
Submitted:
10 July 2025
Posted:
14 July 2025
You are already at the latest version
Abstract
Listeria monocytogenes is often found in pork intestines and can contaminate pork production, posing a risk to consumers. This study aimed to characterize 16 L. monocytogenes isolates from fresh and packaged pork loin, identify their serotypes, and assess antibiotic resistance. To evaluate chitosan susceptibility as a potential strategy to control L. monocytogenes in the pork industry and to determine its effectiveness in a eukaryotic model to demonstrate pathogenicity. Sixteen isolates were evaluated, and two were determined as serotype 1/2a, twelve were 1/2b, and none were serotype 4b. Variations were observed in their pathogenicity factors. All isolates could infect chicken embryos except isolate 1.2, which displayed attenuated pathogenicity. The isolates also demonstrated strong biofilm-forming abilities. It was shown that chitosan inhibits the growth of L. monocytogenes, resulting in a viable alternative against this pathogen, but in this work, one isolate exhibited resistance to chitosan. L. monocytogenes strains present in pork meat represent a public health risk. This study characterized L. monocytogenes isolates from pork, revealing diverse serotypes (1/2a, 1/2b) and strong biofilm-forming abilities. While chitosan exhibited antimicrobial activity against most isolates, resistance was observed in one strain. These findings highlight the potential public health risk L. monocytogenes poses in pork and the need for continued research to develop effective control strategies.
Keywords:
contaminated pork meat
; chitosan
; pathogenesis
1. Introduction
Listeria monocytogenes is a pathogen found in foods and has shown persistence in food-related environments [1]. L. monocytogenes belongs to the genus Listeria, comprising 20 other species, some recently described [2]. Only L. ivanovii and L. monocytogenes are pathogenic to humans and ruminants [3]. Among these species, L. monocytogenes is classified into four genetic lineages, including 14 serotypes. Most food safety authorities worldwide consider all strains of L. monocytogenes virulent, although there is significant genetic variability among isolates obtained from diverse sources [4]. Serotypes 1/2a, 1/2b, 1/2c, and 4b are the most commonly responsible for cases of listeriosis, although they may vary in some geographic regions of the world [5,6,7,8].
The primary food sources of L. monocytogenes that have been reported include fresh, unpasteurized milk and cheeses, ice cream, fresh or processed vegetables and fruits, fresh or undercooked poultry, sausages, hot dogs, deli meats, and fresh or smoked fish and other seafood. L. monocytogenes has also been found in fresh pet food [9]. Ready-to-eat foods represent the primary sources of L. monocytogenes contamination, and contaminated fresh meats also represent a risk. Since they are considered a nutrient-rich matrix that supports the growth of microorganisms, L. monocytogenes can continue to develop and remain viable even at sub-zero temperatures. This highlights its resilience in food storage environments [10].
L. monocytogenes` ability to persist in the processing environment is widely recognized as the primary cause of food contamination. Furthermore, outbreaks of foodborne listeriosis have been associated with contamination of processing facilities by environmental L. monocytogenes [11]. This pathogen in cooling and cutting areas significantly contributes to meat contamination [12].
Human infection with L. monocytogenes can cause a foodborne disease called listeriosis, and this infection is responsible for affecting elderly, immune-compromised individuals, pregnant women, children, and can cause abortions [13]. L. monocytogenes infection occurs after ingestion of food contaminated with this pathogen. The infection is facilitated by proteins encoded by a specific gene cluster. Once in the host, L. monocytogenes escapes from the phagocytic vacuole and initiates multiplication in the host cell cytoplasm. Then, the motility occurs by the induction of actin polymerization at one pole of the bacterial cell. The last step involves motility within cytoplasmic evaginations of the neighboring cells, where the cycle begins again [14]. This intracellular infection/pathogenicity is mediated by some gene products encoded on a six-gene cluster called LIPI-1 (Listeria pathogenicity island), about 9 kb long [15]. This cluster consists of a transcriptional master regulator (prfA); two phospholipases known as plcA and plcB, which are involved in lysing host cell membranes; a hemolysin (Listeriolysin -hlyA) that is necessary for disrupting the host’s phagosome to access the cytoplasm; a metalloprotease that activates the inactive propeptide plcB in the extracellular environment; and the actA gene that encodes the surface protein ActA, which is essential for bacterial motility [16,17]. All these genes and their products are involved in the L. monocytogenes pathogenicity.
When listeriosis occurs, the main set of antibiotics used to treat this illness include trimethoprim with sulfamethoxazole, ampicillin, tetracycline, gentamicin, erythromycin, penicillin, rifampicin, and chloramphenicol [18]. The indiscriminate use of these compounds can force wild-type isolates to develop multiple antibiotic resistance, and its resistance can range from 3 to 13 antibiotics [18]. Listeriosis is a disease that poses a high risk of death among vulnerable individuals. Out of 1,600 people who become ill, 260 die each year (CDC, 2020). The European Union reports a 15.6% fatality rate among reported cases of listeriosis [19].
To minimize the damage caused by L. monocytogenes, various control methods have been developed in the meat processing sector within the industry [20]. It is known that L. monocytogenes develops tolerance to some bactericides such as benzalkonium chloride, alkylbenzyl-dimethyl ammonium chloride, n-alkyldimethylethylbenzyl ammonium chloride, tertiary alkyl-amine, 1,3-propanediamine-N-(3-aminopropyl) N-dodecyl, sodium hypochlorite, and potassium persulphate [20,21,22]. For this reason, alternative methods have been proposed for its control, such as the use of biopolymers with bactericidal effects [23]. In this sense, chitosan is a biopolymer derived from shrimp waste that has been tested as an alternative for the potential development of a product against L. monocytogenes [23].
The aims of this study were to characterize sixteen L. monocytogenes isolates from fresh pork loin and fresh packaged pork loin, identify their serotypes and antibiotic resistance profiles, assess their susceptibility to chitosan as a possible strategy to control L. monocytogenes, and test their pathogenicity in a chicken embryo model.
2. Materials and Methods
2.1. Sampling
Four sampling events, each lasting two months, were conducted at a regulated slaughterhouse south of Sonora, México. Sixteen samples of fresh pork loin were collected in sterile bags, and ten samples of fresh packaged pork loin were collected in their final packaging. Samples were preserved at a low temperature of 4–8 °C and analyzed on the collection day.
2.2. Isolation and Phenotypic Characterization of Listeria monocytogenes
All isolates were obtained from fresh pork loin and packaged fresh pork loin, following the procedure described in Appendix C of NOM 210-SSA-2014, which resembles the method outlined by the International Organization for Standardization in ISO 11290 for L. monocytogenes. Briefly, 25 g of the sample was mixed with 25 mL of phosphate-buffered saline (137 mM NaCl, 10 mM phosphate, 2.7 mM KCl, and pH 7.4) in a sterile blender in aseptic conditions. Then, each 25 mL from each sample was added to 225 mL of Fraser Broth (Biocontrol, Cat. 63017-500) containing Fraser supplements (Sigma-Aldrich, Cat. 90836-10VL; Darmstadt, Germany), and incubated for 48 h at 36 °C. Then the sample was plated on Oxford plates (Condalab, Cat. 1133; Torrejón de Ardoz, Madrid, Spain) with supplements (Oxoid, Cat. SR0140E; Waltham, Massachusetts, USA) and incubated for 48 h at 36 °C. The round grayish colonies surrounded by dark halos were taken to the following tests. The biochemical characterization was carried out according to Momtaz and Yadollahi: Gram staining; catalase and oxidase tests, motility test at 25 °C, acid production from rhamnose and xylose, the β-hemolytic activity on sheep blood agar test, and the CAMP test (Christie, Atkins, Munch-Peterson test) [24]. Staphylococcus aureus ATCC 29213 (SA+) and Rhodococcus equi ATCC 6939 (RE−) were used as controls for the CAMP test. Listeria monocytogenes ATCC 7644 was used as the positive control for all tests. All positive isolates to L. monocytogenes were recovered and stored at − 70 °C with glycerol (15%) until use.
