
Submitted:
10 July 2025
Posted:
11 July 2025
You are already at the latest version
Abstract
Background/Objectives: Methicillin-resistant Staphylococcus aureus (MRSA) remains a critical public health concern due to its multidrug resistance and capacity to form persistent infections, particularly in the context of implanted medical devices. Alternative therapeutic strategies that target bacterial virulence instead of viability are increasingly explored. This study aimed to evaluate the antimicrobial and antivirulence activity of an extract derived from Lacticaseibacillus rhamnosus CRL 2244 against two MRSA strains—USA300 and M86—and to elucidate its effects on bacterial physiology and gene expression under host-mimicking conditions. Methods: Antimicrobial activity was assessed using agar diffusion, MIC, and time-kill assays. Scanning electron microscopy of cells exposed to the extract confirmed decreased cellular density and morphological changes. Phenotypic assays evaluated biofilm formation, staphyloxanthin production, and adhesion to fibronectin. RT-qPCR analyzed transcriptional responses. Viability was assessed in the presence of human serum and type I collagen. Results: The CRL 2244 extract demonstrated bactericidal activity with up to 6-log10 CFU/mL reduction at 1x MIC. In USA300, the extract reduced the expression of hla, lukAB, fnbA, and icaA, correlating with decreased staphyloxanthin levels. In M86, a significant reduction in biofilm formation and repression of lukAB, nucA, and fnbA were observed. Adhesion to fibronectin was impaired in both strains. The extract showed no cytotoxicity in human serum but reduced viability in collagen-enriched conditions. Conclusions: The Lcb. rhamnosus CRL 2244 extract modulates MRSA virulence in a strain-specific manner, targeting key regulatory and structural genes without inducing cytotoxic effects.
Keywords:
Staphylococcus aureus
; lactic acid bacteria
; probiotics
; methicillin-resistant
; transcriptional response
; secreted compounds
1. Introduction
Bacterial resistance to antibiotic represents one of the most pressing global health challenges of the 21st century (WHO, 2024). The increase in infections caused by resistant pathogens not only limits therapeutic options, but also increase morbidity, mortality and healthcare costs. Among these pathogens, methicillin-resistant Staphylococcus aureus (MRSA) has been designated as a critical priority by both the World Health Organization (WHO) and the Centers for Disease Control and Prevention (CDC) due to its high prevalence and clinical impact [1,2,3].
MRSA is responsible for a wide range of acute and chronic infections, both hospital-acquired (HA-MRSA) and community-acquired (CA-MRSA). While HA-MRSA is a major cause of pneumonia, bloodstream infections, and surgical site infections, CA-MRSA is mainly associated with skin and soft tissue infections, but can also cause serious conditions such as osteomyelitis and toxic shock syndrome [4,5,6]. In addition, MRSA plays a key role in chronic lung infections in patients with cystic fibrosis, where its ability to form biofilms and acquire antibiotic-tolerant phenotypes makes it difficult to eradicate and worsens patient outcomes [7,8]. The pathogenic success of S. aureus lies in its diverse arsenal of virulence factors that facilitate colonization, immune evasion and tissue invasion. In particular, Panton-Valentine leukocidin (PVL), which is more frequently associated with CA-MRSA strains, induces neutrophil lysis and contributes to tissue necrosis. Enterotoxins and toxic shock syndrome toxin-1 (TSST-1) are implicated in foodborne illness and systemic toxicity. In addition, adhesin expression and biofilm-forming ability enable persistence in host tissues and medical devices, further enhancing resistance. Protein A, coagulase, and capsular polysaccharides also contribute to immune evasion and virulence [9,10,11,12].
The global propagation of MRSA strains resistant to multiple antibiotic classes has prompted an urgent search for alternative antimicrobial strategies [13,14]. Among these, lactic acid bacteria (LAB) have shown promise due to their Generally Recognized as Safe (GRAS) status, natural antimicrobial activity and their health-promoting effects, particularly in maintaining gut health and modulating the immune system [15,16]. Recent studies have reported the ability of LAB to inhibit multidrug-resistant pathogens, including MRSA [17,18] . Our group recently demonstrated that Lacticaseibacillus rhamnosus CRL 2244 secretes antimicrobial compound(s) with bactericidal activity against carbapenem-resistant Acinetobacter baumannii (CRAB) as well as other clinically relevant pathogens, including MRSA [19]. Notably, these compounds also act synergistically with β-lactam antibiotics and vancomycin, restoring the susceptibility of S. aureus strains resistant to these drugs [19].
Based on these findings, the present study investigates the antimicrobial activity of the extract from Lcb. rhamnosus CRL 2244 against MRSA strains, with emphasis on its potential clinical application. We employed both phenotypic and transcriptomic approaches to assess the bactericidal effects of the extract on two representative MRSA strains: the epidemic community-associated USA300 clone (characterized by SCCmec type IVc and the PVL gene) [20,21] and the hospital-associated strain M86, isolated from a cystic fibrosis patient [22]. In addition, we evaluated its efficacy in physiologically relevant conditions, such as human serum, collagen matrices and, fibronectin-binding assays—targeting a host protein crucial for S. aureus colonization. This study provides new insights into the therapeutic potential of LAB-derived antimicrobials against multidrug-resistant pathogens in clinically complex infection scenarios.
2. Results
2.1. Antimicrobial Activity of Lcb. rhamnosus CRL 2244 Extract Against MRSA Strains
The antimicrobial activity of the extract obtained from Lcb. rhamnosus CRL 2244 against MRSA strains was evaluated by spot-on the lawn assay (Table 1). In CLDE medium, the extract exhibited low to moderate inhibitory activity, with inhibition halos of 10 mm for strain CA-MRSA USA300 strain and 12 mm for HA-MRSA M86. In contrast, high activity was observed on blood agar, with halos of 20 mm for both strains, indicating a significant increase in antimicrobial activity on this medium. This effect could be attributed to the blood agar components that promote diffusion or stability of the active compound. Likewise, the activity of the extract was evaluated against isogenic mutants of USA300 in virulence-related genes, using CLDE plates. Inhibition halos ranging from 15 to 20 mm were observed, with the greatest effect recorded for the Δhla mutant (IDH = 20 mm), suggesting that the response to the extract could be influenced by pathogen-specific virulence factors (Table 1).
The minimum inhibitory concentration (MIC) of the Lcb. rhamnosus CRL 2244 extract was determined against MRSA strains USA300 and M86. The MIC value for both strains was found to be 5 µg/µL, indicating that this concentration was sufficient to inhibit visible bacterial growth after 24 h of incubation.
2.2. Morphological Changes Induced by Lcb. rhamnosus CRL 2244 Extract on S. aureus Cells
In addition, microscopy analysis and colony counting were performed to assess morphological alterations and reductions in cellular density in S. aureus cultures exposed to the extract. Treatment of S. aureus USA300 with the extract at 2x and 4x MIC resulted in noticeable morphological changes and a marked decrease in bacterial cell density, confirming the extract’s ability to impair bacterial growth (Figure 1; Figure S1). Under control conditions, bacterial cells appeared in dense aggregates with smooth surfaces and intact coccoid morphology (Figure 1). In contrast, treatment with the extract at 2x MIC led to a reduction in cell density, with fewer cells (Figure 1; Figure S1). Cell exposed to 2x MIC showed morphological irregularities, suggesting membrane stress or early structural compromise (Figure 1). At 4x MIC, these effects were markedly intensified. Only few cells sparsely distributed were seen, with visible signs of morphological changes, in particular an increase in cell length after exposure to the extract (Figure 1). The progressive loss of integrity and density observed across increasing concentrations supports the extract’s dose-dependent bactericidal effect, further confirmed by quantitative CFU assays (Figure 1; Figure S1).
2.3. Bactericidal Effect of Lcb. rhamnosus CRL 2244 Extract Against MRSA Strains
To evaluate the impact of Lcb. rhamnosus CRL 2244 extract (E) on the viability of the two MRSA strains, USA300 and M86, time-kill assays were performed (Figure 2). In both cases, bacterial cultures treated with 1x MIC of the extract showed a marked decrease in viable cell count over time. In the case of strain USA300, treatment with 1x MIC resulted in a reduction of ~6 log10 CFU/mL after 24 h compared to the untreated control, indicating a potent bactericidal effect (Figure 2). A similar trend was observed for strain M86, with bacterial counts below the limit of detection at 24 h post-treatment. In contrast, treatments with 0.5x MIC of the extract led to a moderate decrease in bacterial load, maintaining bacteriostatic activity for the same period (Figure 2). Untreated controls for both strains showed exponential growth, confirming the antimicrobial efficacy of the CRL 2244 extract in a both a dose and time-dependent manner.
To evaluate the potential synergistic or additive effect of the CRL 2244 extract in combination with a β-lactam antibiotic, a killing assay was performed using subinhibitory concentrations of ampicillin. A standardized inoculum of S. aureus USA300 was cultured in the presence of 0.5x MIC ampicillin alone or in combination with 0.5x MIC of the extract. Bacterial viability was assessed at 0, 2, 4, 6, and 24 hours. As shown in Figure S2A, treatment with ampicillin resulted in a ~ 4 log10 reduction in CFU/mL over time, whereas the combination treatment led to a more pronounced decrease in bacterial viability (Figure S2A), supporting a potential additive interaction between the extract and ampicillin. Interestingly, in addition to reduced viability, cultures treated with the combination of ampicillin and extract also displayed a small colony phenotype (Figure S2B).
