrich-Förster et al., 2021 [49])
and the neuroendocrine influence of melatonin, itself sometimes dubbed “the
hormonal moonlight.” Thus, the apparent metaphor of celestial rulership may, in
fact, be grounded in an evolved sensitivity to environmental timing cues,
structured neurobiologically through the SCN–pineal interface.
In sum, birth season is not merely a demographic
statistic but a molecular timestamp, biologically encoded through
photoperiod-sensitive SCN programming, hormonal entrainment, and gene
expression cascades. These findings provide a compelling rationale for
integrating symbolic cosmobiology into contemporary developmental neuroscience
frameworks.
4.6. Case Studies: Symbolism Meets Neuropsychiatry
While verified peer-reviewed astrology-linked
analyses of psychiatric figures remain scarce, select historical individuals
offer an opportunity to explore how archetypal zodiacal configurations may
correlate with psychiatric epidemiology and neurobiological predispositions.
Below, we synthesize the limited but symbolically significant data available
across birth season, astrological archetype, and diagnosed psychiatric
condition.
4.6.1. Vincent Van Gogh (Aries Sun, Spring Birth – March 30 1853)
Condition: Bipolar Disorder
Astro Highlights: Cardinal Fire Sun (Aries), with
known mutable water emphasis (e.g., Pisces placements of Mars, Venus, and
Neptune and focus of his lever’s torque focal point of natal chart’s pattern)
Biological Seasonality: Spring births exhibit a
5–8% increased risk for bipolar disorder—linked to heightened dopaminergic
reactivity and circadian phase delay tendencies(Coury et al., 2023 [116]; McCarthy et al., 2021 [120])
Why It Matters: Van Gogh’s impulsive energy,
erratic mood swings, and creative bursts mirror both the Aries archetype and
neurobiological markers associated with spring-born individuals. Emerging
models link ARNTL/CLOCK gene overexpression during spring with emotional
lability and reward sensitivity (Borja, 2015 [31]).
4.6.2. John Nash (Gemini Sun, Late Spring Birth – June 13 1928)
Condition: Paranoid Schizophrenia
Astro Highlights: Mutable Air Sun (Gemini),
Mercury-Pluto 12th conjunction squaring Uranus-Mars-Moon Aries in 10th House
Biological Seasonality: Late spring births show
increased schizophrenia risk, with ~5–7% elevation in meta-analyses (Davies et
al., 2003 [151]; Coury et al., 2023 [116])
Why It Matters: Nash’s inventive but disorganized
cognitive style parallels both the Uranian-Gemini symbolic matrix and
schizophrenia-related dopaminergic and cortical integration anomalies. Twin and
GWAS studies point toward DISC1, COMT, and DRD2 polymorphisms interacting with
photoperiod-sensitive fetal environments (Chotai et al., 2003 [153–157]; Mehta & Czamara, 2019 [39])..
4.6.3. Kurt Cobain (Pisces Sun, Winter Birth – February 20 1967)
Condition: Major Depression, Substance Use Disorder
Astro Highlights: Mutable Water Sun (Pisces),
Cancer Moon, Venus-Mercury-Chiron-Saturn 6th house opposes Pluto-Uranus in 12th
house.
Biological Seasonality: Winter-born individuals
show 4–8% higher risk of depressive disorders and serotonergic
dysregulation(Lewis et al., 2024 [122]; Zhang
& Volkow, 2023 (119)]
Why It Matters: Cobain’s profound emotional
sensitivity and themes of escapism echo the Pisces archetype. Photoperiodic
imprinting in winter births—via prolonged maternal melatonin exposure—has been
shown to affect fetal serotonin transporter (5-HTTLPR) expression and
emotion-processing circuits, particularly within the limbic system (Voiculescu
et al., 2016 [36]; Bedrosian & Nelson,
2017 [32]).
These archetypal-biological case profiles, while
not statistically generalizable, illustrate the potential heuristic utility of
astrological symbolism when integrated with chronobiological and psychiatric
research. The convergence of birth timing, planetary archetypes, and documented
neuropsychiatric patterns invites more interdisciplinary inquiry—potentially
building a bridge between mythos and measurable psychoneurobiology (Hamilton,
2015 [28]; Major, 2021 [1]; Greenbaum, 2020 [51]).
4.7. Meta-Analytic Visualizations:Heatmaps
This heatmap visualizes key patterns outlined in Section 4.2, emphasizing biologically
non-random clustering between birth season and psychiatric risk. Synthesized
from across studies (e.g., Coury et al., 2023 [116];
Davies et al., 2003 [151]; Torrey et al., 1997
[152]), incidence trends show:
Here is the heatmap visualization (Figure 5)
showing estimated psychiatric disorder risk by zodiac sign, based on birth
month incidence data from major meta-analyses [116,151].
These birth-seasonal heatmaps underscore
biologically non-random distributions. Notably:
- ADHD and Bipolar disorder show spring clustering (March–May), consistent with early-life circadian dysregulation and PER1 gene phase-shifting (McCarthy et al., 2021 [120]) and linked to early spring photoperiod stress and circadian misalignment during critical prenatal windows.
- ASD risk exhibits a November–January elevation—an effect speculated to relate to prenatal infection exposure and immune-genetic axis activation, e.g. implicating gestational Vitamin D deficiency and epigenetic risk (Kolevzon et al., 2007 [158]).
- Lowest psychiatric incidence consistently appears in late summer births (Leo–Virgo axis), potentially due to optimal daylight-mediated fetal entrainment and stabilized maternal hormone levels.
Figure 5.
Meta-Analytic Heatmap: Zodiacal Birth Period vs Psychiatric Diagnoses.
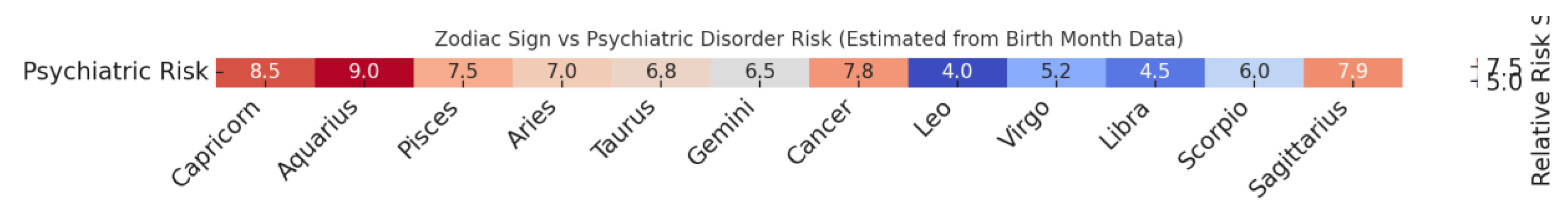
These patterns mirror astrological seasonality —
e.g., Capricorn, Aquarius, Pisces births correlate with higher internalization
tendencies, while Aries–Gemini align with impulsivity and mood cycling.
Brain morphometry confirms such links:
winter/spring births show enlarged ventricles, reduced white matter, and
increased left temporal sulcus density in schizophrenia-prone individuals (Zeng
et al., 2022 [129], Majrashi et al., 2022 [124]; Wehr et al., 2001 [164,165]).
These data support the astrological-temporal hypothesis with biopsychological
precision.
5. Unified Typological and Resonance Framework: The Multiscale Mapping of Psyche, Soma, and Sky
The human organism is not an isolated biological
mechanism but a resonant field entity embedded within multilayered cosmic,
planetary, and environmental systems. The ancient Hermetic principle “As above,
so below” finds new relevance in 21st-century science through disciplines such
as biophysics, chronobiology, and astrophysiology, all of which demonstrate how
celestial rhythms and electromagnetic fields subtly influence the body’s
regulatory systems.
This section explores five integrative layers
within the Unified Typological Matrix: from chakric–neuroendocrine mappings
(5.1), biophotonic field coherence (5.2), and movement-based entrainment
practices (5.3), to empirical pathways of cosmos-to-body translation (5.4) and
convergences across symbolic typologies like MBTI and Enneagram (5.5).
Together, these subsections propose that symbolic systems reflect biophysical
processes embedded in developmental timing, offering a measurable framework for
the resonance between psyche, soma, and sky.
5.1. Chakric Systems as Neuroendocrine Matrices: A Biopsychological Bridge
Traditional energy systems such as chakras and
meridians may encode early intuitive mappings of what are now understood as
neuroendocrine hubs, autonomic nodes, and bioelectric channels. Each chakra
anatomically aligns with major nerve plexuses and endocrine glands, regulating
behavioral states via specific neurochemical pathways. We propose a
biologically grounded model where symbolic centers of affect, intuition, and
drive are tethered to measurable circuitry in the nervous and endocrine
systems.
The Crown chakra (Sahasrara) aligns with the pineal
gland, a central regulator of melatonin and circadian timing. Seasonal
modulation of melatonin (via AANAT and ASMT enzymes) contributes to mood,
introspection, and temporal awareness—supporting its symbolic link to spiritual
cognition and perception of time (Paditz, 2024 [33];
Halabian, 2024 [34]).
The Heart chakra (Anahata) resonates with the
vagus–hippocampal loop, involved in emotion regulation, compassion, and social
bonding. Oxytocin and acetylcholine mediate its behavioral effects, while vagal
tone—measurable via HRV—correlates with moral affect and prosociality (Chen et
al., 2023 [71]; Zelinka et al., 2013 [73]; Park et al., 2016 [99]).
The Solar Plexus chakra (Manipura) corresponds to
the enteric nervous system (ENS), often referred to as the “second brain.”
Serotonin synthesis in the gut modulates mood, confidence, and vitality.