2.3. Genotypic Characterization of Listeria monocytogenes Isolates
2.3.1. DNA Extraction
A colony from a previous culture for each isolate was used to inoculate 5 mL of trypticase soy broth and then incubated at 37°C for 24 h. Subsequently, the cells were harvested by centrifugation at 5000 x g for 3 min, and the cell pellet was used for DNA extraction. Briefly, the cell pellet of each isolate was resuspended in a lysis buffer (0.03 M Tris-HCl, 0.01 M EDTA, and 20 g/L lysozyme) and incubated at 37°C for 30 min. Genomic DNA was extracted with the Qiagen® DNeasy® Blood and Tissue Kit according to the manufacturer’s instructions. The DNA concentration and purity were measured using a Nanodrop 2000c UV–Vis spectrophotometer (ThermoFisher, Inc.;Wilmington, DE, USA).
2.3.2. Detection of Virulence Factors by PCR
The detection of the virulence factors was made by PCR. The components of the reaction were the following: 1× PCR buffer, 2 mM of MgCl2, 0.2μM of dNTPs, 0.2 μM of each oligonucleotide, 1 U of Taq DNA polymerase (Invitrogen™. Cat. No. 18038042), 10 ng of DNA and water until 25uL. All the primers used are listed in Supporting Table S1. A Labnet, MultiGene optiMAX machine (Edison, NJ, USA) was used to run the reactions. The conditions used were an initial denaturation step at 95 °C for 5 min, 30 cycles at 95 °C for 30 s, annealing for 30 s and the melting temperature is indicated in Supporting Table S1 for each primer pair, and extension at 72 °C for 30 s, and a last step at 72 °C for 10 min.
2.3.3. Molecular Serotyping of Wild Isolates of Listeria Monocytogenes
All isolates recovered were serotyped by PCR, according to Doumit et. al. [5]. The set of primers used for serotyping are listed in Table S1. These primer pairs target the three major serovars of L. monocytogenes. The set of lmo0737 amplifies a region of 691 bp of the putative protein gene with an unknown function to detect the serotype 1/2a. The pair ORF2819 amplifies a region of the putative transcriptional regulator generating a 471 bp fragment to detect the serotype 1/2b. For serotype 4b, the set of primers ORF2110 was utilized to detect a fragment of the putative secreted protein, yielding an amplicon of 597 bp. The components of the reaction were the following: 1× PCR buffer, 2.5 mM of MgCl2, 0.8 mM of dNTPs, 0.4 μM of each oligonucleotide, 1 U of Taq DNA polymerase, 10 ng of DNA and water until 12.5 uL. The PCR conditions included an initial denaturation step at 95 °C for 7 min, followed by 30 cycles at 95 °C for 15 s, annealing at 56 °C for 20 s, and extension at 72 °C for 60 s, and a last step at 72 °C for 10 min. PCR products were loaded and electrophoresed in a 1% agarose gel in 0.5 X Trisacetate EDTA buffer, stained with ethidium bromide, and visualized on a Minibis Pro DNR system (Bio-Imaging systems).
2.4. Assessing the Virulence of L. monocytogenes Isolates on Chicken Embryos
2.4.1. Checking for the Dead Embryos
This assay was carried out according to Andersson et al. [25] with modifications. Briefly, this experiment was performed in chicken embryos (Gallus gallus); the eggs were incubated at 37.5 °C with 65% relative moisture for 8-9 days to discard dead embryos.
2.4.2. L. monocytogenes Inoculum
All isolates were grown in Brain Heart Infusion broth for 24 h. After the incubation, 1 mL was taken and centrifuged at 4000 x g for 10 min to collect bacterial cells, which were resuspended in 1 mL of NaCl (0.9 % (w/v)).
2.4.3. Infecting the Chicken Embryos with L. monocytogenes Wild Type Isolates
All the viable embryos selected were injected with 100 µL containing 5x105 CFU. The eggs were perforated carefully with forceps. The infective dose was injected with a Syringe (1 mL, BD Plastipak, no. 300013). Paraffin was applied to the opened egg with cotton, and a small piece of tape was used to cover the opening. The eggs were returned to the incubator. The embryo’s viability was measured every 24h until the end of the experiment (120 h). The experiment was carried out three times with five replicates per treatment. Mortality was calculated as the percentage of the total of embryos inoculated for each bacterial treatment during 5-day incubation.
2.4.4. L. monocytogenes Detection in Chicken Liver Embryos
To verify whether L. monocytogenes had caused an infection, PCR detection was performed in the liver of the chicken embryo. Once the embryos were dead, they were removed from the incubator and analyzed. Each egg was opened on a sterile petri dish. The embryo was separated from the fluids with forceps and washed with 0.9% (w/v) NaCl. The liver was extracted following the instructions of Andersson et al. [25]. The liver was placed in a sterile 1.5 mL Eppendorf tube containing 1 mL of 0.9% (wt/vol) NaCl. Total DNA was extracted with Wizard® Genomic DNA Purification Kit (PROMEGA) following the manufacturer’s instructions for Culture cells and animal tissue. The DNA was quantified using a Nanodrop 2000c UV–Vis spectrophotometer (ThermoFisher, Inc.; Wilmington, DE, USA). The L. monocytogenes detection was carried out for hlyA gene as previously described in a section Molecular detection of virulence factors. Listeria monocytogenes ATCC 15313 and Listeria innocua ATCC 33091 were used as negative infection controls while the strain Listeria monocytogenes 7644 was used as positive infection control.
2.5. Ethical Declaration for the Use of Chicken Embryos
This study was conducted with the approval of the Ethics Committee for Research and Animal Welfare of the Instituto Tecnológico de Sonora (Institutional Approval Number was 2024-09). All necessary precautions were taken to ensure the welfare of the chicken embryos used, in compliance with international and national regulations for the use of animals in scientific research.
