Submitted:
03 July 2025
Posted:
14 July 2025
You are already at the latest version
Abstract
Amphibians are particularly vulnerable to hydric stress due to their permeable skin, biphasic life cycle, and strong dependence on aquatic and moist terrestrial environments. In the North-Western Mediterranean Basin—one of Europe’s most climate-sensitive regions—the intensification of droughts associated with climate change poses a critical threat to amphibian populations. Increased aridification, either due to higher temperatures or to more frequent, prolonged, and severe drought episodes can affect both aquatic and terrestrial life stages, directly altering breeding opportunities, larval development, post-metamorphic survival, and dispersal capacity. This review aims to gather and synthesize current knowledge on the ecological, physiological, and demographic impacts of drought on amphibians of the North-Western Mediterranean across habitat types, including ephemeral ponds, permanent water bodies, lotic systems, and terrestrial landscapes. Drought-induced shifts in hydroperiod can drastically reduce reproductive success and accelerate larval development with fitness consequences, while on land, desiccation risk and habitat degradation could limit access to refugia and fragment populations by reducing structural connectivity. These environmental constraints are compounded by the interactions between drought and emerging infectious diseases. We discuss the current knowledge on how chytrid fungi (Batrachochytrium dendrobatidis and B. salamandrivorans) and ranaviruses may respond to temperature and moisture regimes, and how drought may affect their transmission dynamics, host susceptibility, and pathogen persistence. In these cases, microbiome disruption, pollutant concentration, and increased contact rates between species may amplify disease outbreaks under dry conditions, but a better understanding of the multifactorial effects of drought on amphibian biology and disease ecology is needed for predicting species vulnerability, identifying high-risk populations, and guiding future conservation and management strategies in Mediterranean environments.
Keywords:
drougths
; Amphibia
; lotic
; lentic
; emerging diseases
; climatic change
; North-Western Mediterranean Basin
; hydroperiod
1. Introduction
Climate change due to increases in human greenhouse gas emissions since the beginning of the industrial revolution up to the present day is now considered one of the major threats to biodiversity and one which underlies several reported extinction events [1,2,3]. Climate is rapidly changing, and the trend towards increased global temperatures has been forecasted to continue for the coming decades [4], together with the risk of droughts. Model projections for the 2090s indicate that the proportion of the global land surface in extreme drought is predicted to increase by a factor of 10 to 30. Although species have demonstrated resilience to important climate shifts throughout evolutionary history, the current rate of global changes is challenging the ability of organisms to adapt rapidly. This poses serious risks to wildlife communities, potentially driving biodiversity losses [5]. Importantly, human-induced climate change is not only altering the mean temperature but also the patterns of precipitation and evapotranspiration, including extreme weather events such as heatwaves, floods, wildfires and droughts [6,7,8,9]. In addition, global trends towards aridification can further compromise biodiversity through changes in land use and overexploitation of hydric resources, leading to unpredictable changes in water variability and availability [10,11].
Droughts, characterized by anomalously low precipitation, abnormal soil moisture deficits, and excessive evapotranspiration [8,12,13], are a major environmental stressor with diverse potential effects on wildlife communities. They can impact wildlife directly by inducing physiological stress that affects survival and fecundity. Additionally, they exert indirect effects by altering habitat structure [14], resource availability [15], and interspecific interactions. These can include shifts in microbial soil composition [16], forest die-offs [17], the proliferation of invasive species [18], and the destabilization of species interactions due to changes in competition and trophic dynamics [19,20]. Drought effects depend on its duration and intensity [15,16,17,18,19,20,21]. In this sense, the percentage of land under extreme drought, defined as the 1% most severe drought conditions that occurred between 1950 and 2000, is expected to rise from 1% today to 30% by the end of the 21st century according to climate models [22]. Estimates for the southwestern United States [23] suggest that several ecoregions will see large increases in drought exposure, with 29 out of 84 ecoregions being subject to increases of over 200%, and 19 ecoregions being subject to increases over 400%. Concerning the effects of extreme weather and climate events [15], increases in their frequency, severity and/or duration can trigger resource bottlenecks that act as powerful demographic constraints on terrestrial fauna, often exacerbating other anthropic pressures such as land use change and driving populations to local extinction or marked declines. Bottlenecks caused by drought were found for most taxa, but interestingly they accounted for all three recorded impacts on amphibians.
Climate change is impacting water availability in the regions surrounding the Mediterranean Sea. The Western Mediterranean has been identified [8,24,25,26,27] as one of the regions with a moisture deficit from of oceanic moisture source from the North Atlantic source. Most studies show negative consequences of climate change in soil moisture, aquifer recharge, irrigation demand, hydrological extremes (droughts, low flows), and water and soil quality (nutrient concentration, soil salinity, soil erosion). Projections generally agree on an increased duration, frequency and severity of droughts in the Mediterranean Basin [13,28]. However, recent studies [29] show that precipitation trends over the past one and a half century are negligible, and that the precipitation in the area is dominated by high temporal variability. Despite this, increasing aridity in the region is likely unavoidable and to be driven primarily by rising temperatures and enhanced evapotranspiration. In this sense, Mediterranean climate change has been observed at a magnitude exceeding global means [30]. The regional temperature increase is higher than the global mean temperature increase, with a peak at 1.9°C above 1950 temperature in 2022. Thus, it is important to highlight that, regardless of the presence, or not, of significant trends in precipitation in the Mediterranean Basin, the region is undergoing a progressive process of climatic aridification. This trend is primarily driven by an increase in atmospheric evaporative demand associated with rising temperatures and potentially compounded by changes in precipitation patterns. As a result, the region is experiencing increasingly severe ecological and agricultural droughts [8,12,13].
The Western Mediterranean Basin is not only home to droughts but also to a great use of water resources. For instance, Western Mediterranean countries are among the most productive agricultural supplies mainly thanks to the combination of high temperatures and the irrigation of water-consuming crops. Withdrawals for irrigation are high because of the evaporative demand for crops grown in the Mediterranean climate [31]. The demand for irrigation water in the eastern and southern countries is often over 50% and reaches almost 90% in some countries [32]. In these countries, agriculture is the primary water-consuming sector, with high-demand crops. In addition, the great increase of tourism in the last decades [33], poses an added pressure in the consumption of water resources for personal and recreational activities mainly in summer [33], which is not only the driest and hottest season, but also it is the season whose climate is becoming even more arid [34]. Heatwaves and droughts interact with various water uses, sometimes creating a positive feedback loop. For example, during droughts, agricultural water consumption often increases, further exacerbating water scarcity [34]. Additionally, the increase in forest cover in non-cultivated areas can also decrease the amount of water that reaches streams, ponds, and can become groundwater. In Catalonia (North-Eastern Spain), the average annual temperature has already increased by 1.9 °C and the average annual rainfall has decreased by about 104 mm compared to the middle of the 20th century [35], with an increased duration and intensity of drought periods, while the increase in forest cover has greatly increased. In fact, FOREStime report [36] estimates a 29% reduction of blue water between 1990 and 2014 in the North-Eastern Iberian Peninsula, including Catalonia. All this leads to a scenario of great water scarcity similar to other areas with a similar climate (e.g. California, see [17,36], where less water will be available for different uses, while the increased vegetation cover will demand more water to compensate for a warmer and drier atmosphere.
Amphibians are among the most threatened animal groups, and a recently updated assessment [37] reports that their conservation status keeps worsening globally. Most species lay eggs in the water, which hatch into aquatic larvae that metamorphose into land-living juveniles. Drought can heavily impact amphibian species that rely on water availability for larval development, with direct and indirect effects on larval survival and postmetamorphic fitness [38,39]. Soil moisture is important also for most amphibian postmetamorphic age-classes which rely on underground microrefugia [40], or for many amphibians’ species which obtain water directly from the substrate [41]. Thus, droughts are related to increased mortality rates in amphibian populations [42], altered community compositions [10,43,44], and even decreases in biodiversity and functional diversity [45]. Thus, while amphibian species may have to endure smaller increases of drought episodes compared to the other vertebrate groups [23], this may nevertheless imply much more detrimental effects in amphibians than on other faunal groups.
Drought is expected to have the most severe consequences on amphibians [46]. According to [47], drought events have increased in several areas with high amphibian diversity, including much of the Amazon basin, the Atlantic Forest of Brazil, Madagascar, southwestern United States, northern Mexico, and also continental Europe [47]. Despite this, surprisingly in amphibians the risks associated with more intense or recurrent dry periods remain relatively understudied compared to the other groups [48].
Concerning the North-Western Mediterranean Basin, while local species are adapted to survive and endure these temporarily arid conditions, it remains uncertain whether they can adapt to an increasing frequency and intensity of dry periods under changing water availability conditions [49]. In Europe, the frequency and magnitude of droughts and heatwaves have increased over the last 50 yr, especially in the southern and western regions [50,51]. These events, in addition to the levels of water demand by human activities, will drive water scarcity in the North-Western Mediterranean Basin, determining an important part of the negative effects on the amphibian communities, which will synergistically act together with other factors such as habitat loss and fragmentation, pollutants, emerging diseases, and invasive species.
In this review, we assess the potential impact of the reduction of water availability and the possible increase of severe droughts in the amphibian community of the North-Western Mediterranean Basin, highlighting the risks for this crucial element of the fresh-water biodiversity in the long-term. We focus on how the loss of superficial water and moisture can affect amphibian assemblages in lentic, lotic and terrestrial habitats, and also on how it can interact with the spread of emergent diseases, like chytridiomycosis and ranavirosis, which can be a very important conservation issue in the future.
2. General Susceptibility of the Amphibian Community to Drought
The amphibian community inhabiting North-Western Mediterranean basin is under the domain of the Mediterranean climate with hot and dry summers, mild wet winters and high spatio-temporal variability of precipitation, which is mainly concentrated in autumn and secondarily in spring. Usually, the amphibian community in this area is subject to two well-characterised hydrological phases. A wet period, mainly from autumn to spring, and a dry period concentrated mainly in summer. However, there is great inter-annual variability in the duration and intensity of these periods. The adaptability of the local amphibian community to changes in the wet-dry regime probably depends on many intrinsic and extrinsic factors, determining different susceptibilities (Supplementary Table S1). These different susceptibilities most probably imply a phylogenetic signal, similar to the signal found in thermal tolerances [52].
Since aridity periods are predicted to increase in frequency and intensity [13,28], this will put a strain on those species that are not plastic or adapted enough to face the diminished water availability for the Mediterranean Region [24,26,47,53] identifies Western Europe as one of the regions in which amphibian species are most exposed to droughts (Figure 1).
By the end of the 21st century, the combined effects of rising temperatures, reduced precipitation, and increasing pressure on water resources are expected to drive the expansion of the Mediterranean region’s drier ecosystems into areas further north and at higher altitudes [54]. It has been observed that many species of amphibians in the Iberian Peninsula have already moved up in altitude towards cooler and more moist environments [55]. Changes in predicted water availability to the end of XXI century for the Western Mediterranean (Figure 2, data extracted from [26]), predict a general decrease in water availability and an increase of water demand.
Local anuran larvae are typically primary consumers or opportunistic omnivores, whereas urodela larvae tend to be primarily predatory [39]. More importantly, the length of the larval phase frequently determines species-specific habitat requirements [39,56,57]. Pond drying is one of the main risks for amphibians in these communities. Species have specific requirements regarding the hydroperiod, which defines a trade-off between predation, competition, and desiccation risk [38]. For instance, the preferred hydroperiod can be short to avoid potential predators like fish or large dragonfly larvae but must be long enough to allow larval development to reach completion, if not met, at the cost of high levels of larval mortality and reproductive failure. Thus, prolonged periods of drought most probably will affect the rates of colonization and extinction, driving changes in occupancy and metapopulation dynamics in some species [46,58,59], like for instance in the observed fecundity and survival at different ontogenetic stages in Bombina variegata [60], where population growth is strongly influenced by drought frequency. Decreases in fecundity can also be accompanied by changes in larval survival, larval size, growth and time of metamorphosis [61,62]. At the landscape level, droughts can alter amphibian ranges and breeding phenology with consequences for reproductive success [63].
On the other hand, most pond-breeding adult amphibians spend little time in breeding ponds, and most of their lifetime is spent in terrestrial habitats that may, or not, be directly adjacent to the breeding site. Each species and age-class may show different strategies to survive during severe droughts in terrestrial habitats. The intrinsic dispersal capacity of each species is important for migrating to areas with water, even over large distances. Theoretically, high dispersal rates, high fecundity, and a short lifespan are associated to metapopulations experiencing unpredictable environments and presumably more adapted and resilient to droughts, while a very low dispersal rate, low fecundity and a long lifespan are associated to populations occupying predictable environments [60]. Accordingly, small toadlets and adults of Epidalea calamita migrate distances of several hundred meters within a short time [64,65], which may significantly help prevent local extinction in sink populations. Hyla molleri and Pelobates cultripes are capable of covering accumulated displacements up to 3.5 and 1.8 km respectively [66]. Similarly, Pelophylax perezi has high dispersal capacities [67] that allows the species to maintain genetic structure between far waterbodies. Contrarily however, other amphibian species have low dispersal capacities but may be very resilient thanks to longer lifespans. It has been suggested that high survival of adult D. fuscus [68], coupled with their temporary emigration, may compensate for the negative effects of drought on larvae and facilitate resilience of this species to drought conditions. In fact, long lifespans were identified as the main factor allowing long-term persistence of a population or marbled salamander under the stress of occasional catastrophic reproductive failures due to low precipitation [69]. Similarly, urodeles in the North-Western Mediterranean can live more than 10 years [70,71,72,73], which could be a crucial feature to allow a population to overcome long droughts.
In most amphibian species only a small fraction of the individuals perform long-distance dispersal events while short distance displacements are much more frequent [65,67,74,75]. Usually, this small proportion of the population is enough to maintain genetic and metapopulational structures. However, droughts also produce habitat loss and fragmentation [39], hindering the connectivity along breeding sites, which is an important factor which could determine the survival of metapopulations. Landscape configuration conditions colonization and extinction probabilities and is very important for the persistence of “sink” populations [76,77].
Finally, changes in temperature or precipitation can potentially influence the timing of amphibian reproduction [56,78]. An earlier onset of spring breeding behaviour correlated with a warming climate has been observed among various pond-breeding frogs and toads in Europe [79,80,81,82,83]. [84] through a meta-analysis of global amphibian phenological data, concluded that this group is strongly influenced by climate change. Although they did not identify consistent patterns in demographic parameters such as abundance, survival, breeding success, or morphology, they found that drought and elevated temperatures can significantly alter amphibian phenology—advancing, delaying, or even preventing the onset of breeding. Such shifts in breeding timing can disrupt ecosystem processes and force larval development to occur under suboptimal conditions, leading to increased intraspecific competition and reduced resource availability [83].
3. Potential Effects of Extreme Droughts
3.1. Lakes and Ponds
The hydrology of lakes and ponds, whether from natural or human origin, is generally characterized by being dependent on the hydrological and rainfall regime in one small area. That is, if we exclude large artificial reservoirs and the mountain lakes located in the main West European mountain ranges, most of the lentic environments on the Western Mediterranean area are quite sensitive to long periods of drought due to natural climate variations, anthropogenic climate change, or water overexploitation.
Most amphibians present in the Western Mediterranean can reproduce in lentic environments. It is usually assumed that these communities are structured along a series of abiotic and biotic gradients that determine the abundance of each species [85,86,87]. Among all factors, hydroperiod is arguably considered the most relevant. Studies on amphibian communities in temperate regions agree that pond breeding amphibian communities are organized along the hydroperiod gradient, ranging from permanent to temporary and ephemeral ponds (see e.g. [85,88,89]). While long hydroperiods support more complex faunal communities and promote the selection of predator-avoidance traits in amphibian larvae [58], ephemeral environments select for traits related to rapid development and efficient foraging instead, as the risk of reproductive failure primarily depends on hydroperiod length [90]. In an intermediate situation, temporary ponds show varying levels of desiccation and predation risk, with potentially important competition levels. In Mediterranean regions, temporary ponds are the main breeding habitats for several amphibian species [90,91]. In these environments the alternation of dry and wet periods determines the resident community, as both terrestrial and aquatic species need to have strategies to resist dry periods and droughts, such as the ability to migrate, advance metamorphosis, increase growth rate or resistance behaviours. This combination of factors makes Mediterranean temporary ponds unique in its high biodiversity, becoming specially protected systems listed as priority habitats by the Habitats Directive code 3170 [92].
