
Submitted:
24 September 2024
Posted:
25 September 2024
You are already at the latest version
Abstract
Background/Objectives: Listeria monocytogenes is a foodborne pathogen that can infect humans and animals, causing non-invasive gastrointestinal listeriosis or invasive listeriosis. The objectives of this study were to determine: the genetic diversity of L. monocytogenes; the genes associated with the resistance to antibiotics, benzalkonium chloride (BC), and CdCl2, and biofilm formation. Methods: Seventeen strains of L. monocytogenes isolated fresh fruits and vegetables were selected for this study. Genetic diversity and the genes associated with antibiotic resistance were determined by PCR amplification. Susceptibility to antibiotics were determined using the agar diffusion method. Resistance to BC and CdCl2 was determined using the minimum inhibitory concentration method. Capacity for biofilm formation was evaluated using the crystal violet staining method. Results: This study demonstrates that the isolates of L. monocytogenes belong to serotypes 1/2a (29.4%) and 1/2b (70.5%). Potential hypervirulent strains containing 70.5% Listeria pathogenicity island 1 (LIPI-1-2), 17.6% (LIPI-1-3), and 11.7% (LIPI-1-4) were also identified. Antibiotic susceptibility tests show that L. monocytogenes exhibit six different multiresistant patterns with a MAR index of ³ 0.46 (70.5%); besides, the gen Ide, tetM, and msrA were detected in 52.9%, 29.4%, and 17.6%, respectively, associated with efflux pump Lde, tetracycline and ciprofloxacin resistance. Phenotypic test shows that 58.8% cadmium-resistant L. monocytogenes have a co-resistance to BC of 23.5%. Subsequently, all strains of L. monocytogenes exhibited moderate biofilm. Conclusions: This study contributes to the persistence and genetic diversity of strains L. monocytogenes isolated fresh fruits and vegetables; in addition, resistance to CdCl2 and correlation with co-resistance to BC disinfectant used in the food industry.
Keywords:
Listeria monocytogenes
; virulence factors
; antimicrobial resistance
; benzalkonium chloride
; biofilms
1. Introduction
Listeria monocytogenes is a facultative intracellular pathogen widely distributed in nature and causes non-invasive gastrointestinal listeriosis or invasive listeriosis. Clinical manifestations of invasive listeriosis are septicemia, encephalitis, endocarditis, meningitis, abortions, and fetal death, or non-invasive gastrointestinal listeriosis may be asyntomatic or have flu-like symptoms or a febrile gastroenteritis syndrome [1,2]. Invasive listeriosis affects high-risk groups, including populations such as adults (> 65 years), pregmant women newborns, immunocompromised people and patients with cancer or diabetes [3,4]. The Centers for Disease Control and Prevention (CDC) estimates that there are approximately 1600 cases of listeriosis and 260 deaths annually [3]. In the European Union (EU), invasive listeriosis in humans is (2500 cases annually), but it is the most serious cause of foodborne disease, with a high rate of hospitalization and death, besides, the zoonosis caused by this pathogen in 2022 increased by 15.9% compared to 2021 with, 2738 cases [4]. L. monocytogenes can be spread through the agricultural environments, such as soil and irrigation water these sources may be contaminate fresh produce including fruits and vegetables at various stages of production and processing [5,6,7], creating a public health problem with economic losses for the food industry.
In fact, the Interagency Food Safety Analytics Collaboration (IFSAC) reports that 76% of foodborne L. monocytogenes illnesses (2016-2020) in the United States were associated with three categories in particular with dairy products (37.1%), fruits (24.8%), and vegetable row crops (14.1%) [8]. Recalls, Market Withdrawals, & Safety Alerts in the United States reports recalls in 2023 of fruits and vegetables due to the presence of this pathogen in organic green kiwifruit, organic frozen pineapple, and frozen fruit blend containing organic frozen pineapple, kale, spinach, collard green products, mung bean sprouts [9]. Furthermore, Food Standards Australia-New Zealand (FSANZ) in the period from 2019 to 2023 reports 83 food recalls associated with contamination by foodborne pathogens where L. monocytogenes represents 36% (30 recalls) of these recalls followed by Salmonella spp. (33%; 27 recalls) and Escherichia coli (22%; 18 recalls) the recalls include several food categories such as fruits and vegetables, dairy products, meats and processed meats, etc [10].
Additionally, the prevalence of resistance of L. monocytogenes isolates in food and environment has been associated with the use of antibiotics in medicine, veterinary medicine, and agricultural production systems, with some practices such as soils treated with manure, growth promoters, and misuse of therapeutic treatments for veterinary purposes [11,12,13]. The increase in multi-resistance pathogens is a significant public health problem; besides, the situation becomes increasingly severe as these pathogens spread globally, and acquire new resistance mechanisms, and therefore, there are no alternative therapies for their control [14,15]. In fact, the World Health Organization estimates that bacterial resistance will cause 10 million deaths by 2025 [16]. Moreover, the persistence de L. monocytogenes is linked (i) to resistance to antimicrobials or sanitizing treatments; (ii) the ability of cells to form biofilm on equipment or in the environmental surroundings; (iii) the presence of strains to survive in various food preservation conditions or environmental stresses; or (iv) the inability to remove cells from niches onto the food environment [17,18,19,20]. Therefore, the objectives of the present study were to determine: i) the genetic diversity of L. monocytogenes in fresh fruits and vegetables; ii) the genes associated with resistance antibiotics and multidrug-resistant strains of L. monocytogenes; and iii) the resistance to benzalkonium chloride (BC), cadmium chloride (CdCl2) and formation of biofilm.
2. Results
2.1. Genomic of L. monocytogenes Sublineages and Virulence Genes
The isolates of L. monocytogenes belong to phylogenetic group I.1 (29.4%; serotype 1/2a) and II.2 (70.5%; serotype 1/2b). Additionally, pathogenicity islands were detected in L. monocytogenes, including LIPI-1 and LIPI-2 (100%), LIPI-3 (29.4%) and LIPI-4 (11.7%). The prfA and actA genes were detected in 100% of the isolates. Among the isolates, the following pathogenicity islands were found in significant prevalence: 70.5% (LIPI-1+ LIPI-2), 17.6% (LIPI-1+ LIPI-2+ LIPI-3), and 11.7% (LIPI-1+ LIPI-2+ LIPI-3+LIPI-4) (Table 1).
2.2. Antimicrobial Resistance Gene Profiling
Sixteen isolates were screened for the ciprofloxacin resistance gene Ide (52.9%). Of these, 41.1% showed phenotypic intermediate resistance and 11.7% resistance against ciprofloxacin. The tetracycline resistance gene tetM was detected in 29.4%, whereas five isolates showed phenotypic resistance. Macrolide resistance gen msrA was detected in 17.6%. However, the macrolide resistance gene ermA and the chloramphenicol resistance gene cat were not detected (Table 1).
2.3. Antimicrobials, Sanitizing, Cadmium, and Biofilm
L. monocytogenes was found to be susceptible to ciprofloxacin, tetracycline, vancomycin, chloramphenicol, gentamicin, erythromycin, and trimethoprim-sulfamethoxazole (47-100%) according to Table 2. However, the isolates were resistant to 1st and 3rd generation β-lactam (penicillin, ampicillin, dicloxacillin, cephalothin and cefotaxime) (94.1-100%), clindamycin (52.9%), tetracycline (29.4%), and ciprofloxacin (11.7%). Among the tested L. monocytogenes isolates, six different multiresistant patterns were observed, with the most frequently being PE-CF-AM-CFX-DC (23.5%) and PE-CF-AM-CFX-DC-CLM (35.2%). The MAR index for L. monocytogenes isolates ranged from 0.23 to 0.61, with 70.5% of them presenting a MAR index of ≥ 0.46 (Table 1). Additionally, the clindamycin and ciprofloxacin showed intermediate resistance of 41.1% (Table 2). In addition, MIC of 76.4% of isolates laid between 0.7 and 3.1 µg/mL BC, while the criteria for resistance of BC is CMI ≥ 6 µg/mL, which is at least twice that of the MIC for the predominant number of L. monocytogenes strains (MIC = 3.1 µg/mL) (Table 3). MIC of 41.1% of isolates were to < 70 µg/mL CdCl2 and 58.8% cadmiun-resistance L. monocytogenes (MIC ≥ 70 µg/mL). Finally, all L. monocytogenes were classified as carrying a moderate biofilm (Table 3).