2.6. Assessment of the Antibiotic Resistance Profiles of Wild-Type L. monocytogenes Isolates
To confirm susceptibility/resistance to antibiotics an analysis of minimum inhibitory concentration (MIC, μg/mL) was analyzed on 68 Microscan GN systems according to the Clinical and Laboratory Standard Institute [26]. The inhibition was evaluated based on the breakpoints criteria established in the CLSI guidelines for Staphylococcus aureus because no resistance criteria exist for Listeria monocytogenes susceptibility testing in the CLSI guidelines. Twenty-one antibiotics were tested against all isolates AMC, amoxicillin-clavulanic acid; SAM, ampicillin-sulbactam; AMP, ampicillin; CRO, ceftriaxone; CIP, ciprofloxacin; CLI, clindamycin; DAP, daptomycin; ERY, erythromycin; FOF, fosfomycin; GEN, gentamicin; LVX, levofloxacin; LZD, linezolid; MXF, moxifloxacin; NIT, nitrofurantoin; OXA, oxacillin; PEN, penicillin; RIF, rifampin; SYN, synercid; TET, tetracycline; SXT, trimethoprim-sulfamethoxazole; and VAN, vancomycin. The multiple antibiotic resistance (MAR) index was calculated according to Singh et al. [27]. MAR index MARI = number of resistance antibiotics/total number of antibiotics tested. The strains that resisted at least three antibiotic compounds (>0.143) were considered multiple antibiotic resistance (MAR).
2.7. Antimicrobial Assays with Shrimp Chitosan Against L. monocytogenes Isolates
2.7.1. Chitosan Preparation
Chitosan was obtained following the process of Rodríguez-Núñez et al. [28]. Chitin extraction was carried out on a pilot scale using lactic fermentation, following the procedure outlined by Bueno-Solano et al. [29]. To obtain chitosan with a low molecular weight, the method described by Weska et al. [30] was employed with certain modifications. Initially, the chitin was subjected to a protein removal process by immersing it in a 4.5% (w/v) NaOH solution at 65 °C for 4 h. Subsequently, the resulting solid precipitate underwent a mineral removal step by immersion in a 3.6% (w/w) HCl solution at room temperature for 4 h. The final stage involved an alkaline deacetylation process using a 45% (w/v) NaOH solution at 120 °C for 2 h, followed by thorough rinsing with water and drying at 40 °C for 12 hours. Prior to utilization, the chitosan was finely powdered through milling to achieve a particle size of 180 μm.
2.7.2. Chitosan Antimicrobial Assay
To establish the chitosan concentration and exposure time to inhibit the L. monocytogenes growth, a preliminary experiment was conducted using three concentrations and three exposure times. The concentrations of 0.1, 0.175, and 0.25 % (w/v) diluted in 1% acetic acid were evaluated at 7.5, 26, and 45 min exposure times. The treatments were incubated at 37°C for 45 min, harvesting at 7.5, 26, and 45 min and serial dilutions were made to count the CFU/mL for each treatment. Once the appropriate concentration (0.25%) and time (45 min) were selected and evaluated with the wildtype strains of L. monocytogenes (Figure S1). To discard the effect of acetic acid on bacterial development and only attribute the antimicrobial effect to chitosan, a treatment with only acetic acid was evaluated (1% v/v). Before carrying out the test, each strain was thawed and grown in 5 mL of TSB for 24 h at 37°C. After that, 1 mL was taken and inoculated in a flask with 99 mL of TSB, shaken, and incubated for 7 hours at 37°C. Subsequently, from this 7-hour culture, 1 μL of the culture (approx. 1x106 cells) was taken and inoculated into a 1.5 mL tube containing 999 μL of each treatment. The cell concentrations were expressed in Log of CFU/mL. The cell counts obtained after each treatment were analyzed by one-way ANOVA using STATGRAPHICS Plus 5.0 applying a multiple comparison procedure by the Tukey method (HSD) at 95% confidence.
3. Results
3.1. Identification of L. monocytogenes Isolates
Here, Listeria monocytogenes was detected in sixteen out of 26 samples (Table 1). All isolates obtained showed biochemical behavior as L. monocytogenes and were confirmed by PCR using the hlyA gene, a common gene used for L. monocytogenes confirmation [31]. This finding confirms the presence of the LLO-encoding gene, a crucial toxin associated with the pathogenicity of L. monocytogenes, which indicates the potential virulence of these isolates [31]. Indeed, the hlyA gene plays a crucial role in identifying L. monocytogenes, making it the most frequently selected target among other virulence genes for PCR detection of this pathogen. Molecular detection is less time-consuming than biochemical methods, increasing accuracy and effectiveness [31].
3.2. Prevalence of L. monocytogenes in Fresh Pork Loin and Fresh Packaged Pork Loin
The prevalence is not the central focus of this work, but it is worth mentioning. Sixteen samples out of 26 from fresh pork loin and fresh packaged pork loin were found contaminated with L. monocytogenes, giving 61.5 % of the total prevalence (Table 1). In this case, fresh packaged pork loin samples were highly prevalent (70 %), with seven contaminated samples giving 7 confirmed isolates (Table 1). Of sixteen fresh pork loin samples, 9 were contaminated, and a prevalence of 56.3% was observed, leading to the isolation of nine L. monocytogenes strains (Table 1).
Food safety is a significant global concern. Contamination of food, particularly by L. monocytogenes, can result in serious consequences. L. monocytogenes contamination not only causes economic losses for the food industry but also poses severe health risks, such as miscarriage and food poisoning. Given the persistent nature of this pathogen, it is essential for the food industry to implement regular cleaning and disinfection practices to ensure safety [32]. In Colombian swine, the prevalence of L. monocytogenes in fresh pork meat was reported to be 33.9%, less than this work [33]. In other work about raw pork meat from processing plants, the L. monocytogenes is similar to this work, with a 37 % prevalence reported [34]. Sixteen isolates were obtained from both fresh pork loin and fresh packaged pork loin, confirming the presence of L. monocytogenes in fresh pork meat. The contamination of L. monocytogenes can occur at various stages of the pork production chain, primarily due to inadequate cleaning and disinfection processes. This allows the pathogen to become established in production equipment [12]. These findings of L. monocytogenes levels indicate that the current sanitizing techniques and cleaning agents are inadequate for eliminating this pathogen, emphasizing the need for a more thorough sanitization process.
3.3. Serotype Identification of L. monocytogenes Isolates
This study focused solely on serotypes 1/2a, 1/2b, and 4b, previously reported as the most frequent in human listeriosis cases [35].
Here, serotype 1/2b was the most predominant, with 68.7 % in both types of samples (Table 1). Two isolates were classified as 1/2a, and one isolate was classified as serotype 4b (Table 1). Two isolates could not be classified using the method employed (Table 1). The incidence of listeriosis has been reported to range between 1 and 10 cases per million population annually, with higher rates observed in certain countries [35]. Listeriosis is particularly severe in individuals with weakened immune systems, pregnant women, and newborns, with an estimated mortality rate of 20-30% among clinical cases. To prevent listeriosis, efforts should focus on controlling food production and handling, as well as educating consumers, especially those in high-risk groups, about the risks of exposure and the precautions they should take. Research has examined the prevalence of L. monocytogenes serotypes in pork loins and surfaces throughout the production chain, revealing the presence of serotypes 1/2a (19%) and 1/2b (80.95%) [32]. In pork meat, the predominant serotypes are typically 1/2a and 1/2c, followed by 1/2b [36]. Although serotype 4b has also been isolated from pork products (Prencipe et al., 2012; Zuber et al., 2019), it is not the most commonly occurring serotype. One notable study found the serotypes in pork loins were 1/2a, 1/2b, and 4b, which together represented over 90% of the isolates (Hof et al., 1997). L. monocytogenes serotypes in pork loins are a significant concern throughout the production chain. Serotypes 1/2a, 1/2b, and 4b have been identified, emphasizing the need for targeted interventions to address specific strains of concern. Ongoing surveillance, continuous improvement of food safety practices, and consumer awareness are essential to reduce the burden of listeriosis associated with pork products.