2.4. MRSA Transcriptional Changes Triggered by the Extract from Lcb. rhamnosus CRL 2244
To assess the transcriptional response of MRSA to the CRL 2244 extract, we performed quantitative RT-PCR (qRT-PCR) on the CA-MRSA strain USA300 and the HA-MRSA strain M86 following exposure to sub-inhibitory concentrations (sub-MIC) of the extract (Figure 3). The results revealed significant transcriptional changes of several virulence-associated genes, particularly in USA300. In USA300, the exposure to the extract (USA300 E) resulted in strong upregulation of agrA, sarA, and cflA (p < 0.01 for agrA and sarA), compared to the untreated control (Figure 3A). agrA is part of the agr quorum sensing system that regulates virulence, while sarA plays a major role in regulation of biofilm formation. clfA, which encodes clumping factor A, also showed increased expression, possibly reflecting a compensatory response aimed at enhancing adhesion in the face of biofilm disruption. In contrast, genes associated with cytotoxin production, including hla (α-hemolysin) and lukAB (leucocidin), were significantly downregulated, suggesting that the extract may reduce the cytotoxic potential of S. aureus. nucA, which encodes a thermonuclease involved in biofilm matrix degradation, was also downregulated, possibly reflecting an alteration in biofilm structure. Furthermore, key adhesion-related genes, fnbA and icaA, were significantly downregulated in response to the extract (Figure 3A), indicating that the extract interferes with major biofilm formation pathways, likely weakening S. aureus colonization and persistence. Interestingly, no significant differences were detected in the expression levels of sigB, a general stress response regulator, and spaA, associated with immune evasion, suggesting that the extract primarily affects virulence and biofilm regulation rather than general stress response pathways.
In the HA-MRSA M86 strain (Figure 3B), qRT-PCR analysis revealed a distinct transcriptional response to the extract compared to USA300. Significant upregulation was observed in agrA, spa, and cflA (Figure 3B), while lukAB, nucA, and fnbA were significantly downregulated (Figure 3B). No changes were detected in the transcript’s levels of sigB, sarA, hla, and icaA remained unchanged, indicating a more moderate and strain-specific response compared to USA300.
Taken together, these findings demonstrate that exposure to the CRL 2244 extract modulates key virulence-related genes in both MRSA strains, with USA300 exhibiting a broader and more pronounced transcriptional shift. These suggests that the extract may differentially impact MRSA strains depending on their genetic background and virulence profile, providing insight into its potential use as a strain-specific anti-virulence strategy.
2.5. Effect of the Extract on MRSA Virulence: Inhibition of Biofilm Formation and Staphyloxanthin Production
The biofilm-forming ability of MRSA strains USA300 and M86 was evaluated under untreated and extract-treated conditions (USA300 E and M86 E, respectively). Biofilm quantification, expressed as the OD₅₉₂/OD₆₀₀ ratio, showed that strain M86 produced significantly more biofilm compared to the other conditions analyzed (Figure 4A). However, exposure to the CRL 2244 extract (M86 E) drastically reduced its biofilm formation, bringing levels down to those observed in USA300 and USA300 E (Figure 4A). No significant differences were detected between USA300 and its extract-treated condition (Figure 4A), indicating that the extract has a more pronounced antibiofilm effect on high biofilm-forming strains like M86. These findings suggest that the extract specifically targets and disrupts robust biofilm architectures, likely interfering with structural components or regulatory pathways involved in biofilm development.
These assays were conducted in LB medium, a nutrient-rich environment commonly used for bacterial growth, and the observed effects were consistent when tested in the more enriched Terrific Broth (TB). This consistency across media confirms the robustness and reproducibility of the extract’s antibiofilm activity (data not shown).
Staphyloxanthin production was also evaluated in the same experimental conditions. Compared to the untreated strain, USA300 exposed to the extract (USA300 E) exhibited a significant reduction in pigment production (Figure 4B). In contrast, no significant difference in staphyloxanthin production was observed between M86 and its treated counterpart (M86 E) (Figure 4B). Changes in pigment production may reflect strain-specific metabolic shifts in S. aureus, which may vary depending on the strain’s baseline levels and susceptibility to the treatment. Taken together with the biofilm data, these findings underscore a strain-dependent response to the CRL 2244 extract, with effects observed on multiple virulence-associated phenotypes.
2.6. Impact of the Lcb. rhamnosus CRL 2244 Extract on MRSA Viability in Host-Mimicking Conditions
The antimicrobial activity of the CRL 2244 extract was evaluated under physiologically relevant conditions using MRSA strains USA300 and M86 (Figure 5). In 100% human serum, no significant differences in bacterial counts were observed between untreated and extract-treated cultures for either strain. This lack of activity could be linked to the extract’s reduced solubility soluble in serum, which could interfere with its antimicrobial efficacy. In contrast, exposure to the extract in type I collagen-supplemented medium resulted in a significant reduction in bacterial survival. For strain USA300, extract exposure led in a moderate but statistically significant decrease in CFU/mL (Figure 5A). A more pronounced effect was seen in M86, with approximately a 1-log decrease in CFU/mL (Figure 5B). These findings suggest that the bactericidal effect of the extract is condition-dependent, with enhanced effectiveness in collagen-rich environments. This may be due to interactions between type I collagen and bacterial surface adhesins, potentially facilitating increased susceptibility to the bioactive compounds in the extract. These results support the potential of CRL 2244-derived antimicrobials to function in host-like settings where extracellular matrix components play a key role in bacterial colonization.
2.7. Effect of the Lcb. rhamnosus CRL 2244 Extract on Bacterial Adhesion of MRSA Strains
To assess the impact of the Lcb. rhamnosus CRL 2244 extract on the adhesion of MRSA strains USA300 and M86, fibronectin-coated wells were inoculated with bacterial cultures in the presence of the extract at two different MIC concentrations (co-treatment) and in its absence (control). Simultaneous exposure to the extract at 1x MIC and 2x MIC during the adhesion phase significantly reduced bacterial attachment for both strains (Figure 6). For USA300, adhesion was reduced by 28.27% and 26.86% at 1x MIC and 2x MIC, respectively (Figure 6A). M86 showed similar reductions of 27.62% and 28.98% under the same treatment conditions (Figure 6B). Both strains exhibited maximal adhesion in the untreated control group (100%), which served as a baseline for comparisons in the other experimental conditions (Figure 6A,B).
3. Discussion
In a context of the growing threat posed by infections caused by multidrug-resistant pathogens such as MRSA, compounds derived from LAB have been postulated as innovative therapeutic alternatives. Their safety, metabolic diversity and ability to modulate virulence mechanisms without exerting direct selective pressure on bacterial viability make them attractive candidates for antivirulence strategies [23,24,25,26]. In particular, metabolites produced by LAB have shown inhibitory effects against multidrug-resistant clinical strains by impairing adhesion, interfering with biofilm-forming, and modulating the expression of virulence factor-associated genes, which is especially relevant in clinical settings involving implantable devices, such as heart valves or catheters, which promote colonization and persistent infections [27,28,29].
In this study, we demonstrated that the extract derived from Lcb. rhamnosus CRL 2244 exerts a modulatory effect on MRSA USA300 and M86 strains at both the transcriptional and phenotypic levels, notably reducing staphyloxanthin production and adhesion capacity under host-mimicking conditions. Although the chemical identity of the active compound(s) remains undefined, the findings presented here represent a key step toward functional characterization and support the potential of CRL 2244 extract as an antivirulence agent for preventing or treating MRSA infections in clinical settings.
The extract showed robust antimicrobial activity against both community-associated (USA300) and hospital-associated (M86) MRSA strains, consistent with prior observations in CRAB strains [19,30], supporting the hypothesis of a broad-spectrum mechanism of action. The enhanced inhibition observed on blood agar plates suggests that medium components may influence the diffusion or stability of bioactive molecules, as previously reported [31,32]. Interestingly, the increased susceptibility of the isogenic USA300 Δhla mutant points to a link between virulence factors and extract sensitivity. hla, which encodes α-hemolysin, plays a central role in S. aureus cytotoxicity and immune evasion [33]. Its deletion may reduce the pathogen’s capacity to counteract the stress imposed by the extract, as similarly observed with other antimicrobials targeting virulence factors [34]. Time-kill assays validated that the extract’s bactericidal activity, with reductions of up to 6 log10 CFU/mL after 24 h at 1x MIC in both strains, similar to previous results with CRAB strains [30]. Consistent with this potent bactericidal effect, SEM analysis revealed morphological alterations in S. aureus cells exposed to the extract, including a reduction in cell density and an increase in cell length, features previously observed in other multidrug-resistant pathogens treated with this extract [29]. These structural changes suggest a mechanism of action involving alteration of the cell envelope and were found to be dose-dependent, in agreement with the activity of the extract observed in CFU and MIC assays.