3. Discussion
Listeria monocytogenes is a foodborne pathogen that causes invasive or non-invasive listeriosis in humans; the severity of the pathogenesis is associated with several factors, such as risk group and hazard characterization. In the present study, strains L. monocytogenes isolated from fresh fruits and vegetables belonged to serotypes 1/2a (29.4%) and 1/2b (70.5%). Several authors [5,7,21,22] have reported similar prevalence percentages for serotypes 1/2a – 3a (33-65%) and 1/2b-3b-7 (50-79.6%); but Maćkiw et al. [22] and Chen et al. [23] identified less prevalence for serotypes 1/2a-3a (10.8%) and 1/2b (2%) of isolated L. monocytogenes from ready-to-eat (RTE) foods, fruits, fresh and frozen vegetable. Likewise, Kayode & Okoh [7] does not detect serotype 1/2a but 1/2b (79.61%) and 4b (8.7%) of L. monocytogenes in fruits and vegetables samples. Indeed, serotypes 1/2a, 1/2b, 1/2c and 4b are responsible for 95% of human listeriosis cases and have been frequently isolated from food products and patients [5,24,25]. This suggests that the diversity of L. monocytogenes serotype prevalence may be related to the geographic region of the country, monitoring procedures, and methodologies for detecting of L. monocytogenes in various food categories (fruits, vegetable row crops, dairy, pork, chicken, beef, etc.), specific characteristic of the food (fresh or frozen) or sources of contamination that interact during food production and distribution.
This investigation detected the presence of genes harboring L. monocytogenes pathogenicity islands (LIPIs) to assess the potential risk that L. monocytogenes may pose to public health. All strains of L. monocytogenes isolated from fresh fruits and vegetables were found to harbor the genetic elements of Listeria Pathogenicity Island 1 (LIPI-1; prfA, hly, plcA, plcB, mpl, and actA), which encode virulence factors that promote the growth and spread of L. monocytogenes. Once inside the host cell, the phagocytic vacuole is lysed by listeriolysin O (LLO) encoded by the hly gene, which is a pore-forming toxin that mediates the lysis of bacterial cells in the host cytoplasm and enhances its cytolytic action through phosphatidylinositol-PLC and phosphatidylcholine-PLC that mediate pathogen escape from single and double membrane-bound vacuoles. ActA plays a role in facilitating the motility of the bacterial cell to the host cell’s cytoplasm, and the actin cytoskeleton is hijacked to favor cell-to-cell spread [26,27,28]. The prfA gen was identified in this study, which encodes the PrfA regulatory protein that controls the expression of the pathogenicity determinants of L. monocytogenes [29]. Our results are similar to those reported by several researchers [5,7,30,31]; have shown that prfA (100%), mpl (92-100%), plcA (92-100%), plcB (100%), hly (100%), and actA (84-100%) were detected in L. monocytogenes isolated from fruits and fresh and frozen vegetable, and agricultural environments such as irrigation water and agricultural soil. The genes inlA, inlB, inlC, and inlJ harbor the Listeria Pathogenicity Island 2 (LIPI-2) were detected in all the L. monocytogenes strains in this study. This is in agreement with several investigations where the genes inlA (74.1-100%), inlB (81.5-100%), inlC (70.6-100%), and, inlJ (66.7-100%) were detected, which encode a set of internalins that play a role in the adhesion, invasion de L. monocytogenes cells to the host cell and dissemination of L. monocytogenes [5,7,22,29,30,31]. InlA adheres to and invades intestinal epithelial cells that express the E-cadherina receptor, thereby facilitating the crossing of the intestinal; besides, the InlC and InlJ involved in post-intestinal dissemination of L. monocytogenes infection [6,22,32].
Other islands detected in this study were Listeria Pathogenicity Island-3 (LIPI-3) in 29.4% and Listeria Pathogenicity Island-4 (LIPI-4) in 11.7%. These genes harboring LIPI-3 encode Listeriolysin S (LLS), a bacteriocin with hemolytic and cytotoxic factors that contributes to polymorphonuclear neutrophils survival or alteration of the gut microbiota [22,30,33], and LIPI-4 is involved in the infection of the host’s neuronal and placental tissues; besides, it confers hyper-virulence strains [34,35].
Additionally, all L. monocytogenes isolates exhibited resistance to penicillin, ampicillin, dicloxacillin, cephalothin, cefotaxime, clindamycin, tetracycline and ciprofloxacin being the most frequently encountered. Several studies have reported varying prevalences of antimicrobial resistance to penicillin (2.5-100%), ampicillin (50-100%), gentamicin (20-40%), STX (30%), erythromycin (23.5-100%), tetracycline (90-100%), chloramphenicol (20-70%), cefotaxime (80-100%), clindamycin (57.5-100%), cephalothin (50-100%), and ciplofloxacin (35.2-40%) in Listeria spp. and L. monocytogenes isolates from different categories of food and food processing environments [31,36,37,38]. Phenotypic intermediate resistance to clindamycin (41%), ciprofloxacin (41%), erythromycin (5.8%) and gentamicin (5.8%) L. monocytogenes isolates in this study is in agreement with other investigators [37,38,39] where reported prevalence of intermediate resistance to clindamycin (30%), intermediate to ciprofloxacin (5-64.7%), intermediate gentamicin (2.5%), intermediate tetracyclin (2-5.8%) in L. monocytogenes. Over-prescription of antibiotics in clinical practice, use of antibiotics in animal production, inadequate veterinary treatment to prevent animal disease, and migration and accumulation of residues of veterinary antibiotics in agricultural soils and irrigation water may contribute to the prevalence of resistance and heterogeneity of antimicrobial resistance patterns observed in L. monocytogenes isolates [11,13,38,40].
This study demonstrates that L. monocytogenes exhibit six different multiresistant patterns with a MAR index of ≥ 0.46 (70.5%), indicating a higher risk source exposed to antibiotics. A MAR index of ≥ 0.2 suggests intensive use of antibiotics in the region and a high risk of promoting antibiotic resistance [41,42]. Iwu & Okoh [31] and Maurice Bilung et al. [36] reported similar MAR index (0.31-0.85) for multidrug-resistant Listeria spp. and MAR index (0.2-1) for L. monocytogenes isolates from irrigation water and agricultural soil, suggesting a higher risk source that is constantly exposed to antibiotics, which are used to prevent or treat animal disease and promote animal growth. Agricultural activities such as the use of fertilizers containing antibiotic residues increase resistance or the presence of antibiotic-resistant strains in the soil or the use of water for plant irrigation [12,43].
Furthermore, L. monocytogenes has been found to develop resistance to several antibiotics, tetracycline, ciprofloxacin, erythromycin, clindamycin, penicillin and ampicillin, through the acquisition of genetic elements such as conjugative transposons, and self-transferable and mobilizable plasmids [13,28,44]. Therefore, the detection of Ide, tetM, and msrA in the L. monocytogenes isolates in our study may be related to resistance mechanisms such as efflux pump Lde and the transposon Tn916 harboring the tetM confers to resistance to ciprofloxacin and tetracycline [44,45]. In accordance with the detection of Ide, tetM, and msrA in Listeria spp. and L. monocytogenes isolates from slaughtering and processing environments, food and clinical [7,15,39,46]. Although we did not detected the presence of cat and ermA genes among our isolates, they have already been detected in L. monocytogenes; cat (100%) and ermA (16.9%) Listeria spp. in food and processing environments [15,46]. In fact, these mechanisms of resistance affect the treatment of human listeriosis, drugs such as (i) first-line ampicillin or penicillin G in combination with an aminoclycosid (gentamicin); (ii) second-line trimethoprim in combination with sulfonamide, such as sulfamethoxazole-co-trimoxazole, as well as erythromycin, tetracycline, and vancomycin [28,45].