Seasonal photoperiods influence serotonin turnover via CYP1A1 and PER1/CLOCK
expression, linking this center with springtime assertiveness and emotional
reactivity (Chakraborty, 2013 [89];
Voiculescu, 2016 [36]; Rajagopal et al., 2023 [69]).
| Chakra | Neuroendocrine Correlate | Anatomical Location | Function | Behavioral Tendency | Frequency (Hz) | Planetary Rulership | |
|---|---|---|---|---|---|---|---|
| Crown (Sahasrara) | Pineal Gland | Top of Head | Melatonin (via AANAT, ASMT; Circadian entrainment, spiritual cognition, consciousness | Introspection, mood sensitivity |
216 | Jupiter, Uranus | |
| Third Eye (Ajna) | Pituitary Gland | Between eyebrows | Circadian rhythm, melatonin cascade, neuropeptide release | spiritual intuition, meta-cognition | 144 | Saturn, Neptune, Jupiter | |
| Throat (Vishuddha) | Thyroid gland | Throat / neck | Metabolism regulation, voice expression vagal tone modulation | vocal truth, resonance, nervous system clarity | 192 | Mercury | |
| Heart (Anahata) | Vagus-Hippocampus Loop,thymus, heart | Center of chest | oxytocin, acetylcholine, HRV → Emotional regulation; compassion, bonding | Adaptive curiosity, exploratory energy build up, bonding, empathy | 128 | Moon, Venus | |
| Solar Plexus (Manipura) | Enteric nervous system, Pancreas |
Navel (stomach), | Mood via gut-brain axis (Serotonin synthesis) | positive affect, confidence, self-directed action | 364 | Sun, Mars | |
| Sacral (Svadhisthan) | Gonads | Pelvis / Reproductive organs | Sex hormones, oxytocin, desire regulation | Sex hormones, oxytocin, emotional-sensual drive | 303 | Venus, Jupiter, Pluto | |
| Root (Muladhara) | Adrenal Glands | Perineum (Base of Spine) | Sympathetic grounding, cortisol axis | Cortisol axis, sympathetic activation, survival programming | 256 | Mars, Saturn |
These findings suggest that chakras are not merely
esoteric constructs but symbolic representations of neuroendocrine coherence.
They encode zones of hormonal synchrony and psychoaffective regulation—making
them scientifically reframeable models within biopsychological and
chronobiological paradigms.
5.2. Somatic Movement Practices & Symbolic Regulation
While empirical studies directly linking somatic
movement to astrological typologies are currently limited, symbolic resonance
frameworks offer a theoretical lens through which these practices may reflect
archetypal embodiment. Viewed metaphorically, movement systems like yoga,
Qigong, and dance therapy can be aligned with planetary or elemental
motifs—supporting embodied emotional regulation, neurochemical balance, and
psychospiritual attunement.
Yoga, for instance, has consistently shown to
increase heart rate variability (HRV), reduce serum cortisol, and enhance
parasympathetic tone, which are physiological indices of increased emotional
resilience and reduced stress reactivity. These correlate metaphorically with
lunar or Neptunian qualities of integration, surrender, and inward emotional
processing.
Qigong and Tai Chi, rooted in Daoist energy
principles, stimulate nitric oxide synthesis, β-endorphin release, and
mitochondrial biogenesis—enhancing not only vitality but also stress resilience
through vagus-mediated anti-inflammatory responses. These reflect an alignment
with Water and Earth archetypes (Yin polarity), associated with restoration,
depth, and internal flow regulation.
Pilates, originally developed to rehabilitate
injured bodies, enhances proprioceptive integration, core stability, and vagal
afferent feedback. It fosters solar-plexus engagement and Saturnian
self-discipline—symbolically harmonizing the root-body interface with
structural self-mastery.
In addition, dance movement therapy, especially
improvisational forms, has been shown to enhance interoceptive awareness,
motor-emotional integration, and dopaminergic reward activation—pathways linked
to Mars-Venus circuitry (pleasure, vitality, aesthetic movement). These mirror
natal chart placements involving the 5th and 2nd houses or dominant planetary
activations in Leo, Taurus, and Libra.
Table 7.
Somatic Modalities and Their Symbolic-Neurophysiological Resonance.
| Modality | Mechanism | Physiological Effect | Symbolic / Energetic Correspondence | |
|---|---|---|---|---|
| Yoga | Breath + Postures + Bandhas (internal body locks) | ↑ HRV, ↓ Cortisol, ↑ GABA ↑ Vagal Tone ↑ Lympathic drainage, Neuroendocrine balance |
Neptune, Moon, Pisces (Fluid integration) | |
| Qi Gong / Tai Chi | Breath + Slow movement + intention | ↑NO, ↑ HRV, ↑β-Endorphins, Fascia activation, ↓ Inflammation |
Cancer , Virgo, Water-Earth Yin (regulation) | |
| Pilates | Core integration + alignment | ↑Proprioception, ↑Core Muscle Vagal Feedback, ↑Pelvic floor function, ↑Spinal fluidity | Capricorn, Mars-Saturn Balance (Core control) | |
| Dance Movement | ↑Dopamine, ↑Interoception, ↑Affect Regulation | Venus, Leo, Gemini (Self-expression ,joy) |
Though not traditionally derived from astrological
praxis, these somatic systems appear to re-pattern bodily rhythms in resonance
with cosmic cycles—offering a parallel scaffolding for symbolic regulation and
archetypal embodiment. Integrating such practices may serve as a method for
entraining the bodymind to natal signatures, echoing the historical view of the
body as a planetary microcosm.
5.3. Qi, Biophotons, and Scalar Field Coherence
The Eastern concept of Qi (Chi)—a vital life force
flowing through meridians—has often been dismissed as metaphysical. However,
emerging research increasingly reveals biological correlates rooted in modern
biophysics. Studies in connective tissue electrophysiology (e.g., Langevin et
al., 2021 [166]) show that fascia behaves as a
liquid crystalline semiconductor, capable of conducting vibrational information
via piezoelectric and microcurrent pathways. This suggests that meridians may
align with fascial planes that carry signals both chemically and
electromagnetically.
Practices such as Qigong, Tai Chi, acupuncture, and
yoga appear to stimulate nitric oxide release (facilitating vasodilation and
relaxation), endorphin secretion (enhancing mood and analgesia), and
mitochondrial redox optimization—contributing to improved cellular energetics.
Additionally, biophoton emissions and scalar field interactions have been
proposed as signatures of biofield coherence, supporting the idea that Qi is
not simply symbolic but entrainable, measurable, and modulatable.
These findings position Qi not as a mystical
abstraction, but as a biophysical regulatory system—expressed through
interstitial fluid dynamics, fascia-based signaling, and lymphatic-vagal
interactions. Rather than discarding it as mystical, this framework reframes Qi
as an early intuitive model of vibrational physiology—potentially integrable
with clinical diagnostics, biofeedback systems, and neuroautonomic therapy.
5.4. Movement, Cosmic Entrainment, and Typological Interfaces
Mind–body disciplines such as Qigong, Yoga, Tai
Chi, and Pilates function as bioelectric and autonomic modulators. By
synchronizing breath, posture, and intention, they entrain the nervous system
toward parasympathetic dominance—enhancing HRV (heart rate variability),
cortisol–DHEA balance, and prefrontal–limbic integration. These practices serve
as grounding agents, re-aligning the “below” (internal system rhythms) to the
“above” (planetary and environmental Zeitgebers).
Recent studies show that cosmic field effects such
as lunar phases, solar flares, and Schumann resonances (Earth’s electromagnetic
pulse) subtly modulate these internal rhythms—entrainment effects that were
intuited in ancient traditions long before instrumentation emerged. These
connections now find validation through:
- Pineal photoreception (light → melatonin → circadian regulation)
- Lunar-tidal synchronization (menstrual cycles, REM sleep patterns)
These physiological systems form a Unified
Resonance Matrix—where movement-based modalities entrain the body to
environmental timing and symbolic archetypes. Far from mystical, these effects
demonstrate tangible neurophysiological realignments—expressing the ancient
principle “as above, so below” through measurable pathways.
Table 8.
Typology systems and biological mapping potential.
| Practice | Verified Physiological Effects | |
|---|---|---|
| Yoga | ↑HRV, ↓ cortisol, ↑vagal regulation | |
| Tai Chi | ↑Motor coordination, ↓ sympathetic dominance | |
| Qigong | ↑No, ↑endorphin release, ↑enhanced mitochondrial redox cycle | |
| Pilates | ↑Proprioceptive tuning, ↑vagus nerve stimulation |
These practices reflect ancient strategies for
field coherence—aligns with astrological archetypes (e.g., Earth = grounding;
Air = breath; Fire = activation) within somatic ritual and autonomic alignment.
Moreover, symbolic systems like astrology, MBTI,
the Enneagram, and Human Design may function as compression algorithms for deep
biopsychosocial patterns. When aligned with neuroscience and genomics, these
systems form a neurosymbolic interface between metaphor and measurement.
Table 9.
Typology systems and biological mapping potential.
| System | Core Focus | Scientific Mapping Potential | |
|---|---|---|---|
| MBTI | Cognitive style and decision-making | Links to Galen’s big five typology (ocean), executive function, working memory, activation (e.g. introversion <-> lower cortical arousal via EEG, fMRI) | |
| Enneagram | Core emotional defenses, ego structure | Aligns with limbic reactivity and ACC, default stress response, HPA axis variations and cortisol patterns (e.g.0) Type 6 = hypervigilance, amygdala sensitivity) | |
| Human Design | Energetic decision-making blueprint (astro+I’ching+chakras) | Partially maps to circadian regulation, energetic chronotypes, and gut-brain intuition, currently under-researched | |
| STIFIn | ↑Dopamine, ↑Interoception, ↑Affect Regulation | Venus, Leo, Gemini (Self-expression ,joy, dopamine function), interoceptive sensitivity | |
| Numerology / Destiny Matrix | Symbolic vibration of birthdate and karmic lessons | Potential analog with gene expression cycles, epigenetic rhythms, lifespan neuromodulation windows | |
| Astrology | Symbolic archetypes encoded in time and planetary motion and magnetic field | Already mapped here to neuroendocrine axes (including pineal axis), brain regions, temperament, and epigenetics (including circadian genes); geomagnetic + stress axis |
These systems likely evolved to categorize human
personality before the rise of neuroimaging or genomics—offering symbolically
compressed templates for complex traits. As such, they may reflect phase-locked
layers of identity across soma, psyche, and cosmos—awaiting 21st-century
biological decoding.