3.4. Detection of Virulence Factors in L. monocytogenes Isolates
The most common virulence factors were detected by PCR to determine the virulence profile associated with L. monocytogenes pathogenicity. Other studies had reported similar findings, documenting variability in the virulence genes detection [37,38]. The L. monocytogenes isolates showed different virulence genes presence, most isolates possess all genes evaluated (Table 2). Some of them lack genes; isolates 3.1 and 3.2 lack actA and plcA genes, and isolate 1.2 lacks InlA, InlB, and InlJ. The plcA gene is not present in isolate 7.2, and the InlB is not present in isolate 10.1 (Table 2).
The products of the virulence genes are necessary for L. monocytogenes infection, prfA is the master regulator of most of these genes. The phospholipases (plcA and plcB) are required to lyse the host cell membrane and to escape from host cell’s phagosomes L. monocytogenes expresses the pore-forming sulfhydryl-activate listeriolysin (LLO product of the gene hlyA). The actA gene produces an actin assembly surface protein that is responsible for the movement within host cells [15]. The internalins (lnlA, InlB, InlC, InlJ) are proteins that are found in bacterial membrane involved in the invasion and adhesion to different types of eukaryotic cells [39].
The iap gene is a virulence factor needed for the invasion of L. monocytogenes into host cell, and possesses a murein hydrolase activity involved in bacterial cell division [40]. Most wild L. monocytogenes isolates evaluated in this work possess all the genes necessary for proper infection of the eukaryotic cell. The absence of the gene actA had been reported previously in isolates from meat products in Poland by Kawacka et al. [41], where some isolates showed a rate of 10% to 78% of actA presence. Similar results were obtained in Romania from ready to eat products, the actA gene was not detected in isolates from samples of years 2019 and 2020 [38]. The internalins in some isolates are absent, a study on genome analysis showed differences in the presence of internalin genes.
Here, the authors mention that L. monocytogenes can have different virulence profiles, including the internalin genes [4]. In this work, the gene plcA was absent in three isolates, by Coroneo et al. [37], where the plcA gene was less predominant in 20% of the isolates evaluated. Here, we highlight some genotypic variations according to the presence or absence of virulence factors.
The absence of specific genes could indicate a reduction in virulence; however, in this case, the only isolate that showed reduced virulence was L. monocytogenes 1.2, as indicated by the results in chicken embryos (shown below). The reduction in its virulence could be attributed to the lack of InlA, InlB, and InlJ genes. Most of the isolates obtained in this study (11 isolates: 7 (1/2b), 2 (1/2a), and 2 ND) possess genes that are involved in the production of virulence proteins necessary for causing infections in eukaryotic organisms. This presents a potential risk for individuals who may come into contact with contaminated meat. Additionally, understanding the presence of these genes could be beneficial for several purposes: developing vaccines against these bacteria, creating new antibacterial drugs that target these genes, and implementing strategies for timely detection in food and clinical samples. This information may also aid in the quicker and more accurate identification of listeriosis outbreaks.
3.5. Assessing the Virulence of L. monocytogenes Isolates on Chicken Embryos
Similar survival patterns were observed when comparing the infectivity of genetically different L. monocytogenes strains in chicken embryos and mice. Specifically, strains that exhibited lower infectivity in the chicken embryo model also showed reduced infectivity in the mouse model. The chicken embryo assay is a reliable, viable, and well-described method for evaluating bacterial infection in a eucaryotic model [25]. All isolates were evaluated in chicken embryos to examine whether they could cause an infection.
The negative controls CTL (NaCl 0.9%), L. monocytogenes ATCC 15313, and L. innocua ATCC 33091 did not affect the viability of chicken embryos (Figure 1). Listeria monocytogenes ATCC 7644 triggered the mortality of chicken embryos 48 h after infection.
In this experiment, we highlight that the isolates from fresh packaged pork loin were more aggressive than those obtained from fresh pork loin, reaching 100 % mortality 48 h post-infection. Five (2.3, 3.1, 3.2, 3.3, and 42.1) of the seven isolates from fresh packaged pork loin showed 100% mortality at 48 h post-infection, and the remaining two (2.1 and 42.2) reached 100% at 72 h post-infection (Figure 1). In fresh pork loin, 2 isolates (1.1 and 10.1) showed 100% mortality 48 h after infection. Three isolates (7.1, 7.2, z44) showed 100% mortality up to 72 h, one (7.3) at 96 h, and two (1.3 and 10.2) at 120 h. One isolate (1.2) showed only 80% mortality, leaving 20% viable embryos (Figure 1). To confirm the infection in a chicken embryo, DNA was extracted from the livers of infected embryos. The hylA gene was used to detect L. monocytogenes by PCR. In embryos treated with saline solution (NaCl 0.9 %), L. innocua ATCC 33091, and L. monocytogenes ATCC 15313, the hlyA gene was not detected (Table S2).
L. monocytogenes wild-type isolates showed differences in the virulence pattern. Strains from fresh pork loin were less aggressive than those from packaged pork loin, and most killed the embryos at 48 h. Unlike fresh pork loin, Listeria strains from packaged pork loin were under cold stress. It is documented that the SigB protein can mediate the survival of L. monocytogenes under a wide range of lethal stresses along the food production chain, including low-temperature stress [42]. As part of the operon sigma B (rsb genes), these proteins can sense environmental stress conditions and regulate the SigB signaling pathway. Under adaptive stress conditions, the SigB protein can activate the PrfA transcription factor, activating virulence factors [42]. This cold stress could trigger the aggressiveness of L. monocytogenes isolates from freshly packaged pork loin, killing the embryos in the first 48 h, as observed previously [43,44].
Regarding the virulence factors, these results show that the absence of actA and plcA genes is not a determinant for virulence behavior in the eukaryotic model. However, the absence of InlA, InlB, InlJ may lead to an attenuation of virulence. It has been shown that these proteins are necessary for cellular internalization in the host. As previously demonstrated through mutation assays targeting the internalin A gene, the internalization pathway can be affected, leading to an attenuation of virulence, although it does not completely inhibit it [45]. Possibly, this infection decrease is caused by the attenuated virulence. The virulence shown by L. monocytogenes wild-type isolates suggests that these isolates can effectively evade cellular barriers and reach the liver in chicken embryos, demonstrating their pathogenic nature and potential risk to people who may come in contact with this strain through contaminated foods [44].
In the case of Listeria monocytogenes ATCC 15313, was unable to infect the chicken embryos nor infect a murine model [46]. Listeria monocytogenes ATCC 15313 is an avirulent strain that does not produce listeriolysin O (LLO), a well-established main virulence factor of L. monocytogenes [47,48], and the hlyA gene, which encodes this factor, is absent in this strain, resulting in its avirulent behavior [46]. The hlyA gene was used in PCR to detect the translocation on chicken embryos post-infection as the main virulence factor; all L. monocytogenes strains were detected except Listeria innocua ATCC 33091 and L. monocytogenes ATCC 15313 in liver from chicken embryos.
The results indicated variations in virulence among different isolates, with those derived from packaged pork loin showing greater aggressiveness and faster mortality rates in embryos. This study underscores the significance of considering how cold stress affects the virulence of L. monocytogenes in food products. It also emphasizes the need for strong food safety measures, especially for refrigerated and processed foods.