This investigation suggests that the high prevalence of resistance and intermediate resistance in the isolates of L. monocytogenes may be due to an inadequate use of antimicrobial agents in veterinary medicine, the extensive use animal foodstuff, agricultural production systems or to the intrinsic resistance of L. monocytogenes against cephalosporins and fluoroquinolones, which is associated with the lack or low affinity of the enzyme that catalyzes the final step of cell wall synthesis [28,45]. However, the prevalence of antibiotic resistance reported in different countries is influenced by the health policies related to comprehensive antimicrobial management and the determination of antimicrobial breakpoints specific to veterinary medicine in the methods for antimicrobial susceptibility testing of bacterial pathogens of animal origin and zoonotic bacteria that can affect humans [39,47].
Moreover, L. monocytogenes has demonstrated resistance to non-essential toxic metals, including arsenic and cadmium [48]. In this study, 58.8% of the L. monocytogenes isolates were resistant to CdCl2 (MIC ≥ 70 µg/mL); this was a particularly common occurrence among isolates of serotype 1/2a and 1/2b. Similar findings have been reported by other researchers [30,49,50,51] regarding the prevalence of cadmium resistance (63-90%) in serotypes 1/2a and 1/2b of L. monocytogenes isolated from food and environment. The presence of heavy metal residues in the environment is related the industrial sector (anthropogenic sources) such as agricultural practices such as using of phosphate fertilizers, which represent a significant source of cadmium input to agricultural soil, water and food onto chain food [52]; this way increases the survival potential of L. monocytogenes and the acquisition of mobile genetic elements of heavy metal resistance-determinants in diverse environmental niches. Likewise, Zhang et al. [51] argue that cadmium exerts long-term selective pressure, allowing L. monocytogenes to produce tolerance.
On the other hand, QACs are used in the food industry within disinfection processes to control, reduce, and inactivate foodborne pathogens [53,54,55,56]; however, the prevalence of resistance to QACs, in particular to BC has been detected in L. monocytogenes isolated from food and processing plant environments, and has association to cadmium [57,58,59,60]. Our study shows that four L. monocytogenes isolates (23.5%) were resistant to BC, and ten isolates (58.8%) were resistant to cadmium with co-resistance to BC and cadmium (23.5%), respectively. Ratani et al. [61] showed 14% resistant to BC and 57% to Cd and Xu et al. [50] detected 16.7 % resistant to BC and Cd in L. monocytogenes. However, the cadmiun-resistant L. monocytogenes and BC were not always correlated [49]. Based on the cadmium or BC resistance result, this could be due to the genetic diversity of L. monocytogenes strains associated with genetic determinants cadmium resistance such as cadA1 (plasmid-transposon Tn5422), cadA2 (plasmid pLM80), cadA3 (at chromosome level of L. monocytogenes) and cadC [48,62]; BC resistance genes qacA/B, qacC/D, qacE, qacE1Δ-sul, qacF, qacG, bcrABC, transposon Tn6188 (containing the qacH gene), or mdrL (chromosome and plasmid-borne encodes an efflux pump) [50,60,63,64]. In addition to the various breakpoints specific to determine resistance to disinfectants (MIC = 4 - 32 µg/mL), it may interfere with the prevalence of phenomena of resistance BC in L. monocytogenes as they are established according to the number of L. monocytogenes isolates, origin of strains, medium for susceptibility testing medium, etc. [65].
Additionally, the decrease in the efficiency of QACs is related to (i) environmental niches with sites that are difficult to clean and disinfect, the inability to remove cells, (ii) the presence of organic matter on food contact surfaces, (iii) exposure to sublethal concentrations of QACs on food contact surfaces that allow BC tolerance of L. monocytogenes; besides, this is associated with the persistence of L. monocytogenes in the food industry and the subsequent adaptation and formation of biofilms [18,66,67,68]. Our results indicate that all L. monocytogenes isolates can form a biofilm, which harbors the genes inlA, prfA, plcA, hly, plcB, and actA associated with biofilm formation. Previous research indicated that inlA, inlL, prfA, plcA, actA, Imo0673, bapL, recO, Imo2504, and luxS play a role in the different stages of L. monocytogenes biofilms formation [35,46,69]. Likewise, Price et al. [70] argue that the presence of LIPI-1 genes, hly and prfA, are required for biofilm formation by L. monocytogenes. However, biofilm formation is a complex and dynamic process that is contingent upon a number of factors, including the availability of nutrients in the environment, the origin and biodiversity of the strain, and the quorum sensing (QS) that have activation and regulation of biofilm-associated genes and virulence factors [19]. Several studies have demonstrated that L. monocytogenes forms biofilms exhibit significantly greater resistance to sanitizing and antibiotic compounds than free-floating cells [44,71]. Therefore, it could represent a source of concurrent food contamination, increasing the risk to the consumer and impacting public health, in addition to the economic losses associated with voluntary recalls or damage to equipment within the food industry. Moreover, in this study, L. monocytogenes strains isolated from fresh fruits and vegetables can potentially cause severe human infection; however, the severity of the clinical manifestations of L. monocytogenes is related to genetic diversity, immune system status, or host comorbidities. Indeed, Castañedas-Ruelas et al. [72] argument that the dearth of data concerning the significance of L. monocytogenes in Mexico underscores the necessity to sensitize authorities to the characterization of the risks associated with food and human exposure to L. monocytogenes, thereby facilitating an understanding of the clinical and epidemiological impact of listeriosis in Mexico. Notably, it is essential to incorporate techniques that allow us to determine the biodiversity of L. monocytogenes. This enables the identification of clonal complexes (CCs) and sublineages (SLs), enabling a relationship between the origin of isolation and the infection's severity in this investigation's context. For example, SL121 (CC121), SL9 (CC9), and CC8 + CC16 are associated with Lineage II, which has a food origin, and SL1 (CC1), SL2 (CC2), SL4 (CC4), and SL6 (CC6) with clinical cases of lineage I. CC1 and CC4 are related to invasive forms of listeriosis, including maternal-neonatal and CNS infection [34,73].
4. Materials and Methods
4.1. Bacterial Strains
Seventeen strains of L. monocytogenes were selected for this study from various fresh fruits and vegetables, including Hass avocados, lettuce, parsley, cilantro, broccoli, and cucumber. The strains were confirmed by PCR using hly (Listeriolysin O) and prs (Putative phosphoribosyl pyrophosphate synthetase) [24,25]. Stocks were stored in tryptic soy broth (TSB; Becton Dickinson Bioxon, Le Pont de Claix, France) containing 30% glycerol at – 80 ºC.
4.2. Genomic Characterization: Genes Involved in Pathogenicity Islands, Biofilm Formation and Resistance Antibiotics
L. monocytogenes strains were reactivated in TSB with 0.6% yeast extract (TSBYE) (Sigma-Aldrich, St. Louis, MO, USA) for 24 h at 30 ºC. According to the manufacturer’s instructions, genomic DNA was extracted from L. monocytogenes using a Bacteria DNA Preparation Kit (Jena Bioscience, Jena, Germany). All L. monocytogenes strains were investigated for detection of the genes (prfA, hly, plcA, plcB, mpl, actA, inlA, inlB, inlC, inlJ, llsA, llsG, llsH, llsX, llsB, llsY, llsD, llsP, licC, licB, licA, and glvA) that harbored the L. monocytogenes pathogenicity islands (LIPI’s) by PCR using the protocol of Zhang et al. [21]. Subsequently, phylogenetic group of L. monocytogenes [I.1 (172a-3a), I.2 (1/2c-3c), II.1 (4b-4d-4e), II.2 (1/2b-3b-7), and III (4a-4c)] and the genes associated with resistance antibiotics [efflux pump Ide (Ide), chloramphenicol acetyltransferase (cat), macrolide-lincosamide-streptogramin B efflux pump (msrA), rRNA adenine-N-6-methyltransferase (ermA), and ribosomal protection protein tetM (tetM)] were determined using the protocol of Doumith et al. [24] and Boháčová et al. [39]. After amplification, the products were electrophoresed on 1 % (w/v) agarose gel (UltraPure agarose, Invitrogen, Carlsbad, USA) using SYBR Green (Sigma-Aldrich, St. Louis, MO, USA) and visualized by transillumination under UV light (UVP, DigiDoc-It Darkroom, Upland, CA, USA).