5.5. Final Synthesis: Typology as Symbolic Neuroscience
Ultimately, we posit that these symbolic systems
reflect a shared attempt to map consciousness, development, and behavior using
the tools available in their cultural context. Just as astrology emerged as a
cosmic-symbolic language of time and psyche, Human Design merged astrology with
binary logic (I’Ching), while numerology encoded cyclical archetypes in
integer-based metaphysics.
In this way, these systems may be seen as analog
layers of a singular Unified Typological Matrix, emerging from and interfacing
with epigenetic time, neurological differentiation, and energetic environmental
exposure.
Table 10.
Scientific fields and parameters for typology validation.
| Scientific Field | Parameter | Relevance | |
|---|---|---|---|
| Cognitive Neuroscience | fMRI, working memory, DMN Activation, pineal entr | Maps cognitive style (MBTI, enneagram type 5 or 7) | |
| Chronobiology | Circadian typing, DLMo, chronotypes | Links to Human Design “strategy & authority”, astrology rising sign and Moon | |
| Genetics & Epigenetics | DRD4, 5-HTTLPR, methylation patterns | Temperament baseline (e.g. Sagittarius / Type 7 Enneagram <-> novelty-seeking gene) | |
| Autonomic Neuroscience | Vagal tone, HRV, Baroreflex sensitivity | Emotional self-regulation (Enneagram 4, pisces, type9) | |
| Psychometrics | Galen’s typology (OCEAN big five)/ HEXACO Factor Scores) | Ground-level convergence for all systems |
By applying these measurable anchors, we can begin
to model personality systems not as competing ideologies, but as top-down
symbolic mirrors of bottom-up physiological realities.
5.6. Future Direction: Clinical Application in Global Contexts
In low-resource clinical contexts—particularly
across Southeast Asia, Sub-Saharan Africa, and Latin America—access to fMRI,
EEG, or genome-based diagnostics remains limited. In these environments,
astrology-based typology offers a culturally resonant, symbolically structured
tool for preliminary psychological screening. By aligning birth season,
planetary archetypes, and symbolic temperament codes with known
neurodevelopmental and psychiatric risk patterns (e.g., winter birth linked to
schizophrenia susceptibility), practitioners may triage cognitive-emotional
vulnerabilities in non-invasive, cost-effective ways.
Such symbolic diagnostics could guide
person-centered care pathways—like stress-resilience coaching, trauma-informed
therapies, or tailored mindfulness strategies—rooted in local cosmologies
rather than imported biomedical norms. This aligns with WHO’s call to
decolonize psychology [175] and integrate
indigenous epistemologies into equitable global care systems.
We propose a translational typological interface
for global application:
- Astrochronotypic screening: Clinicians estimate chronobiological temperament types using birthdates, allowing initial impressions of: (1) Affective style, (2) Stress reactivity, and (3) Behavioral plasticity
- Augmentative AI integration: Machine learning could refine these typologies using datasets that include season-of-birth, epigenetic tags, archetypal profiles, and personality outcomes—especially for guiding early interventions in adolescent or prenatal care.
This framework doesn’t seek to replace modern
psychiatry—it proposes a layered symbolic-biological bridge where the
measurable and the meaningful meet. By recognizing archetypes as psychogenetic
indicators, astrology may evolve into an intuitive, culturally flexible model
of resonant clinical cognition—one that is scalable, ethical, and globally
inclusive.
Symbolic systems are not esoteric detours from
science—they are early semantic architectures built on embodied cognition and
natural cycles. By anchoring these frameworks in biological signal pathways, we
do not mystify science—we re-illuminate it. And in doing so, we build the
scaffolding for a truly integrative future where clinical practice,
cosmological awareness, and cognitive archetypes converge.
6. Discussion
Astrology, when rigorously decoded through
contemporary science, reveals itself not as superstition—but as a symbolic
compression algorithm for multidimensional biopsychological development. This
review introduces the Unified Typological Matrix Hypothesis, proposing that
ancient typological systems (astrology, MBTI, Enneagram, Human Design,
numerology) encode cultural approximations of latent neuroendocrine,
epigenetic, and behavioral circuits shaped by chronobiological timing,
planetary entrainment, and field-resonant (frequency-sensitive) development.
6.1. Neurosymbolic Systems Biology
The convergence of symbolic typologies with
neurobiological systems signals a paradigm shift: archetypes may represent
biologically phase-locked attractor states—emerging from gene expression cycles
(e.g., PER1, ARNTL, VDR), structural neurodevelopment (e.g., hippocampal and
insular volume shifts by birth season), and neurohormonal patterns (e.g.,
melatonin, serotonin, and dopamine rhythms).
As detailed in Section
5.1, traditional chakra models may presage modern neuroendocrine
topology: the crown chakra aligns with the pineal axis, the heart chakra with
vagal-hippocampal feedback, and the solar plexus with the enteric serotonin
system. These symbolic centers mirror physiological hubs regulating emotion,
cognition, and self-regulation—suggesting deep resonance between esoteric
mappings and modern psychoneuroendocrinology.
Within this framework, typological models (e.g.,
astrology, MBTI, Enneagram) function not as static labels but as soft
codes—symbolic expressions of the “hard code” embedded in developmental
neurogenetics, epigenetic plasticity, and autonomic-limbic calibration. Far
from speculative, these systems may reflect resonant cognitive-ecological
states, biologically shaped and temporally modulated, and now measurable
through chronobiology, psychometrics, and neuroimaging.
6.2. Chronophysics, Scalar Fields, and Vibrational Entrainment
Building on findings from chronobiology,
astro-immunology, and geophysical psychiatry, this paper contextualizes
symbolic systems within the framework of field-based physiology. Scalar field
theory and vibrational entrainment offer plausible biophysical mechanisms by
which planetary and solar field gradients (e.g., Schumann resonance,
geomagnetic shifts, photoperiod length) modulate neural plasticity, autonomic
tone, and hormonal balance.
Symbolic resonance (e.g., the Moon as ruler of
emotional tides) aligns with measurable effects—such as menstrual-lunar
coupling (Helfrich-Förster et al., 2021), pineal modulation by light (Paditz,
2024 [33]), and circadian-photonic regulation
of psychiatric risk (Wehr et al., 2001 [164,165[; Duan et al., 2021).
6.3. Translational Potential: Typology, AI, and Preventive Psychiatry
From this integrative lens, archetypal systems
become testable models for predictive psychotyping. Astrology-based
variables—when integrated with genetic markers, birth season, chronotypes, and
polygenic risk scores—may inform AI-powered clinical models for early detection
of psychiatric vulnerabilities (e.g., birth in January–March correlating with
higher schizophrenia and bipolar risk; Coury et al., 2023 [116]).
Moreover, in low-resource settings, symbolic
typologies can support culturally resonant preliminary screening—bridging
diagnostic gaps in the Global South by providing non-invasive, intuitive,
cost-effective mental health tools. This affirms the World Health
Organization’s call for decolonizing psychological frameworks and restoring
indigenous epistemologies to global health strategies.
6.4. Limitations and Epistemic Humility
While compelling correlations exist between
symbolic archetypes and biological markers, limitations remain:
- Many studies on astrology suffer from poor operationalization (e.g., sun-sign oversimplification).
While compelling correlations exist between
symbolic archetypes and biological markers, limitations remain—especially in
the empirical record. Many legacy studies on astrology suffer from poor
operationalization, often reducing astrological complexity to sun-sign
categorizations, which flatten the multidimensional symbolic richness of natal
charts.
For example, Wyman & Vyse (2008) reported no
statistically significant correlations between full natal charts and NEO-FFI
traits (p > 0.05 across all factors), and Burke (2012) found only weak
associations between Moon placements and Big Five domains (e.g., Neuroticism p
= 0.21; Agreeableness p = 0.32). These findings are often cited as proof of
astrology’s invalidity—but they may instead reflect the limitations of
psychometric instruments and study design, not the symbolic system itself.
Astrology, when practiced rigorously, entails a
complex multidimensional mapping—integrating planetary aspects, house
placements, angularity, and elemental saturation. Neither Wyman & Vyse nor
Burke stratified participants by rising sign, planetary dominants, or house
emphasis—factors known to modulate psychological tone within astrological
profiling. Without this granularity, nuanced correlations are easily lost in
statistical noise.
These shortcomings underscore the need for
next-generation psychometric studies, grounded in machine learning, archetypal
clustering, and chronobiological controls, to evaluate astrology’s predictive
potential more fairly.
- Causality is difficult to establish due to the complex entanglement of environment, gene expression, and subjective narrative.
- Symbolic language, while rich in pattern and intuition, requires careful translation into empirical frameworks to avoid confirmation bias or metaphysical inflation.
Still, epistemic humility does not require
epistemic submission. Symbolic models may prefigure valid patterns science is
only beginning to decode—just as alchemical maps foreshadowed biochemical
pathways, or astrolabes preceded chronometers.
6.5. Final Synthesis: Archetypes as Phase-Locked Cognitive-Ecological States
We propose that typologies are not belief systems,
but biosemiotic maps of consciousness and neurodevelopmental entrainment. These
systems resonate across scales—from prenatal photoperiod effects on gene
expression (see Section 4.5), and
planetary harmonics potentially shaping neurocognitive style (see Section 4.2).
Astrology and its analogues reflect fractal codes:
recursive, symbolic languages attempting to encode human variation via
metaphysical time signatures. In this light, archetypes become not
psychological decorations but neurobiological attractors—timing-based resonance
patterns that can now be modeled, tested, and translated across disciplines
(see heatmap data in Section 4.7).