The position of a pond along this gradient each year largely depends on short- and long-term rainfall patterns, creating an unpredictability to which many species have adapted. This includes showing several types of phenotypic plasticity in larvae, and an important ability to optimize selection of oviposition sites and flexible strategies in the timing of the onset of breeding in adults [56,90,93,94,95,96,97]. For instance, most temperate amphibians can show antipredator morphologies if there are predators present [98] or accelerate the timing of metamorphosis to escape drying waters. However, all these shifts have trade-offs, and in the case of premature metamorphosis developmental acceleration comes at the cost of reduced body size, altered body proportions, and possible impairments of the immune function in the recently metamorphosed individual [38,58,88], which may even have downstream effects on susceptibility to disease [63,99].
Worryingly, prolonged droughts will directly and repeatedly affect the hydroperiod gradient that underpins this diversity and structures most amphibian communities, and plasticity not only entails costs but also has limits, meaning that several species may be pushed beyond their adaptive capacity if droughts become more frequent and intense [100]. While the Western Mediterranean basin is characterized by pronounced interannual variability in both the amount and timing of rainfall, it is already known that extreme events can strongly shape community composition. During a prolonged period of abnormally low rainfall in Doñana Natural Park (2011–2022), an exacerbated unpredictability in the onset of inundation of the temporary ponds led to interannual differences in amphibian community composition [39]. In the long term, an increased frequency or length of droughts either tend to favour the most dominant species [57,101,102], or leads to a direct loss of the available habitats and to a reduction of abundance and diversity of amphibians [39,59,103,104].
On the positive side, droughts can lead to the disappearance of invasive fish species from many sites, often without subsequent recolonization, which are associated with reduced amphibian occupancy in areas where these exotics are present. This suggests that droughts may offer a window of opportunity for invasive species control which may be one of the main problems for amphibian population dynamics [10,105]. For instance, [106] modelled the effects of various threats on the larval development of Epidalea calamita—a species that breeds mainly in ephemeral and temporary ponds—and found that predation pressure from invasive species had a stronger negative impact on tadpole guilds than changes in temperature or precipitation. This highlights that the presence of invasive predators poses a greater threat to the species than climate change itself. Supporting this, recent monitoring in Catalonian natural parks showed this species increased both in abundance and occupancy during the 2021–2024 drought period [107]. Conversely, if droughts do not reduce the presence of invasive fish and the remaining aquatic habitats continue to host these species, the combined stressors may lead to sharp declines in several amphibian populations. The same study [107], along with others, found that species dependent on longer hydroperiods were negatively affected in the short term by the reduced availability of suitable habitats, in some cases resulting in local extinctions [10,69,108].
Beyond the direct effects of altered hydroperiods (Figure 3), many other ecological factors associated with drying habitats can indirectly affect amphibian behaviour, physiology, and fitness [109]. Amphibians that breed in ephemeral or temporary ponds are especially exposed to fluctuations in temperature and precipitation, which can alter larval density, food availability, and host–pathogen interactions [46,63]. When these changes lead to metamorphosis at smaller body sizes [38,59,89], this will most probably entail costs in terms of survival in the first terrestrial life stages [110]. Shifts in reproductive phenology can also influence the outcome of competitive and predatory interactions, thereby potentially altering community dynamics in assemblages of co-existing species, beyond direct effects on individual mortality and survival [111,112].If multiple species that previously segregated temporally are increasingly forced to breed synchronously, this may intensify both interspecific competition and predation pressure – particularly if predators are also constrained to breed or forage within the shortened periods of inundation – potentially leading to unpredictable shifts in community dynamics.
In coastal freshwater ecosystems, the effects of drought can be particularly severe due to salinization. Proximity to the sea can lead to increased salt concentrations in aquatic habitats during drought periods [113,114,115]. Although salinization also occurs inland—mainly as a result of aquifer overexploitation and prolonged droughts [116,117] in coastal wetlands, it is especially exacerbated by decreasing precipitation, rising temperatures, and seawater intrusion. Drought periods disrupt the balance and dynamics between fresh and saltwater in coastal areas, a process that can drive significant changes in amphibian and aquatic communities [118]. Most amphibians in Western Europe have complex life cycles that include aquatic stages—eggs, larvae, and in some species, even adults—that are frequently exposed to increasingly saline conditions. Some species can reproduce in slightly brackish or low-salinity coastal habitats [114], but amphibians are generally highly susceptible to elevated salinity due to their permeable skin and limited osmoregulatory capacity [119]. Salinity can negatively impact reproductive performance, reducing sperm viability, fecundity, and embryo survival, with early life stages being particularly vulnerable [115,120,121]. Even moderate salinity can prolong development and alter hormone activity in larvae, depressing thyroid function, affecting osmoregulatory hormones, and altering antioxidant responses [119,122,123,124]. These physiological disruptions can lead to slower growth, reduced activity and foraging efficiency, and ultimately extended larval periods [125,126,127,128,129,130,131]. Larvae exposed to salinity may also exhibit reduced swimming performance and speed, increasing their susceptibility to predation [131,132,133,134]. As a result, repeated increases in salinity can compromise the normal survival of cohorts because saline and drought-stressed conditions will coincide with shorter hydroperiods and increased predator density. In Western Europe, certain populations of Pleurodeles waltl, Triturus marmoratus, T. cristatus, Lissotriton helveticus, Discoglossus pictus, D. galganoi, Pelobates cultripes, P. fuscus, Hyla meridionalis, Epidalea calamita, Bufo spinosus, Bufotes balearicus, B. viridis and Pelophylax perezi have demonstrated a degree of physiological adaptation to osmotically stressful environments, allowing some populations to persist in mildly or occasionally saline habitats [113,115,125,135,136,137,138,139], but no amphibian species is known to permanently inhabit highly saline environments.
3.2. Rivers, Streams, and Brooks
Under Mediterranean climate conditions, most watercourses exhibit a seasonal flow regime, with a wet phase occurring during late autumn, winter, and spring, and a dry phase that typically takes place in summer [140,141]. While permanent rivers in the region experience substantial fluctuations in discharge, they rarely dry out completely—the dry phase in these systems usually corresponds to reduced flow or water volume. In contrast, temporary rivers may break into a series of disconnected pools or even dry up entirely for part of the year. At the most extreme end of the gradient, ephemeral streams flow only briefly, typically following rainfall events [141,142]. Most of the research has historically focused on permanent rivers, despite that temporary rivers are common watercourses throughout the world [142,143,144]. They are especially abundant in the Mediterranean basin, where they are currently more studied [145].
It is commonly accepted that temporary rivers support lower overall biodiversity than permanent ones, as the latter tend to exhibit more complex physical structures and contain a wider variety of microhabitats [146]. Temporary rivers are characterized by marked interannual variation in the onset and duration of their dry phases, which can jeopardize the survival of species not adapted to endure prolonged droughts or flash floods. Due to these constraints, temporary rivers can host unique biodiversity and often act as refugia for specialized species [147]. Their intermittent nature may also make them more resistant to biological invasions, such as those by exotic fish [148,149], but see [150,151]. Similarly to temporary ponds, these systems can be frequently considered ‘hotspots’ of regional biodiversity and can play a key role in maintaining the ecological integrity of river networks [147]. Nevertheless, many are under significant pressure due to water overexploitation [152,153]. Climate change models predict a global increase in the extent of intermittent rivers, with formerly perennial rivers in some regions already transitioning to temporary flow regimes [142,154]. However, in this context temporary rivers themselves are becoming increasingly vulnerable. The combined effects of reduced precipitation and growing human water demands are intensifying the frequency and duration of no-flow periods in many areas [155,156]. Intensifying drought conditions are expected to reduce hydrological connectivity, limiting species dispersal and disrupting metapopulation dynamics [157]. Even species typically regarded as resilient may be at risk if extended dry phases—or shifts in their seasonal timing—become decoupled from species’ life-history strategies [158,159].
Although the Western Mediterranean does not host a high number of stream-specialist amphibians, those that do occur typically have high conservation value and occupy unique ecological niches. Newts of the genera Calotriton and Euproctus are typically protected under national or regional conservation frameworks, yet they face increasing threats – that similarly affect other stream-dwelling anurans, such as Rana pyrenaica – from rising water temperatures, more frequent extreme rainfall events, the introduction of non-native fish species into the few stream sections that still retain water , and water overexploitation [160,161,162]. In contrast to species inhabiting lentic habitats, those adapted to lotic environments tend to have more limited dispersal abilities [163,164,165]. This is likely because riverine systems are geologically more stable, and thus, evolutionary pressures have not favoured long-distance dispersal [165,166,167]. As running waters are generally more persistent than small lentic water bodies, populations within them are less reliant on dispersal for survival, often resulting in higher levels of genetic isolation, even in small distances [163,168]. Conversely, species from lentic environments tend to exhibit greater dispersal capabilities due to the unpredictable hydroperiods associated with rainfall dependency and, on average, occupy larger geographical ranges than species restricted to more stable lotic systems [166]. As a result, the increasing disruption of water continuity within and among river systems—primarily driven by more frequent droughts—may have severe consequences for the diversity and persistence of lotic-adapted species. A notable example is the Montseny Brook Newt (Calotriton arnoldi), listed as Critically Endangered, which is currently facing an extremely vulnerable situation in parts of its already restricted range due to markedly reduced stream flows [162]. This decline in water availability is likely the result of interacting pressures, including climate change, land-use transformations, and excessive water abstraction [162]. Moreover, the situation is exacerbated by the species’ extremely limited dispersal capacity and the near-total absence of connectivity among its fragmented populations [168], creating a highly precarious scenario for the long-term survival in some brooks.
On the other hand, for more generalist amphibian species that inhabit both lentic and lotic environments, the main stressors and ecological trade-offs across biotic and abiotic gradients in river systems may broadly mirror those observed in ponds. In lentic habitats, permanent waters typically harbour a higher density of amphibian predators and competitors, while temporary systems are associated with greater desiccation risk [169]. However, unlike the more predictable gradient found in ponds, temporary rivers can also support fish species that are adapted to survive in isolated pools during dry periods [147,170]. Moreover, due to their limited baseflow and rapid hydrologic response to rainfall, temporary streams often experience more pronounced flood peaks than perennial systems [171,172,173]. In a recent study on the distribution of generalist amphibians in a Mediterranean stream system, [169] identified water availability as the primary factor driving both abundance and diversity. Although water availability was positively correlated with fish presence, the study suggests that in temporary rivers, amphibians that tend to avoid fish in pond environments may successfully evade predation by exploiting microhabitats shaped by natural river dynamics—features that are largely absent in ponds. While this points to a potentially favourable scenario for amphibians able to inhabit temporary river systems, the study also found that water availability clearly determined the presence of species typically associated with permanent aquatic habitats, pointing that droughts may affect these species in a manner similar to how they impact fish populations, in some cases even leading to local extinctions [108].
In addition to reductions in water quantity, climate change also poses significant threats to water quality. During dry spells, the capacity of temporary streams and rivers to dilute pollutants is substantially reduced, impairing their ability to buffer the effects of continuous wastewater discharges. While effluent volumes typically remain stable throughout the year, the dilution potential of receiving water bodies is highly variable leading to sharp increases in pollutant concentrations during drought events [174]. This issue is not adequately addressed in current water management frameworks [175]. Among pollutants, glyphosate has received particular attention in amphibian research, with sublethal effects documented across all developmental stages [176]. However, the extent to which glyphosate-based herbicides and other pesticides contribute to amphibian declines remains unclear, largely due to a lack of data on actual exposure levels in natural populations [176]. Although the dynamics of pesticide presence and concentration in Mediterranean freshwater systems are not yet fully understood [177], there is growing concern that prolonged droughts may exacerbate the accumulation and persistence of these substances, thereby intensifying their ecological impact [178].
Finally, it is important to note that even though amphibians also use temporary rivers and in most cases are more able to colonize them than fish, they are not considered yet in the environmental quality indices of rivers and streams (Ecological Quality Ratios -EQRs-), based on ecological status classifications of the Water Framework Directive (WFD), Directive 2000/60/EC of the European Parliament and of the Council of 23 October 2000 (https://eur-lex.europa.eu/eli/dir/2000/60/oj).
3.3. Terrestrial Habitat
Most adult amphibians in the Western Mediterranean spend the majority of the year in terrestrial habitats, returning to aquatic environments only during the breeding season. In terrestrial settings, they are particularly vulnerable to water scarcity due to their permeable skin and reliance on moist microhabitats for hydration and cutaneous respiration. During droughts or extended dry periods, dehydration stress increases, compromising physiological processes such as gas exchange, locomotion, and aerobic activity [41]. Reduced soil moisture and the desiccation of terrestrial refuges can lower survival rates of post-metamorphic and adult individuals. For instance, in Bombina variegata, a severe drought led to a 12% and 10% reduction in juvenile and adult survival, respectively [60]. Similarly, in Rana sylvatica, adult survival was positively correlated with monthly rainfall [61]. In Ambystoma tigrinum [179], male survival during a drought year in breeding ponds was up to 54% lower compared to survival in adjacent forest habitats. Recent modelling studies further underscore the impact of climate change on amphibian activity levels. [46] showed that, under future scenarios combining increased temperatures and drought conditions, the potential activity of different frog ecotypes was markedly reduced. Notably, reduced rainfall had a greater negative effect on activity than temperature increases alone. Stream-dwelling and semiaquatic ecotypes were particularly assessed to be susceptible to desiccation stress, likely due to their higher dependency on continuous water availability compared to arboreal or fossorial species. Droughts will likely also reduce dispersal opportunities by limiting surface activity, thereby decreasing the probability of successful emigration and colonization of new habitats [108].
Beyond the immediate limitation posed by reduced atmospheric moisture, extreme droughts can have long-term impacts on the terrestrial habitats inhabited by adult amphibians. Severe and recurrent drought events lead to widespread tree mortality and alter forest structure [44,180,181] which can significantly affect survival and activity during the most vulnerable life stages of some species [182,183,184]. Certain terrestrial microhabitats—such as dead logs, woody debris, and fallen trees that retain soil moisture—can function as hydrological refuges, reducing mortality among both juveniles and adults [184]. Thus, on a more positive note, some drought episodes may increase the volume of deadwood, a key component of forest maturity. e.g. [185,186]. This deadwood contributes to soil moisture retention and enhances the forest’s capacity to withstand future droughts, while supporting the diversity of understory organisms [187]. However, the broader trend in a context of increasing drought and fire frequency is the replacement of moist forest ecosystems by scrublands, or the shift from drought-sensitive to drought-adapted tree species. These changes typically result in more open habitats, where amphibians are more exposed to desiccation stress [188,189,190]. Among the habitats most critical in this changing landscape are riparian environments, which represent ecotones between terrestrial and aquatic systems and maintain unique moist conditions even within a fully Mediterranean context [191]. These areas provide essential terrestrial refuges for both adults and juveniles, as well as foraging and dispersal routes for many amphibian and non-amphibian species. Riparian habitats are therefore crucial for amphibian persistence during drought periods [192,193] and help buffer the impacts of increasing droughts by enhancing connectivity and ecosystem resilience [194]. In fact, such environments now serve as the last refuges for several populations of Atlantic or montane amphibian species persisting under Mediterranean climatic conditions, such as several locations of Rana iberica or low-elevation populations of Calotriton asper and deserve special attention from a conservation perspective.