At the molecular level, the extract induced transcriptional changes consistent with virulence modulation. In USA300, we observed a broader transcriptional response than M86. USA300 showed overexpression of agrA and sarA, two master regulators of quorum sensing and biofilm formation, respectively. Despite this, expression of critical virulence genes—including hla, lukAB, icaA, and fnbA—was significantly reduced. This apparent decoupling between regulatory elements and effector genes suggests a reprogramming of the virulence profile rather than a global activation, a phenomenon previously reported in response to environmental or metabolic stimuli [34]. Wang et al. (2024) demonstrated that Lcb. rhamnosus P118 can produce tryptophan-derived metabolites (e.g., indole) that modulate host inflammation [35]. While immunomodulatory effects were not assessed in this study, reduced expression of key toxins (hla, lukAB) may imply a diminished capacity to trigger host inflammatory responses.
Phenotypically, USA300 did not exhibit significant biofilm changes, likely due to its low basal biofilm formation under tested conditions. In contrast, staphyloxanthin production was markedly reduced, correlating with hla repression and potentially a less oxidative cellular environment. As a key antioxidant carotenoid, staphyloxanthin protects S. aureus from host-derived stress [36,37]. Reduction of staphyloxanthin, possibly mediated by sarA repression, could increase the vulnerability of the pathogen, reinforcing its value as an antivirulence target [38,39]. In line with this hypothesis, prior studies have shown that natural compounds and anti-inflammatory drugs can disrupt staphyloxanthin biosynthesis [40].
The M86, by contrast, showed a more restricted transcriptional response. While sarA, hla, and icaA expression remained unchanged, we observed increased expression of agrA and spa, and repression of lukAB, nucA, and fnbA. These molecular changes correlated with a pronounced reduction in biofilm formation, suggesting the extract may be particularly effective against high biofilm-forming strains—a trait also observed in CRAB [29]. The downregulation of nucA, involved in extracellular DNA degradation and biofilm dispersal [41], may reflect alterations in biofilm matrix maturation or stability. In contrast to USA300, M86 showed no changes in levels of staphyloxanthin production, likely due to stable expression of biosynthesis-related genes such as sigB or crtM, which were not analyzed in this study.
When tested under host-mimicking conditions, the CRL 2244 extract’s activity varied. In human serum, no significant reduction in bacterial viability was detected, suggesting low cytotoxicity in protein-rich environments. However, a significant decrease in cell viability was observed in the presence of type I collagen, particularly for M86. This indicates that certain components of the tissue microenvironment may sensitize MRSA to the effect of the extract, in agreement with studies showing that interaction with extracellular matrix proteins modulates virulence and antimicrobial susceptibility of S. aureus [9,42]. Adhesion assays using fibronectin-coated surfaces showed that the extract impaired MRSA anchoring, even at subinhibitory concentration, showing a strong effect in the highly adherent M86 strain. This phenotype correlates with reduced expression of key adhesion genes such as fnbA and icaA, supporting the hypothesis that CRL 2244 extract interferes with early colonization—an essential step in staphylococcal pathogenesis and biofilm-associated infections [34].
Finally, like transcriptomic studies by Peng et al. (2023) who showed that Lcb. rhamnosus SCB0119 modulates key genes in E. coli and S. aureus, we observed repression of hla, lukAB, and fnbA, along with functional reductions in adhesion and staphyloxanthin production [43]. These findings support the concept that LAB-derived compounds can reprogram virulence traits in a strain-dependent manner, reinforcing their value as antivirulence rather than bactericidal agents.
4. Materials and Methods
4.1. Bacterial Strains and Growth Conditions
USA300 (CA-MRSA) [20,44] and M86 (HA-MRSA) [22] strains were used in this work. In addition, a total of six S. aureus (SA) USA 300 mutants were used for spot-on-the-lawn assays to test antimicrobial activity (Table 1). The mutant in agrA, sarA, lukS-PV, cflA, fnbA (Nebraska Tn Mutant Library), and hla [45]. The strains were grown on Cystine-Lactose Electrolyte-Deficient (CLED) medium (Beckton Dickinson, Franklin Lakes, NJ, USA) overnight at 37˚C and used within 24 h.
4.2. Preparation of Extracts from Lcb. rhamnosus CRL 2244
To obtain the crude cell-free extract, cell free conditioning media (CFCM) of Lcb. rhamnosus CRL 2244 incubated for 96 h under appropriate growth conditions was used. After incubation, the culture was filtered and immediately subjected to lyophilization using a Virtis 4K benchtop lyophilizer. The lyophilized material was resuspended in 250 mL of deionized water and extracted by liquid-liquid partitioning with ethyl acetate (1:1 v/v) in a 2000 mL separatory funnel. The mixture was shaken vigorously and allowed to separate; the organic phase was collected. The aqueous phase was re-extracted twice with decreasing volumes of ethyl acetate (200 mL and 190 mL). The combined organic extracts were dried with ~30 g anhydrous magnesium sulfate, filtered and concentrated using a rotary evaporator (IKA RV 10 digital V) set at 150 rpm with gradually reduced pressure from 395 mbar to 75 mbar over 20-30 min. The concentrated extract was stored at -20 °C and used without further purification.
4.3. Antimicrobial Susceptibility Assays
The antimicrobial activity of the Lcb. rhamnosus CRL 2244 extract was assessed using a modified version of the well diffusion method outlined in a previous study [19]. Bacterial strains were resuspended in sterile saline solution (0.85% NaCl, w/v) to a concentration of 0.5 McFarland units (1.5 x 108 CFU/mL). These suspensions were then inoculated onto CLED and blood agar plates. A volume of 10 μL of the extract (800 μg/μL) was applied to the surface of the plates. After incubating at 37 °C for 24 hours, the inhibition zone diameters (IDH) were measured. The results were categorized as follows: less active (IDH ≤ 10 mm), moderately active (IDH = 11–14 mm), and highly active (IDH ≥ 15 mm) [46]. All assays were conducted using three independent biological replicates.
The minimum inhibitory concentration (MIC) of the extract was determined against MRSA strains USA300 and M86 by broth microdilution following CLSI guidelines [47]. Briefly, serial two-fold dilutions of the extract were prepared in cation-adjusted Mueller-Hinton (CAMH) broth in 96-well microplates. Bacterial suspensions were adjusted to approximately 5 x 10⁵ CFU/mL and added to each well. Plates were incubated at 37 °C for 24 h, and bacterial growth was assessed visually. The MIC was defined as the lowest extract concentration that completely inhibited visible growth.
4.4. Scanning Electron Microscopy (SEM)
To evaluate the reduction in cellular density and morphological changes induced by the extract, SEM was performed on cultures of strain USA300. A standardized inoculum (0.5 McFarland), prepared from colonies grown overnight on LB agar at 37 °C, was used to inoculate 2 mL of LB broth supplemented with the extract at 2x and 4x MIC. As a control, USA300 was cultured under identical conditions in LB broth without the addition of the extract. After incubation for 18-20 h at 37 °C, the cells were centrifuged at 5000 rpm for 5 min, washed twice with sterile saline solution and fixed in 2% glutaraldehyde solution. Cells were deposited on protamine coated glace slides, dehydrated in an ethanol series and critical point dry for subsequent visualization at the core imaging Lab, Department of Biological Science, CSUF with the JCM-7000 NeoScope™ Benchtop SEM.
Bacterial growth of the samples, prior fixation, was assessed by serial dilution and colony counting.
4.5. Time-Killing Assay
The bactericidal effect of the extract was evaluated against MRSA USA300 and M86 strains for a period of 24 h, following a previously described protocol [19]. The assays were performed in tubes using a bacterial inoculum prepared by 1:10 diluting a 0.5 McFarland (1.5 x 108 CFU/mL) suspension in Luria-Bertani (LB) broth. Cultures were exposed to the extract at concentrations of 0.5x MIC and 1x MIC, while untreated cultures (no extract) were used as controls.
Samples were incubated at 37 °C with shaking, and bacterial viability was assessed at 0, 2, 4, 6 and 24 h by serial dilutions and cultures on CLDE agar. Colony forming units (CFU) were counted after overnight incubation at 37 °C. Each condition was tested in two independent experiments, performed in duplicates. In addition, a time-killing assay with a 0.5x MIC of ampicillin and 0.5x MIC of ampicillin plus 0.5x MIC of the extract was performed to observed synergy or additive effects.