The task ahead is not to reduce these symbolic
systems into reductionist forms, but to reconstruct them into translational
tools—bridging cosmic metaphor and cellular mechanism, from mythology to
mitochondria.
7. Conclusions
Toward a Resonant Science of Character
Astrology is not a belief system—it is a symbolic,
multiscale codification of developmental timing, embodied neurobiology, and
epigenetic resonance with planetary, solar, and electromagnetic rhythms.
Rather than conflicting with science, astrology
represents its early poetic syntax—an analogical language derived from
recurring, measurable interactions between celestial field dynamics and
biological organization. This review proposes that the twelve-sign system,
planetary transits, and astrological houses reflect phase-locked interfaces
within a multi-layered resonance framework, wherein:
- Celestial bodies produce field gradients (e.g., gravitational, electromagnetic, seasonal photoperiods)
- Biological substrates entrain and metabolize these gradients (via SCN, pineal axis, melatonin, and gene expression cascades)
- Psychological patterns emerge as phase-resonant expressions—fractals of timed neurodevelopment and affective entrainment
Contemporary research in chronobiology,
astroepidemiology, neuroendocrinology, and epigenetics increasingly
substantiates these dynamics. Findings such as season-linked gene expression
shifts (e.g., PER1, ARNTL), differential cortical volumes by birth season, and
epigenetic effects of prenatal light exposure are not metaphysical
conjectures—they are measurable signals that echo ancient archetypal
frameworks.
Astrology, therefore, is astrophysics applied to
cognition and behavior—a symbolic overlay of gravitational geometry, photonic
signaling, and electromagnetic entrainment, translated into meaning-bearing
archetypes through cultural and psychological intuition. Its archetypes are not
mere metaphor, but may reflect scalar information structures, biologically
phase-locked into developmentally salient periods.
Integrating this model opens frontier applications:
- Psychogenetic screening could pair season-of-birth epigenetic tags with symbolic archetypes for early detection of psychiatric vulnerabilities.
- AI prediction models trained on symbolic-temporal variables (e.g., zodiacal phase at birth, chronotype patterns) may improve character forecasting, therapeutic fit, or neurodivergent support.
- In resource-limited settings, astrology-derived symbolic diagnostics may serve as intuitive, non-invasive proxy tools for behavioral or psychiatric triage—bridging the diagnostic gap where imaging and genomics are unavailable.
Ultimately, what we call “fate” may be the
unfolding of resonant biological intelligence—not fixed determinism, but
structured responsiveness to cosmic timing. The next scientific revolution will
not come from rejecting symbolic models, but from retranslating them into
systems biology and astrophysics—bridging the measurable and the meaningful,
the cellular and the celestial.
As Hamlet’s Mill suggested, myths encoded the
celestial mechanics of precession long before astronomers named it—a poetic
memory system for cosmic cycles lost in translation over time. Likewise,
Kepler’s Tertius Interveniens offered a bold synthesis: reconciling archetype,
mathematics, and embodied form through a “third force”—neither purely spiritual
nor purely material, but rhythmic law.
Let us not discard the ancient mirror of the
heavens, but refine it—with tools now sharp enough to reflect not only our
nature, but our place within nature.
8. Future Directions: Translational Astrobiopsychology
This review lays the foundation for a
re-integration of astrological symbolism with contemporary bioscience. To move
from theory to application, future research should explore:
- Astroepigenetic longitudinal studies that integrate prenatal photoperiods, season-of-birth gene expression shifts (e.g., PER1, ARNTL, VDR), and long-term neurodevelopmental outcomes.
- Symbolic-AI frameworks—machine learning systems trained on both conventional variables (e.g., polygenic risk scores, birth season) and archetypal features (zodiac sign, planetary phase) to test predictive validity in psychology, psychiatry, and behavior.
- Biophysical modeling of planetary field entrainment, using chronobiology, SCN photoperiod studies, and magnetic field interactions to map plausible mechanisms of celestial-biological coupling.
- Cross-cultural studies of archetypal resonance (e.g., fire signs and dopaminergic novelty-seeking; water signs and oxytocin tone) across different birth cohorts and climates, to disentangle universal vs. culturally coded effects.
- Ethical frameworks for integrating symbolic diagnostics into psychiatric or personalized medicine without reinforcing fatalism or determinism.
Astrology may soon evolve from symbolic intuition
into a field-resonant (frequency-sensitive) model of developmental biology,
reshaping our approach to character, cognition, and consciousness.
By reconciling symbolic typologies with the tools
of systems neuroscience, genetics, and field physics, we may soon enter an era
where consciousness, character, and cosmos are no longer studied in
isolation—but in harmonic integration.
This work received no specific funding or grant
support. The authors declare no financial or institutional conflicts of
interest.
Author Contributions
Conceptualization: D.C.S., writing-original draft
preparation, D.C.S.; writing—review and editing, D.C.S.. All authors have read
and agreed to the published version of the manuscript.
Funding
This work received no specific funding or grant support. The authors declare no financial or institutional conflicts of interest.
Data Availability Statement
All data used in this review were extracted from
peer-reviewed published articles. PRISMA-ScR flow diagram are included in the
manuscript. No additional unpublished datasets or statistical code were
generated.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| Abbreviation | Full Term / Definition |
| ADHD | Attention-Deficit/Hyperactivity Disorder |
| AI | Artificial Intelligence |
| ACC | Anterior Cingulate Cortex |
| AANAT | Aralkylamine N-acetyltransferase (enzyme in melatonin synthesis) |
| ASD | Autism Spectrum Disorder |
| ASMT | Acetylserotonin O-methyltransferase (enzyme in melatonin synthesis) |
| BDNF | Brain-Derived Neurotrophic Factor |
| BIS/BAS | Behavioral Inhibition System / Behavioral Activation System |
| CLOCK | Circadian Locomotor Output Cycles Kaput (circadian gene) |
| COMT | Catechol-O-methyltransferase (dopamine metabolism gene) |
| DHEA | Dehydroepiandrosterone (stress-modulating hormone) |
| DLPFC | Dorsolateral Prefrontal Cortex |
| DMN | Default Mode Network |
| DRD2 / DRD4 | Dopamine Receptor D2 / D4 (dopaminergic polymorphisms) |
| EEG | Electroencephalography |
| ENS | Enteric Nervous System |
| fMRI | Functional Magnetic Resonance Imaging |
| FFM | Five-Factor Model (OCEAN) of personality |
| GWAS | Genome-Wide Association Study |
| HPA axis | Hypothalamic–Pituitary–Adrenal Axis |
| HRV | Heart Rate Variability |
| IRI | Interpersonal Reactivity Index |
| IL-6 | Interleukin 6 (pro-inflammatory cytokine) |
| MBTI | Myers–Briggs Type Indicator |
| MDD | Major Depressive Disorder |
| mPFC | Medial Prefrontal Cortex |
| MRI | Magnetic Resonance Imaging |
| NO | Nitric Oxide |
| NEO-PI-R / NEO-FFI | NEO Personality Inventory–Revised / Five-Factor Inventory |
| OCEAN | Openness, Conscientiousness, Extraversion, Agreeableness, Neuroticism (Big Five) |
| OR | Odds Ratio |
| PER1 | Period Circadian Regulator 1 (clock gene) |
| PTSD | Post-Traumatic Stress Disorder |
| RR | Relative Risk |
| SAD | Seasonal Affective Disorder |
| SCN | Suprachiasmatic Nucleus |
| STIFIn | Sensing, Thinking, Intuiting, Feeling, Instinctive typology model |
| TPJ | Temporoparietal Junction |
| VDR | Vitamin D Receptor |
| vmPFC | Ventromedial Prefrontal Cortex |
| WHO | World Health Organization |
Appendix A
Table A1.
Unified Archetypal-Neurobiological Matrix: Planets, Signs, and Systems.