Droughts also increase the risk of forest fires. Drier and hotter conditions, combined with the expected accumulation of forest deadwood, are likely to raise the frequency and severity of wildfires [4,195,196,197]. In the Western Mediterranean Basin, before 1970, wildfires were primarily driven by fuel accumulation resulting from farmland abandonment and rural depopulation [196]. However, since 1970, wildfires have become more strongly linked to climatic variables associated with drought, particularly high temperatures and low precipitation, with significant fire–climate relationships in the same region in recent decades [198]. The effects of wildfires on amphibian populations have been studied worldwide, but mainly in biomes classified as temperate forests [184,198,199]. Interestingly, in a review in 2021 [199], no evidence of a predictable response to anuran communities to fire in any biome is reported, finding that 20% of studies reported positive effects, 26% observed negative effects and the majority of 47% of the studies did not detect a significant effect of fire on anuran assemblages. Thus, while fire can impact amphibian communities through both direct and indirect mechanisms, the final outcomes may depend on fire- and species-specific singularities. Thus, the impact of wildfires on amphibians should be assessed separately for short- and long-term on individuals, species and habitats [200,201]. While direct mortality occurs during the fire, the main long-term effects result from rapid changes in habitat structure [202]. Fires cause direct mortality of amphibians in their terrestrial phase, indirect mortality or displacement due to sudden habitat alteration, larval mortality due to chemical changes in breeding ponds, or by direct loss of breeding sites through siltation caused by slope erosion [201]. After these initial effects, forest fires reduce soil water retention capacity and interception (green water), and increase soil erosion and evaporation [203], which collectively create a post-fire environment which is similar to aridification or drought conditions. In the Western Mediterranean this usually promotes expansion of scrubland at the expense of forest cover [188]. If fires are occasional, this can lead to an interesting spatial mosaic of burned and unburned habitats hosting different faunal communities [204,205]. However, if fires are repeated or larger in extension, a shift in dominant species may occur, a fact that has already been hinted in amphibian communities of the area, in which Mediterranean amphibian species are promoted by fires in areas limiting with the Atlantic bioregion [202]. This is supported by the close link between vegetation structure and faunal assemblages in burned landscapes [206,207]. For instance, the decline in the number of pond-breeding species in a Mediterranean Area which hosted some vegetation-associated species, was inversely related to the percentage of vegetation burned [201]. Conversely, an increase in species richness and larval densities was observed in unburned areas, likely due to adult migration from nearby burned zones. In the longer term, if the reduction in vegetation cover is permanent can lead to decreased soil moisture, which is detrimental to amphibians that rely on moist environments [48,207,208,209]. Therefore, most probably the main effect of fire in the Western Mediterranean amphibian communities will be to act synergistically with droughts [188,189], accelerating the substitution of habitats and facilitating or accelerating community shifts. In this context, Atlantic or sub-mediterranean species marginally inhabiting the Mediterranean biome might be the most impacted by the increase of forest fires. Species like Lissotriton helveticus or Rana iberica [202], which are of Atlantic affinity but maintain populations under Mediterranean climate, or vegetation- or forest-associated species like Hyla meridionalis [210], or for instance Salamandra salamandra [201], will probably be the most severely affected.
4. Emerging Diseases and Drought
Currently, fifteen emerging diseases have been described in European amphibians [211]. Among these, two are of fungal origin — Batrachochytrium dendrobatidis (Bd, [212]) and Batrachochytrium salamandrivorans (Bsal, [213]) — which cause the disease chytridiomycosis, and one is of viral origin (Ranavirus, [214]). Together, these three pathogens account for the majority of mortality events associated with emerging diseases in European amphibians. In the Western Mediterranean, several chytridiomycosis outbreaks have been historically recorded, related to either Bd (e.g., [215,216]), Bsal [217], or Ranaviruses [218]. The intensity, prevalence, and lethality of these pathogens depend on both intrinsic species-related factors—such as individual susceptibility, microbiota composition, and immune status—and environmental factors, including temperature, precipitation, altitude, and drought, among others (Figure 4). These factors can act independently, but they are often interrelated, acting simultaneously and sometimes synergistically or in a mutually dependent manner. This is not only for the direct effects of diseases but also for the possible synergies with other stressors. For instance, an increase of death rates in adult specimens could result in a truncated age structure, which may erode the capacity of populations to overcome consecutive recruitment failures during long droughts [219]. To date, few studies have investigated how environmental stressors such as drought may interact with infectious diseases to amplify amphibian declines [220].
4.1. Batrachochytrium dendrobatidis (Bd)
The growth and survival of Batrachochytrium dendrobatidis (Bd) are strongly temperature-dependent. Its optimal growth range lies between 17 °C and 25 °C, while temperatures below 10 °C and above 28 °C significantly reduce or halt growth. Zoospores die within 4 hours at 37 °C [221,222,223,224,225]. Bd is also highly sensitive to desiccation, exhibiting 100% mortality after just 3 hours of complete drying. In addition, the pathogen poorly tolerates salinity; exposure to a 5% NaCl solution is lethal [221,226]. Beyond these physical constraints, other environmental factors may influence Bd virulence, prevalence, and host susceptibility [219]. These characteristics, together with intraspecific variability (e.g., [227]) and interspecific or individual differences in host response (e.g., [228]), make it particularly challenging to predict the pathogen’s effects on individual fitness and population dynamics.
Theoretically, ephemeral and temporary ponds should remain free of Bd during extended dry periods, whereas permanent ponds can act as pathogen reservoirs [229]. From this perspective, severe droughts could limit Bd dispersal and persistence. However, zoospores may survive in microscale moist environments—or even in fog or cloud layers under very humid conditions [230,231]—and, most importantly, the fungus can persist on the skin of amphibians during their terrestrial phase, or in/on other aquatic or semi-aquatic animal structures [230,232,233]. This allows Bd to recolonize ponds once water becomes available again, often from nearby aquatic habitats [232]. For instance, recently the painted frog (Discoglossus pictus)—an introduced species in north-eastern Spain and south-eastern France—has been identified as a potential Bd reservoir for ephemeral and temporary aquatic habitats, potentially facilitating the persistence of Bd in such environments in which this species usually breeds. Very high infection loads have been reported in the species [234], and according to [235], the usually syntopic Natterjack toad Epidalea calamita exhibits higher zoospore loads (as estimated by qPCR) when in sympatry with D. pictus.
Interestingly, the dynamics of Bd may differ in lotic systems (streams and rivers) and in more aquatic species, where the pathogen appears to spread more effectively during summer, when anurans concentrate in and around the remaining water sources [236]. Under these conditions, the probability of zoospore transmission increases due to higher animal density, and mass mortality events are more likely, as infection intensity is usually elevated [236,237]. Conversely, during wetter periods, increased water flow in streams may dilute zoospore concentrations, thus reducing transmission efficiency [238]. Nevertheless, the effect of dilution may be counteracted by humid conditions, which could enhance transmission through alternative infection routes [236].
In addition to the pathogen’s capacity to persist and recolonize, the host’s ability to mount an immune response adds further complexity to future disease dynamics. For example, infection load in individuals of Bufo spinosus was positively correlated with the time individuals spent in aquatic environments [239]. Moreover, drought conditions may reduce the abundance of Bd-inhibiting bacteria in the amphibian skin microbiome, rendering toadlets more vulnerable to infection, [240]. Finally, similar to the trade-offs seen with premature metamorphosis—such as smaller body size or reduced immune function—early emergence may increase susceptibility to pathogens [63]. These findings highlight the complex, multifactorial responses of native amphibians to severe drought events in relation to Bd (see e.g., [241]), underlining the need to evaluate multiple, interacting stressors when assessing the drivers of amphibian reproductive and disease patterns.
4.2. Batrachochytrium salamandrivorans Bsal
In Europe, Bsal has been detected in captive Salamandra salamandra populations in Belgium, Germany, Spain, the Netherlands, and the United Kingdom, and in wild populations in some regions of Belgium, Germany, and the Netherlands. In the Western Mediterranean basin, Bsal has so far only been detected in wild populations of Triturus marmoratus and Salamandra salamandra at a single locality in Spain, likely introduced via exotic species [217]. The Western Mediterranean hosts a high diversity of urodeles, many of which show high susceptibility to Batrachochytrium salamandrivorans (Bsal), at least under laboratory conditions (see Table 1).
The outlook for Bsal under rising temperature conditions is less favourable than for Bd, as Bsal’s optimal growth occurs between 10 °C and 15 °C, and its zoospores die at temperatures of 25 °C or higher [242,243]. Under global warming scenarios, it is assumed that under both realistic (+2 °C) and extreme (+4 °C) warming conditions, Bsal will exhibits minimal growth rates during the summer [244]—the driest period in the Mediterranean. Thus, in principle extremely hot and dry periods will be the most detrimental to Bsal survival. However, Bsal has a dual transmission, with environmentally resistant non-motile encysted spores in addition to the motile zoospores which are similar to B. dendrobatidis’ [242,245]. It is still unclear if the resistant encysted spores may display resistance to dry periods, but some data suggests that the pathogen may be a relatively poor disperser [246]. Bsal infection is transmitted through direct contact between individuals or via water. [243] found that the probability of transmission upon contact between infected and uninfected newts was very high (>90%), even at early stages of infection. Although reducing host density and increasing habitat complexity may help reduce transmission, such conditions are rarely met during drought periods, when individuals tend to aggregate in limited moist refuges. For example, [247] modelled Bsal growth for Salamandra salamandra, predicting that optimal pathogen proliferation should occur in summer and autumn—precisely when Mediterranean amphibian activity is typically minimal until the first autumn rains. Under this scenario, species- or population-specific responses to Bsal infection during droughts remain uncertain, and the timing and intensity of possible dispersion events and outbreaks under a Mediterranean climate will likely depend on the composition and connectivity of the local amphibian community, plus the duration and onset of rainfall or drought conditions.
4.3. Ranaviruses
Ranaviruses are also considered emerging pathogens, meaning their impact on amphibian populations is recent and mostly linked to increases in host range, geographic spread, or incidence (e.g., [248]). There is strong evidence of host switching between vertebrate classes, including amphibians, reptiles, and fish [248,249,250], with reports of simultaneous infections in sympatric fish and frog populations [251,252]. In the Iberian Peninsula, the Common Midwife Toad Virus (CMTV), together with Frog Virus 3 (FV3), has caused mass mortality events since the late 1980s. Climate warming may be triggering CMTV outbreaks [253], supporting the hypothesis of its endemic status in the region.
Climate change is widely recognised as a driver of disease emergence, particularly in regions where host–pathogen associations were well established [254,255]. Rising temperatures, increased frequency of droughts, and shifts in seasonal rainfall patterns can significantly alter ranavirus transmission dynamics. Higher temperatures have been associated with increased virus replication, incidence, and mortality [255], and outbreaks have become more frequent during historic warming events such as those in the 1990s. Modelling studies predict that future warming and more frequent droughts will expand the temporal and spatial extent of ranavirus outbreaks, potentially affecting larval recruitment severely [253,254,255]. Interestingly, although outbreaks are generally more frequent at high temperatures [253], some studies report greater lethality of infections at lower temperatures [256,257], indicating that temperature effects may vary depending on host species, life stage, and strain.
Environmental stressors associated with climate change—such as drought, contamination, and altered microbial environments—may further increase susceptibility to ranavirus. Amphibian larvae exposed to pesticides in water are more likely to contract and develop severe infections than those in uncontaminated habitats [258]. Reduced water levels concentrate contaminants, potentially leading to immunosuppression and heightened infection risk [259,260,261,262,263]. In parallel, drought can alter skin microbiome composition, which plays a key role in immune defense. Studies have shown that the stability of the amphibian skin microbiome depends on the environmental microbial reservoir, which can be disrupted by climatic and anthropogenic pressures [264,265]. This disruption may increase inter- and intraspecific variation in susceptibility to ranavirus – or other pathogen – infections across different populations.
Similarly to chytrid fungus, drought may also indirectly promote or hinder host contact by concentrating amphibians in fewer aquatic habitats – thereby facilitating interspecific interactions and increasing the likelihood of host switching – or either diminishing amphibian stays in the water and reducing infections. In Mediterranean regions desiccation risk in temporary and ephemeral ponds can induce premature metamorphosis in most species, but this could increase disease vulnerability in tadpoles at Gosner stages 44–46 stages, which are shown to be particularly vulnerable to ranavirus infection due to intrinsic immunosuppression during metamorphosis [266,267].
Finally, climate warming is also expected to facilitate ranavirus spread to higher altitudes. Rana temporaria, a species living near the southern limit of its range in Mediterranean mountain areas, has been shown to be highly susceptible to ranavirus, and could experience an increase of disease linked mortality in future warming scenarios [268]. Despite well-documented outbreaks and severe population declines in Iberia in recent years [217], the long-term epidemiology of ranavirus is a complex interaction among phenology, population dynamics, habitat heterogeneity, and climate change [218,219,262], which deserves to be further studied.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, A.M. and E.P.-B.; validation, A.M. and E.P.-B.; investigation, A.M. and E.P.-B.; resources, A.M. and E.P.-B.; writing—original draft preparation, A.M. and E.P.-B.; writing—review and editing, A.M. and E.P.-B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript.
| Bd | Batrachochytrium dendrobatidis |
| Bsal | Batrachochytrium salamandrivorans |
References
- Thomas, C.; Cameron, A.; Green, R. et al. Extinction risk from climate change. Nature 2004. 427, 145–148. [CrossRef]
- Parmesan, C. Ecological and Evolutionary Responses to Recent Climate Change. Annual Review of Ecology, Evolution, and Systematics 2006, 37, 637- 669. [CrossRef]
- Thuiller, W.; Lavergne, S.; Roquet, C. et al. Consequences of climate change on the tree of life in Europe. Nature 2011, 470, 531–534. [CrossRef]
- IPCC, 2023: Climate Change 2023: Synthesis Report. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change [Core Writing Team, H. Lee and J. Romero (eds.)]. IPCC, Geneva, Switzerland, 2023, 184 pp. [CrossRef]
- Pörtner, H.O.; Scholes, R.J.; Agard, J.; Archer, E. et al. IPBES-IPCC Co-Sponsored Workshop Report on Biodiversity and Climate Change 2021. [CrossRef]
- Dai. A. Increasing drought under global warming in observations and models. Nature Climate Change 2013, 3, 52–58. [CrossRef]
- Chen, D.; Norris, J.; Thackeray, C.; Hall, A. Increasing precipitation whiplash in climate change hotspots. Environmental Research Letters 2022, 17, 12, 124011. [CrossRef]
- Jézéquel, A.; Faranda, D.; Drobinski, P.; Lionello, P. (), Extreme Event Attribution in the Mediterranean. Int. J. Climatol. 2025, e8799. [CrossRef]
- Swain, D.L.; Prein, A.F.; Abatzoglou, J.T.; Albano, C.M.; Brunner, M.; Diffenbaugh, N.S. et al. Hydroclimate volatility on a warming Earth. Nature Reviews Earth & Environment 2025, 6, 1, 35-50. [CrossRef]
- Moss, W.E.; McDevitt-Galles, T.; Muths, E.; Bobzien, S.; Purificato, J.; Johnson, P. Resilience of native amphibian communities following catastrophic drought: Evidence from a decade of regional-scale monitoring. Biological Conservation 2021, 263, 109352. [CrossRef]
- Zhang, W.; Zhou, T.; Wu, P. Anthropogenic amplification of precipitation variability over the past century. Science 2024, 385, 427-432. [CrossRef]
- Lionello, P.; Scarascia, L. The Relation of Climate Extremes With Global Warming in the Mediterranean Region and Its North Versus South Contrast.” Regional Environmental Change 2020, 20, no. 31. [CrossRef]
- Tramblay, Y.; Koutroulis, A.; Samaniego, L.; Vicente-Serrano, S.M.; Volaire, F.; Boone, A.; Le Page, M.; Llasat, M.C. et al. Challenges for drought assessment in the Mediterranean region under future climate scenarios. Earth-Science Reviews 2020, 210, 103348. [CrossRef]
- Wen, L.; Saintilan, N.; Reid, J.R.; Colloff, M.J. Changes in distribution of waterbirds following prolonged drought reflect habitat availability in coastal and inland regions. Ecol. Evol. 2016, 6, 6672–6689. [CrossRef]
- Maron, M.; McAlpine, C.A.; Watson, J.E.; Maxwell, S.; Barnard, P. Climate-induced resource bottlenecks exacerbate species vulnerability: a review. Divers. Distrib. 2015, 21, 731–743. [CrossRef]
- Canarini, A.; Schmidt, H.; Fuchslueger, L. et al. Ecological memory of recurrent drought modifies soil processes via changes in soil microbial community. Nat. Commun. 2021 12, 5308. [CrossRef]
- Steel, Z. L.; Jones, G.M.; Collins, B.M. et al. Mega-disturbances cause rapid decline of mature conifer forest habitat in California. Ecol. Appl. 2023, 33, e2763. [CrossRef]
- Everard, K.; Seabloom, E.W.; Harpole, W.S.; De Mazancourt, C. Plant water use affects competition for nitrogen: why drought favors invasive species in California. Am. Nat. 2010, 175, 85–97. [CrossRef]
- Cavin, L.; Mountford, E.P.; Peterken, G.F.; Jump, A.S. Extreme drought alters competitive dominance within and between tree species in a mixed forest stand. Funct. Ecol. 2013, 27, 1424–1435. [CrossRef]
- Ledger, M.E.; Brown, L.E.; Edwards, F.K.; Milner, A.M.; Woodward, G. Drought alters the structure and functioning of complex food webs. Nat. Clim. Change 2013, 3, 223–227. [CrossRef]
- Prugh, L.R.; Deguines, N.; Grinath, J.B. et al. Ecological winners and losers of extreme drought in California. Nature Clim Change 2018, 8, 819–824. [CrossRef]
- Burke, E.J.; Brown, S.J.; Christidis, N. Modeling the recent evolution of global drought and projections for the twenty-first century with the Hadley Centre climate model. J. Hydrometeorol. 2006, 7, 1113–1125. [CrossRef]
- Van den Bosch, M.; Costanza, J.K.; Peek, R.A. et al. Climate change scenarios forecast increased drought exposure for terrestrial vertebrates in the contiguous United States. Commun. Earth Environ. 2024, 5, 708. [CrossRef]
- Iglesias, A.; Garrote,L.; Flores, F.; Moneo, M. Challenges to manage the risk of water scarcity and climate change in the Mediterranean, Water Resour. Manage. 2007, 21, 5, 775–788. [CrossRef]
- García-Ruiz, J.M.; López-Moreno, J.I.; Vicente-Serrano, S.M.; Lasanta-Martínez, T.; Beguería. S. Mediterranean water resources in a global change scenario. Earth Sci. Rev. 2011, 105, 3–4, 121–139. [CrossRef]
- Eekhout, J.P.C.; Nunes, J.P.; Tramblay, Y.; de Vente, J. Severe Impacts on Water Resources Projected for the Mediterranean Basin. WIREs Water 2025, 12, e70012. [CrossRef]
- Gimeno-Sotelo, L.; Sorí, R.; Nieto, R. et al. Unravelling the origin of the atmospheric moisture deficit that leads to droughts. Nat Water 2024, 2, 242–253. [CrossRef]
- Barrera-Escoda, A.; Gonçalves, M.; Guerreiro, D. et al. Projections of temperature and precipitation extremes in the North Western Mediterranean Basin by dynamical downscaling of climate scenarios at high resolution (1971–2050). Climatic Change 2014, 122, 567–582. [CrossRef]
- Vicente-Serrano, S. M.; Tramblay, Y.; Reig, F.; González-Hidalgo, J. C.; Beguería, S.; Brunetti, M.; ... Potopová, V. High temporal variability not trend dominates Mediterranean precipitation. Nature 2025, 639, 658-666. [CrossRef]
- Drobinski, P.; B. Azzopardi, H. Ben Janet Allal, et al. 2020. “Energy Transition in the Mediterranean.” In: Climate and Environmental Change in the Mediterranean Basin—Current Situation and Risks for the Future. First Mediterranean Assessment Report, edited by W. Cramer, J. Guiot, and K. Marini, 265–322. Union for the Mediterranean, Plan Bleu, UNEP/MAP. [CrossRef]
- Thivet, G; Fernández, S. Water demand management: the Mediterranean experience. Technical Focus Paper. Global Partnership Group. Plan Bleu. 2020. 74pp. Available on: https://planbleu.org/en/publications/water-demand-management-the-mediterranean-experience/.