Antibiotic resistance of wild-type Listeria monocytogenes isolates. The main concern regarding foodborne pathogens is antibiotic resistance. In cases of listeriosis, penicillin, ampicillin, and gentamicin are the primary antibiotics employed for treatment [49]. L. monocytogenes wild-type isolates showed 100% susceptibility to amoxicillin-clavulanic acid; ampicillin-sulbactam, ampicillin, erythromycin, fosfomycin, gentamicin, moxifloxacin, rifampin, trimethoprim-sulfamethoxazole and vancomycin (Table 3). Here, 100% of isolates were resistant to ceftriaxone, clindamycin, daptomycin, and oxacillin, and 93.3 % were resistant to nitrofurantoin. 53.3% of the isolates showed resistance to ciprofloxacin. Moreover, 20% of the isolates resisted levofloxacin, linezolid, and synercid (quinupristin/dalfopristin) (Table 3). In this research, 13.3% of the isolates resisted penicillin, while only 6.6% resisted tetracycline (Table 3). L. monocytogenes has developed resistance to various antimicrobial compounds.
In bacteria, evolutionary mechanisms can develop that allow them to resist the harmful effects of antibiotics. Among these mechanisms are three very common ones that confer resistance to several groups of antibiotics; although specific for each type of compound, their function is similar: i) efflux pumps, ii) modifications in the ribosome binding sites, and iii) enzymatic inactivation of the antibiotic compound. For tetracycline [50], lincosamides [51], oxazolidinones [52], and streptogramins [53], resistance mechanisms like efflux pumps, ribosomal modifications, and enzymatic inactivation have been described. For β-lactam antibiotics, bacteria can develop another mechanism like membrane modification and changing the cell wall structure [54]. For lipopeptides like daptomycin, microorganisms modified some regulatory networks, cell wall structures, and efflux pumps [55]. For fluoroquinolones, although efflux pumps and inactivation enzymes, bacteria require other modifications that confer resistance, like mutations in the topoisomerase gene [54]. For nitrofurantoin, a specific mutation is required, this is focused on nitroreductase genes avoiding the nitrofurantoin activation [56].
The resistance of Listeria monocytogenes to β-lactam [57], tetracycline [58], lincosamides [59], fluoroquinolones [60], oxazolidinones [59], lipopeptides (daptomycin) [55], streptogramins (Synercid) [61] and nitrofurantoin [62] were previously documented.
Isolate 1.2 showed the highest Multiple Antibiotic Resistance Index (MARI) at 0.476, followed by isolate 2.1 at 0.429, and isolate z44 at 0.381. The lowest value shown for the MAR index was 0.238, which was present in most isolates (Table 4). A reference parameter recently used to assess antibiotic resistance is the MAR Index; in this study, all strains recorded values >0.143 were considered multiple antibiotic-resistant strains. This resistance might have originated due to prolonged exposure to sublethal concentrations of antimicrobial compounds and sanitizers used in food processing facilities, which create stressful conditions and foster the development of antibiotic resistance [63]. MAR isolates might also arise from the indiscriminate use of antimicrobials in pork production. Furthermore, if these MAR isolates contaminate the food processing chain, they can be spread to humans through contaminated food and potentially cause a disease [64].
The emergence of multi-resistant bacteria is a problem of global concern. The impact on human health is significant, particularly due to the lack of effective antibiotics, which can increase mortality rates. Infections will be more difficult to treat, leading to higher economic costs due to prolonged illnesses, which may burden hospitals. These infections could cause a loss of economic productivity in society, with greater effects in developing countries or countries with fewer economic resources. Additionally, these bacteria would heighten the risks associated with medical procedures such as transplants, chemotherapy, and major surgeries because these procedures rely on effective antibiotics to prevent infections. This emergence of multidrug-resistant bacteria in recent years is attributed to the overuse of antimicrobials in humans, agriculture, and animal husbandry. There is a need to raise awareness in the population about the responsible use of antibiotics and to implement monitoring systems to detect the spread of multidrug-resistant bacteria.
Chitosan as an alternative to use against L. monocytogenes in the food industry. Chitosan is a biodegradable and non-toxic biopolymer with antimicrobial and antioxidant properties. There has been an increase in chitosan applications such as wastewater treatment, agriculture, biomedicine, pharmaceuticals, cosmetics, and the food industry, generating a positive impact on the environment by contributing to the reduction of pollution by replacing the indiscriminate use of chemicals [65].
Chitosan is a polysaccharide extracted from chitin that can exert antimicrobial activity against a broad range of foodborne pathogens, and its function depends on the source of chitosan and factors like molecular weight, particle size, pH, temperature, salinity, divalent cations, chitosan solvent, and suspended medium [66]. The chitosan used here had a molecular weight of 114.83 kDa, viscosity of 427.83 cp, deacetylation’s degree of 92.33% and density of 868.34 kg/m3. As reported in other works, a 0.25% of chitosan was used and 45 minutes exposition for elimination of L. monocytogenes (Figure S1) [67,68]. A concentration of 0.25 % at 45 min could inhibit the growth of most isolates. Particularly, the isolate 10.1 was not affected by the polymer, this isolate was evaluated six times to avoid mistakes, and the results were always consistent showing resistance against chitosan (Table 5). Acetic acid did not impact the growth of L. monocytogenes, therefore the observed inhibition is associated with the chitosan used (Table 5). It has been reported that concentrations of 0.02 to 0.5 % of chitosan can inhibit L. monocytogenes [67,68]. In this study, a concentration of 0.25% applied for 45 minutes successfully inhibited L. monocytogenes, decreasing its counts from 6 to 0 Log CFU/mL. Additionally, concentrations of 0.15% have also proven effective against both L. monocytogenes and Salmonella Typhimurium. [69]. Regarding the acetic acid effect, in this work, the concentrations of acetic acid had no effect on L. monocytogenes Log CFU/mL, contrasting with previous work by Ibañez-Peinado, et al. [69], where the acetic acid had a reducing effect on the initial Log CFU/mL used in their experiments, and their findings revealed that higher pH values of the chitosan solutions are less effective than low pH values. Most studies on chitosan against foodborne pathogens address its antimicrobial effect, but the mechanisms by which microorganisms develop resistance to this polymer remain unclear [70]. A study carried out in S. aureus, a Gram-positive bacterium, documented that modifications to cell surface properties can reduce the negative charge of the cell wall and cell membrane, causing a reduced chitosan binding; this suggests that modifications of the cell wall and cell membranes confer resistance to chitosan [70]. Possibly, L. monocytogenes developed a similar mechanism explaining the resistance to chitosan observed in this study.
4. Conclusions
Sixteen isolates were evaluated and characterized as Listeria monocytogenes obtained from pork meat. The prevalence varied between sample types, and serotypes 1/2a and 1/2b were frequent in both types of samples. Regarding virulence factors, it was found that some isolates lacked certain genes involved in the infection process. However, the absence of some main genes can affect pathogenic behavior in the eukaryotic model, as they successfully infected chicken embryos, causing death or attenuating virulence.