4.6. RNA Extraction and Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR) Assays
MRSA strains USA 300 and M86 cells were cultured in the absence or presence of 2.5 µg/µL of extract (sub-MIC concentration= 0.5x MIC) for 18 hours at 37 °C. Total RNA was extracted in triplicate for each condition using a commercial kit (Direct-zol RNA Kit, ZymoResearch), following treatment with 10 mg/mL of Lysostaphin (Sigma-Aldrich, St. Louis, MO, USA) and 50 mM EDTA buffer for 1 hour at 37 °C. DNase-treated RNA was then used for complementary DNA (cDNA) synthesis with the iScript™ Reverse Transcription Supermix (Bio-Rad, Hercules, CA, USA), following the manufacturer’s instructions. The cDNA concentrations were adjusted to 50 ng/µL, and qPCR was performed using qPCRBIO SyGreen Blue Mix Lo-ROX, according to the manufacturer’s guidelines (PCR Biosystems, Wayne, PA, USA). Each qPCR assay included at least three biological replicates of cDNA and was performed in triplicate using the CFX96 Touch™ Real-Time PCR Detection System (Bio-Rad, Hercules, CA, USA). The data were presented as NRQs (normalized relative quantities), calculated using the qBASE method [48,49], with recA genes as normalization controls. Experimental data were obtained from technical triplicates of three independent biological replicates. Statistically significant differences were indicated by asterisks and determined by ANOVA followed by Tukey’s multiple comparison test (P < 0.05), using GraphPad Prism (GraphPad Software).
4.7. Biofilm Formation Assay
Biofilm formation in MRSA strains USA300 and M86 was assessed using Terrific Broth (TB) and Luria-Bertani (LB) media to enhance stress conditions. The assay was adapted from the protocol described by Rodriguez et al 2023 [30], with modifications to perform it in test tubes. Bacterial cultures were prepared under two conditions: a control group consisting of 1.5 mL of LB medium and 150 µL of bacterial suspension adjusted to 0.5 McFarland, and a treatment group with the same inoculum supplemented with the extract at 0.5x MIC. Samples were incubated at 37 °C for 48 h without shaking to promote biofilm development.
After incubation, planktonic cells were removed, and the remaining biofilm was stained with 1% crystal violet solution for 30 minutes. Excess dye was discarded, and tubes were washed twice with 1x PBS. Samples were then air-dried, and the retained stain was solubilized with 30% (v/v) acetic acid for 30 minutes. Optical density was measured at 592 nm. Total biomass was previously determined by OD600 before washing, and results were expressed as the ratio between biofilm biomass and total biomass (OD592/OD600).
All experiments were performed in triplicate. Statistical analysis was conducted using the Mann–Whitney U test in GraphPad Prism (GraphPad Software, San Diego, CA, USA), with a significance threshold set at P < 0.05.
4.8. Staphyloxanthin Production Assay
The production of staphyloxanthin in MRSA strains USA300 and M86 was assessed following the protocol described by Morikawa (2021) [50]. Briefly, the strains were inoculated in terrific broth and Luria-Bertani (LB) broth, with and without exposure to a sub-MIC concentration (2.5 µg/µL) of the extract and incubated at 37 °C for 48 h with agitation. Subsequently, 850 µL of the culture was collected by centrifugation at 10,000 × g for 1 minute, washed with distilled water, and resuspended in 200 µl of methanol. The samples were then heated at 55 °C for 5 minutes and centrifuged at 15,000 × g for 1 minute to remove cellular debris. The extraction process was performed twice, and the resulting extracts were pooled into a single tube, adjusting the final volume to 1 mL with methanol. Finally, absorbance was measured at 465 nm.
The relative absorbance (RA) was calculated using the equation RA = A_treated/A_control. The results were interpreted as follows: RA = 1 indicates no effect of the antimicrobial on staphyloxanthin production; RA < 1 suggests a reduction in its production; and RA > 1 may indicate either an unexpected effect or an antimicrobial-induced increase in production.
4.9. Human Fluids Survival Assay
The effect of the extract on bacterial survival in human fluids was evaluated in MRSA strains USA300 and M86. Assays were conducted using two conditions: 100% human serum (Innovative Research, USA, certified vendor approved by ISO, FDA, USDA, and EPA) and LB broth supplemented with type I collagen (8 µg/mL physiological concentration, CalBiochem Inc). In both cases, cultures were prepared in a final volume of 1 mL, containing 100 µL of an overnight bacterial culture (adjusted to 0.5 McFarland), with or without the addition of extract at 1x MIC (5.85 µg/ml final concentration). Samples were incubated overnight at 37 °C with agitation. Bacterial viability was determined by serial dilution and plating on CLDE agar to calculate CFU/mL. Each condition was tested in two independent experiments, performed in duplicate.
4.10. Adhesion Assay
The effect of the extract on bacterial adhesion of MRSA strains USA300 and M86 was evaluated using 96-well microplates coated with Corning® BioCoat® Fibronectin Clear Flat Bottom TC, following the protocol described by Peacock et al. (2000) [51]. Fibronectin-coated wells were inoculated with 100 µL of bacterial suspensions in LB broth incubated overnight under the following conditions: untreated control (overnight cultures in LB), and co-treatment during adhesion, in which overnight cultures were added together with extract at 1x MIC or 2x MIC directly into the wells. After inoculation, all plates were incubated for 2 h at 37 °C. After incubation, the wells were washed three times with 1x PBS, fixed with 2% glutaraldehyde for 1 h, stained with 1% crystal violet for 5 min, rinsed with distilled water and air dried. The stain was then solubilized with 30% acetic acid and absorbance was measured at 405 nm to quantify adherent biomass. Adhesion was expressed as relative adhesion ratio (AR = A_treated/A_control) and as percentage adhesion relative to control (% Adhesion = [A_treated/A_control] × 100). All conditions were tested in triplicate in at least two independent experiments.
4.11. Statistical Analysis
All experiments were performed in technical duplicate or triplicate and repeated in at least three independent biological replicates, unless otherwise indicated. Data are expressed as mean ± standard deviation (SD). Statistical analyses were performed with GraphPad Prism, applying one-way or two-way ANOVA depending on the experimental design. Where appropriate, Tukey’s multiple comparison test or unpair two-tailed Student’s t-test was used as post hoc analysis. A p-value < 0.05 was considered statistically significant. Significance levels are indicated in the legend of each figure as follows: *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001; ns = not significant.
5. Conclusions
The results of this study underscore the therapeutic potential of compounds produced by LAB, particularly the extract derived from Lcb. rhamnosus CRL 2244. Rather than functioning as a conventional antimicrobial, this extract modulates S. aureus virulence by targeting key regulatory pathways in a strain-dependent manner. Given the increasing prevalence of multidrug-resistant S. aureus infections and the limited number of effective therapeutic options, these findings underscore the importance of developing alternative therapeutic approaches. Further in vivo studies will be essential to assess the clinical efficacy, safety, and potential effect on bacterial adaptation associated with long-term use of this extract.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1. Viability of S. aureus USA300 cultured in LB broth and supplemented with 2x and 4x MIC of the extract determined by serial dilution and CFU count before fixation for SEM analysis. Figure S2. A) Time-kill assay evaluating the effect of CRL 2244 extract in combination with ampicillin against S. aureus USA300. Viability was assessed by CFU enumeration at 0, 2, 4, 6, and 24 h. Data represent the mean ± SD of biological replicates. B) Treatment of USA300 cultures with the combination of ampicillin and the extract resulted in the appearance of small colony variants.
Author Contributions
Conceptualization, C.R. and M.S.R.; methodology, C.R., B.G., V.M., N.T.S. and M.S.R.; investigation, C.R., B.G., V.M., N.T.S. and M.S.R.; data analysis, C.R. and M.S.R.; writing original-draft preparation, C.R. and M.S.R.; writing-review and editing, C.R., R.R.B., G.R., N.T.S. and M.S.R.; funding acquisition, M.S.R. and N.T.S. All authors have read and agreed to the published version of the manuscript.
Funding
The research was supported by the Office of Research and Sponsored Projects 2024-2025 Research, Scholarship, and Creative Activity (RSCA) to MSR and NTS. In addition, the research was partly funded by the U-RISE at Cal State Fullerton grant 5T34 GM149493-02 to support BG. The content of this paper is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgements
The authors are grateful to Steve Karl from at the core imaging Lab, Department of Biological Science, College of Natural Sciences and Mathematics, California State University Fullerton, for his valuable technical assistance and support with the SEM analyses.
Conflicts of Interest
The authors declare no conflict of interest.
References
- (WHO). WHO. Global Priority List of Antibiotic-resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics. 2017.
- CDC. Antibiotic Resistance Threats in the United States. Atlanta, GA: US Department of Health and Human Services, CDC; 2019. 2019.
- Collaborators GBDAR. Global burden of bacterial antimicrobial resistance 1990-2021: a systematic analysis with forecasts to 2050. Lancet. 2024;404:1199-226.
- Defres S, Marwick C, Nathwani D. MRSA as a cause of lung infection including airway infection, community-acquired pneumonia and hospital-acquired pneumonia. Eur Respir J. 2009;34:1470-6.
- Kong EF, Johnson JK, Jabra-Rizk MA. Community-Associated Methicillin-Resistant Staphylococcus aureus: An Enemy amidst Us. PLoS Pathog. 2016;12:e1005837.
- Zhao JC, Zhang XH, Zhang N, Wu ZD, Wang J, Yu QH, et al. Complicated Posterior Cervical Skin and Soft Tissue Infections at a Single Referral Center. Biomed Res Int. 2020;2020:5230763.
- Lu KY, Wagner NJ, Velez AZ, Ceppe A, Conlon BP, Muhlebach MS. Antibiotic Tolerance and Treatment Outcomes in Cystic Fibrosis Methicillin-Resistant Staphylococcus aureus Infections. Microbiol Spectr. 2023;11:e0406122.