- Blinda, M.; Margat, J. Ressources et demandes en eau en région méditerranéenne. Situations et perspectives. Proceedings of Congrès mondial de l’eau 2008. 14pp Available on: https://iwra.org/proceedings/congress/resource/abs326_article.pdf.
- Toth, E.;Neri, M. Tourism water demand modelling in Mediterranean cities under current and future climate, EGU General Assembly 2024, Vienna, Austria, 14–19 Apr 2024, EGU24-18575. [CrossRef]
- Howard, B. California Drought Spurs Groundwater Drilling Boom in Central Valley. National Geographic 2014, Archived from the original on August 16, 2014. Available on: https://www.nationalgeographic.com/culture/article/140815-central-valley-california-drilling-boom-groundwater-drought-wells.
- BAIC. Butlletí Anual d’Indicadors Climàtics (BAIC 2023). Servei Meteorològic de Catalunya 2023, Report, 127pp. https://www.meteo.cat/wpweb/climatologia/butlletins-i-episodis-meteorologics/butlleti-anual-dindicadors-climatics/.
- Blanqué, M; de Cáceres, M.; García-Valdés, M.; Martínez-Vilalta, J.; Roces-Díaz, J.V.; Vayreda, J. FOREStime. Canvis dels serveis ecosistèmics dels boscos de catalunya al llarg dels darrers 25 anys (període 1990-2014). Oficina Catalana del Canvi Climàtic-CREAF-CTFC. Report. 35pp. https://canviclimatic.gencat.cat/web/.content/02_OFICINA/publicacions/publicacions_de_canvi_climatic/Estudis_i_docs_adaptacio/FORESTIME.PDF.
- Luedtke, J.A.; Chanson, J.; Neam, K.; Hobin, L.; Maciel, A.O.; Catenazzi, A.; et al. Ongoing declines for the world’s amphibians in the face of emerging threats. Nature 2023, 622, 308-314. [CrossRef]
- Richter-Boix, A.; Llorente, G.A.; Montori, A. Effects of phenotypic plasticity on post-metamorphic traits during pre-metamorphic stages in the anuran Pelodytes punctatus. Evol. Ecol. Res. 2006a, 8, 309-320. Available on: https://www.evolutionary-ecology.com/issues/v08n02/iiar1861.pdf.
- Díaz-Paniagua, C.; Florencio, M.; de Felipe, M.; Ramírez-Soto, M.; Román, I.; Arribas, R. Groundwater decline has negatively affected the well-preserved amphibian community of Doñana National Park (SW Spain). Amphibia-Reptilia 2024, 45, 2, 205-217. [CrossRef]
- Giacometti, D.; Tattersall, G.J. Putting the energetic-savings hypothesis underground: fossoriality does not affect metabolic rates in amphibians. Evol. Ecol. 2023, 37, 761-777. [CrossRef]
- Hillman, S.S. et al. Ecological and environmental physiology of amphibians. Oxford University Press, Oxford, UK. 2010, Oxford, 2008; online edn, Oxford Academic, 1 Apr. 2010, accessed 9 Apr. 2025. [CrossRef]
- Hillman, J.C.; Hillman, A.K.K.; Mortality of wildlife in Nairobi National Park, during the drought of 1973–1974. African Journal of Ecology 1977, 15, 1–18. [CrossRef]
- McMenamin, S.K.; Hadly, E.A.; Wright, C.K. Climatic change and wetland desiccation cause amphibian decline in Yellowstone National Park, Proc. Natl. Acad. Sci. U.S.A. 2008, 105, 44, 16988-16993. [CrossRef]
- Batllori, E. Lloret, F.; Aakala, T. et al.. Forest and woodland replacement patterns following drought-related mortality. Proc. Natl. Acad. Sci. U.S.A. 2020, 117, 47, 29720-29729.
- Aguirre-Gutiérrez, J.; Malhi, Y.; Lewis, S.L. et al. Long-term droughts may drive drier tropical forests towards increased functional, taxonomic and phylogenetic homogeneity. Nature Communications 2020, 11, 1–10. [CrossRef]
- Walls, S.C.; Barichivich, W.J.; Brown, M.E. Drought, Deluge and Declines: The Impact of Precipitation Extremes on Amphibians in a Changing Climate. Biology, 2013, 2, 1, 399-418. [CrossRef]
- Twomey, E.; Sylvester, F.; Jourdan, J.; Hollert, H.; Schulte, L. M. Quantifying exposure of amphibian species to heat waves, cold spells, and droughts. Conservation Biology 2025, e70074. [CrossRef]
- Wu, N.C.; Bovo, R.P.; Enriquez-Urzelai, U. et al. Global exposure risk of frogs to increasing environmental dryness. Nat. Clim. Chang. 2024, 14, 1314–1322. [CrossRef]
- Parmesan, C.; Root, T.L.;Willig, M.R. Impacts of extreme weather and climate on terrestrial biota. Bulletin of the American Meteorological Society 2000, 81, 443–450.
- Fink, A.H.; Brücher, T.; Krüger, A.; Leckebusch, G.C.; Pinto, J. G. Ulbrich, U. The 2003 European summer heatwaves and drought–synoptic diagnosis and impacts. Weather 2004, 59, 209–216. [CrossRef]
- Vautard, R.; Yiou, P.; D’Andrea, F. et al. Summertime European heat and drought waves induced by wintertime Mediterranean rainfall deficit. Geophysical Research Letters 2007, 34, 1–5. [CrossRef]
- Pottier, P.; Kearney, M.R.; Wu, N.C. et al. Vulnerability of amphibians to global warming. Nature 2025, 639, 954–961. [CrossRef]
- García-Ruiz, J.M.; López-Moreno, J.I.; Vicente-Serrano, S.M.; Lasanta-Martínez, T.; Beguería. S. Mediterranean water resources in a global change scenario. Earth Sci. Rev. 2011, 105, 3–4, 121–139. [CrossRef]
- Peñuelas, J.; Boada, M. A global change-induced biome shift in the Montseny mountains (NE Spain). Global Change Biology 2003, 9, 131-140. [CrossRef]
- Enriquez-Urzelai, U.; Bernardo, N.; Moreno-Rueda, G.; Montori, A.; Llorente, G.A. Are amphibians tracking their climatic niches in response to climate warming? A test with Iberian amphibians. Climatic Change 2019, 154, 289–301. [CrossRef]
- Díaz-Paniagua, C. Variability in timing of larval season in an amphibian community in SW Spain. Ecography 1992, 15, 267-272. https://www.jstor.org/stable/3683156.
- Díaz-Paniagua, C.; Gómez-Rodríguez, C.; Portheault, A.; Florencio, M. Why a system of heterogeneous temporary ponds favours amphibian communities? Amphibians in Doñana National Park, an example of preserved breeding habitats. In: International Conference On Mediterranean Temporary Ponds, 2009, 245-254. Fraga, I.; Arguimbau, P.; Eds, Consell Insular Menorca, Recerca 14. Maó Menorca.
- Richter-Boix, A.; Llorente, G.A.; Montori, A. A comparative study of predator-induced phenotype in tadpoles across a pond permanency gradient. Hydrobiologia 2007b, 583, 43-56. [CrossRef]
- Scheele, B.C.; Driscoll, D.A.; Fischer, J.; Hunter, D.A. Decline of an endangered amphibian during an extreme climatic event. Ecosphere 2012, 3, 11, 101. [CrossRef]
- Cayuela, H.; Arsovski, D.; Bonnaire, E.; Duguet, R.; Joly, P.; Besnard, A. The impact of severe drought on survival, fecundity, and population persistence in an endangered amphibian. Ecosphere 2016, 7, 2, e01246. [CrossRef]
- Berven, K.A. Factors affecting population fluctuations in larval and adult stages of the wood frog (Rana sylvatica). Ecology 1990, 71, 1599–1608. [CrossRef]
- Tejedo, M.; F. Marangoni, C.; Pertoldi, A.; Richter-Boix, A.; Laurila, G.; Orizaola, A. G. Nicieza, D. Alvarez, and I. Gomez-Mestre.. Contrasting effects of environmental factors during larval stage on morphological plasticity in post-metamorphic frogs. Climate research 2010, 43, 31–39.
- Kohli, A.K.; Lindauer, A.L.; Brannelly, L.A.; Ohmer, M.E.B.; Richards-Zawacki, C.; Rollins-Smith, L.; Voyles, J. Disease and the Drying Pond: Examining Possible Links among Drought, Immune Function, and Disease. Development in Amphibians. Physiological and Biochemical Zoology 2019, 92, 3, 339-348. [CrossRef]
- Sinsch, U. Postmetamorphic dispersal and recruitment of first breeders in a Bufo calamita metapopulation. Oecologia 1997, 112, 42–47. [CrossRef]
- Sinsch, U.; Oromi, N.; Miaud, C.; Denton, J.; Sanuy, D. Spatial ecology of natterjack toads. Anim. Conserv. 2012, 15, 388-396. [CrossRef]
- Martínez-Gil, H.; Sánchez-Montes, G.; Montes-Gavilán, P. et al. Fine-scale functional connectivity of two syntopic pond-breeding amphibians with contrasting life-history traits: an integrative assessment of direct and indirect estimates of dispersal. Conserv. Genet. 2023, 24, 361–374. [CrossRef]
- Capellà-Marzo. B.; Sánchez-Montes, G.; Martínez-Solano, I. Contrasting demographic trends and asymmetric migration rates in a spatially structured amphibian population. Integr. Zool. 2020, 15, 482–497. [CrossRef]
- Price, S.J.; Browne, R.A.; Dorcas, M.E. Resistance and Resilience of A Stream Salamander To Supraseasonal Drought. Herpetologica 2012, 68, 3, 312–323. [CrossRef]
- Taylor, B.E.; Scott, D.E.; Gibbons, J.W. Catastrophic reproductive failure, terrestrial survival, and persistence of the marbled salamander. Conserv. Biol. 2006, 20, 792–801. [CrossRef]
- Montori, A.; Herrero, P. Caudata. In: Amphibia-Lissamphibia García-París, M; Montori, A.; Herrero, P. Ramos, M.A. et al. Eds. Museo Nacional de Ciencias Naturales. CSIC, Madrid. ISBN: 84-00-08292-3. Fauna Ibérica 2004, 24, 43-275.
- Grossenbacher, K; Thiesmeier, B. et al. Eds. Handbuch der Reptilien und Amphibien Europas. Band 4/I: Schwanzlurche (Urodela) I. AULA-Verlag. 1999, 409pp.
- Grossenbacher, K; Thiesmeier, B. et al. Eds. Handbuch der Reptilien und Amphibien Europas. Band 4/IIa Schwanzlurche (Urodela) II. AULA-Verlag 2003, 351pp.
- Grossenbacher, K; Thiesmeier, B. et al. Eds. Handbuch der Reptilien und Amphibien Europas. Band 4/IIb Schwanzlurche (Urodela) III. AULA-Verlag 2004, 391pp.
- Sánchez-Montes, G.; Wang, J.; Ariño, A.H.; Martínez-Solano, Í. Mountains as barriers to gene flow in amphibians: Quantifying the differential effect of a major mountain ridge on the genetic structure of four sympatric species with different life history traits. J. Biogeogr. 2018, 45, 318–331. [CrossRef]
- Cayuela, H.; Valenzuela-Sánchez, A.; Teulier, L.; Martínez-Solano, I.; Léna, J-P.; Merilä, J.; Muths, E.; Shine, R.; Quay, L.; Denoël, M.; Clobert, J.; Schmidt, B.R. Determinants and Consequences of Dispersal in Vertebrates with Complex Life Cycles: A Review of Pond-Breeding Amphibians. Q. Rev. Biol. 2020, 95, 1–36. [CrossRef]
- Taylor, P.D.; Fahrig, L.; Henein, K.; Merriam, G. Connectivity Is a Vital Element of Landscape Structure. Oikos 1993, 68, 571. [CrossRef]
- Vogt, P.; Ferrari, J.R.; Lookingbill, T.R.; Gardner, R.H.;, Riitters, K.H.; Ostapowicz, K. Mapping functional connectivity. Ecol. Indic. 2009, 9, 64–71. [CrossRef]
- Blaustein, A.R.; Walls, S.C.; Bancroft, B.A.; Lawler, J.J.; Searle, C.L.; Gervasi, S.S. Direct and Indirect Effects of Climate Change on Amphibian Populations. Diversity 2010, 2, 281-313. [CrossRef]
- Beebee, T.J.C. Amphibian breeding and climate. Nature 1995, 74, 6519, 219-220. [CrossRef]
- Tryjanowski. P.; Mariusz. R.; Sparks, T. Changes in spawning dates of Common Frogs and Common Toads in western Poland in 1978–2002. Annales Zoologici Fennici 2003, 40, 459-464. http://www.jstor.org/stable/23735858.
- Scott, A.W.; Pithart, D.; Adamson, J.K. Long-term United Kingdom trends in the breeding phenology of the common frog, Rana temporaria. Journal of Herpetology 2008, 42,1, 89-96. [CrossRef]
- Prodon, R.; Geniez, P.; Cheylan. M.; Devers, F.; Chuine, I.; Besnard, A. A reversal of the shift towards earlier spring phenology in several Mediterranean reptiles and amphibians during the 1998–2013 warming slowdown. Glob Change Biol. 2017, 23: 5481–5491. [CrossRef]
- Montori, A.; Amat, F. Surviving on the edge: present and future effects of climate warming on the common frog (Rana temporaria) population in the Montseny massif (NE Iberia) PeerJ 2023, 11, :e14527 . [CrossRef]
- Ficetola, G.F.; Maiorano, L. Contrasting effects of temperature and precipitation change on amphibian phenology, abundance and performance. Oecologia 2016, 181, 3, 683-693. [CrossRef]
- Wellborn, G.A.; Skelly, D.K.; Werner, E.E. Mechanisms creating community structure across a freshwater habitat gradient. Ann. Rev. Ecol. Evol. Syst. 1996, 27, 337-363. [CrossRef]
- Babbitt, K.J.; Baber, M.J.; Tarr, T.L. Patterns of larval amphibian distribution along a wetland hydroperiod gradient. Can. J. Zool. 2003, 81, 1539-1552.
- Van Buskirk, J. Habitat partitioning in European and North American pond-breeding frogs and toads. Div. Distrib. 2003, 9, 399-410.