4.3. Phenotypic Characterization for the Persistence of L. monocytogenes
4.3.1. Disinfectant and Heavy Metal Sensitivity
Benzalkonium chloride (BC) (Sigma-Aldrich, St. Louis, MO, USA) was used to determine the sensitivity of L. monocytogenes strains to quaternary ammonium compound (QAC) using the protocol of Gray et al. [30], with modifications. Briefly, L. monocytogenes strains were grown overnight in Mueller Hinton broth (MHB; Becton Dickinson Bioxon, Le Pont de Claix, France) at 30 ºC and diluted to ~108 CFU/mL. The BC stock concentration, 100 µL, was added to the microtiter plates (Corning® 96-Well Assay Microplate, Lowell, MA, USA) with concentrations of 100, 50, 25, 12.5, 6.2, 3.1, 1.5, and 0.7 µg/mL. The microtiter plates were then incubated at 30 ºC / 24 h, and growth was monitored by measuring the OD560 using a Multiskan FC (Thermo Fisher Scientific, Inc., Madison, WI, USA) to determine the minimum inhibitory concentration (MIC). Each assay was performed in triplicate, with positive and negative controls. Cadmium chloride (CdCl2; Sigma-Aldrich, St. Louis, MO, USA) was used to determine the resistance of L. monocytogenes to the heavy metal cadmium. Mueller Hinton Agar (MHA; Becton Dickinson Bioxon, Le Pont de Claix, France) was supplemented with different concentrations of CdCl2 (400, 200, 100, 70, 50, 25, and 12.5 µg/mL). Each L. monocytogenes isolate was adjusted to ~108 CFU/mL and inoculated onto the CdCl2 plates. The plates were then incubated at 37 ºC / 24 h for triplicate. Resistance to cadmium was interpreted as ≥ 70 µg/mL [49,50].
4.3.2. Phenotypic Antibiotic Sensitivity and Resistance Analysis
The resistance and susceptibility of L. monocytogenes strains to antibiotics were determined using the agar diffusion method, following the Clinical and Laboratory Standards Institute (CLSI) [74]. Bacterial suspensions adjusted to 0.5 McFarland were inoculated onto MHA, incorporated the antibiotics, and incubated at 35 ºC / 24 h. Thirteen were selected among eleven classes of antimicrobials: phenicols [chloramphenicol (CL, 30 µg)]; cephalosporines (1st generation) [cephalothin (CF, 30 µg)]; licosamide [clindamycin (CLM, 30 µg)]; sulphonamide [trimethoprim-sulfamethoxazole (SXT, 2.5/23.75 μg)]; cyclic peptides [tetracycline (TE, 30 µg)]; macrolide [erythromycin (E, 15 µg)]; aminoglycoside [gentamicin (GE, 10 µg)]; fluoroquinolone [ciprofloxacin (CPF, 5 μg)]; cephalosporines (3rd generation) [cefotaxime (CFX, 30 µg)]; glycopeptide [vancomycin (VA, 30 µg)]; β-lactam [penicillin (P, 10 U), ampicillin (AM, 10 µg), and dicloxacillin (DC, 1 µg)] (BBLTM Sensi-DiscTM). The inhibition zones were interpreted as Resistance (R), Intermediate resistance (I) and Susceptible (S) according to CLSI [28]. L. monocytogenes ATCC 19111 was used as the positive control. The multiple antibiotic resistance (MAR) index of L. monocytogenes isolates was determined using the methods by Krumperman [41] and Blasco et al. [75].
4.3.3. Biofilm Formation Assay
The strains’ ability to form biofilms was evaluated in polystyrene microtiter plates (Corning® 96-Well Assay Microplate, Lowell, MA, USA) using crystal violet (CV) staining, following the protocol described by Avila-Novoa et al. [76]. For each strain, 230 μL of TSB and 20 μL bacterial suspension (~108 CFU/mL) were added to polystyrene microtiter plates and incubated at 30 °C for 240 h. The planktonic bacteria were removed using 200 μL of phosphate-buffered saline (PBS; 7 mM Na2HPO4, 3 mM NaH2PO4 and 130 mM NaCl, pH 7.4). The biofilm was fixed with 200 μL of methanol for 10 min, dried at 55 ºC for 15 min, and stained with 200 µL of 0.1% crystal violet for 45 min. Excess stain was rinsed off with PBS and resolubilized with 200 μL of 95% ethanol. Absorbance was measured at 595 nm (OD595), using the Multiskan FC. The assay was performed in triplicate, including positive and negative controls. The cutoff OD (ODc) was determined using the protocol described by Stepanović et al. [77].
5. Conclusions
The present study provides data regarding the genetic diversity of L. monocytogenes in fruits and vegetables in Mexico and the possible impact on its population associated with the genetic determinants involved in the severity of the pathology and resistance mechanisms of antibiotics that are used within the therapeutic scheme of the patient or veterinary medicine, in addition to the environmental impact that fertilizer residues have on antimicrobial resistance. This is to raise awareness of a continuous improvement in treatments and sanitary prerequisites such as agricultural practices, food farming practices, and standard operating procedures for sanitation. A recommendation is to validate and rotate disinfectants to reduce the risk of L. monocytogenes niches being established in the environment and tolerance to disinfectants that promote the survival of L. monocytogenes biofilms.
Author Contributions
Conceptualization, methodology, validation, formal analysis, investigation, data curation, writing—original draft preparation and visualization, M.G.A.-N.; methodology and investigation, O.A.S.-V.; investigation, validation and formal analysis, P.J.G.-M., L.M.-C., and N.E.M.-G.; writing— review and editing, supervision, resources, project administration, funding acquisition, and visualization, M.G.-L. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data used to support the findings of this study are available from the corresponding author upon request.
Acknowledgments
The authors would like to thank Daniel Hernández Alvarado for his technical support.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Charlier, C. , Perrodeau, É., Leclercq, A., Cazenave, B., Pilmis, B., Henry, B., Lopes, A., Maury, M. M., Moura, A., Goffinet, F., Dieye, H. B., Thouvenot, P., Ungeheuer, M. N., Tourdjman, M., Goulet, V., de Valk, H., Lortholary, O., Ravaud, P., Lecuit, M., MONALISA study group. Clinical features and prognostic factors of listeriosis: the MONALISA national prospective cohort study. Lancet. Infect. Dis. 2017, 17, 510–519. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Boland, J.A. , Kuhn, M., Berche, P., Chakraborty, T., Domínguez-Bernal, G., Goebel, W., González-Zorn, B., Wehland, J., Kreft, J. Listeria pathogenesis and molecular virulence determinants. Clin. Microbiol. Rev. 2001, 14, 584–640. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Listeria Infection (Listeriosis). Available online: https://www.cdc.gov/listeria/about/index.html (accessed on 19 June 2024).
- European Food Safety Authority (EFSA). Listeria. Available online: https://www.efsa.europa.eu/en/topics/topic/listeria#efsas-role (accessed on 13 August 2024).
- Chen, M. , Chen, Y., Wu, Q., Zhang, J., Cheng, J., Li, F., Zeng, H., Lei, T., Pang, R., Ye, Q., Bai, J., Wang, J., Wei, X., Zhang, Y., Ding, Y. Genetic characteristics and virulence of Listeria monocytogenes isolated from fresh vegetables in China. BMC Microbiol. 2019, 19, 119. [Google Scholar] [CrossRef]
- Gartley, S. , Anderson-Coughlin, B., Sharma, M., Kniel, K.E. Listeria monocytogenes in Irrigation Water: An Assessment of Outbreaks, Sources, Prevalence, and Persistence. Microorganisms 2022, 10, 1319. [Google Scholar] [CrossRef]
- Kayode, A.J. , Okoh, A.I. Incidence and genetic diversity of multi-drug resistant Listeria monocytogenes isolates recovered from fruits and vegetables in the Eastern Cape Province, South Africa. Int. J. Food Microbiol. 2022, 363, 109513. [Google Scholar] [CrossRef] [PubMed]
- Interagency Food Safety Analytics Collaboration (IFSAC). 2022. Foodborne illness source attribution estimates for 2020 for Salmonella, Escherichia coli O157, and Listeria monocytogenes using multi-year outbreak surveillance data, United States. Available online: https://www.cdc.gov/ifsac/media/pdfs/P19-2020-report- TriAgency-508.pdf (accessed on 20 August 2024).