- Jean-Pierre V, Boudet A, Sorlin P, Menetrey Q, Chiron R, Lavigne JP, et al. Biofilm Formation by Staphylococcus aureus in the Specific Context of Cystic Fibrosis. International journal of molecular sciences. 2022;24.
- Foster TJ, Geoghegan JA, Ganesh VK, Hook M. Adhesion, invasion and evasion: the many functions of the surface proteins of Staphylococcus aureus. Nature reviews Microbiology. 2014;12:49-62.
- Tong SY, Davis JS, Eichenberger E, Holland TL, Fowler VG, Jr. Staphylococcus aureus infections: epidemiology, pathophysiology, clinical manifestations, and management. Clin Microbiol Rev. 2015;28:603-61.
- Ford CA, Hurford IM, Cassat JE. Antivirulence Strategies for the Treatment of Staphylococcus aureus Infections: A Mini Review. Front Microbiol. 2020;11:632706.
- Kwiecinski JM, Horswill AR. Staphylococcus aureus bloodstream infections: pathogenesis and regulatory mechanisms. Current opinion in microbiology. 2020;53:51-60.
- Abdullahi IN, Latorre-Fernandez J, Reuben RC, Trabelsi I, Gonzalez-Azcona C, Arfaoui A, et al. Beyond the Wild MRSA: Genetic Features and Phylogenomic Review of mecC-Mediated Methicillin Resistance in Non-aureus Staphylococci and Mammaliicocci. Microorganisms. 2023;12.
- Lan T, Zhang B, Liu JL, Jia Q, Gao J, Cao L, et al. Prevalence and Antibiotic Resistance Patterns of Methicillin-Resistant Staphylococcus aureus (MRSA) in a Hospital Setting: A Retrospective Study from 2018 to 2022. Indian J Microbiol. 2024;64:1035-43.
- Cazorla SI, Maldonado-Galdeano C, Weill R, De Paula J, Perdigon GDV. Oral Administration of Probiotics Increases Paneth Cells and Intestinal Antimicrobial Activity. Front Microbiol. 2018;9:736.
- De Filippis F, Pasolli E, Ercolini D. The food-gut axis: lactic acid bacteria and their link to food, the gut microbiome and human health. FEMS Microbiol Rev. 2020;44:454-89.
- Ladha G, Jeevaratnam K. A novel antibacterial compound produced by Lactobacillus plantarum LJR13 isolated from rumen liquor of goat effectively controls multi-drug resistant human pathogens. Microbiol Res. 2020;241:126563.
- Rastogi S, Mittal V, Singh A. Selection of Potential Probiotic Bacteria from Exclusively Breastfed Infant Faeces with Antagonistic Activity Against Multidrug-Resistant ESKAPE Pathogens. Probiotics Antimicrob Proteins. 2021;13:739-50.
- Rodriguez C, Ramlaoui D, Gasca B, Azis A, Leal C, Lopez C, et al. Lacticaseibacillus rhamnosus CRL 2244 secreted metabolites display killing and antibiotic synergistic activity against multi-drug resistant pathogens. PLoS One. 2024;19:e0306273.
- Diekema DJ, Richter SS, Heilmann KP, Dohrn CL, Riahi F, Tendolkar S, et al. Continued emergence of USA300 methicillin-resistant Staphylococcus aureus in the United States: results from a nationwide surveillance study. Infection control and hospital epidemiology. 2014;35:285-92.
- Jiang JH, Cameron DR, Nethercott C, Aires-de-Sousa M, Peleg AY. Virulence attributes of successful methicillin-resistant Staphylococcus aureus lineages. Clin Microbiol Rev. 2023;36:e0014822.
- Endimiani A, Blackford M, Dasenbrook EC, Reed MD, Bajaksouszian S, Hujer AM, et al. Emergence of linezolid-resistant Staphylococcus aureus after prolonged treatment of cystic fibrosis patients in Cleveland, Ohio. Antimicrob Agents Chemother. 2011;55:1684-92.
- Arena MP, Capozzi V, Russo P, Drider D, Spano G, Fiocco D. Immunobiosis and probiosis: antimicrobial activity of lactic acid bacteria with a focus on their antiviral and antifungal properties. Appl Microbiol Biotechnol. 2018;102:9949-58.
- Sharma A, Gupta G, Ahmad T, Kaur B, Hakeem KR. Tailoring cellular metabolism in lactic acid bacteria through metabolic engineering. J Microbiol Methods. 2020;170:105862.
- Alonso-Roman R, Last A, Mirhakkak MH, Sprague JL, Moller L, Grossmann P, et al. Lactobacillus rhamnosus colonisation antagonizes Candida albicans by forcing metabolic adaptations that compromise pathogenicity. Nat Commun. 2022;13:3192.
- Azami S, Arefian E, Kashef N. Postbiotics of Lactobacillus casei target virulence and biofilm formation of Pseudomonas aeruginosa by modulating quorum sensing. Archives of microbiology. 2022;204:157.
- Patel M, Siddiqui AJ, Hamadou WS, Surti M, Awadelkareem AM, Ashraf SA, et al. Inhibition of Bacterial Adhesion and Antibiofilm Activities of a Glycolipid Biosurfactant from Lactobacillus rhamnosus with Its Physicochemical and Functional Properties. Antibiotics (Basel). 2021;10.
- Scott E, De Paepe K, Van de Wiele T. Postbiotics and Their Health Modulatory Biomolecules. Biomolecules. 2022;12.
- Che J, Shi J, Fang C, Zeng X, Wu Z, Du Q, et al. Elimination of Pathogen Biofilms via Postbiotics from Lactic Acid Bacteria: A Promising Method in Food and Biomedicine. Microorganisms. 2024;12.
- Rodriguez C, Ramlaoui D, Georgeos N, Gasca B, Leal C, Subils T, et al. Antimicrobial activity of the Lacticaseibacillus rhamnosus CRL 2244 and its impact on the phenotypic and transcriptional responses in carbapenem resistant Acinetobacter baumannii. Sci Rep. 2023;13:14323.
- Klancnik A, Piskernik S, Jersek B, Mozina SS. Evaluation of diffusion and dilution methods to determine the antibacterial activity of plant extracts. J Microbiol Methods. 2010;81:121-6.
- Hossain TJ. Methods for screening and evaluation of antimicrobial activity: A review of protocols, advantages, and limitations. Eur J Microbiol Immunol (Bp). 2024;14:97-115.
- Berube BJ, Bubeck Wardenburg J. Staphylococcus aureus alpha-toxin: nearly a century of intrigue. Toxins (Basel). 2013;5:1140-66.
- Otto M. Staphylococcus aureus toxins. Current opinion in microbiology. 2014;17:32-7.
- Yue BWPZJSYZLL. Lacticaseibacillus rhamnosus P118 enhances host tolerance to Salmonella infection by promoting microbe-derived indole metabolites eLige. 2025.
- Clauditz A, Resch A, Wieland KP, Peschel A, Gotz F. Staphyloxanthin plays a role in the fitness of Staphylococcus aureus and its ability to cope with oxidative stress. Infect Immun. 2006;74:4950-3.
- Liu GY, Essex A, Buchanan JT, Datta V, Hoffman HM, Bastian JF, et al. Staphylococcus aureus golden pigment impairs neutrophil killing and promotes virulence through its antioxidant activity. J Exp Med. 2005;202:209-15.
- Elmesseri RA, Saleh SE, Elsherif HM, Yahia IS, Aboshanab KM. Staphyloxanthin as a Potential Novel Target for Deciphering Promising Anti-Staphylococcus aureus Agents. Antibiotics (Basel). 2022;11.
- Elmesseri RA, Saleh SE, Ghobish SA, Majrashi TA, Elsherif HM, Aboshanab KM. Diclofenac and Meloxicam Exhibited Anti-Virulence Activities Targeting Staphyloxanthin Production in Methicillin-Resistant Staphylococcus aureus. Antibiotics (Basel). 2023;12.
- Xue L, Chen YY, Yan Z, Lu W, Wan D, Zhu H. Staphyloxanthin: a potential target for antivirulence therapy. Infect Drug Resist. 2019;12:2151-60.
- Kiedrowski MR, Kavanaugh JS, Malone CL, Mootz JM, Voyich JM, Smeltzer MS, et al. Nuclease modulates biofilm formation in community-associated methicillin-resistant Staphylococcus aureus. PLoS One. 2011;6:e26714.
- Bhattacharya M, Horswill AR. The role of human extracellular matrix proteins in defining Staphylococcus aureus biofilm infections. FEMS Microbiol Rev. 2024;48.
- Peng H, Zhou G, Yang XM, Chen GJ, Chen HB, Liao ZL, et al. Transcriptomic Analysis Revealed Antimicrobial Mechanisms of Lactobacillus rhamnosus SCB0119 against Escherichia coli and Staphylococcus aureus. International journal of molecular sciences. 2022;23.
- Tenover FC, Goering RV. Methicillin-resistant Staphylococcus aureus strain USA300: origin and epidemiology. J Antimicrob Chemother. 2009;64:441-6.