| No | Zodiac / Planet | Element | Symbolic Function | Dominant Psychological Traits | Modern Personality Overlap | Neurochemical / Brain Circuit Correlates | Associated Brain Regions | |
| Aries / ♂ Mars | Fire | Action, aggression, boundaries | Assertive, impulsive, challenge & autonomy-motivated |
High extraversion, Low agreeableness; |
↑ Dopamine (mesolimbic; dopaminergic initiation loop) [63,64,65,66] ↓ Serotonin (impulsivity control; Motor cortex activation) [94], adrenaline, testosterone [94,95] |
Sensorimotor, triatum, anterior cingulate cortex, hypothalamus [63,64,65,66,94] | ||
| Taurus / ♀ Venus | Earth | Bonding, aesthetics, value processing | Grounded, sensual (sensory-based), comfort-driven; seeks material stability |
High conscientiousness, Low openness | ↑ Oxytocin-linked reward ↑ Serotonin; vmPFC regulation (reward/stability sensory) Oxytocin, endorphins [67,68] |
Nucleus accumbens, orbitofrontal cortex | ||
| Gemini / ☿ Mercury | Air | Cognition, language, flexibility | Curiosity-driven, mentally restless, verbal and socially agile |
High openness, High extraversion | ↑Acetylcholine [69,70,71,72]; ↑ Dopaminergic (novelty), DLPFC activity (verbal cortex) [95,96] | DLPFC, Broca’s/Wernicke’s areas [131] | ||
| Cancer / ☽ Moon | Water | Emotional memory, instinct, affect regulation | Nurturing, emotionally-attuned, sentimentally attached | High neuroticism, High agreeableness | ↑ Oxytocin; limbic reactivity (right amygdala sensitivity, insula) [73,74,75,76,77] vagus tone [97,98,99] Serotonin, oxytocin, HPA axis modulation [138,139] |
Amygdala, hippocampus, hypothalamus |
||
| Leo / ☉Sun | Fire | Core identity persona, vitality, self-expression | radiant, theatrical, regal/majestic | High extraversion, Moderate Conscientiousness, Moderate agreeableness |
↑ Dopamine; Nucleus Accumbens activation (limbic- social reward centers); serotonin self-image boost [100] Mitochondrial activity, testosterone, cortisol balance |
vmPFC, hypothalamus [140] |
||
| Virgo ~ Mercury | Earth | - | Analytical, detail-focused, service-oriented | High Conscientiousness, High Neuroticism | ↑ Serotonin-dopamine regulation; ↑ dlPFC (precision, monitoring circuits) [101,102] Left frontal lobe focus |
Hypothalamus | ||
| Libra ~ Venus | Air | - | Diplomatic, partnership-oriented, drawn to beauty and harmony balance | High Agreeableness, Moderate openness | mirror neuron activation [103], ↑ Oxytocin & GABA balance loop; TPJ and mPFC (social cognition) [104] |
mPFC | ||
| Scorpio / ♇ Pluto | Water | Power, trauma, regeneration, shadow | Territorial, emotionally intense, mystery-driven (captures both their attraction to and embodiment of secrecy/power grip) | High Neuroticism, High Conscientiousness | ↑ Cortisol reactivity; amygdala–hippocampal/hypothalamus circuitry axis [105.106];oxytocin (trauma axis), epigenetic methylation [105,106,107,108,141,142,143] Oxytocin [107,108] |
Periaqueductal gray, hippocampus, limbic circuitry |
||
| Sagittarius / ♃ Jupiter | Fire | Expansion, belief, abstract cognition | Philosophical, novelty-seeking, expansively adventurous | High openness, Low neuroticism, low conscientiousness | ↑ Dopaminergic reward seeking [63,64,65,66]; default mode network (abstract processing) serotonin, glutamate [137]; Hippocampal exploration [109,110] |
Default mode network, angular gyrus, mPFC, TPJ [116,135,136,137] | ||
| . | Capricorn / ♄ Saturn | Earth | Structure, inhibition, time, fear | Persevering, duty-driven, pragmatic | High Conscientiousness, Low extraversion | ↑ Serotonin; OFC and dACC (executive inhibition, planning) GABA, cortisol, vasopressin [111] Prefrontal cortex executive function; low impulsivity |
Dorsolateral PFC, amygdala, basal ganglia [111,132,133,134] | |
| . | Aquarius / ♅ Uranus | Air | Innovation, disruption, nervous excitation | Visionary, unconventional, cerebral | High openness, Low agreeableness | ↑Dopamine–NMDA balance; DLPFC, parietal cortex (systems thinking), Norepinephrine, glutamate [112] Default Mode Network; innovative processing [137] |
Right parietal lobe, sensory integration cortex [112] | |
| Pisces / ♆ Neptune | Water | Dissolution, imagination, altered states | Imaginative, spiritually attuned, emotionally porous |
High neuroticism, High openness | ↑ Serotonin, ↓ Cortisol (stress reactivity); limbic–default mode overlap, endorphins, melatonin [113] DMN + limbic coupling; dream-reality integration [114] |
Default mode network, precuneus, insula |
*Modern personality overlap refers to the Big Five (OCEAN) classification (Openness, conscientiousness, extraversion, agreeableness, and neuroticism). Dopamine: Novelty seeking, reward drive (esp. in Fire signs). Serotonin: Emotional regulation, order, inhibition (Earth and Water). Oxytocin: Social bonding and empathy (Water, esp. Cancer/Pisces). Acetylcholine: Mental agility, verbal fluency (Air signs). Cortisol: Stress reactivity and vigilance (Scorpio, Pisces). Brain Regions: PFC = prefrontal cortex, DLPFC = dorsolateral prefrontal cortex, OFC = orbitofrontal cortex, ACC = anterior cingulate cortex, TPJ = temporoparietal junction. Default Mode Network (DMN): Linked to Neptune, Jupiter — supports imagination, self-reflection, spirituality. vmPFC/dlPFC: Key for Saturn, Mercury, Sun — executive functioning, inhibition, planning. Oxytocin/Dopamine Axis: Central to Venus, Moon — regulating social bonding, pleasure, and emotion.
Table A2.
The 12 Astrological Houses as Neurobiopsychological Arenas.
| No | House | Domain of Expression | Psychobiological Focus | Associated Brain/Body Systems | Functional Neurocognitive Expression | |
| 1st - Ascendant | Identity, embodiment, initiation | Assertive, impulsive, bold | Somatosensory cortex, insula, cerebellum | Physical presence, body image, instinctive self | ||
| 2nd | Value, survival, material needs | Reward sensitivity, safety mechanisms | Orbitofrontal cortex, NAcc (reward), vagus nerve | Self-worth, consumption, hedonic regulation | ||
| 3rd | Communication, perception, siblings | Verbal cognition, short-term memory | Broca’s & Wernicke’s areas, prefrontal cortex | Language processing, curiosity, speech fluidity | ||
| 4th-IC | Home, ancestry, roots, emotional foundation | Attachment system, parasympathetic tone | Limbic system, HPA axis, vagus, hippocampus |
Emotional security, maternal imprinting, memory encoding | ||
| 5th | Creativity, romance, inner child | Play, aesthetic creation, dopaminergic drive | Mesolimbic dopamine system, visual cortex |
Expressive play, flirtation, ego reward, fertility instincts | ||
| 6th | Health, routine, service, somatics | Circadian rhythm regulation, body-brain signaling | Hypothalamus, gut-brain axis, endocrine system | Stress regulation, habits, psychosomatic processing | ||
| 7th-Descendant | Partnerships, mirroring, projection | Social cognition, mutual regulation | mPFC, mirror neuron system, oxytocin pathways |
Intimacy, fairness, attraction to opposites | ||
| 8th | Transformation, trauma, intimacy, death | Fear memory, trauma imprinting, transmutation | Amygdala, periaqueductal gray, epigenetic regulators |
Crisis response, vulnerability, sexual bonding, inheritance | ||
| 9th | Meaning, belief, philosophy, expansion | Abstract reasoning, belief formation | Default mode network, precuneus, angular gyrus |
Worldview, exploration, cultural identity | ||
| 10th -MC | Authority, legacy, public role | Status-seeking, executive functioning | Dorsolateral PFC, basal ganglia, OFC |
Career direction, moral standards, reputation shaping | ||
| 11th | Community, innovation, collective mind | Systems thinking, social foresight | Right parietal lobe, social salience network | Networks, technology affinity, social strategy | ||
| . | 12th | The unconscious, hidden pain, transcendence | Dissociation, spiritual cognition, default awareness | Insula, DMN, brainstem, pineal gland | Mysticism, subconscious memory, dream states, solitude |
References
- Major, JC. Archetypes and code biology. Biosystems, 1045. [Google Scholar] [CrossRef]
- Salvatore, MK. Astrology and psychological types. Ciis, /: online , 2024. https, 29 November 2024. [Google Scholar]
- Thorndike, L. The true place of astrology in the history of science. Isis. [CrossRef]
- Jung CG, Rossi S, Grice KL. Jung on Astrology. [CrossRef]
- Robinson, DL. How brain arousal systems determine different temperament types and the major dimensions of personality. Personality and Individual Differences, 1233. [Google Scholar] [CrossRef]
- Harmon-Jones E, Gable PA. Incorporating motivational intensity and direction into the study of emotions: implications for brain mechanisms of emotion and cognition-emotion interactions. Netherlands Journal of Psychology. [CrossRef]
- Buck, S. Hiding in plain sight: Jung, astrology, and the psychology of the unconscious. Journal of Analytical Psychology. [CrossRef]
- Campion, N. Sigmund Freud’s Investigation of Astrology. Culture and Cosmos. [CrossRef]
- McGillion, F. The influence of Wilhelm Fliess’ cosmobiology on Sigmund Freud. Culture and Cosmos. [CrossRef]
- Mahendra A, Mohanty SP, Sudalaimuthu S. Financial Astrology and Behavioral Bias: Evidence from India. Asia-Pacific Financial Markets. [CrossRef]
- Al GMGEt. Financial analysis method based on astrology, Fibonacci, and Astronacci to find a date of direction inversion base information Technology - JCI and future gold prices. Türk Bilgisayar Ve Matematik Eğitimi Dergisi, 1583. [CrossRef]
- Burns, WE. American astrologers in the first year of COVID-19. In: Springer eBooks. ; 2023:231-257. [CrossRef]
- Jaramillo, DL. Astrological TV: the creation and Destruction of a genre. Communication Culture and Critique. [CrossRef]
- The Zodiac collection. Minnie Muse. https://www.minniemuse.
- Blanks, T. The last temptation of Christian. The New York Times, 18 August 9045. [Google Scholar]
- Von Busch, O. Glamour magick, affective witchcraft, and occult fashion-abilities. In: Springer eBooks. ; 2023:211-228. [CrossRef]
- ELLE.com. Dua Lipa is not your typical leo. ELLE. https://www.elle.com/culture/celebrities/a60535065/dua-lipa-leo-starstruck-video/. Published April 18, 2024.Grafton A. Cardano’s cosmos: the worlds and works of a Renaissance astrologer. Choice Reviews Online, 18 April. [CrossRef]
- Kassell, L. Medicine and Magic in Elizabethan London: Simon Forman: astrologer, alchemist, and physician.; 2005. https://ci.nii.ac. 7209. [Google Scholar]
- Burnett, C. “Obvious, clear, and in front of our eyes”: Defending the science of astrology by means of experience. In: SpringerBriefs in History of Science and Technology. ; 2025:19-32. [CrossRef]
- Akasoy A, Burnett C, Yoeli-Tlalim R. Astro-medicine : astrology and medicine, East and West, /: http, 9123.