- Food & Drudg Administration (FDA). 2024. Recalls, Market Withdrawals, & Safety Alerts. Available online: https://www.fda.gov/safety/recalls-market-withdrawals-safety-alerts (accessed on 13 August 2024).
- Food Standard Australia- New Zealand (FSANZ). Australian food recall statistics. Available online: https://www.foodstandards.gov.au/food-recalls/recallstats (accessed on 16 August 2024).
- Hu, X. , Zhou, Q., Luo, Y. Occurrence and source analysis of typical veterinary antibiotics in manure, soil, vegetables and groundwater from organic vegetable bases, northern China. Environ. Pollut. 2010, 158, 2992–2998. [Google Scholar] [CrossRef]
- Popowska, M. , Rzeczycka, M., Miernik, A., Krawczyk-Balska, A., Walsh, F., Duffy, B. Influence of soil use on prevalence of tetracycline,streptomycin, and erythromycin resistance and associated resistance genes. Antimicrob. Agents Chemother. 2012, 56, 1434–1443. [Google Scholar] [CrossRef]
- Soleimani, M. , Sadrabad, E.K., Hamidian, N., Heydari, A., Mohajeri, F.A. Prevalence and Antibiotic Resistance of Listeria monocytogenes in Chicken Meat Retailers in Yazd, Iran. J. Environ. Health Sustain. Dev. 2019, 4, 895–902. [Google Scholar] [CrossRef]
- Broncano-Lavado, A. , Santamaría-Corral, G., Esteban, J., García-Quintanilla, M. Advances in bacteriophage therapy against relevant multidrug-resistant pathogens. Antibiotics 2021, 10, 672. [Google Scholar] [CrossRef]
- Wu, L. , Bao, H., Yang, Z., He, T., Tian, Y., Zhou, Y., Pang, M., Wang, R., Zhang, H. Antimicrobial susceptibility, multilocus sequence typing, and virulence of Listeria isolated from a slaughterhouse in Jiangsu, China. BMC Microbiol. 2021, 21, 327. [Google Scholar] [CrossRef]
- Giono-Cerezo, S. , Santos-Preciado, J.I., Morfín-Otero, M.R., Torres-López, F.J., Alcántar-Curiel, M.D. Resistencia antimicrobiana. Importancia y esfuerzos por contenerla. Gac. Méd. Méx. 2020, 156, 172–180. [Google Scholar] [CrossRef]
- Carpentier, B. , Cerf, O. Review - Persistence of Listeria monocytogenes in food industry equipment and premises. Int. J. Food Microbiol. 2011, 145, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Colagiorgi, A. , Bruini, I., Di Ciccio, P.A., Zanardi, E., Ghidini, S., Ianieri, A. Listeria monocytogenes Biofilms in the wonderland of food industry. Pathogens 2017, 6, 41. [Google Scholar] [CrossRef]
- Yu, T. , Jiang, X., Xu, X., Jiang, C., Kang, R., Jiang, X. Andrographolide Inhibits Biofilm and Virulence in Listeria monocytogenes as a Quorum-Sensing Inhibitor. Molecules 2022, 27, 3234. [Google Scholar] [CrossRef]
- Wiśniewski, P. , Chajęcka-Wierzchowska, W., Zadernowska, A. High-Pressure Processing—Impacts on the Virulence and Antibiotic Resistance of Listeria monocytogenes Isolated from Food and Food Processing Environments. Foods 2023, 12, 3899. [Google Scholar] [CrossRef]
- Zhang, Y. , Dong, S., Chen, H., Chen, J., Zhang, J., Zhang, Z., Yang, Y., Xu, Z., Zhan, L., Mei, L. Prevalence, Genotypic Characteristics and Antibiotic Resistance of Listeria monocytogenes From Retail Foods in Bulk in Zhejiang Province, China. Front. Microbiol. 2019, 10, 1710. [Google Scholar] [CrossRef]
- Maćkiw, E. , Korsak, D., Kowalska, J., Felix, B., Stasiak, M., Kucharek, K., Postupolski, J. Incidence and genetic variability of Listeria monocytogenes isolated from vegetables in Poland. Int. J. Food Microbiol. 2021, 339, 109023. [Google Scholar] [CrossRef]
- Chen, M. , Wu, Q., Zhang, J., Yan, Z., Wang, J. Prevalence and characterization of Listeria monocytogenes isolated from retail-level ready-to-eat foods in South China. Food Control 2014, 38, 1–7. [Google Scholar] [CrossRef]
- Doumith, M. , Buchrieser, C., Glaser, P., Jacquet, C., Martin, P. Differentiation of the major Listeria monocytogenes serovars by multiplex PCR. J. Clin. Microbiol. 2004, 42, 3819–3822. [Google Scholar] [CrossRef]
- Montero, D. , Bodero, M., Riveros, G., Lapierre, L., Gaggero, A., Vidal, R.M., Vidal, M. Molecular epidemiology and genetic diversity of Listeria monocytogenes isolates from a wide variety of ready-to-eat foods and their relationship to clinical strains from listeriosis outbreaks in Chile. Front. Microbiol. 2015, 6, 384. [Google Scholar] [CrossRef]
- Poimenidou, S.V. , Dalmasso, M., Papadimitriou, K., Fox, E.M., Skandamis, P.N., Jordan, K. Virulence gene sequencing highlights similarities and differences in sequences in Listeria monocytogenes serotype 1/2a and 4b strains of clinical and food origin from 3 different geographic locations. Front. Microbiol. 2018, 9, 1103. [Google Scholar] [CrossRef]
- Pizarro-Cerdá, J. , Cossart, P. Microbe profile: Listeria monocytogenes: A paradigm among intracellular bacterial pathogens. Microbiology (Reading) 2019, 165, 719–721. [Google Scholar] [CrossRef] [PubMed]
- Matle, I. , Mbatha, K.R., Madoroba, E. A review of Listeria monocytogenes from meat and meat products: Epidemiology, virulence factors, antimicrobial resistance and diagnosis. Onderstepoort J. Vet. Res. 2020, 87, e1–e20. [Google Scholar] [CrossRef] [PubMed]
- Osman, K.M. , Kappell, A.D., Fox, E.M., Orabi, A., Samir, A. Prevalence, pathogenicity, virulence, antibiotic resistance, and phylogenetic analysis of biofilmproducing Listeria monocytogenes isolated from different ecological niches in Egypt: Food, humans, animals, and environment. Pathogens 2020, 9, 5. [Google Scholar] [CrossRef]
- Gray, J.A. , Chandry, P.S., Kaur, M., Kocharunchitt, C., Bowman, J.P., Fox, E.M. Characterisation of Listeria monocytogenes food-associated isolates to assess environmental fitness and virulence potential. Int. J. Food Microbiol. 2021, 350, 109247. [Google Scholar] [CrossRef]
- Iwu, C.D. , Okoh, A.I. Characterization of antibiogram fingerprints in Listeria monocytogenes recovered from irrigation water and agricultural soil samples. PLoS ONE 2020, 15, e0228956. [Google Scholar] [CrossRef]
- Pournajaf, A. , Rajabnia, R., Sedighi, M., Kassani, A., Moqarabzadeh, V., Lotfollahi, L., Ardebilli, A., Emadi, B., Irajian, G. Prevalence, and virulence determination of Listeria monocytogenes strains isolated from clinical and non-clinical samples by multiplex polymerase chain reaction. Rev. Soc. Bras. Med. Trop. 2016, 49, 624–627. [Google Scholar] [CrossRef]