- Nygaard TK, Pallister KB, DuMont AL, DeWald M, Watkins RL, Pallister EQ, et al. Alpha-toxin induces programmed cell death of human T cells, B cells, and monocytes during USA300 infection. PLoS One. 2012;7:e36532.
- Halder D, Mandal M, Chatterjee SS, Pal NK, Mandal S. Indigenous Probiotic Lactobacillus Isolates Presenting Antibiotic like Activity against Human Pathogenic Bacteria. Biomedicines. 2017;5.
- (CLSI) CLSI. Performance standards for antimicrobial susceptibility testing: Thirty Edition informational supplement. CLSI Document M100-S30:2020. Clinical Lab Standards Institute. 2020.
- Nishimura B, Escalante J, Mezcord V, Tuttobene MR, Subils T, Actis LA, et al. Human serum albumin-induced modification of Ton-B-dependent receptor expression in cefiderocol-exposed carbapenem-resistant Acinetobacter baumannii. Int J Antimicrob Agents. 2023;62:106950.
- Mezcord V, Traglia GM, Pasteran F, Escalante J, Lopez C, Wong O, et al. Characterisation of cefiderocol-resistant spontaneous mutant variants of Klebsiella pneumoniae-producing NDM-5 with a single mutation in cirA. Int J Antimicrob Agents. 2024;63:107131.
- Morikawa K, Maruyama A, Inose Y, Higashide M, Hayashi H, Ohta T. Overexpression of sigma factor, sigma(B), urges Staphylococcus aureus to thicken the cell wall and to resist beta-lactams. Biochem Biophys Res Commun. 2001;288:385-9.
- Peacock SJ, Day NP, Thomas MG, Berendt AR, Foster TJ. Clinical isolates of Staphylococcus aureus exhibit diversity in fnb genes and adhesion to human fibronectin. J Infect. 2000;41:23-31.
Figure 1.
Scanning electron microscopy (SEM) analysis of S. aureus USA300 exposed to the CRL 2244 extract. Representative SEM micrographs of USA300 cells grown under (A) control conditions, (B) treatment with 2x MIC of the extract, and (C) treatment with 4x MIC of the extract. All images were taken at 1,000x, and 5,000x magnification; scale bars represent 10 μm and 5 μm.
Figure 1.
Scanning electron microscopy (SEM) analysis of S. aureus USA300 exposed to the CRL 2244 extract. Representative SEM micrographs of USA300 cells grown under (A) control conditions, (B) treatment with 2x MIC of the extract, and (C) treatment with 4x MIC of the extract. All images were taken at 1,000x, and 5,000x magnification; scale bars represent 10 μm and 5 μm.

Figure 2.
Lcb. rhamnosus CRL 2244 extract killing activity against MRSA. The strains USA300 and M86 were incubated at 37 °C for 24 h in presence of extract at 0.5x MIC and 1x MIC. CFU/mL were determined at different incubation times for a period of 24 h. All assays were carried out in technical duplicates.
Figure 2.
Lcb. rhamnosus CRL 2244 extract killing activity against MRSA. The strains USA300 and M86 were incubated at 37 °C for 24 h in presence of extract at 0.5x MIC and 1x MIC. CFU/mL were determined at different incubation times for a period of 24 h. All assays were carried out in technical duplicates.
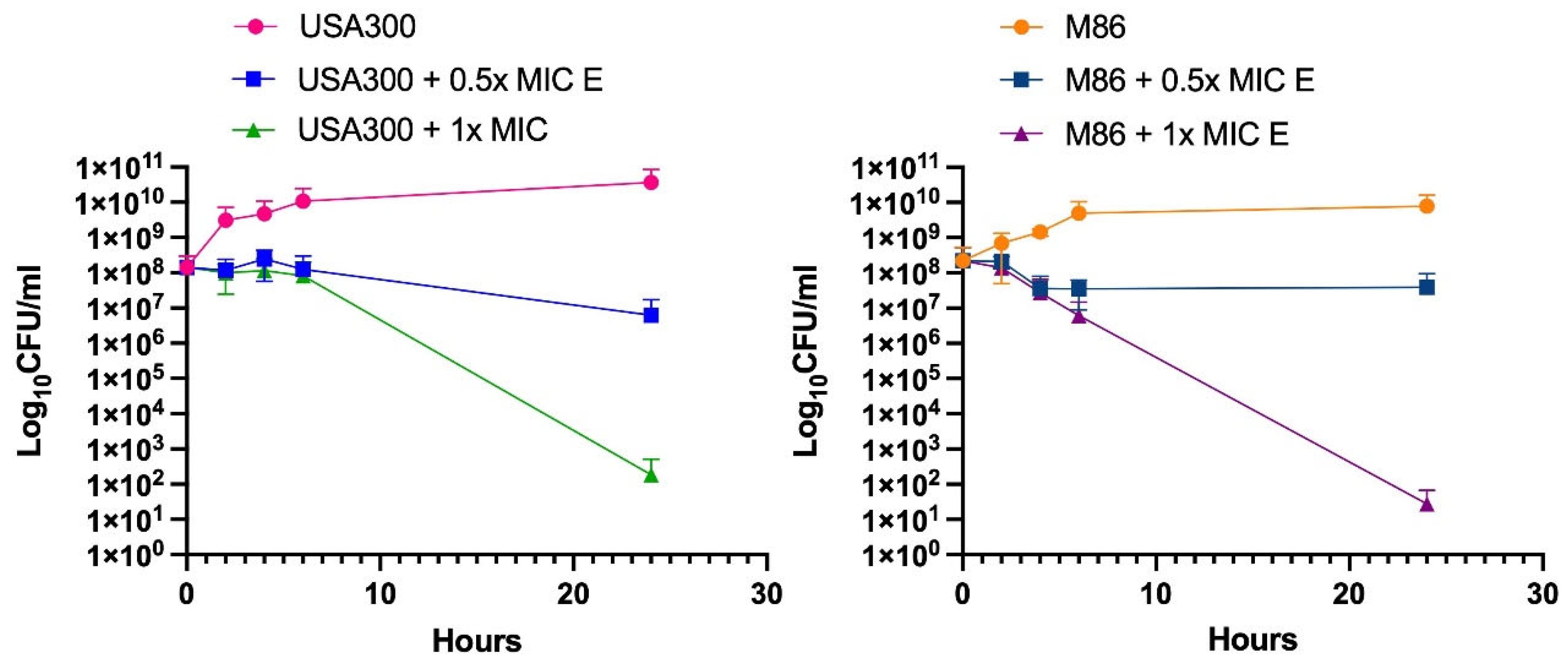
Figure 3.
Differential gene expression profiles of MRSA strains to Lcb. rhamnosus CRL 2244 extract. The clinical isolates USA300 (A) and M86 (B) were grown with and without sub-MIC concentration of extract from Lcb. rhamnosus CRL 2244 for 18 h at 37 °C. The differential gene expression was determined by qRT-PCR of genes involved in quorum sensing (agrA), global transcriptional regulation (sarA and sigB), immune evasion (spaA), cytotoxin production (hla and lukAB), biofilm formation (icaA), surface adhesion (fnbA and clfA), and nucleic acid degradation (nucA), all of which are key factors in S. aureus virulence. The data presented are the mean ± standard deviation (SD) of normalized relative to recA transcript levels calculated using the qBASE method. Statistical significance (P < 0.05) was determined by one-way ANOVA followed by T-test. *, P < 0.05; **, P < 0.005; ns, not significant. Experimental data were obtained from technical triplicates of three independent biological replicates. Error bars indicate SD.
Figure 3.
Differential gene expression profiles of MRSA strains to Lcb. rhamnosus CRL 2244 extract. The clinical isolates USA300 (A) and M86 (B) were grown with and without sub-MIC concentration of extract from Lcb. rhamnosus CRL 2244 for 18 h at 37 °C. The differential gene expression was determined by qRT-PCR of genes involved in quorum sensing (agrA), global transcriptional regulation (sarA and sigB), immune evasion (spaA), cytotoxin production (hla and lukAB), biofilm formation (icaA), surface adhesion (fnbA and clfA), and nucleic acid degradation (nucA), all of which are key factors in S. aureus virulence. The data presented are the mean ± standard deviation (SD) of normalized relative to recA transcript levels calculated using the qBASE method. Statistical significance (P < 0.05) was determined by one-way ANOVA followed by T-test. *, P < 0.05; **, P < 0.005; ns, not significant. Experimental data were obtained from technical triplicates of three independent biological replicates. Error bars indicate SD.