- Richter-Boix, A.; Llorente, G.A.; Montori, A. A comparative analysis of the adaptive developmental plasticity hypothesis in six Mediterranean anuran species along a pond permanency gradient. Evol. Ecol. Res. 2006b, 8, 1139-1154. Available on: https://www.evolutionary-ecology.com/issues/v08n06/ppar1922.pdf.
- Richter-Boix, A.; Llorente, G.A.; Montori, A. Segregación espacial y temporal de una comunidad de anfibios en una región mediterránea. Munibe (Suplemento / Gehigarria) 2007, 25, 120-128.
- Jakob, C.; Poizat, G.; Veith, M.; Seitz, A.; Crivelli, A. Breeding phenology and larval distribution of amphibians in a Mediterranean pond network with unpredictable hydrology. Hydrobiologia 2003, 499, 51-61.
- Blondel, J.; Aronson, J. Biology and Wildlife of the Mediterranean Region. Oxford University Press, Oxford. ISBN: 9780198500353, 1999, 318pp.
- Directive 92/43/EEC - Conservation of natural habitats and of wild fauna and flora- Habitats Directive (21.05.1992). https://eur-lex.europa.eu/eli/dir/1992/43/oj/eng.
- Resetarits Jr, W. J.; Wilbur, H. M. Choice of oviposition site by Hyla chrysoscelis: role of predators and competitors. Ecology 1989, 70, 1, 220-228. [CrossRef]
- Morand, A.; Joly, P. Habitat variability and space utilization by the amphibian communities of the French Upper-Rhone floodplain. Hydrobiologia 1995, 300/301, 249-257. [CrossRef]
- Morand, A.; Joly, P.; Grolet, O. Phenotypic variation in metamorphosis in five anuran species along a gradient of stream influence. C.R. Acad. Sci. Paris 1997, 320, 645-652. [CrossRef]
- Cayuela, H.; Cheylan, M.; Joly, P. The best of a harsh lot in a specialized species: breeding habitat use by the yellow-bellied toad (Bombina variegata) on rocky riverbanks. Amphibia-Reptilia 2011, 32, 4, 533-539. [CrossRef]
- Pujol-Buxó, E.; Riaño, G.M.; Llorente, G.A. Mild segregation in the breeding preferences of an invasive anuran (Discoglossus pictus) and its main native competitor (Epidalea calamita) in ephemeral ponds. Amphibia-Reptilia 2019, 40, 4, 425-435. [CrossRef]
- Relyea, R.A.; Hoverman, J.T. The impact of larval predators and competitors on the morphology and fitness of juvenile treefrogs. Oecologia 2003, 134, 596–604. [CrossRef]
- Gervasi, S.S.; Foufopoulos, J. Costs of plasticity: responses to desiccation decrease post-metamorphic immune function in a pond-breeding amphibian. Functional Ecology 2008, 22: 100-108. [CrossRef]
- Richter-Boix, A, Tejedo, M. Rezende, E.L. Evolution and plasticity of anuran larval development in response to desiccation. A comparative analysis. Ecol Evol. 2011 1, 1,15-25. [CrossRef]
- Gómez-Rodríguez, C.; Díaz-Paniagua, C.; Bustamante, J.; Portheault, A.; Florencio, M. Inter-annual variability in amphibian assemblages: Implications for diversity assessment and conservation in temporary ponds. Aquat. Conserv.: Mar. Freshw. Ecosyst. 2010, 20, 668-677.
- Gómez-Rodríguez, C.; Díaz-Paniagua, C.; Bustamante, J.; Serrano, L.; Portheault, A. Relative importance of dynamic and static environmental variables as predictors of amphibian diversity patterns. Acta Oecol. 2010, 36, 650-658. [CrossRef]
- McMenamin, S. K.; Hadly, E.A.; Wright, C.K. Climatic Change and Wetland Desiccation Cause Amphibian Decline in Yellowstone National Park. Proceedings of the National Academy of Sciences of the United States of America 2008, 105, 44, 16988–16993. [CrossRef]
- Mac Nally, R.; Horrocks, G.F.B.; Lada, H. Anuran responses to pressures from high-amplitude drought–flood–drought sequences under climate change. Climatic Change 2017, 141, 243–257 . [CrossRef]
- Pollard, C.J.; Stockwell, M.P.; Bower, D.S.; Garnham, J.I.; Pickett, E.J.; Darcovich, K.; O'meara, J.; Clulow, J.; Mahony, M.J. Removal of an exotic fish influences amphibian breeding site selection. Jour. Wild. Mgmt. 2017, 81, 720-727. [CrossRef]
- Colomer, M.A. ; Margalida, A.; Sanuy, I.; Llorente, G.A.; Sanuy, D.; Pujol-Buxó, E. A computational model approach to assess the effect of climate change on the growth and development of tadpoles. Ecological Modelling 2021, 461, 109763. [CrossRef]
- Pujol-Buxó, E.; García-Salmerón, A.; Valera-Florensa, J.; Loras-Ortí, F.; Baena-Crespo, O.; Maluquer-Margalef, J.; Mora-Rueda, C.; Martínez-Silvestre, A. Seguiment i diagnosi de les poblacions dʼamfibis a diferents punts dʼaigua de la Xarxa de Parcs Naturals de la Diputació de Barcelona, 2024, Report. Diputació de Barcelona. 58pp.
- Zylstra, E.R.; Swann, D.E.; Hossack, B.R.; Muths, E.; Steidl, R.J. Drought-mediated extinction of an arid-land amphibian: insights from a spatially explicit dynamic occupancy model. Ecological Applications 2019, 29, 3, e01859. [CrossRef]
- Blaustein, A.R.; Urbina, J.; Snyder, P.W.; Reynolds, E.; Dang, T.; Hoverman, J. T.; Han, B.; Olson, D.H.; Searle, C.; Hambalek, N.M. Effects of Emerging Infectious Diseases on Amphibians: A Review of Experimental Studies. Diversity 2018, 10, 3, 81. [CrossRef]
- Székely, D.; Cogălniceanu, D.; Székely, P. et al. How to recover from a bad start: size at metamorphosis affects growth and survival in a tropical amphibian. BMC Ecol 2020, 20, 24. [CrossRef]
- Anderson, T.L.; Rowland, F.E.; Semlitsch, R.D. Variation in phenology and density differentially affects predator–prey interactions between salamanders. Oecologia 2017, 185, 475-486. [CrossRef]
- Jara, F.G.; Thurman, L.L.; Montiglio, P.O.; Sih, A.; Garcia, T.S. Warming-induced shifts in amphibian phenology and behavior lead to altered predator–prey dynamics. Oecologia 2019, 189, 803-813. [CrossRef]
- Díaz-Paniagua, C. Facteurs associés à la reproduction des amphibiens de Doñana. Détermination de l’habitat. Bulletin de la Société Herpétologique de France 1982, 22: 24-26.
- Lorrain-Soligon, L.; Robin, F.; Bertin, X.; Jankovic, M.; Rousseau, P.; Lelong, V.; Brischoux, F. Long-term trends of salinity in coastal wetlands: Effects of climate, extreme weather events, and sea water level. Environmental Research 2023, 237, 1. [CrossRef]
- Lorrain-Soligon, L.; Boudard, L.; Sebastiano, M.; Costantini, D.; Angelier, F.; Ribout, C.; Leclerc, M.; Kato, A.; Robin, F. Brischoux, F. Salty surprises: Developmental and behavioral responses to environmental salinity reveal higher tolerance of inland rather than coastal Bufo spinosus tadpoles. Environmental Research 2025, 264, 2, 120401. [CrossRef]
- Herbert,E.R.; Boon,P.; Burgin,A.J.; Neubauer, S.C.; Franklin, R.B.; Ardón,M.; Hopfensperger, K.N.; Lamers,L.P.M.; Gell, P. A global perspective on wetland salinization: ecological consequences of agrowing threat to freshwater wetlands. Ecosphere 2015, 6:art206–art243. [CrossRef]
- Relyea, R.; Mattes, B.; Schermerhorn, C.; Shepard, I. Freshwater salinization and the evolved tolerance of amphibians. Ecology and Evolution 2024, 14, e11069. [CrossRef]
- Lorrain-Soligon, L.; Brischoux, F.; Pétillon, J. The interactive effects of salt and heat on coastal ectotherms. Trends in Ecology & Evolution 2024, 39, 12, 1076-1079. [CrossRef]
- Gomez-Mestre, I.; Tejedo, M.; Ramayo, E.; Estepa, J. Developmental alterations and osmoregulatory physiology of a larval anuran under osmotic stress. Physiological and Biochemical Zoology 2004, 77, 267–274. [CrossRef]
- Hart, B.T.; Lake, P.S.; Webb, J.A.; Grace, M.R. Ecological risk to aquatic systems from salinity increases. Aust. J. Bot. 2003, 51, 689–702. [CrossRef]
- Walker, R.H.; Belvin, A.C.; Mouser, J.B.; Pennino, A.; Plont, S.; Robinson, C.D.; Smith, L. B.; Thapa, J.; Zipper, C.E.; Angermeier, P.L.; Entrekin, S.A.; Global review reveals how disparate study motivations, analytical designs, and focal ions limit understanding of salinization effects on freshwater animals. Sci. Total Environ. 2023, 164061 . [CrossRef]
- Burraco, P.; Gomez-Mestre, I. Physiological stress responses in amphibian larvae to multiple stressors reveal marked anthropogenic effects even below lethal levels. Physiol. Biochem. Zool. 2016, 89, 462–472. [CrossRef]
- Tornabene, B.J.; Hossack, B.R.; Crespi, E.J.; Breuner, C.W. Corticosterone mediates a growth-survival tradeoff for an amphibian exposed to increased salinity. J. Exp. Zool. Part A: Ecological and Integrative Physiology 2021b, 335, 703–715. [CrossRef]
- Tornabene, B.J.; Crespi, E.J.; Breuner, C.W.; Hossack, B.R.; Testing whether adrenal steroids mediate phenotypic and physiologic effects of elevated salinity on larval tiger salamanders. Integr. Zool. 2022, .18, 27–44. [CrossRef]
- Gomez-Mestre, I.; Tejedo, M. Local adaptation of an anuran amphibian to osmotically stressful environments. Evolution 2003, 57, 1889–1899. [CrossRef]
- Gomez-Mestre, I.; Tejedo, M. Adaptation or exaptation? An experimental test of hypotheses on the origin of salinity tolerance in Bufo calamita. J Evol Biol. 2005, ,18, 4, 847-55. https://10.1111/j.1420-9101.2004.00878.x.
- Wu, C.-S.; Kam, Y.C. Effects of salinity on the survival, growth, development, and metamorphosis of Fejervarya limnocharis tadpoles living in brackish water. Zoological science 2009, 26, 476–482. [CrossRef]
- Bernabò, I.; Bonacci, A.; Coscarelli, F.; Tripepi, M.; Brunelli, E.; Effects of salinity stress on Bufo balearicus and Bufo bufo tadpoles: tolerance, morphological gill alterations and Na+/K+-ATPase localization. Aquatic toxicology (Amsterdam, Netherlands) 2013, 132–133C, 119–133. [CrossRef]
- Wood, L.; Welch, A.M. Assessment of interactive effects of elevated salinity and three pesticides on life history and behavior of southern toad (Anaxyrus terrestris) tadpoles. Environ. Toxicol. Chem. 2015, 34, 667–676. [CrossRef]
- Lukens, E.; Wilcoxen, T.E.; Effects of elevated salinity on Cuban treefrog Osteopilus septontrionalis aldosterone levels, growth, and development. Mar. Freshw. Behav. Physiol. 2020, 53, 99–111. [CrossRef]
- Tornabene, B.J.; Breuner, C.W.; Hossack, B.R. Comparative effects of energy-related saline wastewaters and sodium chloride on hatching, survival, and fitness-associated traits of two amphibian species. Environ. Toxicol. Chem. 2021, 40, 3137–3147. [CrossRef]
- Hall, E.M.; Brady, S.P.; Mattheus, N.M.; Earley, R.L.; Diamond, M.; Crespi, E.J. Physiological consequences of exposure to salinized roadside ponds on wood frog larvae and adults. Biol. Conserv. 2017, 209, 98–106. [CrossRef]
- Haramura, T, Hatching plasticity in response to salinity levels in a rhacophorid frog inhabiting a coastal area. J. Zool 2016, 299, 125–131. [CrossRef]
- Denoël, M.; Bichot, M.; Ficetola, G.F.; Delcourt, J.; Ylieff, M.; Kestemont, P.; Poncin, P. Cumulative effects of road de-icing salt on amphibian behavior. Aquat. Toxicol. 2010, 99, 275–280. [CrossRef]
- Beebee, T.J.C. Salt tolerance of natterjack toad (Bufo calamita) eggs and larvae from coastal and inland populations in Britain. Journal of Herpetology 1985, 1, 14–16.
- Ortiz-Santaliestra, M.E.; Fernández-Benéitez, M.J.; Lizana, M.; Marco, A. Adaptation to osmotic stress provides protection against ammonium nitrate in Pelophylax perezi embryos. Environmental Pollution 2010, 158, 934–940. [CrossRef]
- Bernabò, I.; Bonacci, A.; Coscarelli, F.; Tripepi, M.; Brunelli, E. Effects of salinity stress on Bufo balearicus and Bufo bufo tadpoles: Tolerance, morphological gill alterations and Na+/K+-ATPase localization. Aquatic Toxicology 2013, 132-133, 119–133. [CrossRef]
- Thirion, J.M. Salinity of the reproductive habitats of the Western Spadefoot Toad Pelobates cultripes (Cuvier, 1829), along the Atlantic coast of France (Anura: Pelobatidae). Herpetozoa 2014, 27, 13–20.
- Galán, P.; Rodríguez-Fernández, S. Efecto de los temporales atlánticos invernales sobre la población de Discoglossus galganoi de los acantilados costeros de Galicia. Boletín de la Asociación Herpetológica Española 2018, 29, 1, 70-75.