- Vilchis-Rangel, R.E. , Espinoza-Mellado, M.R., Salinas-Jaramillo, I.J., Martinez-Peña, M.D., Rodas-Suárez, O.R. Association of Listeria monocytogenes LIPI-1 and LIPI-3 marker llsX with invasiveness. Curr. Microbiol. 2019, 76, 637–643. [Google Scholar] [CrossRef]
- Maury, M.M. , Tsai, Y.H., Charlier, C., Touchon, M., Chenal-Francisque, V., Leclercq, A., Criscuolo, A., Gaultier, C., Roussel, S., Brisabois, A., Disson, O., Rocha, E.P.C., Brisse, S., Lecuit, M. Uncovering Listeria monocytogenes hypervirulence by harnessing its biodiversity. Nat. Genet. 2016, 48, 308–313. [Google Scholar] [CrossRef]
- Mafuna, T. , Matle, I., Magwedere, K., Pierneef, R.E., Reva, O.N. Whole Genome-Based Characterization of Listeria monocytogenes Isolates Recovered From the Food Chain in South Africa. Front. Microbiol. 2021, 12, 669287. [Google Scholar] [CrossRef]
- Maurice Bilung, L. , Sin Chai, L., Tahar, A.S., Ted, C.K., Apun, K. Prevalence, Genetic Heterogeneity, and Antibiotic Resistance Profile of Listeria spp. and Listeria monocytogenes at Farm Level: A Highlight of ERIC- and BOX-PCR to Reveal Genetic Diversity. BioMed Res. Int. 2018, 2018, 3067494. [Google Scholar] [CrossRef]
- Wiśniewski, P. , Zakrzewski, A.J., Zadernowska, A., Chajęcka-Wierzchowska, W. Antimicrobial Resistance and Virulence Characterization of Listeria monocytogenes Strains Isolated from Food and Food Processing Environments. Pathogens 2022, 11, 1099. [Google Scholar] [CrossRef] [PubMed]
- Panera-Martínez, S. , Capita, R., García-Fernández, C., Alonso-Calleja, C. Viability and Virulence of Listeria monocytogenes in Poultry. Microorganisms 2023, 11, 2232. [Google Scholar] [CrossRef] [PubMed]
- Boháčová, M. , Zdeňková, K., Tomáštíková, Z., Fuchsová, V., Demnerová, K., Karpíšková, R., Pazlarová, J. Monitoring of resistance genes in Listeria monocytogenes isolates and their presence in the extracellular DNA of biofilms: a case study from the Czech Republic. Folia Microbiol. 2018, 63, 653–664. [Google Scholar] [CrossRef] [PubMed]
- Iwu, C.D. , Okoh, A.I. Preharvest transmission routes of fresh produce associated bacterial pathogens with outbreak potentials: A review. Int. J. Environ. Res. Public Health 2019, 16, 4407. [Google Scholar] [CrossRef] [PubMed]
- Krumperman, P.H. Multiple antibiotic resistance indexing of Escherichia coli to identify high-risk sources of fecal contamination of foods. Appl. Environ. Microbiol. 1983, 46, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Titilawo, Y. , Sibanda, T., Obi, L., Okoh, A. Multiple antibiotic resistance indexing of Escherichia coli to identify high-risk sources of faecal contamination of water. Environ. Sci. Pollut. Res. 2015, 22, 10969–10980. [Google Scholar] [CrossRef]
- Abriouel, H. , Omar, N.B., Molinos, A.C., López, R.L., Grande, M.J., Martínez-Viedma, P., Ortega, E., Cañamero, M.M., Galvez, A. Comparative analysis of genetic diversity and incidence of virulence factors and antibiotic resistance among enterococcal populations from raw fruit and vegetable foods, water and soil, and clinical samples. Int. J. Food Microbiol. 2008, 123, 38–49. [Google Scholar] [CrossRef]
- Matereke, L.T. , Okoh, A.I. Listeria monocytogenes virulence, antimicrobial resistance and environmental persistence: A review. Pathogens 2020, 9, 528. [Google Scholar] [CrossRef]
- Bertrand, S. , Huys, G., Yde, M., D’Haene, K., Tardy, F., Vrints, M., Swings, J., Collard, J.M. Detection and characterization of tet(M) in tetracycline-resistant Listeria strains from human and food-processing origins in Belgium and France. J. Med. Microbiol. 2005, 54, 1151–1156. [Google Scholar] [CrossRef]
- Avila-Novoa, M.G. , González-Torres, B., González-Gómez, J.P., Guerrero-Medina, P.J., Martínez-Chávez, L., Martínez-Gonzáles, N.E., Chaidez, C., Gutiérrez-Lomelí, M. Genomic Insights into Listeria monocytogenes: Organic Acid Interventions for Biofilm Prevention and Control. Int. J. Mol. Sci. 2023, 24, 13108. [Google Scholar] [CrossRef]
- European Committee on Antimicrobial Susceptibility Testing (EUCAST) /Veterinary Committee on Antimicrobial Susceptibility Testing (VetCAST). Available online: https://www.eucast.org/ast_of_veterinary_pathogens (accessed on 13 August 2024).
- Parsons, C. , Lee, S., Kathariou, S. Heavy metal resistance determinants of the foodborne pathogen Listeria monocytogenes. Genes 2019, 10, 11. [Google Scholar] [CrossRef]
- Mullapudi, S. , Siletzky, R.M., Kathariou, S. Heavy-metal and benzalkonium chloride resistance of Listeria monocytogenes isolates from the environment of Turkey-processing plants. Appl. Environ. Microbiol. 2008, 74, 1464–1468. [Google Scholar] [CrossRef]
- Xu, D. , Deng, Y., Fan, R., Shi, L., Bai, J., Yan, H. Coresistance to Benzalkonium Chloride Disinfectant and Heavy Metal Ions in Listeria monocytogenes and Listeria innocua Swine Isolates from China. Foodborne Pathog. Dis. 2019, 16, 696–703. [Google Scholar] [CrossRef]
- Zhang, H. , Zhou, Y., Bao, H., Zhang, L., Wang, R., Zhou, X. Plasmid-borne cadmium resistant determinants are associated with the susceptibility of Listeria monocytogenes to bacteriophage. Microbiol. Res. 2015, 172, 1–6. [Google Scholar] [CrossRef]
- Agency For Toxic Substances and Disease Registry (ATSDR). Cadmium ToxGuide. (Listeriosis). Available online: https://www.atsdr.cdc.gov/toxguides/toxguide-5.pdf (accessed on 19 June 2024).
- Thévenot, D. , Dernburg, A., Vernozy-Rozand, C. An updated review of Listeria monocytogenes in the pork meat industry and its products. J. Appl. Microbiol. 2006, 101, 7–17. [Google Scholar] [CrossRef]
- Paluszak, Z. , Gryń, G., Bauza-Kaszewska, J., Skowron, K.J., Wiktorczyk-Kapischke, N., Korkus, J., Pawlak, M., Szymańska, E., Kraszewska, Z., Buszko, K., Skowron, K. Prevalence and antimicrobialsusceptibility of Listeria monocytogenes strains isolated from a meat processing plant. Ann. Agric. Environ. Med. 2021, 28, 595–604. [Google Scholar] [CrossRef]
- Duze, S.T. , Marimani, M., Patel, M. Tolerance of Listeria monocytogenes to biocides used in food processing environments. Food Microbiol. 2021, 97, 103758. [Google Scholar] [CrossRef]
- Code of Federal Regulations (CFR). Part 178-Indirect Food Additives: Adjuvants, Production Aids, and Sanitizers. Available online: https://www.ecfr.gov/current/title-21/chapter-I/subchapter-B/part-178#178.1010 (accessed on 9 September 2024).