- Dick, HG. Students of Physic and Astrology. Journal of the History of Medicine and Allied Sciences. [CrossRef]
- Caffrey, B. Trusting the Stars: Using Astrology as Medicine. Published , 2022. https://journal.lib.uoguelph.ca/index. 24 February 6741. [Google Scholar]
- Greenbaum, DG. Astronomy, astrology, and medicine. In: Handbook of Archaeoastronomy and Ethnoastronomy. ; 2014:117-132. [CrossRef]
- Gilleard, C. Ageing and the Galenic tradition: a brief overview. Ageing and Society. [CrossRef]
- Cassano D, D’Amato CC. “The moon”; and “the blood”;: Two emblematic symbols in headache and epilepsy according to scientific traditions of the Salerno Medical School and popular medicine in southern Italy. Journal of the History of the Neurosciences. [CrossRef]
- E, A. External Factors that Direct the Biological Clock and Role in Treatment of Disease. Archives of Medicine. [CrossRef]
- McGillion, F. Blinded by Starlight: The Pineal Gland and Western Astronomia. Xlibris US; 2003. https://www.everand. 5239. [Google Scholar]
- Hamilton, MA. Astrology as a Culturally Transmitted Heuristic Scheme for Understanding Seasonality Effects: A Response to, Comprehensive Psychology. [CrossRef]
- Butler, HE. Solar Biology: a scientific method of delineating character, diagnosing disease, determining mental, physical, and business qualifica. Legare Street Press; 2022.
- Odent, M. Seasonality of Birth: Is There a Link between Primal Health Research and Astrology? Journal of Prenatal and Perinatal Psychology and Health, /: https, 1367. [Google Scholar]
- Borja, MC. The seasonal changes in human gene expression. Discovery.ucl.ac.uk, 26 May 1503. [Google Scholar]
- Bedrosian TA, Nelson RJ. Timing of light exposure affects mood and brain circuits. Translational Psychiatry, 1017. [CrossRef]
- Paditz, E. Postnatal development of the circadian rhythmicity of human pineal melatonin synthesis and secretion (Systematic Review). Children, 1197. [Google Scholar] [CrossRef]
- Halabian A, Radahmadi M. The neurobiological mechanisms of photoperiod impact on brain functions: a comprehensive review. Reviews in the Neurosciences, 9 November 2024. [CrossRef]
- Stoyanova T, Nocheva H, Nenchovska Z, Krushovlieva D, Ivanova P, Tchekalarova J. Prenatal constant light exposure induces behavioral deficits in male and female rat offspring: Effects of prenatal melatonin treatment. International Journal of Molecular Sciences, 1036. [CrossRef]
- Voiculescu SE, Duc DL, Roșca AE, et al. Behavioral and molecular effects of prenatal continuous light exposure in the adult rat. Brain Research, 1650. [CrossRef]
- Wang J, Gu R, Kong X, Luan S, Luo YLL. Genome-wide association studies (GWAS) and post-GWAS analyses of impulsivity: A systematic review. Progress in Neuro-Psychopharmacology and Biological Psychiatry, 1109. [CrossRef]
- Fay CR, Toth AL. Is the genetic architecture of behavior exceptionally complex? Current Opinion in Insect Science, 1011. [CrossRef]
- Mehta D, Czamara D. GWAS of behavioral traits. Current Topics in Behavioral Neurosciences, 1: online , 2019, 1 January 2019. [CrossRef]
- Non AL, Cerdeña JP. Considerations, caveats, and suggestions for the use of polygenic scores for social and behavioral traits. Behavior Genetics. [CrossRef]
- Bakken NR, Parker N, Hannigan LJ, et al. Childhood trajectories of emotional and behavioral difficulties are related to polygenic liability for mood and anxiety disorders. Journal of Child Psychology and Psychiatry, 27 October 2024. [CrossRef]
- Joo YY, Lee E, Kim BG, Kim G, Seo J, Cha J. Polygenic architecture of brain structure and function, behaviors, and psychopathologies in children. bioRxiv (Cold Spring Harbor Laboratory), 23 May 2024. [CrossRef]
- Bredström, A. Culture and Context in Mental Health diagnosing: Scrutinizing the DSM-5 revision. Journal of Medical Humanities. [CrossRef]
- Aggarwal, NK. Culture, Communication, and DSM-5 diagnostic Reliability. Journal of the National Medical Association. [CrossRef]
- Andreatta G, Tessmar-Raible K. The still dark side of the Moon: molecular mechanisms of Lunar-Controlled rhythms and clocks. Journal of Molecular Biology, 3525. [CrossRef]
- Cajochen C, Altanay-Ekici S, Münch M, Frey S, Knoblauch V, Wirz-Justice A. Evidence that the Lunar Cycle Influences Human Sleep. Current Biology, 1485. [CrossRef]
- Clauss JA, Avery SN, Blackford JU. The nature of individual differences in inhibited temperament and risk for psychiatric disease: A review and meta-analysis. Progress in Neurobiology. [CrossRef]
- Zimecki, M. The lunar cycle: effects on human and animal behavior and physiology. PubMed, /: https, 1640. [Google Scholar]
- Helfrich-Förster C, Monecke S, Spiousas I, Hovestadt T, Mitesser O, Wehr TA. Women temporarily synchronize their menstrual cycles with the luminance and gravimetric cycles of the Moon. Science Advances. [CrossRef]
- Burnett, C. The Certitude of Astrology: the Scientific Methodology of Al-Qabīsī and Abu MaShar1. Early Science and Medicine. [CrossRef]
- Greenbaum, DG. Hellenistic Astronomy in Medicine. In: BRILL eBooks. ; 2020:350-380. [CrossRef]
- Scofield, B. The nature of astrology: History, Philosophy, and the Science of Self-Organizing Systems. Simon and Schuster; 2023.
- Quinn, VS. Using Astrology to Confront & Discuss Pseudoscience in the Classroom. The American Biology Teacher. [CrossRef]
- Burke, K. Big Five personality traits and astrology: The relationship between the Moon variable and the NEO PI-R. Pacifica Graduate Institute ProQuest Dissertations & Theses, 2012. [Google Scholar]
- Wyman AJ, Vyse S. Science versus the Stars: A Double-Blind test of the validity of the NEO Five-Factor inventory and Computer-Generated Astrological Natal Charts. The Journal of General Psychology. [CrossRef]
- Park SH, Na HJ. A comparative study on the relationship between MBTI personality types and character cards of tarot. Industry Promotion Research, /: https, 2023.
- Case P, Phillipson G. Astrology, Alchemy and Retro-Organization Theory: An Astro-Genealogical critique of the Myers-Briggs Type Indicator®. Organization. [CrossRef]
- Bosch, A. The cycle of the zodiac signs: Archetypes Rooted in Reality. Mijnbestseller.nl; 2022.
- Glas, N. The physiognomy of the temperaments. British Homeopathic Journal. [CrossRef]
- Zhang, X. Traditional Cosmology and Hamlet’s Delay--—Four humours and their activation of his personal character. Studies in Literature and Language. [CrossRef]
- Greenbaum, DG. Temperament: Astrology’s Forgotten Key.
- DeYoung, CG. The neuromodulator of exploration: A unifying theory of the role of dopamine in personality. Frontiers in Human Neuroscience. [CrossRef]
- Pickering A, Gray J. Dopamine, appetitive reinforcement, and the neuropsychology of human learning: An individual differences approach. In: Advances in Individual Differences Research. ; 2001. http://eprints.gold.ac. 8445.
- Humińska-Lisowska, K. Dopamine in Sports: A narrative review on the genetic and epigenetic factors shaping personality and athletic performance. International Journal of Molecular Sciences, 1160. [Google Scholar] [CrossRef]
- Huminska-Lisowska K, Stronska-Pluta A, Suchanecka A, et al. Association study of the Taq1D rs1800498 polymorphism of the DRD2 gene with personality traits in a group of athletes. Baltic Journal of Health and Physical Activity. [CrossRef]
- Apergis-Schoute AM, Van Der Flier FE, Ip SHY, et al. Perseveration and shifting in Obsessive-Compulsive Disorder as a function of uncertainty, punishment, and serotonergic medication. Biological Psychiatry Global Open Science. [CrossRef]
- Marcos-Prieto P, Ordali E, Mariotti V, et al. Dopaminergic and serotonergic genetic variants predict actions and expectations of cooperation and punishment. Scientific Reports. [CrossRef]
- Norden M, Kanarik M, Laugus K, et al. Serotonin release by parachloroamphetamine in rats with high and low sociability: High prefrontal release capacity in sociable females. Journal of Psychopharmacology, 24 September 2024. [CrossRef]
- Rajagopal L, Huang M, Mahjour S, et al. The dopamine D1 receptor positive allosteric modulator, DETQ, improves cognition and social interaction in aged mice and enhances cortical and hippocampal acetylcholine efflux. Behavioural Brain Research, 1147. [CrossRef]
- Rappeneau V, Díaz FC. Convergence of oxytocin and dopamine signalling in neuronal circuits: Insights into the neurobiology of social interactions across species. Neuroscience & Biobehavioral Reviews, 1056. [CrossRef]
- Chen WJ, Chen H, Li ZM, Huang WY, Wu JL. Acetylcholine muscarinic M1 receptors in the rodent prefrontal cortex modulate cognitive abilities to establish social hierarchy. Neuropsychopharmacology. [CrossRef]
- Zhang Y, Karadas M, Liu J, et al. Interaction of acetylcholine and oxytocin neuromodulation in the hippocampus. Neuron, 1862. [CrossRef]
- Zelinka V, Cojan Y, Desseilles M. Hypnosis, Attachment, and Oxytocin:An Integrative perspective1. International Journal of Clinical and Experimental Hypnosis. [CrossRef]
- Linden JH, Sugarman LI, De Benedittis Professor Of G, Varga K. Attachment and hypnosis. Revisiting Our Evolutionary Past to Reconstruct Our Future. The Routledge International Handbook of Clinical Hypnosis.