- Belmar, O.; Velasco, J.; Martinez-Capel, F. Hydrological Classification of Natural Flow Regimes to Support Environmental Flow Assessments in Intensively Regulated Mediterranean Rivers, Segura River Basin (Spain). Environmental Management 2011, 47, 992–1004. [CrossRef]
- Bonada, N.; Cañedo-Argüelles, M.; Gallart, F.; von Schiller, D.; Fortuño, P.; Latron, J.; ... Cid, N. Conservation and management of isolated pools in temporary rivers. Water 2020, 12, 10, 2870. [CrossRef]
- Magand, C.; Alves, M.H.; Calleja, E.; Datry, T.; Dörflinger, G.; England, J.; Gallart, F.; Gómez, R.; Jorda-Capdevila, D.; Marti, E.; Munne, A.; Pastor, V.A.; Stubbington, R.; Tziortzis, I.; Von Schiller, D. Intermittent rivers and ephemeral streams: what water managers need to know. European Cooperation in Science and Technology. Technical report – Cost ACTION CA 15113. 2020. [CrossRef]
- Datry. T.; Larned, S.T.; Tockner, K. Intermittent Rivers: A Challenge for Freshwater Ecology. Bioscience 2014, 64, 3, 229-235 . [CrossRef]
- Leigh, C.; Boulton, A.J.; Courtwright, J.L.; Fritz, K.; May, C.L.; Walker, R.H.; Datry, T. Ecological research and management of intermittent rivers: An historical review and future directions. Freshwater Biology 2016, 61, 1181–1199. [CrossRef]
- Munné, A.; Bonada, N.; Cid, N.; Gallart, F.; Solà, C.; Bardina, M.; Rovira, A.; Sierra, C.; Soria, M.; Fortuño, P.; et al. A Proposal to Classify and Assess Ecological Status in Mediterranean Temporary Rivers: Research Insights to Solve Management Needs. Water 2021, 13, 767. [CrossRef]
- Soria, M.; Leigh, C.; Datry, T.; Bini, L.M.; Bonada, N. Biodiversity in perennial and intermittent rivers: a meta-analysis. Oikos, 2017,126, 1078-1089. [CrossRef]
- Acuña, V.; Hunter, M.; Ruhí, A. Managing temporary streams and rivers as unique rather than second-class ecosystems. Biological Conservation 2017, 211, 12-19. [CrossRef]
- Bernardo, J.M.; Ilhéu, M.; Matono, P.; Costa, A.M. Interannual variation of fish assemblage structure in a Mediterranean river: implications of streamflow on the dominance of native or exotic species. River Res. Applic. 2003, 19, 521-532. [CrossRef]
- Muñoz, I.; García-Berthou, E.; Sabater, S. The Effect of Multiple Stressors on Biological Communities in the Llobregat. In: Sabater, S.; Ginebreda, A.; Barceló, D. (eds) The Llobregat. The Handbook of Environmental Chemistry, 2012, 21, Springer, Berlin, Heidelberg. [CrossRef]
- Matono, P.; Da Silva, J.; Ilhéu, M. How Does an Invasive Cyprinid Benefit from the Hydrological Disturbance of Mediterranean Temporary Streams? Diversity 2018, 10, 47. [CrossRef]
- Guareschi, S.; South, J. Biological invasions in intermittent rivers and streams: current knowledge, and future frontiers. Ecosistemas 2024, 33, 1, 2600. [CrossRef]
- King, A.J.; Townsend, S.A.; Douglas, M.M; Kennard, M.J. Implications of water extraction on the low-flow hydrology and ecology of tropical savannah rivers: an appraisal for northern Australia. Freshwater Science 2015, 34, 2, 741-758. [CrossRef]
- Stefanidis, K.; Panagopoulos, Y.; Psomas, A.; Mimikou, M. Assessment of the natural flow regime in a Mediterranean river impacted from irrigated agriculture. Science of the Total Environment 2016, 573, 1492–1502. [CrossRef]
- Skoulikidis, N.T.; Vardakas, L.; Karaouzas, I; et al. Assessing water stress in Mediterranean lotic systems: insights from an artificially intermittent river in Greece. Aquat Sci 2011, 73, 581–597 . [CrossRef]
- Sabo, J.L. Predicting the river’s blue line for fish conservation, Proc. Natl. Acad. Sci. U.S.A. 2014, 111, 38, 13686-13687. [CrossRef]
- Ruhí, A.; Olden, J.D.; Sabo, J.L. Declining streamflow induces collapse and replacement of native fish in the American Southwest. Front Ecol Environ 2016, 14, 9, 465–472. [CrossRef]
- Jaeger, K.L.; Olden, J.D.; Pelland, N.A. Climate change poised to threaten hydrologic connectivity and endemic fishes in dryland streams, Proc. Natl. Acad. Sci. U.S.A. 2014, 111, 38, 13894-13899. [CrossRef]
- Lytle, D.A.; N.LeR. Poff Adaptation to natural flow regimes. Trends in Ecology & Evolution 2004, 19, 2, 94 – 100. [CrossRef]
- Chessman, B.C. Relationships between lotic macroinvertebrate traits and responses to extreme drought. Freshw. Biol. 2015, 60, 50-63. [CrossRef]
- Serra-Cobo, J.; Marques, T.; Martínez-Rica, J.P.; Ecological segregation between Rana pyrenaica and Rana temporaria, and differential predation of Euproctus asper on their tadpoles. Netherland J. Zoology 2000, 50, 1, 65-73. [CrossRef]
- Colomer, M.À.; Montori, A.; García, E.; Fondevilla, C. Using a bioinspired model to determine the extinction risk of Calotriton asper populations as a result of an increase in extreme rainfall in a scenario of climatic change. Ecological Modelling 2014, 281, 1–14. [CrossRef]
- Guinart, D.; Solórzano, S.; Amat, F.; Grau, J.; Fernández-Guiberteau, D.; Montori, A. Habitat Management of the Endemic and Critical Endangered Montseny Brook Newt (Calotriton arnoldi). Land 2022, 11, 449. [CrossRef]
- Ball, S.E.; Bovero, S.; Sotgiu, G.; Tessa, G.; Angelini, C.; Bielby, J.; Durrant, C.; Favelli, M.; Gazzaniga, E.; Garner, T.W.J. Islands within an island: Population genetic structure of the endemic Sardinian newt, Euproctus platycephalus. Ecol Evol. 2017; 7: 1190–1211. [CrossRef]
- Montori, A.; Llorente, G.A.; Richter-Boix, À. Habitat features affecting the small-scale distribution and longitudinal migration patterns of Calotriton asper in a Pre-Pyrenean population. Amphibia-Reptilia 2008, 29, 3, 371-381. [CrossRef]
- Ribera, I.; Vogler, A. Habitat type as a determinant of species range sizes: the example of lotic-lentic differences in aquatic Coleoptera. Biological Journal of the Linnean Society 2000, 71, 1, 33-52. [CrossRef]
- Ribera, I.; Foster, G.N.; Vogler, A.P. Does habitat use explain large scale species richness patterns of aquatic beetles in Europe? – Ecography 2003, 26: 145–152. https://www.jstor.org/stable/3683429.
- Abellán, P.; Ribera, I. Geographic location and phylogeny are the main determinants of the size of the geographical range in aquatic beetles. BMC Evolutionary Biology 2011, 11:344. http://www.biomedcentral.com/1471-2148/11/344.
- Talavera, A.; Palmada-Flores, M.; Burriel-Carranza, B.; Valbuena-Ureña, E.; Mochales-Riaño, G. et al. Genomic insights into the montseny brook newt (Calotriton arnoldi), a critically endangered glacial relict. iScience 2024, 27,108665. [CrossRef]
- Puig-Gironès, R.; Bel, G.; Cid, N.; Cañedo-Argüelles, M.; Fernández-Calero, J.M.; Quevedo-Ortiz G.; Fortuño, P; Vinyoles, D.; Real, J.; Pujol-Buxó, E.; Bonada, N. Water availability and biological interactions shape amphibian abundance and diversity in Mediterranean temporary rivers. Science of The Total Environment 2024 953, 175917. [CrossRef]
- Kerezsy, A.; Gido, K.; Magalhães, M.F.; Skelton, P.H. The Biota of Intermittent Rivers and Ephemeral Streams: Fishes. Intermittent Rivers and Ephemeral Streams, 2017, 273–298. [CrossRef]
- Huxter, E.H.H.; Van Meerveld, H.J. Intermittent and perennial streamflow regime characteristics in the Okanagan. Canadian Water Resources Journal 2012, 37, 4, 391-414. [CrossRef]
- Fraser, N.; Schumer, R. Intermittency in dust deposition rates around the world. AGU Fall Meeting 2011, San Francisco, CA. https://ui.adsabs.harvard.edu/abs/2011AGUFMPP23B1850F.
- Camarasa-Belmonte AM. Flash-flooding of Ephemeral Streams in the Context of Climate Change. CIG. 2021, 47, 121-142. [CrossRef]
- Jiang, J.; Wang, Z.; Zhang, Z.; Wu, X.; Lai, C.; Zeng, Z.; Chen, X. Extreme drought-heatwave exacerbates water quality deterioration in China. Ecological Indicators 2025, 170, 113008. [CrossRef]
- Corominas, L.; Foley, J.; Guest, J.S.; Hospido, A.; Larsen, H.F.; Morera, S.; Shaw, A. Life cycle assessment applied to wastewater treatment: state of the art. Water Res. 2013, 47, 15, 5480-92. [CrossRef]
- Wagner, N.; Reichenbecher, W.; Teichmann, H.; Tappeser, B.; Lötters, S. Questions concerning the potential impact of glyphosate-based herbicides on amphibians, Environmental Toxicology and Chemistry, 2013, 32, 8, 1688–1700. [CrossRef]
- Chow, R.; Curchod, L.; Davies, E. et al. Seasonal drivers and risks of aquatic pesticide pollution in drought and post-drought conditions in three Mediterranean watersheds. Science of The Total Environment 2023, 858, 2, 159784. [CrossRef]
- Curchod, L.; Oltramare, C.; Junghan, M.; et al. Temporal variation of pesticide mixtures in rivers of three agricultural watersheds during a major drought in the Western Cape, South Africa. Water Research X 2020, 6, 100039. [CrossRef]
- Church, D.R.; Bailey, L.L.; Wilbur, H.M.; Kendall, W.L.; Hines. J.E. Iteroparity in the variable environment of the salamander Ambystoma tigrinum. Ecology 2007, 88, 891–903. [CrossRef]
- Carnicer, J.; Coll, M.; Ninyerola, M.; Pons, X.; Sanchez, X.; Peñuelas, J. Widespread crown condition decline, food web disruption, and amplified tree mortality with increased climate change-type drought. Proceedings of the National Academy of Sciences, 2011, 108, 4, 1474-1478. [CrossRef]
- Carnicer, J.; Vives-Ingla, M.; Blanquer, L.; Méndez-Camps, X.; Rosell, C.; Sabaté, S;, Gutiérrez, E.; Sauras, T.; Peñuelas, J.; Barbeta, A. Forest resilience to global warming is strongly modulated by local-scale topographic, microclimatic and biotic conditions. Journal of Ecology 2021, 109, 9, 3322-3339. [CrossRef]
- Reading, C.J. Linking global warming to amphibian declines through its effects on female body condition and survivorship. Oecologia 2007, 151, 1, 125-131. [CrossRef]
- Daszak, P.; Scott, D.E.; Kilpatrick, A.M.; Faggioni, C..; Gibbons, J.W.; Porter, D. Amphibian population declines at savannah river site are linked to climate, not chytridiomycosis. Ecology 2005, 86, 3232-3237. [CrossRef]
- Hossack, B.R.; Lowe, W.H.; Corn, P.S.; Rapid increases and time-lagged declines in amphibian occupancy after wildfire. Conservation Biology 2013, 27, 219–228. [CrossRef]
- Lombardi, F.; Lasserre, B.; Chirici, G.; Tognetti, R.; Marchetti, M. Deadwood occurrence and forest structure as indicators of old-growth forest conditions in Mediterranean mountainous ecosystems. Ecoscience 2012, 19, 4, 344-355. [CrossRef]
- Badalamenti, E.; La Mantia, T.; La Mantia, G.; Cairone, A.; La Mela Veca, D.S. Living and dead aboveground biomass in Mediterranean forests: Evidence of old-growth traits in a Quercus pubescens Willd. sl stand. Forests 2017, 8, 6, 187. [CrossRef]
- Parisi, F.; Pioli, S.; Lombardi, F.; Fravolini, G.; Marchetti, M.; Tognetti, R. Linking deadwood traits with saproxylic invertebrates and fungi in European forests-a review. iForest-Biogeosciences and Forestry 2018, 11, 3, 423. [CrossRef]
- Batllori, E.; De Cáceres, M.; Brotons, L.; Ackerly, D.D.; Moritz, M.A.; Lloret, F. Cumulative effects of fire and drought in Mediterranean ecosystems. Ecosphere 2017, 8, 8, e01906. [CrossRef]
- Karavani, A.; Boer, M.M.; Baudena, M.; Colinas, C.; Díaz-Sierra, R.; Pemán, J.;… Resco de Dios, V. Fire-induced deforestation in drought-prone Mediterranean forests: drivers and unknowns from leaves to communities. Ecological Monographs 2018, 88, 2, 141-169. [CrossRef]
- Ogaya, R.; Peñuelas, J. Climate change effects in a Mediterranean forest following 21 consecutive years of experimental drought. Forests 2021, 12, 3, 306. [CrossRef]
- Naiman, R.J.; Bechtold, J.S.; Drake, D.C.; Latterell, J.J.; O'Keefe, T.C.; Balian, E.V. Origins, Patterns, and Importance of Heterogeneity in Riparian Systems. In: Lovett, G.M.; Turner, M.G.; Jones, C.G.; Weathers, K.C. (eds) Ecosystem Function in Heterogeneous Landscapes. 2005, Springer, New York, NY. [CrossRef]
- Naiman, R.J.; Decamps, H. The ecology of interfaces: riparian zones. Annu. Rev. Ecol. Syst. 1997, 28, 1, 621–658. [CrossRef]
- Burbrink, F.T.; Phillips, C.A.; Heske, E.J. A riparian zone in Southern Illinois as a potential dispersal corridor for reptiles and amphibians. Biol. Conserv. 1998, 86, 2, 107–115. [CrossRef]
- Aguilar, F.F.; Velo-Antón, G.; Tarroso, P. et al. Fine-scale habitat preferences of riparian ectotherms in a human-influenced landscape: insights from two herptiles endemic to the Iberian Peninsula. Biodivers Conserv 2025. [CrossRef]
- Pausas, J.G.; Fernández-Muñoz, S. Fire regime changes in the Western Mediterranean Basin: from fuel-limited to drought-driven fire regime. Climatic Change 2012, 110, 215–226. [CrossRef]
- Wasserman, T.N.; Mueller, S.E. Climate influences on future fire severity: a synthesis of climate-fire interactions and impacts on fire regimes, high-severity fire, and forests in the western United States. Fire Ecol. 2023, 19, 43. [CrossRef]
- Pausas, J.G. Changes in Fire and Climate in the Eastern Iberian Peninsula (Mediterranean Basin). Climatic Change 2004, 63, 337–350. [CrossRef]
- Dunham, J.B.; Rosenberger, A.E.; Luce, C.H.; Rieman. B.E.; Influences of wildfire and channel reorganization on spatial and temporal variation in stream temperature and the distribution of fish and amphibians. Ecosystems 2007, 10, 335–346. [CrossRef]
- dos Anjos, A.G.; Solé, M.; Benchimol, M. Fire effects on anurans: what we know so far? Forest Ecology and Management 2021, 495, 119338. [CrossRef]
- Montori, A.; Llorente, G.A.; Clivillé, S.; Santos, X.; Carretero, M.A. Efectes de l’incendi forestal de 1994 sobre les poblacions d’amfibis del Parc Natural del Garraf. Monografies Diputació de Barcelona, 2000, 30. 105-108. https://parcs.diba.cat/documents/182160/f3fbe2c5-c98f-4498-b808-abd350256b69.
- Montori, A. Estat de les poblacions d’amfibis del Parc del Garraf 16 anys després de l’incendi forestal de 1994 i efectes dels focs repetitius sobre les comunitats d’amfibis. Unpublished Report. ACOM 2010. Dept. d’innovació, universitats i empreses. Generalitat de Catalunya. 2010, 55pp.
- Chergui, B.; Ayres, C.; Santos, X. Assessing the response of amphibians to wildfire according to forest type and bioregion affinity of species. Basic and Applied Herpetology 2022, 36, 5-17. [CrossRef]
- Carretero, J.M.; Guzmán, E.; Martínez, M. i Úbeda, X. Els incendis forestals a l’àrea del Garraf-Castelldefels. II premi “Castelldefels ambit sostenible” Castelldefels, 2003, 148pp.
- Moretti, M.; Duelli, P.; Obrist, M.K. Biodiversity and resilience of arthropod communities after fire disturbance in temperate forests. Oecologia 2006, 149, 312-327. [CrossRef]
- Santos, X.; Bros, V.; Miño, A. Recolonization of a burned Mediterranean area by terrestrial gastropods. Biodiversity and Conservation 2009, 18, 3153–3165. [CrossRef]
- Moretti, M.; Legg, C. Combining plant and animal traits to assess community functional responses to disturbance. Ecography 2008, 31, 1-11. [CrossRef]
- Burrow, A.; Maerz, J. How plants affect amphibian populations. Biological Reviews 2022, 97, 5, 1749-1767. [CrossRef]
- Haggerty, C.J.; Crisman, T.L; Rohr, J.R. Effects of forestry-driven changes to groundcover and soil moisture on amphibian desiccation, dispersal, and survival. Ecological Applications 2019, 29, 3, e01870. [CrossRef]
- Haggerty, C.J.; Crisman, T.L; Rohr, J.R. Direct and indirect effects of pine silviculture on the larval occupancy and breeding of declining amphibian species. Journal of Applied Ecology 2019, 56, 12, 2652-2662. [CrossRef]
- Muñoz, A.; Felicisimo, A.M.; Santos, X. Assessing the resistance of a breeding amphibian community to a large wildfire. Acta Oecologica 2019, 99, 103439. [CrossRef]
- Martínez-Silvestre, A.; Garcia-Salmeron, A.: Pujol-Buxó, E.: Baena, O. Anàlisi de patògens emergents dels amfibis dins de 7 parcs de la Xarxa de Parcs Naturals de la Diputació de Barcelona. Butll. Soc. Catalana d’Herpetologia. 2022, 29, 44-55.
- Longcore, J. E.; Pessier, A. P.; Nichols, D. K. Batrachochytrium dendrobatidis gen. et sp. nov., a chytrid pathogenic to amphibians. Mycologia 1999, 91, 2, 219-227. [CrossRef]
- Martel, A.; Spitzen-van der Sluijs, A.; Blooi, M.; Bert, W.; Ducatelle, R.; Fisher, M. C.; ... Pasmans, F. Batrachochytrium salamandrivorans sp. nov. causes lethal chytridiomycosis in amphibians. Proceedings of the National Academy of Sciences 2013, 110, 38, 15325-15329. [CrossRef]
- Granoff, A.; Came, P.E.; Breeze, D.C. Viruses and renal carcinoma of Rana pipiens: I. The isolation and properties of virus from normal and tumor tissues. Virology 1966, 29, 133–148 . [CrossRef]
- Bosch, J.; Martı́nez-Solano, I.; Garcı́a-Parı́s, M. Evidence of a chytrid fungus infection involved in the decline of the common midwife toad (Alytes obstetricans) in protected areas of central Spain. Biological conservation, 2001 97(3), 331-337.