- Xu, D. , Li, Y., Shamim Hasan Zahid, M., Yamasaki, S., Shi, L., Li, J.R., Yan, H. Benzalkonium chloride and heavy-metal tolerance in Listeria monocytogenes from retail foods. Int. J. Food Microbiol. 2014, 190, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Minarovičová, J. , Véghová, A., Mikulášová, M., Chovanová, R., Šoltýs, K., Drahovská, H., Kaclíková, E. Benzalkonium chloride tolerance of Listeria monocytogenes strains isolated from a meat processing facility is related to presence of plasmid-borne bcrABC cassette. Antonie van Leeuwenhoek 2018, 111, 1913–1923. [Google Scholar] [CrossRef] [PubMed]
- Haubert, L. , Zehetmeyr, M.L., da Silva, W.P. Resistance to benzalkonium chloride and cadmium chloride in Listeria monocytogenes isolates from food and food-processing environments in southern Brazil. Can. J. Microbiol. 2019, 65, 429–435. [Google Scholar] [CrossRef] [PubMed]
- Cooper, A.L. , Carrillo, C.D., Deschenes, M., Blais, B.W. Genomic markers for quaternary ammonium compound resistance as a persistence indicator for Listeria monocytogenes contamination in food manufacturing environments. J. Food Prot. 2021, 84, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Ratani, S.S. , Siletzky, R.M., Dutta, V., Yildirim, S., Osborne, J.A., Lin, W., Hitchins, A.D., Ward, T.J., Kathariou, S. Heavy metal and disinfectant resistance of Listeria monocytogenes from foods and food processing plants. Appl. Environ. Microbiol. 2012, 78, 6938–6945. [Google Scholar] [CrossRef]
- Mullapudi, S. , Siletzky, R.M., Kathariou, S. Diverse cadmium resistance determinants in listeria monocytogenes isolates from the Turkey processing plant environment. Appl. Environ. Microbiol. 2010, 76, 627–630. [Google Scholar] [CrossRef]
- Romanova, N. , Favrin, S., Griffiths, M.W. Sensitivity of Listeria monocytogenes to sanitizers used in the meat processing industry. Appl. Environ. Microbiol. 2002, 68, 6405–6409. [Google Scholar] [CrossRef]
- López-Alonso, V. , Ortiz, S., Corujo, A., Martínez-Suárez, J.V. Analysis of benzalkonium chloride resistance and potential virulence of Listeria monocytogenes isolates obtained from different stages of a poultry production chain in Spain. J. Food Prot. 2020, 83, 443–451. [Google Scholar] [CrossRef]
- Martínez-Suárez, J.V. , Ortiz, S., López-Alonso, V. Potential impact of the resistance to quaternary ammonium disinfectants on the persistence of Listeria monocytogenes in food processing environments. Front. Microbiol. 2016, 7, 638. [Google Scholar] [CrossRef]
- Fox, E.M. , Leonard, N., Jordan, K. Physiological and transcriptional characterization of persistent and nonpersistent Listeria monocytogenes isolates. Appl. Environ. Microbiol. 2011, 77, 6559–6569. [Google Scholar] [CrossRef] [PubMed]
- Capita, R. , Riesco-Peláez, F., Alonso-Hernando, A., Alonso-Calleja, C. Exposure of Escherichia coli ATCC 12806 to sublethal concentrations of food-grade biocides influences its ability to form biofilm, resistance to antimicrobials, and ultrastructure. Appl. Environ. Microbiol. 2014, 80, 1268–1280. [Google Scholar] [CrossRef]
- Veasey, S. , Muriana, P.M. Evaluation of electrolytically-generated hypochlorous acid (‘electrolyzed water’) for sanitation of meat and meat-contact surfaces. Foods 2016, 5, 42. [Google Scholar] [CrossRef]
- Travier, L. , Guadagnini, S., Gouin, E., Dufour, A., Chenal-Francisque, V., Cossart, P., Olivo-Marin, J.C., Ghigo, J.M., Disson, O., Lecuit, M. ActA Promotes Listeria monocytogenes Aggregation, Intestinal Colonization and Carriage. PLoS Pathog. 2013, 9, e1003131. [Google Scholar] [CrossRef]
- Price, R. , Jayeola, V., Niedermeyer, J., Parsons, C., Kathariou, S. The Listeria monocytogenes key virulence determinants hly and prfa are involved in biofilm formation and aggregation but not colonization of fresh produce. Pathogens 2018, 7, 18. [Google Scholar] [CrossRef]
- Colagiorgi, A. , Di Ciccio, P., Zanardi, E., Ghidini, S., Ianieri, A. A Look inside the Listeria monocytogenes Biofilms Extracellular Matrix. Microorganisms 2016, 4, 22. [Google Scholar] [CrossRef]
- Castañeda-Ruelas, G. , Eslava-Campos, C., Castro-del Campo, N., León-Félix, J., Chaidez-Quiroz, C. Listeriosis en México: importancia clínica y epidemiológica. Salud Pública Méx. 2014, 56, 654–659. [Google Scholar]
- Disson, O. , Moura, A., Lecuit, M. Making Sense of the Biodiversity and Virulence of Listeria monocytogenes. Trends Microbiol. 2021, 29, 811–822. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 26th ed.; CLSI Supplement M100S; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- Blasco, M.D. , Esteve, C., Alcaide, E. Multiresistant waterborne pathogens isolated from water reservoirs and cooling systems. J. Appl. Microbiol. 2008, 105, 469–475. [Google Scholar] [CrossRef]
- Avila-Novoa, M.G. , Navarrete-Sahagún, V., González-Gómez, J.P., Novoa-Valdovinos, C., Guerrero-Medina, P.J., García-Frutos, R., Martínez-Chávez, L., Martínez-Gonzáles, N.E., Gutiérrez-Lomelí, M. Conditions of in vitro biofilm formation by serogroups of Listeria monocytogenes isolated from hass avocados sold at markets in Mexico. Foods 2021, 10, 2097. [Google Scholar]
- Stepanović, S. , Ćirković, I., Ranin, L., Švabić-Vlahović, M. Biofilm formation by Salmonella spp. and Listeria monocytogenes on plastic surface. Lett. Appl. Microbiol. 2004, 38, 428–432. [Google Scholar] [CrossRef]

Table 1.
Genetic analysis and antibiotic resistance of L. monocytogenes isolates.
| Strain no. | Source | Genetic determinants of virulence | Phylogenetic groups | Serotype | CdCl2 | CPF | Antimicrobial resistance genes | Antibiotic Resistance Pattern | MAR index |
|---|---|---|---|---|---|---|---|---|---|
| Lm-11 | Vegetable | LIPI-1 + LIPI-2 | II.2 | 1/2b | R | S | --- | PE-CF-AM-CFX-DC | 0.38 |
| Lm-14 | Vegetable | LIPI-1 + LIPI-2 | II.2 | 1/2b | R | I | Ide | PE-CF-AM-CFX-DC | 0.38 |
| Lm-13 | Vegetable | LIPI-1+ LIPI-2 | II.2 | 1/2b | R | S | msrA + tetM | PE-CF-AM-CFX-DC-TE | 0.46 |
| Lm-17 | Vegetable | LIPI-1 + LIPI-2 | II.2 | 1/2b | R | I | Ide | PE-CF-AM-CFX-DC | 0.38 |
| Lm-18 | Vegetable | LIPI-1 + LIPI-2 | II.2 | 1/2b | R | I | Ide | PE-CF-AM-CFX-DC-CLM | 0.46 |
| Lm-42 | Fruit | LIPI-1+ LIPI-2 | II.2 | 1/2b | S | S | tetM | PE-CF-AM-CFX-DC-CLM-TE | 0.53 |
| Lm-43 | Fruit | LIPI-1+ LIPI-2 | II.2 | 1/2b | S | S | --- | PE-CF-AM-CFX-DC-TE | 0.46 |
| Lm-68 | Fruit | LIPI-1 + LIPI-2 | II.2 | 1/2b | S | I | Ide + msrA | PE-CF-AM-CFX-DC | 0.38 |
| Lm-136 | Fruit | LIPI-1 + LIPI-2 | II.2 | 1/2b | S | I | Ide | PE-CF-AM-CFX-DC-CLM | 0.46 |
| Lm-133 | Fruit | LIPI-1 + LIPI-2 | II.2 | 1/2b | S | S | --- | PE-CF-AM-CFX-DC-CLM | 0.46 |
| Lm-147 | Fruit | LIPI-1 + LIPI-2 | II.2 | 1/2b | S | R | Ide + tetM | PE-CF-AM-CFX-DC-CPF-CLM-TE | 0.61 |
| Lm-138 | Fruit | LIPI-1 + LIPI-2 | II.2 | 1/2b | R | I | Ide | PE-CF-AM-CFX-DC-CLM | 0.46 |
| Lm-19 | Vegetable | LIPI-1+ LIPI-2 + LIPI-3 | I.1 | 1/2a | S | S | tetM | PE-CF-AM-CFX-DC-TE | 0.46 |
| Lm-24 | Vegetable | LIPI-1+ LIPI-2 + LIPI-3 | I.1 | 1/2a | R | S | --- | PE-CF-AM-CFX-DC-CLM | 0.46 |
| Lm-15 | Vegetable | LIPI-1+LIPI-2 + LIPI-3 | I.1 | 1/2a | R | S | msrA | PE-CF-AM-CFX-DC-CLM | 0.46 |
| Lm-27 | Vegetable | LIPI-1 + LIPI-2 + LIPI-3+ LIPI-4 | I.1 | 1/2a | R | R | Ide + tetM | PE-CF-AM-CFX-DC-CPF-CLM-TE | 0.61 |
| Lm-41 | Vegetable | LIPI-1 + LIPI-2+ LIPI-3+ LIPI-4 | I.1 | 1/2a | R | I | Ide | AM-CFX-DC | 0.23 |
LIPI-1: Isolates harboring virulence genes (prfA, hly, plcA, plcB, mpl, actA); LIPI-2: Isolates harboring virulence genes (inlA, inlB, inlC, inlJ); LIPI-3: Isolates harboring virulence genes (llsA, llsG, llsH, llsX, llsB, llsY, llsD, and llsP); LIPI-4: Isolates harboring virulence genes (licC, licB, licA, and glva); I, Intermediate resistance to ciprofloxacin; R, resistance to ciprofloxacin or CdCl2; S, Susceptible to ciprofloxacin or CdCl2; CPF, ciprofloxacin; AM, ampicillin; CLM, clindamycin; CF, cephalothin; CFX, cefotaxime; CL, chloramphenicol; GE, gentamicin; E, erythromycin; TE, tetracycline; VA, vancomycin; SXT, trimethoprim-sulfamethoxazole; PE, penicillin; DC, dicloxacillin.