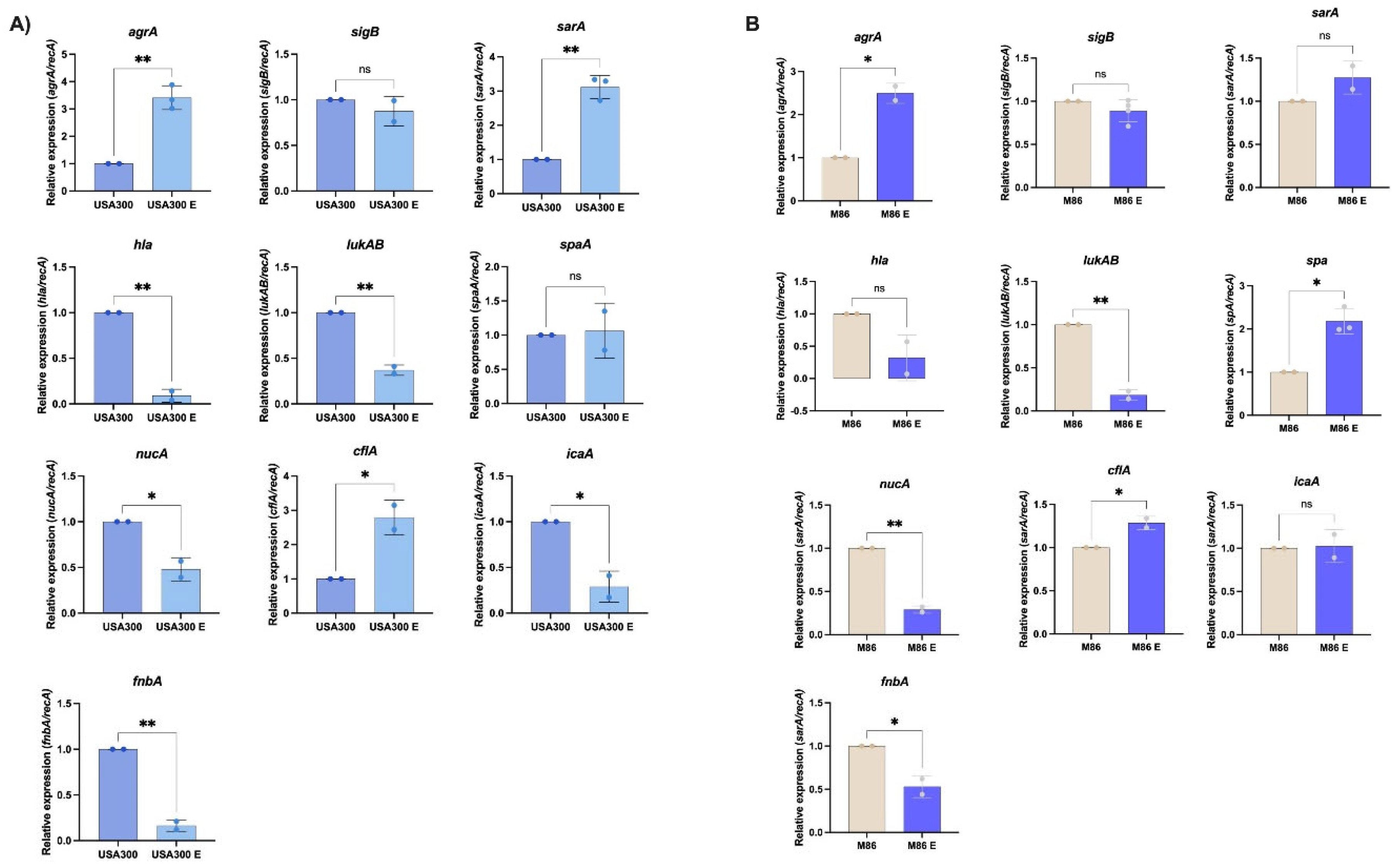
Figure 4.
Effect of Lcb. rhamnosus CRL 2244 extract on virulence features of MRSA strains USA300 and M86. (A) Biofilm formation (B) Relative staphyloxanthin production. Data represent mean ± SD from three independent experiments. Statistical significance was determined by one-way ANOVA with post hoc test (ns = not significant; **p < 0.01; ****p < 0.0001).
Figure 4.
Effect of Lcb. rhamnosus CRL 2244 extract on virulence features of MRSA strains USA300 and M86. (A) Biofilm formation (B) Relative staphyloxanthin production. Data represent mean ± SD from three independent experiments. Statistical significance was determined by one-way ANOVA with post hoc test (ns = not significant; **p < 0.01; ****p < 0.0001).
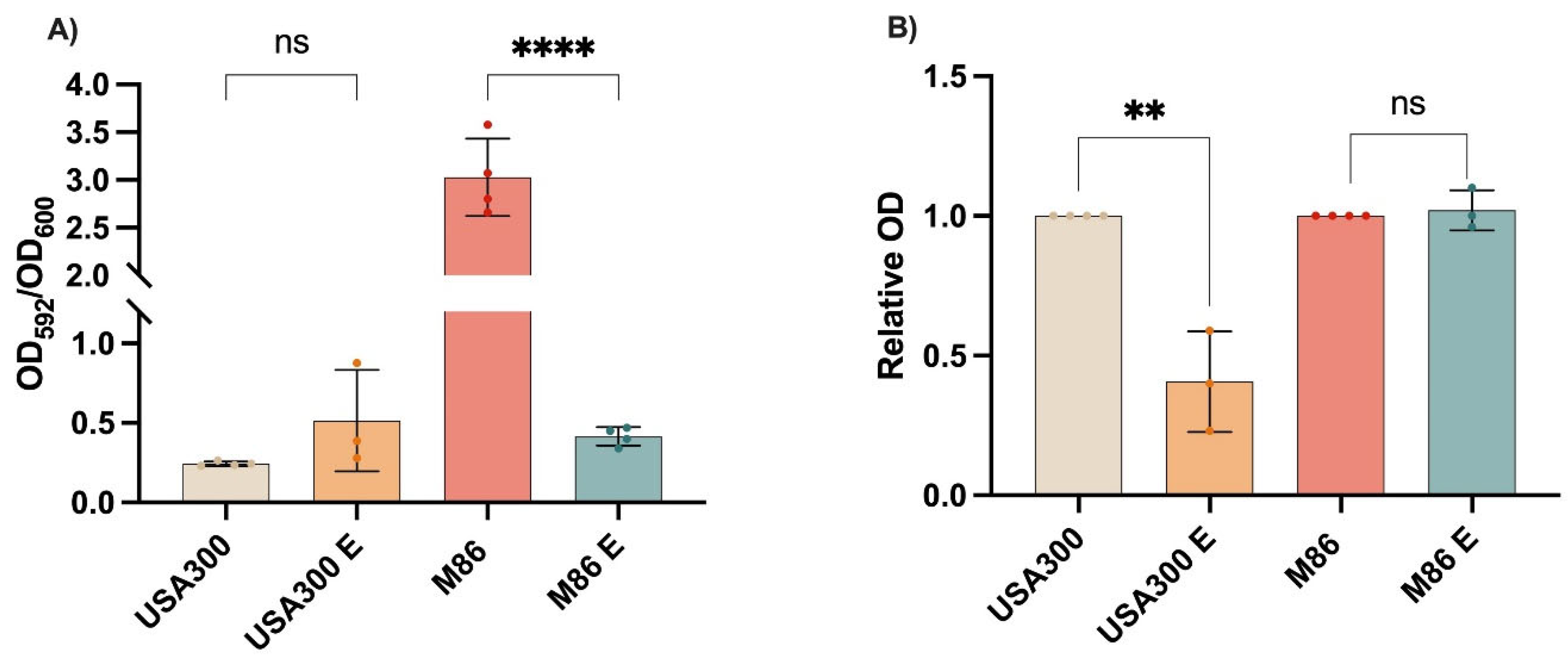
Figure 5.
Effect of Lcb. rhamnosus CRL 2244 extract on MRSA survival in human serum and collagen-supplemented medium. Viability of MRSA strains USA300 (A) and M86 (B) was evaluated after exposure to extract (E) under two physiologically relevant conditions: 100% human serum (HS) and LB broth supplemented with type I collagen (8 µg/mL). Bacterial survival was expressed as log₁₀ CFU/mL. Bars represent mean ± standard deviation of three independent experiments. Statistical significance was determined using a two-tailed unpaired t-test. *p < 0.01; p < 0.05; ns, not significant.
Figure 5.
Effect of Lcb. rhamnosus CRL 2244 extract on MRSA survival in human serum and collagen-supplemented medium. Viability of MRSA strains USA300 (A) and M86 (B) was evaluated after exposure to extract (E) under two physiologically relevant conditions: 100% human serum (HS) and LB broth supplemented with type I collagen (8 µg/mL). Bacterial survival was expressed as log₁₀ CFU/mL. Bars represent mean ± standard deviation of three independent experiments. Statistical significance was determined using a two-tailed unpaired t-test. *p < 0.01; p < 0.05; ns, not significant.