- Walker, S. F.; Bosch, J.; James, T. Y.; Litvintseva, A. P.; Valls, J. A. O.; Piña, S.; ... Fisher, M. C. Invasive pathogens threaten species recovery programs. Current Biology 2008, 18, 18, R853-R854. [CrossRef]
- Martel, A.; Vila-Escale, M.; Fernández-Giberteau, D.; Martinez-Silvestre, A.; Canessa, S.; Van Praet, S.; ... Pasmans, F. Integral chain management of wildlife diseases. Conservation Letters 2020,13 ,2, e12707. [CrossRef]
- Price, S.J.; Garner, T.W.J.; Nichols, R.A.; Balloux, F.; Ayres, C.; Mora-Cabello de Alba, A.; Bosch, J. Collapse of amphibian communities due to an introduced Ranavirus. Current Biology 2014, 24, 21, 2586–2591. [CrossRef]
- Scheele, B.C.; Hunter, D.A.; Banks, S.C.; Pierson, J.C.; Skerratt, L.F.; Webb, R. and Driscoll, D.A. High adult mortality in disease-challenged frog populations increases vulnerability to drought. J Anim Ecol, 2016, 85, 1453-1460. [CrossRef]
- Blaustein, A.R.; Urbina, J.; Snyder, P.W.; Reynolds, E.; Dang, T.; Hoverman, J.T.; Han, B.; Olson, D.H.; Searle, C.; Hambalek, N.M. Effects of Emerging Infectious Diseases on Amphibians: A Review of Experimental Studies. Diversity 2018, 10, 81. [CrossRef]
- Johnson, M.L.; Berger, L.; Philips, L.; Speare, R. Fungicidal effects Batrachochytrium dendrobatidis, UV light, desiccation and heat on the amphibian chytrid Batrachochytrium dendrobatidis. Dis. Aquat. Organ. 2003, 57: 255–260. [CrossRef]
- Piotrowski, J.S.; Annis, S.L.; Longcore, J.E. Physiology of Batrachochytrium dendrobatidis, a chytrid pathogen of amphibians. Mycologia 2004, 96, 9–15. [CrossRef]
- Kilpatrick, A.M.; Briggs, C.J.; Daszak, P. The ecology and impact of chytridiomycosis: an emerging disease of amphibians. Trends in Ecology and Evolution. 2010, 25, 2, 109-118. [CrossRef]
- Van Rooij, P.; Martel, A.; Haesebrouck, F.; Pasmans, F. Amphibian chytridiomycosis: a review with focus on fungus-host interactions. Veterinary Research 2015, 46, 137. [CrossRef]
- .
- Garmyn, A.; Van Rooij, P.; Pasmans, F.; Hellebuyck, T.; Van Den Broeck, W.; Haesebrouck, F.; Martel, A. Waterfowl: potential environmental reservoirs of the chytrid fungus Batrachochytrium dendrobatidis. PLoS One 2012, 7, e35038. [CrossRef]
- Farrer, R.A.; Weinert, L.A.; Bielby, J.; Garner, T.W.; Balloux, F.; Clare, F.; ... Fisher, M.C. Multiple emergences of genetically diverse amphibian-infecting chytrids include a globalized hypervirulent recombinant lineage. Proceedings of the National Academy of Sciences 2011, 108, 46, 18732-18736. [CrossRef]
- Walker, S.F.; Bosch, J.; Gomez, V.; Garner, T.W.; Cunningham, A.A.; Schmeller, D.S.; ... Fisher, M. C. Factors driving pathogenicity vs. prevalence of amphibian panzootic chytridiomycosis in Iberia. Ecology letters 2010, 13, 3, 372-382. [CrossRef]
- Ruggeri, J.; Martins, A.G.D.S.; Domingos, A.H.R.; Santos, I.; Viroomal, I.B.; Toledo, L.F. Seasonal prevalence of the amphibian chytrid in a tropical pond-dwelling tadpole species. Diseases of Aquatic Organisms 2020, 142, 171-176. [CrossRef]
- Kolby, J.E.; Ramirez, S. D.; Berger, L.; Richards-Hrdlicka, K.L.; Jocque, M.; Skerratt, L.F. Terrestrial dispersal and potential environmental transmission of the amphibian chytrid fungus (Batrachochytrium dendrobatidis). PloS one 2015, 10, 4, e0125386. [CrossRef]
- Prado, J. S.; Ernetti, J.R.; Pontes, M.R.; Toledo, L.F. Chytrid in the clouds: an alternative passive transport of a lethal pathogen for amphibians. Hydrobiologia 2023, 850, 9, 2061-2073. [CrossRef]
- Padgett-Flohr, G.E.; Hopkins, R.L. Landscape epidemiology of Batrachochytrium dendrobatidis in central California. Ecography 2010, 33, 4, 688-697. [CrossRef]
- Strauss, A.; Smith, K. G. Why does amphibian chytrid (Batrachochytrium dendrobatidis) not occur everywhere? An exploratory study in Missouri ponds. PloS one 2013, 8, 9, e76035. [CrossRef]
- Martínez-Silvestre, A.; Loras-Ortí, F.; Garcia-Salmeron, A.; Pujol-Buxó, E.; Pérez-Novo, I.; Maluquer-Margalef, J.; Poch, S.; Thumsová, B.; Bosch, J. Introduced Mediterranean painted frogs (Discoglossus pictus) are possible supershedders of the fungus Batrachochytrium dendrobatidis in Catalonia (NE Spain). Amphibia-Reptilia 2023, 44, 2, 257-261. [CrossRef]
- Montori, A.; San Sebastián, O.; Franch, M.; Pujol-Buxó, E.; Llorente, G.A.; Fernández-Loras, A.; Richter-Boix, A.; Bosch, J. Observations on the intensity and prevalence of Batrachochytridium dendrobatidis in sympatric and allopatric Epidalea calamita (native) and Discoglossus pictus (invasive) populations. Basic and Applied Herpetology 2019, 33, 5-17. [CrossRef]
- Ruggeri, J.; de Carvalho-e-Silva, S. P.; James, T. Y.; Toledo, L. F. Amphibian chytrid infection is influenced by rainfall seasonality and water availability. Diseases of Aquatic Organisms 2018, 127, 2, 107-115. [CrossRef]
- Kupferberg, S.J.; Moidu, H.; Adams, A.J.; Catenazzi, A.; Grefsrud, M.; Bobzien, S.; Leidy, R.; Carlson, S.M. Seasonal drought and its effects on frog population dynamics and amphibian disease in intermittent streams. Ecohydrology 2022, 15, 5, e2395. [CrossRef]
- Fernández, S. Factores bióticos y abióticos responsables de la distribución e incidencia de Batrachochytrium dendrobatidis en poblaciones de anfibios de zonas templadas. Doctoral These. Facultad de Ciencias Biológicas. Universidad Complutense de Madrid. 2018, 231pp. https://hdl.handle.net/20.500.14352/16207.
- Bosch, J.; Thumsová, B.; Puschendorf, R.; Bielby, J. Drivers of Batrachochytrium dendrobatidis infection load, with evidence of infection tolerance in adult male toads (Bufo spinosus). Oecologia 2023, 202, 1, 165-174. [CrossRef]
- Buttimer, S.; Moura-Campos, D.; Greenspan, S.E.; Neely, W.J.; Ferrante,L.; Toledo, L.F. et al. Skin microbiome disturbance linked to drought-associated amphibian disease. Ecology Letters 2024, 27, e14372. [CrossRef]
- McDevitt-Galles, T.; Moss, W.E.; Calhoun, D.M.; Briggs, C.J.; Pieter, T. J. How extreme drought events, introduced species, and disease interact to influence threatened amphibian populations. Johnson Freshwater Science 2022, 41, 4, 680-694. [CrossRef]
- More, S.;, Miranda, A.; Bicout, M.; Bøtner, D. et al. Scientific Opinion on the risk of survival, establishment and spread of Batrachochytrium salamandrivorans (Bsal) in the EU. EFSA Journal 2018, 16, 4, 5259, 78 pp. [CrossRef]
- Malagon, D.A.; Melara, L.A.; Prosper, O.F. et al. Host density and habitat structure influence host contact rates and Batrachochytrium salamandrivorans transmission. Sci Rep 2020, 10, 5584. [CrossRef]
- Deiß, F.; Ginal, P.; Rödder, D. Microclimatic Growth Rates of Batrachochytrium salamandrivorans under Current and Future Climates: A Very High Spatial Resolution SDM for Bsal and Salamandra salamandra (Linnaeus, 1758) within Forest Habitats of the European Hotspot Area. Diversity 2024, 16, 510. [CrossRef]
- Stegen, G.; Pasmans, F.; Schmidt, B. R.; Rouffaer, L. O.; Van Praet, S.; Schaub, M.; ... Martel, A. Drivers of salamander extirpation mediated by Batrachochytrium salamandrivorans. Nature 2017, 544, 7650, 353-356. [CrossRef]
- Spitzen - van der Sluijs, A.; Stegen, G.; Bogaerts, S. et al. Post-epizootic salamander persistence in a disease-free refugium suggests poor dispersal ability of Batrachochytrium salamandrivorans. Sci Rep 2018, 8, 3800. [CrossRef]
- Kelly, M.; Cuomo, C.A.; Beukema, W.; Carranza, S.; Erens, J.; Foubert, M. et al. High phenotypic diversity correlated with genomic variation across the European Batrachochytrium salamandrivorans epizootic. PLoS Pathog. 2024, 20, 10, e1012579. [CrossRef]
- Jancovich, J.K.; Davidson, E.W.; Parameswaran, N.; Mao, J.; Chinchar, V.G.; Collins, J.P.; Jacobs, B.L.; Storfer, A. Evidence for emergence of an amphibian iridoviral disease because of human-enhanced spread. Mol. Ecol. 2005, 14, 213–224. [CrossRef]
- Jancovich, J.K.; Bremont, M.; Touchman, J.W.; Jacobs, B.L. Evidence for multiple recent host species shifts among the ranaviruses (family Iridoviridae). J. Virol. 2010, 84, 2636–2647. [CrossRef]
- Bandín, I.; Dopazo, C. Host range, host specificity and hypothesized host shift events among viruses of lower vertebrates. Vet. Res. 2011, 42, 67. [CrossRef]
- Mao, J.; Green, D.E.; Fellers, G. Chinchar, V.G. Molecular characterization of iridoviruses isolated from sympatric amphibians and fish. Virus Res. 1999, 63, 45–52. [CrossRef]
- Bayley, A.E.; Hill, B.J.; Feist, S.W. Susceptibility of the European common frog Rana temporaria to a panel of ranavirus isolates from fish and amphibian hosts. Dis. Aquat. Organ. 2013 103, 171–183. [CrossRef]
- Thumsová, B.; Donaire-Barroso, D.; Mouden El, E.H.; Bosch, J. Fatal chytridiomycosis in the Moroccan midwife toad Alytes maurus and potential distribution of Batrachochytrium dendrobatidis across Morocco. African Journal of Herpetology 2022, 71, 1–11.
- Altizer, S.; Ostfeld, R.S.; Johnson PT, Kutz, S.; Harvell C.D. Climate change and infectious diseases: from evidence to a predictive framework. Science 2013, 341(6145):514-9. [CrossRef]
- Price, S.J.; Leung, W.T.M.; Owen, C.J.; et al. Effects of historic and projected climate change on the range and impacts of an emerging wildlife disease. Glob Change Biol. 2019, 25, 2648–2660. [CrossRef]
- Allender, M. C.; Mitchell, M. A.; Torres, T.; Sekowska, J.; Driskell, E. A. Pathogenicity of Frog Virus 3-like Virus in Red-eared Slider Turtles (Trachemys scripta elegans) at Two Environmental Temperatures. Journal of Comparative Pathology 2013, 149, 2-3, 356–367. [CrossRef]
- Rojas, S.; Richards, K.; Jancovich, J.K.; Davidson, E.W. Influence of temperature on ranavirus infection in larval salamanders, Ambystoma tigrinum. Dis. Aquat. Org. 2005, 63, 95–100. [CrossRef]
- Brunner, J.L.; Storfer, A.; Le Sage, E.H.; Garner, T.W.J.; Gray, M.J.; Hoverman, J.T. Ranavirus Ecology: From Individual Infections to Population Epidemiology to Community Impacts. In: Gray, M.J.; Chinchar, V.G. (eds) Ranaviruses. Springer, Cham. 2025. [CrossRef]
- Cohen, J.M.; Sauer, E.L.; Santiago, O.; Spencer, S.; Rohr, J.R. Divergent impacts of warming weather on wildlife disease risk across climates. Science. 2020, 370, 6519, eabb1702. [CrossRef]
- Daszak, P,; Cunningham, A.A.; Hyatt, A.D. Anthropogenic environmental change and the emergence of infectious diseases in wildlife. Acta Trop. 2001, 78, 2, 103-16. [CrossRef]
- Davis, D.R.; Ferguson, K.J.; Schwarz, M.S.; Kerby, J.L. Effects of agricultural pollutants on stress hormones and viral infection in larval salamanders. Wetlands. 2020, 40, 577-86. [CrossRef]
- Rosa, G.; Sabino-Pinto, J.; Laurentino, T. et al. Impact of asynchronous emergence of two lethal pathogens on amphibian assemblages. Sci. Rep. 2017, 7, 43260. [CrossRef]
- Humphries, J.E.; Lanctôt, C.M.; Robert, J.; McCallum, H.I.; Newell, D.A.; Grogan, L.F. Do immune system changes at metamorphosis predict vulnerability to chytridiomycosis? An update. Dev Comp Immunol. 2022; 136, 104510. [CrossRef]
- Harrison, X.A.; Price, S.J.; Hopkins, K.; Leung, W.T.M.; Sergeant, C.; Garner T.W.J. Diversity-stability dynamics of the amphibian skin microbiome and susceptibility to a lethal viral pathogen. Frontiers in Microbiology, 2019, 10, 2883. [CrossRef]
- Campbell, L.J.; Pawlik, A.H.; Harrison. X.A. Amphibian ranaviruses in Europe: important directions for future research. Facets 2020, 5, 1, 598-614. [CrossRef]
- Haislip, N.A.; Gray, M.J.; Hoverman, J.T.; Miller, D.L. Development and disease: how susceptibility to an emerging pathogen changes through anuran development. PLoS One. 2011, 6, 7, e22307. [CrossRef]
- Kwon, S.; Park, J.; Choi, W.J; Koo, K.S.; Lee, J.G.; Park, D. First case of ranavirus-associated mass mortality in a natural population of the Huanren frog (Rana huanrenensis) tadpoles in South Korea. Anim. Cells Syst. 2017, 21, 5, 358-64. [CrossRef]
- Teacher, A.G.F.; Cunningham, A.A.; Garner, T.W.J. Assessing the long-term impact of Ranavirus infection in wild common frog populations. Animal Conservation 2010, 13, 5, 514–522. [CrossRef]
Figure 1.
Proportion of amphibian species exposed to drought around the world according to [47].
Figure 1.
Proportion of amphibian species exposed to drought around the world according to [47].