The antibiotic resistance shown by the microorganisms studied here is a real concern. One isolate was found to be resistant to as many as ten antibiotics, which poses a latent risk, especially considering these microorganisms were isolated from food and are in close contact with people. Chitosan has been used as a decontamination strategy, but surprisingly, one isolate exhibited resistance to chitosan. This resistance is possibly related to modifications in the cell wall and cell membrane, as demonstrated in S. aureus. This could provide insight into Listeria monocytogenes evolution and highlight the need for caution in using chitosan as an antimicrobial compound, particularly against its indiscriminate application.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Evaluation of chitosan at different exposure times and chitosan concentrations on Listeria monocytogenes ATCC 15313.; Table S1: Oligonucleotides were used for molecular analysis.; Table S2. PCR detection of Listeria monocytogenes on chicken embryos in the virulence assay.
Author Contributions
Conceptualization, A.M.F.L. and E.U.C.S.; methodology, MTGO, NGRM, CEAL, RRR and AGVL; investigation, A.M.F.L.; resources, A.M.F.L.; writing—original draft preparation, A.M.F.L.; writing—review and editing, A.M.F.L. and E.U.C.S; supervision, A.M.F.L. and E.U.C.S.; project administration, A.M.F.L. and E.U.C.S.; funding acquisition, A.M.F.L. and E.U.C.S.
Funding
This research received no external funding.
Acknowledgments
The authors acknowledge the support for this project from the ITSON-PROFAPI program (2023–024).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Carpentier, B.; Cerf, O. Review-Persistence of Listeria Monocytogenes in Food Industry Equipment and Premises. International Journal of Food Microbiology 2011, 145, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Schoder, D.; Pelz, A.; Paulsen, P. Transmission Scenarios of Listeria Monocytogenes on Small Ruminant on-Farm Dairies. Foods 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Schoder, D.; Guldimann, C.; Märtlbauer, E. Asymptomatic Carriage of Listeria Monocytogenes by Animals and Humans and Its Impact on the Food Chain. Foods 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Muchaamba, F.; Eshwar, A.K.; Stevens, M.J.A.; Stephan, R.; Tasara, T. Different Shades of Listeria Monocytogenes: Strain, Serotype, and Lineage-Based Variability in Virulence and Stress Tolerance Profiles. Frontiers in Microbiology 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Doumith, M.; Buchrieser, C.; Glaser, P.; Jacquet, C.; Martin, P. Differentiation of the Major Listeria Monocytogenes Serovars by Multiplex PCR. Journal of Clinical Microbiology 2004, 42, 3819–3822. [Google Scholar] [CrossRef] [PubMed]
- Ducey, Thomas; Usgaard Thomas; Dunn Katherine A. ; Bielawski Joseph P.; Ward, T.J. Multilocus Genotyping Assays for Single Nucleotide Polymorphism-Based Subtyping of Listeria Monocytogenes Isolates. Applied and Environmental Microbiology 2008, 74, 7629–7642. [Google Scholar] [CrossRef] [PubMed]
- Orsi, R.H.; Bakker, H.C. den; Wiedmann, M. Listeria Monocytogenes Lineages: Genomics, Evolution, Ecology, and Phenotypic Characteristics. International Journal of Medical Microbiology 2011, 301, 79–96. [Google Scholar] [CrossRef] [PubMed]
- Parra-Flores, J.; Holý, O.; Bustamante, F.; Lepuschitz, S.; Pietzka, A.; Contreras-Fernández, A.; Castillo, C.; Ovalle, C.; Alarcón-Lavín, M.P.; Cruz-Córdova, A.; et al. Virulence and Antibiotic Resistance Genes in Listeria Monocytogenes Strains Isolated from Ready-to-Eat Foods in Chile. Front. Microbiol. 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Food and Drug Administration Listeria (Listeriosis) Available online: https://www.fda.gov/food/foodborne-pathogens/listeria-listeriosis#:~:text=Past%20listeriosis%20outbreaks%20in%20the,found%20in%20raw%20pet%20food.
- Bridges, D.F.; Bilbao-Sainz, C.; Powell-Palm, M.J.; Williams, T.; Wood, D.; Sinrod, A.J.G.; Ukpai, G.; McHugh, T.H.; Rubinsky, B.; Wu, V.C.H. Viability of and Salmonella Typhimurium after Isochoric Freezing. Journal of Food Safety 2020, 40, e12840. [Google Scholar] [CrossRef]
- Demaître, N.; Rasschaert, G.; De Zutter, L.; Geeraerd, A.; De Reu, K. Genetic Listeria Monocytogenes Types in the Pork Processing Plant Environment: From Occasional Introduction to Plausible Persistence in Harborage Sites. Pathogens 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Thévenot, D.; Dernburg, A.; Vernozy-Rozand, C. An Updated Review of Listeria Monocytogenes in the Pork Meat Industry and Its Products. Journal of Applied Microbiology 2006, 101, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Pricope, L.; Nicolau, A.; Wagner, M.; Rychli, K. The Effect of Sublethal Concentrations of Benzalkonium Chloride on Invasiveness and Intracellular Proliferation of Listeria Monocytogenes. Food Control 2013, 31, 230–235. [Google Scholar] [CrossRef]
- Quereda, J.J.; Morón-García, A.; Palacios-Gorba, C.; Dessaux, C.; García-del Portillo, F.; Pucciarelli, M.G.; Ortega, A.D. Pathogenicity and Virulence of Listeria Monocytogenes: A Trip from Environmental to Medical Microbiology. Virulence 2021, 12, 2509–2545. [Google Scholar] [CrossRef] [PubMed]
- Kayode, A.J.; Igbinosa, E.O.; Okoh, A.I. Overview of Listeriosis in the Southern African Hemisphere—Review. Journal of Food Safety 2020, 40, e12732. [Google Scholar] [CrossRef]
- Vázquez-Boland, J.A.; Domínguez-Bernal, G.; González-Zorn, B.; Kreft, J.; Goebel, W. Pathogenicity Islands and Virulence Evolution in Listeria. Microbes and Infection 2001, 3, 571–584. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Boland, J.A. ; Kuhn Michael; Berche Patrick; Chakraborty Trinad; Domı́nguez-Bernal Gustavo; Goebel Werner; González-Zorn Bruno; Wehland Jürgen; Kreft Jürgen Listeria Pathogenesis and Molecular Virulence Determinants. Clinical Microbiology Reviews 2001, 14, 584–640. [Google Scholar] [CrossRef] [PubMed]
- Ntshanka, Z.; Ekundayo, T.C.; du Plessis, E.M.; Korsten, L.; Okoh, A.I. Occurrence and Molecular Characterization of Multidrug-Resistant Vegetable-Borne Listeria Monocytogenes Isolates. Antibiotics 2022, 11, 1353–1353. [Google Scholar] [CrossRef] [PubMed]
- EFSA The European Union One Health 2018 Zoonoses Report. 2018.