- Rijnders RJP, Van Den Hoogen S, Van Honk J, Terburg D, Kempes MM. Would you? Effects of oxytocin on moral choices in forensic psychopathic patients. Comprehensive Psychoneuroendocrinology, 1002. [CrossRef]
- Zheng X, Wang J, Yang X, et al. Oxytocin, but not vasopressin, decreases willingness to harm others by promoting moral emotions of guilt and shame. Molecular Psychiatry, 3475. [CrossRef]
- Thadea OSA, Putra ST, Putra IGNGS. The relationship between Galen’s personality type theory and emotional intelligence level. Biomolecular and Health Science Journal. [CrossRef]
- Orion, R. Astrology for Dummies (For Dummies (Sports & Hobbies)).; 2007. http://dl.acm.org/citation.cfm? 1212. [Google Scholar]
- Curth, LH. Astrology and physick. In: Manchester University Press eBooks. ; 2018. [CrossRef]
- Eccles, B. Astrological Physiognomy from Ptolemy to the Present Day. Culture and Cosmos. [CrossRef]
- Corredera, VI. Faces and Figures of Fortune: Astrological Physiognomy in Tamburlaine Part 1. Early Modern Literary Studies, /: https, 1492. [Google Scholar]
- Ziegler, J. The matter of character: the case of pre-modern physiognomy. In: The New Synthese Historical Library. ; 2024:53-71. [CrossRef]
- Kai, Z. The Relationship between Personality Traits and Face Shapes in Chinese Traditional Physiognomy. Open Journal of Social Sciences. [CrossRef]
- Physiognomy at the crossroad of magic, science, and the arts.; 2024. [CrossRef]
- Barlow, N. Mars: An Introduction to its Interior, Surface and Atmosphere. Cambridge University Press; 2008.
- Encrenaz T, Coustenis A. Composition and chemistry of the atmospheres of terrestrial planets: Venus, the Earth, Mars, and Titan. In: Springer eBooks. ; 2017:1-28. [CrossRef]
- Solomon, SC. Mercury: the enigmatic innermost planet. Earth and Planetary Science Letters. [CrossRef]
- Charlier B, Namur O. The origin and differentiation of Planet Mercury. Elements. [CrossRef]
- Chakraborty, U. Effects of different phases of the lunar month on humans. Biological Rhythm Research. [CrossRef]
- Chakraborty U, Ghosh T. A study on the physical fitness index, heart rate and blood pressure in different phases of lunar month on male human subjects. International Journal of Biometeorology. [CrossRef]
- Leroy A, Very E, Birmes P, et al. Intrusive experiences in posttraumatic stress disorder: Treatment response induces changes in the directed functional connectivity of the anterior insula. NeuroImage Clinical, 1029. [CrossRef]
- Dominant Planets & Elements in Natal Chart, Astrology Online Calculator. Astro-Seek.com. https://horoscopes.astro-seek.
- Clinical biomechanics in human locomotion.; 2023. [CrossRef]
- Kiive E, Kanarik M, Veidebaum T, Harro J. Neuropeptide Y gene variants and Agreeableness: interaction effect with the birth cohort and the serotonin transporter promoter polymorphism. Acta Neuropsychiatrica. [CrossRef]
- Qiao Y, Song X, Yan J, et al. Neurological activation during verbal fluency task and resting-state functional connectivity abnormalities in obsessive-compulsive disorder: a functional near-infrared spectroscopy study. Frontiers in Psychiatry. [CrossRef]
- Ghanavati E, Salehinejad MA, Nejati V, Nitsche MA. Differential role of prefrontal, temporal and parietal cortices in verbal and figural fluency: Implications for the supramodal contribution of executive functions. Scientific Reports. [CrossRef]
- Porges SW, Davila MI, Lewis GF, et al. Autonomic regulation of preterm infants is enhanced by Family Nurture Intervention. Developmental Psychobiology. [CrossRef]
- Porter CL, Wouden-Miller M, Silva SS, Porter AE. Marital Harmony and Conflict: links to infants’ emotional regulation and cardiac vagal tone. Infancy. [CrossRef]
- Park G, Kappes A, Rho Y, Van Bavel JJ. At the heart of morality lies neuro-visceral integration: lower cardiac vagal tone predicts utilitarian moral judgment. Social Cognitive and Affective Neuroscience, 1588. [CrossRef]
- De Greck M, Rotte M, Paus R, et al. Is our self based on reward? Self-relatedness recruits neural activity in the reward system. NeuroImage, 2066. [CrossRef]
- Petersen J, Ong CW, Hancock AS, Gillam RB, Levin ME, Twohig MP. An examination of the relationship between perfectionism and neurological functioning. Journal of Cognitive Psychotherapy. [CrossRef]
- Wu D, Wang K, Wei D, et al. Perfectionism mediated the relationship between brain structure variation and negative emotion in a nonclinical sample. Cognitive Affective & Behavioral Neuroscience. [CrossRef]
- Enticott PG, Johnston PJ, Herring SE, Hoy KE, Fitzgerald PB. Mirror neuron activation is associated with facial emotion processing. Neuropsychologia, 2851. [CrossRef]
- Van Overwalle, F. Social cognition and the brain: A meta-analysis. Human Brain Mapping. [CrossRef]
- Walther S, Lefebvre S, Conring F, et al. Limbic links to paranoia: increased resting-state functional connectivity between amygdala, hippocampus and orbitofrontal cortex in schizophrenia patients with paranoia. European Archives of Psychiatry and Clinical Neuroscience, 1021. [CrossRef]
- Deng W, Tuominen L, Sussman R, Leathem L, Vinke LN, Holt DJ. Changes in responses of the amygdala and hippocampus during fear conditioning are associated with persecutory beliefs. Scientific Reports. [CrossRef]
- Liu YC, Deng YC, Zhu ZT, et al. Oxytocin modulates inhibitory balance in the prelimbic cortex to support social memory consolidation during REM sleep. Theranostics, 3257. [CrossRef]
- Walia V, Wal P, Mishra S, Agrawal A, Kosey S, Patil AD. Potential role of oxytocin in the regulation of memories and treatment of memory disorders. Peptides, 1712. [CrossRef]
- Plini ERG, Robertson IH, Brosnan MB, Dockree PM. Locus Coeruleus Is Associated with Higher Openness to Experience and IQ: Implications for the Noradrenergic System for Novelty Seeking in Daily Life. Journal of Cognitive Neuroscience, 1: online , 2024, 13 October 2024. [CrossRef]
- Sivashankar Y, Fernandes M, Oudeyer PY, Sauzéon H. The beneficial role of curiosity on route memory in children. Frontiers in Cognition. [CrossRef]
- Chen H, Xie M, Ouyang M, et al. The impact of illness duration on brain activity in goal-directed and habit-learning systems in obsessive-compulsive disorder progression: A resting-state functional imaging study. Neuroscience. [CrossRef]
- Carli M, Invernizzi RW. Serotoninergic and dopaminergic modulation of cortico-striatal circuit in executive and attention deficits induced by NMDA receptor hypofunction in the 5-choice serial reaction time task. Frontiers in Neural Circuits. [CrossRef]
- Chellew K, Evans P, Fornes-Vives J, Pérez G, Garcia-Banda G. The effect of progressive muscle relaxation on daily cortisol secretion. Stress. [CrossRef]
- Afonso RF, Kraft I, Aratanha MA, Kozasa EH. Neural correlates of meditation a review of structural and functional MRI studies. Frontiers in Bioscience-Scholar. [CrossRef]
- Giannotta G, Ruggiero M, Trabacca A. Chronobiology in Paediatric Neurological and Neuropsychiatric Disorders: Harmonizing Care with Biological Clocks. Journal of Clinical Medicine, 7737. [CrossRef]
- Coury SM, Lombroso A, Avila-Quintero VJ, et al. Systematic review and meta-analysis: Season of birth and schizophrenia risk. Schizophrenia Research. [CrossRef]
- Albiñana C, Boelt SG, Cohen AS, et al. Developmental exposure to vitamin D deficiency and subsequent risk of schizophrenia. Schizophrenia Research. [CrossRef]
- Duan C, Jenkins ZM, Castle D. Therapeutic use of melatonin in schizophrenia: A systematic review. World Journal of Psychiatry. [CrossRef]
- Zhang R, Volkow ND. Seasonality of brain function: role in psychiatric disorders. Translational Psychiatry. [CrossRef]
- McCarthy MJ, Gottlieb JF, Gonzalez R, et al. Neurobiological and behavioral mechanisms of circadian rhythm disruption in bipolar disorder: A critical multi-disciplinary literature review and agenda for future research from the ISBD task force on chronobiology. Bipolar Disorders. [CrossRef]
- Luo H, Cheng J, Zhang Z, et al. Seasonal patterns in Chinese population: Validating the seasonal pattern assessment questionnaire and exploring associations with psychiatric diagnoses and biological rhythms. Chronobiology International. [CrossRef]
- Lewis P, Gottlieb JF, Morfeld P, Hellmich M, Erren TC. Perinatal photoperiod associations with bipolar disorder and depression: A systematic literature review and cross-sectional analysis of the UK Biobank database. Psychiatry Research, 1158. [CrossRef]
- McCarty R, Josephs T, Kovtun O, Rosenthal SJ. Correction: Enlightened: addressing circadian and seasonal changes in photoperiod in animal models of bipolar disorder. Translational Psychiatry. [CrossRef]
- Majrashi NA, Alyami AS, Shubayr NA, Alenezi MM, Waiter GD. Amygdala and subregion volumes are associated with photoperiod and seasonal depressive symptoms: A cross-sectional study in the UK Biobank cohort. European Journal of Neuroscience, 1388. [CrossRef]
- Xavier NB, Abreu ACVO, Amando GR, et al. Chronobiological parameters as predictors of early treatment response in major depression. Journal of Affective Disorders. [CrossRef]