Table 2.
Antimicrobial susceptibility test of L. monocytogenes.
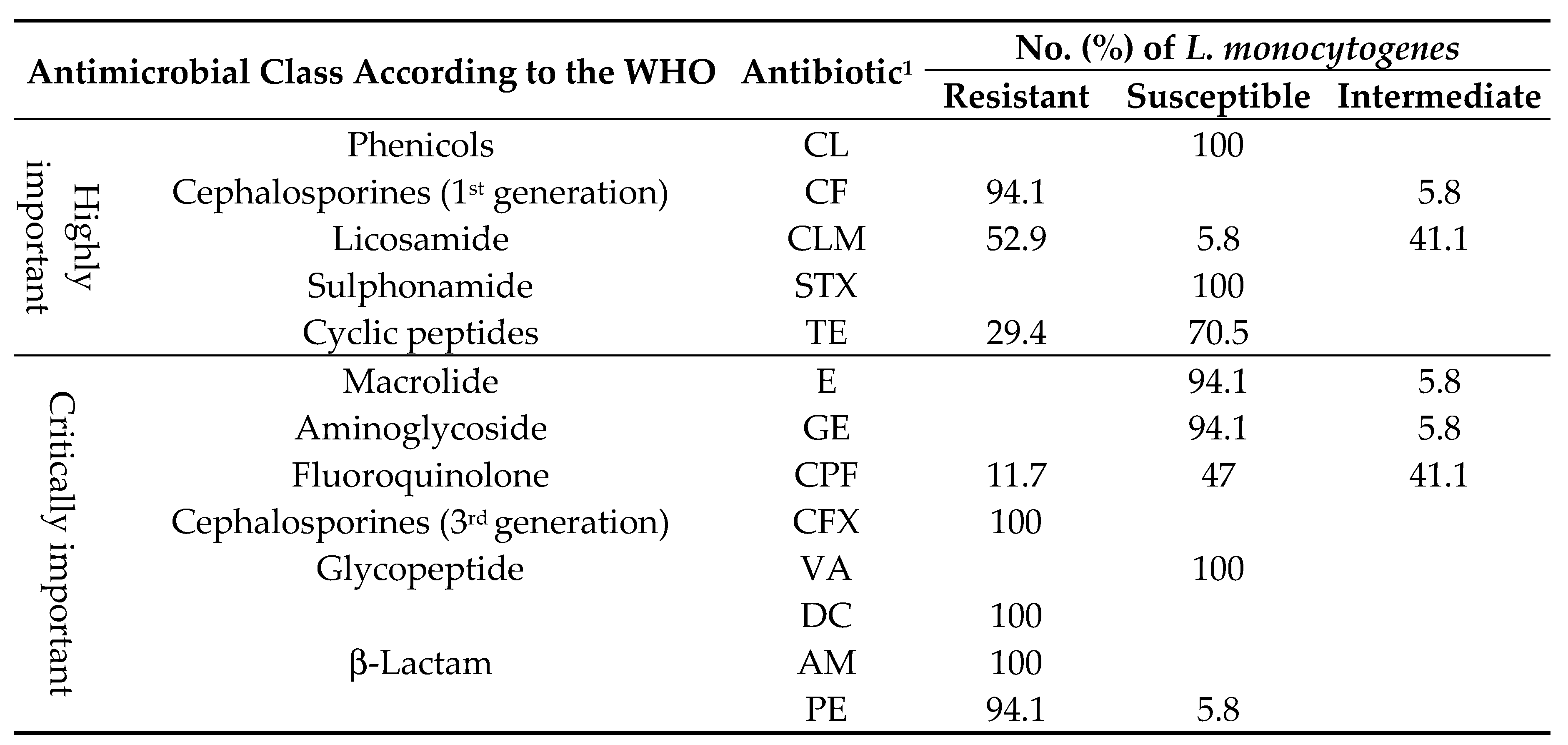
1 AM, ampicillin; CLM, clindamycin; CF, cephalothin; CFX, cefotaxime; CPF, ciprofloxacin; CL, chloramphenicol; GE, gentamicin; E, erythromycin; TE, tetracycline; VA, vancomycin; SXT, trimethoprim-sulfamethoxazole; PE, penicillin; DC, dicloxacillin.
Table 3.
Minimum inhibitory concentration values of BC and CdCl2 in L. monocytogenes strains in relation with biofilm formation.
Table 3.
Minimum inhibitory concentration values of BC and CdCl2 in L. monocytogenes strains in relation with biofilm formation.
| Strain no. | Phylogenetic groups | Serotype | MIC (µg/mL) |
BC | CdCl2 | Biofilm formation (Microtiter plate assays) |
|---|---|---|---|---|---|---|
| Lm-11 | II.2 | 1/2b | 6.2 | █ | █ | Moderate biofilm |
| Lm-14 | II.2 | 1/2b | 6.2 | █ | █ | Moderate biofilm |
| Lm-13 | II.2 | 1/2b | 6.2 | █ | █ | Moderate biofilm |
| Lm-17 | II.2 | 1/2b | 6.2 | █ | █ | Moderate biofilm |
| Lm-18 | II.2 | 1/2b | 3.1 | ☐ | █ | Moderate biofilm |
| Lm-27 | I.1 | 1/2a | 3.1 | ☐ | █ | Moderate biofilm |
| Lm-42 | II.2 | 1/2b | 3.1 | ☐ | ☐ | Moderate biofilm |
| Lm-43 | II.2 | 1/2b | 3.1 | ☐ | ☐ | Moderate biofilm |
| Lm-68 | II.2 | 1/2b | 3.1 | ☐ | ☐ | Moderate biofilm |
| Lm-133 | II.2 | 1/2b | 3.1 | ☐ | ☐ | Moderate biofilm |
| Lm-136 | II.2 | 1/2b | 3.1 | ☐ | ☐ | Moderate biofilm |
| Lm-138 | II.2 | 1/2b | 1.5 | ☐ | █ | Moderate biofilm |
| Lm-41 | I.1 | 1/2a | 0.7 | ☐ | █ | Moderate biofilm |
| Lm-147 | II.2 | 1/2b | 0.7 | ☐ | ☐ | Moderate biofilm |
| Lm-19 | I.1 | 1/2a | 0.7 | ☐ | ☐ | Moderate biofilm |
| Lm-24 | I.1 | 1/2a | 1.5 | ☐ | █ | Moderate biofilm |
| Lm-15 | I.1 | 1/2a | 1.5 | ☐ | █ | Moderate biofilm |
Black squares indicate the resistance to BC (CMI ≥ 6 µg/mL,) or CdCl2 (CMI ≥ 70 µg/mL).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