- Lundén, J.; Autio, T.; Markkula, A.; Hellström, S.; Korkeala, H. Adaptive and Cross-Adaptive Responses of Persistent and Non-Persistent Listeria Monocytogenes Strains to Disinfectants. International Journal of Food Microbiology 2003, 82, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Folsom, J.P.; Frank, J.F. Chlorine Resistance of Listeria Monocytogenes Biofilms and Relationship to Subtype, Cell Density, and Planktonic Cell Chlorine Resistance. Journal of Food Protection 2006, 69, 1292–1296. [Google Scholar] [CrossRef] [PubMed]
- Møretrø, T.; Schirmer, B.C.T.; Heir, E.; Fagerlund, A.; Hjemli, P.; Langsrud, S. Tolerance to Quaternary Ammonium Compound Disinfectants May Enhance Growth of Listeria Monocytogenes in the Food Industry. International Journal of Food Microbiology 2017, 241, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Ardean, C.; Davidescu, C.M.; Nemeş, N.S.; Negrea, A.; Ciopec, M.; Duteanu, N.; Negrea, P.; Duda-seiman, D.; Musta, V. Factors Influencing the Antibacterial Activity of Chitosan and Chitosan Modified by Functionalization. International Journal of Molecular Sciences 2021, Vol. 22, Page 7449 2021, 22, 7449–7449. [Google Scholar] [CrossRef] [PubMed]
- Momtaz, H.; Yadollahi, S. Molecular Characterization of Listeria Monocytogenes Isolated from Fresh Seafood Samples in Iran. Diagnostic Pathology 2013, 8, 149. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Mortality rates in chicken embryos post-inoculation with wild-type Listeria monocytogenes isolates over 120 h. CTL, control solution with NaCl 0.9%. Lm15313, negative infection control Listeria monocytogenes ATCC 15313. Lin 33091, negative infection control Listeria innocua ATCC 33091. Lm 7644, positive infection control Listeria monocytogenes ATCC 7644. Graphs with filled triangles and dotted lines belong to the control strains. Graphs with filled circles belong to isolates from raw pork loin. Graphs with filled circles and dotted lines belong to isolates from raw package pork loin.
Figure 1.
Mortality rates in chicken embryos post-inoculation with wild-type Listeria monocytogenes isolates over 120 h. CTL, control solution with NaCl 0.9%. Lm15313, negative infection control Listeria monocytogenes ATCC 15313. Lin 33091, negative infection control Listeria innocua ATCC 33091. Lm 7644, positive infection control Listeria monocytogenes ATCC 7644. Graphs with filled triangles and dotted lines belong to the control strains. Graphs with filled circles belong to isolates from raw pork loin. Graphs with filled circles and dotted lines belong to isolates from raw package pork loin.
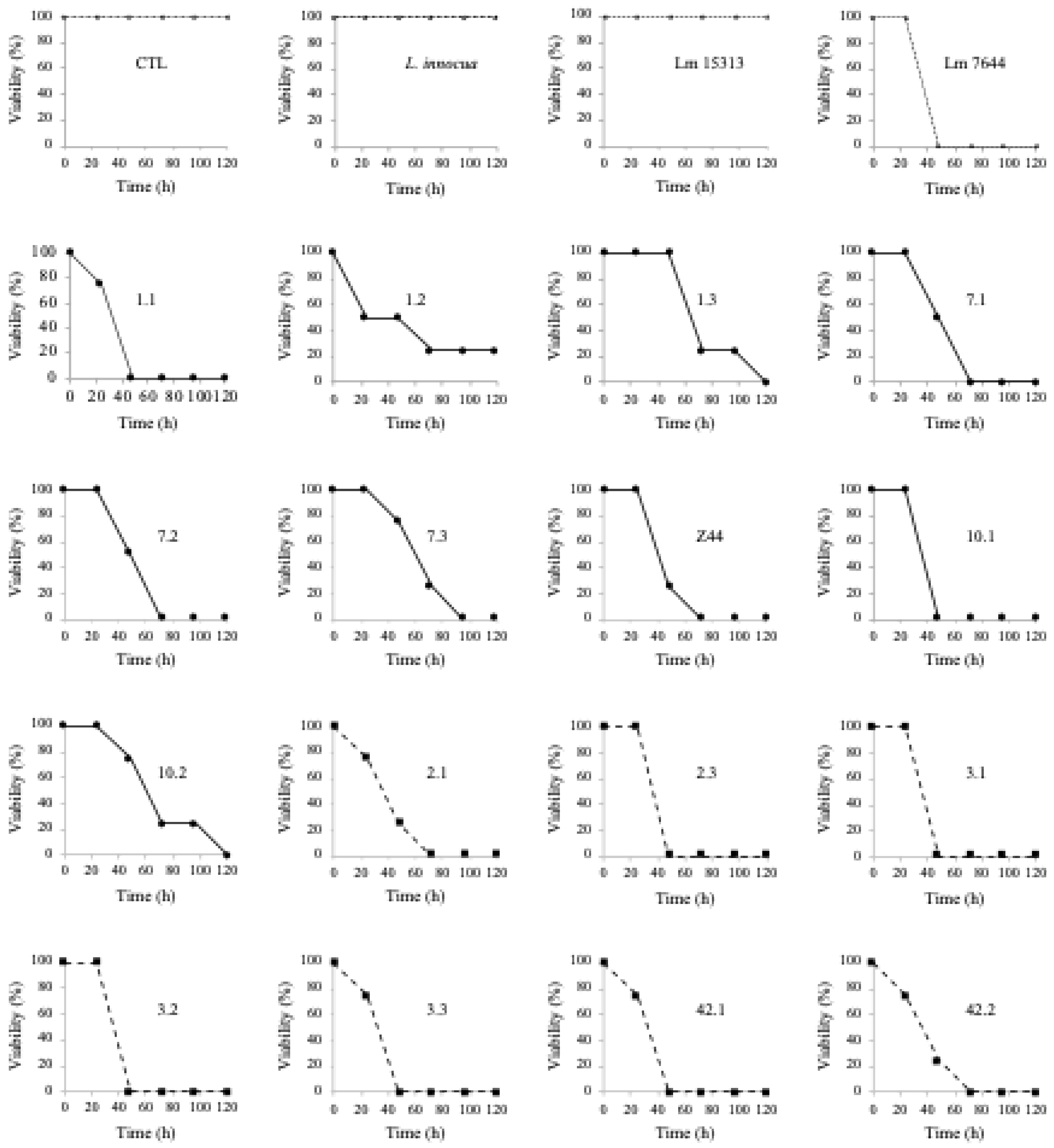

Table 1.
Prevalence and frequency of L. monocytogenes serotypes in raw pork loin and packaged raw pork loin.
Table 1.
Prevalence and frequency of L. monocytogenes serotypes in raw pork loin and packaged raw pork loin.
| Serotypes | |||||||
|---|---|---|---|---|---|---|---|
| Source | Samples | Prevalence | L. monocytogenes isolates obtained | 1/2a | 1/2b | 4b | ND |
| Raw pork loin | 16 | 56.3 % | 9 | 1 (11.1 %) | 7 (77.8 %) | ̶̶̶ | 1 (11.1 %) |
| Raw packaged pork loin | 10 | 70 % | 7 | 1 (14.3 %) | 4 (57.1 %) | 1 (14.3 %) | 1 (14.3 %) |
| TOTAL | 26 | 61.5 % | 16 | 2 (12.5%) | 11 (68.75 %) | 1 (6.25 %) | 2 (12.5 %) |
ND means serotype not defined.
Table 2.
Virulence profiles and their respective serotypes of L. monocytogenes isolates from fresh pork loin and packaged fresh pork loin.
Table 2.