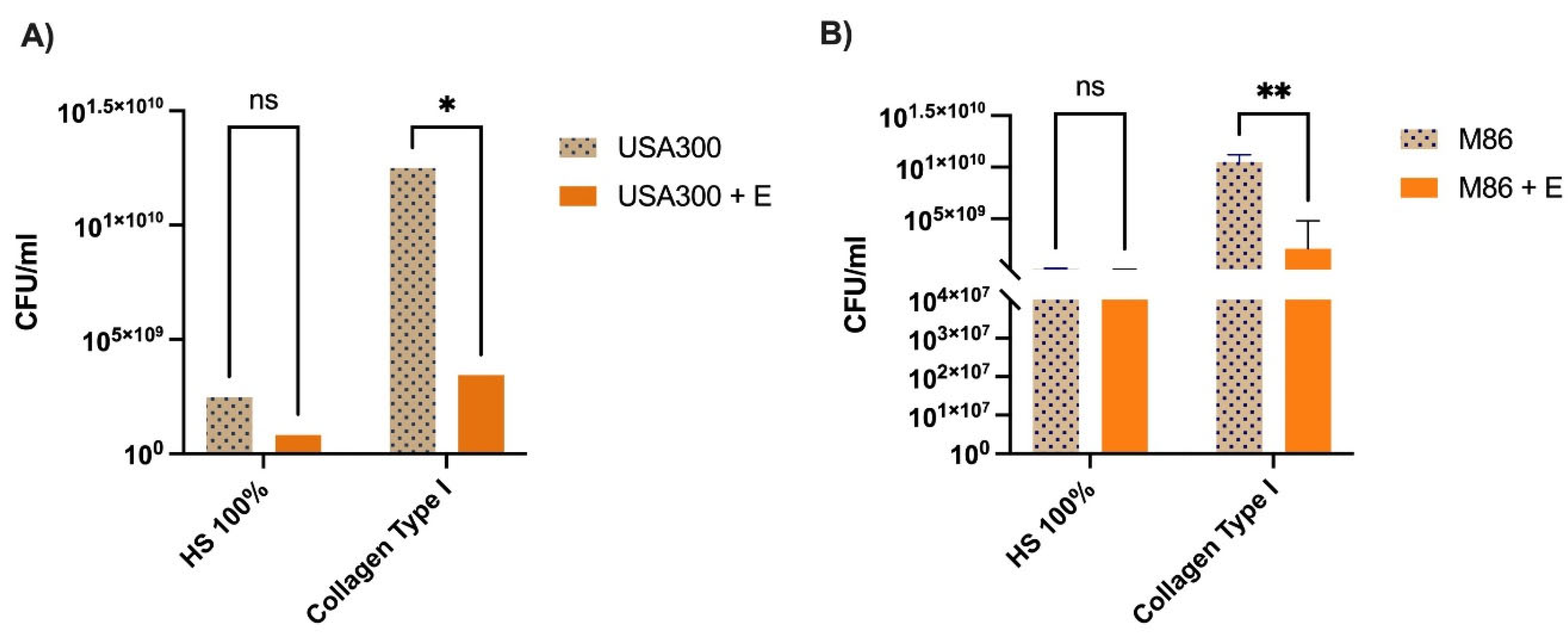
Figure 6.
Effect of the extract on adhesion of MRSA (A) USA300 and (B) M86 strains. Fibronectin-coated wells were inoculated with bacterial cultures incubated under different experimental conditions: co-treatment during the 2 h adhesion assay, in which overnight cultures together with extract at 1x MIC or 2x MIC and untreated control (overnight cultures in LB) were added to the wells. Adhesion was quantified by OD405 measurements after 2 h incubation at 37 °C. The results are expressed as absolute adhesion values (% relative to control). Data represent mean ± SD of three independent experiments. Statistical significance was determined using one-way ANOVA with Tukey’s post hoc test: p < 0.05, **p < 0.01, **p < 0.001, ns = not significant.
Figure 6.
Effect of the extract on adhesion of MRSA (A) USA300 and (B) M86 strains. Fibronectin-coated wells were inoculated with bacterial cultures incubated under different experimental conditions: co-treatment during the 2 h adhesion assay, in which overnight cultures together with extract at 1x MIC or 2x MIC and untreated control (overnight cultures in LB) were added to the wells. Adhesion was quantified by OD405 measurements after 2 h incubation at 37 °C. The results are expressed as absolute adhesion values (% relative to control). Data represent mean ± SD of three independent experiments. Statistical significance was determined using one-way ANOVA with Tukey’s post hoc test: p < 0.05, **p < 0.01, **p < 0.001, ns = not significant.
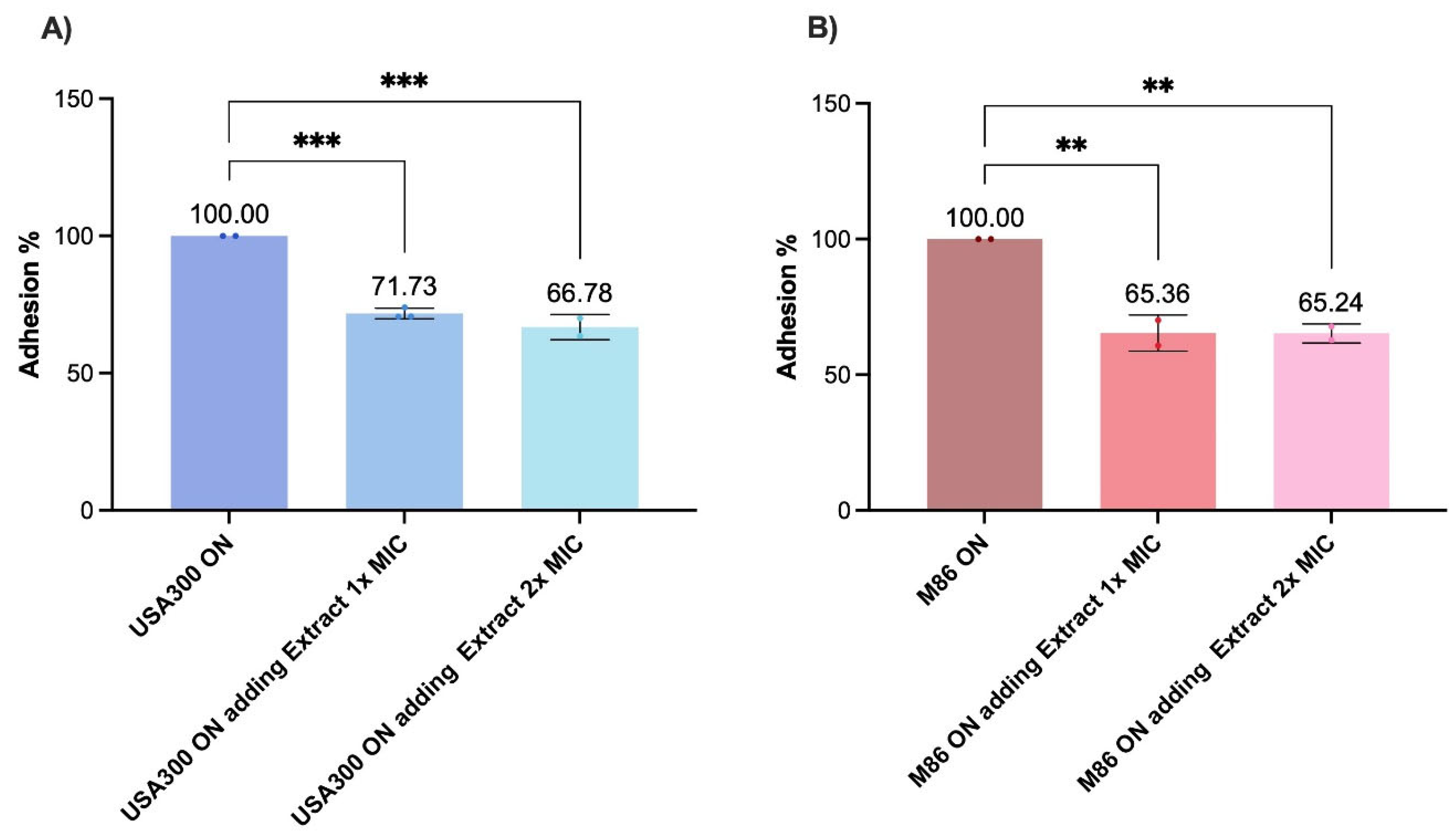

Table 1.
Antimicrobial activity of Lacticaseibacillus rhamnosus CRL 2244 extract against S. aureus strains tested by the spot-on-the-lawn method.
Table 1.
Antimicrobial activity of Lacticaseibacillus rhamnosus CRL 2244 extract against S. aureus strains tested by the spot-on-the-lawn method.
| Strains | Special features | IDH (mm) | ||||
| CLDE agar | Blood agar | |||||
| Staphylococcus aureus | ||||||
| M86 | Linezolid Resistance, PVL+ | 10 | 20 | |||
| USA 300 | mecA, PVL+ | 12 | 20 | |||
| SAUSA 300 mutants | ||||||
| 1992 | ΔagrA (accessory gene regulator protein A) | 16 | 19 | |||
| 0605 | ΔsarA (accessory regulator A) | 15 | nd | |||
| 1382 | ΔlukS-PV (Panton-Valentine leukocidin, LukS-PV | 16 | nd | |||
| 0772 | ΔclfA (clumping factor A) | 18 | nd | |||
| 2441 | ΔfnbA (fibronectin binding protein A) | 16 | nd | |||
| α-toxin | hla (α-toxin) | 20 | 20 | |||
IDH: inhibition diameter halos.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