- Hyndych A, El-Abassi R, Mader EC. The role of sleep and the effects of sleep loss on cognitive, affective, and behavioral processes. Cureus, 16 May 2025. [CrossRef]
- Polat, Y. Examination of interdisciplinary personality profiles in context of Financial Behaviors. DergiPark (Istanbul University), /: online , 2022. https, 31 July 2022. [Google Scholar]
- Raji H, Dinesh S, Sharma S. Inside the impulsive brain: a narrative review on the role of neurobiological, hormonal and genetic factors influencing impulsivity in psychiatric disorders. The Egyptian Journal of Neurology Psychiatry and Neurosurgery. [CrossRef]
- Zeng J, Yan J, Cao H, et al. Neural substrates of reward anticipation and outcome in schizophrenia: a meta-analysis of fMRI findings in the monetary incentive delay task. Translational Psychiatry. [CrossRef]
- Kohlhoff J, Cibralic S, Hawes DJ, Eapen V. Oxytocin receptor gene (OXTR) polymorphisms and social, emotional and behavioral functioning in children and adolescents: A systematic narrative review. Neuroscience & Biobehavioral Reviews, 1045. [CrossRef]
- Ono Y, Zhang X, Noah JA, Dravida S, Hirsch J. Bidirectional connectivity between Broca’s area and Wernicke’s area during interactive verbal communication. Brain Connectivity. [CrossRef]
- Gilbertson H, Fang L, Andrzejewski JA, Carlson JM. Dorsal anterior cingulate cortex intrinsic functional connectivity linked to electrocortical measures of error monitoring. Psychophysiology. [CrossRef]
- Brockett AT, Roesch MR. Anterior cingulate cortex and adaptive control of brain and behavior. International Review of Neurobiology, 2: online , 2020, 16 December 2020. [CrossRef]
- Clairis N, Lopez-Persem A. Debates on the dorsomedial prefrontal/dorsal anterior cingulate cortex: insights for future research. Brain, 4826. [CrossRef]
- Shan Q, Tian Y, Chen H, Lin X, Tian Y. Reduction in the activity of VTA/SNc dopaminergic neurons underlies aging-related decline in novelty seeking. Communications Biology. [CrossRef]
- Hu Y, Pereira AM, Gao X, et al. Right temporoparietal junction underlies avoidance of moral transgression in autism spectrum disorder. Journal of Neuroscience, 1699. [CrossRef]
- Yeshurun Y, Nguyen M, Hasson U. The default mode network: where the idiosyncratic self meets the shared social world. Nature Reviews Neuroscience. [CrossRef]
- Abraham E, Zagoory-Sharon O, Feldman R. Early maternal and paternal caregiving moderates the links between preschoolers’ reactivity and regulation and maturation of the HPA-immune axis. Developmental Psychobiology, 1482. [CrossRef]
- Caria A, Dall’Ò GM. Functional Neuroimaging of Human hypothalamus in Socioemotional Behavior: a Systematic review. Brain Sciences. [CrossRef]
- Evangelou, G. The neuropsychology of free will: An expensive mechanism. Psych-Talk. [CrossRef]
- Schou WS, Ashina S, Amin FM, Goadsby PJ, Ashina M. Calcitonin gene-related peptide and pain: a systematic review. The Journal of Headache and Pain. [CrossRef]
- Popescu ER, Semeniuc S, Hritcu LD, et al. Cortisol and oxytocin could predict covert aggression in some psychotic patients. Medicina. [CrossRef]
- Mbiydzenyuy NE, Qulu LA. Stress, hypothalamic-pituitary-adrenal axis, hypothalamic-pituitary-gonadal axis, and aggression. Metabolic Brain Disease, 1613. [CrossRef]
- Andelin L, Reynolds S, Schoen S. Effectiveness of Occupational therapy using a Sensory Integration Approach: a Multiple-Baseline Design Study. American Journal of Occupational Therapy. [CrossRef]
- Kearney BE, Lanius RA. The brain-body disconnect: A somatic sensory basis for trauma-related disorders. Frontiers in Neuroscience. [CrossRef]
- Checa A, Alvear M, Navas E, et al. RELATIONSHIP BETWEEN HYPOTHALAMUS AND SEXUAL ORIENTATION. MNJ (Malang Neurology Journal). [CrossRef]
- Zhou X, Li A, Mi X, et al. Hyperexcited limbic neurons represent sexual satiety and reduce mating motivation. Science, 6634. [CrossRef]
- Smallwood J, Bernhardt BC, Leech R, Bzdok D, Jefferies E, Margulies DS. The default mode network in cognition: a topographical perspective. Nature Reviews Neuroscience. [CrossRef]
- Saputra RRT, Saputri RAM, Mardiyyah A. Alat Ukur STIFIn untuk Mengukur Perilaku Pemimpin dalam Bisnis. LITERATUS. [CrossRef]
- Poniman, F. A Grand Theory of STIFIn Personality: Basic Functions Theory Revisited. International Journal of Psychosocial Rehabilitation. [CrossRef]
- Davies G, Welham J, Chant D, Torrey EF, McGrath J. A systematic review and meta-analysis of Northern Hemisphere season of birth studies in schizophrenia. Schizophrenia Bulletin. [CrossRef]
- Torrey EF, Miller J, Rawlings R, Yolken RH. Seasonality of births in schizophrenia and bipolar disorder: a review of the literature. Schizophrenia Research. [CrossRef]
- Chotai J, Serretti A, Lattuada E, Lorenzi C, Lilli R. Gene–environment interaction in psychiatric disorders as indicated by season of birth variations in tryptophan hydroxylase (TPH), serotonin transporter (5-HTTLPR) and dopamine receptor (DRD4) gene polymorphisms. Psychiatry Research. [CrossRef]
- Chotai J, Smedh K, Johansson C, Nilsson LG, Adolfsson R. An epidemiological study on gender differences in self-reported seasonal changes in mood and behaviour in a general population of northern Sweden. Nordic Journal of Psychiatry. [CrossRef]
- Chotai J, Lundberg M, Adolfsson R. Variations in personality traits among adolescents and adults according to their season of birth in the general population: further evidence. Personality and Individual Differences. [CrossRef]
- Chotai J, Smedh K, Nilsson LG, Adolfsson R. A dual vulnerability hypothesis for seasonal depression is supported by the seasonal pattern assessment questionnaire in relation to the temperament and character inventory of personality in a general population. Journal of Affective Disorders. [CrossRef]
- Chotai J, Wiseman R. Born lucky? The relationship between feeling lucky and month of birth. Personality and Individual Differences, 1451. [CrossRef]
- Kolevzon A, Gross R, Reichenberg A. Prenatal and perinatal risk factors for autism. Archives of Pediatrics and Adolescent Medicine. [CrossRef]
- Landgraf D, McCarthy MJ, Welsh DK. Circadian clock and stress interactions in the molecular biology of psychiatric disorders. Current Psychiatry Reports. [CrossRef]
- Hühne A, Welsh DK, Landgraf D. Prospects for circadian treatment of mood disorders. Annals of Medicine. [CrossRef]
- Chen YC, Sheen JM, Tiao MM, Tain YL, Huang LT. Roles of melatonin in fetal programming in compromised pregnancies. International Journal of Molecular Sciences, 5380. [CrossRef]
- Smals AGH, Kloppenborg PWC, Benraad ThJ. CIRCANNUAL CYCLE IN PLASMA TESTOSTERONE LEVELS IN MAN. The Journal of Clinical Endocrinology & Metabolism. [CrossRef]
- Blazer, DG. Depression in Late Life: review and commentary. The Journals of Gerontology Series A. [CrossRef]
- Wehr TA, Duncan WC, Sher L, et al. A circadian signal of change of season in patients with seasonal affective disorder. Archives of General Psychiatry, 1108. [CrossRef]
- Wehr TA, Giesen HA, Schulz PM, et al. Contrasts between symptoms of summer depression and winter depression. Journal of Affective Disorders. [CrossRef]
- Langevin HM, Churchill DL, Cipolla MJ. Mechanical signaling through connective tissue: a mechanism for the therapeutic effect of acupuncture. The FASEB Journal, 2275. [CrossRef]
- Somanadhapai S, Yadav SS, Rana S. Application of environics color slides in optimizing the health and well-being. Recent Updates in Disease and Health Research. [CrossRef]
- Priya SP, Rao P, Padmanabhan V, Chaitanya NCSK. Principles and Therapeutics of Yajna. Journal of Pharmacy and Bioallied Sciences, S: 5), 4271. [CrossRef]
- Choudhary, M. Pain Management in Ardhavbhedaka (Migraine Headache) through Adhipati and Krikatika Marma Therapy. International Journal for Research in Applied Science and Engineering Technology. [CrossRef]
- Kantermann T, Sung H, Burgess HJ. Comparing the Morningness-Eveningness questionnaire and Munich ChronoType questionnaire to the Dim Light melatonin onset. Journal of Biological Rhythms. [CrossRef]
- Davies, EA. The interaction of specific frequency bands in the geomagnetic field diurnal spectrum, with specific frequency bands in the human heart rate variability diurnal spectrum. Research Square (Research Square), 26 July 2022. [Google Scholar] [CrossRef]
- Ramishvili A, Janashia K, Tvildiani L. High heart rate variability causes better adaptation to the impact of geomagnetic storms. Atmosphere, 1707. [CrossRef]
- Blanco-Prieto, O. , Catalán, J., Catalán, J., Trujillo-Rojas, L., Peña, A., Rivera del Alamo, M. M., Llavanera, M., Bonet, S., Fernández-Novell, J. M., Yeste, M., & Rodríguez-Gil, J. E. (2020). Red LED Light Acts on the Mitochondrial Electron Chain of Mammalian Sperm via Light-Time Exposure-Dependent Mechanisms. Cells, 912), 2546. [CrossRef]
- Herrera, M. , Caldeira da Silva, C. C., Baptista, M. S., & Kowaltowski, A. J. (2024). Mitochondrial Fatty Acid Oxidation is Stimulated by Red Light Irradiation. [CrossRef]
- Keller, H. Decolonizing psychology as a scientific and ethical necessity. Online Readings in Psychology and Culture. [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