Virulence profiles and their respective serotypes of L. monocytogenes isolates from fresh pork loin and packaged fresh pork loin.
| Source | Bacterial isolate | Virulence factors | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| actA | hlyA | lnlA | InlB | InlC | InlJ | Iap | plcA | plcB | prfA | Serotype | |||
| Raw pork loin | L. monocytogenes ATCC 7644* | + | + | + | + | + | + | + | + | + | + | 1/2a | |
| Lm 1.1 | + | + | + | + | + | + | + | + | + | + | 1/2b | ||
| Lm 1.2 | + | + | ND | ND | + | ND | + | + | + | + | 1/2b | ||
| Lm 1.3 | + | + | + | + | + | + | + | + | + | + | 1/2b | ||
| Lm 7.1 | + | + | + | + | + | + | + | + | + | + | 1/2b | ||
| Lm 7.2 | + | + | + | + | + | + | + | ND | + | + | 1/2b | ||
| Lm 7.3 | + | + | + | + | + | + | + | + | + | + | 1/2b | ||
| Lm Z44 | + | + | + | + | + | + | + | + | + | + | 1/2a | ||
| Lm 10.1 | + | + | + | ND | + | + | + | + | + | + | 1/2b | ||
| Lm 10.2 | + | + | + | + | + | + | + | + | + | + | ND | ||
| Raw packaged pork loin | Lm 2.1 | + | + | + | + | + | + | + | + | + | + | 1/2a | |
| Lm 2.3 | + | + | + | + | + | + | + | + | + | + | 1/2b | ||
| Lm 3.1 | ND | + | + | + | + | + | + | ND | + | + | 4b | ||
| Lm 3.2 | ND | + | + | + | + | + | + | ND | + | + | 1/2b | ||
| Lm 3.3 | + | + | + | + | + | + | + | + | + | + | 1/2b | ||
| Lm 42.1 | + | + | + | + | + | + | + | + | + | + | ND | ||
| Lm 42.2 | + | + | + | + | + | + | + | + | + | + | 1/2b | ||
*L. monocytogenes ATCC 7644 was used as a positive control in PCR assays. ND signifies not detected.
Table 3.
Antibiotic compounds were evaluated against Listeria monocytogenes isolates from pork loin samples.
Table 3.
Antibiotic compounds were evaluated against Listeria monocytogenes isolates from pork loin samples.
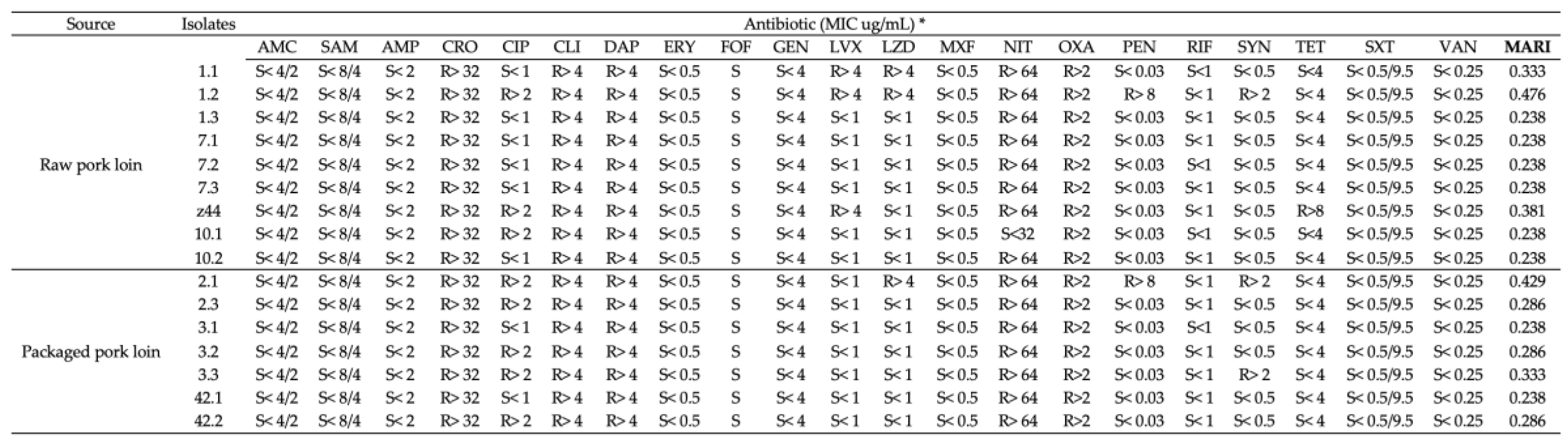 |
*33 Microscan GP systems; MIC = minimum inhibitory concentration according to CLSI criteria. AMC, amoxicillin-clavulanic acid; SAM, ampicillin-sulbactam; AMP, ampicillin; CRO, ceftriaxone; CIP, ciprofloxacin; CLI, clindamycin; DAP, daptomycin; ERY, erythromycin; FOF, fosfomycin; GEN, gentamicin; LVX, levofloxacin; LZD, linezolid; MXF, moxifloxacin; NIT, nitrofurantoin; OXA, oxacillin; PEN, penicillin; RIF, rifampin; SYN, synercid; TET, tetracycline; SXT, trimethoprim-sulfamethoxazole; VAN, vancomycin. R indicates resistance and S indicates sensitivity. MARI, multiple antibiotic resistance index.
Table 5.
Antimicrobial effect of shrimp chitosan against L. monocytogenes isolates from raw and packaged pork loin.
Table 5.
Antimicrobial effect of shrimp chitosan against L. monocytogenes isolates from raw and packaged pork loin.
| Log (CFU/mL) | |||||
|---|---|---|---|---|---|
| Isolate | Initial Inoculum | Control† | Chitosan (0.25%)† | Acetic acid (1%)† | |
| Raw pork loin | Lm 1.1 | 6.36ª | 6.83ª | 0.00b | 6.54ª |
| Lm 1.2 | 6.55ª | 6.76ª | 0.00b | 6.13ª | |
| Lm 1.3 | 6.18ª | 6.35ª | 0.00b | 6.27ª | |
| Lm 7.1 | 6.33ª | 6.70ª | 0.00b | 6.12ª | |
| Lm 7.2 | 6.09ª | 6.59ª | 0.00b | 6.48ª | |
| Lm 7.3 | 6.32ª | 6.71ª | 0.00b | 6.21ª | |
| Lm z44 | 6.49ª | 6.58ª | 0.00b | 6.58ª | |
| Lm 10.1 | 5.87ª | 6.58ª | 4.03ª | 6.50ª | |
| Lm 10.2 | 6.39ª | 6.61ª | 0.00b | 6.33ª | |
| Packaged pork loin | Lm 2.1 | 5.11ª | 5.18ª | 0.00b | 5.11ª |
| Lm 2.3 | 5.13ª | 5.43ª | 0.00b | 5.28ª | |
| Lm 3.1 | 5.36ª | 5.53ª | 0.00b | 5.27ª | |
| Lm 3.2 | 5.55ª | 5.77ª | 0.00b | 5.57ª | |
| Lm 3.3 | 5.17ª | 5.24ª | 0.00b | 5.25ª | |
| Lm 42.1 | 5.61ª | 5.75ª | 0.00b | 5.40ª | |
| Lm 42.2 | 5.26ª | 5.63ª | 0.00b | 5.49ª | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