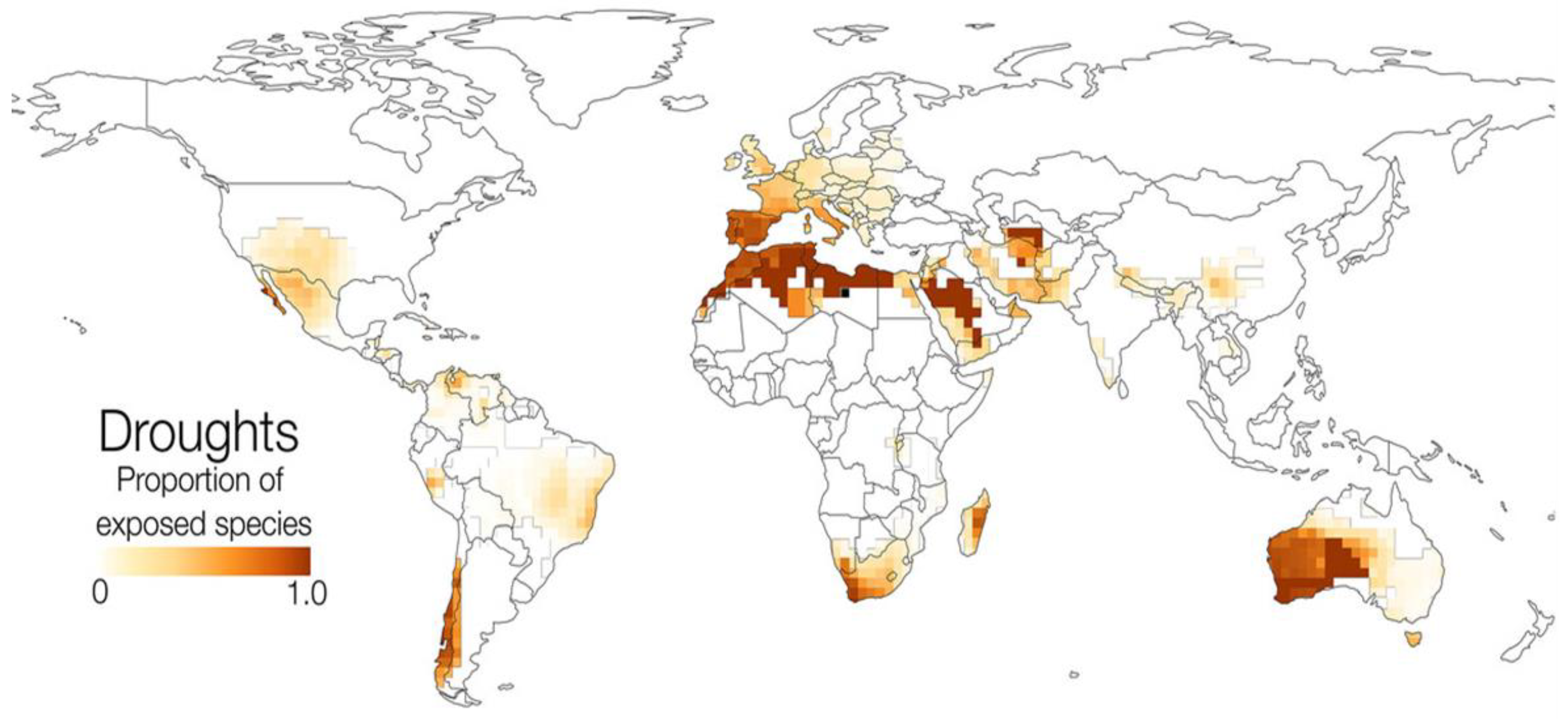
Figure 2.
Predicted change in water resource category (in %) to 2050-2100, from low and high scenarios of greenhouse emissions. AR: Aquifer recharge, R/LI: Reservoir/Lake Inflow, R: Runoff, SM: Soil Moisture, OR: Other Resources and ID: Irrigation Demand. The boxes indicate a confidence interval of 95%. The black horizontal line is the mean. Data include only publications referred to Western Mediterranean Basin gathered by [26], from Supplementary Materials, Tables S1 and S2.
Figure 2.
Predicted change in water resource category (in %) to 2050-2100, from low and high scenarios of greenhouse emissions. AR: Aquifer recharge, R/LI: Reservoir/Lake Inflow, R: Runoff, SM: Soil Moisture, OR: Other Resources and ID: Irrigation Demand. The boxes indicate a confidence interval of 95%. The black horizontal line is the mean. Data include only publications referred to Western Mediterranean Basin gathered by [26], from Supplementary Materials, Tables S1 and S2.
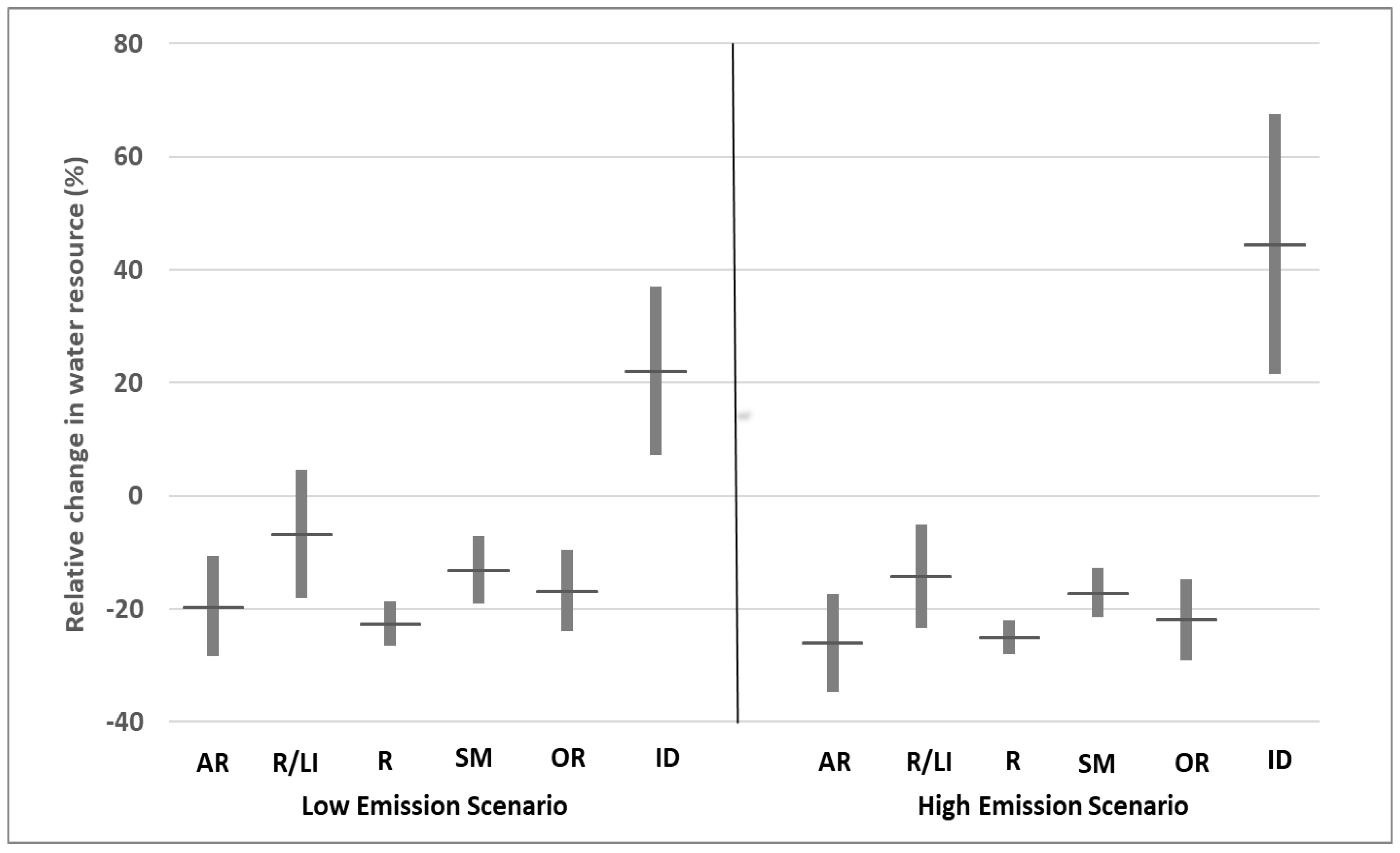
Figure 3.
Schematic diagram of the main pathways via which drought may change tadpole and toadlet fitness. A: Drought reduces hydroperiod forcing tadpoles to modify growth or larval development by means of phenotypic plasticity. B: Drought increases predation and intra- and interspecific competition. C: Increase of water temperatures can favor the outbreak of emergent diseases. D: Drought directly causes amphibian mortality when ponds or stream pools dry out, leading to the loss of entire cohorts. E: The decrease of water volume increases the pollutant concentration. E also has effects on B, C and D.
Figure 3.
Schematic diagram of the main pathways via which drought may change tadpole and toadlet fitness. A: Drought reduces hydroperiod forcing tadpoles to modify growth or larval development by means of phenotypic plasticity. B: Drought increases predation and intra- and interspecific competition. C: Increase of water temperatures can favor the outbreak of emergent diseases. D: Drought directly causes amphibian mortality when ponds or stream pools dry out, leading to the loss of entire cohorts. E: The decrease of water volume increases the pollutant concentration. E also has effects on B, C and D.
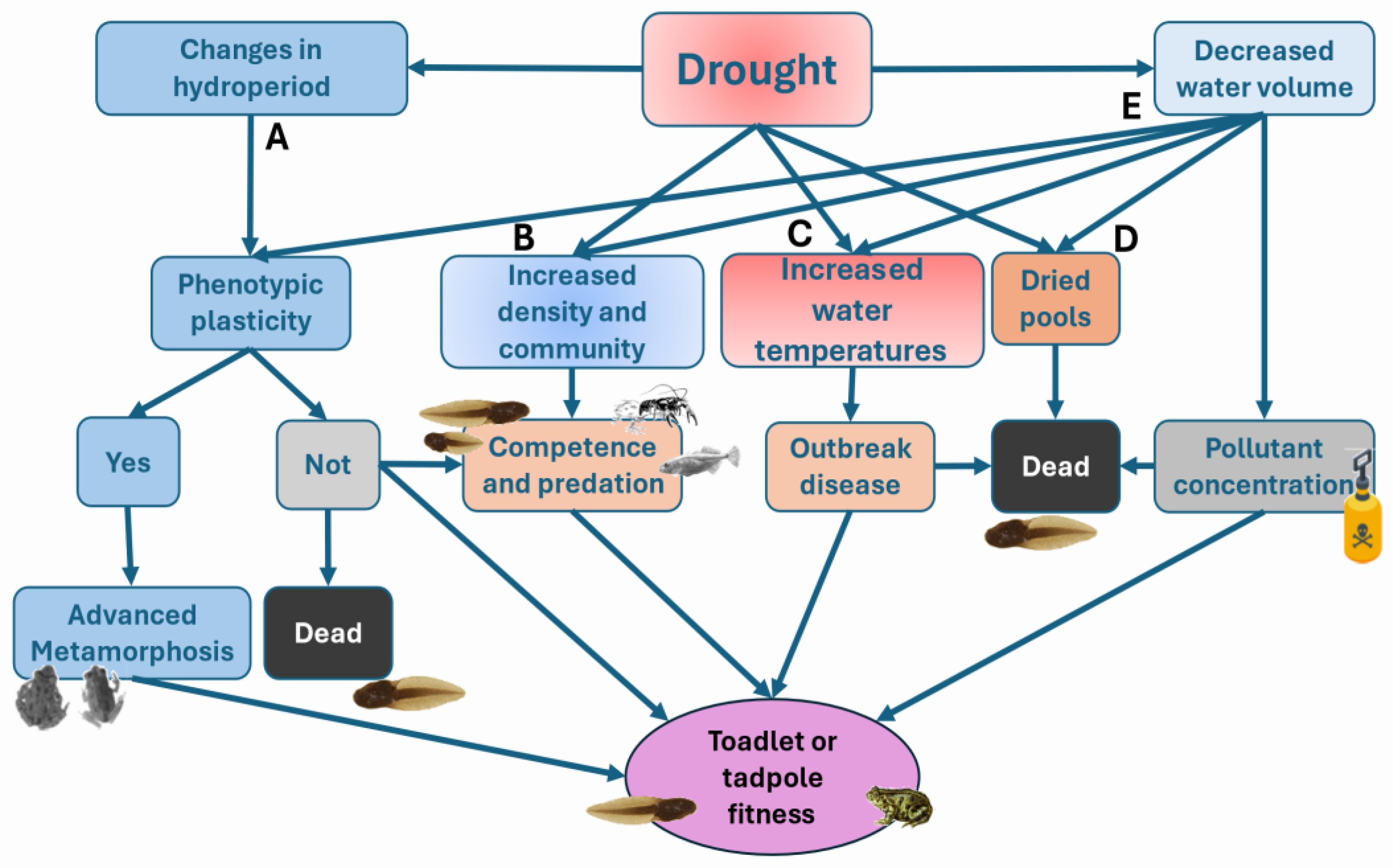
Figure 4.
Schematic diagram of the pathways via which droughts may change host response to disease. A: Drought produces changes in pond communities and alters skin microbiomecompromising host response to the pathogen. B: Higher density of tadpoles increases infection rates. C: Drought increases pollutant concentration affecting host AMP (Antimicrobial peptides) production and host immunity, D: Drought reduces host immunity by means of increased corticosterone. E: The increase of water temperature produced by drought facilitates the outbreak of emerging diseases. Modified from Blaustein et al 2018.
Figure 4.
Schematic diagram of the pathways via which droughts may change host response to disease. A: Drought produces changes in pond communities and alters skin microbiomecompromising host response to the pathogen. B: Higher density of tadpoles increases infection rates. C: Drought increases pollutant concentration affecting host AMP (Antimicrobial peptides) production and host immunity, D: Drought reduces host immunity by means of increased corticosterone. E: The increase of water temperature produced by drought facilitates the outbreak of emerging diseases. Modified from Blaustein et al 2018.
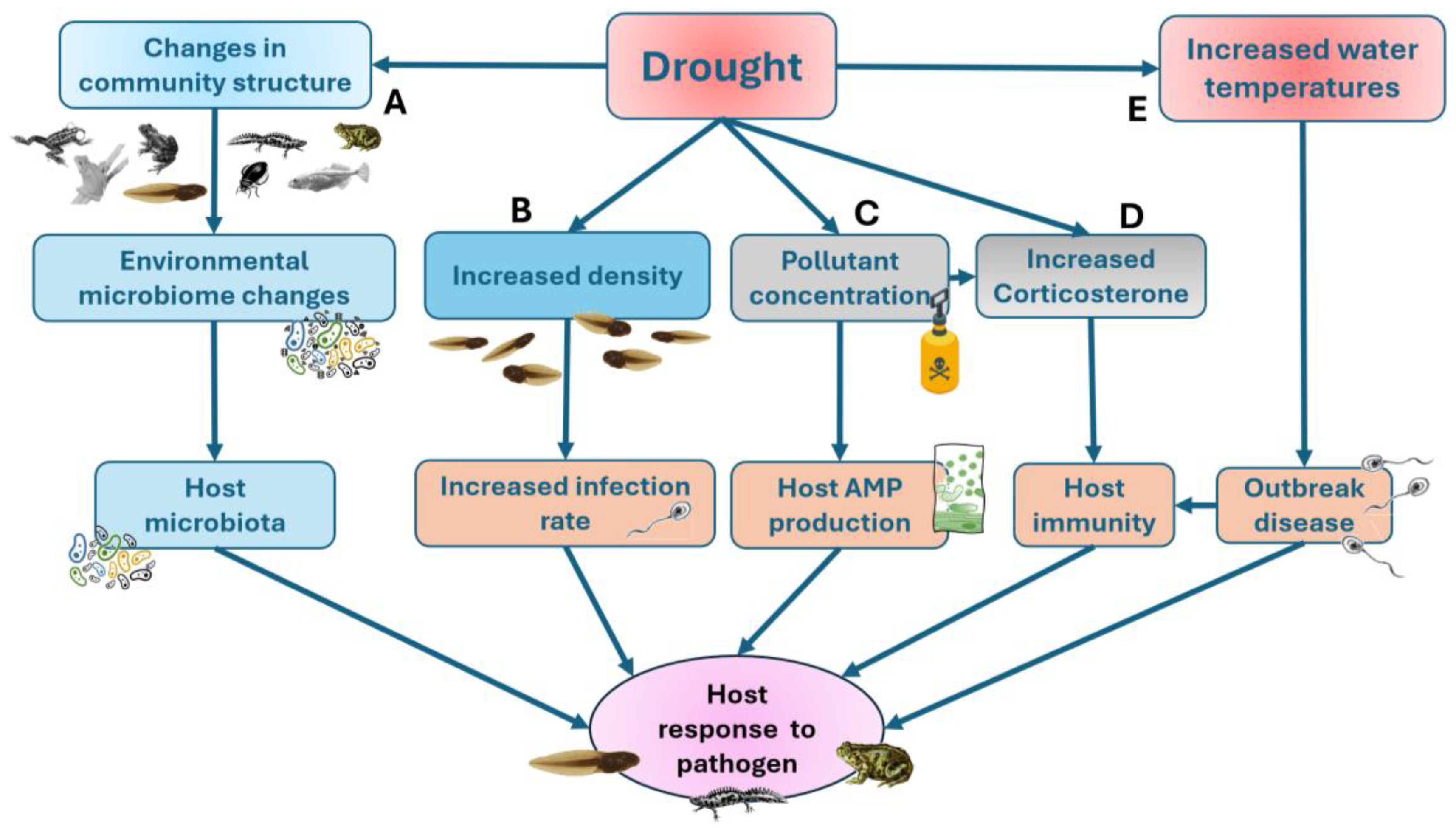
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



